king s research portal fileking s research portal doi: 10.1210/en.2015-1040 document version peer...
TRANSCRIPT

King’s Research Portal
DOI:10.1210/en.2015-1040
Document VersionPeer reviewed version
Link to publication record in King's Research Portal
Citation for published version (APA):Grachev, P., Li, X. F., Goffin, V., & O'Byrne, K. T. (2015). Hypothalamic prolactin regulation of luteinizinghormone secretion in the female rat. Endocrinology, 156(8), 2880-2892. https://doi.org/10.1210/en.2015-1040
Citing this paperPlease note that where the full-text provided on King's Research Portal is the Author Accepted Manuscript or Post-Print version this maydiffer from the final Published version. If citing, it is advised that you check and use the publisher's definitive version for pagination,volume/issue, and date of publication details. And where the final published version is provided on the Research Portal, if citing you areagain advised to check the publisher's website for any subsequent corrections.
General rightsCopyright and moral rights for the publications made accessible in the Research Portal are retained by the authors and/or other copyrightowners and it is a condition of accessing publications that users recognize and abide by the legal requirements associated with these rights.
•Users may download and print one copy of any publication from the Research Portal for the purpose of private study or research.•You may not further distribute the material or use it for any profit-making activity or commercial gain•You may freely distribute the URL identifying the publication in the Research Portal
Take down policyIf you believe that this document breaches copyright please contact [email protected] providing details, and we will remove access tothe work immediately and investigate your claim.
Download date: 14. Jan. 2020

Hypothalamic Prolactin Regulation of LuteinizingHormone Secretion in the Female Rat
Pasha Grachev, Xiao Feng Li, Vincent Goffin, and Kevin T. O’Byrne
Division of Women’s Health (P.G., X.F.L., K.T.O.), School of Medicine, King’s College London SE1 1UL,United Kingdom; and INSERM Unité 1151 (V.G.), Faculté de Médecine, Institut Necker Enfants Malades,75993 Paris, France
Prolactin (PRL) levels increase in response to long-term antipsychotic treatment that disrupts re-productive function. Recent evidence suggests that activation of central PRL receptors (PRLR)inhibits LH secretion and in ovariectomized rats. However, the mechanisms involved, the mode ofLH secretion affected and relevance to hyperprolactinemia remain unknown. We therefore in-vestigated the contribution of central PRL/PRLR signaling to the control of estradiol-induced surgesof LH and PRL and pulsatile LH secretion under basal and hyperprolactinemic conditions. First, bysubjecting ovariectomized estradiol-primed rats intracerebroventricularly administered with PRLto frequent blood sampling, we demonstrated that acute activation of hypothalamic PRLR disruptspulsatile LH secretion. Pretreatment (intracerebroventricularly) with the pure PRLR antagonist,�1-9-G129R-hPRL, or the �-aminobutyric acid receptor type A antagonist, bicuculline, blocked thiseffect. Next, we revealed that sustained blockade of hypothalamic PRLR using �1-9-G129R-hPRLaugmented the magnitude of LH surges induced by estradiol benzoate and progesterone treat-ment and suppressed the concomitant surges of PRL. Finally, we determined that acute antagonismof central PRLR is insufficient to normalize the duration of the LH pulse interval prolonged as aresult of hyperprolactinemia induced by chronic exposure to the atypical antipsychotic sulpiride.These data serve as the first evidence to suggest that PRL signaling through hypothalamic PRLRinhibits pulsatile secretion of LH in a �-aminobutyric acid receptor type A-dependent fashion andtonically restrains the magnitude of the LH surge. Furthermore, our results indicate that transientblockade of hypothalamic PRL/PRLR signaling is not an effective strategy for restoring LH pulsatilityperturbed by chronic hyperprolactinemia. (Endocrinology 156: 0000–0000, 2015)
Prolactin (PRL) is a peptide hormone with diverse reg-ulatory functions in the reproductive, nervous, and
immune systems. It derives its name from its involvementin the control of lactation and mammary gland develop-ment (1). The anterior pituitary is the major origin of PRLand contains abundant lactotropes, specialized cells thatsecrete PRL into systemic circulation (2). The brain is a keytarget of PRL; circulating PRL is actively transportedacross the blood brain barrier into the cerebrospinalfluid (CSF) by PRL receptor (PRLR)-expressing cells ofthe choroid plexus (3). Furthermore, neural as well as
glial cells within circumventricular nuclei have beenshown to internalize and retain immunoreactive PRLfrom the CSF (4).
The plethora of biological effects exerted by PRL areinitiated by the interaction between PRL and its receptor,PRLR, which exists in long and short isoforms in humansand rodents (5). The PRLR is an archetype of type 1 cy-tokine receptors and comprises homodimerized single-pass transmembrane chains of minimal complexity com-pared to other family members (6). Upon PRL binding,conformational changes occurring within the receptor ho-
ISSN Print 0013-7227 ISSN Online 1945-7170Printed in USACopyright © 2015 by the Endocrine SocietyReceived January 14, 2015. Accepted May 14, 2015.First Published Online May 20, 2015
Abbreviations: aCSF, artificial CSF; ARC, arcuate nucleus; AVPV, anteroventral periven-tricular nucleus; CSF, cerebrospinal fluid; D2R, dopamine type 2 receptor; E2, 17�-estradiol;E2B, E2 benzoate; GABA, �-aminobutyric acid; icv, intracerebroventricular; KNDy, kisspep-tin/neurokinin B/dynorphin A; LC, locus coeruleus; OVX, ovariectomized; P4, progesterone;PI3K, phosphatidylinositol-3-kinase; PRL, prolactin; PRLR, PRL receptor; pSTAT5, phos-phorylated STAT5; SCN, suprachiasmatic nucleus; STAT5, signal transducer and activatorof transcription-5; TIDA, tuberoinfundibular dopaminergic.
tapraid4/zee-end/zee-end/zee99915/zee8070-15z xppws S�5 6/2/15 20:33 Art: EN-15-1040 Input-ss
O R I G I N A L R E S E A R C H
doi: 10.1210/en.2015-1040 Endocrinology press.endocrine.org/journal/endo 1
AQ:1–3
AQ: 4
AQ: 5
<zjs;Original Research> • <zjss;><zshorttitle>

modimer trigger a combination of intracellular signalingcascades including Janus kinase-2/signal transducer andactivator of transcription-5 (STAT5), Src kinase/phospha-tidylinositol-3-kinase (PI3K)/Akt, MAPK and Nek3-vav2-Rac1 pathways (5). PRLRs are expressed through-out the hypothalamus in both long- and short-formisoforms (7–10), notably within neurons known to playimportant roles in the control of reproductive hormonesecretion. Curiously, hypothalamic PRLRs have beenshown to be twice as abundant in women than in men (11).
