kirk o. winemiller, department of wildlife and fisheries
TRANSCRIPT

Kirk O. Winemiller, Department of Wildlife and Fisheries Sciences, Texas A&M University,College Station, TX, 77843, USA (e-mail: [email protected])
ABSTRACTDespite the fact that peacock cichlids (Cichla spp.) areim IK>rtafit food and game fishes, little research has beenconducted on native fluvial populations in South America. Here I review ecological infonnation obtained forCichla species based on recent research in Venezuela. Cichla are essentially restricted to waters of hightransparency where they often are dominant diurnal piscivores. Within Venezuela, Cichla orinocensis is themost widely distributed species, and up to four species may coexist in certain rivers of the RIo Casiquiare Basinin Amazonia. In Venezuela, Cichla ocellaris is restricted to the Rio Cuyunf Basin, and Cichla cf. monoculusis restl:icted to the Casiquiare and upper Orinoco Basins. In certain rivers, up to three Cichla species coexistby partitioning available habitats: Cichla intermedia are near structure in the river channel within or near swiftcurrent, C. orinocensis in shallow water near the margins of lagoons and slow river reaches, and Cichlatemensis in somewhat deeper areas of lagoons and the river channel, often near banks. Cichla orinocensis andC. temensis in blackwater rivers of the Amazon region are larger on average when compared to conspecificsin the savanna (llanos) region of the Orinoco Basin. Nonetheless, conspecifics from diverse locations confonnto the same length-weight regression and have similar growth rates. Cichla temensis and C. orinocensis havebeen successfully introduced into Venezuelan reservoirs, where C. temensis tends to inhabit deeper water.
Cichla are substrate nesters with biparental care of eggs and fry lasting up to several weeks. In fluvialsystems, ~uction is initiated during low-water conditions and continues into the flood period. Males ofall species form nuccha1 humps, and both sexes have more intense coloration during the reproductive period.Characiform and perciform fishes are the major prey, and food partitioning has been documented in both
and showed less S~OQal shift in dict composition than C. temensis and C. intermedia in the RIo Cinaruco. InGuri Reservoir, C. temensis feeds more-on characiforms and less on cichlids than C. orinocensis. Peacockcichlid populations seem to be very sensitive to fishing pressure. Given their high abunqance, broad diets, and
blackwater ecosystems.
INTRODUCTIONP~ock cichlids (Cichla spp.) are among the most. widespread and conspicuous predatory fishes of low-
land rivers within the large drainage basins of trQpicalSouth America. They are called pav6~ in Venezuela,tucunar6 in most other parts of South America, andpeacock bass by some English s~. Peacockcichlid is the official commoQ name designated by theAmerican Fisheries Society, however this name isalso used in the aquarium hobby for certain Africanrift lake cichlids. By most accounts, these are thelargest representatives within the Cichlidae, with only
the Lake Tanganyika piscivore, Boulengerochromismicrolepis, possibly attaining a larger maximum sizethan Cichla temensis (>12 kg).
The popularity of peacock cichlids as sportfishesderives from their ecological niche as voracious diur-nal piscivores. The ferocity with which peacock
JOURNAL OF AQUARlCULroRE AND AQVA11C SCIEN~
CICBLID RaEARca: STATE OF THE ARTVOLUME IX
Page 93

appear in Novoa (1993), Ochoalturbe (1993), and Gilet al. (1993). Zaret (1977) and Winemiller (1990)discussed the adaptive function of the caudal ocellus,or eyespot (a dark spot with a well-defmed light ringaround it), in Cichla.
Over the past 12 years, my colleagues and I havestudied fluvial and reservoir populations of peacockcichlids in Venezuela. Much of this research wasconducted in Venezuela's Rio Cinaruco (Orinocodrainage in southern llanos ofEstado Apure) between1989 and present. In addition, we examined Cichla inthe Rio Casiquiare and its tributaries (Rio Negro-Amazon drainage) between 1993-1999, Rio Aguarobetween 1995-97 (Estado Guarico), Guri Reservoir(Rio Carom, Estado Bolfvar) between 1993-4, LasMajaguas Reservoir (Rio Cojedes, Estado Portuguesa)from 1984-1994, and the Modules of Apure (a systemof dikes and ponds in Estado Apure) and adjacentCafioCaicarafrom 1982-1995. Althoughmuchaddi-tional research is needed, it is now possible to summa-rize and compare information to improve conserva-tion of this ecologically and economically importantcichlid.
TAXONOMIC DIVERSITY ANDBIOGEOGRAPHYAccording to Kullander (1986), the genus Cichla isdistinguished by a large mouth with a forward project-ing lower jaw, a large laminar infraorbital bone joinedtightly to the lachrymal; a notched dorsal fin (spinelength increases until the 5th and decreases posteri-orly until the penultimate, the last spine longer andtightly joined with the soft -rayed portion of the dorsalfin which is as high as the spinous portion); micro-gillrakers on both sides of all arches, with spines onexposed surfaces; and scaly fins, except for the pecto-rals. In addition, all Cichla > 1 ()() mm standard length(SL) have a distinct caudal ocellus (Kullander andNijssen, 1989). Stiassny (1982) diagnosed the genuswith four shared, derived characters of skeleton andmusculature, and in a later analysis (Stiassny, 1991)concluded that the neotropical pike cichlids(Crenicichla) were their closest relatives. Kullander's(1998) consensus tree from maximum parsimonyanalysis of the Cichlidae showed the neotropicals aspolyphyletic and grouped Cichla with Crenicichlanear the base of the main neotropical cichlid radiation.A more recent analysis based on mitochondrial DNA(Farias et al., 1999) grouped Cichla with Astronotus
cichlids attack baitfishes and artificial lures and thepower and tenacity of their resistance after beinghooked is now legendary among anglers. With in-
creasing frequency, peacock cichlid sportfishing sto-ries are published in newspapers, magazines, books,and internet web sites worldwide. This popularity hashad the unfortunate consequence of resulting in inten-tional introductions of Cichla species in tropical andsubtropical freshwater systems around the world,sometimes to the detriment of native fish populations.Exotic populations of Cichla have been established inreservoirs of Hawaii (Devick, 1969), Panama (Zaretand Paine, 1973), Puerto Rico (Lilyestrom et al.,1994) and Brasil (Godinhoetal., 1994), and canals inFlorida (Shafland, 1993). Peacock cichlids also havebeen used in native regions to control densities ofexotic tilapia in pond cultures (McGinty, 1984; Fisherand Grant, 1994)
The capacity for exotic peacock cichlids to alter,via predation, communities and entire ecosystems hasbeen demonstrated. In the 1970s, the introduction ofCichla ocellaris into Lake Gat1:in, a man-made reser-voir in Panama, resulted in massive changes in com-munity composition, the structure of the aquatic foodweb, and even components of the terrestrial commu-nity (Zaret and Paine, 1979). A similar scenario ofaquatic community transition followed the introduc-tion of Cichla orinocensis and C. temensis into LasMajaguas Reservoir in Venezuela (Barbarino, 1986;Winemiller, 1989).
Paradoxically, this group of cichlids has been thefocus of relatively little research. Until very recently,most of the published studies reported on exoticpopulations, mostly in reservoirs. Zaret (1980) pre-sented life history information for C. ocellaris in LakeGatUn, Panama. Characteristics of C. ocellaris inHawaiian reservoirs were summarized in technicalreports (Devick 1969, 1972), and life histories ofpeacock cichlids in fish farms were investigated inBrasil (Fontenele, 1950; Braga, 1952,1953). Aspectsof the ecology of peacock cichlids inhabiting nativerivers appear in Lowe-McConnell (1969), Goulding(1980), Goulding et al. (1988), and Calaet al. (1996).Descriptions of populations in Venezuelan reservoirs
JOURNAL OF AQUARICULTURE AND AQUAllC SCIEN~Clcm.w RFSEARCH: STATE OF THE ART
VOLUME IX
Page 94

Figure 1. Cichla species documented in Venezu-eta: C. temensis (male in breeding condition, RioPasimoni, Estado Amazonas) (left top),C. orinocensis (Rio Pasimoni) (left center),C. intermedia (Rio Cinaruco, E. Apure) (left bot-tom), C. cr. monoculus (Rio Emoni, E. Amazonas)(right top) and C. ocellaris (preserved specimen,Rio Cuyuni, E. Bolivar) (right center).
All five recognized species are known from Ven-ezuela (Fig. 1): C. ocellaris Schneider (Cuyuni Ba-sin), C. orinocensis Humboldt (Orinoco, Casiquiare-Negro), C. temensis Humboldt (Orinoco, Casiquiare-Negro), C. intermedia Machado 1971 (Orinoco,Casiquiare-Negro), and C. cf. monoculus Spix(Casiquiare-Negro). The geographic range of eachspecies within Venezuela is illustrated in Figure 2,with populations established in reservoirs representedas dots. Cichla orinocensis and C. temensis are themost widespread within Venezuela, and range through-out the Rio Negro Basin and into other rivers of
rather than the crenicichlines, and also concluded thatneotropicaI cichlids were monophyletic as assumed
previously (Stiassny, 1991).Five Cichla species are currently recognized by
systematists, but several additional species from theAmazon Basin apparently require description(Kullander, 1986; Ku11ander and Nijssen, 1989).Kullander (1986) indicated that five species werevalid, but more recently suggested 11 taxa (K" l1~nderand Nijssen, 1989). To date, the largest number ofcoexisting Cichla species have been found in theupper Rio Orinoco and upper Rio Negro Basins insouthern Venezuela (Machado, 1971; Winemiller un-published data). This region lies on the western fringeof the Guyana Shield and harbors some of the richestaquatic and terrestrial diversity of fauna and flora onearth (Goulding et aI., 1988; Berry et al., 1995).
