lessons learned from a decade of assessment and restoration studies of benthic invertebrates and...
TRANSCRIPT

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, researchlibraries, and research funders in the common goal of maximizing access to critical research.
Lessons Learned from a Decade of Assessment and Restoration Studies of BenthicInvertebrates and Submersed Aquatic Vegetation in Lake PontchartrainAuthor(s): Michael A. Poirrier, Elizabeth A. Spalding, and Carol D. FranzeSource: Journal of Coastal Research, Number 10054:88-100. 2009.Published By: Coastal Education and Research FoundationDOI: http://dx.doi.org/10.2112/SI54-005.1URL: http://www.bioone.org/doi/full/10.2112/SI54-005.1
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, andenvironmental sciences. BioOne provides a sustainable online platform for over 170 journals and books publishedby nonprofit societies, associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance ofBioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiriesor rights and permissions requests should be directed to the individual publisher as copyright holder.

Lessons Learned from a Decade of Assessment andRestoration Studies of Benthic Invertebrates andSubmersed Aquatic Vegetation in Lake Pontchartrain
Michael A. Poirrier{, Elizabeth A. Spalding{, and Carol D. Franze{
{Estuarine Research LaboratoryDepartment of Biological
Sciences and thePontchartrain Institute forEnvironmental Sciences
University of New OrleansLakefront Campus
2000 Lakeshore DriveNew Orleans, LA 70148, [email protected]
{Louisiana Sea Grant Marine Extension/LSU Ag Center Research & Extension
21549 Old Covington HighwayHammond, LA 70403, U.S.A.
ABSTRACT
POIRRIER, M.A.; SPALDING, E.A., and FRANZE, C.D., 2009. Lessons learned from a decade of assessment andrestoration studies of benthic invertebrates and submersed aquatic vegetation in Lake Pontchartrain. Journal of CoastalResearch, SI(54), 88–100. West Palm Beach (Florida), ISSN 0749-0208.
Studies of Lake Pontchartrain benthic invertebrates and submersed aquatic vegetation (SAV) from 1996 through 2005provided the opportunity to determine community responses to severe disturbances including: recovery from shelldredging; effects of anoxia and hypoxia from saltwater intrusion; the 1997 Bonnet Carre Spillway opening; a prolongeddrought resulting from an El Nino Southern Oscillation; and effects of Hurricane Katrina. We included reviews andupdates of our recently published work, an integrated analysis of findings, and restoration recommendations. Duringthis decadal study there were no prolonged periods of ‘‘normal’’ conditions. Instead, benthos and SAV were experiencingor recovering from significant temporal disturbances. This made it difficult to use the abundance of a particular set oforganisms at a point in time to evaluate habitat quality or restoration success without considering past disturbanceeffects. Benthos species diversity and the relative abundance of stress tolerant groups such as annelids were goodindicators of short-term adverse conditions, but rapid changes occurred in response to salinity and dissolved oxygen.Rangia cuneata was a good indicator of the extent of hypoxia and long-term damage from hurricanes. The distributionand abundance of SAV decreased with turbidity and nutrient increases, and Vallisneria americana and other freshwaterSAV decreased with prolonged salinity increases. Resistance to and resilience after disturbances and natural changesduring long term cycles have to be considered in evaluating habitat condition and restoration success.
ADDITIONAL INDEX WORDS: Estuary, shell dredging, spillway, ENSO, drought, hurricane, salinity, turbidity,water clarity, saltwater intrusion, hypoxia, Rangia cuneata, Ischadium recurvum, species diversity, SAV, Ruppiamaritima, Vallisneria Americana.
INTRODUCTION
Lake Pontchartrain is a large, shallow, oligohaline estuary
located in a deteriorating delta of the Mississippi River. The
estuary is disturbed by both natural and human factors. Lake
Pontchartrain (Figure 1) has a mean salinity of 3.9 ppt
(Francis et al., 1994), a mean depth of 3.7 m, and a surface
area of 1630 km2 (Sikora and Kjerfve, 1985). Salinity
increases from west to east (Swenson, 1980). Freshwater
enters via streams to the west and north, outfall canals on the
south shore, and periodic openings and leakage of the Bonnet
Carre Spillway. Higher salinity water enters from eastern
tidal passes and the Inner Harbor Navigation Canal (IHNC),
which is connected to the Mississippi River Gulf Outlet
(MRGO). From 1933 to 1990, accumulated Rangia cuneata
shells were dredged from the lake bottom and used for the
construction of roads, parking lots, levees, and the production
of cement (Sikora and Sikora, 1981, 1982; USACE, 1987).
Current adverse effects include urban and agricultural
runoff, shoreline modification, Mississippi River flood diver-
sions (Penland et al., 2001), and saltwater intrusion that
causes salinity stratification and bottom water anoxia and
hypoxia (Abadie and Poirrier, 2000).
Benthic invertebrates are generally accepted as meaning-
ful indicators of the environmental health of estuaries
(Boesch and Rosenberg, 1981; Diaz and Rosenberg, 1995;
Reish, 1986; Wilson, 1994). Because they cannot move away
from adverse conditions, they are continuously exposed to
environmental stress and therefore must respond to episodic
environmental disturbances (Bilyard, 1987; Moore, 1992).DOI:10.2112/SI54-005.1.
Journal of Coastal Research SI 54 88–100 West Palm Beach, Florida Fall 2009

Also because of their sessile nature, differences in their
temporal and spatial community composition are easily
sampled and quantified. Oligohaline assemblages have low
species richness (Remane and Schleiper, 1971) and are
controlled more by abiotic factors than persistent biotic
interactions (Boesch and Rosenberg, 1981; Gunter, 1960). They
are composed of biologically diverse taxa that provide impor-
tant links in food chains and water quality processes. Rangia
cuneata (Bivalvia:Mactridae) or ‘‘common rangia’’ is the
dominant member of the benthos in oligohaline Gulf Coast
estuaries based on number of individuals (Hopkins and
Andrews, 1970), total biomass (Cain, 1975; Odum, 1967), life
span, and ecosystem interactions (Darnell, 1958; LaSalle and
de la Cruz, 1985; Peterson and Heck, 1999; Vaughn and
Hakenkamp, 2001). It was a principal indicator of the condition
of bottom habitat in our studies.
The first report on Lake Pontchartrain benthic inverte-
brates was by Suttkus, Darnell, and Darnell (1954). They
found an average of 95 large R. cuneata per square meter, and
their study included areas near the mouth of IHNC. In later
studies conducted from 1969 to 1972, Tarver and Dugas
(1973) found that the average density of large R. cuneata in
the lake was 39 clams/m2, and they did not find clams larger
than 16 mm in areas that were continually dredged. Poirrier
(1978) documented salinity stratification and low dissolved
oxygen associated with saltwater intrusion from the IHNC
and mentioned possible adverse effects on benthic organisms.
In an ecological characterization of the benthic community of
Lake Pontchartrain, Sikora and Sikora (1982) found that
total number of species in Lake Pontchartrain was lower than
expected for brackish systems and that all measures of
benthic community structure indicate a stressed community.
Junot, Poirrier, and Soniat (1983) documented an adverse
impact on benthos near the mouth of the IHNC due to low
dissolved oxygen from salinity stratification. Poirrier et al.
(1984) collaborated with Schurtz and St. Pe (1984) in a study
of bottom dissolved oxygen concentrations resulting from
salinity stratification from the IHNC and effects on benthic
invertebrates. They attributed the ‘‘dead zone’’ reported by
Sikora and Sikora (1982) to low dissolved oxygen and other
conditions associated with salinity stratification. The USACE
(1987) prepared a report on the effects of shell dredging that
summarized the literature and possible impact on benthic
organisms. In a study of the Duncan outfall canal, Powers,
Poirrier, and Yund (1992) found that nematodes, oligo-
chaetes, and capitellid polychaetes are abundant in areas
affected by urban runoff, but the densities of the molluscan
species R. cuneata, Texadina sphinctostoma, and Probythi-
nella protera were low and increased with distance from the
source of pollutants. Spalding Walker, and Poirrier (2007)
quantified the benefits of restoring R. cuneata through
complete closure of the MRGO using clam clearance rates of
four phytoplankton species, silts and clays, and Escherichia
coli. They estimated the time it would take R. cuneata to filter
a volume of water equal to the lake before and after
restoration using clam density, relationships between length
and dry weight biomass, and filtration rates.
The reestablishment of historic submersed aquatic vege-
tation (SAV) is a realistic and measurable Lake Pontchar-
train restoration goal. The former dominant species, Vallis-
neria americana, declined by over 50% since first studied in
1953 (Burns and Poirrier, 1996). With SAV abundance
rapidly decreasing, further disturbance, whether it is
natural or anthropogenic, could possibly decimate remaining
populations. Factors that control SAV distribution and
abundance in Lake Pontchartrain are complex, but the
principal direct driver is light availability (Cho and Poirrier,
2005a; Dennison et al., 1993), which is adversely affected by
increased turbidity and algal epiphyte growth. Eutrophica-
tion increases turbidity and epiphyte growth. Thus, SAV is a
good indicator of water clarity, eutrophication trends, and
results of restoration efforts (Cho and Poirrier, 2005a,
2005b).