Although few GnRH neurons, major hypothalamic ef-fectors of the hypothalamo-pituitary gonadal axis, ex-press PRLRs (12), they receive ample projections fromregulatory afferents that are PRLR positive: kisspeptin/neurokinin B/dynorphin A (KNDy) neurons of the arcuatenucleus (ARC), an essential component of the GnRHpulse generator; kisspeptin neurons of the anteroventralperiventricular nucleus (AVPV), which are indispensablefor mediating the positive-feedback effects of estradiol(E2) on GnRH secretion that results in the preovulatoryLH surge; and �-aminobutyric acid (GABA)ergic neuronsof the AVPV that regulate GnRH neuron excitability (12).Indeed, central (13) or peripheral (14) administration ofovine PRL prevents the occurrence of preovulatory LHsurges in ovary-intact rats and down-regulates ARC kiss-peptin expression and LH levels in ovariectomized (OVX)rats (15). Intracerebroventricular (icv) administration ofPRL also dose dependently induces the phosphorylationof STAT5 in the ARC and AVPV neurons of the OVX rat,and this effect is amplified by the replacement of E2 (16).A significant proportion of PRL-receptive ARC neuronsare likely to be KNDy neurons because icv administrationof PRL induced phosphorylated STAT5 expression inmost kisspeptin-immunoreactive cells in this region (15).
Another group of highly PRL-receptive hypothalamicneurons are involved in the negative feedback of PRL onits own secretion. Tuberoinfundibular dopaminergic(TIDA) neurons project from the ARC to the median em-inence, in which they secrete dopamine into the long portalvasculature (17). In addition, rodents possess tuberohy-pophyseal and periventricular-hypophyseal dopamineneurons (18), which innervate and secrete dopamine intoshort portal vessels within the neural and intermediatelobes of the pituitary that supply the anterior lobe (17).Dopamine released by these neuroendocrine systems ac-tivates dopamine type 2 receptors (D2R) on hypophyseallactotropes, tonically inhibiting PRL secretion (19). Cu-riously, KNDy neurons synapse with TIDA (20) andperiventricular-hypophyseal (21) neurons, and earlierstudies have demonstrated projection of ARC dynorphin-ergic neurons to TIDA neurons (22). Moreover, recentevidence suggests that kisspeptin stimulates PRL release
by inhibiting the activity of hypothalamic dopaminergicneurons in an E2-dependent fashion (23). Thus, the hy-pothalamic neuroendocrine systems regulating the secre-tion of gonadotropins and PRL appear to participate inregulatory cross talk that is subject to modulation by cen-tral PRL, albeit through yet unknown mechanisms.
LH surges occur after exposure to elevated E2 concen-trations that stimulate de novo synthesis and release ofprogesterone (P4) by specialized neurons and astrocites inthe diencephalon (24–26). The positive feedback effects ofE2 and P4 are then conveyed to GnRH neurons by kiss-peptin neurons of the AVPV (27) subject to appropriatecircadian disinhibition (28). Several lines of evidence im-plicate central PRL/PRLR signaling in the regulation of theLH surge: 1) hyperprolactinemia causes anovulation inwomen with microprolactinomas by inhibiting not onlypulsatile LH secretion but also the midcycle LH surge (29,30); 2) a high proportion of both ARC and AVPV kiss-peptin neurons express PRLR in female rats (12); 3) lac-tating mice express 90% less kisspeptin mRNA and 60%less immunoreactive kisspeptin in AVPV neurons com-pared with diestrous controls (31, 32). However the roleof central PRL in the regulation of gonadotropin surgesremains obscure. Moreover, it has long been known thatsurges of PRL accompany preovulatory LH surges in ro-dents (33), ruminants (34), and humans (35) alike, andalthough the primary role of the PRL surge is apparentlyluteolytic (36), its potential effects at the hypothalamiclevel have not previously been considered.
To characterize the mechanism by which central PRLsignaling regulates the GnRH pulse generator, we inter-rogated the hypothesis that GABAergic neurons mediatePRL-induced suppression of pulsatile LH secretion inOVX rats primed with low (diestrus d 2) levels of E2
(OVX�E2 rats). Because the involvement of central PRLsignaling in the regulation of gonadotropin surges is cur-rently unknown, we then studied the effect of intracere-broventricularly administered, �1-9-G129R-hPRL, aunique pure PRLR antagonist, on the dynamics of LHsurges induced by 17�-estradiol-3-benzoate (E2B) and P4
treatment (37) in OVX�E2 rats. Finally, we askedwhether acute blockade of central PRLR signaling wouldreverse the inhibitory effect of chronic antipsychotic-in-duced hyperprolactinemia on the LH pulse so as to addressthe hypothesis that chronically elevated PRL acts centrallyto suppress reproductive function.
Materials and Methods
AnimalsAdult female Sprague Dawley rats (200–250 g) obtained
from Harlan were housed under controlled conditions (12 h
tapraid4/zee-end/zee-end/zee99915/zee8070-15z xppws S�5 6/2/15 20:33 Art: EN-15-1040 Input-ss
2 Grachev et al Central PRL Signaling Regulates LH Secretion Endocrinology
AQ: 6
AQ: 7

light, 12 h darkness, with lights on at 7:00 AM; controlled am-bient temperature 22°C � 2°C) and provided with standard lab-oratory chow (RM1; SDS Diets) and water ad libitum. Animalswere group housed (maximum four per enclosure) prior to sur-gery and housed individually after surgery and during experi-mentation. All animal procedures were undertaken in accor-dance with the Animals (Scientific Procedures) Act, 1986 andwere approved by the King’s College London Ethical ReviewPanel Committee.
Surgical proceduresAll surgical procedures were carried out under general anes-
thesia induced by ketamine (100 mg/kg ip; Pharmacia and Up-john) coadministered with xylazine (Rompun; 10 mg/kg ip;Bayer); supplementary injections of ketamine (10 mg, ip) wereadministered to maintain anesthesia as required. Each rat wasinjected sc with 0.4 mL/kg Duphamox LA antibiotic suspension(200 mg/mL procaine benzylpenicillin, 250 mg/mL dihydro-streptomycin-sulfate; Fort Dodge Animal Health) prior to sur-gery. After each surgical procedure, animals were allowed torecover from anesthesia on a heated pad until fully conscious.Postoperative analgesia was provided by means of sc adminis-tration of carprofen (Rimadyl; 4.4 mg/kg; Pfizer Animal Health)daily for 3 days.
