lipids metabolism in plants
TRANSCRIPT

Lipids metabolism
Somayeh Hooshyar

Definition of lipids
• Lipids are a structurally diverse group of hydrophobic molecules that are preferentially soluble in non-aqueous solvents such as chloroform
• Lipids are highly hydrophobic molecules with major roles in membrane structure, storage of carbon and energy reserves, plant protection and cell signaling.

Functions of lipid molecules in plantsLipid types involved Function
Glycerolipids, Sphingolipids, Sterols Membrane structural component
Triacylglycerols, Waxes Storage compounds
Chlorophyll and other pigmentsUbiquinone, plastoquinone
Compounds active in electron transfer reactions
Carotenoids photoprotection
Tocopherols Protection of membranes against damage from free radicals
Long chain and very long chain fatty acids and their derivatives (cutin, suberin, surface waxes)Teriterpens
Water proofing and surface protection
Protein modification
Addition of membrane anchors
Mainly 14:0 and 16:0 fatty acids Acylation
Farnesyl and geranyl geranyl pyrophosphate Prenylation
Phosphatidylinositol, ceramide Other membrane anchor components
Dolicol Glycosilation
Signaling
Abscisic acid, gibberllins, brassinosteroid18:3 fatty acid precursors of jasmonateInositol phosphates, diacylglycerols
Internal
Jasmonate , Volatile insect attractants External
Essential oilsLatex components (rubber, etc)Resin components (terpens)
Defense and antifeeding compounds
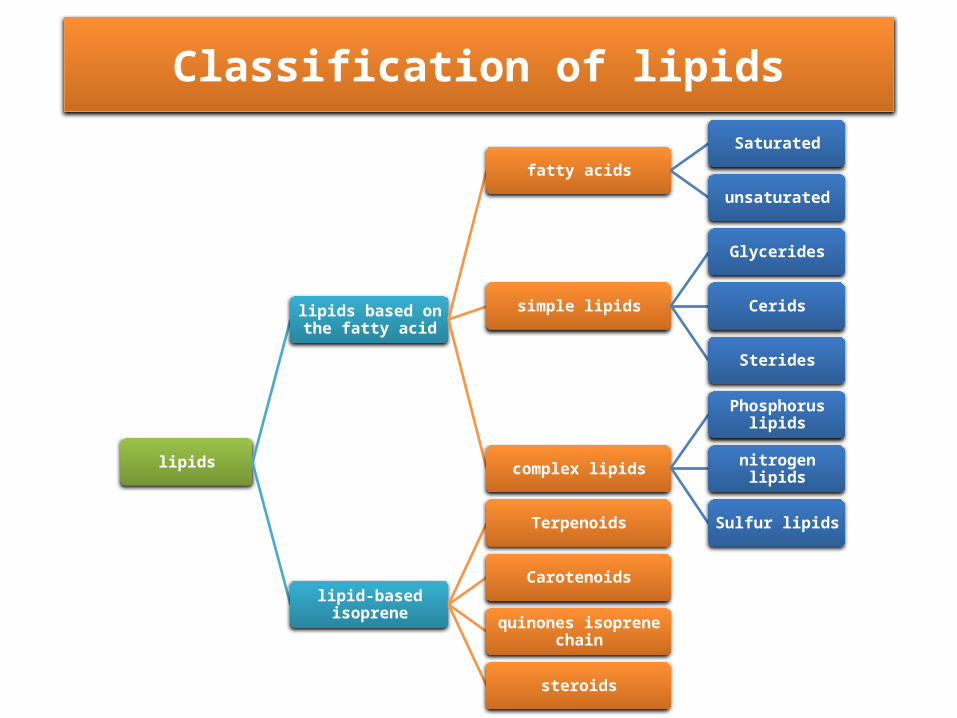
lipids
lipids based on the fatty acid
fatty acids
Saturated
unsaturated
simple lipids
Glycerides
Cerids
Sterides
complex lipids
Phosphorus lipids
nitrogen lipids
Sulfur lipids
lipid-based isoprene
Terpenoids
Carotenoids
quinones isoprene chain
steroids
Classification of lipids



• fatty acids, which are carboxylic acids with long hydrocarbon tails. More than 200 different fatty acids have been identified in plants.
Fatty Acid Synthesis and Export

• Plant fatty acid (FA) synthesis differs from almost all other eukaryotes in two fundamental features.
First, unlike the cytosolic location in other kingdoms, FAs are produced in the plastid compartment of plant cells: chloroplasts in green tissues and proplastids (or leucoplasts) in non-green tissues.
Second, the plant FA synthase (FAS) is a dissociable complex with separate proteins for the acyl carrier protein (ACP) and each enzyme.
• This biosynthetic pathway is not restricted to specific tissues or organs but found in every cell of the plant.
Fatty Acid Synthesis and Export

