leaf traits associated with drought adaptation in faba
TRANSCRIPT

Department of Agricultural Sciences
Faculty of Agriculture and Forestry
University of Helsinki
Doctoral Program in Plant Science
Leaf traits associated with drought adaptation in
faba bean (Vicia faba L.)
Doctoral thesis
Hamid Khazaei
ACADEMIC DISSERTATION
To be presented, with the permission of the Faculty of Agriculture and Forestry, University of
Helsinki, for public examination in lecture room B4 at Viikki campus, Latokartanonkaari 7–9,
Helsinki, on 26 September 2014, at 12 o’clock noon.
Helsinki 2014

2
Supervisors:
Adjunct Professor Frederick L. Stoddard Department of Agricultural Sciences
University of Helsinki
Finland
Professor Mikko J. Sillanpää Department of Mathematical Sciences
Department of Biology
Biocenter Oulu
University of Oulu
Finland
Reviewers: Assistant Professor Petr Smýkal Department of Botany
Palacký University in Olomouc
Czech Republic
Adjunct Professor Simo Hovinen
Opponent:
Professor Wolfgang Link Department of Crop Sciences
Georg-August-University
Germany
Cover figure: Faba bean germplasm grown in the greenhouse. © Hamid Khazaei
ISBN 978-952-10-8905-3 (Print)
ISBN 978-952-10-8906-0 (Online)
ISSN 1798-7407 (Print)
ISSN 1798-744X (Online)
ISSN-L 1798-7407
Department of Agricultural Sciences | Publications | 38
Electronic publication at http://ethesis.helsinki.fi
Unigrafia
Helsinki 2014

3
CONTENTS
List of original publications ........................................................................................................... 5
Abbreviations ................................................................................................................................. 6
ABSTRACT .................................................................................................................................. 7
1 INTRODUCTION ...................................................................................................................... 8
1.1 Faba bean (Vicia faba L.) .................................................................................................... 8
1.2 Drought adaptation .............................................................................................................. 9
1.2.1 Response mechanisms ................................................................................................ 10
1.3 Breeding............................................................................................................................. 13
1.3.1 Conventional breeding ................................................................................................ 13
1.3.2 Genetic resources ........................................................................................................ 14
1.4 Genomics ........................................................................................................................... 16
1.4.1 Marker assisted selection (MAS) ................................................................................ 19
1.4.2 Markers based on DNA .............................................................................................. 19
1.5 Synteny .............................................................................................................................. 20
1.5.1 Medicago truncatula ................................................................................................... 21
1.6 Quantitative trait loci ......................................................................................................... 21
2 AIMS OF THIS STUDY .......................................................................................................... 24
3 MATERIALS AND METHODS ............................................................................................. 25
3.1 Germplasm survey ............................................................................................................. 26
3.1.1 Construction of the FIGS sets ..................................................................................... 26
3.1.2 Germplasm screening ................................................................................................. 26
3.2 Molecular breeding ............................................................................................................ 26
3.2.1 Mapping population development and selection ........................................................ 26
3.2.2 Genotyping ................................................................................................................. 26
3.3 Measurements .................................................................................................................... 27
3.4 Growth conditions and statistical analysis ......................................................................... 28
4 RESULTS AND DISCUSSION ............................................................................................... 31
4.1 Germplasm screening ........................................................................................................ 31
4.2 Molecular breeding ............................................................................................................ 34
4.2.1 Response of parental lines to water deficit ................................................................. 34
4.2.2 Linkage map and chromosome assignment ................................................................ 34
4.2.3 Putative QTLs ............................................................................................................. 36
4.2.4 Synteny ....................................................................................................................... 37

4
4.3 Mendelian traits ................................................................................................................. 38
5 CONCLUSION AND FUTURE PROSPECTS ....................................................................... 40
ACKNOWLEDGMENTS ........................................................................................................... 42
6 REFERENCES ......................................................................................................................... 43

5
List of original publications
This thesis is based on the following publications, which are referred in the text with their
roman numerals.
I. Khazaei H, Street K, Bari A, Mackay M, Stoddard FL (2013) The FIGS (Focused
Identification of Germplasm Strategy) approach identifies traits related to drought
adaptation in Vicia faba genetic resources. PLoS ONE 8:e63107
II. Khazaei H, Street K, Santanen A, Bari A, Stoddard FL (2013) Do faba bean (Vicia
faba L.) accessions from environments with contrasting seasonal moisture
availabilities differ in stomatal characteristics and related traits? Genetic Resources
and Crop Evolution 60:2343–2357
III. Khazaei H, O’Sullivan DM, Sillanpää MJ, Stoddard FL (2014) Use of synteny to
identify candidate genes underlying QTL controlling stomatal traits in faba bean
(Vicia faba L.). Theoretical and Applied Genetics (in press) doi:10.1007/s00122-
014-2383-y
IV. Khazaei H, O’Sullivan DM, Sillanpää MJ, Stoddard FL (2014) Genetic analysis
reveals a novel locus in Vicia faba decoupling pigmentation in the flower from that
in the extra-floral nectaries. Molecular Breeding (in press) doi:10.1007/s11032-014-
0100-9
Statement of contributions on these articles:
Author Contributions I II III IV
Conceived and designed the
experiments
HK KS FLS HK FLS HK FLS DOS HK FLS
Performed the experiments HK HK HK HK
Analysed the data HK AB HK HK DOS HK DOS MJS
Contributed
reagents/materials/analysis tools
HK KS AB
MM FLS
HK KS AB
AS FLS
HK DOS MJS
FLS
HK DOS MJS
FLS
Wrote the paper HK KS AB
FLS
HK KS AS
FLS
HK FLS MJS
DOS
HK FLS MJS
DOS
HK Hamid Khazaei, KS Kenneth Street, FLS Frederick L. Stoddard, AB Abdallah Bari, MM Michael
Mackay, AS Arja Santanen, DOS Donal M. O’Sullivan, MJS Mikko J. Sillanpää
The original publications (II, III and IV) are reprinted with the permission of Springer.

6
Abbreviations
AFLP Amplified fragment length polymorphism
BC Backcross
BSA Bulked sergeant analysis
CGIAR Consultative Group on International Agricultural Research
cM CentiMorgans
EST Expressed sequence tag
EUW Efficient use of water
FIGS Focused identification of germplasm strategy
gs Stomatal conductance
ICARDA International Centre for Agricultural Research in the Dry Areas
ISSR Inter-simple sequence repeats
ITAP Intron-targeted amplified polymorphism
KASP Kompetitive allele specific PCR
LG Linkage group
LOD Logarithm (base 10) of odds
MAS Marker assisted selection
PGR Plant genetic resources
QTL Quantitative trait loci
RAPD Randomly amplified polymorphic DNA
RFLP Restriction fragment length polymorphism
RGA Resistance gene analogue
RIL Recombinant inbred line
SCAR Sequence characterized amplified regions
SNP Single nucleotide polymorphism
SSR Simple sequence repeats
STS Sequence tagged sites
TRAP Target region amplification polymorphism
WUE Water use efficiency

7
ABSTRACT
The potential for use of faba bean (Vicia faba L.) is increasing worldwide due to its positive
environmental impact and nutritional interest, but there are many challenges for faba bean
breeding and cultivation. These include its mixed breeding system, its unknown origin and
wild progenitor, its large genome being the biggest diploid genome among field crops, and
its relative sensitivity to biotic and abiotic stresses (e.g., drought). Little is known about the
ecological adaptation of faba bean germplasm, or about the locations and effects of genes
that influence traits related to drought adaptation, especially stomatal morphology and
function as key characters for gas exchange between plant and atmosphere. Thus, the current
study had two main goals, a) to examine whether faba bean germplasm from drought-prone
and drought-free environments differed in leaf traits related to drought adaptation while
testing a novel genetic resources utilization tool, and b) to screen the genome for regions and
candidate genes controlling morpho-physiological traits related to drought adaptation.
Two sets of faba bean germplasm each containing 201 accessions from dry and wet
regions of the world were chosen according to the principles of FIGS (the Focused
Identification of Germplasm Strategy). Leaf morpho-physiological traits related to drought
response (e.g., stomatal characteristics and water status) were measured under well watered
conditions in a climate-controlled greenhouse. Thereafter, two subsets of 10 accessions each
were exposed to water deficit. The significance of the difference between the two sets
indicated the potential of FIGS to search for traits related to abiotic stress adaptation.
Machine-learning algorithms and multivariate statistics showed that the discrimination
between the two sets could be based on pertinent physiological traits, particularly leaf
temperature and relative water content. When the plants exposed to water deficit, leaf
temperature was the most responsive trait.
Four bi-parental populations were developed, of which Mélodie/2 × ILB938/2 showed
the highest number of polymorphic single nucleotide polymorphisms (SNPs) and was
advanced to the F5 generation, in which 211 individuals were tested for 222 SNPs derived
from Medicago truncatula sequence information. The population was phenotyped for several
morpho-physiological traits during 2013 and 2014. In total, 188 polymorphic SNPs were
assigned to nine linkage groups that covered ~ 928 cM with an average inter-marker distance
of 5.8 cM. The map showed a high degree of synteny with the genome of M. truncatula.
Most of the detected QTLs for stomatal morphology and function were in a single region of
faba bean chromosome II syntenic with a segment of M. truncatula chromosome IV that
harbours receptor-like protein kinase. Furthermore, a novel locus, ssp1, for stipule
pigmentation was mapped in a well conserved region of M. truncatula chromosome V
containing some candidate Myb and bHLH transcription factor genes.
The difference between the leaf temperatures of the two FIGS sets (the wet and dry set)
allowed us to find a reliable and cost-efficient phenotyping tool for screening drought
adaptation related traits in this species. Furthermore, using an appropriate mapping
population and using novel M. truncatula-derived SNPs all brought success to detect the
genetic regions and to indicate candidate genes. Furthermore, our results confirm the
genomic data from model plant species can easily be translated to faba bean. Finally,
breeding faba bean for drought adaptation can be made more straightforward by combining
the use of germplasm tools such as FIGS, rapidly assessed phenotypes such as canopy
temperature, and genomic tools for detecting candidate genes.

8
1 INTRODUCTION
1.1 Faba bean (Vicia faba L.)
Faba bean (Figure 1) also known in English as broad bean, field bean, fava bean or horse
bean is an annual herbaceous species. It is belong to the family Fabaceae, which is the third
largest angiosperm family, including 19,400 species and 740 genera (Lewis et al. 2005; Kew
2014). Faba bean is a member of subfamily Faboideae, tribe Vicieae and genus Vicia L. This
genus is unusual in having a huge variation in genome size, from ~ 1,880 Mbp in V.
monantha Retz. to 13,000 Mbp in V. faba (Raina and Rees 1983).
Figure 1 Faba bean. 1 flower, 2 stigma, 3 pod, 4 seed, 5 and 6 seed profiles, 7 pistil. Source:
Thomé (1885)
Faba bean is on the first domesticated food legumes, probably in the early Neolithic
period (8,000 B.C.). The word faba is originally from one of the roots of Greek verb, φάγέώ,
‘to eat’, indicating its basic role as food in ancient Greek and Rome (Muratova 1931). The
wild progenitor and place of origin of faba bean are still unknown (Cubero 1974; Maxted
1993), and no successful cross between faba bean and other Vicia species has been reported.
A Near or Middle East centre of origin has been proposed for the faba bean (Cubero 1974),
with radiation in four directions from the centre: Europe, along the North Africa coast to
Spain, along the Nile Valley to Ethiopia, and from Mesopotamia to India. In addition,
Afghanistan and Ethiopia have been proposed as secondary centres of diversity (Lawes et al.
1983), and subsequently China as well, since its faba bean gene pool is isolated from those
of Ethiopia and western Asia (Zong et al. 2009). Faba bean is classified into four groups

9
according to seed size: minor (tick bean, ellipsoidal seed; 0.4 – 0.6 g / seed), equina (horse
bean, field bean, flattened seed; 0.6 – 1.0 g / seed), major (broad bean, flattened seed; 1.0 –
3.0 g / seed) and paucijuga (a primitive form likely close to wild progenitor; 0.3 – 0.4 g /
seed) (Muratova 1931; Hanelt 1972; Cubero 1974). The major type apparently emerged
separately in the Mediterranean basin and China, the equina type mostly in North Africa and
Middle East, and the minor and paucijuga types mostly in Ethiopia, northern Europe, Nepal
and Bangladesh (Duc 1997).
In 2012, world production was 4.2 million tons (Tg) from nearly 2.5 million ha (M ha)
(FAOSTAT 2014). China was the major producer, with ~1 M ha (38% of the world planted
area) and 1.4 Tg (33% of world production), and Ethiopia was second (0.57 M ha). In the
European Union, its cultivated area was 0.34 M ha with production of 0.97 Tg (Eurostat
2014). Faba bean is the fourth most important cool-season food legume in the world after
chickpea (Cicer arietinum L.), pea (Pisum sativum L.) and lentil (Lens culinaris Medik.)
(11.6, 9.8 and 4.5 Tg, respectively), but all of these are produced on a much smaller scale
than soya bean (Glycine max [L.] Merr.) (241.8 Tg). Recently, interest in growing faba bean
has greatly increased in Europe, Australia and North America because of its adaptation to
cool climates and its high potential protein yield (Satovic et al. 2013).
Faba bean, like most other legumes, is symbiotic with nodule-forming bacteria with N2-
fixing ability, which provides major benefits to cropping systems and the environment, and
contributes to agricultural sustainability by soil improvement activities (Sprent 2009), so not
only is it free of the need for nitrogen fertilizer, it also contributes to the nitrogen nutrition of
the following crop. Faba bean is considered an excellent candidate to provide nitrogen input
into temperate agricultural systems on account of its wide adaptation and high biomass
productivity (Rispail et al. 2010).
Faba bean seeds are a good source of protein, fibre and nutritional value, and are widely
grown for food and feed (Duc 1997). The protein content in faba bean is 24 ~ 35% of the
seed dry matter and is very rich in lysine. It is one of the main sources of cheap protein for
people in the Middle East, Latin America and Africa (consumed as dry or canned), and for
livestock (mostly pigs and poultry) in many developed countries. Furthermore, the immature
beans are widely used as a vegetable. However, the quality-limiting factors, such as vicine,
convicin and tannins compromise the value of faba bean in many diets. Vicine and convicine
are stored in the cotyledons and are harmful for humans carrying a genetic defect (Glucose-
6-phosphate dehydrogenase deficiency, G6PD) and for both laying hen and broiler
production, while tannins in the seed coat reduce protein and energy digestibility in
monogastric animals (Crépon et al. 2010 and references in). Alleles conferring low tannin
content (zt1 and zt2) and low vicine-convicine content (zvc) have been identified and are
used in breeding programs (Crépon et al. 2010; Torres et al. 2010).
Though the agronomic and economic importance of faba bean is well documented, its
cultivation is still limited due to its susceptibility to several biotic and abiotic stresses such as
drought.
1.2 Drought adaptation
Dry lands cover over 40% of the world and are the home of around 2.1 billion poor people in
nearly 100 countries (Mortimore et al. 2009; United Nations 2011). On a global basis,
drought (temporary water deficit coupled with heat stress) is considered to be the most

10
important abiotic constraint to crop productivity (Fischer and Turner 1978; Huang et al.
2002). Climate change will lead to increased temperatures and changed precipitation patterns
in the world. Models predict that climate change attributed to the emission of greenhouse
gases will increase the frequency and intensity of droughts in agricultural areas worldwide
(Oldfield 2005; Gornall et al. 2010; Dai et al. 2013). In order to improve crop performance,
it is essential to understand how crop species respond and adapt to drought conditions. The
key to drought adaptation for plant breeders and crop physiologists is tailoring the
morphology, physiology and phenology of a crop to its environment in order to manage
water economy. Two key aspects of drought physiology are regulation of water use through
the stomata, and regulation of water uptake through the roots. This thesis focuses on leaf
attributes associated with drought adaptation.
Faba bean, as an important source of protein in those dry areas of developing countries
most likely to be impacted by climate change, is reputed to be relatively sensitive to water
deficit among legume species (McDonald and Paulsen 1997; Khan et al. 2010). It is highly
responsive to irrigation (Oweis et al. 2005), but in most production regions it is seldom if
ever irrigated and mostly depends on stored soil moisture and rainfall for its growth and
development. Thus, development of drought-adapted genotypes is an essential mission to
improve yield and its stability in this crop, but progress has been relatively slow compared to
other crops due to lack of efficient screening methods (Stoddard et al. 2006). Wide genotypic
variation in water stress response has been reported in this species (Heringa et al. 1984;
Grzesiak et al. 1997; Abdelmula et al. 1999; Amede et al. 1999; Link et al. 1999; Ricciardi et
al. 2001; Khan et al. 2007), indicating the potential for breeding of drought-prone
environments.
Field phenotyping for drought response is costly (when rain-exclusion shelters have to be
established and maintained) and time-consuming, and often produces unreliable output due
to seasonal variation, since periods of drought vary in length, timing and intensity (Khan et
al. 2007). In contrast, screening under controlled conditions provides the potential for rapid
response and uniform conditions. An ideal phenotyping method should be fast, low-cost,
non-destructive, accurate and working on several samples (Richards et al. 2010; Monneveux
et al. 2014). Then there should be a link to translate laboratory findings in controlled
conditions to the field, and a novel genotype should also perform well in field conditions in
order for the breeding program to be successful (Passioura 2012). Improvement in new
sensor technologies has allowed the development of automated, high throughput and precise
phenotyping platforms. Although costly to install, these platforms streamline and automate
data collection in large numbers of samples and thus practically reduce manual logistics and
some kinds of experimental errors. The LemnaTec (LemnaTec GmbH, Würselen, Germany;
http://www.lemnatec.com/plant-phenotyping.php), PHENOPSIS (Granier et al. 2006),
GROWSCREEN (Walter et al. 2006) and TraitMill (Reuzeau et al. 2006) are some of these
platforms. Some platforms are specialised for certain types of plants or organs, and GlyPh,
designed for high-throughput measurement of plant water use and growth in soya bean
(Pereyra-Irujo et al. 2012), maybe adaptable for use in other legumes such as faba bean.
1.2.1 Response mechanisms
Three types of plant strategies for coping with drought stress have been characterized,
namely drought avoidance, dehydration tolerance and escape (Ludlow and Muchow 1990;