Two weeks before experiments took place, the rats were bi-laterally OVX and implanted with subcutaneous E2-filledSILASTIC brand capsules (Dow Corning), which produce cir-culating concentrations of E2 within the range observed duringthe diestrus (d 2) phase of the estrous cycle (�35.8 � 1.2 pg/mL),as previously described (38). Rats were then implanted with uni-lateral guide cannulae (22 gauge; Plastics One), stereotaxicallytargeted toward the left lateral cerebral ventricle for microinfu-sion of pharmacological agents. The stereotaxic coordinates forimplantation were 1.5 mm lateral and 0.6 mm posterior tobregma and 4 mm below the surface of the dura (39). Correctcannula placement was confirmed by the observation of gravi-tational meniscus movement upon insertion of an internal injec-tion cannula (Plastics One) with extension tubing preloaded withartificial CSF (aCSF). A 20-mm stainless steel slotted screw (In-stec Laboratories) was affixed to the surface of the skull posteriorto the guide cannula and both were secured using dental cement(Simplex Rapid; Dental Filling). The icv guide cannulae werethen fitted with obturators (Plastics One) to maintain patency.After a 10-day recovery period, the rats were implanted with twocustom-made cardiac catheters via the internal jugular veins toenable simultaneous automated serial blood sampling for pro-filing of LH levels and manual withdrawal of blood for deter-mination of PRL levels (40). The catheters were exteriorized atthe back of the head and enclosed within a 30-cm-long light-weight metal spring tether (Instec Laboratories) secured to theslotted screw cranial attachment with surgical ligature. The dis-tal end of the tether was attached to a dummy swivel (InstecLaboratories), allowing the rat to move freely. After surgery,animals were housed singly. Experimentation commenced aftera further 3 days of recovery from surgery.
Experimental designOn the morning of experimentation, an icv injection cannula
with extension tubing, preloaded with a combination of drugsolutions, was inserted into the guide cannula. The distal end of
the tubing was filled with aCSF. The remainder of the tubing wasfilled with sterile water, with 5 �L air separating the water andaCSF, which allowed the progress of injections to be monitored.The tubing was extended outside the cage and connected via onechannel of a two-channel fluid swivel (Instec Laboratories) to a25-�L syringe (Hamilton) prefilled with sterile water to allowremote microinfusion without disturbing the rats during the ex-periment. One of the two cardiac catheters was then attached viathe second channel of the fluid swivel to a computer-controlledautomated blood sampling system, which enables the intermit-tent withdrawal of small blood samples (25 �L) every 5 minutesfor 4–6 hours without disturbing the rats. After removal of each25-�L blood sample, an equal volume of heparinized saline (50U/mL normal saline; CP Pharmaceuticals) was automatically in-fused to maintain patency of the catheter and blood volume.Once connected, the animals were left undisturbed for 1 hourbefore blood sampling was initiated (between 9:00 and 10:00AM). Drug treatments were administered after a period of controlblood sampling manually over 5 minutes. Additional blood (50�L) was sampled manually via the second cardiac catheter atspecified time points, and each withdrawal was followed by theadministration (iv) of 50 �L heparinized saline. Plasma obtainedby centrifugation of manually withdrawn blood samples wasfrozen at �20°C and later assayed to determine concentrationsof PRL by means of a RIA. Automatically sampled blood wasfrozen at �20°C and later assayed to determine LH concentra-tions by means of RIA. To minimize animal suffering in line withthe Animals (Scientific Procedures) Act, 1986, rats were reusedas part of different treatment groups within experiments, but notbetween experiments, and were allowed to recover for at least 3days between bleeding procedures.
Drugs and hormones�1-9-G129R-hPRL, recombinant human PRL with Gly129
replaced with Arg and nine N-terminal residues deleted, is a purePRLR antagonist (41). �1-9-G129R-hPRL was expressed inEscherichia coli as inclusion bodies, using the pT7L expressionvector, as described previously (42), and purified using an ionexchange column (Hi Trap Q-Sepharose; GE Healthcare LifeSciences). Lyophilized aliquots of �1-9-G129R-hPRL were re-constituted with sterile water and diluted with aCSF. Recombi-nant rat PRL (ABC2856; Source BioScience Life Sciences) wasreconstituted from lyophilized stocks with sterile water and di-luted with aCSF. The selective GABAAR antagonist bicuculline(1(S),9(R)-(�)-bicuculline methbromide; Sigma-Aldrich) wasdissolved in sterile water and diluted with aCSF. The D2R an-tagonist sulpiride [(S)-(�)-sulpiride; sc-258111; Santa Cruz Bio-technology] was solubilized with 1 M tartaric acid (L-(�)-tar-taric acid (sc-218622; Santa Cruz Biotechnology) and dilutedwith saline, the final concentration of tartaric acid being 10 mM.
Effect of central administration of PRL on pulsatileLH secretion in OVX�E2 rats
A total of 7 OVX�E2 rats were used in this experiment; acrossover design ensured that each animal was randomly as-signed to a treatment group and subsequently to its counterpartcontrol group. The treatment groups were as follows: �1-9-G129R-hPRL (antag) � aCSF (n � 4), bicuculline (bic) � aCSF(n � 3), aCSF � PRL (n � 5), antag � PRL (n � 7), and bic �PRL (n � 5). All treatments were administered icv in a volume of
tapraid4/zee-end/zee-end/zee99915/zee8070-15z xppws S�5 6/2/15 20:33 Art: EN-15-1040 Input-ss
doi: 10.1210/en.2015-1040 press.endocrine.org/journal/endo 3

4 �L aCSF. One hour 40 minutes into a 6-hour blood samplingprocedure, rats were pretreated with 20 �g �1-9-G129R-hPRL,500 ng bicuculline, or 4 �L aCSF. Twenty minutes later, theanimals received a further icv injection of 5 �g PRL or 4 �L aCSF.Additional blood samples for measurement of PRL levels werewithdrawn at 60, 135, 150, 180, 240, and 360 minutes relativeto the time when the automated blood sampling was initiated.
Effect of hypothalamic PRLR antagonism on LHand PRL surges induced by E2B and P4 treatmentin OVX�E2 rats
To induce concomitant LH and PRL surges, OVX�E2 ratswere subjected to gonadal steroid priming according to a pro-tocol described previously (43). A total of 10 animals were in-jected (sc) with 2 �g E2B (Sigma-Aldrich) in 100 �L arachis oil(Sigma-Aldrich) at approximately 9:00 AM daily for 2 days. At9:00 AM on the morning of the experiment, the rats received 500�g P4 (Sigma-Aldrich) in 100 �L arachis oil (Sigma-Aldrich), sc.One hour 30 minutes later, a 1-hour control blood samplingprocedure was initiated. Three hours after the P4 administration,the rats were injected (icv) with 20 �g �1-9-G129R-hPRL in 4�L aCSF (n � 7) or 4 �L aCSF (n � 3). One hour later (240 minafter P4 administration), a further 6 hours of serial blood sam-pling was initiated. At the same time, a 3-hour infusion of 60 �g�1-9-G129R-hPRL in 12 �L aCSF (n � 7) or 12 �L aCSF (n �3), driven by a syringe pump (PHD 2000; Harvard Apparatus),was started. Additional blood samples for measurement of PRLlevels were withdrawn at 60, 120, 240, 300, 360, 420, 480, 540,and 600 minutes relative to the time of P4 administration.