Submersed aquatic vegetation distribution in Lake
Pontchartrain was initially described in a report by Suttkus,
Darnell, and Darnell (1954). Vallisneria americana and R.
maritima occurred on the north shore at a site just east of
the Tchefuncte River and from Lewisburg eastward to the
Rigolets. On the southshore, SAV was observed from Indian
Beach to just east of South Point; however, the eastern lobe
of the lake was not surveyed. Montz (1978) surveyed the
lake in 1973 to determine the effects of the Bonnet Carre
Spillway opening on SAV and reported no effects of the
opening on SAV in the lake. The 1973 diversion occurred
coincident with shell dredging in the lake, which maintained
high turbidity and probably prevented cyanobacteria blooms
observed with nutrient introductions after shell dredging
ceased (Brammer et al., 2007). Montz (1978) found two
additional species, Najas guadalupensis and Potamogeton
perfoliatus. Poirrier (personal observation) observed an
exotic species, Myriophyllum spicatum, after the 1973
Spillway opening.
Turner, Darnell, and Bond (1980) compared the surveys by
Suttkus, Darnell, and Darnell (1954), and Montz (1978)
documented a 25% to 35% decrease in SAV. In 1985, Mayer
(1986) conducted a lakewide survey similar to Montz (1978)
Figure 1. Benthic invertebrate and submersed aquatic vegetation survey
sites in Lake Pontchartrain, Louisiana.
Benthic Invertebrates and SAV in Lake Pontchartrain 89
Journal of Coastal Research, Special Issue No. 54, 2009

and found a 50% decrease in SAV between 1973 and 1985.
The maximum depth distribution for SAV in 1985 was 1.2 m
(Mayer, 1986), whereas maximum depth distribution report-
ed from earlier studies was 1.8 m (Montz, 1978; Suttkus,
Darnell, and Darnell, 1954). Burns, Poirrier, and Preston
(1993) conducted SAV surveys in 1991 and 1992. A compar-
ison of their findings to those of Mayer (1986), showed a 17%
decrease from 1985 to 1992. During the 1991–1992 surveys,
SAV foliar cover was greatest in the 0.3- to 0.6-m water depth
contour (Burns, Poirrier, and Preston, 1993). Total areal
cover for SAV in 1991–1992 was 88 ha (24 ha when corrected
for patchiness). Submersed aquatic vegetation foliar cover
along the north shore totaled 76 ha and along the urbanized
south shore it totaled 12 ha (Burns, Poirrier, and Preston,
1993). A comparison of the 1992 and 1993 surveys showed a
74% reduction of SAV areal coverage after passage of
Hurricane Andrew in August 1992 and a 95% reduction on
the south shore after the passage of a severe winter storm in
March 1993 (Burns and Poirrier, 1996). Potamogeton perfo-
liatus, a rare Louisiana plant whose westernmost distribution
is Lake Pontchartrain, was observed at five sites during the
1991 to 1993 surveys (Burns, Poirrier, and Preston, 1995).
Benthos and SAV provide essential habitat for sport and
commercial finfish, shellfish, and wildlife including waterfowl
and the manatee (Cottam, 1939; Darnell, 1958; Ebersole and
Kennedy, 1995; LaSalle and de la Cruz, 1985; Orth, Heck,
and van Montfrans, 1984; Perry and Uhler, 1981, 1988;
Richman and Lovvorn, 2004). In addition to long-term
monitoring and assessment, our studies focused on ecological
factors and environmental stressors that determine the
distribution and abundance of these critical habitats. An
important aspect of our studies was the determination of the
cause of habitat loss and provision of strategies needed for
sustained restoration. Restoration of these essential habitats
will result in expanded fish and wildlife resources, and
aquatic recreational opportunities.
Major research objectives were to distinguish anthropo-
genic stressors from natural abiotic and biotic ecological
drivers. Often anthropogenic perturbations work in concert
with natural environmental drivers, making organisms more
sensitive to natural stress and producing fragile communi-
ties that are unable to rapidly recover from natural
disturbances. Some of these disturbances cause chronic
stress, producing gradual long-term changes; others might
be acute, but episodic, producing periodic severe disturbanc-
es. Our studies focused on the effects of severe natural and
anthropogenic disruptions that included recovery from shell
dredging; effects of anoxia and hypoxia from MRGO
saltwater intrusion; the 1997 opening of the Bonnet Carre
Spillway; a prolonged drought resulting from an El Nino
Southern Oscillation (ENSO); and a severe disturbance by
Hurricane Katrina. Each of these disturbances challenged
the resistance and resilience of the benthic invertebrate and
SAV communities, and provided natural experiments on the
response of species populations and community structure to
disturbance. We present original research incorporated with
reviews and updates of our recently published work to
provide an integrated analysis of findings from 1996 through
2005.
MATERIALS AND METHODS
Benthos
Infaunal invertebrates were sampled November 1996–
November 2005 along an east-to-west transect at five fixed
sites and March 1997–November 2005 along a north-to-south
transect at six fixed sites as part of a long-term monitoring
program (Figure 1). Additional benthic invertebrate samples
were collected at 33 sites along 8 transect lines spaced evenly
from west to east across the lake in 1997, November and April
2004, and 2005 (Figure 1). Salinity and dissolved oxygen were
measured during each sampling episode with a YSI 85 S-C-T,
D.O meter. These sites have been used in our previous studies
of benthic invertebrates (for specific methods see Abadie and
Poirrier, 2000; Brammer et al., 2007; Poirrier, Rodriguez del
Rey, and Spalding, 2008).
Three replicate infaunal samples were obtained with a
15 cm 3 15 cm petite Ponar dredge, rinsed in the field
through a 0.6-mm sieve, and the retained material fixed in a
10% buffered formalin solution and stained with Rose bengal.
In the laboratory, samples were rinsed on a 0.5-mm sieve, and
macrofauna were identified to the lowest feasible taxon and
enumerated using a dissecting microscope. Taxonomic desig-
nations followed the Integrated Taxonomic Information
System. Clams were identified to species, sized into 5-mm
classes, and counted. Because of our inability to visually
distinguish the difference between R. cuneata and Mulinia
lateralis, clams smaller than 6 mm were excluded.
We also conducted studies to quantify possible benefits of
restoring R. cuneata through closure of the MRGO (Spalding,
Walker, and Poirrier, 2007). Static, clam clearance rate
experiments using four different phytoplankton species, silts
and clays, and Escherichia coli were used to quantify effects
of clam filtration.
Submersed Aquatic Vegetation
Surveys of SAV abundance to determine areal coverage
along a littoral depth gradient were conducted during late fall
from 1996 through 2005 at Lacombe, Pointe aux Herbes, and
Fontainebleau State Park. A location at Goose Point was
added in 1999, and Lincoln Beach was added in 2000
(Figure 1). These locations have been used in our previous
studies of SAV depth distribution and areal coverage (for
specific methods see Burns and Poirrier, 1996; Burns,
Poirrier, and Preston, 1993; Cho, 2003; Cho and Poirrier,
2005a, 2005b, 2005c). At each location, five random transects
were surveyed. Transects were perpendicular to the shoreline
and extended from the shoreline to a water depth of
maximum colonization. The line intercept method was used
(Gertz, 1984). While snorkeling, species of SAV intercepted by
the transects were identified and distance covered by each
species was recorded in centimeters by direct observation.
Statistical Analyses
Plymouth Routines in Multivariate Ecology (Clark and
Gorley, 2001) were used to analyze macroinvertebrate
community structure. Univariate analyses of variance (AN-
90 Poirrier, Spalding and Franze
Journal of Coastal Research, Special Issue No. 54, 2009

OVA) were performed using general linear model procedures
in SPSS to test for differences in bottom salinity, Shannon-
Wiener (loge) species diversity, and R. cuneata density from
1996 to 2005. Species diversity data were limited to the period
from May 1998 through August 2002 to determine the effect
of the ENSO-induced drought. Data were tested for meeting
parametric analysis criteria and a level of 0.05 was used to
determine significance. Although species diversity and R.
cuneata density exhibited mild departures from normality,
the ANOVA technique is considered robust, and data
exhibiting mild departures from normality may be considered
acceptable for analysis without transformation (Neter, Was-
serman, and Kutner, 1990). Post hoc pairwise comparisons
were made to determine significant differences in means
using the Bonferroni method.
ArcMap GIS software kriging function was used to spatially
analyze the distribution of R. cuneata biomass and the extent
of dissolved oxygen concentration at the mouth of the IHNC
in August 2006. Measured data values were weighted to
derive a prediction for unmeasured locations based on the
distance between the measured sites; the prediction locations;
and the overall spatial arrangement among the measured
sites. Semivariograms were created and then fitted to the
most appropriate model to analyze the spatial correlation
between sites. Microstructure and measurement error were
estimated using replicate samples at each transect site.
Standard error maps were created and evaluated, and cross
validations were performed.
RESULTS
We integrated published and unpublished data on changes
in benthic invertebrates and SAV communities to examine
trends over a 10-year period. A popular overarching assump-
tion is if environmental conditions in Lake Pontchartrain are
improving, population trends in these communities should be
shifting to the historic distribution and abundance of key
species and over time indices of community structure should
indicate less stress. However, during this study it became
clear that communities were regularly affected by diverse
disturbances that challenged evaluation of long-term trends.
This synthesis examines responses to disruptions and
extracts trends, ecologic drivers, and restoration strategies
from the long-term data set.
Recovery from Shell Dredging
Benthos
Abadie and Poirrier (2000) compared densities of large
($21 mm) R. cuneata in studies before and after 1990 when
shell dredging ended to determine if a density increase had
occurred. In 1954 R. cuneata were abundant (95 clams/m2).
They were less abundant in 1973 (39 clams/m2), 1982 (2
clams/m2), and 1984 (41 clams/m2) as the intensity of shell
dredging increased. Samples obtained in 1996 showed an
increase (126 clams/m2) that was comparable to densities in
1954. Clam densities in 1954 and 1996 were statistically
different from 1973, 1982, and 1984, but not different from
each other. These recovery studies indicated that shell
dredging had a significant impact on the bottom community.
