linkages between flow regime, biota, and ecosystem ...biota, and ecosystem processes: implications...
TRANSCRIPT

REVIEW SUMMARY
HYDROLOGY
Linkages between flow regimebiota and ecosystem processesImplications for river restorationMargaret Palmerdagger and Albert Ruhi
BACKGROUND Early civilizations developedaround seasonal river floodplains and the nat-ural rhythm of rivers remains critical to hu-mans today We use streams and rivers to meetdrinking water irrigation and hydropowerneeds by storing and moving water in complexways at the times and places of our choosingConsequently many of Earthrsquos rivers have flowregimes that are ldquounnaturalrdquo in magnitude fre-quency duration and timing The rise in riverdegradation globally has motivated research onthe link between hydrologic alteration and de-clines in valued biota At the same time largelyfueled by new technologies and methods re-search has expanded to understand the pat-terns in and drivers of riverine processes likeprimary production in both near-pristine anddegraded rivers A third line of research sty-mied by how difficult it has been to restoredegraded rivers has called for process-basedrestoration building on knowledge from theother two research thrusts Todayrsquos hydro-
ecological science seeks to understand themechanisms whereby flow regimes affect biotaand ecosystem processes and the interplay be-tween them in a three-way interaction we callthe flow-biota-ecosystem processes nexus
ADVANCESBy shifting the focus from staticpatterns at sites to dynamic processes alongriver networks advances are being made tounderstand the interactions and feedbacks atthe nexus Fueled by increasingly available time-series data and novel modeling emerging re-search ranges from studies on regime-basedproperties such as flow periodicity and itschange to studies on river network structureand associated spatial variation in flow andwater chemistry These studies demonstratehow flow variability influences long-term per-sistence of riverine assemblages and they aredisentangling the direct effects of flow on com-munities and ecosystem processes from itsindirect effects (eg via species interactions
light-blocking turbidity) Changes in temporalpatterns in flow magnitudes can increase risk ofcommunity collapse and alter key ecosystemprocesses such as primary production Growingresearch shows that storm flowsnot only enhanceinputs and downstream export of terrestriallyderived carbon to rivers but when associatedwith sustainedhydrologic connectivitywith soilsexert particular influence onwater chemistry and
biogeochemical processesthat can influence foodwebs Increasedavailabilityof environmental sensorshas stimulated researchshowing that extreme flowsmay impart disproportion-
ate impacts on stream metabolism but the re-lationship can depend on the predictability ofthose flows Research combining changes inflow patterns with stable isotope analyses isrevealing how temporal fluctuations in habitatand in thequality andquantity of basal resourcesinfluence trophic pathways and resulting food-web structure Evidence suggests that restoringparticular facets of a flow regime can producedesirable conservation outcomes but context isparamountRestoration actions goingbeyonddis-crete flow events and enhancing groundwater-influenced river habitat or redirecting subsurfaceflow paths may be critical in future climates
OUTLOOK Our understanding of the flow-biota-ecosystem processes nexus is still incom-plete and is a frontier research topic Challengesinclude connecting organismal and ecosystem-level processes and understanding the role ofmicrobial communities as intermediaries Cap-turing the effects of watershed-level physicaland biogeochemical heterogeneity and pars-ing out direct indirect or cascading effects offlow alteration on biota and processes wouldalso reduce uncertainty in restoration outcomesparticularly in novel nonstationary environ-ments Understanding how much flow resto-ration alone can achieve in urban watershedsis an urgent need as is translating findingsfrom hydroecology to design green infrastruc-ture and flow release programs from reservoirsThese management tools may offer growingopportunities to experiment with flow regimeswhich will assist in refining process-based riverrestorationBoth solid scienceandeffective trans-lation into practicewill be needed to curb the fastpace of global river ecosystem degradation
RESEARCH
Palmer et al Science 365 1264 (2019) 20 September 2019 1 of 1
The list of author affiliations is available in the full article onlineThese authors contributed equally to this workdaggerCorresponding author Email mpalmerumdeduCite this article as M Palmer and A Ruhi Science 365eaaw2087 (2019) DOI 101126scienceaaw2087
River flow regimes have shaped the life history strategies of plants and animals overevolutionary time scales River regulation and associated alteration of flow and thermalregimes alter organismal development often shifting important events such as insectemergence depicted here by Palingenia mayflies entering their winged flying stage to mate
TOMORROWrsquoS EARTHRead more articles onlineat scimagTomorrowsEarth
PHOTOCHRISS_N
SISTOCKPHOTOCOM
ON OUR WEBSITE
Read the full articleat httpdxdoiorg101126scienceaaw2087
on May 27 2020
httpsciencesciencem
agorgD
ownloaded from
REVIEW
HYDROLOGY
Linkages between flow regimebiota and ecosystem processesImplications for river restorationMargaret Palmer1dagger and Albert Ruhi2
River ecosystems are highly biodiverse influence global biogeochemical cycles andprovide valued services However humans are increasingly degrading fluvial ecosystemsby altering their streamflows Effective river restoration requires advancing ourmechanistic understanding of how flow regimes affect biota and ecosystem processesHere we review emerging advances in hydroecology relevant to this goal Spatiotemporalvariation in flow exerts direct and indirect control on the composition structure anddynamics of communities at local to regional scales Streamflows also influenceecosystem processes such as nutrient uptake and transformation organic matterprocessing and ecosystem metabolism We are deepening our understanding of howbiological processes not just static patterns affect and are affected by stream ecosystemprocesses However research on this nexus of flow-biota-ecosystem processes is atan early stage We illustrate this frontier with evidence from highly altered regulated riversand urban streams We also identify research challenges that should be prioritized toadvance process-based river restoration
Rivers have been critical to human existencesince antiquity and are a central part ofthe biosphere Flowing waters sustain riv-erine terrestrial and marine biodiversityand make important contributions to global
biogeochemical cycles However river ecosystemsare increasingly degraded by dam building di-version or abstraction of water clearing of landand climate change Some rivers that once werehealthy and diverse now only support drought-or pollution-tolerant species others shunt eu-trophic water toward coastal regions or offer newhabitat to non-native species These degradedrivers all have one characteristic in commonsome or all aspects of their flows have been al-tered (Fig 1)Flow regime or the characteristic pattern of
flow variation has long been known to be a keydriver of a riverrsquos structure and functioning (1)The characteristic magnitude frequency dura-tion timing and rate of change in river flows haveshaped awide range of species adaptationsmdashfromlife history strategies to behaviors andmorphol-ogies of both aquatic and riparian organisms (2)A riverrsquos flow regime also influences in-stream andflood-plain ecosystemprocesses including primaryproduction and nutrient cycling Because flowdynamism is central to a riverrsquos functioning andits ability to provide ecosystem services flow alter-ation is rarely inconsequential
When flows are altered a combination ofbiotic and abiotic pathways are triggered Forinstance flash floods that reduce predators canhave cascading biotic effects on primary producersand on associated nutrient dynamics Flows thatincrease suspended sediments may inhibit orga-nismal feeding and reproduction or reduceprimaryproduction These are both examples of indirectabiotic effects Although altered flows are oftennot the proximate mechanism of ecological de-gradation they can exacerbate the impacts of otherabiotic stressors (3) For example extreme lowflows resulting from excessive withdrawals canincrease water temperature and pollutant con-centrations to the point that they exceed tolerablelevels for organismal survival or reproductionThe mechanisms linking flow regime alterationto ecological degradation can be numerous andcomplexGiven themagnitude and global extent of river
degradation it is fitting that the United NationsDecade on Ecosystem Restoration has just beendeclared (4) Science to heal streams and rivershas never been more needed Most restorationpractices have focused on improving channelmorphology or habitat and unfortunately re-covery of biodiversity or species of interest hasproven difficult (5 6) This has prompted increasedcalls by scientists to move frommorphological toecosystem-level ldquoprocess-basedrdquo practices (7) thatfocus on restoring flow regimes as well as otherphysical and ecological processes that sometimescovary with flow and support aquatic commu-nities In this Review we summarize the currentunderstanding of the relationship between riverflow regimes biota and ecosystem processes and
outline how future river restoration can benefitfrom a better understanding of the three-wayinteraction that we call the flow-biota-ecosystemprocesses nexus (Fig 2) We address the follow-ing How does flow variation control river biotadirectly and indirectly How does it control riverecosystem processes When flow regimes are al-tered are changes in ecosystem processes coupledwith changes in biota Can these bodies of re-search inform restoration practice and wherecould advances in hydroecology be better lever-agedWe close by identifying key challenges andopportunities in hydroecological researchThis Review focuses primarily on fundamental
research advances however regulated rivers (iethose with dams) and urban streams help groundfindings in real-world problems These two eco-system types have experienced some of the largestshifts in their flow regimes and are central tonumerous restoration efforts Building off of ad-vances needed to restore these degraded eco-systems we highlight research that can informprocess-based restoration more generally Weshow that despite calls for improving the linkbetween science and restoration in practice thereremains a great need to advance research on theflow-biota-ecosystem processes nexus This needopens exciting opportunities for both fundamen-tal and applied interdisciplinary research
How does flow variation controlriver biota
Flow is essential to river life but can also be asource of stress scouring floods remove organismsdroughts stress organismal physiology and flowconditions dictate the changing type quantity andquality of the physical habitat inwhich organismslive (8) However flow variation also influencesorganisms indirectly by keeping predators com-petitors and invaders at bay (9 10) by control-ling the energy sources that enter the food web(11) or by affecting movement of organisms andmatter across river networks and floodplains(12 13) At the inception of the natural flow re-gime concept (1) the study of flow-ecology rela-tionships focused on static representations of flowand local-scale research Over the past 20 yearsimportant progress has beenmade toward under-standing how ecological communities respond todynamic flow regimes across entire river networks
Organisms respond to patterns offlow variation
Stream ecologists have long known that low-and high-flow events can temporarily reduceabundance and diversity of invertebrates andfishes particularly when droughts fragmentthe riverine habitat or when spates mobilize thestreambed However it is not only discrete eventsthat are important long-term patterns of flowvariability have historically selected for organis-mal life histories related to growth reproductiondispersal and the ability to persist under physicaland chemical stress The increasing availabilityof temporally extensive andor high-frequencydatasets is now spurring the use of spectral meth-ods which allow the identification of dominant
RESEARCH
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 1 of 13
1National Socio-Environmental Synthesis Center University ofMaryland Annapolis MD 21401 USA 2Department ofEnvironmental Science Policy and Management Universityof California Berkeley Berkeley CA 94720 USAThese authors contributed equally to this workdaggerCorresponding author Email mpalmerumdedu
on May 27 2020
httpsciencesciencem
agorgD
ownloaded from
frequencies amplitudes and phases in the envi-ronment (14) These characteristic patterns ofvariation or regimes can then be connected toorganismal dynamics For example a waveletanalysis of discharge and abundance of silvercarp larvae found a strong association betweenspawning and peak flows over a particular mag-nitude (15) Using a similar approach temporalchange in stream invertebrate diversity was foundto be influenced not only by the seasonality ofrainfall but by the reliability of this variabilityie its predictability (16)Flow regimes differ across river types and
climates (1) but can also shift temporally at agiven place as a consequence of changes inclimate land use or flow management (Fig 3)Although changes in dominant frequencies inthe environment can have strong impacts on com-munities studying the effects of time-varying flowregimes is still rare (17) Faster flow cycles mayfilter out species with longer generation timesand a preference for stable environments (18) andincreased frequencies of extreme events (eg re-current unpredictable droughts) may drive com-munities to novel stable states (19) However westill do not know which temporal scales of flowvariationmay entrain organismal phenology andwhich may simply represent a source of stressAlthoughmost work to understand this questionhas used correlational approaches mathemat-ical models based on metabolic theory and bio-mechanical constraints may help in predicting
the effects of changing frequencies of environ-mental fluctuations and extremes on river orga-nisms food webs and ecosystem processes (20)
Spatiotemporal variation in flow controlsbiotic persistence
River network structure and directional stream-flow influence the movement of drifting orga-nisms and materials downstream (21) and themain ways in which communities disperse andassemble Experiments suggest that the local envi-ronment exerts control of community compo-sition in the more isolated headwaters whereasdispersal and environmental influences togetheroperate in the well-connected main stem or pri-mary downstream channel This is known as thenetwork position hypothesis (22) Although findingshave sometimes been inconsistent with respectto this idea a recent study using graph theory andreplicate fishmetacommunities (ie multiple com-munities linked by dispersal) suggests that supportfor this hypothesis may depend on network struc-ture and the degree of environmental heteroge-neity in headwaters relative to themainstem (23)Incorporating network structure into the studyof flow regimes is also important because thedendritic connectivity of streams and rivers con-strains organismal movement and associatedpersistence (24) This is particularly relevant toclimate change because shifts in both wet-channel network structure and habitat qualityacross the network are anticipated to occur In
this vein recent research has demonstrated thatin arid climates downstream sections of riversmay bemore suitable for fish spawning whereasin wetter conditions the most suitable habitatoccurs in the headwaters (25)The spatial coordination of flow regimes across
the river network also has key implications forbiodiversity persistence In sets of populationslinked by dispersal or metapopulations a com-bination of diverse physical and biological fea-tures helpsmaintain asynchronous dynamics thusmaximizing stability at larger scales (ie riskspreading or portfolio effect) Only recently havethe simultaneous effects of spatial and temporalvariation in flows been incorporated into streamecology For example widespread flood eventsmay cause synchronous mortality of early lifestages of salmonids [eg (26)] thus weakeningthe portfolio effect On the flip side river branchescan provide diversity in flow and habitat con-ditions ensuring that populations aremaintainedacross the network Using an agent-based modelvalidatedwith long-term fish time series researchhas shown that population asynchrony tends tobe higher across branches than within branchesthus branching network complexity is key to pre-serving metapopulation stability (27) Given thatdams and climate change are making flow re-gimes more similar synchrony in flow-dependentecological processes could be increasing aswell The phenomenon of environmentally forcedsynchrony has long been studied in terrestrial
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 2 of 13
Fig 1 Impacts of river fragmentation by dams urban areas andwater risk across the worldrsquos watersheds Degree of fragmentation(DOF) represents the effects of dams on longitudinal habitatconnectivity and urban impacts (URB) capture the degree of infra-structure development in riparian and floodplain areas (measured asnightlight intensity in urban areas) (153) We averaged river-reach leveldata at the watershed scale using streams with long-term averagedischarge higher than 1 m3s and we then distributed averaged valuesamong the following categories lt5 (low impact) 5 to 20 (mediumimpact) and gt20 (high impact) Colors represent different impact
combinations at the watershed level and flow lines represent themajor rivers of the world Polygons outlined in gray are areas withldquohighrdquo and ldquoextremely highrdquo exposure to water-related risk anaggregated measure of water quantity quality and regulatory riskproposed by the World Resources Institute (154) Around 23 of theworldrsquos watersheds are subject to water-related risks and 19 ofthe watersheds with flow show substantial impacts (ie in gt5 of theriver network) related to longitudinal habitat fragmentation by damsor urban areas These dimensions capture the major sources offlow-related river ecosystem degradation
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
population ecology [Moran theorem (28)] andcould represent an underappreciated emerg-ing risk in fresh waters
Abiotic and biotic influences of flow
Stream ecologists traditionally interpreted thedynamics of communities as a deterministic resultof biotic interactions or as a stochastic conse-quence of the environment (29) However researchhas shown that these fluctuate in relative impor-tance over space and time andmay even influenceeach other For instance biotic interactions mayoccur only under particular environmental con-ditions (30) Inferences gathered from long-termdata combined with time-series methods importedfrom econometrics can help elucidate when and
where abiotic influences have primacy For exam-ple multivariate autoregressive state-space modelsapplied to long-term flow and fish communitydata showed that interannual variation in flowanomalies were the ultimate driver of the dy-namics of a desert stream fish community (31)Although non-native and native fishes interactedsuch effects were small compared with the directeffects of flows on the native component Bioticinteractions that emerge from known species-specific relationships between components of theflow regime such as flood timing and popula-tion dynamics can also be predicted from modelsimulations of community trajectory (30) A recentapplication of these interaction-neutral modelsfound that flows departing from the natural reg-
ime simplified interaction networks among plantguilds in desert rivers (32)A fundamental challenge for community eco-
logy for decades has been quantifying the rela-tive influence of abiotic (flow-driven) and biotic(flow-mediated) effects This is important for theaccurate design and prediction of restoration out-comes including for example anticipatingwhenflow alteration facilitates species invasions bycreating new niches (33) by enhancing propagulepressure or eroding native enemies (34) or by acombination of thesemechanismsDistinguishingthese could help managers compare the benefitsof eradicating non-natives relative to restoringecologically important facets of the flow regime
Flow alteration filters species and traits
Trait-based approaches are becoming increasinglypopular in hydroecology because they enhancecomparison of flow-ecology relationships acrossclimates (35) they lead to predictions about howcommunity change may affect ecosystem func-tioning (36) and they identify the mechanismsthat filter or promote particular taxa under stress-ful conditions such as drought (37) In the case offlow intermittency traits can be used to identifycommunity tipping points by connecting local en-vironmental conditions to organisms with traitsthat allow them to persist in isolated pools or wetsediment or connecting network-level water con-ditions to the ability of highly resilient organismsto drift or actively recolonize from perennial re-fugia (38) Traits can also be used to assess re-storation trajectoriesmdashfor instance by assessingfluctuations in the share of strategies that representdifferent life-history trade-offs [eg species limitedby reproductive capacity or by resources (39)]By studying a communityrsquos taxonomic char-
acteristics (eg species composition) and func-tional characteristics (eg dispersalmodes feedingstrategies) ecologists can determine to whatextent trait redundancy across speciesmay ensureagainst loss of functional diversity For examplein a study on Alpine stream communities subjectto climate change and flow regulation functionaldiversity increased while functional redundancydecreased (40) This finding indicates that ongoinghydroclimatic change can reduce the ability of acommunity to withstand further alteration evenif trait diversity is high because the persistenceof individual traits may be linked to the fate of afew sensitive species In an experimental studycommunities affected by drought presented com-parable taxonomic and functional decays implyingthat these communities were both taxonomicallyand functionally vulnerable (41) The risk of com-munity collapse in response to flow alterationalso depends on the way in which ldquowinningrdquo andldquolosingrdquo traits co-occur in a given species andacross species in a community as a result of aparticular stress Thus community-wide responsesto a particular alterationmay be stronger if speciesaremaladaptive to that change for instance bybeing large-bodied longer-lived and collector-feeding (filtering particulate organic matter) ina scenario of increased flow variability (18) Under-standing how flow alteration impairs species with
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 3 of 13
Nutrient cycling
Primary production
Energy inputs
Secondary production Dispersal refugia Invasions
Food-web interactionsMetabolic regime
Hydrologic connectivity
Streambed sediments
Water chemistry temperature
Channel form riparian vegetation
Decomposition Species demographic rates
River ecosystem integrity
Altered flow regimes
Restored flow regimes
Time
Restoration toolboxAlter pattern of reservoir releasesImprove stormwater controlReforest and restore watersheds
Disc
harg
e w
ater
leve
l Muted high flows
Storm flows
Altered periodicity
Flow homogenization
Shifting baselines
Extreme low flows
Fig 2 Process-based restoration posits that actions must target processes not patternsthat have been degraded River ecosystems are sustained by a host of environmental conditionsor ecosystem structure (gold rectangles) which in turn influence ecosystem processes (greenrectangles) and biotic processes (blue rectangles) These processes as well as ecosystem structureare all controlled by the flow regime Thus correcting facets of flow regime alteration may enhanceriver ecosystem integrity in both direct and indirect ways
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
traits that have effects on ecosystems (eg byparticular feeding strategies) will help anticipatethe risks of losing major ecosystem functionssuch as detrital decomposition (36)
How does flow variation control riverecosystem processes
Rivers produce transform and store organicmatter (OM) and nutrients Carbon (C) and nu-trient concentrations and fluxes vary longitudi-nally and laterally as river networks expand andcontract with rain These variations influenceand are influenced by stream ecosystem metab-olism which can be described as net ecosystemproduction or the difference between gross pri-mary production (GPP) and ecosystem respiration(ER) The metabolic balance between heterotro-phic and autotrophic production that providesresources for consumers can shift seasonally orwith changing flows as can hydrologic connect-ivity that influences inputs of OM and solutes torivers (42) Increasing availability of sensors andadvances inmodeling (43) havemade the study ofmetabolism OM processes and solute dynamicsripe for linking to biotic processes
Flow variability andecosystem metabolism
Spatiotemporal variability in ecosystem processesis a fundamental characteristic of rivers and flowregime is a major driver of this variability (Fig 4)Recentwork shows thatmany rivers in theUnitedStates have strong seasonal patterns in primary
production with spring peaks for larger riversand summer peaks for smaller rivers generallyGPP is negatively correlated with high variationin daily discharge (44) In regions with little sea-sonality andor continual canopy cover GPP islower and its patterns are less predictable (44)Bernhardt et al (45) provide a comprehensivereview of stream metabolism the factors thatinfluence it and why it is difficult to predict Flowhas the largest effect at the extremes such that atlow flowswhen the streambed begins to dry largedrops in GPP occur (46) Storms also often reduceGPP (45) However patterns may vary as a func-tion of flow predictability For example researchin a river characterized by predictable peaks dur-ing spring snowmelt found over two seasons thatGPP rose during spring snowmelt sampling (47)whereas high flows during the winter and sum-mer reduced GPP Within a year this patternresulted in a switch from net autotrophic to netheterotrophic productionThe paths that water takes as it moves to and
within a streammdasha spatial aspect of its flowregimemdashinfluencesmetabolismBycouplingmetab-olism and chemistry time series with informationonhydrologic connectivity and flowpaths ERwasshown to increase in an organic-rich watershedduring peak flows and remained elevated forweeks (48) Sustained hydrologic connectivitywith soils likely fueled inputs of dissolved orga-nic carbon (DOC) that promoted bacterial respi-ration Such inputs can lead to nonlinear responsesinmetabolism For example increasing flows have
led to decreases in GPP up to some thresholdflow beyond which GPP was suppressed by re-duced water clarity from flow-elevated humicDOC (49) In the same vein higher riverine GPPhas been associated with reduced suspendedsediments and high temperatures during low-flow droughts (50) When flows cease entirely andsections of riverbeds dry up carbon dioxide fluxesto the atmosphere can be far higher than those inflowing sections of the channel (51) presumablybecause of spikes in aerobicmicrobial respirationhowever as is the case with methane in riverspartitioning fluxes of these gases to changes inmetabolism (aerobic or anaerobic) is difficult par-ticularly under variable flows (52)
Hydrologic connectivity drivessolute dynamics
Research is growing on how hydrologic connect-ivity to hillslope soils influences water chemistry(53) and how variation in discharge and riverstage influence the strength of bidirectional waterexchanges that affect nitrogen (N) uptake reten-tion and removal (54) Periods of hydrologic con-nectivity may be followed by long periods withlimited exchange which may differ greatly alongthe stream corridor This can lead to complex CN and phosphorus (P) dynamics that influenceor are influenced by microbial and algal produc-tivity When connectivity with watershed solutesources is extensive inputs of solutes such asDOCmay be so high that they exceed the ability of riverecosystem processes such as microbial uptake toregulate their fluxes (55) Theory posits that largestorms pulse terrestrially derived dissolved or-ganicmatter (DOM) into streams and it is quicklyshunted downstream by high flows where it isprocessed in larger parts of the network (56)However how far DOM in pulsedwater is trans-ported is governed not only by discharge but bythe composition of DOMUnderstanding the rela-tionship between DOM sources and its composi-tion is being advanced by combining time-seriesdata with isotopic information to evaluate spatialvariability in hydrologic flow paths transit timeand chemistry as watermoves from the terrestrialto aquatic realms (53 57) It is difficult tomeasurebiotic uptake in streams during storms howeverrecent work suggests that assuming storm flowstransport DOM too quickly for in-stream process-ing may be a poor assumption if the DOM com-position renders it readily utilizable by biota (58)Although hydrologic connectivity is not a newconcept it is not typically considered part of theflow regime yet it can be characterized by itstiming duration and frequency and then linkedto stream processes or patterns Newmethods forquantifying connectivity [eg (59)] are emergingand the energetic and water quality importanceof network connectivity to soils and diverse land-scape elements (such as wetlands and ponds) isbecoming a major area of research (42 59)
Hydrology influences decompositionvia consumers
Detritus (or decaying OM) plays key roles instream ecosystems and most consumers even
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 4 of 13
33104
10103
32101
10100
3110-2
9810-4
3110-5
1922 1949 1976 2004
4
16
256
4096
Perio
d (d
ays)
1963Closure of Glen Canyon Dam
Fig 3 Flow regime shifts in the Colorado River at Lees Ferry Arizona over the past 97 years(1922ndash2018) The flow regime changed drastically around 1963 with the closure of Glen CanyonDam Natural seasonality was dampened and artificial weekly and subweekly scales emerged Graydashed lines indicate 7-day flow cycles resulting from flow management for hydropower andyearly flow cycles resulting from extended base flows and spring snowmelt floods Colors representwavelet power and confidence level contours identify statistically significant power A very wetperiod in the mid-1980s and the experimental releases that took place after 1996 restored flowevents but not the regime Across dammed rivers of the American Southwest muted flowseasonality has opened niches for non-natives particularly organisms adapted to living in stableresource-limited environments at the expense of many native fishes adapted to highly variable(periodic or stochastic) flow conditions that characterized free-flowing hydrographs (155) Plotcreated with the biwavelet R package (156) using mean daily discharge data from United StatesGeological Survey station 09380000 [Dam logo US National Park Service]
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
apical predators are at least partially supportedby it (60) Macroconsumers break up particulateorganic matter (POM) into small pieces enhanc-ing the overall mineralization rate by bacteriaand fungi and even moderate flows may resultin export of OM particularly fine POM resultingfrommacroconsumer shredding activities Over-all howevermicrobes largely drive C losses throughrespiration (61) and although nutrient availa-bility temperature and POM composition (egleaf type) influence this how flow mediates rela-tionships has been more difficult to pin downRecent work (62) designed to separate the effectsthat different components of the flow regimehave on the mechanisms driving decompositionin perennial streams found that dissolution andmicrobial processing consistently exceeded theeffects of fragmentation and microbial pro-cessing was best explained by daily variabilityin discharge When flow regimes become in-termittent the lack ofwater can impedemicrobialand consumer colonization of POM resulting indecreases in decomposition the extent of whichis related to the duration of the drying (46 63)Consistent water cover is therefore importantto decomposition and thus shifts toward lowerflows when litter inputs are high (eg in thefall) can have a large impact on consumers andassociated food webs (63 64)
When flows are altered are changes inbiota and ecosystem processes linked
To help ground this question in the world ofrestoration practice we begin with a focus ontwo of the most common forms of river flow al-teration damming and urbanization For theformer we emphasize hydropower dams becausethey are growing in number they strongly alterflow regimes and the mitigation of their impactshas spurred the field of environmental flow re-search We also focus on urban streams becausestudies on the impacts of imperviousness havebeen extensive and these streams have been the
target of numerous restoration efforts We summa-rize knowledge on the effects of flow on organ-isms ecosystem processes and their interaction inthese two types of river systems and then describehow this can inform river restoration in general
Urbanization and dams changeflow regimes
With few exceptions flow regimes below hydro-power dams and in urban streams are highlyaltered at one or both ends of the flow spectrumRegimes in impounded rivers are typically lessseasonal than daily rainfall would suggest par-ticularly below large hydropower dams with highwater-storage capacity rather than below smallerless impactful run-of-the-river facilities (65)Hydro-power dams canmute peak river flows but power-generation cycles induce rapidly fluctuating flowpatterns (hydropeaking) that often occur onhuman-relevant scales [ie hours to weeks (18)]tending to bemore pronounced in seasons of highenergy demand Hydropeaking is often accom-panied by spikes in temperature (thermopeak-ing) either cold or warm depending on seasonand reservoir operation (66) Thus downstreamorganisms and ecosystems are generally subjectto multiple periodic stresses In urban streamsbaseflows can be higher or lower depending onwatershed characteristics and infrastructurehowever peak flows are invariably larger thanthose prior to urbanization Understanding theecological impacts of urban stormflow is com-plicated by indirect effects on water quality In-creases in flow magnitude frequency and ratesof change during storms leads to elevated con-centrations of pollutants Despite this studiesseparating the causes of biological impairmenthave reported that the unique effects of alteredurban flows can be significant (67 68) Spikesin stream temperature caused by runoff fromhot pavement (69) add to stress on the biotaHydropeaking and urban stormflows might
be expected to reduce primary production by
scouring algae (70) yet some studies have re-ported elevated GPP or alternating periods ofnet autotrophy and heterotrophy (71ndash73) Lightcan override the effects of flow on metabolismby enhancing algal photosynthesis (74) how-ever if hydropeaking increases turbidity GPPmay decline (73) In urban streams elevatedGPP is often attributed to open canopies andorto elevated nutrient concentrations both of whichoften co-occur with their altered flow regimes(72) As discussed later such shifts in primaryproductivity have been linked to changes introphic structure eg more algal-based foodwebs (75) depending on the relative availabil-ity of terrestrial carbon inputs (both DOM andPOM) (76) Inputs of POM and its quality as afood resource changewith landscape context (egvegetative cover) and catchment hydrology (77)but POM availability can also be limited by flowvariability which can increase decompositionrates and decrease residence times (78 79)As is the case for running-water systems in
general high flows in urban streams bring pulsesof solutes including N DOM and various pol-lutants (80) Solute dynamics and biogeochemicalprocesses in regulated rivers have been insuffi-ciently studied with researchers focusing moreon such processes in reservoirs rather than indownstream river ecosystems We found nostudies attempting to link urban or regulatedflow regimes and solute fluxes to changes infood webs or stream metabolic balance How-ever recent studies show shifts in microbial tax-onomic and functional composition in responseto pulsed urban flows (81) and thesemay help inexploring a potential link but are complicated bythe fact that non-flow stressors affect consumercomposition For example urban and regulatedrivers typically have animal communities thatare less diverse and composed of tolerant groupsoften non-native Hydropeaking and urban stormflows can certainly cause direct mortality via dis-placement and transport to unsuitable habitatsbut many of the organismal effects are realizedthrough indirect pathways For instance hydro-peaking reduces the viability of river-edge egg-laying specialists by limiting their access to suitablespawning habitat disrupting reproductive successand insect emergence (82) Flow regulation caneven create ecological traps if organismal phenol-ogy shiftswith thenovel environmental conditionsFor example dam-induced warming triggeredsummer diapause in a mayfly resulting in theloss of its last generation (83) Dams also isolatepopulations harming organisms that requiremigrations to complete their life cycles whetherthese are between riverine and marine habitatsor within the freshwater domain By analogy al-tered flow regimes in urban streams can also leadto isolated pools in streams with lowered base-flows This can be associated with periods ofhypoxia punctuated by extreme flow pulses inwhich these streams become ldquoscoured or suffo-catedrdquo (74)As illustrated using urban streams and reg-
ulated rivers research is moving forward andhints at the three-way nexus but is clearly still
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 5 of 13
Fig 4 Ecosystem metabolismpatterns across a river networkSpatial patterns in river eco-system metabolism for the Deva-Cares catchment in northernSpain as represented by the ratioof gross primary production(P) to ecosystem respiration (R)A PR gt 1 indicates autotrophicprocesses dominate so theseriver sections are accumulatingor exporting organic carbonPR lt 1 indicates dominanceby heterotrophic processes inriver sections that are receivingorganic C input (eg fromterrestrial sources) The boxesindicate regions of the networkimpacted by human pressures including deforestation (brown box) waste products from urbansites (green box) and a combination of the two (purple box) Such visualizations are usefulto managers in understanding regions of concern that may need the most restoration actions[Reproduced with permission from (157)]
PR ratiolt025
gt150
025 ndash 050 050 ndash 075075 ndash 100100 ndash 150
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
inadequate to meet restoration needs Studieson ecosystem processes below dams are parti-cularly limited and much of what we knowcomes from only a few river systems Furtherpublished work focuses primarily on benthicecosystem processes immediately below damsrather than riverine planktonic realms whereproductivity in large regulated rivers can beimportant
Linking flow biota andecosystem processes
Research on flow regimes that bridges organis-mal and ecosystem ecology has traditionallyrelied on states instead of rates ie nutrientconcentrations rather than uptake or fluxesstanding stocks of algae rather than productivityor metabolism and abundance of organismsrather than secondary productivity This workgenerally shows that flow variability suppressesand stability increases algal standing stock andgrazer densities with peaks in invertebrate bio-mass following those of algae [eg (84 85)]Over the past decade research began to focuson how flow regimes alter functional aspects offoodwebs For example biofilms inMediterranean-climate rivers are thicker andmetabolicallymoreactive downstream of flow-stabilizing dams (71)When flow variability increases below hydro-power dams fluctuations can reduce biofilmdevelopment and its quality as a food resource(86) Using experimental flumes subjected tohydropeaking shifts have been found in periph-yton fatty acid content from important highlyunsaturated compounds to nonessential saturatedones (87)Indirect evidence suggests that flow-induced
changes in dominant resource type (eg algaeversus detritus) resource quantity (biomass) orresource quality (nutrient content) can propa-gate to higher trophic levels via bottom-up ef-fects For instance detritivorous invertebrateswere found to dominate species richness andinvertebrate biomass upstream but not down-stream of an irrigation dam (75) diets of om-nivore insects indicated that flow stabilizationshifted a detrital into an algal-fueled food webSimilar evidence comes from Glen Canyon Damin the Colorado River (88) Near the dam whereprimary production is high (73) food webs aresimplified and gut content analyses have sug-gested algal production supports gt50 of in-vertebrate and gt70 of fish production andthe food web can be easily perturbed by flooddisturbance Farther downriver food webs showincreased reliance on detrital resources havehigher trophic efficiency and are more complexand more resistant to disturbance (76) In con-trast recent work on seasonal tropical riversfound algal productivity and zero-flow distur-bance did not explain food-web structure Inthis case it seems that highly mobile predatorsbuffered the local effects of drought (89) Thisstudy along with earlier ones at the site (90) isof particular interest because it links temporalpatterns in predatory fish with flow-dependentavailability of resources
Only a few studies have provided evidence oftop-down controls on foodwebs being influencedby flow variation Fish in a tropical floodplainriver were found to exert top-down control onprimary producers but only under certain hydro-logic conditions (91) Similarly in aCalifornian riverwinter flood pulses were shown to control insectconsumers late in the season ultimately deter-mining whether fish exerted top-down controlon the lower trophic levels (10) These illustratethe importance of pulsed hydrology on resource-consumer linkages and show that not just standingstocks but also the dominant controls structuringfood webs may fluctuate over time in response toflow variationAn indirect line of evidence supporting the
importance of the flow-biota-ecosystem processesnexus comes from theoretically oriented researchon food-web structure Changes in productivityhabitat size and disturbance underpin the basisof the three main hypotheses of food-chainlength which state that chains should be longerin stable productive large ecosystems (92) No-tably flow regimes influence these three con-trols Because food-chain length determines keyaspects of a river ecosystem such as the risk oftrophic cascades or the pollutant accumulationin top predators understanding how it is af-fected by flow alteration is important How-ever understanding the interplay between thehypotheses of food-chain length and the flow-dependent biological mechanisms that allowfood webs to shift in structure (eg predatorinvasions local extinctions consumers chang-ing diets) is still a relatively recent endeavor(93ndash95) Evidence suggests that zero-flows short-en food chains mainly by keeping streamsfishless (94 96) whereas stabilization of variableflows can lengthen food chains by allowing om-nivores to feed on abundant high trophic levelprey (95)New studies are also focusing on interactions
between drivers of food-web structure that hadbeen previously only been examined in isolationsuch as disturbance regimes and ecosystem sizeboth influenced by flow regime Using metabolictheory predator biomass has been shown toscale with both prey resources and fluctuatingstream habitat size with reductions in preda-tor size leading to declines in predator biomasssupported per unit of prey biomass (97) Movingforward the combination of habitat andmetabolicconstraints on predators may provide a basis toconnect flow-induced disturbance changes inecosystem processes and habitat fluctuationsto riverine food-web structure The metabolictheory of ecology (98) is a unifying frameworkthat allows scaling responses of organisms towhole ecosystems andmay become particularlyuseful in situations where flow alteration affectsorganismal body size or water temperature Insuch cases strong changes in metabolic ratescan be expected affecting resource uptake andorganismal growth to ecosystem-level produc-tion and respirationFinally an important body of food webndashrelated
research that provides strong inferential evidence
for the nexus comes from studies on animal-mediated nutrient cycling and ecological stoi-chiometry Theory suggests detrital and algalfood websmay be linked by cascading effects onnutrient cycling (99) and empirical work showsthat organismal excretion andmigration (whichrelocates C) may be important to this link (100)(Fig 5) A recent study (101) showed that ex-cretion of highly labile dissolved organic C ni-trogen (N) and phosphorus (P) by aquatic insectsmay be sufficient to support a substantial fractionof microbial energy and nutrient needs Giventhat C availability can be too low tomeetmicrobialenergy needs [eg (102 103)] and that microbialactivity enhances detrital quality to consumers(104) a C subsidy from excretory processes couldbe important at least during low-flow periodsAdditionally microbial use of excreted dissolvedorganic nitrogen (DON) and DOP may releasealgae from potential competition with microbesfor inorganic N and P if this is the case GPPshould increase Ecological stoichiometry mayalso be important for understanding constraintson organismal growth rates as rapid growthdisproportionally increases demand of P [as it isused in ribosomal RNA (105)] This could in turnlimit the ability of organisms to grow fast andexploit temporary habitats Interestingly elemen-tal ratios can vary interspecifically in response totemperature or predation pressure (106)mdashfactorsthat increase with low flows and habitat fragmen-tation Overall studies that link biota with bio-geochemical processes remain rare (107) butanalyzing flow regimes as a driver of elementturnover rates may provide a way forward
Can hydroecological advances informrestoration practice
Shortfalls in the outcomes of river restorationhave prompted calls to identify and restore pro-cesses that support and sustain biological com-munities rather than focusing only on rivergeomorphology and habitat In urban streamsscientists have critiqued an overemphasis onstructural engineered approaches (7 108) thatdo little to restore the full flow regime (109) Inlarge regulated rivers identifying and restoringimportant features of the flow regime via damoperations is often possible (albeit expensive)However that does not solve other problemsassociated with dams such as habitat frag-mentation (110) altered sediment and thermalregimes (111 112) or disrupted biogeochemicalprocesses (113) Overall efforts to alter reservoirreleases or improve urban stormwater infra-structure to partially mimic a riverrsquos natural flowregime have great potential Technologies arebeing evaluated for real-time management ofurban flows and novel ways to design flow re-leases from dams are being proposed (114 115)however research still lags far behind the needfor solutions Importantly the paucity of studieson the flow-biota-ecosystem processes nexus pointsto a need for more integrative research connectinghydrologists population and community ecologistsand ecosystem ecologists in the context ofrestoration
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 6 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
Restoring flow regimes ratherthan eventsScientifically informed flow designs may existthat maintain ecosystem integrity in regulatedrivers (116 117) but these have produced mixedresults to date In the dam-altered Putah Creekin California mimicking the natural timing ofhigh and low flows enhanced native fish by im-proving their spawning habitat reducing watertemperatures maintaining flowing conditionsand reducing the abundance of non-native spe-cies (118) However other similar attempts havebeen less successful In some cases non-nativefish density also benefitted from flow patternsmimicking an unregulated regime (119) in othersnon-natives bounced back shortly after the flowrelease (120) These experiences illustrate thateven natural flow features may have inadvertentnegative effects if only part of the community orecosystem is considered Models for the NavajoDam (San Juan River New Mexico) show thatadequately timed releases could simultaneouslyenhance native and minimize non-native fishpopulations (121) thus predicted benefits couldsurpass those of mimicking a natural flow re-gime potentially persisting even during dryyears Another study using the ldquodesignerrdquo flowparadigm (114) attempted to understand howdifferent facets of flow variability control fisheryproduction in the Lower Mekong floodplains Time-
series models combining discharge and fisherycatch suggested that enhanced flood pulses andlong inter-flood intervals may increase fisheryyieldsAlthough environmental flow practices have
mostly focused on re-creating discrete flowevents (eg flood pulses) these can still serve asvaluable experiments An iconic example of alarge-scale flow release is Minute 319 an amend-ment to the United StatesndashMexico treaty allow-ing opening of floodgates in the Lower ColoradoRiver and a one-time rewetting of the parchedColorado River Delta (122) Although this pulsed-flow experiment did not promote establishmentof native woody riparian seedlings such trialscan inform future restoration practices for ex-ample by determining whether the provision ofbare moist substrate enhances seed germina-tion (123) Hypothesis-driven experimental flowreleases combined with deep knowledge of tar-get ecosystems can provide insights into resto-ration actions [eg (124)] Still an outstandingquestion is how flow regime restoration couldbe leveraged to restore other important alteredaspects of the ecosystem such as sediment andthermal regimes For example cold-water releasesby existing dams in climate-vulnerable basinscould prevent organisms that thrive at low tem-peratures from becoming replaced by general-ist warm-tolerant taxa (125)
Although the concept of environmental flowsis generally associated with impounded riversits relevance to urban streams is clear (126)However because frequent high storm flowsare viewed as the problem reducing them gen-erally becomes the only restoration goal This isoften attempted by shifting the spatial distribu-tion of velocities within channels through ma-nipulation of channel sinuosity installation offlow-diverting weirs or adding other structuresThese approaches may reduce local erosion butrarely result in levels of water quality ecosystemprocesses or biodiversity in urban streams thatcome close to those of unimpacted streams(127 128) Focusing on the channel ratherthan the watershed context at large means thatkey ecosystemprocesses can remain compromised(129) A recent study compared time series ofmetabolism in two streamsmdashonewith stormwaterponds in the watershed and the other restoredby altering channel slope banks and in-streamstructural complexity (103) The study found thatpeaks in GPP and ER in the ldquorestoredrdquo streamwere higher more prone to resets induced byflashy storms and that recovery time scales werelonger compared with the stream in a watershedwith stormwater measures (Fig 6) Ponds areone type of nature-inspired green infrastructureused to restore streamflow tomore natural condi-tions tree plantings constructed wetlands andgrass swales can in turn replace gray infrastruc-ture such as concrete tunnels that capture andstore waterDistributing green infrastructure in water-
sheds helps restore streams because it targetsthe cause of streamflow alteration includinginsufficient infiltration groundwater rechargeand more generally limited watershed storagecapacity (129) By tackling the cause rather thanthe symptoms of flow alteration these practicescan reduce ecological impacts that result fromthe cascading effects of altered flows includingpoor water quality and temperature spikes Re-search on the effectiveness of green infrastructureon restoring streams is sparse and limited bylack of data before and after implementationalthough studies are beginning to appear Forexample upland green infrastructure in theform of shallow wetland-like pools that capturestormwater and reconnect it to the groundwaterhas been shown to enhance baseflows while re-ducing peak flows and pollutant loadsmore thanrestoration structures within the mainstem ofthe receiving stream (130) Similar structuresbuilt between stormwater outfalls and perennialstreams did not modulate the timing magnitudeor duration of urban storm responses nor didthey improve water quality or biodiversity (131)illustrating the need for researchers to understandhow exact design and watershed context influ-ence performance
Spatiotemporal variability andrestoration designs
Research showing how local and watershed-scale environmental drivers vary temporally andalong river networks in ways that shape biota
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 7 of 13
Flow pulses (storms) flashy flowsDroughts intermittencyLowstable flowsAltered timing seasonality
DOC N P
DetritusMicrobial
stoichiometric constraints
Excretion exudates
Detrital pathway
Algal pathway
Algal stoichiometric constraints
Organismal dispersal source-sink dynamics
Algae
Community replacement
ndash ndash +
ndash ndash + ndash
+
+ndash +
Invasions
Turnover
Consumer stoichiometric constraints
Fig 5 Flow variation affects biota and ecosystem processes and food webs integrate bothHydrologic connectivity along river networks is a first landscape-level filter constraining dispersal oforganisms in river networks Extreme flow variation (floods and droughts) are a source of disturbanceto all organisms (gray box) but particularly to the higher trophic levels given their higher metabolicdemands In turn flow variation controls inputs of nutrients and terrestrial leaf litter to the stream whichinfluences microbial activity and the relative contributions of the detrital (brown) and algal (green)pathways in sustaining aquatic consumers Not just quantity but also food quality (eg CN and CPratios) influence dietary choices of consumers and thus trophic transfer efficiency Finally alteration offlow regimes may influence all these compartments and may also open niches for new players Ifbiological invasions occur at the top of the food web these have the potential to divert energy away fromnative predators and to alter food-web structure through top-down controls
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
and ecosystem processes can help prioritizeselection of sites for restoration For exampleusing the concept of river network saturation(55) to identify river sections where in-streamremoval of nutrients cannot keep up with highsource inputs of nutrients suggests that restorationactions targeting those sections may dispropor-tionately improve water quality Identification ofsubcatchments that disproportionately influencestream chemistry may also be informed by com-bining methods from landscape ecology andcatchment hydrology focusing on the spatialscale of variance in water chemistry (132) Head-waters are often prioritized for restoration be-
cause poor water quality in them can havedownstream impacts Research showing thatlocal environmental constraints on river com-munities are important in headwaters as well asin the larger channels but dispersal constraintsare much less important in the former (133 134)implies that headwaters should be prioritizedHowever research quantifying time-varying con-tributions of local communities to river-wide(network) biodiversity can also inform prioritiza-tion by identifying sites that host a dispropor-tionate share of species or that host uniquespeciesmdashfor example because they shelter sensi-tive taxa during critical flow bottlenecks This idea
encapsulates the concept of keystone habitatsor local sites that strongly influence metacom-munity dynamics (135) This has been appliedto aquatic invertebrates across intermittent rivernetworks by tracking time-varying site-specificcontributions to community dissimilarity (or betadiversity) (136)Other approaches to prioritize restoration ac-
tions may be based on contributions to system-wide stability rather than numbers of individualsor species For example the restoration of fall-run Chinook salmon portfolios may be assessedvia the contributions that different periods andlocationsmake tometapopulation-wide stabilityowing to spatiotemporal variation in habitatconditions (137) Additionally interventionscould promote conditions that are not impor-tant now but could be important in the future[ie proto-refugia (138)] For example resto-ration of groundwater-influenced habitats couldmitigate future stress to drought-sensitive taxaunder scenarios of increased intermittencyPatches of undisturbed land that protect flowregimes in streams within a larger catchmentthat is otherwise urbanized can be sources ofcolonists to restored sites Priorities that focuson present degradation of river sectionsmay failto address site contribution to impairment orrecovery at broader scales or under differentscenarios These insights indicate a great poten-tial for new methods to prioritize restoration ofboth urban and regulated streamsWe know little about how altered flow regimes
propagate through entire river networks butwork is starting to assess the cumulative effectsof dams on flow regimes as well as dam-levelcontributions to alteration Although these ap-proaches have largely focused on dam cascades[eg (139)] if expanded they could help identifyfavorable sections in river networks to developenvironmental flow operations Here exper-imentation could again help with testing ofnew strategies For instance large-scale flowexperiments could be designed that coordinatereleases among dams or between dams and free-flowing tributaries to enhance the restorativeeffects of natural pulses (140)
Hydrologic connectivityinfluences restoration
Recovering critical flow paths and their connec-tivity to rivers is important but it remains asubstantial challenge for flow-degraded riversand streams Whereas regulated rivers tend tohave reduced lateral and longitudinal hydro-logic connectivity urban streams have unnaturallevels of connectivity because pipes and over-land runoff route water quickly In both cases fun-damental hydroecological insights into streambedstructure landscape storage capacity and ground-water connectivity can help to envision novel re-storation approaches (130 141 142) More generallyhydrologic connectivity is becoming a frameworkto identify alternative ways to restore streams viastormwater management (143) For example arecent study suggested that reducing road densityand hydrologic connectivity between roads
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 8 of 13
ER
Disc
harg
e (f
t3 s)
GPPG
PP o
r ER
(g O
2 m-2
day
-1)
Discharge
Watershed BMPs
In-stream restoration
0
ndash4
4
0
ndash4
4
4
2
6
4
2
6
Jan Sept July May Mar Nov Jan
A
B C
Fig 6 Green infrastructure as a means to restore stream ecosystems (A) Discharge andecosystem respiration (ER more-negative values mean higher respiration) and gross primaryproduction (GPP) for two urban streams in which restoration efforts involved (top) modifications inthe stream channel and (bottom) traditional stormwater best management practices (BMPs)[Reproduced with permission from (103)] (B and C) Example of a comprehensive greeninfrastructure approach to ecologically restore an urban headwater stream that also supports thelifestyle of resident urban communities (B) Stream at the onset of restoration and (C) afterrestoration [Reproduced with permission from Greenworks Portland OR]
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 9 of 13
Table 1 Research challenges and opportunities Despite the progress that has been made in understanding how flow regimes affect biota and ecosystem
processes major challenges persist that prevent a complete understanding of the flow-biota-ecosystem processes nexus Making progress on these
challenges requires a mix of fundamental and applied research
Topic Research challenge Opportunity
Connecting
organismal to
ecosystem-level
processes
Research on the effects of flow on organisms
and ecosystem processes has followed parallel
tracksmdashhindering our mechanistic
understanding of the flow-biota-ecosystem
processes nexus
Generate theory that connects concepts and frameworks
that are well accepted in river community and ecosystem
ecology For example the notion of environmental filters
that connect regional to local composition via a combination
of dispersal and environmental conditions (community
ecology) with analogous theory on solute transport versus
reactivity (ecosystem ecology)
Our current understanding of the links between
phenology of biota and ecosystem processes
is precursory
Use high-frequency sensors to characterize metabolic
regimes at sites with ongoing organismal monitoring
Leverage citizen-science biodiversity data to detect
spatiotemporal shifts in animal phenology
(eg insect emergence fish spawning)
Use remote sensing tools to better understand
links between hydrology and aquatic and riparian
plant phenology
The link between stoichiometry and flow
variation remains largely unexplored
Repurpose data that is already being collected to test
flow-stoichiometry relationships eg studies using C and
N stable isotopes to describe CN ratios across flow
regime types Perform field experiments to quantify
indirect effects of invertebrate and fish consumption
on ecosystem metabolism
Ecological
consequences of
hydrologic
connectivity
The importance of anaerobic metabolism in
carbon processing as flows cease and parts of
streambeds dry is a major frontier research area
Create networks of sites across different geologies
and flow regimes that collect time-series data on
methane nitrous oxide and carbon dioxide fluxes
from intermittent streams
We ignore how biota in the wet phase is
influenced by processes and patterns
from the dry phase
Research the functions of dry riverbeds Use camera traps and
drone technology to investigate the role of dry riverbeds
as a dispersal corridor metabarcoding to quantify
ldquodark biodiversityrdquo in the hyporheic
Wetlands and their links to stream networks are poorly
delineated globally Even in the United States and
Europe current wetland inventories do not capture
these links yet understanding how their connectivity
to river systems influences network-scale ecosystem
processes is needed
Analyze time series from high-frequency sensors distributed in
river networks varying in wetland connectivity Further
development and application of time-series methods
used by neuroscientists
We need to identify the relationship between
varying flows and solute dynamics
associated with changes in catchment-scale
hydrologic exchanges
Design collaborative work among catchment hydrologists
and stream ecologists combining methods to estimate
transit times residence times and identify sources
across heterogeneous landscapes Link these properties to
land management and water quality aspects
Scaling and
transferring flow-
ecology relationships
across space and time
We ignore how biodiversity responds to
propagation of flow regime alteration
across river networks
Leverage extensive hydrometric networks (eg United States
Geological Survey National Water Information System)
to perform spatially replicated time-series analyses
on flow and use these to generate expectations
on how river network position should mediate
biodiversity responses to flow alteration
Flow-ecology relationships do not account for
nonstationarity and abrupt change
Use process-based models that account for mechanisms
and associated uncertainties When using time-series
data and models select methods that are robust
to nonstationarity Linear flow-ecology relationships
should not be assumed
Current flow-ecology relationships assume that
responses of species do not change over time
Consider flow regime alteration as a process with ecological and
evolutionary consequences Include intraspecific variation
in traits and potential for rapid evolution in models
continued on next page
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
and streams may improve water quality morethan other restoration options (70) Additionallyresearch on the age and lability of DOC enteringstreams has shown how precipitation or land dis-turbance can change watershed flow paths (53)and this could inspire new restoration approachesfocused on land use and soil properties Manystudies on hydrologic connectivity and networkstructure suggest a need for restoration to focuson landscape heterogeneity especially on thepresence ofwetlands lakes and ponds (144) Suchfeaturesare frequently lost inurbanizedcatchmentsand outcomes could be improved by restoring orincreasing connectivitywith these ecosystems (145)Hydrologic connectivity associated with ground-water inputs or abstractions land-use manage-ment and return flows fromwastewater treatmentplants also need to be considered in these initia-tives (146)Restoring hydrologic connectivity is also key
to achieving biodiversity outcomesmdashparticularlyin dammed systems Longitudinal connectivityfor fish inhabiting dam-altered rivers can beenhanced by the construction of fishways andsophisticated passage facilities However moni-toring data on the effectiveness of such inter-
ventions are still scarce and available evidencesuggests that passage effectivity largely dependson technology used and species-specific fish be-havior (147 148) Although trap-and-transportprograms for migratory fishes can help augmentthreatened populations (149) these practicesoften increase straying and thus the potentialfor genetic homogenization as seen from ju-venile salmon collected at dams and transporteddownstream to increase out-migration success(150) Overall a combination of engineering andmanagement solutions may be best to restorefunctional connectivity in dam-altered riversThe link between hydrologic connectivity andecological restoration is also central to con-siderations on where to build remove and re-operate hydrologic infrastructure Hundredsof dams have been intentionally removed sincethe 1970s and removal of large dams such asthe Elwha River Dam (Washington USA) hasshown that restoring longitudinal connectivitycan have immediate geomorphic effects with com-plex temporal dynamics (151) Infrastructure canalso be designed or reoperated to manage forrestoring lateral connectivity and enhanced flood-plain processes For example the Yolo Bypass in
the Sacramento River California has becomea prime rearing and migration site for chinooksalmon showing that flood control and habitatfor fish and wildlife do not need to be at odds(152) All these examples show the importanceof integrating hydrologic connectivity in ecolog-ical restoration whether the goal is to minimizethe ecological effects of fragmentation restorefree-flowing watercourses or operate green infra-structure to enhance infiltration floodplain con-nectivity or other ecological processes
Research challenges and opportunities
Two decades of concerted efforts to research theeffects of flow regime alteration have tremen-dously advanced our understanding of the waysin which streamflow influences biota and eco-system processes However important challengesremain These aremainly related to the scarcity ofscience connecting organismal to ecosystem-levelprocesses through for example microbial linksthe difficulty of studying watershed-level phys-ical and biogeochemical heterogeneity and itsinfluence on riverine processes and biota thecomplexity of parsing out the direct effects offlow from flow-mediated effects and co-occurring
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 10 of 13
Topic Research challenge Opportunity
Cascading
implications of
hydrologic
alteration
We have a poor understanding of the
interactions among physical drivers
Build on large-scale flow experiments to test hypotheses on the
combined effects of altered flow sediment and thermal
regimes Experiment with multilevel intakes in reservoirs
Promote collection of non-flow variables (eg temperature
water quality parameters) at hydrometric stations
At low flows biota are being exposed to high
concentrations of new classes of pollutants
whose impacts are poorly known especially if
biotic impacts interact with impacts on
ecosystem processes
Design realistic experimental studies to advance knowledge
on the ecological effects of multiple stressors
Collect long-term data at sites varying in contaminant
inputs and hydrologic regimes Link microbial to
organismal and ecosystem-level responses
Using hydroecological
science to inform
river ecosystem
restoration
Environmental flow science largely ignores
human dimensions
Use methods that allow identifying causal networks
in complex socioenvironmental systems Include
human needs and thresholds in research on
flow-ecology relationships
Environmental flows are generally prescribed
at the scale of single infrastructures restoration
designs and assessment are generally at reach scales
Promote coordinated large-scale flow experiments
and restoration designs at the river basin scale
Extend assessment of green infrastructure
effectiveness beyond individual structures to
infrastructure networks and link both
to river outcomes
Environmental flows in impounded rivers
generally target the conservation of some
particular species rather than outcomes at the
food-web or ecosystem level
Advance fundamental research on microbialndash
ecosystem process linkages Expand the notion
of designer flows to food-web structure and
ecosystem services when possible
Environmental flow science has largely
ignored surface waterndashgroundwater
interactions
Expand science to understand the link between
restoration actions groundwater recharge and
inputs to streams Provide science relevant to
the implementation of new legislation on
groundwater management (eg Sustainable
Groundwater Management Act in California)
Uncertainty regarding future
hydroclimate and
river flows is growing
Refine regional climate models Promote research on
the interaction between land-use change and
evapotranspiration Incorporate groundwater
influence in macroscale hydrology models
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
stressors (particularly in urban streams) and thechallenge of predicting outcomeswhen both thephysical and the biological environments arechangingAchieving a predictive understanding of hy-
droecological relationships is essential to design-ing successful self-sustaining restoration actionsin river ecosystems However translating basichydroecological insights into restoration practicerequires understanding the complexities of work-ing within a human-dominated water cycle Al-though it is rarely possible to restore all facetsof a natural flow regime ecologically informeddesigns may help mimic key aspects of the nat-ural flow regime (eg periodic pulse flows) orcreate regimes that are not natural but maximizeoutcomes given socioeconomic constraints InTable 1 we identify persisting and emergingchallenges in hydroecological research We paireach challenge with an opportunity (related toresearch design technology or implementation)We contend that these challenges should beprioritized to advance process-based river eco-system restoration
REFERENCES AND NOTES
1 N L Poff et al The natural flow regime Bioscience 47769ndash784 (1997) doi 1023071313099
2 D A Lytle N L Poff Adaptation to natural flow regimesTrends Ecol Evol 19 94ndash100 (2004) doi 101016jtree200310002 pmid 16701235
3 S Sabater A Elosegi R Ludwig in Multiple Stressors in RiverEcosystems (Elsevier 2019) pp 1ndash22
4 United Nations New UN Decade on Ecosystem Restorationoffers unparalleled opportunity for job creation food securityand addressing climate change (2019) httpswwwunenvironmentorgnews-and-storiespress-releasenew-un-decade-ecosystem-restoration-offers-unparalleled-opportunity
5 A W Lorenz P Haase K Januschke A SundermannD Hering Revisiting restored river reaches - Assessingchange of aquatic and riparian communities afterfive years Sci Total Environ 613-614 1185ndash1195(2018) doi 101016jscitotenv201709188pmid 28954379
6 M A Palmer K L Hondula B J Koch Ecological Restorationof Streams and Rivers Shifting Strategies and ShiftingGoals (2014) vol 45
7 T J Beechie et al Process-based principles for restoringriver ecosystems Bioscience 60 209ndash222 (2010)doi 101525bio20106037
8 S E Bunn A H Arthington Basic principles and ecologicalconsequences of altered flow regimes for aquaticbiodiversity Environ Manage 30 492ndash507 (2002)doi 101007s00267-002-2737-0 pmid 12481916
9 K D Fausch Y Taniguchi S Nakano G D GrossmanC R Townsend Flood disturbance regimes influence rainbowtrout invasion success among five holarctic regions EcolAppl 11 1438ndash1455 (2001) doi 1018901051-0761(2001)011[1438FDRIRT]20CO2
10 M E Power M S Parker W E Dietrich Seasonalreassembly of a river food web Floods droughts andimpacts of fish Ecol Monogr 78 263ndash282 (2008)doi 10189006-09021
11 V Acuntildea A Giorgi I Muntildeoz U R S Uehlinger S SabaterFlow extremes and benthic organic matter shape themetabolism of a headwater Mediterranean streamFreshwater Biol 49 960ndash971 (2004) doi 101111j1365-2427200401239x
12 K Tockner F Malard J V Ward An extension of the floodpulse concept Hydrol Processes 14 2861ndash2883 (2000)doi 1010021099-1085(20001112)141617lt2861AID-HYP124gt30CO2-F
13 J S Perkin et al Fragmentation and dewatering transformGreat Plains stream fish communities Ecol Monogr 8573ndash92 (2015) doi 10189014-01211
14 J L Sabo D M Post Quantifying periodic stochastic andcatastrophic environmental variation Ecol Monogr 7819ndash40 (2008) doi 10189006-13401
15 F Shuai et al Silver carp larva abundance in response toriver flow rate revealed by cross-wavelet modellingEcol Modell 383 98ndash105 (2018) doi 101016jecolmodel201805020
16 J D Tonkin M T Bogan N Bonada B Rios-ToumaD A Lytle Seasonality and predictability shape temporalspecies diversity Ecology 98 1201ndash1216 (2017)doi 101002ecy1761 pmid 28144975
17 N L Poff Beyond the natural flow regime Broadening thehydro‐ecological foundation to meet environmental flowschallenges in a non‐stationary world Freshwater Biol 631011ndash1021 (2018) doi 101111fwb13038
18 A Ruhi X Dong C H McDaniel D P Batzer J L SaboDetrimental effects of a novel flow regime on the functionaltrajectory of an aquatic invertebrate metacommunity GlobChange Biol 24 3749ndash3765 (2018) doi 101111gcb14133pmid 29665147
19 M T Bogan D A Lytle Severe drought drives novelcommunity trajectories in desert stream pools FreshwaterBiol 56 2070ndash2081 (2011) doi 101111j1365-2427201102638x
20 G Woodward et al The effects of climatic fluctuations andextreme events on running water ecosystems Philos TransR Soc London Ser B 371 20150274 (2016) pmid 27114576
21 M A Palmer C M Swan K Nelson P Silver R AlvestadStreambed landscapes Evidence that stream invertebratesrespond to the type and spatial arrangement of patchesLandsc Ecol 15 563ndash576 (2000) doi 101023A1008194130695
22 D Schmera et al Does isolation influence the relative role ofenvironmental and dispersal‐related processes in streamnetworks An empirical test of the network positionhypothesis using multiple taxa Freshwater Biol 63 74ndash85(2018) doi 101111fwb12973
23 R Henriques‐Silva et al A comprehensive examination of thenetwork position hypothesis across multiple rivermetacommunities Ecography 42 284ndash294 (2019)doi 101111ecog03908
24 K L Jaeger J D Olden N A Pelland Climate change poisedto threaten hydrologic connectivity and endemic fishes indryland streams Proc Natl Acad Sci USA 11113894ndash13899 (2014) doi 101073pnas1320890111pmid 25136090
25 S Garbin E A Celegon P Fanton G Botter Hydrologicalcontrols on river network connectivity R Soc Open Sci 6181428 (2019) doi 101098rsos181428 pmid 30891270
26 F Cattaneacuteo B Hugueny N Lamouroux Synchrony in browntrout Salmo trutta population dynamics A lsquoMoran effectrsquo onearly‐life stages Oikos 100 43ndash54 (2003) doi 101034j1600-0706200311912x
27 A Terui et al Metapopulation stability in branching rivernetworks Proc Natl Acad Sci USA 115 E5963ndashE5969(2018) doi 101073pnas1800060115 pmid 29895695
28 P A P Moran The statistical analysis of the Canadian Lynxcycle Aust J Zool 1 291ndash298 (1953) doi 101071ZO9530291
29 G D Grossman P B Moyle J O Whitaker Jr Stochasticityin structural and functional characteristics of an Indianastream fish assemblage A test of community theoryAm Nat 120 423ndash454 (1982) doi 101086284004
30 D A Lytle D M Merritt J D Tonkin J D OldenL V Reynolds Linking river flow regimes to riparian plantguilds A community-wide modeling approach Ecol Appl 271338ndash1350 (2017) doi 101002eap1528 pmid 28263426
31 A Ruhiacute E E Holmes J N Rinne J L Sabo Anomalousdroughts not invasion decrease persistence of native fishesin a desert river Glob Change Biol 21 1482ndash1496 (2015)doi 101111gcb12780 pmid 25430731
32 J D Tonkin D M Merritt J D Olden L V ReynoldsD A Lytle Flow regime alteration degrades ecologicalnetworks in riparian ecosystems Nat Ecol Evol 2 86ndash93(2018) doi 101038s41559-017-0379-0 pmid 29180707
33 M C Mims J D Olden Fish assemblages respond toaltered flow regimes via ecological filtering of lifehistory strategies Freshwater Biol 58 50ndash62 (2013)doi 101111fwb12037
34 P T J Johnson J D Olden M J Vander Zanden Daminvaders Impoundments facilitate biological invasions intofreshwaters Front Ecol Environ 6 357ndash363 (2008)doi 101890070156
35 N Bonada S Doledec B Statzner Taxonomic and biologicaltrait differences of stream macroinvertebrate communitiesbetween mediterranean and temperate regions Implicationsfor future climatic scenarios Glob Change Biol 131658ndash1671 (2007) doi 101111j1365-2486200701375x
36 M I Pyne N L Poff Vulnerability of stream communitycomposition and function to projected thermal warming andhydrologic change across ecoregions in the western UnitedStates Glob Change Biol 23 77ndash93 (2017) doi 101111gcb13437 pmid 27429092
37 D B Herbst S D Cooper R B Medhurst S W WisemanC T Hunsaker Drought ecohydrology alters the structureand function of benthic invertebrate communities inmountain streams Freshwater Biol 64 886ndash902 (2019)doi 101111fwb13270
38 M T Bogan E T Chester T Datry A L MurphyB J Robson A Ruhi R Stubbington J E Whitney inResistance Resilience and Community Recovery in IntermittentRivers and Ephemeral Streams T Datry N Bonada A BoultonEds (Academic Press 2017) pp 349ndash376
39 S Houmlckendorff et al Characterizing fish responses to a riverrestoration over 21 years based on speciesrsquo traits ConservBiol 31 1098ndash1108 (2017) doi 101111cobi12908pmid 28218807
40 D Bruno et al Structural and functional responses ofinvertebrate communities to climate change andflow regulation in alpine catchments Glob Change Biol25 1612ndash1628 (2019) doi 101111gcb14581pmid 30698905
41 T W H Aspin et al Extreme drought pushes streaminvertebrate communities over functional thresholds GlobChange Biol 25 230ndash244 (2019) doi 101111gcb14495pmid 30346098
42 S E Tank J B Fellman E Hood E S Kritzberg Beyondrespiration Controls on lateral carbon fluxes across theterrestrial-aquatic interface Limnol Oceanogr Lett 3 76ndash88(2018) doi 101002lol210065
43 A P Appling et al The metabolic regimes of 356 rivers inthe United States Sci Data 5 180292 (2018) doi 101038sdata2018292 pmid 30532078
44 P Savoy et al Metabolic rhythms in flowing watersAn approach for classifying river productivity regimesLimnol Oceanogr (2019) doi 101002lno11154
45 E S Bernhardt et al The metabolic regimes of flowingwaters Limnol Oceanogr 63 S99ndashS118 (2018)doi 101002lno10726
46 M Colls X Timoner S Sabater V Acuntildea Effects of durationand frequency of the non-flow period on stream metabolismEcosystems 101007s10021-019-00345-1 (2019)doi 101007s10021-019-00345-1
47 A J Ulseth E Bertuzzo G A Singer J Schelker T J BattinClimate-induced changes in spring snowmelt impactecosystem metabolism and carbon fluxes in an Alpine streamnetwork Ecosystems 21 373ndash390 (2018) doi 101007s10021-017-0155-7
48 B O L Demars Hydrological pulses and burning of dissolvedorganic carbon by stream respiration Limnol Oceanogr 64406ndash421 (2019) doi 101002lno11048
49 R T Hensley L Kirk M Spangler M N Gooseff M J CohenFlow extremes as spatiotemporal control points on riversolute fluxes and metabolism J Geophys Res Biogeosci124 537ndash555 (2019) doi 1010292018JG004738
50 J D Hosen et al Enhancement of primary production duringdrought in a temperate watershed is greater in larger riversthan headwater streams Limnol Oceanogr 64 1458ndash1472(2019) doi 101002lno11127
51 L Goacutemez-Gener et al When water vanishes Magnitude andregulation of carbon dioxide emissions from dry temporarystreams Ecosystems 19 710ndash723 (2016) doi 101007s10021-016-9963-4
52 E H Stanley et al The ecology of methane in streams andrivers Patterns controls and global significance EcolMonogr 86 146ndash171 (2016) doi 10189015-1027
53 R T Barnes D E Butman H F Wilson P A RaymondRiverine export of aged carbon driven by flow path depth andresidence time Environ Sci Technol 52 1028ndash1035 (2018)doi 101021acsest7b04717 pmid 29313674
54 G Pinay et al Riparian corridors A new conceptualframework for assessing nitrogen buffering across biomesFront Environ Sci 6 47 (2018) doi 103389fenvs201800047
55 W M Wollheim et al River network saturation conceptFactors influencing the balance of biogeochemical supply and
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 11 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
demand of river networks Biogeochemistry 141 503ndash521(2018) doi 101007s10533-018-0488-0
56 P A Raymond J E Saiers W V Sobczak Hydrological andbiogeochemical controls on watershed dissolved organicmatter transport Pulse-shunt concept Ecology 97 5ndash16(2016) doi 10189014-16841 pmid 27008769
57 C Tunaley D Tetzlaff J Lessels C Soulsby Linking high-frequency DOC dynamics to the age of connected watersources Water Resour Res 52 5232ndash5247 (2016)doi 1010022015WR018419
58 S Bernal et al Supply demand and in-stream retention ofdissolved organic carbon and nitrate during storms inMediterranean forested headwater streams Front EnvironSci 7 60 (2019) doi 103389fenvs201900060
59 J Harvey et al How hydrologic connectivity regulates waterquality in river corridors J Am Water Resour Assoc 55369ndash381 (2019) doi 1011111752-168812691
60 J C Moore et al Detritus trophic dynamics and biodiversityEcol Lett 7 584ndash600 (2004) doi 101111j1461-0248200400606x
61 A D Rosemond et al Freshwater ecology Experimentalnutrient additions accelerate terrestrial carbon loss fromstream ecosystems Science 347 1142ndash1145 (2015)doi 101126scienceaaa1958 pmid 25745171
62 A C Y Yeung J L Musetta-Lambert D P KreutzweiserP K Sibley J S Richardson Relations of interannualdifferences in stream litter breakdown with dischargeBioassessment implications Ecosphere 9 e02423 (2018)doi 101002ecs22423
63 R M Northington J R Webster Experimental reductions instream flow alter litter processing and consumer subsidies inheadwater streams Freshwater Biol 62 737ndash750 (2017)doi 101111fwb12898
64 E Mas-Martiacute I Sanpera-Calbet I Muntildeoz Bottom-up effectsof streambed drying on consumer performance throughchanges in resource quality Aquat Sci 79 719ndash731 (2017)doi 101007s00027-017-0531-6
65 B D Richter G A Thomas Restoring environmental flows bymodifying dam operations Ecol Soc 12 12 (2007)doi 105751ES-02014-120112
66 G Zolezzi A Siviglia M Toffolon B MaioliniThermopeaking in Alpine streams Event characterizationand time scales Ecohydrology 4 564ndash576 (2011)doi 101002eco132
67 T Meiszligner B Sures C K Feld Multiple stressors and therole of hydrology on benthic invertebrates in mountainousstreams Sci Total Environ 663 841ndash851 (2019)doi 101016jscitotenv201901288 pmid 30738264
68 I R Waite et al Effects of urban multi-stressors on threestream biotic assemblages Sci Total Environ 6601472ndash1485 (2019) doi 101016jscitotenv201812240pmid 30743940
69 K C Nelson M A Palmer Stream temperature surges underurbanization and climate change Data models andresponses J Am Water Resour Assoc 43 440ndash452 (2007)doi 101111j1752-1688200700034x
70 J R Blaszczak J M Delesantro Y Zhong D L UrbanE S Bernhardt Watershed urban development controls onurban streamwater chemistry variability Biogeochemistry144 61ndash84 (2019) doi 101007s10533-019-00572-7
71 L Ponsatiacute et al Biofilm responses to flow regulation by damsin Mediterranean rivers River Res Appl 31 1003ndash1016(2015) doi 101002rra2807
72 J M Alberts J J Beaulieu I Buffam Watershed land useand seasonal variation constrain the influence of ripariancanopy cover on stream ecosystem metabolism Ecosystems20 553ndash567 (2017) doi 101007s10021-016-0040-9pmid 30416371
73 R O Hall Jr et al Turbidity light temperature andhydropeaking control primary productivity in the ColoradoRiver Grand Canyon Limnol Oceanogr 60 512ndash526 (2015)doi 101002lno10031
74 J R Blaszczak J M Delesantro D L Urban M W DoyleE S Bernhardt Scoured or suffocated Urban streamecosystems oscillate between hydrologic and dissolvedoxygen extremes Limnol Oceanogr 64 877ndash894 (2019)doi 101002lno11081
75 J-R Mor et al Dam regulation and riverine food-webstructure in a Mediterranean river Sci Total Environ 625301ndash310 (2018) doi 101016jscitotenv201712296pmid 29289778
76 H A Wellard Kelly et al Macroinvertebrate diets reflecttributary inputs and turbidity-driven changes in food
availability in the Colorado River downstream of GlenCanyon Dam Freshw Sci 32 397ndash410 (2013)doi 10189912-0881
77 S J Imberger R M Thompson M R Grace Urbancatchment hydrology overwhelms reach scale effects ofriparian vegetation on organic matter dynamics FreshwaterBiol 56 1370ndash1389 (2011) doi 101111j1365-2427201102575x
78 R T Martins A S Melo J F Gonccedilalves Jr N HamadaLeaf-litter breakdown in urban streams of Central AmazoniaDirect and indirect effects of physical chemical andbiological factors Freshw Sci 34 716ndash726 (2015)doi 101086681086
79 C M Yule J Y Gan T Jinggut K V Lee Urbanizationaffects food webs and leaf-litter decomposition in a tropicalstream in Malaysia Freshw Sci 34 702ndash715 (2015)doi 101086681252
80 T A Newcomer et al Influence of natural and novel organiccarbon sources on denitrification in forest degraded urbanand restored streams Ecol Monogr 82 449ndash466 (2012)doi 10189012-04581
81 A Chaudhary I Kauser A Ray R Poretsky Taxon-drivenfunctional shifts associated with storm flow in an urbanstream microbial community MSphere 3 e00194-18 (2018)doi 101128mSphere00194-18 pmid 29976643
82 T A Kennedy et al Flow management for hydropowerextirpates aquatic insects undermining river food websBioscience 66 561ndash575 (2016) doi 101093bioscibiw059
83 A Glazaczow D Orwin M Bogdziewicz Increasedtemperature delays the late-season phenology of multivoltineinsect Sci Rep 6 38022 (2016) doi 101038srep38022pmid 27905493
84 S G Fisher L J Gray N B Grimm D E Busch Temporalsuccession in a desert stream ecosystem following flashflooding Ecol Monogr 52 93ndash110 (1982) doi 1023072937346
85 G J Scrimgeour M J Winterbourn Effects of floods onepilithon and benthic macroinvertebrate populations in anunstable New Zealand river Hydrobiologia 171 33ndash44 (1989)doi 101007BF00005722
86 E Bondar-Kunze S Maier D Schoumlnauer N Bahl T HeinAntagonistic and synergistic effects on a stream periphytoncommunity under the influence of pulsed flow velocityincrease and nutrient enrichment Sci Total Environ 573594ndash602 (2016) doi 101016jscitotenv201608158pmid 27585428
87 M J Cashman G L Harvey G Wharton M C Bruno Woodmitigates the effect of hydropeaking scour on periphytonbiomass and nutritional quality in semi-natural flumesimulations Aquat Sci 79 459ndash471 (2017) doi 101007s00027-016-0510-3
88 W F Cross et al Food-web dynamics in a large riverdiscontinuum Ecol Monogr 83 311ndash337 (2013)doi 10189012-17271
89 D M Warfe et al Productivity disturbance and ecosystemsize have no influence on food chain length in seasonallyconnected rivers PLOS ONE 8 e66240 (2013)doi 101371journalpone0066240
90 T D Jardine et al Consumer-resource coupling in wet-drytropical rivers J Anim Ecol 81 310ndash322 (2012)doi 101111j1365-2656201101925x pmid 22103689
91 K O Winemiller et al Pulsing hydrology determines top‐down control of basal resources in a tropical riverndashfloodplainecosystem Ecol Monogr 84 621ndash635 (2014)doi 10189013-18221
92 D M Post The long and short of food-chain length TrendsEcol Evol 17 269ndash277 (2002) doi 101016S0169-5347(02)02455-2
93 D M Post G Takimoto Proximate structural mechanismsfor variation in food‐chain length Oikos 116 775ndash782 (2007)doi 101111j0030-1299200715552x
94 P A McHugh A R McIntosh P G Jellyman Dual influencesof ecosystem size and disturbance on food chain length instreams Ecol Lett 13 881ndash890 (2010) doi 101111j1461-0248201001484x pmid 20482579
95 A Ruhiacute et al Flow regulation increases food-chain lengththrough omnivory mechanisms in a Mediterranean rivernetwork Freshwater Biol 61 1536ndash1549 (2016) doi 101111fwb12794
96 J L Sabo J C Finlay T Kennedy D M Post The role ofdischarge variation in scaling of drainage area and food chainlength in rivers Science 330 965ndash967 (2010) doi 101126science1196005 pmid 20947729
97 A R McIntosh et al Capacity to support predators scaleswith habitat size Sci Adv 4 eaap7523 (2018) doi 101126sciadvaap7523 pmid 29978038
98 J H Brown J F Gillooly A P Allen V M Savage G B WestToward a metabolic theory of ecology Ecology 851771ndash1789 (2004) doi 10189003-9000
99 K Zou E Theacutebault G Lacroix S Barot Interactions betweenthe green and brown food web determine ecosystemfunctioning Funct Ecol 30 1454ndash1465 (2016) doi 1011111365-243512626
100 M J Vanni P B McIntyre Predicting nutrient excretion ofaquatic animals with metabolic ecology and ecologicalstoichiometry A global synthesis Ecology 97 3460ndash3471(2016) doi 101002ecy1582 pmid 27912023
101 T B Parr K A Capps S P Inamdar K A Metcalf Animal-mediated organic matter transformation Aquatic insects as asource of microbially bioavailable organic nutrients andenergy Funct Ecol 33 524ndash535 (2019) doi 1011111365-243513242
102 C P Arango et al Urban infrastructure influences dissolvedorganic matter quality and bacterial metabolism in an urbanstream network Freshwater Biol 62 1917ndash1928 (2017)
103 L G Larsen J W Harvey Disrupted carbon cycling inrestored and unrestored urban streams Critical timescalesand controls Limnol Oceanogr 62 S160ndashS182 (2017)doi 101002lno10613
104 H M Halvorson G White J T Scott M A Evans-WhiteDietary and taxonomic controls on incorporation of microbialcarbon and phosphorus by detritivorous caddisfliesOecologia 180 567ndash579 (2016) doi 101007s00442-015-3464-6 pmid 26497125
105 J J Elser et al Biological stoichiometry from genes toecosystems Ecol Lett 3 540ndash550 (2000) doi 101046j1461-0248200000185x
106 E K Moody et al Consumption explains intraspecificvariation in nutrient recycling stoichiometry in a desert fishEcology 99 1552ndash1561 (2018) doi 101002ecy2372pmid 29882955
107 N Welti et al Bridging food webs ecosystem metabolismand biogeochemistry using ecological stoichiometry theoryFront Microbiol 8 1298 (2017) doi 103389fmicb201701298 pmid 28747904
108 M A Palmer S Filoso R M Fanelli From ecosystems toecosystem services Stream restoration as ecologicalengineering Ecol Eng 65 62ndash70 (2014) doi 101016jecoleng201307059
109 M J Burns T D Fletcher C J Walsh A R LadsonB E Hatt Hydrologic shortcomings of conventional urbanstormwater management and opportunities for reformLandsc Urban Plan 105 230ndash240 (2012) doi 101016jlandurbplan201112012
110 C R Liermann C Nilsson J Robertson R Y NgImplications of dam obstruction for global freshwater fishdiversity Bioscience 62 539ndash548 (2012) doi 101525bio20126265
111 J D Olden R J Naiman Incorporating thermal regimes intoenvironmental flows assessments Modifying dam operationsto restore freshwater ecosystem integrity Freshwater Biol55 86ndash107 (2010) doi 101111j1365-2427200902179x
112 D B Osmundson R J Ryel V L Lamarra J Pitlick Flowndashsedimentndashbiota relations Implications for river regulationeffects on native fish abundance Ecol Appl 12 1719ndash1739(2002)
113 D von Schiller et al Regulation causes nitrogen cyclingdiscontinuities in Mediterranean rivers Sci Total Environ540 168ndash177 (2016) doi 101016jscitotenv201507017pmid 26233271
114 J L Sabo et al Designing river flows to improve foodsecurity futures in the Lower Mekong Basin Science 358eaao1053 (2017) doi 101126scienceaao1053pmid 29217541
115 W Xu et al Improving the multi-objective performance ofrainwater harvesting systems using real-time controltechnology Water 10 147 (2018) doi 103390w10020147
116 M Acreman et al Environmental flows for naturalhybrid and novel riverine ecosystems in a changingworld Front Ecol Environ 12 466ndash473 (2014)doi 101890130134
117 N L Poff et al Sustainable water management under futureuncertainty with eco-engineering decision scaling Nat ClimChang 6 25ndash34 (2016) doi 101038nclimate2765
118 J D Kiernan P B Moyle P K Crain Restoring native fishassemblages to a regulated California stream using the
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 12 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
natural flow regime concept Ecol Appl 22 1472ndash1482(2012) doi 10189011-04801 pmid 22908707
119 D L Propst K B Gido Responses of native andnonnative fishes to natural flow regime mimicry in theSan Juan River Trans Am Fish Soc 133 922ndash931 (2004)doi 101577T03-0571
120 T K Pool J D Olden Assessing long‐term fish responses andshort‐term solutions to flow regulation in a dryland river basinEcol Freshwat Fish 24 56ndash66 (2015) doi 101111eff12125
121 W Chen J D Olden Designing flows to resolve human andenvironmental water needs in a dam-regulated riverNat Commun 8 2158 (2017) doi 101038s41467-017-02226-4 pmid 29255194
122 S M Nelson et al A history of the 2014 Minute 319environmental pulse flow as documented by fieldmeasurements and satellite imagery Ecol Eng 106733ndash748 (2017) doi 101016jecoleng201610040
123 P B Shafroth et al A large-scale environmental flowexperiment for riparian restoration in the Colorado RiverDelta Ecol Eng 106 645ndash660 (2017) doi 101016jecoleng201702016
124 E P Glenn P L Nagler P B Shafroth C J JarchowEffectiveness of environmental flows for riparian restorationin arid regions A tale of four rivers Ecol Eng 106 695ndash703(2017) doi 101016jecoleng201701009
125 S Hotaling D S Finn J Joseph Giersch D W WeisrockD Jacobsen Climate change and alpine stream biologyProgress challenges and opportunities for the futureBiol Rev Camb Philos Soc 92 2024ndash2045 (2017)doi 101111brv12319 pmid 28105701
126 C J Walsh T D Fletcher M J Burns Urban stormwaterrunoff A new class of environmental flow problem PLOS ONE7 e45814 (2012) doi 101371journalpone0045814pmid 23029257
127 A Selvakumar T P OrsquoConnor S D Struck Role of streamrestoration on improving benthic macroinvertebrates and in-stream water quality in an urban watershed Case studyJ Environ Eng 136 127ndash139 (2010) doi 101061(ASCE)EE1943-78700000116
128 R W Lammers B P Bledsoe What role does streamrestoration play in nutrient management Crit Rev EnvironSci Technol 47 335ndash371 (2017) doi 1010801064338920171318618
129 H E Golden N Hoghooghi Green infrastructure and itscatchment-scale effects An emerging science WIREs Water5 e1254 (2018) doi 101002wat21254 pmid 29682288
130 M R Williams G Bhatt S Filoso G Yactayo Streamrestoration performance and its contribution to theChesapeake Bay TMDL Challenges posed by climate changein urban areas Estuaries Coasts 40 1227ndash1246 (2017)doi 101007s12237-017-0226-1
131 R M Fanelli K Prestegaard M A Palmer Freshw Sci101086706072 (2019)
132 B W Abbott et al Unexpected spatial stability of waterchemistry in headwater stream networks Ecol Lett 21296ndash308 (2018) doi 101111ele12897 pmid 29282860
133 B L Brown C M Swan Dendritic network structureconstrains metacommunity properties in riverineecosystems J Anim Ecol 79 571ndash580 (2010) doi 101111j1365-2656201001668x pmid 20180874
134 C M Swan B L Brown Metacommunity theory meetsrestoration Isolation may mediate how ecologicalcommunities respond to stream restoration Ecol Appl 272209ndash2219 (2017) doi 101002eap1602 pmid 28718193
135 N Mouquet D Gravel F Massol V Calcagno Extending theconcept of keystone species to communities and ecosystemsEcol Lett 16 1ndash8 (2013) doi 101111ele12014 pmid 23062191
136 A Ruhiacute T Datry J L Sabo Interpreting beta-diversitycomponents over time to conserve metacommunities inhighly dynamic ecosystems Conserv Biol 31 1459ndash1468(2017) doi 101111cobi12906 pmid 28188969
137 L Yamane L W Botsford D P Kilduff Tracking restorationof population diversity via the portfolio effect J Appl Ecol55 472ndash481 (2018) doi 1011111365-266412978
138 D W Braidwood M A Taggart M Smith R AndersenTranslocations conservation and climate change Use ofrestoration sites as protorefuges and protorefugia RestorEcol 26 20ndash28 (2018) doi 101111rec12642
139 Y Wang D Wang Q W Lewis J Wu F Huang A frameworkto assess the cumulative impacts of dams on hydrologicalregime A case study of the Yangtze River Hydrol Processes31 3045ndash3055 (2017) doi 101002hyp11239
140 J L Sabo et al Pulsed flows tributary inputs and food-webstructure in a highly regulated river J Appl Ecol 551884ndash1895 (2018) doi 1011111365-266413109
141 C Magliozzi G Coro R C Grabowski A I PackmanS Krause A multiscale statistical method to identifypotential areas of hyporheic exchange for river restorationplanning Environ Model Softw 111 311ndash323 (2019)doi 101016jenvsoft201809006
142 S S Schwartz B Smith Restoring hydrologic function inurban landscapes with suburban subsoiling J Hydrol 543770ndash781 (2016) doi 101016jjhydrol201610051
143 C J Walsh T D Fletcher A R Ladson Stream restoration inurban catchments through redesigning stormwater systemsLooking to the catchment to save the stream J N AmBenthol Soc 24 690ndash705 (2005) doi 10189904-0201
144 N M Schmadel et al Thresholds of lake and reservoirconnectivity in river networks control nitrogen removalNat Commun 9 2779 (2018) doi 101038s41467-018-05156-x pmid 30018449
145 C D Sayer Conservation of aquatic landscapes Pondslakes and rivers as integrated systems WIREs Water 1573ndash585 (2014) doi 101002wat21045
146 R G Luthy D L Sedlak M H Plumlee D Austin V H ReshWastewater‐effluent‐dominated streams as ecosystem‐
management tools in a drier climate Front Ecol Environ 13477ndash485 (2015) doi 101890150038
147 D W Roscoe S G Hinch Effectiveness monitoring of fishpassage facilities Historical trends geographic patterns andfuture directions Fish Fish 11 12ndash33 (2010) doi 101111j1467-2979200900333x
148 C M Bunt T Castro-Santos A Haro Performanceof fish passage structures at upstream barriers tomigration River Res Appl 28 457ndash478 (2012)doi 101002rra1565
149 D Weigel et al Evaluation of a trap-and-transport programfor a threatened population of steelhead (Oncorhynchusmykiss) Conserv Genet 20 1195ndash1199 (2019) doi 101007s10592-019-01200-5
150 M H Bond et al Combined effects of barge transportationriver environment and rearing location on straying andmigration of adult Snake River fall-run Chinook SalmonTrans Am Fish Soc 146 60ndash73 (2017) doi 1010800002848720161235614
151 A C Ritchie et al Morphodynamic evolution followingsediment release from the worldrsquos largest dam removalSci Rep 8 13279 (2018) doi 101038s41598-018-30817-8pmid 30185796
152 T Sommer et al Californiarsquos Yolo Bypass Evidence thatflood control can be compatible with fisheries wetlandswildlife and agriculture Fisheries 26 6ndash16 (2001)doi 1015771548-8446(2001)026lt0006CYBgt20CO2
153 G Grill et al Mapping the worldrsquos free-flowing riversNature 569 215ndash221 (2019) doi 101038s41586-019-1111-9pmid 31068722
154 G Gassert M Luck M Landis P Reig T Shiao ldquoAqueductglobal maps 21 Constructing decision-relevant global waterrisk indicatorsrdquo Working paper World Resources Institute(2015) wwwwriorgpublicationaqueduct-global-maps-21-indicators
155 A Ruhiacute J D Olden J L Sabo Declining streamflow inducescollapse and replacement of native fishes in the AmericanSouthwest Front Ecol Environ 14 465ndash472 (2016)doi 101002fee1424
156 T C Gouhier A Grinsted V Simko R package Biwaveletconduct univariate and bivariate wavelet analyses (2018)
157 T Rodriacuteguez-Castillo E Esteacutevez A M Gonzaacutelez-FerrerasJ Barquiacuten Estimating ecosystem metabolism toentire river networks Ecosystems (2018) doi 101007s10021-018-0311-8
ACKNOWLEDGMENTS
We greatly appreciate those who read and provided importantinsights on drafts of this paper A Armstrong K HondulaJ Hosen K Leathers G de Mendoza C Maietta I MunozJ Tonkin and two anonymous reviewers We also thank T Tu forhelp with Fig 1 and S Carlson L Comte T Datry J HosenM Kondolf N Jones D Lytle E Moody J Olden M PowerJ Sabo and M Sanchez-Montoya for valuable discussionsCompeting interests The authors have no competing interestsFunding MAP was supported by funds from the NSF (1639145)and AR was supported by funds from the University of CaliforniaBerkeley and NSF (1802714)
101126scienceaaw2087
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 13 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
restorationLinkages between flow regime biota and ecosystem processes Implications for river
Margaret Palmer and Albert Ruhi
DOI 101126scienceaaw2087 (6459) eaaw2087365Science
this issue p eaaw2087Scienceflow variability and achieve hydrological connectivity between a river and its surroundingsand ecosystem processes To be successful such efforts must go beyond accounting for flood pulses to restore natural mimic ecologically important aspects of natural flow regimes guided by insights into how variations in flow affect biotawater quality and ecological processes In a Review Palmer and Ruhi explain how restoration designs now attempt to
Human activities have altered the flow regimes of many of Earths rivers with negative impacts on biodiversityRiver restoration guided by research
ARTICLE TOOLS httpsciencesciencemagorgcontent3656459eaaw2087
REFERENCES
httpsciencesciencemagorgcontent3656459eaaw2087BIBLThis article cites 147 articles 7 of which you can access for free
PERMISSIONS httpwwwsciencemagorghelpreprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAASScienceScience 1200 New York Avenue NW Washington DC 20005 The title (print ISSN 0036-8075 online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
Science No claim to original US Government WorksCopyright copy 2019 The Authors some rights reserved exclusive licensee American Association for the Advancement of
on May 27 2020
httpsciencesciencem
agorgD
ownloaded from

REVIEW
HYDROLOGY
Linkages between flow regimebiota and ecosystem processesImplications for river restorationMargaret Palmer1dagger and Albert Ruhi2
River ecosystems are highly biodiverse influence global biogeochemical cycles andprovide valued services However humans are increasingly degrading fluvial ecosystemsby altering their streamflows Effective river restoration requires advancing ourmechanistic understanding of how flow regimes affect biota and ecosystem processesHere we review emerging advances in hydroecology relevant to this goal Spatiotemporalvariation in flow exerts direct and indirect control on the composition structure anddynamics of communities at local to regional scales Streamflows also influenceecosystem processes such as nutrient uptake and transformation organic matterprocessing and ecosystem metabolism We are deepening our understanding of howbiological processes not just static patterns affect and are affected by stream ecosystemprocesses However research on this nexus of flow-biota-ecosystem processes is atan early stage We illustrate this frontier with evidence from highly altered regulated riversand urban streams We also identify research challenges that should be prioritized toadvance process-based river restoration
Rivers have been critical to human existencesince antiquity and are a central part ofthe biosphere Flowing waters sustain riv-erine terrestrial and marine biodiversityand make important contributions to global
biogeochemical cycles However river ecosystemsare increasingly degraded by dam building di-version or abstraction of water clearing of landand climate change Some rivers that once werehealthy and diverse now only support drought-or pollution-tolerant species others shunt eu-trophic water toward coastal regions or offer newhabitat to non-native species These degradedrivers all have one characteristic in commonsome or all aspects of their flows have been al-tered (Fig 1)Flow regime or the characteristic pattern of