Effect of hypothalamic PRLR antagonism onpulsatile LH secretion in chronicallyhyperprolactinemic OVX�E2 rats
A total of 15 animals were used in this experiment. To renderOVX�E2 rats chronically hyperprolactinemic, sc injections ofsulpiride (5 mg/kg in 450 �L 10 mM tartaric acid) were given at9:00 AM and 5:00 PM hours daily for at least 11 days prior toexperimentation as well as on the day of the experiment (n � 8).This protocol has been previously demonstrated to inducechronic hyperprolactinemia in OVX�E2 rats (16). Control rats(n � 7) received 450 �L 10 mM tartaric acid (sc) at 9:00 am and5:00 PM daily for at least 11 days prior to experimentation as wellas on the day of the experiment. Rats were subjected to a 4-hourserial blood sampling procedure that commenced at 10:00 AM.The animals received four consecutive icv injections of 20 �g�1-9-G129R-hPRL in 4 �L aCSF or 4 �L aCSF at 20-minuteintervals between 12:00 and 3:00 PM. Additional blood samplesfor measurement of PRL levels were withdrawn hourly between9:30 AM and 5:30 PM. The experiment followed a crossover de-sign whereby all animals received �1-9-G129R-hPRL and six ofthe eight sulpiride-treated rats and five of the seven control ratswere randomly selected to receive aCSF during separate trials atleast 3 days apart.
Hormone RIADouble-antibody RIAs supplied by the National Hormone
and Peptide Program (Harbor-UCLA Medical Center, Torrance,California) were used to determine LH and PRL concentrationsin the 25-�L whole-blood samples and 10-�L plasma samples,
respectively. Values are expressed in terms of the standard prep-arations NIDDK-rPRL-RP-3 and NIDDK-rLH-RP-3. The limitof sensitivity of the LH RIA was 0.093 ng/mL and the inter- andintraassay variations were 6.8% and 8.0%, respectively. Thelimit of sensitivity of the PRL RIA was 4.9 ng/mL. The intraassayvariation was 8.9% and all PRL values were obtained from asingle assay.
Data analysisDetection of LH pulses was facilitated by the use of the al-
gorithm ULTRA (44). Two intraassay coefficients of variation(2 � cv) of rat plasma pools, with LH concentrations fallingwithin the log phase of the LH RIA standard curve were used asthe reference threshold for pulse detection. The effect of treat-ments on LH pulse frequency was analyzed by comparing themean LH pulse interval in the 2-hour period preceding treatmentwith that in 2-hour posttreatment period(s). The period duration(in minutes) was divided by the number of LH pulses detected ineach of these periods to give the appropriate LH pulse interval.When there were no LH pulses evident during the first 2-hourposttreatment period, the LH pulse interval assigned to this pe-riod was taken as the interval from the onset of treatment to thefirst LH pulse in the second 2-hour posttreatment period. Thesignificance of the effect of treatments on LH pulse frequencywas compared with the respective negative control group at thesame time points as well as with the mean pulse interval duringthe 2-hour pretreatment period in the same treatment group.Statistical significance was tested using a one-way ANOVA withBonferroni post hoc test. P .05 was considered statisticallysignificant. Values given in the text and figures represent mean(�SEM).
PRL levels in all experiments, as well as surge-like LH levels,were compared using a two-way ANOVA with repeated mea-sures (influence of time and treatment) with Holm-Sidak pair-wise multiple comparison. Overall significance threshold wasP .05. In addition, cumulative changes in PRL and LH levelswere expressed as area under the curve for individual treatmentgroups, which were compared using a paired, two-tailed Stu-dent’s t test (two groups) or a one-way ANOVA with Bonferronipost hoc test (more than two groups). P .05 was consideredstatistically significant. Values given in the text and figures rep-resent mean (�SEM).
Results
Effect of central administration of PRL on pulsatileLH secretion in OVX�E2 rats
No effect of icv �1-9-G129R-hPRL pretreatment onthe LH pulse was observed in rats administered with aCSF(representative example in Figure 1A), at least during thefirst 4 hours 20 minutes after the injection. Bicuculline alsohad no significant effect on pulsatile LH secretion (repre-sentative example in Figure 1B). Administration (icv) ofPRL suppressed pulsatile LH secretion for a period of 1–2hours (representative examples in Figure 1, C and D). NoLH pulses were detectable during the period of PRL-in-duced suppression, the onset of which was immediate
tapraid4/zee-end/zee-end/zee99915/zee8070-15z xppws S�5 6/2/15 20:33 Art: EN-15-1040 Input-ss
4 Grachev et al Central PRL Signaling Regulates LH Secretion Endocrinology
F1
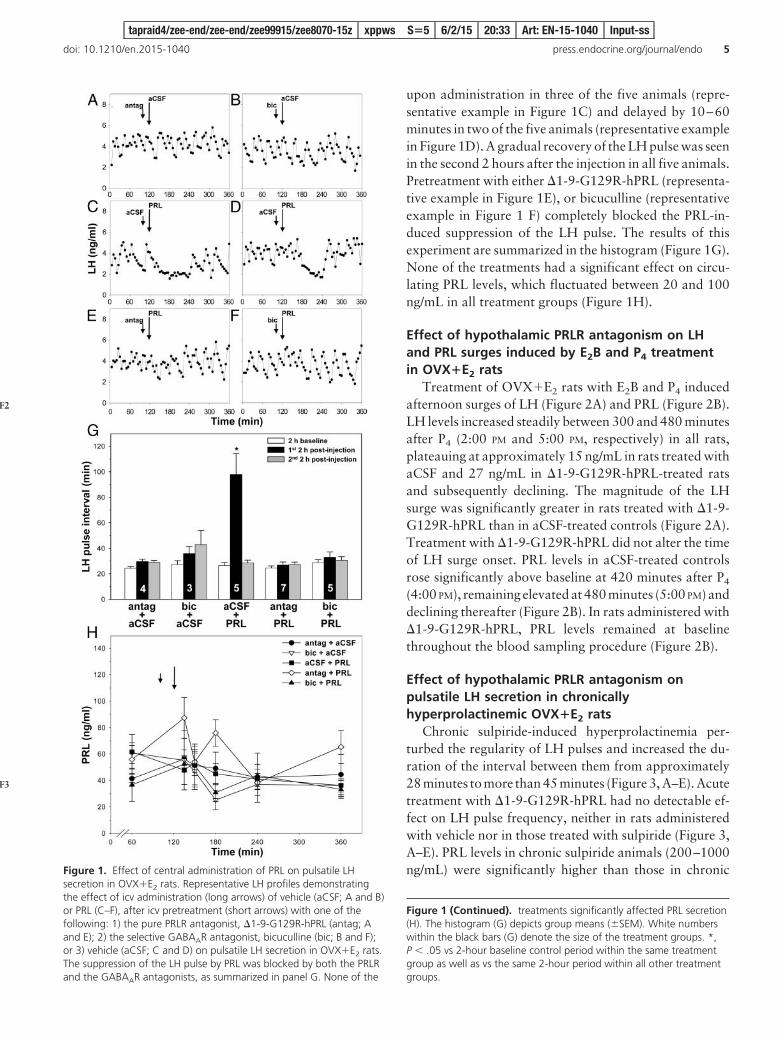
upon administration in three of the five animals (repre-sentative example in Figure 1C) and delayed by 10–60minutes in two of the five animals (representative examplein Figure 1D). A gradual recovery of the LH pulse was seenin the second 2 hours after the injection in all five animals.Pretreatment with either �1-9-G129R-hPRL (representa-tive example in Figure 1E), or bicuculline (representativeexample in Figure 1 F) completely blocked the PRL-in-duced suppression of the LH pulse. The results of thisexperiment are summarized in the histogram (Figure 1G).None of the treatments had a significant effect on circu-lating PRL levels, which fluctuated between 20 and 100ng/mL in all treatment groups (Figure 1H).