11
Kramer and Boyer 1995). Each of these strategies has its own complex morphological and
physiological mechanisms that involve complex genetic control.
Drought avoidance refers to mechanisms related to keeping high plant water status under
water-limited conditions by minimizing water loss and / or maximizing water uptake through
root characteristics. Stomatal closure is the most important drought avoidance mechanism
and is affected by abscisic acid (ABA) transported from the roots (Schachtman and Goodger
2008). The guard cells are the structures and they open and close the stomata. Stomatal traits
such as morphology (e.g., density and size) and function (e.g., conductance, gs) are
considered key determinants of plant growth and water status (reviewed in publication I).
Faba bean accessions with higher stomatal density were shown to have lower yield and less
resistance to water stress, while lower stomatal density was associated with better drought
avoidance (Ricciardi 1989). Appropriate stomatal activity might be the key role for
improving drought adaptation in faba beans by reducing water loss and increasing
transpiration efficiency (Dawish and Fahmy 1997). The gs was found to be the most
important trait determining water use in relatively small sets of faba bean accessions (Nerkar
et al. 1981; Khan et al. 2007). Carbon isotope discrimination (Δ13
C) has been proposed as a
physiological criterion to select C3 crops, correlated with yield and water use efficiency, as a
result of the balance between photosynthetic activity and gs (e.g., Condon et al. 2004). Khan
et al. (2007) found wide genetic variation for Δ13
C in faba bean, and reported a negative
correlation between transpiration efficiency and Δ13
C under well watered conditions.
Nevertheless, Δ13
C is an expensive measurement, so it is suitable for important subsets of
samples rather than large germplasm surveys. Leaf temperature has been shown to be a rapid
and cost-efficient surrogate for measuring gs and water status in several plant species (Blum
2011a) including faba bean (Khan et al. 2010). Water use efficiency (WUE) has often been
proposed as one the most important determinant of crop productivity under water-limited
conditions and a target for plant drought adaptation. Blum (2009) argued, however, that
selection for high WUE under drought conditions leads to dramatic losses in yield and
drought adaptation. To overcome this, two layers of selection are needed: one for higher
Δ13
C and the other for shoot biomass (Krishnamurthy et al. 2013). Efficient use of water, i.e.,
maximizing soil moisture capture through transpiration, is the target for improvement of
crop yield under drought conditions (Blum 2009).
Cuticular wax protects leaves against non-transpiration water loss. Water stress improved
the deposition of cuticular wax and subsequently enhanced drought avoidance in transgenic
alfalfa (Medicago sativa L.) (Zhang et al. 2005). In the same way, accumulation of cuticular
waxes significantly increased in soya bean plants exposed to water deficit (Kim et al. 2007)
and cuticular wax accumulation is considered an important drought avoidance strategy in
many plants (Kosma et al. 2009; Zhou et al. 2013). However, abundant wax is not always
correlated with drought tolerance (Ristic and Jenks 2002). Variation in the wax content, and
in its synthesis in response to drought stress, have not yet been documented in faba bean.
Root characteristics have been shown to be an important contributor to drought
avoidance in many crop species (Blum 2011a; Monneveux et al. 2014), including legume
species such as groundnut (Arachis hypogaea L.), pigeon pea (Cajanus cajan (L.) Millsp.),
cowpea (Vigna unguiculata (L.) Walp.), soya bean, common bean (Phaseolus vulgaris L.)
(Purushothaman et al. 2013 and references in), chickpea (Kashiwagi et al. 2006; 2008a) and
lentil (Sarker et al. 2005; Kumar et al. 2012; Singh et al. 2013). The uptake of water can be
maximized and dehydration postponed by appropriate root characteristics. The first of these

12
is rooting depth, as deeper roots can continue to extract water from deep in the soil profile
when shallow roots have already exhausted the water available to them (Blum 2011b). Faba
bean is sometimes considered to be relatively shallow rooted, with maximum rooting depth
of 50 – 90 cm, depending on soil type (Manschadi et al. 1998). Extensive variation in rooting
depth exists in faba bean germplasm, for example, rooting depth of eight accessions at 35
days after sowing ranged from 32 to 45 cm (Khan and Stoddard, unpublished data, EU-Faba
project). Previous work on faba bean, which has been focused on traits related to stomatal
activity and other aspects of drought adaptation, has demonstrated the necessary need for
research on roots in this species (Khan et al. 2007; 2010).
Dehydration tolerance refers to maintaining metabolic activities at low tissue water
potential (surviving internal water deficits). Keeping high tissue turgor by osmotic
adjustment (OA), antioxidant defence systems and changed dynamics of plant hormones are
the major physiological stress-adaptive traits to minimize the detrimental effects of water
deficit (Morgan 1984). OA maintains cell water content by increasing the osmotic force that
can be exerted by cells on their surroundings and thus increasing water uptake (Blum
2011a). OA has not been deeply been investigated in faba bean, but it has been demonstrated
in many other legumes (Morgan et al. 1991; Ashraf and Iram 2005; Turner et al. 2007;
Silvente et al. 2012). In chickpea, OA was not related to changes in carbohydrate
composition or leaf gas exchange under drought conditions (Basu et al. 2007). A wide
variation was found in pea genotypes for OA when they were exposed to water stress, with
some producing considerable amounts of soluble sugars and proline (Sánchez et al. 1998).
Concerning faba bean, Amede et al. (1999) and Khan et al. (2007) reported a decline in
osmotic potential in 19 and six inbred lines, respectively, under water stress conditions, but
there was no evidence of osmotic adjustment. A wider spectrum of faba bean germplasm
needs to be surveyed for the presence of osmotic adjustment.
Drought escape is the ability of a crop to complete its life cycle before the onset of
drought or unfavourable conditions. The matching of crop growth to the pattern of rainfall is
one of the most important breeding goals for drought-prone environments, particularly those
prone to terminal drought. Early flowering is the most important phenological trait when
terminal drought is likely. Consequently, early sowing along with early seedling vigour and
better root establishment in dry environments allows rapid ground coverage and as a result
reduces evaporation from the soil (Loss et al. 1997). Terminal drought escape via early
production is a successful breeding strategy in Mediterranean environments (Araus et al.
2004 and references in). Although earliness allows crops to avoid terminal drought, it can
lead to a yield penalty under well watered conditions (Monneveux et al. 2012). There is
considerable variation in days to onset of flowering in faba bean accessions (Stoddard 1993;
Khazaei et al. 2012) and developmental responses to temperature and day length (Iannucci et
al. 2008; Patrick and Stoddard 2010). Earliness is also a desirable attribute for often short
season climates such as boreal zone (Stoddard and Hämäläinen 2011).
At the end it should be noticed that combinations of these traits, not a single trait, are
required to improve yield and its stability under drought conditions. For example, combined
selection for earliness, increased harvest index and phenotypic plasticity has been proposed
as a methodto stabilize legume yield under Mediterranean conditions (Siddique et al. 2003).

13
1.3 Breeding
The situation of crop production in the 21th century has been analysed by many researchers
and does not look bright (Pinstrup-Andersen et al. 1999). Furthermore many biotic and
abiotic stresses significantly reduce crop yields. Plant breeding plays an important role for
increasing crop yield and stress adaptation (Moose and Mumm 2008). Plant breeding
describes methods for the creation, selection, and fixation of superior plant phenotypes for
the development of improved cultivars. The primary goals of faba bean breeding are, those
of like any other crop, to improved yields, seed quality (e.g., low vicine-convicine and
tannin-free), adaptation to biotic and abiotic stresses, and other agronomic traits (Torres et al.
2006; 2010; 2012). Relatively little effort has been made to breed faba bean for adaptation to
abiotic stresses, especially using morpho-physiological traits related to drought adaptation.
1.3.1 Conventional breeding
Faba bean is partially cross-pollinated (allogamous) at frequencies ranging from 4-84%, with
the value determined by the interaction between the plant genotype, its environment, and the
population of pollinators (Bond and Poulsen 1983; Stoddard and Bond 1987). The mixed
mating system means that breeding methods for self-pollinated or cross-pollinated crops are
either not appropriate, or need adjustment. Two main breeding strategies are employed,
population development with recurrent selection, and line breeding with management of the
degree of cross pollination by isolation strategies (Gnanasambandam et al. 2012).
Conventional faba bean breeding may take around 1 decade, from crossing inbred lines,
population development, multi-environment evaluation of progenies for desired traits
(including yield and its stability) and then selecting the superior genotypes. Faba bean can
show high levels of heterosis (Bond 1989a; Link et al. 1996, 1997; Dieckmann and Link
2010; Meitzel et al. 2011). Part of this can be captured in a composite cultivar, where 2-3
inbreed lines are allowed to interbreed freely, and this was the main breeding strategy for
several decades (Bond 1989a). Cytoplasmic male sterility (CMS) systems were developed,
but suffered from instability, as an unacceptably high proportion of male-sterile individuals
reverted to fertility (Bond 1989b; Link et al. 1997). ICARDA maintains two different
categories of faba bean material, ILB (International Legume Bean) accessions from different
countries, and BPL (Bean Pure Line) accessions that are derived through selfing from each
chosen ILB line depending on the variation in it (Saxena and Varma 1985). There is
potential to use homozygous BPLs as parents in faba bean breeding programmes to ensure
the consistency of progeny.
Conventional breeding involves crossing whole genomes followed by selection of the
superior recombinants from among the many segregation products. This procedure is
laborious and time consuming, involving many crosses, several generations, and careful
phenotyping, and linkage drag (tight linkage of undesired traits with the desired loci) may
make it difficult to achieve the desired breeding outcome. DNA marker technologies, the
development of several types of molecular markers and molecular breeding strategies offer
possibilities to plant breeders and geneticists to overcome many of the problems faced
during conventional breeding. Drought adaptation is a complex quantitative trait (Khan et al.
2010) controlled by several genes or QTLs of small effects, so many authors hold that
breeding for it requires the combination of conventional and molecular strategies (Chaves et
al. 2003; Blum 2011a).

14
The total area occupied for faba bean production worldwide dropped by 57% (5.4 to 2.3
M ha) from 1961 to 2012, but average yield has increased around 2-fold (0.9 to 1.7 t/ha)
during the same time (Figure 2). The yield trend is still upward, showing how conventional
breeding efforts have been successful in this crop (Bond et al. 1994).
Figure 2 World faba bean area harvested and yield from 1961 to 2012 (FAOSTAT 2014)
Faba bean yields are, however, considered to be unstable, which has been attributed to
many causes, including the partial dependence on insect pollinators, and the general lack of
improvement in breeding for resistance to numerous environmental constraints (biotic and
abiotic stresses). Highly significant genotype × environment interaction for yield has been
reported in faba bean accessions (Annicchiarico and Iannucci 2008; Flores et al. 2013).
Breeding for adaptation to biotic and abiotic (particularly drought adaptation) stresses were
summarized by Sillero et al. (2010) and Khan et al. (2010).
1.3.2 Genetic resources
There is considerable variation in plant genetic resources (PGR) collections conserved ex
situ in agricultural genebanks, so screening genetic resources would be the first sound step of
any pre-breeding program. Plant breeders find useful genes within these collections that can
have impressive impacts on crop yields and adaptation to biotic and abiotic stresses. The
efficient use of PGR is essential to conserve biodiversity, respond to future challenges
including climate change, and thus pay big dividends to agriculture and environment
(Johnson 2008).
A wealth of germplasm is available, comprising over 7.4 million accessions conserved ex
situ in more than 1,700 national and international agricultural genebanks worldwide, and
about 18% of the accessions are food legumes. Concerning faba bean, 43,695 accessions
(accessed on 15 March 2013 from ICARDA genebank) are conserved mostly within 37
global genebanks. The ICARDA collection exceeds 9,000 accessions (21% of global
0
1
2
3
4
5
6
7
0.0
0.5
1.0
1.5
2.0
1960 1965 1970 1975 1980 1985 1990 1995 2000 2005 2010 2015
To
tal
are
a h
arv
este
d (
M h
a)
Yie
ld (t
/ha)
Year

15
collection), followed by the Institute of Crop Germplasm Resources of the Chinese Academy
of Agricultural Sciences, ICGR-CAAS (10%) and the Australian Temperate Field Crops
Collection, ATFCC (6%) (FAO 2010). Faba bean is fifth among grain legumes for number
of conserved ex situ accessions (after soya bean, ~ 230,000; chickpea, ~ 98,000; pea, ~
94,000 and lentil, ~ 58,000 accessions).
Faba bean is represented in the collections by only the cultivated forms. Both botanical
and molecular outcomes suggest that the wild ancestor of faba bean either has not yet been
discovered or has vanished (Duc et al. 2010). Modernization of agriculture also reduces
genetic diversity in crops, so ex situ collection saved in genebanks as well as in situ
conservation by farmers in their agro-ecosystems are all the more crucial for present and
future faba bean breeding programs. The next challenge concerning faba bean conservation
in genebanks, is that due to the partial allogamy status of this species, maintenance activities
are costly and difficult. Half of the keepers reported that their faba bean collections were
maintained with open pollination (Suso et al. 2005).
Wide genetic variation has already been reported in faba bean genetic resources for agro-
morphological traits such as morphology (Robertson and El-Sherbeeny 1991; Perrino et al.
1991; Suso et al. 1993; Lang et al. 1993; Terzopoulos et al. 2003, 2004; Karaköy et al.
2014), seed size (Nadal et al. 2003), seed composition (Gresta et al. 2010; Khamassi et al.
2013; Karaköy et al. 2014), and yield and its components (Polignano et al. 1993; Keneni et
al. 2005; Terzopoulos et al. 2008). Concerning abiotic stresses, high levels of adaptation
have been reported for winter hardiness (Arbaoui et al. 2008a; Link et al. 2010) and drought
(Amede et al. 1999; Khan et al. 2007). This enormous variability is expected in this ancient
domesticated crop with its wide adaptability to various latitudes and altitudes, its ability to
grow from spring and autumn sowings, and its various usages for food and feed.
In addition, molecular markers as a powerful and more reliable tool have been used for
characterising genetic diversity in faba bean genetic resources. Different marker types have
been used for this purpose such as RAPDs (Link et al. 2005), AFLPs (Zeid et al. 2001; 2003;
Zong et al. 2009; 2010), TRAPs (Kwon et al. 2010), ISSRs (Terzopoulos and Bebeli 2008;
Alghamdi et al. 2011; Wang et al. 2012) and SNPs (Kaur et al. 2014a). Exceptionally wide
genetic diversity was found in accessions from North China, while those from Central China
had less diversity and European and North African germplasm sets were genetically close to
each other. This high level of genetic diversity, confirmed by both phenotyping and
molecular markers, is a key to successful crop improvement.
Since agricultural genebanks conserve large numbers of accessions in overlapping
collections, identifying suitable material is like searching for the proverbial needle in a
haystack. Thus an efficient and rapid tool for mining genebanks is needed. Core collections
were introduced as a way to work with fewer accessions that would represent most of the
genetic variation of a large collection in a relatively small number (5–10 % of the total
collection) of chosen accessions (Frankel 1984). Nevertheless, core collections are not
constructed to highlight or identify materials that are most likely to contain the specific
adaptive traits desired in a particular search (see publication I and references in). In other
words, the core collection concept focuses on genetic diversity, not adaptive phenotype.
The concept of centres of diversity that established the association between diversity and
eco-geographic distribution was proposed in the early 1900s by Nicholas I. Vavilov (1887–
1943), who recognized the importance of locating genetic variation based on eco-climatic

16
conditions (Vavilov 1922). Much earlier, in the 1600s, farmers in eastern North America
were apparently aware of the need for ecological adaption and imported wheats from
western Europe with better resistance to cold and pests (Olmstead and Rhode 2011). The
Focused Identification of Germplasm Strategy (FIGS) was proposed by Mackay and Street
(2004) as a new approach to utilize PGR. FIGS uses available environmental data
(geographic and agro-climatic information) of collection sites to identify manageable sets of
accessions “most likely” and “least likely” to express the trait of interest (Figure 3). This
approach allows rapid reduction of the thousands of accessions to a manageable number with
a high probability of carrying desired traits for the breeder’s strategy (Mackay and Street
2004; Publication I). Odong et al. (2013) classified core collections into three categories,
namely those representing individuals (CC-I), extremes (CC-X) and distribution (CC-D).
Among them, CC-X is similar to FIGS in the sense of considering geographical selection,
but it still does not consider adaptive traits.
Figure 3 Distribution of core collection and FIGS sets in the overall spectrum of genetic
diversity
Novel statistical approaches are becoming available to screen plant genetic resources
more efficiently and precisely, such as machine learning algorithms (Bari et al. 2012; Bari et
al. 2014a, b) and Bayesian mathematics (Dayanandan et al. 2013). The work of this thesis
has run in conjunction with the testing of these methods for identifying potential sources of
abiotic stress in faba bean (Bari et al. 2014c). At the same time, genotyping is becoming
faster and cheaper. Next generation sequencing, NGS (i.e. high-throughput genotyping) is
providing new tools to analyse PGR, such as identifying patterns of genetic diversity,
mapping quantitative traits, and mining novel alleles that are not discovered yet (Kilian and
Graner 2012). This would be a suitable approach for a species such as faba bean with its
several breeding challenges.
1.4 Genomics
Faba bean is a diploid (2n = 12) with six remarkably large chromosomes, which makes it a
suitable tool for cytogenetic studies. Its genome is one of the largest of any diploid field
crops (~13,000 Mbp) and contains more than 95% of repetitive DNA. The faba bean genome
size is 2.6, 3.2, 17.6 and 26 times larger than those of pea, lentil, chickpea and Medicago
truncatula, respectively (Sato et al. 2010) and 4-fold larger than the human (Homo sapiens)
FIGS
Core collection
FIGS

17
genome (Figure 4). Hence, certain kinds of investigation are less advanced in faba bean than
in many other species. Sequence information is available on only 20,550 ESTs in the public
databases, in contrast to nearly 1.5 million for soya bean (GenBank®,
http://www.ncbi.nlm.nih.gov, 20.05.2014). Hence, functional genomic studies, identification
of important genes and development of a highly saturated map are relatively limited in this
species. Nevertheless, significant progress has been made to enrich its genetic and genomic
resources in the last two decades. The lack of genome compatibility of faba bean with other
Vicia species may explain why it is reproductively isolated from them. The mitochondrial
genome sequence of faba bean (cv. Broad Windsor) was estimated at 588,000 bp. It
comprised 52 genes, of which 32 encoded proteins, 3 rRNA, and 17 tRNA. Of the 114
unique open reading frames (ORFs) 36 lacked significant homology to known proteins
(Negruk 2013).
Moreover, advances in NGS combined with techniques for the quantification of gene
expression (e.g., SuperSAGE), are facilitating genome-wide transcriptome studies and target
gene identification in organisms such as faba bean, from which massive nucleotide sequence
information is not yet available. As a result, possible regulators revealed by SuperSAGE
analysis and associated with the ascochyta-faba bean and ascochyta-lentil interactions have
been genotyped and tentatively located on the crop genetic maps (Madrid et al. 2013) in
order to identify potential targets for molecular breeding for ascochyta resistance.
Furthermore, large-scale SSR marker development was successfully achieved using NGS on
the faba bean genome (Yang et al. 2012).
Figure 4 Genome size of faba bean compared to those of some other organisms
0
2000
4000
6000
8000
10000
12000
14000
Arabidopsis Medicago
truncatula
Chickpea Soybean Human Lentil Pea Faba bean
Gen
om
e si
ze (
1C
, M
bp
)