JOURNAL OF AQUARICULnJRE AND AQUAnC SCIENCES
ClCBLID RESEARCH: STATE 01' 1m: ART
VOLUME IX
Page 95
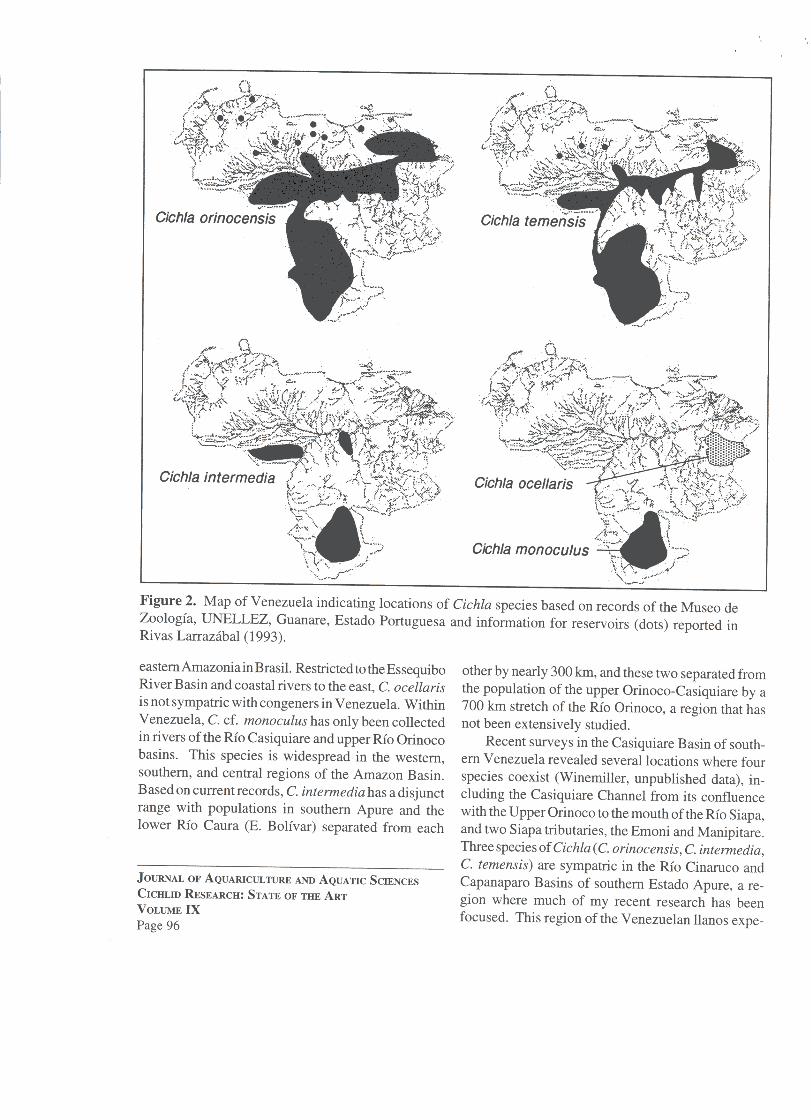

riences markedly seasonal rainfall and contains sandy,nutrient-poor soils, savanna vegetation. and clearacidic streams. The same three Cichla species are alsofound in blackwater rivers of Venezuela's EstadoAmazonas. Following Sioli (1975), blackwater riversare stained with dissolved organic compounds andhave low pH. conductivity, dissolved inorganic nutri-ent concentrations, and concentrations of suspendedsediments. The annual distribution of rainfall isgreater and less seasonal and the landscape moredominated by evergreen tropical forests as one travelssouthward through Venezuelan Amazonia. Both ofthese areas enjoy special protected status: the Cinaruco/Capanaparo as a National park and faunal reserve, andthe upper Orinoco/upper Rio Negro region containsnational parks and reserves for indigenous peoples.Because of the remoteness of the area. the fish stocksin southern Amazonas are less exploited.
A detailed description of C. ocellaris appears inKullander and Nijssen (1989). Venezuelan speci-mens of C. ocellaris are rare in museum collections,but all Cichla examined from the Rio Cuyuni conformto this species. Cichlaocellaris bas a moderately deepbody relative to known congeners. Like all Cichla,coloration is variable depending on ontogenetic stageand reproductive state. Sexual dichromatism has notbeen documented in this species. In live adults, thebackground varies from grey-green to yellow or ol-ive-green, the dorsUm darker than the flanks and thebelly very light. Three black vertical bars are on theflanks, the first two with a faint extension of the barthat reaches further ventrally. In adults, the dorsalportion of each bar consists of irregular dark blotches,lined by bright yellow or gold spots. Small irregularblack spots are generally present on the post orbital,cheek. and opercular regions. In most individuals,black blotches and spots are sometimes present on theflanks, especially near the belly and on the caudalpeduncle. Juveniles «50 mm SL) lack vertical barsbut have three large black spots on the flanks. Theyalso lack the caudal ocellus but have a large black spotat the caudal fin base (adults of all Cichla species havethe caudal ocellus, and small juveniles of all specieslack it). Colors, especially the orange-red of thethroat. pelvic fins, anal fin and lower half of the caudalfin, are more intense in breeding individuals of bothsexes (Zaret, 1980). This situation holds true for theother four species studied in Venezuela (all five havethe orange-red coloration in the same areas), and
males of all five species fonD a nucchal hump (abulging accumulation of fatty tissue on the forehead)just prior to and during the reproductive period. Inaddition, the iris of both sexes of all four Orinoco-Casiquiare Cichla species turns blood red during thereproductive period, and this presumably is a charac-teristic of all members of the genus.
Kullander (1986) provides a detailed descriptionof C. monoculus, and the species identified here as C.cf monoculus appears to agree with this description,although given its disjunct geographic distributionwith Amazonian populations, further investigation iswarranted. This species is present in the upper Orinocoand upper-mid reaches of the Rio Casiquiare andtributaries, but no records of C. monoculus exist forthe extreme blackwaters of the lower Casiquiare (in-cluding the Rio Pasimoni and tributaries) and upperRio Negro in Venezuela and Brasil. Relative to C.ocellaris, C. monoculus is more slender in adulthood,has a discontinuous rather than continuous lateral line,wider vertical bars on the flanks that extend ventrallyfrom the dorsum rather than beginning slightly belowthe dorsal fin base. To date, the only significant studyof C. monoculus ecology was conducted in the Co-lombian Amazon (Cala et al., 1996).
Machado ( 1971) gave the taxonomic descriptionsof C. intermedia, a species only know from Venezu-elan territory, and also described general features ofC. orinocensis (using the name C. ocellaris) and C.temensis. Cichla intermedia, a vividly colored fish, issometimes referred to as the royal or blackstripepeacock cichlid, or pav6n royal. Depending on envi-ronmental conditions and reproductive state, the back-ground color of this species varies from a dull grey-green to a brilliant turquoise-green. The ventral halfof the head and body is bright yellow in some indi-viduals. There is a very distinct black lateral band,actually composed of a series of interconnected verti-cally-elongate blotches, along the entire length of thebody. These blotches sometimes show a faint verticalbar extending dorsally to just below the base of thedorsal fin and ventrally toward the white belly. Smallblack flecks often appear on the post orbital region,cheeks, opercles, and especially the flanks. Juveniles
JOURNAL OF AQUARICULTURE AND AQUAnC ScIENCES
CICBLID RFSEAaca: STATE OF THE ART
VOWME IX
Page 97

Table 1. Maximum documented length, maximum age, mean growth rate, and minimum size ofmaturation reported for Cichla species from Venezuela (from Jepsenet aI., 1999; Winemilleret aI., 1997;Winemiller, unpublished data).
Maximum Agel
(yr)
Maximum SL
(mm)Mean Growth Rate
(mm/yr)
Minimum SL
Maturation (mm)
C. orinocensis 455 rivers550 reservoirs810 rivers621 reservoirs429 rivers
'"( 28.5 310 male270 female360 male325 female320 male270 female
C. temensis 9 46.0
C. intermedia t) 19,6
225 rivers 2346 rivers 3 [5+] 4
C. ocellarisC. cr. monoculus [230] 4
i estimates based on annuli and assuming fishes spawn for the fIrst time at age 1 (from Jepsen et aI.,
1999)2 Kullander (1986) reported maximum SL= 410 mm for C. ocellaris in Guyana.3 CaIa et aI. (1996) reported maximum SL= 504 rnm for C. monoculus in Peru.4 estimate from the Peruvian Amazon based on a length-frequency method of age estimation (CaIa
et aI., 1996)
Small juveniles «50 rom SL) have two black spots onthe flanks anteriorly, and a black lateral stripe on theposterior third of the body (Fig. 3). In exuemelyblackwaters, such as in Guri Reservoir, C. orinocensissometimes has a bronze background coloration andnumerous, small white spots on the body and fms.
Coloration of C. temensis is highly variable de-pending on ontogenetic stage, state of reproductivematuration, and breeding preparedness. Juveniles«50 rom SL) have a black horizontal stripe extendingfrom the shoulder to the base of the caudal fin (Fig. 3).Larger juveniles, subadults, and nonbreeding adultsare bronze with a broad, dark lateral band that extendsthe length of the body and tIU"ee broad vertical barsspaced apart evenly below the length of the dorsal fm.The body may be covered with white or cream spots,some diffuse, and others arranged in horizontal dashesabove and below the 18teral band. The head, dorsalfin, and caudal fin also have white spotting. The headhas large, irregular black blotches on the postorbitaland dorsal part of the opercular region. This blackpigmentation seems to be larger and extends furtherventrally on the head in adult females, but this requiressystematic study for verification. Smaller, scattered
have a more uniform lateral band than adults, and thefaint vertical bars often are apparent
Cichla orinocensis is similar in appearance to C.ocellaris, indeed, the species has been erroneouslyidentified as that species in most of the literature priorto Kullander (1982) and much confusion remainseven today. Cichla orinocensis has a discontinuouslateral line and generally has a more brilliant back-ground coloration than C. ocellaris that ranges fromyellow to green. Adults lack distinct vertical bars(some smaller individuals may show a very faintpattern of vertical baring above and below the spots)but instead have three large, sometimes irregular,black spots along the flanks. Similar to the caudalocellus, each spot has a bright yellow or gold outline,which gives rise to the common name three-star orbutterfly peacock (pav6n tres estrellas, pav6n mari-posa). Cichla orinocensis essentially lacks any blackmarkings on the head that are present in congeners.
JOURNAL OF AQUARICULTURE AND AQuADC SCIEN<D
CICBLID RFSEARCB: STATE OF THE ART
VOLUME IX
Page 98
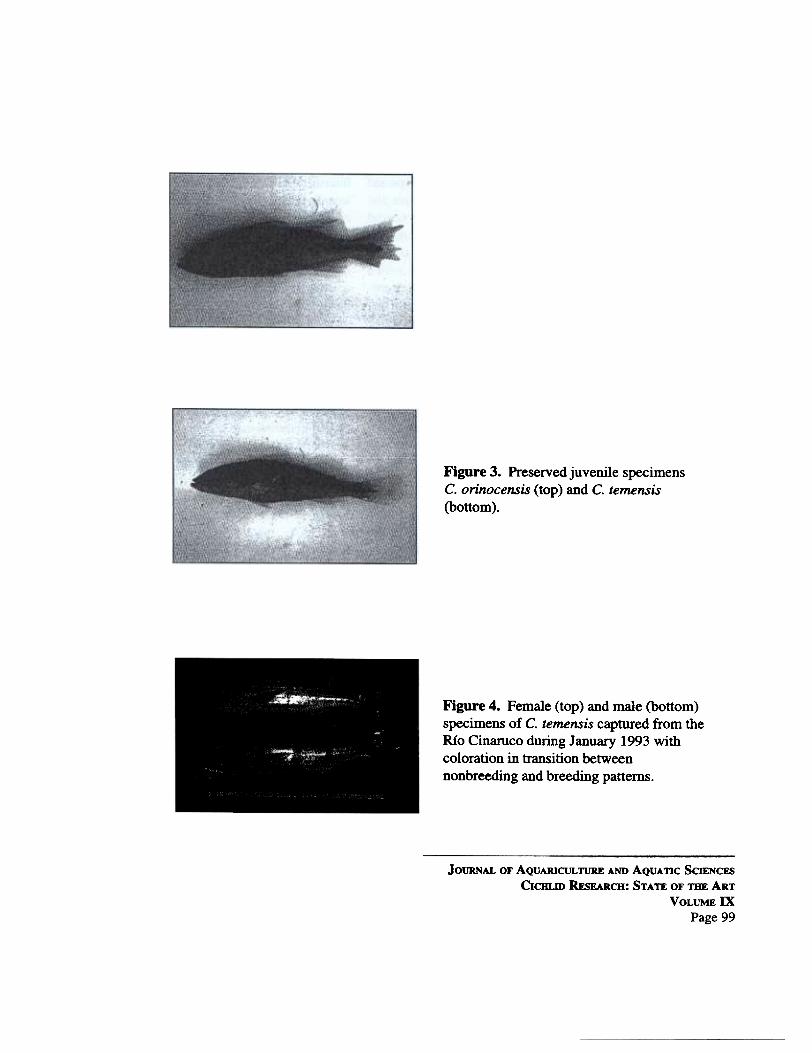
Figure 3. Preserved juvenile specimensC. orinocensis (top) and C. temensis(bottom).
Figure 4. Female (top) and male (bottom)specimens of C. temensis captured from theRio Cinaruco during January 1993 withcoloration in transition betweennonbreeding and breeding patterns.
JOURNAL OF AQUARICULTURE AND AQUAnC SCIENCES
CIcm.m RESEARCH: STATE OF THE ART
VOLUME IX
Page 99

black spots also appear on the cheek and opercle ofboth sexes. Breeding adults turn green with thedorsum darker than the ventrum. and the red-orangecoloration of the throat, anal fin, and lower half of thecaudal fin intensifies in the manner typical of allCichla. The orange coloration can extend onto thebelly and ventral flanks in some individuals. Ripe andbreeding adults also have three broad black bars thatextend from the base of the dorsal fm to the ventralflanks, which normally are white. Fish in transitionbetween the nonbreeding and ripe/breeding colora-tion pattern are encountered frequently, often with adull olive-green background coloration, black verti-cal bars, and white spotting on the head, body, and fms(Fig. 4). The highly variable coloration pattern of C.temensis causes considerable confusion among sportanglers, many of whom erroneously conclude thatmultiple species are represented. Among the variouslocal names used for this species are "pav6ncinchado","pav6n trucha" and "lapa" in Venezuela, and "tucunarepinima" in Brasil. Banded and speckled peacock aretwo of the most common English names used for this
species.
HABITAT ASSOCIA nONSAlthough peacock cichlids collectively inhabit a di-versity of habitats ranging from streams to largereservoirs, each species has fairly distinct habitataffinities which are particularly apparent in regionswere two or more species coexist. The universalrequirement of all Cichla species appears to be highwater transparency, warm temperatures, and access tolentic habitats for feeding and/or reproduction. Theneed for clear water apparently is tied to the fact thatCichla are strictly diurnal predators that rely on visionand rapid pursuit to capture prey. In all of my fieldexperiences, I have never found peacock cichlids inturbid waters associated with whitewater systems(i.e., neutral pH, high conductivity, high concentra-tions of suspended clays and other sediments, andhigh nutrient concentrations). Undoubtedly, they areable to traverse stretches of turbid water within the RioOrinoco in order to disperse among the many mar-ginallagoons and clearwater tributaries (the latter
having few suspended particulates, high transpar-ency, and low to neutral pH). The broader distributionof C. orinocensis relative to its congeners suggeststhat this species is more tolerant of turbidity. Forexample, C. orinocensis is well established in CafioCaicara (E. Apure), a river with characteristics that areintermediate between whitewater and blackwater sys-tems (e.g., moderate-high transparency, staining fromorganic compounds, pH 6-6.5, and floating aquaticmacrophytes). It is unclear if the species has a historicpresence in this river, which forms the northwesternlimit of its geographic range, or if it colonized from theadjacent modules, a dike/pond system constructedduring the 1970s. In other regions of the llanos, C.orinocensis is sometimes found in excavated pondsand shallow impoundments with moderate transpar-
ency.The five Cichla species in Venezuela show con-
sistent patterns of association with waters of differentphysicochemical characteristics according to Sioli' s(1975) classification scheme for tropical rivers. Asmentioned above, C. orinocensis seems to be mosttolerant of a wide range of conditions ranging fromextreme blackwaters, to clear waters, to waters withcharacteristics intermediate between them and thoseof white waters. Even so, there is no question that C.orinocensis achieves greatest population densitiesunder classic clearwater (e.g., Aguaro, Cinaruco) andblackwater (e.g., Pasimoni) conditions. Cichlaorinocensis penetrates further into headwaters than C.temensis, and occurs more frequently in small tribu-tary streams that flow directly into large rivers. Ap-parently, these two species are eliminated from a largeregion of the Guyana Shield (corresponding to most ofEstado Bolivar and eastern Estado Amazonas) by thepresence of cataracts. Cichla temensis seems to beentirely restricted to clearwater and blackwater rivers,and C. intennedia and C. cf. monoculus apparently arerestricted to clearwater rivers of the upper Orinoco-Casiquiare region (plus the moderate blackwater RioCapanaparo and blackwater Rio Caura as northern-most limits in the case of C. intennedia).
Recent studies (Jepsen et al., 1997; Winemiller etal., 1997) based on extensive sampling with hook andline and nets described the habitat associations of thethree peacock cichlid species in the Rfo Cinaruco.Both studies discovered a high degree of habitatseparation among the species (Fig. 5). Cichlaintennedia is essentially restricted to the main chan-
JOURNAL OF AQUARICULTURE AND AQUATIC SCIEN~
CICBLID RESEARCH: STATE OF mE ART
VOLUME IX
Page 100
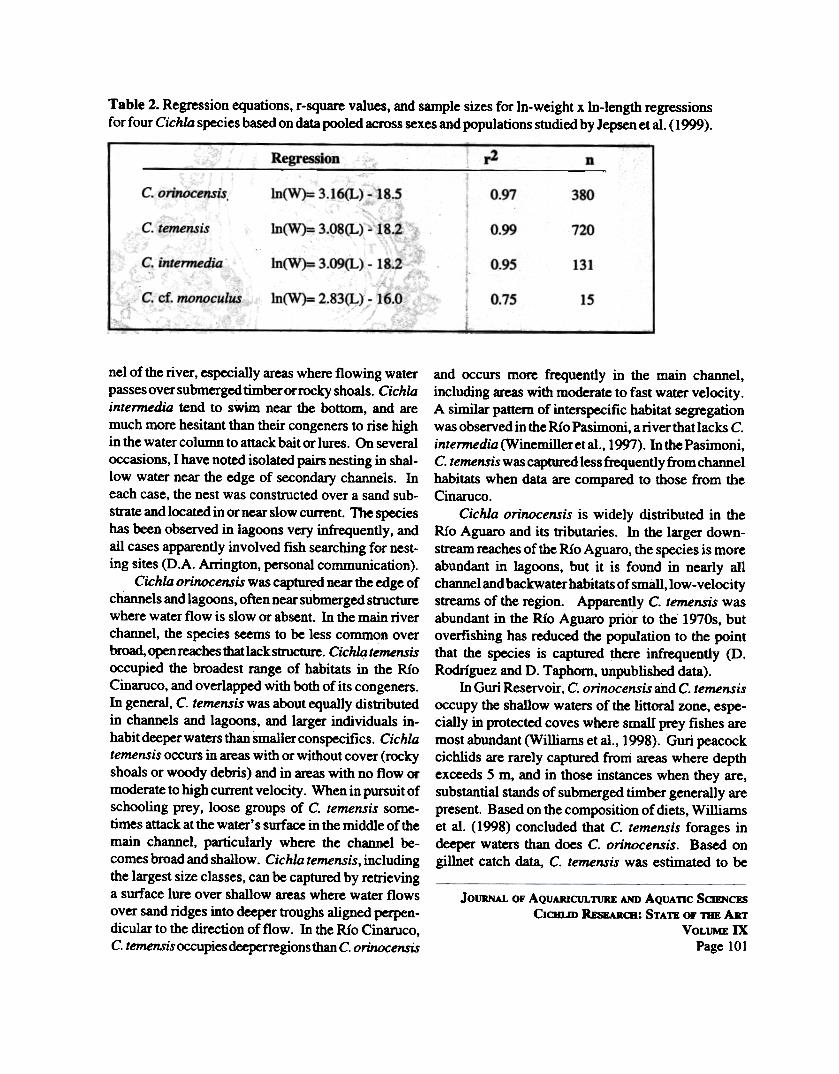
Table 2. Regression equations, r-square values, and sample sizes for In-weight x In-length regressionsfor four Cichla species based on data pooled across sexes and populations studied by Jepsen et aI. (1999).
r2Regression n
C. orinocensis In(W)= 3.16(L) - 18.5 0.97 380
1n(W)= 3.08(L) - 18.2 0.99 720C. temensis
C. intermedia In(W)= 3.09(L 18.2 0.95 13k
C. ct. monoculus In(W)= 2.83(L) - 16.0 O.7S 15
nel of the river, especially areas where flowing waterpasses over submerged timber or rocky shoals. Cichlaintennedia tend to swim near the bottom, aDd aremuch more hesitant than their congeners to rise highin the water column to attack bait or lures. On severaloccasions, I have noted isolated pairs nesting in shal-low water near the edge of secondary channels. Ineach case, the nest was constructed over a sand sub-strate and located in or near slow current. The specieshas been observed in lagoons very infrequently, andall cases apparently involved fish searching for nest-ing sites (D.A. Arrington, personal communication).
Cichla orinocensis was c~ near the edge ofchannels and lagoons, often near submerged structurewhere water flow is slow or absent. In the main riverchannel, the species seems to be less common overbroad. open reaches that lack structure. Cichla temensisoccupied the broadest range of habitats in the RioCinaruco, and overlapped with both of its congeners.In general, C. temensis was about equally distributedin channels and lagoons, and larger individuals in-habit deeper waters than smaller conspecifics. Cichlatemensis occurs in areas with or without cover (rockyshoals or woody debris) aDd in areas with no flow ormoderate to high current velocity. When in pursuit ofschooling prey, loose groups of C. temensis some-times attack at the water's surface in the middle of themain channel, particularly where the channel be-comes broad and shallow. Cichla temensis, includingthe largest size classes, can be captured by retrievinga surface lure over shallow areas where water flowsover sand ridges into deeper troughs aligned perpen-dicular to the direction of flow. In the Rio Cinaruco,C. temensis occupies deeperregions than C. orlnocensis
and occurs more frequently in the main channel,including areas with moderate to fast water velocity .A similar pattern of interspecific habitat segregationwas observed in the RIo Pasimoni, a river that lacks C.intennedia (Winemiller et aI., 1997). In the Pasimoni,C. temensis was captured less frequently from channelhabitats when data are compared to those from theCinaruco.
Cichla orinocensis is widely distributed in theRIo Aguaro and its tributaries. In the larger down-stream reaches of the RIo Aguaro, the species is moreabundant in lagoons, but it is found in nearly allchannel and backwater habitats of small, low-velocitystreams of the region. Apparently C. temensis wasabundant in the RIo Aguaro prior to the 1970s, butoverfishing has reduced the population to the pointthat the species is captured there infrequently (D.Rodriguez and D. Taphorn, unpublished data).
In Guri Reservoir, C. orinocensis and C. temensisoccupy the shallow waters of the littoral zone, espe-cially in protected coves where small prey fishes aremost abundant (Williams et aI., 1998). Guri peacockcichlids are rarely captured from areas where depthexceeds 5 m, and in those instances when they are,substantial stands of submerged timber generally arepresent. Based on the composition of diets, Williamset aI. (1998) concluded that C. temensis forages indeeper waters than does C. orinocensis. Based ongillnet catch da~ C. temensis was estimated to be
JOURNAL OF AQUARlCULroRI: AND AQUADC SCIENCES
CICHLID RDEARca: STATE 01' THE ART
VOLUME IX
Page 101

'e~.'1.:
\.' ~
~~~ ~..~.. -~ ""'.
.,:-;~'.~~
/
Figure S. Schematic diagramof a segment of the Rio Cinaruco
channel showing habitataffmities of Cichla species
(I = C. intennedia,0 = C. orinocensis,
T = C. temensis).Arrows indicate regions
of strongest current velocity,thin lines indicate
depth contours.
.~
~
more abundant in Guri, and both species have de-clined in abundance between the mid-1980s andpresent. This decline probably has been due to agradual decline in aquaiic production in this olig-otrophic reservoir, rather than fishing pressure.
FEEDING ECOLOGY AND SPE~INTERA cnONSThe vast majority of Cichla are piscivorous, withadult size classes only occasionally consuming shrimpand other ~uatic invertebrates. The exception ap-pears to be certain populations inhabiting reseryoirs.In Las Majaguas, Barbarino found significant frac-tions of shrimp in the diets of both C. orinocensis andC. temensis (Table 3). Small juveniles consumeaquatic microcrustacea. and fishes >50 mm consumefishes, aquatic and terrestrial insects, and shrimp, withfishes being consumed increasingly by larger sizeclasses. In the Rio Cinaruco, the system where we
JOURNAL OF AQVARlCULnJRE AND AQUADC SClFSca
CICIn.ID RaEARCB: STATE OF THE ART
VOLUME IX
Page 102
have the most dietary data for Cichla, fishes com-prised from 80-100% of the diet of peacock cichlidsbased on the volume of stomach contents. Resourcepartitioning was documented, especially during thelow and rising water periods. During the dry season,Cinaruco C. orinocensis and C. temensis feed heavilyon small characifonn fishes, and C. intennedia con-sume a variety of fishes, including small catfishes. Aswater levels rose, C. temensis fed more on juvenileCichla and silurifonn fishes, C. intennedia fed moreon insects and crustacea, and C. orinocensis fed on awide variety of fish prey. Cichla orinocensis had abroader diet overall, and showed less seasonal shift indiet composition than C. temensis and C. intennedia.This can perhaps be explained by the fact that C.orinocensis forages in shallow water near the inter-face with terrestrial vegetation throughout the year,whereas the other two species tend to be in the channelor funherfrom shore during the falling and low-waterperiods, but move into these areas during the floodperiod. Table 3 summarizes Cichla diets based onstudies conducted in four systems.
In Guri Reservoir, C. temensis consume more

characiform fishes, and C. orinocensis feed more oncichlids(Giletal., 1993; Novoaet al., 1993; Williamset al., 1998). In Gun, these two species showed nodifference in the size of prey consumed (Williams etal., 1998), but in the Cinaruco C. temensis consumedsignificantly larger prey than C. orinocensis (Jepsenet al., 1997). When data were adjusted for predatorsize, there was no difference among prey sizes con-sumed by the three peacock cichlid species of theCinaruco, which is to say the species are capable ofconsuming equal-size prey in relation to predatorlength. In rivers containing large C. temensis, mostfish taxa probably are vulnerable to predation. I havereceived accounts from sport anglers of cases in whichC. orinocensis fighting on the end of a fishing linehave been consumed by large C. temensis. I have alsoobserved large C. temensis attacking schools of largecharaciform fishes (e.g., Chalceus macrolepidorus,Semaprochilodus kneri, Boulengerella spp.) near thesurface of the water. These attacks are quite explo-sive, and fishes that are not grasped on an initial attackmay hover at the surface, stunned and subject tosubsequent attacks. The explosiveness and power ofthis surface feeding behavior is a primary cause for theinternational sportfishiog industry's interest in this
species.The demonstrated prowess of peacock cichlids as
piscivores, when combined with their typically highabundance, suggests a potential to influence preypopulations. The experiences from reservoirs andexotic peacock cichlid populations indicate a poten-tial role for peacock cichlids as keystone predators.Keystone predators can suppress the population den-sities of competitively superior prey species, and as aresult, foster higher levels of local species diversity(paine, 1966). In Lake GatUo. the introduction of C.ocellaris was followed by radical changes in commu-nity composition and the structure of the aquatic foodweb (Zaret and Paine, 1972). The evidence suggeststhat Cichla predation eliminated or greatly reducedformerly dominant prey populations which in turohada cascading effect on other aquatic species and evencertain terrestrial species. For example, the elimina-tion of small invertebrate-feeding fishes resulted inmore mosquito larvae and a higher incidence of ma-laria around the lake. Cichla greatly reduced thepopulation of pelagic atherinid fishes which wasfollowed by a reduction in terns. This case involvedC. ocellaris interacting as an exotic species in a
reservoir, an artificial habitat. Could peacock cichlidsfunction as keystone predators within native fluvialcommunities?
Given the dominance of Cichla as diurnalpiscivores in their native river ecosystems, it seemscertain that many strong food-web effects, both direct(as keystone predators) or indirect (e.g., competitiveinteractions, or effects on species at nonadjacent lev-els in a food chain) are influenced by changes in eitherthe numerical abundance or the size structure ofCichla populations. Tropical aquatic food webs arevery complex, and even basic descriptive data arelacking for systems containing peacock cichlids. Inrecent years, my colleagues and I have been studyingvarious aspects of the fishes and ecology of theCinaruco.
Jepsen et al. (1997), Winemiller et al. (1997), andWinemiller and Jepsen (1999) described how C.temensis feed on Semaprochilodus men", an abundantcharaciform that migrates between the nutrient-poorCinaruco and the nutrient-rich floodplains of theOrinoco to the east. Each year during the rainy season,the migratory fishes spawn on the rich floodplains,then adults and juveniles migrate into the Cinarucoduring the falling-water period (Nov.-Jan.) and re-main there until the next rains (May). Cichla temensisfeed heavily on juvenile S. knen" during their massmigrations up the Cinaruco. This period of migrationis also the time when C. temensis show the lowestfrequency of empty stomachs, fastest growth, andimprovement in body condition. During the monthsbetween the end of the upstream migrations and thestart of the next rainy season, Cichla reduce theirfeeding activity and undergo gonadal maturation inpreparation for spawning. Therefore, the peacockcichlid population is nutritionally subsidized by bio-mass produced on the whitewater Orinoco floodplainin the form of migratory fish.
Analysis of stable isotope data indicates thatnearly half of the carbon assimi1.atp.Q by large C.temensis is derived from the whitewater system(Winemiller and Jepsen. in press). Cichla fitness isenhanced and its population density is increased as aresult of feeding on this imported production. Since
JOURNAL OF AQUARlCULnJRE AND AQUATIC SclENca
CICID..ID RFSEARCB: STATE OF THE ART
VOLUME IX
Page 103

1990; Oil et aI., 1993; Jepsen et aI., 1997).Whereas large adult C. temensis probably have
relatively few natural enemies (e.g., river otters),small size classes are vulnerable to predation by agreat variety of piscivorous fishes, but to date the onlydirect reports are for large Cichla feeding on smallerCichla.
nearly all Cinaruco fishes, including peacock cichlids,are year-round residents of the oligotrophic system,this nutritional subsidy for a large predator with abroad diet may have ramifications for the populationdynamics of resident prey populations. Research thatattempts to model the food web dynamics of thisspatial subsidy is ongoing in my lab. Other olig-otrophic rivers with peacock cichlids also supportlarge populations of migratory prochilodontids (e.g.,Rio Negro).
There are relatively few data to identify the majorpredators of peacock cichlids. Freshwater dolphins(lnia geoffrensis) are common in most rivers withpeacock cichlids, but the limited dietary informationavailable for dolphins indicates that they feed mostlyon small fishes (McGuire and Winemiller, 1998). Inthe Rio Cinaruco, dolphins sometimes capture pea-cock cichlids when they are released by sport anglers.Given that this has not been observed in other riverswhere sportfishing is less common, the behavior ap-pears to be learned. It seems doubtful that dolphins areable to capture and subdue large peacock cichlids.
Piranhas (serrasalmine characids) feed on pea-cockcichlid fins, especially when the cichlid is strug-gling on the end of a fishing line. Fin nips areextremely common on all species of Cichla in habitatswhere piranhas are common, ie. virtually all fluvialhabitats. Winemiller (1990) hypothesized that findamage from piranhas has been a sufficient selectivepressure to result in the evolution of the caudal ocellus(eyespot) as a deterrent. Based on field and laboratoryexperiments withAstronotus ocellatus, another preda-tory cichlid with a distinct caudal ocellus, WineIniller(1990; Winemiller and Kelso-Winemiller 1993) pre-sented evidence in support of the theory that thecaudal eyespot (false eye) in piscivorous cichlids is anadaptation that confuses small fin-nipping piranhasby caudal mimicry of the head. This is an alternative
. to Zaret's (1977) earlier hypothesis that the caudal
ocellus in C. acellatus functions as an intraspecificrecognition signal to avert cannibalism in this piscivore.Zaret ( 1977) stated that cannibalism does not occur inC. ocellatus, however many cases have been docu-mented among other Cichla species (Winemiller,
POPULA nON STRUCTUREThe density and size structure of peacock cichlidpopulations varies greatly among locations and spe-cies. In addition, both the density and size structure ofsome populations appear to have changed over time inresponse to fishing pressure and/or ecosystem matu-ration, the latter having been observed in new reser-voirs. The case for Guri Reservoir was already cited,and other Venezuelan reservoirs have shown similarpatterns of community and ecosystem development
following impoundment.The initial fish community of Las Majaguas Res-
ervoir contained a high biomass of local river species,with piranhas being the dominant piscivores(Fernandez Yepez and Ant6n, 1966). With time, theinflux of nutrients from the flooded terrestrial ecosys-tem waned, and fish biomass declined. During thissame period, peacock cichlids (C. orinocensis, C.temensis) and other cichlid species from other loca-tionsin the llanos were introduced into the reservoir.Control of piranhas was a major motivation for theintroduction of peacock cichlids ( A. Barbarino, peTs.comm.). The fish community now is dominated bycichlids (Winemiller 1989), with peacock cichlids asthe dominant piscivores. A similar situation appearsto be underway in the Bocon6 Reservoir (E.Portuguesa) , constructed in 1985 (pers. COInm., D.Bermudez and D. Taphorn, UNELLEZ). In the caseof Las Majaguas, very large peacock cichlids werecaptured during the mid 1980s, a period when thepeacock cichlid population appeared to have reachedits peak density. For example, I captured a 550 mmSL, 5 kg female C. orinocensis from the shore of the
lake in 1984. InLasMajaguas,areservoirconstructedin a whitewater region, C. temensis does not achievethe same size as conspecifics in rivers or clearwaterand blackwater reservoirs lying further east. This isfurther evidence that C. temensis is not as well adaptedphysiologically as C. orinocensis to variation in waterchemistry. Likewise, C. orinocensis from nativerivers never attain such large sizes, which indicates
JOURNAL OF AQUARICULTURE AND AQUATIC SClENCF3
Clcmm RFSEARCH: STATE OF THE ART
VOLUME IX
Page 104

Table 3. Comparison of dietary data for Cichla orinocensis, C. temensis, and C. intennedia from fourlocations based on results reported in IGil et aI., 1993; 2Williams et aI., 1998; 3Jepsen et aI. 1997;4Winemiller et aI., 1997; 5Winemiller, 1990; and 6Barbarino, 1996.
Data %wt. % vol. % vol. % freq , % vol. % rreq.
C. orinocensiscrustacea
characiforms
perciformssiluriformsunidentified/misc
0
35.1
40.6
o.~
31.9
66.3
00
3.6
43.7
4.7
()
81..5
0
32.0
46.0
022.0
44.8
2.2
43.9
024.3
024.3
6.6
41.407.4
0
66.0
19.8
0.1
70.7-
21.2
0
5.5
Q
57.1
17.1
4.521.2
0
53.8
15.4
3.826.9
0
0
22.60
19.3
C. temensiscrustacea
characiforms
perciformssilurifonnsunidentified/misc
014..2
C. intennedia
crustacea
characiforms
perciformssiluriforms
unidentified/miSt
20.6
12.4
23.2
5.7
50.3
()
61.5
1.1
15.4
0
the floodplain of the Rio Capanaparo to the north(LagunaBrava). Because it was based on a larger totalsample, they felt their best estimates were those for thetwo species combined: 148/ba for Laguna Larga, and178/ha for Laguna Brava. Consistent with theseestimates, angling catch per unit effort was higher inLaguna Brava (2.1 Cichlalperson/hr) than LagunaLarga (1.65 Cichla/person/hr).
that biotic interactions in rivers. possibly in the fOnDof competition with C. temensis. limit the maximumsize of this species in rivers. The population sizestructure of peacock cicblids in Las Majaguas hasdeclined markedly over the past 15 years. and capturerates by local fishers have declined as well (Barbarino.
1996).Our best estimates of population density and
structure comes from research on the Rio Cinaruco.Taphom and Barbarino (1993) used mark and recap-ture method to estimate the abundance of C. temensisand C. orinocensis in a 7.3 ha lagoon connected to theRio Cinaruco (Laguna Larga) and a 3.8 ha lagoon in
JOURNAL OF AQUARlCULTURE AND AQUA11C SCIEN~
CICBLID RESEARCH: STATE 01' THE ART
VOLUME IX
Page 105

Cichla temensis are larger than C. orinocensis andC. intermedia in the Cinaruco (Jepsen et al., 1997;Winemiller et al., 1997). In a study of age and growth,we found that C. temensis grows significantly fasterthan its two congeners (Jepsen et al., 1999). The studyby Jepsen et al. also compared maximum ages andsizes among river and reservoir populations (selectedfindings summarized in Table 1). Their analysisconfirmed that C. orinocensis attains larger maximumsizes in reservoirs, but also that C. temensis attains itslargest sizes in rivers. Based on the otolith method ofage determination, the maximum age in their sampleswas 8 years for C; temensis from the Rio Pasimoni(Amazonia). Using the otolith method, Winemiller etal. (1997) estimated a 750 mm SL, 9 kg C. temensisfrom the same river at 9 years. They concluded thatthis population was dominated by fish of 9-11 years.Their estimate of annual growth by Pasimoni C.temensis was based on limited data and an inappropri-ate model, and their estimate of 70 mm/yr should bereplaced by Jepsen et al.'s value of 46 mm/yr.
Although Pasimoni peacock cichlids tend to belarger than conspecifics in the Cinaruco, their weight-length relationships are virtually identical in the caseof C. temensis and very similar for C. orinocensis(Jepsen et aI., 1997; Winemiller et al., 1997). Theslope for the log-weight x log-length regression isused as an indicator of fish relative condition. For C.orinocensis, populations with the lowest slopes werefrom the Rio Aguaro and Amazonia, and the highestwas Las Majaguas Reservoir. For C. temensis, theslope was nearly the same for all river and reservoirpopulations examined. Table 2 presents weight xlength regression equations for four Cichla speciesbased on available data.
Repeated sampling indicates that the size struc-ture of the peacock cichlid populations in the RioCinaruco has not changed over the past ten years. Thisriver receives moderate fishing pressure, mostly fromsport anglers who practice catch and release. Also,based on standing densities of fishes during the low-water season, the Rio Cinaruco appears to be moreproductive than the more extreme blackwater rivers ofthe Guyana Shield and Amazonia. In contrast, our
samples from the Rio Pasimoni indicate a downwardshift in the size distribution of the C. temensis popu-lation over the past six years. We first surveyed thisriver during 1993 just after it had been discovered by
groups conducting sportfishing tourism. Althoughthe sport anglers practiced catch and release, this newcommercial enterprise stimulated greater fishing ac-tivity by local subsistence and commercial fishers,some of whom, with their increased income, were alsoable to buy more gasoline for the journey to thisremote area. If special precautions are not taken, catchand release can result in high mortality from piranhasand river dolphins.
Nearly all Pasimoni C. temensis in our January1993 sample were extremely large, with only oneindividual <560 mm SL. Using angling as our pri-mary collecting method, we did not observe or captureany small immature individuals of either Cichla spe-cies, an unusual occurrence. Our seine samples andvisual surveys also revealed no juvenile peacockcichlids. Presumably, the natural mortality amongvery young fish (larvae and young of the year) is veryhigh, and recruitment of young fish into the adultpopulations is very low under normal conditions.This is probably due to extremely low natural produc-tivity near the food web base wi~ the Pasimoni' sacidic, nutrient-poor waters, and also the high diver-sity of natural predators in the local fish assemblage.Surveys of the Pasimoni during January 1997, 1998and 1999 resulted in captures of smaller maximumsizes and many more juveniles and small adults.Because we used the same methods, we interpret thisdifference as reflecting a real change in populationsize structure. The removal of large adults via thefishery may have resulted in increased foraging op-portunities for juveniles and lower predation by larger
conspecifics.In the Rio Aguaro, more than two decades of
commercial and subsistence fishing has resulted inextremely low densities of C. temensis, and a C.orinocensis population comprised almost entirely ofage-l fish (Jepsen et al. 1997). Venezuelans longinvolved in peacock cichlid sportfishing claim that C.temensis formerly were abundant, with individuals 5-8 kg common in the Aguaro. The information avail-able at this time indicate a consistent pattern thatresults from overexploitation of fluvial populations ofpeacock cichlids, in which the elimination of largeadults is followed by an increase in the relative abun-
JOURNAL OF AQUARICULTURE AND AQUAnC SCIENCESCICHLm ~CH: STATE OF THE ART
VOLUME IX
Page 106

dance of small size classes and slow replacement ofthese large size classes. Cichla temensis appears to beparticularly vulnerable to such changes, and this maybe a function of its association with nutrient-poorecosystems and cannibalism by large adults causing arecruitment bottleneck.
REPRODUCTIONReproduction by peacock cichlids follows the typicalpattern of substrate spawning cichlids in which bothsexes defend eggs and fry. Nesting, spawning, andbrood care in C. ()cellaris was described by Braga(1952), Devick(1 %9), andZaret (1980), however theidentity of their species requires confirmation in lightof recent taxonomic work. AccordingtoZaret(1980),C. ocellaris guards its brood for a period of up to 10weeks, at which time the juveniles are 60-70 mm SL.Zaret described how the parents move the newlyhatched fry in their mouths from the oviposition nestto nest depressions that lie up to 2 maway. Theparentsessentially cease feeding during the period of broodcare, and all attention is focused on protecting thebrood from the multitude of egg and larval predatorsin their surroundings (ca. 250 fish species have beendocumented in the Rio Cinaruco). Anglers catchbrooding peacock cichlids, and presumably thesestrikes are born of aggression for brood defense ratherthan hunger: Jepsen et al. (1999) detennined thatotolith annuli are formed during the period just priorto and during nesting (end of the low-water period intothe start of the annual rains), which indicates reducednutritional status from lower food intake during thereproductive period. Zaret also discussed sexualselection in C. ocellaris. Since only males form largenucchal humps derived from fat deposition, this char-acter may serve as an honest advertisement to femalesthat indicates relative foraging success by the maleduring the previous growing season (and see Barlow,
1999).The number of eggs counted in five C. ocellaris
nests in Hawaii ranged from 4,457 to 5,744 (Devick,1969). Zaret (1980) claimed that C. ocellaris nests inPanama contained up to 10,000 eggs. Cala et al.(1996) estimated mean fecundity from 10 C. monocu-lus at 7,400. Based on counts from preserved ovaries,the mean fecundity of three ripe female C. intermedia
fromtheCinaruco(270-305mmSL)was 1,711;meanfecundity of three female C. temensis from the Cinaruco(330-430 mm SL) was 5,725. A single female C.
orinocensis from the Pasimoni contained 1,694 ripeova. These estimates for fishes from Venezuela arebased on relatively small adults. As in most batch-spawning fishes, fecundity increases with size inCichla.
My colleagues and I have only made informalobservations of nesting and brood care by peacockcichlids in Venezuela. In the Rio Pasimoni (January1993), we found six nests of C. orinocensis withfertilized eggs. All of these nests were constructed onrock substrates in shallow water (ca. 30 cm) near theshore of lagoons. During a period of unusually lowwater levels (January 1998), we captured many C.temensis in advanced stages of spawning prepared-ness from a single lagoon that was connected to themain channel of the Pasimoni. The fish appeared to beat highest densities over broad shallow reaches 3-15 mfrom the shoreline. I concluded that the fish wereeither nesting or preparing for nesting; During Janu-ary 1998, we caught seven male C. temensis withnucchal humps and bright coloration and 9 adultslacking humps (presumed females) from a cut-offpool (ca. 450 m2) in the Rio Siapa, a Casiquiaretributary. When we released one of these capturedfish into the river channel, it quickly returned 10 thelagoon through a shallow (15 cm), narrow (ca. 3 m)channel in the sandbank.
In the Rio Cinaruco, sport anglers target a broadshallow section of one particular lagoon during thelate dry season, because C. temensis are know to formnesting aggregations there each year. Zaret (1980)described how male C. ocellaris behave aggressivelytoward each other during the period of nest siteselection and courtship. Therefore, it appears likelythat C. temensis form nesting aggregations as afunc-tion of limited suitable nesting habitat rather thansocial instincts. I have observed nesting C. orinocensisas isolated pairs in several rivers and reservoirs, andD. Rodriguez (unpublished data) reported C.orinocensis nesting aggregations in lagoons of the
Aguaro.Most studies report that spawning Cichla can be
found during any time of the year, but that spawningpeaks generally occur just prior to and during the early
JOURNAL OF AQUARICULTURE AND AQUAnC SCIENCES
CJCHLm ~RCH: STATE OF THE ART
VOLUME IX
Page 107

part of the rainy season (Fontenele, 1950; Lowe-McConnell, 1969; Zaret, 1980; Jepsen et al., 1999). Ingeneral, it seems that spawning is more synchronizedin more seasonal habitats. For example, Cichla of theRio Cinaruco, an intensely seasonal floodplain river,show gradual gonadal maturation over the course ofthe falling-water season (December-May) and evi-dence of nesting and spawning during the final weeksof the dry season (Jepsen et al., 1999). Lowe-McConnell (1969) reported identical spawning dy-namics for C. ocellaris of the Rupununi savannaregion of Guyana.
Presumably, nesting and mate choice are achievedmore easily under conditions of low water and highfish density. Likewise, larval feeding and brooddefense may be achieved more effectively within theinundated floodplain. The same pattern of reproduc-tive seasonality is observed in large piscivorous cic:h1ids(Serranochromis spp.) of the seasonal Zambezi Riverfloodplain (Winemiller, 1991). In less seasonalhabi-tats, such as reservoirs, Cichla spawning probably ismuch less synchronized. The low rate of detection ofannuli (annual growth checks) on the otoliths ofCichla from Guri Reservoir (Jepsen et al., 1999)supports this contention. Zaret (.1980) reported spawn-ing activity during all months of the year in LakeGatlin, however two periods of peak activity werenoted. In the Peruvian Amazon, where rainfall is notas seasonal as the llanos, ripe C. monoculus werefound during 9 of 12 months surveyed, and spawningbegan during the annual period of increasing rainfalland peaked about one month prior to the period ofpeak rainfall (Cala et al., 1996).
CONSERV A nONDespite the fact that they are often the most abundantand conspicuous piscivores in South American aquaticsystems. pc.acock cichlids seem to be very sensitive tooverharvest. In the Rio Pasimoni of Venezuela' sremote Amazonian region. the population size struc-ture of C. temensis underwent a significant change just5-6 years following the initiation of organizedsportfishing. Numerous independent accounts reveal
that the Rio Aguaro (central llanos) used to support afishery for trophy C. temensis, which together with C.orinocensis was abundant 20-30 years ago. Now C.temensis are relatively uncommon in this river, andthe C. orinocensis population is dominated by age-lfish. From the late 19808 to the early 1990s, GuriReservoir could boast one of the premiere peacockcichlid fisheries in the world. Although large peacockcichlids still can be found in this giant blackwaterreservoir, much of the sP9rtfishing activity has beenshifted to the upper portion of the impoundmentwhere the discharge from the Rio Cuyunf and RioParagua is first received and aquatic primary produc-tion is higher.
In short, it appears that peacock cichlid popula-tions are ~erable to impacts from a variety ofcauses ranging from watershed changes to overhar-vest. Whencompared to the accumulated knowledgethat provides the basis for management of sportfishstocks in North America or Europe, it seems there isvery much that remains to be learned about the ecol-ogy of peacock cichlids. We have more informationfor non-native populations of peacock cichlids thannative stocks in South American waters. For speciessuch as C. monoculus and those in Brasil yet todescribed, we have little or no ecological information.
Within pristine native habitats, peacock cichlidsappear to live a decade or more, so that, once removed,old large individuals are not replaced for severalyears. The very successful largemouth bass fisheriesof North America might provide a management model,and size-based catch limits (including slot limits thatallow possession of fish smaller or larger than aprescribed size interval) might facilitate sustainedharvest of the largest size classes. Obviously, en-forcement is critical to the success of catch limits.However, enforcement has been lacking in manyregions of South America where commercial andsubsistence netting of peacock cichlids is conductedillegally {a detailed discussion of Venezuelan regula-tions and management recommendations appears inTaphorn and Barbarino, 1993).
Some features of peacock cichlid ecology and theaquatic ecosystems they inhabit make over harvestinga resource problem that transcends just fishery con-cerns. The popularity of peacock cichlids as sportfishesobviously stems from their ecological niche as vora-cious, diurnal piscivores. Because of the foreignexchange generated by the international sportfishery,
JOURNAL OF AQUARICULTURE AND AQUAnC SCIEN~
CICHLID RF8EARCH: STATE OF THE ART
VOLUME IX
Page 108

A CKN 0 WLEDG MENTSI thank the many colleagues who have collaboratedwith me on studies of Cichla ecology in Venezuela.especially Don Taphom, David Jepsen, AnielloBarbarino, Douglas Rodriguez, John Williams, ChrisPeterson, and Albrey Arrington. I also thank thoseindividuals and institUtions that in various ways facili-tated and supported our field efforts in Venezuela.These include Glenn Webb and Clayton Lofgren ofthe Cinaruco Fishing Lodge, Carol Marzuola of TheCinaruco Fishing Club, Luis Balbas of EDELCA-Guri, Hector Escande I of SAD A -AMAZONAS, Basil
Stergios of UNE'_.J.EZ, Octavio Santaella of SanCarlos del Rio Negro, and the staffs ofBIOCENTRO,REUNEI_1.F.Z, INPARQUES, and SARPA.
it remains in the best interest of South Americancountries to manage these fisheries on a sustainableyield basis.
In addition to having economic relevance, pea-cock cichlids may influence the community dynamicsof oligotrophic aquatic ecosystems with low in situproduction. As already noted, the introduction ofexotic peacock cichlids into Lake Gat11n, Panamaresulted in large-scale changes in the aquatic foodweb. Similar scenarios of community transition fol-lowing the introduction of Cichla into Venezuelanreservoirs were noted as well. Experiences based onthe introduction of peacock cichlids into impound-ments suggests that these predators can significantlyinfluence the structure and function of aquatic ecosys-tems. Because tropical rivers that contain peacockcichlids have not been studied as well as reservoirs,the issue deserves further attention. Peacock cichlidshave all the characteristics of keystone predators andtheoretically could function as promoters of biologi-cal diversity in tropical rivers via negative impacts oncompetitive dominants, thus facilitating coexistenceamong diverse prey taxa.
REFERENCESBarbarino, A. 1986. Estado ActUal del RecursoPesquero Recreacional y de la Eutroficacion Cul-tural en el Embalse Las Majaguas. Undergraduatethesis, Universidad Nacional Experimental de losllanos Occidentales, Guanare, Portuguesa,Venezuela.
CONCLUSIONSOver the past 10 years, research on peacock cichlidsin Venezuela has greatly increased our knowledge ofthe ecology of this important group of cichlids withintheir native geographic distribution. Prior to theseefforts, most ecological information was derived fromresearch conducted in aquaculture facilities and onnon-native populations introduced into reservoirs.We still have much to learn. Systematic relationships,taxonomy, and biogeography of the genus Cichlaremains poorly known, with several apparentlyundescribed taxa in Brasil (peTs. comm., S. Kullander).Basic ecological information, including habitats,trophic ecology and species interactions, is lacking formost native Cichla -populations. Because of theirappeal as sportfish and novelty fish for the aquariumtrade, Cichla are being transported to locations theworld over with increasing frequency. In tropicalregions, these piscivores have great potential to im-pact native faunas, a lesson learned from the scenarioin Lake Gatl1n, Panama. Yet in their native SouthAmerican rivers, peacock cichlids may play an impor-tant role in regulating high community diversity.Given the great ecological and economic importanceof these cichlids, considerably greater investment inresearch is clearly warranted.
Barbarino, A. 1996. Diagnostico del RecursoPesquero como base para su Reglamentacion en elEmblase "Las Majaguas", Estado Portuguesa, Ven-ezuela. Master's thesis, Universidad Nacional Ex-perimental de los Llanos Occidentales, Guanare,Portuguesa. Venezuela, 93 pp.
Barlow, G.W. 1998. Sexual-selection models forexaggerated traits are useful but constraining.American Zoologist, 38: 59-69.
Berry, P .E., B.K. Holst. and K. Yatskievych. 1995Flora of the Venezuelan Guayana: Vol. 1: Intro-duction. Missouri Botanical Garden, St. Louis,Missouri. 320 pp.
Braga, R. 1952. Ninhos de tucunares Cichlatemensis Humboldt e Cichla ocellaris Bloch &Schneider. Revista Brasileira Biologica, 12: 273-278.
JOURNAL OF AQUARlCULTlIRE AND AQUAnC SClENCFS
ClCHLID RESEARCH: STATE OF THE ART
VOLUME IX
Page l~

Braga, R. 1953. Crecimiento de tucunare pinima,Cichla temensis Humboldt, em cativeiro(Actinopterygii, Cichlidae). Dusenia, 4: 41-47.
Oil, C.E., J. Andrada, E. Mendez, andJ.M. Salazar.1993. Estudio preliminar sobre alimentaci6n encautiverio y contenido estomacal de Cichlatemensis del embalse Guri, Estado Bolivar, Venezu-ela. Natura, 96:42-47.Cala, P., E. Gonzales, and M.P. Varona. 1996.
Aspectos bio16gicos y taxon6micos deltucunare, Cichla monoculus (Pisces: Cichlidae).Dahlia, 1: 23-37.
Godinho, A.L., M. Tavares da Fonseca, and L.Marcia de Araujo. 1994. The ecology ofpredator fish introductions: the case of Rio DoceValley lakes. p. 77-83 in R. M. Pinto-Coelho, A.Giani, and E. von Sperling, eds.Ecology andHuman Impact on Lakes and Reservoirs of MinasGerais with Special Reference to FutureDevelopment and Management Strategies.SEGRAC, Belo Horizonte, Brazil.
Devick; W.S. 1969. Life history study of thetucunare, Cichla ocellaris. Federal Aid inSportfish Restoration Project F-4R-17, JobCompletion Report, Hawaii Department ofLand and Natural Resources, Honolulu. 34 pp.
Goulding, M. 1980. The Fishes and the Forest.University of California Press, Berkeley. 280 pp.
Devick. W.S. 1972. Life history study of thetucunare Cichla ocellaris. FederalAid inSportfish Restoration Project F-9-1. JobCompletion Report. Hawaii Department of Landand Natural Resources, Honolulu. 32 pp. Goulding, M., M.L. Carvalho, and E.G. FeITeira.
1988. Rio Negro, Rich Life in Poor water: Amazo-nian Diversity and Foodchain Ecology as seenthrough Fish Communities. SPB Academic Pub-lishing, The Hague, The Netherlands. 200 pp.
Farias, I.P., G. Ortf, I. Sampaio, H. Schneider, andA. Meyer. 1999. Mitochondrial DNA phylogenyof the family Cichlidae: monophyly and fastmolecular evolution of th~ neotropical assemblage.Molecular Evolution, 48: 703-711. Jepsen, D.B., K.O. Winemiller, and D.C. Taphorn.
1997. Temporal patterns of resource partitioningamong Cichla species in a Venezuelan black-waterriver. JournaI ofFish Biology, 51: 1085-1108.
Fernandez Yepez, A., and J. Ant6n. 1966. Estudio(analisis) Las Majaguas. Hoya de los rios CojedesSarare. Ministerio deObras Publicas, Caracas. 93pp. Jepsen, D.B., K.O. Winemiller, D.C. Taphom, and
D. Rodriguez-Olarte. '1999. Variation in age struc-ture and growth of ~cock cichlids from rivers andreservoirs of Venezuela. Journal cf Fish Biology,55: 433-450.
Fisher, F;W., andW.E. Grant.. 1994. Use ofana-.tive predator to control overcrowding inWanD-water polyculture ponds: simulation of atucunare (Cichla monoculus) tilapia '
(Oreochromis niloticus} system; Ecologicalmodelling, 72: 205-227.
Kullander, S.D. 1983. A Revision of the SouthAmerican Cichlid Genus Cichlasoma. SwedishMuseum of Natural History, Stockholm, Sweden.296 pp.Fontenele, O. 1950. Contribucao para 0
conhecimiento de biologiados tucunares(Actinopterygii,Cichlidae) em captiviero. Aprelhlode reproducao. Habitosde desova e incubacao.Revista Brasileira Biologica, 10: 503-519.
Kullander, S.O. 1986. Cichlid Fishes of the Ama-zon River Drainage of Peru. Swedish Museum ofNatural History, Stockholm, Sweden. 431 pp.
JOURNAL OF AQUARICULTURE AND AQUATIC SCIENCFS
CICHLm ~EARCH: STATE OF THE ART
VOLUME IXPage 110

t
Kullander, S.O. 1998. A phylogeny and classifica-tion of the South American Cichlidae (Teleostei:Percifonnes). p. 461-498 in L.R. Malabarba, R.E.Reis, R.P. Vari, S.M.S. Lucena, and C.A.S. Lucena,eds. Phylogeny and Classification of Neo- TropicalFishes. Editora Universitaria-EDIPUCRS, PortoAlegre. Brazil.
estrategias de ordenamiento pesquero. Sociedad deCiencias Naturales La Salle, 49: 159-197.
Ochoa lturbe, R. 1993. Los pavones enCamatagua. Natura, 96: 40-41.
Paine, R. T. 1966. Food web complexity and spe-cies diversity. American Naturalist, 100: 65-75.
Kullander, S. and H. Nijssen. 1989. The Cichlidsof Surinam: Teleostei, Labroidei. EJ. Brill,Leiden, The Netherlands. 256 pp.
Rivas LarrWbal, L. 1993. Distribuci6n depavones en Venezuela. Natura, 96: 30-33.
Shafland, P.L. 1993. An overview of Rorida's in-troduced butterfly peacock bass (Cichla ocellaris)sportfishery. Natura, 96: 26-29.
Lilyestrom, C.G., P. Quinones, and G. Oliveras.1994. Peacock and largemouth bass competition inLa Plata Reservoir. Federal Aid in Sportfish Resto-ration Project F-30. Job Completion Report, PuertoRico Deparbnent of Natural and Environmental Re-sources, San Juan. 32 pp.
Sioli, H. 1975. Amazon tributaries and drainagebasins. p. 199-213 in A. D. Hasler, ed. Coupling ofLand and Water Systems. Springer Verlag, NewYork.Lowe-McConnell, R.H. 1969. The cichlid fishes
of Guyana, South America, with notes on theirecology and breeding behavior. Zoological Journalof the Linnaean Society, 48: 255-302.
Stiassny, M.L.]. 1982. The relationships of theneotropical genus Cichla (percifonnes, Cichlidae):a phyletic analysis including some functional con-siderations. Journal of Zoology, London 197: 427-453.
Machado-Allison, A. 1971. Contribuci6n a1conocimiento de la taxonomia del genera Cichla(percifonnes Cichlidae) en Venezuela, Parte I.Acta Biologica Venezolana, 7: 459-497. Stiassny, M.L.J. 1991. Phylogenetic
intrarelationsbips of the family Cicblidae: an over-view. p. 1-35 in M.H.A. Keenleyside, ed. CichlidFishes: Behaviour, Ecology and Evolution.Chapman & Hall, London.
McGinty, A.S. 1984. Effects of predation bytucunare (Cichla ocellaris) on Tilapia nilotica inponds. Journal of Agriculture of the University ofPuerto Rico, 68: 101-105.
Taphorn, D.C., and A. Barbarino. 1993.Evaluacion de la situacion actual de los pavones,Cichla spp., en el Parque Nacional Capanaparo-CinalUco, Estado Apure, Venezuela. Natura, 9610-25.
McGuire, T.L., and K.O. Winemiller. 1998. Habi-tat affinity and population structure of freshwaterdolphins,lnia geoffrensis, in the Cinaroco River,Venezuela. Biotropica, 30: 625-638.
Novoa. D.P. 1993. Aspectos generales sobre labiologfa. pesqueria. manejo y cultivo del pav6n(Cichla orinocensis y C. temensis) en ellago deGuri y otras areas de la regi6n Guayana. Natura.96:34-39.
Williams, J.D., K.O. Winemiller, D.C. Taphom,and L. Balbas. 1998. Ecology and status ofpiscivores in Guri, an oligotrophic tropical reser-voir. North American Journal of Fisheries Man-agement, 18: 274-285.
JOURNAL OF AQUARICULTURE AND AQUAnC SCIENCES
ClCHLID RESEARCH: STATE OF THE ART
VOLUME IX
Page III
Novoa, D.F., J. Koonce, A. Locci, and F. Ramos.1989. La ictiofauna dellago de Guri:composicion, abundancia y potencial pesquero. ll.Evaluacion del potencial pesquero dellage Guri y

Winemiller, K.O. and D.B. Jepsen. In press. Mi-gratory neotropical fish subsidize food webs of oli-gotrophic blackwater rivers. in G.A. Polis, M.E.Power, and G. Huxel, eds. Food Webs at the Land-scape Level, University of Chicago Press, Chicago.
Winemiller, K.O. 1989. Patterns of variation inlife history among South American fishes in sea-sonal environments. Oecologia, 81: 225-241.
Winemiller, KO. 1990. Caudal eyespots as deter-rents against fm predation in the neotropical cichlidAstronotus ocellatus. Copeia, 1990: 665-673. Zaret, T .M. 1977. Inhibition of cannibalism in
Cichla ocellaris and hypothesis of predator mim-icry among South American fishes. Evolution, 31421-437.
Winemiller, K.O. 1991. Comparative ecology ofSerranochromis species (Teleostei: Cichlidae) inthe Upper Zambezi River. Journal of Fish Biology,39: 617-639. Zaret, T .M. 1980. Life history and growth rela-
tionships of Cichla ocellaris, a predatory SouthAmerican cichlid. Biotropica, 12: 144-157.Winemiller, K.O.andL.C. Kelso-Winemiller.
1993. Predatory response of piranhas to alternativeprey. National Geographic Research, 9: 344-357. Zaret, T.M., and R.T. Paine.. 1973. Species intro-
duction into a tropical lake. Science, 182: 449-455Winemiller, K.O., L.C. Kelso-Winemiller, and A.L.Brenkert. 1995. Ecological and morphological di'-versification in fluvial cichlid fishes. Environnuon-tal Biology of Fishes, 44: 235-261.
Winemiller. K.O.. D.C. Taphom, and A. BarbarinoDuque. 1997.. The ecology of Cichla (Cichlidae)in two blackwater rivers of southern Venezuela.Copeia. 1997: 690-696.
Winemiller, K.O. and D.B. Jepsen. 1998. Effectsof seasonality and fish movement on tropical riverfood webs. Journal of Fish Biology, 53 (Supple-ment A): 267-2%.
JOURNAL OF AQUARICULTURE AND AQUAllC SCIEN~
CICHLID RFSEARCH: STATE OF mE ART
VOLUME IX
Page 112