Effect of hypothalamic PRLR antagonism on LHand PRL surges induced by E2B and P4 treatmentin OVX�E2 rats
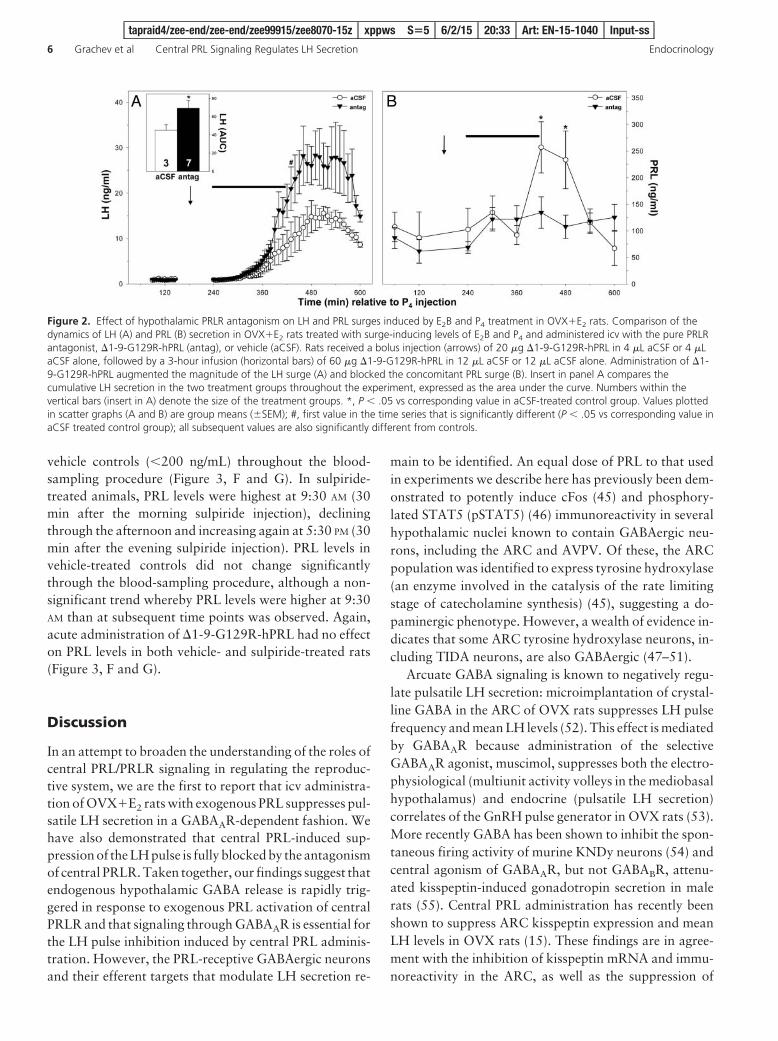
Treatment of OVX�E2 rats with E2B and P4 inducedafternoon surges of LH (Figure 2A) and PRL (Figure 2B).LH levels increased steadily between 300 and 480 minutesafter P4 (2:00 PM and 5:00 PM, respectively) in all rats,plateauing at approximately 15 ng/mL in rats treated withaCSF and 27 ng/mL in �1-9-G129R-hPRL-treated ratsand subsequently declining. The magnitude of the LHsurge was significantly greater in rats treated with �1-9-G129R-hPRL than in aCSF-treated controls (Figure 2A).Treatment with �1-9-G129R-hPRL did not alter the timeof LH surge onset. PRL levels in aCSF-treated controlsrose significantly above baseline at 420 minutes after P4
(4:00 PM), remaining elevated at 480 minutes (5:00 PM) anddeclining thereafter (Figure 2B). In rats administered with�1-9-G129R-hPRL, PRL levels remained at baselinethroughout the blood sampling procedure (Figure 2B).
Effect of hypothalamic PRLR antagonism onpulsatile LH secretion in chronicallyhyperprolactinemic OVX�E2 rats
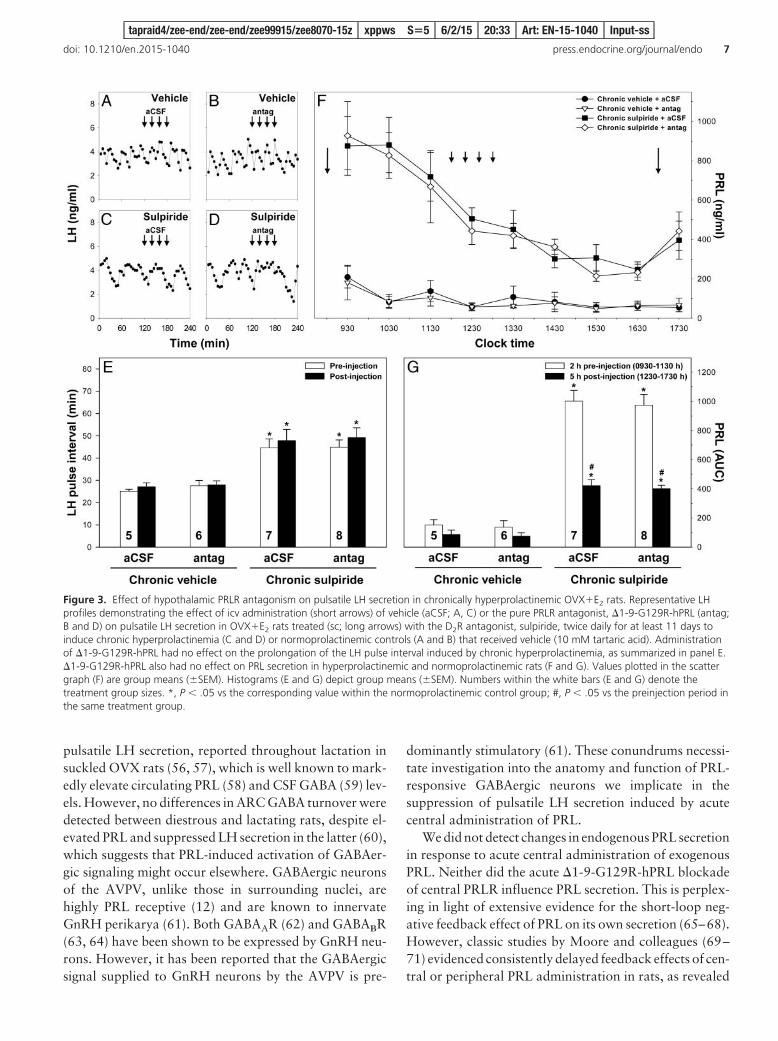
Chronic sulpiride-induced hyperprolactinemia per-turbed the regularity of LH pulses and increased the du-ration of the interval between them from approximately28 minutes to more than 45 minutes (Figure 3, A–E). Acutetreatment with �1-9-G129R-hPRL had no detectable ef-fect on LH pulse frequency, neither in rats administeredwith vehicle nor in those treated with sulpiride (Figure 3,A–E). PRL levels in chronic sulpiride animals (200–1000ng/mL) were significantly higher than those in chronicFigure 1. Effect of central administration of PRL on pulsatile LH
secretion in OVX�E2 rats. Representative LH profiles demonstratingthe effect of icv administration (long arrows) of vehicle (aCSF; A and B)or PRL (C–F), after icv pretreatment (short arrows) with one of thefollowing: 1) the pure PRLR antagonist, �1-9-G129R-hPRL (antag; Aand E); 2) the selective GABAAR antagonist, bicuculline (bic; B and F);or 3) vehicle (aCSF; C and D) on pulsatile LH secretion in OVX�E2 rats.The suppression of the LH pulse by PRL was blocked by both the PRLRand the GABAAR antagonists, as summarized in panel G. None of the
Figure 1 (Continued). treatments significantly affected PRL secretion(H). The histogram (G) depicts group means (�SEM). White numberswithin the black bars (G) denote the size of the treatment groups. *,P .05 vs 2-hour baseline control period within the same treatmentgroup as well as vs the same 2-hour period within all other treatmentgroups.