18
Table 1 History of molecular marker application for constructing genetic linkage map in
faba bean
Marker type / total marker Mapping
population / type
Population
size
Total map
length
(cM)
Average inter-
marker
distance (cM)
No. of LGs
(Assigned to
chromosome)
Reference Application
7 RFLPs, 4 morpho, 3 iso, 3
RAPDs (17)
BC – 231.4 – 7 (–) van de Ven et
al. (1991)
M
43 RAPDs, 7 iso, 1 RFLP
(51)
2 F2 families 20+44 300-350 – 11 (1) Torres et al.
(1993)
M
8 Morpho, 7 RFLPs, 4 iso, 4
RAPDs (23)
BC – 300 – 7 (–) Ramsay et al.
(1995)
M, Q
147 RAPDs, 9 iso, 1 morpho
(157)
7 F2 families 813 850 – 48 (6) Satovic et al.
(1996)
M, T, C
105 RAPDs, 7 iso, 3 seed
protein genes, 1 morpho
(116)
3 F2 families 175 ~1,200 20 13 (7) Vaz Patto et
al. (1999)
M, T, Q, C
77 RAPDs (77) F7, RIL 57 973.2 14.66 14 (–) Surahman
(2001)
M
117 RAPDs, 2 iso, 2 seed
protein genes (121)
F2 196 1,445.5 13.72 16 (9) Román et al.
(2002; 2003)
M, Q
176 RAPDs, 6 iso, 4 SSRs, 3
seed protein genes, 2
morpho (191)
11 F2 families 654 1,559 8 14 (5) Román et al.
(2004)
M, T, C
94 RAPDs, 4 iso, 3 SSRs, 2
seed protein genes (103)
F2:3 families 159 1,308 – 18 (6) Avila et al.
(2004; 2005)
Q
135 ITAPs (135) F6, RIL 94 1,685.8 14.6 12 (–) Elwood et al.
(2008)
M, *
131 RAPDs (131) F6, RIL 101 1,635.39 14.73 21 (–) Arbaoui et al.
(2008b)
M, Q
238 RAPDs, 21 Intron-
spanning marker, 6 SSRs, 5
ESTs, 4 iso, 2 STSs, 1
SCAR (277)
F6, RIL 165 2,856.7 12.72 21 (9) Díaz-Ruiz et
al. (2009 a,b;
2010)
Q
167 ESTs, 71 RAPDs, 11
SSRs, 3 RGAs, 3 seed
protein genes, 2 iso, 1
morpho (258)
F6, RIL 124 1,875 7.26 16 (8) Cruz-
Izquierdo et
al. (2012)
M, Q, *
110 SSRs, 9 faba EST-SSRs,
5 grass pea EST-SSRs, 3 pea
EST-SSRs, 1 ITAP (128)
F2 129 1,587 12.5 15 (–) Ma et al.
(2013)
M
121 RAPDs, 38 ESTs, 6
SSRs, 5 RGAs, 1 defense-
related gene, 1 seed protein
gene (172)
F7:8, RIL 119 1,402.1 9.87 29 (15) Gutiérrez et
al. (2013)
Q, C
729 markers in total 3 RILs (RIL1 Cruz-
Izquierdo et al.
(2012), RIL2 Díaz-
Ruiz et al. (2009
a,b), RIL3
Gutiérrez et al.
(2013)
408 4,613 6 43 (13) Satovic et al.
(2013)
M, Q, C
465 SNPs, 57 EST-derived
SSRs (522)
F5:6, RIL 95 1,216.8 2.3 12 (12) Kaur et al.
(2014b)
M, Q, *
LG linkage group, morpho morphological marker, iso isozymes, T Assignation of linkage groups to
chromosomes by trisomic segregation, M Development of a map, Q QTL, C composite map, *
comparative map

19
1.4.1 Marker assisted selection (MAS)
The use of DNA markers in plant breeding is called marker-assisted selection (MAS) and
has opened a new gate in agricultural sciences, ‘Molecular breeding’, since the early 1980s.
Use of new biotechnological methods holds promise to provide tools that assist plant
breeders to develop new cultivars with greater reliability and cost-efficiency. There are three
major types of genetic markers, namely morphological, biochemical and DNA markers.
Three main advantages of selection based on DNA markers over conventional phenotyped-
based selection are the relative simplicity of MAS that may save time and resources, the
potential to use it at earlier stages of plant growth than phenotyping, and the lack of effect of
environment (Collard et al. 2005; Collard and Mackill 2008). MAS is based on the selection
of molecular markers in a segregating population and/or pyramiding desirable alleles at the
target loci. This approach was first used at faba bean breeding in the early 1990s and has had
numerous applications since, such as construction of linkage maps (Table 1), assessing the
level of genetic diversity within germplasm (see Alghamdi et al. 2012; Kaur et al. 2014a and
references in), quantitative trait loci (QTL) mapping, and detection of economically
important genes (Table 2). The current constraints of using MAS in faba bean breeding are
the early stages of its DNA marker technology development, the reliability and accuracy of
QTL mapping studies, and most importantly, the limited range of markers and limited
polymorphism of markers in breeding material, in contrast to some other species.
Sirks (1931) was the earliest to report on the faba bean genome, identifying 19 genetic
factors that formed four linkage groups. His material was lost during World War II. Four
decades later, Sjödin (1971) used translocation lines in the assignment of different loci
(morphological observations) to their respective chromosomes. Several morphological
markers and seven isozyme loci were subsequently added into the faba bean map (Suso et al.
1993; Torres et al. 1995 and references in). Primary trisomics were also used to assign genes
and linkage groups to specific chromosomes (Torres et al. 1998; Van Patto et al. 1999). The
first DNA-base linkage map in this species was constructed with just 17 markers, of which 10
were DNA-based (van de Ven et al. 1991). The faba bean genetic linkage map was initially
developed using morphological, isozyme, RFLP, RAPD and seed protein gene markers that
were limited in number, transferability or synteny with model plant species. The first SSR
markers were described in this species by Požárková et al. (2002) and soon enriched a
composite map (Román et al. 2004). The first gene-base map was reported in 2008 (Elwood
et al. 2008) and more recently enriched by using 511 genic markers (Kaur et al. 2014b) with
2.3 cM average inter-marker density (Table 1). Satovic et al. (2013) published a reference
consensus genetic map that covered 4,062 cM in six main linkage groups corresponding to the
six chromosomes of faba bean.
1.4.2 Markers based on DNA
DNA markers linked to desired traits are a valuable selection tool in modern plant breeding.
Molecular markers have been sought for both simple and complex traits (reviewed in Torres
et al. 2010; 2012). In the case of major genes, the bulked segregant analysis (BSA) method
has been used for the identification of markers that are tightly linked to the genes underlying
parameters such as tannin content (controlled by zt1 and zt2, two independent but
complementary genes), vicine-convicine content (Gutiérrez et al. 2006; 2007; 2008),
hypersensitive response to rust (Avila et al. 2003), and indeterminate growth habit (Vf-TFL1)
(Avila et al. 2006; 2007). The performance of these markers is still not completely

20
satisfactory because of their genetic distance from the trait and their lack of conservation in
all genetic backgrounds. Compared with BSA, the use of a candidate gene approach allows
the development of diagnostic or perfect markers that are completely linked to or within the
selected trait. This approach is possible only when the target gene is well characterized in the
target species or in a related species carrying orthologous genes.
Genic molecular markers are developed from the coding sequences such as ESTs or fully
characterized genes, frequently with known functions, in contrast to most of the previously
developed molecular markers (e.g., RAPDs and AFLPs) that are directly from the total
genomic DNA without any information on function (Varshney et al. 2007). The efficacy of
genic molecular markers may thus be superior to DNA-based markers in molecular breeding.
Gene-based SNPs are the most abundant class of polymorphic sites in any organism’s
genome, providing low genotyping cost per data point, locus specificity (accuracy and
reproducibility), co-dominance, simple documentation and potential for high-throughput
analysis. Thus they have become a powerful tool in genetic mapping, association studies,
assessing genetic diversity and positional cloning (Rafalski 2010; Semagn et al. 2014). The
discovery of SNPs in candidate genes or transcript sequences (ESTs) has been a recurrent
strategy in plant genomics, mainly because SNPs on which the markers are based could
themselves be causative for trait differences (Galeano et al. 2012). SNPs were recently used
to assess genetic diversity in faba bean germplasm (Cottage et al. 2012a; Kaur et al. 2014a).
Elwood et al. (2008) and Kaur et al. (2014b) have developed exclusively gene-based genetic
linkage maps in faba bean. Less progress has been made to develop and characterize EST-
derived SSRs in this species by screening the NCBI database (Gong et al. 2010; Ma et al.
2011; 2013; Akash and Myers 2012) than has been achieved in the other cool-season food
legumes, lentil (e.g., Kaur et al. 2011; Sharpe et al. 2013), chickpea (e.g., Garg et al. 2011;
Hiremath et al. 2011; Jhanwar et al. 2012; Stephens et al. 2014) and pea (e.g., Franssen et al.
2011; Kaur et al. 2012).
1.5 Synteny
Functional genomics in faba bean is more poorly developed than in many other field crops,
especially soya bean, common bean and chickpea among legumes, largely because of its
large genome. An indirect approach such as synteny with a model species may overcome
this shortcoming. Comprehensive comparative genomics to assess synteny can assist back-
and-forth use of genomic resources between different legume species, and also help to
decrease cost and increase the efficiency in genetics and crop breeding by speeding up gene
discovery (Ellwood et al. 2008). Comparative genomic analysis has established the
correspondence between faba bean chromosomes and those of the model legumes (Ellwood
et al. 2008; Cruz-Izquierdo et al. 2012).
Arabidopsis thaliana as a well known model species has played an important role in
illuminating the function of many genes, especially in monocotyledons (Meinke et al. 1998).
Nevertheless, the genomic information of A. thaliana may not be easily translated for use in
legumes, since it is susceptible to different pathogens, is incapable of biological nitrogen
fixation, and lacks many of the secondary pathways and quality traits that are widespread in
legumes (Handberg and Stougaard 1992). Hence, M. truncatula and Lotus japonicus have
been developed as model legumes (Sato et al. 2007; 2010), as both are diploid, with haploid
chromosome numbers of 8 and 6, respectively. The genomes are less than 4% of the size of

21
the faba bean genome (500 Mbp and 472 Mbp, respectively) (Sato et al. 2010). In addition,
they are small self-pollinating plants with short life cycles and able to be transformed by
such methods as using Agrobacterium tumefaciens or A. rhizogenes, making them ideal
model legume species (Rispail et al. 2010). Nearly 264 and 315.1 Mb of M. truncatula
(http://www.medicagohapmap.org/?genome) and L. japonicus
(http://www.kazusa.or.jp/lotus/) have been sequenced and there are more than 260,000 and
240,000 ESTs available in the public domain (Benson et al. 2013, GenBank®,
http://www.ncbi.nlm.nih.gov, last visit 15.05.2014).
Considerable synteny among legume genomes has been reported (e.g., Choi et al. 2004;
Kalo et al. 2004; Zhu et al. 2005; Phan et al. 2006; 2007; Elwood et al. 2008; Kalo et al.
2011; Cruz-Izquierdo et al. 2012). The recent soya bean (Schmutz et al. 2010) and chickpea
(Varshney et al. 2013) whole genome sequences are also being exploited in comparative
mapping studies, in order to identify orthologous genes and further develop efficient
molecular tools in other species such as faba bean.
1.5.1 Medicago truncatula
Comparative genomics studies have shown that M. truncatula (also known as: barrel medic,
barrel medick or barrel clover) has high collinearity with faba bean (Ellwood et al. 2008;
Cruz-Izquierdo et al. 2012; Kaur et al. 2012; 2014b). The high level of conservation in gene
order indicates that M. truncatula can be used to facilitate faba bean molecular breeding
(Rispail et al. 2010). An NCBI BLAST search showed that about 45% of faba bean
mitochondrial DNA sequence was homologous to M. truncatula nuclear sequences (Negruk
2013). The high synteny between M. truncatula and faba bean can be used in orthogonal
gene identification, MAS, and constructing highly saturated genetic linkage maps or physical
maps. Identification of QTLs and then candidate genes also may be used to produce
transgenic elite lines for specific breeding purpose by genetic engineering.
From use of synteny between M. truncatula and pea, 5,460 pea unigenes were identified
through comparative mapping and placed in the consensus map of pea (Bordat et al. 2011).
A high syntenic resolution map between M. truncatula and faba bean using 768 SNPs is in
progress to construct a consensus genetic map in faba bean (Webb et al. in preparation).
Both faba bean and M. truncatula are cool-season legumes. M. truncatula is considered
to be relatively drought-adapted, compared to other legumes (González et al. 1995) and pea
(Gálvez et al. 2005). The genetic similarity and synteny between M. truncatula and faba
bean may help in identifying genes responsible for drought adaptation in faba bean and
accelerate breeding for drought-prone environments. ESTs of genes involved in drought
adaptation in chickpea, pea and M. truncatula have been already identified (Jayashree et al.
2005; Buhariwalla et al. 2005).
1.6 Quantitative trait loci
Many agriculturally important traits (e.g., yield, quality factors and many forms of disease
resistance) are controlled by many genes and are called quantitative traits. Detecting the
major loci of these traits, QTLs, is one of the most important aspects of MAS application in
plant breeding. In QTL mapping, it is generally assumed that there is a small number of
major genes detectable in relatively small samples. Combining phenotypic data and allelic
marker segregation along a genome allows the detection of QTLs, which increases our

22
knowledge on their inheritance and gene action. The summary of detected QTLs in faba
bean is presented in Table 2.
The first QTL mapping in this species was reported by Ramsay et al. (1995), who
detected several loci for morphological and biochemical traits. Mapping QTL in faba bean
for biotic stresses, such as resistance to parasitic plants or pathogenic fungi, has been
attempted (Table 2). Two of the major disease constraints, namely ascochyta blight (caused
by Ascochyta fabae Speg.,) and crenate broomrape, have been widely subjected to QTL
studies on F2 and RIL populations. The QTLs accounting for significant proportions of these
resistances have been identified, mapped and validated in multi-environment trials (Roman
et al. 2003; Avila et al. 2004; Diaz-Ruiz et al. 2009a, b; 2010; Gutiérrez et al. 2013; Kaur et
al. 2014b). In addition, some efforts has been given to rust resistance (Avila et al. 2003), but
relatively little for chocolate spot (Botrytis fabae Sard.) in spite of the importance and
widespread nature of this disease. QTLs for these traits have been assigned to 5 of the 6
chromosomes (I, II, III, V and VI), but no linkage group had been assigned to chromosome
IV until recently, when Ruiz-Rodriguez et al. (2014) assigned linkage group I.B (previously
reported by Cruz-Izquierdo et al. 2012) to this chromosome.
A little progress has also been made in identifying QTLs for abiotic stresses such as frost
tolerance (Arbaoui et al. 2008b), yield components, and early flowering time associated with
drought avoidance during the reproductive stage (Vaz Patto et al. 1999; Avila et al. 2005;
Cruz-Izquierdo et al. 2012), but outcomes still are far behind those gained in biotic stresses.
Saturation of the genomic regions associated with target regions and QTL validation in
multiple environments and genetic backgrounds are prerequisites to uncover reliable marker-
trait associations. Nevertheless, marker density in faba bean remains rather low for the
identification of responsible QTLs / genes and use of MAS approaches. Considerably fewer
QTLs have been reported in this species than in other grain legumes (Kumar et al. 2011). At
present, linkage groups detected in faba bean (mostly by AM Torres’s research group, see
Ruiz-Rodriguez et al. 2014) have been anchored to all faba bean chromosomes, which
provides the source for integration of genetic and physical maps that will facilitate fine QTL
mapping and gene identification in this species.

23
Table 2 Summary of QTL / loci detected in faba bean for morphological traits, biotic and
abiotic stresses and quality traits
Trait No. of
QTL/loci
Chromosome Main output Reference
Morphology
Morphological trait – unknown The first QTL map in faba bean including
morphological and biochemical traits
Ramsay et al.
(1995)
Growth habit 1 V The first molecular marker (CAPs) for
growth habit (ti) by using candidate gene
approach (F2)
Avila et al.
(2006)
1 V Complete gene sequence of Vf-TFL1
using different genotypes
Avila et al.
(2007)
Seed coat colour Sc II Black seed coat colour on chromosome II Cruz-
Izquierdo et
al. (2012)
Biotic stress
Broomrape resistance
Orobanche crenata
Forsk.
3 I, II, VI QTLs Oc1, Oc2, Oc3 explained more than
70% of phenotypic variation
Román et al.
(2002)
4 I, II, VI QTLs Oc2, Oc3, Oc4, Oc5 in 2 years and
3 environments
Díaz-Ruiz et
al. (2010)
7 VI 2 new QTLs added, Oc7 and 8 Gutiérrez et
al. (2013)
Orobanche foetida
Poir.
2 I, III Mapping Of1 and Of2 Díaz-Ruiz et
al. (2009b)
3 V Co-localization of Oc8 and Of3 in
chromosome V confirms a common
resistance against O. crenata and O.
foetida.
Gutiérrez et
al. (2013)
Ascochyta fabae Speg., 2 II, III QTLs associated to ascochyta blight
resistance, Af1 and Af2 explained ~ 50%
of phenotypic variation
Román et al.
(2003)
Díaz-Ruiz et
al. (2009a)
6 III Isolate and organ-specific QTLs for
ascochyta blight resistance, Af3 to Af8
Avila et al.
(2004)
4 I, II, VI Detecting genes associated to ascochyta
blight resistance using SNPs
Kaur et al.
(2014b)
Uromyces viciae-fabae
(Pers.) J. Shört.
1 unknown Identifying the first molecular markers
linked to Uvf-1 gene conferring
hypersensitive resistance against rust (F2)
Avila et al.
(2003)
Abiotic stress
Frost tolerance 5 unknown Frost tolerance and physiologically related
traits
Arbaoui et al.
(2008b)
Yield and its
components
7 VI First putative QTLs for seed weight,
explained 30% of phenotypic variation
Vaz Patto et
al. (1999)
15 unknown Mapping floral characters, yield
distribution and yield components
Avila et al.
(2005)
12 II, V, VI QTLs for days to flowering, flowering
length, pod length, No. seeds and ovules
per pod
Cruz-
Izquierdo et
al. (2012)
Quality traits
Low vicine-convicine vc unknown RAPD markers linked to low vicine-
convicine content and their conversion to
SCARs (F2)
Gutierrez et
al. (2006)
Zero tannin zt1 unknown RAPD markers linked to zt1 and their
conversion to SCARs (F2)
Gutierrez et
al. (2007)
zt2 unknown RAPD markers linked to zt2 and their
conversion to SCARs (F2)
Gutierrez et
al. (2008)

24
2 AIMS OF THIS STUDY
The main aim of this study was to investigate the genetic potential for drought adaptation in
faba bean by developing tools for germplasm utilization and marker assistant selection
(MAS) that can be used in pre-breeding. To reach this end, two collections of faba bean,
each of 201 accessions, originating from the wet and dry regions of the world and chosen
using a novel approach (FIGS), were screened for leaf traits related to drought adaptation
(stomatal morphology and function, and leaf water relations). Advanced biparental
populations were developed to detect QTLs governing these traits, and synteny with M.
truncatula as a model legume species was employed to identify candidate genes. During
population development, the genetic control of some Mendelian characters (e.g., stipule spot
pigmentation, SSP) was also considered.
The specific aims of this study were:
1) to compare the drought adaption-related traits of two faba bean sets originating from
environments with contrasting seasonal moisture availabilities, selected using machine-
learning algorithms. The primary hypothesis was that ecotypic differences occurred so traits
associated with plant moisture regulation and lifecycle would differ between the two sets
(Publication I).
2) to test whether faba bean germplasm from drought-prone and non-drought-prone
environments differed in drought adaption related traits under both well watered and water
deficit conditions. In addition, inter-relationships between studied traits were investigated
(Publication II).
3) a, to develop a gene-based linkage map using SNPs; b, to identify QTLs controlling
drought adaption-related traits; and c, to exploit synteny between faba bean and M.
truncatula to examine the putative gene content of the detected QTLs for candidate genes
(Publication III).
4) to investigate the genetic control of SSP and to exploit synteny between faba bean and
M. truncatula to detect putative candidate genes (Publication IV).