flow variation has long been known to be a keydriver of a riverrsquos structure and functioning (1)The characteristic magnitude frequency dura-tion timing and rate of change in river flows haveshaped awide range of species adaptationsmdashfromlife history strategies to behaviors andmorphol-ogies of both aquatic and riparian organisms (2)A riverrsquos flow regime also influences in-stream andflood-plain ecosystemprocesses including primaryproduction and nutrient cycling Because flowdynamism is central to a riverrsquos functioning andits ability to provide ecosystem services flow alter-ation is rarely inconsequential
When flows are altered a combination ofbiotic and abiotic pathways are triggered Forinstance flash floods that reduce predators canhave cascading biotic effects on primary producersand on associated nutrient dynamics Flows thatincrease suspended sediments may inhibit orga-nismal feeding and reproduction or reduceprimaryproduction These are both examples of indirectabiotic effects Although altered flows are oftennot the proximate mechanism of ecological de-gradation they can exacerbate the impacts of otherabiotic stressors (3) For example extreme lowflows resulting from excessive withdrawals canincrease water temperature and pollutant con-centrations to the point that they exceed tolerablelevels for organismal survival or reproductionThe mechanisms linking flow regime alterationto ecological degradation can be numerous andcomplexGiven themagnitude and global extent of river
degradation it is fitting that the United NationsDecade on Ecosystem Restoration has just beendeclared (4) Science to heal streams and rivershas never been more needed Most restorationpractices have focused on improving channelmorphology or habitat and unfortunately re-covery of biodiversity or species of interest hasproven difficult (5 6) This has prompted increasedcalls by scientists to move frommorphological toecosystem-level ldquoprocess-basedrdquo practices (7) thatfocus on restoring flow regimes as well as otherphysical and ecological processes that sometimescovary with flow and support aquatic commu-nities In this Review we summarize the currentunderstanding of the relationship between riverflow regimes biota and ecosystem processes and
outline how future river restoration can benefitfrom a better understanding of the three-wayinteraction that we call the flow-biota-ecosystemprocesses nexus (Fig 2) We address the follow-ing How does flow variation control river biotadirectly and indirectly How does it control riverecosystem processes When flow regimes are al-tered are changes in ecosystem processes coupledwith changes in biota Can these bodies of re-search inform restoration practice and wherecould advances in hydroecology be better lever-agedWe close by identifying key challenges andopportunities in hydroecological researchThis Review focuses primarily on fundamental
research advances however regulated rivers (iethose with dams) and urban streams help groundfindings in real-world problems These two eco-system types have experienced some of the largestshifts in their flow regimes and are central tonumerous restoration efforts Building off of ad-vances needed to restore these degraded eco-systems we highlight research that can informprocess-based restoration more generally Weshow that despite calls for improving the linkbetween science and restoration in practice thereremains a great need to advance research on theflow-biota-ecosystem processes nexus This needopens exciting opportunities for both fundamen-tal and applied interdisciplinary research
How does flow variation controlriver biota
Flow is essential to river life but can also be asource of stress scouring floods remove organismsdroughts stress organismal physiology and flowconditions dictate the changing type quantity andquality of the physical habitat inwhich organismslive (8) However flow variation also influencesorganisms indirectly by keeping predators com-petitors and invaders at bay (9 10) by control-ling the energy sources that enter the food web(11) or by affecting movement of organisms andmatter across river networks and floodplains(12 13) At the inception of the natural flow re-gime concept (1) the study of flow-ecology rela-tionships focused on static representations of flowand local-scale research Over the past 20 yearsimportant progress has beenmade toward under-standing how ecological communities respond todynamic flow regimes across entire river networks
Organisms respond to patterns offlow variation
Stream ecologists have long known that low-and high-flow events can temporarily reduceabundance and diversity of invertebrates andfishes particularly when droughts fragmentthe riverine habitat or when spates mobilize thestreambed However it is not only discrete eventsthat are important long-term patterns of flowvariability have historically selected for organis-mal life histories related to growth reproductiondispersal and the ability to persist under physicaland chemical stress The increasing availabilityof temporally extensive andor high-frequencydatasets is now spurring the use of spectral meth-ods which allow the identification of dominant
RESEARCH
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 1 of 13
1National Socio-Environmental Synthesis Center University ofMaryland Annapolis MD 21401 USA 2Department ofEnvironmental Science Policy and Management Universityof California Berkeley Berkeley CA 94720 USAThese authors contributed equally to this workdaggerCorresponding author Email mpalmerumdedu
on May 27 2020
httpsciencesciencem
agorgD
ownloaded from
frequencies amplitudes and phases in the envi-ronment (14) These characteristic patterns ofvariation or regimes can then be connected toorganismal dynamics For example a waveletanalysis of discharge and abundance of silvercarp larvae found a strong association betweenspawning and peak flows over a particular mag-nitude (15) Using a similar approach temporalchange in stream invertebrate diversity was foundto be influenced not only by the seasonality ofrainfall but by the reliability of this variabilityie its predictability (16)Flow regimes differ across river types and
climates (1) but can also shift temporally at agiven place as a consequence of changes inclimate land use or flow management (Fig 3)Although changes in dominant frequencies inthe environment can have strong impacts on com-munities studying the effects of time-varying flowregimes is still rare (17) Faster flow cycles mayfilter out species with longer generation timesand a preference for stable environments (18) andincreased frequencies of extreme events (eg re-current unpredictable droughts) may drive com-munities to novel stable states (19) However westill do not know which temporal scales of flowvariationmay entrain organismal phenology andwhich may simply represent a source of stressAlthoughmost work to understand this questionhas used correlational approaches mathemat-ical models based on metabolic theory and bio-mechanical constraints may help in predicting
the effects of changing frequencies of environ-mental fluctuations and extremes on river orga-nisms food webs and ecosystem processes (20)
Spatiotemporal variation in flow controlsbiotic persistence
River network structure and directional stream-flow influence the movement of drifting orga-nisms and materials downstream (21) and themain ways in which communities disperse andassemble Experiments suggest that the local envi-ronment exerts control of community compo-sition in the more isolated headwaters whereasdispersal and environmental influences togetheroperate in the well-connected main stem or pri-mary downstream channel This is known as thenetwork position hypothesis (22) Although findingshave sometimes been inconsistent with respectto this idea a recent study using graph theory andreplicate fishmetacommunities (ie multiple com-munities linked by dispersal) suggests that supportfor this hypothesis may depend on network struc-ture and the degree of environmental heteroge-neity in headwaters relative to themainstem (23)Incorporating network structure into the studyof flow regimes is also important because thedendritic connectivity of streams and rivers con-strains organismal movement and associatedpersistence (24) This is particularly relevant toclimate change because shifts in both wet-channel network structure and habitat qualityacross the network are anticipated to occur In
this vein recent research has demonstrated thatin arid climates downstream sections of riversmay bemore suitable for fish spawning whereasin wetter conditions the most suitable habitatoccurs in the headwaters (25)The spatial coordination of flow regimes across
the river network also has key implications forbiodiversity persistence In sets of populationslinked by dispersal or metapopulations a com-bination of diverse physical and biological fea-tures helpsmaintain asynchronous dynamics thusmaximizing stability at larger scales (ie riskspreading or portfolio effect) Only recently havethe simultaneous effects of spatial and temporalvariation in flows been incorporated into streamecology For example widespread flood eventsmay cause synchronous mortality of early lifestages of salmonids [eg (26)] thus weakeningthe portfolio effect On the flip side river branchescan provide diversity in flow and habitat con-ditions ensuring that populations aremaintainedacross the network Using an agent-based modelvalidatedwith long-term fish time series researchhas shown that population asynchrony tends tobe higher across branches than within branchesthus branching network complexity is key to pre-serving metapopulation stability (27) Given thatdams and climate change are making flow re-gimes more similar synchrony in flow-dependentecological processes could be increasing aswell The phenomenon of environmentally forcedsynchrony has long been studied in terrestrial
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 2 of 13
Fig 1 Impacts of river fragmentation by dams urban areas andwater risk across the worldrsquos watersheds Degree of fragmentation(DOF) represents the effects of dams on longitudinal habitatconnectivity and urban impacts (URB) capture the degree of infra-structure development in riparian and floodplain areas (measured asnightlight intensity in urban areas) (153) We averaged river-reach leveldata at the watershed scale using streams with long-term averagedischarge higher than 1 m3s and we then distributed averaged valuesamong the following categories lt5 (low impact) 5 to 20 (mediumimpact) and gt20 (high impact) Colors represent different impact
combinations at the watershed level and flow lines represent themajor rivers of the world Polygons outlined in gray are areas withldquohighrdquo and ldquoextremely highrdquo exposure to water-related risk anaggregated measure of water quantity quality and regulatory riskproposed by the World Resources Institute (154) Around 23 of theworldrsquos watersheds are subject to water-related risks and 19 ofthe watersheds with flow show substantial impacts (ie in gt5 of theriver network) related to longitudinal habitat fragmentation by damsor urban areas These dimensions capture the major sources offlow-related river ecosystem degradation
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
population ecology [Moran theorem (28)] andcould represent an underappreciated emerg-ing risk in fresh waters
Abiotic and biotic influences of flow
Stream ecologists traditionally interpreted thedynamics of communities as a deterministic resultof biotic interactions or as a stochastic conse-quence of the environment (29) However researchhas shown that these fluctuate in relative impor-tance over space and time andmay even influenceeach other For instance biotic interactions mayoccur only under particular environmental con-ditions (30) Inferences gathered from long-termdata combined with time-series methods importedfrom econometrics can help elucidate when and
where abiotic influences have primacy For exam-ple multivariate autoregressive state-space modelsapplied to long-term flow and fish communitydata showed that interannual variation in flowanomalies were the ultimate driver of the dy-namics of a desert stream fish community (31)Although non-native and native fishes interactedsuch effects were small compared with the directeffects of flows on the native component Bioticinteractions that emerge from known species-specific relationships between components of theflow regime such as flood timing and popula-tion dynamics can also be predicted from modelsimulations of community trajectory (30) A recentapplication of these interaction-neutral modelsfound that flows departing from the natural reg-
ime simplified interaction networks among plantguilds in desert rivers (32)A fundamental challenge for community eco-
logy for decades has been quantifying the rela-tive influence of abiotic (flow-driven) and biotic(flow-mediated) effects This is important for theaccurate design and prediction of restoration out-comes including for example anticipatingwhenflow alteration facilitates species invasions bycreating new niches (33) by enhancing propagulepressure or eroding native enemies (34) or by acombination of thesemechanismsDistinguishingthese could help managers compare the benefitsof eradicating non-natives relative to restoringecologically important facets of the flow regime
Flow alteration filters species and traits
Trait-based approaches are becoming increasinglypopular in hydroecology because they enhancecomparison of flow-ecology relationships acrossclimates (35) they lead to predictions about howcommunity change may affect ecosystem func-tioning (36) and they identify the mechanismsthat filter or promote particular taxa under stress-ful conditions such as drought (37) In the case offlow intermittency traits can be used to identifycommunity tipping points by connecting local en-vironmental conditions to organisms with traitsthat allow them to persist in isolated pools or wetsediment or connecting network-level water con-ditions to the ability of highly resilient organismsto drift or actively recolonize from perennial re-fugia (38) Traits can also be used to assess re-storation trajectoriesmdashfor instance by assessingfluctuations in the share of strategies that representdifferent life-history trade-offs [eg species limitedby reproductive capacity or by resources (39)]By studying a communityrsquos taxonomic char-
acteristics (eg species composition) and func-tional characteristics (eg dispersalmodes feedingstrategies) ecologists can determine to whatextent trait redundancy across speciesmay ensureagainst loss of functional diversity For examplein a study on Alpine stream communities subjectto climate change and flow regulation functionaldiversity increased while functional redundancydecreased (40) This finding indicates that ongoinghydroclimatic change can reduce the ability of acommunity to withstand further alteration evenif trait diversity is high because the persistenceof individual traits may be linked to the fate of afew sensitive species In an experimental studycommunities affected by drought presented com-parable taxonomic and functional decays implyingthat these communities were both taxonomicallyand functionally vulnerable (41) The risk of com-munity collapse in response to flow alterationalso depends on the way in which ldquowinningrdquo andldquolosingrdquo traits co-occur in a given species andacross species in a community as a result of aparticular stress Thus community-wide responsesto a particular alterationmay be stronger if speciesaremaladaptive to that change for instance bybeing large-bodied longer-lived and collector-feeding (filtering particulate organic matter) ina scenario of increased flow variability (18) Under-standing how flow alteration impairs species with
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 3 of 13
Nutrient cycling
Primary production
Energy inputs
Secondary production Dispersal refugia Invasions
Food-web interactionsMetabolic regime
Hydrologic connectivity
Streambed sediments
Water chemistry temperature
Channel form riparian vegetation
Decomposition Species demographic rates
River ecosystem integrity
Altered flow regimes
Restored flow regimes
Time
Restoration toolboxAlter pattern of reservoir releasesImprove stormwater controlReforest and restore watersheds
Disc
harg
e w
ater
leve
l Muted high flows
Storm flows
Altered periodicity
Flow homogenization
Shifting baselines
Extreme low flows
Fig 2 Process-based restoration posits that actions must target processes not patternsthat have been degraded River ecosystems are sustained by a host of environmental conditionsor ecosystem structure (gold rectangles) which in turn influence ecosystem processes (greenrectangles) and biotic processes (blue rectangles) These processes as well as ecosystem structureare all controlled by the flow regime Thus correcting facets of flow regime alteration may enhanceriver ecosystem integrity in both direct and indirect ways
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
traits that have effects on ecosystems (eg byparticular feeding strategies) will help anticipatethe risks of losing major ecosystem functionssuch as detrital decomposition (36)
How does flow variation control riverecosystem processes
Rivers produce transform and store organicmatter (OM) and nutrients Carbon (C) and nu-trient concentrations and fluxes vary longitudi-nally and laterally as river networks expand andcontract with rain These variations influenceand are influenced by stream ecosystem metab-olism which can be described as net ecosystemproduction or the difference between gross pri-mary production (GPP) and ecosystem respiration(ER) The metabolic balance between heterotro-phic and autotrophic production that providesresources for consumers can shift seasonally orwith changing flows as can hydrologic connect-ivity that influences inputs of OM and solutes torivers (42) Increasing availability of sensors andadvances inmodeling (43) havemade the study ofmetabolism OM processes and solute dynamicsripe for linking to biotic processes
Flow variability andecosystem metabolism
Spatiotemporal variability in ecosystem processesis a fundamental characteristic of rivers and flowregime is a major driver of this variability (Fig 4)Recentwork shows thatmany rivers in theUnitedStates have strong seasonal patterns in primary
production with spring peaks for larger riversand summer peaks for smaller rivers generallyGPP is negatively correlated with high variationin daily discharge (44) In regions with little sea-sonality andor continual canopy cover GPP islower and its patterns are less predictable (44)Bernhardt et al (45) provide a comprehensivereview of stream metabolism the factors thatinfluence it and why it is difficult to predict Flowhas the largest effect at the extremes such that atlow flowswhen the streambed begins to dry largedrops in GPP occur (46) Storms also often reduceGPP (45) However patterns may vary as a func-tion of flow predictability For example researchin a river characterized by predictable peaks dur-ing spring snowmelt found over two seasons thatGPP rose during spring snowmelt sampling (47)whereas high flows during the winter and sum-mer reduced GPP Within a year this patternresulted in a switch from net autotrophic to netheterotrophic productionThe paths that water takes as it moves to and
within a streammdasha spatial aspect of its flowregimemdashinfluencesmetabolismBycouplingmetab-olism and chemistry time series with informationonhydrologic connectivity and flowpaths ERwasshown to increase in an organic-rich watershedduring peak flows and remained elevated forweeks (48) Sustained hydrologic connectivitywith soils likely fueled inputs of dissolved orga-nic carbon (DOC) that promoted bacterial respi-ration Such inputs can lead to nonlinear responsesinmetabolism For example increasing flows have
led to decreases in GPP up to some thresholdflow beyond which GPP was suppressed by re-duced water clarity from flow-elevated humicDOC (49) In the same vein higher riverine GPPhas been associated with reduced suspendedsediments and high temperatures during low-flow droughts (50) When flows cease entirely andsections of riverbeds dry up carbon dioxide fluxesto the atmosphere can be far higher than those inflowing sections of the channel (51) presumablybecause of spikes in aerobicmicrobial respirationhowever as is the case with methane in riverspartitioning fluxes of these gases to changes inmetabolism (aerobic or anaerobic) is difficult par-ticularly under variable flows (52)
Hydrologic connectivity drivessolute dynamics
Research is growing on how hydrologic connect-ivity to hillslope soils influences water chemistry(53) and how variation in discharge and riverstage influence the strength of bidirectional waterexchanges that affect nitrogen (N) uptake reten-tion and removal (54) Periods of hydrologic con-nectivity may be followed by long periods withlimited exchange which may differ greatly alongthe stream corridor This can lead to complex CN and phosphorus (P) dynamics that influenceor are influenced by microbial and algal produc-tivity When connectivity with watershed solutesources is extensive inputs of solutes such asDOCmay be so high that they exceed the ability of riverecosystem processes such as microbial uptake toregulate their fluxes (55) Theory posits that largestorms pulse terrestrially derived dissolved or-ganicmatter (DOM) into streams and it is quicklyshunted downstream by high flows where it isprocessed in larger parts of the network (56)However how far DOM in pulsedwater is trans-ported is governed not only by discharge but bythe composition of DOMUnderstanding the rela-tionship between DOM sources and its composi-tion is being advanced by combining time-seriesdata with isotopic information to evaluate spatialvariability in hydrologic flow paths transit timeand chemistry as watermoves from the terrestrialto aquatic realms (53 57) It is difficult tomeasurebiotic uptake in streams during storms howeverrecent work suggests that assuming storm flowstransport DOM too quickly for in-stream process-ing may be a poor assumption if the DOM com-position renders it readily utilizable by biota (58)Although hydrologic connectivity is not a newconcept it is not typically considered part of theflow regime yet it can be characterized by itstiming duration and frequency and then linkedto stream processes or patterns Newmethods forquantifying connectivity [eg (59)] are emergingand the energetic and water quality importanceof network connectivity to soils and diverse land-scape elements (such as wetlands and ponds) isbecoming a major area of research (42 59)
Hydrology influences decompositionvia consumers
Detritus (or decaying OM) plays key roles instream ecosystems and most consumers even
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 4 of 13
33104
10103
32101
10100
3110-2
9810-4
3110-5
1922 1949 1976 2004
4
16
256
4096
Perio
d (d
ays)
1963Closure of Glen Canyon Dam
Fig 3 Flow regime shifts in the Colorado River at Lees Ferry Arizona over the past 97 years(1922ndash2018) The flow regime changed drastically around 1963 with the closure of Glen CanyonDam Natural seasonality was dampened and artificial weekly and subweekly scales emerged Graydashed lines indicate 7-day flow cycles resulting from flow management for hydropower andyearly flow cycles resulting from extended base flows and spring snowmelt floods Colors representwavelet power and confidence level contours identify statistically significant power A very wetperiod in the mid-1980s and the experimental releases that took place after 1996 restored flowevents but not the regime Across dammed rivers of the American Southwest muted flowseasonality has opened niches for non-natives particularly organisms adapted to living in stableresource-limited environments at the expense of many native fishes adapted to highly variable(periodic or stochastic) flow conditions that characterized free-flowing hydrographs (155) Plotcreated with the biwavelet R package (156) using mean daily discharge data from United StatesGeological Survey station 09380000 [Dam logo US National Park Service]
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
apical predators are at least partially supportedby it (60) Macroconsumers break up particulateorganic matter (POM) into small pieces enhanc-ing the overall mineralization rate by bacteriaand fungi and even moderate flows may resultin export of OM particularly fine POM resultingfrommacroconsumer shredding activities Over-all howevermicrobes largely drive C losses throughrespiration (61) and although nutrient availa-bility temperature and POM composition (egleaf type) influence this how flow mediates rela-tionships has been more difficult to pin downRecent work (62) designed to separate the effectsthat different components of the flow regimehave on the mechanisms driving decompositionin perennial streams found that dissolution andmicrobial processing consistently exceeded theeffects of fragmentation and microbial pro-cessing was best explained by daily variabilityin discharge When flow regimes become in-termittent the lack ofwater can impedemicrobialand consumer colonization of POM resulting indecreases in decomposition the extent of whichis related to the duration of the drying (46 63)Consistent water cover is therefore importantto decomposition and thus shifts toward lowerflows when litter inputs are high (eg in thefall) can have a large impact on consumers andassociated food webs (63 64)
When flows are altered are changes inbiota and ecosystem processes linked
To help ground this question in the world ofrestoration practice we begin with a focus ontwo of the most common forms of river flow al-teration damming and urbanization For theformer we emphasize hydropower dams becausethey are growing in number they strongly alterflow regimes and the mitigation of their impactshas spurred the field of environmental flow re-search We also focus on urban streams becausestudies on the impacts of imperviousness havebeen extensive and these streams have been the
target of numerous restoration efforts We summa-rize knowledge on the effects of flow on organ-isms ecosystem processes and their interaction inthese two types of river systems and then describehow this can inform river restoration in general
Urbanization and dams changeflow regimes
With few exceptions flow regimes below hydro-power dams and in urban streams are highlyaltered at one or both ends of the flow spectrumRegimes in impounded rivers are typically lessseasonal than daily rainfall would suggest par-ticularly below large hydropower dams with highwater-storage capacity rather than below smallerless impactful run-of-the-river facilities (65)Hydro-power dams canmute peak river flows but power-generation cycles induce rapidly fluctuating flowpatterns (hydropeaking) that often occur onhuman-relevant scales [ie hours to weeks (18)]tending to bemore pronounced in seasons of highenergy demand Hydropeaking is often accom-panied by spikes in temperature (thermopeak-ing) either cold or warm depending on seasonand reservoir operation (66) Thus downstreamorganisms and ecosystems are generally subjectto multiple periodic stresses In urban streamsbaseflows can be higher or lower depending onwatershed characteristics and infrastructurehowever peak flows are invariably larger thanthose prior to urbanization Understanding theecological impacts of urban stormflow is com-plicated by indirect effects on water quality In-creases in flow magnitude frequency and ratesof change during storms leads to elevated con-centrations of pollutants Despite this studiesseparating the causes of biological impairmenthave reported that the unique effects of alteredurban flows can be significant (67 68) Spikesin stream temperature caused by runoff fromhot pavement (69) add to stress on the biotaHydropeaking and urban stormflows might
be expected to reduce primary production by
scouring algae (70) yet some studies have re-ported elevated GPP or alternating periods ofnet autotrophy and heterotrophy (71ndash73) Lightcan override the effects of flow on metabolismby enhancing algal photosynthesis (74) how-ever if hydropeaking increases turbidity GPPmay decline (73) In urban streams elevatedGPP is often attributed to open canopies andorto elevated nutrient concentrations both of whichoften co-occur with their altered flow regimes(72) As discussed later such shifts in primaryproductivity have been linked to changes introphic structure eg more algal-based foodwebs (75) depending on the relative availabil-ity of terrestrial carbon inputs (both DOM andPOM) (76) Inputs of POM and its quality as afood resource changewith landscape context (egvegetative cover) and catchment hydrology (77)but POM availability can also be limited by flowvariability which can increase decompositionrates and decrease residence times (78 79)As is the case for running-water systems in
general high flows in urban streams bring pulsesof solutes including N DOM and various pol-lutants (80) Solute dynamics and biogeochemicalprocesses in regulated rivers have been insuffi-ciently studied with researchers focusing moreon such processes in reservoirs rather than indownstream river ecosystems We found nostudies attempting to link urban or regulatedflow regimes and solute fluxes to changes infood webs or stream metabolic balance How-ever recent studies show shifts in microbial tax-onomic and functional composition in responseto pulsed urban flows (81) and thesemay help inexploring a potential link but are complicated bythe fact that non-flow stressors affect consumercomposition For example urban and regulatedrivers typically have animal communities thatare less diverse and composed of tolerant groupsoften non-native Hydropeaking and urban stormflows can certainly cause direct mortality via dis-placement and transport to unsuitable habitatsbut many of the organismal effects are realizedthrough indirect pathways For instance hydro-peaking reduces the viability of river-edge egg-laying specialists by limiting their access to suitablespawning habitat disrupting reproductive successand insect emergence (82) Flow regulation caneven create ecological traps if organismal phenol-ogy shiftswith thenovel environmental conditionsFor example dam-induced warming triggeredsummer diapause in a mayfly resulting in theloss of its last generation (83) Dams also isolatepopulations harming organisms that requiremigrations to complete their life cycles whetherthese are between riverine and marine habitatsor within the freshwater domain By analogy al-tered flow regimes in urban streams can also leadto isolated pools in streams with lowered base-flows This can be associated with periods ofhypoxia punctuated by extreme flow pulses inwhich these streams become ldquoscoured or suffo-catedrdquo (74)As illustrated using urban streams and reg-
ulated rivers research is moving forward andhints at the three-way nexus but is clearly still
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 5 of 13
Fig 4 Ecosystem metabolismpatterns across a river networkSpatial patterns in river eco-system metabolism for the Deva-Cares catchment in northernSpain as represented by the ratioof gross primary production(P) to ecosystem respiration (R)A PR gt 1 indicates autotrophicprocesses dominate so theseriver sections are accumulatingor exporting organic carbonPR lt 1 indicates dominanceby heterotrophic processes inriver sections that are receivingorganic C input (eg fromterrestrial sources) The boxesindicate regions of the networkimpacted by human pressures including deforestation (brown box) waste products from urbansites (green box) and a combination of the two (purple box) Such visualizations are usefulto managers in understanding regions of concern that may need the most restoration actions[Reproduced with permission from (157)]
PR ratiolt025
gt150
025 ndash 050 050 ndash 075075 ndash 100100 ndash 150
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
inadequate to meet restoration needs Studieson ecosystem processes below dams are parti-cularly limited and much of what we knowcomes from only a few river systems Furtherpublished work focuses primarily on benthicecosystem processes immediately below damsrather than riverine planktonic realms whereproductivity in large regulated rivers can beimportant
Linking flow biota andecosystem processes
Research on flow regimes that bridges organis-mal and ecosystem ecology has traditionallyrelied on states instead of rates ie nutrientconcentrations rather than uptake or fluxesstanding stocks of algae rather than productivityor metabolism and abundance of organismsrather than secondary productivity This workgenerally shows that flow variability suppressesand stability increases algal standing stock andgrazer densities with peaks in invertebrate bio-mass following those of algae [eg (84 85)]Over the past decade research began to focuson how flow regimes alter functional aspects offoodwebs For example biofilms inMediterranean-climate rivers are thicker andmetabolicallymoreactive downstream of flow-stabilizing dams (71)When flow variability increases below hydro-power dams fluctuations can reduce biofilmdevelopment and its quality as a food resource(86) Using experimental flumes subjected tohydropeaking shifts have been found in periph-yton fatty acid content from important highlyunsaturated compounds to nonessential saturatedones (87)Indirect evidence suggests that flow-induced
changes in dominant resource type (eg algaeversus detritus) resource quantity (biomass) orresource quality (nutrient content) can propa-gate to higher trophic levels via bottom-up ef-fects For instance detritivorous invertebrateswere found to dominate species richness andinvertebrate biomass upstream but not down-stream of an irrigation dam (75) diets of om-nivore insects indicated that flow stabilizationshifted a detrital into an algal-fueled food webSimilar evidence comes from Glen Canyon Damin the Colorado River (88) Near the dam whereprimary production is high (73) food webs aresimplified and gut content analyses have sug-gested algal production supports gt50 of in-vertebrate and gt70 of fish production andthe food web can be easily perturbed by flooddisturbance Farther downriver food webs showincreased reliance on detrital resources havehigher trophic efficiency and are more complexand more resistant to disturbance (76) In con-trast recent work on seasonal tropical riversfound algal productivity and zero-flow distur-bance did not explain food-web structure Inthis case it seems that highly mobile predatorsbuffered the local effects of drought (89) Thisstudy along with earlier ones at the site (90) isof particular interest because it links temporalpatterns in predatory fish with flow-dependentavailability of resources
Only a few studies have provided evidence oftop-down controls on foodwebs being influencedby flow variation Fish in a tropical floodplainriver were found to exert top-down control onprimary producers but only under certain hydro-logic conditions (91) Similarly in aCalifornian riverwinter flood pulses were shown to control insectconsumers late in the season ultimately deter-mining whether fish exerted top-down controlon the lower trophic levels (10) These illustratethe importance of pulsed hydrology on resource-consumer linkages and show that not just standingstocks but also the dominant controls structuringfood webs may fluctuate over time in response toflow variationAn indirect line of evidence supporting the
importance of the flow-biota-ecosystem processesnexus comes from theoretically oriented researchon food-web structure Changes in productivityhabitat size and disturbance underpin the basisof the three main hypotheses of food-chainlength which state that chains should be longerin stable productive large ecosystems (92) No-tably flow regimes influence these three con-trols Because food-chain length determines keyaspects of a river ecosystem such as the risk oftrophic cascades or the pollutant accumulationin top predators understanding how it is af-fected by flow alteration is important How-ever understanding the interplay between thehypotheses of food-chain length and the flow-dependent biological mechanisms that allowfood webs to shift in structure (eg predatorinvasions local extinctions consumers chang-ing diets) is still a relatively recent endeavor(93ndash95) Evidence suggests that zero-flows short-en food chains mainly by keeping streamsfishless (94 96) whereas stabilization of variableflows can lengthen food chains by allowing om-nivores to feed on abundant high trophic levelprey (95)New studies are also focusing on interactions
between drivers of food-web structure that hadbeen previously only been examined in isolationsuch as disturbance regimes and ecosystem sizeboth influenced by flow regime Using metabolictheory predator biomass has been shown toscale with both prey resources and fluctuatingstream habitat size with reductions in preda-tor size leading to declines in predator biomasssupported per unit of prey biomass (97) Movingforward the combination of habitat andmetabolicconstraints on predators may provide a basis toconnect flow-induced disturbance changes inecosystem processes and habitat fluctuationsto riverine food-web structure The metabolictheory of ecology (98) is a unifying frameworkthat allows scaling responses of organisms towhole ecosystems andmay become particularlyuseful in situations where flow alteration affectsorganismal body size or water temperature Insuch cases strong changes in metabolic ratescan be expected affecting resource uptake andorganismal growth to ecosystem-level produc-tion and respirationFinally an important body of food webndashrelated
research that provides strong inferential evidence
for the nexus comes from studies on animal-mediated nutrient cycling and ecological stoi-chiometry Theory suggests detrital and algalfood websmay be linked by cascading effects onnutrient cycling (99) and empirical work showsthat organismal excretion andmigration (whichrelocates C) may be important to this link (100)(Fig 5) A recent study (101) showed that ex-cretion of highly labile dissolved organic C ni-trogen (N) and phosphorus (P) by aquatic insectsmay be sufficient to support a substantial fractionof microbial energy and nutrient needs Giventhat C availability can be too low tomeetmicrobialenergy needs [eg (102 103)] and that microbialactivity enhances detrital quality to consumers(104) a C subsidy from excretory processes couldbe important at least during low-flow periodsAdditionally microbial use of excreted dissolvedorganic nitrogen (DON) and DOP may releasealgae from potential competition with microbesfor inorganic N and P if this is the case GPPshould increase Ecological stoichiometry mayalso be important for understanding constraintson organismal growth rates as rapid growthdisproportionally increases demand of P [as it isused in ribosomal RNA (105)] This could in turnlimit the ability of organisms to grow fast andexploit temporary habitats Interestingly elemen-tal ratios can vary interspecifically in response totemperature or predation pressure (106)mdashfactorsthat increase with low flows and habitat fragmen-tation Overall studies that link biota with bio-geochemical processes remain rare (107) butanalyzing flow regimes as a driver of elementturnover rates may provide a way forward
Can hydroecological advances informrestoration practice
Shortfalls in the outcomes of river restorationhave prompted calls to identify and restore pro-cesses that support and sustain biological com-munities rather than focusing only on rivergeomorphology and habitat In urban streamsscientists have critiqued an overemphasis onstructural engineered approaches (7 108) thatdo little to restore the full flow regime (109) Inlarge regulated rivers identifying and restoringimportant features of the flow regime via damoperations is often possible (albeit expensive)However that does not solve other problemsassociated with dams such as habitat frag-mentation (110) altered sediment and thermalregimes (111 112) or disrupted biogeochemicalprocesses (113) Overall efforts to alter reservoirreleases or improve urban stormwater infra-structure to partially mimic a riverrsquos natural flowregime have great potential Technologies arebeing evaluated for real-time management ofurban flows and novel ways to design flow re-leases from dams are being proposed (114 115)however research still lags far behind the needfor solutions Importantly the paucity of studieson the flow-biota-ecosystem processes nexus pointsto a need for more integrative research connectinghydrologists population and community ecologistsand ecosystem ecologists in the context ofrestoration
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 6 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
Restoring flow regimes ratherthan eventsScientifically informed flow designs may existthat maintain ecosystem integrity in regulatedrivers (116 117) but these have produced mixedresults to date In the dam-altered Putah Creekin California mimicking the natural timing ofhigh and low flows enhanced native fish by im-proving their spawning habitat reducing watertemperatures maintaining flowing conditionsand reducing the abundance of non-native spe-cies (118) However other similar attempts havebeen less successful In some cases non-nativefish density also benefitted from flow patternsmimicking an unregulated regime (119) in othersnon-natives bounced back shortly after the flowrelease (120) These experiences illustrate thateven natural flow features may have inadvertentnegative effects if only part of the community orecosystem is considered Models for the NavajoDam (San Juan River New Mexico) show thatadequately timed releases could simultaneouslyenhance native and minimize non-native fishpopulations (121) thus predicted benefits couldsurpass those of mimicking a natural flow re-gime potentially persisting even during dryyears Another study using the ldquodesignerrdquo flowparadigm (114) attempted to understand howdifferent facets of flow variability control fisheryproduction in the Lower Mekong floodplains Time-
series models combining discharge and fisherycatch suggested that enhanced flood pulses andlong inter-flood intervals may increase fisheryyieldsAlthough environmental flow practices have
mostly focused on re-creating discrete flowevents (eg flood pulses) these can still serve asvaluable experiments An iconic example of alarge-scale flow release is Minute 319 an amend-ment to the United StatesndashMexico treaty allow-ing opening of floodgates in the Lower ColoradoRiver and a one-time rewetting of the parchedColorado River Delta (122) Although this pulsed-flow experiment did not promote establishmentof native woody riparian seedlings such trialscan inform future restoration practices for ex-ample by determining whether the provision ofbare moist substrate enhances seed germina-tion (123) Hypothesis-driven experimental flowreleases combined with deep knowledge of tar-get ecosystems can provide insights into resto-ration actions [eg (124)] Still an outstandingquestion is how flow regime restoration couldbe leveraged to restore other important alteredaspects of the ecosystem such as sediment andthermal regimes For example cold-water releasesby existing dams in climate-vulnerable basinscould prevent organisms that thrive at low tem-peratures from becoming replaced by general-ist warm-tolerant taxa (125)
Although the concept of environmental flowsis generally associated with impounded riversits relevance to urban streams is clear (126)However because frequent high storm flowsare viewed as the problem reducing them gen-erally becomes the only restoration goal This isoften attempted by shifting the spatial distribu-tion of velocities within channels through ma-nipulation of channel sinuosity installation offlow-diverting weirs or adding other structuresThese approaches may reduce local erosion butrarely result in levels of water quality ecosystemprocesses or biodiversity in urban streams thatcome close to those of unimpacted streams(127 128) Focusing on the channel ratherthan the watershed context at large means thatkey ecosystemprocesses can remain compromised(129) A recent study compared time series ofmetabolism in two streamsmdashonewith stormwaterponds in the watershed and the other restoredby altering channel slope banks and in-streamstructural complexity (103) The study found thatpeaks in GPP and ER in the ldquorestoredrdquo streamwere higher more prone to resets induced byflashy storms and that recovery time scales werelonger compared with the stream in a watershedwith stormwater measures (Fig 6) Ponds areone type of nature-inspired green infrastructureused to restore streamflow tomore natural condi-tions tree plantings constructed wetlands andgrass swales can in turn replace gray infrastruc-ture such as concrete tunnels that capture andstore waterDistributing green infrastructure in water-
sheds helps restore streams because it targetsthe cause of streamflow alteration includinginsufficient infiltration groundwater rechargeand more generally limited watershed storagecapacity (129) By tackling the cause rather thanthe symptoms of flow alteration these practicescan reduce ecological impacts that result fromthe cascading effects of altered flows includingpoor water quality and temperature spikes Re-search on the effectiveness of green infrastructureon restoring streams is sparse and limited bylack of data before and after implementationalthough studies are beginning to appear Forexample upland green infrastructure in theform of shallow wetland-like pools that capturestormwater and reconnect it to the groundwaterhas been shown to enhance baseflows while re-ducing peak flows and pollutant loadsmore thanrestoration structures within the mainstem ofthe receiving stream (130) Similar structuresbuilt between stormwater outfalls and perennialstreams did not modulate the timing magnitudeor duration of urban storm responses nor didthey improve water quality or biodiversity (131)illustrating the need for researchers to understandhow exact design and watershed context influ-ence performance
Spatiotemporal variability andrestoration designs
Research showing how local and watershed-scale environmental drivers vary temporally andalong river networks in ways that shape biota
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 7 of 13
Flow pulses (storms) flashy flowsDroughts intermittencyLowstable flowsAltered timing seasonality
DOC N P
DetritusMicrobial
stoichiometric constraints
Excretion exudates
Detrital pathway
Algal pathway
Algal stoichiometric constraints
Organismal dispersal source-sink dynamics
Algae
Community replacement
ndash ndash +
ndash ndash + ndash
+
+ndash +
Invasions
Turnover
Consumer stoichiometric constraints
Fig 5 Flow variation affects biota and ecosystem processes and food webs integrate bothHydrologic connectivity along river networks is a first landscape-level filter constraining dispersal oforganisms in river networks Extreme flow variation (floods and droughts) are a source of disturbanceto all organisms (gray box) but particularly to the higher trophic levels given their higher metabolicdemands In turn flow variation controls inputs of nutrients and terrestrial leaf litter to the stream whichinfluences microbial activity and the relative contributions of the detrital (brown) and algal (green)pathways in sustaining aquatic consumers Not just quantity but also food quality (eg CN and CPratios) influence dietary choices of consumers and thus trophic transfer efficiency Finally alteration offlow regimes may influence all these compartments and may also open niches for new players Ifbiological invasions occur at the top of the food web these have the potential to divert energy away fromnative predators and to alter food-web structure through top-down controls
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
and ecosystem processes can help prioritizeselection of sites for restoration For exampleusing the concept of river network saturation(55) to identify river sections where in-streamremoval of nutrients cannot keep up with highsource inputs of nutrients suggests that restorationactions targeting those sections may dispropor-tionately improve water quality Identification ofsubcatchments that disproportionately influencestream chemistry may also be informed by com-bining methods from landscape ecology andcatchment hydrology focusing on the spatialscale of variance in water chemistry (132) Head-waters are often prioritized for restoration be-
cause poor water quality in them can havedownstream impacts Research showing thatlocal environmental constraints on river com-munities are important in headwaters as well asin the larger channels but dispersal constraintsare much less important in the former (133 134)implies that headwaters should be prioritizedHowever research quantifying time-varying con-tributions of local communities to river-wide(network) biodiversity can also inform prioritiza-tion by identifying sites that host a dispropor-tionate share of species or that host uniquespeciesmdashfor example because they shelter sensi-tive taxa during critical flow bottlenecks This idea
encapsulates the concept of keystone habitatsor local sites that strongly influence metacom-munity dynamics (135) This has been appliedto aquatic invertebrates across intermittent rivernetworks by tracking time-varying site-specificcontributions to community dissimilarity (or betadiversity) (136)Other approaches to prioritize restoration ac-
tions may be based on contributions to system-wide stability rather than numbers of individualsor species For example the restoration of fall-run Chinook salmon portfolios may be assessedvia the contributions that different periods andlocationsmake tometapopulation-wide stabilityowing to spatiotemporal variation in habitatconditions (137) Additionally interventionscould promote conditions that are not impor-tant now but could be important in the future[ie proto-refugia (138)] For example resto-ration of groundwater-influenced habitats couldmitigate future stress to drought-sensitive taxaunder scenarios of increased intermittencyPatches of undisturbed land that protect flowregimes in streams within a larger catchmentthat is otherwise urbanized can be sources ofcolonists to restored sites Priorities that focuson present degradation of river sectionsmay failto address site contribution to impairment orrecovery at broader scales or under differentscenarios These insights indicate a great poten-tial for new methods to prioritize restoration ofboth urban and regulated streamsWe know little about how altered flow regimes
propagate through entire river networks butwork is starting to assess the cumulative effectsof dams on flow regimes as well as dam-levelcontributions to alteration Although these ap-proaches have largely focused on dam cascades[eg (139)] if expanded they could help identifyfavorable sections in river networks to developenvironmental flow operations Here exper-imentation could again help with testing ofnew strategies For instance large-scale flowexperiments could be designed that coordinatereleases among dams or between dams and free-flowing tributaries to enhance the restorativeeffects of natural pulses (140)
Hydrologic connectivityinfluences restoration
Recovering critical flow paths and their connec-tivity to rivers is important but it remains asubstantial challenge for flow-degraded riversand streams Whereas regulated rivers tend tohave reduced lateral and longitudinal hydro-logic connectivity urban streams have unnaturallevels of connectivity because pipes and over-land runoff route water quickly In both cases fun-damental hydroecological insights into streambedstructure landscape storage capacity and ground-water connectivity can help to envision novel re-storation approaches (130 141 142) More generallyhydrologic connectivity is becoming a frameworkto identify alternative ways to restore streams viastormwater management (143) For example arecent study suggested that reducing road densityand hydrologic connectivity between roads
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 8 of 13
ER
Disc
harg
e (f
t3 s)
GPPG
PP o
r ER
(g O
2 m-2
day
-1)
Discharge
Watershed BMPs
In-stream restoration
0
ndash4
4
0
ndash4
4
4
2
6
4
2
6
Jan Sept July May Mar Nov Jan
A
B C
Fig 6 Green infrastructure as a means to restore stream ecosystems (A) Discharge andecosystem respiration (ER more-negative values mean higher respiration) and gross primaryproduction (GPP) for two urban streams in which restoration efforts involved (top) modifications inthe stream channel and (bottom) traditional stormwater best management practices (BMPs)[Reproduced with permission from (103)] (B and C) Example of a comprehensive greeninfrastructure approach to ecologically restore an urban headwater stream that also supports thelifestyle of resident urban communities (B) Stream at the onset of restoration and (C) afterrestoration [Reproduced with permission from Greenworks Portland OR]
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 9 of 13
Table 1 Research challenges and opportunities Despite the progress that has been made in understanding how flow regimes affect biota and ecosystem
processes major challenges persist that prevent a complete understanding of the flow-biota-ecosystem processes nexus Making progress on these
challenges requires a mix of fundamental and applied research
Topic Research challenge Opportunity
Connecting
organismal to
ecosystem-level
processes
Research on the effects of flow on organisms
and ecosystem processes has followed parallel
tracksmdashhindering our mechanistic
understanding of the flow-biota-ecosystem
processes nexus
Generate theory that connects concepts and frameworks
that are well accepted in river community and ecosystem
ecology For example the notion of environmental filters
that connect regional to local composition via a combination
of dispersal and environmental conditions (community
ecology) with analogous theory on solute transport versus
reactivity (ecosystem ecology)
Our current understanding of the links between
phenology of biota and ecosystem processes
is precursory
Use high-frequency sensors to characterize metabolic
regimes at sites with ongoing organismal monitoring
Leverage citizen-science biodiversity data to detect
spatiotemporal shifts in animal phenology
(eg insect emergence fish spawning)
Use remote sensing tools to better understand
links between hydrology and aquatic and riparian
plant phenology
The link between stoichiometry and flow
variation remains largely unexplored
Repurpose data that is already being collected to test
flow-stoichiometry relationships eg studies using C and
N stable isotopes to describe CN ratios across flow
regime types Perform field experiments to quantify
indirect effects of invertebrate and fish consumption
on ecosystem metabolism
Ecological
consequences of
hydrologic
connectivity
The importance of anaerobic metabolism in
carbon processing as flows cease and parts of
streambeds dry is a major frontier research area
Create networks of sites across different geologies
and flow regimes that collect time-series data on
methane nitrous oxide and carbon dioxide fluxes
from intermittent streams
We ignore how biota in the wet phase is
influenced by processes and patterns
from the dry phase
Research the functions of dry riverbeds Use camera traps and
drone technology to investigate the role of dry riverbeds
as a dispersal corridor metabarcoding to quantify
ldquodark biodiversityrdquo in the hyporheic
Wetlands and their links to stream networks are poorly
delineated globally Even in the United States and
Europe current wetland inventories do not capture
these links yet understanding how their connectivity
to river systems influences network-scale ecosystem
processes is needed
Analyze time series from high-frequency sensors distributed in
river networks varying in wetland connectivity Further
development and application of time-series methods
used by neuroscientists
We need to identify the relationship between
varying flows and solute dynamics
associated with changes in catchment-scale
hydrologic exchanges
Design collaborative work among catchment hydrologists
and stream ecologists combining methods to estimate
transit times residence times and identify sources
across heterogeneous landscapes Link these properties to
land management and water quality aspects
Scaling and
transferring flow-
ecology relationships
across space and time
We ignore how biodiversity responds to
propagation of flow regime alteration
across river networks
Leverage extensive hydrometric networks (eg United States
Geological Survey National Water Information System)
to perform spatially replicated time-series analyses
on flow and use these to generate expectations
on how river network position should mediate
biodiversity responses to flow alteration
Flow-ecology relationships do not account for
nonstationarity and abrupt change
Use process-based models that account for mechanisms
and associated uncertainties When using time-series
data and models select methods that are robust
to nonstationarity Linear flow-ecology relationships
should not be assumed
Current flow-ecology relationships assume that
responses of species do not change over time
Consider flow regime alteration as a process with ecological and
evolutionary consequences Include intraspecific variation
in traits and potential for rapid evolution in models
continued on next page
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
and streams may improve water quality morethan other restoration options (70) Additionallyresearch on the age and lability of DOC enteringstreams has shown how precipitation or land dis-turbance can change watershed flow paths (53)and this could inspire new restoration approachesfocused on land use and soil properties Manystudies on hydrologic connectivity and networkstructure suggest a need for restoration to focuson landscape heterogeneity especially on thepresence ofwetlands lakes and ponds (144) Suchfeaturesare frequently lost inurbanizedcatchmentsand outcomes could be improved by restoring orincreasing connectivitywith these ecosystems (145)Hydrologic connectivity associated with ground-water inputs or abstractions land-use manage-ment and return flows fromwastewater treatmentplants also need to be considered in these initia-tives (146)Restoring hydrologic connectivity is also key
to achieving biodiversity outcomesmdashparticularlyin dammed systems Longitudinal connectivityfor fish inhabiting dam-altered rivers can beenhanced by the construction of fishways andsophisticated passage facilities However moni-toring data on the effectiveness of such inter-
ventions are still scarce and available evidencesuggests that passage effectivity largely dependson technology used and species-specific fish be-havior (147 148) Although trap-and-transportprograms for migratory fishes can help augmentthreatened populations (149) these practicesoften increase straying and thus the potentialfor genetic homogenization as seen from ju-venile salmon collected at dams and transporteddownstream to increase out-migration success(150) Overall a combination of engineering andmanagement solutions may be best to restorefunctional connectivity in dam-altered riversThe link between hydrologic connectivity andecological restoration is also central to con-siderations on where to build remove and re-operate hydrologic infrastructure Hundredsof dams have been intentionally removed sincethe 1970s and removal of large dams such asthe Elwha River Dam (Washington USA) hasshown that restoring longitudinal connectivitycan have immediate geomorphic effects with com-plex temporal dynamics (151) Infrastructure canalso be designed or reoperated to manage forrestoring lateral connectivity and enhanced flood-plain processes For example the Yolo Bypass in
the Sacramento River California has becomea prime rearing and migration site for chinooksalmon showing that flood control and habitatfor fish and wildlife do not need to be at odds(152) All these examples show the importanceof integrating hydrologic connectivity in ecolog-ical restoration whether the goal is to minimizethe ecological effects of fragmentation restorefree-flowing watercourses or operate green infra-structure to enhance infiltration floodplain con-nectivity or other ecological processes
Research challenges and opportunities
Two decades of concerted efforts to research theeffects of flow regime alteration have tremen-dously advanced our understanding of the waysin which streamflow influences biota and eco-system processes However important challengesremain These aremainly related to the scarcity ofscience connecting organismal to ecosystem-levelprocesses through for example microbial linksthe difficulty of studying watershed-level phys-ical and biogeochemical heterogeneity and itsinfluence on riverine processes and biota thecomplexity of parsing out the direct effects offlow from flow-mediated effects and co-occurring
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 10 of 13
Topic Research challenge Opportunity
Cascading
implications of
hydrologic
alteration
We have a poor understanding of the
interactions among physical drivers
Build on large-scale flow experiments to test hypotheses on the
combined effects of altered flow sediment and thermal
regimes Experiment with multilevel intakes in reservoirs
Promote collection of non-flow variables (eg temperature
water quality parameters) at hydrometric stations
At low flows biota are being exposed to high
concentrations of new classes of pollutants
whose impacts are poorly known especially if
biotic impacts interact with impacts on
ecosystem processes
Design realistic experimental studies to advance knowledge
on the ecological effects of multiple stressors
Collect long-term data at sites varying in contaminant
inputs and hydrologic regimes Link microbial to
organismal and ecosystem-level responses
Using hydroecological
science to inform
river ecosystem
restoration
Environmental flow science largely ignores
human dimensions
Use methods that allow identifying causal networks
in complex socioenvironmental systems Include
human needs and thresholds in research on
flow-ecology relationships
Environmental flows are generally prescribed
at the scale of single infrastructures restoration
designs and assessment are generally at reach scales
Promote coordinated large-scale flow experiments
and restoration designs at the river basin scale
Extend assessment of green infrastructure
effectiveness beyond individual structures to
infrastructure networks and link both
to river outcomes
Environmental flows in impounded rivers
generally target the conservation of some
particular species rather than outcomes at the
food-web or ecosystem level
Advance fundamental research on microbialndash
ecosystem process linkages Expand the notion
of designer flows to food-web structure and
ecosystem services when possible
Environmental flow science has largely
ignored surface waterndashgroundwater
interactions
Expand science to understand the link between
restoration actions groundwater recharge and
inputs to streams Provide science relevant to
the implementation of new legislation on
groundwater management (eg Sustainable
Groundwater Management Act in California)
Uncertainty regarding future
hydroclimate and
river flows is growing
Refine regional climate models Promote research on
the interaction between land-use change and
evapotranspiration Incorporate groundwater
influence in macroscale hydrology models
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
stressors (particularly in urban streams) and thechallenge of predicting outcomeswhen both thephysical and the biological environments arechangingAchieving a predictive understanding of hy-
droecological relationships is essential to design-ing successful self-sustaining restoration actionsin river ecosystems However translating basichydroecological insights into restoration practicerequires understanding the complexities of work-ing within a human-dominated water cycle Al-though it is rarely possible to restore all facetsof a natural flow regime ecologically informeddesigns may help mimic key aspects of the nat-ural flow regime (eg periodic pulse flows) orcreate regimes that are not natural but maximizeoutcomes given socioeconomic constraints InTable 1 we identify persisting and emergingchallenges in hydroecological research We paireach challenge with an opportunity (related toresearch design technology or implementation)We contend that these challenges should beprioritized to advance process-based river eco-system restoration
REFERENCES AND NOTES
1 N L Poff et al The natural flow regime Bioscience 47769ndash784 (1997) doi 1023071313099
2 D A Lytle N L Poff Adaptation to natural flow regimesTrends Ecol Evol 19 94ndash100 (2004) doi 101016jtree200310002 pmid 16701235
3 S Sabater A Elosegi R Ludwig in Multiple Stressors in RiverEcosystems (Elsevier 2019) pp 1ndash22
4 United Nations New UN Decade on Ecosystem Restorationoffers unparalleled opportunity for job creation food securityand addressing climate change (2019) httpswwwunenvironmentorgnews-and-storiespress-releasenew-un-decade-ecosystem-restoration-offers-unparalleled-opportunity
5 A W Lorenz P Haase K Januschke A SundermannD Hering Revisiting restored river reaches - Assessingchange of aquatic and riparian communities afterfive years Sci Total Environ 613-614 1185ndash1195(2018) doi 101016jscitotenv201709188pmid 28954379
6 M A Palmer K L Hondula B J Koch Ecological Restorationof Streams and Rivers Shifting Strategies and ShiftingGoals (2014) vol 45
7 T J Beechie et al Process-based principles for restoringriver ecosystems Bioscience 60 209ndash222 (2010)doi 101525bio20106037
8 S E Bunn A H Arthington Basic principles and ecologicalconsequences of altered flow regimes for aquaticbiodiversity Environ Manage 30 492ndash507 (2002)doi 101007s00267-002-2737-0 pmid 12481916
9 K D Fausch Y Taniguchi S Nakano G D GrossmanC R Townsend Flood disturbance regimes influence rainbowtrout invasion success among five holarctic regions EcolAppl 11 1438ndash1455 (2001) doi 1018901051-0761(2001)011[1438FDRIRT]20CO2
10 M E Power M S Parker W E Dietrich Seasonalreassembly of a river food web Floods droughts andimpacts of fish Ecol Monogr 78 263ndash282 (2008)doi 10189006-09021
11 V Acuntildea A Giorgi I Muntildeoz U R S Uehlinger S SabaterFlow extremes and benthic organic matter shape themetabolism of a headwater Mediterranean streamFreshwater Biol 49 960ndash971 (2004) doi 101111j1365-2427200401239x
12 K Tockner F Malard J V Ward An extension of the floodpulse concept Hydrol Processes 14 2861ndash2883 (2000)doi 1010021099-1085(20001112)141617lt2861AID-HYP124gt30CO2-F
13 J S Perkin et al Fragmentation and dewatering transformGreat Plains stream fish communities Ecol Monogr 8573ndash92 (2015) doi 10189014-01211
14 J L Sabo D M Post Quantifying periodic stochastic andcatastrophic environmental variation Ecol Monogr 7819ndash40 (2008) doi 10189006-13401
15 F Shuai et al Silver carp larva abundance in response toriver flow rate revealed by cross-wavelet modellingEcol Modell 383 98ndash105 (2018) doi 101016jecolmodel201805020
16 J D Tonkin M T Bogan N Bonada B Rios-ToumaD A Lytle Seasonality and predictability shape temporalspecies diversity Ecology 98 1201ndash1216 (2017)doi 101002ecy1761 pmid 28144975
17 N L Poff Beyond the natural flow regime Broadening thehydro‐ecological foundation to meet environmental flowschallenges in a non‐stationary world Freshwater Biol 631011ndash1021 (2018) doi 101111fwb13038
18 A Ruhi X Dong C H McDaniel D P Batzer J L SaboDetrimental effects of a novel flow regime on the functionaltrajectory of an aquatic invertebrate metacommunity GlobChange Biol 24 3749ndash3765 (2018) doi 101111gcb14133pmid 29665147
19 M T Bogan D A Lytle Severe drought drives novelcommunity trajectories in desert stream pools FreshwaterBiol 56 2070ndash2081 (2011) doi 101111j1365-2427201102638x
20 G Woodward et al The effects of climatic fluctuations andextreme events on running water ecosystems Philos TransR Soc London Ser B 371 20150274 (2016) pmid 27114576
21 M A Palmer C M Swan K Nelson P Silver R AlvestadStreambed landscapes Evidence that stream invertebratesrespond to the type and spatial arrangement of patchesLandsc Ecol 15 563ndash576 (2000) doi 101023A1008194130695
22 D Schmera et al Does isolation influence the relative role ofenvironmental and dispersal‐related processes in streamnetworks An empirical test of the network positionhypothesis using multiple taxa Freshwater Biol 63 74ndash85(2018) doi 101111fwb12973
23 R Henriques‐Silva et al A comprehensive examination of thenetwork position hypothesis across multiple rivermetacommunities Ecography 42 284ndash294 (2019)doi 101111ecog03908
24 K L Jaeger J D Olden N A Pelland Climate change poisedto threaten hydrologic connectivity and endemic fishes indryland streams Proc Natl Acad Sci USA 11113894ndash13899 (2014) doi 101073pnas1320890111pmid 25136090
25 S Garbin E A Celegon P Fanton G Botter Hydrologicalcontrols on river network connectivity R Soc Open Sci 6181428 (2019) doi 101098rsos181428 pmid 30891270
26 F Cattaneacuteo B Hugueny N Lamouroux Synchrony in browntrout Salmo trutta population dynamics A lsquoMoran effectrsquo onearly‐life stages Oikos 100 43ndash54 (2003) doi 101034j1600-0706200311912x
27 A Terui et al Metapopulation stability in branching rivernetworks Proc Natl Acad Sci USA 115 E5963ndashE5969(2018) doi 101073pnas1800060115 pmid 29895695
28 P A P Moran The statistical analysis of the Canadian Lynxcycle Aust J Zool 1 291ndash298 (1953) doi 101071ZO9530291
29 G D Grossman P B Moyle J O Whitaker Jr Stochasticityin structural and functional characteristics of an Indianastream fish assemblage A test of community theoryAm Nat 120 423ndash454 (1982) doi 101086284004
30 D A Lytle D M Merritt J D Tonkin J D OldenL V Reynolds Linking river flow regimes to riparian plantguilds A community-wide modeling approach Ecol Appl 271338ndash1350 (2017) doi 101002eap1528 pmid 28263426
31 A Ruhiacute E E Holmes J N Rinne J L Sabo Anomalousdroughts not invasion decrease persistence of native fishesin a desert river Glob Change Biol 21 1482ndash1496 (2015)doi 101111gcb12780 pmid 25430731
32 J D Tonkin D M Merritt J D Olden L V ReynoldsD A Lytle Flow regime alteration degrades ecologicalnetworks in riparian ecosystems Nat Ecol Evol 2 86ndash93(2018) doi 101038s41559-017-0379-0 pmid 29180707
33 M C Mims J D Olden Fish assemblages respond toaltered flow regimes via ecological filtering of lifehistory strategies Freshwater Biol 58 50ndash62 (2013)doi 101111fwb12037
34 P T J Johnson J D Olden M J Vander Zanden Daminvaders Impoundments facilitate biological invasions intofreshwaters Front Ecol Environ 6 357ndash363 (2008)doi 101890070156
35 N Bonada S Doledec B Statzner Taxonomic and biologicaltrait differences of stream macroinvertebrate communitiesbetween mediterranean and temperate regions Implicationsfor future climatic scenarios Glob Change Biol 131658ndash1671 (2007) doi 101111j1365-2486200701375x
36 M I Pyne N L Poff Vulnerability of stream communitycomposition and function to projected thermal warming andhydrologic change across ecoregions in the western UnitedStates Glob Change Biol 23 77ndash93 (2017) doi 101111gcb13437 pmid 27429092
37 D B Herbst S D Cooper R B Medhurst S W WisemanC T Hunsaker Drought ecohydrology alters the structureand function of benthic invertebrate communities inmountain streams Freshwater Biol 64 886ndash902 (2019)doi 101111fwb13270
38 M T Bogan E T Chester T Datry A L MurphyB J Robson A Ruhi R Stubbington J E Whitney inResistance Resilience and Community Recovery in IntermittentRivers and Ephemeral Streams T Datry N Bonada A BoultonEds (Academic Press 2017) pp 349ndash376
39 S Houmlckendorff et al Characterizing fish responses to a riverrestoration over 21 years based on speciesrsquo traits ConservBiol 31 1098ndash1108 (2017) doi 101111cobi12908pmid 28218807
40 D Bruno et al Structural and functional responses ofinvertebrate communities to climate change andflow regulation in alpine catchments Glob Change Biol25 1612ndash1628 (2019) doi 101111gcb14581pmid 30698905
41 T W H Aspin et al Extreme drought pushes streaminvertebrate communities over functional thresholds GlobChange Biol 25 230ndash244 (2019) doi 101111gcb14495pmid 30346098
42 S E Tank J B Fellman E Hood E S Kritzberg Beyondrespiration Controls on lateral carbon fluxes across theterrestrial-aquatic interface Limnol Oceanogr Lett 3 76ndash88(2018) doi 101002lol210065
43 A P Appling et al The metabolic regimes of 356 rivers inthe United States Sci Data 5 180292 (2018) doi 101038sdata2018292 pmid 30532078
44 P Savoy et al Metabolic rhythms in flowing watersAn approach for classifying river productivity regimesLimnol Oceanogr (2019) doi 101002lno11154
45 E S Bernhardt et al The metabolic regimes of flowingwaters Limnol Oceanogr 63 S99ndashS118 (2018)doi 101002lno10726
46 M Colls X Timoner S Sabater V Acuntildea Effects of durationand frequency of the non-flow period on stream metabolismEcosystems 101007s10021-019-00345-1 (2019)doi 101007s10021-019-00345-1
47 A J Ulseth E Bertuzzo G A Singer J Schelker T J BattinClimate-induced changes in spring snowmelt impactecosystem metabolism and carbon fluxes in an Alpine streamnetwork Ecosystems 21 373ndash390 (2018) doi 101007s10021-017-0155-7
48 B O L Demars Hydrological pulses and burning of dissolvedorganic carbon by stream respiration Limnol Oceanogr 64406ndash421 (2019) doi 101002lno11048
49 R T Hensley L Kirk M Spangler M N Gooseff M J CohenFlow extremes as spatiotemporal control points on riversolute fluxes and metabolism J Geophys Res Biogeosci124 537ndash555 (2019) doi 1010292018JG004738
50 J D Hosen et al Enhancement of primary production duringdrought in a temperate watershed is greater in larger riversthan headwater streams Limnol Oceanogr 64 1458ndash1472(2019) doi 101002lno11127
51 L Goacutemez-Gener et al When water vanishes Magnitude andregulation of carbon dioxide emissions from dry temporarystreams Ecosystems 19 710ndash723 (2016) doi 101007s10021-016-9963-4
52 E H Stanley et al The ecology of methane in streams andrivers Patterns controls and global significance EcolMonogr 86 146ndash171 (2016) doi 10189015-1027
53 R T Barnes D E Butman H F Wilson P A RaymondRiverine export of aged carbon driven by flow path depth andresidence time Environ Sci Technol 52 1028ndash1035 (2018)doi 101021acsest7b04717 pmid 29313674
54 G Pinay et al Riparian corridors A new conceptualframework for assessing nitrogen buffering across biomesFront Environ Sci 6 47 (2018) doi 103389fenvs201800047
55 W M Wollheim et al River network saturation conceptFactors influencing the balance of biogeochemical supply and
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 11 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
demand of river networks Biogeochemistry 141 503ndash521(2018) doi 101007s10533-018-0488-0
56 P A Raymond J E Saiers W V Sobczak Hydrological andbiogeochemical controls on watershed dissolved organicmatter transport Pulse-shunt concept Ecology 97 5ndash16(2016) doi 10189014-16841 pmid 27008769
57 C Tunaley D Tetzlaff J Lessels C Soulsby Linking high-frequency DOC dynamics to the age of connected watersources Water Resour Res 52 5232ndash5247 (2016)doi 1010022015WR018419
58 S Bernal et al Supply demand and in-stream retention ofdissolved organic carbon and nitrate during storms inMediterranean forested headwater streams Front EnvironSci 7 60 (2019) doi 103389fenvs201900060
59 J Harvey et al How hydrologic connectivity regulates waterquality in river corridors J Am Water Resour Assoc 55369ndash381 (2019) doi 1011111752-168812691
60 J C Moore et al Detritus trophic dynamics and biodiversityEcol Lett 7 584ndash600 (2004) doi 101111j1461-0248200400606x
61 A D Rosemond et al Freshwater ecology Experimentalnutrient additions accelerate terrestrial carbon loss fromstream ecosystems Science 347 1142ndash1145 (2015)doi 101126scienceaaa1958 pmid 25745171
62 A C Y Yeung J L Musetta-Lambert D P KreutzweiserP K Sibley J S Richardson Relations of interannualdifferences in stream litter breakdown with dischargeBioassessment implications Ecosphere 9 e02423 (2018)doi 101002ecs22423
63 R M Northington J R Webster Experimental reductions instream flow alter litter processing and consumer subsidies inheadwater streams Freshwater Biol 62 737ndash750 (2017)doi 101111fwb12898
64 E Mas-Martiacute I Sanpera-Calbet I Muntildeoz Bottom-up effectsof streambed drying on consumer performance throughchanges in resource quality Aquat Sci 79 719ndash731 (2017)doi 101007s00027-017-0531-6
65 B D Richter G A Thomas Restoring environmental flows bymodifying dam operations Ecol Soc 12 12 (2007)doi 105751ES-02014-120112
66 G Zolezzi A Siviglia M Toffolon B MaioliniThermopeaking in Alpine streams Event characterizationand time scales Ecohydrology 4 564ndash576 (2011)doi 101002eco132
67 T Meiszligner B Sures C K Feld Multiple stressors and therole of hydrology on benthic invertebrates in mountainousstreams Sci Total Environ 663 841ndash851 (2019)doi 101016jscitotenv201901288 pmid 30738264
68 I R Waite et al Effects of urban multi-stressors on threestream biotic assemblages Sci Total Environ 6601472ndash1485 (2019) doi 101016jscitotenv201812240pmid 30743940
69 K C Nelson M A Palmer Stream temperature surges underurbanization and climate change Data models andresponses J Am Water Resour Assoc 43 440ndash452 (2007)doi 101111j1752-1688200700034x
70 J R Blaszczak J M Delesantro Y Zhong D L UrbanE S Bernhardt Watershed urban development controls onurban streamwater chemistry variability Biogeochemistry144 61ndash84 (2019) doi 101007s10533-019-00572-7
71 L Ponsatiacute et al Biofilm responses to flow regulation by damsin Mediterranean rivers River Res Appl 31 1003ndash1016(2015) doi 101002rra2807
72 J M Alberts J J Beaulieu I Buffam Watershed land useand seasonal variation constrain the influence of ripariancanopy cover on stream ecosystem metabolism Ecosystems20 553ndash567 (2017) doi 101007s10021-016-0040-9pmid 30416371
73 R O Hall Jr et al Turbidity light temperature andhydropeaking control primary productivity in the ColoradoRiver Grand Canyon Limnol Oceanogr 60 512ndash526 (2015)doi 101002lno10031
74 J R Blaszczak J M Delesantro D L Urban M W DoyleE S Bernhardt Scoured or suffocated Urban streamecosystems oscillate between hydrologic and dissolvedoxygen extremes Limnol Oceanogr 64 877ndash894 (2019)doi 101002lno11081
75 J-R Mor et al Dam regulation and riverine food-webstructure in a Mediterranean river Sci Total Environ 625301ndash310 (2018) doi 101016jscitotenv201712296pmid 29289778
76 H A Wellard Kelly et al Macroinvertebrate diets reflecttributary inputs and turbidity-driven changes in food
availability in the Colorado River downstream of GlenCanyon Dam Freshw Sci 32 397ndash410 (2013)doi 10189912-0881
77 S J Imberger R M Thompson M R Grace Urbancatchment hydrology overwhelms reach scale effects ofriparian vegetation on organic matter dynamics FreshwaterBiol 56 1370ndash1389 (2011) doi 101111j1365-2427201102575x
78 R T Martins A S Melo J F Gonccedilalves Jr N HamadaLeaf-litter breakdown in urban streams of Central AmazoniaDirect and indirect effects of physical chemical andbiological factors Freshw Sci 34 716ndash726 (2015)doi 101086681086
79 C M Yule J Y Gan T Jinggut K V Lee Urbanizationaffects food webs and leaf-litter decomposition in a tropicalstream in Malaysia Freshw Sci 34 702ndash715 (2015)doi 101086681252
80 T A Newcomer et al Influence of natural and novel organiccarbon sources on denitrification in forest degraded urbanand restored streams Ecol Monogr 82 449ndash466 (2012)doi 10189012-04581
81 A Chaudhary I Kauser A Ray R Poretsky Taxon-drivenfunctional shifts associated with storm flow in an urbanstream microbial community MSphere 3 e00194-18 (2018)doi 101128mSphere00194-18 pmid 29976643
82 T A Kennedy et al Flow management for hydropowerextirpates aquatic insects undermining river food websBioscience 66 561ndash575 (2016) doi 101093bioscibiw059
83 A Glazaczow D Orwin M Bogdziewicz Increasedtemperature delays the late-season phenology of multivoltineinsect Sci Rep 6 38022 (2016) doi 101038srep38022pmid 27905493
84 S G Fisher L J Gray N B Grimm D E Busch Temporalsuccession in a desert stream ecosystem following flashflooding Ecol Monogr 52 93ndash110 (1982) doi 1023072937346
85 G J Scrimgeour M J Winterbourn Effects of floods onepilithon and benthic macroinvertebrate populations in anunstable New Zealand river Hydrobiologia 171 33ndash44 (1989)doi 101007BF00005722
86 E Bondar-Kunze S Maier D Schoumlnauer N Bahl T HeinAntagonistic and synergistic effects on a stream periphytoncommunity under the influence of pulsed flow velocityincrease and nutrient enrichment Sci Total Environ 573594ndash602 (2016) doi 101016jscitotenv201608158pmid 27585428
87 M J Cashman G L Harvey G Wharton M C Bruno Woodmitigates the effect of hydropeaking scour on periphytonbiomass and nutritional quality in semi-natural flumesimulations Aquat Sci 79 459ndash471 (2017) doi 101007s00027-016-0510-3
88 W F Cross et al Food-web dynamics in a large riverdiscontinuum Ecol Monogr 83 311ndash337 (2013)doi 10189012-17271
89 D M Warfe et al Productivity disturbance and ecosystemsize have no influence on food chain length in seasonallyconnected rivers PLOS ONE 8 e66240 (2013)doi 101371journalpone0066240
90 T D Jardine et al Consumer-resource coupling in wet-drytropical rivers J Anim Ecol 81 310ndash322 (2012)doi 101111j1365-2656201101925x pmid 22103689
91 K O Winemiller et al Pulsing hydrology determines top‐down control of basal resources in a tropical riverndashfloodplainecosystem Ecol Monogr 84 621ndash635 (2014)doi 10189013-18221
92 D M Post The long and short of food-chain length TrendsEcol Evol 17 269ndash277 (2002) doi 101016S0169-5347(02)02455-2
93 D M Post G Takimoto Proximate structural mechanismsfor variation in food‐chain length Oikos 116 775ndash782 (2007)doi 101111j0030-1299200715552x
94 P A McHugh A R McIntosh P G Jellyman Dual influencesof ecosystem size and disturbance on food chain length instreams Ecol Lett 13 881ndash890 (2010) doi 101111j1461-0248201001484x pmid 20482579
95 A Ruhiacute et al Flow regulation increases food-chain lengththrough omnivory mechanisms in a Mediterranean rivernetwork Freshwater Biol 61 1536ndash1549 (2016) doi 101111fwb12794
96 J L Sabo J C Finlay T Kennedy D M Post The role ofdischarge variation in scaling of drainage area and food chainlength in rivers Science 330 965ndash967 (2010) doi 101126science1196005 pmid 20947729
97 A R McIntosh et al Capacity to support predators scaleswith habitat size Sci Adv 4 eaap7523 (2018) doi 101126sciadvaap7523 pmid 29978038
98 J H Brown J F Gillooly A P Allen V M Savage G B WestToward a metabolic theory of ecology Ecology 851771ndash1789 (2004) doi 10189003-9000
99 K Zou E Theacutebault G Lacroix S Barot Interactions betweenthe green and brown food web determine ecosystemfunctioning Funct Ecol 30 1454ndash1465 (2016) doi 1011111365-243512626
100 M J Vanni P B McIntyre Predicting nutrient excretion ofaquatic animals with metabolic ecology and ecologicalstoichiometry A global synthesis Ecology 97 3460ndash3471(2016) doi 101002ecy1582 pmid 27912023
101 T B Parr K A Capps S P Inamdar K A Metcalf Animal-mediated organic matter transformation Aquatic insects as asource of microbially bioavailable organic nutrients andenergy Funct Ecol 33 524ndash535 (2019) doi 1011111365-243513242
102 C P Arango et al Urban infrastructure influences dissolvedorganic matter quality and bacterial metabolism in an urbanstream network Freshwater Biol 62 1917ndash1928 (2017)
103 L G Larsen J W Harvey Disrupted carbon cycling inrestored and unrestored urban streams Critical timescalesand controls Limnol Oceanogr 62 S160ndashS182 (2017)doi 101002lno10613
104 H M Halvorson G White J T Scott M A Evans-WhiteDietary and taxonomic controls on incorporation of microbialcarbon and phosphorus by detritivorous caddisfliesOecologia 180 567ndash579 (2016) doi 101007s00442-015-3464-6 pmid 26497125
105 J J Elser et al Biological stoichiometry from genes toecosystems Ecol Lett 3 540ndash550 (2000) doi 101046j1461-0248200000185x
106 E K Moody et al Consumption explains intraspecificvariation in nutrient recycling stoichiometry in a desert fishEcology 99 1552ndash1561 (2018) doi 101002ecy2372pmid 29882955
107 N Welti et al Bridging food webs ecosystem metabolismand biogeochemistry using ecological stoichiometry theoryFront Microbiol 8 1298 (2017) doi 103389fmicb201701298 pmid 28747904
108 M A Palmer S Filoso R M Fanelli From ecosystems toecosystem services Stream restoration as ecologicalengineering Ecol Eng 65 62ndash70 (2014) doi 101016jecoleng201307059
109 M J Burns T D Fletcher C J Walsh A R LadsonB E Hatt Hydrologic shortcomings of conventional urbanstormwater management and opportunities for reformLandsc Urban Plan 105 230ndash240 (2012) doi 101016jlandurbplan201112012
110 C R Liermann C Nilsson J Robertson R Y NgImplications of dam obstruction for global freshwater fishdiversity Bioscience 62 539ndash548 (2012) doi 101525bio20126265
111 J D Olden R J Naiman Incorporating thermal regimes intoenvironmental flows assessments Modifying dam operationsto restore freshwater ecosystem integrity Freshwater Biol55 86ndash107 (2010) doi 101111j1365-2427200902179x
112 D B Osmundson R J Ryel V L Lamarra J Pitlick Flowndashsedimentndashbiota relations Implications for river regulationeffects on native fish abundance Ecol Appl 12 1719ndash1739(2002)
113 D von Schiller et al Regulation causes nitrogen cyclingdiscontinuities in Mediterranean rivers Sci Total Environ540 168ndash177 (2016) doi 101016jscitotenv201507017pmid 26233271
114 J L Sabo et al Designing river flows to improve foodsecurity futures in the Lower Mekong Basin Science 358eaao1053 (2017) doi 101126scienceaao1053pmid 29217541
115 W Xu et al Improving the multi-objective performance ofrainwater harvesting systems using real-time controltechnology Water 10 147 (2018) doi 103390w10020147
116 M Acreman et al Environmental flows for naturalhybrid and novel riverine ecosystems in a changingworld Front Ecol Environ 12 466ndash473 (2014)doi 101890130134
117 N L Poff et al Sustainable water management under futureuncertainty with eco-engineering decision scaling Nat ClimChang 6 25ndash34 (2016) doi 101038nclimate2765
118 J D Kiernan P B Moyle P K Crain Restoring native fishassemblages to a regulated California stream using the
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 12 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
natural flow regime concept Ecol Appl 22 1472ndash1482(2012) doi 10189011-04801 pmid 22908707
119 D L Propst K B Gido Responses of native andnonnative fishes to natural flow regime mimicry in theSan Juan River Trans Am Fish Soc 133 922ndash931 (2004)doi 101577T03-0571
120 T K Pool J D Olden Assessing long‐term fish responses andshort‐term solutions to flow regulation in a dryland river basinEcol Freshwat Fish 24 56ndash66 (2015) doi 101111eff12125
121 W Chen J D Olden Designing flows to resolve human andenvironmental water needs in a dam-regulated riverNat Commun 8 2158 (2017) doi 101038s41467-017-02226-4 pmid 29255194
122 S M Nelson et al A history of the 2014 Minute 319environmental pulse flow as documented by fieldmeasurements and satellite imagery Ecol Eng 106733ndash748 (2017) doi 101016jecoleng201610040
123 P B Shafroth et al A large-scale environmental flowexperiment for riparian restoration in the Colorado RiverDelta Ecol Eng 106 645ndash660 (2017) doi 101016jecoleng201702016
124 E P Glenn P L Nagler P B Shafroth C J JarchowEffectiveness of environmental flows for riparian restorationin arid regions A tale of four rivers Ecol Eng 106 695ndash703(2017) doi 101016jecoleng201701009
125 S Hotaling D S Finn J Joseph Giersch D W WeisrockD Jacobsen Climate change and alpine stream biologyProgress challenges and opportunities for the futureBiol Rev Camb Philos Soc 92 2024ndash2045 (2017)doi 101111brv12319 pmid 28105701
126 C J Walsh T D Fletcher M J Burns Urban stormwaterrunoff A new class of environmental flow problem PLOS ONE7 e45814 (2012) doi 101371journalpone0045814pmid 23029257
127 A Selvakumar T P OrsquoConnor S D Struck Role of streamrestoration on improving benthic macroinvertebrates and in-stream water quality in an urban watershed Case studyJ Environ Eng 136 127ndash139 (2010) doi 101061(ASCE)EE1943-78700000116
128 R W Lammers B P Bledsoe What role does streamrestoration play in nutrient management Crit Rev EnvironSci Technol 47 335ndash371 (2017) doi 1010801064338920171318618
129 H E Golden N Hoghooghi Green infrastructure and itscatchment-scale effects An emerging science WIREs Water5 e1254 (2018) doi 101002wat21254 pmid 29682288
130 M R Williams G Bhatt S Filoso G Yactayo Streamrestoration performance and its contribution to theChesapeake Bay TMDL Challenges posed by climate changein urban areas Estuaries Coasts 40 1227ndash1246 (2017)doi 101007s12237-017-0226-1
131 R M Fanelli K Prestegaard M A Palmer Freshw Sci101086706072 (2019)
132 B W Abbott et al Unexpected spatial stability of waterchemistry in headwater stream networks Ecol Lett 21296ndash308 (2018) doi 101111ele12897 pmid 29282860
133 B L Brown C M Swan Dendritic network structureconstrains metacommunity properties in riverineecosystems J Anim Ecol 79 571ndash580 (2010) doi 101111j1365-2656201001668x pmid 20180874
134 C M Swan B L Brown Metacommunity theory meetsrestoration Isolation may mediate how ecologicalcommunities respond to stream restoration Ecol Appl 272209ndash2219 (2017) doi 101002eap1602 pmid 28718193
135 N Mouquet D Gravel F Massol V Calcagno Extending theconcept of keystone species to communities and ecosystemsEcol Lett 16 1ndash8 (2013) doi 101111ele12014 pmid 23062191
136 A Ruhiacute T Datry J L Sabo Interpreting beta-diversitycomponents over time to conserve metacommunities inhighly dynamic ecosystems Conserv Biol 31 1459ndash1468(2017) doi 101111cobi12906 pmid 28188969
137 L Yamane L W Botsford D P Kilduff Tracking restorationof population diversity via the portfolio effect J Appl Ecol55 472ndash481 (2018) doi 1011111365-266412978
138 D W Braidwood M A Taggart M Smith R AndersenTranslocations conservation and climate change Use ofrestoration sites as protorefuges and protorefugia RestorEcol 26 20ndash28 (2018) doi 101111rec12642
139 Y Wang D Wang Q W Lewis J Wu F Huang A frameworkto assess the cumulative impacts of dams on hydrologicalregime A case study of the Yangtze River Hydrol Processes31 3045ndash3055 (2017) doi 101002hyp11239
140 J L Sabo et al Pulsed flows tributary inputs and food-webstructure in a highly regulated river J Appl Ecol 551884ndash1895 (2018) doi 1011111365-266413109
141 C Magliozzi G Coro R C Grabowski A I PackmanS Krause A multiscale statistical method to identifypotential areas of hyporheic exchange for river restorationplanning Environ Model Softw 111 311ndash323 (2019)doi 101016jenvsoft201809006
142 S S Schwartz B Smith Restoring hydrologic function inurban landscapes with suburban subsoiling J Hydrol 543770ndash781 (2016) doi 101016jjhydrol201610051
143 C J Walsh T D Fletcher A R Ladson Stream restoration inurban catchments through redesigning stormwater systemsLooking to the catchment to save the stream J N AmBenthol Soc 24 690ndash705 (2005) doi 10189904-0201
144 N M Schmadel et al Thresholds of lake and reservoirconnectivity in river networks control nitrogen removalNat Commun 9 2779 (2018) doi 101038s41467-018-05156-x pmid 30018449
145 C D Sayer Conservation of aquatic landscapes Pondslakes and rivers as integrated systems WIREs Water 1573ndash585 (2014) doi 101002wat21045
146 R G Luthy D L Sedlak M H Plumlee D Austin V H ReshWastewater‐effluent‐dominated streams as ecosystem‐
management tools in a drier climate Front Ecol Environ 13477ndash485 (2015) doi 101890150038
147 D W Roscoe S G Hinch Effectiveness monitoring of fishpassage facilities Historical trends geographic patterns andfuture directions Fish Fish 11 12ndash33 (2010) doi 101111j1467-2979200900333x
148 C M Bunt T Castro-Santos A Haro Performanceof fish passage structures at upstream barriers tomigration River Res Appl 28 457ndash478 (2012)doi 101002rra1565
149 D Weigel et al Evaluation of a trap-and-transport programfor a threatened population of steelhead (Oncorhynchusmykiss) Conserv Genet 20 1195ndash1199 (2019) doi 101007s10592-019-01200-5
150 M H Bond et al Combined effects of barge transportationriver environment and rearing location on straying andmigration of adult Snake River fall-run Chinook SalmonTrans Am Fish Soc 146 60ndash73 (2017) doi 1010800002848720161235614
151 A C Ritchie et al Morphodynamic evolution followingsediment release from the worldrsquos largest dam removalSci Rep 8 13279 (2018) doi 101038s41598-018-30817-8pmid 30185796
152 T Sommer et al Californiarsquos Yolo Bypass Evidence thatflood control can be compatible with fisheries wetlandswildlife and agriculture Fisheries 26 6ndash16 (2001)doi 1015771548-8446(2001)026lt0006CYBgt20CO2
153 G Grill et al Mapping the worldrsquos free-flowing riversNature 569 215ndash221 (2019) doi 101038s41586-019-1111-9pmid 31068722
154 G Gassert M Luck M Landis P Reig T Shiao ldquoAqueductglobal maps 21 Constructing decision-relevant global waterrisk indicatorsrdquo Working paper World Resources Institute(2015) wwwwriorgpublicationaqueduct-global-maps-21-indicators
155 A Ruhiacute J D Olden J L Sabo Declining streamflow inducescollapse and replacement of native fishes in the AmericanSouthwest Front Ecol Environ 14 465ndash472 (2016)doi 101002fee1424
156 T C Gouhier A Grinsted V Simko R package Biwaveletconduct univariate and bivariate wavelet analyses (2018)
157 T Rodriacuteguez-Castillo E Esteacutevez A M Gonzaacutelez-FerrerasJ Barquiacuten Estimating ecosystem metabolism toentire river networks Ecosystems (2018) doi 101007s10021-018-0311-8
ACKNOWLEDGMENTS
We greatly appreciate those who read and provided importantinsights on drafts of this paper A Armstrong K HondulaJ Hosen K Leathers G de Mendoza C Maietta I MunozJ Tonkin and two anonymous reviewers We also thank T Tu forhelp with Fig 1 and S Carlson L Comte T Datry J HosenM Kondolf N Jones D Lytle E Moody J Olden M PowerJ Sabo and M Sanchez-Montoya for valuable discussionsCompeting interests The authors have no competing interestsFunding MAP was supported by funds from the NSF (1639145)and AR was supported by funds from the University of CaliforniaBerkeley and NSF (1802714)
101126scienceaaw2087
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 13 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
restorationLinkages between flow regime biota and ecosystem processes Implications for river
Margaret Palmer and Albert Ruhi
DOI 101126scienceaaw2087 (6459) eaaw2087365Science
this issue p eaaw2087Scienceflow variability and achieve hydrological connectivity between a river and its surroundingsand ecosystem processes To be successful such efforts must go beyond accounting for flood pulses to restore natural mimic ecologically important aspects of natural flow regimes guided by insights into how variations in flow affect biotawater quality and ecological processes In a Review Palmer and Ruhi explain how restoration designs now attempt to
Human activities have altered the flow regimes of many of Earths rivers with negative impacts on biodiversityRiver restoration guided by research
ARTICLE TOOLS httpsciencesciencemagorgcontent3656459eaaw2087
REFERENCES
httpsciencesciencemagorgcontent3656459eaaw2087BIBLThis article cites 147 articles 7 of which you can access for free
PERMISSIONS httpwwwsciencemagorghelpreprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAASScienceScience 1200 New York Avenue NW Washington DC 20005 The title (print ISSN 0036-8075 online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
Science No claim to original US Government WorksCopyright copy 2019 The Authors some rights reserved exclusive licensee American Association for the Advancement of
on May 27 2020
httpsciencesciencem
agorgD
ownloaded from

frequencies amplitudes and phases in the envi-ronment (14) These characteristic patterns ofvariation or regimes can then be connected toorganismal dynamics For example a waveletanalysis of discharge and abundance of silvercarp larvae found a strong association betweenspawning and peak flows over a particular mag-nitude (15) Using a similar approach temporalchange in stream invertebrate diversity was foundto be influenced not only by the seasonality ofrainfall but by the reliability of this variabilityie its predictability (16)Flow regimes differ across river types and
climates (1) but can also shift temporally at agiven place as a consequence of changes inclimate land use or flow management (Fig 3)Although changes in dominant frequencies inthe environment can have strong impacts on com-munities studying the effects of time-varying flowregimes is still rare (17) Faster flow cycles mayfilter out species with longer generation timesand a preference for stable environments (18) andincreased frequencies of extreme events (eg re-current unpredictable droughts) may drive com-munities to novel stable states (19) However westill do not know which temporal scales of flowvariationmay entrain organismal phenology andwhich may simply represent a source of stressAlthoughmost work to understand this questionhas used correlational approaches mathemat-ical models based on metabolic theory and bio-mechanical constraints may help in predicting
the effects of changing frequencies of environ-mental fluctuations and extremes on river orga-nisms food webs and ecosystem processes (20)
Spatiotemporal variation in flow controlsbiotic persistence
River network structure and directional stream-flow influence the movement of drifting orga-nisms and materials downstream (21) and themain ways in which communities disperse andassemble Experiments suggest that the local envi-ronment exerts control of community compo-sition in the more isolated headwaters whereasdispersal and environmental influences togetheroperate in the well-connected main stem or pri-mary downstream channel This is known as thenetwork position hypothesis (22) Although findingshave sometimes been inconsistent with respectto this idea a recent study using graph theory andreplicate fishmetacommunities (ie multiple com-munities linked by dispersal) suggests that supportfor this hypothesis may depend on network struc-ture and the degree of environmental heteroge-neity in headwaters relative to themainstem (23)Incorporating network structure into the studyof flow regimes is also important because thedendritic connectivity of streams and rivers con-strains organismal movement and associatedpersistence (24) This is particularly relevant toclimate change because shifts in both wet-channel network structure and habitat qualityacross the network are anticipated to occur In
this vein recent research has demonstrated thatin arid climates downstream sections of riversmay bemore suitable for fish spawning whereasin wetter conditions the most suitable habitatoccurs in the headwaters (25)The spatial coordination of flow regimes across
the river network also has key implications forbiodiversity persistence In sets of populationslinked by dispersal or metapopulations a com-bination of diverse physical and biological fea-tures helpsmaintain asynchronous dynamics thusmaximizing stability at larger scales (ie riskspreading or portfolio effect) Only recently havethe simultaneous effects of spatial and temporalvariation in flows been incorporated into streamecology For example widespread flood eventsmay cause synchronous mortality of early lifestages of salmonids [eg (26)] thus weakeningthe portfolio effect On the flip side river branchescan provide diversity in flow and habitat con-ditions ensuring that populations aremaintainedacross the network Using an agent-based modelvalidatedwith long-term fish time series researchhas shown that population asynchrony tends tobe higher across branches than within branchesthus branching network complexity is key to pre-serving metapopulation stability (27) Given thatdams and climate change are making flow re-gimes more similar synchrony in flow-dependentecological processes could be increasing aswell The phenomenon of environmentally forcedsynchrony has long been studied in terrestrial
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 2 of 13
Fig 1 Impacts of river fragmentation by dams urban areas andwater risk across the worldrsquos watersheds Degree of fragmentation(DOF) represents the effects of dams on longitudinal habitatconnectivity and urban impacts (URB) capture the degree of infra-structure development in riparian and floodplain areas (measured asnightlight intensity in urban areas) (153) We averaged river-reach leveldata at the watershed scale using streams with long-term averagedischarge higher than 1 m3s and we then distributed averaged valuesamong the following categories lt5 (low impact) 5 to 20 (mediumimpact) and gt20 (high impact) Colors represent different impact
combinations at the watershed level and flow lines represent themajor rivers of the world Polygons outlined in gray are areas withldquohighrdquo and ldquoextremely highrdquo exposure to water-related risk anaggregated measure of water quantity quality and regulatory riskproposed by the World Resources Institute (154) Around 23 of theworldrsquos watersheds are subject to water-related risks and 19 ofthe watersheds with flow show substantial impacts (ie in gt5 of theriver network) related to longitudinal habitat fragmentation by damsor urban areas These dimensions capture the major sources offlow-related river ecosystem degradation
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
population ecology [Moran theorem (28)] andcould represent an underappreciated emerg-ing risk in fresh waters
Abiotic and biotic influences of flow
Stream ecologists traditionally interpreted thedynamics of communities as a deterministic resultof biotic interactions or as a stochastic conse-quence of the environment (29) However researchhas shown that these fluctuate in relative impor-tance over space and time andmay even influenceeach other For instance biotic interactions mayoccur only under particular environmental con-ditions (30) Inferences gathered from long-termdata combined with time-series methods importedfrom econometrics can help elucidate when and
where abiotic influences have primacy For exam-ple multivariate autoregressive state-space modelsapplied to long-term flow and fish communitydata showed that interannual variation in flowanomalies were the ultimate driver of the dy-namics of a desert stream fish community (31)Although non-native and native fishes interactedsuch effects were small compared with the directeffects of flows on the native component Bioticinteractions that emerge from known species-specific relationships between components of theflow regime such as flood timing and popula-tion dynamics can also be predicted from modelsimulations of community trajectory (30) A recentapplication of these interaction-neutral modelsfound that flows departing from the natural reg-
ime simplified interaction networks among plantguilds in desert rivers (32)A fundamental challenge for community eco-
logy for decades has been quantifying the rela-tive influence of abiotic (flow-driven) and biotic(flow-mediated) effects This is important for theaccurate design and prediction of restoration out-comes including for example anticipatingwhenflow alteration facilitates species invasions bycreating new niches (33) by enhancing propagulepressure or eroding native enemies (34) or by acombination of thesemechanismsDistinguishingthese could help managers compare the benefitsof eradicating non-natives relative to restoringecologically important facets of the flow regime
Flow alteration filters species and traits
Trait-based approaches are becoming increasinglypopular in hydroecology because they enhancecomparison of flow-ecology relationships acrossclimates (35) they lead to predictions about howcommunity change may affect ecosystem func-tioning (36) and they identify the mechanismsthat filter or promote particular taxa under stress-ful conditions such as drought (37) In the case offlow intermittency traits can be used to identifycommunity tipping points by connecting local en-vironmental conditions to organisms with traitsthat allow them to persist in isolated pools or wetsediment or connecting network-level water con-ditions to the ability of highly resilient organismsto drift or actively recolonize from perennial re-fugia (38) Traits can also be used to assess re-storation trajectoriesmdashfor instance by assessingfluctuations in the share of strategies that representdifferent life-history trade-offs [eg species limitedby reproductive capacity or by resources (39)]By studying a communityrsquos taxonomic char-