tapraid4/zee-end/zee-end/zee99915/zee8070-15z xppws S�5 6/2/15 20:33 Art: EN-15-1040 Input-ss
doi: 10.1210/en.2015-1040 press.endocrine.org/journal/endo 5
F2
F3
vehicle controls (200 ng/mL) throughout the blood-sampling procedure (Figure 3, F and G). In sulpiride-treated animals, PRL levels were highest at 9:30 AM (30min after the morning sulpiride injection), decliningthrough the afternoon and increasing again at 5:30 PM (30min after the evening sulpiride injection). PRL levels invehicle-treated controls did not change significantlythrough the blood-sampling procedure, although a non-significant trend whereby PRL levels were higher at 9:30AM than at subsequent time points was observed. Again,acute administration of �1-9-G129R-hPRL had no effecton PRL levels in both vehicle- and sulpiride-treated rats(Figure 3, F and G).
Discussion
In an attempt to broaden the understanding of the roles ofcentral PRL/PRLR signaling in regulating the reproduc-tive system, we are the first to report that icv administra-tion of OVX�E2 rats with exogenous PRL suppresses pul-satile LH secretion in a GABAAR-dependent fashion. Wehave also demonstrated that central PRL-induced sup-pression of the LH pulse is fully blocked by the antagonismof central PRLR. Taken together, our findings suggest thatendogenous hypothalamic GABA release is rapidly trig-gered in response to exogenous PRL activation of centralPRLR and that signaling through GABAAR is essential forthe LH pulse inhibition induced by central PRL adminis-tration. However, the PRL-receptive GABAergic neuronsand their efferent targets that modulate LH secretion re-
main to be identified. An equal dose of PRL to that usedin experiments we describe here has previously been dem-onstrated to potently induce cFos (45) and phosphory-lated STAT5 (pSTAT5) (46) immunoreactivity in severalhypothalamic nuclei known to contain GABAergic neu-rons, including the ARC and AVPV. Of these, the ARCpopulation was identified to express tyrosine hydroxylase(an enzyme involved in the catalysis of the rate limitingstage of catecholamine synthesis) (45), suggesting a do-paminergic phenotype. However, a wealth of evidence in-dicates that some ARC tyrosine hydroxylase neurons, in-cluding TIDA neurons, are also GABAergic (47–51).
Arcuate GABA signaling is known to negatively regu-late pulsatile LH secretion: microimplantation of crystal-line GABA in the ARC of OVX rats suppresses LH pulsefrequency and mean LH levels (52). This effect is mediatedby GABAAR because administration of the selectiveGABAAR agonist, muscimol, suppresses both the electro-physiological (multiunit activity volleys in the mediobasalhypothalamus) and endocrine (pulsatile LH secretion)correlates of the GnRH pulse generator in OVX rats (53).More recently GABA has been shown to inhibit the spon-taneous firing activity of murine KNDy neurons (54) andcentral agonism of GABAAR, but not GABABR, attenu-ated kisspeptin-induced gonadotropin secretion in malerats (55). Central PRL administration has recently beenshown to suppress ARC kisspeptin expression and meanLH levels in OVX rats (15). These findings are in agree-ment with the inhibition of kisspeptin mRNA and immu-noreactivity in the ARC, as well as the suppression of
Figure 2. Effect of hypothalamic PRLR antagonism on LH and PRL surges induced by E2B and P4 treatment in OVX�E2 rats. Comparison of thedynamics of LH (A) and PRL (B) secretion in OVX�E2 rats treated with surge-inducing levels of E2B and P4 and administered icv with the pure PRLRantagonist, �1-9-G129R-hPRL (antag), or vehicle (aCSF). Rats received a bolus injection (arrows) of 20 �g �1-9-G129R-hPRL in 4 �L aCSF or 4 �LaCSF alone, followed by a 3-hour infusion (horizontal bars) of 60 �g �1-9-G129R-hPRL in 12 �L aCSF or 12 �L aCSF alone. Administration of �1-9-G129R-hPRL augmented the magnitude of the LH surge (A) and blocked the concomitant PRL surge (B). Insert in panel A compares thecumulative LH secretion in the two treatment groups throughout the experiment, expressed as the area under the curve. Numbers within thevertical bars (insert in A) denote the size of the treatment groups. *, P .05 vs corresponding value in aCSF-treated control group. Values plottedin scatter graphs (A and B) are group means (�SEM); #, first value in the time series that is significantly different (P .05 vs corresponding value inaCSF treated control group); all subsequent values are also significantly different from controls.
tapraid4/zee-end/zee-end/zee99915/zee8070-15z xppws S�5 6/2/15 20:33 Art: EN-15-1040 Input-ss
6 Grachev et al Central PRL Signaling Regulates LH Secretion Endocrinology
pulsatile LH secretion, reported throughout lactation insuckled OVX rats (56, 57), which is well known to mark-edly elevate circulating PRL (58) and CSF GABA (59) lev-els. However, no differences in ARC GABA turnover weredetected between diestrous and lactating rats, despite el-evated PRL and suppressed LH secretion in the latter (60),which suggests that PRL-induced activation of GABAer-gic signaling might occur elsewhere. GABAergic neuronsof the AVPV, unlike those in surrounding nuclei, arehighly PRL receptive (12) and are known to innervateGnRH perikarya (61). Both GABAAR (62) and GABABR(63, 64) have been shown to be expressed by GnRH neu-rons. However, it has been reported that the GABAergicsignal supplied to GnRH neurons by the AVPV is pre-
dominantly stimulatory (61). These conundrums necessi-tate investigation into the anatomy and function of PRL-responsive GABAergic neurons we implicate in thesuppression of pulsatile LH secretion induced by acutecentral administration of PRL.