25
3 MATERIALS AND METHODS
The experimental part of the work is described here as a general outline.
Table 3 Summary of methods used in this work are listed below. More details are described
in the original publications (I − IV) and following text
Method Publication
Focused Identification of Germplasm Strategy (FIGS) I, II
Machine-learning algorithms* I
Screening drought adaptation-related phenotypes I, II, III
Evaluation of water deficit response II, III
Multivariate statistics II
SNP genotyping III, IV
DNA extraction* III, IV
QTL analysis III
Synteny analyses with M. truncatula III, IV
Developing a genetic linkage map III
Crossing and population development III, IV
KASPar assay* III, IV
Backcrossing IV
Microsatellite loci screening (confirmation of hybrids) IV
Methods marked with asterisk were not carried out by HK

26
3.1 Germplasm survey
3.1.1 Construction of the FIGS sets
The “dry” and “wet” sets each comprised 200 accessions (listed in supplementary Table 1
publications I and II), plus one of known response from earlier investigations (ILB 938/2 and
Aurora/2, respectively) (Khan et al. 2007; Khazaei et al. 2010). Collection sites were
identified where the long-term average annual rainfall was between 300 and 550 mm/year
(dry set) and over 800 mm / year (wet set), respectively. After the sites were identified, one
accession per site was chosen at random. A hierarchical cluster analysis was performed using
the collection site agro-climatic parameters (long-term yearly precipitation, long-term yearly
aridity index, long-term yearly minimum temperature, long-term yearly maximum
temperature, temperature seasonality, precipitation seasonality, precipitation of wettest
quarter and precipitation of coldest quarter; http://www.worldclim.org/bioclim) in order to
achieve a balanced representation of sites (more details can be found in publication I and II).
3.1.2 Germplasm screening
The two sets were evaluated using randomized complete block design with 4 replicates for
some morphological, physiological and phenological traits during 2010−2011 under well
watered conditions (measurements listed in Table 4). In the next step, on the basis of
principal component analysis (PCA) on studied traits, ten representative accessions from
each set (wet and dry, called “subsets”) were selected for exposure to transient moisture
stress following Khan et al. (2007) (publication II).
3.2 Molecular breeding
3.2.1 Mapping population development and selection
Four parental lines [Aurora/2 (drought-sensitive), Mélodie/2 (High EUW), Disco/2 (zt2) and
ILB 938/2 (high WUE)] differing in water-deficit response and other valuable traits were
chosen at the start of the project for crossing and development of RIL populations. Crosses
Mélodie/2 × ILB 938/2, Mélodie/2 × Disco/2, ILB 938/2 × Aurora/2 and ILB 938/2 ×
Disco/2 were prepared by hand, along with their reciprocals, in the insect-proof greenhouse.
The standard sets of quantitative-genetics generations (F1, F´1, F2, and BC1F1) were prepared.
The F2 seeds were advanced to the next generation (F3–F4) by single seed descent, each
comprising ~ 350 individuals. The parental lines were in a collection of 35 accessions used
in the development of 756 SNPs that have been validated in an international panel (Cottage
et al. 2012b; Webb et al. in preparation). Mélodie/2 and ILB 938/2 showed the highest
genetic distance from each other, with 222 polymorphic SNPs, so this cross was advanced to
F5 and chosen for deeper investigation (publication III and IV). Mélodie/2 was selected from
a French low vicine-convicine cultivar with high EUW, while ILB 938/2 was a selection
from an Ecuadorian landrace with relatively high WUE.
3.2.2 Genotyping
High throughput genomic DNA was extracted by LGC Genomics laboratory (LGC
Genomics, Hoddesdon, UK) according to the manufacturer’s instructions

27
(http://www.lgcgenomics.com/nucleic-acid-extraction/services/dna_extraction_services/).
SNP genotyping was accomplished using the KASPar™ (Kompetitive Allele Specific PCR)
assay (KBioscience, UK) platform, a single-plex SNP genotyping methodology using allele-
specific amplification followed by fluorescence detection for genotyping
(http://www.kbioscience.co.uk/reagents/KASP_manual.pdf). The SNP sequences and
specific alleles for parental lines used in this study a listed in Supplementary Table 2,
publication III.
3.2.3 Phenotyping
The F5 population (Mélodie/2 × ILB 938/2) and its with parental lines were phenotyped in a
climate-controlled glasshouse during 2013−2014 for some morphological and physiological
traits (see Table 4) using a completely randomized design with 3 replicates under well
watered and water deficit conditions (publication III). Furthermore, the population was
phenotyped for vicine and convicine content (data not shown in this thesis).
3.2.4 QTL mapping, synteny and candidate genes
Details of genetic linkage map construction and QTL analysis are presented in Table 5. SNP
segregation was subjected to the chi square goodness-of-fit test to assess deviations from the
expected Mendelian segregation ratio (1:1). SNPs showing normal diploid segregation (P ≥
0.05) were chosen for the linkage map construction. The Kosambi function was used to
calculate the map distance in centiMorgans (cM) (Kosambi 1943). Composite interval
mapping (CIM) was used to detect the relationship between each linkage group and putative
QTL locations. Significant QTLs were analysed with CIM. Cofactors were determined using
the forward and backward method in the standard CIM model with the probability in and out
of 0.1. The 95% significance threshold for QTL detection was determined by the phenotype
permutation using 1,000 permutations at an experiment-wise P < 0.05.
The overall level of collinearity between the detected putative QTLs (using sequences
flanking SNPs) and M. truncatula was examined. Reciprocal best BLAST hit E-values were
employed to determine the strength of the orthologous relationship. Gene family,
phylogenetic context, and tissue-specific gene expression profiles of the orthologous genes
from model plant species were examined using the LegumeIP database
(http://plantgrn.noble.org/LegumeIP/) (M. truncatula, Gene Model, Mt3.5v3).
3.3 Measurements
Details of measurements are listed in Table 4. For the leaf morphology (stomatal
morphology and leaflet area), gas exchange traits and leaf temperatures the youngest, fully
expanded leaflets of 8-week-old plants were used in all experiment (publications I, II and
III).
3.3.1 Induction of water deficit (Publications II and III)
For publication II, transient drought response (subsets in 2011), 2 L plastic pots were filled
with 1.42 kg of the mixture (sand / peat, 3:1 v/v) that had a water holding capacity (WHC) of
20% (w/w), using a completely randomized factorial design with 4 replicates. Each pot was
brought to WHC by adding 285 ml of water. Pots were weighed every 2 days and amounts of
water equal to the loss in weight were added. Ten days after sowing, 60 g of perlite was

28
added to the top of each pot (to reduce soil evaporation). In the well watered treatment, the
pots were irrigated as described above until harvesting at 13 weeks (when the main tiller of
the plants turned yellow). Half of the plants were exposed to a gradual and uniform water
stress starting at 5 weeks by reducing water by 2% (w/w) of available water per every two
days to bring the moisture level down from the field capacity (20% w/w) to moisture stress
(2–4% w/w). Pots were weighed and where water use exceeded 2%, irrigation was applied.
Measurements were done 2 weeks after the gradual drying was started.
For publication III in 2013, 4 L pots were filled with 2.48 kg of the mixture (sand / peat,
2:1 v/v) that had a WHC of 24% (w/w). Each pot was brought to WHC by adding 595 ml of
water. Pots were weighed three times per week and amounts of water equal to the loss in
weight were added to maintain soil moisture level at field capacity. Immediately after 8-
week-old plants were phenotyped under well watered conditions, a uniform water stress was
started by reducing relative available water by 2% (w/w) every day to bring the moisture
level down from the field capacity (24% w/w). Pots were weighed every day and where
water use exceeded 2%, irrigation was applied (Khan et al. 2007). When the soil moisture
content had reached 2−4%, 10 days after the induction of water deficit was started, canopy
temperature was measured. Thereafter normal irrigation was continued until harvest.
3.4 Growth conditions and statistical analysis
All experiments (germplasm survey and mapping population phenotyping) were conducted
in the climate-controlled glasshouse of the Department of Agricultural Sciences, University
of Helsinki, Finland. Soil moisture level was maintained at field capacity with automatic
irrigation for all plants in the well watered experiments. Seeds of all accessions were
inoculated with Rhizobium leguminosarum biovar. viciae (faba bean strain, Elomestari Oy,
Tornio, Finland) before sowing. A mixture of sand (size: 0.5–1.2 mm, Saint-Gobain Weber
Oy Ab, Helsinki, Finland) and peat (White 420 W, Kekkilä Oy, Vantaa, Finland) containing
all essential nutrients was used as the potting medium. For all experiments, at three and five
weeks after sowing, 70 ml of nitrogen-free fertilizer (equivalent to 20 kg of P and 24 kg of K
per ha) was added to each pot. Photoperiod was adjusted to 14 h light and 10 h dark, and the
temperature was maintained at ~ 21°C day/15°C night ±2°C. Photosynthetic photon flux
density (PPFD) was approximately 300 µmol m-2
s-1
at the canopy level. Relative humidity
was maintained at 60%. The temperature, humidity and light conditions were automatically
recorded throughout the experiments. For the germplasm survey experiments, 2 L pots used
with mixture of sand and peat (3:1 v/v) was used. In the population phenotyping experiment,
4 L plastic pots filled by sand and peat (2:1 v/v) used. Statistical analysis and software used
to analyse data in this thesis are presented in Table 5.

29
Table 4 Measurements and methods used in the experiments
SD stomatal density, SL stomatal length, SW stomatal width, SA stomatal area, SAAL stomatal area per
unit area of leaflet, Anet photosynthetic rate, gs stomatal conductance, E transpiration rate, WUEi
Intrinsic water use efficiency (Anet/gs), PPFD photosynthesis photon flux density
a In publications I and II, the canopy temperature differences from air temperature and leaflet
temperature differences from air temperature were calculated
Measurements Germplasm survey Mapping
population
Instrument / Method / Reference Publication
Stomatal
morphology
Yes (SD, SL, SW, SA,
SAAL)
Yes (SD, SL) Impression method (Wang and Clarke
1993), using abaxial surface of leaflet
I, II, III
Leaflet area Yes No LI-6200 (LI-COR Inc., Lincoln, NE,
USA)
I, II
Number of tillers Yes No Counting I
Gas exchange traits Yes (Anet, gs, E, WUEi) Yes (gs) LI-6400 (LI-COR Inc.), equipped
with a 2×3 cm leaf chamber with a
LED light source (6400-02B, 90% red
and 10% blue), reference CO2
concentration = 400 µmol mol-1
, flow
rate = 400 µmol s-1
, PPFD = 1,000
μmol m-2
s-1
I, II, III
Leaf temperaturea Yes No LI-6400 (LI-COR Inc.) I, II
Canopy temperaturea Yes Yes Infrared thermometer (IRT, FLUKE®
thermometer gun 574, Everett, WA,
USA)
http://www.plantstress.com/methods/I
RT_protocol.htm
I, II, III
Water use subsets No Weighing pots (according to Khan et
al. 2007)
II
Biomass WUE subsets No Total biomass / water used II
Relative water
content
Yes No According to Barrs and Weatherley
(1962)
I, II
Days to flowering Yes No Number of days to the onset of
flowering
II
Seed weight No Yes Weighing 10 mature dried seeds per
plant
III
Stipule spot
pigmentation
No Yes Scoring the absence / presence of
pigmentation
IV
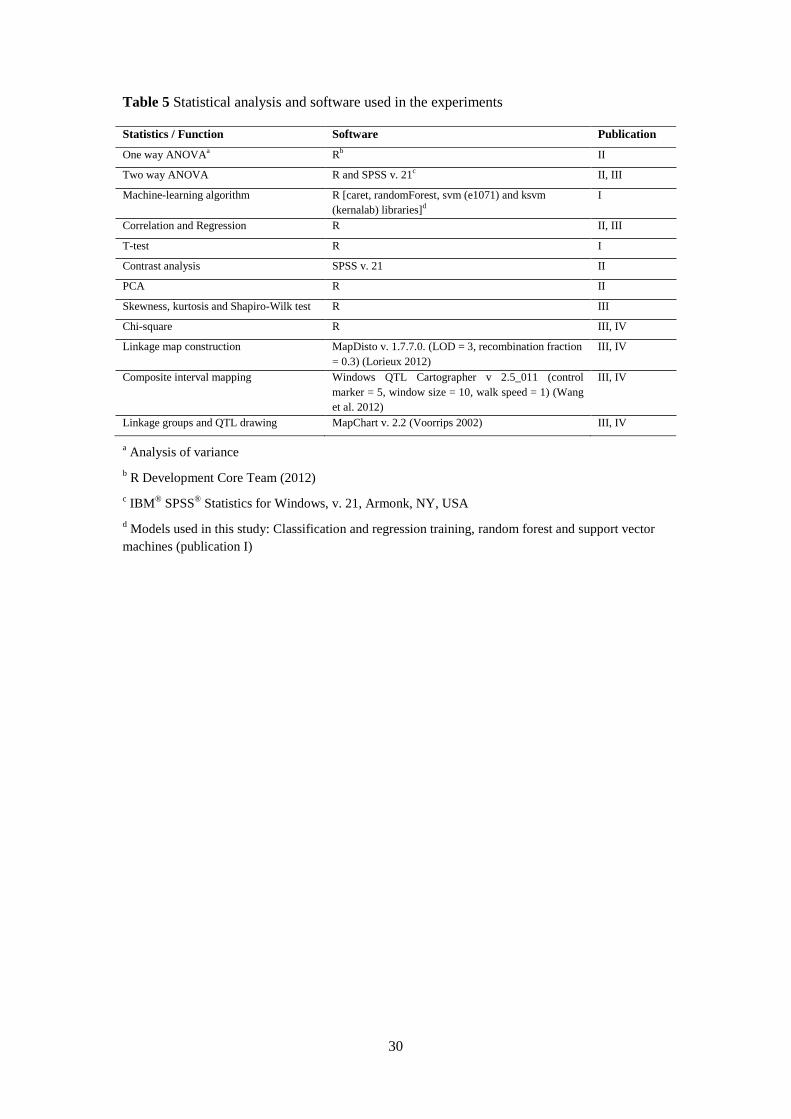
30
Table 5 Statistical analysis and software used in the experiments
Statistics / Function Software Publication
One way ANOVAa R
b II
Two way ANOVA R and SPSS v. 21c II, III
Machine-learning algorithm R [caret, randomForest, svm (e1071) and ksvm
(kernalab) libraries]d
I
Correlation and Regression R II, III
T-test R I
Contrast analysis SPSS v. 21 II
PCA R II
Skewness, kurtosis and Shapiro-Wilk test R III
Chi-square R III, IV
Linkage map construction MapDisto v. 1.7.7.0. (LOD = 3, recombination fraction
= 0.3) (Lorieux 2012)
III, IV
Composite interval mapping Windows QTL Cartographer v 2.5_011 (control
marker = 5, window size = 10, walk speed = 1) (Wang
et al. 2012)
III, IV
Linkage groups and QTL drawing MapChart v. 2.2 (Voorrips 2002) III, IV
a Analysis of variance
b R Development Core Team (2012)
c IBM
® SPSS
® Statistics for Windows, v. 21, Armonk, NY, USA
d Models used in this study: Classification and regression training, random forest and support vector
machines (publication I)

31
4 RESULTS AND DISCUSSION
4.1 Germplasm screening
The morpho-physiological and phenological measurements of the dry and wet FIGS sets
differed significantly under well watered conditions. The dry set had significantly larger
stomata, greater stomatal area per unit area of leaflet, higher relative water content (RWC),
leaflet area and transpiration rate, and in contrast lower leaflet and canopy temperature,
fewer fertile tillers and fewer days to flowering than the wet set. Greater variation for most
traits was observed within the wet set. Stomatal density and stomatal area per unit area of
leaflet were negatively correlated with gas exchange parameters and positively correlated
with intrinsic water use efficiency, while stomatal size showed the opposite trends
(publications I and II).
Three machine-learning algorithms (classification and regression training, random forest
and support vector machines) were employed to examine difference between the two FIGS
sets (publication I). The outcome revealed that sets were classified into two groups based on
the phenotypic evaluation with little or no overlap between sets, showing a high degree of
discrimination between sets. Among the traits, leaflet and canopy temperature and RWC
contributed most to the discrimination between sets (Table 5 in publication I). Machine-
learning algorithms have been little used in this context, so their results were confirmed by
using a well known multivariate statistical methods, principal component analysis, that
further assessed the pattern of variation in the two sets and confirmed the results of the
machine-learning algorithms results. The first four principal components (PC1-4) explained
81.3% of the total variation, of which the first component (PC1) accounted for around half.
Gas exchange traits (photosynthetic rate, gs, intrinsic WUE and transpiration rate) made the
highest contribution to PC1. The second PC was highly dependent on leaflet and canopy
temperature along with RWC. These traits contributed more than other measurements to
discriminate the sets from each other (Figure 5), and also made the greatest contribution to
separating the two sets according to machine-learning algorithms (see publication I). The
next components were mostly based on stomatal morphology. Distribution of the two FIGS
sets shows how FIGS successfully demonstrates that ecotypic differentiation driven by
moisture availability occurred in faba bean germplasm (Figure 5). Similarly, several salt-
tolerant accessions in wild Phaseolus species were identified from arid, coastal, or saline
areas close to their centre of diversity in Central and South America (Bayuelo-Jiménez et al.
2002). When applied to Lathyrus species, FIGS also allowed selection of manageable
subsets with a high probability of finding accessions adapted to drought and heat (Shehadeh
2011). Furthermore, FIGS has been used to detect numerous sources of resistance to biotic
stresses in wheat (Triticum ssp.) and barley (Hordeum vulgare L.) (e.g., Kaur et al. 2008;
Bhullar et al. 2009; Endresen et al. 2011; Bari et al. 2012, 2014b).