Fatty Acid Synthesis and Export
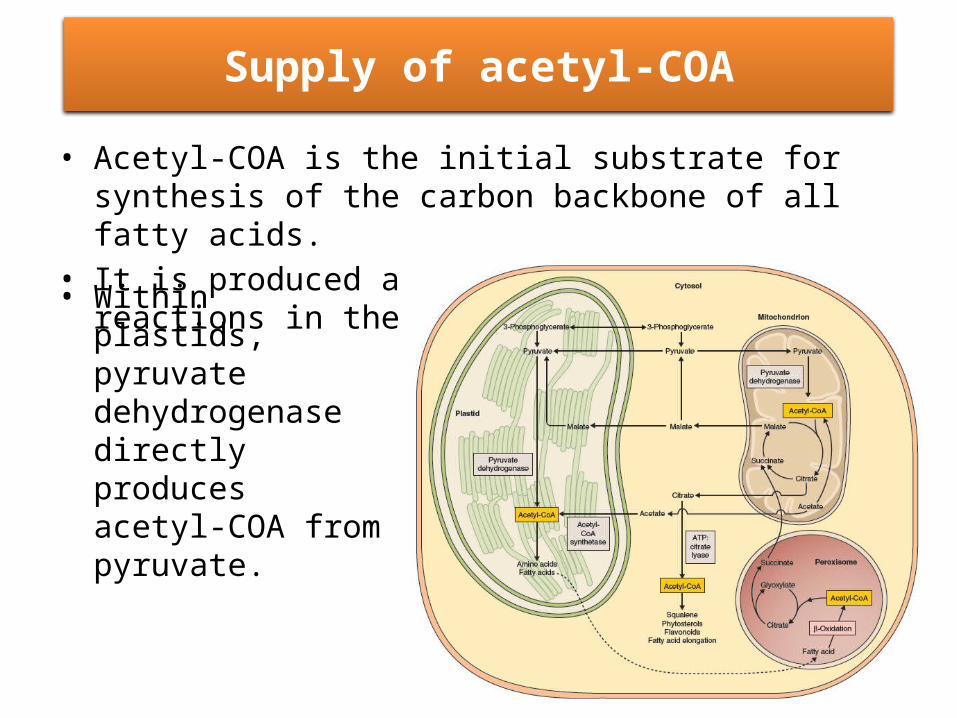
Supply of acetyl-COA
• Acetyl-COA is the initial substrate for synthesis of the carbon backbone of all fatty acids.
• It is produced and consumed by many of reactions in the cell.
• Within plastids, pyruvate dehydrogenase directly produces acetyl-COA from pyruvate.

Supply of acetyl-COA

Pyruvate Dehydrogenase Complex
• The PDHC contains three components: E1 (pyruvate dehydrogenase, PDH, composed of E1α and E1β
subunits), E2 (dihydrolipoyl acyltransferase, DHLAT) E3 (dihydrolipoamide dehydrogenase, LPD)

PDH reaction
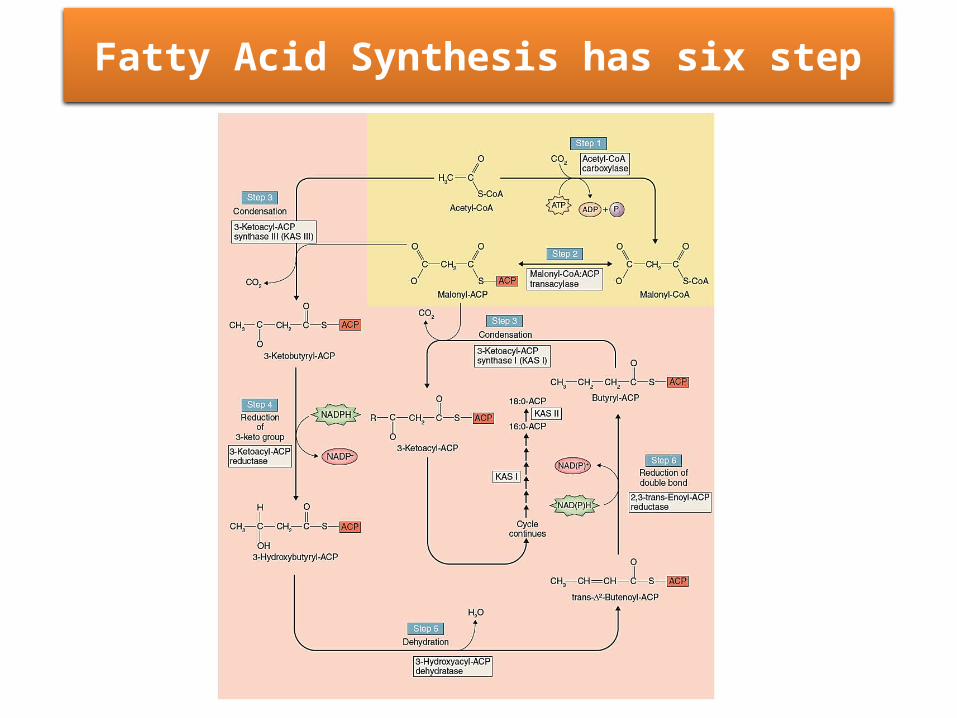
Fatty Acid Synthesis has six step

Step 1

• The first step in fatty acid synthesis is the formation of malonyl-CoA from acetyl-CoA and bicarbonate by acetyl-CoA carboxylase (ACCase).
• In most plants the plastid form of ACCase has four subunits:
1. Biotin carboxyl carrier protein (BCCP)2. Biotin carboxylase (BC)3. Alpha subunit of carboxyltransferase (α-CT)4. Beta subunit of carboxyltransferase (b-CT)
Step 1

ACCase reaction

ACCase reaction

• Three of the subunits are encoded by nuclear genes, whereas the fourth is encoded in the chloroplast genome
• Acetyl-CoA carboxylase (ACCase) is a key rate-determining step that controls FA biosynthesis. ACCase activity is under complex regulation by light, phosphorylation, thioredoxin, PII protein, and product feedback control.
• Two different forms of ACCase occur in plants: A homomeric form (multisubunit type) located in plastids of
all plants except grass family A heteromeric form (multifunctional type) located in cytosol
of all plants and plastids of grass family
Acetyl-CoA carboxylase (ACCase)

• Fatty acid synthase (FAS) refers to all enzyme activities in fatty acid biosynthesis except ACCase.
• Two different types of FAS are found in nature: Type I: in animals and yeasto A single multifunctional enzyme complex characterized by
large subunits (250 KDa)o Functions like a protein complex Type II: in plants and most bacteriao In which each enzyme activity resides on an individual protein
that can be readily separated from the other activities participating in fatty acid synthesis.
o Type II FAS also includes ACP.o Functions like a metabolic pathway.
Fatty acid synthase

Step 2

• Before entering the fatty acid synthesis pathway, the malonyl group of malonyl-CoA produced by ACC has to be transferred from CoA to ACP.
• This transfer is catalyzed by a malonyl-CoA:ACP malonyltransferase (MCMT).
Step 2

• Acyl carrier protein (ACP) is a small protein about 80 amino acids long and contains a phosphopantethine prostetic group covalently linked to a serine.
Acyl carrier protein (ACP)

• Fatty acids are grown by sequential condensation of two-carbon units by enzymes of the fatty acid synthase complex.
• During each cycle, four reactions occur: Condensation Reduction Dehydration Reduction • Synthesis of a C16 fatty acid requires that the cycle be
repeated seven times.

Step 3: condensation

• The condensation reaction to form a new C-C bond is catalyzed by 3-ketoacyl-ACP synthase (KAS)
• All plants contain three isoforms of KAS KASI is most active with C4-C10 acyl-ACP KASII accepts only longer chain C10-C16 acyl-ACP KASIII is most active with acetyl-CoA rather than acyl-
ACP• In arabidopsis During the first turn of the cycle, the
condensation reaction is catalyzed by ketoacyl-ACP synthase (KAS) III.
• For the next six turns of the cycle, the condensation reaction is catalyzed by isoform I of KAS. Finally, KAS II is used during the conversion of 16:0 to 18:0.
Step 3: condensation

Step 4: Reduction

Step 5: Dehydration

Step 6: Reduction

• Regulation of transcriptional level a single transcription factor that directly controls the
transcriptional activation of the fatty acid biosynthetic pathway: it is called WRINKLED1 (WRI1)
• Regulation of optimization of enzyme activity the PII/AtGLB1 protein was shown to interact with BCCP
subunits of heteromeric HtACCase in a 2-oxoglutarate-dependent manner and to control ACCase activity by reducing the Vmax of the enzyme.
Control of the fatty acid biosynthetic pathway

Fatty Acid Elongation, Desaturation, and Export From Plastid
• C16:0 fatty acids produced by the fatty acid synthesis pathway can enter three possible reactions.
First, they can be elongated by an additional cycle of fatty acid synthesis. In these cases, KAS II is used during the conversion of 16:0 to 18:0.
Alternatively, C16:0 can enter the prokaryotic glycerolipid pathway.
Finally, 16:0-ACP can be hydrolyzed by thioesterase enzymes to release free fatty acids that are exported from the plastid.

Fatty Acid Desaturation

• Plants have two type of fatty acyl desaturases:• A soluble, plastid localized desaturase called Δ9 stearoyl-
ACP desaturase (SAD)• Most fatty acyl desaturases are unsoluble integral
membrane proteins localized in plastid or ER • FAD2-8 and FAB2 are fatty acid desaturases of
arabidopsis• Temperature factor determine glycerolipid desaturation in
most plant.
Fatty Acid Desaturation

Fatty Acid Export From Plastid
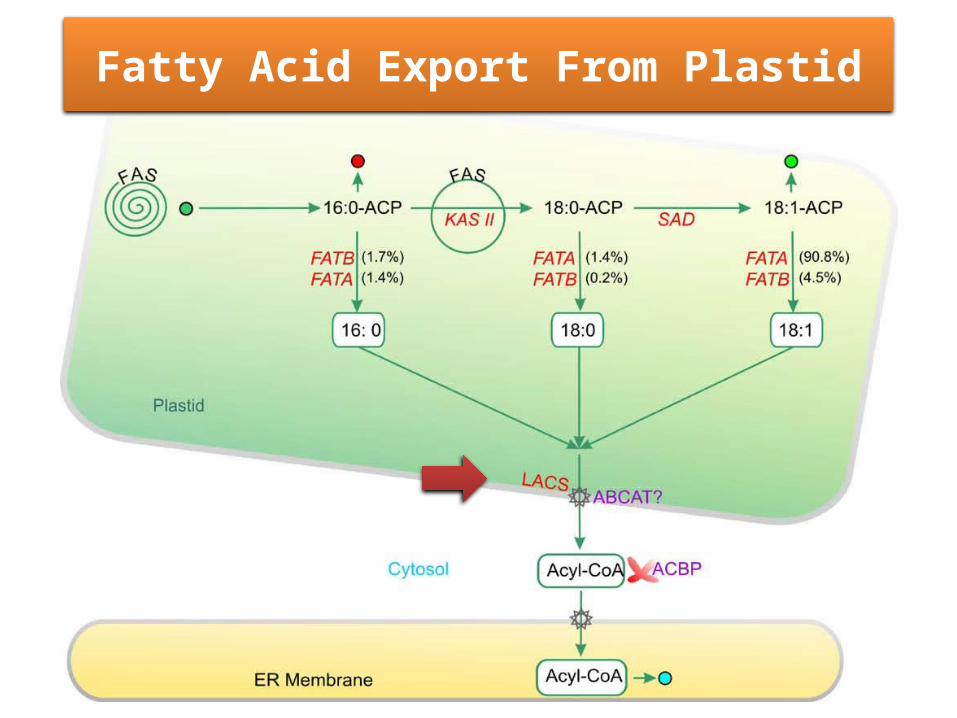
Fatty Acid Export From Plastid

Fatty Acid Export From Plastid

• Organisms store excess energy and release it when they suffer from energy deprivation. For most eukaryotes, the preferred storage compounds are lipids in the form of triacylglycerols (TAGs).
Synthesis of storage lipids
• Triacylglycerols are reserves of energy and carbon, found mostly in seeds and pollen. They are strongly non-polar. a triacylglycerol molecule consists of a glycerol molecule with one fatty acid molecule esterified to each of its three carbons.

Comparison of energy yield from metabolism of fatty acids and carbohydrates to CO2 and H2O

• Two pathways are recognized for TAG synthesis in ER of plants:
Acyl-CoA-dependent pathway (The Kennedy pathway)
Acyl-CoA-independent pathway
TAG biosynthesis in plants

Acyl-CoA-dependent pathway (The Kennedy pathway)
ER

Acyl-CoA-independent pathway
ER

• Numerous transcription factors are involved in a complex and hierarchical system integrating TAG production with other aspects of seed and embryo development.
• WRI1 plays a major role in controlling expression of genes involved in seed oil biosynthesis.
• LEC1 act upstream of WRI1.
Regulation of TAG biosynthesis

• Triacylglycerols accumulate in discrete subcellular organelles called oil bodies.
• These lipid particles are present in seeds, flowers, pollen and fruit of higher plants.
• Each OB has a matrix of TAGs surrounded by a layer of phospholipids (PLs) and different proteins.
• The most abundant of these are the oleosins, but others such as caleosins and steroleosins are also present.
Oil body (oleosome)

• Oleosins in seeds are small proteins of about 15 to 26 kDa that contain a sequence of 70-80 hydrophobic amino acids toward the middle of the protein and have more hydrophilic N- and C- terminal domains.
• They completely cover the surface of the subcellular OB.• Caleosins also appear to play a role in TAG mobilization
during germination, possibly by facilitating interactions with vacuoles.
• Steroleosins, in addition to an oil body–anchoring domain, possess a sterol-binding dehydrogenase that might play a role in signal transduction.
Oil body (oleosome)

Oil body (oleosome)
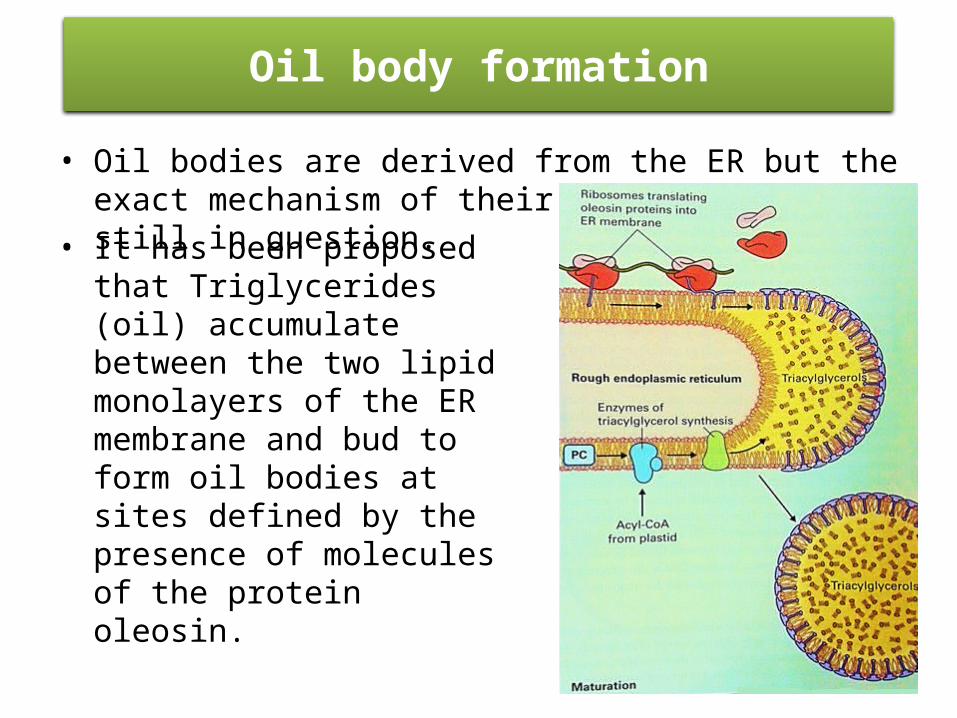
• Oil bodies are derived from the ER but the exact mechanism of their biogenesis is still in question.
Oil body formation
• It has been proposed that Triglycerides (oil) accumulate between the two lipid monolayers of the ER membrane and bud to form oil bodies at sites defined by the presence of molecules of the protein oleosin.

• There are special oil bodies in tapetum cells of anthers in Arabidopsis and Brassica.
Oil body (tapetosome)
• These oleosomes are present in a novel, neutral lipid-containing organelle, which has been termed the tapetosome because of its unique presence in tapetum

• The breakdown of stored triacylglycerols in oil-storing seeds takes place in the oil body and peroxisome.
• In the oil body, triacylglycerols are degraded by lipase (TAGL) to glycerol and fatty acids.
• Fatty acids are transported to the peroxisome where they are further degraded to acetyl-CoA by the β-oxidation pathway.
Catabolism of storage lipids

• In plants, the oxidation of fatty acids takes place in peroxisomes of germinating seeds and leaves.
• Fatty acids are activated by the addition of coenzyme A (CoA) to form fatty acyl-CoA molecules. The fatty acyl-CoA is broken down to two carbon acetyl units in a cyclic process. In each pass through the sequence, one acetyl residue is removed in the form of acetyl-CoA from the carboxyl end of an acyl-CoA.
• Seven passes through the cycle are required to oxidize a C16 fatty acid to eight molecules of acetyl-CoA.
β-oxidation pathway

Refrences







Thank you