We did not detect changes in endogenous PRL secretionin response to acute central administration of exogenousPRL. Neither did the acute �1-9-G129R-hPRL blockadeof central PRLR influence PRL secretion. This is perplex-ing in light of extensive evidence for the short-loop neg-ative feedback effect of PRL on its own secretion (65–68).However, classic studies by Moore and colleagues (69–71) evidenced consistently delayed feedback effects of cen-tral or peripheral PRL administration in rats, as revealed
Figure 3. Effect of hypothalamic PRLR antagonism on pulsatile LH secretion in chronically hyperprolactinemic OVX�E2 rats. Representative LHprofiles demonstrating the effect of icv administration (short arrows) of vehicle (aCSF; A, C) or the pure PRLR antagonist, �1-9-G129R-hPRL (antag;B and D) on pulsatile LH secretion in OVX�E2 rats treated (sc; long arrows) with the D2R antagonist, sulpiride, twice daily for at least 11 days toinduce chronic hyperprolactinemia (C and D) or normoprolactinemic controls (A and B) that received vehicle (10 mM tartaric acid). Administrationof �1-9-G129R-hPRL had no effect on the prolongation of the LH pulse interval induced by chronic hyperprolactinemia, as summarized in panel E.�1-9-G129R-hPRL also had no effect on PRL secretion in hyperprolactinemic and normoprolactinemic rats (F and G). Values plotted in the scattergraph (F) are group means (�SEM). Histograms (E and G) depict group means (�SEM). Numbers within the white bars (E and G) denote thetreatment group sizes. *, P .05 vs the corresponding value within the normoprolactinemic control group; #, P .05 vs the preinjection period inthe same treatment group.
tapraid4/zee-end/zee-end/zee99915/zee8070-15z xppws S�5 6/2/15 20:33 Art: EN-15-1040 Input-ss
doi: 10.1210/en.2015-1040 press.endocrine.org/journal/endo 7

by median eminence dopamine synthesis and turnover in-creasing significantly as long as 12–24 hours after treat-ment. Moore and colleagues (72, 73) subsequently re-vealed a more rapid activation of PRL feedback in animalswith basal PRL secretion suppressed by bromocriptine(D2R agonist) treatment or hypophysectomy, althougheven that was detectable only after 4 hours. Thus, theabsence of a short-loop negative feedback effect of centralPRLR activation, or that of their blockade, on circulatingPRL levels in normoprolactinemic OVX�E2 rats withinthe examined time frame is expected.
Having established a tentative mechanism for PRL-in-duced suppression of pulsatile LH secretion and con-firmed the pure antagonistic properties of �1-9-G129R-hPRL in our experimental setting, we interrogated thehypothesis that PRL found endogenously within the cen-tral nervous system modulates LH and PRL surges. Therehave been several prior attempts to investigate the involve-ment of central PRL signaling in the regulation of the LHsurge (16, 74, 75). However, these studies involved theaugmentation of PRL signaling beyond that found underphysiologic conditions. Here we demonstrate that block-ade of central PRLR augments the magnitude of the LHsurges induced by E2B and P4 treatment, without alteringthe time of its onset, and blocks the concomitant PRLsurge in normoprolactinemic OVX�E2 rats. Our findingsraise the possibility that the endogenous PRL surge pro-vides restraint of the LH surge by hindering hyperactiva-tion of hypothalamic circuitry involved in its induction.Indeed, there appears to be an inverse relationship be-tween the magnitudes of the LH and PRL surges, as re-vealed by pharmacological manipulation of P4 signaling(76). However, complete suppression of endogenous PRLsurges by treatment with bromocriptine had no effect onthe concomitant LH surge in proestrous rats (77). Hence,the disinhibition of LH surge magnitude after central �1-9-G129R-hPRL administration is unlikely to be conse-quential to PRL surge suppression per se but is perhapsrather the effect of restricted access of residual endogenousPRL to central PRLR.
Because few GnRH neurons express PRLR (12), thedisinhibition of LH surge magnitude resulting fromthe acute antagonism of central PRLR could involve theblockade of direct prolactinergic suppression of AVPVkisspeptin neurons and/or that of prolactinergic stimula-tion of afferents that regulate the excitability of GnRHneurons, eg, GABAergic AVPV neurons (12, 78, 79). Al-ternatively, blockade of central PRLR (and/or that of thePRL surge) by �1-9-G129R-hPRL could augment themagnitudeof theLHsurgebydiminishingPRL-stimulateddopaminergic activity in the hypothalamus (80). Indeed,more than a third of GnRH neurons have been demon-
strated to be subject to tonic inhibition by dopamine (78)that could originate from ARC TIDA neurons (81, 82) orGABAergic kisspeptin neurons in the AVPV (79, 83). Fur-thermore, P4 is known to allosterically potentiate the ef-fects of GABA on GABAAR (84). Although all animalsreceived the same dose of exogenous P4, antagonism ofcentral PRLR could have interfered with E2B-induced ste-roidogenesis by hypothalamic astrocytes (24–26), effec-tively enhancing the amplitude of the LH surge (76). In-deed, neurosteroid biosynthesis has been shown to beregulated by PRL in newts (85), although this phenome-non has not yet been explored in mammals. Notwith-standing, studies in mammals implicate GABA/GABAARsignaling in modulating neurosteroidogenesis (86, 87). Be-cause we witnessed the essential role of hypothalamicGABA/GABAAR signaling in mediating the effects of PRLon LH secretion, evidence substantiated by the apparentlyPRL-receptive nature of hypothalamic GABA neurons(12), it is logical to speculate that PRL regulation of denovo progesterone synthesis by neurons and glia could bemediated by GABAergic afferents (87).
Another potential mechanism that might underlie the�1-9-G129R-hPRL-induced amplification of the LHsurge involves changes in the sensitivity of noradrenergicneurons of the locus coeruleus (LC) to P4. These neuronsdown-regulate their expression of P4 receptor on proestrusafternoon (88), concomitant with the PRL surge and in-creased cFos expression in the LC (89), and are known toprovide stimulatory noradrenergic input to GnRH neu-rons during the LH surge (90, 91). Because no data areavailable on the PRL sensitivity of the noradrenergic LC,it is unclear at what level PRL or �1-9-G129R-hPRLwould be able to modulate these pathways. However, re-cent evidence again implicates the highly PRL-receptive(12) AVPV neurons (92). These hypotheses require urgentinvestigation.