acteristics (eg species composition) and func-tional characteristics (eg dispersalmodes feedingstrategies) ecologists can determine to whatextent trait redundancy across speciesmay ensureagainst loss of functional diversity For examplein a study on Alpine stream communities subjectto climate change and flow regulation functionaldiversity increased while functional redundancydecreased (40) This finding indicates that ongoinghydroclimatic change can reduce the ability of acommunity to withstand further alteration evenif trait diversity is high because the persistenceof individual traits may be linked to the fate of afew sensitive species In an experimental studycommunities affected by drought presented com-parable taxonomic and functional decays implyingthat these communities were both taxonomicallyand functionally vulnerable (41) The risk of com-munity collapse in response to flow alterationalso depends on the way in which ldquowinningrdquo andldquolosingrdquo traits co-occur in a given species andacross species in a community as a result of aparticular stress Thus community-wide responsesto a particular alterationmay be stronger if speciesaremaladaptive to that change for instance bybeing large-bodied longer-lived and collector-feeding (filtering particulate organic matter) ina scenario of increased flow variability (18) Under-standing how flow alteration impairs species with
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 3 of 13
Nutrient cycling
Primary production
Energy inputs
Secondary production Dispersal refugia Invasions
Food-web interactionsMetabolic regime
Hydrologic connectivity
Streambed sediments
Water chemistry temperature
Channel form riparian vegetation
Decomposition Species demographic rates
River ecosystem integrity
Altered flow regimes
Restored flow regimes
Time
Restoration toolboxAlter pattern of reservoir releasesImprove stormwater controlReforest and restore watersheds
Disc
harg
e w
ater
leve
l Muted high flows
Storm flows
Altered periodicity
Flow homogenization
Shifting baselines
Extreme low flows
Fig 2 Process-based restoration posits that actions must target processes not patternsthat have been degraded River ecosystems are sustained by a host of environmental conditionsor ecosystem structure (gold rectangles) which in turn influence ecosystem processes (greenrectangles) and biotic processes (blue rectangles) These processes as well as ecosystem structureare all controlled by the flow regime Thus correcting facets of flow regime alteration may enhanceriver ecosystem integrity in both direct and indirect ways
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
traits that have effects on ecosystems (eg byparticular feeding strategies) will help anticipatethe risks of losing major ecosystem functionssuch as detrital decomposition (36)
How does flow variation control riverecosystem processes
Rivers produce transform and store organicmatter (OM) and nutrients Carbon (C) and nu-trient concentrations and fluxes vary longitudi-nally and laterally as river networks expand andcontract with rain These variations influenceand are influenced by stream ecosystem metab-olism which can be described as net ecosystemproduction or the difference between gross pri-mary production (GPP) and ecosystem respiration(ER) The metabolic balance between heterotro-phic and autotrophic production that providesresources for consumers can shift seasonally orwith changing flows as can hydrologic connect-ivity that influences inputs of OM and solutes torivers (42) Increasing availability of sensors andadvances inmodeling (43) havemade the study ofmetabolism OM processes and solute dynamicsripe for linking to biotic processes
Flow variability andecosystem metabolism
Spatiotemporal variability in ecosystem processesis a fundamental characteristic of rivers and flowregime is a major driver of this variability (Fig 4)Recentwork shows thatmany rivers in theUnitedStates have strong seasonal patterns in primary
production with spring peaks for larger riversand summer peaks for smaller rivers generallyGPP is negatively correlated with high variationin daily discharge (44) In regions with little sea-sonality andor continual canopy cover GPP islower and its patterns are less predictable (44)Bernhardt et al (45) provide a comprehensivereview of stream metabolism the factors thatinfluence it and why it is difficult to predict Flowhas the largest effect at the extremes such that atlow flowswhen the streambed begins to dry largedrops in GPP occur (46) Storms also often reduceGPP (45) However patterns may vary as a func-tion of flow predictability For example researchin a river characterized by predictable peaks dur-ing spring snowmelt found over two seasons thatGPP rose during spring snowmelt sampling (47)whereas high flows during the winter and sum-mer reduced GPP Within a year this patternresulted in a switch from net autotrophic to netheterotrophic productionThe paths that water takes as it moves to and
within a streammdasha spatial aspect of its flowregimemdashinfluencesmetabolismBycouplingmetab-olism and chemistry time series with informationonhydrologic connectivity and flowpaths ERwasshown to increase in an organic-rich watershedduring peak flows and remained elevated forweeks (48) Sustained hydrologic connectivitywith soils likely fueled inputs of dissolved orga-nic carbon (DOC) that promoted bacterial respi-ration Such inputs can lead to nonlinear responsesinmetabolism For example increasing flows have
led to decreases in GPP up to some thresholdflow beyond which GPP was suppressed by re-duced water clarity from flow-elevated humicDOC (49) In the same vein higher riverine GPPhas been associated with reduced suspendedsediments and high temperatures during low-flow droughts (50) When flows cease entirely andsections of riverbeds dry up carbon dioxide fluxesto the atmosphere can be far higher than those inflowing sections of the channel (51) presumablybecause of spikes in aerobicmicrobial respirationhowever as is the case with methane in riverspartitioning fluxes of these gases to changes inmetabolism (aerobic or anaerobic) is difficult par-ticularly under variable flows (52)
Hydrologic connectivity drivessolute dynamics
Research is growing on how hydrologic connect-ivity to hillslope soils influences water chemistry(53) and how variation in discharge and riverstage influence the strength of bidirectional waterexchanges that affect nitrogen (N) uptake reten-tion and removal (54) Periods of hydrologic con-nectivity may be followed by long periods withlimited exchange which may differ greatly alongthe stream corridor This can lead to complex CN and phosphorus (P) dynamics that influenceor are influenced by microbial and algal produc-tivity When connectivity with watershed solutesources is extensive inputs of solutes such asDOCmay be so high that they exceed the ability of riverecosystem processes such as microbial uptake toregulate their fluxes (55) Theory posits that largestorms pulse terrestrially derived dissolved or-ganicmatter (DOM) into streams and it is quicklyshunted downstream by high flows where it isprocessed in larger parts of the network (56)However how far DOM in pulsedwater is trans-ported is governed not only by discharge but bythe composition of DOMUnderstanding the rela-tionship between DOM sources and its composi-tion is being advanced by combining time-seriesdata with isotopic information to evaluate spatialvariability in hydrologic flow paths transit timeand chemistry as watermoves from the terrestrialto aquatic realms (53 57) It is difficult tomeasurebiotic uptake in streams during storms howeverrecent work suggests that assuming storm flowstransport DOM too quickly for in-stream process-ing may be a poor assumption if the DOM com-position renders it readily utilizable by biota (58)Although hydrologic connectivity is not a newconcept it is not typically considered part of theflow regime yet it can be characterized by itstiming duration and frequency and then linkedto stream processes or patterns Newmethods forquantifying connectivity [eg (59)] are emergingand the energetic and water quality importanceof network connectivity to soils and diverse land-scape elements (such as wetlands and ponds) isbecoming a major area of research (42 59)
Hydrology influences decompositionvia consumers
Detritus (or decaying OM) plays key roles instream ecosystems and most consumers even
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 4 of 13
33104
10103
32101
10100
3110-2
9810-4
3110-5
1922 1949 1976 2004
4
16
256
4096
Perio
d (d
ays)
1963Closure of Glen Canyon Dam
Fig 3 Flow regime shifts in the Colorado River at Lees Ferry Arizona over the past 97 years(1922ndash2018) The flow regime changed drastically around 1963 with the closure of Glen CanyonDam Natural seasonality was dampened and artificial weekly and subweekly scales emerged Graydashed lines indicate 7-day flow cycles resulting from flow management for hydropower andyearly flow cycles resulting from extended base flows and spring snowmelt floods Colors representwavelet power and confidence level contours identify statistically significant power A very wetperiod in the mid-1980s and the experimental releases that took place after 1996 restored flowevents but not the regime Across dammed rivers of the American Southwest muted flowseasonality has opened niches for non-natives particularly organisms adapted to living in stableresource-limited environments at the expense of many native fishes adapted to highly variable(periodic or stochastic) flow conditions that characterized free-flowing hydrographs (155) Plotcreated with the biwavelet R package (156) using mean daily discharge data from United StatesGeological Survey station 09380000 [Dam logo US National Park Service]
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
apical predators are at least partially supportedby it (60) Macroconsumers break up particulateorganic matter (POM) into small pieces enhanc-ing the overall mineralization rate by bacteriaand fungi and even moderate flows may resultin export of OM particularly fine POM resultingfrommacroconsumer shredding activities Over-all howevermicrobes largely drive C losses throughrespiration (61) and although nutrient availa-bility temperature and POM composition (egleaf type) influence this how flow mediates rela-tionships has been more difficult to pin downRecent work (62) designed to separate the effectsthat different components of the flow regimehave on the mechanisms driving decompositionin perennial streams found that dissolution andmicrobial processing consistently exceeded theeffects of fragmentation and microbial pro-cessing was best explained by daily variabilityin discharge When flow regimes become in-termittent the lack ofwater can impedemicrobialand consumer colonization of POM resulting indecreases in decomposition the extent of whichis related to the duration of the drying (46 63)Consistent water cover is therefore importantto decomposition and thus shifts toward lowerflows when litter inputs are high (eg in thefall) can have a large impact on consumers andassociated food webs (63 64)
When flows are altered are changes inbiota and ecosystem processes linked
To help ground this question in the world ofrestoration practice we begin with a focus ontwo of the most common forms of river flow al-teration damming and urbanization For theformer we emphasize hydropower dams becausethey are growing in number they strongly alterflow regimes and the mitigation of their impactshas spurred the field of environmental flow re-search We also focus on urban streams becausestudies on the impacts of imperviousness havebeen extensive and these streams have been the
target of numerous restoration efforts We summa-rize knowledge on the effects of flow on organ-isms ecosystem processes and their interaction inthese two types of river systems and then describehow this can inform river restoration in general
Urbanization and dams changeflow regimes
With few exceptions flow regimes below hydro-power dams and in urban streams are highlyaltered at one or both ends of the flow spectrumRegimes in impounded rivers are typically lessseasonal than daily rainfall would suggest par-ticularly below large hydropower dams with highwater-storage capacity rather than below smallerless impactful run-of-the-river facilities (65)Hydro-power dams canmute peak river flows but power-generation cycles induce rapidly fluctuating flowpatterns (hydropeaking) that often occur onhuman-relevant scales [ie hours to weeks (18)]tending to bemore pronounced in seasons of highenergy demand Hydropeaking is often accom-panied by spikes in temperature (thermopeak-ing) either cold or warm depending on seasonand reservoir operation (66) Thus downstreamorganisms and ecosystems are generally subjectto multiple periodic stresses In urban streamsbaseflows can be higher or lower depending onwatershed characteristics and infrastructurehowever peak flows are invariably larger thanthose prior to urbanization Understanding theecological impacts of urban stormflow is com-plicated by indirect effects on water quality In-creases in flow magnitude frequency and ratesof change during storms leads to elevated con-centrations of pollutants Despite this studiesseparating the causes of biological impairmenthave reported that the unique effects of alteredurban flows can be significant (67 68) Spikesin stream temperature caused by runoff fromhot pavement (69) add to stress on the biotaHydropeaking and urban stormflows might
be expected to reduce primary production by
scouring algae (70) yet some studies have re-ported elevated GPP or alternating periods ofnet autotrophy and heterotrophy (71ndash73) Lightcan override the effects of flow on metabolismby enhancing algal photosynthesis (74) how-ever if hydropeaking increases turbidity GPPmay decline (73) In urban streams elevatedGPP is often attributed to open canopies andorto elevated nutrient concentrations both of whichoften co-occur with their altered flow regimes(72) As discussed later such shifts in primaryproductivity have been linked to changes introphic structure eg more algal-based foodwebs (75) depending on the relative availabil-ity of terrestrial carbon inputs (both DOM andPOM) (76) Inputs of POM and its quality as afood resource changewith landscape context (egvegetative cover) and catchment hydrology (77)but POM availability can also be limited by flowvariability which can increase decompositionrates and decrease residence times (78 79)As is the case for running-water systems in
general high flows in urban streams bring pulsesof solutes including N DOM and various pol-lutants (80) Solute dynamics and biogeochemicalprocesses in regulated rivers have been insuffi-ciently studied with researchers focusing moreon such processes in reservoirs rather than indownstream river ecosystems We found nostudies attempting to link urban or regulatedflow regimes and solute fluxes to changes infood webs or stream metabolic balance How-ever recent studies show shifts in microbial tax-onomic and functional composition in responseto pulsed urban flows (81) and thesemay help inexploring a potential link but are complicated bythe fact that non-flow stressors affect consumercomposition For example urban and regulatedrivers typically have animal communities thatare less diverse and composed of tolerant groupsoften non-native Hydropeaking and urban stormflows can certainly cause direct mortality via dis-placement and transport to unsuitable habitatsbut many of the organismal effects are realizedthrough indirect pathways For instance hydro-peaking reduces the viability of river-edge egg-laying specialists by limiting their access to suitablespawning habitat disrupting reproductive successand insect emergence (82) Flow regulation caneven create ecological traps if organismal phenol-ogy shiftswith thenovel environmental conditionsFor example dam-induced warming triggeredsummer diapause in a mayfly resulting in theloss of its last generation (83) Dams also isolatepopulations harming organisms that requiremigrations to complete their life cycles whetherthese are between riverine and marine habitatsor within the freshwater domain By analogy al-tered flow regimes in urban streams can also leadto isolated pools in streams with lowered base-flows This can be associated with periods ofhypoxia punctuated by extreme flow pulses inwhich these streams become ldquoscoured or suffo-catedrdquo (74)As illustrated using urban streams and reg-
ulated rivers research is moving forward andhints at the three-way nexus but is clearly still
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 5 of 13
Fig 4 Ecosystem metabolismpatterns across a river networkSpatial patterns in river eco-system metabolism for the Deva-Cares catchment in northernSpain as represented by the ratioof gross primary production(P) to ecosystem respiration (R)A PR gt 1 indicates autotrophicprocesses dominate so theseriver sections are accumulatingor exporting organic carbonPR lt 1 indicates dominanceby heterotrophic processes inriver sections that are receivingorganic C input (eg fromterrestrial sources) The boxesindicate regions of the networkimpacted by human pressures including deforestation (brown box) waste products from urbansites (green box) and a combination of the two (purple box) Such visualizations are usefulto managers in understanding regions of concern that may need the most restoration actions[Reproduced with permission from (157)]
PR ratiolt025
gt150
025 ndash 050 050 ndash 075075 ndash 100100 ndash 150
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
inadequate to meet restoration needs Studieson ecosystem processes below dams are parti-cularly limited and much of what we knowcomes from only a few river systems Furtherpublished work focuses primarily on benthicecosystem processes immediately below damsrather than riverine planktonic realms whereproductivity in large regulated rivers can beimportant
Linking flow biota andecosystem processes
Research on flow regimes that bridges organis-mal and ecosystem ecology has traditionallyrelied on states instead of rates ie nutrientconcentrations rather than uptake or fluxesstanding stocks of algae rather than productivityor metabolism and abundance of organismsrather than secondary productivity This workgenerally shows that flow variability suppressesand stability increases algal standing stock andgrazer densities with peaks in invertebrate bio-mass following those of algae [eg (84 85)]Over the past decade research began to focuson how flow regimes alter functional aspects offoodwebs For example biofilms inMediterranean-climate rivers are thicker andmetabolicallymoreactive downstream of flow-stabilizing dams (71)When flow variability increases below hydro-power dams fluctuations can reduce biofilmdevelopment and its quality as a food resource(86) Using experimental flumes subjected tohydropeaking shifts have been found in periph-yton fatty acid content from important highlyunsaturated compounds to nonessential saturatedones (87)Indirect evidence suggests that flow-induced
changes in dominant resource type (eg algaeversus detritus) resource quantity (biomass) orresource quality (nutrient content) can propa-gate to higher trophic levels via bottom-up ef-fects For instance detritivorous invertebrateswere found to dominate species richness andinvertebrate biomass upstream but not down-stream of an irrigation dam (75) diets of om-nivore insects indicated that flow stabilizationshifted a detrital into an algal-fueled food webSimilar evidence comes from Glen Canyon Damin the Colorado River (88) Near the dam whereprimary production is high (73) food webs aresimplified and gut content analyses have sug-gested algal production supports gt50 of in-vertebrate and gt70 of fish production andthe food web can be easily perturbed by flooddisturbance Farther downriver food webs showincreased reliance on detrital resources havehigher trophic efficiency and are more complexand more resistant to disturbance (76) In con-trast recent work on seasonal tropical riversfound algal productivity and zero-flow distur-bance did not explain food-web structure Inthis case it seems that highly mobile predatorsbuffered the local effects of drought (89) Thisstudy along with earlier ones at the site (90) isof particular interest because it links temporalpatterns in predatory fish with flow-dependentavailability of resources
Only a few studies have provided evidence oftop-down controls on foodwebs being influencedby flow variation Fish in a tropical floodplainriver were found to exert top-down control onprimary producers but only under certain hydro-logic conditions (91) Similarly in aCalifornian riverwinter flood pulses were shown to control insectconsumers late in the season ultimately deter-mining whether fish exerted top-down controlon the lower trophic levels (10) These illustratethe importance of pulsed hydrology on resource-consumer linkages and show that not just standingstocks but also the dominant controls structuringfood webs may fluctuate over time in response toflow variationAn indirect line of evidence supporting the
importance of the flow-biota-ecosystem processesnexus comes from theoretically oriented researchon food-web structure Changes in productivityhabitat size and disturbance underpin the basisof the three main hypotheses of food-chainlength which state that chains should be longerin stable productive large ecosystems (92) No-tably flow regimes influence these three con-trols Because food-chain length determines keyaspects of a river ecosystem such as the risk oftrophic cascades or the pollutant accumulationin top predators understanding how it is af-fected by flow alteration is important How-ever understanding the interplay between thehypotheses of food-chain length and the flow-dependent biological mechanisms that allowfood webs to shift in structure (eg predatorinvasions local extinctions consumers chang-ing diets) is still a relatively recent endeavor(93ndash95) Evidence suggests that zero-flows short-en food chains mainly by keeping streamsfishless (94 96) whereas stabilization of variableflows can lengthen food chains by allowing om-nivores to feed on abundant high trophic levelprey (95)New studies are also focusing on interactions
between drivers of food-web structure that hadbeen previously only been examined in isolationsuch as disturbance regimes and ecosystem sizeboth influenced by flow regime Using metabolictheory predator biomass has been shown toscale with both prey resources and fluctuatingstream habitat size with reductions in preda-tor size leading to declines in predator biomasssupported per unit of prey biomass (97) Movingforward the combination of habitat andmetabolicconstraints on predators may provide a basis toconnect flow-induced disturbance changes inecosystem processes and habitat fluctuationsto riverine food-web structure The metabolictheory of ecology (98) is a unifying frameworkthat allows scaling responses of organisms towhole ecosystems andmay become particularlyuseful in situations where flow alteration affectsorganismal body size or water temperature Insuch cases strong changes in metabolic ratescan be expected affecting resource uptake andorganismal growth to ecosystem-level produc-tion and respirationFinally an important body of food webndashrelated
research that provides strong inferential evidence
for the nexus comes from studies on animal-mediated nutrient cycling and ecological stoi-chiometry Theory suggests detrital and algalfood websmay be linked by cascading effects onnutrient cycling (99) and empirical work showsthat organismal excretion andmigration (whichrelocates C) may be important to this link (100)(Fig 5) A recent study (101) showed that ex-cretion of highly labile dissolved organic C ni-trogen (N) and phosphorus (P) by aquatic insectsmay be sufficient to support a substantial fractionof microbial energy and nutrient needs Giventhat C availability can be too low tomeetmicrobialenergy needs [eg (102 103)] and that microbialactivity enhances detrital quality to consumers(104) a C subsidy from excretory processes couldbe important at least during low-flow periodsAdditionally microbial use of excreted dissolvedorganic nitrogen (DON) and DOP may releasealgae from potential competition with microbesfor inorganic N and P if this is the case GPPshould increase Ecological stoichiometry mayalso be important for understanding constraintson organismal growth rates as rapid growthdisproportionally increases demand of P [as it isused in ribosomal RNA (105)] This could in turnlimit the ability of organisms to grow fast andexploit temporary habitats Interestingly elemen-tal ratios can vary interspecifically in response totemperature or predation pressure (106)mdashfactorsthat increase with low flows and habitat fragmen-tation Overall studies that link biota with bio-geochemical processes remain rare (107) butanalyzing flow regimes as a driver of elementturnover rates may provide a way forward
Can hydroecological advances informrestoration practice
Shortfalls in the outcomes of river restorationhave prompted calls to identify and restore pro-cesses that support and sustain biological com-munities rather than focusing only on rivergeomorphology and habitat In urban streamsscientists have critiqued an overemphasis onstructural engineered approaches (7 108) thatdo little to restore the full flow regime (109) Inlarge regulated rivers identifying and restoringimportant features of the flow regime via damoperations is often possible (albeit expensive)However that does not solve other problemsassociated with dams such as habitat frag-mentation (110) altered sediment and thermalregimes (111 112) or disrupted biogeochemicalprocesses (113) Overall efforts to alter reservoirreleases or improve urban stormwater infra-structure to partially mimic a riverrsquos natural flowregime have great potential Technologies arebeing evaluated for real-time management ofurban flows and novel ways to design flow re-leases from dams are being proposed (114 115)however research still lags far behind the needfor solutions Importantly the paucity of studieson the flow-biota-ecosystem processes nexus pointsto a need for more integrative research connectinghydrologists population and community ecologistsand ecosystem ecologists in the context ofrestoration
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 6 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
Restoring flow regimes ratherthan eventsScientifically informed flow designs may existthat maintain ecosystem integrity in regulatedrivers (116 117) but these have produced mixedresults to date In the dam-altered Putah Creekin California mimicking the natural timing ofhigh and low flows enhanced native fish by im-proving their spawning habitat reducing watertemperatures maintaining flowing conditionsand reducing the abundance of non-native spe-cies (118) However other similar attempts havebeen less successful In some cases non-nativefish density also benefitted from flow patternsmimicking an unregulated regime (119) in othersnon-natives bounced back shortly after the flowrelease (120) These experiences illustrate thateven natural flow features may have inadvertentnegative effects if only part of the community orecosystem is considered Models for the NavajoDam (San Juan River New Mexico) show thatadequately timed releases could simultaneouslyenhance native and minimize non-native fishpopulations (121) thus predicted benefits couldsurpass those of mimicking a natural flow re-gime potentially persisting even during dryyears Another study using the ldquodesignerrdquo flowparadigm (114) attempted to understand howdifferent facets of flow variability control fisheryproduction in the Lower Mekong floodplains Time-
series models combining discharge and fisherycatch suggested that enhanced flood pulses andlong inter-flood intervals may increase fisheryyieldsAlthough environmental flow practices have
mostly focused on re-creating discrete flowevents (eg flood pulses) these can still serve asvaluable experiments An iconic example of alarge-scale flow release is Minute 319 an amend-ment to the United StatesndashMexico treaty allow-ing opening of floodgates in the Lower ColoradoRiver and a one-time rewetting of the parchedColorado River Delta (122) Although this pulsed-flow experiment did not promote establishmentof native woody riparian seedlings such trialscan inform future restoration practices for ex-ample by determining whether the provision ofbare moist substrate enhances seed germina-tion (123) Hypothesis-driven experimental flowreleases combined with deep knowledge of tar-get ecosystems can provide insights into resto-ration actions [eg (124)] Still an outstandingquestion is how flow regime restoration couldbe leveraged to restore other important alteredaspects of the ecosystem such as sediment andthermal regimes For example cold-water releasesby existing dams in climate-vulnerable basinscould prevent organisms that thrive at low tem-peratures from becoming replaced by general-ist warm-tolerant taxa (125)
Although the concept of environmental flowsis generally associated with impounded riversits relevance to urban streams is clear (126)However because frequent high storm flowsare viewed as the problem reducing them gen-erally becomes the only restoration goal This isoften attempted by shifting the spatial distribu-tion of velocities within channels through ma-nipulation of channel sinuosity installation offlow-diverting weirs or adding other structuresThese approaches may reduce local erosion butrarely result in levels of water quality ecosystemprocesses or biodiversity in urban streams thatcome close to those of unimpacted streams(127 128) Focusing on the channel ratherthan the watershed context at large means thatkey ecosystemprocesses can remain compromised(129) A recent study compared time series ofmetabolism in two streamsmdashonewith stormwaterponds in the watershed and the other restoredby altering channel slope banks and in-streamstructural complexity (103) The study found thatpeaks in GPP and ER in the ldquorestoredrdquo streamwere higher more prone to resets induced byflashy storms and that recovery time scales werelonger compared with the stream in a watershedwith stormwater measures (Fig 6) Ponds areone type of nature-inspired green infrastructureused to restore streamflow tomore natural condi-tions tree plantings constructed wetlands andgrass swales can in turn replace gray infrastruc-ture such as concrete tunnels that capture andstore waterDistributing green infrastructure in water-
sheds helps restore streams because it targetsthe cause of streamflow alteration includinginsufficient infiltration groundwater rechargeand more generally limited watershed storagecapacity (129) By tackling the cause rather thanthe symptoms of flow alteration these practicescan reduce ecological impacts that result fromthe cascading effects of altered flows includingpoor water quality and temperature spikes Re-search on the effectiveness of green infrastructureon restoring streams is sparse and limited bylack of data before and after implementationalthough studies are beginning to appear Forexample upland green infrastructure in theform of shallow wetland-like pools that capturestormwater and reconnect it to the groundwaterhas been shown to enhance baseflows while re-ducing peak flows and pollutant loadsmore thanrestoration structures within the mainstem ofthe receiving stream (130) Similar structuresbuilt between stormwater outfalls and perennialstreams did not modulate the timing magnitudeor duration of urban storm responses nor didthey improve water quality or biodiversity (131)illustrating the need for researchers to understandhow exact design and watershed context influ-ence performance
Spatiotemporal variability andrestoration designs
Research showing how local and watershed-scale environmental drivers vary temporally andalong river networks in ways that shape biota
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 7 of 13
Flow pulses (storms) flashy flowsDroughts intermittencyLowstable flowsAltered timing seasonality
DOC N P
DetritusMicrobial
stoichiometric constraints
Excretion exudates
Detrital pathway
Algal pathway
Algal stoichiometric constraints
Organismal dispersal source-sink dynamics
Algae
Community replacement
ndash ndash +
ndash ndash + ndash
+
+ndash +
Invasions
Turnover
Consumer stoichiometric constraints
Fig 5 Flow variation affects biota and ecosystem processes and food webs integrate bothHydrologic connectivity along river networks is a first landscape-level filter constraining dispersal oforganisms in river networks Extreme flow variation (floods and droughts) are a source of disturbanceto all organisms (gray box) but particularly to the higher trophic levels given their higher metabolicdemands In turn flow variation controls inputs of nutrients and terrestrial leaf litter to the stream whichinfluences microbial activity and the relative contributions of the detrital (brown) and algal (green)pathways in sustaining aquatic consumers Not just quantity but also food quality (eg CN and CPratios) influence dietary choices of consumers and thus trophic transfer efficiency Finally alteration offlow regimes may influence all these compartments and may also open niches for new players Ifbiological invasions occur at the top of the food web these have the potential to divert energy away fromnative predators and to alter food-web structure through top-down controls
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
and ecosystem processes can help prioritizeselection of sites for restoration For exampleusing the concept of river network saturation(55) to identify river sections where in-streamremoval of nutrients cannot keep up with highsource inputs of nutrients suggests that restorationactions targeting those sections may dispropor-tionately improve water quality Identification ofsubcatchments that disproportionately influencestream chemistry may also be informed by com-bining methods from landscape ecology andcatchment hydrology focusing on the spatialscale of variance in water chemistry (132) Head-waters are often prioritized for restoration be-
cause poor water quality in them can havedownstream impacts Research showing thatlocal environmental constraints on river com-munities are important in headwaters as well asin the larger channels but dispersal constraintsare much less important in the former (133 134)implies that headwaters should be prioritizedHowever research quantifying time-varying con-tributions of local communities to river-wide(network) biodiversity can also inform prioritiza-tion by identifying sites that host a dispropor-tionate share of species or that host uniquespeciesmdashfor example because they shelter sensi-tive taxa during critical flow bottlenecks This idea
encapsulates the concept of keystone habitatsor local sites that strongly influence metacom-munity dynamics (135) This has been appliedto aquatic invertebrates across intermittent rivernetworks by tracking time-varying site-specificcontributions to community dissimilarity (or betadiversity) (136)Other approaches to prioritize restoration ac-
tions may be based on contributions to system-wide stability rather than numbers of individualsor species For example the restoration of fall-run Chinook salmon portfolios may be assessedvia the contributions that different periods andlocationsmake tometapopulation-wide stabilityowing to spatiotemporal variation in habitatconditions (137) Additionally interventionscould promote conditions that are not impor-tant now but could be important in the future[ie proto-refugia (138)] For example resto-ration of groundwater-influenced habitats couldmitigate future stress to drought-sensitive taxaunder scenarios of increased intermittencyPatches of undisturbed land that protect flowregimes in streams within a larger catchmentthat is otherwise urbanized can be sources ofcolonists to restored sites Priorities that focuson present degradation of river sectionsmay failto address site contribution to impairment orrecovery at broader scales or under differentscenarios These insights indicate a great poten-tial for new methods to prioritize restoration ofboth urban and regulated streamsWe know little about how altered flow regimes
propagate through entire river networks butwork is starting to assess the cumulative effectsof dams on flow regimes as well as dam-levelcontributions to alteration Although these ap-proaches have largely focused on dam cascades[eg (139)] if expanded they could help identifyfavorable sections in river networks to developenvironmental flow operations Here exper-imentation could again help with testing ofnew strategies For instance large-scale flowexperiments could be designed that coordinatereleases among dams or between dams and free-flowing tributaries to enhance the restorativeeffects of natural pulses (140)
Hydrologic connectivityinfluences restoration
Recovering critical flow paths and their connec-tivity to rivers is important but it remains asubstantial challenge for flow-degraded riversand streams Whereas regulated rivers tend tohave reduced lateral and longitudinal hydro-logic connectivity urban streams have unnaturallevels of connectivity because pipes and over-land runoff route water quickly In both cases fun-damental hydroecological insights into streambedstructure landscape storage capacity and ground-water connectivity can help to envision novel re-storation approaches (130 141 142) More generallyhydrologic connectivity is becoming a frameworkto identify alternative ways to restore streams viastormwater management (143) For example arecent study suggested that reducing road densityand hydrologic connectivity between roads
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 8 of 13
ER
Disc
harg
e (f
t3 s)
GPPG
PP o
r ER
(g O
2 m-2
day
-1)
Discharge
Watershed BMPs
In-stream restoration
0
ndash4
4
0
ndash4
4
4
2
6
4
2
6
Jan Sept July May Mar Nov Jan
A
B C
Fig 6 Green infrastructure as a means to restore stream ecosystems (A) Discharge andecosystem respiration (ER more-negative values mean higher respiration) and gross primaryproduction (GPP) for two urban streams in which restoration efforts involved (top) modifications inthe stream channel and (bottom) traditional stormwater best management practices (BMPs)[Reproduced with permission from (103)] (B and C) Example of a comprehensive greeninfrastructure approach to ecologically restore an urban headwater stream that also supports thelifestyle of resident urban communities (B) Stream at the onset of restoration and (C) afterrestoration [Reproduced with permission from Greenworks Portland OR]
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 9 of 13
Table 1 Research challenges and opportunities Despite the progress that has been made in understanding how flow regimes affect biota and ecosystem
processes major challenges persist that prevent a complete understanding of the flow-biota-ecosystem processes nexus Making progress on these
challenges requires a mix of fundamental and applied research
Topic Research challenge Opportunity
Connecting
organismal to
ecosystem-level
processes
Research on the effects of flow on organisms
and ecosystem processes has followed parallel
tracksmdashhindering our mechanistic
understanding of the flow-biota-ecosystem
processes nexus
Generate theory that connects concepts and frameworks
that are well accepted in river community and ecosystem
ecology For example the notion of environmental filters
that connect regional to local composition via a combination
of dispersal and environmental conditions (community
ecology) with analogous theory on solute transport versus
reactivity (ecosystem ecology)
Our current understanding of the links between
phenology of biota and ecosystem processes
is precursory
Use high-frequency sensors to characterize metabolic
regimes at sites with ongoing organismal monitoring
Leverage citizen-science biodiversity data to detect
spatiotemporal shifts in animal phenology
(eg insect emergence fish spawning)
Use remote sensing tools to better understand
links between hydrology and aquatic and riparian
plant phenology
The link between stoichiometry and flow
variation remains largely unexplored
Repurpose data that is already being collected to test
flow-stoichiometry relationships eg studies using C and
N stable isotopes to describe CN ratios across flow
regime types Perform field experiments to quantify
indirect effects of invertebrate and fish consumption
on ecosystem metabolism
Ecological
consequences of
hydrologic
connectivity
The importance of anaerobic metabolism in
carbon processing as flows cease and parts of
streambeds dry is a major frontier research area
Create networks of sites across different geologies
and flow regimes that collect time-series data on
methane nitrous oxide and carbon dioxide fluxes
from intermittent streams
We ignore how biota in the wet phase is
influenced by processes and patterns
from the dry phase
Research the functions of dry riverbeds Use camera traps and
drone technology to investigate the role of dry riverbeds
as a dispersal corridor metabarcoding to quantify
ldquodark biodiversityrdquo in the hyporheic
Wetlands and their links to stream networks are poorly
delineated globally Even in the United States and
Europe current wetland inventories do not capture
these links yet understanding how their connectivity
to river systems influences network-scale ecosystem
processes is needed
Analyze time series from high-frequency sensors distributed in
river networks varying in wetland connectivity Further
development and application of time-series methods
used by neuroscientists
We need to identify the relationship between
varying flows and solute dynamics
associated with changes in catchment-scale
hydrologic exchanges
Design collaborative work among catchment hydrologists
and stream ecologists combining methods to estimate
transit times residence times and identify sources
across heterogeneous landscapes Link these properties to
land management and water quality aspects
Scaling and
transferring flow-
ecology relationships
across space and time
We ignore how biodiversity responds to
propagation of flow regime alteration
across river networks
Leverage extensive hydrometric networks (eg United States
Geological Survey National Water Information System)
to perform spatially replicated time-series analyses
on flow and use these to generate expectations
on how river network position should mediate
biodiversity responses to flow alteration
Flow-ecology relationships do not account for
nonstationarity and abrupt change
Use process-based models that account for mechanisms
and associated uncertainties When using time-series
data and models select methods that are robust
to nonstationarity Linear flow-ecology relationships
should not be assumed
Current flow-ecology relationships assume that
responses of species do not change over time
Consider flow regime alteration as a process with ecological and
evolutionary consequences Include intraspecific variation
in traits and potential for rapid evolution in models
continued on next page
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
and streams may improve water quality morethan other restoration options (70) Additionallyresearch on the age and lability of DOC enteringstreams has shown how precipitation or land dis-turbance can change watershed flow paths (53)and this could inspire new restoration approachesfocused on land use and soil properties Manystudies on hydrologic connectivity and networkstructure suggest a need for restoration to focuson landscape heterogeneity especially on thepresence ofwetlands lakes and ponds (144) Suchfeaturesare frequently lost inurbanizedcatchmentsand outcomes could be improved by restoring orincreasing connectivitywith these ecosystems (145)Hydrologic connectivity associated with ground-water inputs or abstractions land-use manage-ment and return flows fromwastewater treatmentplants also need to be considered in these initia-tives (146)Restoring hydrologic connectivity is also key
to achieving biodiversity outcomesmdashparticularlyin dammed systems Longitudinal connectivityfor fish inhabiting dam-altered rivers can beenhanced by the construction of fishways andsophisticated passage facilities However moni-toring data on the effectiveness of such inter-
ventions are still scarce and available evidencesuggests that passage effectivity largely dependson technology used and species-specific fish be-havior (147 148) Although trap-and-transportprograms for migratory fishes can help augmentthreatened populations (149) these practicesoften increase straying and thus the potentialfor genetic homogenization as seen from ju-venile salmon collected at dams and transporteddownstream to increase out-migration success(150) Overall a combination of engineering andmanagement solutions may be best to restorefunctional connectivity in dam-altered riversThe link between hydrologic connectivity andecological restoration is also central to con-siderations on where to build remove and re-operate hydrologic infrastructure Hundredsof dams have been intentionally removed sincethe 1970s and removal of large dams such asthe Elwha River Dam (Washington USA) hasshown that restoring longitudinal connectivitycan have immediate geomorphic effects with com-plex temporal dynamics (151) Infrastructure canalso be designed or reoperated to manage forrestoring lateral connectivity and enhanced flood-plain processes For example the Yolo Bypass in
the Sacramento River California has becomea prime rearing and migration site for chinooksalmon showing that flood control and habitatfor fish and wildlife do not need to be at odds(152) All these examples show the importanceof integrating hydrologic connectivity in ecolog-ical restoration whether the goal is to minimizethe ecological effects of fragmentation restorefree-flowing watercourses or operate green infra-structure to enhance infiltration floodplain con-nectivity or other ecological processes
Research challenges and opportunities
Two decades of concerted efforts to research theeffects of flow regime alteration have tremen-dously advanced our understanding of the waysin which streamflow influences biota and eco-system processes However important challengesremain These aremainly related to the scarcity ofscience connecting organismal to ecosystem-levelprocesses through for example microbial linksthe difficulty of studying watershed-level phys-ical and biogeochemical heterogeneity and itsinfluence on riverine processes and biota thecomplexity of parsing out the direct effects offlow from flow-mediated effects and co-occurring
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 10 of 13
Topic Research challenge Opportunity
Cascading
implications of
hydrologic
alteration
We have a poor understanding of the
interactions among physical drivers
Build on large-scale flow experiments to test hypotheses on the
combined effects of altered flow sediment and thermal
regimes Experiment with multilevel intakes in reservoirs
Promote collection of non-flow variables (eg temperature
water quality parameters) at hydrometric stations
At low flows biota are being exposed to high
concentrations of new classes of pollutants
whose impacts are poorly known especially if
biotic impacts interact with impacts on
ecosystem processes
Design realistic experimental studies to advance knowledge
on the ecological effects of multiple stressors
Collect long-term data at sites varying in contaminant
inputs and hydrologic regimes Link microbial to
organismal and ecosystem-level responses
Using hydroecological
science to inform
river ecosystem
restoration
Environmental flow science largely ignores
human dimensions
Use methods that allow identifying causal networks
in complex socioenvironmental systems Include
human needs and thresholds in research on
flow-ecology relationships
Environmental flows are generally prescribed
at the scale of single infrastructures restoration
designs and assessment are generally at reach scales
Promote coordinated large-scale flow experiments
and restoration designs at the river basin scale
Extend assessment of green infrastructure
effectiveness beyond individual structures to
infrastructure networks and link both
to river outcomes
Environmental flows in impounded rivers
generally target the conservation of some
particular species rather than outcomes at the
food-web or ecosystem level
Advance fundamental research on microbialndash
ecosystem process linkages Expand the notion
of designer flows to food-web structure and
ecosystem services when possible
Environmental flow science has largely
ignored surface waterndashgroundwater
interactions
Expand science to understand the link between
restoration actions groundwater recharge and
inputs to streams Provide science relevant to
the implementation of new legislation on
groundwater management (eg Sustainable
Groundwater Management Act in California)
Uncertainty regarding future
hydroclimate and
river flows is growing
Refine regional climate models Promote research on
the interaction between land-use change and
evapotranspiration Incorporate groundwater
influence in macroscale hydrology models
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
stressors (particularly in urban streams) and thechallenge of predicting outcomeswhen both thephysical and the biological environments arechangingAchieving a predictive understanding of hy-
droecological relationships is essential to design-ing successful self-sustaining restoration actionsin river ecosystems However translating basichydroecological insights into restoration practicerequires understanding the complexities of work-ing within a human-dominated water cycle Al-though it is rarely possible to restore all facetsof a natural flow regime ecologically informeddesigns may help mimic key aspects of the nat-ural flow regime (eg periodic pulse flows) orcreate regimes that are not natural but maximizeoutcomes given socioeconomic constraints InTable 1 we identify persisting and emergingchallenges in hydroecological research We paireach challenge with an opportunity (related toresearch design technology or implementation)We contend that these challenges should beprioritized to advance process-based river eco-system restoration
REFERENCES AND NOTES
1 N L Poff et al The natural flow regime Bioscience 47769ndash784 (1997) doi 1023071313099
2 D A Lytle N L Poff Adaptation to natural flow regimesTrends Ecol Evol 19 94ndash100 (2004) doi 101016jtree200310002 pmid 16701235
3 S Sabater A Elosegi R Ludwig in Multiple Stressors in RiverEcosystems (Elsevier 2019) pp 1ndash22
4 United Nations New UN Decade on Ecosystem Restorationoffers unparalleled opportunity for job creation food securityand addressing climate change (2019) httpswwwunenvironmentorgnews-and-storiespress-releasenew-un-decade-ecosystem-restoration-offers-unparalleled-opportunity
5 A W Lorenz P Haase K Januschke A SundermannD Hering Revisiting restored river reaches - Assessingchange of aquatic and riparian communities afterfive years Sci Total Environ 613-614 1185ndash1195(2018) doi 101016jscitotenv201709188pmid 28954379
6 M A Palmer K L Hondula B J Koch Ecological Restorationof Streams and Rivers Shifting Strategies and ShiftingGoals (2014) vol 45
7 T J Beechie et al Process-based principles for restoringriver ecosystems Bioscience 60 209ndash222 (2010)doi 101525bio20106037
8 S E Bunn A H Arthington Basic principles and ecologicalconsequences of altered flow regimes for aquaticbiodiversity Environ Manage 30 492ndash507 (2002)doi 101007s00267-002-2737-0 pmid 12481916
9 K D Fausch Y Taniguchi S Nakano G D GrossmanC R Townsend Flood disturbance regimes influence rainbowtrout invasion success among five holarctic regions EcolAppl 11 1438ndash1455 (2001) doi 1018901051-0761(2001)011[1438FDRIRT]20CO2
10 M E Power M S Parker W E Dietrich Seasonalreassembly of a river food web Floods droughts andimpacts of fish Ecol Monogr 78 263ndash282 (2008)doi 10189006-09021
11 V Acuntildea A Giorgi I Muntildeoz U R S Uehlinger S SabaterFlow extremes and benthic organic matter shape themetabolism of a headwater Mediterranean streamFreshwater Biol 49 960ndash971 (2004) doi 101111j1365-2427200401239x
12 K Tockner F Malard J V Ward An extension of the floodpulse concept Hydrol Processes 14 2861ndash2883 (2000)doi 1010021099-1085(20001112)141617lt2861AID-HYP124gt30CO2-F
13 J S Perkin et al Fragmentation and dewatering transformGreat Plains stream fish communities Ecol Monogr 8573ndash92 (2015) doi 10189014-01211
14 J L Sabo D M Post Quantifying periodic stochastic andcatastrophic environmental variation Ecol Monogr 7819ndash40 (2008) doi 10189006-13401
15 F Shuai et al Silver carp larva abundance in response toriver flow rate revealed by cross-wavelet modellingEcol Modell 383 98ndash105 (2018) doi 101016jecolmodel201805020
16 J D Tonkin M T Bogan N Bonada B Rios-ToumaD A Lytle Seasonality and predictability shape temporalspecies diversity Ecology 98 1201ndash1216 (2017)doi 101002ecy1761 pmid 28144975
17 N L Poff Beyond the natural flow regime Broadening thehydro‐ecological foundation to meet environmental flowschallenges in a non‐stationary world Freshwater Biol 631011ndash1021 (2018) doi 101111fwb13038
18 A Ruhi X Dong C H McDaniel D P Batzer J L SaboDetrimental effects of a novel flow regime on the functionaltrajectory of an aquatic invertebrate metacommunity GlobChange Biol 24 3749ndash3765 (2018) doi 101111gcb14133pmid 29665147
19 M T Bogan D A Lytle Severe drought drives novelcommunity trajectories in desert stream pools FreshwaterBiol 56 2070ndash2081 (2011) doi 101111j1365-2427201102638x
20 G Woodward et al The effects of climatic fluctuations andextreme events on running water ecosystems Philos TransR Soc London Ser B 371 20150274 (2016) pmid 27114576
21 M A Palmer C M Swan K Nelson P Silver R AlvestadStreambed landscapes Evidence that stream invertebratesrespond to the type and spatial arrangement of patchesLandsc Ecol 15 563ndash576 (2000) doi 101023A1008194130695
22 D Schmera et al Does isolation influence the relative role ofenvironmental and dispersal‐related processes in streamnetworks An empirical test of the network positionhypothesis using multiple taxa Freshwater Biol 63 74ndash85(2018) doi 101111fwb12973
23 R Henriques‐Silva et al A comprehensive examination of thenetwork position hypothesis across multiple rivermetacommunities Ecography 42 284ndash294 (2019)doi 101111ecog03908
24 K L Jaeger J D Olden N A Pelland Climate change poisedto threaten hydrologic connectivity and endemic fishes indryland streams Proc Natl Acad Sci USA 11113894ndash13899 (2014) doi 101073pnas1320890111pmid 25136090
25 S Garbin E A Celegon P Fanton G Botter Hydrologicalcontrols on river network connectivity R Soc Open Sci 6181428 (2019) doi 101098rsos181428 pmid 30891270
26 F Cattaneacuteo B Hugueny N Lamouroux Synchrony in browntrout Salmo trutta population dynamics A lsquoMoran effectrsquo onearly‐life stages Oikos 100 43ndash54 (2003) doi 101034j1600-0706200311912x
27 A Terui et al Metapopulation stability in branching rivernetworks Proc Natl Acad Sci USA 115 E5963ndashE5969(2018) doi 101073pnas1800060115 pmid 29895695
28 P A P Moran The statistical analysis of the Canadian Lynxcycle Aust J Zool 1 291ndash298 (1953) doi 101071ZO9530291
29 G D Grossman P B Moyle J O Whitaker Jr Stochasticityin structural and functional characteristics of an Indianastream fish assemblage A test of community theoryAm Nat 120 423ndash454 (1982) doi 101086284004
30 D A Lytle D M Merritt J D Tonkin J D OldenL V Reynolds Linking river flow regimes to riparian plantguilds A community-wide modeling approach Ecol Appl 271338ndash1350 (2017) doi 101002eap1528 pmid 28263426
31 A Ruhiacute E E Holmes J N Rinne J L Sabo Anomalousdroughts not invasion decrease persistence of native fishesin a desert river Glob Change Biol 21 1482ndash1496 (2015)doi 101111gcb12780 pmid 25430731
32 J D Tonkin D M Merritt J D Olden L V ReynoldsD A Lytle Flow regime alteration degrades ecologicalnetworks in riparian ecosystems Nat Ecol Evol 2 86ndash93(2018) doi 101038s41559-017-0379-0 pmid 29180707
33 M C Mims J D Olden Fish assemblages respond toaltered flow regimes via ecological filtering of lifehistory strategies Freshwater Biol 58 50ndash62 (2013)doi 101111fwb12037
34 P T J Johnson J D Olden M J Vander Zanden Daminvaders Impoundments facilitate biological invasions intofreshwaters Front Ecol Environ 6 357ndash363 (2008)doi 101890070156
35 N Bonada S Doledec B Statzner Taxonomic and biologicaltrait differences of stream macroinvertebrate communitiesbetween mediterranean and temperate regions Implicationsfor future climatic scenarios Glob Change Biol 131658ndash1671 (2007) doi 101111j1365-2486200701375x
36 M I Pyne N L Poff Vulnerability of stream communitycomposition and function to projected thermal warming andhydrologic change across ecoregions in the western UnitedStates Glob Change Biol 23 77ndash93 (2017) doi 101111gcb13437 pmid 27429092
37 D B Herbst S D Cooper R B Medhurst S W WisemanC T Hunsaker Drought ecohydrology alters the structureand function of benthic invertebrate communities inmountain streams Freshwater Biol 64 886ndash902 (2019)doi 101111fwb13270
38 M T Bogan E T Chester T Datry A L MurphyB J Robson A Ruhi R Stubbington J E Whitney inResistance Resilience and Community Recovery in IntermittentRivers and Ephemeral Streams T Datry N Bonada A BoultonEds (Academic Press 2017) pp 349ndash376
39 S Houmlckendorff et al Characterizing fish responses to a riverrestoration over 21 years based on speciesrsquo traits ConservBiol 31 1098ndash1108 (2017) doi 101111cobi12908pmid 28218807
40 D Bruno et al Structural and functional responses ofinvertebrate communities to climate change andflow regulation in alpine catchments Glob Change Biol25 1612ndash1628 (2019) doi 101111gcb14581pmid 30698905
41 T W H Aspin et al Extreme drought pushes streaminvertebrate communities over functional thresholds GlobChange Biol 25 230ndash244 (2019) doi 101111gcb14495pmid 30346098
42 S E Tank J B Fellman E Hood E S Kritzberg Beyondrespiration Controls on lateral carbon fluxes across theterrestrial-aquatic interface Limnol Oceanogr Lett 3 76ndash88(2018) doi 101002lol210065
43 A P Appling et al The metabolic regimes of 356 rivers inthe United States Sci Data 5 180292 (2018) doi 101038sdata2018292 pmid 30532078
44 P Savoy et al Metabolic rhythms in flowing watersAn approach for classifying river productivity regimesLimnol Oceanogr (2019) doi 101002lno11154
45 E S Bernhardt et al The metabolic regimes of flowingwaters Limnol Oceanogr 63 S99ndashS118 (2018)doi 101002lno10726
46 M Colls X Timoner S Sabater V Acuntildea Effects of durationand frequency of the non-flow period on stream metabolismEcosystems 101007s10021-019-00345-1 (2019)doi 101007s10021-019-00345-1
47 A J Ulseth E Bertuzzo G A Singer J Schelker T J BattinClimate-induced changes in spring snowmelt impactecosystem metabolism and carbon fluxes in an Alpine streamnetwork Ecosystems 21 373ndash390 (2018) doi 101007s10021-017-0155-7
48 B O L Demars Hydrological pulses and burning of dissolvedorganic carbon by stream respiration Limnol Oceanogr 64406ndash421 (2019) doi 101002lno11048
49 R T Hensley L Kirk M Spangler M N Gooseff M J CohenFlow extremes as spatiotemporal control points on riversolute fluxes and metabolism J Geophys Res Biogeosci124 537ndash555 (2019) doi 1010292018JG004738
50 J D Hosen et al Enhancement of primary production duringdrought in a temperate watershed is greater in larger riversthan headwater streams Limnol Oceanogr 64 1458ndash1472(2019) doi 101002lno11127
51 L Goacutemez-Gener et al When water vanishes Magnitude andregulation of carbon dioxide emissions from dry temporarystreams Ecosystems 19 710ndash723 (2016) doi 101007s10021-016-9963-4
52 E H Stanley et al The ecology of methane in streams andrivers Patterns controls and global significance EcolMonogr 86 146ndash171 (2016) doi 10189015-1027
53 R T Barnes D E Butman H F Wilson P A RaymondRiverine export of aged carbon driven by flow path depth andresidence time Environ Sci Technol 52 1028ndash1035 (2018)doi 101021acsest7b04717 pmid 29313674
54 G Pinay et al Riparian corridors A new conceptualframework for assessing nitrogen buffering across biomesFront Environ Sci 6 47 (2018) doi 103389fenvs201800047
55 W M Wollheim et al River network saturation conceptFactors influencing the balance of biogeochemical supply and
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 11 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
demand of river networks Biogeochemistry 141 503ndash521(2018) doi 101007s10533-018-0488-0
56 P A Raymond J E Saiers W V Sobczak Hydrological andbiogeochemical controls on watershed dissolved organicmatter transport Pulse-shunt concept Ecology 97 5ndash16(2016) doi 10189014-16841 pmid 27008769
57 C Tunaley D Tetzlaff J Lessels C Soulsby Linking high-frequency DOC dynamics to the age of connected watersources Water Resour Res 52 5232ndash5247 (2016)doi 1010022015WR018419
58 S Bernal et al Supply demand and in-stream retention ofdissolved organic carbon and nitrate during storms inMediterranean forested headwater streams Front EnvironSci 7 60 (2019) doi 103389fenvs201900060
59 J Harvey et al How hydrologic connectivity regulates waterquality in river corridors J Am Water Resour Assoc 55369ndash381 (2019) doi 1011111752-168812691
60 J C Moore et al Detritus trophic dynamics and biodiversityEcol Lett 7 584ndash600 (2004) doi 101111j1461-0248200400606x
61 A D Rosemond et al Freshwater ecology Experimentalnutrient additions accelerate terrestrial carbon loss fromstream ecosystems Science 347 1142ndash1145 (2015)doi 101126scienceaaa1958 pmid 25745171
62 A C Y Yeung J L Musetta-Lambert D P KreutzweiserP K Sibley J S Richardson Relations of interannualdifferences in stream litter breakdown with dischargeBioassessment implications Ecosphere 9 e02423 (2018)doi 101002ecs22423
63 R M Northington J R Webster Experimental reductions instream flow alter litter processing and consumer subsidies inheadwater streams Freshwater Biol 62 737ndash750 (2017)doi 101111fwb12898
64 E Mas-Martiacute I Sanpera-Calbet I Muntildeoz Bottom-up effectsof streambed drying on consumer performance throughchanges in resource quality Aquat Sci 79 719ndash731 (2017)doi 101007s00027-017-0531-6
65 B D Richter G A Thomas Restoring environmental flows bymodifying dam operations Ecol Soc 12 12 (2007)doi 105751ES-02014-120112
66 G Zolezzi A Siviglia M Toffolon B MaioliniThermopeaking in Alpine streams Event characterizationand time scales Ecohydrology 4 564ndash576 (2011)doi 101002eco132
67 T Meiszligner B Sures C K Feld Multiple stressors and therole of hydrology on benthic invertebrates in mountainousstreams Sci Total Environ 663 841ndash851 (2019)doi 101016jscitotenv201901288 pmid 30738264
68 I R Waite et al Effects of urban multi-stressors on threestream biotic assemblages Sci Total Environ 6601472ndash1485 (2019) doi 101016jscitotenv201812240pmid 30743940
69 K C Nelson M A Palmer Stream temperature surges underurbanization and climate change Data models andresponses J Am Water Resour Assoc 43 440ndash452 (2007)doi 101111j1752-1688200700034x
70 J R Blaszczak J M Delesantro Y Zhong D L UrbanE S Bernhardt Watershed urban development controls onurban streamwater chemistry variability Biogeochemistry144 61ndash84 (2019) doi 101007s10533-019-00572-7
71 L Ponsatiacute et al Biofilm responses to flow regulation by damsin Mediterranean rivers River Res Appl 31 1003ndash1016(2015) doi 101002rra2807
72 J M Alberts J J Beaulieu I Buffam Watershed land useand seasonal variation constrain the influence of ripariancanopy cover on stream ecosystem metabolism Ecosystems20 553ndash567 (2017) doi 101007s10021-016-0040-9pmid 30416371
73 R O Hall Jr et al Turbidity light temperature andhydropeaking control primary productivity in the ColoradoRiver Grand Canyon Limnol Oceanogr 60 512ndash526 (2015)doi 101002lno10031
74 J R Blaszczak J M Delesantro D L Urban M W DoyleE S Bernhardt Scoured or suffocated Urban streamecosystems oscillate between hydrologic and dissolvedoxygen extremes Limnol Oceanogr 64 877ndash894 (2019)doi 101002lno11081
75 J-R Mor et al Dam regulation and riverine food-webstructure in a Mediterranean river Sci Total Environ 625301ndash310 (2018) doi 101016jscitotenv201712296pmid 29289778
76 H A Wellard Kelly et al Macroinvertebrate diets reflecttributary inputs and turbidity-driven changes in food
availability in the Colorado River downstream of GlenCanyon Dam Freshw Sci 32 397ndash410 (2013)doi 10189912-0881
77 S J Imberger R M Thompson M R Grace Urbancatchment hydrology overwhelms reach scale effects ofriparian vegetation on organic matter dynamics FreshwaterBiol 56 1370ndash1389 (2011) doi 101111j1365-2427201102575x
78 R T Martins A S Melo J F Gonccedilalves Jr N HamadaLeaf-litter breakdown in urban streams of Central AmazoniaDirect and indirect effects of physical chemical andbiological factors Freshw Sci 34 716ndash726 (2015)doi 101086681086
79 C M Yule J Y Gan T Jinggut K V Lee Urbanizationaffects food webs and leaf-litter decomposition in a tropicalstream in Malaysia Freshw Sci 34 702ndash715 (2015)doi 101086681252
80 T A Newcomer et al Influence of natural and novel organiccarbon sources on denitrification in forest degraded urbanand restored streams Ecol Monogr 82 449ndash466 (2012)doi 10189012-04581
81 A Chaudhary I Kauser A Ray R Poretsky Taxon-drivenfunctional shifts associated with storm flow in an urbanstream microbial community MSphere 3 e00194-18 (2018)doi 101128mSphere00194-18 pmid 29976643
82 T A Kennedy et al Flow management for hydropowerextirpates aquatic insects undermining river food websBioscience 66 561ndash575 (2016) doi 101093bioscibiw059
83 A Glazaczow D Orwin M Bogdziewicz Increasedtemperature delays the late-season phenology of multivoltineinsect Sci Rep 6 38022 (2016) doi 101038srep38022pmid 27905493
84 S G Fisher L J Gray N B Grimm D E Busch Temporalsuccession in a desert stream ecosystem following flashflooding Ecol Monogr 52 93ndash110 (1982) doi 1023072937346
85 G J Scrimgeour M J Winterbourn Effects of floods onepilithon and benthic macroinvertebrate populations in anunstable New Zealand river Hydrobiologia 171 33ndash44 (1989)doi 101007BF00005722
86 E Bondar-Kunze S Maier D Schoumlnauer N Bahl T HeinAntagonistic and synergistic effects on a stream periphytoncommunity under the influence of pulsed flow velocityincrease and nutrient enrichment Sci Total Environ 573594ndash602 (2016) doi 101016jscitotenv201608158pmid 27585428
87 M J Cashman G L Harvey G Wharton M C Bruno Woodmitigates the effect of hydropeaking scour on periphytonbiomass and nutritional quality in semi-natural flumesimulations Aquat Sci 79 459ndash471 (2017) doi 101007s00027-016-0510-3
88 W F Cross et al Food-web dynamics in a large riverdiscontinuum Ecol Monogr 83 311ndash337 (2013)doi 10189012-17271
89 D M Warfe et al Productivity disturbance and ecosystemsize have no influence on food chain length in seasonallyconnected rivers PLOS ONE 8 e66240 (2013)doi 101371journalpone0066240
90 T D Jardine et al Consumer-resource coupling in wet-drytropical rivers J Anim Ecol 81 310ndash322 (2012)doi 101111j1365-2656201101925x pmid 22103689
91 K O Winemiller et al Pulsing hydrology determines top‐down control of basal resources in a tropical riverndashfloodplainecosystem Ecol Monogr 84 621ndash635 (2014)doi 10189013-18221
92 D M Post The long and short of food-chain length TrendsEcol Evol 17 269ndash277 (2002) doi 101016S0169-5347(02)02455-2
93 D M Post G Takimoto Proximate structural mechanismsfor variation in food‐chain length Oikos 116 775ndash782 (2007)doi 101111j0030-1299200715552x
94 P A McHugh A R McIntosh P G Jellyman Dual influencesof ecosystem size and disturbance on food chain length instreams Ecol Lett 13 881ndash890 (2010) doi 101111j1461-0248201001484x pmid 20482579
95 A Ruhiacute et al Flow regulation increases food-chain lengththrough omnivory mechanisms in a Mediterranean rivernetwork Freshwater Biol 61 1536ndash1549 (2016) doi 101111fwb12794
96 J L Sabo J C Finlay T Kennedy D M Post The role ofdischarge variation in scaling of drainage area and food chainlength in rivers Science 330 965ndash967 (2010) doi 101126science1196005 pmid 20947729
97 A R McIntosh et al Capacity to support predators scaleswith habitat size Sci Adv 4 eaap7523 (2018) doi 101126sciadvaap7523 pmid 29978038
98 J H Brown J F Gillooly A P Allen V M Savage G B WestToward a metabolic theory of ecology Ecology 851771ndash1789 (2004) doi 10189003-9000
99 K Zou E Theacutebault G Lacroix S Barot Interactions betweenthe green and brown food web determine ecosystemfunctioning Funct Ecol 30 1454ndash1465 (2016) doi 1011111365-243512626
100 M J Vanni P B McIntyre Predicting nutrient excretion ofaquatic animals with metabolic ecology and ecologicalstoichiometry A global synthesis Ecology 97 3460ndash3471(2016) doi 101002ecy1582 pmid 27912023
101 T B Parr K A Capps S P Inamdar K A Metcalf Animal-mediated organic matter transformation Aquatic insects as asource of microbially bioavailable organic nutrients andenergy Funct Ecol 33 524ndash535 (2019) doi 1011111365-243513242
102 C P Arango et al Urban infrastructure influences dissolvedorganic matter quality and bacterial metabolism in an urbanstream network Freshwater Biol 62 1917ndash1928 (2017)
103 L G Larsen J W Harvey Disrupted carbon cycling inrestored and unrestored urban streams Critical timescalesand controls Limnol Oceanogr 62 S160ndashS182 (2017)doi 101002lno10613
104 H M Halvorson G White J T Scott M A Evans-WhiteDietary and taxonomic controls on incorporation of microbialcarbon and phosphorus by detritivorous caddisfliesOecologia 180 567ndash579 (2016) doi 101007s00442-015-3464-6 pmid 26497125
105 J J Elser et al Biological stoichiometry from genes toecosystems Ecol Lett 3 540ndash550 (2000) doi 101046j1461-0248200000185x
106 E K Moody et al Consumption explains intraspecificvariation in nutrient recycling stoichiometry in a desert fishEcology 99 1552ndash1561 (2018) doi 101002ecy2372pmid 29882955
107 N Welti et al Bridging food webs ecosystem metabolismand biogeochemistry using ecological stoichiometry theoryFront Microbiol 8 1298 (2017) doi 103389fmicb201701298 pmid 28747904
108 M A Palmer S Filoso R M Fanelli From ecosystems toecosystem services Stream restoration as ecologicalengineering Ecol Eng 65 62ndash70 (2014) doi 101016jecoleng201307059
109 M J Burns T D Fletcher C J Walsh A R LadsonB E Hatt Hydrologic shortcomings of conventional urbanstormwater management and opportunities for reformLandsc Urban Plan 105 230ndash240 (2012) doi 101016jlandurbplan201112012
110 C R Liermann C Nilsson J Robertson R Y NgImplications of dam obstruction for global freshwater fishdiversity Bioscience 62 539ndash548 (2012) doi 101525bio20126265
111 J D Olden R J Naiman Incorporating thermal regimes intoenvironmental flows assessments Modifying dam operationsto restore freshwater ecosystem integrity Freshwater Biol55 86ndash107 (2010) doi 101111j1365-2427200902179x
112 D B Osmundson R J Ryel V L Lamarra J Pitlick Flowndashsedimentndashbiota relations Implications for river regulationeffects on native fish abundance Ecol Appl 12 1719ndash1739(2002)
113 D von Schiller et al Regulation causes nitrogen cyclingdiscontinuities in Mediterranean rivers Sci Total Environ540 168ndash177 (2016) doi 101016jscitotenv201507017pmid 26233271
114 J L Sabo et al Designing river flows to improve foodsecurity futures in the Lower Mekong Basin Science 358eaao1053 (2017) doi 101126scienceaao1053pmid 29217541
115 W Xu et al Improving the multi-objective performance ofrainwater harvesting systems using real-time controltechnology Water 10 147 (2018) doi 103390w10020147
116 M Acreman et al Environmental flows for naturalhybrid and novel riverine ecosystems in a changingworld Front Ecol Environ 12 466ndash473 (2014)doi 101890130134
117 N L Poff et al Sustainable water management under futureuncertainty with eco-engineering decision scaling Nat ClimChang 6 25ndash34 (2016) doi 101038nclimate2765
118 J D Kiernan P B Moyle P K Crain Restoring native fishassemblages to a regulated California stream using the
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 12 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
natural flow regime concept Ecol Appl 22 1472ndash1482(2012) doi 10189011-04801 pmid 22908707
119 D L Propst K B Gido Responses of native andnonnative fishes to natural flow regime mimicry in theSan Juan River Trans Am Fish Soc 133 922ndash931 (2004)doi 101577T03-0571
120 T K Pool J D Olden Assessing long‐term fish responses andshort‐term solutions to flow regulation in a dryland river basinEcol Freshwat Fish 24 56ndash66 (2015) doi 101111eff12125
121 W Chen J D Olden Designing flows to resolve human andenvironmental water needs in a dam-regulated riverNat Commun 8 2158 (2017) doi 101038s41467-017-02226-4 pmid 29255194
122 S M Nelson et al A history of the 2014 Minute 319environmental pulse flow as documented by fieldmeasurements and satellite imagery Ecol Eng 106733ndash748 (2017) doi 101016jecoleng201610040
123 P B Shafroth et al A large-scale environmental flowexperiment for riparian restoration in the Colorado RiverDelta Ecol Eng 106 645ndash660 (2017) doi 101016jecoleng201702016
124 E P Glenn P L Nagler P B Shafroth C J JarchowEffectiveness of environmental flows for riparian restorationin arid regions A tale of four rivers Ecol Eng 106 695ndash703(2017) doi 101016jecoleng201701009
125 S Hotaling D S Finn J Joseph Giersch D W WeisrockD Jacobsen Climate change and alpine stream biologyProgress challenges and opportunities for the futureBiol Rev Camb Philos Soc 92 2024ndash2045 (2017)doi 101111brv12319 pmid 28105701
126 C J Walsh T D Fletcher M J Burns Urban stormwaterrunoff A new class of environmental flow problem PLOS ONE7 e45814 (2012) doi 101371journalpone0045814pmid 23029257
127 A Selvakumar T P OrsquoConnor S D Struck Role of streamrestoration on improving benthic macroinvertebrates and in-stream water quality in an urban watershed Case studyJ Environ Eng 136 127ndash139 (2010) doi 101061(ASCE)EE1943-78700000116
128 R W Lammers B P Bledsoe What role does streamrestoration play in nutrient management Crit Rev EnvironSci Technol 47 335ndash371 (2017) doi 1010801064338920171318618
129 H E Golden N Hoghooghi Green infrastructure and itscatchment-scale effects An emerging science WIREs Water5 e1254 (2018) doi 101002wat21254 pmid 29682288
130 M R Williams G Bhatt S Filoso G Yactayo Streamrestoration performance and its contribution to theChesapeake Bay TMDL Challenges posed by climate changein urban areas Estuaries Coasts 40 1227ndash1246 (2017)doi 101007s12237-017-0226-1
131 R M Fanelli K Prestegaard M A Palmer Freshw Sci101086706072 (2019)
132 B W Abbott et al Unexpected spatial stability of waterchemistry in headwater stream networks Ecol Lett 21296ndash308 (2018) doi 101111ele12897 pmid 29282860
133 B L Brown C M Swan Dendritic network structureconstrains metacommunity properties in riverineecosystems J Anim Ecol 79 571ndash580 (2010) doi 101111j1365-2656201001668x pmid 20180874
134 C M Swan B L Brown Metacommunity theory meetsrestoration Isolation may mediate how ecologicalcommunities respond to stream restoration Ecol Appl 272209ndash2219 (2017) doi 101002eap1602 pmid 28718193
135 N Mouquet D Gravel F Massol V Calcagno Extending theconcept of keystone species to communities and ecosystemsEcol Lett 16 1ndash8 (2013) doi 101111ele12014 pmid 23062191
136 A Ruhiacute T Datry J L Sabo Interpreting beta-diversitycomponents over time to conserve metacommunities inhighly dynamic ecosystems Conserv Biol 31 1459ndash1468(2017) doi 101111cobi12906 pmid 28188969
137 L Yamane L W Botsford D P Kilduff Tracking restorationof population diversity via the portfolio effect J Appl Ecol55 472ndash481 (2018) doi 1011111365-266412978
138 D W Braidwood M A Taggart M Smith R AndersenTranslocations conservation and climate change Use ofrestoration sites as protorefuges and protorefugia RestorEcol 26 20ndash28 (2018) doi 101111rec12642
139 Y Wang D Wang Q W Lewis J Wu F Huang A frameworkto assess the cumulative impacts of dams on hydrologicalregime A case study of the Yangtze River Hydrol Processes31 3045ndash3055 (2017) doi 101002hyp11239
140 J L Sabo et al Pulsed flows tributary inputs and food-webstructure in a highly regulated river J Appl Ecol 551884ndash1895 (2018) doi 1011111365-266413109
141 C Magliozzi G Coro R C Grabowski A I PackmanS Krause A multiscale statistical method to identifypotential areas of hyporheic exchange for river restorationplanning Environ Model Softw 111 311ndash323 (2019)doi 101016jenvsoft201809006
142 S S Schwartz B Smith Restoring hydrologic function inurban landscapes with suburban subsoiling J Hydrol 543770ndash781 (2016) doi 101016jjhydrol201610051
143 C J Walsh T D Fletcher A R Ladson Stream restoration inurban catchments through redesigning stormwater systemsLooking to the catchment to save the stream J N AmBenthol Soc 24 690ndash705 (2005) doi 10189904-0201
144 N M Schmadel et al Thresholds of lake and reservoirconnectivity in river networks control nitrogen removalNat Commun 9 2779 (2018) doi 101038s41467-018-05156-x pmid 30018449
145 C D Sayer Conservation of aquatic landscapes Pondslakes and rivers as integrated systems WIREs Water 1573ndash585 (2014) doi 101002wat21045
146 R G Luthy D L Sedlak M H Plumlee D Austin V H ReshWastewater‐effluent‐dominated streams as ecosystem‐
management tools in a drier climate Front Ecol Environ 13477ndash485 (2015) doi 101890150038
147 D W Roscoe S G Hinch Effectiveness monitoring of fishpassage facilities Historical trends geographic patterns andfuture directions Fish Fish 11 12ndash33 (2010) doi 101111j1467-2979200900333x
148 C M Bunt T Castro-Santos A Haro Performanceof fish passage structures at upstream barriers tomigration River Res Appl 28 457ndash478 (2012)doi 101002rra1565
149 D Weigel et al Evaluation of a trap-and-transport programfor a threatened population of steelhead (Oncorhynchusmykiss) Conserv Genet 20 1195ndash1199 (2019) doi 101007s10592-019-01200-5
150 M H Bond et al Combined effects of barge transportationriver environment and rearing location on straying andmigration of adult Snake River fall-run Chinook SalmonTrans Am Fish Soc 146 60ndash73 (2017) doi 1010800002848720161235614
151 A C Ritchie et al Morphodynamic evolution followingsediment release from the worldrsquos largest dam removalSci Rep 8 13279 (2018) doi 101038s41598-018-30817-8pmid 30185796
152 T Sommer et al Californiarsquos Yolo Bypass Evidence thatflood control can be compatible with fisheries wetlandswildlife and agriculture Fisheries 26 6ndash16 (2001)doi 1015771548-8446(2001)026lt0006CYBgt20CO2
153 G Grill et al Mapping the worldrsquos free-flowing riversNature 569 215ndash221 (2019) doi 101038s41586-019-1111-9pmid 31068722
154 G Gassert M Luck M Landis P Reig T Shiao ldquoAqueductglobal maps 21 Constructing decision-relevant global waterrisk indicatorsrdquo Working paper World Resources Institute(2015) wwwwriorgpublicationaqueduct-global-maps-21-indicators
155 A Ruhiacute J D Olden J L Sabo Declining streamflow inducescollapse and replacement of native fishes in the AmericanSouthwest Front Ecol Environ 14 465ndash472 (2016)doi 101002fee1424
156 T C Gouhier A Grinsted V Simko R package Biwaveletconduct univariate and bivariate wavelet analyses (2018)
157 T Rodriacuteguez-Castillo E Esteacutevez A M Gonzaacutelez-FerrerasJ Barquiacuten Estimating ecosystem metabolism toentire river networks Ecosystems (2018) doi 101007s10021-018-0311-8
ACKNOWLEDGMENTS
We greatly appreciate those who read and provided importantinsights on drafts of this paper A Armstrong K HondulaJ Hosen K Leathers G de Mendoza C Maietta I MunozJ Tonkin and two anonymous reviewers We also thank T Tu forhelp with Fig 1 and S Carlson L Comte T Datry J HosenM Kondolf N Jones D Lytle E Moody J Olden M PowerJ Sabo and M Sanchez-Montoya for valuable discussionsCompeting interests The authors have no competing interestsFunding MAP was supported by funds from the NSF (1639145)and AR was supported by funds from the University of CaliforniaBerkeley and NSF (1802714)
101126scienceaaw2087
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 13 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
restorationLinkages between flow regime biota and ecosystem processes Implications for river
Margaret Palmer and Albert Ruhi
DOI 101126scienceaaw2087 (6459) eaaw2087365Science
this issue p eaaw2087Scienceflow variability and achieve hydrological connectivity between a river and its surroundingsand ecosystem processes To be successful such efforts must go beyond accounting for flood pulses to restore natural mimic ecologically important aspects of natural flow regimes guided by insights into how variations in flow affect biotawater quality and ecological processes In a Review Palmer and Ruhi explain how restoration designs now attempt to
Human activities have altered the flow regimes of many of Earths rivers with negative impacts on biodiversityRiver restoration guided by research
ARTICLE TOOLS httpsciencesciencemagorgcontent3656459eaaw2087
REFERENCES
httpsciencesciencemagorgcontent3656459eaaw2087BIBLThis article cites 147 articles 7 of which you can access for free
PERMISSIONS httpwwwsciencemagorghelpreprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAASScienceScience 1200 New York Avenue NW Washington DC 20005 The title (print ISSN 0036-8075 online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
Science No claim to original US Government WorksCopyright copy 2019 The Authors some rights reserved exclusive licensee American Association for the Advancement of
on May 27 2020
httpsciencesciencem
agorgD
ownloaded from

population ecology [Moran theorem (28)] andcould represent an underappreciated emerg-ing risk in fresh waters
Abiotic and biotic influences of flow
Stream ecologists traditionally interpreted thedynamics of communities as a deterministic resultof biotic interactions or as a stochastic conse-quence of the environment (29) However researchhas shown that these fluctuate in relative impor-tance over space and time andmay even influenceeach other For instance biotic interactions mayoccur only under particular environmental con-ditions (30) Inferences gathered from long-termdata combined with time-series methods importedfrom econometrics can help elucidate when and
where abiotic influences have primacy For exam-ple multivariate autoregressive state-space modelsapplied to long-term flow and fish communitydata showed that interannual variation in flowanomalies were the ultimate driver of the dy-namics of a desert stream fish community (31)Although non-native and native fishes interactedsuch effects were small compared with the directeffects of flows on the native component Bioticinteractions that emerge from known species-specific relationships between components of theflow regime such as flood timing and popula-tion dynamics can also be predicted from modelsimulations of community trajectory (30) A recentapplication of these interaction-neutral modelsfound that flows departing from the natural reg-
ime simplified interaction networks among plantguilds in desert rivers (32)A fundamental challenge for community eco-
logy for decades has been quantifying the rela-tive influence of abiotic (flow-driven) and biotic(flow-mediated) effects This is important for theaccurate design and prediction of restoration out-comes including for example anticipatingwhenflow alteration facilitates species invasions bycreating new niches (33) by enhancing propagulepressure or eroding native enemies (34) or by acombination of thesemechanismsDistinguishingthese could help managers compare the benefitsof eradicating non-natives relative to restoringecologically important facets of the flow regime
Flow alteration filters species and traits
Trait-based approaches are becoming increasinglypopular in hydroecology because they enhancecomparison of flow-ecology relationships acrossclimates (35) they lead to predictions about howcommunity change may affect ecosystem func-tioning (36) and they identify the mechanismsthat filter or promote particular taxa under stress-ful conditions such as drought (37) In the case offlow intermittency traits can be used to identifycommunity tipping points by connecting local en-vironmental conditions to organisms with traitsthat allow them to persist in isolated pools or wetsediment or connecting network-level water con-ditions to the ability of highly resilient organismsto drift or actively recolonize from perennial re-fugia (38) Traits can also be used to assess re-storation trajectoriesmdashfor instance by assessingfluctuations in the share of strategies that representdifferent life-history trade-offs [eg species limitedby reproductive capacity or by resources (39)]By studying a communityrsquos taxonomic char-
acteristics (eg species composition) and func-tional characteristics (eg dispersalmodes feedingstrategies) ecologists can determine to whatextent trait redundancy across speciesmay ensureagainst loss of functional diversity For examplein a study on Alpine stream communities subjectto climate change and flow regulation functionaldiversity increased while functional redundancydecreased (40) This finding indicates that ongoinghydroclimatic change can reduce the ability of acommunity to withstand further alteration evenif trait diversity is high because the persistenceof individual traits may be linked to the fate of afew sensitive species In an experimental studycommunities affected by drought presented com-parable taxonomic and functional decays implyingthat these communities were both taxonomicallyand functionally vulnerable (41) The risk of com-munity collapse in response to flow alterationalso depends on the way in which ldquowinningrdquo andldquolosingrdquo traits co-occur in a given species andacross species in a community as a result of aparticular stress Thus community-wide responsesto a particular alterationmay be stronger if speciesaremaladaptive to that change for instance bybeing large-bodied longer-lived and collector-feeding (filtering particulate organic matter) ina scenario of increased flow variability (18) Under-standing how flow alteration impairs species with
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 3 of 13
Nutrient cycling
Primary production
Energy inputs
Secondary production Dispersal refugia Invasions
Food-web interactionsMetabolic regime
Hydrologic connectivity
Streambed sediments
Water chemistry temperature
Channel form riparian vegetation
Decomposition Species demographic rates
River ecosystem integrity
Altered flow regimes
Restored flow regimes
Time
Restoration toolboxAlter pattern of reservoir releasesImprove stormwater controlReforest and restore watersheds
Disc
harg
e w
ater
leve
l Muted high flows
Storm flows
Altered periodicity
Flow homogenization
Shifting baselines
Extreme low flows
Fig 2 Process-based restoration posits that actions must target processes not patternsthat have been degraded River ecosystems are sustained by a host of environmental conditionsor ecosystem structure (gold rectangles) which in turn influence ecosystem processes (greenrectangles) and biotic processes (blue rectangles) These processes as well as ecosystem structureare all controlled by the flow regime Thus correcting facets of flow regime alteration may enhanceriver ecosystem integrity in both direct and indirect ways
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
traits that have effects on ecosystems (eg byparticular feeding strategies) will help anticipatethe risks of losing major ecosystem functionssuch as detrital decomposition (36)
How does flow variation control riverecosystem processes
Rivers produce transform and store organicmatter (OM) and nutrients Carbon (C) and nu-trient concentrations and fluxes vary longitudi-nally and laterally as river networks expand andcontract with rain These variations influenceand are influenced by stream ecosystem metab-olism which can be described as net ecosystemproduction or the difference between gross pri-mary production (GPP) and ecosystem respiration(ER) The metabolic balance between heterotro-phic and autotrophic production that providesresources for consumers can shift seasonally orwith changing flows as can hydrologic connect-ivity that influences inputs of OM and solutes torivers (42) Increasing availability of sensors andadvances inmodeling (43) havemade the study ofmetabolism OM processes and solute dynamicsripe for linking to biotic processes
Flow variability andecosystem metabolism
Spatiotemporal variability in ecosystem processesis a fundamental characteristic of rivers and flowregime is a major driver of this variability (Fig 4)Recentwork shows thatmany rivers in theUnitedStates have strong seasonal patterns in primary
production with spring peaks for larger riversand summer peaks for smaller rivers generallyGPP is negatively correlated with high variationin daily discharge (44) In regions with little sea-sonality andor continual canopy cover GPP islower and its patterns are less predictable (44)Bernhardt et al (45) provide a comprehensivereview of stream metabolism the factors thatinfluence it and why it is difficult to predict Flowhas the largest effect at the extremes such that atlow flowswhen the streambed begins to dry largedrops in GPP occur (46) Storms also often reduceGPP (45) However patterns may vary as a func-tion of flow predictability For example researchin a river characterized by predictable peaks dur-ing spring snowmelt found over two seasons thatGPP rose during spring snowmelt sampling (47)whereas high flows during the winter and sum-mer reduced GPP Within a year this patternresulted in a switch from net autotrophic to netheterotrophic productionThe paths that water takes as it moves to and
within a streammdasha spatial aspect of its flowregimemdashinfluencesmetabolismBycouplingmetab-olism and chemistry time series with informationonhydrologic connectivity and flowpaths ERwasshown to increase in an organic-rich watershedduring peak flows and remained elevated forweeks (48) Sustained hydrologic connectivitywith soils likely fueled inputs of dissolved orga-nic carbon (DOC) that promoted bacterial respi-ration Such inputs can lead to nonlinear responsesinmetabolism For example increasing flows have
led to decreases in GPP up to some thresholdflow beyond which GPP was suppressed by re-duced water clarity from flow-elevated humicDOC (49) In the same vein higher riverine GPPhas been associated with reduced suspendedsediments and high temperatures during low-flow droughts (50) When flows cease entirely andsections of riverbeds dry up carbon dioxide fluxesto the atmosphere can be far higher than those inflowing sections of the channel (51) presumablybecause of spikes in aerobicmicrobial respirationhowever as is the case with methane in riverspartitioning fluxes of these gases to changes inmetabolism (aerobic or anaerobic) is difficult par-ticularly under variable flows (52)
Hydrologic connectivity drivessolute dynamics
Research is growing on how hydrologic connect-ivity to hillslope soils influences water chemistry(53) and how variation in discharge and riverstage influence the strength of bidirectional waterexchanges that affect nitrogen (N) uptake reten-tion and removal (54) Periods of hydrologic con-nectivity may be followed by long periods withlimited exchange which may differ greatly alongthe stream corridor This can lead to complex CN and phosphorus (P) dynamics that influenceor are influenced by microbial and algal produc-tivity When connectivity with watershed solutesources is extensive inputs of solutes such asDOCmay be so high that they exceed the ability of riverecosystem processes such as microbial uptake toregulate their fluxes (55) Theory posits that largestorms pulse terrestrially derived dissolved or-ganicmatter (DOM) into streams and it is quicklyshunted downstream by high flows where it isprocessed in larger parts of the network (56)However how far DOM in pulsedwater is trans-ported is governed not only by discharge but bythe composition of DOMUnderstanding the rela-tionship between DOM sources and its composi-tion is being advanced by combining time-seriesdata with isotopic information to evaluate spatialvariability in hydrologic flow paths transit timeand chemistry as watermoves from the terrestrialto aquatic realms (53 57) It is difficult tomeasurebiotic uptake in streams during storms howeverrecent work suggests that assuming storm flowstransport DOM too quickly for in-stream process-ing may be a poor assumption if the DOM com-position renders it readily utilizable by biota (58)Although hydrologic connectivity is not a newconcept it is not typically considered part of theflow regime yet it can be characterized by itstiming duration and frequency and then linkedto stream processes or patterns Newmethods forquantifying connectivity [eg (59)] are emergingand the energetic and water quality importanceof network connectivity to soils and diverse land-scape elements (such as wetlands and ponds) isbecoming a major area of research (42 59)
Hydrology influences decompositionvia consumers
Detritus (or decaying OM) plays key roles instream ecosystems and most consumers even
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 4 of 13
33104
10103
32101
10100
3110-2
9810-4
3110-5
1922 1949 1976 2004
4
16
256
4096
Perio
d (d
ays)
1963Closure of Glen Canyon Dam
Fig 3 Flow regime shifts in the Colorado River at Lees Ferry Arizona over the past 97 years(1922ndash2018) The flow regime changed drastically around 1963 with the closure of Glen CanyonDam Natural seasonality was dampened and artificial weekly and subweekly scales emerged Graydashed lines indicate 7-day flow cycles resulting from flow management for hydropower andyearly flow cycles resulting from extended base flows and spring snowmelt floods Colors representwavelet power and confidence level contours identify statistically significant power A very wetperiod in the mid-1980s and the experimental releases that took place after 1996 restored flowevents but not the regime Across dammed rivers of the American Southwest muted flowseasonality has opened niches for non-natives particularly organisms adapted to living in stableresource-limited environments at the expense of many native fishes adapted to highly variable(periodic or stochastic) flow conditions that characterized free-flowing hydrographs (155) Plotcreated with the biwavelet R package (156) using mean daily discharge data from United StatesGeological Survey station 09380000 [Dam logo US National Park Service]
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
apical predators are at least partially supportedby it (60) Macroconsumers break up particulateorganic matter (POM) into small pieces enhanc-ing the overall mineralization rate by bacteriaand fungi and even moderate flows may resultin export of OM particularly fine POM resultingfrommacroconsumer shredding activities Over-all howevermicrobes largely drive C losses throughrespiration (61) and although nutrient availa-bility temperature and POM composition (egleaf type) influence this how flow mediates rela-tionships has been more difficult to pin downRecent work (62) designed to separate the effectsthat different components of the flow regimehave on the mechanisms driving decompositionin perennial streams found that dissolution andmicrobial processing consistently exceeded theeffects of fragmentation and microbial pro-cessing was best explained by daily variabilityin discharge When flow regimes become in-termittent the lack ofwater can impedemicrobialand consumer colonization of POM resulting indecreases in decomposition the extent of whichis related to the duration of the drying (46 63)Consistent water cover is therefore importantto decomposition and thus shifts toward lowerflows when litter inputs are high (eg in thefall) can have a large impact on consumers andassociated food webs (63 64)
When flows are altered are changes inbiota and ecosystem processes linked
To help ground this question in the world ofrestoration practice we begin with a focus ontwo of the most common forms of river flow al-teration damming and urbanization For theformer we emphasize hydropower dams becausethey are growing in number they strongly alterflow regimes and the mitigation of their impactshas spurred the field of environmental flow re-search We also focus on urban streams becausestudies on the impacts of imperviousness havebeen extensive and these streams have been the
target of numerous restoration efforts We summa-rize knowledge on the effects of flow on organ-isms ecosystem processes and their interaction inthese two types of river systems and then describehow this can inform river restoration in general
Urbanization and dams changeflow regimes
With few exceptions flow regimes below hydro-power dams and in urban streams are highlyaltered at one or both ends of the flow spectrumRegimes in impounded rivers are typically lessseasonal than daily rainfall would suggest par-ticularly below large hydropower dams with highwater-storage capacity rather than below smallerless impactful run-of-the-river facilities (65)Hydro-power dams canmute peak river flows but power-generation cycles induce rapidly fluctuating flowpatterns (hydropeaking) that often occur onhuman-relevant scales [ie hours to weeks (18)]tending to bemore pronounced in seasons of highenergy demand Hydropeaking is often accom-panied by spikes in temperature (thermopeak-ing) either cold or warm depending on seasonand reservoir operation (66) Thus downstreamorganisms and ecosystems are generally subjectto multiple periodic stresses In urban streamsbaseflows can be higher or lower depending onwatershed characteristics and infrastructurehowever peak flows are invariably larger thanthose prior to urbanization Understanding theecological impacts of urban stormflow is com-plicated by indirect effects on water quality In-creases in flow magnitude frequency and ratesof change during storms leads to elevated con-centrations of pollutants Despite this studiesseparating the causes of biological impairmenthave reported that the unique effects of alteredurban flows can be significant (67 68) Spikesin stream temperature caused by runoff fromhot pavement (69) add to stress on the biotaHydropeaking and urban stormflows might
be expected to reduce primary production by
scouring algae (70) yet some studies have re-ported elevated GPP or alternating periods ofnet autotrophy and heterotrophy (71ndash73) Lightcan override the effects of flow on metabolismby enhancing algal photosynthesis (74) how-ever if hydropeaking increases turbidity GPPmay decline (73) In urban streams elevatedGPP is often attributed to open canopies andorto elevated nutrient concentrations both of whichoften co-occur with their altered flow regimes(72) As discussed later such shifts in primaryproductivity have been linked to changes introphic structure eg more algal-based foodwebs (75) depending on the relative availabil-ity of terrestrial carbon inputs (both DOM andPOM) (76) Inputs of POM and its quality as afood resource changewith landscape context (egvegetative cover) and catchment hydrology (77)but POM availability can also be limited by flowvariability which can increase decompositionrates and decrease residence times (78 79)As is the case for running-water systems in
general high flows in urban streams bring pulsesof solutes including N DOM and various pol-lutants (80) Solute dynamics and biogeochemicalprocesses in regulated rivers have been insuffi-ciently studied with researchers focusing moreon such processes in reservoirs rather than indownstream river ecosystems We found nostudies attempting to link urban or regulatedflow regimes and solute fluxes to changes infood webs or stream metabolic balance How-ever recent studies show shifts in microbial tax-onomic and functional composition in responseto pulsed urban flows (81) and thesemay help inexploring a potential link but are complicated bythe fact that non-flow stressors affect consumercomposition For example urban and regulatedrivers typically have animal communities thatare less diverse and composed of tolerant groupsoften non-native Hydropeaking and urban stormflows can certainly cause direct mortality via dis-placement and transport to unsuitable habitatsbut many of the organismal effects are realizedthrough indirect pathways For instance hydro-peaking reduces the viability of river-edge egg-laying specialists by limiting their access to suitablespawning habitat disrupting reproductive successand insect emergence (82) Flow regulation caneven create ecological traps if organismal phenol-ogy shiftswith thenovel environmental conditionsFor example dam-induced warming triggeredsummer diapause in a mayfly resulting in theloss of its last generation (83) Dams also isolatepopulations harming organisms that requiremigrations to complete their life cycles whetherthese are between riverine and marine habitatsor within the freshwater domain By analogy al-tered flow regimes in urban streams can also leadto isolated pools in streams with lowered base-flows This can be associated with periods ofhypoxia punctuated by extreme flow pulses inwhich these streams become ldquoscoured or suffo-catedrdquo (74)As illustrated using urban streams and reg-
ulated rivers research is moving forward andhints at the three-way nexus but is clearly still
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 5 of 13
Fig 4 Ecosystem metabolismpatterns across a river networkSpatial patterns in river eco-system metabolism for the Deva-Cares catchment in northernSpain as represented by the ratioof gross primary production(P) to ecosystem respiration (R)A PR gt 1 indicates autotrophicprocesses dominate so theseriver sections are accumulatingor exporting organic carbonPR lt 1 indicates dominanceby heterotrophic processes inriver sections that are receivingorganic C input (eg fromterrestrial sources) The boxesindicate regions of the networkimpacted by human pressures including deforestation (brown box) waste products from urbansites (green box) and a combination of the two (purple box) Such visualizations are usefulto managers in understanding regions of concern that may need the most restoration actions[Reproduced with permission from (157)]
PR ratiolt025
gt150
025 ndash 050 050 ndash 075075 ndash 100100 ndash 150
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
inadequate to meet restoration needs Studieson ecosystem processes below dams are parti-cularly limited and much of what we knowcomes from only a few river systems Furtherpublished work focuses primarily on benthicecosystem processes immediately below damsrather than riverine planktonic realms whereproductivity in large regulated rivers can beimportant
Linking flow biota andecosystem processes
Research on flow regimes that bridges organis-mal and ecosystem ecology has traditionallyrelied on states instead of rates ie nutrientconcentrations rather than uptake or fluxesstanding stocks of algae rather than productivityor metabolism and abundance of organismsrather than secondary productivity This workgenerally shows that flow variability suppressesand stability increases algal standing stock andgrazer densities with peaks in invertebrate bio-mass following those of algae [eg (84 85)]Over the past decade research began to focuson how flow regimes alter functional aspects offoodwebs For example biofilms inMediterranean-climate rivers are thicker andmetabolicallymoreactive downstream of flow-stabilizing dams (71)When flow variability increases below hydro-power dams fluctuations can reduce biofilmdevelopment and its quality as a food resource(86) Using experimental flumes subjected tohydropeaking shifts have been found in periph-yton fatty acid content from important highlyunsaturated compounds to nonessential saturatedones (87)Indirect evidence suggests that flow-induced
changes in dominant resource type (eg algaeversus detritus) resource quantity (biomass) orresource quality (nutrient content) can propa-gate to higher trophic levels via bottom-up ef-fects For instance detritivorous invertebrateswere found to dominate species richness andinvertebrate biomass upstream but not down-stream of an irrigation dam (75) diets of om-nivore insects indicated that flow stabilizationshifted a detrital into an algal-fueled food webSimilar evidence comes from Glen Canyon Damin the Colorado River (88) Near the dam whereprimary production is high (73) food webs aresimplified and gut content analyses have sug-gested algal production supports gt50 of in-vertebrate and gt70 of fish production andthe food web can be easily perturbed by flooddisturbance Farther downriver food webs showincreased reliance on detrital resources havehigher trophic efficiency and are more complexand more resistant to disturbance (76) In con-trast recent work on seasonal tropical riversfound algal productivity and zero-flow distur-bance did not explain food-web structure Inthis case it seems that highly mobile predatorsbuffered the local effects of drought (89) Thisstudy along with earlier ones at the site (90) isof particular interest because it links temporalpatterns in predatory fish with flow-dependentavailability of resources
Only a few studies have provided evidence oftop-down controls on foodwebs being influencedby flow variation Fish in a tropical floodplainriver were found to exert top-down control onprimary producers but only under certain hydro-logic conditions (91) Similarly in aCalifornian riverwinter flood pulses were shown to control insectconsumers late in the season ultimately deter-mining whether fish exerted top-down controlon the lower trophic levels (10) These illustratethe importance of pulsed hydrology on resource-consumer linkages and show that not just standingstocks but also the dominant controls structuringfood webs may fluctuate over time in response toflow variationAn indirect line of evidence supporting the
importance of the flow-biota-ecosystem processesnexus comes from theoretically oriented researchon food-web structure Changes in productivityhabitat size and disturbance underpin the basisof the three main hypotheses of food-chainlength which state that chains should be longerin stable productive large ecosystems (92) No-tably flow regimes influence these three con-trols Because food-chain length determines keyaspects of a river ecosystem such as the risk oftrophic cascades or the pollutant accumulationin top predators understanding how it is af-fected by flow alteration is important How-ever understanding the interplay between thehypotheses of food-chain length and the flow-dependent biological mechanisms that allowfood webs to shift in structure (eg predatorinvasions local extinctions consumers chang-ing diets) is still a relatively recent endeavor(93ndash95) Evidence suggests that zero-flows short-en food chains mainly by keeping streamsfishless (94 96) whereas stabilization of variableflows can lengthen food chains by allowing om-nivores to feed on abundant high trophic levelprey (95)New studies are also focusing on interactions
between drivers of food-web structure that hadbeen previously only been examined in isolationsuch as disturbance regimes and ecosystem sizeboth influenced by flow regime Using metabolictheory predator biomass has been shown toscale with both prey resources and fluctuatingstream habitat size with reductions in preda-tor size leading to declines in predator biomasssupported per unit of prey biomass (97) Movingforward the combination of habitat andmetabolicconstraints on predators may provide a basis toconnect flow-induced disturbance changes inecosystem processes and habitat fluctuationsto riverine food-web structure The metabolictheory of ecology (98) is a unifying frameworkthat allows scaling responses of organisms towhole ecosystems andmay become particularlyuseful in situations where flow alteration affectsorganismal body size or water temperature Insuch cases strong changes in metabolic ratescan be expected affecting resource uptake andorganismal growth to ecosystem-level produc-tion and respirationFinally an important body of food webndashrelated
research that provides strong inferential evidence
for the nexus comes from studies on animal-mediated nutrient cycling and ecological stoi-chiometry Theory suggests detrital and algalfood websmay be linked by cascading effects onnutrient cycling (99) and empirical work showsthat organismal excretion andmigration (whichrelocates C) may be important to this link (100)(Fig 5) A recent study (101) showed that ex-cretion of highly labile dissolved organic C ni-trogen (N) and phosphorus (P) by aquatic insectsmay be sufficient to support a substantial fractionof microbial energy and nutrient needs Giventhat C availability can be too low tomeetmicrobialenergy needs [eg (102 103)] and that microbialactivity enhances detrital quality to consumers(104) a C subsidy from excretory processes couldbe important at least during low-flow periodsAdditionally microbial use of excreted dissolvedorganic nitrogen (DON) and DOP may releasealgae from potential competition with microbesfor inorganic N and P if this is the case GPPshould increase Ecological stoichiometry mayalso be important for understanding constraintson organismal growth rates as rapid growthdisproportionally increases demand of P [as it isused in ribosomal RNA (105)] This could in turnlimit the ability of organisms to grow fast andexploit temporary habitats Interestingly elemen-tal ratios can vary interspecifically in response totemperature or predation pressure (106)mdashfactorsthat increase with low flows and habitat fragmen-tation Overall studies that link biota with bio-geochemical processes remain rare (107) butanalyzing flow regimes as a driver of elementturnover rates may provide a way forward
Can hydroecological advances informrestoration practice
Shortfalls in the outcomes of river restorationhave prompted calls to identify and restore pro-cesses that support and sustain biological com-munities rather than focusing only on rivergeomorphology and habitat In urban streamsscientists have critiqued an overemphasis onstructural engineered approaches (7 108) thatdo little to restore the full flow regime (109) Inlarge regulated rivers identifying and restoringimportant features of the flow regime via damoperations is often possible (albeit expensive)However that does not solve other problemsassociated with dams such as habitat frag-mentation (110) altered sediment and thermalregimes (111 112) or disrupted biogeochemicalprocesses (113) Overall efforts to alter reservoirreleases or improve urban stormwater infra-structure to partially mimic a riverrsquos natural flowregime have great potential Technologies arebeing evaluated for real-time management ofurban flows and novel ways to design flow re-leases from dams are being proposed (114 115)however research still lags far behind the needfor solutions Importantly the paucity of studieson the flow-biota-ecosystem processes nexus pointsto a need for more integrative research connectinghydrologists population and community ecologistsand ecosystem ecologists in the context ofrestoration
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 6 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
Restoring flow regimes ratherthan eventsScientifically informed flow designs may existthat maintain ecosystem integrity in regulatedrivers (116 117) but these have produced mixedresults to date In the dam-altered Putah Creekin California mimicking the natural timing ofhigh and low flows enhanced native fish by im-proving their spawning habitat reducing watertemperatures maintaining flowing conditionsand reducing the abundance of non-native spe-cies (118) However other similar attempts havebeen less successful In some cases non-nativefish density also benefitted from flow patternsmimicking an unregulated regime (119) in othersnon-natives bounced back shortly after the flowrelease (120) These experiences illustrate thateven natural flow features may have inadvertentnegative effects if only part of the community orecosystem is considered Models for the NavajoDam (San Juan River New Mexico) show thatadequately timed releases could simultaneouslyenhance native and minimize non-native fishpopulations (121) thus predicted benefits couldsurpass those of mimicking a natural flow re-gime potentially persisting even during dryyears Another study using the ldquodesignerrdquo flowparadigm (114) attempted to understand howdifferent facets of flow variability control fisheryproduction in the Lower Mekong floodplains Time-
series models combining discharge and fisherycatch suggested that enhanced flood pulses andlong inter-flood intervals may increase fisheryyieldsAlthough environmental flow practices have
mostly focused on re-creating discrete flowevents (eg flood pulses) these can still serve asvaluable experiments An iconic example of alarge-scale flow release is Minute 319 an amend-ment to the United StatesndashMexico treaty allow-ing opening of floodgates in the Lower ColoradoRiver and a one-time rewetting of the parchedColorado River Delta (122) Although this pulsed-flow experiment did not promote establishmentof native woody riparian seedlings such trialscan inform future restoration practices for ex-ample by determining whether the provision ofbare moist substrate enhances seed germina-tion (123) Hypothesis-driven experimental flowreleases combined with deep knowledge of tar-get ecosystems can provide insights into resto-ration actions [eg (124)] Still an outstandingquestion is how flow regime restoration couldbe leveraged to restore other important alteredaspects of the ecosystem such as sediment andthermal regimes For example cold-water releasesby existing dams in climate-vulnerable basinscould prevent organisms that thrive at low tem-peratures from becoming replaced by general-ist warm-tolerant taxa (125)
Although the concept of environmental flowsis generally associated with impounded riversits relevance to urban streams is clear (126)However because frequent high storm flowsare viewed as the problem reducing them gen-erally becomes the only restoration goal This isoften attempted by shifting the spatial distribu-tion of velocities within channels through ma-nipulation of channel sinuosity installation offlow-diverting weirs or adding other structuresThese approaches may reduce local erosion butrarely result in levels of water quality ecosystemprocesses or biodiversity in urban streams thatcome close to those of unimpacted streams(127 128) Focusing on the channel ratherthan the watershed context at large means thatkey ecosystemprocesses can remain compromised(129) A recent study compared time series ofmetabolism in two streamsmdashonewith stormwaterponds in the watershed and the other restoredby altering channel slope banks and in-streamstructural complexity (103) The study found thatpeaks in GPP and ER in the ldquorestoredrdquo streamwere higher more prone to resets induced byflashy storms and that recovery time scales werelonger compared with the stream in a watershedwith stormwater measures (Fig 6) Ponds areone type of nature-inspired green infrastructureused to restore streamflow tomore natural condi-tions tree plantings constructed wetlands andgrass swales can in turn replace gray infrastruc-ture such as concrete tunnels that capture andstore waterDistributing green infrastructure in water-
sheds helps restore streams because it targetsthe cause of streamflow alteration includinginsufficient infiltration groundwater rechargeand more generally limited watershed storagecapacity (129) By tackling the cause rather thanthe symptoms of flow alteration these practicescan reduce ecological impacts that result fromthe cascading effects of altered flows includingpoor water quality and temperature spikes Re-search on the effectiveness of green infrastructureon restoring streams is sparse and limited bylack of data before and after implementationalthough studies are beginning to appear Forexample upland green infrastructure in theform of shallow wetland-like pools that capturestormwater and reconnect it to the groundwaterhas been shown to enhance baseflows while re-ducing peak flows and pollutant loadsmore thanrestoration structures within the mainstem ofthe receiving stream (130) Similar structuresbuilt between stormwater outfalls and perennialstreams did not modulate the timing magnitudeor duration of urban storm responses nor didthey improve water quality or biodiversity (131)illustrating the need for researchers to understandhow exact design and watershed context influ-ence performance
Spatiotemporal variability andrestoration designs
Research showing how local and watershed-scale environmental drivers vary temporally andalong river networks in ways that shape biota
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 7 of 13
Flow pulses (storms) flashy flowsDroughts intermittencyLowstable flowsAltered timing seasonality
DOC N P
DetritusMicrobial
stoichiometric constraints
Excretion exudates
Detrital pathway
Algal pathway
Algal stoichiometric constraints
Organismal dispersal source-sink dynamics
Algae
Community replacement
ndash ndash +
ndash ndash + ndash
+
+ndash +
Invasions
Turnover
Consumer stoichiometric constraints
Fig 5 Flow variation affects biota and ecosystem processes and food webs integrate bothHydrologic connectivity along river networks is a first landscape-level filter constraining dispersal oforganisms in river networks Extreme flow variation (floods and droughts) are a source of disturbanceto all organisms (gray box) but particularly to the higher trophic levels given their higher metabolicdemands In turn flow variation controls inputs of nutrients and terrestrial leaf litter to the stream whichinfluences microbial activity and the relative contributions of the detrital (brown) and algal (green)pathways in sustaining aquatic consumers Not just quantity but also food quality (eg CN and CPratios) influence dietary choices of consumers and thus trophic transfer efficiency Finally alteration offlow regimes may influence all these compartments and may also open niches for new players Ifbiological invasions occur at the top of the food web these have the potential to divert energy away fromnative predators and to alter food-web structure through top-down controls
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
and ecosystem processes can help prioritizeselection of sites for restoration For exampleusing the concept of river network saturation(55) to identify river sections where in-streamremoval of nutrients cannot keep up with highsource inputs of nutrients suggests that restorationactions targeting those sections may dispropor-tionately improve water quality Identification ofsubcatchments that disproportionately influencestream chemistry may also be informed by com-bining methods from landscape ecology andcatchment hydrology focusing on the spatialscale of variance in water chemistry (132) Head-waters are often prioritized for restoration be-
cause poor water quality in them can havedownstream impacts Research showing thatlocal environmental constraints on river com-munities are important in headwaters as well asin the larger channels but dispersal constraintsare much less important in the former (133 134)implies that headwaters should be prioritizedHowever research quantifying time-varying con-tributions of local communities to river-wide(network) biodiversity can also inform prioritiza-tion by identifying sites that host a dispropor-tionate share of species or that host uniquespeciesmdashfor example because they shelter sensi-tive taxa during critical flow bottlenecks This idea
encapsulates the concept of keystone habitatsor local sites that strongly influence metacom-munity dynamics (135) This has been appliedto aquatic invertebrates across intermittent rivernetworks by tracking time-varying site-specificcontributions to community dissimilarity (or betadiversity) (136)Other approaches to prioritize restoration ac-
tions may be based on contributions to system-wide stability rather than numbers of individualsor species For example the restoration of fall-run Chinook salmon portfolios may be assessedvia the contributions that different periods andlocationsmake tometapopulation-wide stabilityowing to spatiotemporal variation in habitatconditions (137) Additionally interventionscould promote conditions that are not impor-tant now but could be important in the future[ie proto-refugia (138)] For example resto-ration of groundwater-influenced habitats couldmitigate future stress to drought-sensitive taxaunder scenarios of increased intermittencyPatches of undisturbed land that protect flowregimes in streams within a larger catchmentthat is otherwise urbanized can be sources ofcolonists to restored sites Priorities that focuson present degradation of river sectionsmay failto address site contribution to impairment orrecovery at broader scales or under differentscenarios These insights indicate a great poten-tial for new methods to prioritize restoration ofboth urban and regulated streamsWe know little about how altered flow regimes
propagate through entire river networks butwork is starting to assess the cumulative effectsof dams on flow regimes as well as dam-levelcontributions to alteration Although these ap-proaches have largely focused on dam cascades[eg (139)] if expanded they could help identifyfavorable sections in river networks to developenvironmental flow operations Here exper-imentation could again help with testing ofnew strategies For instance large-scale flowexperiments could be designed that coordinatereleases among dams or between dams and free-flowing tributaries to enhance the restorativeeffects of natural pulses (140)
Hydrologic connectivityinfluences restoration
Recovering critical flow paths and their connec-tivity to rivers is important but it remains asubstantial challenge for flow-degraded riversand streams Whereas regulated rivers tend tohave reduced lateral and longitudinal hydro-logic connectivity urban streams have unnaturallevels of connectivity because pipes and over-land runoff route water quickly In both cases fun-damental hydroecological insights into streambedstructure landscape storage capacity and ground-water connectivity can help to envision novel re-storation approaches (130 141 142) More generallyhydrologic connectivity is becoming a frameworkto identify alternative ways to restore streams viastormwater management (143) For example arecent study suggested that reducing road densityand hydrologic connectivity between roads
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 8 of 13
ER
Disc
harg
e (f
t3 s)
GPPG
PP o
r ER
(g O
2 m-2
day
-1)
Discharge
Watershed BMPs
In-stream restoration
0
ndash4
4
0
ndash4
4
4
2
6
4
2
6
Jan Sept July May Mar Nov Jan
A
B C
Fig 6 Green infrastructure as a means to restore stream ecosystems (A) Discharge andecosystem respiration (ER more-negative values mean higher respiration) and gross primaryproduction (GPP) for two urban streams in which restoration efforts involved (top) modifications inthe stream channel and (bottom) traditional stormwater best management practices (BMPs)[Reproduced with permission from (103)] (B and C) Example of a comprehensive greeninfrastructure approach to ecologically restore an urban headwater stream that also supports thelifestyle of resident urban communities (B) Stream at the onset of restoration and (C) afterrestoration [Reproduced with permission from Greenworks Portland OR]
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 9 of 13
Table 1 Research challenges and opportunities Despite the progress that has been made in understanding how flow regimes affect biota and ecosystem
processes major challenges persist that prevent a complete understanding of the flow-biota-ecosystem processes nexus Making progress on these
challenges requires a mix of fundamental and applied research
Topic Research challenge Opportunity
Connecting
organismal to
ecosystem-level
processes
Research on the effects of flow on organisms
and ecosystem processes has followed parallel
tracksmdashhindering our mechanistic
understanding of the flow-biota-ecosystem
processes nexus
Generate theory that connects concepts and frameworks
that are well accepted in river community and ecosystem
ecology For example the notion of environmental filters
that connect regional to local composition via a combination
of dispersal and environmental conditions (community
ecology) with analogous theory on solute transport versus
reactivity (ecosystem ecology)
Our current understanding of the links between
phenology of biota and ecosystem processes
is precursory
Use high-frequency sensors to characterize metabolic
regimes at sites with ongoing organismal monitoring
Leverage citizen-science biodiversity data to detect
spatiotemporal shifts in animal phenology
(eg insect emergence fish spawning)
Use remote sensing tools to better understand
links between hydrology and aquatic and riparian
plant phenology
The link between stoichiometry and flow
variation remains largely unexplored
Repurpose data that is already being collected to test
flow-stoichiometry relationships eg studies using C and
N stable isotopes to describe CN ratios across flow
regime types Perform field experiments to quantify
indirect effects of invertebrate and fish consumption
on ecosystem metabolism
Ecological
consequences of
hydrologic
connectivity
The importance of anaerobic metabolism in
carbon processing as flows cease and parts of
streambeds dry is a major frontier research area
Create networks of sites across different geologies
and flow regimes that collect time-series data on
methane nitrous oxide and carbon dioxide fluxes
from intermittent streams
We ignore how biota in the wet phase is
influenced by processes and patterns
from the dry phase
Research the functions of dry riverbeds Use camera traps and
drone technology to investigate the role of dry riverbeds
as a dispersal corridor metabarcoding to quantify
ldquodark biodiversityrdquo in the hyporheic
Wetlands and their links to stream networks are poorly
delineated globally Even in the United States and
Europe current wetland inventories do not capture
these links yet understanding how their connectivity
to river systems influences network-scale ecosystem
processes is needed
Analyze time series from high-frequency sensors distributed in
river networks varying in wetland connectivity Further
development and application of time-series methods
used by neuroscientists
We need to identify the relationship between
varying flows and solute dynamics
associated with changes in catchment-scale
hydrologic exchanges
Design collaborative work among catchment hydrologists
and stream ecologists combining methods to estimate
transit times residence times and identify sources
across heterogeneous landscapes Link these properties to
land management and water quality aspects
Scaling and
transferring flow-
ecology relationships
across space and time
We ignore how biodiversity responds to
propagation of flow regime alteration
across river networks
Leverage extensive hydrometric networks (eg United States
Geological Survey National Water Information System)
to perform spatially replicated time-series analyses
on flow and use these to generate expectations
on how river network position should mediate
biodiversity responses to flow alteration
Flow-ecology relationships do not account for
nonstationarity and abrupt change
Use process-based models that account for mechanisms
and associated uncertainties When using time-series
data and models select methods that are robust
to nonstationarity Linear flow-ecology relationships
should not be assumed
Current flow-ecology relationships assume that
responses of species do not change over time
Consider flow regime alteration as a process with ecological and
evolutionary consequences Include intraspecific variation
in traits and potential for rapid evolution in models
continued on next page
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
and streams may improve water quality morethan other restoration options (70) Additionallyresearch on the age and lability of DOC enteringstreams has shown how precipitation or land dis-turbance can change watershed flow paths (53)and this could inspire new restoration approachesfocused on land use and soil properties Manystudies on hydrologic connectivity and networkstructure suggest a need for restoration to focuson landscape heterogeneity especially on thepresence ofwetlands lakes and ponds (144) Suchfeaturesare frequently lost inurbanizedcatchmentsand outcomes could be improved by restoring orincreasing connectivitywith these ecosystems (145)Hydrologic connectivity associated with ground-water inputs or abstractions land-use manage-ment and return flows fromwastewater treatmentplants also need to be considered in these initia-tives (146)Restoring hydrologic connectivity is also key
to achieving biodiversity outcomesmdashparticularlyin dammed systems Longitudinal connectivityfor fish inhabiting dam-altered rivers can beenhanced by the construction of fishways andsophisticated passage facilities However moni-toring data on the effectiveness of such inter-
ventions are still scarce and available evidencesuggests that passage effectivity largely dependson technology used and species-specific fish be-havior (147 148) Although trap-and-transportprograms for migratory fishes can help augmentthreatened populations (149) these practicesoften increase straying and thus the potentialfor genetic homogenization as seen from ju-venile salmon collected at dams and transporteddownstream to increase out-migration success(150) Overall a combination of engineering andmanagement solutions may be best to restorefunctional connectivity in dam-altered riversThe link between hydrologic connectivity andecological restoration is also central to con-siderations on where to build remove and re-operate hydrologic infrastructure Hundredsof dams have been intentionally removed sincethe 1970s and removal of large dams such asthe Elwha River Dam (Washington USA) hasshown that restoring longitudinal connectivitycan have immediate geomorphic effects with com-plex temporal dynamics (151) Infrastructure canalso be designed or reoperated to manage forrestoring lateral connectivity and enhanced flood-plain processes For example the Yolo Bypass in
the Sacramento River California has becomea prime rearing and migration site for chinooksalmon showing that flood control and habitatfor fish and wildlife do not need to be at odds(152) All these examples show the importanceof integrating hydrologic connectivity in ecolog-ical restoration whether the goal is to minimizethe ecological effects of fragmentation restorefree-flowing watercourses or operate green infra-structure to enhance infiltration floodplain con-nectivity or other ecological processes
Research challenges and opportunities
Two decades of concerted efforts to research theeffects of flow regime alteration have tremen-dously advanced our understanding of the waysin which streamflow influences biota and eco-system processes However important challengesremain These aremainly related to the scarcity ofscience connecting organismal to ecosystem-levelprocesses through for example microbial linksthe difficulty of studying watershed-level phys-ical and biogeochemical heterogeneity and itsinfluence on riverine processes and biota thecomplexity of parsing out the direct effects offlow from flow-mediated effects and co-occurring
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 10 of 13
Topic Research challenge Opportunity
Cascading
implications of
hydrologic
alteration
We have a poor understanding of the
interactions among physical drivers
Build on large-scale flow experiments to test hypotheses on the
combined effects of altered flow sediment and thermal
regimes Experiment with multilevel intakes in reservoirs
Promote collection of non-flow variables (eg temperature
water quality parameters) at hydrometric stations
At low flows biota are being exposed to high
concentrations of new classes of pollutants
whose impacts are poorly known especially if
biotic impacts interact with impacts on
ecosystem processes
Design realistic experimental studies to advance knowledge
on the ecological effects of multiple stressors
Collect long-term data at sites varying in contaminant
inputs and hydrologic regimes Link microbial to
organismal and ecosystem-level responses
Using hydroecological
science to inform
river ecosystem
restoration
Environmental flow science largely ignores
human dimensions
Use methods that allow identifying causal networks
in complex socioenvironmental systems Include
human needs and thresholds in research on
flow-ecology relationships
Environmental flows are generally prescribed
at the scale of single infrastructures restoration
designs and assessment are generally at reach scales
Promote coordinated large-scale flow experiments
and restoration designs at the river basin scale
Extend assessment of green infrastructure
effectiveness beyond individual structures to
infrastructure networks and link both
to river outcomes
Environmental flows in impounded rivers
generally target the conservation of some
particular species rather than outcomes at the
food-web or ecosystem level
Advance fundamental research on microbialndash
ecosystem process linkages Expand the notion
of designer flows to food-web structure and
ecosystem services when possible
Environmental flow science has largely
ignored surface waterndashgroundwater
interactions
Expand science to understand the link between
restoration actions groundwater recharge and
inputs to streams Provide science relevant to
the implementation of new legislation on
groundwater management (eg Sustainable
Groundwater Management Act in California)
Uncertainty regarding future
hydroclimate and
river flows is growing
Refine regional climate models Promote research on
the interaction between land-use change and
evapotranspiration Incorporate groundwater
influence in macroscale hydrology models
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
stressors (particularly in urban streams) and thechallenge of predicting outcomeswhen both thephysical and the biological environments arechangingAchieving a predictive understanding of hy-
droecological relationships is essential to design-ing successful self-sustaining restoration actionsin river ecosystems However translating basichydroecological insights into restoration practicerequires understanding the complexities of work-ing within a human-dominated water cycle Al-though it is rarely possible to restore all facetsof a natural flow regime ecologically informeddesigns may help mimic key aspects of the nat-ural flow regime (eg periodic pulse flows) orcreate regimes that are not natural but maximizeoutcomes given socioeconomic constraints InTable 1 we identify persisting and emergingchallenges in hydroecological research We paireach challenge with an opportunity (related toresearch design technology or implementation)We contend that these challenges should beprioritized to advance process-based river eco-system restoration
REFERENCES AND NOTES
1 N L Poff et al The natural flow regime Bioscience 47769ndash784 (1997) doi 1023071313099
2 D A Lytle N L Poff Adaptation to natural flow regimesTrends Ecol Evol 19 94ndash100 (2004) doi 101016jtree200310002 pmid 16701235
3 S Sabater A Elosegi R Ludwig in Multiple Stressors in RiverEcosystems (Elsevier 2019) pp 1ndash22
4 United Nations New UN Decade on Ecosystem Restorationoffers unparalleled opportunity for job creation food securityand addressing climate change (2019) httpswwwunenvironmentorgnews-and-storiespress-releasenew-un-decade-ecosystem-restoration-offers-unparalleled-opportunity
5 A W Lorenz P Haase K Januschke A SundermannD Hering Revisiting restored river reaches - Assessingchange of aquatic and riparian communities afterfive years Sci Total Environ 613-614 1185ndash1195(2018) doi 101016jscitotenv201709188pmid 28954379
6 M A Palmer K L Hondula B J Koch Ecological Restorationof Streams and Rivers Shifting Strategies and ShiftingGoals (2014) vol 45
7 T J Beechie et al Process-based principles for restoringriver ecosystems Bioscience 60 209ndash222 (2010)doi 101525bio20106037
8 S E Bunn A H Arthington Basic principles and ecologicalconsequences of altered flow regimes for aquaticbiodiversity Environ Manage 30 492ndash507 (2002)doi 101007s00267-002-2737-0 pmid 12481916
9 K D Fausch Y Taniguchi S Nakano G D GrossmanC R Townsend Flood disturbance regimes influence rainbowtrout invasion success among five holarctic regions EcolAppl 11 1438ndash1455 (2001) doi 1018901051-0761(2001)011[1438FDRIRT]20CO2
10 M E Power M S Parker W E Dietrich Seasonalreassembly of a river food web Floods droughts andimpacts of fish Ecol Monogr 78 263ndash282 (2008)doi 10189006-09021
11 V Acuntildea A Giorgi I Muntildeoz U R S Uehlinger S SabaterFlow extremes and benthic organic matter shape themetabolism of a headwater Mediterranean streamFreshwater Biol 49 960ndash971 (2004) doi 101111j1365-2427200401239x
12 K Tockner F Malard J V Ward An extension of the floodpulse concept Hydrol Processes 14 2861ndash2883 (2000)doi 1010021099-1085(20001112)141617lt2861AID-HYP124gt30CO2-F
13 J S Perkin et al Fragmentation and dewatering transformGreat Plains stream fish communities Ecol Monogr 8573ndash92 (2015) doi 10189014-01211
14 J L Sabo D M Post Quantifying periodic stochastic andcatastrophic environmental variation Ecol Monogr 7819ndash40 (2008) doi 10189006-13401
15 F Shuai et al Silver carp larva abundance in response toriver flow rate revealed by cross-wavelet modellingEcol Modell 383 98ndash105 (2018) doi 101016jecolmodel201805020
16 J D Tonkin M T Bogan N Bonada B Rios-ToumaD A Lytle Seasonality and predictability shape temporalspecies diversity Ecology 98 1201ndash1216 (2017)doi 101002ecy1761 pmid 28144975
17 N L Poff Beyond the natural flow regime Broadening thehydro‐ecological foundation to meet environmental flowschallenges in a non‐stationary world Freshwater Biol 631011ndash1021 (2018) doi 101111fwb13038
18 A Ruhi X Dong C H McDaniel D P Batzer J L SaboDetrimental effects of a novel flow regime on the functionaltrajectory of an aquatic invertebrate metacommunity GlobChange Biol 24 3749ndash3765 (2018) doi 101111gcb14133pmid 29665147
19 M T Bogan D A Lytle Severe drought drives novelcommunity trajectories in desert stream pools FreshwaterBiol 56 2070ndash2081 (2011) doi 101111j1365-2427201102638x
20 G Woodward et al The effects of climatic fluctuations andextreme events on running water ecosystems Philos TransR Soc London Ser B 371 20150274 (2016) pmid 27114576
21 M A Palmer C M Swan K Nelson P Silver R AlvestadStreambed landscapes Evidence that stream invertebratesrespond to the type and spatial arrangement of patchesLandsc Ecol 15 563ndash576 (2000) doi 101023A1008194130695
22 D Schmera et al Does isolation influence the relative role ofenvironmental and dispersal‐related processes in streamnetworks An empirical test of the network positionhypothesis using multiple taxa Freshwater Biol 63 74ndash85(2018) doi 101111fwb12973
23 R Henriques‐Silva et al A comprehensive examination of thenetwork position hypothesis across multiple rivermetacommunities Ecography 42 284ndash294 (2019)doi 101111ecog03908
24 K L Jaeger J D Olden N A Pelland Climate change poisedto threaten hydrologic connectivity and endemic fishes indryland streams Proc Natl Acad Sci USA 11113894ndash13899 (2014) doi 101073pnas1320890111pmid 25136090
25 S Garbin E A Celegon P Fanton G Botter Hydrologicalcontrols on river network connectivity R Soc Open Sci 6181428 (2019) doi 101098rsos181428 pmid 30891270
26 F Cattaneacuteo B Hugueny N Lamouroux Synchrony in browntrout Salmo trutta population dynamics A lsquoMoran effectrsquo onearly‐life stages Oikos 100 43ndash54 (2003) doi 101034j1600-0706200311912x
27 A Terui et al Metapopulation stability in branching rivernetworks Proc Natl Acad Sci USA 115 E5963ndashE5969(2018) doi 101073pnas1800060115 pmid 29895695
28 P A P Moran The statistical analysis of the Canadian Lynxcycle Aust J Zool 1 291ndash298 (1953) doi 101071ZO9530291
29 G D Grossman P B Moyle J O Whitaker Jr Stochasticityin structural and functional characteristics of an Indianastream fish assemblage A test of community theoryAm Nat 120 423ndash454 (1982) doi 101086284004
30 D A Lytle D M Merritt J D Tonkin J D OldenL V Reynolds Linking river flow regimes to riparian plantguilds A community-wide modeling approach Ecol Appl 271338ndash1350 (2017) doi 101002eap1528 pmid 28263426
31 A Ruhiacute E E Holmes J N Rinne J L Sabo Anomalousdroughts not invasion decrease persistence of native fishesin a desert river Glob Change Biol 21 1482ndash1496 (2015)doi 101111gcb12780 pmid 25430731
32 J D Tonkin D M Merritt J D Olden L V ReynoldsD A Lytle Flow regime alteration degrades ecologicalnetworks in riparian ecosystems Nat Ecol Evol 2 86ndash93(2018) doi 101038s41559-017-0379-0 pmid 29180707
33 M C Mims J D Olden Fish assemblages respond toaltered flow regimes via ecological filtering of lifehistory strategies Freshwater Biol 58 50ndash62 (2013)doi 101111fwb12037
34 P T J Johnson J D Olden M J Vander Zanden Daminvaders Impoundments facilitate biological invasions intofreshwaters Front Ecol Environ 6 357ndash363 (2008)doi 101890070156
35 N Bonada S Doledec B Statzner Taxonomic and biologicaltrait differences of stream macroinvertebrate communitiesbetween mediterranean and temperate regions Implicationsfor future climatic scenarios Glob Change Biol 131658ndash1671 (2007) doi 101111j1365-2486200701375x
36 M I Pyne N L Poff Vulnerability of stream communitycomposition and function to projected thermal warming andhydrologic change across ecoregions in the western UnitedStates Glob Change Biol 23 77ndash93 (2017) doi 101111gcb13437 pmid 27429092
37 D B Herbst S D Cooper R B Medhurst S W WisemanC T Hunsaker Drought ecohydrology alters the structureand function of benthic invertebrate communities inmountain streams Freshwater Biol 64 886ndash902 (2019)doi 101111fwb13270
38 M T Bogan E T Chester T Datry A L MurphyB J Robson A Ruhi R Stubbington J E Whitney inResistance Resilience and Community Recovery in IntermittentRivers and Ephemeral Streams T Datry N Bonada A BoultonEds (Academic Press 2017) pp 349ndash376
39 S Houmlckendorff et al Characterizing fish responses to a riverrestoration over 21 years based on speciesrsquo traits ConservBiol 31 1098ndash1108 (2017) doi 101111cobi12908pmid 28218807
40 D Bruno et al Structural and functional responses ofinvertebrate communities to climate change andflow regulation in alpine catchments Glob Change Biol25 1612ndash1628 (2019) doi 101111gcb14581pmid 30698905
41 T W H Aspin et al Extreme drought pushes streaminvertebrate communities over functional thresholds GlobChange Biol 25 230ndash244 (2019) doi 101111gcb14495pmid 30346098
42 S E Tank J B Fellman E Hood E S Kritzberg Beyondrespiration Controls on lateral carbon fluxes across theterrestrial-aquatic interface Limnol Oceanogr Lett 3 76ndash88(2018) doi 101002lol210065
43 A P Appling et al The metabolic regimes of 356 rivers inthe United States Sci Data 5 180292 (2018) doi 101038sdata2018292 pmid 30532078
44 P Savoy et al Metabolic rhythms in flowing watersAn approach for classifying river productivity regimesLimnol Oceanogr (2019) doi 101002lno11154
45 E S Bernhardt et al The metabolic regimes of flowingwaters Limnol Oceanogr 63 S99ndashS118 (2018)doi 101002lno10726
46 M Colls X Timoner S Sabater V Acuntildea Effects of durationand frequency of the non-flow period on stream metabolismEcosystems 101007s10021-019-00345-1 (2019)doi 101007s10021-019-00345-1
47 A J Ulseth E Bertuzzo G A Singer J Schelker T J BattinClimate-induced changes in spring snowmelt impactecosystem metabolism and carbon fluxes in an Alpine streamnetwork Ecosystems 21 373ndash390 (2018) doi 101007s10021-017-0155-7
48 B O L Demars Hydrological pulses and burning of dissolvedorganic carbon by stream respiration Limnol Oceanogr 64406ndash421 (2019) doi 101002lno11048
49 R T Hensley L Kirk M Spangler M N Gooseff M J CohenFlow extremes as spatiotemporal control points on riversolute fluxes and metabolism J Geophys Res Biogeosci124 537ndash555 (2019) doi 1010292018JG004738
50 J D Hosen et al Enhancement of primary production duringdrought in a temperate watershed is greater in larger riversthan headwater streams Limnol Oceanogr 64 1458ndash1472(2019) doi 101002lno11127
51 L Goacutemez-Gener et al When water vanishes Magnitude andregulation of carbon dioxide emissions from dry temporarystreams Ecosystems 19 710ndash723 (2016) doi 101007s10021-016-9963-4
52 E H Stanley et al The ecology of methane in streams andrivers Patterns controls and global significance EcolMonogr 86 146ndash171 (2016) doi 10189015-1027
53 R T Barnes D E Butman H F Wilson P A RaymondRiverine export of aged carbon driven by flow path depth andresidence time Environ Sci Technol 52 1028ndash1035 (2018)doi 101021acsest7b04717 pmid 29313674
54 G Pinay et al Riparian corridors A new conceptualframework for assessing nitrogen buffering across biomesFront Environ Sci 6 47 (2018) doi 103389fenvs201800047
55 W M Wollheim et al River network saturation conceptFactors influencing the balance of biogeochemical supply and
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 11 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
demand of river networks Biogeochemistry 141 503ndash521(2018) doi 101007s10533-018-0488-0
56 P A Raymond J E Saiers W V Sobczak Hydrological andbiogeochemical controls on watershed dissolved organicmatter transport Pulse-shunt concept Ecology 97 5ndash16(2016) doi 10189014-16841 pmid 27008769
57 C Tunaley D Tetzlaff J Lessels C Soulsby Linking high-frequency DOC dynamics to the age of connected watersources Water Resour Res 52 5232ndash5247 (2016)doi 1010022015WR018419
58 S Bernal et al Supply demand and in-stream retention ofdissolved organic carbon and nitrate during storms inMediterranean forested headwater streams Front EnvironSci 7 60 (2019) doi 103389fenvs201900060
59 J Harvey et al How hydrologic connectivity regulates waterquality in river corridors J Am Water Resour Assoc 55369ndash381 (2019) doi 1011111752-168812691
60 J C Moore et al Detritus trophic dynamics and biodiversityEcol Lett 7 584ndash600 (2004) doi 101111j1461-0248200400606x
61 A D Rosemond et al Freshwater ecology Experimentalnutrient additions accelerate terrestrial carbon loss fromstream ecosystems Science 347 1142ndash1145 (2015)doi 101126scienceaaa1958 pmid 25745171
62 A C Y Yeung J L Musetta-Lambert D P KreutzweiserP K Sibley J S Richardson Relations of interannualdifferences in stream litter breakdown with dischargeBioassessment implications Ecosphere 9 e02423 (2018)doi 101002ecs22423
63 R M Northington J R Webster Experimental reductions instream flow alter litter processing and consumer subsidies inheadwater streams Freshwater Biol 62 737ndash750 (2017)doi 101111fwb12898
64 E Mas-Martiacute I Sanpera-Calbet I Muntildeoz Bottom-up effectsof streambed drying on consumer performance throughchanges in resource quality Aquat Sci 79 719ndash731 (2017)doi 101007s00027-017-0531-6
65 B D Richter G A Thomas Restoring environmental flows bymodifying dam operations Ecol Soc 12 12 (2007)doi 105751ES-02014-120112
66 G Zolezzi A Siviglia M Toffolon B MaioliniThermopeaking in Alpine streams Event characterizationand time scales Ecohydrology 4 564ndash576 (2011)doi 101002eco132
67 T Meiszligner B Sures C K Feld Multiple stressors and therole of hydrology on benthic invertebrates in mountainousstreams Sci Total Environ 663 841ndash851 (2019)doi 101016jscitotenv201901288 pmid 30738264
68 I R Waite et al Effects of urban multi-stressors on threestream biotic assemblages Sci Total Environ 6601472ndash1485 (2019) doi 101016jscitotenv201812240pmid 30743940
69 K C Nelson M A Palmer Stream temperature surges underurbanization and climate change Data models andresponses J Am Water Resour Assoc 43 440ndash452 (2007)doi 101111j1752-1688200700034x
70 J R Blaszczak J M Delesantro Y Zhong D L UrbanE S Bernhardt Watershed urban development controls onurban streamwater chemistry variability Biogeochemistry144 61ndash84 (2019) doi 101007s10533-019-00572-7
71 L Ponsatiacute et al Biofilm responses to flow regulation by damsin Mediterranean rivers River Res Appl 31 1003ndash1016(2015) doi 101002rra2807
72 J M Alberts J J Beaulieu I Buffam Watershed land useand seasonal variation constrain the influence of ripariancanopy cover on stream ecosystem metabolism Ecosystems20 553ndash567 (2017) doi 101007s10021-016-0040-9pmid 30416371
73 R O Hall Jr et al Turbidity light temperature andhydropeaking control primary productivity in the ColoradoRiver Grand Canyon Limnol Oceanogr 60 512ndash526 (2015)doi 101002lno10031
74 J R Blaszczak J M Delesantro D L Urban M W DoyleE S Bernhardt Scoured or suffocated Urban streamecosystems oscillate between hydrologic and dissolvedoxygen extremes Limnol Oceanogr 64 877ndash894 (2019)doi 101002lno11081
75 J-R Mor et al Dam regulation and riverine food-webstructure in a Mediterranean river Sci Total Environ 625301ndash310 (2018) doi 101016jscitotenv201712296pmid 29289778
76 H A Wellard Kelly et al Macroinvertebrate diets reflecttributary inputs and turbidity-driven changes in food
availability in the Colorado River downstream of GlenCanyon Dam Freshw Sci 32 397ndash410 (2013)doi 10189912-0881
77 S J Imberger R M Thompson M R Grace Urbancatchment hydrology overwhelms reach scale effects ofriparian vegetation on organic matter dynamics FreshwaterBiol 56 1370ndash1389 (2011) doi 101111j1365-2427201102575x
78 R T Martins A S Melo J F Gonccedilalves Jr N HamadaLeaf-litter breakdown in urban streams of Central AmazoniaDirect and indirect effects of physical chemical andbiological factors Freshw Sci 34 716ndash726 (2015)doi 101086681086
79 C M Yule J Y Gan T Jinggut K V Lee Urbanizationaffects food webs and leaf-litter decomposition in a tropicalstream in Malaysia Freshw Sci 34 702ndash715 (2015)doi 101086681252
80 T A Newcomer et al Influence of natural and novel organiccarbon sources on denitrification in forest degraded urbanand restored streams Ecol Monogr 82 449ndash466 (2012)doi 10189012-04581
81 A Chaudhary I Kauser A Ray R Poretsky Taxon-drivenfunctional shifts associated with storm flow in an urbanstream microbial community MSphere 3 e00194-18 (2018)doi 101128mSphere00194-18 pmid 29976643
82 T A Kennedy et al Flow management for hydropowerextirpates aquatic insects undermining river food websBioscience 66 561ndash575 (2016) doi 101093bioscibiw059
83 A Glazaczow D Orwin M Bogdziewicz Increasedtemperature delays the late-season phenology of multivoltineinsect Sci Rep 6 38022 (2016) doi 101038srep38022pmid 27905493
84 S G Fisher L J Gray N B Grimm D E Busch Temporalsuccession in a desert stream ecosystem following flashflooding Ecol Monogr 52 93ndash110 (1982) doi 1023072937346
85 G J Scrimgeour M J Winterbourn Effects of floods onepilithon and benthic macroinvertebrate populations in anunstable New Zealand river Hydrobiologia 171 33ndash44 (1989)doi 101007BF00005722
86 E Bondar-Kunze S Maier D Schoumlnauer N Bahl T HeinAntagonistic and synergistic effects on a stream periphytoncommunity under the influence of pulsed flow velocityincrease and nutrient enrichment Sci Total Environ 573594ndash602 (2016) doi 101016jscitotenv201608158pmid 27585428
87 M J Cashman G L Harvey G Wharton M C Bruno Woodmitigates the effect of hydropeaking scour on periphytonbiomass and nutritional quality in semi-natural flumesimulations Aquat Sci 79 459ndash471 (2017) doi 101007s00027-016-0510-3
88 W F Cross et al Food-web dynamics in a large riverdiscontinuum Ecol Monogr 83 311ndash337 (2013)doi 10189012-17271
89 D M Warfe et al Productivity disturbance and ecosystemsize have no influence on food chain length in seasonallyconnected rivers PLOS ONE 8 e66240 (2013)doi 101371journalpone0066240
90 T D Jardine et al Consumer-resource coupling in wet-drytropical rivers J Anim Ecol 81 310ndash322 (2012)doi 101111j1365-2656201101925x pmid 22103689
91 K O Winemiller et al Pulsing hydrology determines top‐down control of basal resources in a tropical riverndashfloodplainecosystem Ecol Monogr 84 621ndash635 (2014)doi 10189013-18221
92 D M Post The long and short of food-chain length TrendsEcol Evol 17 269ndash277 (2002) doi 101016S0169-5347(02)02455-2
93 D M Post G Takimoto Proximate structural mechanismsfor variation in food‐chain length Oikos 116 775ndash782 (2007)doi 101111j0030-1299200715552x
94 P A McHugh A R McIntosh P G Jellyman Dual influencesof ecosystem size and disturbance on food chain length instreams Ecol Lett 13 881ndash890 (2010) doi 101111j1461-0248201001484x pmid 20482579
95 A Ruhiacute et al Flow regulation increases food-chain lengththrough omnivory mechanisms in a Mediterranean rivernetwork Freshwater Biol 61 1536ndash1549 (2016) doi 101111fwb12794
96 J L Sabo J C Finlay T Kennedy D M Post The role ofdischarge variation in scaling of drainage area and food chainlength in rivers Science 330 965ndash967 (2010) doi 101126science1196005 pmid 20947729
97 A R McIntosh et al Capacity to support predators scaleswith habitat size Sci Adv 4 eaap7523 (2018) doi 101126sciadvaap7523 pmid 29978038
98 J H Brown J F Gillooly A P Allen V M Savage G B WestToward a metabolic theory of ecology Ecology 851771ndash1789 (2004) doi 10189003-9000
99 K Zou E Theacutebault G Lacroix S Barot Interactions betweenthe green and brown food web determine ecosystemfunctioning Funct Ecol 30 1454ndash1465 (2016) doi 1011111365-243512626
100 M J Vanni P B McIntyre Predicting nutrient excretion ofaquatic animals with metabolic ecology and ecologicalstoichiometry A global synthesis Ecology 97 3460ndash3471(2016) doi 101002ecy1582 pmid 27912023
101 T B Parr K A Capps S P Inamdar K A Metcalf Animal-mediated organic matter transformation Aquatic insects as asource of microbially bioavailable organic nutrients andenergy Funct Ecol 33 524ndash535 (2019) doi 1011111365-243513242
102 C P Arango et al Urban infrastructure influences dissolvedorganic matter quality and bacterial metabolism in an urbanstream network Freshwater Biol 62 1917ndash1928 (2017)
103 L G Larsen J W Harvey Disrupted carbon cycling inrestored and unrestored urban streams Critical timescalesand controls Limnol Oceanogr 62 S160ndashS182 (2017)doi 101002lno10613
104 H M Halvorson G White J T Scott M A Evans-WhiteDietary and taxonomic controls on incorporation of microbialcarbon and phosphorus by detritivorous caddisfliesOecologia 180 567ndash579 (2016) doi 101007s00442-015-3464-6 pmid 26497125
105 J J Elser et al Biological stoichiometry from genes toecosystems Ecol Lett 3 540ndash550 (2000) doi 101046j1461-0248200000185x
106 E K Moody et al Consumption explains intraspecificvariation in nutrient recycling stoichiometry in a desert fishEcology 99 1552ndash1561 (2018) doi 101002ecy2372pmid 29882955
107 N Welti et al Bridging food webs ecosystem metabolismand biogeochemistry using ecological stoichiometry theoryFront Microbiol 8 1298 (2017) doi 103389fmicb201701298 pmid 28747904
108 M A Palmer S Filoso R M Fanelli From ecosystems toecosystem services Stream restoration as ecologicalengineering Ecol Eng 65 62ndash70 (2014) doi 101016jecoleng201307059
109 M J Burns T D Fletcher C J Walsh A R LadsonB E Hatt Hydrologic shortcomings of conventional urbanstormwater management and opportunities for reformLandsc Urban Plan 105 230ndash240 (2012) doi 101016jlandurbplan201112012
110 C R Liermann C Nilsson J Robertson R Y NgImplications of dam obstruction for global freshwater fishdiversity Bioscience 62 539ndash548 (2012) doi 101525bio20126265
111 J D Olden R J Naiman Incorporating thermal regimes intoenvironmental flows assessments Modifying dam operationsto restore freshwater ecosystem integrity Freshwater Biol55 86ndash107 (2010) doi 101111j1365-2427200902179x
112 D B Osmundson R J Ryel V L Lamarra J Pitlick Flowndashsedimentndashbiota relations Implications for river regulationeffects on native fish abundance Ecol Appl 12 1719ndash1739(2002)
113 D von Schiller et al Regulation causes nitrogen cyclingdiscontinuities in Mediterranean rivers Sci Total Environ540 168ndash177 (2016) doi 101016jscitotenv201507017pmid 26233271
114 J L Sabo et al Designing river flows to improve foodsecurity futures in the Lower Mekong Basin Science 358eaao1053 (2017) doi 101126scienceaao1053pmid 29217541
115 W Xu et al Improving the multi-objective performance ofrainwater harvesting systems using real-time controltechnology Water 10 147 (2018) doi 103390w10020147
116 M Acreman et al Environmental flows for naturalhybrid and novel riverine ecosystems in a changingworld Front Ecol Environ 12 466ndash473 (2014)doi 101890130134
117 N L Poff et al Sustainable water management under futureuncertainty with eco-engineering decision scaling Nat ClimChang 6 25ndash34 (2016) doi 101038nclimate2765
118 J D Kiernan P B Moyle P K Crain Restoring native fishassemblages to a regulated California stream using the
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 12 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
natural flow regime concept Ecol Appl 22 1472ndash1482(2012) doi 10189011-04801 pmid 22908707
119 D L Propst K B Gido Responses of native andnonnative fishes to natural flow regime mimicry in theSan Juan River Trans Am Fish Soc 133 922ndash931 (2004)doi 101577T03-0571
120 T K Pool J D Olden Assessing long‐term fish responses andshort‐term solutions to flow regulation in a dryland river basinEcol Freshwat Fish 24 56ndash66 (2015) doi 101111eff12125
121 W Chen J D Olden Designing flows to resolve human andenvironmental water needs in a dam-regulated riverNat Commun 8 2158 (2017) doi 101038s41467-017-02226-4 pmid 29255194
122 S M Nelson et al A history of the 2014 Minute 319environmental pulse flow as documented by fieldmeasurements and satellite imagery Ecol Eng 106733ndash748 (2017) doi 101016jecoleng201610040
123 P B Shafroth et al A large-scale environmental flowexperiment for riparian restoration in the Colorado RiverDelta Ecol Eng 106 645ndash660 (2017) doi 101016jecoleng201702016
124 E P Glenn P L Nagler P B Shafroth C J JarchowEffectiveness of environmental flows for riparian restorationin arid regions A tale of four rivers Ecol Eng 106 695ndash703(2017) doi 101016jecoleng201701009
125 S Hotaling D S Finn J Joseph Giersch D W WeisrockD Jacobsen Climate change and alpine stream biologyProgress challenges and opportunities for the futureBiol Rev Camb Philos Soc 92 2024ndash2045 (2017)doi 101111brv12319 pmid 28105701
126 C J Walsh T D Fletcher M J Burns Urban stormwaterrunoff A new class of environmental flow problem PLOS ONE7 e45814 (2012) doi 101371journalpone0045814pmid 23029257
127 A Selvakumar T P OrsquoConnor S D Struck Role of streamrestoration on improving benthic macroinvertebrates and in-stream water quality in an urban watershed Case studyJ Environ Eng 136 127ndash139 (2010) doi 101061(ASCE)EE1943-78700000116
128 R W Lammers B P Bledsoe What role does streamrestoration play in nutrient management Crit Rev EnvironSci Technol 47 335ndash371 (2017) doi 1010801064338920171318618
129 H E Golden N Hoghooghi Green infrastructure and itscatchment-scale effects An emerging science WIREs Water5 e1254 (2018) doi 101002wat21254 pmid 29682288
130 M R Williams G Bhatt S Filoso G Yactayo Streamrestoration performance and its contribution to theChesapeake Bay TMDL Challenges posed by climate changein urban areas Estuaries Coasts 40 1227ndash1246 (2017)doi 101007s12237-017-0226-1
131 R M Fanelli K Prestegaard M A Palmer Freshw Sci101086706072 (2019)
132 B W Abbott et al Unexpected spatial stability of waterchemistry in headwater stream networks Ecol Lett 21296ndash308 (2018) doi 101111ele12897 pmid 29282860
133 B L Brown C M Swan Dendritic network structureconstrains metacommunity properties in riverineecosystems J Anim Ecol 79 571ndash580 (2010) doi 101111j1365-2656201001668x pmid 20180874
134 C M Swan B L Brown Metacommunity theory meetsrestoration Isolation may mediate how ecologicalcommunities respond to stream restoration Ecol Appl 272209ndash2219 (2017) doi 101002eap1602 pmid 28718193
135 N Mouquet D Gravel F Massol V Calcagno Extending theconcept of keystone species to communities and ecosystemsEcol Lett 16 1ndash8 (2013) doi 101111ele12014 pmid 23062191
136 A Ruhiacute T Datry J L Sabo Interpreting beta-diversitycomponents over time to conserve metacommunities inhighly dynamic ecosystems Conserv Biol 31 1459ndash1468(2017) doi 101111cobi12906 pmid 28188969
137 L Yamane L W Botsford D P Kilduff Tracking restorationof population diversity via the portfolio effect J Appl Ecol55 472ndash481 (2018) doi 1011111365-266412978
138 D W Braidwood M A Taggart M Smith R AndersenTranslocations conservation and climate change Use ofrestoration sites as protorefuges and protorefugia RestorEcol 26 20ndash28 (2018) doi 101111rec12642
139 Y Wang D Wang Q W Lewis J Wu F Huang A frameworkto assess the cumulative impacts of dams on hydrologicalregime A case study of the Yangtze River Hydrol Processes31 3045ndash3055 (2017) doi 101002hyp11239
140 J L Sabo et al Pulsed flows tributary inputs and food-webstructure in a highly regulated river J Appl Ecol 551884ndash1895 (2018) doi 1011111365-266413109
141 C Magliozzi G Coro R C Grabowski A I PackmanS Krause A multiscale statistical method to identifypotential areas of hyporheic exchange for river restorationplanning Environ Model Softw 111 311ndash323 (2019)doi 101016jenvsoft201809006
142 S S Schwartz B Smith Restoring hydrologic function inurban landscapes with suburban subsoiling J Hydrol 543770ndash781 (2016) doi 101016jjhydrol201610051
143 C J Walsh T D Fletcher A R Ladson Stream restoration inurban catchments through redesigning stormwater systemsLooking to the catchment to save the stream J N AmBenthol Soc 24 690ndash705 (2005) doi 10189904-0201
144 N M Schmadel et al Thresholds of lake and reservoirconnectivity in river networks control nitrogen removalNat Commun 9 2779 (2018) doi 101038s41467-018-05156-x pmid 30018449
145 C D Sayer Conservation of aquatic landscapes Pondslakes and rivers as integrated systems WIREs Water 1573ndash585 (2014) doi 101002wat21045
146 R G Luthy D L Sedlak M H Plumlee D Austin V H ReshWastewater‐effluent‐dominated streams as ecosystem‐
management tools in a drier climate Front Ecol Environ 13477ndash485 (2015) doi 101890150038
147 D W Roscoe S G Hinch Effectiveness monitoring of fishpassage facilities Historical trends geographic patterns andfuture directions Fish Fish 11 12ndash33 (2010) doi 101111j1467-2979200900333x
148 C M Bunt T Castro-Santos A Haro Performanceof fish passage structures at upstream barriers tomigration River Res Appl 28 457ndash478 (2012)doi 101002rra1565
149 D Weigel et al Evaluation of a trap-and-transport programfor a threatened population of steelhead (Oncorhynchusmykiss) Conserv Genet 20 1195ndash1199 (2019) doi 101007s10592-019-01200-5
150 M H Bond et al Combined effects of barge transportationriver environment and rearing location on straying andmigration of adult Snake River fall-run Chinook SalmonTrans Am Fish Soc 146 60ndash73 (2017) doi 1010800002848720161235614
151 A C Ritchie et al Morphodynamic evolution followingsediment release from the worldrsquos largest dam removalSci Rep 8 13279 (2018) doi 101038s41598-018-30817-8pmid 30185796
152 T Sommer et al Californiarsquos Yolo Bypass Evidence thatflood control can be compatible with fisheries wetlandswildlife and agriculture Fisheries 26 6ndash16 (2001)doi 1015771548-8446(2001)026lt0006CYBgt20CO2
153 G Grill et al Mapping the worldrsquos free-flowing riversNature 569 215ndash221 (2019) doi 101038s41586-019-1111-9pmid 31068722
154 G Gassert M Luck M Landis P Reig T Shiao ldquoAqueductglobal maps 21 Constructing decision-relevant global waterrisk indicatorsrdquo Working paper World Resources Institute(2015) wwwwriorgpublicationaqueduct-global-maps-21-indicators
155 A Ruhiacute J D Olden J L Sabo Declining streamflow inducescollapse and replacement of native fishes in the AmericanSouthwest Front Ecol Environ 14 465ndash472 (2016)doi 101002fee1424
156 T C Gouhier A Grinsted V Simko R package Biwaveletconduct univariate and bivariate wavelet analyses (2018)
157 T Rodriacuteguez-Castillo E Esteacutevez A M Gonzaacutelez-FerrerasJ Barquiacuten Estimating ecosystem metabolism toentire river networks Ecosystems (2018) doi 101007s10021-018-0311-8
ACKNOWLEDGMENTS
We greatly appreciate those who read and provided importantinsights on drafts of this paper A Armstrong K HondulaJ Hosen K Leathers G de Mendoza C Maietta I MunozJ Tonkin and two anonymous reviewers We also thank T Tu forhelp with Fig 1 and S Carlson L Comte T Datry J HosenM Kondolf N Jones D Lytle E Moody J Olden M PowerJ Sabo and M Sanchez-Montoya for valuable discussionsCompeting interests The authors have no competing interestsFunding MAP was supported by funds from the NSF (1639145)and AR was supported by funds from the University of CaliforniaBerkeley and NSF (1802714)
101126scienceaaw2087
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 13 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
restorationLinkages between flow regime biota and ecosystem processes Implications for river
Margaret Palmer and Albert Ruhi
DOI 101126scienceaaw2087 (6459) eaaw2087365Science
this issue p eaaw2087Scienceflow variability and achieve hydrological connectivity between a river and its surroundingsand ecosystem processes To be successful such efforts must go beyond accounting for flood pulses to restore natural mimic ecologically important aspects of natural flow regimes guided by insights into how variations in flow affect biotawater quality and ecological processes In a Review Palmer and Ruhi explain how restoration designs now attempt to
Human activities have altered the flow regimes of many of Earths rivers with negative impacts on biodiversityRiver restoration guided by research
ARTICLE TOOLS httpsciencesciencemagorgcontent3656459eaaw2087
REFERENCES
httpsciencesciencemagorgcontent3656459eaaw2087BIBLThis article cites 147 articles 7 of which you can access for free
PERMISSIONS httpwwwsciencemagorghelpreprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAASScienceScience 1200 New York Avenue NW Washington DC 20005 The title (print ISSN 0036-8075 online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
Science No claim to original US Government WorksCopyright copy 2019 The Authors some rights reserved exclusive licensee American Association for the Advancement of
on May 27 2020
httpsciencesciencem
agorgD
ownloaded from
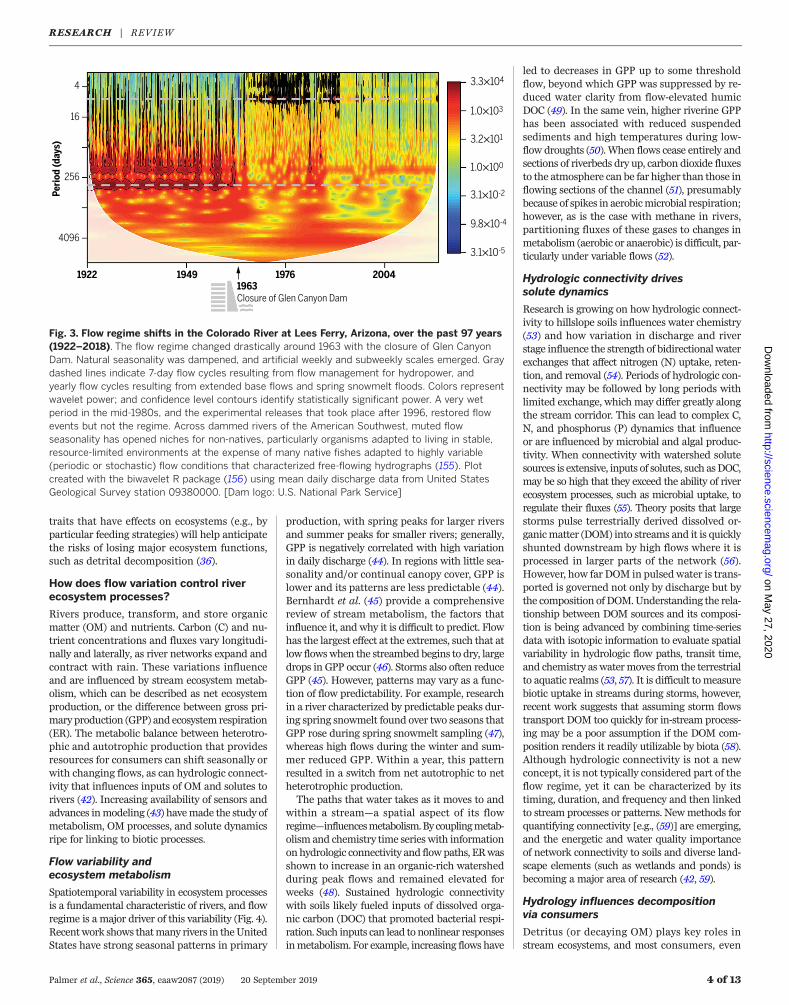
traits that have effects on ecosystems (eg byparticular feeding strategies) will help anticipatethe risks of losing major ecosystem functionssuch as detrital decomposition (36)
How does flow variation control riverecosystem processes
Rivers produce transform and store organicmatter (OM) and nutrients Carbon (C) and nu-trient concentrations and fluxes vary longitudi-nally and laterally as river networks expand andcontract with rain These variations influenceand are influenced by stream ecosystem metab-olism which can be described as net ecosystemproduction or the difference between gross pri-mary production (GPP) and ecosystem respiration(ER) The metabolic balance between heterotro-phic and autotrophic production that providesresources for consumers can shift seasonally orwith changing flows as can hydrologic connect-ivity that influences inputs of OM and solutes torivers (42) Increasing availability of sensors andadvances inmodeling (43) havemade the study ofmetabolism OM processes and solute dynamicsripe for linking to biotic processes
Flow variability andecosystem metabolism
Spatiotemporal variability in ecosystem processesis a fundamental characteristic of rivers and flowregime is a major driver of this variability (Fig 4)Recentwork shows thatmany rivers in theUnitedStates have strong seasonal patterns in primary
production with spring peaks for larger riversand summer peaks for smaller rivers generallyGPP is negatively correlated with high variationin daily discharge (44) In regions with little sea-sonality andor continual canopy cover GPP islower and its patterns are less predictable (44)Bernhardt et al (45) provide a comprehensivereview of stream metabolism the factors thatinfluence it and why it is difficult to predict Flowhas the largest effect at the extremes such that atlow flowswhen the streambed begins to dry largedrops in GPP occur (46) Storms also often reduceGPP (45) However patterns may vary as a func-tion of flow predictability For example researchin a river characterized by predictable peaks dur-ing spring snowmelt found over two seasons thatGPP rose during spring snowmelt sampling (47)whereas high flows during the winter and sum-mer reduced GPP Within a year this patternresulted in a switch from net autotrophic to netheterotrophic productionThe paths that water takes as it moves to and
within a streammdasha spatial aspect of its flowregimemdashinfluencesmetabolismBycouplingmetab-olism and chemistry time series with informationonhydrologic connectivity and flowpaths ERwasshown to increase in an organic-rich watershedduring peak flows and remained elevated forweeks (48) Sustained hydrologic connectivitywith soils likely fueled inputs of dissolved orga-nic carbon (DOC) that promoted bacterial respi-ration Such inputs can lead to nonlinear responsesinmetabolism For example increasing flows have
led to decreases in GPP up to some thresholdflow beyond which GPP was suppressed by re-duced water clarity from flow-elevated humicDOC (49) In the same vein higher riverine GPPhas been associated with reduced suspendedsediments and high temperatures during low-flow droughts (50) When flows cease entirely andsections of riverbeds dry up carbon dioxide fluxesto the atmosphere can be far higher than those inflowing sections of the channel (51) presumablybecause of spikes in aerobicmicrobial respirationhowever as is the case with methane in riverspartitioning fluxes of these gases to changes inmetabolism (aerobic or anaerobic) is difficult par-ticularly under variable flows (52)
Hydrologic connectivity drivessolute dynamics
Research is growing on how hydrologic connect-ivity to hillslope soils influences water chemistry(53) and how variation in discharge and riverstage influence the strength of bidirectional waterexchanges that affect nitrogen (N) uptake reten-tion and removal (54) Periods of hydrologic con-nectivity may be followed by long periods withlimited exchange which may differ greatly alongthe stream corridor This can lead to complex CN and phosphorus (P) dynamics that influenceor are influenced by microbial and algal produc-tivity When connectivity with watershed solutesources is extensive inputs of solutes such asDOCmay be so high that they exceed the ability of riverecosystem processes such as microbial uptake toregulate their fluxes (55) Theory posits that largestorms pulse terrestrially derived dissolved or-ganicmatter (DOM) into streams and it is quicklyshunted downstream by high flows where it isprocessed in larger parts of the network (56)However how far DOM in pulsedwater is trans-ported is governed not only by discharge but bythe composition of DOMUnderstanding the rela-tionship between DOM sources and its composi-tion is being advanced by combining time-seriesdata with isotopic information to evaluate spatialvariability in hydrologic flow paths transit timeand chemistry as watermoves from the terrestrialto aquatic realms (53 57) It is difficult tomeasurebiotic uptake in streams during storms howeverrecent work suggests that assuming storm flowstransport DOM too quickly for in-stream process-ing may be a poor assumption if the DOM com-position renders it readily utilizable by biota (58)Although hydrologic connectivity is not a newconcept it is not typically considered part of theflow regime yet it can be characterized by itstiming duration and frequency and then linkedto stream processes or patterns Newmethods forquantifying connectivity [eg (59)] are emergingand the energetic and water quality importanceof network connectivity to soils and diverse land-scape elements (such as wetlands and ponds) isbecoming a major area of research (42 59)
Hydrology influences decompositionvia consumers
Detritus (or decaying OM) plays key roles instream ecosystems and most consumers even
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 4 of 13
33104
10103
32101
10100
3110-2
9810-4
3110-5
1922 1949 1976 2004
4
16
256
4096
Perio
d (d
ays)
1963Closure of Glen Canyon Dam
Fig 3 Flow regime shifts in the Colorado River at Lees Ferry Arizona over the past 97 years(1922ndash2018) The flow regime changed drastically around 1963 with the closure of Glen CanyonDam Natural seasonality was dampened and artificial weekly and subweekly scales emerged Graydashed lines indicate 7-day flow cycles resulting from flow management for hydropower andyearly flow cycles resulting from extended base flows and spring snowmelt floods Colors representwavelet power and confidence level contours identify statistically significant power A very wetperiod in the mid-1980s and the experimental releases that took place after 1996 restored flowevents but not the regime Across dammed rivers of the American Southwest muted flowseasonality has opened niches for non-natives particularly organisms adapted to living in stableresource-limited environments at the expense of many native fishes adapted to highly variable(periodic or stochastic) flow conditions that characterized free-flowing hydrographs (155) Plotcreated with the biwavelet R package (156) using mean daily discharge data from United StatesGeological Survey station 09380000 [Dam logo US National Park Service]
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
apical predators are at least partially supportedby it (60) Macroconsumers break up particulateorganic matter (POM) into small pieces enhanc-ing the overall mineralization rate by bacteriaand fungi and even moderate flows may resultin export of OM particularly fine POM resultingfrommacroconsumer shredding activities Over-all howevermicrobes largely drive C losses throughrespiration (61) and although nutrient availa-bility temperature and POM composition (egleaf type) influence this how flow mediates rela-tionships has been more difficult to pin downRecent work (62) designed to separate the effectsthat different components of the flow regimehave on the mechanisms driving decompositionin perennial streams found that dissolution andmicrobial processing consistently exceeded theeffects of fragmentation and microbial pro-cessing was best explained by daily variabilityin discharge When flow regimes become in-termittent the lack ofwater can impedemicrobialand consumer colonization of POM resulting indecreases in decomposition the extent of whichis related to the duration of the drying (46 63)Consistent water cover is therefore importantto decomposition and thus shifts toward lowerflows when litter inputs are high (eg in thefall) can have a large impact on consumers andassociated food webs (63 64)
When flows are altered are changes inbiota and ecosystem processes linked
To help ground this question in the world ofrestoration practice we begin with a focus ontwo of the most common forms of river flow al-teration damming and urbanization For theformer we emphasize hydropower dams becausethey are growing in number they strongly alterflow regimes and the mitigation of their impactshas spurred the field of environmental flow re-search We also focus on urban streams becausestudies on the impacts of imperviousness havebeen extensive and these streams have been the
target of numerous restoration efforts We summa-rize knowledge on the effects of flow on organ-isms ecosystem processes and their interaction inthese two types of river systems and then describehow this can inform river restoration in general
Urbanization and dams changeflow regimes
With few exceptions flow regimes below hydro-power dams and in urban streams are highlyaltered at one or both ends of the flow spectrumRegimes in impounded rivers are typically lessseasonal than daily rainfall would suggest par-ticularly below large hydropower dams with highwater-storage capacity rather than below smallerless impactful run-of-the-river facilities (65)Hydro-power dams canmute peak river flows but power-generation cycles induce rapidly fluctuating flowpatterns (hydropeaking) that often occur onhuman-relevant scales [ie hours to weeks (18)]tending to bemore pronounced in seasons of highenergy demand Hydropeaking is often accom-panied by spikes in temperature (thermopeak-ing) either cold or warm depending on seasonand reservoir operation (66) Thus downstreamorganisms and ecosystems are generally subjectto multiple periodic stresses In urban streamsbaseflows can be higher or lower depending onwatershed characteristics and infrastructurehowever peak flows are invariably larger thanthose prior to urbanization Understanding theecological impacts of urban stormflow is com-plicated by indirect effects on water quality In-creases in flow magnitude frequency and ratesof change during storms leads to elevated con-centrations of pollutants Despite this studiesseparating the causes of biological impairmenthave reported that the unique effects of alteredurban flows can be significant (67 68) Spikesin stream temperature caused by runoff fromhot pavement (69) add to stress on the biotaHydropeaking and urban stormflows might
be expected to reduce primary production by
scouring algae (70) yet some studies have re-ported elevated GPP or alternating periods ofnet autotrophy and heterotrophy (71ndash73) Lightcan override the effects of flow on metabolismby enhancing algal photosynthesis (74) how-ever if hydropeaking increases turbidity GPPmay decline (73) In urban streams elevatedGPP is often attributed to open canopies andorto elevated nutrient concentrations both of whichoften co-occur with their altered flow regimes(72) As discussed later such shifts in primaryproductivity have been linked to changes introphic structure eg more algal-based foodwebs (75) depending on the relative availabil-ity of terrestrial carbon inputs (both DOM andPOM) (76) Inputs of POM and its quality as afood resource changewith landscape context (egvegetative cover) and catchment hydrology (77)but POM availability can also be limited by flowvariability which can increase decompositionrates and decrease residence times (78 79)As is the case for running-water systems in
general high flows in urban streams bring pulsesof solutes including N DOM and various pol-lutants (80) Solute dynamics and biogeochemicalprocesses in regulated rivers have been insuffi-ciently studied with researchers focusing moreon such processes in reservoirs rather than indownstream river ecosystems We found nostudies attempting to link urban or regulatedflow regimes and solute fluxes to changes infood webs or stream metabolic balance How-ever recent studies show shifts in microbial tax-onomic and functional composition in responseto pulsed urban flows (81) and thesemay help inexploring a potential link but are complicated bythe fact that non-flow stressors affect consumercomposition For example urban and regulatedrivers typically have animal communities thatare less diverse and composed of tolerant groupsoften non-native Hydropeaking and urban stormflows can certainly cause direct mortality via dis-placement and transport to unsuitable habitatsbut many of the organismal effects are realizedthrough indirect pathways For instance hydro-peaking reduces the viability of river-edge egg-laying specialists by limiting their access to suitablespawning habitat disrupting reproductive successand insect emergence (82) Flow regulation caneven create ecological traps if organismal phenol-ogy shiftswith thenovel environmental conditionsFor example dam-induced warming triggeredsummer diapause in a mayfly resulting in theloss of its last generation (83) Dams also isolatepopulations harming organisms that requiremigrations to complete their life cycles whetherthese are between riverine and marine habitatsor within the freshwater domain By analogy al-tered flow regimes in urban streams can also leadto isolated pools in streams with lowered base-flows This can be associated with periods ofhypoxia punctuated by extreme flow pulses inwhich these streams become ldquoscoured or suffo-catedrdquo (74)As illustrated using urban streams and reg-
ulated rivers research is moving forward andhints at the three-way nexus but is clearly still
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 5 of 13
Fig 4 Ecosystem metabolismpatterns across a river networkSpatial patterns in river eco-system metabolism for the Deva-Cares catchment in northernSpain as represented by the ratioof gross primary production(P) to ecosystem respiration (R)A PR gt 1 indicates autotrophicprocesses dominate so theseriver sections are accumulatingor exporting organic carbonPR lt 1 indicates dominanceby heterotrophic processes inriver sections that are receivingorganic C input (eg fromterrestrial sources) The boxesindicate regions of the networkimpacted by human pressures including deforestation (brown box) waste products from urbansites (green box) and a combination of the two (purple box) Such visualizations are usefulto managers in understanding regions of concern that may need the most restoration actions[Reproduced with permission from (157)]
PR ratiolt025
gt150
025 ndash 050 050 ndash 075075 ndash 100100 ndash 150
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
inadequate to meet restoration needs Studieson ecosystem processes below dams are parti-cularly limited and much of what we knowcomes from only a few river systems Furtherpublished work focuses primarily on benthicecosystem processes immediately below damsrather than riverine planktonic realms whereproductivity in large regulated rivers can beimportant
Linking flow biota andecosystem processes
Research on flow regimes that bridges organis-mal and ecosystem ecology has traditionallyrelied on states instead of rates ie nutrientconcentrations rather than uptake or fluxesstanding stocks of algae rather than productivityor metabolism and abundance of organismsrather than secondary productivity This workgenerally shows that flow variability suppressesand stability increases algal standing stock andgrazer densities with peaks in invertebrate bio-mass following those of algae [eg (84 85)]Over the past decade research began to focuson how flow regimes alter functional aspects offoodwebs For example biofilms inMediterranean-climate rivers are thicker andmetabolicallymoreactive downstream of flow-stabilizing dams (71)When flow variability increases below hydro-power dams fluctuations can reduce biofilmdevelopment and its quality as a food resource(86) Using experimental flumes subjected tohydropeaking shifts have been found in periph-yton fatty acid content from important highlyunsaturated compounds to nonessential saturatedones (87)Indirect evidence suggests that flow-induced
changes in dominant resource type (eg algaeversus detritus) resource quantity (biomass) orresource quality (nutrient content) can propa-gate to higher trophic levels via bottom-up ef-fects For instance detritivorous invertebrateswere found to dominate species richness andinvertebrate biomass upstream but not down-stream of an irrigation dam (75) diets of om-nivore insects indicated that flow stabilizationshifted a detrital into an algal-fueled food webSimilar evidence comes from Glen Canyon Damin the Colorado River (88) Near the dam whereprimary production is high (73) food webs aresimplified and gut content analyses have sug-gested algal production supports gt50 of in-vertebrate and gt70 of fish production andthe food web can be easily perturbed by flooddisturbance Farther downriver food webs showincreased reliance on detrital resources havehigher trophic efficiency and are more complexand more resistant to disturbance (76) In con-trast recent work on seasonal tropical riversfound algal productivity and zero-flow distur-bance did not explain food-web structure Inthis case it seems that highly mobile predatorsbuffered the local effects of drought (89) Thisstudy along with earlier ones at the site (90) isof particular interest because it links temporalpatterns in predatory fish with flow-dependentavailability of resources
Only a few studies have provided evidence oftop-down controls on foodwebs being influencedby flow variation Fish in a tropical floodplainriver were found to exert top-down control onprimary producers but only under certain hydro-logic conditions (91) Similarly in aCalifornian riverwinter flood pulses were shown to control insectconsumers late in the season ultimately deter-mining whether fish exerted top-down controlon the lower trophic levels (10) These illustratethe importance of pulsed hydrology on resource-consumer linkages and show that not just standingstocks but also the dominant controls structuringfood webs may fluctuate over time in response toflow variationAn indirect line of evidence supporting the
importance of the flow-biota-ecosystem processesnexus comes from theoretically oriented researchon food-web structure Changes in productivityhabitat size and disturbance underpin the basisof the three main hypotheses of food-chainlength which state that chains should be longerin stable productive large ecosystems (92) No-tably flow regimes influence these three con-trols Because food-chain length determines keyaspects of a river ecosystem such as the risk oftrophic cascades or the pollutant accumulationin top predators understanding how it is af-fected by flow alteration is important How-ever understanding the interplay between thehypotheses of food-chain length and the flow-dependent biological mechanisms that allowfood webs to shift in structure (eg predatorinvasions local extinctions consumers chang-ing diets) is still a relatively recent endeavor(93ndash95) Evidence suggests that zero-flows short-en food chains mainly by keeping streamsfishless (94 96) whereas stabilization of variableflows can lengthen food chains by allowing om-nivores to feed on abundant high trophic levelprey (95)New studies are also focusing on interactions
between drivers of food-web structure that hadbeen previously only been examined in isolationsuch as disturbance regimes and ecosystem sizeboth influenced by flow regime Using metabolictheory predator biomass has been shown toscale with both prey resources and fluctuatingstream habitat size with reductions in preda-tor size leading to declines in predator biomasssupported per unit of prey biomass (97) Movingforward the combination of habitat andmetabolicconstraints on predators may provide a basis toconnect flow-induced disturbance changes inecosystem processes and habitat fluctuationsto riverine food-web structure The metabolictheory of ecology (98) is a unifying frameworkthat allows scaling responses of organisms towhole ecosystems andmay become particularlyuseful in situations where flow alteration affectsorganismal body size or water temperature Insuch cases strong changes in metabolic ratescan be expected affecting resource uptake andorganismal growth to ecosystem-level produc-tion and respirationFinally an important body of food webndashrelated
research that provides strong inferential evidence
for the nexus comes from studies on animal-mediated nutrient cycling and ecological stoi-chiometry Theory suggests detrital and algalfood websmay be linked by cascading effects onnutrient cycling (99) and empirical work showsthat organismal excretion andmigration (whichrelocates C) may be important to this link (100)(Fig 5) A recent study (101) showed that ex-cretion of highly labile dissolved organic C ni-trogen (N) and phosphorus (P) by aquatic insectsmay be sufficient to support a substantial fractionof microbial energy and nutrient needs Giventhat C availability can be too low tomeetmicrobialenergy needs [eg (102 103)] and that microbialactivity enhances detrital quality to consumers(104) a C subsidy from excretory processes couldbe important at least during low-flow periodsAdditionally microbial use of excreted dissolvedorganic nitrogen (DON) and DOP may releasealgae from potential competition with microbesfor inorganic N and P if this is the case GPPshould increase Ecological stoichiometry mayalso be important for understanding constraintson organismal growth rates as rapid growthdisproportionally increases demand of P [as it isused in ribosomal RNA (105)] This could in turnlimit the ability of organisms to grow fast andexploit temporary habitats Interestingly elemen-tal ratios can vary interspecifically in response totemperature or predation pressure (106)mdashfactorsthat increase with low flows and habitat fragmen-tation Overall studies that link biota with bio-geochemical processes remain rare (107) butanalyzing flow regimes as a driver of elementturnover rates may provide a way forward
Can hydroecological advances informrestoration practice
Shortfalls in the outcomes of river restorationhave prompted calls to identify and restore pro-cesses that support and sustain biological com-munities rather than focusing only on rivergeomorphology and habitat In urban streamsscientists have critiqued an overemphasis onstructural engineered approaches (7 108) thatdo little to restore the full flow regime (109) Inlarge regulated rivers identifying and restoringimportant features of the flow regime via damoperations is often possible (albeit expensive)However that does not solve other problemsassociated with dams such as habitat frag-mentation (110) altered sediment and thermalregimes (111 112) or disrupted biogeochemicalprocesses (113) Overall efforts to alter reservoirreleases or improve urban stormwater infra-structure to partially mimic a riverrsquos natural flowregime have great potential Technologies arebeing evaluated for real-time management ofurban flows and novel ways to design flow re-leases from dams are being proposed (114 115)however research still lags far behind the needfor solutions Importantly the paucity of studieson the flow-biota-ecosystem processes nexus pointsto a need for more integrative research connectinghydrologists population and community ecologistsand ecosystem ecologists in the context ofrestoration
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 6 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
Restoring flow regimes ratherthan eventsScientifically informed flow designs may existthat maintain ecosystem integrity in regulatedrivers (116 117) but these have produced mixedresults to date In the dam-altered Putah Creekin California mimicking the natural timing ofhigh and low flows enhanced native fish by im-proving their spawning habitat reducing watertemperatures maintaining flowing conditionsand reducing the abundance of non-native spe-cies (118) However other similar attempts havebeen less successful In some cases non-nativefish density also benefitted from flow patternsmimicking an unregulated regime (119) in othersnon-natives bounced back shortly after the flowrelease (120) These experiences illustrate thateven natural flow features may have inadvertentnegative effects if only part of the community orecosystem is considered Models for the NavajoDam (San Juan River New Mexico) show thatadequately timed releases could simultaneouslyenhance native and minimize non-native fishpopulations (121) thus predicted benefits couldsurpass those of mimicking a natural flow re-gime potentially persisting even during dryyears Another study using the ldquodesignerrdquo flowparadigm (114) attempted to understand howdifferent facets of flow variability control fisheryproduction in the Lower Mekong floodplains Time-
series models combining discharge and fisherycatch suggested that enhanced flood pulses andlong inter-flood intervals may increase fisheryyieldsAlthough environmental flow practices have
mostly focused on re-creating discrete flowevents (eg flood pulses) these can still serve asvaluable experiments An iconic example of alarge-scale flow release is Minute 319 an amend-ment to the United StatesndashMexico treaty allow-ing opening of floodgates in the Lower ColoradoRiver and a one-time rewetting of the parchedColorado River Delta (122) Although this pulsed-flow experiment did not promote establishmentof native woody riparian seedlings such trialscan inform future restoration practices for ex-ample by determining whether the provision ofbare moist substrate enhances seed germina-tion (123) Hypothesis-driven experimental flowreleases combined with deep knowledge of tar-get ecosystems can provide insights into resto-ration actions [eg (124)] Still an outstandingquestion is how flow regime restoration couldbe leveraged to restore other important alteredaspects of the ecosystem such as sediment andthermal regimes For example cold-water releasesby existing dams in climate-vulnerable basinscould prevent organisms that thrive at low tem-peratures from becoming replaced by general-ist warm-tolerant taxa (125)
Although the concept of environmental flowsis generally associated with impounded riversits relevance to urban streams is clear (126)However because frequent high storm flowsare viewed as the problem reducing them gen-erally becomes the only restoration goal This isoften attempted by shifting the spatial distribu-tion of velocities within channels through ma-nipulation of channel sinuosity installation offlow-diverting weirs or adding other structuresThese approaches may reduce local erosion butrarely result in levels of water quality ecosystemprocesses or biodiversity in urban streams thatcome close to those of unimpacted streams(127 128) Focusing on the channel ratherthan the watershed context at large means thatkey ecosystemprocesses can remain compromised(129) A recent study compared time series ofmetabolism in two streamsmdashonewith stormwaterponds in the watershed and the other restoredby altering channel slope banks and in-streamstructural complexity (103) The study found thatpeaks in GPP and ER in the ldquorestoredrdquo streamwere higher more prone to resets induced byflashy storms and that recovery time scales werelonger compared with the stream in a watershedwith stormwater measures (Fig 6) Ponds areone type of nature-inspired green infrastructureused to restore streamflow tomore natural condi-tions tree plantings constructed wetlands andgrass swales can in turn replace gray infrastruc-ture such as concrete tunnels that capture andstore waterDistributing green infrastructure in water-
sheds helps restore streams because it targetsthe cause of streamflow alteration includinginsufficient infiltration groundwater rechargeand more generally limited watershed storagecapacity (129) By tackling the cause rather thanthe symptoms of flow alteration these practicescan reduce ecological impacts that result fromthe cascading effects of altered flows includingpoor water quality and temperature spikes Re-search on the effectiveness of green infrastructureon restoring streams is sparse and limited bylack of data before and after implementationalthough studies are beginning to appear Forexample upland green infrastructure in theform of shallow wetland-like pools that capturestormwater and reconnect it to the groundwaterhas been shown to enhance baseflows while re-ducing peak flows and pollutant loadsmore thanrestoration structures within the mainstem ofthe receiving stream (130) Similar structuresbuilt between stormwater outfalls and perennialstreams did not modulate the timing magnitudeor duration of urban storm responses nor didthey improve water quality or biodiversity (131)illustrating the need for researchers to understandhow exact design and watershed context influ-ence performance
Spatiotemporal variability andrestoration designs
Research showing how local and watershed-scale environmental drivers vary temporally andalong river networks in ways that shape biota
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 7 of 13
Flow pulses (storms) flashy flowsDroughts intermittencyLowstable flowsAltered timing seasonality
DOC N P
DetritusMicrobial
stoichiometric constraints
Excretion exudates
Detrital pathway
Algal pathway
Algal stoichiometric constraints
Organismal dispersal source-sink dynamics
Algae
Community replacement
ndash ndash +
ndash ndash + ndash
+
+ndash +
Invasions
Turnover
Consumer stoichiometric constraints
Fig 5 Flow variation affects biota and ecosystem processes and food webs integrate bothHydrologic connectivity along river networks is a first landscape-level filter constraining dispersal oforganisms in river networks Extreme flow variation (floods and droughts) are a source of disturbanceto all organisms (gray box) but particularly to the higher trophic levels given their higher metabolicdemands In turn flow variation controls inputs of nutrients and terrestrial leaf litter to the stream whichinfluences microbial activity and the relative contributions of the detrital (brown) and algal (green)pathways in sustaining aquatic consumers Not just quantity but also food quality (eg CN and CPratios) influence dietary choices of consumers and thus trophic transfer efficiency Finally alteration offlow regimes may influence all these compartments and may also open niches for new players Ifbiological invasions occur at the top of the food web these have the potential to divert energy away fromnative predators and to alter food-web structure through top-down controls
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
and ecosystem processes can help prioritizeselection of sites for restoration For exampleusing the concept of river network saturation(55) to identify river sections where in-streamremoval of nutrients cannot keep up with highsource inputs of nutrients suggests that restorationactions targeting those sections may dispropor-tionately improve water quality Identification ofsubcatchments that disproportionately influencestream chemistry may also be informed by com-bining methods from landscape ecology andcatchment hydrology focusing on the spatialscale of variance in water chemistry (132) Head-waters are often prioritized for restoration be-
cause poor water quality in them can havedownstream impacts Research showing thatlocal environmental constraints on river com-munities are important in headwaters as well asin the larger channels but dispersal constraintsare much less important in the former (133 134)implies that headwaters should be prioritizedHowever research quantifying time-varying con-tributions of local communities to river-wide(network) biodiversity can also inform prioritiza-tion by identifying sites that host a dispropor-tionate share of species or that host uniquespeciesmdashfor example because they shelter sensi-tive taxa during critical flow bottlenecks This idea
encapsulates the concept of keystone habitatsor local sites that strongly influence metacom-munity dynamics (135) This has been appliedto aquatic invertebrates across intermittent rivernetworks by tracking time-varying site-specificcontributions to community dissimilarity (or betadiversity) (136)Other approaches to prioritize restoration ac-
tions may be based on contributions to system-wide stability rather than numbers of individualsor species For example the restoration of fall-run Chinook salmon portfolios may be assessedvia the contributions that different periods andlocationsmake tometapopulation-wide stabilityowing to spatiotemporal variation in habitatconditions (137) Additionally interventionscould promote conditions that are not impor-tant now but could be important in the future[ie proto-refugia (138)] For example resto-ration of groundwater-influenced habitats couldmitigate future stress to drought-sensitive taxaunder scenarios of increased intermittencyPatches of undisturbed land that protect flowregimes in streams within a larger catchmentthat is otherwise urbanized can be sources ofcolonists to restored sites Priorities that focuson present degradation of river sectionsmay failto address site contribution to impairment orrecovery at broader scales or under differentscenarios These insights indicate a great poten-tial for new methods to prioritize restoration ofboth urban and regulated streamsWe know little about how altered flow regimes
propagate through entire river networks butwork is starting to assess the cumulative effectsof dams on flow regimes as well as dam-levelcontributions to alteration Although these ap-proaches have largely focused on dam cascades[eg (139)] if expanded they could help identifyfavorable sections in river networks to developenvironmental flow operations Here exper-imentation could again help with testing ofnew strategies For instance large-scale flowexperiments could be designed that coordinatereleases among dams or between dams and free-flowing tributaries to enhance the restorativeeffects of natural pulses (140)
Hydrologic connectivityinfluences restoration
Recovering critical flow paths and their connec-tivity to rivers is important but it remains asubstantial challenge for flow-degraded riversand streams Whereas regulated rivers tend tohave reduced lateral and longitudinal hydro-logic connectivity urban streams have unnaturallevels of connectivity because pipes and over-land runoff route water quickly In both cases fun-damental hydroecological insights into streambedstructure landscape storage capacity and ground-water connectivity can help to envision novel re-storation approaches (130 141 142) More generallyhydrologic connectivity is becoming a frameworkto identify alternative ways to restore streams viastormwater management (143) For example arecent study suggested that reducing road densityand hydrologic connectivity between roads
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 8 of 13
ER
Disc
harg
e (f
t3 s)
GPPG
PP o
r ER
(g O
2 m-2
day
-1)
Discharge
Watershed BMPs
In-stream restoration
0
ndash4
4
0
ndash4
4
4
2
6
4
2
6
Jan Sept July May Mar Nov Jan
A
B C
Fig 6 Green infrastructure as a means to restore stream ecosystems (A) Discharge andecosystem respiration (ER more-negative values mean higher respiration) and gross primaryproduction (GPP) for two urban streams in which restoration efforts involved (top) modifications inthe stream channel and (bottom) traditional stormwater best management practices (BMPs)[Reproduced with permission from (103)] (B and C) Example of a comprehensive greeninfrastructure approach to ecologically restore an urban headwater stream that also supports thelifestyle of resident urban communities (B) Stream at the onset of restoration and (C) afterrestoration [Reproduced with permission from Greenworks Portland OR]
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 9 of 13
Table 1 Research challenges and opportunities Despite the progress that has been made in understanding how flow regimes affect biota and ecosystem
processes major challenges persist that prevent a complete understanding of the flow-biota-ecosystem processes nexus Making progress on these
challenges requires a mix of fundamental and applied research
Topic Research challenge Opportunity
Connecting
organismal to
ecosystem-level
processes
Research on the effects of flow on organisms
and ecosystem processes has followed parallel
tracksmdashhindering our mechanistic
understanding of the flow-biota-ecosystem
processes nexus
Generate theory that connects concepts and frameworks
that are well accepted in river community and ecosystem
ecology For example the notion of environmental filters
that connect regional to local composition via a combination
of dispersal and environmental conditions (community
ecology) with analogous theory on solute transport versus
reactivity (ecosystem ecology)
Our current understanding of the links between
phenology of biota and ecosystem processes
is precursory
Use high-frequency sensors to characterize metabolic
regimes at sites with ongoing organismal monitoring
Leverage citizen-science biodiversity data to detect
spatiotemporal shifts in animal phenology
(eg insect emergence fish spawning)
Use remote sensing tools to better understand
links between hydrology and aquatic and riparian
plant phenology
The link between stoichiometry and flow
variation remains largely unexplored
Repurpose data that is already being collected to test
flow-stoichiometry relationships eg studies using C and
N stable isotopes to describe CN ratios across flow
regime types Perform field experiments to quantify
indirect effects of invertebrate and fish consumption
on ecosystem metabolism
Ecological
consequences of
hydrologic
connectivity
The importance of anaerobic metabolism in
carbon processing as flows cease and parts of
streambeds dry is a major frontier research area
Create networks of sites across different geologies
and flow regimes that collect time-series data on
methane nitrous oxide and carbon dioxide fluxes
from intermittent streams
We ignore how biota in the wet phase is
influenced by processes and patterns
from the dry phase
Research the functions of dry riverbeds Use camera traps and
drone technology to investigate the role of dry riverbeds
as a dispersal corridor metabarcoding to quantify
ldquodark biodiversityrdquo in the hyporheic
Wetlands and their links to stream networks are poorly
delineated globally Even in the United States and
Europe current wetland inventories do not capture
these links yet understanding how their connectivity
to river systems influences network-scale ecosystem
processes is needed
Analyze time series from high-frequency sensors distributed in
river networks varying in wetland connectivity Further
development and application of time-series methods
used by neuroscientists
We need to identify the relationship between
varying flows and solute dynamics
associated with changes in catchment-scale
hydrologic exchanges
Design collaborative work among catchment hydrologists
and stream ecologists combining methods to estimate
transit times residence times and identify sources
across heterogeneous landscapes Link these properties to
land management and water quality aspects
Scaling and
transferring flow-
ecology relationships
across space and time
We ignore how biodiversity responds to
propagation of flow regime alteration
across river networks
Leverage extensive hydrometric networks (eg United States
Geological Survey National Water Information System)
to perform spatially replicated time-series analyses
on flow and use these to generate expectations
on how river network position should mediate
biodiversity responses to flow alteration
Flow-ecology relationships do not account for
nonstationarity and abrupt change
Use process-based models that account for mechanisms
and associated uncertainties When using time-series
data and models select methods that are robust
to nonstationarity Linear flow-ecology relationships
should not be assumed
Current flow-ecology relationships assume that
responses of species do not change over time
Consider flow regime alteration as a process with ecological and
evolutionary consequences Include intraspecific variation
in traits and potential for rapid evolution in models
continued on next page
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
and streams may improve water quality morethan other restoration options (70) Additionallyresearch on the age and lability of DOC enteringstreams has shown how precipitation or land dis-turbance can change watershed flow paths (53)and this could inspire new restoration approachesfocused on land use and soil properties Manystudies on hydrologic connectivity and networkstructure suggest a need for restoration to focuson landscape heterogeneity especially on thepresence ofwetlands lakes and ponds (144) Suchfeaturesare frequently lost inurbanizedcatchmentsand outcomes could be improved by restoring orincreasing connectivitywith these ecosystems (145)Hydrologic connectivity associated with ground-water inputs or abstractions land-use manage-ment and return flows fromwastewater treatmentplants also need to be considered in these initia-tives (146)Restoring hydrologic connectivity is also key
to achieving biodiversity outcomesmdashparticularlyin dammed systems Longitudinal connectivityfor fish inhabiting dam-altered rivers can beenhanced by the construction of fishways andsophisticated passage facilities However moni-toring data on the effectiveness of such inter-
ventions are still scarce and available evidencesuggests that passage effectivity largely dependson technology used and species-specific fish be-havior (147 148) Although trap-and-transportprograms for migratory fishes can help augmentthreatened populations (149) these practicesoften increase straying and thus the potentialfor genetic homogenization as seen from ju-venile salmon collected at dams and transporteddownstream to increase out-migration success(150) Overall a combination of engineering andmanagement solutions may be best to restorefunctional connectivity in dam-altered riversThe link between hydrologic connectivity andecological restoration is also central to con-siderations on where to build remove and re-operate hydrologic infrastructure Hundredsof dams have been intentionally removed sincethe 1970s and removal of large dams such asthe Elwha River Dam (Washington USA) hasshown that restoring longitudinal connectivitycan have immediate geomorphic effects with com-plex temporal dynamics (151) Infrastructure canalso be designed or reoperated to manage forrestoring lateral connectivity and enhanced flood-plain processes For example the Yolo Bypass in
the Sacramento River California has becomea prime rearing and migration site for chinooksalmon showing that flood control and habitatfor fish and wildlife do not need to be at odds(152) All these examples show the importanceof integrating hydrologic connectivity in ecolog-ical restoration whether the goal is to minimizethe ecological effects of fragmentation restorefree-flowing watercourses or operate green infra-structure to enhance infiltration floodplain con-nectivity or other ecological processes
Research challenges and opportunities
Two decades of concerted efforts to research theeffects of flow regime alteration have tremen-dously advanced our understanding of the waysin which streamflow influences biota and eco-system processes However important challengesremain These aremainly related to the scarcity ofscience connecting organismal to ecosystem-levelprocesses through for example microbial linksthe difficulty of studying watershed-level phys-ical and biogeochemical heterogeneity and itsinfluence on riverine processes and biota thecomplexity of parsing out the direct effects offlow from flow-mediated effects and co-occurring
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 10 of 13
Topic Research challenge Opportunity
Cascading
implications of
hydrologic
alteration
We have a poor understanding of the
interactions among physical drivers
Build on large-scale flow experiments to test hypotheses on the
combined effects of altered flow sediment and thermal
regimes Experiment with multilevel intakes in reservoirs
Promote collection of non-flow variables (eg temperature
water quality parameters) at hydrometric stations
At low flows biota are being exposed to high
concentrations of new classes of pollutants
whose impacts are poorly known especially if
biotic impacts interact with impacts on
ecosystem processes
Design realistic experimental studies to advance knowledge
on the ecological effects of multiple stressors
Collect long-term data at sites varying in contaminant
inputs and hydrologic regimes Link microbial to
organismal and ecosystem-level responses
Using hydroecological
science to inform
river ecosystem
restoration
Environmental flow science largely ignores
human dimensions
Use methods that allow identifying causal networks
in complex socioenvironmental systems Include
human needs and thresholds in research on
flow-ecology relationships
Environmental flows are generally prescribed
at the scale of single infrastructures restoration
designs and assessment are generally at reach scales
Promote coordinated large-scale flow experiments
and restoration designs at the river basin scale
Extend assessment of green infrastructure
effectiveness beyond individual structures to
infrastructure networks and link both
to river outcomes
Environmental flows in impounded rivers
generally target the conservation of some
particular species rather than outcomes at the
food-web or ecosystem level
Advance fundamental research on microbialndash
ecosystem process linkages Expand the notion
of designer flows to food-web structure and
ecosystem services when possible
Environmental flow science has largely
ignored surface waterndashgroundwater
interactions
Expand science to understand the link between
restoration actions groundwater recharge and
inputs to streams Provide science relevant to
the implementation of new legislation on
groundwater management (eg Sustainable
Groundwater Management Act in California)
Uncertainty regarding future
hydroclimate and
river flows is growing
Refine regional climate models Promote research on
the interaction between land-use change and
evapotranspiration Incorporate groundwater
influence in macroscale hydrology models
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
stressors (particularly in urban streams) and thechallenge of predicting outcomeswhen both thephysical and the biological environments arechangingAchieving a predictive understanding of hy-
droecological relationships is essential to design-ing successful self-sustaining restoration actionsin river ecosystems However translating basichydroecological insights into restoration practicerequires understanding the complexities of work-ing within a human-dominated water cycle Al-though it is rarely possible to restore all facetsof a natural flow regime ecologically informeddesigns may help mimic key aspects of the nat-ural flow regime (eg periodic pulse flows) orcreate regimes that are not natural but maximizeoutcomes given socioeconomic constraints InTable 1 we identify persisting and emergingchallenges in hydroecological research We paireach challenge with an opportunity (related toresearch design technology or implementation)We contend that these challenges should beprioritized to advance process-based river eco-system restoration
REFERENCES AND NOTES
1 N L Poff et al The natural flow regime Bioscience 47769ndash784 (1997) doi 1023071313099
2 D A Lytle N L Poff Adaptation to natural flow regimesTrends Ecol Evol 19 94ndash100 (2004) doi 101016jtree200310002 pmid 16701235
3 S Sabater A Elosegi R Ludwig in Multiple Stressors in RiverEcosystems (Elsevier 2019) pp 1ndash22
4 United Nations New UN Decade on Ecosystem Restorationoffers unparalleled opportunity for job creation food securityand addressing climate change (2019) httpswwwunenvironmentorgnews-and-storiespress-releasenew-un-decade-ecosystem-restoration-offers-unparalleled-opportunity
5 A W Lorenz P Haase K Januschke A SundermannD Hering Revisiting restored river reaches - Assessingchange of aquatic and riparian communities afterfive years Sci Total Environ 613-614 1185ndash1195(2018) doi 101016jscitotenv201709188pmid 28954379
6 M A Palmer K L Hondula B J Koch Ecological Restorationof Streams and Rivers Shifting Strategies and ShiftingGoals (2014) vol 45
7 T J Beechie et al Process-based principles for restoringriver ecosystems Bioscience 60 209ndash222 (2010)doi 101525bio20106037
8 S E Bunn A H Arthington Basic principles and ecologicalconsequences of altered flow regimes for aquaticbiodiversity Environ Manage 30 492ndash507 (2002)doi 101007s00267-002-2737-0 pmid 12481916
9 K D Fausch Y Taniguchi S Nakano G D GrossmanC R Townsend Flood disturbance regimes influence rainbowtrout invasion success among five holarctic regions EcolAppl 11 1438ndash1455 (2001) doi 1018901051-0761(2001)011[1438FDRIRT]20CO2
10 M E Power M S Parker W E Dietrich Seasonalreassembly of a river food web Floods droughts andimpacts of fish Ecol Monogr 78 263ndash282 (2008)doi 10189006-09021
11 V Acuntildea A Giorgi I Muntildeoz U R S Uehlinger S SabaterFlow extremes and benthic organic matter shape themetabolism of a headwater Mediterranean streamFreshwater Biol 49 960ndash971 (2004) doi 101111j1365-2427200401239x
12 K Tockner F Malard J V Ward An extension of the floodpulse concept Hydrol Processes 14 2861ndash2883 (2000)doi 1010021099-1085(20001112)141617lt2861AID-HYP124gt30CO2-F
13 J S Perkin et al Fragmentation and dewatering transformGreat Plains stream fish communities Ecol Monogr 8573ndash92 (2015) doi 10189014-01211
14 J L Sabo D M Post Quantifying periodic stochastic andcatastrophic environmental variation Ecol Monogr 7819ndash40 (2008) doi 10189006-13401
15 F Shuai et al Silver carp larva abundance in response toriver flow rate revealed by cross-wavelet modellingEcol Modell 383 98ndash105 (2018) doi 101016jecolmodel201805020
16 J D Tonkin M T Bogan N Bonada B Rios-ToumaD A Lytle Seasonality and predictability shape temporalspecies diversity Ecology 98 1201ndash1216 (2017)doi 101002ecy1761 pmid 28144975
17 N L Poff Beyond the natural flow regime Broadening thehydro‐ecological foundation to meet environmental flowschallenges in a non‐stationary world Freshwater Biol 631011ndash1021 (2018) doi 101111fwb13038
18 A Ruhi X Dong C H McDaniel D P Batzer J L SaboDetrimental effects of a novel flow regime on the functionaltrajectory of an aquatic invertebrate metacommunity GlobChange Biol 24 3749ndash3765 (2018) doi 101111gcb14133pmid 29665147
19 M T Bogan D A Lytle Severe drought drives novelcommunity trajectories in desert stream pools FreshwaterBiol 56 2070ndash2081 (2011) doi 101111j1365-2427201102638x
20 G Woodward et al The effects of climatic fluctuations andextreme events on running water ecosystems Philos TransR Soc London Ser B 371 20150274 (2016) pmid 27114576
21 M A Palmer C M Swan K Nelson P Silver R AlvestadStreambed landscapes Evidence that stream invertebratesrespond to the type and spatial arrangement of patchesLandsc Ecol 15 563ndash576 (2000) doi 101023A1008194130695
22 D Schmera et al Does isolation influence the relative role ofenvironmental and dispersal‐related processes in streamnetworks An empirical test of the network positionhypothesis using multiple taxa Freshwater Biol 63 74ndash85(2018) doi 101111fwb12973
23 R Henriques‐Silva et al A comprehensive examination of thenetwork position hypothesis across multiple rivermetacommunities Ecography 42 284ndash294 (2019)doi 101111ecog03908
24 K L Jaeger J D Olden N A Pelland Climate change poisedto threaten hydrologic connectivity and endemic fishes indryland streams Proc Natl Acad Sci USA 11113894ndash13899 (2014) doi 101073pnas1320890111pmid 25136090
25 S Garbin E A Celegon P Fanton G Botter Hydrologicalcontrols on river network connectivity R Soc Open Sci 6181428 (2019) doi 101098rsos181428 pmid 30891270
26 F Cattaneacuteo B Hugueny N Lamouroux Synchrony in browntrout Salmo trutta population dynamics A lsquoMoran effectrsquo onearly‐life stages Oikos 100 43ndash54 (2003) doi 101034j1600-0706200311912x
27 A Terui et al Metapopulation stability in branching rivernetworks Proc Natl Acad Sci USA 115 E5963ndashE5969(2018) doi 101073pnas1800060115 pmid 29895695
28 P A P Moran The statistical analysis of the Canadian Lynxcycle Aust J Zool 1 291ndash298 (1953) doi 101071ZO9530291
29 G D Grossman P B Moyle J O Whitaker Jr Stochasticityin structural and functional characteristics of an Indianastream fish assemblage A test of community theoryAm Nat 120 423ndash454 (1982) doi 101086284004
30 D A Lytle D M Merritt J D Tonkin J D OldenL V Reynolds Linking river flow regimes to riparian plantguilds A community-wide modeling approach Ecol Appl 271338ndash1350 (2017) doi 101002eap1528 pmid 28263426
31 A Ruhiacute E E Holmes J N Rinne J L Sabo Anomalousdroughts not invasion decrease persistence of native fishesin a desert river Glob Change Biol 21 1482ndash1496 (2015)doi 101111gcb12780 pmid 25430731
32 J D Tonkin D M Merritt J D Olden L V ReynoldsD A Lytle Flow regime alteration degrades ecologicalnetworks in riparian ecosystems Nat Ecol Evol 2 86ndash93(2018) doi 101038s41559-017-0379-0 pmid 29180707
33 M C Mims J D Olden Fish assemblages respond toaltered flow regimes via ecological filtering of lifehistory strategies Freshwater Biol 58 50ndash62 (2013)doi 101111fwb12037
34 P T J Johnson J D Olden M J Vander Zanden Daminvaders Impoundments facilitate biological invasions intofreshwaters Front Ecol Environ 6 357ndash363 (2008)doi 101890070156
35 N Bonada S Doledec B Statzner Taxonomic and biologicaltrait differences of stream macroinvertebrate communitiesbetween mediterranean and temperate regions Implicationsfor future climatic scenarios Glob Change Biol 131658ndash1671 (2007) doi 101111j1365-2486200701375x
36 M I Pyne N L Poff Vulnerability of stream communitycomposition and function to projected thermal warming andhydrologic change across ecoregions in the western UnitedStates Glob Change Biol 23 77ndash93 (2017) doi 101111gcb13437 pmid 27429092
37 D B Herbst S D Cooper R B Medhurst S W WisemanC T Hunsaker Drought ecohydrology alters the structureand function of benthic invertebrate communities inmountain streams Freshwater Biol 64 886ndash902 (2019)doi 101111fwb13270
38 M T Bogan E T Chester T Datry A L MurphyB J Robson A Ruhi R Stubbington J E Whitney inResistance Resilience and Community Recovery in IntermittentRivers and Ephemeral Streams T Datry N Bonada A BoultonEds (Academic Press 2017) pp 349ndash376
39 S Houmlckendorff et al Characterizing fish responses to a riverrestoration over 21 years based on speciesrsquo traits ConservBiol 31 1098ndash1108 (2017) doi 101111cobi12908pmid 28218807
40 D Bruno et al Structural and functional responses ofinvertebrate communities to climate change andflow regulation in alpine catchments Glob Change Biol25 1612ndash1628 (2019) doi 101111gcb14581pmid 30698905
41 T W H Aspin et al Extreme drought pushes streaminvertebrate communities over functional thresholds GlobChange Biol 25 230ndash244 (2019) doi 101111gcb14495pmid 30346098
42 S E Tank J B Fellman E Hood E S Kritzberg Beyondrespiration Controls on lateral carbon fluxes across theterrestrial-aquatic interface Limnol Oceanogr Lett 3 76ndash88(2018) doi 101002lol210065
43 A P Appling et al The metabolic regimes of 356 rivers inthe United States Sci Data 5 180292 (2018) doi 101038sdata2018292 pmid 30532078
44 P Savoy et al Metabolic rhythms in flowing watersAn approach for classifying river productivity regimesLimnol Oceanogr (2019) doi 101002lno11154
45 E S Bernhardt et al The metabolic regimes of flowingwaters Limnol Oceanogr 63 S99ndashS118 (2018)doi 101002lno10726
46 M Colls X Timoner S Sabater V Acuntildea Effects of durationand frequency of the non-flow period on stream metabolismEcosystems 101007s10021-019-00345-1 (2019)doi 101007s10021-019-00345-1
47 A J Ulseth E Bertuzzo G A Singer J Schelker T J BattinClimate-induced changes in spring snowmelt impactecosystem metabolism and carbon fluxes in an Alpine streamnetwork Ecosystems 21 373ndash390 (2018) doi 101007s10021-017-0155-7
48 B O L Demars Hydrological pulses and burning of dissolvedorganic carbon by stream respiration Limnol Oceanogr 64406ndash421 (2019) doi 101002lno11048
49 R T Hensley L Kirk M Spangler M N Gooseff M J CohenFlow extremes as spatiotemporal control points on riversolute fluxes and metabolism J Geophys Res Biogeosci124 537ndash555 (2019) doi 1010292018JG004738
50 J D Hosen et al Enhancement of primary production duringdrought in a temperate watershed is greater in larger riversthan headwater streams Limnol Oceanogr 64 1458ndash1472(2019) doi 101002lno11127
51 L Goacutemez-Gener et al When water vanishes Magnitude andregulation of carbon dioxide emissions from dry temporarystreams Ecosystems 19 710ndash723 (2016) doi 101007s10021-016-9963-4
52 E H Stanley et al The ecology of methane in streams andrivers Patterns controls and global significance EcolMonogr 86 146ndash171 (2016) doi 10189015-1027
53 R T Barnes D E Butman H F Wilson P A RaymondRiverine export of aged carbon driven by flow path depth andresidence time Environ Sci Technol 52 1028ndash1035 (2018)doi 101021acsest7b04717 pmid 29313674
54 G Pinay et al Riparian corridors A new conceptualframework for assessing nitrogen buffering across biomesFront Environ Sci 6 47 (2018) doi 103389fenvs201800047
55 W M Wollheim et al River network saturation conceptFactors influencing the balance of biogeochemical supply and
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 11 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
demand of river networks Biogeochemistry 141 503ndash521(2018) doi 101007s10533-018-0488-0
56 P A Raymond J E Saiers W V Sobczak Hydrological andbiogeochemical controls on watershed dissolved organicmatter transport Pulse-shunt concept Ecology 97 5ndash16(2016) doi 10189014-16841 pmid 27008769
57 C Tunaley D Tetzlaff J Lessels C Soulsby Linking high-frequency DOC dynamics to the age of connected watersources Water Resour Res 52 5232ndash5247 (2016)doi 1010022015WR018419
58 S Bernal et al Supply demand and in-stream retention ofdissolved organic carbon and nitrate during storms inMediterranean forested headwater streams Front EnvironSci 7 60 (2019) doi 103389fenvs201900060
59 J Harvey et al How hydrologic connectivity regulates waterquality in river corridors J Am Water Resour Assoc 55369ndash381 (2019) doi 1011111752-168812691
60 J C Moore et al Detritus trophic dynamics and biodiversityEcol Lett 7 584ndash600 (2004) doi 101111j1461-0248200400606x
61 A D Rosemond et al Freshwater ecology Experimentalnutrient additions accelerate terrestrial carbon loss fromstream ecosystems Science 347 1142ndash1145 (2015)doi 101126scienceaaa1958 pmid 25745171
62 A C Y Yeung J L Musetta-Lambert D P KreutzweiserP K Sibley J S Richardson Relations of interannualdifferences in stream litter breakdown with dischargeBioassessment implications Ecosphere 9 e02423 (2018)doi 101002ecs22423
63 R M Northington J R Webster Experimental reductions instream flow alter litter processing and consumer subsidies inheadwater streams Freshwater Biol 62 737ndash750 (2017)doi 101111fwb12898
64 E Mas-Martiacute I Sanpera-Calbet I Muntildeoz Bottom-up effectsof streambed drying on consumer performance throughchanges in resource quality Aquat Sci 79 719ndash731 (2017)doi 101007s00027-017-0531-6
65 B D Richter G A Thomas Restoring environmental flows bymodifying dam operations Ecol Soc 12 12 (2007)doi 105751ES-02014-120112
66 G Zolezzi A Siviglia M Toffolon B MaioliniThermopeaking in Alpine streams Event characterizationand time scales Ecohydrology 4 564ndash576 (2011)doi 101002eco132
67 T Meiszligner B Sures C K Feld Multiple stressors and therole of hydrology on benthic invertebrates in mountainousstreams Sci Total Environ 663 841ndash851 (2019)doi 101016jscitotenv201901288 pmid 30738264
68 I R Waite et al Effects of urban multi-stressors on threestream biotic assemblages Sci Total Environ 6601472ndash1485 (2019) doi 101016jscitotenv201812240pmid 30743940
69 K C Nelson M A Palmer Stream temperature surges underurbanization and climate change Data models andresponses J Am Water Resour Assoc 43 440ndash452 (2007)doi 101111j1752-1688200700034x
70 J R Blaszczak J M Delesantro Y Zhong D L UrbanE S Bernhardt Watershed urban development controls onurban streamwater chemistry variability Biogeochemistry144 61ndash84 (2019) doi 101007s10533-019-00572-7
71 L Ponsatiacute et al Biofilm responses to flow regulation by damsin Mediterranean rivers River Res Appl 31 1003ndash1016(2015) doi 101002rra2807
72 J M Alberts J J Beaulieu I Buffam Watershed land useand seasonal variation constrain the influence of ripariancanopy cover on stream ecosystem metabolism Ecosystems20 553ndash567 (2017) doi 101007s10021-016-0040-9pmid 30416371
73 R O Hall Jr et al Turbidity light temperature andhydropeaking control primary productivity in the ColoradoRiver Grand Canyon Limnol Oceanogr 60 512ndash526 (2015)doi 101002lno10031
74 J R Blaszczak J M Delesantro D L Urban M W DoyleE S Bernhardt Scoured or suffocated Urban streamecosystems oscillate between hydrologic and dissolvedoxygen extremes Limnol Oceanogr 64 877ndash894 (2019)doi 101002lno11081
75 J-R Mor et al Dam regulation and riverine food-webstructure in a Mediterranean river Sci Total Environ 625301ndash310 (2018) doi 101016jscitotenv201712296pmid 29289778
76 H A Wellard Kelly et al Macroinvertebrate diets reflecttributary inputs and turbidity-driven changes in food
availability in the Colorado River downstream of GlenCanyon Dam Freshw Sci 32 397ndash410 (2013)doi 10189912-0881
77 S J Imberger R M Thompson M R Grace Urbancatchment hydrology overwhelms reach scale effects ofriparian vegetation on organic matter dynamics FreshwaterBiol 56 1370ndash1389 (2011) doi 101111j1365-2427201102575x
78 R T Martins A S Melo J F Gonccedilalves Jr N HamadaLeaf-litter breakdown in urban streams of Central AmazoniaDirect and indirect effects of physical chemical andbiological factors Freshw Sci 34 716ndash726 (2015)doi 101086681086
79 C M Yule J Y Gan T Jinggut K V Lee Urbanizationaffects food webs and leaf-litter decomposition in a tropicalstream in Malaysia Freshw Sci 34 702ndash715 (2015)doi 101086681252
80 T A Newcomer et al Influence of natural and novel organiccarbon sources on denitrification in forest degraded urbanand restored streams Ecol Monogr 82 449ndash466 (2012)doi 10189012-04581
81 A Chaudhary I Kauser A Ray R Poretsky Taxon-drivenfunctional shifts associated with storm flow in an urbanstream microbial community MSphere 3 e00194-18 (2018)doi 101128mSphere00194-18 pmid 29976643
82 T A Kennedy et al Flow management for hydropowerextirpates aquatic insects undermining river food websBioscience 66 561ndash575 (2016) doi 101093bioscibiw059
83 A Glazaczow D Orwin M Bogdziewicz Increasedtemperature delays the late-season phenology of multivoltineinsect Sci Rep 6 38022 (2016) doi 101038srep38022pmid 27905493
84 S G Fisher L J Gray N B Grimm D E Busch Temporalsuccession in a desert stream ecosystem following flashflooding Ecol Monogr 52 93ndash110 (1982) doi 1023072937346
85 G J Scrimgeour M J Winterbourn Effects of floods onepilithon and benthic macroinvertebrate populations in anunstable New Zealand river Hydrobiologia 171 33ndash44 (1989)doi 101007BF00005722
86 E Bondar-Kunze S Maier D Schoumlnauer N Bahl T HeinAntagonistic and synergistic effects on a stream periphytoncommunity under the influence of pulsed flow velocityincrease and nutrient enrichment Sci Total Environ 573594ndash602 (2016) doi 101016jscitotenv201608158pmid 27585428
87 M J Cashman G L Harvey G Wharton M C Bruno Woodmitigates the effect of hydropeaking scour on periphytonbiomass and nutritional quality in semi-natural flumesimulations Aquat Sci 79 459ndash471 (2017) doi 101007s00027-016-0510-3
88 W F Cross et al Food-web dynamics in a large riverdiscontinuum Ecol Monogr 83 311ndash337 (2013)doi 10189012-17271
89 D M Warfe et al Productivity disturbance and ecosystemsize have no influence on food chain length in seasonallyconnected rivers PLOS ONE 8 e66240 (2013)doi 101371journalpone0066240
90 T D Jardine et al Consumer-resource coupling in wet-drytropical rivers J Anim Ecol 81 310ndash322 (2012)doi 101111j1365-2656201101925x pmid 22103689
91 K O Winemiller et al Pulsing hydrology determines top‐down control of basal resources in a tropical riverndashfloodplainecosystem Ecol Monogr 84 621ndash635 (2014)doi 10189013-18221
92 D M Post The long and short of food-chain length TrendsEcol Evol 17 269ndash277 (2002) doi 101016S0169-5347(02)02455-2
93 D M Post G Takimoto Proximate structural mechanismsfor variation in food‐chain length Oikos 116 775ndash782 (2007)doi 101111j0030-1299200715552x
94 P A McHugh A R McIntosh P G Jellyman Dual influencesof ecosystem size and disturbance on food chain length instreams Ecol Lett 13 881ndash890 (2010) doi 101111j1461-0248201001484x pmid 20482579
95 A Ruhiacute et al Flow regulation increases food-chain lengththrough omnivory mechanisms in a Mediterranean rivernetwork Freshwater Biol 61 1536ndash1549 (2016) doi 101111fwb12794
96 J L Sabo J C Finlay T Kennedy D M Post The role ofdischarge variation in scaling of drainage area and food chainlength in rivers Science 330 965ndash967 (2010) doi 101126science1196005 pmid 20947729
97 A R McIntosh et al Capacity to support predators scaleswith habitat size Sci Adv 4 eaap7523 (2018) doi 101126sciadvaap7523 pmid 29978038
98 J H Brown J F Gillooly A P Allen V M Savage G B WestToward a metabolic theory of ecology Ecology 851771ndash1789 (2004) doi 10189003-9000
99 K Zou E Theacutebault G Lacroix S Barot Interactions betweenthe green and brown food web determine ecosystemfunctioning Funct Ecol 30 1454ndash1465 (2016) doi 1011111365-243512626
100 M J Vanni P B McIntyre Predicting nutrient excretion ofaquatic animals with metabolic ecology and ecologicalstoichiometry A global synthesis Ecology 97 3460ndash3471(2016) doi 101002ecy1582 pmid 27912023
101 T B Parr K A Capps S P Inamdar K A Metcalf Animal-mediated organic matter transformation Aquatic insects as asource of microbially bioavailable organic nutrients andenergy Funct Ecol 33 524ndash535 (2019) doi 1011111365-243513242
102 C P Arango et al Urban infrastructure influences dissolvedorganic matter quality and bacterial metabolism in an urbanstream network Freshwater Biol 62 1917ndash1928 (2017)
103 L G Larsen J W Harvey Disrupted carbon cycling inrestored and unrestored urban streams Critical timescalesand controls Limnol Oceanogr 62 S160ndashS182 (2017)doi 101002lno10613
104 H M Halvorson G White J T Scott M A Evans-WhiteDietary and taxonomic controls on incorporation of microbialcarbon and phosphorus by detritivorous caddisfliesOecologia 180 567ndash579 (2016) doi 101007s00442-015-3464-6 pmid 26497125
105 J J Elser et al Biological stoichiometry from genes toecosystems Ecol Lett 3 540ndash550 (2000) doi 101046j1461-0248200000185x
106 E K Moody et al Consumption explains intraspecificvariation in nutrient recycling stoichiometry in a desert fishEcology 99 1552ndash1561 (2018) doi 101002ecy2372pmid 29882955
107 N Welti et al Bridging food webs ecosystem metabolismand biogeochemistry using ecological stoichiometry theoryFront Microbiol 8 1298 (2017) doi 103389fmicb201701298 pmid 28747904
108 M A Palmer S Filoso R M Fanelli From ecosystems toecosystem services Stream restoration as ecologicalengineering Ecol Eng 65 62ndash70 (2014) doi 101016jecoleng201307059
109 M J Burns T D Fletcher C J Walsh A R LadsonB E Hatt Hydrologic shortcomings of conventional urbanstormwater management and opportunities for reformLandsc Urban Plan 105 230ndash240 (2012) doi 101016jlandurbplan201112012
110 C R Liermann C Nilsson J Robertson R Y NgImplications of dam obstruction for global freshwater fishdiversity Bioscience 62 539ndash548 (2012) doi 101525bio20126265
111 J D Olden R J Naiman Incorporating thermal regimes intoenvironmental flows assessments Modifying dam operationsto restore freshwater ecosystem integrity Freshwater Biol55 86ndash107 (2010) doi 101111j1365-2427200902179x
112 D B Osmundson R J Ryel V L Lamarra J Pitlick Flowndashsedimentndashbiota relations Implications for river regulationeffects on native fish abundance Ecol Appl 12 1719ndash1739(2002)
113 D von Schiller et al Regulation causes nitrogen cyclingdiscontinuities in Mediterranean rivers Sci Total Environ540 168ndash177 (2016) doi 101016jscitotenv201507017pmid 26233271
114 J L Sabo et al Designing river flows to improve foodsecurity futures in the Lower Mekong Basin Science 358eaao1053 (2017) doi 101126scienceaao1053pmid 29217541
115 W Xu et al Improving the multi-objective performance ofrainwater harvesting systems using real-time controltechnology Water 10 147 (2018) doi 103390w10020147
116 M Acreman et al Environmental flows for naturalhybrid and novel riverine ecosystems in a changingworld Front Ecol Environ 12 466ndash473 (2014)doi 101890130134
117 N L Poff et al Sustainable water management under futureuncertainty with eco-engineering decision scaling Nat ClimChang 6 25ndash34 (2016) doi 101038nclimate2765
118 J D Kiernan P B Moyle P K Crain Restoring native fishassemblages to a regulated California stream using the
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 12 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
natural flow regime concept Ecol Appl 22 1472ndash1482(2012) doi 10189011-04801 pmid 22908707
119 D L Propst K B Gido Responses of native andnonnative fishes to natural flow regime mimicry in theSan Juan River Trans Am Fish Soc 133 922ndash931 (2004)doi 101577T03-0571
120 T K Pool J D Olden Assessing long‐term fish responses andshort‐term solutions to flow regulation in a dryland river basinEcol Freshwat Fish 24 56ndash66 (2015) doi 101111eff12125
121 W Chen J D Olden Designing flows to resolve human andenvironmental water needs in a dam-regulated riverNat Commun 8 2158 (2017) doi 101038s41467-017-02226-4 pmid 29255194
122 S M Nelson et al A history of the 2014 Minute 319environmental pulse flow as documented by fieldmeasurements and satellite imagery Ecol Eng 106733ndash748 (2017) doi 101016jecoleng201610040
123 P B Shafroth et al A large-scale environmental flowexperiment for riparian restoration in the Colorado RiverDelta Ecol Eng 106 645ndash660 (2017) doi 101016jecoleng201702016
124 E P Glenn P L Nagler P B Shafroth C J JarchowEffectiveness of environmental flows for riparian restorationin arid regions A tale of four rivers Ecol Eng 106 695ndash703(2017) doi 101016jecoleng201701009
125 S Hotaling D S Finn J Joseph Giersch D W WeisrockD Jacobsen Climate change and alpine stream biologyProgress challenges and opportunities for the futureBiol Rev Camb Philos Soc 92 2024ndash2045 (2017)doi 101111brv12319 pmid 28105701
126 C J Walsh T D Fletcher M J Burns Urban stormwaterrunoff A new class of environmental flow problem PLOS ONE7 e45814 (2012) doi 101371journalpone0045814pmid 23029257
127 A Selvakumar T P OrsquoConnor S D Struck Role of streamrestoration on improving benthic macroinvertebrates and in-stream water quality in an urban watershed Case studyJ Environ Eng 136 127ndash139 (2010) doi 101061(ASCE)EE1943-78700000116
128 R W Lammers B P Bledsoe What role does streamrestoration play in nutrient management Crit Rev EnvironSci Technol 47 335ndash371 (2017) doi 1010801064338920171318618
129 H E Golden N Hoghooghi Green infrastructure and itscatchment-scale effects An emerging science WIREs Water5 e1254 (2018) doi 101002wat21254 pmid 29682288
130 M R Williams G Bhatt S Filoso G Yactayo Streamrestoration performance and its contribution to theChesapeake Bay TMDL Challenges posed by climate changein urban areas Estuaries Coasts 40 1227ndash1246 (2017)doi 101007s12237-017-0226-1
131 R M Fanelli K Prestegaard M A Palmer Freshw Sci101086706072 (2019)
132 B W Abbott et al Unexpected spatial stability of waterchemistry in headwater stream networks Ecol Lett 21296ndash308 (2018) doi 101111ele12897 pmid 29282860
133 B L Brown C M Swan Dendritic network structureconstrains metacommunity properties in riverineecosystems J Anim Ecol 79 571ndash580 (2010) doi 101111j1365-2656201001668x pmid 20180874
134 C M Swan B L Brown Metacommunity theory meetsrestoration Isolation may mediate how ecologicalcommunities respond to stream restoration Ecol Appl 272209ndash2219 (2017) doi 101002eap1602 pmid 28718193
135 N Mouquet D Gravel F Massol V Calcagno Extending theconcept of keystone species to communities and ecosystemsEcol Lett 16 1ndash8 (2013) doi 101111ele12014 pmid 23062191
136 A Ruhiacute T Datry J L Sabo Interpreting beta-diversitycomponents over time to conserve metacommunities inhighly dynamic ecosystems Conserv Biol 31 1459ndash1468(2017) doi 101111cobi12906 pmid 28188969
137 L Yamane L W Botsford D P Kilduff Tracking restorationof population diversity via the portfolio effect J Appl Ecol55 472ndash481 (2018) doi 1011111365-266412978
138 D W Braidwood M A Taggart M Smith R AndersenTranslocations conservation and climate change Use ofrestoration sites as protorefuges and protorefugia RestorEcol 26 20ndash28 (2018) doi 101111rec12642
139 Y Wang D Wang Q W Lewis J Wu F Huang A frameworkto assess the cumulative impacts of dams on hydrologicalregime A case study of the Yangtze River Hydrol Processes31 3045ndash3055 (2017) doi 101002hyp11239
140 J L Sabo et al Pulsed flows tributary inputs and food-webstructure in a highly regulated river J Appl Ecol 551884ndash1895 (2018) doi 1011111365-266413109
141 C Magliozzi G Coro R C Grabowski A I PackmanS Krause A multiscale statistical method to identifypotential areas of hyporheic exchange for river restorationplanning Environ Model Softw 111 311ndash323 (2019)doi 101016jenvsoft201809006
142 S S Schwartz B Smith Restoring hydrologic function inurban landscapes with suburban subsoiling J Hydrol 543770ndash781 (2016) doi 101016jjhydrol201610051
143 C J Walsh T D Fletcher A R Ladson Stream restoration inurban catchments through redesigning stormwater systemsLooking to the catchment to save the stream J N AmBenthol Soc 24 690ndash705 (2005) doi 10189904-0201
144 N M Schmadel et al Thresholds of lake and reservoirconnectivity in river networks control nitrogen removalNat Commun 9 2779 (2018) doi 101038s41467-018-05156-x pmid 30018449
145 C D Sayer Conservation of aquatic landscapes Pondslakes and rivers as integrated systems WIREs Water 1573ndash585 (2014) doi 101002wat21045
146 R G Luthy D L Sedlak M H Plumlee D Austin V H ReshWastewater‐effluent‐dominated streams as ecosystem‐
management tools in a drier climate Front Ecol Environ 13477ndash485 (2015) doi 101890150038
147 D W Roscoe S G Hinch Effectiveness monitoring of fishpassage facilities Historical trends geographic patterns andfuture directions Fish Fish 11 12ndash33 (2010) doi 101111j1467-2979200900333x
148 C M Bunt T Castro-Santos A Haro Performanceof fish passage structures at upstream barriers tomigration River Res Appl 28 457ndash478 (2012)doi 101002rra1565
149 D Weigel et al Evaluation of a trap-and-transport programfor a threatened population of steelhead (Oncorhynchusmykiss) Conserv Genet 20 1195ndash1199 (2019) doi 101007s10592-019-01200-5
150 M H Bond et al Combined effects of barge transportationriver environment and rearing location on straying andmigration of adult Snake River fall-run Chinook SalmonTrans Am Fish Soc 146 60ndash73 (2017) doi 1010800002848720161235614
151 A C Ritchie et al Morphodynamic evolution followingsediment release from the worldrsquos largest dam removalSci Rep 8 13279 (2018) doi 101038s41598-018-30817-8pmid 30185796
152 T Sommer et al Californiarsquos Yolo Bypass Evidence thatflood control can be compatible with fisheries wetlandswildlife and agriculture Fisheries 26 6ndash16 (2001)doi 1015771548-8446(2001)026lt0006CYBgt20CO2
153 G Grill et al Mapping the worldrsquos free-flowing riversNature 569 215ndash221 (2019) doi 101038s41586-019-1111-9pmid 31068722
154 G Gassert M Luck M Landis P Reig T Shiao ldquoAqueductglobal maps 21 Constructing decision-relevant global waterrisk indicatorsrdquo Working paper World Resources Institute(2015) wwwwriorgpublicationaqueduct-global-maps-21-indicators
155 A Ruhiacute J D Olden J L Sabo Declining streamflow inducescollapse and replacement of native fishes in the AmericanSouthwest Front Ecol Environ 14 465ndash472 (2016)doi 101002fee1424
156 T C Gouhier A Grinsted V Simko R package Biwaveletconduct univariate and bivariate wavelet analyses (2018)
157 T Rodriacuteguez-Castillo E Esteacutevez A M Gonzaacutelez-FerrerasJ Barquiacuten Estimating ecosystem metabolism toentire river networks Ecosystems (2018) doi 101007s10021-018-0311-8
ACKNOWLEDGMENTS
We greatly appreciate those who read and provided importantinsights on drafts of this paper A Armstrong K HondulaJ Hosen K Leathers G de Mendoza C Maietta I MunozJ Tonkin and two anonymous reviewers We also thank T Tu forhelp with Fig 1 and S Carlson L Comte T Datry J HosenM Kondolf N Jones D Lytle E Moody J Olden M PowerJ Sabo and M Sanchez-Montoya for valuable discussionsCompeting interests The authors have no competing interestsFunding MAP was supported by funds from the NSF (1639145)and AR was supported by funds from the University of CaliforniaBerkeley and NSF (1802714)
101126scienceaaw2087
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 13 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
restorationLinkages between flow regime biota and ecosystem processes Implications for river
Margaret Palmer and Albert Ruhi
DOI 101126scienceaaw2087 (6459) eaaw2087365Science
this issue p eaaw2087Scienceflow variability and achieve hydrological connectivity between a river and its surroundingsand ecosystem processes To be successful such efforts must go beyond accounting for flood pulses to restore natural mimic ecologically important aspects of natural flow regimes guided by insights into how variations in flow affect biotawater quality and ecological processes In a Review Palmer and Ruhi explain how restoration designs now attempt to
Human activities have altered the flow regimes of many of Earths rivers with negative impacts on biodiversityRiver restoration guided by research
ARTICLE TOOLS httpsciencesciencemagorgcontent3656459eaaw2087
REFERENCES
httpsciencesciencemagorgcontent3656459eaaw2087BIBLThis article cites 147 articles 7 of which you can access for free
PERMISSIONS httpwwwsciencemagorghelpreprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAASScienceScience 1200 New York Avenue NW Washington DC 20005 The title (print ISSN 0036-8075 online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
Science No claim to original US Government WorksCopyright copy 2019 The Authors some rights reserved exclusive licensee American Association for the Advancement of
on May 27 2020
httpsciencesciencem
agorgD
ownloaded from

apical predators are at least partially supportedby it (60) Macroconsumers break up particulateorganic matter (POM) into small pieces enhanc-ing the overall mineralization rate by bacteriaand fungi and even moderate flows may resultin export of OM particularly fine POM resultingfrommacroconsumer shredding activities Over-all howevermicrobes largely drive C losses throughrespiration (61) and although nutrient availa-bility temperature and POM composition (egleaf type) influence this how flow mediates rela-tionships has been more difficult to pin downRecent work (62) designed to separate the effectsthat different components of the flow regimehave on the mechanisms driving decompositionin perennial streams found that dissolution andmicrobial processing consistently exceeded theeffects of fragmentation and microbial pro-cessing was best explained by daily variabilityin discharge When flow regimes become in-termittent the lack ofwater can impedemicrobialand consumer colonization of POM resulting indecreases in decomposition the extent of whichis related to the duration of the drying (46 63)Consistent water cover is therefore importantto decomposition and thus shifts toward lowerflows when litter inputs are high (eg in thefall) can have a large impact on consumers andassociated food webs (63 64)
When flows are altered are changes inbiota and ecosystem processes linked
To help ground this question in the world ofrestoration practice we begin with a focus ontwo of the most common forms of river flow al-teration damming and urbanization For theformer we emphasize hydropower dams becausethey are growing in number they strongly alterflow regimes and the mitigation of their impactshas spurred the field of environmental flow re-search We also focus on urban streams becausestudies on the impacts of imperviousness havebeen extensive and these streams have been the
target of numerous restoration efforts We summa-rize knowledge on the effects of flow on organ-isms ecosystem processes and their interaction inthese two types of river systems and then describehow this can inform river restoration in general
Urbanization and dams changeflow regimes
With few exceptions flow regimes below hydro-power dams and in urban streams are highlyaltered at one or both ends of the flow spectrumRegimes in impounded rivers are typically lessseasonal than daily rainfall would suggest par-ticularly below large hydropower dams with highwater-storage capacity rather than below smallerless impactful run-of-the-river facilities (65)Hydro-power dams canmute peak river flows but power-generation cycles induce rapidly fluctuating flowpatterns (hydropeaking) that often occur onhuman-relevant scales [ie hours to weeks (18)]tending to bemore pronounced in seasons of highenergy demand Hydropeaking is often accom-panied by spikes in temperature (thermopeak-ing) either cold or warm depending on seasonand reservoir operation (66) Thus downstreamorganisms and ecosystems are generally subjectto multiple periodic stresses In urban streamsbaseflows can be higher or lower depending onwatershed characteristics and infrastructurehowever peak flows are invariably larger thanthose prior to urbanization Understanding theecological impacts of urban stormflow is com-plicated by indirect effects on water quality In-creases in flow magnitude frequency and ratesof change during storms leads to elevated con-centrations of pollutants Despite this studiesseparating the causes of biological impairmenthave reported that the unique effects of alteredurban flows can be significant (67 68) Spikesin stream temperature caused by runoff fromhot pavement (69) add to stress on the biotaHydropeaking and urban stormflows might
be expected to reduce primary production by
scouring algae (70) yet some studies have re-ported elevated GPP or alternating periods ofnet autotrophy and heterotrophy (71ndash73) Lightcan override the effects of flow on metabolismby enhancing algal photosynthesis (74) how-ever if hydropeaking increases turbidity GPPmay decline (73) In urban streams elevatedGPP is often attributed to open canopies andorto elevated nutrient concentrations both of whichoften co-occur with their altered flow regimes(72) As discussed later such shifts in primaryproductivity have been linked to changes introphic structure eg more algal-based foodwebs (75) depending on the relative availabil-ity of terrestrial carbon inputs (both DOM andPOM) (76) Inputs of POM and its quality as afood resource changewith landscape context (egvegetative cover) and catchment hydrology (77)but POM availability can also be limited by flowvariability which can increase decompositionrates and decrease residence times (78 79)As is the case for running-water systems in
general high flows in urban streams bring pulsesof solutes including N DOM and various pol-lutants (80) Solute dynamics and biogeochemicalprocesses in regulated rivers have been insuffi-ciently studied with researchers focusing moreon such processes in reservoirs rather than indownstream river ecosystems We found nostudies attempting to link urban or regulatedflow regimes and solute fluxes to changes infood webs or stream metabolic balance How-ever recent studies show shifts in microbial tax-onomic and functional composition in responseto pulsed urban flows (81) and thesemay help inexploring a potential link but are complicated bythe fact that non-flow stressors affect consumercomposition For example urban and regulatedrivers typically have animal communities thatare less diverse and composed of tolerant groupsoften non-native Hydropeaking and urban stormflows can certainly cause direct mortality via dis-placement and transport to unsuitable habitatsbut many of the organismal effects are realizedthrough indirect pathways For instance hydro-peaking reduces the viability of river-edge egg-laying specialists by limiting their access to suitablespawning habitat disrupting reproductive successand insect emergence (82) Flow regulation caneven create ecological traps if organismal phenol-ogy shiftswith thenovel environmental conditionsFor example dam-induced warming triggeredsummer diapause in a mayfly resulting in theloss of its last generation (83) Dams also isolatepopulations harming organisms that requiremigrations to complete their life cycles whetherthese are between riverine and marine habitatsor within the freshwater domain By analogy al-tered flow regimes in urban streams can also leadto isolated pools in streams with lowered base-flows This can be associated with periods ofhypoxia punctuated by extreme flow pulses inwhich these streams become ldquoscoured or suffo-catedrdquo (74)As illustrated using urban streams and reg-
ulated rivers research is moving forward andhints at the three-way nexus but is clearly still
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 5 of 13
Fig 4 Ecosystem metabolismpatterns across a river networkSpatial patterns in river eco-system metabolism for the Deva-Cares catchment in northernSpain as represented by the ratioof gross primary production(P) to ecosystem respiration (R)A PR gt 1 indicates autotrophicprocesses dominate so theseriver sections are accumulatingor exporting organic carbonPR lt 1 indicates dominanceby heterotrophic processes inriver sections that are receivingorganic C input (eg fromterrestrial sources) The boxesindicate regions of the networkimpacted by human pressures including deforestation (brown box) waste products from urbansites (green box) and a combination of the two (purple box) Such visualizations are usefulto managers in understanding regions of concern that may need the most restoration actions[Reproduced with permission from (157)]
PR ratiolt025
gt150
025 ndash 050 050 ndash 075075 ndash 100100 ndash 150
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
inadequate to meet restoration needs Studieson ecosystem processes below dams are parti-cularly limited and much of what we knowcomes from only a few river systems Furtherpublished work focuses primarily on benthicecosystem processes immediately below damsrather than riverine planktonic realms whereproductivity in large regulated rivers can beimportant
Linking flow biota andecosystem processes
Research on flow regimes that bridges organis-mal and ecosystem ecology has traditionallyrelied on states instead of rates ie nutrientconcentrations rather than uptake or fluxesstanding stocks of algae rather than productivityor metabolism and abundance of organismsrather than secondary productivity This workgenerally shows that flow variability suppressesand stability increases algal standing stock andgrazer densities with peaks in invertebrate bio-mass following those of algae [eg (84 85)]Over the past decade research began to focuson how flow regimes alter functional aspects offoodwebs For example biofilms inMediterranean-climate rivers are thicker andmetabolicallymoreactive downstream of flow-stabilizing dams (71)When flow variability increases below hydro-power dams fluctuations can reduce biofilmdevelopment and its quality as a food resource(86) Using experimental flumes subjected tohydropeaking shifts have been found in periph-yton fatty acid content from important highlyunsaturated compounds to nonessential saturatedones (87)Indirect evidence suggests that flow-induced
changes in dominant resource type (eg algaeversus detritus) resource quantity (biomass) orresource quality (nutrient content) can propa-gate to higher trophic levels via bottom-up ef-fects For instance detritivorous invertebrateswere found to dominate species richness andinvertebrate biomass upstream but not down-stream of an irrigation dam (75) diets of om-nivore insects indicated that flow stabilizationshifted a detrital into an algal-fueled food webSimilar evidence comes from Glen Canyon Damin the Colorado River (88) Near the dam whereprimary production is high (73) food webs aresimplified and gut content analyses have sug-gested algal production supports gt50 of in-vertebrate and gt70 of fish production andthe food web can be easily perturbed by flooddisturbance Farther downriver food webs showincreased reliance on detrital resources havehigher trophic efficiency and are more complexand more resistant to disturbance (76) In con-trast recent work on seasonal tropical riversfound algal productivity and zero-flow distur-bance did not explain food-web structure Inthis case it seems that highly mobile predatorsbuffered the local effects of drought (89) Thisstudy along with earlier ones at the site (90) isof particular interest because it links temporalpatterns in predatory fish with flow-dependentavailability of resources
Only a few studies have provided evidence oftop-down controls on foodwebs being influencedby flow variation Fish in a tropical floodplainriver were found to exert top-down control onprimary producers but only under certain hydro-logic conditions (91) Similarly in aCalifornian riverwinter flood pulses were shown to control insectconsumers late in the season ultimately deter-mining whether fish exerted top-down controlon the lower trophic levels (10) These illustratethe importance of pulsed hydrology on resource-consumer linkages and show that not just standingstocks but also the dominant controls structuringfood webs may fluctuate over time in response toflow variationAn indirect line of evidence supporting the
importance of the flow-biota-ecosystem processesnexus comes from theoretically oriented researchon food-web structure Changes in productivityhabitat size and disturbance underpin the basisof the three main hypotheses of food-chainlength which state that chains should be longerin stable productive large ecosystems (92) No-tably flow regimes influence these three con-trols Because food-chain length determines keyaspects of a river ecosystem such as the risk oftrophic cascades or the pollutant accumulationin top predators understanding how it is af-fected by flow alteration is important How-ever understanding the interplay between thehypotheses of food-chain length and the flow-dependent biological mechanisms that allowfood webs to shift in structure (eg predatorinvasions local extinctions consumers chang-ing diets) is still a relatively recent endeavor(93ndash95) Evidence suggests that zero-flows short-en food chains mainly by keeping streamsfishless (94 96) whereas stabilization of variableflows can lengthen food chains by allowing om-nivores to feed on abundant high trophic levelprey (95)New studies are also focusing on interactions
between drivers of food-web structure that hadbeen previously only been examined in isolationsuch as disturbance regimes and ecosystem sizeboth influenced by flow regime Using metabolictheory predator biomass has been shown toscale with both prey resources and fluctuatingstream habitat size with reductions in preda-tor size leading to declines in predator biomasssupported per unit of prey biomass (97) Movingforward the combination of habitat andmetabolicconstraints on predators may provide a basis toconnect flow-induced disturbance changes inecosystem processes and habitat fluctuationsto riverine food-web structure The metabolictheory of ecology (98) is a unifying frameworkthat allows scaling responses of organisms towhole ecosystems andmay become particularlyuseful in situations where flow alteration affectsorganismal body size or water temperature Insuch cases strong changes in metabolic ratescan be expected affecting resource uptake andorganismal growth to ecosystem-level produc-tion and respirationFinally an important body of food webndashrelated
research that provides strong inferential evidence
for the nexus comes from studies on animal-mediated nutrient cycling and ecological stoi-chiometry Theory suggests detrital and algalfood websmay be linked by cascading effects onnutrient cycling (99) and empirical work showsthat organismal excretion andmigration (whichrelocates C) may be important to this link (100)(Fig 5) A recent study (101) showed that ex-cretion of highly labile dissolved organic C ni-trogen (N) and phosphorus (P) by aquatic insectsmay be sufficient to support a substantial fractionof microbial energy and nutrient needs Giventhat C availability can be too low tomeetmicrobialenergy needs [eg (102 103)] and that microbialactivity enhances detrital quality to consumers(104) a C subsidy from excretory processes couldbe important at least during low-flow periodsAdditionally microbial use of excreted dissolvedorganic nitrogen (DON) and DOP may releasealgae from potential competition with microbesfor inorganic N and P if this is the case GPPshould increase Ecological stoichiometry mayalso be important for understanding constraintson organismal growth rates as rapid growthdisproportionally increases demand of P [as it isused in ribosomal RNA (105)] This could in turnlimit the ability of organisms to grow fast andexploit temporary habitats Interestingly elemen-tal ratios can vary interspecifically in response totemperature or predation pressure (106)mdashfactorsthat increase with low flows and habitat fragmen-tation Overall studies that link biota with bio-geochemical processes remain rare (107) butanalyzing flow regimes as a driver of elementturnover rates may provide a way forward
Can hydroecological advances informrestoration practice
Shortfalls in the outcomes of river restorationhave prompted calls to identify and restore pro-cesses that support and sustain biological com-munities rather than focusing only on rivergeomorphology and habitat In urban streamsscientists have critiqued an overemphasis onstructural engineered approaches (7 108) thatdo little to restore the full flow regime (109) Inlarge regulated rivers identifying and restoringimportant features of the flow regime via damoperations is often possible (albeit expensive)However that does not solve other problemsassociated with dams such as habitat frag-mentation (110) altered sediment and thermalregimes (111 112) or disrupted biogeochemicalprocesses (113) Overall efforts to alter reservoirreleases or improve urban stormwater infra-structure to partially mimic a riverrsquos natural flowregime have great potential Technologies arebeing evaluated for real-time management ofurban flows and novel ways to design flow re-leases from dams are being proposed (114 115)however research still lags far behind the needfor solutions Importantly the paucity of studieson the flow-biota-ecosystem processes nexus pointsto a need for more integrative research connectinghydrologists population and community ecologistsand ecosystem ecologists in the context ofrestoration
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 6 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
Restoring flow regimes ratherthan eventsScientifically informed flow designs may existthat maintain ecosystem integrity in regulatedrivers (116 117) but these have produced mixedresults to date In the dam-altered Putah Creekin California mimicking the natural timing ofhigh and low flows enhanced native fish by im-proving their spawning habitat reducing watertemperatures maintaining flowing conditionsand reducing the abundance of non-native spe-cies (118) However other similar attempts havebeen less successful In some cases non-nativefish density also benefitted from flow patternsmimicking an unregulated regime (119) in othersnon-natives bounced back shortly after the flowrelease (120) These experiences illustrate thateven natural flow features may have inadvertentnegative effects if only part of the community orecosystem is considered Models for the NavajoDam (San Juan River New Mexico) show thatadequately timed releases could simultaneouslyenhance native and minimize non-native fishpopulations (121) thus predicted benefits couldsurpass those of mimicking a natural flow re-gime potentially persisting even during dryyears Another study using the ldquodesignerrdquo flowparadigm (114) attempted to understand howdifferent facets of flow variability control fisheryproduction in the Lower Mekong floodplains Time-
series models combining discharge and fisherycatch suggested that enhanced flood pulses andlong inter-flood intervals may increase fisheryyieldsAlthough environmental flow practices have
mostly focused on re-creating discrete flowevents (eg flood pulses) these can still serve asvaluable experiments An iconic example of alarge-scale flow release is Minute 319 an amend-ment to the United StatesndashMexico treaty allow-ing opening of floodgates in the Lower ColoradoRiver and a one-time rewetting of the parchedColorado River Delta (122) Although this pulsed-flow experiment did not promote establishmentof native woody riparian seedlings such trialscan inform future restoration practices for ex-ample by determining whether the provision ofbare moist substrate enhances seed germina-tion (123) Hypothesis-driven experimental flowreleases combined with deep knowledge of tar-get ecosystems can provide insights into resto-ration actions [eg (124)] Still an outstandingquestion is how flow regime restoration couldbe leveraged to restore other important alteredaspects of the ecosystem such as sediment andthermal regimes For example cold-water releasesby existing dams in climate-vulnerable basinscould prevent organisms that thrive at low tem-peratures from becoming replaced by general-ist warm-tolerant taxa (125)
Although the concept of environmental flowsis generally associated with impounded riversits relevance to urban streams is clear (126)However because frequent high storm flowsare viewed as the problem reducing them gen-erally becomes the only restoration goal This isoften attempted by shifting the spatial distribu-tion of velocities within channels through ma-nipulation of channel sinuosity installation offlow-diverting weirs or adding other structuresThese approaches may reduce local erosion butrarely result in levels of water quality ecosystemprocesses or biodiversity in urban streams thatcome close to those of unimpacted streams(127 128) Focusing on the channel ratherthan the watershed context at large means thatkey ecosystemprocesses can remain compromised(129) A recent study compared time series ofmetabolism in two streamsmdashonewith stormwaterponds in the watershed and the other restoredby altering channel slope banks and in-streamstructural complexity (103) The study found thatpeaks in GPP and ER in the ldquorestoredrdquo streamwere higher more prone to resets induced byflashy storms and that recovery time scales werelonger compared with the stream in a watershedwith stormwater measures (Fig 6) Ponds areone type of nature-inspired green infrastructureused to restore streamflow tomore natural condi-tions tree plantings constructed wetlands andgrass swales can in turn replace gray infrastruc-ture such as concrete tunnels that capture andstore waterDistributing green infrastructure in water-
sheds helps restore streams because it targetsthe cause of streamflow alteration includinginsufficient infiltration groundwater rechargeand more generally limited watershed storagecapacity (129) By tackling the cause rather thanthe symptoms of flow alteration these practicescan reduce ecological impacts that result fromthe cascading effects of altered flows includingpoor water quality and temperature spikes Re-search on the effectiveness of green infrastructureon restoring streams is sparse and limited bylack of data before and after implementationalthough studies are beginning to appear Forexample upland green infrastructure in theform of shallow wetland-like pools that capturestormwater and reconnect it to the groundwaterhas been shown to enhance baseflows while re-ducing peak flows and pollutant loadsmore thanrestoration structures within the mainstem ofthe receiving stream (130) Similar structuresbuilt between stormwater outfalls and perennialstreams did not modulate the timing magnitudeor duration of urban storm responses nor didthey improve water quality or biodiversity (131)illustrating the need for researchers to understandhow exact design and watershed context influ-ence performance
Spatiotemporal variability andrestoration designs
Research showing how local and watershed-scale environmental drivers vary temporally andalong river networks in ways that shape biota
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 7 of 13
Flow pulses (storms) flashy flowsDroughts intermittencyLowstable flowsAltered timing seasonality
DOC N P
DetritusMicrobial
stoichiometric constraints
Excretion exudates
Detrital pathway
Algal pathway
Algal stoichiometric constraints
Organismal dispersal source-sink dynamics
Algae
Community replacement
ndash ndash +
ndash ndash + ndash
+
+ndash +
Invasions
Turnover
Consumer stoichiometric constraints
Fig 5 Flow variation affects biota and ecosystem processes and food webs integrate bothHydrologic connectivity along river networks is a first landscape-level filter constraining dispersal oforganisms in river networks Extreme flow variation (floods and droughts) are a source of disturbanceto all organisms (gray box) but particularly to the higher trophic levels given their higher metabolicdemands In turn flow variation controls inputs of nutrients and terrestrial leaf litter to the stream whichinfluences microbial activity and the relative contributions of the detrital (brown) and algal (green)pathways in sustaining aquatic consumers Not just quantity but also food quality (eg CN and CPratios) influence dietary choices of consumers and thus trophic transfer efficiency Finally alteration offlow regimes may influence all these compartments and may also open niches for new players Ifbiological invasions occur at the top of the food web these have the potential to divert energy away fromnative predators and to alter food-web structure through top-down controls
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
and ecosystem processes can help prioritizeselection of sites for restoration For exampleusing the concept of river network saturation(55) to identify river sections where in-streamremoval of nutrients cannot keep up with highsource inputs of nutrients suggests that restorationactions targeting those sections may dispropor-tionately improve water quality Identification ofsubcatchments that disproportionately influencestream chemistry may also be informed by com-bining methods from landscape ecology andcatchment hydrology focusing on the spatialscale of variance in water chemistry (132) Head-waters are often prioritized for restoration be-
cause poor water quality in them can havedownstream impacts Research showing thatlocal environmental constraints on river com-munities are important in headwaters as well asin the larger channels but dispersal constraintsare much less important in the former (133 134)implies that headwaters should be prioritizedHowever research quantifying time-varying con-tributions of local communities to river-wide(network) biodiversity can also inform prioritiza-tion by identifying sites that host a dispropor-tionate share of species or that host uniquespeciesmdashfor example because they shelter sensi-tive taxa during critical flow bottlenecks This idea
encapsulates the concept of keystone habitatsor local sites that strongly influence metacom-munity dynamics (135) This has been appliedto aquatic invertebrates across intermittent rivernetworks by tracking time-varying site-specificcontributions to community dissimilarity (or betadiversity) (136)Other approaches to prioritize restoration ac-
tions may be based on contributions to system-wide stability rather than numbers of individualsor species For example the restoration of fall-run Chinook salmon portfolios may be assessedvia the contributions that different periods andlocationsmake tometapopulation-wide stabilityowing to spatiotemporal variation in habitatconditions (137) Additionally interventionscould promote conditions that are not impor-tant now but could be important in the future[ie proto-refugia (138)] For example resto-ration of groundwater-influenced habitats couldmitigate future stress to drought-sensitive taxaunder scenarios of increased intermittencyPatches of undisturbed land that protect flowregimes in streams within a larger catchmentthat is otherwise urbanized can be sources ofcolonists to restored sites Priorities that focuson present degradation of river sectionsmay failto address site contribution to impairment orrecovery at broader scales or under differentscenarios These insights indicate a great poten-tial for new methods to prioritize restoration ofboth urban and regulated streamsWe know little about how altered flow regimes
propagate through entire river networks butwork is starting to assess the cumulative effectsof dams on flow regimes as well as dam-levelcontributions to alteration Although these ap-proaches have largely focused on dam cascades[eg (139)] if expanded they could help identifyfavorable sections in river networks to developenvironmental flow operations Here exper-imentation could again help with testing ofnew strategies For instance large-scale flowexperiments could be designed that coordinatereleases among dams or between dams and free-flowing tributaries to enhance the restorativeeffects of natural pulses (140)
Hydrologic connectivityinfluences restoration
Recovering critical flow paths and their connec-tivity to rivers is important but it remains asubstantial challenge for flow-degraded riversand streams Whereas regulated rivers tend tohave reduced lateral and longitudinal hydro-logic connectivity urban streams have unnaturallevels of connectivity because pipes and over-land runoff route water quickly In both cases fun-damental hydroecological insights into streambedstructure landscape storage capacity and ground-water connectivity can help to envision novel re-storation approaches (130 141 142) More generallyhydrologic connectivity is becoming a frameworkto identify alternative ways to restore streams viastormwater management (143) For example arecent study suggested that reducing road densityand hydrologic connectivity between roads
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 8 of 13
ER
Disc
harg
e (f
t3 s)
GPPG
PP o
r ER
(g O
2 m-2
day
-1)
Discharge
Watershed BMPs
In-stream restoration
0
ndash4
4
0
ndash4
4
4
2
6
4
2
6
Jan Sept July May Mar Nov Jan
A
B C
Fig 6 Green infrastructure as a means to restore stream ecosystems (A) Discharge andecosystem respiration (ER more-negative values mean higher respiration) and gross primaryproduction (GPP) for two urban streams in which restoration efforts involved (top) modifications inthe stream channel and (bottom) traditional stormwater best management practices (BMPs)[Reproduced with permission from (103)] (B and C) Example of a comprehensive greeninfrastructure approach to ecologically restore an urban headwater stream that also supports thelifestyle of resident urban communities (B) Stream at the onset of restoration and (C) afterrestoration [Reproduced with permission from Greenworks Portland OR]
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 9 of 13
Table 1 Research challenges and opportunities Despite the progress that has been made in understanding how flow regimes affect biota and ecosystem
processes major challenges persist that prevent a complete understanding of the flow-biota-ecosystem processes nexus Making progress on these
challenges requires a mix of fundamental and applied research
Topic Research challenge Opportunity
Connecting
organismal to
ecosystem-level
processes
Research on the effects of flow on organisms
and ecosystem processes has followed parallel
tracksmdashhindering our mechanistic
understanding of the flow-biota-ecosystem
processes nexus
Generate theory that connects concepts and frameworks
that are well accepted in river community and ecosystem
ecology For example the notion of environmental filters
that connect regional to local composition via a combination
of dispersal and environmental conditions (community
ecology) with analogous theory on solute transport versus
reactivity (ecosystem ecology)
Our current understanding of the links between
phenology of biota and ecosystem processes
is precursory
Use high-frequency sensors to characterize metabolic
regimes at sites with ongoing organismal monitoring
Leverage citizen-science biodiversity data to detect
spatiotemporal shifts in animal phenology
(eg insect emergence fish spawning)
Use remote sensing tools to better understand
links between hydrology and aquatic and riparian
plant phenology
The link between stoichiometry and flow
variation remains largely unexplored
Repurpose data that is already being collected to test
flow-stoichiometry relationships eg studies using C and
N stable isotopes to describe CN ratios across flow
regime types Perform field experiments to quantify
indirect effects of invertebrate and fish consumption
on ecosystem metabolism
Ecological
consequences of
hydrologic
connectivity
The importance of anaerobic metabolism in
carbon processing as flows cease and parts of
streambeds dry is a major frontier research area
Create networks of sites across different geologies
and flow regimes that collect time-series data on
methane nitrous oxide and carbon dioxide fluxes
from intermittent streams
We ignore how biota in the wet phase is
influenced by processes and patterns
from the dry phase
Research the functions of dry riverbeds Use camera traps and
drone technology to investigate the role of dry riverbeds
as a dispersal corridor metabarcoding to quantify
ldquodark biodiversityrdquo in the hyporheic
Wetlands and their links to stream networks are poorly
delineated globally Even in the United States and
Europe current wetland inventories do not capture
these links yet understanding how their connectivity
to river systems influences network-scale ecosystem
processes is needed
Analyze time series from high-frequency sensors distributed in
river networks varying in wetland connectivity Further
development and application of time-series methods
used by neuroscientists
We need to identify the relationship between
varying flows and solute dynamics
associated with changes in catchment-scale
hydrologic exchanges
Design collaborative work among catchment hydrologists
and stream ecologists combining methods to estimate
transit times residence times and identify sources
across heterogeneous landscapes Link these properties to
land management and water quality aspects
Scaling and
transferring flow-
ecology relationships
across space and time
We ignore how biodiversity responds to
propagation of flow regime alteration
across river networks
Leverage extensive hydrometric networks (eg United States
Geological Survey National Water Information System)
to perform spatially replicated time-series analyses
on flow and use these to generate expectations
on how river network position should mediate
biodiversity responses to flow alteration
Flow-ecology relationships do not account for
nonstationarity and abrupt change
Use process-based models that account for mechanisms
and associated uncertainties When using time-series
data and models select methods that are robust
to nonstationarity Linear flow-ecology relationships
should not be assumed
Current flow-ecology relationships assume that
responses of species do not change over time
Consider flow regime alteration as a process with ecological and
evolutionary consequences Include intraspecific variation
in traits and potential for rapid evolution in models
continued on next page
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
and streams may improve water quality morethan other restoration options (70) Additionallyresearch on the age and lability of DOC enteringstreams has shown how precipitation or land dis-turbance can change watershed flow paths (53)and this could inspire new restoration approachesfocused on land use and soil properties Manystudies on hydrologic connectivity and networkstructure suggest a need for restoration to focuson landscape heterogeneity especially on thepresence ofwetlands lakes and ponds (144) Suchfeaturesare frequently lost inurbanizedcatchmentsand outcomes could be improved by restoring orincreasing connectivitywith these ecosystems (145)Hydrologic connectivity associated with ground-water inputs or abstractions land-use manage-ment and return flows fromwastewater treatmentplants also need to be considered in these initia-tives (146)Restoring hydrologic connectivity is also key
to achieving biodiversity outcomesmdashparticularlyin dammed systems Longitudinal connectivityfor fish inhabiting dam-altered rivers can beenhanced by the construction of fishways andsophisticated passage facilities However moni-toring data on the effectiveness of such inter-
ventions are still scarce and available evidencesuggests that passage effectivity largely dependson technology used and species-specific fish be-havior (147 148) Although trap-and-transportprograms for migratory fishes can help augmentthreatened populations (149) these practicesoften increase straying and thus the potentialfor genetic homogenization as seen from ju-venile salmon collected at dams and transporteddownstream to increase out-migration success(150) Overall a combination of engineering andmanagement solutions may be best to restorefunctional connectivity in dam-altered riversThe link between hydrologic connectivity andecological restoration is also central to con-siderations on where to build remove and re-operate hydrologic infrastructure Hundredsof dams have been intentionally removed sincethe 1970s and removal of large dams such asthe Elwha River Dam (Washington USA) hasshown that restoring longitudinal connectivitycan have immediate geomorphic effects with com-plex temporal dynamics (151) Infrastructure canalso be designed or reoperated to manage forrestoring lateral connectivity and enhanced flood-plain processes For example the Yolo Bypass in
the Sacramento River California has becomea prime rearing and migration site for chinooksalmon showing that flood control and habitatfor fish and wildlife do not need to be at odds(152) All these examples show the importanceof integrating hydrologic connectivity in ecolog-ical restoration whether the goal is to minimizethe ecological effects of fragmentation restorefree-flowing watercourses or operate green infra-structure to enhance infiltration floodplain con-nectivity or other ecological processes
Research challenges and opportunities
Two decades of concerted efforts to research theeffects of flow regime alteration have tremen-dously advanced our understanding of the waysin which streamflow influences biota and eco-system processes However important challengesremain These aremainly related to the scarcity ofscience connecting organismal to ecosystem-levelprocesses through for example microbial linksthe difficulty of studying watershed-level phys-ical and biogeochemical heterogeneity and itsinfluence on riverine processes and biota thecomplexity of parsing out the direct effects offlow from flow-mediated effects and co-occurring
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 10 of 13
Topic Research challenge Opportunity
Cascading
implications of
hydrologic
alteration
We have a poor understanding of the
interactions among physical drivers
Build on large-scale flow experiments to test hypotheses on the
combined effects of altered flow sediment and thermal
regimes Experiment with multilevel intakes in reservoirs
Promote collection of non-flow variables (eg temperature
water quality parameters) at hydrometric stations
At low flows biota are being exposed to high
concentrations of new classes of pollutants
whose impacts are poorly known especially if
biotic impacts interact with impacts on
ecosystem processes
Design realistic experimental studies to advance knowledge
on the ecological effects of multiple stressors
Collect long-term data at sites varying in contaminant
inputs and hydrologic regimes Link microbial to
organismal and ecosystem-level responses
Using hydroecological
science to inform
river ecosystem
restoration
Environmental flow science largely ignores
human dimensions
Use methods that allow identifying causal networks
in complex socioenvironmental systems Include
human needs and thresholds in research on
flow-ecology relationships
Environmental flows are generally prescribed
at the scale of single infrastructures restoration
designs and assessment are generally at reach scales
Promote coordinated large-scale flow experiments
and restoration designs at the river basin scale
Extend assessment of green infrastructure
effectiveness beyond individual structures to
infrastructure networks and link both
to river outcomes
Environmental flows in impounded rivers
generally target the conservation of some
particular species rather than outcomes at the
food-web or ecosystem level
Advance fundamental research on microbialndash
ecosystem process linkages Expand the notion
of designer flows to food-web structure and
ecosystem services when possible
Environmental flow science has largely
ignored surface waterndashgroundwater
interactions
Expand science to understand the link between
restoration actions groundwater recharge and
inputs to streams Provide science relevant to
the implementation of new legislation on
groundwater management (eg Sustainable
Groundwater Management Act in California)
Uncertainty regarding future
hydroclimate and
river flows is growing
Refine regional climate models Promote research on
the interaction between land-use change and
evapotranspiration Incorporate groundwater
influence in macroscale hydrology models
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
stressors (particularly in urban streams) and thechallenge of predicting outcomeswhen both thephysical and the biological environments arechangingAchieving a predictive understanding of hy-
droecological relationships is essential to design-ing successful self-sustaining restoration actionsin river ecosystems However translating basichydroecological insights into restoration practicerequires understanding the complexities of work-ing within a human-dominated water cycle Al-though it is rarely possible to restore all facetsof a natural flow regime ecologically informeddesigns may help mimic key aspects of the nat-ural flow regime (eg periodic pulse flows) orcreate regimes that are not natural but maximizeoutcomes given socioeconomic constraints InTable 1 we identify persisting and emergingchallenges in hydroecological research We paireach challenge with an opportunity (related toresearch design technology or implementation)We contend that these challenges should beprioritized to advance process-based river eco-system restoration
REFERENCES AND NOTES
1 N L Poff et al The natural flow regime Bioscience 47769ndash784 (1997) doi 1023071313099
2 D A Lytle N L Poff Adaptation to natural flow regimesTrends Ecol Evol 19 94ndash100 (2004) doi 101016jtree200310002 pmid 16701235
3 S Sabater A Elosegi R Ludwig in Multiple Stressors in RiverEcosystems (Elsevier 2019) pp 1ndash22
4 United Nations New UN Decade on Ecosystem Restorationoffers unparalleled opportunity for job creation food securityand addressing climate change (2019) httpswwwunenvironmentorgnews-and-storiespress-releasenew-un-decade-ecosystem-restoration-offers-unparalleled-opportunity
5 A W Lorenz P Haase K Januschke A SundermannD Hering Revisiting restored river reaches - Assessingchange of aquatic and riparian communities afterfive years Sci Total Environ 613-614 1185ndash1195(2018) doi 101016jscitotenv201709188pmid 28954379
6 M A Palmer K L Hondula B J Koch Ecological Restorationof Streams and Rivers Shifting Strategies and ShiftingGoals (2014) vol 45
7 T J Beechie et al Process-based principles for restoringriver ecosystems Bioscience 60 209ndash222 (2010)doi 101525bio20106037
8 S E Bunn A H Arthington Basic principles and ecologicalconsequences of altered flow regimes for aquaticbiodiversity Environ Manage 30 492ndash507 (2002)doi 101007s00267-002-2737-0 pmid 12481916
9 K D Fausch Y Taniguchi S Nakano G D GrossmanC R Townsend Flood disturbance regimes influence rainbowtrout invasion success among five holarctic regions EcolAppl 11 1438ndash1455 (2001) doi 1018901051-0761(2001)011[1438FDRIRT]20CO2
10 M E Power M S Parker W E Dietrich Seasonalreassembly of a river food web Floods droughts andimpacts of fish Ecol Monogr 78 263ndash282 (2008)doi 10189006-09021
11 V Acuntildea A Giorgi I Muntildeoz U R S Uehlinger S SabaterFlow extremes and benthic organic matter shape themetabolism of a headwater Mediterranean streamFreshwater Biol 49 960ndash971 (2004) doi 101111j1365-2427200401239x
12 K Tockner F Malard J V Ward An extension of the floodpulse concept Hydrol Processes 14 2861ndash2883 (2000)doi 1010021099-1085(20001112)141617lt2861AID-HYP124gt30CO2-F
13 J S Perkin et al Fragmentation and dewatering transformGreat Plains stream fish communities Ecol Monogr 8573ndash92 (2015) doi 10189014-01211
14 J L Sabo D M Post Quantifying periodic stochastic andcatastrophic environmental variation Ecol Monogr 7819ndash40 (2008) doi 10189006-13401
15 F Shuai et al Silver carp larva abundance in response toriver flow rate revealed by cross-wavelet modellingEcol Modell 383 98ndash105 (2018) doi 101016jecolmodel201805020
16 J D Tonkin M T Bogan N Bonada B Rios-ToumaD A Lytle Seasonality and predictability shape temporalspecies diversity Ecology 98 1201ndash1216 (2017)doi 101002ecy1761 pmid 28144975
17 N L Poff Beyond the natural flow regime Broadening thehydro‐ecological foundation to meet environmental flowschallenges in a non‐stationary world Freshwater Biol 631011ndash1021 (2018) doi 101111fwb13038
18 A Ruhi X Dong C H McDaniel D P Batzer J L SaboDetrimental effects of a novel flow regime on the functionaltrajectory of an aquatic invertebrate metacommunity GlobChange Biol 24 3749ndash3765 (2018) doi 101111gcb14133pmid 29665147
19 M T Bogan D A Lytle Severe drought drives novelcommunity trajectories in desert stream pools FreshwaterBiol 56 2070ndash2081 (2011) doi 101111j1365-2427201102638x
20 G Woodward et al The effects of climatic fluctuations andextreme events on running water ecosystems Philos TransR Soc London Ser B 371 20150274 (2016) pmid 27114576
21 M A Palmer C M Swan K Nelson P Silver R AlvestadStreambed landscapes Evidence that stream invertebratesrespond to the type and spatial arrangement of patchesLandsc Ecol 15 563ndash576 (2000) doi 101023A1008194130695
22 D Schmera et al Does isolation influence the relative role ofenvironmental and dispersal‐related processes in streamnetworks An empirical test of the network positionhypothesis using multiple taxa Freshwater Biol 63 74ndash85(2018) doi 101111fwb12973
23 R Henriques‐Silva et al A comprehensive examination of thenetwork position hypothesis across multiple rivermetacommunities Ecography 42 284ndash294 (2019)doi 101111ecog03908
24 K L Jaeger J D Olden N A Pelland Climate change poisedto threaten hydrologic connectivity and endemic fishes indryland streams Proc Natl Acad Sci USA 11113894ndash13899 (2014) doi 101073pnas1320890111pmid 25136090
25 S Garbin E A Celegon P Fanton G Botter Hydrologicalcontrols on river network connectivity R Soc Open Sci 6181428 (2019) doi 101098rsos181428 pmid 30891270
26 F Cattaneacuteo B Hugueny N Lamouroux Synchrony in browntrout Salmo trutta population dynamics A lsquoMoran effectrsquo onearly‐life stages Oikos 100 43ndash54 (2003) doi 101034j1600-0706200311912x
27 A Terui et al Metapopulation stability in branching rivernetworks Proc Natl Acad Sci USA 115 E5963ndashE5969(2018) doi 101073pnas1800060115 pmid 29895695
28 P A P Moran The statistical analysis of the Canadian Lynxcycle Aust J Zool 1 291ndash298 (1953) doi 101071ZO9530291
29 G D Grossman P B Moyle J O Whitaker Jr Stochasticityin structural and functional characteristics of an Indianastream fish assemblage A test of community theoryAm Nat 120 423ndash454 (1982) doi 101086284004
30 D A Lytle D M Merritt J D Tonkin J D OldenL V Reynolds Linking river flow regimes to riparian plantguilds A community-wide modeling approach Ecol Appl 271338ndash1350 (2017) doi 101002eap1528 pmid 28263426
31 A Ruhiacute E E Holmes J N Rinne J L Sabo Anomalousdroughts not invasion decrease persistence of native fishesin a desert river Glob Change Biol 21 1482ndash1496 (2015)doi 101111gcb12780 pmid 25430731
32 J D Tonkin D M Merritt J D Olden L V ReynoldsD A Lytle Flow regime alteration degrades ecologicalnetworks in riparian ecosystems Nat Ecol Evol 2 86ndash93(2018) doi 101038s41559-017-0379-0 pmid 29180707
33 M C Mims J D Olden Fish assemblages respond toaltered flow regimes via ecological filtering of lifehistory strategies Freshwater Biol 58 50ndash62 (2013)doi 101111fwb12037
34 P T J Johnson J D Olden M J Vander Zanden Daminvaders Impoundments facilitate biological invasions intofreshwaters Front Ecol Environ 6 357ndash363 (2008)doi 101890070156
35 N Bonada S Doledec B Statzner Taxonomic and biologicaltrait differences of stream macroinvertebrate communitiesbetween mediterranean and temperate regions Implicationsfor future climatic scenarios Glob Change Biol 131658ndash1671 (2007) doi 101111j1365-2486200701375x
36 M I Pyne N L Poff Vulnerability of stream communitycomposition and function to projected thermal warming andhydrologic change across ecoregions in the western UnitedStates Glob Change Biol 23 77ndash93 (2017) doi 101111gcb13437 pmid 27429092
37 D B Herbst S D Cooper R B Medhurst S W WisemanC T Hunsaker Drought ecohydrology alters the structureand function of benthic invertebrate communities inmountain streams Freshwater Biol 64 886ndash902 (2019)doi 101111fwb13270
38 M T Bogan E T Chester T Datry A L MurphyB J Robson A Ruhi R Stubbington J E Whitney inResistance Resilience and Community Recovery in IntermittentRivers and Ephemeral Streams T Datry N Bonada A BoultonEds (Academic Press 2017) pp 349ndash376
39 S Houmlckendorff et al Characterizing fish responses to a riverrestoration over 21 years based on speciesrsquo traits ConservBiol 31 1098ndash1108 (2017) doi 101111cobi12908pmid 28218807
40 D Bruno et al Structural and functional responses ofinvertebrate communities to climate change andflow regulation in alpine catchments Glob Change Biol25 1612ndash1628 (2019) doi 101111gcb14581pmid 30698905
41 T W H Aspin et al Extreme drought pushes streaminvertebrate communities over functional thresholds GlobChange Biol 25 230ndash244 (2019) doi 101111gcb14495pmid 30346098
42 S E Tank J B Fellman E Hood E S Kritzberg Beyondrespiration Controls on lateral carbon fluxes across theterrestrial-aquatic interface Limnol Oceanogr Lett 3 76ndash88(2018) doi 101002lol210065
43 A P Appling et al The metabolic regimes of 356 rivers inthe United States Sci Data 5 180292 (2018) doi 101038sdata2018292 pmid 30532078
44 P Savoy et al Metabolic rhythms in flowing watersAn approach for classifying river productivity regimesLimnol Oceanogr (2019) doi 101002lno11154
45 E S Bernhardt et al The metabolic regimes of flowingwaters Limnol Oceanogr 63 S99ndashS118 (2018)doi 101002lno10726
46 M Colls X Timoner S Sabater V Acuntildea Effects of durationand frequency of the non-flow period on stream metabolismEcosystems 101007s10021-019-00345-1 (2019)doi 101007s10021-019-00345-1
47 A J Ulseth E Bertuzzo G A Singer J Schelker T J BattinClimate-induced changes in spring snowmelt impactecosystem metabolism and carbon fluxes in an Alpine streamnetwork Ecosystems 21 373ndash390 (2018) doi 101007s10021-017-0155-7
48 B O L Demars Hydrological pulses and burning of dissolvedorganic carbon by stream respiration Limnol Oceanogr 64406ndash421 (2019) doi 101002lno11048
49 R T Hensley L Kirk M Spangler M N Gooseff M J CohenFlow extremes as spatiotemporal control points on riversolute fluxes and metabolism J Geophys Res Biogeosci124 537ndash555 (2019) doi 1010292018JG004738
50 J D Hosen et al Enhancement of primary production duringdrought in a temperate watershed is greater in larger riversthan headwater streams Limnol Oceanogr 64 1458ndash1472(2019) doi 101002lno11127
51 L Goacutemez-Gener et al When water vanishes Magnitude andregulation of carbon dioxide emissions from dry temporarystreams Ecosystems 19 710ndash723 (2016) doi 101007s10021-016-9963-4
52 E H Stanley et al The ecology of methane in streams andrivers Patterns controls and global significance EcolMonogr 86 146ndash171 (2016) doi 10189015-1027
53 R T Barnes D E Butman H F Wilson P A RaymondRiverine export of aged carbon driven by flow path depth andresidence time Environ Sci Technol 52 1028ndash1035 (2018)doi 101021acsest7b04717 pmid 29313674
54 G Pinay et al Riparian corridors A new conceptualframework for assessing nitrogen buffering across biomesFront Environ Sci 6 47 (2018) doi 103389fenvs201800047
55 W M Wollheim et al River network saturation conceptFactors influencing the balance of biogeochemical supply and
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 11 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
demand of river networks Biogeochemistry 141 503ndash521(2018) doi 101007s10533-018-0488-0
56 P A Raymond J E Saiers W V Sobczak Hydrological andbiogeochemical controls on watershed dissolved organicmatter transport Pulse-shunt concept Ecology 97 5ndash16(2016) doi 10189014-16841 pmid 27008769
57 C Tunaley D Tetzlaff J Lessels C Soulsby Linking high-frequency DOC dynamics to the age of connected watersources Water Resour Res 52 5232ndash5247 (2016)doi 1010022015WR018419
58 S Bernal et al Supply demand and in-stream retention ofdissolved organic carbon and nitrate during storms inMediterranean forested headwater streams Front EnvironSci 7 60 (2019) doi 103389fenvs201900060
59 J Harvey et al How hydrologic connectivity regulates waterquality in river corridors J Am Water Resour Assoc 55369ndash381 (2019) doi 1011111752-168812691
60 J C Moore et al Detritus trophic dynamics and biodiversityEcol Lett 7 584ndash600 (2004) doi 101111j1461-0248200400606x
61 A D Rosemond et al Freshwater ecology Experimentalnutrient additions accelerate terrestrial carbon loss fromstream ecosystems Science 347 1142ndash1145 (2015)doi 101126scienceaaa1958 pmid 25745171
62 A C Y Yeung J L Musetta-Lambert D P KreutzweiserP K Sibley J S Richardson Relations of interannualdifferences in stream litter breakdown with dischargeBioassessment implications Ecosphere 9 e02423 (2018)doi 101002ecs22423
63 R M Northington J R Webster Experimental reductions instream flow alter litter processing and consumer subsidies inheadwater streams Freshwater Biol 62 737ndash750 (2017)doi 101111fwb12898
64 E Mas-Martiacute I Sanpera-Calbet I Muntildeoz Bottom-up effectsof streambed drying on consumer performance throughchanges in resource quality Aquat Sci 79 719ndash731 (2017)doi 101007s00027-017-0531-6
65 B D Richter G A Thomas Restoring environmental flows bymodifying dam operations Ecol Soc 12 12 (2007)doi 105751ES-02014-120112
66 G Zolezzi A Siviglia M Toffolon B MaioliniThermopeaking in Alpine streams Event characterizationand time scales Ecohydrology 4 564ndash576 (2011)doi 101002eco132
67 T Meiszligner B Sures C K Feld Multiple stressors and therole of hydrology on benthic invertebrates in mountainousstreams Sci Total Environ 663 841ndash851 (2019)doi 101016jscitotenv201901288 pmid 30738264
68 I R Waite et al Effects of urban multi-stressors on threestream biotic assemblages Sci Total Environ 6601472ndash1485 (2019) doi 101016jscitotenv201812240pmid 30743940
69 K C Nelson M A Palmer Stream temperature surges underurbanization and climate change Data models andresponses J Am Water Resour Assoc 43 440ndash452 (2007)doi 101111j1752-1688200700034x
70 J R Blaszczak J M Delesantro Y Zhong D L UrbanE S Bernhardt Watershed urban development controls onurban streamwater chemistry variability Biogeochemistry144 61ndash84 (2019) doi 101007s10533-019-00572-7
71 L Ponsatiacute et al Biofilm responses to flow regulation by damsin Mediterranean rivers River Res Appl 31 1003ndash1016(2015) doi 101002rra2807
72 J M Alberts J J Beaulieu I Buffam Watershed land useand seasonal variation constrain the influence of ripariancanopy cover on stream ecosystem metabolism Ecosystems20 553ndash567 (2017) doi 101007s10021-016-0040-9pmid 30416371
73 R O Hall Jr et al Turbidity light temperature andhydropeaking control primary productivity in the ColoradoRiver Grand Canyon Limnol Oceanogr 60 512ndash526 (2015)doi 101002lno10031
74 J R Blaszczak J M Delesantro D L Urban M W DoyleE S Bernhardt Scoured or suffocated Urban streamecosystems oscillate between hydrologic and dissolvedoxygen extremes Limnol Oceanogr 64 877ndash894 (2019)doi 101002lno11081
75 J-R Mor et al Dam regulation and riverine food-webstructure in a Mediterranean river Sci Total Environ 625301ndash310 (2018) doi 101016jscitotenv201712296pmid 29289778
76 H A Wellard Kelly et al Macroinvertebrate diets reflecttributary inputs and turbidity-driven changes in food
availability in the Colorado River downstream of GlenCanyon Dam Freshw Sci 32 397ndash410 (2013)doi 10189912-0881
77 S J Imberger R M Thompson M R Grace Urbancatchment hydrology overwhelms reach scale effects ofriparian vegetation on organic matter dynamics FreshwaterBiol 56 1370ndash1389 (2011) doi 101111j1365-2427201102575x
78 R T Martins A S Melo J F Gonccedilalves Jr N HamadaLeaf-litter breakdown in urban streams of Central AmazoniaDirect and indirect effects of physical chemical andbiological factors Freshw Sci 34 716ndash726 (2015)doi 101086681086
79 C M Yule J Y Gan T Jinggut K V Lee Urbanizationaffects food webs and leaf-litter decomposition in a tropicalstream in Malaysia Freshw Sci 34 702ndash715 (2015)doi 101086681252
80 T A Newcomer et al Influence of natural and novel organiccarbon sources on denitrification in forest degraded urbanand restored streams Ecol Monogr 82 449ndash466 (2012)doi 10189012-04581
81 A Chaudhary I Kauser A Ray R Poretsky Taxon-drivenfunctional shifts associated with storm flow in an urbanstream microbial community MSphere 3 e00194-18 (2018)doi 101128mSphere00194-18 pmid 29976643
82 T A Kennedy et al Flow management for hydropowerextirpates aquatic insects undermining river food websBioscience 66 561ndash575 (2016) doi 101093bioscibiw059
83 A Glazaczow D Orwin M Bogdziewicz Increasedtemperature delays the late-season phenology of multivoltineinsect Sci Rep 6 38022 (2016) doi 101038srep38022pmid 27905493
84 S G Fisher L J Gray N B Grimm D E Busch Temporalsuccession in a desert stream ecosystem following flashflooding Ecol Monogr 52 93ndash110 (1982) doi 1023072937346
85 G J Scrimgeour M J Winterbourn Effects of floods onepilithon and benthic macroinvertebrate populations in anunstable New Zealand river Hydrobiologia 171 33ndash44 (1989)doi 101007BF00005722
86 E Bondar-Kunze S Maier D Schoumlnauer N Bahl T HeinAntagonistic and synergistic effects on a stream periphytoncommunity under the influence of pulsed flow velocityincrease and nutrient enrichment Sci Total Environ 573594ndash602 (2016) doi 101016jscitotenv201608158pmid 27585428
87 M J Cashman G L Harvey G Wharton M C Bruno Woodmitigates the effect of hydropeaking scour on periphytonbiomass and nutritional quality in semi-natural flumesimulations Aquat Sci 79 459ndash471 (2017) doi 101007s00027-016-0510-3
88 W F Cross et al Food-web dynamics in a large riverdiscontinuum Ecol Monogr 83 311ndash337 (2013)doi 10189012-17271
89 D M Warfe et al Productivity disturbance and ecosystemsize have no influence on food chain length in seasonallyconnected rivers PLOS ONE 8 e66240 (2013)doi 101371journalpone0066240
90 T D Jardine et al Consumer-resource coupling in wet-drytropical rivers J Anim Ecol 81 310ndash322 (2012)doi 101111j1365-2656201101925x pmid 22103689
91 K O Winemiller et al Pulsing hydrology determines top‐down control of basal resources in a tropical riverndashfloodplainecosystem Ecol Monogr 84 621ndash635 (2014)doi 10189013-18221
92 D M Post The long and short of food-chain length TrendsEcol Evol 17 269ndash277 (2002) doi 101016S0169-5347(02)02455-2
93 D M Post G Takimoto Proximate structural mechanismsfor variation in food‐chain length Oikos 116 775ndash782 (2007)doi 101111j0030-1299200715552x
94 P A McHugh A R McIntosh P G Jellyman Dual influencesof ecosystem size and disturbance on food chain length instreams Ecol Lett 13 881ndash890 (2010) doi 101111j1461-0248201001484x pmid 20482579
95 A Ruhiacute et al Flow regulation increases food-chain lengththrough omnivory mechanisms in a Mediterranean rivernetwork Freshwater Biol 61 1536ndash1549 (2016) doi 101111fwb12794
96 J L Sabo J C Finlay T Kennedy D M Post The role ofdischarge variation in scaling of drainage area and food chainlength in rivers Science 330 965ndash967 (2010) doi 101126science1196005 pmid 20947729
97 A R McIntosh et al Capacity to support predators scaleswith habitat size Sci Adv 4 eaap7523 (2018) doi 101126sciadvaap7523 pmid 29978038
98 J H Brown J F Gillooly A P Allen V M Savage G B WestToward a metabolic theory of ecology Ecology 851771ndash1789 (2004) doi 10189003-9000
99 K Zou E Theacutebault G Lacroix S Barot Interactions betweenthe green and brown food web determine ecosystemfunctioning Funct Ecol 30 1454ndash1465 (2016) doi 1011111365-243512626
100 M J Vanni P B McIntyre Predicting nutrient excretion ofaquatic animals with metabolic ecology and ecologicalstoichiometry A global synthesis Ecology 97 3460ndash3471(2016) doi 101002ecy1582 pmid 27912023
101 T B Parr K A Capps S P Inamdar K A Metcalf Animal-mediated organic matter transformation Aquatic insects as asource of microbially bioavailable organic nutrients andenergy Funct Ecol 33 524ndash535 (2019) doi 1011111365-243513242
102 C P Arango et al Urban infrastructure influences dissolvedorganic matter quality and bacterial metabolism in an urbanstream network Freshwater Biol 62 1917ndash1928 (2017)
103 L G Larsen J W Harvey Disrupted carbon cycling inrestored and unrestored urban streams Critical timescalesand controls Limnol Oceanogr 62 S160ndashS182 (2017)doi 101002lno10613
104 H M Halvorson G White J T Scott M A Evans-WhiteDietary and taxonomic controls on incorporation of microbialcarbon and phosphorus by detritivorous caddisfliesOecologia 180 567ndash579 (2016) doi 101007s00442-015-3464-6 pmid 26497125
105 J J Elser et al Biological stoichiometry from genes toecosystems Ecol Lett 3 540ndash550 (2000) doi 101046j1461-0248200000185x
106 E K Moody et al Consumption explains intraspecificvariation in nutrient recycling stoichiometry in a desert fishEcology 99 1552ndash1561 (2018) doi 101002ecy2372pmid 29882955
107 N Welti et al Bridging food webs ecosystem metabolismand biogeochemistry using ecological stoichiometry theoryFront Microbiol 8 1298 (2017) doi 103389fmicb201701298 pmid 28747904
108 M A Palmer S Filoso R M Fanelli From ecosystems toecosystem services Stream restoration as ecologicalengineering Ecol Eng 65 62ndash70 (2014) doi 101016jecoleng201307059
109 M J Burns T D Fletcher C J Walsh A R LadsonB E Hatt Hydrologic shortcomings of conventional urbanstormwater management and opportunities for reformLandsc Urban Plan 105 230ndash240 (2012) doi 101016jlandurbplan201112012
110 C R Liermann C Nilsson J Robertson R Y NgImplications of dam obstruction for global freshwater fishdiversity Bioscience 62 539ndash548 (2012) doi 101525bio20126265
111 J D Olden R J Naiman Incorporating thermal regimes intoenvironmental flows assessments Modifying dam operationsto restore freshwater ecosystem integrity Freshwater Biol55 86ndash107 (2010) doi 101111j1365-2427200902179x
112 D B Osmundson R J Ryel V L Lamarra J Pitlick Flowndashsedimentndashbiota relations Implications for river regulationeffects on native fish abundance Ecol Appl 12 1719ndash1739(2002)
113 D von Schiller et al Regulation causes nitrogen cyclingdiscontinuities in Mediterranean rivers Sci Total Environ540 168ndash177 (2016) doi 101016jscitotenv201507017pmid 26233271
114 J L Sabo et al Designing river flows to improve foodsecurity futures in the Lower Mekong Basin Science 358eaao1053 (2017) doi 101126scienceaao1053pmid 29217541
115 W Xu et al Improving the multi-objective performance ofrainwater harvesting systems using real-time controltechnology Water 10 147 (2018) doi 103390w10020147
116 M Acreman et al Environmental flows for naturalhybrid and novel riverine ecosystems in a changingworld Front Ecol Environ 12 466ndash473 (2014)doi 101890130134
117 N L Poff et al Sustainable water management under futureuncertainty with eco-engineering decision scaling Nat ClimChang 6 25ndash34 (2016) doi 101038nclimate2765
118 J D Kiernan P B Moyle P K Crain Restoring native fishassemblages to a regulated California stream using the
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 12 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
natural flow regime concept Ecol Appl 22 1472ndash1482(2012) doi 10189011-04801 pmid 22908707
119 D L Propst K B Gido Responses of native andnonnative fishes to natural flow regime mimicry in theSan Juan River Trans Am Fish Soc 133 922ndash931 (2004)doi 101577T03-0571
120 T K Pool J D Olden Assessing long‐term fish responses andshort‐term solutions to flow regulation in a dryland river basinEcol Freshwat Fish 24 56ndash66 (2015) doi 101111eff12125
121 W Chen J D Olden Designing flows to resolve human andenvironmental water needs in a dam-regulated riverNat Commun 8 2158 (2017) doi 101038s41467-017-02226-4 pmid 29255194
122 S M Nelson et al A history of the 2014 Minute 319environmental pulse flow as documented by fieldmeasurements and satellite imagery Ecol Eng 106733ndash748 (2017) doi 101016jecoleng201610040
123 P B Shafroth et al A large-scale environmental flowexperiment for riparian restoration in the Colorado RiverDelta Ecol Eng 106 645ndash660 (2017) doi 101016jecoleng201702016
124 E P Glenn P L Nagler P B Shafroth C J JarchowEffectiveness of environmental flows for riparian restorationin arid regions A tale of four rivers Ecol Eng 106 695ndash703(2017) doi 101016jecoleng201701009
125 S Hotaling D S Finn J Joseph Giersch D W WeisrockD Jacobsen Climate change and alpine stream biologyProgress challenges and opportunities for the futureBiol Rev Camb Philos Soc 92 2024ndash2045 (2017)doi 101111brv12319 pmid 28105701
126 C J Walsh T D Fletcher M J Burns Urban stormwaterrunoff A new class of environmental flow problem PLOS ONE7 e45814 (2012) doi 101371journalpone0045814pmid 23029257
127 A Selvakumar T P OrsquoConnor S D Struck Role of streamrestoration on improving benthic macroinvertebrates and in-stream water quality in an urban watershed Case studyJ Environ Eng 136 127ndash139 (2010) doi 101061(ASCE)EE1943-78700000116
128 R W Lammers B P Bledsoe What role does streamrestoration play in nutrient management Crit Rev EnvironSci Technol 47 335ndash371 (2017) doi 1010801064338920171318618
129 H E Golden N Hoghooghi Green infrastructure and itscatchment-scale effects An emerging science WIREs Water5 e1254 (2018) doi 101002wat21254 pmid 29682288
130 M R Williams G Bhatt S Filoso G Yactayo Streamrestoration performance and its contribution to theChesapeake Bay TMDL Challenges posed by climate changein urban areas Estuaries Coasts 40 1227ndash1246 (2017)doi 101007s12237-017-0226-1
131 R M Fanelli K Prestegaard M A Palmer Freshw Sci101086706072 (2019)
132 B W Abbott et al Unexpected spatial stability of waterchemistry in headwater stream networks Ecol Lett 21296ndash308 (2018) doi 101111ele12897 pmid 29282860
133 B L Brown C M Swan Dendritic network structureconstrains metacommunity properties in riverineecosystems J Anim Ecol 79 571ndash580 (2010) doi 101111j1365-2656201001668x pmid 20180874
134 C M Swan B L Brown Metacommunity theory meetsrestoration Isolation may mediate how ecologicalcommunities respond to stream restoration Ecol Appl 272209ndash2219 (2017) doi 101002eap1602 pmid 28718193
135 N Mouquet D Gravel F Massol V Calcagno Extending theconcept of keystone species to communities and ecosystemsEcol Lett 16 1ndash8 (2013) doi 101111ele12014 pmid 23062191
136 A Ruhiacute T Datry J L Sabo Interpreting beta-diversitycomponents over time to conserve metacommunities inhighly dynamic ecosystems Conserv Biol 31 1459ndash1468(2017) doi 101111cobi12906 pmid 28188969
137 L Yamane L W Botsford D P Kilduff Tracking restorationof population diversity via the portfolio effect J Appl Ecol55 472ndash481 (2018) doi 1011111365-266412978
138 D W Braidwood M A Taggart M Smith R AndersenTranslocations conservation and climate change Use ofrestoration sites as protorefuges and protorefugia RestorEcol 26 20ndash28 (2018) doi 101111rec12642
139 Y Wang D Wang Q W Lewis J Wu F Huang A frameworkto assess the cumulative impacts of dams on hydrologicalregime A case study of the Yangtze River Hydrol Processes31 3045ndash3055 (2017) doi 101002hyp11239
140 J L Sabo et al Pulsed flows tributary inputs and food-webstructure in a highly regulated river J Appl Ecol 551884ndash1895 (2018) doi 1011111365-266413109
141 C Magliozzi G Coro R C Grabowski A I PackmanS Krause A multiscale statistical method to identifypotential areas of hyporheic exchange for river restorationplanning Environ Model Softw 111 311ndash323 (2019)doi 101016jenvsoft201809006
142 S S Schwartz B Smith Restoring hydrologic function inurban landscapes with suburban subsoiling J Hydrol 543770ndash781 (2016) doi 101016jjhydrol201610051
143 C J Walsh T D Fletcher A R Ladson Stream restoration inurban catchments through redesigning stormwater systemsLooking to the catchment to save the stream J N AmBenthol Soc 24 690ndash705 (2005) doi 10189904-0201
144 N M Schmadel et al Thresholds of lake and reservoirconnectivity in river networks control nitrogen removalNat Commun 9 2779 (2018) doi 101038s41467-018-05156-x pmid 30018449
145 C D Sayer Conservation of aquatic landscapes Pondslakes and rivers as integrated systems WIREs Water 1573ndash585 (2014) doi 101002wat21045
146 R G Luthy D L Sedlak M H Plumlee D Austin V H ReshWastewater‐effluent‐dominated streams as ecosystem‐
management tools in a drier climate Front Ecol Environ 13477ndash485 (2015) doi 101890150038
147 D W Roscoe S G Hinch Effectiveness monitoring of fishpassage facilities Historical trends geographic patterns andfuture directions Fish Fish 11 12ndash33 (2010) doi 101111j1467-2979200900333x
148 C M Bunt T Castro-Santos A Haro Performanceof fish passage structures at upstream barriers tomigration River Res Appl 28 457ndash478 (2012)doi 101002rra1565
149 D Weigel et al Evaluation of a trap-and-transport programfor a threatened population of steelhead (Oncorhynchusmykiss) Conserv Genet 20 1195ndash1199 (2019) doi 101007s10592-019-01200-5
150 M H Bond et al Combined effects of barge transportationriver environment and rearing location on straying andmigration of adult Snake River fall-run Chinook SalmonTrans Am Fish Soc 146 60ndash73 (2017) doi 1010800002848720161235614
151 A C Ritchie et al Morphodynamic evolution followingsediment release from the worldrsquos largest dam removalSci Rep 8 13279 (2018) doi 101038s41598-018-30817-8pmid 30185796
152 T Sommer et al Californiarsquos Yolo Bypass Evidence thatflood control can be compatible with fisheries wetlandswildlife and agriculture Fisheries 26 6ndash16 (2001)doi 1015771548-8446(2001)026lt0006CYBgt20CO2
153 G Grill et al Mapping the worldrsquos free-flowing riversNature 569 215ndash221 (2019) doi 101038s41586-019-1111-9pmid 31068722
154 G Gassert M Luck M Landis P Reig T Shiao ldquoAqueductglobal maps 21 Constructing decision-relevant global waterrisk indicatorsrdquo Working paper World Resources Institute(2015) wwwwriorgpublicationaqueduct-global-maps-21-indicators
155 A Ruhiacute J D Olden J L Sabo Declining streamflow inducescollapse and replacement of native fishes in the AmericanSouthwest Front Ecol Environ 14 465ndash472 (2016)doi 101002fee1424
156 T C Gouhier A Grinsted V Simko R package Biwaveletconduct univariate and bivariate wavelet analyses (2018)
157 T Rodriacuteguez-Castillo E Esteacutevez A M Gonzaacutelez-FerrerasJ Barquiacuten Estimating ecosystem metabolism toentire river networks Ecosystems (2018) doi 101007s10021-018-0311-8
ACKNOWLEDGMENTS
We greatly appreciate those who read and provided importantinsights on drafts of this paper A Armstrong K HondulaJ Hosen K Leathers G de Mendoza C Maietta I MunozJ Tonkin and two anonymous reviewers We also thank T Tu forhelp with Fig 1 and S Carlson L Comte T Datry J HosenM Kondolf N Jones D Lytle E Moody J Olden M PowerJ Sabo and M Sanchez-Montoya for valuable discussionsCompeting interests The authors have no competing interestsFunding MAP was supported by funds from the NSF (1639145)and AR was supported by funds from the University of CaliforniaBerkeley and NSF (1802714)
101126scienceaaw2087
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 13 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
restorationLinkages between flow regime biota and ecosystem processes Implications for river
Margaret Palmer and Albert Ruhi
DOI 101126scienceaaw2087 (6459) eaaw2087365Science
this issue p eaaw2087Scienceflow variability and achieve hydrological connectivity between a river and its surroundingsand ecosystem processes To be successful such efforts must go beyond accounting for flood pulses to restore natural mimic ecologically important aspects of natural flow regimes guided by insights into how variations in flow affect biotawater quality and ecological processes In a Review Palmer and Ruhi explain how restoration designs now attempt to
Human activities have altered the flow regimes of many of Earths rivers with negative impacts on biodiversityRiver restoration guided by research
ARTICLE TOOLS httpsciencesciencemagorgcontent3656459eaaw2087
REFERENCES
httpsciencesciencemagorgcontent3656459eaaw2087BIBLThis article cites 147 articles 7 of which you can access for free
PERMISSIONS httpwwwsciencemagorghelpreprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAASScienceScience 1200 New York Avenue NW Washington DC 20005 The title (print ISSN 0036-8075 online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
Science No claim to original US Government WorksCopyright copy 2019 The Authors some rights reserved exclusive licensee American Association for the Advancement of
on May 27 2020
httpsciencesciencem
agorgD
ownloaded from

inadequate to meet restoration needs Studieson ecosystem processes below dams are parti-cularly limited and much of what we knowcomes from only a few river systems Furtherpublished work focuses primarily on benthicecosystem processes immediately below damsrather than riverine planktonic realms whereproductivity in large regulated rivers can beimportant
Linking flow biota andecosystem processes
Research on flow regimes that bridges organis-mal and ecosystem ecology has traditionallyrelied on states instead of rates ie nutrientconcentrations rather than uptake or fluxesstanding stocks of algae rather than productivityor metabolism and abundance of organismsrather than secondary productivity This workgenerally shows that flow variability suppressesand stability increases algal standing stock andgrazer densities with peaks in invertebrate bio-mass following those of algae [eg (84 85)]Over the past decade research began to focuson how flow regimes alter functional aspects offoodwebs For example biofilms inMediterranean-climate rivers are thicker andmetabolicallymoreactive downstream of flow-stabilizing dams (71)When flow variability increases below hydro-power dams fluctuations can reduce biofilmdevelopment and its quality as a food resource(86) Using experimental flumes subjected tohydropeaking shifts have been found in periph-yton fatty acid content from important highlyunsaturated compounds to nonessential saturatedones (87)Indirect evidence suggests that flow-induced
changes in dominant resource type (eg algaeversus detritus) resource quantity (biomass) orresource quality (nutrient content) can propa-gate to higher trophic levels via bottom-up ef-fects For instance detritivorous invertebrateswere found to dominate species richness andinvertebrate biomass upstream but not down-stream of an irrigation dam (75) diets of om-nivore insects indicated that flow stabilizationshifted a detrital into an algal-fueled food webSimilar evidence comes from Glen Canyon Damin the Colorado River (88) Near the dam whereprimary production is high (73) food webs aresimplified and gut content analyses have sug-gested algal production supports gt50 of in-vertebrate and gt70 of fish production andthe food web can be easily perturbed by flooddisturbance Farther downriver food webs showincreased reliance on detrital resources havehigher trophic efficiency and are more complexand more resistant to disturbance (76) In con-trast recent work on seasonal tropical riversfound algal productivity and zero-flow distur-bance did not explain food-web structure Inthis case it seems that highly mobile predatorsbuffered the local effects of drought (89) Thisstudy along with earlier ones at the site (90) isof particular interest because it links temporalpatterns in predatory fish with flow-dependentavailability of resources
Only a few studies have provided evidence oftop-down controls on foodwebs being influencedby flow variation Fish in a tropical floodplainriver were found to exert top-down control onprimary producers but only under certain hydro-logic conditions (91) Similarly in aCalifornian riverwinter flood pulses were shown to control insectconsumers late in the season ultimately deter-mining whether fish exerted top-down controlon the lower trophic levels (10) These illustratethe importance of pulsed hydrology on resource-consumer linkages and show that not just standingstocks but also the dominant controls structuringfood webs may fluctuate over time in response toflow variationAn indirect line of evidence supporting the
importance of the flow-biota-ecosystem processesnexus comes from theoretically oriented researchon food-web structure Changes in productivityhabitat size and disturbance underpin the basisof the three main hypotheses of food-chainlength which state that chains should be longerin stable productive large ecosystems (92) No-tably flow regimes influence these three con-trols Because food-chain length determines keyaspects of a river ecosystem such as the risk oftrophic cascades or the pollutant accumulationin top predators understanding how it is af-fected by flow alteration is important How-ever understanding the interplay between thehypotheses of food-chain length and the flow-dependent biological mechanisms that allowfood webs to shift in structure (eg predatorinvasions local extinctions consumers chang-ing diets) is still a relatively recent endeavor(93ndash95) Evidence suggests that zero-flows short-en food chains mainly by keeping streamsfishless (94 96) whereas stabilization of variableflows can lengthen food chains by allowing om-nivores to feed on abundant high trophic levelprey (95)New studies are also focusing on interactions
between drivers of food-web structure that hadbeen previously only been examined in isolationsuch as disturbance regimes and ecosystem sizeboth influenced by flow regime Using metabolictheory predator biomass has been shown toscale with both prey resources and fluctuatingstream habitat size with reductions in preda-tor size leading to declines in predator biomasssupported per unit of prey biomass (97) Movingforward the combination of habitat andmetabolicconstraints on predators may provide a basis toconnect flow-induced disturbance changes inecosystem processes and habitat fluctuationsto riverine food-web structure The metabolictheory of ecology (98) is a unifying frameworkthat allows scaling responses of organisms towhole ecosystems andmay become particularlyuseful in situations where flow alteration affectsorganismal body size or water temperature Insuch cases strong changes in metabolic ratescan be expected affecting resource uptake andorganismal growth to ecosystem-level produc-tion and respirationFinally an important body of food webndashrelated
research that provides strong inferential evidence
for the nexus comes from studies on animal-mediated nutrient cycling and ecological stoi-chiometry Theory suggests detrital and algalfood websmay be linked by cascading effects onnutrient cycling (99) and empirical work showsthat organismal excretion andmigration (whichrelocates C) may be important to this link (100)(Fig 5) A recent study (101) showed that ex-cretion of highly labile dissolved organic C ni-trogen (N) and phosphorus (P) by aquatic insectsmay be sufficient to support a substantial fractionof microbial energy and nutrient needs Giventhat C availability can be too low tomeetmicrobialenergy needs [eg (102 103)] and that microbialactivity enhances detrital quality to consumers(104) a C subsidy from excretory processes couldbe important at least during low-flow periodsAdditionally microbial use of excreted dissolvedorganic nitrogen (DON) and DOP may releasealgae from potential competition with microbesfor inorganic N and P if this is the case GPPshould increase Ecological stoichiometry mayalso be important for understanding constraintson organismal growth rates as rapid growthdisproportionally increases demand of P [as it isused in ribosomal RNA (105)] This could in turnlimit the ability of organisms to grow fast andexploit temporary habitats Interestingly elemen-tal ratios can vary interspecifically in response totemperature or predation pressure (106)mdashfactorsthat increase with low flows and habitat fragmen-tation Overall studies that link biota with bio-geochemical processes remain rare (107) butanalyzing flow regimes as a driver of elementturnover rates may provide a way forward
Can hydroecological advances informrestoration practice
Shortfalls in the outcomes of river restorationhave prompted calls to identify and restore pro-cesses that support and sustain biological com-munities rather than focusing only on rivergeomorphology and habitat In urban streamsscientists have critiqued an overemphasis onstructural engineered approaches (7 108) thatdo little to restore the full flow regime (109) Inlarge regulated rivers identifying and restoringimportant features of the flow regime via damoperations is often possible (albeit expensive)However that does not solve other problemsassociated with dams such as habitat frag-mentation (110) altered sediment and thermalregimes (111 112) or disrupted biogeochemicalprocesses (113) Overall efforts to alter reservoirreleases or improve urban stormwater infra-structure to partially mimic a riverrsquos natural flowregime have great potential Technologies arebeing evaluated for real-time management ofurban flows and novel ways to design flow re-leases from dams are being proposed (114 115)however research still lags far behind the needfor solutions Importantly the paucity of studieson the flow-biota-ecosystem processes nexus pointsto a need for more integrative research connectinghydrologists population and community ecologistsand ecosystem ecologists in the context ofrestoration
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 6 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
Restoring flow regimes ratherthan eventsScientifically informed flow designs may existthat maintain ecosystem integrity in regulatedrivers (116 117) but these have produced mixedresults to date In the dam-altered Putah Creekin California mimicking the natural timing ofhigh and low flows enhanced native fish by im-proving their spawning habitat reducing watertemperatures maintaining flowing conditionsand reducing the abundance of non-native spe-cies (118) However other similar attempts havebeen less successful In some cases non-nativefish density also benefitted from flow patternsmimicking an unregulated regime (119) in othersnon-natives bounced back shortly after the flowrelease (120) These experiences illustrate thateven natural flow features may have inadvertentnegative effects if only part of the community orecosystem is considered Models for the NavajoDam (San Juan River New Mexico) show thatadequately timed releases could simultaneouslyenhance native and minimize non-native fishpopulations (121) thus predicted benefits couldsurpass those of mimicking a natural flow re-gime potentially persisting even during dryyears Another study using the ldquodesignerrdquo flowparadigm (114) attempted to understand howdifferent facets of flow variability control fisheryproduction in the Lower Mekong floodplains Time-
series models combining discharge and fisherycatch suggested that enhanced flood pulses andlong inter-flood intervals may increase fisheryyieldsAlthough environmental flow practices have
mostly focused on re-creating discrete flowevents (eg flood pulses) these can still serve asvaluable experiments An iconic example of alarge-scale flow release is Minute 319 an amend-ment to the United StatesndashMexico treaty allow-ing opening of floodgates in the Lower ColoradoRiver and a one-time rewetting of the parchedColorado River Delta (122) Although this pulsed-flow experiment did not promote establishmentof native woody riparian seedlings such trialscan inform future restoration practices for ex-ample by determining whether the provision ofbare moist substrate enhances seed germina-tion (123) Hypothesis-driven experimental flowreleases combined with deep knowledge of tar-get ecosystems can provide insights into resto-ration actions [eg (124)] Still an outstandingquestion is how flow regime restoration couldbe leveraged to restore other important alteredaspects of the ecosystem such as sediment andthermal regimes For example cold-water releasesby existing dams in climate-vulnerable basinscould prevent organisms that thrive at low tem-peratures from becoming replaced by general-ist warm-tolerant taxa (125)
Although the concept of environmental flowsis generally associated with impounded riversits relevance to urban streams is clear (126)However because frequent high storm flowsare viewed as the problem reducing them gen-erally becomes the only restoration goal This isoften attempted by shifting the spatial distribu-tion of velocities within channels through ma-nipulation of channel sinuosity installation offlow-diverting weirs or adding other structuresThese approaches may reduce local erosion butrarely result in levels of water quality ecosystemprocesses or biodiversity in urban streams thatcome close to those of unimpacted streams(127 128) Focusing on the channel ratherthan the watershed context at large means thatkey ecosystemprocesses can remain compromised(129) A recent study compared time series ofmetabolism in two streamsmdashonewith stormwaterponds in the watershed and the other restoredby altering channel slope banks and in-streamstructural complexity (103) The study found thatpeaks in GPP and ER in the ldquorestoredrdquo streamwere higher more prone to resets induced byflashy storms and that recovery time scales werelonger compared with the stream in a watershedwith stormwater measures (Fig 6) Ponds areone type of nature-inspired green infrastructureused to restore streamflow tomore natural condi-tions tree plantings constructed wetlands andgrass swales can in turn replace gray infrastruc-ture such as concrete tunnels that capture andstore waterDistributing green infrastructure in water-
sheds helps restore streams because it targetsthe cause of streamflow alteration includinginsufficient infiltration groundwater rechargeand more generally limited watershed storagecapacity (129) By tackling the cause rather thanthe symptoms of flow alteration these practicescan reduce ecological impacts that result fromthe cascading effects of altered flows includingpoor water quality and temperature spikes Re-search on the effectiveness of green infrastructureon restoring streams is sparse and limited bylack of data before and after implementationalthough studies are beginning to appear Forexample upland green infrastructure in theform of shallow wetland-like pools that capturestormwater and reconnect it to the groundwaterhas been shown to enhance baseflows while re-ducing peak flows and pollutant loadsmore thanrestoration structures within the mainstem ofthe receiving stream (130) Similar structuresbuilt between stormwater outfalls and perennialstreams did not modulate the timing magnitudeor duration of urban storm responses nor didthey improve water quality or biodiversity (131)illustrating the need for researchers to understandhow exact design and watershed context influ-ence performance
Spatiotemporal variability andrestoration designs
Research showing how local and watershed-scale environmental drivers vary temporally andalong river networks in ways that shape biota
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 7 of 13
Flow pulses (storms) flashy flowsDroughts intermittencyLowstable flowsAltered timing seasonality
DOC N P
DetritusMicrobial
stoichiometric constraints
Excretion exudates
Detrital pathway
Algal pathway
Algal stoichiometric constraints
Organismal dispersal source-sink dynamics
Algae
Community replacement
ndash ndash +
ndash ndash + ndash
+
+ndash +
Invasions
Turnover
Consumer stoichiometric constraints
Fig 5 Flow variation affects biota and ecosystem processes and food webs integrate bothHydrologic connectivity along river networks is a first landscape-level filter constraining dispersal oforganisms in river networks Extreme flow variation (floods and droughts) are a source of disturbanceto all organisms (gray box) but particularly to the higher trophic levels given their higher metabolicdemands In turn flow variation controls inputs of nutrients and terrestrial leaf litter to the stream whichinfluences microbial activity and the relative contributions of the detrital (brown) and algal (green)pathways in sustaining aquatic consumers Not just quantity but also food quality (eg CN and CPratios) influence dietary choices of consumers and thus trophic transfer efficiency Finally alteration offlow regimes may influence all these compartments and may also open niches for new players Ifbiological invasions occur at the top of the food web these have the potential to divert energy away fromnative predators and to alter food-web structure through top-down controls
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
and ecosystem processes can help prioritizeselection of sites for restoration For exampleusing the concept of river network saturation(55) to identify river sections where in-streamremoval of nutrients cannot keep up with highsource inputs of nutrients suggests that restorationactions targeting those sections may dispropor-tionately improve water quality Identification ofsubcatchments that disproportionately influencestream chemistry may also be informed by com-bining methods from landscape ecology andcatchment hydrology focusing on the spatialscale of variance in water chemistry (132) Head-waters are often prioritized for restoration be-
cause poor water quality in them can havedownstream impacts Research showing thatlocal environmental constraints on river com-munities are important in headwaters as well asin the larger channels but dispersal constraintsare much less important in the former (133 134)implies that headwaters should be prioritizedHowever research quantifying time-varying con-tributions of local communities to river-wide(network) biodiversity can also inform prioritiza-tion by identifying sites that host a dispropor-tionate share of species or that host uniquespeciesmdashfor example because they shelter sensi-tive taxa during critical flow bottlenecks This idea
encapsulates the concept of keystone habitatsor local sites that strongly influence metacom-munity dynamics (135) This has been appliedto aquatic invertebrates across intermittent rivernetworks by tracking time-varying site-specificcontributions to community dissimilarity (or betadiversity) (136)Other approaches to prioritize restoration ac-
tions may be based on contributions to system-wide stability rather than numbers of individualsor species For example the restoration of fall-run Chinook salmon portfolios may be assessedvia the contributions that different periods andlocationsmake tometapopulation-wide stabilityowing to spatiotemporal variation in habitatconditions (137) Additionally interventionscould promote conditions that are not impor-tant now but could be important in the future[ie proto-refugia (138)] For example resto-ration of groundwater-influenced habitats couldmitigate future stress to drought-sensitive taxaunder scenarios of increased intermittencyPatches of undisturbed land that protect flowregimes in streams within a larger catchmentthat is otherwise urbanized can be sources ofcolonists to restored sites Priorities that focuson present degradation of river sectionsmay failto address site contribution to impairment orrecovery at broader scales or under differentscenarios These insights indicate a great poten-tial for new methods to prioritize restoration ofboth urban and regulated streamsWe know little about how altered flow regimes
propagate through entire river networks butwork is starting to assess the cumulative effectsof dams on flow regimes as well as dam-levelcontributions to alteration Although these ap-proaches have largely focused on dam cascades[eg (139)] if expanded they could help identifyfavorable sections in river networks to developenvironmental flow operations Here exper-imentation could again help with testing ofnew strategies For instance large-scale flowexperiments could be designed that coordinatereleases among dams or between dams and free-flowing tributaries to enhance the restorativeeffects of natural pulses (140)
Hydrologic connectivityinfluences restoration
Recovering critical flow paths and their connec-tivity to rivers is important but it remains asubstantial challenge for flow-degraded riversand streams Whereas regulated rivers tend tohave reduced lateral and longitudinal hydro-logic connectivity urban streams have unnaturallevels of connectivity because pipes and over-land runoff route water quickly In both cases fun-damental hydroecological insights into streambedstructure landscape storage capacity and ground-water connectivity can help to envision novel re-storation approaches (130 141 142) More generallyhydrologic connectivity is becoming a frameworkto identify alternative ways to restore streams viastormwater management (143) For example arecent study suggested that reducing road densityand hydrologic connectivity between roads
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 8 of 13
ER
Disc
harg
e (f
t3 s)
GPPG
PP o
r ER
(g O
2 m-2
day
-1)
Discharge
Watershed BMPs
In-stream restoration
0
ndash4
4
0
ndash4
4
4
2
6
4
2
6
Jan Sept July May Mar Nov Jan
A
B C
Fig 6 Green infrastructure as a means to restore stream ecosystems (A) Discharge andecosystem respiration (ER more-negative values mean higher respiration) and gross primaryproduction (GPP) for two urban streams in which restoration efforts involved (top) modifications inthe stream channel and (bottom) traditional stormwater best management practices (BMPs)[Reproduced with permission from (103)] (B and C) Example of a comprehensive greeninfrastructure approach to ecologically restore an urban headwater stream that also supports thelifestyle of resident urban communities (B) Stream at the onset of restoration and (C) afterrestoration [Reproduced with permission from Greenworks Portland OR]
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 9 of 13
Table 1 Research challenges and opportunities Despite the progress that has been made in understanding how flow regimes affect biota and ecosystem
processes major challenges persist that prevent a complete understanding of the flow-biota-ecosystem processes nexus Making progress on these
challenges requires a mix of fundamental and applied research
Topic Research challenge Opportunity
Connecting
organismal to
ecosystem-level
processes
Research on the effects of flow on organisms
and ecosystem processes has followed parallel
tracksmdashhindering our mechanistic
understanding of the flow-biota-ecosystem
processes nexus
Generate theory that connects concepts and frameworks
that are well accepted in river community and ecosystem
ecology For example the notion of environmental filters
that connect regional to local composition via a combination
of dispersal and environmental conditions (community
ecology) with analogous theory on solute transport versus
reactivity (ecosystem ecology)
Our current understanding of the links between
phenology of biota and ecosystem processes
is precursory
Use high-frequency sensors to characterize metabolic
regimes at sites with ongoing organismal monitoring
Leverage citizen-science biodiversity data to detect
spatiotemporal shifts in animal phenology
(eg insect emergence fish spawning)
Use remote sensing tools to better understand
links between hydrology and aquatic and riparian
plant phenology
The link between stoichiometry and flow
variation remains largely unexplored
Repurpose data that is already being collected to test
flow-stoichiometry relationships eg studies using C and
N stable isotopes to describe CN ratios across flow
regime types Perform field experiments to quantify
indirect effects of invertebrate and fish consumption
on ecosystem metabolism
Ecological
consequences of
hydrologic
connectivity
The importance of anaerobic metabolism in
carbon processing as flows cease and parts of
streambeds dry is a major frontier research area
Create networks of sites across different geologies
and flow regimes that collect time-series data on
methane nitrous oxide and carbon dioxide fluxes
from intermittent streams
We ignore how biota in the wet phase is
influenced by processes and patterns
from the dry phase
Research the functions of dry riverbeds Use camera traps and
drone technology to investigate the role of dry riverbeds
as a dispersal corridor metabarcoding to quantify
ldquodark biodiversityrdquo in the hyporheic
Wetlands and their links to stream networks are poorly
delineated globally Even in the United States and
Europe current wetland inventories do not capture
these links yet understanding how their connectivity
to river systems influences network-scale ecosystem
processes is needed
Analyze time series from high-frequency sensors distributed in
river networks varying in wetland connectivity Further
development and application of time-series methods
used by neuroscientists
We need to identify the relationship between
varying flows and solute dynamics
associated with changes in catchment-scale
hydrologic exchanges
Design collaborative work among catchment hydrologists
and stream ecologists combining methods to estimate
transit times residence times and identify sources
across heterogeneous landscapes Link these properties to
land management and water quality aspects
Scaling and
transferring flow-
ecology relationships
across space and time
We ignore how biodiversity responds to
propagation of flow regime alteration
across river networks
Leverage extensive hydrometric networks (eg United States
Geological Survey National Water Information System)
to perform spatially replicated time-series analyses
on flow and use these to generate expectations
on how river network position should mediate
biodiversity responses to flow alteration
Flow-ecology relationships do not account for
nonstationarity and abrupt change
Use process-based models that account for mechanisms
and associated uncertainties When using time-series
data and models select methods that are robust
to nonstationarity Linear flow-ecology relationships
should not be assumed
Current flow-ecology relationships assume that
responses of species do not change over time
Consider flow regime alteration as a process with ecological and
evolutionary consequences Include intraspecific variation
in traits and potential for rapid evolution in models
continued on next page
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
and streams may improve water quality morethan other restoration options (70) Additionallyresearch on the age and lability of DOC enteringstreams has shown how precipitation or land dis-turbance can change watershed flow paths (53)and this could inspire new restoration approachesfocused on land use and soil properties Manystudies on hydrologic connectivity and networkstructure suggest a need for restoration to focuson landscape heterogeneity especially on thepresence ofwetlands lakes and ponds (144) Suchfeaturesare frequently lost inurbanizedcatchmentsand outcomes could be improved by restoring orincreasing connectivitywith these ecosystems (145)Hydrologic connectivity associated with ground-water inputs or abstractions land-use manage-ment and return flows fromwastewater treatmentplants also need to be considered in these initia-tives (146)Restoring hydrologic connectivity is also key
to achieving biodiversity outcomesmdashparticularlyin dammed systems Longitudinal connectivityfor fish inhabiting dam-altered rivers can beenhanced by the construction of fishways andsophisticated passage facilities However moni-toring data on the effectiveness of such inter-
ventions are still scarce and available evidencesuggests that passage effectivity largely dependson technology used and species-specific fish be-havior (147 148) Although trap-and-transportprograms for migratory fishes can help augmentthreatened populations (149) these practicesoften increase straying and thus the potentialfor genetic homogenization as seen from ju-venile salmon collected at dams and transporteddownstream to increase out-migration success(150) Overall a combination of engineering andmanagement solutions may be best to restorefunctional connectivity in dam-altered riversThe link between hydrologic connectivity andecological restoration is also central to con-siderations on where to build remove and re-operate hydrologic infrastructure Hundredsof dams have been intentionally removed sincethe 1970s and removal of large dams such asthe Elwha River Dam (Washington USA) hasshown that restoring longitudinal connectivitycan have immediate geomorphic effects with com-plex temporal dynamics (151) Infrastructure canalso be designed or reoperated to manage forrestoring lateral connectivity and enhanced flood-plain processes For example the Yolo Bypass in
the Sacramento River California has becomea prime rearing and migration site for chinooksalmon showing that flood control and habitatfor fish and wildlife do not need to be at odds(152) All these examples show the importanceof integrating hydrologic connectivity in ecolog-ical restoration whether the goal is to minimizethe ecological effects of fragmentation restorefree-flowing watercourses or operate green infra-structure to enhance infiltration floodplain con-nectivity or other ecological processes
Research challenges and opportunities
Two decades of concerted efforts to research theeffects of flow regime alteration have tremen-dously advanced our understanding of the waysin which streamflow influences biota and eco-system processes However important challengesremain These aremainly related to the scarcity ofscience connecting organismal to ecosystem-levelprocesses through for example microbial linksthe difficulty of studying watershed-level phys-ical and biogeochemical heterogeneity and itsinfluence on riverine processes and biota thecomplexity of parsing out the direct effects offlow from flow-mediated effects and co-occurring
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 10 of 13
Topic Research challenge Opportunity
Cascading
implications of
hydrologic
alteration
We have a poor understanding of the
interactions among physical drivers
Build on large-scale flow experiments to test hypotheses on the
combined effects of altered flow sediment and thermal
regimes Experiment with multilevel intakes in reservoirs
Promote collection of non-flow variables (eg temperature
water quality parameters) at hydrometric stations
At low flows biota are being exposed to high
concentrations of new classes of pollutants
whose impacts are poorly known especially if
biotic impacts interact with impacts on
ecosystem processes
Design realistic experimental studies to advance knowledge
on the ecological effects of multiple stressors
Collect long-term data at sites varying in contaminant
inputs and hydrologic regimes Link microbial to
organismal and ecosystem-level responses
Using hydroecological
science to inform
river ecosystem
restoration
Environmental flow science largely ignores
human dimensions
Use methods that allow identifying causal networks
in complex socioenvironmental systems Include
human needs and thresholds in research on
flow-ecology relationships
Environmental flows are generally prescribed
at the scale of single infrastructures restoration
designs and assessment are generally at reach scales
Promote coordinated large-scale flow experiments
and restoration designs at the river basin scale
Extend assessment of green infrastructure
effectiveness beyond individual structures to
infrastructure networks and link both
to river outcomes
Environmental flows in impounded rivers
generally target the conservation of some
particular species rather than outcomes at the
food-web or ecosystem level
Advance fundamental research on microbialndash
ecosystem process linkages Expand the notion
of designer flows to food-web structure and
ecosystem services when possible
Environmental flow science has largely
ignored surface waterndashgroundwater
interactions
Expand science to understand the link between
restoration actions groundwater recharge and
inputs to streams Provide science relevant to
the implementation of new legislation on
groundwater management (eg Sustainable
Groundwater Management Act in California)
Uncertainty regarding future
hydroclimate and
river flows is growing
Refine regional climate models Promote research on
the interaction between land-use change and
evapotranspiration Incorporate groundwater
influence in macroscale hydrology models
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
stressors (particularly in urban streams) and thechallenge of predicting outcomeswhen both thephysical and the biological environments arechangingAchieving a predictive understanding of hy-
droecological relationships is essential to design-ing successful self-sustaining restoration actionsin river ecosystems However translating basichydroecological insights into restoration practicerequires understanding the complexities of work-ing within a human-dominated water cycle Al-though it is rarely possible to restore all facetsof a natural flow regime ecologically informeddesigns may help mimic key aspects of the nat-ural flow regime (eg periodic pulse flows) orcreate regimes that are not natural but maximizeoutcomes given socioeconomic constraints InTable 1 we identify persisting and emergingchallenges in hydroecological research We paireach challenge with an opportunity (related toresearch design technology or implementation)We contend that these challenges should beprioritized to advance process-based river eco-system restoration
REFERENCES AND NOTES
1 N L Poff et al The natural flow regime Bioscience 47769ndash784 (1997) doi 1023071313099
2 D A Lytle N L Poff Adaptation to natural flow regimesTrends Ecol Evol 19 94ndash100 (2004) doi 101016jtree200310002 pmid 16701235
3 S Sabater A Elosegi R Ludwig in Multiple Stressors in RiverEcosystems (Elsevier 2019) pp 1ndash22
4 United Nations New UN Decade on Ecosystem Restorationoffers unparalleled opportunity for job creation food securityand addressing climate change (2019) httpswwwunenvironmentorgnews-and-storiespress-releasenew-un-decade-ecosystem-restoration-offers-unparalleled-opportunity
5 A W Lorenz P Haase K Januschke A SundermannD Hering Revisiting restored river reaches - Assessingchange of aquatic and riparian communities afterfive years Sci Total Environ 613-614 1185ndash1195(2018) doi 101016jscitotenv201709188pmid 28954379
6 M A Palmer K L Hondula B J Koch Ecological Restorationof Streams and Rivers Shifting Strategies and ShiftingGoals (2014) vol 45
7 T J Beechie et al Process-based principles for restoringriver ecosystems Bioscience 60 209ndash222 (2010)doi 101525bio20106037
8 S E Bunn A H Arthington Basic principles and ecologicalconsequences of altered flow regimes for aquaticbiodiversity Environ Manage 30 492ndash507 (2002)doi 101007s00267-002-2737-0 pmid 12481916
9 K D Fausch Y Taniguchi S Nakano G D GrossmanC R Townsend Flood disturbance regimes influence rainbowtrout invasion success among five holarctic regions EcolAppl 11 1438ndash1455 (2001) doi 1018901051-0761(2001)011[1438FDRIRT]20CO2
10 M E Power M S Parker W E Dietrich Seasonalreassembly of a river food web Floods droughts andimpacts of fish Ecol Monogr 78 263ndash282 (2008)doi 10189006-09021
11 V Acuntildea A Giorgi I Muntildeoz U R S Uehlinger S SabaterFlow extremes and benthic organic matter shape themetabolism of a headwater Mediterranean streamFreshwater Biol 49 960ndash971 (2004) doi 101111j1365-2427200401239x
12 K Tockner F Malard J V Ward An extension of the floodpulse concept Hydrol Processes 14 2861ndash2883 (2000)doi 1010021099-1085(20001112)141617lt2861AID-HYP124gt30CO2-F
13 J S Perkin et al Fragmentation and dewatering transformGreat Plains stream fish communities Ecol Monogr 8573ndash92 (2015) doi 10189014-01211
14 J L Sabo D M Post Quantifying periodic stochastic andcatastrophic environmental variation Ecol Monogr 7819ndash40 (2008) doi 10189006-13401
15 F Shuai et al Silver carp larva abundance in response toriver flow rate revealed by cross-wavelet modellingEcol Modell 383 98ndash105 (2018) doi 101016jecolmodel201805020
16 J D Tonkin M T Bogan N Bonada B Rios-ToumaD A Lytle Seasonality and predictability shape temporalspecies diversity Ecology 98 1201ndash1216 (2017)doi 101002ecy1761 pmid 28144975
17 N L Poff Beyond the natural flow regime Broadening thehydro‐ecological foundation to meet environmental flowschallenges in a non‐stationary world Freshwater Biol 631011ndash1021 (2018) doi 101111fwb13038
18 A Ruhi X Dong C H McDaniel D P Batzer J L SaboDetrimental effects of a novel flow regime on the functionaltrajectory of an aquatic invertebrate metacommunity GlobChange Biol 24 3749ndash3765 (2018) doi 101111gcb14133pmid 29665147
19 M T Bogan D A Lytle Severe drought drives novelcommunity trajectories in desert stream pools FreshwaterBiol 56 2070ndash2081 (2011) doi 101111j1365-2427201102638x
20 G Woodward et al The effects of climatic fluctuations andextreme events on running water ecosystems Philos TransR Soc London Ser B 371 20150274 (2016) pmid 27114576
21 M A Palmer C M Swan K Nelson P Silver R AlvestadStreambed landscapes Evidence that stream invertebratesrespond to the type and spatial arrangement of patchesLandsc Ecol 15 563ndash576 (2000) doi 101023A1008194130695
22 D Schmera et al Does isolation influence the relative role ofenvironmental and dispersal‐related processes in streamnetworks An empirical test of the network positionhypothesis using multiple taxa Freshwater Biol 63 74ndash85(2018) doi 101111fwb12973
23 R Henriques‐Silva et al A comprehensive examination of thenetwork position hypothesis across multiple rivermetacommunities Ecography 42 284ndash294 (2019)doi 101111ecog03908
24 K L Jaeger J D Olden N A Pelland Climate change poisedto threaten hydrologic connectivity and endemic fishes indryland streams Proc Natl Acad Sci USA 11113894ndash13899 (2014) doi 101073pnas1320890111pmid 25136090
25 S Garbin E A Celegon P Fanton G Botter Hydrologicalcontrols on river network connectivity R Soc Open Sci 6181428 (2019) doi 101098rsos181428 pmid 30891270
26 F Cattaneacuteo B Hugueny N Lamouroux Synchrony in browntrout Salmo trutta population dynamics A lsquoMoran effectrsquo onearly‐life stages Oikos 100 43ndash54 (2003) doi 101034j1600-0706200311912x
27 A Terui et al Metapopulation stability in branching rivernetworks Proc Natl Acad Sci USA 115 E5963ndashE5969(2018) doi 101073pnas1800060115 pmid 29895695
28 P A P Moran The statistical analysis of the Canadian Lynxcycle Aust J Zool 1 291ndash298 (1953) doi 101071ZO9530291
29 G D Grossman P B Moyle J O Whitaker Jr Stochasticityin structural and functional characteristics of an Indianastream fish assemblage A test of community theoryAm Nat 120 423ndash454 (1982) doi 101086284004
30 D A Lytle D M Merritt J D Tonkin J D OldenL V Reynolds Linking river flow regimes to riparian plantguilds A community-wide modeling approach Ecol Appl 271338ndash1350 (2017) doi 101002eap1528 pmid 28263426
31 A Ruhiacute E E Holmes J N Rinne J L Sabo Anomalousdroughts not invasion decrease persistence of native fishesin a desert river Glob Change Biol 21 1482ndash1496 (2015)doi 101111gcb12780 pmid 25430731
32 J D Tonkin D M Merritt J D Olden L V ReynoldsD A Lytle Flow regime alteration degrades ecologicalnetworks in riparian ecosystems Nat Ecol Evol 2 86ndash93(2018) doi 101038s41559-017-0379-0 pmid 29180707
33 M C Mims J D Olden Fish assemblages respond toaltered flow regimes via ecological filtering of lifehistory strategies Freshwater Biol 58 50ndash62 (2013)doi 101111fwb12037
34 P T J Johnson J D Olden M J Vander Zanden Daminvaders Impoundments facilitate biological invasions intofreshwaters Front Ecol Environ 6 357ndash363 (2008)doi 101890070156
35 N Bonada S Doledec B Statzner Taxonomic and biologicaltrait differences of stream macroinvertebrate communitiesbetween mediterranean and temperate regions Implicationsfor future climatic scenarios Glob Change Biol 131658ndash1671 (2007) doi 101111j1365-2486200701375x
36 M I Pyne N L Poff Vulnerability of stream communitycomposition and function to projected thermal warming andhydrologic change across ecoregions in the western UnitedStates Glob Change Biol 23 77ndash93 (2017) doi 101111gcb13437 pmid 27429092
37 D B Herbst S D Cooper R B Medhurst S W WisemanC T Hunsaker Drought ecohydrology alters the structureand function of benthic invertebrate communities inmountain streams Freshwater Biol 64 886ndash902 (2019)doi 101111fwb13270
38 M T Bogan E T Chester T Datry A L MurphyB J Robson A Ruhi R Stubbington J E Whitney inResistance Resilience and Community Recovery in IntermittentRivers and Ephemeral Streams T Datry N Bonada A BoultonEds (Academic Press 2017) pp 349ndash376
39 S Houmlckendorff et al Characterizing fish responses to a riverrestoration over 21 years based on speciesrsquo traits ConservBiol 31 1098ndash1108 (2017) doi 101111cobi12908pmid 28218807
40 D Bruno et al Structural and functional responses ofinvertebrate communities to climate change andflow regulation in alpine catchments Glob Change Biol25 1612ndash1628 (2019) doi 101111gcb14581pmid 30698905
41 T W H Aspin et al Extreme drought pushes streaminvertebrate communities over functional thresholds GlobChange Biol 25 230ndash244 (2019) doi 101111gcb14495pmid 30346098
42 S E Tank J B Fellman E Hood E S Kritzberg Beyondrespiration Controls on lateral carbon fluxes across theterrestrial-aquatic interface Limnol Oceanogr Lett 3 76ndash88(2018) doi 101002lol210065
43 A P Appling et al The metabolic regimes of 356 rivers inthe United States Sci Data 5 180292 (2018) doi 101038sdata2018292 pmid 30532078
44 P Savoy et al Metabolic rhythms in flowing watersAn approach for classifying river productivity regimesLimnol Oceanogr (2019) doi 101002lno11154
45 E S Bernhardt et al The metabolic regimes of flowingwaters Limnol Oceanogr 63 S99ndashS118 (2018)doi 101002lno10726
46 M Colls X Timoner S Sabater V Acuntildea Effects of durationand frequency of the non-flow period on stream metabolismEcosystems 101007s10021-019-00345-1 (2019)doi 101007s10021-019-00345-1
47 A J Ulseth E Bertuzzo G A Singer J Schelker T J BattinClimate-induced changes in spring snowmelt impactecosystem metabolism and carbon fluxes in an Alpine streamnetwork Ecosystems 21 373ndash390 (2018) doi 101007s10021-017-0155-7
48 B O L Demars Hydrological pulses and burning of dissolvedorganic carbon by stream respiration Limnol Oceanogr 64406ndash421 (2019) doi 101002lno11048
49 R T Hensley L Kirk M Spangler M N Gooseff M J CohenFlow extremes as spatiotemporal control points on riversolute fluxes and metabolism J Geophys Res Biogeosci124 537ndash555 (2019) doi 1010292018JG004738
50 J D Hosen et al Enhancement of primary production duringdrought in a temperate watershed is greater in larger riversthan headwater streams Limnol Oceanogr 64 1458ndash1472(2019) doi 101002lno11127
51 L Goacutemez-Gener et al When water vanishes Magnitude andregulation of carbon dioxide emissions from dry temporarystreams Ecosystems 19 710ndash723 (2016) doi 101007s10021-016-9963-4
52 E H Stanley et al The ecology of methane in streams andrivers Patterns controls and global significance EcolMonogr 86 146ndash171 (2016) doi 10189015-1027
53 R T Barnes D E Butman H F Wilson P A RaymondRiverine export of aged carbon driven by flow path depth andresidence time Environ Sci Technol 52 1028ndash1035 (2018)doi 101021acsest7b04717 pmid 29313674
54 G Pinay et al Riparian corridors A new conceptualframework for assessing nitrogen buffering across biomesFront Environ Sci 6 47 (2018) doi 103389fenvs201800047
55 W M Wollheim et al River network saturation conceptFactors influencing the balance of biogeochemical supply and
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 11 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
demand of river networks Biogeochemistry 141 503ndash521(2018) doi 101007s10533-018-0488-0
56 P A Raymond J E Saiers W V Sobczak Hydrological andbiogeochemical controls on watershed dissolved organicmatter transport Pulse-shunt concept Ecology 97 5ndash16(2016) doi 10189014-16841 pmid 27008769
57 C Tunaley D Tetzlaff J Lessels C Soulsby Linking high-frequency DOC dynamics to the age of connected watersources Water Resour Res 52 5232ndash5247 (2016)doi 1010022015WR018419
58 S Bernal et al Supply demand and in-stream retention ofdissolved organic carbon and nitrate during storms inMediterranean forested headwater streams Front EnvironSci 7 60 (2019) doi 103389fenvs201900060
59 J Harvey et al How hydrologic connectivity regulates waterquality in river corridors J Am Water Resour Assoc 55369ndash381 (2019) doi 1011111752-168812691
60 J C Moore et al Detritus trophic dynamics and biodiversityEcol Lett 7 584ndash600 (2004) doi 101111j1461-0248200400606x
61 A D Rosemond et al Freshwater ecology Experimentalnutrient additions accelerate terrestrial carbon loss fromstream ecosystems Science 347 1142ndash1145 (2015)doi 101126scienceaaa1958 pmid 25745171
62 A C Y Yeung J L Musetta-Lambert D P KreutzweiserP K Sibley J S Richardson Relations of interannualdifferences in stream litter breakdown with dischargeBioassessment implications Ecosphere 9 e02423 (2018)doi 101002ecs22423
63 R M Northington J R Webster Experimental reductions instream flow alter litter processing and consumer subsidies inheadwater streams Freshwater Biol 62 737ndash750 (2017)doi 101111fwb12898
64 E Mas-Martiacute I Sanpera-Calbet I Muntildeoz Bottom-up effectsof streambed drying on consumer performance throughchanges in resource quality Aquat Sci 79 719ndash731 (2017)doi 101007s00027-017-0531-6
65 B D Richter G A Thomas Restoring environmental flows bymodifying dam operations Ecol Soc 12 12 (2007)doi 105751ES-02014-120112
66 G Zolezzi A Siviglia M Toffolon B MaioliniThermopeaking in Alpine streams Event characterizationand time scales Ecohydrology 4 564ndash576 (2011)doi 101002eco132
67 T Meiszligner B Sures C K Feld Multiple stressors and therole of hydrology on benthic invertebrates in mountainousstreams Sci Total Environ 663 841ndash851 (2019)doi 101016jscitotenv201901288 pmid 30738264
68 I R Waite et al Effects of urban multi-stressors on threestream biotic assemblages Sci Total Environ 6601472ndash1485 (2019) doi 101016jscitotenv201812240pmid 30743940
69 K C Nelson M A Palmer Stream temperature surges underurbanization and climate change Data models andresponses J Am Water Resour Assoc 43 440ndash452 (2007)doi 101111j1752-1688200700034x
70 J R Blaszczak J M Delesantro Y Zhong D L UrbanE S Bernhardt Watershed urban development controls onurban streamwater chemistry variability Biogeochemistry144 61ndash84 (2019) doi 101007s10533-019-00572-7
71 L Ponsatiacute et al Biofilm responses to flow regulation by damsin Mediterranean rivers River Res Appl 31 1003ndash1016(2015) doi 101002rra2807
72 J M Alberts J J Beaulieu I Buffam Watershed land useand seasonal variation constrain the influence of ripariancanopy cover on stream ecosystem metabolism Ecosystems20 553ndash567 (2017) doi 101007s10021-016-0040-9pmid 30416371
73 R O Hall Jr et al Turbidity light temperature andhydropeaking control primary productivity in the ColoradoRiver Grand Canyon Limnol Oceanogr 60 512ndash526 (2015)doi 101002lno10031
74 J R Blaszczak J M Delesantro D L Urban M W DoyleE S Bernhardt Scoured or suffocated Urban streamecosystems oscillate between hydrologic and dissolvedoxygen extremes Limnol Oceanogr 64 877ndash894 (2019)doi 101002lno11081
75 J-R Mor et al Dam regulation and riverine food-webstructure in a Mediterranean river Sci Total Environ 625301ndash310 (2018) doi 101016jscitotenv201712296pmid 29289778
76 H A Wellard Kelly et al Macroinvertebrate diets reflecttributary inputs and turbidity-driven changes in food
availability in the Colorado River downstream of GlenCanyon Dam Freshw Sci 32 397ndash410 (2013)doi 10189912-0881
77 S J Imberger R M Thompson M R Grace Urbancatchment hydrology overwhelms reach scale effects ofriparian vegetation on organic matter dynamics FreshwaterBiol 56 1370ndash1389 (2011) doi 101111j1365-2427201102575x
78 R T Martins A S Melo J F Gonccedilalves Jr N HamadaLeaf-litter breakdown in urban streams of Central AmazoniaDirect and indirect effects of physical chemical andbiological factors Freshw Sci 34 716ndash726 (2015)doi 101086681086
79 C M Yule J Y Gan T Jinggut K V Lee Urbanizationaffects food webs and leaf-litter decomposition in a tropicalstream in Malaysia Freshw Sci 34 702ndash715 (2015)doi 101086681252
80 T A Newcomer et al Influence of natural and novel organiccarbon sources on denitrification in forest degraded urbanand restored streams Ecol Monogr 82 449ndash466 (2012)doi 10189012-04581
81 A Chaudhary I Kauser A Ray R Poretsky Taxon-drivenfunctional shifts associated with storm flow in an urbanstream microbial community MSphere 3 e00194-18 (2018)doi 101128mSphere00194-18 pmid 29976643
82 T A Kennedy et al Flow management for hydropowerextirpates aquatic insects undermining river food websBioscience 66 561ndash575 (2016) doi 101093bioscibiw059
83 A Glazaczow D Orwin M Bogdziewicz Increasedtemperature delays the late-season phenology of multivoltineinsect Sci Rep 6 38022 (2016) doi 101038srep38022pmid 27905493
84 S G Fisher L J Gray N B Grimm D E Busch Temporalsuccession in a desert stream ecosystem following flashflooding Ecol Monogr 52 93ndash110 (1982) doi 1023072937346
85 G J Scrimgeour M J Winterbourn Effects of floods onepilithon and benthic macroinvertebrate populations in anunstable New Zealand river Hydrobiologia 171 33ndash44 (1989)doi 101007BF00005722
86 E Bondar-Kunze S Maier D Schoumlnauer N Bahl T HeinAntagonistic and synergistic effects on a stream periphytoncommunity under the influence of pulsed flow velocityincrease and nutrient enrichment Sci Total Environ 573594ndash602 (2016) doi 101016jscitotenv201608158pmid 27585428
87 M J Cashman G L Harvey G Wharton M C Bruno Woodmitigates the effect of hydropeaking scour on periphytonbiomass and nutritional quality in semi-natural flumesimulations Aquat Sci 79 459ndash471 (2017) doi 101007s00027-016-0510-3
88 W F Cross et al Food-web dynamics in a large riverdiscontinuum Ecol Monogr 83 311ndash337 (2013)doi 10189012-17271
89 D M Warfe et al Productivity disturbance and ecosystemsize have no influence on food chain length in seasonallyconnected rivers PLOS ONE 8 e66240 (2013)doi 101371journalpone0066240
90 T D Jardine et al Consumer-resource coupling in wet-drytropical rivers J Anim Ecol 81 310ndash322 (2012)doi 101111j1365-2656201101925x pmid 22103689
91 K O Winemiller et al Pulsing hydrology determines top‐down control of basal resources in a tropical riverndashfloodplainecosystem Ecol Monogr 84 621ndash635 (2014)doi 10189013-18221
92 D M Post The long and short of food-chain length TrendsEcol Evol 17 269ndash277 (2002) doi 101016S0169-5347(02)02455-2
93 D M Post G Takimoto Proximate structural mechanismsfor variation in food‐chain length Oikos 116 775ndash782 (2007)doi 101111j0030-1299200715552x
94 P A McHugh A R McIntosh P G Jellyman Dual influencesof ecosystem size and disturbance on food chain length instreams Ecol Lett 13 881ndash890 (2010) doi 101111j1461-0248201001484x pmid 20482579
95 A Ruhiacute et al Flow regulation increases food-chain lengththrough omnivory mechanisms in a Mediterranean rivernetwork Freshwater Biol 61 1536ndash1549 (2016) doi 101111fwb12794
96 J L Sabo J C Finlay T Kennedy D M Post The role ofdischarge variation in scaling of drainage area and food chainlength in rivers Science 330 965ndash967 (2010) doi 101126science1196005 pmid 20947729
97 A R McIntosh et al Capacity to support predators scaleswith habitat size Sci Adv 4 eaap7523 (2018) doi 101126sciadvaap7523 pmid 29978038
98 J H Brown J F Gillooly A P Allen V M Savage G B WestToward a metabolic theory of ecology Ecology 851771ndash1789 (2004) doi 10189003-9000
99 K Zou E Theacutebault G Lacroix S Barot Interactions betweenthe green and brown food web determine ecosystemfunctioning Funct Ecol 30 1454ndash1465 (2016) doi 1011111365-243512626
100 M J Vanni P B McIntyre Predicting nutrient excretion ofaquatic animals with metabolic ecology and ecologicalstoichiometry A global synthesis Ecology 97 3460ndash3471(2016) doi 101002ecy1582 pmid 27912023
101 T B Parr K A Capps S P Inamdar K A Metcalf Animal-mediated organic matter transformation Aquatic insects as asource of microbially bioavailable organic nutrients andenergy Funct Ecol 33 524ndash535 (2019) doi 1011111365-243513242
102 C P Arango et al Urban infrastructure influences dissolvedorganic matter quality and bacterial metabolism in an urbanstream network Freshwater Biol 62 1917ndash1928 (2017)
103 L G Larsen J W Harvey Disrupted carbon cycling inrestored and unrestored urban streams Critical timescalesand controls Limnol Oceanogr 62 S160ndashS182 (2017)doi 101002lno10613
104 H M Halvorson G White J T Scott M A Evans-WhiteDietary and taxonomic controls on incorporation of microbialcarbon and phosphorus by detritivorous caddisfliesOecologia 180 567ndash579 (2016) doi 101007s00442-015-3464-6 pmid 26497125
105 J J Elser et al Biological stoichiometry from genes toecosystems Ecol Lett 3 540ndash550 (2000) doi 101046j1461-0248200000185x
106 E K Moody et al Consumption explains intraspecificvariation in nutrient recycling stoichiometry in a desert fishEcology 99 1552ndash1561 (2018) doi 101002ecy2372pmid 29882955
107 N Welti et al Bridging food webs ecosystem metabolismand biogeochemistry using ecological stoichiometry theoryFront Microbiol 8 1298 (2017) doi 103389fmicb201701298 pmid 28747904
108 M A Palmer S Filoso R M Fanelli From ecosystems toecosystem services Stream restoration as ecologicalengineering Ecol Eng 65 62ndash70 (2014) doi 101016jecoleng201307059
109 M J Burns T D Fletcher C J Walsh A R LadsonB E Hatt Hydrologic shortcomings of conventional urbanstormwater management and opportunities for reformLandsc Urban Plan 105 230ndash240 (2012) doi 101016jlandurbplan201112012
110 C R Liermann C Nilsson J Robertson R Y NgImplications of dam obstruction for global freshwater fishdiversity Bioscience 62 539ndash548 (2012) doi 101525bio20126265
111 J D Olden R J Naiman Incorporating thermal regimes intoenvironmental flows assessments Modifying dam operationsto restore freshwater ecosystem integrity Freshwater Biol55 86ndash107 (2010) doi 101111j1365-2427200902179x
112 D B Osmundson R J Ryel V L Lamarra J Pitlick Flowndashsedimentndashbiota relations Implications for river regulationeffects on native fish abundance Ecol Appl 12 1719ndash1739(2002)
113 D von Schiller et al Regulation causes nitrogen cyclingdiscontinuities in Mediterranean rivers Sci Total Environ540 168ndash177 (2016) doi 101016jscitotenv201507017pmid 26233271
114 J L Sabo et al Designing river flows to improve foodsecurity futures in the Lower Mekong Basin Science 358eaao1053 (2017) doi 101126scienceaao1053pmid 29217541
115 W Xu et al Improving the multi-objective performance ofrainwater harvesting systems using real-time controltechnology Water 10 147 (2018) doi 103390w10020147
116 M Acreman et al Environmental flows for naturalhybrid and novel riverine ecosystems in a changingworld Front Ecol Environ 12 466ndash473 (2014)doi 101890130134
117 N L Poff et al Sustainable water management under futureuncertainty with eco-engineering decision scaling Nat ClimChang 6 25ndash34 (2016) doi 101038nclimate2765
118 J D Kiernan P B Moyle P K Crain Restoring native fishassemblages to a regulated California stream using the
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 12 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
natural flow regime concept Ecol Appl 22 1472ndash1482(2012) doi 10189011-04801 pmid 22908707
119 D L Propst K B Gido Responses of native andnonnative fishes to natural flow regime mimicry in theSan Juan River Trans Am Fish Soc 133 922ndash931 (2004)doi 101577T03-0571
120 T K Pool J D Olden Assessing long‐term fish responses andshort‐term solutions to flow regulation in a dryland river basinEcol Freshwat Fish 24 56ndash66 (2015) doi 101111eff12125
121 W Chen J D Olden Designing flows to resolve human andenvironmental water needs in a dam-regulated riverNat Commun 8 2158 (2017) doi 101038s41467-017-02226-4 pmid 29255194
122 S M Nelson et al A history of the 2014 Minute 319environmental pulse flow as documented by fieldmeasurements and satellite imagery Ecol Eng 106733ndash748 (2017) doi 101016jecoleng201610040
123 P B Shafroth et al A large-scale environmental flowexperiment for riparian restoration in the Colorado RiverDelta Ecol Eng 106 645ndash660 (2017) doi 101016jecoleng201702016
124 E P Glenn P L Nagler P B Shafroth C J JarchowEffectiveness of environmental flows for riparian restorationin arid regions A tale of four rivers Ecol Eng 106 695ndash703(2017) doi 101016jecoleng201701009
125 S Hotaling D S Finn J Joseph Giersch D W WeisrockD Jacobsen Climate change and alpine stream biologyProgress challenges and opportunities for the futureBiol Rev Camb Philos Soc 92 2024ndash2045 (2017)doi 101111brv12319 pmid 28105701
126 C J Walsh T D Fletcher M J Burns Urban stormwaterrunoff A new class of environmental flow problem PLOS ONE7 e45814 (2012) doi 101371journalpone0045814pmid 23029257
127 A Selvakumar T P OrsquoConnor S D Struck Role of streamrestoration on improving benthic macroinvertebrates and in-stream water quality in an urban watershed Case studyJ Environ Eng 136 127ndash139 (2010) doi 101061(ASCE)EE1943-78700000116
128 R W Lammers B P Bledsoe What role does streamrestoration play in nutrient management Crit Rev EnvironSci Technol 47 335ndash371 (2017) doi 1010801064338920171318618
129 H E Golden N Hoghooghi Green infrastructure and itscatchment-scale effects An emerging science WIREs Water5 e1254 (2018) doi 101002wat21254 pmid 29682288
130 M R Williams G Bhatt S Filoso G Yactayo Streamrestoration performance and its contribution to theChesapeake Bay TMDL Challenges posed by climate changein urban areas Estuaries Coasts 40 1227ndash1246 (2017)doi 101007s12237-017-0226-1
131 R M Fanelli K Prestegaard M A Palmer Freshw Sci101086706072 (2019)
132 B W Abbott et al Unexpected spatial stability of waterchemistry in headwater stream networks Ecol Lett 21296ndash308 (2018) doi 101111ele12897 pmid 29282860
133 B L Brown C M Swan Dendritic network structureconstrains metacommunity properties in riverineecosystems J Anim Ecol 79 571ndash580 (2010) doi 101111j1365-2656201001668x pmid 20180874
134 C M Swan B L Brown Metacommunity theory meetsrestoration Isolation may mediate how ecologicalcommunities respond to stream restoration Ecol Appl 272209ndash2219 (2017) doi 101002eap1602 pmid 28718193
135 N Mouquet D Gravel F Massol V Calcagno Extending theconcept of keystone species to communities and ecosystemsEcol Lett 16 1ndash8 (2013) doi 101111ele12014 pmid 23062191
136 A Ruhiacute T Datry J L Sabo Interpreting beta-diversitycomponents over time to conserve metacommunities inhighly dynamic ecosystems Conserv Biol 31 1459ndash1468(2017) doi 101111cobi12906 pmid 28188969
137 L Yamane L W Botsford D P Kilduff Tracking restorationof population diversity via the portfolio effect J Appl Ecol55 472ndash481 (2018) doi 1011111365-266412978
138 D W Braidwood M A Taggart M Smith R AndersenTranslocations conservation and climate change Use ofrestoration sites as protorefuges and protorefugia RestorEcol 26 20ndash28 (2018) doi 101111rec12642
139 Y Wang D Wang Q W Lewis J Wu F Huang A frameworkto assess the cumulative impacts of dams on hydrologicalregime A case study of the Yangtze River Hydrol Processes31 3045ndash3055 (2017) doi 101002hyp11239
140 J L Sabo et al Pulsed flows tributary inputs and food-webstructure in a highly regulated river J Appl Ecol 551884ndash1895 (2018) doi 1011111365-266413109
141 C Magliozzi G Coro R C Grabowski A I PackmanS Krause A multiscale statistical method to identifypotential areas of hyporheic exchange for river restorationplanning Environ Model Softw 111 311ndash323 (2019)doi 101016jenvsoft201809006
142 S S Schwartz B Smith Restoring hydrologic function inurban landscapes with suburban subsoiling J Hydrol 543770ndash781 (2016) doi 101016jjhydrol201610051
143 C J Walsh T D Fletcher A R Ladson Stream restoration inurban catchments through redesigning stormwater systemsLooking to the catchment to save the stream J N AmBenthol Soc 24 690ndash705 (2005) doi 10189904-0201
144 N M Schmadel et al Thresholds of lake and reservoirconnectivity in river networks control nitrogen removalNat Commun 9 2779 (2018) doi 101038s41467-018-05156-x pmid 30018449
145 C D Sayer Conservation of aquatic landscapes Pondslakes and rivers as integrated systems WIREs Water 1573ndash585 (2014) doi 101002wat21045
146 R G Luthy D L Sedlak M H Plumlee D Austin V H ReshWastewater‐effluent‐dominated streams as ecosystem‐
management tools in a drier climate Front Ecol Environ 13477ndash485 (2015) doi 101890150038
147 D W Roscoe S G Hinch Effectiveness monitoring of fishpassage facilities Historical trends geographic patterns andfuture directions Fish Fish 11 12ndash33 (2010) doi 101111j1467-2979200900333x
148 C M Bunt T Castro-Santos A Haro Performanceof fish passage structures at upstream barriers tomigration River Res Appl 28 457ndash478 (2012)doi 101002rra1565
149 D Weigel et al Evaluation of a trap-and-transport programfor a threatened population of steelhead (Oncorhynchusmykiss) Conserv Genet 20 1195ndash1199 (2019) doi 101007s10592-019-01200-5
150 M H Bond et al Combined effects of barge transportationriver environment and rearing location on straying andmigration of adult Snake River fall-run Chinook SalmonTrans Am Fish Soc 146 60ndash73 (2017) doi 1010800002848720161235614
151 A C Ritchie et al Morphodynamic evolution followingsediment release from the worldrsquos largest dam removalSci Rep 8 13279 (2018) doi 101038s41598-018-30817-8pmid 30185796
152 T Sommer et al Californiarsquos Yolo Bypass Evidence thatflood control can be compatible with fisheries wetlandswildlife and agriculture Fisheries 26 6ndash16 (2001)doi 1015771548-8446(2001)026lt0006CYBgt20CO2
153 G Grill et al Mapping the worldrsquos free-flowing riversNature 569 215ndash221 (2019) doi 101038s41586-019-1111-9pmid 31068722
154 G Gassert M Luck M Landis P Reig T Shiao ldquoAqueductglobal maps 21 Constructing decision-relevant global waterrisk indicatorsrdquo Working paper World Resources Institute(2015) wwwwriorgpublicationaqueduct-global-maps-21-indicators
155 A Ruhiacute J D Olden J L Sabo Declining streamflow inducescollapse and replacement of native fishes in the AmericanSouthwest Front Ecol Environ 14 465ndash472 (2016)doi 101002fee1424
156 T C Gouhier A Grinsted V Simko R package Biwaveletconduct univariate and bivariate wavelet analyses (2018)
157 T Rodriacuteguez-Castillo E Esteacutevez A M Gonzaacutelez-FerrerasJ Barquiacuten Estimating ecosystem metabolism toentire river networks Ecosystems (2018) doi 101007s10021-018-0311-8
ACKNOWLEDGMENTS
We greatly appreciate those who read and provided importantinsights on drafts of this paper A Armstrong K HondulaJ Hosen K Leathers G de Mendoza C Maietta I MunozJ Tonkin and two anonymous reviewers We also thank T Tu forhelp with Fig 1 and S Carlson L Comte T Datry J HosenM Kondolf N Jones D Lytle E Moody J Olden M PowerJ Sabo and M Sanchez-Montoya for valuable discussionsCompeting interests The authors have no competing interestsFunding MAP was supported by funds from the NSF (1639145)and AR was supported by funds from the University of CaliforniaBerkeley and NSF (1802714)
101126scienceaaw2087
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 13 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
restorationLinkages between flow regime biota and ecosystem processes Implications for river
Margaret Palmer and Albert Ruhi
DOI 101126scienceaaw2087 (6459) eaaw2087365Science
this issue p eaaw2087Scienceflow variability and achieve hydrological connectivity between a river and its surroundingsand ecosystem processes To be successful such efforts must go beyond accounting for flood pulses to restore natural mimic ecologically important aspects of natural flow regimes guided by insights into how variations in flow affect biotawater quality and ecological processes In a Review Palmer and Ruhi explain how restoration designs now attempt to
Human activities have altered the flow regimes of many of Earths rivers with negative impacts on biodiversityRiver restoration guided by research
ARTICLE TOOLS httpsciencesciencemagorgcontent3656459eaaw2087
REFERENCES
httpsciencesciencemagorgcontent3656459eaaw2087BIBLThis article cites 147 articles 7 of which you can access for free
PERMISSIONS httpwwwsciencemagorghelpreprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAASScienceScience 1200 New York Avenue NW Washington DC 20005 The title (print ISSN 0036-8075 online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
Science No claim to original US Government WorksCopyright copy 2019 The Authors some rights reserved exclusive licensee American Association for the Advancement of
on May 27 2020
httpsciencesciencem
agorgD
ownloaded from

Restoring flow regimes ratherthan eventsScientifically informed flow designs may existthat maintain ecosystem integrity in regulatedrivers (116 117) but these have produced mixedresults to date In the dam-altered Putah Creekin California mimicking the natural timing ofhigh and low flows enhanced native fish by im-proving their spawning habitat reducing watertemperatures maintaining flowing conditionsand reducing the abundance of non-native spe-cies (118) However other similar attempts havebeen less successful In some cases non-nativefish density also benefitted from flow patternsmimicking an unregulated regime (119) in othersnon-natives bounced back shortly after the flowrelease (120) These experiences illustrate thateven natural flow features may have inadvertentnegative effects if only part of the community orecosystem is considered Models for the NavajoDam (San Juan River New Mexico) show thatadequately timed releases could simultaneouslyenhance native and minimize non-native fishpopulations (121) thus predicted benefits couldsurpass those of mimicking a natural flow re-gime potentially persisting even during dryyears Another study using the ldquodesignerrdquo flowparadigm (114) attempted to understand howdifferent facets of flow variability control fisheryproduction in the Lower Mekong floodplains Time-
series models combining discharge and fisherycatch suggested that enhanced flood pulses andlong inter-flood intervals may increase fisheryyieldsAlthough environmental flow practices have
mostly focused on re-creating discrete flowevents (eg flood pulses) these can still serve asvaluable experiments An iconic example of alarge-scale flow release is Minute 319 an amend-ment to the United StatesndashMexico treaty allow-ing opening of floodgates in the Lower ColoradoRiver and a one-time rewetting of the parchedColorado River Delta (122) Although this pulsed-flow experiment did not promote establishmentof native woody riparian seedlings such trialscan inform future restoration practices for ex-ample by determining whether the provision ofbare moist substrate enhances seed germina-tion (123) Hypothesis-driven experimental flowreleases combined with deep knowledge of tar-get ecosystems can provide insights into resto-ration actions [eg (124)] Still an outstandingquestion is how flow regime restoration couldbe leveraged to restore other important alteredaspects of the ecosystem such as sediment andthermal regimes For example cold-water releasesby existing dams in climate-vulnerable basinscould prevent organisms that thrive at low tem-peratures from becoming replaced by general-ist warm-tolerant taxa (125)
Although the concept of environmental flowsis generally associated with impounded riversits relevance to urban streams is clear (126)However because frequent high storm flowsare viewed as the problem reducing them gen-erally becomes the only restoration goal This isoften attempted by shifting the spatial distribu-tion of velocities within channels through ma-nipulation of channel sinuosity installation offlow-diverting weirs or adding other structuresThese approaches may reduce local erosion butrarely result in levels of water quality ecosystemprocesses or biodiversity in urban streams thatcome close to those of unimpacted streams(127 128) Focusing on the channel ratherthan the watershed context at large means thatkey ecosystemprocesses can remain compromised(129) A recent study compared time series ofmetabolism in two streamsmdashonewith stormwaterponds in the watershed and the other restoredby altering channel slope banks and in-streamstructural complexity (103) The study found thatpeaks in GPP and ER in the ldquorestoredrdquo streamwere higher more prone to resets induced byflashy storms and that recovery time scales werelonger compared with the stream in a watershedwith stormwater measures (Fig 6) Ponds areone type of nature-inspired green infrastructureused to restore streamflow tomore natural condi-tions tree plantings constructed wetlands andgrass swales can in turn replace gray infrastruc-ture such as concrete tunnels that capture andstore waterDistributing green infrastructure in water-
sheds helps restore streams because it targetsthe cause of streamflow alteration includinginsufficient infiltration groundwater rechargeand more generally limited watershed storagecapacity (129) By tackling the cause rather thanthe symptoms of flow alteration these practicescan reduce ecological impacts that result fromthe cascading effects of altered flows includingpoor water quality and temperature spikes Re-search on the effectiveness of green infrastructureon restoring streams is sparse and limited bylack of data before and after implementationalthough studies are beginning to appear Forexample upland green infrastructure in theform of shallow wetland-like pools that capturestormwater and reconnect it to the groundwaterhas been shown to enhance baseflows while re-ducing peak flows and pollutant loadsmore thanrestoration structures within the mainstem ofthe receiving stream (130) Similar structuresbuilt between stormwater outfalls and perennialstreams did not modulate the timing magnitudeor duration of urban storm responses nor didthey improve water quality or biodiversity (131)illustrating the need for researchers to understandhow exact design and watershed context influ-ence performance
Spatiotemporal variability andrestoration designs
Research showing how local and watershed-scale environmental drivers vary temporally andalong river networks in ways that shape biota
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 7 of 13
Flow pulses (storms) flashy flowsDroughts intermittencyLowstable flowsAltered timing seasonality
DOC N P
DetritusMicrobial
stoichiometric constraints
Excretion exudates
Detrital pathway
Algal pathway
Algal stoichiometric constraints
Organismal dispersal source-sink dynamics
Algae
Community replacement
ndash ndash +
ndash ndash + ndash
+
+ndash +
Invasions
Turnover
Consumer stoichiometric constraints
Fig 5 Flow variation affects biota and ecosystem processes and food webs integrate bothHydrologic connectivity along river networks is a first landscape-level filter constraining dispersal oforganisms in river networks Extreme flow variation (floods and droughts) are a source of disturbanceto all organisms (gray box) but particularly to the higher trophic levels given their higher metabolicdemands In turn flow variation controls inputs of nutrients and terrestrial leaf litter to the stream whichinfluences microbial activity and the relative contributions of the detrital (brown) and algal (green)pathways in sustaining aquatic consumers Not just quantity but also food quality (eg CN and CPratios) influence dietary choices of consumers and thus trophic transfer efficiency Finally alteration offlow regimes may influence all these compartments and may also open niches for new players Ifbiological invasions occur at the top of the food web these have the potential to divert energy away fromnative predators and to alter food-web structure through top-down controls
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
and ecosystem processes can help prioritizeselection of sites for restoration For exampleusing the concept of river network saturation(55) to identify river sections where in-streamremoval of nutrients cannot keep up with highsource inputs of nutrients suggests that restorationactions targeting those sections may dispropor-tionately improve water quality Identification ofsubcatchments that disproportionately influencestream chemistry may also be informed by com-bining methods from landscape ecology andcatchment hydrology focusing on the spatialscale of variance in water chemistry (132) Head-waters are often prioritized for restoration be-
cause poor water quality in them can havedownstream impacts Research showing thatlocal environmental constraints on river com-munities are important in headwaters as well asin the larger channels but dispersal constraintsare much less important in the former (133 134)implies that headwaters should be prioritizedHowever research quantifying time-varying con-tributions of local communities to river-wide(network) biodiversity can also inform prioritiza-tion by identifying sites that host a dispropor-tionate share of species or that host uniquespeciesmdashfor example because they shelter sensi-tive taxa during critical flow bottlenecks This idea
encapsulates the concept of keystone habitatsor local sites that strongly influence metacom-munity dynamics (135) This has been appliedto aquatic invertebrates across intermittent rivernetworks by tracking time-varying site-specificcontributions to community dissimilarity (or betadiversity) (136)Other approaches to prioritize restoration ac-
tions may be based on contributions to system-wide stability rather than numbers of individualsor species For example the restoration of fall-run Chinook salmon portfolios may be assessedvia the contributions that different periods andlocationsmake tometapopulation-wide stabilityowing to spatiotemporal variation in habitatconditions (137) Additionally interventionscould promote conditions that are not impor-tant now but could be important in the future[ie proto-refugia (138)] For example resto-ration of groundwater-influenced habitats couldmitigate future stress to drought-sensitive taxaunder scenarios of increased intermittencyPatches of undisturbed land that protect flowregimes in streams within a larger catchmentthat is otherwise urbanized can be sources ofcolonists to restored sites Priorities that focuson present degradation of river sectionsmay failto address site contribution to impairment orrecovery at broader scales or under differentscenarios These insights indicate a great poten-tial for new methods to prioritize restoration ofboth urban and regulated streamsWe know little about how altered flow regimes
propagate through entire river networks butwork is starting to assess the cumulative effectsof dams on flow regimes as well as dam-levelcontributions to alteration Although these ap-proaches have largely focused on dam cascades[eg (139)] if expanded they could help identifyfavorable sections in river networks to developenvironmental flow operations Here exper-imentation could again help with testing ofnew strategies For instance large-scale flowexperiments could be designed that coordinatereleases among dams or between dams and free-flowing tributaries to enhance the restorativeeffects of natural pulses (140)
Hydrologic connectivityinfluences restoration
Recovering critical flow paths and their connec-tivity to rivers is important but it remains asubstantial challenge for flow-degraded riversand streams Whereas regulated rivers tend tohave reduced lateral and longitudinal hydro-logic connectivity urban streams have unnaturallevels of connectivity because pipes and over-land runoff route water quickly In both cases fun-damental hydroecological insights into streambedstructure landscape storage capacity and ground-water connectivity can help to envision novel re-storation approaches (130 141 142) More generallyhydrologic connectivity is becoming a frameworkto identify alternative ways to restore streams viastormwater management (143) For example arecent study suggested that reducing road densityand hydrologic connectivity between roads
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 8 of 13
ER
Disc
harg
e (f
t3 s)
GPPG
PP o
r ER
(g O
2 m-2
day
-1)
Discharge
Watershed BMPs
In-stream restoration
0
ndash4
4
0
ndash4
4
4
2
6
4
2
6
Jan Sept July May Mar Nov Jan
A
B C
Fig 6 Green infrastructure as a means to restore stream ecosystems (A) Discharge andecosystem respiration (ER more-negative values mean higher respiration) and gross primaryproduction (GPP) for two urban streams in which restoration efforts involved (top) modifications inthe stream channel and (bottom) traditional stormwater best management practices (BMPs)[Reproduced with permission from (103)] (B and C) Example of a comprehensive greeninfrastructure approach to ecologically restore an urban headwater stream that also supports thelifestyle of resident urban communities (B) Stream at the onset of restoration and (C) afterrestoration [Reproduced with permission from Greenworks Portland OR]
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 9 of 13
Table 1 Research challenges and opportunities Despite the progress that has been made in understanding how flow regimes affect biota and ecosystem
processes major challenges persist that prevent a complete understanding of the flow-biota-ecosystem processes nexus Making progress on these
challenges requires a mix of fundamental and applied research
Topic Research challenge Opportunity
Connecting
organismal to
ecosystem-level
processes
Research on the effects of flow on organisms
and ecosystem processes has followed parallel
tracksmdashhindering our mechanistic
understanding of the flow-biota-ecosystem
processes nexus
Generate theory that connects concepts and frameworks
that are well accepted in river community and ecosystem
ecology For example the notion of environmental filters
that connect regional to local composition via a combination
of dispersal and environmental conditions (community
ecology) with analogous theory on solute transport versus
reactivity (ecosystem ecology)
Our current understanding of the links between
phenology of biota and ecosystem processes
is precursory
Use high-frequency sensors to characterize metabolic
regimes at sites with ongoing organismal monitoring
Leverage citizen-science biodiversity data to detect
spatiotemporal shifts in animal phenology
(eg insect emergence fish spawning)
Use remote sensing tools to better understand
links between hydrology and aquatic and riparian
plant phenology
The link between stoichiometry and flow
variation remains largely unexplored
Repurpose data that is already being collected to test
flow-stoichiometry relationships eg studies using C and
N stable isotopes to describe CN ratios across flow
regime types Perform field experiments to quantify
indirect effects of invertebrate and fish consumption
on ecosystem metabolism
Ecological
consequences of
hydrologic
connectivity
The importance of anaerobic metabolism in
carbon processing as flows cease and parts of
streambeds dry is a major frontier research area
Create networks of sites across different geologies
and flow regimes that collect time-series data on
methane nitrous oxide and carbon dioxide fluxes
from intermittent streams
We ignore how biota in the wet phase is
influenced by processes and patterns
from the dry phase
Research the functions of dry riverbeds Use camera traps and
drone technology to investigate the role of dry riverbeds
as a dispersal corridor metabarcoding to quantify
ldquodark biodiversityrdquo in the hyporheic
Wetlands and their links to stream networks are poorly
delineated globally Even in the United States and
Europe current wetland inventories do not capture
these links yet understanding how their connectivity
to river systems influences network-scale ecosystem
processes is needed
Analyze time series from high-frequency sensors distributed in
river networks varying in wetland connectivity Further
development and application of time-series methods
used by neuroscientists
We need to identify the relationship between
varying flows and solute dynamics
associated with changes in catchment-scale
hydrologic exchanges
Design collaborative work among catchment hydrologists
and stream ecologists combining methods to estimate
transit times residence times and identify sources
across heterogeneous landscapes Link these properties to
land management and water quality aspects
Scaling and
transferring flow-
ecology relationships
across space and time
We ignore how biodiversity responds to
propagation of flow regime alteration
across river networks
Leverage extensive hydrometric networks (eg United States
Geological Survey National Water Information System)
to perform spatially replicated time-series analyses
on flow and use these to generate expectations
on how river network position should mediate
biodiversity responses to flow alteration
Flow-ecology relationships do not account for
nonstationarity and abrupt change
Use process-based models that account for mechanisms
and associated uncertainties When using time-series
data and models select methods that are robust
to nonstationarity Linear flow-ecology relationships
should not be assumed
Current flow-ecology relationships assume that
responses of species do not change over time
Consider flow regime alteration as a process with ecological and
evolutionary consequences Include intraspecific variation
in traits and potential for rapid evolution in models
continued on next page
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
and streams may improve water quality morethan other restoration options (70) Additionallyresearch on the age and lability of DOC enteringstreams has shown how precipitation or land dis-turbance can change watershed flow paths (53)and this could inspire new restoration approachesfocused on land use and soil properties Manystudies on hydrologic connectivity and networkstructure suggest a need for restoration to focuson landscape heterogeneity especially on thepresence ofwetlands lakes and ponds (144) Suchfeaturesare frequently lost inurbanizedcatchmentsand outcomes could be improved by restoring orincreasing connectivitywith these ecosystems (145)Hydrologic connectivity associated with ground-water inputs or abstractions land-use manage-ment and return flows fromwastewater treatmentplants also need to be considered in these initia-tives (146)Restoring hydrologic connectivity is also key
to achieving biodiversity outcomesmdashparticularlyin dammed systems Longitudinal connectivityfor fish inhabiting dam-altered rivers can beenhanced by the construction of fishways andsophisticated passage facilities However moni-toring data on the effectiveness of such inter-
ventions are still scarce and available evidencesuggests that passage effectivity largely dependson technology used and species-specific fish be-havior (147 148) Although trap-and-transportprograms for migratory fishes can help augmentthreatened populations (149) these practicesoften increase straying and thus the potentialfor genetic homogenization as seen from ju-venile salmon collected at dams and transporteddownstream to increase out-migration success(150) Overall a combination of engineering andmanagement solutions may be best to restorefunctional connectivity in dam-altered riversThe link between hydrologic connectivity andecological restoration is also central to con-siderations on where to build remove and re-operate hydrologic infrastructure Hundredsof dams have been intentionally removed sincethe 1970s and removal of large dams such asthe Elwha River Dam (Washington USA) hasshown that restoring longitudinal connectivitycan have immediate geomorphic effects with com-plex temporal dynamics (151) Infrastructure canalso be designed or reoperated to manage forrestoring lateral connectivity and enhanced flood-plain processes For example the Yolo Bypass in
the Sacramento River California has becomea prime rearing and migration site for chinooksalmon showing that flood control and habitatfor fish and wildlife do not need to be at odds(152) All these examples show the importanceof integrating hydrologic connectivity in ecolog-ical restoration whether the goal is to minimizethe ecological effects of fragmentation restorefree-flowing watercourses or operate green infra-structure to enhance infiltration floodplain con-nectivity or other ecological processes
Research challenges and opportunities
Two decades of concerted efforts to research theeffects of flow regime alteration have tremen-dously advanced our understanding of the waysin which streamflow influences biota and eco-system processes However important challengesremain These aremainly related to the scarcity ofscience connecting organismal to ecosystem-levelprocesses through for example microbial linksthe difficulty of studying watershed-level phys-ical and biogeochemical heterogeneity and itsinfluence on riverine processes and biota thecomplexity of parsing out the direct effects offlow from flow-mediated effects and co-occurring
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 10 of 13
Topic Research challenge Opportunity
Cascading
implications of
hydrologic
alteration
We have a poor understanding of the
interactions among physical drivers
Build on large-scale flow experiments to test hypotheses on the
combined effects of altered flow sediment and thermal
regimes Experiment with multilevel intakes in reservoirs
Promote collection of non-flow variables (eg temperature
water quality parameters) at hydrometric stations
At low flows biota are being exposed to high
concentrations of new classes of pollutants
whose impacts are poorly known especially if
biotic impacts interact with impacts on
ecosystem processes
Design realistic experimental studies to advance knowledge
on the ecological effects of multiple stressors
Collect long-term data at sites varying in contaminant
inputs and hydrologic regimes Link microbial to
organismal and ecosystem-level responses
Using hydroecological
science to inform
river ecosystem
restoration
Environmental flow science largely ignores
human dimensions
Use methods that allow identifying causal networks
in complex socioenvironmental systems Include
human needs and thresholds in research on
flow-ecology relationships
Environmental flows are generally prescribed
at the scale of single infrastructures restoration
designs and assessment are generally at reach scales
Promote coordinated large-scale flow experiments
and restoration designs at the river basin scale
Extend assessment of green infrastructure
effectiveness beyond individual structures to
infrastructure networks and link both
to river outcomes
Environmental flows in impounded rivers
generally target the conservation of some
particular species rather than outcomes at the
food-web or ecosystem level
Advance fundamental research on microbialndash
ecosystem process linkages Expand the notion
of designer flows to food-web structure and
ecosystem services when possible
Environmental flow science has largely
ignored surface waterndashgroundwater
interactions
Expand science to understand the link between
restoration actions groundwater recharge and
inputs to streams Provide science relevant to
the implementation of new legislation on
groundwater management (eg Sustainable
Groundwater Management Act in California)
Uncertainty regarding future
hydroclimate and
river flows is growing
Refine regional climate models Promote research on
the interaction between land-use change and
evapotranspiration Incorporate groundwater
influence in macroscale hydrology models
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
stressors (particularly in urban streams) and thechallenge of predicting outcomeswhen both thephysical and the biological environments arechangingAchieving a predictive understanding of hy-
droecological relationships is essential to design-ing successful self-sustaining restoration actionsin river ecosystems However translating basichydroecological insights into restoration practicerequires understanding the complexities of work-ing within a human-dominated water cycle Al-though it is rarely possible to restore all facetsof a natural flow regime ecologically informeddesigns may help mimic key aspects of the nat-ural flow regime (eg periodic pulse flows) orcreate regimes that are not natural but maximizeoutcomes given socioeconomic constraints InTable 1 we identify persisting and emergingchallenges in hydroecological research We paireach challenge with an opportunity (related toresearch design technology or implementation)We contend that these challenges should beprioritized to advance process-based river eco-system restoration
REFERENCES AND NOTES
1 N L Poff et al The natural flow regime Bioscience 47769ndash784 (1997) doi 1023071313099
2 D A Lytle N L Poff Adaptation to natural flow regimesTrends Ecol Evol 19 94ndash100 (2004) doi 101016jtree200310002 pmid 16701235
3 S Sabater A Elosegi R Ludwig in Multiple Stressors in RiverEcosystems (Elsevier 2019) pp 1ndash22
4 United Nations New UN Decade on Ecosystem Restorationoffers unparalleled opportunity for job creation food securityand addressing climate change (2019) httpswwwunenvironmentorgnews-and-storiespress-releasenew-un-decade-ecosystem-restoration-offers-unparalleled-opportunity
5 A W Lorenz P Haase K Januschke A SundermannD Hering Revisiting restored river reaches - Assessingchange of aquatic and riparian communities afterfive years Sci Total Environ 613-614 1185ndash1195(2018) doi 101016jscitotenv201709188pmid 28954379
6 M A Palmer K L Hondula B J Koch Ecological Restorationof Streams and Rivers Shifting Strategies and ShiftingGoals (2014) vol 45
7 T J Beechie et al Process-based principles for restoringriver ecosystems Bioscience 60 209ndash222 (2010)doi 101525bio20106037
8 S E Bunn A H Arthington Basic principles and ecologicalconsequences of altered flow regimes for aquaticbiodiversity Environ Manage 30 492ndash507 (2002)doi 101007s00267-002-2737-0 pmid 12481916
9 K D Fausch Y Taniguchi S Nakano G D GrossmanC R Townsend Flood disturbance regimes influence rainbowtrout invasion success among five holarctic regions EcolAppl 11 1438ndash1455 (2001) doi 1018901051-0761(2001)011[1438FDRIRT]20CO2
10 M E Power M S Parker W E Dietrich Seasonalreassembly of a river food web Floods droughts andimpacts of fish Ecol Monogr 78 263ndash282 (2008)doi 10189006-09021
11 V Acuntildea A Giorgi I Muntildeoz U R S Uehlinger S SabaterFlow extremes and benthic organic matter shape themetabolism of a headwater Mediterranean streamFreshwater Biol 49 960ndash971 (2004) doi 101111j1365-2427200401239x
12 K Tockner F Malard J V Ward An extension of the floodpulse concept Hydrol Processes 14 2861ndash2883 (2000)doi 1010021099-1085(20001112)141617lt2861AID-HYP124gt30CO2-F
13 J S Perkin et al Fragmentation and dewatering transformGreat Plains stream fish communities Ecol Monogr 8573ndash92 (2015) doi 10189014-01211
14 J L Sabo D M Post Quantifying periodic stochastic andcatastrophic environmental variation Ecol Monogr 7819ndash40 (2008) doi 10189006-13401
15 F Shuai et al Silver carp larva abundance in response toriver flow rate revealed by cross-wavelet modellingEcol Modell 383 98ndash105 (2018) doi 101016jecolmodel201805020
16 J D Tonkin M T Bogan N Bonada B Rios-ToumaD A Lytle Seasonality and predictability shape temporalspecies diversity Ecology 98 1201ndash1216 (2017)doi 101002ecy1761 pmid 28144975
17 N L Poff Beyond the natural flow regime Broadening thehydro‐ecological foundation to meet environmental flowschallenges in a non‐stationary world Freshwater Biol 631011ndash1021 (2018) doi 101111fwb13038
18 A Ruhi X Dong C H McDaniel D P Batzer J L SaboDetrimental effects of a novel flow regime on the functionaltrajectory of an aquatic invertebrate metacommunity GlobChange Biol 24 3749ndash3765 (2018) doi 101111gcb14133pmid 29665147
19 M T Bogan D A Lytle Severe drought drives novelcommunity trajectories in desert stream pools FreshwaterBiol 56 2070ndash2081 (2011) doi 101111j1365-2427201102638x
20 G Woodward et al The effects of climatic fluctuations andextreme events on running water ecosystems Philos TransR Soc London Ser B 371 20150274 (2016) pmid 27114576
21 M A Palmer C M Swan K Nelson P Silver R AlvestadStreambed landscapes Evidence that stream invertebratesrespond to the type and spatial arrangement of patchesLandsc Ecol 15 563ndash576 (2000) doi 101023A1008194130695
22 D Schmera et al Does isolation influence the relative role ofenvironmental and dispersal‐related processes in streamnetworks An empirical test of the network positionhypothesis using multiple taxa Freshwater Biol 63 74ndash85(2018) doi 101111fwb12973
23 R Henriques‐Silva et al A comprehensive examination of thenetwork position hypothesis across multiple rivermetacommunities Ecography 42 284ndash294 (2019)doi 101111ecog03908
24 K L Jaeger J D Olden N A Pelland Climate change poisedto threaten hydrologic connectivity and endemic fishes indryland streams Proc Natl Acad Sci USA 11113894ndash13899 (2014) doi 101073pnas1320890111pmid 25136090
25 S Garbin E A Celegon P Fanton G Botter Hydrologicalcontrols on river network connectivity R Soc Open Sci 6181428 (2019) doi 101098rsos181428 pmid 30891270
26 F Cattaneacuteo B Hugueny N Lamouroux Synchrony in browntrout Salmo trutta population dynamics A lsquoMoran effectrsquo onearly‐life stages Oikos 100 43ndash54 (2003) doi 101034j1600-0706200311912x
27 A Terui et al Metapopulation stability in branching rivernetworks Proc Natl Acad Sci USA 115 E5963ndashE5969(2018) doi 101073pnas1800060115 pmid 29895695
28 P A P Moran The statistical analysis of the Canadian Lynxcycle Aust J Zool 1 291ndash298 (1953) doi 101071ZO9530291
29 G D Grossman P B Moyle J O Whitaker Jr Stochasticityin structural and functional characteristics of an Indianastream fish assemblage A test of community theoryAm Nat 120 423ndash454 (1982) doi 101086284004
30 D A Lytle D M Merritt J D Tonkin J D OldenL V Reynolds Linking river flow regimes to riparian plantguilds A community-wide modeling approach Ecol Appl 271338ndash1350 (2017) doi 101002eap1528 pmid 28263426
31 A Ruhiacute E E Holmes J N Rinne J L Sabo Anomalousdroughts not invasion decrease persistence of native fishesin a desert river Glob Change Biol 21 1482ndash1496 (2015)doi 101111gcb12780 pmid 25430731
32 J D Tonkin D M Merritt J D Olden L V ReynoldsD A Lytle Flow regime alteration degrades ecologicalnetworks in riparian ecosystems Nat Ecol Evol 2 86ndash93(2018) doi 101038s41559-017-0379-0 pmid 29180707
33 M C Mims J D Olden Fish assemblages respond toaltered flow regimes via ecological filtering of lifehistory strategies Freshwater Biol 58 50ndash62 (2013)doi 101111fwb12037
34 P T J Johnson J D Olden M J Vander Zanden Daminvaders Impoundments facilitate biological invasions intofreshwaters Front Ecol Environ 6 357ndash363 (2008)doi 101890070156
35 N Bonada S Doledec B Statzner Taxonomic and biologicaltrait differences of stream macroinvertebrate communitiesbetween mediterranean and temperate regions Implicationsfor future climatic scenarios Glob Change Biol 131658ndash1671 (2007) doi 101111j1365-2486200701375x
36 M I Pyne N L Poff Vulnerability of stream communitycomposition and function to projected thermal warming andhydrologic change across ecoregions in the western UnitedStates Glob Change Biol 23 77ndash93 (2017) doi 101111gcb13437 pmid 27429092
37 D B Herbst S D Cooper R B Medhurst S W WisemanC T Hunsaker Drought ecohydrology alters the structureand function of benthic invertebrate communities inmountain streams Freshwater Biol 64 886ndash902 (2019)doi 101111fwb13270
38 M T Bogan E T Chester T Datry A L MurphyB J Robson A Ruhi R Stubbington J E Whitney inResistance Resilience and Community Recovery in IntermittentRivers and Ephemeral Streams T Datry N Bonada A BoultonEds (Academic Press 2017) pp 349ndash376
39 S Houmlckendorff et al Characterizing fish responses to a riverrestoration over 21 years based on speciesrsquo traits ConservBiol 31 1098ndash1108 (2017) doi 101111cobi12908pmid 28218807
40 D Bruno et al Structural and functional responses ofinvertebrate communities to climate change andflow regulation in alpine catchments Glob Change Biol25 1612ndash1628 (2019) doi 101111gcb14581pmid 30698905
41 T W H Aspin et al Extreme drought pushes streaminvertebrate communities over functional thresholds GlobChange Biol 25 230ndash244 (2019) doi 101111gcb14495pmid 30346098
42 S E Tank J B Fellman E Hood E S Kritzberg Beyondrespiration Controls on lateral carbon fluxes across theterrestrial-aquatic interface Limnol Oceanogr Lett 3 76ndash88(2018) doi 101002lol210065
43 A P Appling et al The metabolic regimes of 356 rivers inthe United States Sci Data 5 180292 (2018) doi 101038sdata2018292 pmid 30532078
44 P Savoy et al Metabolic rhythms in flowing watersAn approach for classifying river productivity regimesLimnol Oceanogr (2019) doi 101002lno11154
45 E S Bernhardt et al The metabolic regimes of flowingwaters Limnol Oceanogr 63 S99ndashS118 (2018)doi 101002lno10726
46 M Colls X Timoner S Sabater V Acuntildea Effects of durationand frequency of the non-flow period on stream metabolismEcosystems 101007s10021-019-00345-1 (2019)doi 101007s10021-019-00345-1
47 A J Ulseth E Bertuzzo G A Singer J Schelker T J BattinClimate-induced changes in spring snowmelt impactecosystem metabolism and carbon fluxes in an Alpine streamnetwork Ecosystems 21 373ndash390 (2018) doi 101007s10021-017-0155-7
48 B O L Demars Hydrological pulses and burning of dissolvedorganic carbon by stream respiration Limnol Oceanogr 64406ndash421 (2019) doi 101002lno11048
49 R T Hensley L Kirk M Spangler M N Gooseff M J CohenFlow extremes as spatiotemporal control points on riversolute fluxes and metabolism J Geophys Res Biogeosci124 537ndash555 (2019) doi 1010292018JG004738
50 J D Hosen et al Enhancement of primary production duringdrought in a temperate watershed is greater in larger riversthan headwater streams Limnol Oceanogr 64 1458ndash1472(2019) doi 101002lno11127
51 L Goacutemez-Gener et al When water vanishes Magnitude andregulation of carbon dioxide emissions from dry temporarystreams Ecosystems 19 710ndash723 (2016) doi 101007s10021-016-9963-4
52 E H Stanley et al The ecology of methane in streams andrivers Patterns controls and global significance EcolMonogr 86 146ndash171 (2016) doi 10189015-1027
53 R T Barnes D E Butman H F Wilson P A RaymondRiverine export of aged carbon driven by flow path depth andresidence time Environ Sci Technol 52 1028ndash1035 (2018)doi 101021acsest7b04717 pmid 29313674
54 G Pinay et al Riparian corridors A new conceptualframework for assessing nitrogen buffering across biomesFront Environ Sci 6 47 (2018) doi 103389fenvs201800047
55 W M Wollheim et al River network saturation conceptFactors influencing the balance of biogeochemical supply and
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 11 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
demand of river networks Biogeochemistry 141 503ndash521(2018) doi 101007s10533-018-0488-0
56 P A Raymond J E Saiers W V Sobczak Hydrological andbiogeochemical controls on watershed dissolved organicmatter transport Pulse-shunt concept Ecology 97 5ndash16(2016) doi 10189014-16841 pmid 27008769
57 C Tunaley D Tetzlaff J Lessels C Soulsby Linking high-frequency DOC dynamics to the age of connected watersources Water Resour Res 52 5232ndash5247 (2016)doi 1010022015WR018419
58 S Bernal et al Supply demand and in-stream retention ofdissolved organic carbon and nitrate during storms inMediterranean forested headwater streams Front EnvironSci 7 60 (2019) doi 103389fenvs201900060
59 J Harvey et al How hydrologic connectivity regulates waterquality in river corridors J Am Water Resour Assoc 55369ndash381 (2019) doi 1011111752-168812691
60 J C Moore et al Detritus trophic dynamics and biodiversityEcol Lett 7 584ndash600 (2004) doi 101111j1461-0248200400606x
61 A D Rosemond et al Freshwater ecology Experimentalnutrient additions accelerate terrestrial carbon loss fromstream ecosystems Science 347 1142ndash1145 (2015)doi 101126scienceaaa1958 pmid 25745171
62 A C Y Yeung J L Musetta-Lambert D P KreutzweiserP K Sibley J S Richardson Relations of interannualdifferences in stream litter breakdown with dischargeBioassessment implications Ecosphere 9 e02423 (2018)doi 101002ecs22423
63 R M Northington J R Webster Experimental reductions instream flow alter litter processing and consumer subsidies inheadwater streams Freshwater Biol 62 737ndash750 (2017)doi 101111fwb12898
64 E Mas-Martiacute I Sanpera-Calbet I Muntildeoz Bottom-up effectsof streambed drying on consumer performance throughchanges in resource quality Aquat Sci 79 719ndash731 (2017)doi 101007s00027-017-0531-6
65 B D Richter G A Thomas Restoring environmental flows bymodifying dam operations Ecol Soc 12 12 (2007)doi 105751ES-02014-120112
66 G Zolezzi A Siviglia M Toffolon B MaioliniThermopeaking in Alpine streams Event characterizationand time scales Ecohydrology 4 564ndash576 (2011)doi 101002eco132
67 T Meiszligner B Sures C K Feld Multiple stressors and therole of hydrology on benthic invertebrates in mountainousstreams Sci Total Environ 663 841ndash851 (2019)doi 101016jscitotenv201901288 pmid 30738264
68 I R Waite et al Effects of urban multi-stressors on threestream biotic assemblages Sci Total Environ 6601472ndash1485 (2019) doi 101016jscitotenv201812240pmid 30743940
69 K C Nelson M A Palmer Stream temperature surges underurbanization and climate change Data models andresponses J Am Water Resour Assoc 43 440ndash452 (2007)doi 101111j1752-1688200700034x
70 J R Blaszczak J M Delesantro Y Zhong D L UrbanE S Bernhardt Watershed urban development controls onurban streamwater chemistry variability Biogeochemistry144 61ndash84 (2019) doi 101007s10533-019-00572-7
71 L Ponsatiacute et al Biofilm responses to flow regulation by damsin Mediterranean rivers River Res Appl 31 1003ndash1016(2015) doi 101002rra2807
72 J M Alberts J J Beaulieu I Buffam Watershed land useand seasonal variation constrain the influence of ripariancanopy cover on stream ecosystem metabolism Ecosystems20 553ndash567 (2017) doi 101007s10021-016-0040-9pmid 30416371
73 R O Hall Jr et al Turbidity light temperature andhydropeaking control primary productivity in the ColoradoRiver Grand Canyon Limnol Oceanogr 60 512ndash526 (2015)doi 101002lno10031
74 J R Blaszczak J M Delesantro D L Urban M W DoyleE S Bernhardt Scoured or suffocated Urban streamecosystems oscillate between hydrologic and dissolvedoxygen extremes Limnol Oceanogr 64 877ndash894 (2019)doi 101002lno11081
75 J-R Mor et al Dam regulation and riverine food-webstructure in a Mediterranean river Sci Total Environ 625301ndash310 (2018) doi 101016jscitotenv201712296pmid 29289778
76 H A Wellard Kelly et al Macroinvertebrate diets reflecttributary inputs and turbidity-driven changes in food
availability in the Colorado River downstream of GlenCanyon Dam Freshw Sci 32 397ndash410 (2013)doi 10189912-0881
77 S J Imberger R M Thompson M R Grace Urbancatchment hydrology overwhelms reach scale effects ofriparian vegetation on organic matter dynamics FreshwaterBiol 56 1370ndash1389 (2011) doi 101111j1365-2427201102575x
78 R T Martins A S Melo J F Gonccedilalves Jr N HamadaLeaf-litter breakdown in urban streams of Central AmazoniaDirect and indirect effects of physical chemical andbiological factors Freshw Sci 34 716ndash726 (2015)doi 101086681086
79 C M Yule J Y Gan T Jinggut K V Lee Urbanizationaffects food webs and leaf-litter decomposition in a tropicalstream in Malaysia Freshw Sci 34 702ndash715 (2015)doi 101086681252
80 T A Newcomer et al Influence of natural and novel organiccarbon sources on denitrification in forest degraded urbanand restored streams Ecol Monogr 82 449ndash466 (2012)doi 10189012-04581
81 A Chaudhary I Kauser A Ray R Poretsky Taxon-drivenfunctional shifts associated with storm flow in an urbanstream microbial community MSphere 3 e00194-18 (2018)doi 101128mSphere00194-18 pmid 29976643
82 T A Kennedy et al Flow management for hydropowerextirpates aquatic insects undermining river food websBioscience 66 561ndash575 (2016) doi 101093bioscibiw059
83 A Glazaczow D Orwin M Bogdziewicz Increasedtemperature delays the late-season phenology of multivoltineinsect Sci Rep 6 38022 (2016) doi 101038srep38022pmid 27905493
84 S G Fisher L J Gray N B Grimm D E Busch Temporalsuccession in a desert stream ecosystem following flashflooding Ecol Monogr 52 93ndash110 (1982) doi 1023072937346
85 G J Scrimgeour M J Winterbourn Effects of floods onepilithon and benthic macroinvertebrate populations in anunstable New Zealand river Hydrobiologia 171 33ndash44 (1989)doi 101007BF00005722
86 E Bondar-Kunze S Maier D Schoumlnauer N Bahl T HeinAntagonistic and synergistic effects on a stream periphytoncommunity under the influence of pulsed flow velocityincrease and nutrient enrichment Sci Total Environ 573594ndash602 (2016) doi 101016jscitotenv201608158pmid 27585428
87 M J Cashman G L Harvey G Wharton M C Bruno Woodmitigates the effect of hydropeaking scour on periphytonbiomass and nutritional quality in semi-natural flumesimulations Aquat Sci 79 459ndash471 (2017) doi 101007s00027-016-0510-3
88 W F Cross et al Food-web dynamics in a large riverdiscontinuum Ecol Monogr 83 311ndash337 (2013)doi 10189012-17271
89 D M Warfe et al Productivity disturbance and ecosystemsize have no influence on food chain length in seasonallyconnected rivers PLOS ONE 8 e66240 (2013)doi 101371journalpone0066240
90 T D Jardine et al Consumer-resource coupling in wet-drytropical rivers J Anim Ecol 81 310ndash322 (2012)doi 101111j1365-2656201101925x pmid 22103689
91 K O Winemiller et al Pulsing hydrology determines top‐down control of basal resources in a tropical riverndashfloodplainecosystem Ecol Monogr 84 621ndash635 (2014)doi 10189013-18221
92 D M Post The long and short of food-chain length TrendsEcol Evol 17 269ndash277 (2002) doi 101016S0169-5347(02)02455-2
93 D M Post G Takimoto Proximate structural mechanismsfor variation in food‐chain length Oikos 116 775ndash782 (2007)doi 101111j0030-1299200715552x
94 P A McHugh A R McIntosh P G Jellyman Dual influencesof ecosystem size and disturbance on food chain length instreams Ecol Lett 13 881ndash890 (2010) doi 101111j1461-0248201001484x pmid 20482579
95 A Ruhiacute et al Flow regulation increases food-chain lengththrough omnivory mechanisms in a Mediterranean rivernetwork Freshwater Biol 61 1536ndash1549 (2016) doi 101111fwb12794
96 J L Sabo J C Finlay T Kennedy D M Post The role ofdischarge variation in scaling of drainage area and food chainlength in rivers Science 330 965ndash967 (2010) doi 101126science1196005 pmid 20947729
97 A R McIntosh et al Capacity to support predators scaleswith habitat size Sci Adv 4 eaap7523 (2018) doi 101126sciadvaap7523 pmid 29978038
98 J H Brown J F Gillooly A P Allen V M Savage G B WestToward a metabolic theory of ecology Ecology 851771ndash1789 (2004) doi 10189003-9000
99 K Zou E Theacutebault G Lacroix S Barot Interactions betweenthe green and brown food web determine ecosystemfunctioning Funct Ecol 30 1454ndash1465 (2016) doi 1011111365-243512626
100 M J Vanni P B McIntyre Predicting nutrient excretion ofaquatic animals with metabolic ecology and ecologicalstoichiometry A global synthesis Ecology 97 3460ndash3471(2016) doi 101002ecy1582 pmid 27912023
101 T B Parr K A Capps S P Inamdar K A Metcalf Animal-mediated organic matter transformation Aquatic insects as asource of microbially bioavailable organic nutrients andenergy Funct Ecol 33 524ndash535 (2019) doi 1011111365-243513242
102 C P Arango et al Urban infrastructure influences dissolvedorganic matter quality and bacterial metabolism in an urbanstream network Freshwater Biol 62 1917ndash1928 (2017)
103 L G Larsen J W Harvey Disrupted carbon cycling inrestored and unrestored urban streams Critical timescalesand controls Limnol Oceanogr 62 S160ndashS182 (2017)doi 101002lno10613
104 H M Halvorson G White J T Scott M A Evans-WhiteDietary and taxonomic controls on incorporation of microbialcarbon and phosphorus by detritivorous caddisfliesOecologia 180 567ndash579 (2016) doi 101007s00442-015-3464-6 pmid 26497125
105 J J Elser et al Biological stoichiometry from genes toecosystems Ecol Lett 3 540ndash550 (2000) doi 101046j1461-0248200000185x
106 E K Moody et al Consumption explains intraspecificvariation in nutrient recycling stoichiometry in a desert fishEcology 99 1552ndash1561 (2018) doi 101002ecy2372pmid 29882955
107 N Welti et al Bridging food webs ecosystem metabolismand biogeochemistry using ecological stoichiometry theoryFront Microbiol 8 1298 (2017) doi 103389fmicb201701298 pmid 28747904
108 M A Palmer S Filoso R M Fanelli From ecosystems toecosystem services Stream restoration as ecologicalengineering Ecol Eng 65 62ndash70 (2014) doi 101016jecoleng201307059
109 M J Burns T D Fletcher C J Walsh A R LadsonB E Hatt Hydrologic shortcomings of conventional urbanstormwater management and opportunities for reformLandsc Urban Plan 105 230ndash240 (2012) doi 101016jlandurbplan201112012
110 C R Liermann C Nilsson J Robertson R Y NgImplications of dam obstruction for global freshwater fishdiversity Bioscience 62 539ndash548 (2012) doi 101525bio20126265
111 J D Olden R J Naiman Incorporating thermal regimes intoenvironmental flows assessments Modifying dam operationsto restore freshwater ecosystem integrity Freshwater Biol55 86ndash107 (2010) doi 101111j1365-2427200902179x
112 D B Osmundson R J Ryel V L Lamarra J Pitlick Flowndashsedimentndashbiota relations Implications for river regulationeffects on native fish abundance Ecol Appl 12 1719ndash1739(2002)
113 D von Schiller et al Regulation causes nitrogen cyclingdiscontinuities in Mediterranean rivers Sci Total Environ540 168ndash177 (2016) doi 101016jscitotenv201507017pmid 26233271
114 J L Sabo et al Designing river flows to improve foodsecurity futures in the Lower Mekong Basin Science 358eaao1053 (2017) doi 101126scienceaao1053pmid 29217541
115 W Xu et al Improving the multi-objective performance ofrainwater harvesting systems using real-time controltechnology Water 10 147 (2018) doi 103390w10020147
116 M Acreman et al Environmental flows for naturalhybrid and novel riverine ecosystems in a changingworld Front Ecol Environ 12 466ndash473 (2014)doi 101890130134
117 N L Poff et al Sustainable water management under futureuncertainty with eco-engineering decision scaling Nat ClimChang 6 25ndash34 (2016) doi 101038nclimate2765
118 J D Kiernan P B Moyle P K Crain Restoring native fishassemblages to a regulated California stream using the
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 12 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
natural flow regime concept Ecol Appl 22 1472ndash1482(2012) doi 10189011-04801 pmid 22908707
119 D L Propst K B Gido Responses of native andnonnative fishes to natural flow regime mimicry in theSan Juan River Trans Am Fish Soc 133 922ndash931 (2004)doi 101577T03-0571
120 T K Pool J D Olden Assessing long‐term fish responses andshort‐term solutions to flow regulation in a dryland river basinEcol Freshwat Fish 24 56ndash66 (2015) doi 101111eff12125
121 W Chen J D Olden Designing flows to resolve human andenvironmental water needs in a dam-regulated riverNat Commun 8 2158 (2017) doi 101038s41467-017-02226-4 pmid 29255194
122 S M Nelson et al A history of the 2014 Minute 319environmental pulse flow as documented by fieldmeasurements and satellite imagery Ecol Eng 106733ndash748 (2017) doi 101016jecoleng201610040
123 P B Shafroth et al A large-scale environmental flowexperiment for riparian restoration in the Colorado RiverDelta Ecol Eng 106 645ndash660 (2017) doi 101016jecoleng201702016
124 E P Glenn P L Nagler P B Shafroth C J JarchowEffectiveness of environmental flows for riparian restorationin arid regions A tale of four rivers Ecol Eng 106 695ndash703(2017) doi 101016jecoleng201701009
125 S Hotaling D S Finn J Joseph Giersch D W WeisrockD Jacobsen Climate change and alpine stream biologyProgress challenges and opportunities for the futureBiol Rev Camb Philos Soc 92 2024ndash2045 (2017)doi 101111brv12319 pmid 28105701
126 C J Walsh T D Fletcher M J Burns Urban stormwaterrunoff A new class of environmental flow problem PLOS ONE7 e45814 (2012) doi 101371journalpone0045814pmid 23029257
127 A Selvakumar T P OrsquoConnor S D Struck Role of streamrestoration on improving benthic macroinvertebrates and in-stream water quality in an urban watershed Case studyJ Environ Eng 136 127ndash139 (2010) doi 101061(ASCE)EE1943-78700000116
128 R W Lammers B P Bledsoe What role does streamrestoration play in nutrient management Crit Rev EnvironSci Technol 47 335ndash371 (2017) doi 1010801064338920171318618
129 H E Golden N Hoghooghi Green infrastructure and itscatchment-scale effects An emerging science WIREs Water5 e1254 (2018) doi 101002wat21254 pmid 29682288
130 M R Williams G Bhatt S Filoso G Yactayo Streamrestoration performance and its contribution to theChesapeake Bay TMDL Challenges posed by climate changein urban areas Estuaries Coasts 40 1227ndash1246 (2017)doi 101007s12237-017-0226-1
131 R M Fanelli K Prestegaard M A Palmer Freshw Sci101086706072 (2019)
132 B W Abbott et al Unexpected spatial stability of waterchemistry in headwater stream networks Ecol Lett 21296ndash308 (2018) doi 101111ele12897 pmid 29282860
133 B L Brown C M Swan Dendritic network structureconstrains metacommunity properties in riverineecosystems J Anim Ecol 79 571ndash580 (2010) doi 101111j1365-2656201001668x pmid 20180874
134 C M Swan B L Brown Metacommunity theory meetsrestoration Isolation may mediate how ecologicalcommunities respond to stream restoration Ecol Appl 272209ndash2219 (2017) doi 101002eap1602 pmid 28718193
135 N Mouquet D Gravel F Massol V Calcagno Extending theconcept of keystone species to communities and ecosystemsEcol Lett 16 1ndash8 (2013) doi 101111ele12014 pmid 23062191
136 A Ruhiacute T Datry J L Sabo Interpreting beta-diversitycomponents over time to conserve metacommunities inhighly dynamic ecosystems Conserv Biol 31 1459ndash1468(2017) doi 101111cobi12906 pmid 28188969
137 L Yamane L W Botsford D P Kilduff Tracking restorationof population diversity via the portfolio effect J Appl Ecol55 472ndash481 (2018) doi 1011111365-266412978
138 D W Braidwood M A Taggart M Smith R AndersenTranslocations conservation and climate change Use ofrestoration sites as protorefuges and protorefugia RestorEcol 26 20ndash28 (2018) doi 101111rec12642
139 Y Wang D Wang Q W Lewis J Wu F Huang A frameworkto assess the cumulative impacts of dams on hydrologicalregime A case study of the Yangtze River Hydrol Processes31 3045ndash3055 (2017) doi 101002hyp11239
140 J L Sabo et al Pulsed flows tributary inputs and food-webstructure in a highly regulated river J Appl Ecol 551884ndash1895 (2018) doi 1011111365-266413109
141 C Magliozzi G Coro R C Grabowski A I PackmanS Krause A multiscale statistical method to identifypotential areas of hyporheic exchange for river restorationplanning Environ Model Softw 111 311ndash323 (2019)doi 101016jenvsoft201809006
142 S S Schwartz B Smith Restoring hydrologic function inurban landscapes with suburban subsoiling J Hydrol 543770ndash781 (2016) doi 101016jjhydrol201610051
143 C J Walsh T D Fletcher A R Ladson Stream restoration inurban catchments through redesigning stormwater systemsLooking to the catchment to save the stream J N AmBenthol Soc 24 690ndash705 (2005) doi 10189904-0201
144 N M Schmadel et al Thresholds of lake and reservoirconnectivity in river networks control nitrogen removalNat Commun 9 2779 (2018) doi 101038s41467-018-05156-x pmid 30018449
145 C D Sayer Conservation of aquatic landscapes Pondslakes and rivers as integrated systems WIREs Water 1573ndash585 (2014) doi 101002wat21045
146 R G Luthy D L Sedlak M H Plumlee D Austin V H ReshWastewater‐effluent‐dominated streams as ecosystem‐
management tools in a drier climate Front Ecol Environ 13477ndash485 (2015) doi 101890150038
147 D W Roscoe S G Hinch Effectiveness monitoring of fishpassage facilities Historical trends geographic patterns andfuture directions Fish Fish 11 12ndash33 (2010) doi 101111j1467-2979200900333x
148 C M Bunt T Castro-Santos A Haro Performanceof fish passage structures at upstream barriers tomigration River Res Appl 28 457ndash478 (2012)doi 101002rra1565
149 D Weigel et al Evaluation of a trap-and-transport programfor a threatened population of steelhead (Oncorhynchusmykiss) Conserv Genet 20 1195ndash1199 (2019) doi 101007s10592-019-01200-5
150 M H Bond et al Combined effects of barge transportationriver environment and rearing location on straying andmigration of adult Snake River fall-run Chinook SalmonTrans Am Fish Soc 146 60ndash73 (2017) doi 1010800002848720161235614
151 A C Ritchie et al Morphodynamic evolution followingsediment release from the worldrsquos largest dam removalSci Rep 8 13279 (2018) doi 101038s41598-018-30817-8pmid 30185796
152 T Sommer et al Californiarsquos Yolo Bypass Evidence thatflood control can be compatible with fisheries wetlandswildlife and agriculture Fisheries 26 6ndash16 (2001)doi 1015771548-8446(2001)026lt0006CYBgt20CO2
153 G Grill et al Mapping the worldrsquos free-flowing riversNature 569 215ndash221 (2019) doi 101038s41586-019-1111-9pmid 31068722
154 G Gassert M Luck M Landis P Reig T Shiao ldquoAqueductglobal maps 21 Constructing decision-relevant global waterrisk indicatorsrdquo Working paper World Resources Institute(2015) wwwwriorgpublicationaqueduct-global-maps-21-indicators
155 A Ruhiacute J D Olden J L Sabo Declining streamflow inducescollapse and replacement of native fishes in the AmericanSouthwest Front Ecol Environ 14 465ndash472 (2016)doi 101002fee1424
156 T C Gouhier A Grinsted V Simko R package Biwaveletconduct univariate and bivariate wavelet analyses (2018)
157 T Rodriacuteguez-Castillo E Esteacutevez A M Gonzaacutelez-FerrerasJ Barquiacuten Estimating ecosystem metabolism toentire river networks Ecosystems (2018) doi 101007s10021-018-0311-8
ACKNOWLEDGMENTS
We greatly appreciate those who read and provided importantinsights on drafts of this paper A Armstrong K HondulaJ Hosen K Leathers G de Mendoza C Maietta I MunozJ Tonkin and two anonymous reviewers We also thank T Tu forhelp with Fig 1 and S Carlson L Comte T Datry J HosenM Kondolf N Jones D Lytle E Moody J Olden M PowerJ Sabo and M Sanchez-Montoya for valuable discussionsCompeting interests The authors have no competing interestsFunding MAP was supported by funds from the NSF (1639145)and AR was supported by funds from the University of CaliforniaBerkeley and NSF (1802714)
101126scienceaaw2087
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 13 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
restorationLinkages between flow regime biota and ecosystem processes Implications for river
Margaret Palmer and Albert Ruhi
DOI 101126scienceaaw2087 (6459) eaaw2087365Science
this issue p eaaw2087Scienceflow variability and achieve hydrological connectivity between a river and its surroundingsand ecosystem processes To be successful such efforts must go beyond accounting for flood pulses to restore natural mimic ecologically important aspects of natural flow regimes guided by insights into how variations in flow affect biotawater quality and ecological processes In a Review Palmer and Ruhi explain how restoration designs now attempt to
Human activities have altered the flow regimes of many of Earths rivers with negative impacts on biodiversityRiver restoration guided by research
ARTICLE TOOLS httpsciencesciencemagorgcontent3656459eaaw2087
REFERENCES
httpsciencesciencemagorgcontent3656459eaaw2087BIBLThis article cites 147 articles 7 of which you can access for free
PERMISSIONS httpwwwsciencemagorghelpreprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAASScienceScience 1200 New York Avenue NW Washington DC 20005 The title (print ISSN 0036-8075 online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
Science No claim to original US Government WorksCopyright copy 2019 The Authors some rights reserved exclusive licensee American Association for the Advancement of
on May 27 2020
httpsciencesciencem
agorgD
ownloaded from

and ecosystem processes can help prioritizeselection of sites for restoration For exampleusing the concept of river network saturation(55) to identify river sections where in-streamremoval of nutrients cannot keep up with highsource inputs of nutrients suggests that restorationactions targeting those sections may dispropor-tionately improve water quality Identification ofsubcatchments that disproportionately influencestream chemistry may also be informed by com-bining methods from landscape ecology andcatchment hydrology focusing on the spatialscale of variance in water chemistry (132) Head-waters are often prioritized for restoration be-
cause poor water quality in them can havedownstream impacts Research showing thatlocal environmental constraints on river com-munities are important in headwaters as well asin the larger channels but dispersal constraintsare much less important in the former (133 134)implies that headwaters should be prioritizedHowever research quantifying time-varying con-tributions of local communities to river-wide(network) biodiversity can also inform prioritiza-tion by identifying sites that host a dispropor-tionate share of species or that host uniquespeciesmdashfor example because they shelter sensi-tive taxa during critical flow bottlenecks This idea
encapsulates the concept of keystone habitatsor local sites that strongly influence metacom-munity dynamics (135) This has been appliedto aquatic invertebrates across intermittent rivernetworks by tracking time-varying site-specificcontributions to community dissimilarity (or betadiversity) (136)Other approaches to prioritize restoration ac-
tions may be based on contributions to system-wide stability rather than numbers of individualsor species For example the restoration of fall-run Chinook salmon portfolios may be assessedvia the contributions that different periods andlocationsmake tometapopulation-wide stabilityowing to spatiotemporal variation in habitatconditions (137) Additionally interventionscould promote conditions that are not impor-tant now but could be important in the future[ie proto-refugia (138)] For example resto-ration of groundwater-influenced habitats couldmitigate future stress to drought-sensitive taxaunder scenarios of increased intermittencyPatches of undisturbed land that protect flowregimes in streams within a larger catchmentthat is otherwise urbanized can be sources ofcolonists to restored sites Priorities that focuson present degradation of river sectionsmay failto address site contribution to impairment orrecovery at broader scales or under differentscenarios These insights indicate a great poten-tial for new methods to prioritize restoration ofboth urban and regulated streamsWe know little about how altered flow regimes
propagate through entire river networks butwork is starting to assess the cumulative effectsof dams on flow regimes as well as dam-levelcontributions to alteration Although these ap-proaches have largely focused on dam cascades[eg (139)] if expanded they could help identifyfavorable sections in river networks to developenvironmental flow operations Here exper-imentation could again help with testing ofnew strategies For instance large-scale flowexperiments could be designed that coordinatereleases among dams or between dams and free-flowing tributaries to enhance the restorativeeffects of natural pulses (140)
Hydrologic connectivityinfluences restoration
Recovering critical flow paths and their connec-tivity to rivers is important but it remains asubstantial challenge for flow-degraded riversand streams Whereas regulated rivers tend tohave reduced lateral and longitudinal hydro-logic connectivity urban streams have unnaturallevels of connectivity because pipes and over-land runoff route water quickly In both cases fun-damental hydroecological insights into streambedstructure landscape storage capacity and ground-water connectivity can help to envision novel re-storation approaches (130 141 142) More generallyhydrologic connectivity is becoming a frameworkto identify alternative ways to restore streams viastormwater management (143) For example arecent study suggested that reducing road densityand hydrologic connectivity between roads
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 8 of 13
ER
Disc
harg
e (f
t3 s)
GPPG
PP o
r ER
(g O
2 m-2
day
-1)
Discharge
Watershed BMPs
In-stream restoration
0
ndash4
4
0
ndash4
4
4
2
6
4
2
6
Jan Sept July May Mar Nov Jan
A
B C
Fig 6 Green infrastructure as a means to restore stream ecosystems (A) Discharge andecosystem respiration (ER more-negative values mean higher respiration) and gross primaryproduction (GPP) for two urban streams in which restoration efforts involved (top) modifications inthe stream channel and (bottom) traditional stormwater best management practices (BMPs)[Reproduced with permission from (103)] (B and C) Example of a comprehensive greeninfrastructure approach to ecologically restore an urban headwater stream that also supports thelifestyle of resident urban communities (B) Stream at the onset of restoration and (C) afterrestoration [Reproduced with permission from Greenworks Portland OR]
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 9 of 13
Table 1 Research challenges and opportunities Despite the progress that has been made in understanding how flow regimes affect biota and ecosystem
processes major challenges persist that prevent a complete understanding of the flow-biota-ecosystem processes nexus Making progress on these
challenges requires a mix of fundamental and applied research
Topic Research challenge Opportunity
Connecting
organismal to
ecosystem-level
processes
Research on the effects of flow on organisms
and ecosystem processes has followed parallel
tracksmdashhindering our mechanistic
understanding of the flow-biota-ecosystem
processes nexus
Generate theory that connects concepts and frameworks
that are well accepted in river community and ecosystem
ecology For example the notion of environmental filters
that connect regional to local composition via a combination
of dispersal and environmental conditions (community
ecology) with analogous theory on solute transport versus
reactivity (ecosystem ecology)
Our current understanding of the links between
phenology of biota and ecosystem processes
is precursory
Use high-frequency sensors to characterize metabolic
regimes at sites with ongoing organismal monitoring
Leverage citizen-science biodiversity data to detect
spatiotemporal shifts in animal phenology
(eg insect emergence fish spawning)
Use remote sensing tools to better understand
links between hydrology and aquatic and riparian
plant phenology
The link between stoichiometry and flow
variation remains largely unexplored
Repurpose data that is already being collected to test
flow-stoichiometry relationships eg studies using C and
N stable isotopes to describe CN ratios across flow
regime types Perform field experiments to quantify
indirect effects of invertebrate and fish consumption
on ecosystem metabolism
Ecological
consequences of
hydrologic
connectivity
The importance of anaerobic metabolism in
carbon processing as flows cease and parts of
streambeds dry is a major frontier research area
Create networks of sites across different geologies
and flow regimes that collect time-series data on
methane nitrous oxide and carbon dioxide fluxes
from intermittent streams
We ignore how biota in the wet phase is
influenced by processes and patterns
from the dry phase
Research the functions of dry riverbeds Use camera traps and
drone technology to investigate the role of dry riverbeds
as a dispersal corridor metabarcoding to quantify
ldquodark biodiversityrdquo in the hyporheic
Wetlands and their links to stream networks are poorly
delineated globally Even in the United States and
Europe current wetland inventories do not capture
these links yet understanding how their connectivity
to river systems influences network-scale ecosystem
processes is needed
Analyze time series from high-frequency sensors distributed in
river networks varying in wetland connectivity Further
development and application of time-series methods
used by neuroscientists
We need to identify the relationship between
varying flows and solute dynamics
associated with changes in catchment-scale
hydrologic exchanges
Design collaborative work among catchment hydrologists
and stream ecologists combining methods to estimate
transit times residence times and identify sources
across heterogeneous landscapes Link these properties to
land management and water quality aspects
Scaling and
transferring flow-
ecology relationships
across space and time
We ignore how biodiversity responds to
propagation of flow regime alteration
across river networks
Leverage extensive hydrometric networks (eg United States
Geological Survey National Water Information System)
to perform spatially replicated time-series analyses
on flow and use these to generate expectations
on how river network position should mediate
biodiversity responses to flow alteration
Flow-ecology relationships do not account for
nonstationarity and abrupt change
Use process-based models that account for mechanisms
and associated uncertainties When using time-series
data and models select methods that are robust
to nonstationarity Linear flow-ecology relationships
should not be assumed
Current flow-ecology relationships assume that
responses of species do not change over time
Consider flow regime alteration as a process with ecological and
evolutionary consequences Include intraspecific variation
in traits and potential for rapid evolution in models
continued on next page
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
and streams may improve water quality morethan other restoration options (70) Additionallyresearch on the age and lability of DOC enteringstreams has shown how precipitation or land dis-turbance can change watershed flow paths (53)and this could inspire new restoration approachesfocused on land use and soil properties Manystudies on hydrologic connectivity and networkstructure suggest a need for restoration to focuson landscape heterogeneity especially on thepresence ofwetlands lakes and ponds (144) Suchfeaturesare frequently lost inurbanizedcatchmentsand outcomes could be improved by restoring orincreasing connectivitywith these ecosystems (145)Hydrologic connectivity associated with ground-water inputs or abstractions land-use manage-ment and return flows fromwastewater treatmentplants also need to be considered in these initia-tives (146)Restoring hydrologic connectivity is also key
to achieving biodiversity outcomesmdashparticularlyin dammed systems Longitudinal connectivityfor fish inhabiting dam-altered rivers can beenhanced by the construction of fishways andsophisticated passage facilities However moni-toring data on the effectiveness of such inter-
ventions are still scarce and available evidencesuggests that passage effectivity largely dependson technology used and species-specific fish be-havior (147 148) Although trap-and-transportprograms for migratory fishes can help augmentthreatened populations (149) these practicesoften increase straying and thus the potentialfor genetic homogenization as seen from ju-venile salmon collected at dams and transporteddownstream to increase out-migration success(150) Overall a combination of engineering andmanagement solutions may be best to restorefunctional connectivity in dam-altered riversThe link between hydrologic connectivity andecological restoration is also central to con-siderations on where to build remove and re-operate hydrologic infrastructure Hundredsof dams have been intentionally removed sincethe 1970s and removal of large dams such asthe Elwha River Dam (Washington USA) hasshown that restoring longitudinal connectivitycan have immediate geomorphic effects with com-plex temporal dynamics (151) Infrastructure canalso be designed or reoperated to manage forrestoring lateral connectivity and enhanced flood-plain processes For example the Yolo Bypass in
the Sacramento River California has becomea prime rearing and migration site for chinooksalmon showing that flood control and habitatfor fish and wildlife do not need to be at odds(152) All these examples show the importanceof integrating hydrologic connectivity in ecolog-ical restoration whether the goal is to minimizethe ecological effects of fragmentation restorefree-flowing watercourses or operate green infra-structure to enhance infiltration floodplain con-nectivity or other ecological processes
Research challenges and opportunities
Two decades of concerted efforts to research theeffects of flow regime alteration have tremen-dously advanced our understanding of the waysin which streamflow influences biota and eco-system processes However important challengesremain These aremainly related to the scarcity ofscience connecting organismal to ecosystem-levelprocesses through for example microbial linksthe difficulty of studying watershed-level phys-ical and biogeochemical heterogeneity and itsinfluence on riverine processes and biota thecomplexity of parsing out the direct effects offlow from flow-mediated effects and co-occurring
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 10 of 13
Topic Research challenge Opportunity
Cascading
implications of
hydrologic
alteration
We have a poor understanding of the
interactions among physical drivers
Build on large-scale flow experiments to test hypotheses on the
combined effects of altered flow sediment and thermal
regimes Experiment with multilevel intakes in reservoirs
Promote collection of non-flow variables (eg temperature
water quality parameters) at hydrometric stations
At low flows biota are being exposed to high
concentrations of new classes of pollutants
whose impacts are poorly known especially if
biotic impacts interact with impacts on
ecosystem processes
Design realistic experimental studies to advance knowledge
on the ecological effects of multiple stressors
Collect long-term data at sites varying in contaminant
inputs and hydrologic regimes Link microbial to
organismal and ecosystem-level responses
Using hydroecological
science to inform
river ecosystem
restoration
Environmental flow science largely ignores
human dimensions
Use methods that allow identifying causal networks
in complex socioenvironmental systems Include
human needs and thresholds in research on
flow-ecology relationships
Environmental flows are generally prescribed
at the scale of single infrastructures restoration
designs and assessment are generally at reach scales
Promote coordinated large-scale flow experiments
and restoration designs at the river basin scale
Extend assessment of green infrastructure
effectiveness beyond individual structures to
infrastructure networks and link both
to river outcomes
Environmental flows in impounded rivers
generally target the conservation of some
particular species rather than outcomes at the
food-web or ecosystem level
Advance fundamental research on microbialndash
ecosystem process linkages Expand the notion
of designer flows to food-web structure and
ecosystem services when possible
Environmental flow science has largely
ignored surface waterndashgroundwater
interactions
Expand science to understand the link between
restoration actions groundwater recharge and
inputs to streams Provide science relevant to
the implementation of new legislation on
groundwater management (eg Sustainable
Groundwater Management Act in California)
Uncertainty regarding future
hydroclimate and
river flows is growing
Refine regional climate models Promote research on
the interaction between land-use change and
evapotranspiration Incorporate groundwater
influence in macroscale hydrology models
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
stressors (particularly in urban streams) and thechallenge of predicting outcomeswhen both thephysical and the biological environments arechangingAchieving a predictive understanding of hy-
droecological relationships is essential to design-ing successful self-sustaining restoration actionsin river ecosystems However translating basichydroecological insights into restoration practicerequires understanding the complexities of work-ing within a human-dominated water cycle Al-though it is rarely possible to restore all facetsof a natural flow regime ecologically informeddesigns may help mimic key aspects of the nat-ural flow regime (eg periodic pulse flows) orcreate regimes that are not natural but maximizeoutcomes given socioeconomic constraints InTable 1 we identify persisting and emergingchallenges in hydroecological research We paireach challenge with an opportunity (related toresearch design technology or implementation)We contend that these challenges should beprioritized to advance process-based river eco-system restoration
REFERENCES AND NOTES
1 N L Poff et al The natural flow regime Bioscience 47769ndash784 (1997) doi 1023071313099
2 D A Lytle N L Poff Adaptation to natural flow regimesTrends Ecol Evol 19 94ndash100 (2004) doi 101016jtree200310002 pmid 16701235
3 S Sabater A Elosegi R Ludwig in Multiple Stressors in RiverEcosystems (Elsevier 2019) pp 1ndash22
4 United Nations New UN Decade on Ecosystem Restorationoffers unparalleled opportunity for job creation food securityand addressing climate change (2019) httpswwwunenvironmentorgnews-and-storiespress-releasenew-un-decade-ecosystem-restoration-offers-unparalleled-opportunity
5 A W Lorenz P Haase K Januschke A SundermannD Hering Revisiting restored river reaches - Assessingchange of aquatic and riparian communities afterfive years Sci Total Environ 613-614 1185ndash1195(2018) doi 101016jscitotenv201709188pmid 28954379
6 M A Palmer K L Hondula B J Koch Ecological Restorationof Streams and Rivers Shifting Strategies and ShiftingGoals (2014) vol 45
7 T J Beechie et al Process-based principles for restoringriver ecosystems Bioscience 60 209ndash222 (2010)doi 101525bio20106037
8 S E Bunn A H Arthington Basic principles and ecologicalconsequences of altered flow regimes for aquaticbiodiversity Environ Manage 30 492ndash507 (2002)doi 101007s00267-002-2737-0 pmid 12481916
9 K D Fausch Y Taniguchi S Nakano G D GrossmanC R Townsend Flood disturbance regimes influence rainbowtrout invasion success among five holarctic regions EcolAppl 11 1438ndash1455 (2001) doi 1018901051-0761(2001)011[1438FDRIRT]20CO2
10 M E Power M S Parker W E Dietrich Seasonalreassembly of a river food web Floods droughts andimpacts of fish Ecol Monogr 78 263ndash282 (2008)doi 10189006-09021
11 V Acuntildea A Giorgi I Muntildeoz U R S Uehlinger S SabaterFlow extremes and benthic organic matter shape themetabolism of a headwater Mediterranean streamFreshwater Biol 49 960ndash971 (2004) doi 101111j1365-2427200401239x
12 K Tockner F Malard J V Ward An extension of the floodpulse concept Hydrol Processes 14 2861ndash2883 (2000)doi 1010021099-1085(20001112)141617lt2861AID-HYP124gt30CO2-F
13 J S Perkin et al Fragmentation and dewatering transformGreat Plains stream fish communities Ecol Monogr 8573ndash92 (2015) doi 10189014-01211
14 J L Sabo D M Post Quantifying periodic stochastic andcatastrophic environmental variation Ecol Monogr 7819ndash40 (2008) doi 10189006-13401
15 F Shuai et al Silver carp larva abundance in response toriver flow rate revealed by cross-wavelet modellingEcol Modell 383 98ndash105 (2018) doi 101016jecolmodel201805020
16 J D Tonkin M T Bogan N Bonada B Rios-ToumaD A Lytle Seasonality and predictability shape temporalspecies diversity Ecology 98 1201ndash1216 (2017)doi 101002ecy1761 pmid 28144975
17 N L Poff Beyond the natural flow regime Broadening thehydro‐ecological foundation to meet environmental flowschallenges in a non‐stationary world Freshwater Biol 631011ndash1021 (2018) doi 101111fwb13038
18 A Ruhi X Dong C H McDaniel D P Batzer J L SaboDetrimental effects of a novel flow regime on the functionaltrajectory of an aquatic invertebrate metacommunity GlobChange Biol 24 3749ndash3765 (2018) doi 101111gcb14133pmid 29665147
19 M T Bogan D A Lytle Severe drought drives novelcommunity trajectories in desert stream pools FreshwaterBiol 56 2070ndash2081 (2011) doi 101111j1365-2427201102638x
20 G Woodward et al The effects of climatic fluctuations andextreme events on running water ecosystems Philos TransR Soc London Ser B 371 20150274 (2016) pmid 27114576
21 M A Palmer C M Swan K Nelson P Silver R AlvestadStreambed landscapes Evidence that stream invertebratesrespond to the type and spatial arrangement of patchesLandsc Ecol 15 563ndash576 (2000) doi 101023A1008194130695
22 D Schmera et al Does isolation influence the relative role ofenvironmental and dispersal‐related processes in streamnetworks An empirical test of the network positionhypothesis using multiple taxa Freshwater Biol 63 74ndash85(2018) doi 101111fwb12973
23 R Henriques‐Silva et al A comprehensive examination of thenetwork position hypothesis across multiple rivermetacommunities Ecography 42 284ndash294 (2019)doi 101111ecog03908
24 K L Jaeger J D Olden N A Pelland Climate change poisedto threaten hydrologic connectivity and endemic fishes indryland streams Proc Natl Acad Sci USA 11113894ndash13899 (2014) doi 101073pnas1320890111pmid 25136090
25 S Garbin E A Celegon P Fanton G Botter Hydrologicalcontrols on river network connectivity R Soc Open Sci 6181428 (2019) doi 101098rsos181428 pmid 30891270
26 F Cattaneacuteo B Hugueny N Lamouroux Synchrony in browntrout Salmo trutta population dynamics A lsquoMoran effectrsquo onearly‐life stages Oikos 100 43ndash54 (2003) doi 101034j1600-0706200311912x
27 A Terui et al Metapopulation stability in branching rivernetworks Proc Natl Acad Sci USA 115 E5963ndashE5969(2018) doi 101073pnas1800060115 pmid 29895695
28 P A P Moran The statistical analysis of the Canadian Lynxcycle Aust J Zool 1 291ndash298 (1953) doi 101071ZO9530291
29 G D Grossman P B Moyle J O Whitaker Jr Stochasticityin structural and functional characteristics of an Indianastream fish assemblage A test of community theoryAm Nat 120 423ndash454 (1982) doi 101086284004
30 D A Lytle D M Merritt J D Tonkin J D OldenL V Reynolds Linking river flow regimes to riparian plantguilds A community-wide modeling approach Ecol Appl 271338ndash1350 (2017) doi 101002eap1528 pmid 28263426
31 A Ruhiacute E E Holmes J N Rinne J L Sabo Anomalousdroughts not invasion decrease persistence of native fishesin a desert river Glob Change Biol 21 1482ndash1496 (2015)doi 101111gcb12780 pmid 25430731
32 J D Tonkin D M Merritt J D Olden L V ReynoldsD A Lytle Flow regime alteration degrades ecologicalnetworks in riparian ecosystems Nat Ecol Evol 2 86ndash93(2018) doi 101038s41559-017-0379-0 pmid 29180707
33 M C Mims J D Olden Fish assemblages respond toaltered flow regimes via ecological filtering of lifehistory strategies Freshwater Biol 58 50ndash62 (2013)doi 101111fwb12037
34 P T J Johnson J D Olden M J Vander Zanden Daminvaders Impoundments facilitate biological invasions intofreshwaters Front Ecol Environ 6 357ndash363 (2008)doi 101890070156
35 N Bonada S Doledec B Statzner Taxonomic and biologicaltrait differences of stream macroinvertebrate communitiesbetween mediterranean and temperate regions Implicationsfor future climatic scenarios Glob Change Biol 131658ndash1671 (2007) doi 101111j1365-2486200701375x
36 M I Pyne N L Poff Vulnerability of stream communitycomposition and function to projected thermal warming andhydrologic change across ecoregions in the western UnitedStates Glob Change Biol 23 77ndash93 (2017) doi 101111gcb13437 pmid 27429092
37 D B Herbst S D Cooper R B Medhurst S W WisemanC T Hunsaker Drought ecohydrology alters the structureand function of benthic invertebrate communities inmountain streams Freshwater Biol 64 886ndash902 (2019)doi 101111fwb13270
38 M T Bogan E T Chester T Datry A L MurphyB J Robson A Ruhi R Stubbington J E Whitney inResistance Resilience and Community Recovery in IntermittentRivers and Ephemeral Streams T Datry N Bonada A BoultonEds (Academic Press 2017) pp 349ndash376
39 S Houmlckendorff et al Characterizing fish responses to a riverrestoration over 21 years based on speciesrsquo traits ConservBiol 31 1098ndash1108 (2017) doi 101111cobi12908pmid 28218807
40 D Bruno et al Structural and functional responses ofinvertebrate communities to climate change andflow regulation in alpine catchments Glob Change Biol25 1612ndash1628 (2019) doi 101111gcb14581pmid 30698905
41 T W H Aspin et al Extreme drought pushes streaminvertebrate communities over functional thresholds GlobChange Biol 25 230ndash244 (2019) doi 101111gcb14495pmid 30346098
42 S E Tank J B Fellman E Hood E S Kritzberg Beyondrespiration Controls on lateral carbon fluxes across theterrestrial-aquatic interface Limnol Oceanogr Lett 3 76ndash88(2018) doi 101002lol210065
43 A P Appling et al The metabolic regimes of 356 rivers inthe United States Sci Data 5 180292 (2018) doi 101038sdata2018292 pmid 30532078
44 P Savoy et al Metabolic rhythms in flowing watersAn approach for classifying river productivity regimesLimnol Oceanogr (2019) doi 101002lno11154
45 E S Bernhardt et al The metabolic regimes of flowingwaters Limnol Oceanogr 63 S99ndashS118 (2018)doi 101002lno10726
46 M Colls X Timoner S Sabater V Acuntildea Effects of durationand frequency of the non-flow period on stream metabolismEcosystems 101007s10021-019-00345-1 (2019)doi 101007s10021-019-00345-1
47 A J Ulseth E Bertuzzo G A Singer J Schelker T J BattinClimate-induced changes in spring snowmelt impactecosystem metabolism and carbon fluxes in an Alpine streamnetwork Ecosystems 21 373ndash390 (2018) doi 101007s10021-017-0155-7
48 B O L Demars Hydrological pulses and burning of dissolvedorganic carbon by stream respiration Limnol Oceanogr 64406ndash421 (2019) doi 101002lno11048
49 R T Hensley L Kirk M Spangler M N Gooseff M J CohenFlow extremes as spatiotemporal control points on riversolute fluxes and metabolism J Geophys Res Biogeosci124 537ndash555 (2019) doi 1010292018JG004738
50 J D Hosen et al Enhancement of primary production duringdrought in a temperate watershed is greater in larger riversthan headwater streams Limnol Oceanogr 64 1458ndash1472(2019) doi 101002lno11127
51 L Goacutemez-Gener et al When water vanishes Magnitude andregulation of carbon dioxide emissions from dry temporarystreams Ecosystems 19 710ndash723 (2016) doi 101007s10021-016-9963-4
52 E H Stanley et al The ecology of methane in streams andrivers Patterns controls and global significance EcolMonogr 86 146ndash171 (2016) doi 10189015-1027
53 R T Barnes D E Butman H F Wilson P A RaymondRiverine export of aged carbon driven by flow path depth andresidence time Environ Sci Technol 52 1028ndash1035 (2018)doi 101021acsest7b04717 pmid 29313674
54 G Pinay et al Riparian corridors A new conceptualframework for assessing nitrogen buffering across biomesFront Environ Sci 6 47 (2018) doi 103389fenvs201800047
55 W M Wollheim et al River network saturation conceptFactors influencing the balance of biogeochemical supply and
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 11 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
demand of river networks Biogeochemistry 141 503ndash521(2018) doi 101007s10533-018-0488-0
56 P A Raymond J E Saiers W V Sobczak Hydrological andbiogeochemical controls on watershed dissolved organicmatter transport Pulse-shunt concept Ecology 97 5ndash16(2016) doi 10189014-16841 pmid 27008769
57 C Tunaley D Tetzlaff J Lessels C Soulsby Linking high-frequency DOC dynamics to the age of connected watersources Water Resour Res 52 5232ndash5247 (2016)doi 1010022015WR018419
58 S Bernal et al Supply demand and in-stream retention ofdissolved organic carbon and nitrate during storms inMediterranean forested headwater streams Front EnvironSci 7 60 (2019) doi 103389fenvs201900060
59 J Harvey et al How hydrologic connectivity regulates waterquality in river corridors J Am Water Resour Assoc 55369ndash381 (2019) doi 1011111752-168812691
60 J C Moore et al Detritus trophic dynamics and biodiversityEcol Lett 7 584ndash600 (2004) doi 101111j1461-0248200400606x
61 A D Rosemond et al Freshwater ecology Experimentalnutrient additions accelerate terrestrial carbon loss fromstream ecosystems Science 347 1142ndash1145 (2015)doi 101126scienceaaa1958 pmid 25745171
62 A C Y Yeung J L Musetta-Lambert D P KreutzweiserP K Sibley J S Richardson Relations of interannualdifferences in stream litter breakdown with dischargeBioassessment implications Ecosphere 9 e02423 (2018)doi 101002ecs22423
63 R M Northington J R Webster Experimental reductions instream flow alter litter processing and consumer subsidies inheadwater streams Freshwater Biol 62 737ndash750 (2017)doi 101111fwb12898
64 E Mas-Martiacute I Sanpera-Calbet I Muntildeoz Bottom-up effectsof streambed drying on consumer performance throughchanges in resource quality Aquat Sci 79 719ndash731 (2017)doi 101007s00027-017-0531-6
65 B D Richter G A Thomas Restoring environmental flows bymodifying dam operations Ecol Soc 12 12 (2007)doi 105751ES-02014-120112
66 G Zolezzi A Siviglia M Toffolon B MaioliniThermopeaking in Alpine streams Event characterizationand time scales Ecohydrology 4 564ndash576 (2011)doi 101002eco132
67 T Meiszligner B Sures C K Feld Multiple stressors and therole of hydrology on benthic invertebrates in mountainousstreams Sci Total Environ 663 841ndash851 (2019)doi 101016jscitotenv201901288 pmid 30738264
68 I R Waite et al Effects of urban multi-stressors on threestream biotic assemblages Sci Total Environ 6601472ndash1485 (2019) doi 101016jscitotenv201812240pmid 30743940
69 K C Nelson M A Palmer Stream temperature surges underurbanization and climate change Data models andresponses J Am Water Resour Assoc 43 440ndash452 (2007)doi 101111j1752-1688200700034x
70 J R Blaszczak J M Delesantro Y Zhong D L UrbanE S Bernhardt Watershed urban development controls onurban streamwater chemistry variability Biogeochemistry144 61ndash84 (2019) doi 101007s10533-019-00572-7
71 L Ponsatiacute et al Biofilm responses to flow regulation by damsin Mediterranean rivers River Res Appl 31 1003ndash1016(2015) doi 101002rra2807
72 J M Alberts J J Beaulieu I Buffam Watershed land useand seasonal variation constrain the influence of ripariancanopy cover on stream ecosystem metabolism Ecosystems20 553ndash567 (2017) doi 101007s10021-016-0040-9pmid 30416371
73 R O Hall Jr et al Turbidity light temperature andhydropeaking control primary productivity in the ColoradoRiver Grand Canyon Limnol Oceanogr 60 512ndash526 (2015)doi 101002lno10031
74 J R Blaszczak J M Delesantro D L Urban M W DoyleE S Bernhardt Scoured or suffocated Urban streamecosystems oscillate between hydrologic and dissolvedoxygen extremes Limnol Oceanogr 64 877ndash894 (2019)doi 101002lno11081
75 J-R Mor et al Dam regulation and riverine food-webstructure in a Mediterranean river Sci Total Environ 625301ndash310 (2018) doi 101016jscitotenv201712296pmid 29289778
76 H A Wellard Kelly et al Macroinvertebrate diets reflecttributary inputs and turbidity-driven changes in food
availability in the Colorado River downstream of GlenCanyon Dam Freshw Sci 32 397ndash410 (2013)doi 10189912-0881
77 S J Imberger R M Thompson M R Grace Urbancatchment hydrology overwhelms reach scale effects ofriparian vegetation on organic matter dynamics FreshwaterBiol 56 1370ndash1389 (2011) doi 101111j1365-2427201102575x
78 R T Martins A S Melo J F Gonccedilalves Jr N HamadaLeaf-litter breakdown in urban streams of Central AmazoniaDirect and indirect effects of physical chemical andbiological factors Freshw Sci 34 716ndash726 (2015)doi 101086681086
79 C M Yule J Y Gan T Jinggut K V Lee Urbanizationaffects food webs and leaf-litter decomposition in a tropicalstream in Malaysia Freshw Sci 34 702ndash715 (2015)doi 101086681252
80 T A Newcomer et al Influence of natural and novel organiccarbon sources on denitrification in forest degraded urbanand restored streams Ecol Monogr 82 449ndash466 (2012)doi 10189012-04581
81 A Chaudhary I Kauser A Ray R Poretsky Taxon-drivenfunctional shifts associated with storm flow in an urbanstream microbial community MSphere 3 e00194-18 (2018)doi 101128mSphere00194-18 pmid 29976643
82 T A Kennedy et al Flow management for hydropowerextirpates aquatic insects undermining river food websBioscience 66 561ndash575 (2016) doi 101093bioscibiw059
83 A Glazaczow D Orwin M Bogdziewicz Increasedtemperature delays the late-season phenology of multivoltineinsect Sci Rep 6 38022 (2016) doi 101038srep38022pmid 27905493
84 S G Fisher L J Gray N B Grimm D E Busch Temporalsuccession in a desert stream ecosystem following flashflooding Ecol Monogr 52 93ndash110 (1982) doi 1023072937346
85 G J Scrimgeour M J Winterbourn Effects of floods onepilithon and benthic macroinvertebrate populations in anunstable New Zealand river Hydrobiologia 171 33ndash44 (1989)doi 101007BF00005722
86 E Bondar-Kunze S Maier D Schoumlnauer N Bahl T HeinAntagonistic and synergistic effects on a stream periphytoncommunity under the influence of pulsed flow velocityincrease and nutrient enrichment Sci Total Environ 573594ndash602 (2016) doi 101016jscitotenv201608158pmid 27585428
87 M J Cashman G L Harvey G Wharton M C Bruno Woodmitigates the effect of hydropeaking scour on periphytonbiomass and nutritional quality in semi-natural flumesimulations Aquat Sci 79 459ndash471 (2017) doi 101007s00027-016-0510-3
88 W F Cross et al Food-web dynamics in a large riverdiscontinuum Ecol Monogr 83 311ndash337 (2013)doi 10189012-17271
89 D M Warfe et al Productivity disturbance and ecosystemsize have no influence on food chain length in seasonallyconnected rivers PLOS ONE 8 e66240 (2013)doi 101371journalpone0066240
90 T D Jardine et al Consumer-resource coupling in wet-drytropical rivers J Anim Ecol 81 310ndash322 (2012)doi 101111j1365-2656201101925x pmid 22103689
91 K O Winemiller et al Pulsing hydrology determines top‐down control of basal resources in a tropical riverndashfloodplainecosystem Ecol Monogr 84 621ndash635 (2014)doi 10189013-18221
92 D M Post The long and short of food-chain length TrendsEcol Evol 17 269ndash277 (2002) doi 101016S0169-5347(02)02455-2
93 D M Post G Takimoto Proximate structural mechanismsfor variation in food‐chain length Oikos 116 775ndash782 (2007)doi 101111j0030-1299200715552x
94 P A McHugh A R McIntosh P G Jellyman Dual influencesof ecosystem size and disturbance on food chain length instreams Ecol Lett 13 881ndash890 (2010) doi 101111j1461-0248201001484x pmid 20482579
95 A Ruhiacute et al Flow regulation increases food-chain lengththrough omnivory mechanisms in a Mediterranean rivernetwork Freshwater Biol 61 1536ndash1549 (2016) doi 101111fwb12794
96 J L Sabo J C Finlay T Kennedy D M Post The role ofdischarge variation in scaling of drainage area and food chainlength in rivers Science 330 965ndash967 (2010) doi 101126science1196005 pmid 20947729
97 A R McIntosh et al Capacity to support predators scaleswith habitat size Sci Adv 4 eaap7523 (2018) doi 101126sciadvaap7523 pmid 29978038
98 J H Brown J F Gillooly A P Allen V M Savage G B WestToward a metabolic theory of ecology Ecology 851771ndash1789 (2004) doi 10189003-9000
99 K Zou E Theacutebault G Lacroix S Barot Interactions betweenthe green and brown food web determine ecosystemfunctioning Funct Ecol 30 1454ndash1465 (2016) doi 1011111365-243512626
100 M J Vanni P B McIntyre Predicting nutrient excretion ofaquatic animals with metabolic ecology and ecologicalstoichiometry A global synthesis Ecology 97 3460ndash3471(2016) doi 101002ecy1582 pmid 27912023
101 T B Parr K A Capps S P Inamdar K A Metcalf Animal-mediated organic matter transformation Aquatic insects as asource of microbially bioavailable organic nutrients andenergy Funct Ecol 33 524ndash535 (2019) doi 1011111365-243513242
102 C P Arango et al Urban infrastructure influences dissolvedorganic matter quality and bacterial metabolism in an urbanstream network Freshwater Biol 62 1917ndash1928 (2017)
103 L G Larsen J W Harvey Disrupted carbon cycling inrestored and unrestored urban streams Critical timescalesand controls Limnol Oceanogr 62 S160ndashS182 (2017)doi 101002lno10613
104 H M Halvorson G White J T Scott M A Evans-WhiteDietary and taxonomic controls on incorporation of microbialcarbon and phosphorus by detritivorous caddisfliesOecologia 180 567ndash579 (2016) doi 101007s00442-015-3464-6 pmid 26497125
105 J J Elser et al Biological stoichiometry from genes toecosystems Ecol Lett 3 540ndash550 (2000) doi 101046j1461-0248200000185x
106 E K Moody et al Consumption explains intraspecificvariation in nutrient recycling stoichiometry in a desert fishEcology 99 1552ndash1561 (2018) doi 101002ecy2372pmid 29882955
107 N Welti et al Bridging food webs ecosystem metabolismand biogeochemistry using ecological stoichiometry theoryFront Microbiol 8 1298 (2017) doi 103389fmicb201701298 pmid 28747904
108 M A Palmer S Filoso R M Fanelli From ecosystems toecosystem services Stream restoration as ecologicalengineering Ecol Eng 65 62ndash70 (2014) doi 101016jecoleng201307059
109 M J Burns T D Fletcher C J Walsh A R LadsonB E Hatt Hydrologic shortcomings of conventional urbanstormwater management and opportunities for reformLandsc Urban Plan 105 230ndash240 (2012) doi 101016jlandurbplan201112012
110 C R Liermann C Nilsson J Robertson R Y NgImplications of dam obstruction for global freshwater fishdiversity Bioscience 62 539ndash548 (2012) doi 101525bio20126265
111 J D Olden R J Naiman Incorporating thermal regimes intoenvironmental flows assessments Modifying dam operationsto restore freshwater ecosystem integrity Freshwater Biol55 86ndash107 (2010) doi 101111j1365-2427200902179x
112 D B Osmundson R J Ryel V L Lamarra J Pitlick Flowndashsedimentndashbiota relations Implications for river regulationeffects on native fish abundance Ecol Appl 12 1719ndash1739(2002)
113 D von Schiller et al Regulation causes nitrogen cyclingdiscontinuities in Mediterranean rivers Sci Total Environ540 168ndash177 (2016) doi 101016jscitotenv201507017pmid 26233271
114 J L Sabo et al Designing river flows to improve foodsecurity futures in the Lower Mekong Basin Science 358eaao1053 (2017) doi 101126scienceaao1053pmid 29217541
115 W Xu et al Improving the multi-objective performance ofrainwater harvesting systems using real-time controltechnology Water 10 147 (2018) doi 103390w10020147
116 M Acreman et al Environmental flows for naturalhybrid and novel riverine ecosystems in a changingworld Front Ecol Environ 12 466ndash473 (2014)doi 101890130134
117 N L Poff et al Sustainable water management under futureuncertainty with eco-engineering decision scaling Nat ClimChang 6 25ndash34 (2016) doi 101038nclimate2765
118 J D Kiernan P B Moyle P K Crain Restoring native fishassemblages to a regulated California stream using the
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 12 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
natural flow regime concept Ecol Appl 22 1472ndash1482(2012) doi 10189011-04801 pmid 22908707
119 D L Propst K B Gido Responses of native andnonnative fishes to natural flow regime mimicry in theSan Juan River Trans Am Fish Soc 133 922ndash931 (2004)doi 101577T03-0571
120 T K Pool J D Olden Assessing long‐term fish responses andshort‐term solutions to flow regulation in a dryland river basinEcol Freshwat Fish 24 56ndash66 (2015) doi 101111eff12125
121 W Chen J D Olden Designing flows to resolve human andenvironmental water needs in a dam-regulated riverNat Commun 8 2158 (2017) doi 101038s41467-017-02226-4 pmid 29255194
122 S M Nelson et al A history of the 2014 Minute 319environmental pulse flow as documented by fieldmeasurements and satellite imagery Ecol Eng 106733ndash748 (2017) doi 101016jecoleng201610040
123 P B Shafroth et al A large-scale environmental flowexperiment for riparian restoration in the Colorado RiverDelta Ecol Eng 106 645ndash660 (2017) doi 101016jecoleng201702016
124 E P Glenn P L Nagler P B Shafroth C J JarchowEffectiveness of environmental flows for riparian restorationin arid regions A tale of four rivers Ecol Eng 106 695ndash703(2017) doi 101016jecoleng201701009
125 S Hotaling D S Finn J Joseph Giersch D W WeisrockD Jacobsen Climate change and alpine stream biologyProgress challenges and opportunities for the futureBiol Rev Camb Philos Soc 92 2024ndash2045 (2017)doi 101111brv12319 pmid 28105701
126 C J Walsh T D Fletcher M J Burns Urban stormwaterrunoff A new class of environmental flow problem PLOS ONE7 e45814 (2012) doi 101371journalpone0045814pmid 23029257
127 A Selvakumar T P OrsquoConnor S D Struck Role of streamrestoration on improving benthic macroinvertebrates and in-stream water quality in an urban watershed Case studyJ Environ Eng 136 127ndash139 (2010) doi 101061(ASCE)EE1943-78700000116
128 R W Lammers B P Bledsoe What role does streamrestoration play in nutrient management Crit Rev EnvironSci Technol 47 335ndash371 (2017) doi 1010801064338920171318618
129 H E Golden N Hoghooghi Green infrastructure and itscatchment-scale effects An emerging science WIREs Water5 e1254 (2018) doi 101002wat21254 pmid 29682288
130 M R Williams G Bhatt S Filoso G Yactayo Streamrestoration performance and its contribution to theChesapeake Bay TMDL Challenges posed by climate changein urban areas Estuaries Coasts 40 1227ndash1246 (2017)doi 101007s12237-017-0226-1
131 R M Fanelli K Prestegaard M A Palmer Freshw Sci101086706072 (2019)
132 B W Abbott et al Unexpected spatial stability of waterchemistry in headwater stream networks Ecol Lett 21296ndash308 (2018) doi 101111ele12897 pmid 29282860
133 B L Brown C M Swan Dendritic network structureconstrains metacommunity properties in riverineecosystems J Anim Ecol 79 571ndash580 (2010) doi 101111j1365-2656201001668x pmid 20180874
134 C M Swan B L Brown Metacommunity theory meetsrestoration Isolation may mediate how ecologicalcommunities respond to stream restoration Ecol Appl 272209ndash2219 (2017) doi 101002eap1602 pmid 28718193
135 N Mouquet D Gravel F Massol V Calcagno Extending theconcept of keystone species to communities and ecosystemsEcol Lett 16 1ndash8 (2013) doi 101111ele12014 pmid 23062191
136 A Ruhiacute T Datry J L Sabo Interpreting beta-diversitycomponents over time to conserve metacommunities inhighly dynamic ecosystems Conserv Biol 31 1459ndash1468(2017) doi 101111cobi12906 pmid 28188969
137 L Yamane L W Botsford D P Kilduff Tracking restorationof population diversity via the portfolio effect J Appl Ecol55 472ndash481 (2018) doi 1011111365-266412978
138 D W Braidwood M A Taggart M Smith R AndersenTranslocations conservation and climate change Use ofrestoration sites as protorefuges and protorefugia RestorEcol 26 20ndash28 (2018) doi 101111rec12642
139 Y Wang D Wang Q W Lewis J Wu F Huang A frameworkto assess the cumulative impacts of dams on hydrologicalregime A case study of the Yangtze River Hydrol Processes31 3045ndash3055 (2017) doi 101002hyp11239
140 J L Sabo et al Pulsed flows tributary inputs and food-webstructure in a highly regulated river J Appl Ecol 551884ndash1895 (2018) doi 1011111365-266413109
141 C Magliozzi G Coro R C Grabowski A I PackmanS Krause A multiscale statistical method to identifypotential areas of hyporheic exchange for river restorationplanning Environ Model Softw 111 311ndash323 (2019)doi 101016jenvsoft201809006
142 S S Schwartz B Smith Restoring hydrologic function inurban landscapes with suburban subsoiling J Hydrol 543770ndash781 (2016) doi 101016jjhydrol201610051
143 C J Walsh T D Fletcher A R Ladson Stream restoration inurban catchments through redesigning stormwater systemsLooking to the catchment to save the stream J N AmBenthol Soc 24 690ndash705 (2005) doi 10189904-0201
144 N M Schmadel et al Thresholds of lake and reservoirconnectivity in river networks control nitrogen removalNat Commun 9 2779 (2018) doi 101038s41467-018-05156-x pmid 30018449
145 C D Sayer Conservation of aquatic landscapes Pondslakes and rivers as integrated systems WIREs Water 1573ndash585 (2014) doi 101002wat21045
146 R G Luthy D L Sedlak M H Plumlee D Austin V H ReshWastewater‐effluent‐dominated streams as ecosystem‐
management tools in a drier climate Front Ecol Environ 13477ndash485 (2015) doi 101890150038
147 D W Roscoe S G Hinch Effectiveness monitoring of fishpassage facilities Historical trends geographic patterns andfuture directions Fish Fish 11 12ndash33 (2010) doi 101111j1467-2979200900333x
148 C M Bunt T Castro-Santos A Haro Performanceof fish passage structures at upstream barriers tomigration River Res Appl 28 457ndash478 (2012)doi 101002rra1565
149 D Weigel et al Evaluation of a trap-and-transport programfor a threatened population of steelhead (Oncorhynchusmykiss) Conserv Genet 20 1195ndash1199 (2019) doi 101007s10592-019-01200-5
150 M H Bond et al Combined effects of barge transportationriver environment and rearing location on straying andmigration of adult Snake River fall-run Chinook SalmonTrans Am Fish Soc 146 60ndash73 (2017) doi 1010800002848720161235614
151 A C Ritchie et al Morphodynamic evolution followingsediment release from the worldrsquos largest dam removalSci Rep 8 13279 (2018) doi 101038s41598-018-30817-8pmid 30185796
152 T Sommer et al Californiarsquos Yolo Bypass Evidence thatflood control can be compatible with fisheries wetlandswildlife and agriculture Fisheries 26 6ndash16 (2001)doi 1015771548-8446(2001)026lt0006CYBgt20CO2
153 G Grill et al Mapping the worldrsquos free-flowing riversNature 569 215ndash221 (2019) doi 101038s41586-019-1111-9pmid 31068722
154 G Gassert M Luck M Landis P Reig T Shiao ldquoAqueductglobal maps 21 Constructing decision-relevant global waterrisk indicatorsrdquo Working paper World Resources Institute(2015) wwwwriorgpublicationaqueduct-global-maps-21-indicators
155 A Ruhiacute J D Olden J L Sabo Declining streamflow inducescollapse and replacement of native fishes in the AmericanSouthwest Front Ecol Environ 14 465ndash472 (2016)doi 101002fee1424
156 T C Gouhier A Grinsted V Simko R package Biwaveletconduct univariate and bivariate wavelet analyses (2018)
157 T Rodriacuteguez-Castillo E Esteacutevez A M Gonzaacutelez-FerrerasJ Barquiacuten Estimating ecosystem metabolism toentire river networks Ecosystems (2018) doi 101007s10021-018-0311-8
ACKNOWLEDGMENTS
We greatly appreciate those who read and provided importantinsights on drafts of this paper A Armstrong K HondulaJ Hosen K Leathers G de Mendoza C Maietta I MunozJ Tonkin and two anonymous reviewers We also thank T Tu forhelp with Fig 1 and S Carlson L Comte T Datry J HosenM Kondolf N Jones D Lytle E Moody J Olden M PowerJ Sabo and M Sanchez-Montoya for valuable discussionsCompeting interests The authors have no competing interestsFunding MAP was supported by funds from the NSF (1639145)and AR was supported by funds from the University of CaliforniaBerkeley and NSF (1802714)
101126scienceaaw2087
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 13 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
restorationLinkages between flow regime biota and ecosystem processes Implications for river
Margaret Palmer and Albert Ruhi
DOI 101126scienceaaw2087 (6459) eaaw2087365Science
this issue p eaaw2087Scienceflow variability and achieve hydrological connectivity between a river and its surroundingsand ecosystem processes To be successful such efforts must go beyond accounting for flood pulses to restore natural mimic ecologically important aspects of natural flow regimes guided by insights into how variations in flow affect biotawater quality and ecological processes In a Review Palmer and Ruhi explain how restoration designs now attempt to
Human activities have altered the flow regimes of many of Earths rivers with negative impacts on biodiversityRiver restoration guided by research
ARTICLE TOOLS httpsciencesciencemagorgcontent3656459eaaw2087
REFERENCES
httpsciencesciencemagorgcontent3656459eaaw2087BIBLThis article cites 147 articles 7 of which you can access for free
PERMISSIONS httpwwwsciencemagorghelpreprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAASScienceScience 1200 New York Avenue NW Washington DC 20005 The title (print ISSN 0036-8075 online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
Science No claim to original US Government WorksCopyright copy 2019 The Authors some rights reserved exclusive licensee American Association for the Advancement of
on May 27 2020
httpsciencesciencem
agorgD
ownloaded from

Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 9 of 13
Table 1 Research challenges and opportunities Despite the progress that has been made in understanding how flow regimes affect biota and ecosystem
processes major challenges persist that prevent a complete understanding of the flow-biota-ecosystem processes nexus Making progress on these
challenges requires a mix of fundamental and applied research
Topic Research challenge Opportunity
Connecting
organismal to
ecosystem-level
processes
Research on the effects of flow on organisms
and ecosystem processes has followed parallel
tracksmdashhindering our mechanistic
understanding of the flow-biota-ecosystem
processes nexus
Generate theory that connects concepts and frameworks
that are well accepted in river community and ecosystem
ecology For example the notion of environmental filters
that connect regional to local composition via a combination
of dispersal and environmental conditions (community
ecology) with analogous theory on solute transport versus
reactivity (ecosystem ecology)
Our current understanding of the links between
phenology of biota and ecosystem processes
is precursory
Use high-frequency sensors to characterize metabolic
regimes at sites with ongoing organismal monitoring
Leverage citizen-science biodiversity data to detect
spatiotemporal shifts in animal phenology
(eg insect emergence fish spawning)
Use remote sensing tools to better understand
links between hydrology and aquatic and riparian
plant phenology
The link between stoichiometry and flow
variation remains largely unexplored
Repurpose data that is already being collected to test
flow-stoichiometry relationships eg studies using C and
N stable isotopes to describe CN ratios across flow
regime types Perform field experiments to quantify
indirect effects of invertebrate and fish consumption
on ecosystem metabolism
Ecological
consequences of
hydrologic
connectivity
The importance of anaerobic metabolism in
carbon processing as flows cease and parts of
streambeds dry is a major frontier research area
Create networks of sites across different geologies
and flow regimes that collect time-series data on
methane nitrous oxide and carbon dioxide fluxes
from intermittent streams
We ignore how biota in the wet phase is
influenced by processes and patterns
from the dry phase
Research the functions of dry riverbeds Use camera traps and
drone technology to investigate the role of dry riverbeds
as a dispersal corridor metabarcoding to quantify
ldquodark biodiversityrdquo in the hyporheic
Wetlands and their links to stream networks are poorly
delineated globally Even in the United States and
Europe current wetland inventories do not capture
these links yet understanding how their connectivity
to river systems influences network-scale ecosystem
processes is needed
Analyze time series from high-frequency sensors distributed in
river networks varying in wetland connectivity Further
development and application of time-series methods
used by neuroscientists
We need to identify the relationship between
varying flows and solute dynamics
associated with changes in catchment-scale
hydrologic exchanges
Design collaborative work among catchment hydrologists
and stream ecologists combining methods to estimate
transit times residence times and identify sources
across heterogeneous landscapes Link these properties to
land management and water quality aspects
Scaling and
transferring flow-
ecology relationships
across space and time
We ignore how biodiversity responds to
propagation of flow regime alteration
across river networks
Leverage extensive hydrometric networks (eg United States
Geological Survey National Water Information System)
to perform spatially replicated time-series analyses
on flow and use these to generate expectations
on how river network position should mediate
biodiversity responses to flow alteration
Flow-ecology relationships do not account for
nonstationarity and abrupt change
Use process-based models that account for mechanisms
and associated uncertainties When using time-series
data and models select methods that are robust
to nonstationarity Linear flow-ecology relationships
should not be assumed
Current flow-ecology relationships assume that
responses of species do not change over time
Consider flow regime alteration as a process with ecological and
evolutionary consequences Include intraspecific variation
in traits and potential for rapid evolution in models
continued on next page
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
and streams may improve water quality morethan other restoration options (70) Additionallyresearch on the age and lability of DOC enteringstreams has shown how precipitation or land dis-turbance can change watershed flow paths (53)and this could inspire new restoration approachesfocused on land use and soil properties Manystudies on hydrologic connectivity and networkstructure suggest a need for restoration to focuson landscape heterogeneity especially on thepresence ofwetlands lakes and ponds (144) Suchfeaturesare frequently lost inurbanizedcatchmentsand outcomes could be improved by restoring orincreasing connectivitywith these ecosystems (145)Hydrologic connectivity associated with ground-water inputs or abstractions land-use manage-ment and return flows fromwastewater treatmentplants also need to be considered in these initia-tives (146)Restoring hydrologic connectivity is also key
to achieving biodiversity outcomesmdashparticularlyin dammed systems Longitudinal connectivityfor fish inhabiting dam-altered rivers can beenhanced by the construction of fishways andsophisticated passage facilities However moni-toring data on the effectiveness of such inter-
ventions are still scarce and available evidencesuggests that passage effectivity largely dependson technology used and species-specific fish be-havior (147 148) Although trap-and-transportprograms for migratory fishes can help augmentthreatened populations (149) these practicesoften increase straying and thus the potentialfor genetic homogenization as seen from ju-venile salmon collected at dams and transporteddownstream to increase out-migration success(150) Overall a combination of engineering andmanagement solutions may be best to restorefunctional connectivity in dam-altered riversThe link between hydrologic connectivity andecological restoration is also central to con-siderations on where to build remove and re-operate hydrologic infrastructure Hundredsof dams have been intentionally removed sincethe 1970s and removal of large dams such asthe Elwha River Dam (Washington USA) hasshown that restoring longitudinal connectivitycan have immediate geomorphic effects with com-plex temporal dynamics (151) Infrastructure canalso be designed or reoperated to manage forrestoring lateral connectivity and enhanced flood-plain processes For example the Yolo Bypass in
the Sacramento River California has becomea prime rearing and migration site for chinooksalmon showing that flood control and habitatfor fish and wildlife do not need to be at odds(152) All these examples show the importanceof integrating hydrologic connectivity in ecolog-ical restoration whether the goal is to minimizethe ecological effects of fragmentation restorefree-flowing watercourses or operate green infra-structure to enhance infiltration floodplain con-nectivity or other ecological processes
Research challenges and opportunities
Two decades of concerted efforts to research theeffects of flow regime alteration have tremen-dously advanced our understanding of the waysin which streamflow influences biota and eco-system processes However important challengesremain These aremainly related to the scarcity ofscience connecting organismal to ecosystem-levelprocesses through for example microbial linksthe difficulty of studying watershed-level phys-ical and biogeochemical heterogeneity and itsinfluence on riverine processes and biota thecomplexity of parsing out the direct effects offlow from flow-mediated effects and co-occurring
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 10 of 13
Topic Research challenge Opportunity
Cascading
implications of
hydrologic
alteration
We have a poor understanding of the
interactions among physical drivers
Build on large-scale flow experiments to test hypotheses on the
combined effects of altered flow sediment and thermal
regimes Experiment with multilevel intakes in reservoirs
Promote collection of non-flow variables (eg temperature
water quality parameters) at hydrometric stations
At low flows biota are being exposed to high
concentrations of new classes of pollutants
whose impacts are poorly known especially if
biotic impacts interact with impacts on
ecosystem processes
Design realistic experimental studies to advance knowledge
on the ecological effects of multiple stressors
Collect long-term data at sites varying in contaminant
inputs and hydrologic regimes Link microbial to
organismal and ecosystem-level responses
Using hydroecological
science to inform
river ecosystem
restoration
Environmental flow science largely ignores
human dimensions
Use methods that allow identifying causal networks
in complex socioenvironmental systems Include
human needs and thresholds in research on
flow-ecology relationships
Environmental flows are generally prescribed
at the scale of single infrastructures restoration
designs and assessment are generally at reach scales
Promote coordinated large-scale flow experiments
and restoration designs at the river basin scale
Extend assessment of green infrastructure
effectiveness beyond individual structures to
infrastructure networks and link both
to river outcomes
Environmental flows in impounded rivers
generally target the conservation of some
particular species rather than outcomes at the
food-web or ecosystem level
Advance fundamental research on microbialndash
ecosystem process linkages Expand the notion
of designer flows to food-web structure and
ecosystem services when possible
Environmental flow science has largely
ignored surface waterndashgroundwater
interactions
Expand science to understand the link between
restoration actions groundwater recharge and
inputs to streams Provide science relevant to
the implementation of new legislation on
groundwater management (eg Sustainable
Groundwater Management Act in California)
Uncertainty regarding future
hydroclimate and
river flows is growing
Refine regional climate models Promote research on
the interaction between land-use change and
evapotranspiration Incorporate groundwater
influence in macroscale hydrology models
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
stressors (particularly in urban streams) and thechallenge of predicting outcomeswhen both thephysical and the biological environments arechangingAchieving a predictive understanding of hy-
droecological relationships is essential to design-ing successful self-sustaining restoration actionsin river ecosystems However translating basichydroecological insights into restoration practicerequires understanding the complexities of work-ing within a human-dominated water cycle Al-though it is rarely possible to restore all facetsof a natural flow regime ecologically informeddesigns may help mimic key aspects of the nat-ural flow regime (eg periodic pulse flows) orcreate regimes that are not natural but maximizeoutcomes given socioeconomic constraints InTable 1 we identify persisting and emergingchallenges in hydroecological research We paireach challenge with an opportunity (related toresearch design technology or implementation)We contend that these challenges should beprioritized to advance process-based river eco-system restoration
REFERENCES AND NOTES
1 N L Poff et al The natural flow regime Bioscience 47769ndash784 (1997) doi 1023071313099
2 D A Lytle N L Poff Adaptation to natural flow regimesTrends Ecol Evol 19 94ndash100 (2004) doi 101016jtree200310002 pmid 16701235
3 S Sabater A Elosegi R Ludwig in Multiple Stressors in RiverEcosystems (Elsevier 2019) pp 1ndash22
4 United Nations New UN Decade on Ecosystem Restorationoffers unparalleled opportunity for job creation food securityand addressing climate change (2019) httpswwwunenvironmentorgnews-and-storiespress-releasenew-un-decade-ecosystem-restoration-offers-unparalleled-opportunity
5 A W Lorenz P Haase K Januschke A SundermannD Hering Revisiting restored river reaches - Assessingchange of aquatic and riparian communities afterfive years Sci Total Environ 613-614 1185ndash1195(2018) doi 101016jscitotenv201709188pmid 28954379
6 M A Palmer K L Hondula B J Koch Ecological Restorationof Streams and Rivers Shifting Strategies and ShiftingGoals (2014) vol 45
7 T J Beechie et al Process-based principles for restoringriver ecosystems Bioscience 60 209ndash222 (2010)doi 101525bio20106037
8 S E Bunn A H Arthington Basic principles and ecologicalconsequences of altered flow regimes for aquaticbiodiversity Environ Manage 30 492ndash507 (2002)doi 101007s00267-002-2737-0 pmid 12481916
9 K D Fausch Y Taniguchi S Nakano G D GrossmanC R Townsend Flood disturbance regimes influence rainbowtrout invasion success among five holarctic regions EcolAppl 11 1438ndash1455 (2001) doi 1018901051-0761(2001)011[1438FDRIRT]20CO2
10 M E Power M S Parker W E Dietrich Seasonalreassembly of a river food web Floods droughts andimpacts of fish Ecol Monogr 78 263ndash282 (2008)doi 10189006-09021
11 V Acuntildea A Giorgi I Muntildeoz U R S Uehlinger S SabaterFlow extremes and benthic organic matter shape themetabolism of a headwater Mediterranean streamFreshwater Biol 49 960ndash971 (2004) doi 101111j1365-2427200401239x
12 K Tockner F Malard J V Ward An extension of the floodpulse concept Hydrol Processes 14 2861ndash2883 (2000)doi 1010021099-1085(20001112)141617lt2861AID-HYP124gt30CO2-F
13 J S Perkin et al Fragmentation and dewatering transformGreat Plains stream fish communities Ecol Monogr 8573ndash92 (2015) doi 10189014-01211
14 J L Sabo D M Post Quantifying periodic stochastic andcatastrophic environmental variation Ecol Monogr 7819ndash40 (2008) doi 10189006-13401
15 F Shuai et al Silver carp larva abundance in response toriver flow rate revealed by cross-wavelet modellingEcol Modell 383 98ndash105 (2018) doi 101016jecolmodel201805020
16 J D Tonkin M T Bogan N Bonada B Rios-ToumaD A Lytle Seasonality and predictability shape temporalspecies diversity Ecology 98 1201ndash1216 (2017)doi 101002ecy1761 pmid 28144975
17 N L Poff Beyond the natural flow regime Broadening thehydro‐ecological foundation to meet environmental flowschallenges in a non‐stationary world Freshwater Biol 631011ndash1021 (2018) doi 101111fwb13038
18 A Ruhi X Dong C H McDaniel D P Batzer J L SaboDetrimental effects of a novel flow regime on the functionaltrajectory of an aquatic invertebrate metacommunity GlobChange Biol 24 3749ndash3765 (2018) doi 101111gcb14133pmid 29665147
19 M T Bogan D A Lytle Severe drought drives novelcommunity trajectories in desert stream pools FreshwaterBiol 56 2070ndash2081 (2011) doi 101111j1365-2427201102638x
20 G Woodward et al The effects of climatic fluctuations andextreme events on running water ecosystems Philos TransR Soc London Ser B 371 20150274 (2016) pmid 27114576
21 M A Palmer C M Swan K Nelson P Silver R AlvestadStreambed landscapes Evidence that stream invertebratesrespond to the type and spatial arrangement of patchesLandsc Ecol 15 563ndash576 (2000) doi 101023A1008194130695
22 D Schmera et al Does isolation influence the relative role ofenvironmental and dispersal‐related processes in streamnetworks An empirical test of the network positionhypothesis using multiple taxa Freshwater Biol 63 74ndash85(2018) doi 101111fwb12973
23 R Henriques‐Silva et al A comprehensive examination of thenetwork position hypothesis across multiple rivermetacommunities Ecography 42 284ndash294 (2019)doi 101111ecog03908
24 K L Jaeger J D Olden N A Pelland Climate change poisedto threaten hydrologic connectivity and endemic fishes indryland streams Proc Natl Acad Sci USA 11113894ndash13899 (2014) doi 101073pnas1320890111pmid 25136090
25 S Garbin E A Celegon P Fanton G Botter Hydrologicalcontrols on river network connectivity R Soc Open Sci 6181428 (2019) doi 101098rsos181428 pmid 30891270
26 F Cattaneacuteo B Hugueny N Lamouroux Synchrony in browntrout Salmo trutta population dynamics A lsquoMoran effectrsquo onearly‐life stages Oikos 100 43ndash54 (2003) doi 101034j1600-0706200311912x
27 A Terui et al Metapopulation stability in branching rivernetworks Proc Natl Acad Sci USA 115 E5963ndashE5969(2018) doi 101073pnas1800060115 pmid 29895695
28 P A P Moran The statistical analysis of the Canadian Lynxcycle Aust J Zool 1 291ndash298 (1953) doi 101071ZO9530291
29 G D Grossman P B Moyle J O Whitaker Jr Stochasticityin structural and functional characteristics of an Indianastream fish assemblage A test of community theoryAm Nat 120 423ndash454 (1982) doi 101086284004
30 D A Lytle D M Merritt J D Tonkin J D OldenL V Reynolds Linking river flow regimes to riparian plantguilds A community-wide modeling approach Ecol Appl 271338ndash1350 (2017) doi 101002eap1528 pmid 28263426
31 A Ruhiacute E E Holmes J N Rinne J L Sabo Anomalousdroughts not invasion decrease persistence of native fishesin a desert river Glob Change Biol 21 1482ndash1496 (2015)doi 101111gcb12780 pmid 25430731
32 J D Tonkin D M Merritt J D Olden L V ReynoldsD A Lytle Flow regime alteration degrades ecologicalnetworks in riparian ecosystems Nat Ecol Evol 2 86ndash93(2018) doi 101038s41559-017-0379-0 pmid 29180707
33 M C Mims J D Olden Fish assemblages respond toaltered flow regimes via ecological filtering of lifehistory strategies Freshwater Biol 58 50ndash62 (2013)doi 101111fwb12037
34 P T J Johnson J D Olden M J Vander Zanden Daminvaders Impoundments facilitate biological invasions intofreshwaters Front Ecol Environ 6 357ndash363 (2008)doi 101890070156
35 N Bonada S Doledec B Statzner Taxonomic and biologicaltrait differences of stream macroinvertebrate communitiesbetween mediterranean and temperate regions Implicationsfor future climatic scenarios Glob Change Biol 131658ndash1671 (2007) doi 101111j1365-2486200701375x
36 M I Pyne N L Poff Vulnerability of stream communitycomposition and function to projected thermal warming andhydrologic change across ecoregions in the western UnitedStates Glob Change Biol 23 77ndash93 (2017) doi 101111gcb13437 pmid 27429092
37 D B Herbst S D Cooper R B Medhurst S W WisemanC T Hunsaker Drought ecohydrology alters the structureand function of benthic invertebrate communities inmountain streams Freshwater Biol 64 886ndash902 (2019)doi 101111fwb13270
38 M T Bogan E T Chester T Datry A L MurphyB J Robson A Ruhi R Stubbington J E Whitney inResistance Resilience and Community Recovery in IntermittentRivers and Ephemeral Streams T Datry N Bonada A BoultonEds (Academic Press 2017) pp 349ndash376
39 S Houmlckendorff et al Characterizing fish responses to a riverrestoration over 21 years based on speciesrsquo traits ConservBiol 31 1098ndash1108 (2017) doi 101111cobi12908pmid 28218807
40 D Bruno et al Structural and functional responses ofinvertebrate communities to climate change andflow regulation in alpine catchments Glob Change Biol25 1612ndash1628 (2019) doi 101111gcb14581pmid 30698905
41 T W H Aspin et al Extreme drought pushes streaminvertebrate communities over functional thresholds GlobChange Biol 25 230ndash244 (2019) doi 101111gcb14495pmid 30346098
42 S E Tank J B Fellman E Hood E S Kritzberg Beyondrespiration Controls on lateral carbon fluxes across theterrestrial-aquatic interface Limnol Oceanogr Lett 3 76ndash88(2018) doi 101002lol210065
43 A P Appling et al The metabolic regimes of 356 rivers inthe United States Sci Data 5 180292 (2018) doi 101038sdata2018292 pmid 30532078
44 P Savoy et al Metabolic rhythms in flowing watersAn approach for classifying river productivity regimesLimnol Oceanogr (2019) doi 101002lno11154
45 E S Bernhardt et al The metabolic regimes of flowingwaters Limnol Oceanogr 63 S99ndashS118 (2018)doi 101002lno10726
46 M Colls X Timoner S Sabater V Acuntildea Effects of durationand frequency of the non-flow period on stream metabolismEcosystems 101007s10021-019-00345-1 (2019)doi 101007s10021-019-00345-1
47 A J Ulseth E Bertuzzo G A Singer J Schelker T J BattinClimate-induced changes in spring snowmelt impactecosystem metabolism and carbon fluxes in an Alpine streamnetwork Ecosystems 21 373ndash390 (2018) doi 101007s10021-017-0155-7
48 B O L Demars Hydrological pulses and burning of dissolvedorganic carbon by stream respiration Limnol Oceanogr 64406ndash421 (2019) doi 101002lno11048
49 R T Hensley L Kirk M Spangler M N Gooseff M J CohenFlow extremes as spatiotemporal control points on riversolute fluxes and metabolism J Geophys Res Biogeosci124 537ndash555 (2019) doi 1010292018JG004738
50 J D Hosen et al Enhancement of primary production duringdrought in a temperate watershed is greater in larger riversthan headwater streams Limnol Oceanogr 64 1458ndash1472(2019) doi 101002lno11127
51 L Goacutemez-Gener et al When water vanishes Magnitude andregulation of carbon dioxide emissions from dry temporarystreams Ecosystems 19 710ndash723 (2016) doi 101007s10021-016-9963-4
52 E H Stanley et al The ecology of methane in streams andrivers Patterns controls and global significance EcolMonogr 86 146ndash171 (2016) doi 10189015-1027
53 R T Barnes D E Butman H F Wilson P A RaymondRiverine export of aged carbon driven by flow path depth andresidence time Environ Sci Technol 52 1028ndash1035 (2018)doi 101021acsest7b04717 pmid 29313674
54 G Pinay et al Riparian corridors A new conceptualframework for assessing nitrogen buffering across biomesFront Environ Sci 6 47 (2018) doi 103389fenvs201800047
55 W M Wollheim et al River network saturation conceptFactors influencing the balance of biogeochemical supply and
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 11 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
demand of river networks Biogeochemistry 141 503ndash521(2018) doi 101007s10533-018-0488-0
56 P A Raymond J E Saiers W V Sobczak Hydrological andbiogeochemical controls on watershed dissolved organicmatter transport Pulse-shunt concept Ecology 97 5ndash16(2016) doi 10189014-16841 pmid 27008769
57 C Tunaley D Tetzlaff J Lessels C Soulsby Linking high-frequency DOC dynamics to the age of connected watersources Water Resour Res 52 5232ndash5247 (2016)doi 1010022015WR018419
58 S Bernal et al Supply demand and in-stream retention ofdissolved organic carbon and nitrate during storms inMediterranean forested headwater streams Front EnvironSci 7 60 (2019) doi 103389fenvs201900060
59 J Harvey et al How hydrologic connectivity regulates waterquality in river corridors J Am Water Resour Assoc 55369ndash381 (2019) doi 1011111752-168812691
60 J C Moore et al Detritus trophic dynamics and biodiversityEcol Lett 7 584ndash600 (2004) doi 101111j1461-0248200400606x
61 A D Rosemond et al Freshwater ecology Experimentalnutrient additions accelerate terrestrial carbon loss fromstream ecosystems Science 347 1142ndash1145 (2015)doi 101126scienceaaa1958 pmid 25745171
62 A C Y Yeung J L Musetta-Lambert D P KreutzweiserP K Sibley J S Richardson Relations of interannualdifferences in stream litter breakdown with dischargeBioassessment implications Ecosphere 9 e02423 (2018)doi 101002ecs22423
63 R M Northington J R Webster Experimental reductions instream flow alter litter processing and consumer subsidies inheadwater streams Freshwater Biol 62 737ndash750 (2017)doi 101111fwb12898
64 E Mas-Martiacute I Sanpera-Calbet I Muntildeoz Bottom-up effectsof streambed drying on consumer performance throughchanges in resource quality Aquat Sci 79 719ndash731 (2017)doi 101007s00027-017-0531-6
65 B D Richter G A Thomas Restoring environmental flows bymodifying dam operations Ecol Soc 12 12 (2007)doi 105751ES-02014-120112
66 G Zolezzi A Siviglia M Toffolon B MaioliniThermopeaking in Alpine streams Event characterizationand time scales Ecohydrology 4 564ndash576 (2011)doi 101002eco132
67 T Meiszligner B Sures C K Feld Multiple stressors and therole of hydrology on benthic invertebrates in mountainousstreams Sci Total Environ 663 841ndash851 (2019)doi 101016jscitotenv201901288 pmid 30738264
68 I R Waite et al Effects of urban multi-stressors on threestream biotic assemblages Sci Total Environ 6601472ndash1485 (2019) doi 101016jscitotenv201812240pmid 30743940
69 K C Nelson M A Palmer Stream temperature surges underurbanization and climate change Data models andresponses J Am Water Resour Assoc 43 440ndash452 (2007)doi 101111j1752-1688200700034x
70 J R Blaszczak J M Delesantro Y Zhong D L UrbanE S Bernhardt Watershed urban development controls onurban streamwater chemistry variability Biogeochemistry144 61ndash84 (2019) doi 101007s10533-019-00572-7
71 L Ponsatiacute et al Biofilm responses to flow regulation by damsin Mediterranean rivers River Res Appl 31 1003ndash1016(2015) doi 101002rra2807
72 J M Alberts J J Beaulieu I Buffam Watershed land useand seasonal variation constrain the influence of ripariancanopy cover on stream ecosystem metabolism Ecosystems20 553ndash567 (2017) doi 101007s10021-016-0040-9pmid 30416371
73 R O Hall Jr et al Turbidity light temperature andhydropeaking control primary productivity in the ColoradoRiver Grand Canyon Limnol Oceanogr 60 512ndash526 (2015)doi 101002lno10031
74 J R Blaszczak J M Delesantro D L Urban M W DoyleE S Bernhardt Scoured or suffocated Urban streamecosystems oscillate between hydrologic and dissolvedoxygen extremes Limnol Oceanogr 64 877ndash894 (2019)doi 101002lno11081
75 J-R Mor et al Dam regulation and riverine food-webstructure in a Mediterranean river Sci Total Environ 625301ndash310 (2018) doi 101016jscitotenv201712296pmid 29289778
76 H A Wellard Kelly et al Macroinvertebrate diets reflecttributary inputs and turbidity-driven changes in food
availability in the Colorado River downstream of GlenCanyon Dam Freshw Sci 32 397ndash410 (2013)doi 10189912-0881
77 S J Imberger R M Thompson M R Grace Urbancatchment hydrology overwhelms reach scale effects ofriparian vegetation on organic matter dynamics FreshwaterBiol 56 1370ndash1389 (2011) doi 101111j1365-2427201102575x
78 R T Martins A S Melo J F Gonccedilalves Jr N HamadaLeaf-litter breakdown in urban streams of Central AmazoniaDirect and indirect effects of physical chemical andbiological factors Freshw Sci 34 716ndash726 (2015)doi 101086681086
79 C M Yule J Y Gan T Jinggut K V Lee Urbanizationaffects food webs and leaf-litter decomposition in a tropicalstream in Malaysia Freshw Sci 34 702ndash715 (2015)doi 101086681252
80 T A Newcomer et al Influence of natural and novel organiccarbon sources on denitrification in forest degraded urbanand restored streams Ecol Monogr 82 449ndash466 (2012)doi 10189012-04581
81 A Chaudhary I Kauser A Ray R Poretsky Taxon-drivenfunctional shifts associated with storm flow in an urbanstream microbial community MSphere 3 e00194-18 (2018)doi 101128mSphere00194-18 pmid 29976643
82 T A Kennedy et al Flow management for hydropowerextirpates aquatic insects undermining river food websBioscience 66 561ndash575 (2016) doi 101093bioscibiw059
83 A Glazaczow D Orwin M Bogdziewicz Increasedtemperature delays the late-season phenology of multivoltineinsect Sci Rep 6 38022 (2016) doi 101038srep38022pmid 27905493
84 S G Fisher L J Gray N B Grimm D E Busch Temporalsuccession in a desert stream ecosystem following flashflooding Ecol Monogr 52 93ndash110 (1982) doi 1023072937346
85 G J Scrimgeour M J Winterbourn Effects of floods onepilithon and benthic macroinvertebrate populations in anunstable New Zealand river Hydrobiologia 171 33ndash44 (1989)doi 101007BF00005722
86 E Bondar-Kunze S Maier D Schoumlnauer N Bahl T HeinAntagonistic and synergistic effects on a stream periphytoncommunity under the influence of pulsed flow velocityincrease and nutrient enrichment Sci Total Environ 573594ndash602 (2016) doi 101016jscitotenv201608158pmid 27585428
87 M J Cashman G L Harvey G Wharton M C Bruno Woodmitigates the effect of hydropeaking scour on periphytonbiomass and nutritional quality in semi-natural flumesimulations Aquat Sci 79 459ndash471 (2017) doi 101007s00027-016-0510-3
88 W F Cross et al Food-web dynamics in a large riverdiscontinuum Ecol Monogr 83 311ndash337 (2013)doi 10189012-17271
89 D M Warfe et al Productivity disturbance and ecosystemsize have no influence on food chain length in seasonallyconnected rivers PLOS ONE 8 e66240 (2013)doi 101371journalpone0066240
90 T D Jardine et al Consumer-resource coupling in wet-drytropical rivers J Anim Ecol 81 310ndash322 (2012)doi 101111j1365-2656201101925x pmid 22103689
91 K O Winemiller et al Pulsing hydrology determines top‐down control of basal resources in a tropical riverndashfloodplainecosystem Ecol Monogr 84 621ndash635 (2014)doi 10189013-18221
92 D M Post The long and short of food-chain length TrendsEcol Evol 17 269ndash277 (2002) doi 101016S0169-5347(02)02455-2
93 D M Post G Takimoto Proximate structural mechanismsfor variation in food‐chain length Oikos 116 775ndash782 (2007)doi 101111j0030-1299200715552x
94 P A McHugh A R McIntosh P G Jellyman Dual influencesof ecosystem size and disturbance on food chain length instreams Ecol Lett 13 881ndash890 (2010) doi 101111j1461-0248201001484x pmid 20482579
95 A Ruhiacute et al Flow regulation increases food-chain lengththrough omnivory mechanisms in a Mediterranean rivernetwork Freshwater Biol 61 1536ndash1549 (2016) doi 101111fwb12794
96 J L Sabo J C Finlay T Kennedy D M Post The role ofdischarge variation in scaling of drainage area and food chainlength in rivers Science 330 965ndash967 (2010) doi 101126science1196005 pmid 20947729
97 A R McIntosh et al Capacity to support predators scaleswith habitat size Sci Adv 4 eaap7523 (2018) doi 101126sciadvaap7523 pmid 29978038
98 J H Brown J F Gillooly A P Allen V M Savage G B WestToward a metabolic theory of ecology Ecology 851771ndash1789 (2004) doi 10189003-9000
99 K Zou E Theacutebault G Lacroix S Barot Interactions betweenthe green and brown food web determine ecosystemfunctioning Funct Ecol 30 1454ndash1465 (2016) doi 1011111365-243512626
100 M J Vanni P B McIntyre Predicting nutrient excretion ofaquatic animals with metabolic ecology and ecologicalstoichiometry A global synthesis Ecology 97 3460ndash3471(2016) doi 101002ecy1582 pmid 27912023
101 T B Parr K A Capps S P Inamdar K A Metcalf Animal-mediated organic matter transformation Aquatic insects as asource of microbially bioavailable organic nutrients andenergy Funct Ecol 33 524ndash535 (2019) doi 1011111365-243513242
102 C P Arango et al Urban infrastructure influences dissolvedorganic matter quality and bacterial metabolism in an urbanstream network Freshwater Biol 62 1917ndash1928 (2017)
103 L G Larsen J W Harvey Disrupted carbon cycling inrestored and unrestored urban streams Critical timescalesand controls Limnol Oceanogr 62 S160ndashS182 (2017)doi 101002lno10613
104 H M Halvorson G White J T Scott M A Evans-WhiteDietary and taxonomic controls on incorporation of microbialcarbon and phosphorus by detritivorous caddisfliesOecologia 180 567ndash579 (2016) doi 101007s00442-015-3464-6 pmid 26497125
105 J J Elser et al Biological stoichiometry from genes toecosystems Ecol Lett 3 540ndash550 (2000) doi 101046j1461-0248200000185x
106 E K Moody et al Consumption explains intraspecificvariation in nutrient recycling stoichiometry in a desert fishEcology 99 1552ndash1561 (2018) doi 101002ecy2372pmid 29882955
107 N Welti et al Bridging food webs ecosystem metabolismand biogeochemistry using ecological stoichiometry theoryFront Microbiol 8 1298 (2017) doi 103389fmicb201701298 pmid 28747904
108 M A Palmer S Filoso R M Fanelli From ecosystems toecosystem services Stream restoration as ecologicalengineering Ecol Eng 65 62ndash70 (2014) doi 101016jecoleng201307059
109 M J Burns T D Fletcher C J Walsh A R LadsonB E Hatt Hydrologic shortcomings of conventional urbanstormwater management and opportunities for reformLandsc Urban Plan 105 230ndash240 (2012) doi 101016jlandurbplan201112012
110 C R Liermann C Nilsson J Robertson R Y NgImplications of dam obstruction for global freshwater fishdiversity Bioscience 62 539ndash548 (2012) doi 101525bio20126265
111 J D Olden R J Naiman Incorporating thermal regimes intoenvironmental flows assessments Modifying dam operationsto restore freshwater ecosystem integrity Freshwater Biol55 86ndash107 (2010) doi 101111j1365-2427200902179x
112 D B Osmundson R J Ryel V L Lamarra J Pitlick Flowndashsedimentndashbiota relations Implications for river regulationeffects on native fish abundance Ecol Appl 12 1719ndash1739(2002)
113 D von Schiller et al Regulation causes nitrogen cyclingdiscontinuities in Mediterranean rivers Sci Total Environ540 168ndash177 (2016) doi 101016jscitotenv201507017pmid 26233271
114 J L Sabo et al Designing river flows to improve foodsecurity futures in the Lower Mekong Basin Science 358eaao1053 (2017) doi 101126scienceaao1053pmid 29217541
115 W Xu et al Improving the multi-objective performance ofrainwater harvesting systems using real-time controltechnology Water 10 147 (2018) doi 103390w10020147
116 M Acreman et al Environmental flows for naturalhybrid and novel riverine ecosystems in a changingworld Front Ecol Environ 12 466ndash473 (2014)doi 101890130134
117 N L Poff et al Sustainable water management under futureuncertainty with eco-engineering decision scaling Nat ClimChang 6 25ndash34 (2016) doi 101038nclimate2765
118 J D Kiernan P B Moyle P K Crain Restoring native fishassemblages to a regulated California stream using the
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 12 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
natural flow regime concept Ecol Appl 22 1472ndash1482(2012) doi 10189011-04801 pmid 22908707
119 D L Propst K B Gido Responses of native andnonnative fishes to natural flow regime mimicry in theSan Juan River Trans Am Fish Soc 133 922ndash931 (2004)doi 101577T03-0571
120 T K Pool J D Olden Assessing long‐term fish responses andshort‐term solutions to flow regulation in a dryland river basinEcol Freshwat Fish 24 56ndash66 (2015) doi 101111eff12125
121 W Chen J D Olden Designing flows to resolve human andenvironmental water needs in a dam-regulated riverNat Commun 8 2158 (2017) doi 101038s41467-017-02226-4 pmid 29255194
122 S M Nelson et al A history of the 2014 Minute 319environmental pulse flow as documented by fieldmeasurements and satellite imagery Ecol Eng 106733ndash748 (2017) doi 101016jecoleng201610040
123 P B Shafroth et al A large-scale environmental flowexperiment for riparian restoration in the Colorado RiverDelta Ecol Eng 106 645ndash660 (2017) doi 101016jecoleng201702016
124 E P Glenn P L Nagler P B Shafroth C J JarchowEffectiveness of environmental flows for riparian restorationin arid regions A tale of four rivers Ecol Eng 106 695ndash703(2017) doi 101016jecoleng201701009
125 S Hotaling D S Finn J Joseph Giersch D W WeisrockD Jacobsen Climate change and alpine stream biologyProgress challenges and opportunities for the futureBiol Rev Camb Philos Soc 92 2024ndash2045 (2017)doi 101111brv12319 pmid 28105701
126 C J Walsh T D Fletcher M J Burns Urban stormwaterrunoff A new class of environmental flow problem PLOS ONE7 e45814 (2012) doi 101371journalpone0045814pmid 23029257
127 A Selvakumar T P OrsquoConnor S D Struck Role of streamrestoration on improving benthic macroinvertebrates and in-stream water quality in an urban watershed Case studyJ Environ Eng 136 127ndash139 (2010) doi 101061(ASCE)EE1943-78700000116
128 R W Lammers B P Bledsoe What role does streamrestoration play in nutrient management Crit Rev EnvironSci Technol 47 335ndash371 (2017) doi 1010801064338920171318618
129 H E Golden N Hoghooghi Green infrastructure and itscatchment-scale effects An emerging science WIREs Water5 e1254 (2018) doi 101002wat21254 pmid 29682288
130 M R Williams G Bhatt S Filoso G Yactayo Streamrestoration performance and its contribution to theChesapeake Bay TMDL Challenges posed by climate changein urban areas Estuaries Coasts 40 1227ndash1246 (2017)doi 101007s12237-017-0226-1
131 R M Fanelli K Prestegaard M A Palmer Freshw Sci101086706072 (2019)
132 B W Abbott et al Unexpected spatial stability of waterchemistry in headwater stream networks Ecol Lett 21296ndash308 (2018) doi 101111ele12897 pmid 29282860
133 B L Brown C M Swan Dendritic network structureconstrains metacommunity properties in riverineecosystems J Anim Ecol 79 571ndash580 (2010) doi 101111j1365-2656201001668x pmid 20180874
134 C M Swan B L Brown Metacommunity theory meetsrestoration Isolation may mediate how ecologicalcommunities respond to stream restoration Ecol Appl 272209ndash2219 (2017) doi 101002eap1602 pmid 28718193
135 N Mouquet D Gravel F Massol V Calcagno Extending theconcept of keystone species to communities and ecosystemsEcol Lett 16 1ndash8 (2013) doi 101111ele12014 pmid 23062191
136 A Ruhiacute T Datry J L Sabo Interpreting beta-diversitycomponents over time to conserve metacommunities inhighly dynamic ecosystems Conserv Biol 31 1459ndash1468(2017) doi 101111cobi12906 pmid 28188969
137 L Yamane L W Botsford D P Kilduff Tracking restorationof population diversity via the portfolio effect J Appl Ecol55 472ndash481 (2018) doi 1011111365-266412978
138 D W Braidwood M A Taggart M Smith R AndersenTranslocations conservation and climate change Use ofrestoration sites as protorefuges and protorefugia RestorEcol 26 20ndash28 (2018) doi 101111rec12642
139 Y Wang D Wang Q W Lewis J Wu F Huang A frameworkto assess the cumulative impacts of dams on hydrologicalregime A case study of the Yangtze River Hydrol Processes31 3045ndash3055 (2017) doi 101002hyp11239
140 J L Sabo et al Pulsed flows tributary inputs and food-webstructure in a highly regulated river J Appl Ecol 551884ndash1895 (2018) doi 1011111365-266413109
141 C Magliozzi G Coro R C Grabowski A I PackmanS Krause A multiscale statistical method to identifypotential areas of hyporheic exchange for river restorationplanning Environ Model Softw 111 311ndash323 (2019)doi 101016jenvsoft201809006
142 S S Schwartz B Smith Restoring hydrologic function inurban landscapes with suburban subsoiling J Hydrol 543770ndash781 (2016) doi 101016jjhydrol201610051
143 C J Walsh T D Fletcher A R Ladson Stream restoration inurban catchments through redesigning stormwater systemsLooking to the catchment to save the stream J N AmBenthol Soc 24 690ndash705 (2005) doi 10189904-0201
144 N M Schmadel et al Thresholds of lake and reservoirconnectivity in river networks control nitrogen removalNat Commun 9 2779 (2018) doi 101038s41467-018-05156-x pmid 30018449
145 C D Sayer Conservation of aquatic landscapes Pondslakes and rivers as integrated systems WIREs Water 1573ndash585 (2014) doi 101002wat21045
146 R G Luthy D L Sedlak M H Plumlee D Austin V H ReshWastewater‐effluent‐dominated streams as ecosystem‐
management tools in a drier climate Front Ecol Environ 13477ndash485 (2015) doi 101890150038
147 D W Roscoe S G Hinch Effectiveness monitoring of fishpassage facilities Historical trends geographic patterns andfuture directions Fish Fish 11 12ndash33 (2010) doi 101111j1467-2979200900333x
148 C M Bunt T Castro-Santos A Haro Performanceof fish passage structures at upstream barriers tomigration River Res Appl 28 457ndash478 (2012)doi 101002rra1565
149 D Weigel et al Evaluation of a trap-and-transport programfor a threatened population of steelhead (Oncorhynchusmykiss) Conserv Genet 20 1195ndash1199 (2019) doi 101007s10592-019-01200-5
150 M H Bond et al Combined effects of barge transportationriver environment and rearing location on straying andmigration of adult Snake River fall-run Chinook SalmonTrans Am Fish Soc 146 60ndash73 (2017) doi 1010800002848720161235614
151 A C Ritchie et al Morphodynamic evolution followingsediment release from the worldrsquos largest dam removalSci Rep 8 13279 (2018) doi 101038s41598-018-30817-8pmid 30185796
152 T Sommer et al Californiarsquos Yolo Bypass Evidence thatflood control can be compatible with fisheries wetlandswildlife and agriculture Fisheries 26 6ndash16 (2001)doi 1015771548-8446(2001)026lt0006CYBgt20CO2
153 G Grill et al Mapping the worldrsquos free-flowing riversNature 569 215ndash221 (2019) doi 101038s41586-019-1111-9pmid 31068722
154 G Gassert M Luck M Landis P Reig T Shiao ldquoAqueductglobal maps 21 Constructing decision-relevant global waterrisk indicatorsrdquo Working paper World Resources Institute(2015) wwwwriorgpublicationaqueduct-global-maps-21-indicators
155 A Ruhiacute J D Olden J L Sabo Declining streamflow inducescollapse and replacement of native fishes in the AmericanSouthwest Front Ecol Environ 14 465ndash472 (2016)doi 101002fee1424
156 T C Gouhier A Grinsted V Simko R package Biwaveletconduct univariate and bivariate wavelet analyses (2018)
157 T Rodriacuteguez-Castillo E Esteacutevez A M Gonzaacutelez-FerrerasJ Barquiacuten Estimating ecosystem metabolism toentire river networks Ecosystems (2018) doi 101007s10021-018-0311-8
ACKNOWLEDGMENTS
We greatly appreciate those who read and provided importantinsights on drafts of this paper A Armstrong K HondulaJ Hosen K Leathers G de Mendoza C Maietta I MunozJ Tonkin and two anonymous reviewers We also thank T Tu forhelp with Fig 1 and S Carlson L Comte T Datry J HosenM Kondolf N Jones D Lytle E Moody J Olden M PowerJ Sabo and M Sanchez-Montoya for valuable discussionsCompeting interests The authors have no competing interestsFunding MAP was supported by funds from the NSF (1639145)and AR was supported by funds from the University of CaliforniaBerkeley and NSF (1802714)
101126scienceaaw2087
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 13 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
restorationLinkages between flow regime biota and ecosystem processes Implications for river
Margaret Palmer and Albert Ruhi
DOI 101126scienceaaw2087 (6459) eaaw2087365Science
this issue p eaaw2087Scienceflow variability and achieve hydrological connectivity between a river and its surroundingsand ecosystem processes To be successful such efforts must go beyond accounting for flood pulses to restore natural mimic ecologically important aspects of natural flow regimes guided by insights into how variations in flow affect biotawater quality and ecological processes In a Review Palmer and Ruhi explain how restoration designs now attempt to
Human activities have altered the flow regimes of many of Earths rivers with negative impacts on biodiversityRiver restoration guided by research
ARTICLE TOOLS httpsciencesciencemagorgcontent3656459eaaw2087
REFERENCES
httpsciencesciencemagorgcontent3656459eaaw2087BIBLThis article cites 147 articles 7 of which you can access for free
PERMISSIONS httpwwwsciencemagorghelpreprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAASScienceScience 1200 New York Avenue NW Washington DC 20005 The title (print ISSN 0036-8075 online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
Science No claim to original US Government WorksCopyright copy 2019 The Authors some rights reserved exclusive licensee American Association for the Advancement of
on May 27 2020
httpsciencesciencem
agorgD
ownloaded from

and streams may improve water quality morethan other restoration options (70) Additionallyresearch on the age and lability of DOC enteringstreams has shown how precipitation or land dis-turbance can change watershed flow paths (53)and this could inspire new restoration approachesfocused on land use and soil properties Manystudies on hydrologic connectivity and networkstructure suggest a need for restoration to focuson landscape heterogeneity especially on thepresence ofwetlands lakes and ponds (144) Suchfeaturesare frequently lost inurbanizedcatchmentsand outcomes could be improved by restoring orincreasing connectivitywith these ecosystems (145)Hydrologic connectivity associated with ground-water inputs or abstractions land-use manage-ment and return flows fromwastewater treatmentplants also need to be considered in these initia-tives (146)Restoring hydrologic connectivity is also key
to achieving biodiversity outcomesmdashparticularlyin dammed systems Longitudinal connectivityfor fish inhabiting dam-altered rivers can beenhanced by the construction of fishways andsophisticated passage facilities However moni-toring data on the effectiveness of such inter-
ventions are still scarce and available evidencesuggests that passage effectivity largely dependson technology used and species-specific fish be-havior (147 148) Although trap-and-transportprograms for migratory fishes can help augmentthreatened populations (149) these practicesoften increase straying and thus the potentialfor genetic homogenization as seen from ju-venile salmon collected at dams and transporteddownstream to increase out-migration success(150) Overall a combination of engineering andmanagement solutions may be best to restorefunctional connectivity in dam-altered riversThe link between hydrologic connectivity andecological restoration is also central to con-siderations on where to build remove and re-operate hydrologic infrastructure Hundredsof dams have been intentionally removed sincethe 1970s and removal of large dams such asthe Elwha River Dam (Washington USA) hasshown that restoring longitudinal connectivitycan have immediate geomorphic effects with com-plex temporal dynamics (151) Infrastructure canalso be designed or reoperated to manage forrestoring lateral connectivity and enhanced flood-plain processes For example the Yolo Bypass in
the Sacramento River California has becomea prime rearing and migration site for chinooksalmon showing that flood control and habitatfor fish and wildlife do not need to be at odds(152) All these examples show the importanceof integrating hydrologic connectivity in ecolog-ical restoration whether the goal is to minimizethe ecological effects of fragmentation restorefree-flowing watercourses or operate green infra-structure to enhance infiltration floodplain con-nectivity or other ecological processes
Research challenges and opportunities
Two decades of concerted efforts to research theeffects of flow regime alteration have tremen-dously advanced our understanding of the waysin which streamflow influences biota and eco-system processes However important challengesremain These aremainly related to the scarcity ofscience connecting organismal to ecosystem-levelprocesses through for example microbial linksthe difficulty of studying watershed-level phys-ical and biogeochemical heterogeneity and itsinfluence on riverine processes and biota thecomplexity of parsing out the direct effects offlow from flow-mediated effects and co-occurring
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 10 of 13
Topic Research challenge Opportunity
Cascading
implications of
hydrologic
alteration
We have a poor understanding of the
interactions among physical drivers
Build on large-scale flow experiments to test hypotheses on the
combined effects of altered flow sediment and thermal
regimes Experiment with multilevel intakes in reservoirs
Promote collection of non-flow variables (eg temperature
water quality parameters) at hydrometric stations
At low flows biota are being exposed to high
concentrations of new classes of pollutants
whose impacts are poorly known especially if
biotic impacts interact with impacts on
ecosystem processes
Design realistic experimental studies to advance knowledge
on the ecological effects of multiple stressors
Collect long-term data at sites varying in contaminant
inputs and hydrologic regimes Link microbial to
organismal and ecosystem-level responses
Using hydroecological
science to inform
river ecosystem
restoration
Environmental flow science largely ignores
human dimensions
Use methods that allow identifying causal networks
in complex socioenvironmental systems Include
human needs and thresholds in research on
flow-ecology relationships
Environmental flows are generally prescribed
at the scale of single infrastructures restoration
designs and assessment are generally at reach scales
Promote coordinated large-scale flow experiments
and restoration designs at the river basin scale
Extend assessment of green infrastructure
effectiveness beyond individual structures to
infrastructure networks and link both
to river outcomes
Environmental flows in impounded rivers
generally target the conservation of some
particular species rather than outcomes at the
food-web or ecosystem level
Advance fundamental research on microbialndash
ecosystem process linkages Expand the notion
of designer flows to food-web structure and
ecosystem services when possible
Environmental flow science has largely
ignored surface waterndashgroundwater
interactions
Expand science to understand the link between
restoration actions groundwater recharge and
inputs to streams Provide science relevant to
the implementation of new legislation on
groundwater management (eg Sustainable
Groundwater Management Act in California)
Uncertainty regarding future
hydroclimate and
river flows is growing
Refine regional climate models Promote research on
the interaction between land-use change and
evapotranspiration Incorporate groundwater
influence in macroscale hydrology models
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
stressors (particularly in urban streams) and thechallenge of predicting outcomeswhen both thephysical and the biological environments arechangingAchieving a predictive understanding of hy-
droecological relationships is essential to design-ing successful self-sustaining restoration actionsin river ecosystems However translating basichydroecological insights into restoration practicerequires understanding the complexities of work-ing within a human-dominated water cycle Al-though it is rarely possible to restore all facetsof a natural flow regime ecologically informeddesigns may help mimic key aspects of the nat-ural flow regime (eg periodic pulse flows) orcreate regimes that are not natural but maximizeoutcomes given socioeconomic constraints InTable 1 we identify persisting and emergingchallenges in hydroecological research We paireach challenge with an opportunity (related toresearch design technology or implementation)We contend that these challenges should beprioritized to advance process-based river eco-system restoration
REFERENCES AND NOTES
1 N L Poff et al The natural flow regime Bioscience 47769ndash784 (1997) doi 1023071313099
2 D A Lytle N L Poff Adaptation to natural flow regimesTrends Ecol Evol 19 94ndash100 (2004) doi 101016jtree200310002 pmid 16701235
3 S Sabater A Elosegi R Ludwig in Multiple Stressors in RiverEcosystems (Elsevier 2019) pp 1ndash22
4 United Nations New UN Decade on Ecosystem Restorationoffers unparalleled opportunity for job creation food securityand addressing climate change (2019) httpswwwunenvironmentorgnews-and-storiespress-releasenew-un-decade-ecosystem-restoration-offers-unparalleled-opportunity
5 A W Lorenz P Haase K Januschke A SundermannD Hering Revisiting restored river reaches - Assessingchange of aquatic and riparian communities afterfive years Sci Total Environ 613-614 1185ndash1195(2018) doi 101016jscitotenv201709188pmid 28954379
6 M A Palmer K L Hondula B J Koch Ecological Restorationof Streams and Rivers Shifting Strategies and ShiftingGoals (2014) vol 45
7 T J Beechie et al Process-based principles for restoringriver ecosystems Bioscience 60 209ndash222 (2010)doi 101525bio20106037
8 S E Bunn A H Arthington Basic principles and ecologicalconsequences of altered flow regimes for aquaticbiodiversity Environ Manage 30 492ndash507 (2002)doi 101007s00267-002-2737-0 pmid 12481916
9 K D Fausch Y Taniguchi S Nakano G D GrossmanC R Townsend Flood disturbance regimes influence rainbowtrout invasion success among five holarctic regions EcolAppl 11 1438ndash1455 (2001) doi 1018901051-0761(2001)011[1438FDRIRT]20CO2
10 M E Power M S Parker W E Dietrich Seasonalreassembly of a river food web Floods droughts andimpacts of fish Ecol Monogr 78 263ndash282 (2008)doi 10189006-09021
11 V Acuntildea A Giorgi I Muntildeoz U R S Uehlinger S SabaterFlow extremes and benthic organic matter shape themetabolism of a headwater Mediterranean streamFreshwater Biol 49 960ndash971 (2004) doi 101111j1365-2427200401239x
12 K Tockner F Malard J V Ward An extension of the floodpulse concept Hydrol Processes 14 2861ndash2883 (2000)doi 1010021099-1085(20001112)141617lt2861AID-HYP124gt30CO2-F
13 J S Perkin et al Fragmentation and dewatering transformGreat Plains stream fish communities Ecol Monogr 8573ndash92 (2015) doi 10189014-01211
14 J L Sabo D M Post Quantifying periodic stochastic andcatastrophic environmental variation Ecol Monogr 7819ndash40 (2008) doi 10189006-13401
15 F Shuai et al Silver carp larva abundance in response toriver flow rate revealed by cross-wavelet modellingEcol Modell 383 98ndash105 (2018) doi 101016jecolmodel201805020
16 J D Tonkin M T Bogan N Bonada B Rios-ToumaD A Lytle Seasonality and predictability shape temporalspecies diversity Ecology 98 1201ndash1216 (2017)doi 101002ecy1761 pmid 28144975
17 N L Poff Beyond the natural flow regime Broadening thehydro‐ecological foundation to meet environmental flowschallenges in a non‐stationary world Freshwater Biol 631011ndash1021 (2018) doi 101111fwb13038
18 A Ruhi X Dong C H McDaniel D P Batzer J L SaboDetrimental effects of a novel flow regime on the functionaltrajectory of an aquatic invertebrate metacommunity GlobChange Biol 24 3749ndash3765 (2018) doi 101111gcb14133pmid 29665147
19 M T Bogan D A Lytle Severe drought drives novelcommunity trajectories in desert stream pools FreshwaterBiol 56 2070ndash2081 (2011) doi 101111j1365-2427201102638x
20 G Woodward et al The effects of climatic fluctuations andextreme events on running water ecosystems Philos TransR Soc London Ser B 371 20150274 (2016) pmid 27114576
21 M A Palmer C M Swan K Nelson P Silver R AlvestadStreambed landscapes Evidence that stream invertebratesrespond to the type and spatial arrangement of patchesLandsc Ecol 15 563ndash576 (2000) doi 101023A1008194130695
22 D Schmera et al Does isolation influence the relative role ofenvironmental and dispersal‐related processes in streamnetworks An empirical test of the network positionhypothesis using multiple taxa Freshwater Biol 63 74ndash85(2018) doi 101111fwb12973
23 R Henriques‐Silva et al A comprehensive examination of thenetwork position hypothesis across multiple rivermetacommunities Ecography 42 284ndash294 (2019)doi 101111ecog03908
24 K L Jaeger J D Olden N A Pelland Climate change poisedto threaten hydrologic connectivity and endemic fishes indryland streams Proc Natl Acad Sci USA 11113894ndash13899 (2014) doi 101073pnas1320890111pmid 25136090
25 S Garbin E A Celegon P Fanton G Botter Hydrologicalcontrols on river network connectivity R Soc Open Sci 6181428 (2019) doi 101098rsos181428 pmid 30891270
26 F Cattaneacuteo B Hugueny N Lamouroux Synchrony in browntrout Salmo trutta population dynamics A lsquoMoran effectrsquo onearly‐life stages Oikos 100 43ndash54 (2003) doi 101034j1600-0706200311912x
27 A Terui et al Metapopulation stability in branching rivernetworks Proc Natl Acad Sci USA 115 E5963ndashE5969(2018) doi 101073pnas1800060115 pmid 29895695
28 P A P Moran The statistical analysis of the Canadian Lynxcycle Aust J Zool 1 291ndash298 (1953) doi 101071ZO9530291
29 G D Grossman P B Moyle J O Whitaker Jr Stochasticityin structural and functional characteristics of an Indianastream fish assemblage A test of community theoryAm Nat 120 423ndash454 (1982) doi 101086284004
30 D A Lytle D M Merritt J D Tonkin J D OldenL V Reynolds Linking river flow regimes to riparian plantguilds A community-wide modeling approach Ecol Appl 271338ndash1350 (2017) doi 101002eap1528 pmid 28263426
31 A Ruhiacute E E Holmes J N Rinne J L Sabo Anomalousdroughts not invasion decrease persistence of native fishesin a desert river Glob Change Biol 21 1482ndash1496 (2015)doi 101111gcb12780 pmid 25430731
32 J D Tonkin D M Merritt J D Olden L V ReynoldsD A Lytle Flow regime alteration degrades ecologicalnetworks in riparian ecosystems Nat Ecol Evol 2 86ndash93(2018) doi 101038s41559-017-0379-0 pmid 29180707
33 M C Mims J D Olden Fish assemblages respond toaltered flow regimes via ecological filtering of lifehistory strategies Freshwater Biol 58 50ndash62 (2013)doi 101111fwb12037
34 P T J Johnson J D Olden M J Vander Zanden Daminvaders Impoundments facilitate biological invasions intofreshwaters Front Ecol Environ 6 357ndash363 (2008)doi 101890070156
35 N Bonada S Doledec B Statzner Taxonomic and biologicaltrait differences of stream macroinvertebrate communitiesbetween mediterranean and temperate regions Implicationsfor future climatic scenarios Glob Change Biol 131658ndash1671 (2007) doi 101111j1365-2486200701375x
36 M I Pyne N L Poff Vulnerability of stream communitycomposition and function to projected thermal warming andhydrologic change across ecoregions in the western UnitedStates Glob Change Biol 23 77ndash93 (2017) doi 101111gcb13437 pmid 27429092
37 D B Herbst S D Cooper R B Medhurst S W WisemanC T Hunsaker Drought ecohydrology alters the structureand function of benthic invertebrate communities inmountain streams Freshwater Biol 64 886ndash902 (2019)doi 101111fwb13270
38 M T Bogan E T Chester T Datry A L MurphyB J Robson A Ruhi R Stubbington J E Whitney inResistance Resilience and Community Recovery in IntermittentRivers and Ephemeral Streams T Datry N Bonada A BoultonEds (Academic Press 2017) pp 349ndash376
39 S Houmlckendorff et al Characterizing fish responses to a riverrestoration over 21 years based on speciesrsquo traits ConservBiol 31 1098ndash1108 (2017) doi 101111cobi12908pmid 28218807
40 D Bruno et al Structural and functional responses ofinvertebrate communities to climate change andflow regulation in alpine catchments Glob Change Biol25 1612ndash1628 (2019) doi 101111gcb14581pmid 30698905
41 T W H Aspin et al Extreme drought pushes streaminvertebrate communities over functional thresholds GlobChange Biol 25 230ndash244 (2019) doi 101111gcb14495pmid 30346098
42 S E Tank J B Fellman E Hood E S Kritzberg Beyondrespiration Controls on lateral carbon fluxes across theterrestrial-aquatic interface Limnol Oceanogr Lett 3 76ndash88(2018) doi 101002lol210065
43 A P Appling et al The metabolic regimes of 356 rivers inthe United States Sci Data 5 180292 (2018) doi 101038sdata2018292 pmid 30532078
44 P Savoy et al Metabolic rhythms in flowing watersAn approach for classifying river productivity regimesLimnol Oceanogr (2019) doi 101002lno11154
45 E S Bernhardt et al The metabolic regimes of flowingwaters Limnol Oceanogr 63 S99ndashS118 (2018)doi 101002lno10726
46 M Colls X Timoner S Sabater V Acuntildea Effects of durationand frequency of the non-flow period on stream metabolismEcosystems 101007s10021-019-00345-1 (2019)doi 101007s10021-019-00345-1
47 A J Ulseth E Bertuzzo G A Singer J Schelker T J BattinClimate-induced changes in spring snowmelt impactecosystem metabolism and carbon fluxes in an Alpine streamnetwork Ecosystems 21 373ndash390 (2018) doi 101007s10021-017-0155-7
48 B O L Demars Hydrological pulses and burning of dissolvedorganic carbon by stream respiration Limnol Oceanogr 64406ndash421 (2019) doi 101002lno11048
49 R T Hensley L Kirk M Spangler M N Gooseff M J CohenFlow extremes as spatiotemporal control points on riversolute fluxes and metabolism J Geophys Res Biogeosci124 537ndash555 (2019) doi 1010292018JG004738
50 J D Hosen et al Enhancement of primary production duringdrought in a temperate watershed is greater in larger riversthan headwater streams Limnol Oceanogr 64 1458ndash1472(2019) doi 101002lno11127
51 L Goacutemez-Gener et al When water vanishes Magnitude andregulation of carbon dioxide emissions from dry temporarystreams Ecosystems 19 710ndash723 (2016) doi 101007s10021-016-9963-4
52 E H Stanley et al The ecology of methane in streams andrivers Patterns controls and global significance EcolMonogr 86 146ndash171 (2016) doi 10189015-1027
53 R T Barnes D E Butman H F Wilson P A RaymondRiverine export of aged carbon driven by flow path depth andresidence time Environ Sci Technol 52 1028ndash1035 (2018)doi 101021acsest7b04717 pmid 29313674
54 G Pinay et al Riparian corridors A new conceptualframework for assessing nitrogen buffering across biomesFront Environ Sci 6 47 (2018) doi 103389fenvs201800047
55 W M Wollheim et al River network saturation conceptFactors influencing the balance of biogeochemical supply and
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 11 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
demand of river networks Biogeochemistry 141 503ndash521(2018) doi 101007s10533-018-0488-0
56 P A Raymond J E Saiers W V Sobczak Hydrological andbiogeochemical controls on watershed dissolved organicmatter transport Pulse-shunt concept Ecology 97 5ndash16(2016) doi 10189014-16841 pmid 27008769
57 C Tunaley D Tetzlaff J Lessels C Soulsby Linking high-frequency DOC dynamics to the age of connected watersources Water Resour Res 52 5232ndash5247 (2016)doi 1010022015WR018419
58 S Bernal et al Supply demand and in-stream retention ofdissolved organic carbon and nitrate during storms inMediterranean forested headwater streams Front EnvironSci 7 60 (2019) doi 103389fenvs201900060
59 J Harvey et al How hydrologic connectivity regulates waterquality in river corridors J Am Water Resour Assoc 55369ndash381 (2019) doi 1011111752-168812691
60 J C Moore et al Detritus trophic dynamics and biodiversityEcol Lett 7 584ndash600 (2004) doi 101111j1461-0248200400606x
61 A D Rosemond et al Freshwater ecology Experimentalnutrient additions accelerate terrestrial carbon loss fromstream ecosystems Science 347 1142ndash1145 (2015)doi 101126scienceaaa1958 pmid 25745171
62 A C Y Yeung J L Musetta-Lambert D P KreutzweiserP K Sibley J S Richardson Relations of interannualdifferences in stream litter breakdown with dischargeBioassessment implications Ecosphere 9 e02423 (2018)doi 101002ecs22423
63 R M Northington J R Webster Experimental reductions instream flow alter litter processing and consumer subsidies inheadwater streams Freshwater Biol 62 737ndash750 (2017)doi 101111fwb12898
64 E Mas-Martiacute I Sanpera-Calbet I Muntildeoz Bottom-up effectsof streambed drying on consumer performance throughchanges in resource quality Aquat Sci 79 719ndash731 (2017)doi 101007s00027-017-0531-6
65 B D Richter G A Thomas Restoring environmental flows bymodifying dam operations Ecol Soc 12 12 (2007)doi 105751ES-02014-120112
66 G Zolezzi A Siviglia M Toffolon B MaioliniThermopeaking in Alpine streams Event characterizationand time scales Ecohydrology 4 564ndash576 (2011)doi 101002eco132
67 T Meiszligner B Sures C K Feld Multiple stressors and therole of hydrology on benthic invertebrates in mountainousstreams Sci Total Environ 663 841ndash851 (2019)doi 101016jscitotenv201901288 pmid 30738264
68 I R Waite et al Effects of urban multi-stressors on threestream biotic assemblages Sci Total Environ 6601472ndash1485 (2019) doi 101016jscitotenv201812240pmid 30743940
69 K C Nelson M A Palmer Stream temperature surges underurbanization and climate change Data models andresponses J Am Water Resour Assoc 43 440ndash452 (2007)doi 101111j1752-1688200700034x
70 J R Blaszczak J M Delesantro Y Zhong D L UrbanE S Bernhardt Watershed urban development controls onurban streamwater chemistry variability Biogeochemistry144 61ndash84 (2019) doi 101007s10533-019-00572-7
71 L Ponsatiacute et al Biofilm responses to flow regulation by damsin Mediterranean rivers River Res Appl 31 1003ndash1016(2015) doi 101002rra2807
72 J M Alberts J J Beaulieu I Buffam Watershed land useand seasonal variation constrain the influence of ripariancanopy cover on stream ecosystem metabolism Ecosystems20 553ndash567 (2017) doi 101007s10021-016-0040-9pmid 30416371
73 R O Hall Jr et al Turbidity light temperature andhydropeaking control primary productivity in the ColoradoRiver Grand Canyon Limnol Oceanogr 60 512ndash526 (2015)doi 101002lno10031
74 J R Blaszczak J M Delesantro D L Urban M W DoyleE S Bernhardt Scoured or suffocated Urban streamecosystems oscillate between hydrologic and dissolvedoxygen extremes Limnol Oceanogr 64 877ndash894 (2019)doi 101002lno11081
75 J-R Mor et al Dam regulation and riverine food-webstructure in a Mediterranean river Sci Total Environ 625301ndash310 (2018) doi 101016jscitotenv201712296pmid 29289778
76 H A Wellard Kelly et al Macroinvertebrate diets reflecttributary inputs and turbidity-driven changes in food
availability in the Colorado River downstream of GlenCanyon Dam Freshw Sci 32 397ndash410 (2013)doi 10189912-0881
77 S J Imberger R M Thompson M R Grace Urbancatchment hydrology overwhelms reach scale effects ofriparian vegetation on organic matter dynamics FreshwaterBiol 56 1370ndash1389 (2011) doi 101111j1365-2427201102575x
78 R T Martins A S Melo J F Gonccedilalves Jr N HamadaLeaf-litter breakdown in urban streams of Central AmazoniaDirect and indirect effects of physical chemical andbiological factors Freshw Sci 34 716ndash726 (2015)doi 101086681086
79 C M Yule J Y Gan T Jinggut K V Lee Urbanizationaffects food webs and leaf-litter decomposition in a tropicalstream in Malaysia Freshw Sci 34 702ndash715 (2015)doi 101086681252
80 T A Newcomer et al Influence of natural and novel organiccarbon sources on denitrification in forest degraded urbanand restored streams Ecol Monogr 82 449ndash466 (2012)doi 10189012-04581
81 A Chaudhary I Kauser A Ray R Poretsky Taxon-drivenfunctional shifts associated with storm flow in an urbanstream microbial community MSphere 3 e00194-18 (2018)doi 101128mSphere00194-18 pmid 29976643
82 T A Kennedy et al Flow management for hydropowerextirpates aquatic insects undermining river food websBioscience 66 561ndash575 (2016) doi 101093bioscibiw059
83 A Glazaczow D Orwin M Bogdziewicz Increasedtemperature delays the late-season phenology of multivoltineinsect Sci Rep 6 38022 (2016) doi 101038srep38022pmid 27905493
84 S G Fisher L J Gray N B Grimm D E Busch Temporalsuccession in a desert stream ecosystem following flashflooding Ecol Monogr 52 93ndash110 (1982) doi 1023072937346
85 G J Scrimgeour M J Winterbourn Effects of floods onepilithon and benthic macroinvertebrate populations in anunstable New Zealand river Hydrobiologia 171 33ndash44 (1989)doi 101007BF00005722
86 E Bondar-Kunze S Maier D Schoumlnauer N Bahl T HeinAntagonistic and synergistic effects on a stream periphytoncommunity under the influence of pulsed flow velocityincrease and nutrient enrichment Sci Total Environ 573594ndash602 (2016) doi 101016jscitotenv201608158pmid 27585428
87 M J Cashman G L Harvey G Wharton M C Bruno Woodmitigates the effect of hydropeaking scour on periphytonbiomass and nutritional quality in semi-natural flumesimulations Aquat Sci 79 459ndash471 (2017) doi 101007s00027-016-0510-3
88 W F Cross et al Food-web dynamics in a large riverdiscontinuum Ecol Monogr 83 311ndash337 (2013)doi 10189012-17271
89 D M Warfe et al Productivity disturbance and ecosystemsize have no influence on food chain length in seasonallyconnected rivers PLOS ONE 8 e66240 (2013)doi 101371journalpone0066240
90 T D Jardine et al Consumer-resource coupling in wet-drytropical rivers J Anim Ecol 81 310ndash322 (2012)doi 101111j1365-2656201101925x pmid 22103689
91 K O Winemiller et al Pulsing hydrology determines top‐down control of basal resources in a tropical riverndashfloodplainecosystem Ecol Monogr 84 621ndash635 (2014)doi 10189013-18221
92 D M Post The long and short of food-chain length TrendsEcol Evol 17 269ndash277 (2002) doi 101016S0169-5347(02)02455-2
93 D M Post G Takimoto Proximate structural mechanismsfor variation in food‐chain length Oikos 116 775ndash782 (2007)doi 101111j0030-1299200715552x
94 P A McHugh A R McIntosh P G Jellyman Dual influencesof ecosystem size and disturbance on food chain length instreams Ecol Lett 13 881ndash890 (2010) doi 101111j1461-0248201001484x pmid 20482579
95 A Ruhiacute et al Flow regulation increases food-chain lengththrough omnivory mechanisms in a Mediterranean rivernetwork Freshwater Biol 61 1536ndash1549 (2016) doi 101111fwb12794
96 J L Sabo J C Finlay T Kennedy D M Post The role ofdischarge variation in scaling of drainage area and food chainlength in rivers Science 330 965ndash967 (2010) doi 101126science1196005 pmid 20947729
97 A R McIntosh et al Capacity to support predators scaleswith habitat size Sci Adv 4 eaap7523 (2018) doi 101126sciadvaap7523 pmid 29978038
98 J H Brown J F Gillooly A P Allen V M Savage G B WestToward a metabolic theory of ecology Ecology 851771ndash1789 (2004) doi 10189003-9000
99 K Zou E Theacutebault G Lacroix S Barot Interactions betweenthe green and brown food web determine ecosystemfunctioning Funct Ecol 30 1454ndash1465 (2016) doi 1011111365-243512626
100 M J Vanni P B McIntyre Predicting nutrient excretion ofaquatic animals with metabolic ecology and ecologicalstoichiometry A global synthesis Ecology 97 3460ndash3471(2016) doi 101002ecy1582 pmid 27912023
101 T B Parr K A Capps S P Inamdar K A Metcalf Animal-mediated organic matter transformation Aquatic insects as asource of microbially bioavailable organic nutrients andenergy Funct Ecol 33 524ndash535 (2019) doi 1011111365-243513242
102 C P Arango et al Urban infrastructure influences dissolvedorganic matter quality and bacterial metabolism in an urbanstream network Freshwater Biol 62 1917ndash1928 (2017)
103 L G Larsen J W Harvey Disrupted carbon cycling inrestored and unrestored urban streams Critical timescalesand controls Limnol Oceanogr 62 S160ndashS182 (2017)doi 101002lno10613
104 H M Halvorson G White J T Scott M A Evans-WhiteDietary and taxonomic controls on incorporation of microbialcarbon and phosphorus by detritivorous caddisfliesOecologia 180 567ndash579 (2016) doi 101007s00442-015-3464-6 pmid 26497125
105 J J Elser et al Biological stoichiometry from genes toecosystems Ecol Lett 3 540ndash550 (2000) doi 101046j1461-0248200000185x
106 E K Moody et al Consumption explains intraspecificvariation in nutrient recycling stoichiometry in a desert fishEcology 99 1552ndash1561 (2018) doi 101002ecy2372pmid 29882955
107 N Welti et al Bridging food webs ecosystem metabolismand biogeochemistry using ecological stoichiometry theoryFront Microbiol 8 1298 (2017) doi 103389fmicb201701298 pmid 28747904
108 M A Palmer S Filoso R M Fanelli From ecosystems toecosystem services Stream restoration as ecologicalengineering Ecol Eng 65 62ndash70 (2014) doi 101016jecoleng201307059
109 M J Burns T D Fletcher C J Walsh A R LadsonB E Hatt Hydrologic shortcomings of conventional urbanstormwater management and opportunities for reformLandsc Urban Plan 105 230ndash240 (2012) doi 101016jlandurbplan201112012
110 C R Liermann C Nilsson J Robertson R Y NgImplications of dam obstruction for global freshwater fishdiversity Bioscience 62 539ndash548 (2012) doi 101525bio20126265
111 J D Olden R J Naiman Incorporating thermal regimes intoenvironmental flows assessments Modifying dam operationsto restore freshwater ecosystem integrity Freshwater Biol55 86ndash107 (2010) doi 101111j1365-2427200902179x
112 D B Osmundson R J Ryel V L Lamarra J Pitlick Flowndashsedimentndashbiota relations Implications for river regulationeffects on native fish abundance Ecol Appl 12 1719ndash1739(2002)
113 D von Schiller et al Regulation causes nitrogen cyclingdiscontinuities in Mediterranean rivers Sci Total Environ540 168ndash177 (2016) doi 101016jscitotenv201507017pmid 26233271
114 J L Sabo et al Designing river flows to improve foodsecurity futures in the Lower Mekong Basin Science 358eaao1053 (2017) doi 101126scienceaao1053pmid 29217541
115 W Xu et al Improving the multi-objective performance ofrainwater harvesting systems using real-time controltechnology Water 10 147 (2018) doi 103390w10020147
116 M Acreman et al Environmental flows for naturalhybrid and novel riverine ecosystems in a changingworld Front Ecol Environ 12 466ndash473 (2014)doi 101890130134
117 N L Poff et al Sustainable water management under futureuncertainty with eco-engineering decision scaling Nat ClimChang 6 25ndash34 (2016) doi 101038nclimate2765
118 J D Kiernan P B Moyle P K Crain Restoring native fishassemblages to a regulated California stream using the
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 12 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
natural flow regime concept Ecol Appl 22 1472ndash1482(2012) doi 10189011-04801 pmid 22908707
119 D L Propst K B Gido Responses of native andnonnative fishes to natural flow regime mimicry in theSan Juan River Trans Am Fish Soc 133 922ndash931 (2004)doi 101577T03-0571
120 T K Pool J D Olden Assessing long‐term fish responses andshort‐term solutions to flow regulation in a dryland river basinEcol Freshwat Fish 24 56ndash66 (2015) doi 101111eff12125
121 W Chen J D Olden Designing flows to resolve human andenvironmental water needs in a dam-regulated riverNat Commun 8 2158 (2017) doi 101038s41467-017-02226-4 pmid 29255194
122 S M Nelson et al A history of the 2014 Minute 319environmental pulse flow as documented by fieldmeasurements and satellite imagery Ecol Eng 106733ndash748 (2017) doi 101016jecoleng201610040
123 P B Shafroth et al A large-scale environmental flowexperiment for riparian restoration in the Colorado RiverDelta Ecol Eng 106 645ndash660 (2017) doi 101016jecoleng201702016
124 E P Glenn P L Nagler P B Shafroth C J JarchowEffectiveness of environmental flows for riparian restorationin arid regions A tale of four rivers Ecol Eng 106 695ndash703(2017) doi 101016jecoleng201701009
125 S Hotaling D S Finn J Joseph Giersch D W WeisrockD Jacobsen Climate change and alpine stream biologyProgress challenges and opportunities for the futureBiol Rev Camb Philos Soc 92 2024ndash2045 (2017)doi 101111brv12319 pmid 28105701
126 C J Walsh T D Fletcher M J Burns Urban stormwaterrunoff A new class of environmental flow problem PLOS ONE7 e45814 (2012) doi 101371journalpone0045814pmid 23029257
127 A Selvakumar T P OrsquoConnor S D Struck Role of streamrestoration on improving benthic macroinvertebrates and in-stream water quality in an urban watershed Case studyJ Environ Eng 136 127ndash139 (2010) doi 101061(ASCE)EE1943-78700000116
128 R W Lammers B P Bledsoe What role does streamrestoration play in nutrient management Crit Rev EnvironSci Technol 47 335ndash371 (2017) doi 1010801064338920171318618
129 H E Golden N Hoghooghi Green infrastructure and itscatchment-scale effects An emerging science WIREs Water5 e1254 (2018) doi 101002wat21254 pmid 29682288
130 M R Williams G Bhatt S Filoso G Yactayo Streamrestoration performance and its contribution to theChesapeake Bay TMDL Challenges posed by climate changein urban areas Estuaries Coasts 40 1227ndash1246 (2017)doi 101007s12237-017-0226-1
131 R M Fanelli K Prestegaard M A Palmer Freshw Sci101086706072 (2019)
132 B W Abbott et al Unexpected spatial stability of waterchemistry in headwater stream networks Ecol Lett 21296ndash308 (2018) doi 101111ele12897 pmid 29282860
133 B L Brown C M Swan Dendritic network structureconstrains metacommunity properties in riverineecosystems J Anim Ecol 79 571ndash580 (2010) doi 101111j1365-2656201001668x pmid 20180874
134 C M Swan B L Brown Metacommunity theory meetsrestoration Isolation may mediate how ecologicalcommunities respond to stream restoration Ecol Appl 272209ndash2219 (2017) doi 101002eap1602 pmid 28718193
135 N Mouquet D Gravel F Massol V Calcagno Extending theconcept of keystone species to communities and ecosystemsEcol Lett 16 1ndash8 (2013) doi 101111ele12014 pmid 23062191
136 A Ruhiacute T Datry J L Sabo Interpreting beta-diversitycomponents over time to conserve metacommunities inhighly dynamic ecosystems Conserv Biol 31 1459ndash1468(2017) doi 101111cobi12906 pmid 28188969
137 L Yamane L W Botsford D P Kilduff Tracking restorationof population diversity via the portfolio effect J Appl Ecol55 472ndash481 (2018) doi 1011111365-266412978
138 D W Braidwood M A Taggart M Smith R AndersenTranslocations conservation and climate change Use ofrestoration sites as protorefuges and protorefugia RestorEcol 26 20ndash28 (2018) doi 101111rec12642
139 Y Wang D Wang Q W Lewis J Wu F Huang A frameworkto assess the cumulative impacts of dams on hydrologicalregime A case study of the Yangtze River Hydrol Processes31 3045ndash3055 (2017) doi 101002hyp11239
140 J L Sabo et al Pulsed flows tributary inputs and food-webstructure in a highly regulated river J Appl Ecol 551884ndash1895 (2018) doi 1011111365-266413109
141 C Magliozzi G Coro R C Grabowski A I PackmanS Krause A multiscale statistical method to identifypotential areas of hyporheic exchange for river restorationplanning Environ Model Softw 111 311ndash323 (2019)doi 101016jenvsoft201809006
142 S S Schwartz B Smith Restoring hydrologic function inurban landscapes with suburban subsoiling J Hydrol 543770ndash781 (2016) doi 101016jjhydrol201610051
143 C J Walsh T D Fletcher A R Ladson Stream restoration inurban catchments through redesigning stormwater systemsLooking to the catchment to save the stream J N AmBenthol Soc 24 690ndash705 (2005) doi 10189904-0201
144 N M Schmadel et al Thresholds of lake and reservoirconnectivity in river networks control nitrogen removalNat Commun 9 2779 (2018) doi 101038s41467-018-05156-x pmid 30018449
145 C D Sayer Conservation of aquatic landscapes Pondslakes and rivers as integrated systems WIREs Water 1573ndash585 (2014) doi 101002wat21045
146 R G Luthy D L Sedlak M H Plumlee D Austin V H ReshWastewater‐effluent‐dominated streams as ecosystem‐
management tools in a drier climate Front Ecol Environ 13477ndash485 (2015) doi 101890150038
147 D W Roscoe S G Hinch Effectiveness monitoring of fishpassage facilities Historical trends geographic patterns andfuture directions Fish Fish 11 12ndash33 (2010) doi 101111j1467-2979200900333x
148 C M Bunt T Castro-Santos A Haro Performanceof fish passage structures at upstream barriers tomigration River Res Appl 28 457ndash478 (2012)doi 101002rra1565
149 D Weigel et al Evaluation of a trap-and-transport programfor a threatened population of steelhead (Oncorhynchusmykiss) Conserv Genet 20 1195ndash1199 (2019) doi 101007s10592-019-01200-5
150 M H Bond et al Combined effects of barge transportationriver environment and rearing location on straying andmigration of adult Snake River fall-run Chinook SalmonTrans Am Fish Soc 146 60ndash73 (2017) doi 1010800002848720161235614
151 A C Ritchie et al Morphodynamic evolution followingsediment release from the worldrsquos largest dam removalSci Rep 8 13279 (2018) doi 101038s41598-018-30817-8pmid 30185796
152 T Sommer et al Californiarsquos Yolo Bypass Evidence thatflood control can be compatible with fisheries wetlandswildlife and agriculture Fisheries 26 6ndash16 (2001)doi 1015771548-8446(2001)026lt0006CYBgt20CO2
153 G Grill et al Mapping the worldrsquos free-flowing riversNature 569 215ndash221 (2019) doi 101038s41586-019-1111-9pmid 31068722
154 G Gassert M Luck M Landis P Reig T Shiao ldquoAqueductglobal maps 21 Constructing decision-relevant global waterrisk indicatorsrdquo Working paper World Resources Institute(2015) wwwwriorgpublicationaqueduct-global-maps-21-indicators
155 A Ruhiacute J D Olden J L Sabo Declining streamflow inducescollapse and replacement of native fishes in the AmericanSouthwest Front Ecol Environ 14 465ndash472 (2016)doi 101002fee1424
156 T C Gouhier A Grinsted V Simko R package Biwaveletconduct univariate and bivariate wavelet analyses (2018)
157 T Rodriacuteguez-Castillo E Esteacutevez A M Gonzaacutelez-FerrerasJ Barquiacuten Estimating ecosystem metabolism toentire river networks Ecosystems (2018) doi 101007s10021-018-0311-8
ACKNOWLEDGMENTS
We greatly appreciate those who read and provided importantinsights on drafts of this paper A Armstrong K HondulaJ Hosen K Leathers G de Mendoza C Maietta I MunozJ Tonkin and two anonymous reviewers We also thank T Tu forhelp with Fig 1 and S Carlson L Comte T Datry J HosenM Kondolf N Jones D Lytle E Moody J Olden M PowerJ Sabo and M Sanchez-Montoya for valuable discussionsCompeting interests The authors have no competing interestsFunding MAP was supported by funds from the NSF (1639145)and AR was supported by funds from the University of CaliforniaBerkeley and NSF (1802714)
101126scienceaaw2087
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 13 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
restorationLinkages between flow regime biota and ecosystem processes Implications for river
Margaret Palmer and Albert Ruhi
DOI 101126scienceaaw2087 (6459) eaaw2087365Science
this issue p eaaw2087Scienceflow variability and achieve hydrological connectivity between a river and its surroundingsand ecosystem processes To be successful such efforts must go beyond accounting for flood pulses to restore natural mimic ecologically important aspects of natural flow regimes guided by insights into how variations in flow affect biotawater quality and ecological processes In a Review Palmer and Ruhi explain how restoration designs now attempt to
Human activities have altered the flow regimes of many of Earths rivers with negative impacts on biodiversityRiver restoration guided by research
ARTICLE TOOLS httpsciencesciencemagorgcontent3656459eaaw2087
REFERENCES
httpsciencesciencemagorgcontent3656459eaaw2087BIBLThis article cites 147 articles 7 of which you can access for free
PERMISSIONS httpwwwsciencemagorghelpreprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAASScienceScience 1200 New York Avenue NW Washington DC 20005 The title (print ISSN 0036-8075 online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
Science No claim to original US Government WorksCopyright copy 2019 The Authors some rights reserved exclusive licensee American Association for the Advancement of
on May 27 2020
httpsciencesciencem
agorgD
ownloaded from

stressors (particularly in urban streams) and thechallenge of predicting outcomeswhen both thephysical and the biological environments arechangingAchieving a predictive understanding of hy-
droecological relationships is essential to design-ing successful self-sustaining restoration actionsin river ecosystems However translating basichydroecological insights into restoration practicerequires understanding the complexities of work-ing within a human-dominated water cycle Al-though it is rarely possible to restore all facetsof a natural flow regime ecologically informeddesigns may help mimic key aspects of the nat-ural flow regime (eg periodic pulse flows) orcreate regimes that are not natural but maximizeoutcomes given socioeconomic constraints InTable 1 we identify persisting and emergingchallenges in hydroecological research We paireach challenge with an opportunity (related toresearch design technology or implementation)We contend that these challenges should beprioritized to advance process-based river eco-system restoration
REFERENCES AND NOTES
1 N L Poff et al The natural flow regime Bioscience 47769ndash784 (1997) doi 1023071313099
2 D A Lytle N L Poff Adaptation to natural flow regimesTrends Ecol Evol 19 94ndash100 (2004) doi 101016jtree200310002 pmid 16701235
3 S Sabater A Elosegi R Ludwig in Multiple Stressors in RiverEcosystems (Elsevier 2019) pp 1ndash22
4 United Nations New UN Decade on Ecosystem Restorationoffers unparalleled opportunity for job creation food securityand addressing climate change (2019) httpswwwunenvironmentorgnews-and-storiespress-releasenew-un-decade-ecosystem-restoration-offers-unparalleled-opportunity
5 A W Lorenz P Haase K Januschke A SundermannD Hering Revisiting restored river reaches - Assessingchange of aquatic and riparian communities afterfive years Sci Total Environ 613-614 1185ndash1195(2018) doi 101016jscitotenv201709188pmid 28954379
6 M A Palmer K L Hondula B J Koch Ecological Restorationof Streams and Rivers Shifting Strategies and ShiftingGoals (2014) vol 45
7 T J Beechie et al Process-based principles for restoringriver ecosystems Bioscience 60 209ndash222 (2010)doi 101525bio20106037
8 S E Bunn A H Arthington Basic principles and ecologicalconsequences of altered flow regimes for aquaticbiodiversity Environ Manage 30 492ndash507 (2002)doi 101007s00267-002-2737-0 pmid 12481916
9 K D Fausch Y Taniguchi S Nakano G D GrossmanC R Townsend Flood disturbance regimes influence rainbowtrout invasion success among five holarctic regions EcolAppl 11 1438ndash1455 (2001) doi 1018901051-0761(2001)011[1438FDRIRT]20CO2
10 M E Power M S Parker W E Dietrich Seasonalreassembly of a river food web Floods droughts andimpacts of fish Ecol Monogr 78 263ndash282 (2008)doi 10189006-09021
11 V Acuntildea A Giorgi I Muntildeoz U R S Uehlinger S SabaterFlow extremes and benthic organic matter shape themetabolism of a headwater Mediterranean streamFreshwater Biol 49 960ndash971 (2004) doi 101111j1365-2427200401239x
12 K Tockner F Malard J V Ward An extension of the floodpulse concept Hydrol Processes 14 2861ndash2883 (2000)doi 1010021099-1085(20001112)141617lt2861AID-HYP124gt30CO2-F
13 J S Perkin et al Fragmentation and dewatering transformGreat Plains stream fish communities Ecol Monogr 8573ndash92 (2015) doi 10189014-01211
14 J L Sabo D M Post Quantifying periodic stochastic andcatastrophic environmental variation Ecol Monogr 7819ndash40 (2008) doi 10189006-13401
15 F Shuai et al Silver carp larva abundance in response toriver flow rate revealed by cross-wavelet modellingEcol Modell 383 98ndash105 (2018) doi 101016jecolmodel201805020
16 J D Tonkin M T Bogan N Bonada B Rios-ToumaD A Lytle Seasonality and predictability shape temporalspecies diversity Ecology 98 1201ndash1216 (2017)doi 101002ecy1761 pmid 28144975
17 N L Poff Beyond the natural flow regime Broadening thehydro‐ecological foundation to meet environmental flowschallenges in a non‐stationary world Freshwater Biol 631011ndash1021 (2018) doi 101111fwb13038
18 A Ruhi X Dong C H McDaniel D P Batzer J L SaboDetrimental effects of a novel flow regime on the functionaltrajectory of an aquatic invertebrate metacommunity GlobChange Biol 24 3749ndash3765 (2018) doi 101111gcb14133pmid 29665147
19 M T Bogan D A Lytle Severe drought drives novelcommunity trajectories in desert stream pools FreshwaterBiol 56 2070ndash2081 (2011) doi 101111j1365-2427201102638x
20 G Woodward et al The effects of climatic fluctuations andextreme events on running water ecosystems Philos TransR Soc London Ser B 371 20150274 (2016) pmid 27114576
21 M A Palmer C M Swan K Nelson P Silver R AlvestadStreambed landscapes Evidence that stream invertebratesrespond to the type and spatial arrangement of patchesLandsc Ecol 15 563ndash576 (2000) doi 101023A1008194130695
22 D Schmera et al Does isolation influence the relative role ofenvironmental and dispersal‐related processes in streamnetworks An empirical test of the network positionhypothesis using multiple taxa Freshwater Biol 63 74ndash85(2018) doi 101111fwb12973
23 R Henriques‐Silva et al A comprehensive examination of thenetwork position hypothesis across multiple rivermetacommunities Ecography 42 284ndash294 (2019)doi 101111ecog03908
24 K L Jaeger J D Olden N A Pelland Climate change poisedto threaten hydrologic connectivity and endemic fishes indryland streams Proc Natl Acad Sci USA 11113894ndash13899 (2014) doi 101073pnas1320890111pmid 25136090
25 S Garbin E A Celegon P Fanton G Botter Hydrologicalcontrols on river network connectivity R Soc Open Sci 6181428 (2019) doi 101098rsos181428 pmid 30891270
26 F Cattaneacuteo B Hugueny N Lamouroux Synchrony in browntrout Salmo trutta population dynamics A lsquoMoran effectrsquo onearly‐life stages Oikos 100 43ndash54 (2003) doi 101034j1600-0706200311912x
27 A Terui et al Metapopulation stability in branching rivernetworks Proc Natl Acad Sci USA 115 E5963ndashE5969(2018) doi 101073pnas1800060115 pmid 29895695
28 P A P Moran The statistical analysis of the Canadian Lynxcycle Aust J Zool 1 291ndash298 (1953) doi 101071ZO9530291
29 G D Grossman P B Moyle J O Whitaker Jr Stochasticityin structural and functional characteristics of an Indianastream fish assemblage A test of community theoryAm Nat 120 423ndash454 (1982) doi 101086284004
30 D A Lytle D M Merritt J D Tonkin J D OldenL V Reynolds Linking river flow regimes to riparian plantguilds A community-wide modeling approach Ecol Appl 271338ndash1350 (2017) doi 101002eap1528 pmid 28263426
31 A Ruhiacute E E Holmes J N Rinne J L Sabo Anomalousdroughts not invasion decrease persistence of native fishesin a desert river Glob Change Biol 21 1482ndash1496 (2015)doi 101111gcb12780 pmid 25430731
32 J D Tonkin D M Merritt J D Olden L V ReynoldsD A Lytle Flow regime alteration degrades ecologicalnetworks in riparian ecosystems Nat Ecol Evol 2 86ndash93(2018) doi 101038s41559-017-0379-0 pmid 29180707
33 M C Mims J D Olden Fish assemblages respond toaltered flow regimes via ecological filtering of lifehistory strategies Freshwater Biol 58 50ndash62 (2013)doi 101111fwb12037
34 P T J Johnson J D Olden M J Vander Zanden Daminvaders Impoundments facilitate biological invasions intofreshwaters Front Ecol Environ 6 357ndash363 (2008)doi 101890070156
35 N Bonada S Doledec B Statzner Taxonomic and biologicaltrait differences of stream macroinvertebrate communitiesbetween mediterranean and temperate regions Implicationsfor future climatic scenarios Glob Change Biol 131658ndash1671 (2007) doi 101111j1365-2486200701375x
36 M I Pyne N L Poff Vulnerability of stream communitycomposition and function to projected thermal warming andhydrologic change across ecoregions in the western UnitedStates Glob Change Biol 23 77ndash93 (2017) doi 101111gcb13437 pmid 27429092
37 D B Herbst S D Cooper R B Medhurst S W WisemanC T Hunsaker Drought ecohydrology alters the structureand function of benthic invertebrate communities inmountain streams Freshwater Biol 64 886ndash902 (2019)doi 101111fwb13270
38 M T Bogan E T Chester T Datry A L MurphyB J Robson A Ruhi R Stubbington J E Whitney inResistance Resilience and Community Recovery in IntermittentRivers and Ephemeral Streams T Datry N Bonada A BoultonEds (Academic Press 2017) pp 349ndash376
39 S Houmlckendorff et al Characterizing fish responses to a riverrestoration over 21 years based on speciesrsquo traits ConservBiol 31 1098ndash1108 (2017) doi 101111cobi12908pmid 28218807
40 D Bruno et al Structural and functional responses ofinvertebrate communities to climate change andflow regulation in alpine catchments Glob Change Biol25 1612ndash1628 (2019) doi 101111gcb14581pmid 30698905
41 T W H Aspin et al Extreme drought pushes streaminvertebrate communities over functional thresholds GlobChange Biol 25 230ndash244 (2019) doi 101111gcb14495pmid 30346098
42 S E Tank J B Fellman E Hood E S Kritzberg Beyondrespiration Controls on lateral carbon fluxes across theterrestrial-aquatic interface Limnol Oceanogr Lett 3 76ndash88(2018) doi 101002lol210065
43 A P Appling et al The metabolic regimes of 356 rivers inthe United States Sci Data 5 180292 (2018) doi 101038sdata2018292 pmid 30532078
44 P Savoy et al Metabolic rhythms in flowing watersAn approach for classifying river productivity regimesLimnol Oceanogr (2019) doi 101002lno11154
45 E S Bernhardt et al The metabolic regimes of flowingwaters Limnol Oceanogr 63 S99ndashS118 (2018)doi 101002lno10726
46 M Colls X Timoner S Sabater V Acuntildea Effects of durationand frequency of the non-flow period on stream metabolismEcosystems 101007s10021-019-00345-1 (2019)doi 101007s10021-019-00345-1
47 A J Ulseth E Bertuzzo G A Singer J Schelker T J BattinClimate-induced changes in spring snowmelt impactecosystem metabolism and carbon fluxes in an Alpine streamnetwork Ecosystems 21 373ndash390 (2018) doi 101007s10021-017-0155-7
48 B O L Demars Hydrological pulses and burning of dissolvedorganic carbon by stream respiration Limnol Oceanogr 64406ndash421 (2019) doi 101002lno11048
49 R T Hensley L Kirk M Spangler M N Gooseff M J CohenFlow extremes as spatiotemporal control points on riversolute fluxes and metabolism J Geophys Res Biogeosci124 537ndash555 (2019) doi 1010292018JG004738
50 J D Hosen et al Enhancement of primary production duringdrought in a temperate watershed is greater in larger riversthan headwater streams Limnol Oceanogr 64 1458ndash1472(2019) doi 101002lno11127
51 L Goacutemez-Gener et al When water vanishes Magnitude andregulation of carbon dioxide emissions from dry temporarystreams Ecosystems 19 710ndash723 (2016) doi 101007s10021-016-9963-4
52 E H Stanley et al The ecology of methane in streams andrivers Patterns controls and global significance EcolMonogr 86 146ndash171 (2016) doi 10189015-1027
53 R T Barnes D E Butman H F Wilson P A RaymondRiverine export of aged carbon driven by flow path depth andresidence time Environ Sci Technol 52 1028ndash1035 (2018)doi 101021acsest7b04717 pmid 29313674
54 G Pinay et al Riparian corridors A new conceptualframework for assessing nitrogen buffering across biomesFront Environ Sci 6 47 (2018) doi 103389fenvs201800047
55 W M Wollheim et al River network saturation conceptFactors influencing the balance of biogeochemical supply and
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 11 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
demand of river networks Biogeochemistry 141 503ndash521(2018) doi 101007s10533-018-0488-0
56 P A Raymond J E Saiers W V Sobczak Hydrological andbiogeochemical controls on watershed dissolved organicmatter transport Pulse-shunt concept Ecology 97 5ndash16(2016) doi 10189014-16841 pmid 27008769
57 C Tunaley D Tetzlaff J Lessels C Soulsby Linking high-frequency DOC dynamics to the age of connected watersources Water Resour Res 52 5232ndash5247 (2016)doi 1010022015WR018419
58 S Bernal et al Supply demand and in-stream retention ofdissolved organic carbon and nitrate during storms inMediterranean forested headwater streams Front EnvironSci 7 60 (2019) doi 103389fenvs201900060
59 J Harvey et al How hydrologic connectivity regulates waterquality in river corridors J Am Water Resour Assoc 55369ndash381 (2019) doi 1011111752-168812691
60 J C Moore et al Detritus trophic dynamics and biodiversityEcol Lett 7 584ndash600 (2004) doi 101111j1461-0248200400606x
61 A D Rosemond et al Freshwater ecology Experimentalnutrient additions accelerate terrestrial carbon loss fromstream ecosystems Science 347 1142ndash1145 (2015)doi 101126scienceaaa1958 pmid 25745171
62 A C Y Yeung J L Musetta-Lambert D P KreutzweiserP K Sibley J S Richardson Relations of interannualdifferences in stream litter breakdown with dischargeBioassessment implications Ecosphere 9 e02423 (2018)doi 101002ecs22423
63 R M Northington J R Webster Experimental reductions instream flow alter litter processing and consumer subsidies inheadwater streams Freshwater Biol 62 737ndash750 (2017)doi 101111fwb12898
64 E Mas-Martiacute I Sanpera-Calbet I Muntildeoz Bottom-up effectsof streambed drying on consumer performance throughchanges in resource quality Aquat Sci 79 719ndash731 (2017)doi 101007s00027-017-0531-6
65 B D Richter G A Thomas Restoring environmental flows bymodifying dam operations Ecol Soc 12 12 (2007)doi 105751ES-02014-120112
66 G Zolezzi A Siviglia M Toffolon B MaioliniThermopeaking in Alpine streams Event characterizationand time scales Ecohydrology 4 564ndash576 (2011)doi 101002eco132
67 T Meiszligner B Sures C K Feld Multiple stressors and therole of hydrology on benthic invertebrates in mountainousstreams Sci Total Environ 663 841ndash851 (2019)doi 101016jscitotenv201901288 pmid 30738264
68 I R Waite et al Effects of urban multi-stressors on threestream biotic assemblages Sci Total Environ 6601472ndash1485 (2019) doi 101016jscitotenv201812240pmid 30743940
69 K C Nelson M A Palmer Stream temperature surges underurbanization and climate change Data models andresponses J Am Water Resour Assoc 43 440ndash452 (2007)doi 101111j1752-1688200700034x
70 J R Blaszczak J M Delesantro Y Zhong D L UrbanE S Bernhardt Watershed urban development controls onurban streamwater chemistry variability Biogeochemistry144 61ndash84 (2019) doi 101007s10533-019-00572-7
71 L Ponsatiacute et al Biofilm responses to flow regulation by damsin Mediterranean rivers River Res Appl 31 1003ndash1016(2015) doi 101002rra2807
72 J M Alberts J J Beaulieu I Buffam Watershed land useand seasonal variation constrain the influence of ripariancanopy cover on stream ecosystem metabolism Ecosystems20 553ndash567 (2017) doi 101007s10021-016-0040-9pmid 30416371
73 R O Hall Jr et al Turbidity light temperature andhydropeaking control primary productivity in the ColoradoRiver Grand Canyon Limnol Oceanogr 60 512ndash526 (2015)doi 101002lno10031
74 J R Blaszczak J M Delesantro D L Urban M W DoyleE S Bernhardt Scoured or suffocated Urban streamecosystems oscillate between hydrologic and dissolvedoxygen extremes Limnol Oceanogr 64 877ndash894 (2019)doi 101002lno11081
75 J-R Mor et al Dam regulation and riverine food-webstructure in a Mediterranean river Sci Total Environ 625301ndash310 (2018) doi 101016jscitotenv201712296pmid 29289778
76 H A Wellard Kelly et al Macroinvertebrate diets reflecttributary inputs and turbidity-driven changes in food
availability in the Colorado River downstream of GlenCanyon Dam Freshw Sci 32 397ndash410 (2013)doi 10189912-0881
77 S J Imberger R M Thompson M R Grace Urbancatchment hydrology overwhelms reach scale effects ofriparian vegetation on organic matter dynamics FreshwaterBiol 56 1370ndash1389 (2011) doi 101111j1365-2427201102575x
78 R T Martins A S Melo J F Gonccedilalves Jr N HamadaLeaf-litter breakdown in urban streams of Central AmazoniaDirect and indirect effects of physical chemical andbiological factors Freshw Sci 34 716ndash726 (2015)doi 101086681086
79 C M Yule J Y Gan T Jinggut K V Lee Urbanizationaffects food webs and leaf-litter decomposition in a tropicalstream in Malaysia Freshw Sci 34 702ndash715 (2015)doi 101086681252
80 T A Newcomer et al Influence of natural and novel organiccarbon sources on denitrification in forest degraded urbanand restored streams Ecol Monogr 82 449ndash466 (2012)doi 10189012-04581
81 A Chaudhary I Kauser A Ray R Poretsky Taxon-drivenfunctional shifts associated with storm flow in an urbanstream microbial community MSphere 3 e00194-18 (2018)doi 101128mSphere00194-18 pmid 29976643
82 T A Kennedy et al Flow management for hydropowerextirpates aquatic insects undermining river food websBioscience 66 561ndash575 (2016) doi 101093bioscibiw059
83 A Glazaczow D Orwin M Bogdziewicz Increasedtemperature delays the late-season phenology of multivoltineinsect Sci Rep 6 38022 (2016) doi 101038srep38022pmid 27905493
84 S G Fisher L J Gray N B Grimm D E Busch Temporalsuccession in a desert stream ecosystem following flashflooding Ecol Monogr 52 93ndash110 (1982) doi 1023072937346
85 G J Scrimgeour M J Winterbourn Effects of floods onepilithon and benthic macroinvertebrate populations in anunstable New Zealand river Hydrobiologia 171 33ndash44 (1989)doi 101007BF00005722
86 E Bondar-Kunze S Maier D Schoumlnauer N Bahl T HeinAntagonistic and synergistic effects on a stream periphytoncommunity under the influence of pulsed flow velocityincrease and nutrient enrichment Sci Total Environ 573594ndash602 (2016) doi 101016jscitotenv201608158pmid 27585428
87 M J Cashman G L Harvey G Wharton M C Bruno Woodmitigates the effect of hydropeaking scour on periphytonbiomass and nutritional quality in semi-natural flumesimulations Aquat Sci 79 459ndash471 (2017) doi 101007s00027-016-0510-3
88 W F Cross et al Food-web dynamics in a large riverdiscontinuum Ecol Monogr 83 311ndash337 (2013)doi 10189012-17271
89 D M Warfe et al Productivity disturbance and ecosystemsize have no influence on food chain length in seasonallyconnected rivers PLOS ONE 8 e66240 (2013)doi 101371journalpone0066240
90 T D Jardine et al Consumer-resource coupling in wet-drytropical rivers J Anim Ecol 81 310ndash322 (2012)doi 101111j1365-2656201101925x pmid 22103689
91 K O Winemiller et al Pulsing hydrology determines top‐down control of basal resources in a tropical riverndashfloodplainecosystem Ecol Monogr 84 621ndash635 (2014)doi 10189013-18221
92 D M Post The long and short of food-chain length TrendsEcol Evol 17 269ndash277 (2002) doi 101016S0169-5347(02)02455-2
93 D M Post G Takimoto Proximate structural mechanismsfor variation in food‐chain length Oikos 116 775ndash782 (2007)doi 101111j0030-1299200715552x
94 P A McHugh A R McIntosh P G Jellyman Dual influencesof ecosystem size and disturbance on food chain length instreams Ecol Lett 13 881ndash890 (2010) doi 101111j1461-0248201001484x pmid 20482579
95 A Ruhiacute et al Flow regulation increases food-chain lengththrough omnivory mechanisms in a Mediterranean rivernetwork Freshwater Biol 61 1536ndash1549 (2016) doi 101111fwb12794
96 J L Sabo J C Finlay T Kennedy D M Post The role ofdischarge variation in scaling of drainage area and food chainlength in rivers Science 330 965ndash967 (2010) doi 101126science1196005 pmid 20947729
97 A R McIntosh et al Capacity to support predators scaleswith habitat size Sci Adv 4 eaap7523 (2018) doi 101126sciadvaap7523 pmid 29978038
98 J H Brown J F Gillooly A P Allen V M Savage G B WestToward a metabolic theory of ecology Ecology 851771ndash1789 (2004) doi 10189003-9000
99 K Zou E Theacutebault G Lacroix S Barot Interactions betweenthe green and brown food web determine ecosystemfunctioning Funct Ecol 30 1454ndash1465 (2016) doi 1011111365-243512626
100 M J Vanni P B McIntyre Predicting nutrient excretion ofaquatic animals with metabolic ecology and ecologicalstoichiometry A global synthesis Ecology 97 3460ndash3471(2016) doi 101002ecy1582 pmid 27912023
101 T B Parr K A Capps S P Inamdar K A Metcalf Animal-mediated organic matter transformation Aquatic insects as asource of microbially bioavailable organic nutrients andenergy Funct Ecol 33 524ndash535 (2019) doi 1011111365-243513242
102 C P Arango et al Urban infrastructure influences dissolvedorganic matter quality and bacterial metabolism in an urbanstream network Freshwater Biol 62 1917ndash1928 (2017)
103 L G Larsen J W Harvey Disrupted carbon cycling inrestored and unrestored urban streams Critical timescalesand controls Limnol Oceanogr 62 S160ndashS182 (2017)doi 101002lno10613
104 H M Halvorson G White J T Scott M A Evans-WhiteDietary and taxonomic controls on incorporation of microbialcarbon and phosphorus by detritivorous caddisfliesOecologia 180 567ndash579 (2016) doi 101007s00442-015-3464-6 pmid 26497125
105 J J Elser et al Biological stoichiometry from genes toecosystems Ecol Lett 3 540ndash550 (2000) doi 101046j1461-0248200000185x
106 E K Moody et al Consumption explains intraspecificvariation in nutrient recycling stoichiometry in a desert fishEcology 99 1552ndash1561 (2018) doi 101002ecy2372pmid 29882955
107 N Welti et al Bridging food webs ecosystem metabolismand biogeochemistry using ecological stoichiometry theoryFront Microbiol 8 1298 (2017) doi 103389fmicb201701298 pmid 28747904
108 M A Palmer S Filoso R M Fanelli From ecosystems toecosystem services Stream restoration as ecologicalengineering Ecol Eng 65 62ndash70 (2014) doi 101016jecoleng201307059
109 M J Burns T D Fletcher C J Walsh A R LadsonB E Hatt Hydrologic shortcomings of conventional urbanstormwater management and opportunities for reformLandsc Urban Plan 105 230ndash240 (2012) doi 101016jlandurbplan201112012
110 C R Liermann C Nilsson J Robertson R Y NgImplications of dam obstruction for global freshwater fishdiversity Bioscience 62 539ndash548 (2012) doi 101525bio20126265
111 J D Olden R J Naiman Incorporating thermal regimes intoenvironmental flows assessments Modifying dam operationsto restore freshwater ecosystem integrity Freshwater Biol55 86ndash107 (2010) doi 101111j1365-2427200902179x
112 D B Osmundson R J Ryel V L Lamarra J Pitlick Flowndashsedimentndashbiota relations Implications for river regulationeffects on native fish abundance Ecol Appl 12 1719ndash1739(2002)
113 D von Schiller et al Regulation causes nitrogen cyclingdiscontinuities in Mediterranean rivers Sci Total Environ540 168ndash177 (2016) doi 101016jscitotenv201507017pmid 26233271
114 J L Sabo et al Designing river flows to improve foodsecurity futures in the Lower Mekong Basin Science 358eaao1053 (2017) doi 101126scienceaao1053pmid 29217541
115 W Xu et al Improving the multi-objective performance ofrainwater harvesting systems using real-time controltechnology Water 10 147 (2018) doi 103390w10020147
116 M Acreman et al Environmental flows for naturalhybrid and novel riverine ecosystems in a changingworld Front Ecol Environ 12 466ndash473 (2014)doi 101890130134
117 N L Poff et al Sustainable water management under futureuncertainty with eco-engineering decision scaling Nat ClimChang 6 25ndash34 (2016) doi 101038nclimate2765
118 J D Kiernan P B Moyle P K Crain Restoring native fishassemblages to a regulated California stream using the
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 12 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
natural flow regime concept Ecol Appl 22 1472ndash1482(2012) doi 10189011-04801 pmid 22908707
119 D L Propst K B Gido Responses of native andnonnative fishes to natural flow regime mimicry in theSan Juan River Trans Am Fish Soc 133 922ndash931 (2004)doi 101577T03-0571
120 T K Pool J D Olden Assessing long‐term fish responses andshort‐term solutions to flow regulation in a dryland river basinEcol Freshwat Fish 24 56ndash66 (2015) doi 101111eff12125
121 W Chen J D Olden Designing flows to resolve human andenvironmental water needs in a dam-regulated riverNat Commun 8 2158 (2017) doi 101038s41467-017-02226-4 pmid 29255194
122 S M Nelson et al A history of the 2014 Minute 319environmental pulse flow as documented by fieldmeasurements and satellite imagery Ecol Eng 106733ndash748 (2017) doi 101016jecoleng201610040
123 P B Shafroth et al A large-scale environmental flowexperiment for riparian restoration in the Colorado RiverDelta Ecol Eng 106 645ndash660 (2017) doi 101016jecoleng201702016
124 E P Glenn P L Nagler P B Shafroth C J JarchowEffectiveness of environmental flows for riparian restorationin arid regions A tale of four rivers Ecol Eng 106 695ndash703(2017) doi 101016jecoleng201701009
125 S Hotaling D S Finn J Joseph Giersch D W WeisrockD Jacobsen Climate change and alpine stream biologyProgress challenges and opportunities for the futureBiol Rev Camb Philos Soc 92 2024ndash2045 (2017)doi 101111brv12319 pmid 28105701
126 C J Walsh T D Fletcher M J Burns Urban stormwaterrunoff A new class of environmental flow problem PLOS ONE7 e45814 (2012) doi 101371journalpone0045814pmid 23029257
127 A Selvakumar T P OrsquoConnor S D Struck Role of streamrestoration on improving benthic macroinvertebrates and in-stream water quality in an urban watershed Case studyJ Environ Eng 136 127ndash139 (2010) doi 101061(ASCE)EE1943-78700000116
128 R W Lammers B P Bledsoe What role does streamrestoration play in nutrient management Crit Rev EnvironSci Technol 47 335ndash371 (2017) doi 1010801064338920171318618
129 H E Golden N Hoghooghi Green infrastructure and itscatchment-scale effects An emerging science WIREs Water5 e1254 (2018) doi 101002wat21254 pmid 29682288
130 M R Williams G Bhatt S Filoso G Yactayo Streamrestoration performance and its contribution to theChesapeake Bay TMDL Challenges posed by climate changein urban areas Estuaries Coasts 40 1227ndash1246 (2017)doi 101007s12237-017-0226-1
131 R M Fanelli K Prestegaard M A Palmer Freshw Sci101086706072 (2019)
132 B W Abbott et al Unexpected spatial stability of waterchemistry in headwater stream networks Ecol Lett 21296ndash308 (2018) doi 101111ele12897 pmid 29282860
133 B L Brown C M Swan Dendritic network structureconstrains metacommunity properties in riverineecosystems J Anim Ecol 79 571ndash580 (2010) doi 101111j1365-2656201001668x pmid 20180874
134 C M Swan B L Brown Metacommunity theory meetsrestoration Isolation may mediate how ecologicalcommunities respond to stream restoration Ecol Appl 272209ndash2219 (2017) doi 101002eap1602 pmid 28718193
135 N Mouquet D Gravel F Massol V Calcagno Extending theconcept of keystone species to communities and ecosystemsEcol Lett 16 1ndash8 (2013) doi 101111ele12014 pmid 23062191
136 A Ruhiacute T Datry J L Sabo Interpreting beta-diversitycomponents over time to conserve metacommunities inhighly dynamic ecosystems Conserv Biol 31 1459ndash1468(2017) doi 101111cobi12906 pmid 28188969
137 L Yamane L W Botsford D P Kilduff Tracking restorationof population diversity via the portfolio effect J Appl Ecol55 472ndash481 (2018) doi 1011111365-266412978
138 D W Braidwood M A Taggart M Smith R AndersenTranslocations conservation and climate change Use ofrestoration sites as protorefuges and protorefugia RestorEcol 26 20ndash28 (2018) doi 101111rec12642
139 Y Wang D Wang Q W Lewis J Wu F Huang A frameworkto assess the cumulative impacts of dams on hydrologicalregime A case study of the Yangtze River Hydrol Processes31 3045ndash3055 (2017) doi 101002hyp11239
140 J L Sabo et al Pulsed flows tributary inputs and food-webstructure in a highly regulated river J Appl Ecol 551884ndash1895 (2018) doi 1011111365-266413109
141 C Magliozzi G Coro R C Grabowski A I PackmanS Krause A multiscale statistical method to identifypotential areas of hyporheic exchange for river restorationplanning Environ Model Softw 111 311ndash323 (2019)doi 101016jenvsoft201809006
142 S S Schwartz B Smith Restoring hydrologic function inurban landscapes with suburban subsoiling J Hydrol 543770ndash781 (2016) doi 101016jjhydrol201610051
143 C J Walsh T D Fletcher A R Ladson Stream restoration inurban catchments through redesigning stormwater systemsLooking to the catchment to save the stream J N AmBenthol Soc 24 690ndash705 (2005) doi 10189904-0201
144 N M Schmadel et al Thresholds of lake and reservoirconnectivity in river networks control nitrogen removalNat Commun 9 2779 (2018) doi 101038s41467-018-05156-x pmid 30018449
145 C D Sayer Conservation of aquatic landscapes Pondslakes and rivers as integrated systems WIREs Water 1573ndash585 (2014) doi 101002wat21045
146 R G Luthy D L Sedlak M H Plumlee D Austin V H ReshWastewater‐effluent‐dominated streams as ecosystem‐
management tools in a drier climate Front Ecol Environ 13477ndash485 (2015) doi 101890150038
147 D W Roscoe S G Hinch Effectiveness monitoring of fishpassage facilities Historical trends geographic patterns andfuture directions Fish Fish 11 12ndash33 (2010) doi 101111j1467-2979200900333x
148 C M Bunt T Castro-Santos A Haro Performanceof fish passage structures at upstream barriers tomigration River Res Appl 28 457ndash478 (2012)doi 101002rra1565
149 D Weigel et al Evaluation of a trap-and-transport programfor a threatened population of steelhead (Oncorhynchusmykiss) Conserv Genet 20 1195ndash1199 (2019) doi 101007s10592-019-01200-5
150 M H Bond et al Combined effects of barge transportationriver environment and rearing location on straying andmigration of adult Snake River fall-run Chinook SalmonTrans Am Fish Soc 146 60ndash73 (2017) doi 1010800002848720161235614
151 A C Ritchie et al Morphodynamic evolution followingsediment release from the worldrsquos largest dam removalSci Rep 8 13279 (2018) doi 101038s41598-018-30817-8pmid 30185796
152 T Sommer et al Californiarsquos Yolo Bypass Evidence thatflood control can be compatible with fisheries wetlandswildlife and agriculture Fisheries 26 6ndash16 (2001)doi 1015771548-8446(2001)026lt0006CYBgt20CO2
153 G Grill et al Mapping the worldrsquos free-flowing riversNature 569 215ndash221 (2019) doi 101038s41586-019-1111-9pmid 31068722
154 G Gassert M Luck M Landis P Reig T Shiao ldquoAqueductglobal maps 21 Constructing decision-relevant global waterrisk indicatorsrdquo Working paper World Resources Institute(2015) wwwwriorgpublicationaqueduct-global-maps-21-indicators
155 A Ruhiacute J D Olden J L Sabo Declining streamflow inducescollapse and replacement of native fishes in the AmericanSouthwest Front Ecol Environ 14 465ndash472 (2016)doi 101002fee1424
156 T C Gouhier A Grinsted V Simko R package Biwaveletconduct univariate and bivariate wavelet analyses (2018)
157 T Rodriacuteguez-Castillo E Esteacutevez A M Gonzaacutelez-FerrerasJ Barquiacuten Estimating ecosystem metabolism toentire river networks Ecosystems (2018) doi 101007s10021-018-0311-8
ACKNOWLEDGMENTS
We greatly appreciate those who read and provided importantinsights on drafts of this paper A Armstrong K HondulaJ Hosen K Leathers G de Mendoza C Maietta I MunozJ Tonkin and two anonymous reviewers We also thank T Tu forhelp with Fig 1 and S Carlson L Comte T Datry J HosenM Kondolf N Jones D Lytle E Moody J Olden M PowerJ Sabo and M Sanchez-Montoya for valuable discussionsCompeting interests The authors have no competing interestsFunding MAP was supported by funds from the NSF (1639145)and AR was supported by funds from the University of CaliforniaBerkeley and NSF (1802714)
101126scienceaaw2087
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 13 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
restorationLinkages between flow regime biota and ecosystem processes Implications for river
Margaret Palmer and Albert Ruhi
DOI 101126scienceaaw2087 (6459) eaaw2087365Science
this issue p eaaw2087Scienceflow variability and achieve hydrological connectivity between a river and its surroundingsand ecosystem processes To be successful such efforts must go beyond accounting for flood pulses to restore natural mimic ecologically important aspects of natural flow regimes guided by insights into how variations in flow affect biotawater quality and ecological processes In a Review Palmer and Ruhi explain how restoration designs now attempt to
Human activities have altered the flow regimes of many of Earths rivers with negative impacts on biodiversityRiver restoration guided by research
ARTICLE TOOLS httpsciencesciencemagorgcontent3656459eaaw2087
REFERENCES
httpsciencesciencemagorgcontent3656459eaaw2087BIBLThis article cites 147 articles 7 of which you can access for free
PERMISSIONS httpwwwsciencemagorghelpreprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAASScienceScience 1200 New York Avenue NW Washington DC 20005 The title (print ISSN 0036-8075 online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
Science No claim to original US Government WorksCopyright copy 2019 The Authors some rights reserved exclusive licensee American Association for the Advancement of
on May 27 2020
httpsciencesciencem
agorgD
ownloaded from

demand of river networks Biogeochemistry 141 503ndash521(2018) doi 101007s10533-018-0488-0
56 P A Raymond J E Saiers W V Sobczak Hydrological andbiogeochemical controls on watershed dissolved organicmatter transport Pulse-shunt concept Ecology 97 5ndash16(2016) doi 10189014-16841 pmid 27008769
57 C Tunaley D Tetzlaff J Lessels C Soulsby Linking high-frequency DOC dynamics to the age of connected watersources Water Resour Res 52 5232ndash5247 (2016)doi 1010022015WR018419
58 S Bernal et al Supply demand and in-stream retention ofdissolved organic carbon and nitrate during storms inMediterranean forested headwater streams Front EnvironSci 7 60 (2019) doi 103389fenvs201900060
59 J Harvey et al How hydrologic connectivity regulates waterquality in river corridors J Am Water Resour Assoc 55369ndash381 (2019) doi 1011111752-168812691
60 J C Moore et al Detritus trophic dynamics and biodiversityEcol Lett 7 584ndash600 (2004) doi 101111j1461-0248200400606x
61 A D Rosemond et al Freshwater ecology Experimentalnutrient additions accelerate terrestrial carbon loss fromstream ecosystems Science 347 1142ndash1145 (2015)doi 101126scienceaaa1958 pmid 25745171
62 A C Y Yeung J L Musetta-Lambert D P KreutzweiserP K Sibley J S Richardson Relations of interannualdifferences in stream litter breakdown with dischargeBioassessment implications Ecosphere 9 e02423 (2018)doi 101002ecs22423
63 R M Northington J R Webster Experimental reductions instream flow alter litter processing and consumer subsidies inheadwater streams Freshwater Biol 62 737ndash750 (2017)doi 101111fwb12898
64 E Mas-Martiacute I Sanpera-Calbet I Muntildeoz Bottom-up effectsof streambed drying on consumer performance throughchanges in resource quality Aquat Sci 79 719ndash731 (2017)doi 101007s00027-017-0531-6
65 B D Richter G A Thomas Restoring environmental flows bymodifying dam operations Ecol Soc 12 12 (2007)doi 105751ES-02014-120112
66 G Zolezzi A Siviglia M Toffolon B MaioliniThermopeaking in Alpine streams Event characterizationand time scales Ecohydrology 4 564ndash576 (2011)doi 101002eco132
67 T Meiszligner B Sures C K Feld Multiple stressors and therole of hydrology on benthic invertebrates in mountainousstreams Sci Total Environ 663 841ndash851 (2019)doi 101016jscitotenv201901288 pmid 30738264
68 I R Waite et al Effects of urban multi-stressors on threestream biotic assemblages Sci Total Environ 6601472ndash1485 (2019) doi 101016jscitotenv201812240pmid 30743940
69 K C Nelson M A Palmer Stream temperature surges underurbanization and climate change Data models andresponses J Am Water Resour Assoc 43 440ndash452 (2007)doi 101111j1752-1688200700034x
70 J R Blaszczak J M Delesantro Y Zhong D L UrbanE S Bernhardt Watershed urban development controls onurban streamwater chemistry variability Biogeochemistry144 61ndash84 (2019) doi 101007s10533-019-00572-7
71 L Ponsatiacute et al Biofilm responses to flow regulation by damsin Mediterranean rivers River Res Appl 31 1003ndash1016(2015) doi 101002rra2807
72 J M Alberts J J Beaulieu I Buffam Watershed land useand seasonal variation constrain the influence of ripariancanopy cover on stream ecosystem metabolism Ecosystems20 553ndash567 (2017) doi 101007s10021-016-0040-9pmid 30416371
73 R O Hall Jr et al Turbidity light temperature andhydropeaking control primary productivity in the ColoradoRiver Grand Canyon Limnol Oceanogr 60 512ndash526 (2015)doi 101002lno10031
74 J R Blaszczak J M Delesantro D L Urban M W DoyleE S Bernhardt Scoured or suffocated Urban streamecosystems oscillate between hydrologic and dissolvedoxygen extremes Limnol Oceanogr 64 877ndash894 (2019)doi 101002lno11081
75 J-R Mor et al Dam regulation and riverine food-webstructure in a Mediterranean river Sci Total Environ 625301ndash310 (2018) doi 101016jscitotenv201712296pmid 29289778
76 H A Wellard Kelly et al Macroinvertebrate diets reflecttributary inputs and turbidity-driven changes in food
availability in the Colorado River downstream of GlenCanyon Dam Freshw Sci 32 397ndash410 (2013)doi 10189912-0881
77 S J Imberger R M Thompson M R Grace Urbancatchment hydrology overwhelms reach scale effects ofriparian vegetation on organic matter dynamics FreshwaterBiol 56 1370ndash1389 (2011) doi 101111j1365-2427201102575x
78 R T Martins A S Melo J F Gonccedilalves Jr N HamadaLeaf-litter breakdown in urban streams of Central AmazoniaDirect and indirect effects of physical chemical andbiological factors Freshw Sci 34 716ndash726 (2015)doi 101086681086
79 C M Yule J Y Gan T Jinggut K V Lee Urbanizationaffects food webs and leaf-litter decomposition in a tropicalstream in Malaysia Freshw Sci 34 702ndash715 (2015)doi 101086681252
80 T A Newcomer et al Influence of natural and novel organiccarbon sources on denitrification in forest degraded urbanand restored streams Ecol Monogr 82 449ndash466 (2012)doi 10189012-04581
81 A Chaudhary I Kauser A Ray R Poretsky Taxon-drivenfunctional shifts associated with storm flow in an urbanstream microbial community MSphere 3 e00194-18 (2018)doi 101128mSphere00194-18 pmid 29976643
82 T A Kennedy et al Flow management for hydropowerextirpates aquatic insects undermining river food websBioscience 66 561ndash575 (2016) doi 101093bioscibiw059
83 A Glazaczow D Orwin M Bogdziewicz Increasedtemperature delays the late-season phenology of multivoltineinsect Sci Rep 6 38022 (2016) doi 101038srep38022pmid 27905493
84 S G Fisher L J Gray N B Grimm D E Busch Temporalsuccession in a desert stream ecosystem following flashflooding Ecol Monogr 52 93ndash110 (1982) doi 1023072937346
85 G J Scrimgeour M J Winterbourn Effects of floods onepilithon and benthic macroinvertebrate populations in anunstable New Zealand river Hydrobiologia 171 33ndash44 (1989)doi 101007BF00005722
86 E Bondar-Kunze S Maier D Schoumlnauer N Bahl T HeinAntagonistic and synergistic effects on a stream periphytoncommunity under the influence of pulsed flow velocityincrease and nutrient enrichment Sci Total Environ 573594ndash602 (2016) doi 101016jscitotenv201608158pmid 27585428
87 M J Cashman G L Harvey G Wharton M C Bruno Woodmitigates the effect of hydropeaking scour on periphytonbiomass and nutritional quality in semi-natural flumesimulations Aquat Sci 79 459ndash471 (2017) doi 101007s00027-016-0510-3
88 W F Cross et al Food-web dynamics in a large riverdiscontinuum Ecol Monogr 83 311ndash337 (2013)doi 10189012-17271
89 D M Warfe et al Productivity disturbance and ecosystemsize have no influence on food chain length in seasonallyconnected rivers PLOS ONE 8 e66240 (2013)doi 101371journalpone0066240
90 T D Jardine et al Consumer-resource coupling in wet-drytropical rivers J Anim Ecol 81 310ndash322 (2012)doi 101111j1365-2656201101925x pmid 22103689
91 K O Winemiller et al Pulsing hydrology determines top‐down control of basal resources in a tropical riverndashfloodplainecosystem Ecol Monogr 84 621ndash635 (2014)doi 10189013-18221
92 D M Post The long and short of food-chain length TrendsEcol Evol 17 269ndash277 (2002) doi 101016S0169-5347(02)02455-2
93 D M Post G Takimoto Proximate structural mechanismsfor variation in food‐chain length Oikos 116 775ndash782 (2007)doi 101111j0030-1299200715552x
94 P A McHugh A R McIntosh P G Jellyman Dual influencesof ecosystem size and disturbance on food chain length instreams Ecol Lett 13 881ndash890 (2010) doi 101111j1461-0248201001484x pmid 20482579
95 A Ruhiacute et al Flow regulation increases food-chain lengththrough omnivory mechanisms in a Mediterranean rivernetwork Freshwater Biol 61 1536ndash1549 (2016) doi 101111fwb12794
96 J L Sabo J C Finlay T Kennedy D M Post The role ofdischarge variation in scaling of drainage area and food chainlength in rivers Science 330 965ndash967 (2010) doi 101126science1196005 pmid 20947729
97 A R McIntosh et al Capacity to support predators scaleswith habitat size Sci Adv 4 eaap7523 (2018) doi 101126sciadvaap7523 pmid 29978038
98 J H Brown J F Gillooly A P Allen V M Savage G B WestToward a metabolic theory of ecology Ecology 851771ndash1789 (2004) doi 10189003-9000
99 K Zou E Theacutebault G Lacroix S Barot Interactions betweenthe green and brown food web determine ecosystemfunctioning Funct Ecol 30 1454ndash1465 (2016) doi 1011111365-243512626
100 M J Vanni P B McIntyre Predicting nutrient excretion ofaquatic animals with metabolic ecology and ecologicalstoichiometry A global synthesis Ecology 97 3460ndash3471(2016) doi 101002ecy1582 pmid 27912023
101 T B Parr K A Capps S P Inamdar K A Metcalf Animal-mediated organic matter transformation Aquatic insects as asource of microbially bioavailable organic nutrients andenergy Funct Ecol 33 524ndash535 (2019) doi 1011111365-243513242
102 C P Arango et al Urban infrastructure influences dissolvedorganic matter quality and bacterial metabolism in an urbanstream network Freshwater Biol 62 1917ndash1928 (2017)
103 L G Larsen J W Harvey Disrupted carbon cycling inrestored and unrestored urban streams Critical timescalesand controls Limnol Oceanogr 62 S160ndashS182 (2017)doi 101002lno10613
104 H M Halvorson G White J T Scott M A Evans-WhiteDietary and taxonomic controls on incorporation of microbialcarbon and phosphorus by detritivorous caddisfliesOecologia 180 567ndash579 (2016) doi 101007s00442-015-3464-6 pmid 26497125
105 J J Elser et al Biological stoichiometry from genes toecosystems Ecol Lett 3 540ndash550 (2000) doi 101046j1461-0248200000185x
106 E K Moody et al Consumption explains intraspecificvariation in nutrient recycling stoichiometry in a desert fishEcology 99 1552ndash1561 (2018) doi 101002ecy2372pmid 29882955
107 N Welti et al Bridging food webs ecosystem metabolismand biogeochemistry using ecological stoichiometry theoryFront Microbiol 8 1298 (2017) doi 103389fmicb201701298 pmid 28747904
108 M A Palmer S Filoso R M Fanelli From ecosystems toecosystem services Stream restoration as ecologicalengineering Ecol Eng 65 62ndash70 (2014) doi 101016jecoleng201307059
109 M J Burns T D Fletcher C J Walsh A R LadsonB E Hatt Hydrologic shortcomings of conventional urbanstormwater management and opportunities for reformLandsc Urban Plan 105 230ndash240 (2012) doi 101016jlandurbplan201112012
110 C R Liermann C Nilsson J Robertson R Y NgImplications of dam obstruction for global freshwater fishdiversity Bioscience 62 539ndash548 (2012) doi 101525bio20126265
111 J D Olden R J Naiman Incorporating thermal regimes intoenvironmental flows assessments Modifying dam operationsto restore freshwater ecosystem integrity Freshwater Biol55 86ndash107 (2010) doi 101111j1365-2427200902179x
112 D B Osmundson R J Ryel V L Lamarra J Pitlick Flowndashsedimentndashbiota relations Implications for river regulationeffects on native fish abundance Ecol Appl 12 1719ndash1739(2002)
113 D von Schiller et al Regulation causes nitrogen cyclingdiscontinuities in Mediterranean rivers Sci Total Environ540 168ndash177 (2016) doi 101016jscitotenv201507017pmid 26233271
114 J L Sabo et al Designing river flows to improve foodsecurity futures in the Lower Mekong Basin Science 358eaao1053 (2017) doi 101126scienceaao1053pmid 29217541
115 W Xu et al Improving the multi-objective performance ofrainwater harvesting systems using real-time controltechnology Water 10 147 (2018) doi 103390w10020147
116 M Acreman et al Environmental flows for naturalhybrid and novel riverine ecosystems in a changingworld Front Ecol Environ 12 466ndash473 (2014)doi 101890130134
117 N L Poff et al Sustainable water management under futureuncertainty with eco-engineering decision scaling Nat ClimChang 6 25ndash34 (2016) doi 101038nclimate2765
118 J D Kiernan P B Moyle P K Crain Restoring native fishassemblages to a regulated California stream using the
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 12 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
natural flow regime concept Ecol Appl 22 1472ndash1482(2012) doi 10189011-04801 pmid 22908707
119 D L Propst K B Gido Responses of native andnonnative fishes to natural flow regime mimicry in theSan Juan River Trans Am Fish Soc 133 922ndash931 (2004)doi 101577T03-0571
120 T K Pool J D Olden Assessing long‐term fish responses andshort‐term solutions to flow regulation in a dryland river basinEcol Freshwat Fish 24 56ndash66 (2015) doi 101111eff12125
121 W Chen J D Olden Designing flows to resolve human andenvironmental water needs in a dam-regulated riverNat Commun 8 2158 (2017) doi 101038s41467-017-02226-4 pmid 29255194
122 S M Nelson et al A history of the 2014 Minute 319environmental pulse flow as documented by fieldmeasurements and satellite imagery Ecol Eng 106733ndash748 (2017) doi 101016jecoleng201610040
123 P B Shafroth et al A large-scale environmental flowexperiment for riparian restoration in the Colorado RiverDelta Ecol Eng 106 645ndash660 (2017) doi 101016jecoleng201702016
124 E P Glenn P L Nagler P B Shafroth C J JarchowEffectiveness of environmental flows for riparian restorationin arid regions A tale of four rivers Ecol Eng 106 695ndash703(2017) doi 101016jecoleng201701009
125 S Hotaling D S Finn J Joseph Giersch D W WeisrockD Jacobsen Climate change and alpine stream biologyProgress challenges and opportunities for the futureBiol Rev Camb Philos Soc 92 2024ndash2045 (2017)doi 101111brv12319 pmid 28105701
126 C J Walsh T D Fletcher M J Burns Urban stormwaterrunoff A new class of environmental flow problem PLOS ONE7 e45814 (2012) doi 101371journalpone0045814pmid 23029257
127 A Selvakumar T P OrsquoConnor S D Struck Role of streamrestoration on improving benthic macroinvertebrates and in-stream water quality in an urban watershed Case studyJ Environ Eng 136 127ndash139 (2010) doi 101061(ASCE)EE1943-78700000116
128 R W Lammers B P Bledsoe What role does streamrestoration play in nutrient management Crit Rev EnvironSci Technol 47 335ndash371 (2017) doi 1010801064338920171318618
129 H E Golden N Hoghooghi Green infrastructure and itscatchment-scale effects An emerging science WIREs Water5 e1254 (2018) doi 101002wat21254 pmid 29682288
130 M R Williams G Bhatt S Filoso G Yactayo Streamrestoration performance and its contribution to theChesapeake Bay TMDL Challenges posed by climate changein urban areas Estuaries Coasts 40 1227ndash1246 (2017)doi 101007s12237-017-0226-1
131 R M Fanelli K Prestegaard M A Palmer Freshw Sci101086706072 (2019)
132 B W Abbott et al Unexpected spatial stability of waterchemistry in headwater stream networks Ecol Lett 21296ndash308 (2018) doi 101111ele12897 pmid 29282860
133 B L Brown C M Swan Dendritic network structureconstrains metacommunity properties in riverineecosystems J Anim Ecol 79 571ndash580 (2010) doi 101111j1365-2656201001668x pmid 20180874
134 C M Swan B L Brown Metacommunity theory meetsrestoration Isolation may mediate how ecologicalcommunities respond to stream restoration Ecol Appl 272209ndash2219 (2017) doi 101002eap1602 pmid 28718193
135 N Mouquet D Gravel F Massol V Calcagno Extending theconcept of keystone species to communities and ecosystemsEcol Lett 16 1ndash8 (2013) doi 101111ele12014 pmid 23062191
136 A Ruhiacute T Datry J L Sabo Interpreting beta-diversitycomponents over time to conserve metacommunities inhighly dynamic ecosystems Conserv Biol 31 1459ndash1468(2017) doi 101111cobi12906 pmid 28188969
137 L Yamane L W Botsford D P Kilduff Tracking restorationof population diversity via the portfolio effect J Appl Ecol55 472ndash481 (2018) doi 1011111365-266412978
138 D W Braidwood M A Taggart M Smith R AndersenTranslocations conservation and climate change Use ofrestoration sites as protorefuges and protorefugia RestorEcol 26 20ndash28 (2018) doi 101111rec12642
139 Y Wang D Wang Q W Lewis J Wu F Huang A frameworkto assess the cumulative impacts of dams on hydrologicalregime A case study of the Yangtze River Hydrol Processes31 3045ndash3055 (2017) doi 101002hyp11239
140 J L Sabo et al Pulsed flows tributary inputs and food-webstructure in a highly regulated river J Appl Ecol 551884ndash1895 (2018) doi 1011111365-266413109
141 C Magliozzi G Coro R C Grabowski A I PackmanS Krause A multiscale statistical method to identifypotential areas of hyporheic exchange for river restorationplanning Environ Model Softw 111 311ndash323 (2019)doi 101016jenvsoft201809006
142 S S Schwartz B Smith Restoring hydrologic function inurban landscapes with suburban subsoiling J Hydrol 543770ndash781 (2016) doi 101016jjhydrol201610051
143 C J Walsh T D Fletcher A R Ladson Stream restoration inurban catchments through redesigning stormwater systemsLooking to the catchment to save the stream J N AmBenthol Soc 24 690ndash705 (2005) doi 10189904-0201
144 N M Schmadel et al Thresholds of lake and reservoirconnectivity in river networks control nitrogen removalNat Commun 9 2779 (2018) doi 101038s41467-018-05156-x pmid 30018449
145 C D Sayer Conservation of aquatic landscapes Pondslakes and rivers as integrated systems WIREs Water 1573ndash585 (2014) doi 101002wat21045
146 R G Luthy D L Sedlak M H Plumlee D Austin V H ReshWastewater‐effluent‐dominated streams as ecosystem‐
management tools in a drier climate Front Ecol Environ 13477ndash485 (2015) doi 101890150038
147 D W Roscoe S G Hinch Effectiveness monitoring of fishpassage facilities Historical trends geographic patterns andfuture directions Fish Fish 11 12ndash33 (2010) doi 101111j1467-2979200900333x
148 C M Bunt T Castro-Santos A Haro Performanceof fish passage structures at upstream barriers tomigration River Res Appl 28 457ndash478 (2012)doi 101002rra1565
149 D Weigel et al Evaluation of a trap-and-transport programfor a threatened population of steelhead (Oncorhynchusmykiss) Conserv Genet 20 1195ndash1199 (2019) doi 101007s10592-019-01200-5
150 M H Bond et al Combined effects of barge transportationriver environment and rearing location on straying andmigration of adult Snake River fall-run Chinook SalmonTrans Am Fish Soc 146 60ndash73 (2017) doi 1010800002848720161235614
151 A C Ritchie et al Morphodynamic evolution followingsediment release from the worldrsquos largest dam removalSci Rep 8 13279 (2018) doi 101038s41598-018-30817-8pmid 30185796
152 T Sommer et al Californiarsquos Yolo Bypass Evidence thatflood control can be compatible with fisheries wetlandswildlife and agriculture Fisheries 26 6ndash16 (2001)doi 1015771548-8446(2001)026lt0006CYBgt20CO2
153 G Grill et al Mapping the worldrsquos free-flowing riversNature 569 215ndash221 (2019) doi 101038s41586-019-1111-9pmid 31068722
154 G Gassert M Luck M Landis P Reig T Shiao ldquoAqueductglobal maps 21 Constructing decision-relevant global waterrisk indicatorsrdquo Working paper World Resources Institute(2015) wwwwriorgpublicationaqueduct-global-maps-21-indicators
155 A Ruhiacute J D Olden J L Sabo Declining streamflow inducescollapse and replacement of native fishes in the AmericanSouthwest Front Ecol Environ 14 465ndash472 (2016)doi 101002fee1424
156 T C Gouhier A Grinsted V Simko R package Biwaveletconduct univariate and bivariate wavelet analyses (2018)
157 T Rodriacuteguez-Castillo E Esteacutevez A M Gonzaacutelez-FerrerasJ Barquiacuten Estimating ecosystem metabolism toentire river networks Ecosystems (2018) doi 101007s10021-018-0311-8
ACKNOWLEDGMENTS
We greatly appreciate those who read and provided importantinsights on drafts of this paper A Armstrong K HondulaJ Hosen K Leathers G de Mendoza C Maietta I MunozJ Tonkin and two anonymous reviewers We also thank T Tu forhelp with Fig 1 and S Carlson L Comte T Datry J HosenM Kondolf N Jones D Lytle E Moody J Olden M PowerJ Sabo and M Sanchez-Montoya for valuable discussionsCompeting interests The authors have no competing interestsFunding MAP was supported by funds from the NSF (1639145)and AR was supported by funds from the University of CaliforniaBerkeley and NSF (1802714)
101126scienceaaw2087
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 13 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
restorationLinkages between flow regime biota and ecosystem processes Implications for river
Margaret Palmer and Albert Ruhi
DOI 101126scienceaaw2087 (6459) eaaw2087365Science
this issue p eaaw2087Scienceflow variability and achieve hydrological connectivity between a river and its surroundingsand ecosystem processes To be successful such efforts must go beyond accounting for flood pulses to restore natural mimic ecologically important aspects of natural flow regimes guided by insights into how variations in flow affect biotawater quality and ecological processes In a Review Palmer and Ruhi explain how restoration designs now attempt to
Human activities have altered the flow regimes of many of Earths rivers with negative impacts on biodiversityRiver restoration guided by research
ARTICLE TOOLS httpsciencesciencemagorgcontent3656459eaaw2087
REFERENCES
httpsciencesciencemagorgcontent3656459eaaw2087BIBLThis article cites 147 articles 7 of which you can access for free
PERMISSIONS httpwwwsciencemagorghelpreprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAASScienceScience 1200 New York Avenue NW Washington DC 20005 The title (print ISSN 0036-8075 online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
Science No claim to original US Government WorksCopyright copy 2019 The Authors some rights reserved exclusive licensee American Association for the Advancement of
on May 27 2020
httpsciencesciencem
agorgD
ownloaded from

natural flow regime concept Ecol Appl 22 1472ndash1482(2012) doi 10189011-04801 pmid 22908707
119 D L Propst K B Gido Responses of native andnonnative fishes to natural flow regime mimicry in theSan Juan River Trans Am Fish Soc 133 922ndash931 (2004)doi 101577T03-0571
120 T K Pool J D Olden Assessing long‐term fish responses andshort‐term solutions to flow regulation in a dryland river basinEcol Freshwat Fish 24 56ndash66 (2015) doi 101111eff12125
121 W Chen J D Olden Designing flows to resolve human andenvironmental water needs in a dam-regulated riverNat Commun 8 2158 (2017) doi 101038s41467-017-02226-4 pmid 29255194
122 S M Nelson et al A history of the 2014 Minute 319environmental pulse flow as documented by fieldmeasurements and satellite imagery Ecol Eng 106733ndash748 (2017) doi 101016jecoleng201610040
123 P B Shafroth et al A large-scale environmental flowexperiment for riparian restoration in the Colorado RiverDelta Ecol Eng 106 645ndash660 (2017) doi 101016jecoleng201702016
124 E P Glenn P L Nagler P B Shafroth C J JarchowEffectiveness of environmental flows for riparian restorationin arid regions A tale of four rivers Ecol Eng 106 695ndash703(2017) doi 101016jecoleng201701009
125 S Hotaling D S Finn J Joseph Giersch D W WeisrockD Jacobsen Climate change and alpine stream biologyProgress challenges and opportunities for the futureBiol Rev Camb Philos Soc 92 2024ndash2045 (2017)doi 101111brv12319 pmid 28105701
126 C J Walsh T D Fletcher M J Burns Urban stormwaterrunoff A new class of environmental flow problem PLOS ONE7 e45814 (2012) doi 101371journalpone0045814pmid 23029257
127 A Selvakumar T P OrsquoConnor S D Struck Role of streamrestoration on improving benthic macroinvertebrates and in-stream water quality in an urban watershed Case studyJ Environ Eng 136 127ndash139 (2010) doi 101061(ASCE)EE1943-78700000116
128 R W Lammers B P Bledsoe What role does streamrestoration play in nutrient management Crit Rev EnvironSci Technol 47 335ndash371 (2017) doi 1010801064338920171318618
129 H E Golden N Hoghooghi Green infrastructure and itscatchment-scale effects An emerging science WIREs Water5 e1254 (2018) doi 101002wat21254 pmid 29682288
130 M R Williams G Bhatt S Filoso G Yactayo Streamrestoration performance and its contribution to theChesapeake Bay TMDL Challenges posed by climate changein urban areas Estuaries Coasts 40 1227ndash1246 (2017)doi 101007s12237-017-0226-1
131 R M Fanelli K Prestegaard M A Palmer Freshw Sci101086706072 (2019)
132 B W Abbott et al Unexpected spatial stability of waterchemistry in headwater stream networks Ecol Lett 21296ndash308 (2018) doi 101111ele12897 pmid 29282860
133 B L Brown C M Swan Dendritic network structureconstrains metacommunity properties in riverineecosystems J Anim Ecol 79 571ndash580 (2010) doi 101111j1365-2656201001668x pmid 20180874
134 C M Swan B L Brown Metacommunity theory meetsrestoration Isolation may mediate how ecologicalcommunities respond to stream restoration Ecol Appl 272209ndash2219 (2017) doi 101002eap1602 pmid 28718193
135 N Mouquet D Gravel F Massol V Calcagno Extending theconcept of keystone species to communities and ecosystemsEcol Lett 16 1ndash8 (2013) doi 101111ele12014 pmid 23062191
136 A Ruhiacute T Datry J L Sabo Interpreting beta-diversitycomponents over time to conserve metacommunities inhighly dynamic ecosystems Conserv Biol 31 1459ndash1468(2017) doi 101111cobi12906 pmid 28188969
137 L Yamane L W Botsford D P Kilduff Tracking restorationof population diversity via the portfolio effect J Appl Ecol55 472ndash481 (2018) doi 1011111365-266412978
138 D W Braidwood M A Taggart M Smith R AndersenTranslocations conservation and climate change Use ofrestoration sites as protorefuges and protorefugia RestorEcol 26 20ndash28 (2018) doi 101111rec12642
139 Y Wang D Wang Q W Lewis J Wu F Huang A frameworkto assess the cumulative impacts of dams on hydrologicalregime A case study of the Yangtze River Hydrol Processes31 3045ndash3055 (2017) doi 101002hyp11239
140 J L Sabo et al Pulsed flows tributary inputs and food-webstructure in a highly regulated river J Appl Ecol 551884ndash1895 (2018) doi 1011111365-266413109
141 C Magliozzi G Coro R C Grabowski A I PackmanS Krause A multiscale statistical method to identifypotential areas of hyporheic exchange for river restorationplanning Environ Model Softw 111 311ndash323 (2019)doi 101016jenvsoft201809006
142 S S Schwartz B Smith Restoring hydrologic function inurban landscapes with suburban subsoiling J Hydrol 543770ndash781 (2016) doi 101016jjhydrol201610051
143 C J Walsh T D Fletcher A R Ladson Stream restoration inurban catchments through redesigning stormwater systemsLooking to the catchment to save the stream J N AmBenthol Soc 24 690ndash705 (2005) doi 10189904-0201
144 N M Schmadel et al Thresholds of lake and reservoirconnectivity in river networks control nitrogen removalNat Commun 9 2779 (2018) doi 101038s41467-018-05156-x pmid 30018449
145 C D Sayer Conservation of aquatic landscapes Pondslakes and rivers as integrated systems WIREs Water 1573ndash585 (2014) doi 101002wat21045
146 R G Luthy D L Sedlak M H Plumlee D Austin V H ReshWastewater‐effluent‐dominated streams as ecosystem‐
management tools in a drier climate Front Ecol Environ 13477ndash485 (2015) doi 101890150038
147 D W Roscoe S G Hinch Effectiveness monitoring of fishpassage facilities Historical trends geographic patterns andfuture directions Fish Fish 11 12ndash33 (2010) doi 101111j1467-2979200900333x
148 C M Bunt T Castro-Santos A Haro Performanceof fish passage structures at upstream barriers tomigration River Res Appl 28 457ndash478 (2012)doi 101002rra1565
149 D Weigel et al Evaluation of a trap-and-transport programfor a threatened population of steelhead (Oncorhynchusmykiss) Conserv Genet 20 1195ndash1199 (2019) doi 101007s10592-019-01200-5
150 M H Bond et al Combined effects of barge transportationriver environment and rearing location on straying andmigration of adult Snake River fall-run Chinook SalmonTrans Am Fish Soc 146 60ndash73 (2017) doi 1010800002848720161235614
151 A C Ritchie et al Morphodynamic evolution followingsediment release from the worldrsquos largest dam removalSci Rep 8 13279 (2018) doi 101038s41598-018-30817-8pmid 30185796
152 T Sommer et al Californiarsquos Yolo Bypass Evidence thatflood control can be compatible with fisheries wetlandswildlife and agriculture Fisheries 26 6ndash16 (2001)doi 1015771548-8446(2001)026lt0006CYBgt20CO2
153 G Grill et al Mapping the worldrsquos free-flowing riversNature 569 215ndash221 (2019) doi 101038s41586-019-1111-9pmid 31068722
154 G Gassert M Luck M Landis P Reig T Shiao ldquoAqueductglobal maps 21 Constructing decision-relevant global waterrisk indicatorsrdquo Working paper World Resources Institute(2015) wwwwriorgpublicationaqueduct-global-maps-21-indicators
155 A Ruhiacute J D Olden J L Sabo Declining streamflow inducescollapse and replacement of native fishes in the AmericanSouthwest Front Ecol Environ 14 465ndash472 (2016)doi 101002fee1424
156 T C Gouhier A Grinsted V Simko R package Biwaveletconduct univariate and bivariate wavelet analyses (2018)
157 T Rodriacuteguez-Castillo E Esteacutevez A M Gonzaacutelez-FerrerasJ Barquiacuten Estimating ecosystem metabolism toentire river networks Ecosystems (2018) doi 101007s10021-018-0311-8
ACKNOWLEDGMENTS
We greatly appreciate those who read and provided importantinsights on drafts of this paper A Armstrong K HondulaJ Hosen K Leathers G de Mendoza C Maietta I MunozJ Tonkin and two anonymous reviewers We also thank T Tu forhelp with Fig 1 and S Carlson L Comte T Datry J HosenM Kondolf N Jones D Lytle E Moody J Olden M PowerJ Sabo and M Sanchez-Montoya for valuable discussionsCompeting interests The authors have no competing interestsFunding MAP was supported by funds from the NSF (1639145)and AR was supported by funds from the University of CaliforniaBerkeley and NSF (1802714)
101126scienceaaw2087
Palmer et al Science 365 eaaw2087 (2019) 20 September 2019 13 of 13
RESEARCH | REVIEWon M
ay 27 2020
httpsciencesciencemagorg
Dow
nloaded from
restorationLinkages between flow regime biota and ecosystem processes Implications for river
Margaret Palmer and Albert Ruhi
DOI 101126scienceaaw2087 (6459) eaaw2087365Science
this issue p eaaw2087Scienceflow variability and achieve hydrological connectivity between a river and its surroundingsand ecosystem processes To be successful such efforts must go beyond accounting for flood pulses to restore natural mimic ecologically important aspects of natural flow regimes guided by insights into how variations in flow affect biotawater quality and ecological processes In a Review Palmer and Ruhi explain how restoration designs now attempt to
Human activities have altered the flow regimes of many of Earths rivers with negative impacts on biodiversityRiver restoration guided by research
ARTICLE TOOLS httpsciencesciencemagorgcontent3656459eaaw2087
REFERENCES
httpsciencesciencemagorgcontent3656459eaaw2087BIBLThis article cites 147 articles 7 of which you can access for free
PERMISSIONS httpwwwsciencemagorghelpreprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAASScienceScience 1200 New York Avenue NW Washington DC 20005 The title (print ISSN 0036-8075 online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
Science No claim to original US Government WorksCopyright copy 2019 The Authors some rights reserved exclusive licensee American Association for the Advancement of
on May 27 2020
httpsciencesciencem
agorgD
ownloaded from

restorationLinkages between flow regime biota and ecosystem processes Implications for river
Margaret Palmer and Albert Ruhi
DOI 101126scienceaaw2087 (6459) eaaw2087365Science
this issue p eaaw2087Scienceflow variability and achieve hydrological connectivity between a river and its surroundingsand ecosystem processes To be successful such efforts must go beyond accounting for flood pulses to restore natural mimic ecologically important aspects of natural flow regimes guided by insights into how variations in flow affect biotawater quality and ecological processes In a Review Palmer and Ruhi explain how restoration designs now attempt to
Human activities have altered the flow regimes of many of Earths rivers with negative impacts on biodiversityRiver restoration guided by research
ARTICLE TOOLS httpsciencesciencemagorgcontent3656459eaaw2087
REFERENCES
httpsciencesciencemagorgcontent3656459eaaw2087BIBLThis article cites 147 articles 7 of which you can access for free
PERMISSIONS httpwwwsciencemagorghelpreprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAASScienceScience 1200 New York Avenue NW Washington DC 20005 The title (print ISSN 0036-8075 online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
Science No claim to original US Government WorksCopyright copy 2019 The Authors some rights reserved exclusive licensee American Association for the Advancement of
on May 27 2020
httpsciencesciencem
agorgD
ownloaded from