32
Figure 5 Principal component analysis scatter diagram based on morpho-physiological traits
in the wet set (black circles) and dry set (grey circles) of faba bean germplasm
Faba bean has been grown for hundreds or thousands of years in a wide range of
environments, including those subjected to often severe drought constraints (Duc et al.
2010), so we expected notable variation between sets from contrasting seasonal moisture
availabilities. Ecosystem disturbance has been shown to add further selection pressure in
annual plant evolution (Peleg et al. 2005). Other methods demonstrated the existence of
ecological adaptation in several plant species (De Micco and Aronne 2012; Sarris and
Koutsias 2014). All of these results support the hypothesis that accessions with desirable
adaptive traits can be identified through the process of predicting the environmental profiles
that select for these traits, and the question was whether FIGS would adequately predict
these profiles, and would do so better than earlier methods. Analysis of natural genetic
variation leads to the discovery of novel genes and alleles, especially in the field of plant
adaptation to the environment (Shindo et al. 2007). Crop wild relatives are the most
promising sources of novel alleles to improve drought adaptation in crops (Nevo et al. 2002),
but in the case of faba bean, there is no wild relative available, so novel methods for utilizing
PGR such as FIGS play a key task in this important aspect. In contrast, the use of core
collections often misses adaptive and rare alleles (Gepts 2006; Pessoa-Filho et al. 2010).
Evaluation of drought adaptation requires phenotyping by exposing plants to contrasting
water regimes. So, the next step sought natural variation within and between the wet and dry
sets for response to water deficit using manageable subsets, 10 accessions representing the
average of each set based on PCA. A moderate, transient, mid-season water deficit was used,
as this is typical of what a crop may experience in the field almost anywhere in the world
(Araus et al. 2002). Water deficit affected most traits and a significant set × water stress
interaction was observed (publication II). Most traits, including canopy temperature
depression (Figure 6), clearly separated two sets under well watered conditions, and the
-6 -4 -2 0 2 4 6-4
-2
0
2
4
PC
2 (
16
%)
PC 1 (46.2%)

33
response of most traits to water deficit also clearly differed between the two sets. The most
outstanding drought adaptation capacity was found in accessions from the dry set, directing
future exploration to these ecosystems. Genetic variation has been shown in leaf temperature
RWC, Δ13
C and gs of a small set of drought-adapted and drought-sensitive lines of faba bean
(Khan et al. 2007). Our result showed that water stress caused stomatal densities to increase
significantly in the dry set while stomatal size decreased in both sets. Stomatal area and
density were negatively correlated to each other in all experiments, so the stomatal area per
unit of leaflet remained fairly constant in the two sets. Stomata do not move. The results
relates partly to reduced leaf size in both sets during the drought interval, due to reduced cell
elongation from low turgour. Differences in stomatal density between the sets suggests that
something else occurred, either more differentiation of guard cells in the dry set, or greater
noise in one set that obscured the differences. These findings were in agreement with those
on several other species (Franks and Farquhar 2007).
Under water deficit conditions, gas exchange fell strongly. The clearest differences
between sets were in leaf temperatures and RWC. The dry set accessions had cooler
canopies under well watered conditions and their leaf temperatures increased more under
water deficit than those of the wet set (Figure 6). This demonstrates that accessions from dry
regions have more effective stomatal control than those from wet regions in both wet and dry
conditions. Overall, these results confirm the key role of stomatal function in water relations
in this species. In the field, lower canopy temperature under drought stress conditions
indicates a better capacity to take up water from deep soil and hence better water status
(Blum 2011a). Fast and accurate phenotyping criteria are currently seen as a major issue for
improvement of drought adaptation in crops (Richards et al. 2010). Canopy temperature can
be measured by an infrared thermometer (IRT) as a rapid and cost-efficient alternative for
preliminary screening for drought adaption under controlled and uniform conditions in faba
bean (Khan et al. 2007, 2010). Canopy temperature has also been shown as an efficient field
phenotyping tool for drought adaptation in other legume species (e.g., soya bean, McKinney
et al. 1998; chickpea, Kashiwagi et al. 2008b; cowpea, Hall 2012). Canopy temperature
measured by IRT as a screening technique for dehydration avoidance was first reported by
Blum et al. (1982). Under drought conditions, this is the most drought adaptive trait
associated with yield performance and has high heritability (Olivares-Villegas et al. 2007).
Good practice in using the IRT, such as those protocol at
http://www.plantstress.com/methods/IRT_protocol.htm, should be followed carefully to
avoid large error variance and non-repeatable results. Water stress caused a reduction in
RWC in both sets, but it was greater reduction in the wet set (27%) than in the dry set (19%).
The ability of genotypes to maintain RWC when moisture is limiting is a good indicator of
drought adaptation (Barrs and Weatherley 1962; Anyia and Herzog 2004; Blum 2011a).
In the current study we screened a large number of faba bean accessions (402), which
showed a wider range of variation for morpho-physiological measurements than previous
reports, suggesting a high potential for breeding for these traits. For example, in our
germplasm gs ranged from ~ 0.040 to 0.620 mol m-2
s-1
, in contrast to the range of 0.125 to
0.176 mol m-2
s-1
in six accessions known to differ for drought response (Khan et al. 2007).
Similarly, ten faba bean accessions (Tanzarella et al. 1984) showed a much narrower range
(77% less) for stomatal density than our work. These comparisons clearly show the enhanced
identification of genotypic variation that results from the large germplasm set analysed in
this work, and from using FIGS to seek extremes in a trait.

34
Figure 6 Effects of water treatments on the canopy temperature depression in dry and wet
set: well watered (white column); water deficit (gray column). Error bars show mean ± 1
SEM
4.2 Molecular breeding
4.2.1 Response of parental lines to water deficit
The parents used for map construction showed different responses to water deficit.
Mélodie/2 had a cooler canopy under well watered conditions and a sizeably higher increase
in canopy temperature under water deficit conditions than ILB 938/2. The gs showed the
reverse trend. Water deficit had 3-fold more effect on biomass production in Mélodie/2 than
in ILB 938/2 but biomass in Mélodie/2 under water deficit conditions was the same as ILB
938/2 under well watered conditions (publication III). These results show the ability of ILB
938/2 to maintain higher water status under water deficit conditions, as it was selected for
high WUE with relatively low yield. In contrast Mélodie/2 with higher EUW showed better
productivity under drought conditions than ILB 938/2, with better water uptake ability
through a well developed root system (Khazaei and Stoddard unpublished data). These two
parental lines nicely explain the concept of WUE and EUW in sense of Blum (2009).
Selection of parental lines, which are genetically and geographically far enough from each
other, is the key point to build a promising RIL populations for successful genetic mapping
and QTL detection (Würschum 2012).
4.2.2 Linkage map and chromosome assignment
Two hundred and eleven individuals were randomly chosen (of 350) from the population
Mélodie/2 × ILB 938/2 for phenotyping and genotyping (publication III). After considering
Wet set
Dry set
-2.0 -1.5 -1.0 -0.5 0.0 0.5
***
Canopy temperature depression (C)
***

35
rejected markers based on the grounds of excess missing data (> 5% missing data per
marker) or excessive expected chi-squared goodness of fit ratio, 188 SNPs, as along with
seed coat colour (publication III), hilum colour and SSP (publication IV) were successfully
mapped in nine linkage groups that covered ~ 928 cM of faba bean genome. The linkage
groups were composed of 2–43 loci and their length ranged from 3.07 to 223.02 cM, with an
average inter-marker density of 5.8 cM. The map showed a high degree of synteny with the
genome of M. truncatula (publication III). High-throughput SNPs have greatly accelerated
the development of high density linkage maps, with 2.3 cM average marker density having
been achieved in faba bean recently (Kaur et al. 2014b; more information in Table 1).
The number of genetic linkage groups should be the same as chromosome number in the
species, so six LGs is expected in faba bean. Nevertheless, the number of LGs generally
exceeds the chromosome number, ranging from 7 to 48 in faba bean, and map length varied
from 231.4 to 4,613 cM (Table 1). Our genetic map had seven main linkage groups and two
fragments which is slightly more than the appropriate number, and the map was slightly
smaller than some others (Table 1). These differences may be related to factors such as the
large size of the genome, the number and types of markers, the software analysis used for
construction of the genetic map (e.g., recombinant fraction and LOD value), the number and
generation of progeny, and especially the genetic background and genetic distance of the
parents (Young 2000; Broman 2006; Semagn et al. 2006; Cheema and Dicks 2009).
The correspondence of linkage groups to their respective chromosomes was performed
using markers with identified chromosome location or linkage according to Cruz-Izquierdo
et al. (2012) and the current consensus genetic map published by Satovic et al. (2013). The
assignments made here are consistent with those possible using the higher syntenic
resolution possible using a 768-locus consensus genetic map (Webb et al. in preparation)
(Figure 7). Seven of the nine linkage groups were assigned to the six faba bean
chromosomes (I–VI) (publication III).

36
Figure 7 Synteny between M. truncatula (Mt) and faba bean (Vf) chromosomes. This figure
was extracted from Webb et al. (in preparation)
4.2.3 Putative QTLs
There is an urgent need to identify chromosome regions associated with the climate change-
related traits in crops in order to assure food security in the near future. Progress in MAS for
a biotic stresses such as drought in faba bean is relatively modest compared to that on biotic
stresses, although some progress have been made for yield and its components (Table 2 and
Torres et al. 2010). There is no corresponding previous report on morpho-physiological traits
related to drought adaptation such as stomatal morphology and function, so the QTLs
detected in this study would be a good starting point for future studies. QTLs were identified
on two of the six faba bean chromosomes, II and IV. Fourteen QTLs were detected from five
studied traits (stomatal density and length, gs, canopy temperature and seed weight) under
well watered conditions over two years (2013 and 2014) and one QTL for canopy
temperature under drought conditions in 2013. The LOD threshold at P = 0.05 varied from
2.8 to 3.0. The proportion of phenotypic variation explained by each individual QTL ranged
from 5.7% to 9.3%. Thirteen QTLs were located on chromosome II, with eight of them
clustered in the same genomic region (qSD-2013-1, qSD-2014, qSL-2013, qSL-2014, qgs-
2013, qCT-2012, qCT-2013 and qCT-2014), in the marker interval, Vf_Mt4g014430_001.
These traits also showed strong phenotypic correlations.
All traits were phenotyped in two years under well watered conditions, and the detected
QTLs remained stable between years except that for gs. Canopy temperature was also

37
phenotyped under water deficit conditions in 2013 and its QTL (qCTd-2013) was mapped in
the same chromosome, chromosome II, ~ 125 cM from those found under well watered
conditions (publication III). The direction of additive effect illustrates that alleles driving
warmer leaves under well watered conditions probably came from Mélodie/2 and those
conferring cooler leaves under stress conditions were from ILB938/2, which is in agreement
with the concept of WUE and EUW (Blum 2009).
A QTL was detected in chromosome II for seed weight, which remained stable during
the two years of phenotyping (qSW-2013-1 and qSW-2014-1). The second stable QTL
governing seed weight was located on chromosome IV (qSW-2014-1 and qSW-2014-2).
QTLs for this trait have been previously reported in chromosome VI (Vaz Patto et al. 1999).
This difference is at least partially attributable to the different parents used, with the study by
Vaz Patto et al. (1999) using paucijuga and major lines whereas the present study used
equina (ILB 938/2) and minor (Mélodie/2) lines, and partly to the different distribution of
markers in the two studies.
Many QTLs governing flowering time and reproductive traits associated with drought
avoidance in faba bean were already mapped on chromosomes I, IV, V and VI (Cruz-
Izquierdo et al. 2012; Satovic et al. 2013; Table 1). In this work, however, transient rather
than terminal drought was the objective, and it was a glasshouse-based experiments,
assessment of grain yield was not considered appropriate. Yield is related to high gs,
photosynthetic rate and cooler canopies in several crops (Fischer et al. 1998; Khan et al.
2010), so the region in chromosome II that made a significant contribution to stomatal
function warrants further investigations.
A short-term mapping population (e.g., F2 and BCs) developed through the BSA
approach is considered a good starting point for molecular mapping, while a long-term
mapping population such as one comprised advanced RIL is suitable for characterization of
traits of importance. Several QTLs in this species were detected at the F2 generation,
especially before 2008, and some of them were later validated at advanced generations, e.g.
ascochyta blight resistance (Díaz-Ruiz et al. 2009a) (see Tables 1 and 2). In more recent
studies, mapping populations are generally advanced to the F6 or F7 generation (Table 1 and
2). In this study, we used an F5 RIL population and Takuno et al. (2012) pointed out that
QTL mapping does not necessarily require RILs at F6 or higher generations, and that F4 (or
even F3) populations would be almost as useful as highly advanced populations.
Two QTLs governing stomatal density and gs (qSD-2013-2 and qgs-2014) were mapped
in regions of distorted SNPs (publication III). The precision of the linkage map (map
distances and marker ordering) may be biased in regions of many distorted markers (Lorieux
et al. 1995a, b). Additionally, QTL regions often overlap with regions undergoing
segregation distortion, because favourable QTL alleles are in excess frequencies in those
regions (Boopathi 2013). Therefore, QTLs found in these regions are more uncertain than
those located elsewhere.
4.2.4 Synteny
Despite the large difference between the genome size of faba bean and M. truncatula, a tight
relationship was found between the two genomes (Figure 7), with strong evidence for
extensive collinearily between linkage groups as it suggested by Rispail et al. (2010). Based
on annotated genic SNPs derived from M. truncatula, candidate genes were found within

38
detected QTL intervals. Candidate genes for M. truncatula and Arabidopsis were almost
identical, and were mostly related to growth and development, stress and transport
(publication III). A segment of M. truncatula chromosome IV that harbours receptor-like
protein kinase (RLKs) was consistently linked with most of the detected QTLs, including
those for stomatal density and length, gs and canopy temperature under well watered
conditions. The candidate gene for canopy temperature under water deficit was ribose-
phosphate pyrophosphokinase 4 in M. truncatula chromosome VI. RLKs are the best known
stress-induced genes and control a wide range of physiological responses such as growth
regulation and specific adaptations in several plant species (e.g., Casson and Gray 2008;
Marshall et al. 2012; Osakabe et al. 2013). Gene ontology for both intervals with detected
QTLs for seed weight were related to regulation of seed germination (Arginyl-tRNA-protein
transferase 1) (Manahan and App 1973; Holman et al. 2009) and embryonic development
ending in seed dormancy (Splicing factor 3B subunit 4) (Raab and Hoth 2007; Aghamirzaie
et al. 2013; http://www.medicagohapmap.org/?genome).
There is another success story for candidate gene discovery in faba bean. An ortholog of
CEN/TFL1-like genes was recognized to be responsible for determinate growth habit in faba
bean, and this was confirmed in several plant species and later used to develop a CAPs
marker for use in MAS (Avila et al. 2007).
4.3 Mendelian traits
During population development, we noticed that parental lines used for population
development also differed in several Mendelian traits, such as seed coat, hilum and flower
colour, and stipule spot pigmentation (SSP). These characters were recorded during
population development (F1 to F5) as well as in backcrosses, their heritability was
investigated and they were mapped along with SNPs. The most detailed findings were on
genetic control of SSP by using four segregating populations in the F1 to F5 generations
(Mélodie/2 × ILB 938/2, Mélodie/2 × Disco/2, ILB 938/2 × Aurora/2 and ILB 938/2 ×
Disco/2) as well as reciprocal crosses. The results revealed that two unlinked genes, ssp1 and
ssp2, determined this trait. A novel locus, ssp1, decoupling pigmentation in the flower from
that in the extra-floral nectarines in ILB 938/2 was detected and mapped onto chromosome I
along with hilum colour using the Mélodie/2 × ILB 938/2 population (publication IV).
Another gene, ssp2 (zt2) in Disco/2 was already mapped in this species (Gutierrez et al.
2008). In addition, the SSP as an early-expressed morphological marker confirmed hybridity
in the crosses, so this trait can be recommended for use as a simple marker in programs
focused on low tannin or vicine-convicine along with white flower and colourless hilum.
Since ssp1 is recessive, but rare (it was present in 6% of the 402 faba bean accessions), it
could be used in the long run if a sufficient number of parents carry it. Another practical
application might be to use the ssp1 gene as a co-marker for visual evidence of outcrossing
during pedigreed seed production if a particular cultivar carries ssp1 as a homozygote. If the
same cultivar were homozygous recessive for a trait that was much more difficult to track,
for example, low vicine-convicine, the appearance of nectary spots would be a simple co-
marker in many situations. If this were desirable, a closely linked molecular marker could be
used to design crosses appropriately for pyramiding because heterozygotes and homozygotes
would be distinguishable. The ssp1 locus was tightly linked into a well conserved region of
M. truncatula chromosome VI with some candidate Myb and bHLH (basic helix-loop-helix)
transcription factor families that have been shown to regulate the anthocyanin pigmentation

39
pathway in many species (e.g., Quattrocchio et al. 1998; Tai et al. 2013). Green seed coat
colour was mapped on faba bean chromosome IV (publication III), while black seed coat
colour was shown to be on faba bean chromosome II (Sjödin 1971; Díaz-Ruiz et al. 2010;
Cruz-Izquierdo et al. 2012).

40
5 CONCLUSION AND FUTURE PROSPECTS
This study has combined the novel use of FIGS for identifying sources of abiotic stress
resources and synteny with M. truncatula for gene mapping and discovery in faba bean. An
appropriate phenotyping method, mapping population and suitable marker system are the
key requirement for a successful molecular breeding program. The differences between the
two FIGS sets (drought-adapted and non-drought-adapted) allowed demonstration of a
reliable and cost-efficient phenotyping tool for screening traits related to drought adaptation
(i.e. canopy temperatures). In the next step, using a mapping population with wide variation
at both DNA sequence and phenotypic level and using M. truncatula-derived SNPs, all
brought success in detecting the genetic regions and even candidate genes (e.g., RLKs) for
studied traits with potential for further functional genomics analysis. RLKs have already
been widely shown to be related to abiotic stress responses in model plants as well as in
several crop species. These results confirmed that genetic information from model species
can be easily translated to faba bean as a “genome orphan”.
When traits other than yield associated with drought adaptation are considered for
mapping, poor phenotyping can often cause failures (Varshney et al. 2011) and hence
generate confusing results. A fast and reliable phenotyping method is needed for the
particular crop in the specific drought profile. Significant correlations allow utilizing simpler
traits as indirect selection criteria for yield and drought adaptation. In this species, leaf
temperatures seems a reliable phenotyping method but it is highly recommended that a
subset of samples be subjected to a more accurate screening method such as Δ13
C. The most
severe droughts mostly occur in poor countries with little access to expensive equipment
such as sensor technologies or even Δ13
C, so economical tools like IRT are exceptionally
important.
Our findings support the assertion that FIGS can be an effective tool in pre-breeding
programmes for traits related to drought adaption in faba bean, and this may be extruded to
other traits related to abiotic stresses and adaptation to climate change. The core-collection
approach asks "what is the range of variation in the species" while the FIGS approach asks
"where can we find the extremes of variation in this trait in the species". Thus FIGS is
complementary to core collections. The major demand for germplasm collections from
agricultural genebanks is mostly for adaptive traits (Kenneth Street personal
communication), so in contrast to core collections, FIGS represents a dynamic, direct and
practical approach that focuses on the sought-after trait rather than on generalized measures
of genetic diversity. Overall, CGIAR suggests the FIGS approach, alongside current core
collection activities, should be part of breeding strategies. Several FIGS sets developed by
ICARDA are available to speed up research on climate change adaptive traits
(http://www.icarda.org/figs-%E2%80%93-new-approach-mining-agricultural-genebanks-
speed-pace-innovation-and-food-security).
This work was carried out under climate-controlled greenhouse conditions and these
findings should be confirmed under field conditions. When preparing to introduce detected
putative QTLs as MAS tools, they should be validated in different environments. The
identification of candidate genes takes away some of the apparent complexity of drought
adaptation in faba bean. Co-localisation of these genes with known QTLs involved in
drought adaptation may help select regulatory genes involved in this respect and to define a
rational approach to developing drought-adapted cultivars. Candidate genes also can be

41
transferred to elite lines of faba bean by MAS or genetic transformation. Limited progress
has been shown on gene transfer or pyramiding using MAS in legumes, but some progress
has been made in soya bean, pea and common bean (Dwivedi et al. 2007).
Research on the root, the hidden part of plant, lags behind that on shoots. Physiologists
and breeders are becoming positioned to breed crops with better rooting systems that
improve productivity under water-limited conditions. Preliminary results on root
phenotyping showed that drought-adapted accessions had relatively better rooting systems
(branching, density and architect) than the wet adapted set, but root length did not differ.
More research needs to be conducted on root morphology and function and its relation to
shoot characteristics in this species, since drought adaptation in rice, for example, was
associated with a more extensive root system as well as reduced stomatal density and
improved WUE (Yu et al. 2013).

42
ACKNOWLEDGMENTS
I am very lucky to have Dr. Frederick L. Stoddard as my main supervisor. His profound
knowledge in crop science, breeding, genetics and statistics along with his kind support,
guidance and encouragements culminated this work. I am very grateful to my co-supervisor
Professor Mikko J. Sillanpää for his advices and supports during molecular biology work.
Many thanks to Professor Pirjo Mäkelä leader of crop science group, for her support for my
PhD course management, thesis review and pre-reviewer suggestions and also Dr. Arja
Santanen for her advice and practical help during lab work. My PhD monitoring team,
Professor Teemu Teeri and Dr. Mervi Seppänen are acknowledged for their valuable
comments and suggestions. I would like to thank Dr. Petr Smýkal and Dr. Simo Hovinen,
pre-reviewers of the thesis for their constructive comments.
Many interesting ideas to develop the current work came from our international
collaboration for developing the FIGS sets by Dr. Kenneth Street, Dr. Abdallah Bari
(ICARDA) and Dr. Michael Mackay (The University of Queensland, Australia), and for
developing the SNPs platform by Professor Donal M. O’Sullivan (University of Reading,
UK). I wish to thank Professor Wolfgang Link (Georg-August-University, Germany) for
providing the seeds of the parental lines for population development.
I owe great gratitude to Markku Tykkyläinen, Sanna M. Peltola, Sini Lindström and
Daniel Richterich, technical staff of the glasshouses of the University of Helsinki, for their
kind assistance during the experiments.
I would like also to acknowledge Damian Wach, Guillermo Mínguez Vélaz, Sandro
Maria Cavaleri, and Stefano Zanotto, visiting students, and Yao Lu, MSc student of this
university, for their for their kind assistance at different stages of this project. Many thanks
to all friends and colleagues at the crop science group, HY.
This work was financially supported by CIMO (the Centre for International Mobility),
the Emil Aaltonen Foundation (Emil Aaltosen Säätiö), Niemi Säätiö, Department of
Agricultural Sciences, University of Helsinki and the Doctoral Program in Plant Science.
Part of the research reported here was conducted in conjunction with "Legume Futures
(Legume-supported cropping systems for Europe)", a collaborative research project funding
from the European Union's Seventh Programme for research, technological development and
demonstration under grant agreement No. 245216.
I give special thanks to my wife, Aida for being patient and kind during the work and
thesis writing, and to my parents for their encouragements during my studies which led me
to the doctoral degree.
Helsinki, September 2014
Hamid Khazaei

43
6 REFERENCES
Abdelmula AA, Link W, Kittlitz EV, Stelling D (1999) Heterosis and inheritance of drought
tolerance in faba bean, Vicia faba L. Plant Breed 118:485–490
Aghamirzaie D, Nabiyouni M, Fang Y, Klumas C, Heath LS, Grene R, Collakova E (2013)
Changes in RNA splicing in developing soybean (Glycine max) embryos. Biology 2:1311–
1337
Akash M, Myers GO (2012) The development of faba bean expressed sequence tag-simple
sequence repeats (EST-SSRs) and their validity in diversity analysis. Plant Breed 131:522–
530
Alghamdi SS, Al-Faifi SA, Migdadi HM, Ammar MH, Siddique K (2011) Inter-simple sequence
repeat (ISSR)-based diversity assessment among faba bean genotypes. Crop Pasture Sci
62:755–760
Alghamdi SS, Migdadi HM, Ammar MH, Paull JG, Siddique KHM (2012) Faba bean genomics:
current status and future prospects. Euphytica 186:609–624
Amede T, von Kittlitz E, Schubert S (1999) Differential drought responses of faba bean (Vicia
faba L.) inbred lines. J Agron Crop Sci 183:35–45
Annicchiarico P, Iannucci A (2008) Breeding strategy for faba bean in Southern Europe based on
cultivar responses across climatically contrasting environments. Crop Sci 48:983–991
Anyia AO, Herzog H (2004) Water-use efficiency, leaf area and leaf gas exchange of cowpeas
under midseason drought. Eur J Agron 20:327–339
Araus JL, Slafer GA, Reynolds MP, Royo C (2002) Plant breeding and drought in C3 cereals:
what should we breed for? Ann Bot 89:925–940
Araus JL, Slafer GA, Reynolds MP, Royo C (2004) Physiology of yield and adaptation in wheat
and barley breeding. In: Nguyen HT, Blum A (eds) Physiology and biotechnology
integration for plant breeding. Marcel Dekker, New York, USA. pp 1–49
Arbaoui M, Balko C, Link W (2008a) Study of faba bean (Vicia faba L.) winter-hardiness and
development of screening methods. Field Crops Res 106:60–67
Arbaoui M, Link W, Satovic Z, Torres AM (2008b) Quantitative trait loci of frost tolerance and
physiologically related traits in faba bean (Vicia faba L.). Euphytica 164:93–104
Ashraf MY, Iram A (2005) Drought stress induced changes in some organic substances in
nodules and other plant parts of two potential legumes differing in salt tolerance. Flora
200:535–546
Avila CM, Sillero JC, Rubiales D, Moreno MT, Torres AM (2003) Identification of RAPD
markers linked to Uvf-1 gene conferring hypersensitive resistance against rust (Uromyces
viciae-fabae) in Vicia faba L. Theor Appl Genet 107:353–358
Avila CM, Satovic Z, Sillero JC, Rubiales D, Moreno MT, Torres AM (2004) Isolate and organ-
specific QTLs for ascochyta blight resistance in faba bean. Theor Appl Genet 108:1071–
1078
Avila CM, Satovic Z, Sillero JC, Rubiales D, Moreno MT, Torres AM (2005) QTL detection for
agronomic traits in faba bean (Vicia faba L.). Agric Conspec Sci 70:65–73

44
Avila CM, Nadal S, Moreno MT, Torres AM (2006) Development of a simple PCR-based
marker for the determination of growth habit in Vicia faba L. using a candidate gene
approach. Mol Breed 17:185–190
Avila CM, Atienza SG, Moreno MT, Torres AM (2007) Development of a new diagnostic
marker for growth habit selection in faba bean (Vicia faba L.) breeding. Theor Appl Genet
115:1075–1082
Bari A, Street K, Mackay M, Endresen DTF, De Pauw E, Amri A (2012) Focused identification
of germplasm strategy (FIGS) detects wheat stem rust resistance linked to environmental
variables. Genet Resour Crop Evol 59:1465–1481
Bari A, Khazaei H, Street K, Stoddard FL, Chaubey YP, Dayanandan S, De Pauw E, Ouabbou H,
Maatougui M, Jilal A, Nachit M (2014a) Assessing plant genetic resources for climate
change adaptive traits. In: Præbel A, Groeneveld LF (eds) Genetic resources for food and
agriculture in a changing climate. January 27–29, 2014. Lillehammer, Norway
Bari A, Amri A, Street K, Mackay M, De Pauw E, Sanders R, Nazari K, Humeid B, Konopka J,
Alo F (2014b) Predicting resistance to stripe (yellow) rust (Puccinia striiformis) in wheat
genetic resources using focused identification of germplasm strategy. J Agr Sci (in press)
doi:10.1017/S0021859613000543
Bari A, Khazaei H, Street K, Sillanpää MJ, Endresen DTF, De Pauw E, Chaubey YP,
Dayanandan S, Stoddard FL, Damania AD (2014c) In silico evaluation of plant genetic
resources to search for traits for adaptation to climate change. (Submitted manuscript)
Barrs HD, Weatherley PE (1962) A re-examination of the relative turgidity technique for
estimating water deficit in leaves. Aust J Biol Sci 15:413–428
Basu PS, Berger JD, Turner NC, Chaturvedi SK, Ali M, Siddique KHM (2007) Osmotic
adjustment of chickpea (Cicer arietinum) is not associated with changes in carbohydrate
composition or leaf gas exchange under drought. Ann Appl Biol 150:217–225
Bayuelo-Jiménez JS, Debouck DG, Lynch JP (2002) Salinity tolerance in Phaseolus species
during early vegetative growth. Crop Sci 42:2184–2192
Benson DA, Cavanaugh M, Clark K, Karsch-Mizrachi I, Lipman DJ, Ostell J, Sayers EW (2013)
GeneBank. Nucl Acids Res 41(D1):D36–D42
Bhullar NK, Street K, Mackay M, Yahiaoui N, Keller B (2009) Unlocking wheat genetic
resources for the molecular identification of previously undescribed functional alleles at the
Pm3 resistance locus. Proc Natl Acad Sci USA 106:9519–9524
Blum A (2009) Effective use of water (EUW) and not water-use efficiency (WUE) is the target
of crop yield improvement under drought stress. Field Crops Res 112:119–123
Blum A (2011a) Plant breeding for water limited environments. Springer-Verlag, New York. 252
p
Blum A (2011b) Drought resistance – is it really a complex trait? Funct Plant Biol 38:753–757
Blum A, Mayer J, Gozlan G (1982) Infrared thermal sensing of plant canopies as a screening
technique for dehydration avoidance in wheat. Field Crops Res 5:137–146
Bond DA (1989a) Prospects for commercialisation of F1 hybrid field beans Vicia faba L.
Euphytica 41:81–86

45
Bond DA (1989b) A short review of research on male sterility and prospects for F1 hybrid
varieties in field beans (Vicia faba L.). Euphytica 41:87–90
Bond DA, Poulsen MH (1983) Pollination. In: Hebblethwaite PD (ed) The faba bean (Vicia faba
L.). Butterworths, London, UK. pp 77–101
Bond DA, Jellis GJ, Rowland GG, Le Guen J, Robertson LD, Khalil SA, Li-Juan L (1994)
Present status and future strategy in breeding faba beans (Vicia faba L.) for resistance to
biotic and abiotic stresses. Euphytica 73:151–166
Boopathi NM (2013) Genetic mapping and marker assisted selection: Basics, practice and
benefits. Springer India
Bordat A, Savois V, Nicolas M, Salse J, Chauveau A, Bourgeois M, Potier J, Houtin H, Rond C,
Murat F, Marget P, Aubert G, Burstin J (2011) Translational genomics in legumes allowed
placing in silico 5460 unigenes on the pea functional map and identified candidate genes in
Pisum sativum L. Genes Genome Genet 1:93–103
Broman KW (2006) The genomes of recombinant inbred lines. Genetics 169:1133–1146
Buhariwalla HK, Jayashree B, Crouch JH (2005) Development of ESTs from chickpea roots and
their use in diversity analysis of the Cicer genus. BMC Plant Biol 5:16
Casson S, Gray JE (2008) Influence of environmental factors on stomatal development. New
Phytol 178:9–23
Chaves MM, Maroco JP, Pereira JS (2003) Understanding plant responses to drought – from
genes to the whole plant. Funct Plant Biol 30:239–264
Cheema J, Dicks J (2009) Computational approaches and software tools for genetic linkage map
estimation in plants. Brief Bioinform 10:595–608
Choi HK, Kim DJ, Uhm T, Limpens E, Baek JM, Lim H, Kalo P, Penmetsa RV, Seres A,
Kulikova O, Bisseling T, Kiss GB, Cook DR (2004) A sequence based genetic map of
Medicago truncatula and comparison of marker colinearity with Medicago sativa. Genetics
166:1463–1502
Collard BCY, Mackill, DJ (2008) Marker-assisted selection: an approach for precision plant
breeding in the twenty-first century. Philos Trans R Soc Lond B Biol Sci 363:557–572
Collard BCY, Jahufer MZZ, Brouwer JB, Pang ECK (2005) An introduction to markers,
quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement:
the basic concepts. Euphytica 142:169–196
Condon AG, Richards RA, Rebetzke GJ, Farquhar GD (2004) Breeding for high water-use
efficiency. J Exp Bot 55:2447–2460
Cottage A, Gostkiewicz K, Thomas JE, Borrows R, Torres AM, O’Sullivan DM (2012a)
Heterozygosity and diversity analysis using mapped SNPs in a faba bean inbreeding
programme. Mol Breed 30:1799–1809
Cottage A, Webb A, Hobbs D, Khamassi K, Maalouf F, Ogbannaya F, Stoddard FL, Duc G, Link
W, Thomas JE, O’Sullivan DM (2012b) SNP discovery and validation for genomic-assisted
breeding of faba bean (Vicia faba L.). In: VI International conference on legume genetics
and genomics (ICLGG). October 2–7, 2012. Hyderabad, India
Crépon K, Marget P, Peyronnet C, Carrouée B, Arese P, Duc G (2010) Nutritional value of faba
bean (Vicia faba L.) seeds for feed and food. Field Crops Res 115:329–339

46
Cruz-Izquierdo S, Avila CM, Satovic Z, Palomino C, Gutierrez N, Ellwood SR, Phan HTT,
Cubero JI, Torres AM (2012) Comparative genomics to bridge Vicia faba with model and
closely-related legume species: stability of QTLs for flowering and yield-related traits. Theor
Appl Genet 125:1767–1782
Cubero J (1974) On the evolution of Vicia faba L. Theor Appl Genet 45:47–51
Dai A (2013) Increasing drought under global warming in observations and models. Nat Clim
Change 3:52–58
Darwish DS, Fahmy GM (1997) Transpiration decline curves and stomatal characteristics of faba
bean genotypes. Biol Plant 39:243–249
Dayanandan S, Bari A, Chaubey YP, Khazaei H, Street K, Stoddard FL, Mackay M, Matthews
D, Endresen DTF, De Pauw E, Kehel Z (2013) Searching for climate change adaptive traits
in crops to sustain agriculture: applied mathematics, phenomics and genomics in dryland
systems. In: 2013 QCBS Symposium. 11-13 December 2013. Montreal, Canada.
http://colloque.qcbs.ca/speaker/s.dayanandan#.Uu1rq_uWwvl
De Micco V, Aronne G (2012) Morpho-anatomical traits for plant adaptation to drought. In:
Aroca R (ed) Plant responses to drought stress. Berlin, Springer-Verlag. pp 37–61
Díaz-Ruiz R, Satovic Z, Avila CM, Alfaro CM, Gutierrez MV, Torres AM, Román B (2009a)
Confirmation of QTLs controlling Ascochyta fabae resistance in different generations of faba
bean (Vicia faba L.). Crop Pasture Sci 60:353–361
Díaz-Ruiz R, Torres A, Gutierrez MV, Rubiales D, Cubero JI, Kharrat M, Satovic Z, Román B
(2009b) Mapping of quantitative trait loci controlling Orobanche foetida Poir. resistance in
faba bean (Vicia faba L.). Afr J Biotechnol 8:2718–2724
Díaz-Ruiz R, Torres AM, Satovic Z, Gutierrez MV, Cubero JI, Román B (2010) Validation of
QTLs for Orobanche crenata resistance in faba bean (Vicia faba L.) across environments
and generations. Theor Appl Genet 120:909–919
Dieckmann S, Link W (2010) Quantitative genetic analysis of embryo heterosis in faba bean
(Vicia faba L.). Theor Appl Genet 120:261–270
Duc G (1997) Faba bean (Vicia faba L.). Field Crops Res 53:99–109
Duc G, Bao S, Baumc M, Redden B, Sadiki M, Suso MJ, Vishniakova M, Zong X (2010)
Diversity maintenance and use of Vicia faba L. genetic resources. Field Crops Res 115:270–
278
Dwivedi SL, Crouch JH, Mackill DJ, Xu Y, Blair MW, Ragot M, Upadhyaya HD, Ortiz R (2007)
The molecularization of public sector crop breeding: Progress, problems, and prospects.
Adv Agron 95:163–318
Ellwood SR, Phan HTT, Jordan M, Torres AM, Avila CM, Cruz-Izquierdo S, Oliver RP (2008)
Construction of a comparative genetic map in faba bean (Vicia faba L.); conservation of
genome structure with Lens culinaris. BMC Genom 9:380
Endresen DTF, Street K, Mackay M, Bari A, De Pauw E (2011) Predictive association between
biotic stress traits and ecogeographic data for wheat and barley landraces. Crop Sci
51:2036–2055
Eurostat (2014) European statistics. European commission. http://epp.eurostat.ec.europa.eu

47
FAO (2010) The second report on the state of the world’s plant genetic resources for food and
agriculture. Rome, Italy. 398 p
FAO (2014) FAOSTAT. Food and agriculture organisation of the United Nations
http://faostat.fao.org/faostat
Fischer RA, Turner NC (1978) Plant productivity in the arid and semiarid zone. Annu Rev Plant
Physiol 29:277–317
Fischer RA, Rees D, Sayre KD, Lu ZM, Condon AG, Saavedra AL (1998) Wheat yield progress
associated with higher stomatal conductance and photosynthetic rate, and cooler canopies.
Crop Sci 38:1467–1475
Flores F, Hybl M, Knudsen JC, Marget P, Muel F, Nadal S, Narits L, Raffiot B, Sass O, Solis I,
Winkler J, Stoddard FL, Rubiales D (2013) Adaptation of spring faba bean types across
European climates. Field Crops Res 145:1–9
Frankel OH (1984) Genetic perspectives of germplasm conservation. In: Arber W, Llimensee K,
Peacock WJ, Starlinger P (eds) Genetic manipulation: Impact on man and society.
Cambridge University Press, Cambridge, UK. pp 161–170
Franks PJ, Farquhar GD (2007) The mechanical diversity of stomata and its significance in gas
exchange control. Plant Physiol 146:729–736
Franssen S, Shrestha RP, Bräutigam A, Bornberg-Bauer E, Weber APM (2011) Comprehensive
transcriptome analysis of the highly complex Pisum sativum genome using next generation
sequencing. BMC Genom 12:227
Galeano CH, Cortés AJ, Fernández AC, Soler A, Franco-Herrera N, Makunde G, Vanderleyden
J, Blair MW (2012) Gene-based single nucleotide polymorphism markers for genetic and
association mapping in common bean. BMC Genet 13:48
Gálvez L, González EM, Arrese-Igor C (2005) Evidence for carbon flux shortage and strong
carbon/nitrogen interactions in pea nodules at early stages of water stress. J Exp Bot
56:2551–2561
Garg R, Patel RK, Tyagi AK, Jain M (2011) De novo assembly of chickpea transcriptome using
short reads for gene discovery and marker identification. DNA Res 18:53–63
Gepts P (2006) Plant genetic resources conservation and utilization: the accomplishments and
future of a societal insurance policy. Crop Sci 46:2278–2292
Gnanasambandam A, Paull J, Torres A, Kaur S, Leonforte T, Li H, Zong X, Yang T, Materne M
(2012) Impact of molecular technologies on faba bean (Vicia faba L.) breeding strategies.
Agronomy 2:132–166
Gong Y-M, Xu S-C, Mao W-H, Hu Q-Z, Zhang G-W, Ding J, Li Z-Y (2010) Generation and
characterization of 11 novel EST derived microsatellites from Vicia faba (Fabaceae). Am J
Bot 97:e69–e71
González EM, Gordon AJ, James CL, Arrese-Igor C (1995) The role of sucrose synthase in the
response of soybean nodules to drought. J Exp Bot 46:1515–1523
Gornall J, Betts R, Burke E, Clark R, Camp J, Willett K, Wiltshire A (2010) Implications of
climate change for agricultural productivity in the early twenty-first century. Philos Trans
Royal Soc B Biol Sci 365:2973–2989

48
Granier C, Aguirrezabal L, Chenu K, Cookson SJ, Dauzat M, Hamard P, Thioux JJ, Rolland G,
Bouchier-Combaud S, Lebaudy A, Muller B, Simonneau T, Tardieu F (2006) PHENOPSIS,
an automated platform for reproducible phenotyping of plant responses to soil water deficit
in Arabidopsis thaliana permitted the identification of an accession with low sensitivity to
soil water deficit. New Phytol 169:623–635
Gresta F, Avola G, Albertini E, Raggi L, Abbate V (2010) A study of variability in the Sicilian
faba bean landrace ‘Larga di Leonforte’. Genet Resour Crop Evol 57:523–531
Grzesiak S, Iijima M, Kono Y, Yamauchi A (1997) Differences in drought tolerance between
cultivars of field bean and field pea. A comparison of droughtresistant and drought-sensitive
cultivars. Acta Physiol Plant 19:349–357
Gutierrez N, Avila CM, Duc G, Marget P, Suso MJ, Moreno MT, Torres AM (2006) CAPs
markers to assist selection for low vicine and convicine content in faba bean (Vicia faba L.).
Theor Appl Genet 114:59–66
Gutierrez N, Avila CM, Rodriguez-Suarez, Moreno MT, Torres AM (2007) Development of
SCAR markers linked to a gene controlling absence of tannins in faba bean. Mol Breed
19:305–314
Gutierrez N, Avila CM, Moreno MT, Torres AM (2008) Development of SCAR markers linked
to zt-2, one of the genes controlling absence of tannins in faba bean. Aust J Agri Res 59:62–
68
Gutiérrez N, Palomino C, Satovic Z, Ruiz-Rodríguez MD, Vitale S, Gutiérrez MV, Rubiales D,
Kharrat M, Amri M, Emeran A, Cubero JI, Atienza SG, Torres AM, Avila CM (2013) QTLs
for Orobanche spp. resistance in faba bean: Identification and validation across different
environments. Mol Breed 23:909–922
Hall AE (2012) Phenotyping cowpeas for adaptation to drought. Front Physiol 3:155
Handberg K, Stougaard J (1992) Lotus japonicus, an autogamous, diploid legume species for
classical and molecular genetics. Plant J 2:487–496
Hanelt P (1972) Die infraspezitische variabilitaet von Vicia faba und ihre Gliederung.
Kulturpflanze 20:75–128
Heringa RJ, Norel A, Post J (1984) Searching for tolerance to drought in Vicia faba. FABIS
Newslett 8:13–14
Hiremath P, Farmer A, Cannon SB, Woodward J, Kudapa H, Tuteja R, Kumar A, Bhanuprakash
A, Mulaosmanovic B, Gujaria N, Krishnamurthy L, Gaur PM, Kavikishor PB, Shah T,
Srinivasan R, Lohse M, Xiao Y, Town CD, Cook DR, May GD, Varshney RK (2011)
Large-scale transcriptome analysis in chickpea (Cicer arietinum L.), an orphan legume crop
of the semi-arid tropics of Asia and Africa. Plant Biotechnol J 9:922–931
Holman TJ, Jones PD, Russell L, Medhurst A, Ubeda Toma´s S, Talloji P, Marquez J, Schmuths
H, Tung SA, Taylor I, Footitt S, Bachmair A, Theodoulou FL, Holdsworth MJ (2009) The
N-end rule pathway promotes seed germination and establishment through removal of ABA
sensitivity in Arabidopsis. Proc Natl Acad Sci USA 106:13618–13623
Huang J, Pray C, Rozelle S (2002) Enhancing the crops to feed the poor. Nature 418:678–684
Iannucci A, Terribile MR, Martiniello P (2008) Effects of temperature and photoperiod on
flowering time of forage legumes in a Mediterranean environment. Field Crops Res
106:156–162

49
Jayashree B, Buhariwalla HK, Shinde S, Kumar PV, Crouch JH (2005) A legume genomics
resource: the chickpea root expressed sequence tag database and bioinformatics tools.
Electron J Biotechnol 8:128–133
Jhanwar S, Priya P, Garg R, Parida SK, Tyagi AK, Jain M (2012) Transcriptome sequencing of
wild chickpea as a rich resource for marker development. Plant Biotechnol J 10:690–702
Johnson RC (2008) Gene banks pay big dividends to agriculture, the environment, and human
welfare. PLoS Biol 6:e148
Kalo P, Seres A, Taylor SA, Jakab J, Kevei Z, Kereszt A, Endre G, Ellis TH, Kiss GB (2004)
Comparative mapping between Medicago sativa and Pisum sativum. Mol Genet Genom
272:235–246
Kalo P, Choi HK, Ellis N, Kiss GB (2011) Synteny and comparative genomics between model
and cool season grain legumes. In: Kole C (series editor), P rez de la Vega M, Torres AM,
Cubero JI, Kole C (eds) Genetics, genomics and breeding in crop plants: cool season food
legumes. Science Pubs Inc, New Hampshire. pp 285–302
Karaköy T, Baloch FS, Toklu F, Özkan H (2014) Variation for selected morphological and
quality-related traits among 178 faba bean landraces collected from Turkey. Plant Genet
Resour12:5–13
Kashiwagi J, Krishnamurthy L, Crouch JH, Serraj (2006) Variability of root length density and
its contributions to seed yield in chickpea (Cicer arietinum L.) under terminal drought
stress. Field Crops Res 95:171–181
Kashiwagi J, Krishnamurthy L, Gaur PM, Chandra S, Upadhyaya HD (2008a) Estimation of
gene effects of the drought avoidance root characteristics in chickpea (C. arietinum L.).
Field Crops Res 105:64–69
Kashiwagi J, Krishnamurthy L, Upadhyaya HD, Gaur PM (2008b) Rapid screening technique for
canopy temperature status and its relevance to drought tolerance improvement in chickpea. J
SAT Agri Res 6:1–4
Kaur N, Street K, Mackay M, Yahiaoui N, Keller B (2008) Molecular approaches for
characterization and use of natural disease resistance in wheat. Eur J Plant Pathol 121:387–
397
Kaur S, Cogan NOI, Pembleton LW, Shinozuka M, Savin KW, Materne M, Forster JW (2011)
Transcriptome sequencing of lentil based on second-generation technology permits large-
scale unigene assembly and SSR marker discovery. BMC Genom 12:265
Kaur S, Pembleton L, Cogan NOI, Savin K, Leonforte T, Paull J, Materne M, Forster J (2012)
Transcriptome sequencing of field pea and faba bean for discovery and validation of SSR
genetic markers. BMC Genom 13:104
Kaur S, Cogan NOI, Forster JW, Paull JG (2014a) Assessment of genetic diversity in faba bean
based on single nucleotide polymorphism. Diversity 6:88–101
Kaur S, Kimber RBE, Cogan NOI, Materne M, Forster JW, Paull J (2014b) SNP discovery and
high-density genetic mapping in faba bean (Vicia faba L.) permits identification of QTLs for
ascochyta blight resistance. Plant Sci 217-218:47–55
Keneni G, Jarso M, Wolabu T, Dino G (2005) Extent and pattern of genetic diversity for
morpho-agronomic traits in Ethiopian highland pulse landraces. II. Faba bean (Vicia faba
L.). Genet Resour Crop Evol 52:551–561

50
Kew (2014) Leguminosae. Royal Botanic Gardens, Kew, UK. http://www.kew.org/science-
research-data/directory/teams/leguminosae/index.htm. Accessed 3 March 2014.
Khamassi K, Jeddi FB, Hobbs D, Irigoyen J, Stoddard FL, O'Sullivan DM, Jones H (2013) A
baseline study of vicine–convicine levels in faba bean (Vicia faba L.) germplasm. Plant
Genet Resour 11:250–257
Khan HR, Link W, Hocking TJH, Stoddard FL (2007) Evaluation of physiological traits for
improving drought tolerance in faba bean (Vicia faba L.). Plant Soil 292:205–217
Khan HR, Paull JG, Siddique KHM, Stoddard FL (2010) Faba bean breeding for drought-
affected environments: A physiological and agronomic perspective. Field Crops Res
115:279–286
Khazaei H, Stoddard FL, Santanen A (2010) Leaf traits associated with drought response in faba
bean (Vicia faba L.). In: Legumes for global health: legume crops and products for food,
feed and environmental benefits. April 26–30, 2010. Antalya, Turkey
Khazaei H, Stoddard FL, Lizarazo C, Street K (2012) New sources of earliness for Finnish faba
bean breeding. In: Maataloustieteen päivät 2012.
http://www.smts.fi/Kasvintuotanto_jalostuu/Khazaei_New%20sources.pdf
Kilian B, Graner A (2012) NGS technologies for analysing germplasm diversity in genebanks.
Brief Funct Genom 11:38–50
Kim KS, Park SH, Kim DK, Jenks MA (2007) Influence of water deficit on leaf cuticular waxes
of soybean (Glycine max). Int J Plant Sci 168:307–316
Kosambi DD (1943) The estimation of map distance from recombination values. Ann Eugen
12:172–175
Kosma DK, Bourdenx B, Bernard A, Parsons EP, Lü S, Joubès J, Jenks MA (2009) The impact
of water deficiency on leaf cuticle lipids of Arabidopsis. Plant Physiol 151:1918–1929
Kramer PJ, Boyer JS (1995) Water relations of plants and soils. Academic Press, New York,
USA
Krishnamurthy L, Kashiwagi J, Tobita S, Ito O, Upadhyaya HD, Gowda CLL, Gaur PM,
Sheshshayee MS, Singh S, Vadez V, Varshney RK (2013) Variation in carbon isotope
discrimination and its relationship with harvest index in the reference collection of chickpea
germplasm. Funct Plant Biol 40:1350–1361
Kumar J, Basu PS, Srivastava E, Chaturvedi SK, Nadarajan N, Kumar S (2005) Phenotyping of
traits imparting drought tolerance in lentil. Crop Pasture Sci 63:547–554
Kumar J, Choudhary AK, Solanki RK, Pratap A (2011) Towards marker-assisted selection in
pulses: a review. Plant Breed 130:297–313
Kwon S, Hu J, Coyne CJ (2010) Genetic diversity and relationship among faba bean (Vicia faba
L.) germplasm entries as revealed by TRAP markers. Plant Genet Resour 8:204–213
Lawes DA, Bond DA, Poulsen MH (1983) Classification, origin, breeding methods and
objectives. In: Hebblethwaite PD (ed) The faba bean (Vicia faba L.). Butterworths, London,
UK. pp 23–76
Lewis G, Schrire B, Mackinder B, Lock M (2005) Legumes of the world. Royal Botanic
Gardens, Kew Publishing, Richmond, UK. 577 p

51
Lang L-J, Yu Z-H, Zheng Z-J, Xu M-S, Ying H-Q (1993) Study and utilization of faba bean
germplasm resources. In: Sears LJ (ed) Faba bean in China: State-of-the-art review.
ICARDA Press. pp 51–63
Link W, Dixkens C, Singh M, Schwall M, Melchinger AE (1995) Genetic diversity in European
and Mediterranean faba bean germ plasm revealed by RAPD markers. Theor Appl Genet
90:27–32
Link W, Schill B, Kittlitz EV (1996) Breeding for wide adaptation in faba bean. Euphytica
92:185–190
Link W, Ederer W, Gumber RK, Melchinger AE (1997) Detection and characterization of two
new CMS systems in faba bean (Vicia faba). Plant Breed 116:158–162
Link W, Abdelmula AA, Kittlitz EV, Bruns S, Riemer H, Stelling D (1999) Genotypic variation
for drought tolerance in Vicia faba. Plant Breed 118:477–483
Link W, Balko C, Stoddard FL (2010) Winter hardiness in faba bean: Physiology and breeding.
Field Crops Res 115:287–296
Lorieux M (2012) MapDisto: fast and efficient computation of genetic linkage map. Mol Breed
30:1231–1235
Lorieux M, Goffinet B, Perrier X, González de León D, Lanaud C (1995a) Maximum likelihood
models for mapping genetic markers showing segregation distortion. 1. Backcross
populations. Theor Appl Genet 90:73–80
Lorieux M, Perrier X, Goffinet B, Lanaud C, González de León D (1995b) Maximum likelihood
models for mapping genetic markers showing segregation distortion. 2. F2 populations.
Theor Appl Genet 90:81–89
Loss SP, Siddique KHM, Tennant D (1997) Adaptation of faba bean (Vicia faba L.) to dryland
Mediterranean-type environments. III. Water use and water-use efficiency. Field Crops Res
54:153–162
Ludlow MM, Muchow RC (1990) A critical evaluation of traits for improving crop yield in
water-limited environments. Adv Agron 43:107–153
Ma Y, Yang T, Guan J, Wang S, Wang H, Sun X, Zong X (2010) Development and
characterization of 21 EST-derived microsatellite markers in Vicia faba (fava bean). Am J
Bot 98:e22–e24
Ma Y, Bao S, Yang T, Hu J, Guan J, He Y, Wang X, Wan Y, Sun X, Jiang J, Gong C, Zong X
(2013) Genetic linkage map of Chinese native variety faba bean (Vicia faba L.) based on
simple sequence repeat markers. Plant Breed 132:397–400
Mackay M, Street K (2004) Focused identification of germplasm strategy—FIGS. In: Black CK,
Panozzo JF, Rebetzke GJ (eds) Proceedings of the 54th Australian cereal chemistry
conference and the 11th wheat breeders assembly. September 21–24, 2004, Canberra, ACT,
Australia. Cereal chemistry division, Royal Australian Chemical Institute (RACI),
Melbourne, Victoria, Australia. pp 138–141
Madrid E, Palomino C, Jüngling R, Frank A, Horres R, Rotter B, Winter P, Kahl G, Torres AM
(2013) DeepSuperSage analysis of the Vicia faba transcriptome in response to Ascochyta
fabae infection. Phytopathol Mediterr 52:166−182
Manahan CO, App AA (1973) An arginyl-transfer ribonucleic acid protein transferase from
cereal embryos. Plant Physiol 52:13–16

52
Manschadi AM, Sauerborn J, Stützel H, Göbel W, Saxena MC (1998) Simulation of faba bean
(Vicia faba L.) root system development under Mediterranean conditions. Eur J Agron
9:259–272
Marshall A, Aalen RB, Audenaert D, Beeckman T, Broadley MR, Butenko MA, Caño-Delgado
A, De Vries S, Dresselhaus T, Felix G, Graham NS, Foulkes J, Granier C, Greb T,
Grossniklaus U, Hammond JP, Heidstra R, Hodgman C, Hothorn M, Inzé D, Østergaard L,
Russinova E, Simon R, Skirycz A, Stahl Y, Zipfel C, De Smete I (2012) Tackling drought
stress: receptor-like kinase presents new approaches. Plant Cell 24:2262–2278
Maxted N (1993) A phenetic investigation of Vicia L. subgenus Vicia (Leguminosae, Vicieae).
Bot J Linn Soc 111:155–182
McDonald GK, Paulsen GM (1997) High temperature effects on photosynthesis and water
relations of grain legumes. Plant Soil 196:47–58
McKinney NV, Schapaugh WT, Kanemasu ET (1989) Selection for canopy temperature
differential in six populations of soybean. Crop Sci 29:255–259
Meinke DW, Cherry JM, Dean C, Rounsley SD, Koornneef M (1998) Arabidopsis thaliana: A
model plant for genome analysis. Science 282:662–682
Meitzel T, Radchuk R, Nunes-Nesi A, Fernie AR, Link W, Weschke W, Weber H (2011) Hybrid
embryos of Vicia faba develop enhanced sink strength, which is established during early
development. Plant J 65:517–531
Monneveux P, Jing R, Misra SC (2012) Phenotyping for drought adaptation in wheat using
physiological traits. Front Physiol 3:429
Monneveux P, Ribaut J-M, Okono A (2014) Frontiers Research Topics. Drought phenotyping in
crops: from theory to practice. Frontiers in Physiology, Lausanne, Switzerland. doi:
10.3389/978-2-88919-181-9. (Originally published in 2011). Freely available at:
http://www.generationcp.org/phenotyping-book-methods-applications
Moose SP, Mumm RH (2008) Molecular plant breeding as the foundation for 21st century crop
improvement. Plant Physiol 147:969–977
Morgan JM (1984) Osmoregulation and water stress in higher plants. Ann Rev Plant Physiol
35:299–319
Morgan JM, Rodriguezmaribona B, Knights EJ (1991) Adaptation to water-deficit in chickpea
breeding lines by osmoregulation: relationship to grain-yields in the field. Field Crops Res
27:61–70
Mortimore M, Anderson S, Cotula L, Davies J, Faccer K, Hesse C, Morton J, Nyangena W,
Skinner J, Wolfangel C (2009). Dryland Opportunities: A new paradigm for people,
ecosystems and development, IUCN, Gland, Switzerland; IIED, London, UK and
UNDP/DDC, Nairobi, Kenya. 86 p
Muratova V (1931) Common Beans (Vicia faba). Bull Appl Bot Genet Plant Breed 50:1–298
Nadal S, Suso MJ, Moreno MT (2003) Management of Vicia faba genetic resources: changes
associated to the selfing process in the major, equina and minor groups. Genet Resour Crop
Evol 50:183–192
Negruk V (2013) Mitochondrial genome sequence of the legume Vicia faba. Front Plant Sci
4:128

53
Nerkar YS, Wilson D, Lawes DA (1981) Genetic variation in stomatal characteristics and
behaviour, water use and growth of five Vicia faba L. genotypes under contrasting soil
moisture regimes. Euphytica 30:335–345
Nevo E, Korol AB, Beiles A, Fahima T (2002) Evolution of wild emmer and wheat
improvement: population genetics, genetic resources, and genome organization of wheat's
progenitor, Triticum dicoccoides. Springer-Verlag, Berlin, Germany. 364 p
Odong TL, Jansen J, van Eeuwijk FA, van Hintum TJL (2013) Quality of core collections for
effective utilisation of genetic resources review, discussion and interpretation. Theor Appl
Genet 126:289–305
Oldfield F (2005) Environmental change: key issues and alternative perspectives. Cambridge
University Press, Cambridge, UK. pp 259–261
Olivares-Villegas JJ, Reynolds MP, McDonald GK (2007) Drought-adaptive attributes in the
Seri/Babax hexaploid wheat population. Funct Plant Biol 34:189–203
Olmstead AL, Rhode PW (2011) Adapting North American wheat production to climatic
challenges, 1839–2009. Proc Natl Acad Sci USA 108:480–485
Osakabe Y, Yamaguchi-Shinozaki K, Shinozaki K, Tran LP (2013) Sensing the environment:
key roles of membrane-localized kinases in plant perception and response to abiotic stress. J
Exp Bot 64:445–458
Oweis T, Hachum A, Pala M (2005) Faba bean productivity under rainfed and supplemental
irrigation in northern Syria. Agr Water Manag 73:57–72
Passioura JB (2012) Phenotyping for drought tolerance in grain crops: when is it useful to
breeders? Funct Plant Biol 39:851–859
Patrick JW Stoddard FL (2010) Physiology of flowering and grain filling in faba bean. Field
Crops Res 115:234–242
Peleg Z, Fahima T, Abbo S, Krugman T, Nevo E, Yakir D, Saranga Y (2005) Genetic diversity
for drought resistance in wild emmer wheat and its ecogeographical associations. Plant Cell
Environ 28:176–191
Pereyra-Irujo GA, Gasco ED, Peirone LS, Aguirrezábal LAN (2012) GlyPh: a low-cost platform
for phenotyping plant growth and water use. Funct Plant Biolol 39:905–913
Perrino P, Robertson LD, Solh MB (1991) Maintenance, evaluation and use of faba bean
germplasm collections: problems and prospects. Options Mediterr 10:21–31
Pessoa-Filho M, Rangel PHN, Ferreira ME (2010) Extracting samples of high diversity from
thematic collections of large gene banks using a genetic-distance based approach. BMC
Plant Biol 10:127
Phan HTT, Ellwood SR, Ford R, Thomas S, Oliver RP (2006) Differences in syntenic complexity
between Medicago truncatula with Lens culinaris and Lupinus albus. Funct Plant Biol
33:775–782
Phan HT, Ellwood SR, Hane JK, Ford R, Materne M, Oliver RP (2007) Extensive macrosynteny
between Medicago truncatula and Lens culinaris ssp. culinaris. Theor Appl Genet 114:549–
558

54
Pinstrup-Andersen P, Pandya-Lorch R, Rosegrant MW (1999) World food prospects: critical
issues for the early twenty-first century. International Food Policy Research Institute,
Washington DC, USA
Polignano GB, Uggenti P, Scippa G (1993) The pattern of genetic diversity in faba bean
collections from Ethiopia and Afghanistan. Genet Resour Crop Evol 40:71–75
Požárková D, Koblížková A, Román B, Torres AM, Lucretti S, Lysák M, Doležel J, Macas J
(2002) Development and characterization of microsatellite markers from chromosome 1-
specific DNA libraries of Vicia faba. Biol Plant 45:337–345
Purushothaman R, Zaman-Allah M, Mallikarjuna N, Pannirselvam R, Krishnamurthy L, Gowda
CLL (2013) Root anatomical traits and their possible contribution to drought tolerance in
grain legumes. Plant Prod Sci 16:1–8
Quattrocchio F, Wing JF, van der Woude K, Mol JNM, Koes R (1998) Analysis of bHLH and
MYB-domain proteins: Species-specific regulatory differences are caused by divergent
evolution of target anthocyanin genes. Plant J 13:475–488
R Development Core Team (2012) R: A language and environment for statistical computing. R
Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL
http://www.R-project.org
Raab S, Hoth S (2007) A mutation in the AtPRP4 splicing factor gene suppresses seed
development in Arabidopsis. Plant Biol 9:447–452
Rafalski JA (2010) Association genetics in crop improvement. Curr Opin Plant Biol 13:174–180
Raina SN, Rees H (1983) DNA variation between and within chromosome complements in Vicia
species. Heredity 51:335–346
Ramsay G, van de Ven W, Waugh R, Griffiths DW, Powel W (1995) Mapping quantitative trait
loci in faba beans. In: Improving production and utilization of grain legumes. Edited by
AEP. Copenhagen, Denmark: 2nd European conference on grain legumes. pp 444–445
Reuzeau C, Frankard V, Hatzfeld Y, Sanz A, Van Camp W, Lejeune P, De Wilde C, Lievens K,
de Wolf J, Vranken E, Peerbolte R, Broekaert W (2006) TraitmillTM
: a functional genomics
platform for the phenotypic analysis of cereals. Plant Genet Resour 4:20–24
Ricciardi L (1989) Plant breeding for resistance to drought. II. Relationship between stomata and
agronomic traits in Vicia faba L. genotypes. Agri Mediterr 119:424–434
Ricciardi L, Polignano GB, De Giovanni C (2001) Genotypic response of faba bean to water
stress. Euphytica 118:39–46
Richards RA, Rebetzke GJ, Watt M, Condon AG, Spielmeyer W, Dolferus R (2010) Breeding
for improved water productivity in temperate cereals: phenotyping, quantitative trait loci,
markers and the selection environment. Funct Plant Biol 37:85–97
Rispail N, Kaló P, Kiss GB, Ellis TH, Gallardo K, Thompson RD, Prats E, Larrainzar E, Ladrera
R, González, EM, Arrese-Igor C, Ferguson BJ, Gresshoff PM, Rubiales D (2010) Model
legumes contribute to faba bean breeding. Field Crops Res 115:253–269
Ristic Z, Jenks MA (2002) Leaf cuticle and water loss in maize lines differing in dehydration
avoidance. J Plant Physiol 159:645–651
Robertson LD, El-Sherbeeny M (1991) Distribution of discreetly scored descriptors in a pure line
faba bean (Vicia faba L.) germplasm collection. Euphytica 57:83–92

55
Román B, Torres AM, Rubiales D, Cubero JI, Satovic Z (2002) Mapping of quantitative trait loci
controlling broomrape (Orobanche crenata Forsk.) resistance in faba bean (Vicia faba L.).
Genome 45:1057–1063
Román B, Satovic Z, Avila CM, Rubiales D, Moreno MT, Torres AM (2003) Locating genes
associated with Ascochyta fabae resistance in Vicia faba L. Aust J Agric Res 54:85–90
Román B, Satovic Z, Pozarkova D, Macas J, Dolezel J, Cubero JI, Torres AM (2004)
Development of a composite map in Vicia faba L. breeding applications and future
prospects. Theor Appl Genet 108:1079–1088
Ruiz-Rodriguez MD, Avila CM, Torres AM, Fuchs J, Schubert I (2014) Anchoring of genetic
linkage maps to the chromosome complement of Vicia faba L. Mol Breed 33:743–748
Sánchez FJ, Manzanares M, Andrés EF de, Tenorio JL, Ayerbe L (1998) Turgor maintenance,
osmotic adjustment and soluble sugar and proline accumulation in 49 pea cultivars in
response to water stress. Field Crops Res 59:225–235
Sarker A, Erskine W, Singh M (2005) Variation in shoot and root characteristics and their
association with drought tolerance in lentil landraces. Genet Resour Crop Evol 52:89–97
Sarris D, Koutsias N (2014) Ecological adaptations of plants to drought influencing the recent
fire regime in the Mediterranean. Agric For Meteorol 184:158–169
Sato S, Nakamura Y, Asamizu E, Isobe S, Tabata S (2007) Genome sequencing and genome
resources in model legumes. Plant Physiol 144:588–593
Sato S, Isobe S, Tabata S (2010) Structural analyses of the genomes in legumes. Curr Opin Plant
Biol 13:146–152
Satovic Z, Torres AM, Cubero JI (1996) Genetic mapping of new morphological, isozyme and
RAPD markers in Vicia faba L. using trisomics. Theor Appl Genet 93:1130–1138
Satovic Z, Avila CM, Cruz-Izquierdo S, Díaz-Ruíz R, García-Ruíz GM, Palomino C, Gutiérrez
N, Ocaña-Moral S, Gutiérrez MV, Cubero JI, Torres AM (2013) A reference consensus
genetic map for molecular markers and economically important traits in faba bean (Vicia
faba L.). BMC Genom 14:932
Saxena MC, Varma S (1985) Faba bears, kabuli chickpeas, and lentils in the 1980s. An
international workshop. May 16–20, 1983; proceedings. International Centre for
Agricultural Research in the Dry Areas. Aleppo, Syria. 395 p
Schachtman DP, Goodger JQ (2008) Chemical root to shoot signaling under drought. Trends
Plant Sci 13:281–287
Schmutz J, Cannon SB, Schlueter J, Ma J, Mitros T, Nelson W, Hyten DL, Song Q, Thelen JJ,
Cheng J, Xu D, Hellsten U, May GD, Yu Y, Sakurai T, Ume-zawa T, Bhattacharyya MK,
Sandhu D, Valliyodan B, Lindquist E, Peto M, Grant D, Shu S, Goodstein D, Barry K,
Futrell-Griggs M, Abernathy B, Du J, Tian Z, Zhu L, Gill N, Joshi T, Libault M,
Sethuraman A, Zhang XC, Shinozaki K, Nguyen HT, Wing RA, Cregan P, Specht J,
Grimwood J, Rokhsar D, Stacey G, Shoemaker RC, Jackson SA (2010) Genome sequence
of the palaeopolyploid soybean. Nature 463:178–183
Semagn K, Bjornstad A, Ndjiondjop MN (2006) Principles, requirements and prospects of
genetic mapping in plants. Afr J Biotechnol 5:2569–2587

56
Semagn K, Babu R, Hearne S, Olsen M (2014) Single nucleotide polymorphism genotyping
using Kompetitive Allele Specific PCR (KASP): overview of the technology and its
application in crop improvement. Mol Breed 33:1–14
Sharpe A, Ramsay L, Sanderson L-A, Fedoruk MJ, Clarke WE, Li R, Kagale S, Vijayan P,
Vandenberg A, Bett KE (2013) Ancient orphan crop joins modern era: gene-based SNP
discovery and mapping in lentil. BMC Genom 14:192
Shehadeh A (2011) Ecogeographic, genetic and taxonomic studies of the genus Lathyrus L. PhD
Thesis. School of Biosciences. University of Birmingham, UK
Shindo C, Bernasconi G, Hardtke CS (2007) Natural genetic variation in Arabidopsis: tools, traits
and prospects for evolutionary ecology. Ann Bot 99:1043–1054
Siddique KHM, Loss SP, Thomson BD (2003) Cool season grain legumes in dryland
Mediterranean environments of Western Australia: significance of early flowering. In:
Saxena NP (ed) Management of agricultural drought. Science publishers, Enfield (NH),
USA. pp 151–161
Sillero JC, Villegas-Fernández AM, Thomas J, Rojas-Molina MM, Emeran AA, Fernández-
Aparicio M, Rubiales D (2010) Faba bean breeding for disease resistance. Field Crops Res
115:297–307
Silvente S, Sobolev AP, Lara M (2012) Metabolite adjustments in drought tolerant and sensitive
soybean genotypes in response to water stress. PLoS One 7:e38554
Singh D, Dikshit HK, Singh R (2013) A new phenotyping technique for screening for drought
tolerance in lentil (Lens culinaris Medik.). Plant Breed 132:185–190
Sirks MJ (1931) Beitrage zur einer genotypischen Analyse der Ackerbohne, Vicia faba L.
Genetica 13:209−636
Sjödin J (1971) Induced morphological variation in Vicia faba L. Hereditas 67:155–180
Sprent J (2009) Legume nodulation: A global perspective. Wiley-Blackwell, Chichester, UK.
http://dx.doi.org/10.1002/9781444316384
Stephens A, Lombardi M, Cogan NOI, Forster J, Hobson K, Materne M, Kaur S (2014) Genetic
marker discovery, intraspecific linkage map construction and QTL analysis of ascochyta
blight resistance in chickpea (Cicer arietinum L.). Mol Breed 33:297–313
Stoddard FL (1993) Limits to retention of fertilized flowers in faba beans (Vicia faba L.). J
Agron Crop Sci 171:251–259
Stoddard FL, Bond DA (1987) The pollination requirements of the faba bean. Bee World
68:144–152
Stoddard FL, Hämäläinen K (2011) Towards the world’s earliest maturing faba beans. Grain
Legumes 56:9–10
Stoddard FL, Balko C, Erskine W, Khan HR, Link W, Sarker A (2006) Screening techniques and
sources of resistance to abiotic stresses in cool-season food legumes. Euphytica 147:167–
186
Surahman M (2001) The construction of genetic map of faba bean using RAPD markers. Bul
Agron 29:31–33
Suso MJ, Moreno MT, Cubero JI (1993) Variation among Spanish faba bean cultivars: taxonomy
and evolutionary implications. Genet Resour Crop Evol 40:105–111

57
Suso MJ, Vishnyakova M, Ramos A, Ambrose M, Duc G, Maggioni L (2005) Report of a survey
on conservation, management and regeneration of grain legume genetic resources. 111 p
Tai HH, Goyer C, Murphy AM (2013) Potato MYB and bHLH transcription factors associated
with anthocyanin intensity and common scab resistance. Botany 91:722–730
Takuno S, Terauchi R, Innan H (2012) The power of QTL mapping with RILs. PLoS One
7:e46545
Tanzarella OA, Pace C De, Filippetti A (1984) Stomatal frequency and size in Vicia faba L. Crop
Sci 24:1070–1076
Terzopoulos PJ, Bebeli PJ (2008) Genetic diversity of Mediterranean faba bean (Vicia faba L.)
with ISSR markers. Field Crops Res 108:39–44
Terzopoulos PJ, Kaltsikes PJ, Bebeli PJ (2003) Collection, evaluation and classification of Greek
populations of faba bean (Vicia faba L.). Genet Resour Crop Evol 50:375–381
Terzopoulos PJ, Kaltsikes PJ, Bebeli PJ (2004) Characterization of Greek populations of faba
bean (Vicia faba L.) and their evaluation using a new parameter. Genet Resour Crop Evol
51:655–662
Terzopoulos PJ, Kalitsikes PJ, Bebeli PJ (2008) Determining the sources of heterogeneity in
Greek faba bean local populations. Field Crops Res 105:124–130
Thomé OW (1885) Flora von Deutschland, Österreich und der Schweiz. Gera, Germany.
(available from http://caliban.mpiz-koeln.mpg.de/thome/band3/tafel_132.html)
Torres AM, Weeden NF, Martín A (1993) Linkage among isozyme, RFLP and RAPD markers in
Vicia faba. Theor Appl Genet 85:937–945
Torres AM, Satovic Z, Cánovas J, Cobos S, Cubero JI (1995) Genetics and mapping of new
isozyme loci in Vicia faba L. using trisomics. Theor Appl Genet 91:783–789
Torres AM, Vaz Patto MC, Satovic Z, Cubero JI (1998) New isozyme loci in faba bean (Vicia
faba L.): Genetic analysis and mapping using trisomics. J Hered 89:271–272
Torres AM, Román B, Avila CM, Satovic Z, Rubiales D, Sillero JC, Cubero JI, Moreno MT
(2006) Faba bean breeding for resistance against biotic stresses: towards application of
marker technology. Euphytica 147:67–80
Torres AM, Avila CM, Gutierrez N, Palomino C, Moreno MT, Cubero JI (2010) Marker-assisted
selection in faba bean (Vicia faba L.). Field Crops Res 115:243–252
Torres AM, Avila CM, Stoddard FL Cubero JI (2012) Faba bean. In: Pérez de la Vega M, Torres
AM, Cubero JI, Kole C (eds) Genetics, genomics and breeding in crop plants: cool season
food legumes. Science Pubs Inc, New Hampshire. pp 50–97
Turner NC, Shahal A, Berger JD, Chaturvedi SK, French RJ, Ludwig C, Mannur DM, Singh SJ,
Yadava HS (2007) Osmotic adjustment in chickpea (Cicer arietinum L.) results in no yield
benefit under terminal drought. J Exp Bot 58:187–194
United Nations (2011) Global drylands: A UN system-wide response. Prepared by the
Environment Management Group. 132 p. Available at:
http://www.zaragoza.es/contenidos/medioambiente/onu//issue07/1107-eng.pdf
van de Ven WTG, Waugh R, Duncan N, Ramsay G, Dow N, Powell W (1991) Development of a
genetic linkage map in Vicia faba using molecular and biochemical techniques. Aspects
Appl Biol 27:49–54

58
Varshney RK, Mahender T, Aggarwal RK, Börner A (2007) Genetic molecular markers in
plants: development and applications. In: Varshney RK, Tuberosa R (eds) Genomic assisted
crop improvement: genomics approaches and platforms. Vol. 1. The Netherlands, Springer.
pp 13–30
Varshney RK, Bansal KC, Aggarwal PK, Datta SK, Craufurd PQ (2011) Agricultural
biotechnology for crop improvement in a variable climate: hope or hype? Trends Plant Sci
16:363–371
Varshney RK, Song C, Saxena RK, Azam S, Yu S, Sharpe AG, Cannon S, Baek J, Rosen BD,
Tar’an B, Millan T, Zhang X, Ramsay LD, Iwata A, Wang Y, Nelson W, Farmer AD, Gaur
PM, Soderlund C, Penmetsa RV, Xu C, Bharti AK, He W, Winter P, Zhao S, Hane JK,
Carrasquilla-Garcia N, Condie JA, Upadhyaya HD, Luo M-C, et al (2013) Draft genome
sequence of kabuli chickpea (Cicer arietinum): genetic structure and breeding constraints
for crop improvement. Nat Biotech 31:240–246
Vavilov NI (1922) The law of homologous series in variation. J Genet 12:47–89
Vaz Patto MC, Torres AM, Koblizkova A, Macas J, Cubero JI (1999) Development of a genetic
composite map of Vicia faba using F2 populations derived from trisomic plants. Theor Appl
Genet 98:736–743
Voorrips RE (2002) MapChart: Software for the graphical presentation of linkage maps and
QTLs. J Hered 93:77–78
Walter A, Scharr H, Gilmer F, Zierer R, Nagel KA, Ernst M, Wiese A, Virnich O, Christ MM,
Uhlig B, Jünger S, Schurr U (2006) Dynamics of seedling growth acclimation towards
altered light conditions can be quantified via GROWSCREEN: a setup and procedure
designed for rapid optical phenotyping of different plant species. New Phytol 174:447–455
Wang H, Clarke JM (1993) Genotypic, intraplant, and environmental variation in stomatal
frequency and size in wheat. Can J Plant Sci 73:671–678
Wang H, Zong X, Guan J, Yang T, Sun X, Ma Y, Redden R (2012) Genetic diversity and
relationship of global faba bean (Vicia faba L.) germplasm revealed by ISSR markers.
Theor Appl Genet 124:789–797
Wang S, Basten CJ, Zeng Z-B (2012) Windows QTL Cartographer 2.5. Department of Statistics,
North Carolina State University, Raleigh, NC,
http://statgen.ncsu.edu/qtlcart/WQTLCart.htm
Webb A, O'Sullivan DM, Khazaei H, Stoddard FL, et al (in preparation) A 768-locus genetic
map of faba bean.
Würschum T (2012) Mapping QTL for agronomic traits in breeding populations. Theor Appl
Genet 125:201–210
Yang T, Bao S-Y, Ford R, Jia T-J, Guan J-P, He Y-H, Sun X-L, Jiang J-Y, Hao J-J, Zhang, X-Y,
Zong, X-X (2012) High-throughput novel microsatellite marker of faba bean via next
generation sequencing. BMC Genom 13:602
Young DN (2000) Constructing a plant genetic linkage map with DNA markers. In: Philips RL,
Vasil JK (eds) DNA-based markers in plants. Kluwer Academic Publishers, Netherlands. pp
31–47

59
Yu L, Chen X, Wang Z, Wang S, Wang Y, Zhu Q, Li S, Xiang C (2013) Arabidopsis enhanced
drought tolerance1/HOMEODOMAIN GLABROUS11 confers drought tolerance in
transgenic rice without yield penalty. Plant Physiol 162:1378–1391
Zeid M, Schon CC, Link W (2001) Genetic diversity in a group of recent elite faba bean lines.
Czech J Genet Plant Breed 37:34–40
Zeid M, Schon CC, Link W (2003) Genetic diversity in recent elite faba bean lines using AFLP
markers. Theor Appl Genet 107:1304–1314
Zhang JY, Broeckling CD, Blancaflor EB, Sledge MK, Sumner LW, Wang ZY (2005)
Overexpression of WXP1, a putative Medicago truncatula AP2 domain containing
transcription factor gene, increases cuticular wax accumulation and enhanced drought
tolerance in transgenic alfalfa (Medicago sativa). Plant J 42:689–707
Zhou L, Ni E, Yang J, Zhou H, Liang H, Li J, Jiang D, Wang Z, Liu Z, Zhuang C (2013) Rice
OsGL1-6 is involved in leaf cuticular wax accumulation and drought resistance. PLoS One
8:e65139
Zhu H, Choi H, Cook DR, Shoemaker RC (2005) Bridging model and crop legumes through
comparative genomics. Plant Physiol 137:1189–1196
Zong X, Liu X, Guan J, Wang S, Liu Q, Paull JG, Redden R (2009) Molecular variation among
Chinese and global winter faba bean germplasm. Theor Appl Genet 118:971–978
Zong X, Ren J, Guan J, Wang S, Liu Q, Paull JG, Redden R (2010) Molecular variation among
Chinese and global germplasm in spring faba bean areas. Plant Breed 129:508–513