Although pretreatment with �1-9-G129R-hPRL aug-mented the magnitude of the LH surge, it had no effect onits time of onset. Similar observations were reported inGnRH-primed, prostaglandin F2�-treated lactating yaks,in which suppression of PRL secretion with quinagolide(another D2R agonist) augmented the magnitude of LHsurges induced by GnRH administration but did not affectits onset (93). The onset of the preovulatory LH surge innocturnal rodents is timed to coincide with the reductionin daylight by the circadian pacemaker within the supra-chiasmatic nucleus (SCN) the hypothalamus (94). Argi-nine vasopressin neurons of the SCN markedly increasethe sensitivity of GnRH neurons to stimulation by AVPVkisspeptin neurons and disinhibit GnRH secretion fromtonic suppression by RF-amide related peptide 3 neuronsin a time-gated fashion (95). We expect the association
tapraid4/zee-end/zee-end/zee99915/zee8070-15z xppws S�5 6/2/15 20:33 Art: EN-15-1040 Input-ss
8 Grachev et al Central PRL Signaling Regulates LH Secretion Endocrinology
AQ: 8
AQ: 9

between disruption of central PRL/PRLR signaling andaugmentation of the magnitude of the LH surge without achange in its timing, which we and others (93) have dem-onstrated, to reflect the suppressive effect of a centrallypenetrating PRL tone on the ability of AVPV kisspeptinneurons to elicit GnRH release during a time window gov-erned by neurons of the SCN, without affecting the time-restricting function of the SCN neurons per se. Indeed,AVPV kisspeptin neurons (12), but not SCN neurons (96),have been shown to express PRLR, confirming that thecircadian pacemaker is not subject to regulation by centralprolactin.
The absence of PRL surges in �1-9-G129R-hPRL-treated animals is perhaps best explained by the inabilityof centrally transported PRL to activate the positive oxy-tocin-PRL feedback loop recently suggested by Kennettand McKee (97). In our experiments, both control and�1-9-G129R-hPRL-treated rats were exposed to P4 levelscapable of suppressing TIDA neuron dopamine outputand thereby permissive to PRL surge generation (98–101).However, blockade of central PRLR presumably extendedto those expressed by hypothalamic oxytocin neurons(102) that secrete oxytocin in response to elevated E2 (103,104) and PRL (97, 105) levels. Oxytocin is a potent PRLsecretagogue (106, 107), its stimulatory effect on PRL se-cretion is augmented during proestrus (108), and oxytocinantagonists block PRL surges (109–111). Thus, �1-9-G129R-hPRL-treated, but not control rats, were probablydeprived of oxytocinergic stimulation of surge release ofPRL. An alternative hypothesis implicates GABA/GABAAR signaling: central administration of bicucullinehas been shown to block afternoon surges of PRL by sup-pressing the activity of TIDA neurons (112) and to aug-ment LH surge magnitude without advancing its onset(113) in rats. Furthermore, we have shown that icv pre-treatment with either �1-9-G129R-hPRL or bicucullinerestores LH pulsatility suppressed by acute central PRLadministration, confirming the close functional relation-ship between hypothalamic PRL/PRLR and GABA/GABAAR signaling implied by the PRL-receptive anatomyof GABAergic afferents to GnRH neurons (12).
These findings are consistent with a role for endoge-nous PRL in attenuating the extent of GnRH neurosecre-tion underlying surges of LH by directly stimulating hy-pothalamic release of GABA and highlight the importanceof central PRL/PRLR signaling in stimulating its own se-cretion through positive-feedback effects on hypotha-lamic GABA neurons (that suppress TIDA inhibition ofPRL secretion) and/or oxytocin neurons (which stimulatepituitary PRL release). Future studies should confirmwhether the mechanism of �1-9-G129R-hPRL-inducedblockade of the PRL surge involves the disruption of pos-
itive oxytocin-PRL feedback and/or that of GABAergicdisinhibition of PRL secretion from the dopamine tone.
Hyperprolactinemia, including that induced by anti-psychotic treatment, suppresses reproductive functionand, paradoxically, the only effective means of treatmentare dopamine receptor agonists, which exacerbate psy-chotic symptoms and are therefore contraindicated (114–116). Having demonstrated that central administration ofPRL inhibits the pulsatile secretion of LH in normopro-lactinemic OVX�E2 rats, we reasoned that central PRLRmight resemble a useful therapeutic target for the man-agement of reproductive defects caused by antipsychotic-induced hyperprolactinemia. To test this hypothesis, weadministered OVX�E2 rats receiving long-term twice-daily sulpiride treatment with �1-9-G129R-hPRL into thecerebroventricular system. We confirmed previous obser-vations (16) that chronic administration of sulpiride is aneffective means of inducing hyperprolactinemia and re-ducing the frequency of LH pulses in OVX�E2 rats. How-ever, four consecutive injections of 20 �g �1-9-G129R-hPRL given every 20 minutes neither altered the PRL levelselevated by chronic exposure to sulpiride nor reversed theinhibitory effect of hyperprolactinemia on LH pulse fre-quency. These data demonstrate that acute blockade ofcentral PRLR is not sufficient to reverse the effect of long-term hyperprolactinemia on pulsatile LH secretion.
Relative to the long-term changes in the chemistry ofARC neurons induced by chronic hyperprolactinemia(117), the limited duration of PRLR antagonism providedby acute icv administration of �1-9-G129R-hPRL mightbe sufficient to transiently disinhibit ARC kisspeptin ex-pression from tonic PRL-induced suppression (15). How-ever, if that were the case, such changes would be detect-able only several hours after treatment and would notnecessarily restore the LH pulse to frequencies observed innormoprolactinemic controls. In the context of the widerliterature, these data suggest that the mechanism by whichchronic hyperprolactinemia perturbs reproductive func-tion involves long-term genomic changes in the cross talkbetween populations of PRL-sensitive hypothalamic neu-rons, which are not reversible by the acute blockade ofcentral PRLR. To further address the contribution of cen-tral PRLR to the suppressive effects of hyperprolactinemiaon reproductive function, selective inducible knockdownof PRLR within discrete hypothalamic nuclei in chroni-cally hyperprolactinemic rats is advised.
In summary, we have shown that the activation of cen-tral PRLR leads to the acute suppression of LH pulsatilityand demonstrated that GABA/GABAAR signaling is anindispensable part of the mechanism involved. These dataraise the possibility that PRL-receptive GABAergic neu-rons may mediate inhibition of fertility attributed to ele-
tapraid4/zee-end/zee-end/zee99915/zee8070-15z xppws S�5 6/2/15 20:33 Art: EN-15-1040 Input-ss
doi: 10.1210/en.2015-1040 press.endocrine.org/journal/endo 9

vated PRL. Moreover, we provide the first evidence thatcentral PRL signaling provides tonic inhibition of the LHsurge and is necessary for the generation of the PRL surgethat tends to coincide with it. Finally, we report a lack ofeffect of acute blockade of central PRLR on pulsatile LHsecretion and PRL levels in hyperprolactinemic rats. