linoleic acid, butnot casein - pnas · linoleic acid, butnotcortisol, ... factors, and other agents...
TRANSCRIPT

Proc. Natl. Acad. Sci. USAVol. 84, pp. 8448-8452, December 1987Cell Biology
Linoleic acid, but not cortisol, stimulates accumulation of casein bymouse mammary epithelial cells in serum-free collagen gel culture
(explant culture/coculture/ELISA/prolactin/glucocorticoid)
BRETT K. LEVAY-YOUNG*, GAUTAM K. BANDYOPADHYAY, AND SATYABRATA NANDICancer Research Laboratory and Department of Zoology, University of California, Berkeley, CA 94720
Communicated by Howard A. Bern, July 24, 1987 (received for review April 24, 1987)
ABSTRACT A two-step culture system has been developedto analyze the role of hormones in casein accumulation bymammary epithelial cells obtained from adrenaiectomized andovariectomized adult virgin mice. In the first step cells aregrown inside collagen gel in medium containing insulin, epi-dermal growth factor (EGF), and linoleic acid for 9 days; theseconditions stimulate very little casein accumulation. Followingthis growth phase the gels are released to float in mediumcontaining insulin, prolactin, and linoleic acid. During thissecond phase the mammary cells will accumulate large amountsof casein, but only in the simultaneous presence of insulin,prolactin, and linoleic acid; in the absence of linoleic acid caseinaccumulation is greatly reduced. The casein accumulation isnot dependent on the presenceofthe glucocorticoid cortisol andwill occur in spite of the presence of the antiglucocorticoidagent RU 38 486. To determine if the response to cortisolobserved in organ culture by other investigators might bemediated by stromal cells, epithelial cells were grown incollagen gel under fatty acid-free conditions and then cocul-tured with explants of mammary fat pads from adult virginmice with or without mammary parenchyma. The cocultureswere performed in fatty acid-free medium containing insulinand prolactin with or without cortisol. In the majority ofexperiments the mammary epithelial cells in the collagen gelaccumulate more casein in the presence of cortisol than in itsabsence, irrespective of the presence ofmammary parenchymain the explant. Thus, mammary epithelial cells are directlydependent on insulin and prolactin for casein accumulation andindirectly dependent on cortisol by means of its effect on thestromal cells. This cortisol effect may be to cause release intothe medium of linoleic acid or a metabolic product of linoleicacid from the stromal cells.
Despite its long history, the study of the effects of hormonesand growth factors on mammary epithelium in culture has notfully defined the roles of these substances in mammarydifferentiation (for review, see refs. 1 and 2). Extensivestudies (3-10) in fragment organ culture using mammarytissue from either mouse or rat under defined, serum-freeconditions demonstrated that insulin (5, 6), prolactin (3-10),and a glucocorticoid, usually cortisol (7-10), were requiredfor the accumulation of casein protein and mRNA. Wholegland organ culture studies (11-13) have substantiated thisrequirement for the three hormones.Fewer experiments to examine this requirement for glu-
cocorticoid have been performed in cell culture, even thoughcell culture has become widely used to study differentiationin mammary epithelial cells. Casein accumulation and secre-tion in cell culture were demonstrated by Emerman et al. in1977 (14) using cells obtained from midpregnant mice andcultured on top of collagen gel. Casein secretion in this
system has been repeatedly shown to be dependent on thepresence of insulin, prolactin, and glucocorticoid, whencompared to medium containing insulin alone (14-16). Therequirement for the three hormones has been recapitulated incell culture in a variety of culture systems, using cellsobtained from normal virgin or midpregnant animals (17-23).Although some of these investigations have attempted toassess the requirement for or the role of glucocorticoid incasein accumulation or secretion (18, 23), these attemptshave been hampered by the inclusion of serum in the culturemedium and/or the long-term nature of the cultures oftenrequired to obtain detectable differentiation.Our aim has been to develop defined culture conditions
that will allow unambiguous interpretation of experimentalresults in analysis of the role of individual hormones, growthfactors, and other agents in milk product synthesis bymammary epithelial cells. Toward this end we have attempt-ed to determine the minimal requirement for casein accumu-lation by mammary epithelial cells from adrenalectomizedand ovariectomized virgin mice using the serum-free collagengel culture system developed in this laboratory (24-28). SinceDNA synthesis has been shown to be a prerequisite tohormone-induced casein accumulation by mammary epithe-lial cells from virgin mice (29), we have used a two-stepsystem in our studies. In the first step, or growth phase,regressed mammary cells from adrenalectomized and ovar-iectomized mice were subjected to proliferation-stimulatingagents in the absence of lactogenic hormones. In the secondphase the cells were allowed to differentiate in the presenceof lactogenic hormones. Using this system we show that cellsfrom adrenalectomized and ovariectomized mice cultured incollagen gel and exposed to insulin and prolactin are depen-dent on linoleic acid to a much greater extent than on theglucocorticoid cortisol for casein accumulation. However,cortisol regains its effectiveness for enhancing casein accu-mulation if the mammary epithelial cells in collagen gel arecocultured with explants containing mammary stroma inmedium without added linoleic acid.
MATERIALS AND METHODSAnimals. Adult female virgin BALB/c mice (4 months old)
were obtained from the University of California CancerResearch Laboratory breeding colony and adrenalectomizedand ovariectomized by way of middorsal incision underphenobarbital anesthesia. The ablated mice were maintainedfor at least 1 month, but no more than 6 weeks, on 0.9% NaClsolution and Purina Rat Chow ad libitum. Completeness ofoperation was assessed visually when the mammary tissuewas taken for dissociation, and mice with obvious accessoryor residual adrenals were excluded.
Abbreviations: EGF, epidermal growth factor; BSA, bovine serumalbumin.*To whom reprint requests should be addressed.
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
8448

Proc. Natl. Acad. Sci. USA 84 (1987) 8449
Epithelial Cell Isolation. The cell dissociation was essen-tially as described (24, 30). Briefly, the mammary glands wereminced and dissociated with collagenase (0.075% CLS II,
Worthington) and further digested with 0.01% Pronase(Calbiochem-Behring). The epithelial cells were separatedfrom stromal cells and cell fragments by centrifugation onpreformed 42% Percoll (Pharmacia) density gradients andused directly for collagen gel culture.
Collagen Gel Culture. Collagen gel culture of mammaryepithelial cells has been extensively described and reviewedelsewhere (24, 30). The serum-free medium, which has alsobeen described (27, 28), was a 1:1 (vol/vol) mixture ofHam'sF12 medium/Dulbecco's modified Eagle's medium (bothfrom GIBCO) containing NaHCO3 at 1.2 g/liter and bufferedwith Hepes at 20 mM. This medium was supplemented withtransferrin, soybean trypsin inhibitor, D-a-tocopherol (allfrom Sigma), trace element mixture, antibiotics, 10 pg ofinsulin per ml (Sigma), and fatty acid-free bovine serumalbumin (BSA) (Armour, Tarrytown, NY) alone or conjugat-ed to the sodium salt of linoleic acid [linoleate-BSA; ref. 28(Sigma)]. These media, with and without linoleic acid, will bereferred to as the basal media. The final concentration ofBSAwas 160 ,g/ml in all cultures, and, when used, the concen-tration of fatty acid was 10 ttg/ml. The medium was changedevery 2 days.The cells were cultured in the basal medium with epidermal
growth factor (EGF) at 10 ng/ml (growth medium) for 9 days.This was followed by rimming the gels with a small spatula torelease them to float in the medium (31) and replacement ofthe growth medium with basal medium containing prolactinat 1 pug/ml and the supplements noted below (differentiationmedium). EGF was from Collaborative Research (Waltham,MA), ovine prolactin was NIADDK oPrl PS-16, a gift of theNational Institute of Arthritis, Diabetes, and Digestive andKidney Diseases and the National Hormone and PituitaryProgram, cortisol was from Sigma, and RU 38 486 was thegenerous gift of R. Deraedt (Roussel, Paris). Cultures wereterminated at the time of release and after 6 days in differ-entiation medium by blotting the collagen gel on a stack ofpaper towels to remove trapped medium (32) and freezing theresulting membrane-like collagen/cell combination at -70°Cfor future assay.
Explant Preparation and Coculture. Explants of mousemammary fat pads were prepared from 4- to 6-month-oldfemale virgin BALB/c mice as described (3). The animalsused were either normal intact virgin mice or virgin mice thathad the parenchyma surgically removed at 3 weeks of age,leaving a fat pad with only stromal tissue (cleared fat pads;ref. 33). The fat pads were aseptically removed and cut to1-mm3 fragments (explants). These explants were placed onstainless steel grids at the medium/gas interface, over thecollagen gels.Assay Procedures. The effect of all culture treatments was
determined on triplicate cultures in each experiment. Eachexperiment was repeated 3-10 times. Data shown are from a
typical experiment. Prior to assay the frozen gels were
homogenized in phosphate-buffered saline (PBS, 6.6 mMphosphate, pH 7.4/145 mM NaCl) with 5 mM EDTA/1%Triton X-100 at 4°C by 5 sec of insonation from a BransonS-75 sonicator. The homogenate was centrifuged at 12,000 x
g for 30 min at 4°C. The clear supernatant was used fora-casein ELISA and NaDodSO4/PAGE. The collagen pelletswere dissolved in PBS with 1 M acetic acid, and the insolublematerial was pelleted by centrifugation at 12,000 x g for 10min at 25°C. This pellet was assayed for DNA content bydiaminobenzoic acid (Aldrich) assay after the method ofHinegardner (34). Because the cells in primary culture are
overwhelmingly diploid, the DNA determination reflects cellnumber (35).
The ELISA was developed from the method of Engvall etal. (36). Purified a-casein (37) was coated onto Costar EIAplates at 1 ng per well in 0.056 M carbonate buffer (pH 9.8).After blocking excess binding capacity with BSA, dilutions ofstandard a-casein or the unknown samples were made in abuffer of distilled water/1% Triton X-100/0.1% BSA andadded to the plates at 100 1.l per well. The antiserum waspreviously developed in this laboratory to a-casein (37) andwas made up at 1:100,000 dilution in a buffer of 2x PBS/2%normal goat serum/0.05% Tween 20. To begin the assay, 100Aul of this solution was added to each well (final antiserumdilution, 1:200,000). The plates were incubated for 44 hr at40C, washed, and incubated with alkaline phosphatase-con-jugated second antibody (Sigma or Miles) at 1:1000 inPBS/0.1% BSA/0.05% Tween 20 for 2 hr at 250C. Afterwashing, the color reaction was carried out for 1-2 hr at 250Cwith p-nitrophenyl phosphate in 1 M diethanolamine buffer(pH 9.8). The reaction product was measured by absorbanceat 405 nm on a Bio-Tek (Burlington, VT) EL305 ELISA platereader.
Protein blotting was done according to the method ofTowbin et al. (38). Aliquots of the culture homogenatescontaining 25 jig of protein were precipitated with metha-nol/acetone (1:1) and dissolved in 8 M urea sample buffer(18). Electrophoresis was performed on 12.5% acrylamidegels (39), after which the proteins were electrophoreticallyblotted to nitrocellulose. The primary antibody was a ratmonoclonal anti-,8-casein (ref. 20; gift of F. Stockdale); thiswas followed by horseradish peroxidase-conjugated rabbitanti-rat antiserum (Boehringer Mannheim). The position ofthe bound antibody was revealed by incubation with 4-chloro-1-naphthol (Sigma) and H202.
RESULTSEffect of Linoleic Acid-Containing Medium. Cells obtained
from adrenalectomized and ovariectomized virgin mice grewin serum-free collagen gel culture in medium containinginsulin, EGF, and linoleate-BSA. Cells assayed before cul-ture contain virtually undetectable amounts of casein. Table1 shows that after a growth phase of 9 days cultured cellscontain small amounts of a-casein. When the collagen gelsare released to float in the medium and the supplements arechanged to insulin, linoleate-BSA, and prolactin, much largeramounts of casein accumulate in the cells. This accumulationis not reproducibly increased by the addition of cortisol, asseen in Table 1. Because the cells come from adrenalecto-
Table 1. Growth and casein accumulation in cells fromadrenalectomized and ovariectomized virgin mice aftergrowth in linoleic acid-containing medium
CellGrowth Differentiation number a-Casein, ngmedium medium x i0-5 per 106 cells
* - 1.47 ± 0.38 NDI, EGF 3.28 ± 1.2 9.02 ± 6.25I, EGF I 3.98 ± 1.2 4.95 ± 3.7I, EGF I, PRL 4.32 ± 0.5 781 ± 141I, EGF I, PRL, F 4.26 + 0.7 519 ± 111
Cells obtained by collagenase digestion were either frozen imme-diately or grown in collagen gel with insulin and EGF for 9 days. Allcultures contained linoleate-BSA. One set ofcultures was terminatedat the end of the growth phase, and the remainder were released andthe medium supplements were changed to the agents listed underDifferentiation medium, again combined with linoleate-BSA. After 6more days, the cultures were terminated and cell number and caseincontent were determined. All values are presented as mean ± SD; n= 3. ND, not detectable. I, insulin; PRL, prolactin; F, cortisol.*Cells frozen immediately.tCulture terminated at the end of growth phase.
Cell Biology: Levay-Young et al.
8450 Cell Biology: Levay-Young et al.
Table 2. Casein accumulation in cells from adrenalectomized andovariectomized virgin mice after growth in fatty acid-free mediumor linoleic acid-containing medium
Growth Differentiation a-Casein, ngmedium medium per 106cells
I, EGF * 0.888 ± 0.29I, EGF I 0.307 ± 0.24I, EGF I, PRL 13.9 ± 9.4I, EGF I, PRL, F 13.3 ± 13.3I, EGF I, PRL, Lin 245 ± 1741, EGF, Lin I, PRL, Lin 781 ± 141
Cells were grown in collagen gel with insulin and EGF with orwithout linoleate-BSA for 9 days (Growth medium). At the end ofthegrowth phase, one set of cultures was terminated, and the remainderwere released and the medium supplements were changed to theagents listed under Differentiation medium. After 6 more days, thecultures were terminated and casein content was determined. Allvalues are presented as mean ± SD; n = 3. I, insulin; PRL, prolactin;F, cortisol; Lin, linoleic acid.*Culture terminated at the end of growth phase.
mized animals and have been cultured in glucocorticoid-freemedium for 9 days it seems unlikely that the accumulationinduced in this system by insulin, prolactin, and linoleic acidis dependent on cortisol or another glucocorticoid.Table 2 shows that if linoleic acid is omitted throughout the
culture, very little casein accumulates in the cells. Cortisoldid not have any effect under these conditions either. Theeffect of linoleic acid can also be demonstrated in cells grownunder fatty acid-free conditions with insulin and EGF; thiswas followed by gel release and exposure to insulin, prolac-tin, and linoleate-BSA. These data are also presented inTable 2 and clearly show that the cells are still capable ofresponding to linoleic acid after growth in its absence.
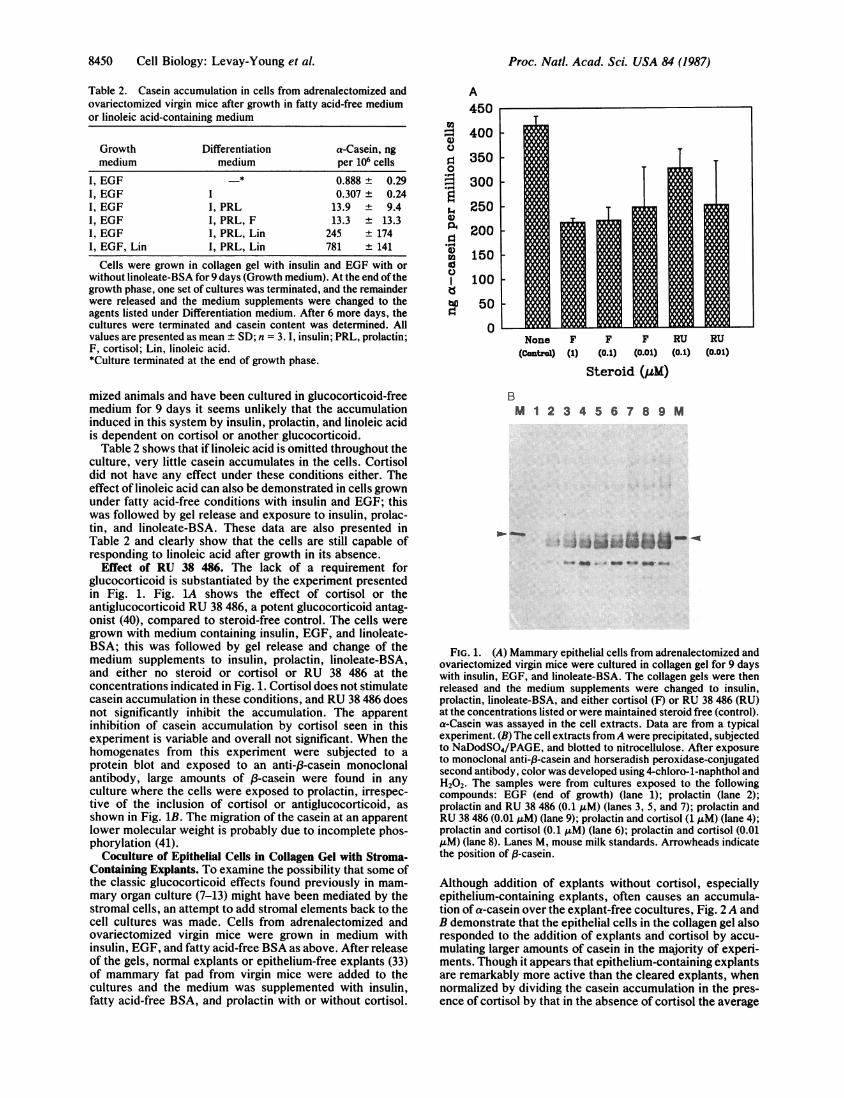
Effect of RU 38 486. The lack of a requirement forglucocorticoid is substantiated by the experiment presentedin Fig. 1. Fig. 1A shows the effect of cortisol or theantiglucocorticoid RU 38 486, a potent glucocorticoid antag-onist (40), compared to steroid-free control. The cells weregrown with medium containing insulin, EGF, and linoleate-BSA; this was followed by gel release and change of themedium supplements to insulin, prolactin, linoleate-BSA,and either no steroid or cortisol or RU 38 486 at theconcentrations indicated in Fig. 1. Cortisol does not stimulatecasein accumulation in these conditions, and RU 38 486 doesnot significantly inhibit the accumulation. The apparentinhibition of casein accumulation by cortisol seen in thisexperiment is variable and overall not significant. When thehomogenates from this experiment were subjected to aprotein blot and exposed to an anti-/3-casein monoclonalantibody, large amounts of f3-casein were found in anyculture where the cells were exposed to prolactin, irrespec-tive of the inclusion of cortisol or antiglucocorticoid, asshown in Fig. 1B. The migration of the casein at an apparentlower molecular weight is probably due to incomplete phos-phorylation (41).
Coculture of Epithelial Cells in Collagen Gel with Stroma-Containing Explants. To examine the possibility that some ofthe classic glucocorticoid effects found previously in mam-mary organ culture (7-13) might have been mediated by thestromal cells, an attempt to add stromal elements back to thecell cultures was made. Cells from adrenalectomized andovariectomized virgin mice were grown in medium withinsulin, EGF, and fatty acid-free BSA as above. After releaseof the gels, normal explants or epithelium-free explants (33)of mammary fat pad from virgin mice were added to thecultures and the medium was supplemented with insulin,fatty acid-free BSA, and prolactin with or without cortisol.
A450
I- 400-
C), 3500
; 300
A, 250
0n 200
°12 150
C.)100
to 50
0None F F F RU RU
(Contyol) (1) (0.1) (0.01) (0.1) (0.01)
Steroid (ILM)
BM 1 2 3 4 5 6 7 8 9 M
- * v40 - 0 QO
FIG. 1. (A) Mammary epithelial cells from adrenalectomized andovariectomized virgin mice were cultured in collagen gel for 9 dayswith insulin, EGF, and linoleate-BSA. The collagen gels were thenreleased and the medium supplements were changed to insulin,prolactin, linoleate-BSA, and either cortisol (F) or RU 38 486 (RU)at the concentrations listed or were maintained steroid free (control).a-Casein was assayed in the cell extracts. Data are from a typicalexperiment. (B) The cell extracts fromA were precipitated, subjectedto NaDodSO4/PAGE, and blotted to nitrocellulose. After exposureto monoclonal anti-f3-casein and horseradish peroxidase-conjugatedsecond antibody, color was developed using 4-chloro-1-naphthol andH202. The samples were from cultures exposed to the followingcompounds: EGF (end of growth) (lane 1); prolactin (lane 2);prolactin and RU 38 486 (0.1 AM) (lanes 3, 5, and 7); prolactin andRU 38 486 (0.01 AuM) (lane 9); prolactin and cortisol (1 MM) (lane 4);prolactin and cortisol (0.1 MM) (lane 6); prolactin and cortisol (0.01MM) (lane 8). Lanes M, mouse milk standards. Arrowheads indicatethe position of 8-casein.
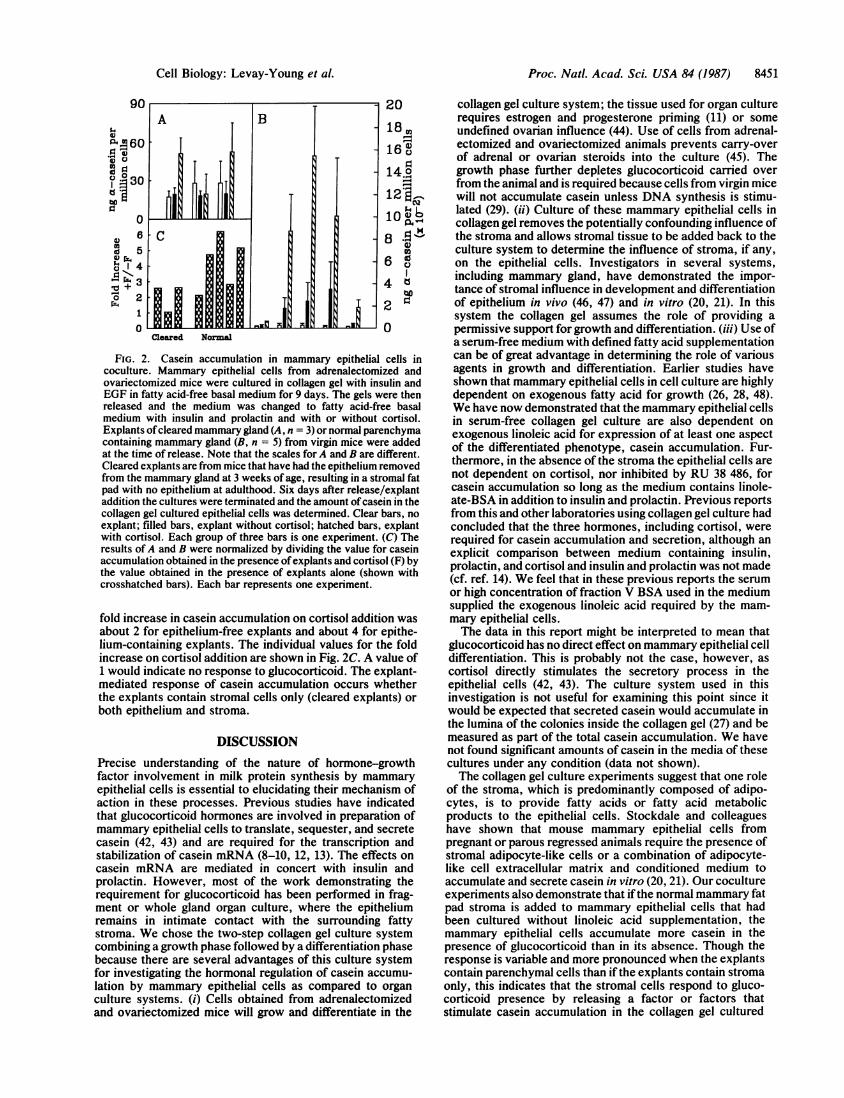
Although addition of explants without cortisol, especiallyepithelium-containing explants, often causes an accumula-tion of a-casein over the explant-free cocultures, Fig. 2A andB demonstrate that the epithelial cells in the collagen gel alsoresponded to the addition of explants and cortisol by accu-mulating larger amounts of casein in the majority of experi-ments. Though it appears that epithelium-containing explantsare remarkably more active than the cleared explants, whennormalized by dividing the casein accumulation in the pres-ence of cortisol by that in the absence of cortisol the average
Proc. Natl. Acad. Sci. USA 84 (1987)
Proc. Natl. Acad. Sci. USA 84 (1987) 8451
B
=1i-I .EN-wiQh NFil hlj
20
18 o16 0
14.o120
U2
04 60 N
6 o
FIG. 2. Casein accumulation in mammary epithelial cells incoculture. Mammary epithelial cells from adrenalectomized andovariectomized mice were cultured in collagen gel with insulin andEGF in fatty acid-free basal medium for 9 days. The gels were thenreleased and the medium was changed to fatty acid-free basalmedium with insulin and prolactin and with or without cortisol.Explants ofcleared mammary gland (A, n = 3) or normal parenchymacontaining mammary gland (B, n = 5) from virgin mice were addedat the time of release. Note that the scales for A and B are different.Cleared explants are from mice that have had the epithelium removedfrom the mammary gland at 3 weeks of age, resulting in a stromal fatpad with no epithelium at adulthood. Six days after release/explantaddition the cultures were terminated and the amount ofcasein in thecollagen gel cultured epithelial cells was determined. Clear bars, noexplant; filled bars, explant without cortisol; hatched bars, explantwith cortisol. Each group of three bars is one experiment. (C) Theresults of A and B were normalized by dividing the value for caseinaccumulation obtained in the presence ofexplants and cortisol (F) bythe value obtained in the presence of explants alone (shown withcrosshatched bars). Each bar represents one experiment.
fold increase in casein accumulation on cortisol addition wasabout 2 for epithelium-free explants and about 4 for epithe-lium-containing explants. The individual values for the foldincrease on cortisol addition are shown in Fig. 2C. A value of1 would indicate no response to glucocorticoid. The explant-mediated response of casein accumulation occurs whetherthe explants contain stromal cells only (cleared explants) orboth epithelium and stroma.
DISCUSSIONPrecise understanding of the nature of hormone-growthfactor involvement in milk protein synthesis by mammaryepithelial cells is essential to elucidating their mechanism ofaction in these processes. Previous studies have indicatedthat glucocorticoid hormones are involved in preparation ofmammary epithelial cells to translate, sequester, and secretecasein (42, 43) and are required for the transcription andstabilization of casein mRNA (8-10, 12, 13). The effects oncasein mRNA are mediated in concert with insulin andprolactin. However, most of the work demonstrating therequirement for glucocorticoid has been performed in frag-ment or whole gland organ culture, where the epitheliumremains in intimate contact with the surrounding fattystroma. We chose the two-step collagen gel culture systemcombining a growth phase followed by a differentiation phasebecause there are several advantages of this culture systemfor investigating the hormonal regulation of casein accumu-lation by mammary epithelial cells as compared to organculture systems. (i) Cells obtained from adrenalectomizedand ovariectomized mice will grow and differentiate in the
collagen gel culture system; the tissue used for organ culturerequires estrogen and progesterone priming (11) or someundefined ovarian influence (44). Use of cells from adrenal-ectomized and ovariectomized animals prevents carry-overof adrenal or ovarian steroids into the culture (45). Thegrowth phase further depletes glucocorticoid carried overfrom the animal and is required because cells from virgin micewill not accumulate casein unless DNA synthesis is stimu-lated (29). (ii) Culture of these mammary epithelial cells incollagen gel removes the potentially confounding influence ofthe stroma and allows stromal tissue to be added back to theculture system to determine the influence of stroma, if any,on the epithelial cells. Investigators in several systems,including mammary gland, have demonstrated the impor-tance of stromal influence in development and differentiationof epithelium in vivo (46, 47) and in vitro (20, 21). In thissystem the collagen gel assumes the role of providing apermissive support for growth and differentiation. (iii) Use ofa serum-free medium with defined fatty acid supplementationcan be of great advantage in determining the role of variousagents in growth and differentiation. Earlier studies haveshown that mammary epithelial cells in cell culture are highlydependent on exogenous fatty acid for growth (26, 28, 48).We have now demonstrated that the mammary epithelial cellsin serum-free collagen gel culture are also dependent onexogenous linoleic acid for expression of at least one aspectof the differentiated phenotype, casein accumulation. Fur-thermore, in the absence of the stroma the epithelial cells arenot dependent on cortisol, nor inhibited by RU 38 486, forcasein accumulation so long as the medium contains linole-ate-BSA in addition to insulin and prolactin. Previous reportsfrom this and other laboratories using collagen gel culture hadconcluded that the three hormones, including cortisol, wererequired for casein accumulation and secretion, although anexplicit comparison between medium containing insulin,prolactin, and cortisol and insulin and prolactin was not made(cf. ref. 14). We feel that in these previous reports the serumor high concentration of fraction V BSA used in the mediumsupplied the exogenous linoleic acid required by the mam-mary epithelial cells.The data in this report might be interpreted to mean that
glucocorticoid has no direct effect on mammary epithelial celldifferentiation. This is probably not the case, however, ascortisol directly stimulates the secretory process in theepithelial cells (42, 43). The culture system used in thisinvestigation is not useful for examining this point since itwould be expected that secreted casein would accumulate inthe lumina of the colonies inside the collagen gel (27) and bemeasured as part of the total casein accumulation. We havenot found significant amounts of casein in the media of thesecultures under any condition (data not shown).The collagen gel culture experiments suggest that one role
of the stroma, which is predominantly composed of adipo-cytes, is to provide fatty acids or fatty acid metabolicproducts to the epithelial cells. Stockdale and colleagueshave shown that mouse mammary epithelial cells frompregnant or parous regressed animals require the presence ofstromal adipocyte-like cells or a combination of adipocyte-like cell extracellular matrix and conditioned medium toaccumulate and secrete casein in vitro (20, 21). Our cocultureexperiments also demonstrate that if the normal mammary fatpad stroma is added to mammary epithelial cells that hadbeen cultured without linoleic acid supplementation, themammary epithelial cells accumulate more casein in thepresence of glucocorticoid than in its absence. Though theresponse is variable and more pronounced when the explantscontain parenchymal cells than ifthe explants contain stromaonly, this indicates that the stromal cells respond to gluco-corticoid presence by releasing a factor or factors thatstimulate casein accumulation in the collagen gel cultured
90
51.4VaP' 60
o 0- °30
0
c 5
.40 2
1
0
Cleared Normal-wN__AXitLi
Cell Biology: Levay-Young et al.
l1

8452 Cell Biology: Levay-Young et al.
epithelial cells. Is this potential mediator of glucocortcoidaction linoleic acid? We feel that this is a possibility forseveral reasons. Linoleic acid is directly effective on theisolated mammary epithelial cells as shown above. Recentwork by Fielder and Talamantes has shown that the syntheticglucocorticoid dexamethasone combined with mouse prolac-tin induces lipolysis in mouse ovarian fat pad tissue (49). Asimilar effect is seen in prolactin-treated rat mammaryexplants (50). Since linoleic acid is a significant component ofdepot fat in mice (51), we surmise that these stroma-derivedfactor(s) induced by glucocorticoid in this system may belipid in nature and even perhaps linoleic acid or one of itsmetabolic products.The large difference in casein accumulation between
cocultures with cleared and epithelium-containing explants isdifficult to explain. It is possible that there is an interactionbetween the epithelial and stromal cells that causes a largerrelease of linoleic acid or increased production of the pro-posed active metabolic products of linoleic acid. Sincecleared explants have a reduced vascularity compared toepithelium-containing explants (52), alternatively it is possi-ble that one or more of the vascular cell types may beinvolved in this process. Another explanation could bemodeled from the work of Kidwell and Shaffer (50) who haveproposed that the mammary epithelial cells on exposure toprolactin release a factor that induces the mammary glandmast cells to stimulate lipolysis in the mammary adipocytes.Since the epithelial cells are absent from the cleared explantsthey would not be present to induce the mast cells. Alterna-tively it is possible that cleared explants would have a muchsmaller population of these potentially important mast cells.At this time we are not sure whether one or more of thesealternatives are responsible for the reduced effectiveness ofthe cleared explants.
This report is dedicated to the memory of Roy E. Young. Wegratefully acknowledge the assistance of F. Stockdale, who suppliedthe monoclonal antibody, and R. Deraedt and Roussel UCLAF, whodonated the RU 38 486. We thank W. Imagawa for extensivediscussions on fatty acids, John Underhill for assistance withphotography, and W. Imagawa, J. Richards, and R. Guzman forreview of the manuscript. This investigation was supported by PublicHealth Service Grants CA09041, CA05388, and CA40160 awarded bythe National Cancer Institute, Department of Health and HumanServices.
1. Topper, Y. J. & Freeman, C. S. (1980) Physiol. Rev. 60,1049-1106.
2. Elias, J. J. (1980) in Hormonal Proteins and Peptides, ed. Li,C. H. (Academic, New York), Vol. 8, pp. 37-74.
3. Elias, J. J. (1957) Science 126, 842-844.4. Oka, T. & Perry, J. W. (1974) J. Biol. Chem. 249, 3586-3591.5. Bolander, F. F., Jr., Nicholas, K. R., Van Wyk, J. J. &
Topper, Y. J. (1981) Proc. Natl. Acad. Sci. USA 78,5682-5684.
6. Chomczynski, P., Qasba, P. & Topper, Y. J. (1984) Science226, 1326-1328.
7. Juergens, W. G., Stockdale, F. E., Topper, Y. J. & Elias, J. J.(1965) Proc. Natl. Acad. Sci. USA 54, 629-634.
8. Nagaiah, K., Bolander, F. F., Jr., Nicholas, K. R., Takemoto,T. & Topper, Y. J. (1981) Biochem. Biophys. Res. Commun.98, 380-387.
9. Kulski, J. K., Topper, Y. J., Chomczynski, P. & Qasba, P.(1983) Biochem. Biophys. Res. Commun. 114, 380-387.
10. Chomczynski, P., Qasba, P. & Topper, Y. J. (1986) Biochem.Biophys. Res. Commun. 134, 812-818.
11. Ichinose, R. R. & Nandi, S. (1964) Science 145, 497-498.12. Terry, P. M., Baneree, M. R. & Lui, R. M. (1977) Proc. Natl.
Acad. Sci. USA 74, 2241-2245.
13. Mehta, N. M., Ganguly, N., Ganguly, R. & Banerjee, M. R.(1980) J. Biol. Chem. 255, 4430-4434.
14. Emerman, J. T., Enami, J., Pitelka, D. R. & Nandi, S. (1977)Proc. Natl. Acad. Sci. USA 74, 4466-4470.
15. Lee, E. Y.-H., Parry, G. & Bissell, M. J. (1984) J. Cell Biol.98, 146-155.
16. Taketani, Y. & Oka, T. (1986) Horm. Metab. Res. 18, 119-125.17. Flynn, D., Yang, J. & Nandi, S. (1982) Differentiation 22,
191-194.18. Tonelli, Q. J. & Sorof, S. (1982) Differentiation 22, 195-200.19. Durban, E. M., Medina, D. & Butel, J. S. (1985) Dev. Biol.
109, 288-298.20. Levine, J. F. & Stockdale, F. E. (1985) J. Cell Biol. 100,
1415-1422.21. Weins, D., Park, C. S. & Stockdale, F. E. (1987) Dev. Biol.
120, 245-258.22. Wicha, M. S., Lowrie, G., Kohn, E., Bagavandoss, P. &
Mann, T. (1982) Proc. Natl. Acad. Sci. USA 79, 3213-3217.23. Ray, D. B., Jansen, R. W., Horst, I. A., Mills, N. C. &
Kowal, J. (1986) Endocrinology 118, 393-407.24. Yang, J., Richards, J., Guzman, R., Imagawa, W. & Nandi, S.
(1980) Proc. Natl. Acad. Sci. USA 77, 2088-2092.25. Yang, J., Guzman, R., Richards, J., Imagawa, W., Mc-
Cormick, K. & Nandi, S. (1980) Endocrinology 107, 35-41.26. Imagawa, W., Tomooka, Y. & Nandi, S. (1982) Proc. Natl.
Acad. Sci. USA 79, 4074-4077.27. Imagawa, W., Tomooka, Y., Hamamoto, S. & Nandi, S.
(1985) Endocrinology 116, 1514-1524.28. Bandyopadhyay, G., Imagawa, W., Wallace, D. R. & Nandi,
S. (1987) J. Biol. Chem. 262, 2750-2756.29. Stockdale, F. E. & Topper, Y. J. (1966) Proc. Natl. Acad. Sci.
USA 56, 1283-1289.30. Richards, J., Larson, L., Yang, J., Guzman, R., Tomooka, Y.,
Osborn, R., Imagawa, W. & Nandi, S. (1983) J. Tissue CultureMethods 8, 31-36.
31. Emerman, J. T. & Pitelka, D. R. (1977) In Vitro 13, 316-328.32. Edery, M., Imagawa, W., Larson, L. & Nandi, S. (1985)
Endocrinology 116, 105-112.33. DeOme, K. B., Faulkin, L. J., Jr., Bern, H. A. & Blair, P. B.
(1959) Cancer Res. 19, 515-520.34. Hinegardner, R. T. (1971) Anal. Biochem. 39, 197-201.35. McGrath, M., Palmer, S. & Nandi, S. (1985) J. Cell. Physiol.
125, 182-191.36. Engvall, E. & Carlsson, H. E. (1976) in First International
Symposium on Immunoenzymatic Techniques, INSERM Sym-posium No. 2, eds., Feldman, G., Druet, P., Bignon, J. &Avrameas, S. (North-Holland, Amsterdam), pp. 135-147.
37. Enami, J. & Nandi, S. (1977) J. Immunol. Methods 18,235-244.
38. Towbin, H., Staehelin, T. & Gordon, J. (1979) Proc. Natl.Acad. Sci. USA 76, 4350-4354.
39. Laemmli, U. K. (1970) Nature (London) 227, 680-685.40. Gagne, D., Pons, M. & Philibert, D. (1985) J. Steroid Biochem.
23, 247-251.41. Rocha, V., Ringo, D. L. & Read, D. B. (1985) Exp. Cell Res.
159, 201-210.42. Mills, E. S. & Topper, Y. J. (1970) J. Cell Biol. 44, 310-328.43. Oka, T. & Topper, Y. J. (1971) J. Biol. Chem. 246, 7701-7707.44. Bolander, F. F., Jr., & Topper, Y. J. (1980) Endocrinology
107, 1281-1285.45. Bolander, F. F., Jr., Nicholas, K. R. & Topper, Y. J. (1979)
Biochem. Biophys. Res. Commun. 91, 247-252.46. Kratochwil, K. (1969) Dev. Biol. 20, 46-71.47. Sakakura, T., Nishizuka, Y. & Dawe, C. J. (1976) Science 194,
1439-1441.48. Wicha, M. S., Liotta, L. A. & Kidwell, W. F. (1979) Cancer
Res. 39, 426-435.49. Fielder, P. J. & Talamantes, F. (1987) Endocrinology 121,
493-497.50. Kidwell, W. R. & Shaffer, J. (1984) J. Am. Oil Chem. Soc. 61,
1900-1904.51. Smith, S., Watts, R. & Dils, R. (1968) J. Lipid Res. 9, 52-57.52. Soemarwoto, I. N. & Bern, H. A. (1958) Am. J. Anat. 103,
403-436.
Proc. Natl. Acad. Sci. USA 84 (1987)