Submersed Aquatic Vegetation
Submersed aquatic vegetation distribution and abundance
were much lower when this study began in 1996 (Figure 2)
than historic levels (Suttkus, Darnell, and Darnell, 1954).
Hurricane Andrew damaged north shore beds in 1992 and a
winter storm damaged south shore beds in 1993. Recovery
from these weather events occurred, but beds did not increase
to 1991 levels (Burns and Poirrier, 1996). Surveys in 1996 did
not indicate an increase in SAV due to cessation of shell
dredging (Figure 2), and an overall increase in SAV did not
occur until 1998 (Cho and Poirrier, 2001).
Effects of Anoxia and Hypoxia from MRGOSaltwater Intrusion
Benthos
Abadie and Poirrier (2000) demonstrated increased abun-
dance of large R. cuneata after shell dredging ended and also
found that large R. cuneata ($21 mm) were absent from a large
area north of the IHNC. Later studies by Abadie and Poirrier
(2001a, 2001b) determined that the affected area was approx-
imately 250 km2 (100 mi2) and was caused by episodes of low
dissolved oxygen due to salinity stratification from more saline
water entering from the MRGO through connections with the
IHNC. Over the course of this study, episodic hypoxia has been
documented as far north as site NS2, 24 km (15 mi) from the
mouth of the IHNC, and hypoxia has occurred regularly at site
NS3B, 17 km (10.5 mi) north of the IHNC (Figure 3). Periodic
samples of benthic invertebrates from north–south transects
through the ‘‘dead zone’’ from 1997 through 2006 demonstrated
the regular occurrence of Shannon-Wiener species diversity
values often in the range of 0.0 to 1.0 (Figure 4). Anoxia and
hypoxia typically is found at the mouth of the IHNC as in
August 2006 (Figure 5). Abadie and Poirrier (2001a) compared
the abundance of large R. cuneata with other indicator taxa and
concluded that these could be used to monitor the area affected
Figure 2. Average annual abundance of submersed aquatic vegetation
(m) by dominant species from fall 1996 to spring 2005 (1996 to 1998, n 5
15; 1999, n 5 20; 2000 to 2006, n 5 25).
Benthic Invertebrates and SAV in Lake Pontchartrain 91
Journal of Coastal Research, Special Issue No. 54, 2009
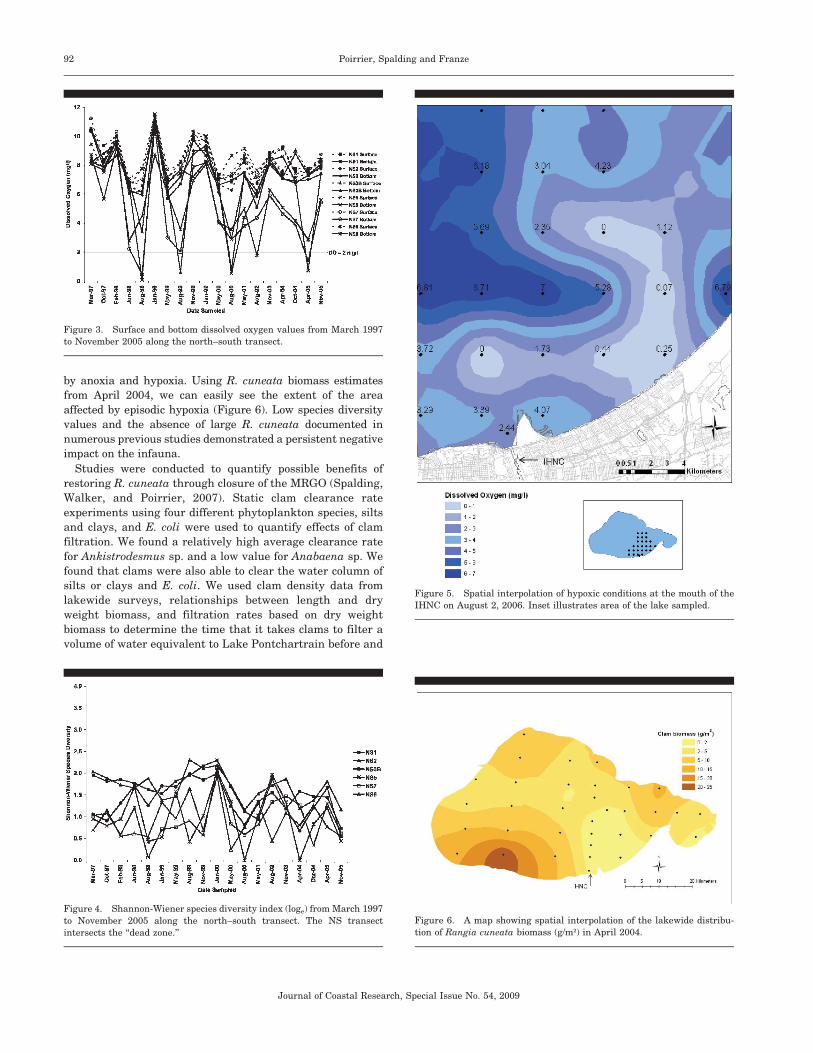
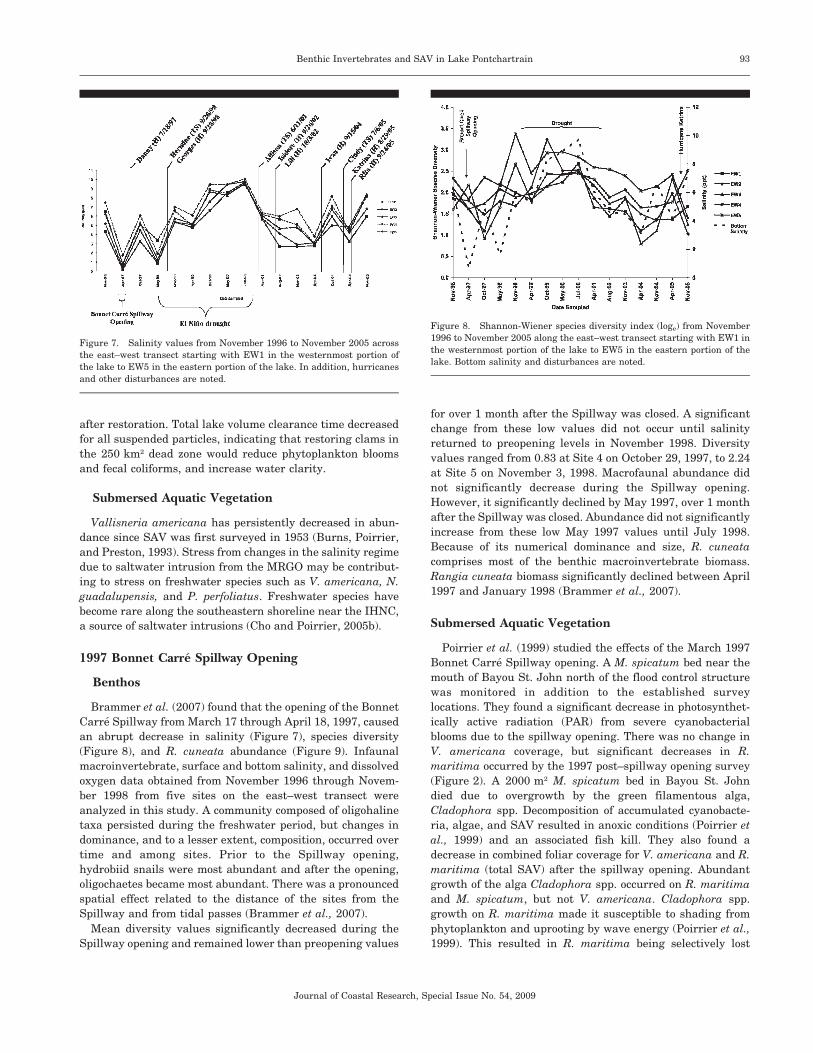
by anoxia and hypoxia. Using R. cuneata biomass estimates
from April 2004, we can easily see the extent of the area
affected by episodic hypoxia (Figure 6). Low species diversity
values and the absence of large R. cuneata documented in
numerous previous studies demonstrated a persistent negative
impact on the infauna.
Studies were conducted to quantify possible benefits of
restoring R. cuneata through closure of the MRGO (Spalding,
Walker, and Poirrier, 2007). Static clam clearance rate
experiments using four different phytoplankton species, silts
and clays, and E. coli were used to quantify effects of clam
filtration. We found a relatively high average clearance rate
for Ankistrodesmus sp. and a low value for Anabaena sp. We
found that clams were also able to clear the water column of
silts or clays and E. coli. We used clam density data from
lakewide surveys, relationships between length and dry
weight biomass, and filtration rates based on dry weight
biomass to determine the time that it takes clams to filter a
volume of water equivalent to Lake Pontchartrain before and
Figure 3. Surface and bottom dissolved oxygen values from March 1997
to November 2005 along the north–south transect.
Figure 4. Shannon-Wiener species diversity index (loge) from March 1997
to November 2005 along the north–south transect. The NS transect
intersects the ‘‘dead zone.’’
Figure 5. Spatial interpolation of hypoxic conditions at the mouth of the
IHNC on August 2, 2006. Inset illustrates area of the lake sampled.
Figure 6. A map showing spatial interpolation of the lakewide distribu-
tion of Rangia cuneata biomass (g/m2) in April 2004.
92 Poirrier, Spalding and Franze
Journal of Coastal Research, Special Issue No. 54, 2009
after restoration. Total lake volume clearance time decreased
for all suspended particles, indicating that restoring clams in
the 250 km2 dead zone would reduce phytoplankton blooms
and fecal coliforms, and increase water clarity.
Submersed Aquatic Vegetation
Vallisneria americana has persistently decreased in abun-
dance since SAV was first surveyed in 1953 (Burns, Poirrier,
and Preston, 1993). Stress from changes in the salinity regime
due to saltwater intrusion from the MRGO may be contribut-
ing to stress on freshwater species such as V. americana, N.
guadalupensis, and P. perfoliatus. Freshwater species have
become rare along the southeastern shoreline near the IHNC,
a source of saltwater intrusions (Cho and Poirrier, 2005b).
1997 Bonnet Carre Spillway Opening
Benthos
Brammer et al. (2007) found that the opening of the Bonnet
Carre Spillway from March 17 through April 18, 1997, caused
an abrupt decrease in salinity (Figure 7), species diversity
(Figure 8), and R. cuneata abundance (Figure 9). Infaunal
macroinvertebrate, surface and bottom salinity, and dissolved
oxygen data obtained from November 1996 through Novem-
ber 1998 from five sites on the east–west transect were
analyzed in this study. A community composed of oligohaline
taxa persisted during the freshwater period, but changes in
dominance, and to a lesser extent, composition, occurred over
time and among sites. Prior to the Spillway opening,
hydrobiid snails were most abundant and after the opening,
oligochaetes became most abundant. There was a pronounced
spatial effect related to the distance of the sites from the
Spillway and from tidal passes (Brammer et al., 2007).
Mean diversity values significantly decreased during the
Spillway opening and remained lower than preopening values
for over 1 month after the Spillway was closed. A significant
change from these low values did not occur until salinity
returned to preopening levels in November 1998. Diversity
values ranged from 0.83 at Site 4 on October 29, 1997, to 2.24
at Site 5 on November 3, 1998. Macrofaunal abundance did
not significantly decrease during the Spillway opening.
However, it significantly declined by May 1997, over 1 month
after the Spillway was closed. Abundance did not significantly
increase from these low May 1997 values until July 1998.
Because of its numerical dominance and size, R. cuneata
comprises most of the benthic macroinvertebrate biomass.
Rangia cuneata biomass significantly declined between April
1997 and January 1998 (Brammer et al., 2007).
Submersed Aquatic Vegetation
Poirrier et al. (1999) studied the effects of the March 1997
Bonnet Carre Spillway opening. A M. spicatum bed near the
mouth of Bayou St. John north of the flood control structure
was monitored in addition to the established survey
locations. They found a significant decrease in photosynthet-
ically active radiation (PAR) from severe cyanobacterial
blooms due to the spillway opening. There was no change in
V. americana coverage, but significant decreases in R.
maritima occurred by the 1997 post–spillway opening survey
(Figure 2). A 2000 m2 M. spicatum bed in Bayou St. John
died due to overgrowth by the green filamentous alga,
Cladophora spp. Decomposition of accumulated cyanobacte-
ria, algae, and SAV resulted in anoxic conditions (Poirrier et
al., 1999) and an associated fish kill. They also found a
decrease in combined foliar coverage for V. americana and R.
maritima (total SAV) after the spillway opening. Abundant
growth of the alga Cladophora spp. occurred on R. maritima
and M. spicatum, but not V. americana. Cladophora spp.
growth on R. maritima made it susceptible to shading from
phytoplankton and uprooting by wave energy (Poirrier et al.,
1999). This resulted in R. maritima being selectively lost
Figure 7. Salinity values from November 1996 to November 2005 across
the east–west transect starting with EW1 in the westernmost portion of
the lake to EW5 in the eastern portion of the lake. In addition, hurricanes
and other disturbances are noted.
Figure 8. Shannon-Wiener species diversity index (loge) from November
1996 to November 2005 along the east–west transect starting with EW1 in
the westernmost portion of the lake to EW5 in the eastern portion of the
lake. Bottom salinity and disturbances are noted.
Benthic Invertebrates and SAV in Lake Pontchartrain 93
Journal of Coastal Research, Special Issue No. 54, 2009

from grass beds. In September 1998, Hurricane Georges
struck east of Lake Pontchartrain, and salinity in the lake
increased from the tidal surge (Figure 7). Comparisons of
pre- and poststorm surveys showed no statistical differences
in SAV abundance (Cho and Poirrier, 2001). High salinity
persisted after the passage of Hurricane Georges due to lack
of rainfall from a drought (Cho and Poirrier, 2001; Zheng et
al., 2003).
1999–2001 Drought resulting from El Nino
Benthos (Cho, 2003)
A strong ENSO event occurred between 1997 and 2001
(Zheng et al., 2003) producing a drought from 1999–2001 in
southern Louisiana. Bottom salinity varied significantly over
time (Figure 7; F(15,63) 5 33.20; p , 0.001). The drought
resulted in a significant lakewide salinity increase across the
east–west transects through fall 2000. Post hoc pairwise
comparisons indicated salinity levels in October 1999, July
2000, and October 2000 were significantly different from all
other values with the exception of salinity levels after
Hurricane Katrina. During July 2000 and through April
2001, salinity values were nearly the same across the east–
west transect (Figure 7). The hooked mussel Ischadium
recurvum and other high salinity species increased as salinity
increased during the drought. Shannon-Wiener species
diversity from May 1998 to August 2002 (Figure 8) differed
significantly over time (F(9,98) 5 4.07; p , 0.001) but not site
(F(4,98) 5 1.170; p 5 0.33). Species diversity values in the
western portion of the lake increased and became similar to
those of the eastern portion. Density of R. cuneata signifi-
cantly differed over time (F(14,149) 5 21.83; p , 0.001). Post
hoc pairwise comparisons indicated clam density values in
April 2001, August 2002, and November 2003 were signifi-
cantly different from all other values with the exception April
2004 and after Hurricane Katrina.
With a decrease in salinity, a decrease in species diversity
is expected, but the low values that occurred in April 2001 in
all but the easternmost portion of the lake are indicative of
other sources of stress. This is supported by the concomitant
shift in species composition to an annelid-dominated benthic
community and a severe loss of R. cuneata during April 2001
and August 2002 indicating severe lakewide stress.
Prior to 1999, small numbers of I. recurvum were confined
to the easternmost area of the lake (density ranged from 0–20
mussels/m2). As the density of R. cuneata decreased, I.
recurvum increased lakewide, including the western portion
of the lake. This resulted in the rapid increase of not only
small mussels (,5 mm) but also the establishment and
growth of larger mussels (.35 mm) on R. cuneata shells that
formed small spherical reefs about 30 cm in diameter. We
found I. recurvum at a mean lakewide density of 2284
mussels/m2 in April 2001. Ischadium recurvum gradually
decreased as salinity in the lake decreased.
Submersed Aquatic Vegetation
Cho and Poirrier (2005b) determined the effects of an ENSO
event on SAV. Relatively low levels of R. maritima occurred
Figure 9. Average Rangia cuneata density (clams/m2) 6 SE of clams greater than or equal to 21 mm across the east–west transect from November 1996 to
November 2005 (n 5 15).
94 Poirrier, Spalding and Franze
Journal of Coastal Research, Special Issue No. 54, 2009

from 1996 through 1998, but a significant increase occurred in
1999 that persisted through 2002 (Figure 2). They discovered
causal links between the El Nino to La Nina climate shift and
SAV change. Reduced rainfall increased salinity (Figure 7) and
water clarity, which produced a rapid increase in the
euryhaline species R. maritima in deeper water and historic
locations where SAV had not been found since 1953. Increased
salinity caused a decrease in the freshwater species V.
americana and loss of N. guadalupensis and P. perfoliatus.
There was a shift in species dominance from V. americana to R.
maritima from 1997 through 2002. Although V. americana and
N. guadalupensis increased significantly between 2001 and
2002, R. maritima continued to dominate in 2002 because of its
dramatic increase at Goose Point (eightfold increase) and
Lacombe (fivefold increase). The increase and abundance of
SAV during these years rivaled the historic distribution of SAV
(Suttkus, Darnell, and Darnell, 1954).
Data obtained in the study were used to develop a model to
predict potential SAV habitat with changes in water clarity
(Cho and Poirrier, 2005a). Three drivers of potential SAV
habitat were (1) water clarity controls SAV colonization depth;
(2) fluctuation in annual mean water level and wave mixing
determines SAV minimum colonization depth; and (3) differ-
ences in shoreface slope determine SAV areal coverage under
comparable water quality conditions. They compared empirical
data to values predicted by the model to validate the model.
Hurricane Katrina
Benthos
A major disturbance of benthic community structure was
caused by Hurricane Katrina, which made landfall as a
category 3 hurricane near eastern Lake Pontchartrain on
August 29, 2005, and produced a storm surge of 3.7 to 4.9 m
along the northeastern shore, which decreased to 1.5 to 3 m
along the western shore (Poirrier, Rodriguez del Rey, and
Spalding, 2008). It caused an increase in salinity (Figure 7),
decrease in Shannon-Wiener species diversity at most sites
(Figures 4 and 8), and a decrease in density of R. cuneata
(Figure 8). Salinity stratification and low dissolved oxygen
were present at three north shore sites on September 30, 2005
(Poirrier, personal observation). Large (.21 mm) R. cuneata
that recently died were a major component of newly formed
shoals and ridges along the shoreline.
Benthic invertebrate samples obtained in November 2004
prior to Hurricane Katrina were compared with posthurri-
cane November 2005 samples to assess damage (Poirrier,
Rodriguez del Rey, and Spalding, 2008). There was a major
shift in the rank order of dominant species. After Hurricane
Katrina, the usual dominant species including the hydrobiid
snails Probythinella protera and Texadina sphinctoma, the
isopod Cerapus benthophilis, and the polychaete Amphicteis
floridus were reduced to less than 1% of the total number of
individuals present. Oligocheates and the polychaete Stre-
blospio benedicti became dominants. Rangia cuneata de-
creased from the most abundant species prior to Hurricane
Katrina to the fifth most abundant species after the
hurricane. A comparison of the number of species, the
number of individuals, species diversity, and R. cuneata
biomass between 2004 and 2005 samples using ANOVA found
significant site and time interactions. This indicates that
Hurricane Katrina caused changes in all major measures of
community structure. The widespread defaunation caused by
Hurricane Katrina is best demonstrated by the loss of R.
cuneata and other community dominants from 50% (800 km2)
of the lake bottom at depths greater than approximately the
3.7-m depth contour. Bottom disturbance from past shell
dredging and enhanced saltwater intrusion from the MRGO
may have contributed to the severity of this impact.
Submersed Aquatic Vegetation
Hurricane Katrina produced a storm tidal surge in eastern
Lake Pontchartrain that increased salinity and damaged
littoral habitats. Surveys conducted in October 2005 found
major decreases in SAV aboveground biomass at all study
locations (Figure 2). SAV beds were reduced to small patches
consisting mainly of V. americana. SAV was eliminated at all
locations but Lacombe. Spring 2006 surveys indicated recovery
of R. maritima, but not V. americana. After Hurricane Katrina,
relatively high salinity due to decreased rainfall adversely
affected V. americana, a freshwater species, but did not affect
R. maritima, a euryhaline species that grows rapidly from
seeds (Cho and Poirrier, 2005c). Submersed aquatics in streams
north of Lake Pontchartrain and marshes in the path of the
storm were also severely affected by Hurricane Katrina (M.A.
Poirrier, personal observation).
DISCUSSION
The analyses of benthic invertebrate distribution and
abundance from 1996 through 2005 indicated large fluctua-
tions in benthic invertebrates (Figures 4 and 8) and SAV
(Figure 2) populations. During the 10-year study there was
no prolonged period that could be regarded as ‘‘normal’’
conditions. Instead, these communities were always experi-
encing or recovering from a significant temporal disturbance.
In addition, persistent spatial perturbations and other
chronic disturbances with greater time, but lesser impact
scales were present. These persistent long-term perturba-
tions included natural deterioration of the Bernard Delta;
disruption of wetland hydrology including Mississippi River
inflow, eutrophication, background toxic pollution from
agriculture and urban runoff; and changes in the general
habitat quality of the watershed, lower basin, and the Gulf of
Mexico (Penland et al., 2001). These multiple stressors might
have produced more fragile communities with reduced
diversity, resistance, and resiliency.
Benthic invertebrate species diversity values from 1996
through 2005 fluctuated as a result of changes in salinity,
hypoxia, and damage from a severe hurricane (Figures 3 and
4). In coastal marine systems, diversity increases with
salinity, and low salinity estuaries, such as Lake Pontchar-
train, are characterized by relatively low diversity (Moore,
1992; Remane and Schleiper, 1971). Because salinity gener-
ally increases from west to east in Lake Pontchartrain, there
is a corresponding increase in species diversity. Exceptions to
Benthic Invertebrates and SAV in Lake Pontchartrain 95
Journal of Coastal Research, Special Issue No. 54, 2009

this pattern occurred during the 1997 Bonnet Carre Spillway
opening and during July 2000 when salinities throughout the
lake were similar (Figures 7 and 8). In 1997, salinity across
the lake was the same due to the massive influx of Mississippi
River water and in 2000 due to the lack of stream flow from a
severe drought. Species diversity also generally increased
along our north–south transect with distance from the mouth
of the INHC even though salinity decreased (Figure 4). This
was due to stress from anoxia and hypoxia that had a greater
effect on the community structure than salinity. This pattern
rapidly changed when stratification was disrupted and
recruitment of higher salinity opportunistic species occurred.
The presence, absence, and relative abundance of indicator
taxa showed strong correlations with shifts in salinity and
dissolved oxygen. Most species are small and opportunistic,
forming different assemblages that rapidly recover from
disturbances. Large R. cuneata require several years to
recover from episodes of harmful dissolved oxygen. Therefore,
clam distribution can be used in annual surveys to assess
areas affected by anoxia and hypoxia.
Recovery from Shell Dredging
Benthos
Our recovery studies conducted after shell dredging ended
in 1990 indicate that shell dredging decreased large clam
density and water clarity (Abadie and Poirrier, 2000, 2001b).
After shell dredging ended, a significant increase in the
density of large R. cuneata indicated recovery to levels
equaling the 1954 baseline data. A study of water clarity
based on Secchi disc visibility observations from three sites on
the Lake Pontchartrain Causeway found that significant
visibility increases occurred at the north and midlake sites,
but no change occurred at the south shore site (Francis and
Poirrier, 1999). This increase in visibility appears to be
related to the increase in R. cuneata biomass after shell
dredging ceased. This is supported by the low density of clams
near the south shore due to saltwater intrusion, where water
clarity did not increase; by the increase in water clarity that
peaked 3 years after shell dredging was stopped, the length of
time that it takes for clams to mature; and by a recent study
(Spalding, Walker, and Poirrier, 2007) indicating that high
clam biomass has a significant effect on water clarity.
Although recovery of large R. cuneata from shell dredging
occurred, episodes of anoxia and hypoxia associated with
saltwater intrusion from the MRGO also causes loss of large
clams (Figure 6). Because high salinity bottom water flows
along depth contours, shallow channels cut by shell dredges
may have facilitated more widespread movement of water low
in dissolved oxygen in the lake.
Submersed Aquatic Vegetation
There was no immediate increase in SAV abundance on the
north shore associated with the water clarity increase in 1993
after shell dredging was stopped (Francis and Poirrier, 1999).
A significant SAV increase did occur in 1998 (Cho and
Poirrier, 2005b). This increase may have been delayed due to
damage from Hurricane Andrew in 1992 (Burns and Poirrier,
1996). Water clarity dynamics in the littoral zone probably
differed from the pelagic zone where water clarity was
studied. Littoral habitats may have been degraded from
watershed increases in nutrients and turbidity. Even if
turbidity from suspended silts and clays decreased, algal
epiphyte growth would increase and shade SAV without
nutrient reduction. These effects persisted after shell dredg-
ing ended, but decreased during the drought. Low runoff
during the drought resulted in relatively clear, low nutrient
conditions and a rapid expansion of SAV as evidenced by the
increase of R. maritima to near historic levels during the
spring of 1999 (Cho and Poirrier, 2005b).
Effects of Anoxia and Hypoxia from MRGOSaltwater Intrusion
Benthos
A major and continuing impact to Lake Pontchartrain is
saltwater intrusion via the IHNC from the MRGO. Unnatu-
rally high salinity bottom water has entered Lake Pontchar-
train through the MRGO since it was completed in 1963. This
saline water produces stratification and periodic bottom
anoxia and hypoxia (Junot, Poirrier, and Soniat, 1983;
Poirrier, 1978). Following the recovery of large R. cuneata
after shell dredging ceased, clams greater than 21 mm
remained absent from a 250 km2 area as a result of anoxia
and hypoxia facilitated by salinity stratification (Abadie and
Poirrier, 2001a). This area has become known as the ‘‘dead
zone’’ (Figure 6). Episodes of high salinity and low dissolved
oxygen may extend beyond this zone and affect benthic
invertebrate populations lakewide. Earlier studies (Sikora
and Sikora, 1982) attributed the stressed benthic community
to toxic substances from urban runoff and chemical spills.
Later studies demonstrated that outfall canals have a
localized effect (Poirrier et al., 1984; Powers, Poirrier, and
Yund, 1992), and the principal cause of stress is low bottom
dissolved oxygen due to saltwater intrusion.
Our study of R. cuneata restoration through closure of the
MRGO found that clam restoration would have widespread
positive contributions to the holistic rehabilitation of Lake
Pontchartrain (Spalding, Walker, and Poirrier, 2007). In
addition to direct benefits of reduction of phytoplankton
blooms, turbidity, and fecal coliforms by increased clam
filtration, increased water clarity would increase SAV. Clam
biomass restoration would add shells that would stabilize the
mud bottom and reduce shoreline erosion. Clam restoration
will also provide more food for fish, crabs, and waterfowl
(Darnell, 1958; Ebersole and Kennedy, 1995; Richman and
Lovvorn, 2004) and increase nutrient cycling through clam
excretion, biodeposition of feces and pseudofeces, and biotur-
bation of sediment (Dame, 1996; Newell, 2004; Vaughn and
Hakenkamp, 2001). Overall, R. cuneata restoration would
lessen eutrophication, improve recreational water quality,
and greatly expand essential fish habitat. Work is in progress
to close the MRGO. Natural recruitment of R. cuneata will
occur in areas that are currently adversely affected. The
annual increase in clam abundance and size classes will
provide restoration milestones and the presence of mature,
96 Poirrier, Spalding and Franze
Journal of Coastal Research, Special Issue No. 54, 2009

large clams will provide a measureable endpoint in the
restoration process.
Submersed Aquatic Vegetation
Elevated salinities from saltwater intrusion have affected
SAV community structure (Figure 2) by favoring the growth
of the opportunistic, euryhaline species, R. maritima, and
contributing to stress on freshwater species, including the
rare species P. perfoliatus (Cho and Poirrier, 2005b). A major
benefit of closing the MGRO will be SAV restoration through
improved water clarity on the southeastern shore due to clam
restoration. Monitoring should be continued to document the
recovery of SAV and clams after the MRGO closure.
1997 Bonnet Carre Spillway Opening
Benthos and SAV
The 1997 spillway opening appeared to affect both benthos
and SAV. Based on decreases in species diversity, abundance,
and the number of taxa, we concluded that the 1997 spillway
opening had a deleterious impact on the benthos (Brammer et
al., 2007). The cause of these changes cannot be attributed
with certainty to any one factor, but we speculate that they
were the result of reduced salinity, cyanobacterial blooms,
and hypoxia or anoxia. Although recovery of benthos
occurred, based on overall changes in macroinvertebrate
abundance and R. cuneata biomass, no increase in benthic
invertebrate productivity was detected. The decline of overall
SAV abundance, especially R. maritima, and the loss of M.
spicatum near the mouth of Bayou St. John after the 1997
spillway opening indicated that the opening also had a
negative effect on SAV (Poirrier et al., 1999). These results
should be considered in the operation of the Bonnet Carre
Spillway and the design of high volume diversions that
discharge directly into Lake Pontchartrain.
1999–2001 Drought Resulting from an El NinoSouthern Oscillation
Benthos
The increase in species diversity, the decrease in R. cuneata,
and the increase in I. recurvum associated with an increase in
salinity during the 1999–2001 drought demonstrated that
climate shifts can affect benthic community structure and
dominant bivalve abundance. However, it is unlikely that
increased salinity alone was responsible for the R. cuneata
decline. The decline occurred during a period of decreasing, not
increasing salinity, and at this time, species diversity was low
and samples were dominated by oligochaetes. The salinity was
well within the known salinity tolerance of R. cuneata, which
can reproduce and grow at salinities up to 15 ppt (Cain, 1973,
1974) and survive salinities as high as 25 ppt (Hopkins,
Anderson, and Horvath, 1973; LaSalle and de la Cruz, 1985;
Swingle and Bland, 1974). The underlying factor causing the
decline was the interaction of stress from episodic hypoxia and
abrupt salinity changes. Organic material and plant nutrients
may have accumulated in the watershed during the drought
and were flushed into the lake with rains, which rapidly
decreased salinity after the drought. Henry, Mangum, and
Webb (1980) reported 50% mortality of R. cuneata after 5–7 d
under deoxygenated conditions in both 2 and 20 ppt salinities.
Those that survived initially did not recover after being
returned to normoxic conditions. In addition, they found that
survival decreased to 3 d when given a hypo- or hypersaline
shock (18 ppt change).
Ischadium recurvum are commonly found in more saline
conditions than R. cuneata (Holland, Mountford, and Mi-
hursky, 1977; Read, 1964). The average low salinity levels in
Lake Pontchartrain are not conducive to recruitment,
establishment, and growth of I. recurvum. Establishment of
I. recurvum was also aided by differences in life history
strategies, which gave this species a competitive edge over R.
cuneata. Ischadium recurvum are epifaunal, attaching to
hard substratum (Holland, Mountford, and Mihursky, 1977),
while R. cuneata are infaunal, burrowing in the sediment.
During periods of hypoxia, it is likely that I. recurvum were
not as severely affected as R. cuneata by low oxygen because
of their higher position in the water column. The die-off of R.
cuneata ended competition from R. cuneata for food and other
resources. This resulted in the rapid increase of not only
small mussels (,5 mm) but also the establishment and
growth of larger mussels (.35 mm). The number of I.
recurvum rapidly decreased with decreasing salinity, and R.
cuneata recovery was slow, indicating that factors other than
salinity affected recovery.
Submersed Aquatic Vegetation
Based on SAV dynamics observed during the drought, it
appears that climate shifts may cause cyclic changes in SAV
with salinity controlling species composition and water
clarity controlling SAV distribution and abundance (Cho
and Poirrier, 2005b). SAV restoration could be accomplished
by improving water clarity. We also learned that the response
of R. maritima to improved water clarity is rapid. The
increase in SAV observed during 1 year of this study was
nearly equal to SAV loss in 40 years of decline. Comparable
natural shifts in SAV likely occurred in the past. However,
this increase in SAV probably would not have occurred if shell
dredging had continued because widespread high-turbid
waters would have continued to shade the littoral zone
(Francis and Poirrier, 1999). The model we developed can be
used to quantitatively predict potential SAV habitat changes
caused by changes in water clarity (Cho and Poirrier, 2005a).
Hurricane Katrina
Benthos
Sampling water quality immediately after Hurricane
Katrina was difficult. However, Pardue et al. (2005) obtained
samples of New Orleans storm water in early September
2005. They found that Hurricane Katrina floodwaters were
typical of storm water with the exception of elevated levels of
lead. Our studies of the effects of Hurricane Katrina
demonstrated that severe hurricanes have direct and indirect
effects on benthos (Poirrier, Rodriguez del Rey, and Spalding,
Benthic Invertebrates and SAV in Lake Pontchartrain 97
Journal of Coastal Research, Special Issue No. 54, 2009

2008). Direct effects included the storm surge and wave
energy disrupting the bottom and displacing and burying
benthic organisms. Indirect effects included the defaunation
of over 800 km2 of lake bottom extending from the deepest
water near the center of the lake to about the 3.7-m contour.
We postulate that this indirect effect was caused by
widespread, higher-salinity bottom water, which formed a
nonmixing stratified layer that became deoxygenated due to
the input of organic material. This further damaged the
benthic community in the entire lake at depths greater than
3.7 m. Some recovery by opportunistic species has occurred,
but R. cuneata recovery has been slow due to high salinity
conditions produced by the storm surge and low rainfall after
Hurricane Katrina.
Submersed Aquatic Vegetation
We found that severe hurricanes such as Hurricane Katrina
directly destroy SAV and recovery rate depends on conditions
after the storm. Ruppia maritima and N. guadalupensis are
more susceptible to damage from wave energy because they
have a bushy growth form with branches and numerous
separate leaves, while V. americana has tapelike leaves.
Ruppia maritima, however, recovered more rapidly than V.
americana because of abundant seeds in the seed bank and
better tolerance of higher post-Hurricane Katrina salinities
than V. americana. Burns and Poirrier (1996) reported rapid
recovery of north shore SAV species after damage from
Hurricane Andrew in August 1992. Hurricane Georges, which
made landfall in October 1998, did not directly damage SAV,
and R. maritima increased in abundance after Georges (Cho
and Poirrier, 2001). Chabreck and Palmisano (1973) also
reported an increase in R. maritima in the Mississippi River
delta marshes after the passage of Hurricane Camille. Poirrer
and Handley (2007) attributed major temporal changes in
seagrass populations in Chandeleur Island Sound mainly to
the direct effects of tropical storms and hurricanes.
Both benthic invertebrates and SAV can serve as endpoints
for the evaluation of habitat quality and restoration status.
However, the distribution and abundance of these oligohaline
organisms does not always follow paradigms established in
higher salinity systems or convey a specific set of ‘‘good or bad’’
values. While the open ecosystem dynamic nature of estuaries
causes frequents disruptions, it also provides mechanisms for
rapid recovery. Unraveling the effects of multiple stressors in
these complex systems is difficult and will require decades of
further study. However, the more we learn about the biology of
indicator species and communities, the better they will serve as
indicators of environmental change.
CONCLUSION
The overall lesson learned in this study is that an increase or
decrease in a particular set of benthic organisms were not
simply determined by persistent, long-term anthropogenic
changes in habitat quality, but were often driven by short-
term disturbances and natural cycles. Measures of community
structure and indicator species such as R. cuneata and V.
americana were useful, but periodic disturbances made using
them to evaluate environmental health during a particular
period difficult. The effects of past disturbances and the degree
and nature of recovery have to be considered. The oligohaline
estuarine zone is composed of opportunistic species that
respond rapidly to dynamic abiotic factors such as salinity
and dissolved oxygen, and populations can rapidly recover by
recruitment from adjacent coastal waters. Resistance and
resilience to periodic habitat disturbances, the presence of
natural cycles in distribution and abundance, and the occur-
rence of alternate community states have to be considered in
evaluating habitat quality and restoration success.
Specific lessons learned pertaining to benthic inverte-
brates:
N Shell dredging had a significant, disruptive effect on
benthic organisms and water clarity, and it should
continue to be banned.
N Saltwater intrusion from the MRGO through the IHNC
and associated episodic anoxia and hypoxia are the
principal causes of stress in the benthic community.
N The widespread stress observed in the benthic community
in earlier studies is due to saltwater intrusion and
associated anoxia and hypoxia, and not toxic substances
from urban runoff. Discernable effects of runoff on the
benthic community are limited to sites near discharges.
N Complete closure of the MRGO would reduce salinity
stratification and associated anoxia and hypoxia and result
in the return of R. cuneata to a 250 km2 (100 mi2 area).
Clam restoration would reduce eutrophication effects,
improve recreational water quality, and greatly expand
essential fish habitat.
N Large R. cuneata take several years to recover from
episodes of harmful dissolved oxygen. Therefore their
distribution and abundance can be used in annual surveys
to assess areas affected by anoxia and hypoxia.
N The 1997 Bonnet Carre Spillway opening disrupted the
benthic invertebrate community by abruptly lowering
salinity, facilitating cyanobacterial blooms, and decreasing
dissolved oxygen. Contrary to common management
paradigms, there was no increase in benthic invertebrate
productivity associated with the 1997 diversion.
N The 1999–2001 ENSO drought produced high bottom
salinity throughout the lake and episodic hypoxia was also
widespread. These hypoxic events coupled with increased
salinity resulted in lakewide changes in species diversity, a
decrease in R. cuneata clams, and an increase in I. recurvum.
N Hurricane Katrina directly killed benthic organisms and
pushed high salinity bottom water that was low in
dissolved oxygen up to the 3.7-m contour, severely
disrupting the structure of the benthic community.
Specific lessons learned pertaining to SAV:
N Light availability is the primary factor that controls SAV
distribution and abundance in the lake. Turbidity reduc-
tion is thus required for SAV restoration. Natural increases
in abundance would not have occurred if shell dredging
had not been banned and associated turbidity decreased.
N Potential SAV area gained from increased water clarity can
be quantitatively predicted using our SAV habitat model.
98 Poirrier, Spalding and Franze
Journal of Coastal Research, Special Issue No. 54, 2009

N Increased R. cuneata density as a result of the MRGO
closure would improve water clarity. Clams remove
suspended solids from the water column, thus increasing
SAV distribution and abundance.
N Closure of the MRGO will reduce episodes of high salinity,
increase the abundance of V. americana and other
freshwater SAV in the southeastern lake, and may help
restore P. perfoliatus.
N Nutrients increase phytoplankton and epiphyte growth,
which limit light availability resulting in a decrease in
SAV. This should be considered in the design of freshwater
diversions.
N Salinity controls SAV species composition by limiting the
distribution of freshwater species. Potamogeton perfoliatus
was extirpated during the drought from Lake Pontchar-
train, the only known habitat in Louisiana.
N Maintaining and increasing SAV ‘‘friendly shorelines’’ and
source populations through V. americana culture and
transplanting will enhance SAV restoration.
N Seeds are important in the life cycle of the pioneer species
R. maritima. Seeds should be incorporated in restoration
strategies.
N Hurricanes can directly damage SAV, but recovery de-
pends upon species adaptations and posthurricane effects.
Ruppia maritima is known to increase after hurricanes.
ACKNOWLEDGMENTS
The preparation of this paper and associated analyses were
funded by grants from the National Oceanic and Atmospheric
Administration through the Pontchartrain Restoration Pro-
gram. Freeport-McMoran, Inc., funded benthic invertebrate
sample collections and preliminary data analyses prior to
2001. The Louisiana Department of Wildlife and Fisheries
and the Lake Pontchartrain Basin Foundation provided funds
for SAV monitoring and restoration studies prior to 2001. We
would like to thank the following individuals who assisted in
these studies: Chip Crews, Jesse Hobson, Wendy Hobson,
Ryan Poirrier, Zoe Rodriguez del Rey, and Ashley Walker.
LITERATURE CITED
Abadie, S.W. and Poirrier, M.A., 2000. Increased density of largerangia clams in Lake Pontchartrain after the cessation of shelldredging. Journal of Shellfish Research, 19(1), 481–485.
Abadie, S.W. and Poirrier, M.A., 2001a. Rangia clams as an indicatorof hypoxia in Lake Pontchartrain. In: Penland, S., Beall, A., andWaters, J. (eds.), Environmental Atlas of the Lake PontchartrainBasin. New Orleans, Louisiana: Lake Pontchartrain Basin Foun-dation, 166p.
Abadie, S.W. and Poirrier, M.A., 2001b. Recent trends in water clarityand clam abundance. In: Penland, S., Beall, A., and Waters, J.(eds.), Environmental Atlas of the Lake Pontchartrain Basin. NewOrleans, Louisiana: Lake Pontchartrain Basin Foundation, 165p.
Bilyard, G.R., 1987. The value of benthic infauna in marine pollutionmonitoring studies. Marine Pollution Bulletin, 18(11), 581–585.
Boesch, D.F. and Rosenberg, R., 1981. Response to stress in marinebenthic communities. In: Barrett, G.W. and Rosenberg, R. (eds.),Stress Effects on Natural Ecosystems. New York: Wiley-Intersci-ence, pp. 179–200.
Brammer, A.; Rodriguez del Rey, Z.; Spalding, E.A., and Poirrier,M.A., 2007. Effects of the 1997 Bonnet Carre Spillway opening on
infaunal macroinvertebrates in Lake Pontchartrain, Louisiana.Journal of Coastal Research, 23(5), 1292–1303.
Burns, J.W. and Poirrier, M.A., 1996. The Restoration of SubmersedAquatic Vegetation in the Lake Pontchartrain Estuary, Louisiana.Metairie, Louisiana: Lake Pontchartrain Basin Foundation, 94p.
Burns, J.W.; Poirrier, M.A., and Preston, K.P., 1993. Effects of urbanrunoff on the environmental quality of Lake Pontchartrain,Louisiana. Effects of New Orleans urban runoff on the distributionand structure of submerged aquatic vegetation communities inLake Pontchartrain, Louisiana. Interim Report. New Orleans:Urban Waste Management and Research Center, University ofNew Orleans, Report No. 92-05, 137p.
Burns, J.W.; Poirrier, M.A., and Preston, K.P., 1995. The status ofPotamogeton perfoliatus (Potamogetonaceae) in Lake Pontchar-train, Louisiana. Sida, 16(4), 757–763.
Cain, T.D., 1973. The combined effects of temperature and salinity onembryos and larvae of the clam Rangia cuneata. Marine Biology, 21, 1–6.
Cain, T.D., 1974. Combined effects of changes in temperature andsalinity on early stages of Rangia cuneata. The Virginia Journal ofScience, 25, 30–31.
Cain, T.D., 1975. Reproduction and recruitment of brackish waterclam Rangia cuneata in James River, Virginia. Fishery Bulletin,73(2), 412–430.
Chabreck, R.H. and Palmisano, A.W., 1973. The effects of HurricaneCamille on the marshes of the Mississippi River delta. Ecology,54(5), 1118–1123.
Cho, H.J., 2003. Ecology of Submersed Aquatic Vegetation in LakePontchartrain, Louisiana. New Orleans, Louisiana: University ofNew Orleans, Doctoral thesis, 153p.
Cho, H.J. and Poirrier, M.A., 2001. Spillway, hurricane, and droughteffects on SAV (grassbeds). In: Penland, S.; Beall, A., and Waters, J.(eds.), Environmental Atlas of the Lake Pontchartrain Basin. NewOrleans, Louisiana: Lake Pontchartrain Basin Foundation, 126p.
Cho, H.J. and Poirrier, M.A., 2005a. A model to estimate potentialSAV (submersed aquatic vegetation) habitat based on studies inLake Pontchartrain. Restoration Ecology, 13(4), 623–629.
Cho, H.J. and Poirrier, M.A., 2005b. Response of SAV to the 1997–2001 El Nino Southern Oscillation shifts in Lake Pontchartrain,Louisiana. Estuaries, 28(2), 215–225.
Cho, H.J. and Poirrier, M.A., 2005c. Seasonal growth and reproduc-tion of Ruppia maritima L. s.l. in Lake Pontchartrain, Louisiana,USA. Aquatic Botany, 81, 37–49.
Clark, K.R. and Gorley, R.N., 2001. PRIMER (Plymouth Routines inMultivariate Ecological Research). Issue, Plymouth, UK: Primer-E Ltd.
Cottam, C., 1939. Food habits of North American diving ducks. USDATechnical Bulletin, 643.
Dame, R.F., 1996. Ecology of Marine Bivalves: An EcosystemApproach. Boca Raton, Florida: CRC Press, 254p.
Darnell, R.M., 1958. Food habits of fishes and larger invertebrates ofLake Pontchartrain, Louisiana, an estuarine community. Instituteof Marine Science, 5, 353–416.
Dennison, W.C.; Orth, R.J.; Moore, K.A.; Stevenson, J.C.; Carter, V.;Kollar, S.; Bergstrom, P.W., and Batiuk, R., 1993. Assessing waterquality with submersed aquatic vegetation. Bioscience, 43, 86–91.
Diaz, R.J. and Rosenberg, R., 1995. Marine benthic hypoxia: a reviewof its ecological effects and the behavioral responses of benthicmacrofauna. Oceanography and Marine Biology, 33, 245–303.
Ebersole, E.L. and Kennedy, V.S., 1995. Prey preferences of bluecrabs Callinectes sapidus on three bivalve species. Marine EcologyProgress Series, 118(1–3), 167–177.
Francis, J.C. and Poirrier, M.A., 1999. Recent trends in water clarityof Lake Pontchartrain. Gulf Coast Research Reports, 11, 1–5.
Francis, J.C.; Poirrier, M.A.; Barbe, D.E.; Wijesundera, V., andMulino, M.M., 1994. Historic trends in the Secchi disk transparencyof Lake Pontchartrain. Gulf Research Reports, 9(1), 1–16.
Gertz, S.M., 1984. Biostatistical aspects of macrophyton sampling. In:Dennis, W.M. and Isom, B.G. (eds.), Ecological Assessment ofMacrophyton: Collection, Use, and Meaning of Data, ASTM STP843. American Society for Testing and Materials, pp. 28–35.
Gunter, G., 1960. Some relations of estuarine organisms to salinity.Limnology and Oceanography, 6(2), 182–190.
Henry, R.P.; Mangum, C.P., and Webb, K.L., 1980. Salt and water
Benthic Invertebrates and SAV in Lake Pontchartrain 99
Journal of Coastal Research, Special Issue No. 54, 2009

balance in the oligohaline clam, Rangia cuneata 2. Accumulation ofintracellular free amino-acids during high salinity adaptation.Journal of Experimental Zoology, 211(1), 11–24.
Holland, A.F.; Mountford, N.K., and Mihursky, J.A., 1977. Temporalvariation in upper bay mesohaline benthic communities: I. The 9-mmud habitat. Chesapeake Science, 18(4), 370–378.
Hopkins, S.H.; Anderson, J.W., and Horvath, K., 1973. The brackishwater clam Rangia cuneata as indicator of ecological effects ofsalinity changes in coastal waters. Contract Report H-73-1, 250p.
Hopkins, S.H. and Andrews, J.D., 1970. Rangia cuneata on the EastCoast: thousand mile range extension, or resurgence? Science,167(3919), 868–869.
Junot, J.A.; Poirrier, M.A., and Soniat, T.M., 1983. Effects of salt-water intrusion from the Inner Harbor Navigation Canal on thebenthos of Lake Pontchartrain, Louisiana. Gulf Research Reports,7(3), 247–254.
LaSalle, M.W. and de la Cruz, A.A., 1985. Species profiles: lifehistories and environmental requirements of coastal fisheries andinvertebrates (Gulf of Mexico)—common rangia. In: Species Profile:Life Histories and Environmental Requirements of Coastal Fisher-ies and Invertebrates (Gulf of Mexico). U.S. Fish and WildlifeService Biological Report 82(11.31). U.S. Army Corps of Engineers,TR EL-82-4, 16p.
Mayer, M.S., 1986. The Submerged Aquatic Vegetation of the LakePontchartrain Estuarine System. New Orleans, Louisiana: Univer-sity of New Orleans, Master’s Thesis, 119p.
Montz, G.N., 1978. The submerged vegetation of Lake Pontchartrain,Louisiana. Castanea, 43(2), 115–128.
Moore, C.G., 1992. Low-salinity back bays and lagoons. In Hackney,C.T., Adams, S.M., and Martin, W.H. (eds.), Biodiversity of theSoutheastern United States. New York: John Wiley, pp. 541–613.
Neter, J.; Wasserman, W., and Kutner, M.H., 1990. Applied LinearStatistical Models. Boston, Massachusetts: Irwin, 1184p.
Newell, R.I.E., 2004. Ecosystem influences of natural and cultivatedpopulations of suspension-feeding bivalve molluscs: a review.Journal of Shellfish Research, 23(1), 51–61.
Odum, H.T., 1967. Biological circuits and the marine systems ofTexas. In: Olson, T.A. and Burgess F.J. (eds.), Pollution and MarineEcology. New York: John Wiley, pp. 99–157.
Orth, R.J.; Heck, J.K.L., and van Montfrans, J., 1984. Faunalcommunities in seagrass beds: a review of the influence of plantstructure and prey characteristics on predator-prey relationships.Estuaries, 7(4A), 339–384.
Pardue, J.H.; Moe, W.M.; McInis, D.; Thibodeaux, L.J.; Valsaraj, K.T.;Maciasz, E.; Van Heerden, I.; Korevec, N., and Yuan, Q.Z., 2005.Chemical and microbiological parameters in New Orleans floodwa-ter following hurricane Katrina. Environmental Science andTechnology, 39(22), 8591–8599.
Penland, S.; McCarty, P.; Beall, A., and Maygarden, D., 2001.Regional description of the Lake Pontchartrain Basin. In: Penland,S., Beall, A., and Waters, J. (eds.), Environmental Atlas of the LakePontchartrain Basin. New Orleans, Louisiana: Lake PontchartrainBasin Foundation, pp. 3–6.
Perry, M.C. and Uhler, F.M., 1981. Asiatic clam (Corbicula mani-lensis) and other foods used by waterfowl in the James River,Virginia. Estuaries, 4(3), 229–233.
Perry, M.C. and Uhler, F.M., 1988. Food habits and distribution ofwintering canvasbacks, Aythya valisineria, on Chesapeake Bay.Estuaries, 11(1), 57–67.
Peterson, B.J. and Heck, K.L.J., 1999. The potential for suspensionfeeding bivalves to increase seagrass productivity. Journal ofExperimental Marine Biology and Ecology, 240, 37–52.
Poirrier, M.A., 1978. Studies of salinity stratification in southernLake Pontchartrain near the Inner Harbor Navigational Canal. TheProceedings of the Louisiana Academy of Sciences, XLI, 26–35.
Poirrier, M.A. and Handley, L.R., 2007. Statewide summary of Louisiana.Seagrass Status and Trends in the Northern Gulf of Mexico: 1940–2002. Gulf of Mexico Program, EPA, USGS Scientific InvestigationsReport 2006-5287 and USEPA 855-R-04-003, pp. 60–71.
Poirrier, M.A.; Maglic, B.; Francis, J.C.; Franze, C.D., and Cho, H.J.,1999. Effects of the 1997 Bonnet Carre Spillway opening on LakePontchartrain submersed aquatic vegetation. In: Recent Research
in Coastal Louisiana: Natural System Function and Response toHuman Influence, pp. 123–133.
Poirrier, M.A.; Rodriguez del Rey, Z., and Spalding, E.A., 2008. Acutedisturbance of Lake Pontchartrain benthic communities by Hurri-cane Katrina. Estuaries and Coasts, 31(6), 1221–1228.
Poirrier, M.A.; Soniat, T.M.; King, Y., and Smith, L., 1984. Anevaluation of the southern Lake Pontchartrain benthos community.New Orleans, Louisiana: Louisiana Department of EnvironmentalQuality Office of Water Resources Water Pollution ControlDivision, 64003-84-05, pp. 1–79.
Powers, S.P.; Poirrier, M.A., and Yund, P.O., 1992. Effects of urbanrunoff on infaunal invertebrates in southern Lake Pontchartrain.Water Environment Federation 65th Annual Conference andExposition, pp. 15–18.
Read, K.R.H., 1964. Ecology and environmental physiology of somePuerto Rican bivalve molluscs and a comparison with boreal forms.Caribbean Journal of Science, 4(4), 459–465.
Reish, D.J., 1986. Benthic invertebrates as indicators of marinepollution: 35 years of study. Oceans, pp. 885–888.
Remane, A. and Schleiper, D., 1971. Biology of Brackish Water. NewYork: John Wiley, 372p.
Richman, S.E. and Lovvorn, J.R., 2004. Relative foraging value tolesser scaup ducks on native and exotic clams from San FranciscoBay. Ecological Applications, 14(4), 1217–1231.
Schurtz, M.H. and St. Pe, K.M., 1984. Water quality investigation ofenvironmental conditions in Lake Pontchartrain. In: Report onInterim Findings to Louisiana Department of Environmental Quality.Baton Rouge, Louisiana: LDEQ, Water Pollution Division, 85p.
Sikora, W.B. and Kjerfve, B., 1985. Factors influencing the salinityregime of Lake Pontchartrain, Louisiana, a shallow coastal lagoon:analysis of a long-term data set. Estuaries, 8(2A), 170–180.
Sikora, W.B. and Sikora, J.P., 1981. Environmental effects ofhydraulic dredging for clam shells in Lake Pontchartrain, Louisi-ana. Baton Rouge, Louisiana: Coastal Ecology Laboratory, Centerfor Wetland Resources, LSU-CEL-81-18, 140p.
Sikora, W.B. and Sikora, J.P., 1982. Ecological Characterization of theBenthic Community of Lake Pontchartrain, Louisiana. NewOrleans District: U.S. Army Corps of Engineers, 214p.
Spalding, E.A.; Walker, A.E., and Poirrier, M.A., 2007. Restoration of 100square miles of shellfish habitat in Lake Pontchartrain. New Orleans,Louisiana: Gulf of Mexico Program, EPA, MX974852-03-0, 30p.
Suttkus, R.D.; Darnell, R.M., and Darnell, J.H., 1954. BiologicalStudy of Lake Pontchartrain. New Orleans, Louisiana: TulaneUniversity, 59p.
Swenson, E.M., 1980. General hydrography of Lake Pontchartrain,Louisiana. In: Environmental Analysis of Lake Pontchartrain,Louisiana, Its Surrounding Wetlands, and Selected Land Uses.New Orleans, Louisiana: U. S. Army Engineer District, DACW29-77-C-0253, pp. 57–156.
Swingle, H.A. and Bland, D.G., 1974. Distribution of the estuarineclam Rangia cuneata in coastal waters of Alabama. AlabamaMarine Resource Bulletin, 10, 9–16.
Tarver, J.W. and Dugas, R.J., 1973. A study of the clam, Rangiacuneata, in Lake Pontchartrain and Lake Maurepas, Louisiana.Louisiana Wildlife and Fisheries Commission, Technical BulletinNo. 5, 97p.
Turner, R.E.; Darnell, J.H., and Bond, J., 1980. Changes in thesubmerged macrophytes of Lake Pontchartrain (Louisiana): 1954–1973. Northeast Gulf Science, 4, 44–49.
USACE, 1987. Clam shell dredging in Lakes Pontchartrain andMaurepas, Louisiana. In: Draft Environmental Impact Statementand Appendixes, United States Army Corps of Engineers, NewOrleans District. New Orleans, Louisiana: U.S. Army Corps ofEngineers, New Orleans District, 151p.
Vaughn, C.C. and Hakenkamp, C.C., 2001. The functional role of burrowingbivalves in freshwater ecosystems. Freshwater Biology, 46, 1431–1446.
Wilson, J.G., 1994. The role of bioindicators in estuarine manage-ment. Estuaries, 17(1A), 94–101.
Zheng, D.; Ding, X.; Zhou, Y., and Chen, Y., 2003. Earth rotation andENSO events: combined excitation of interannual LOD variationsby multiscale atmospheric oscillations. Global and PlanetaryChange, 36, 89–97.
100 Poirrier, Spalding and Franze
Journal of Coastal Research, Special Issue No. 54, 2009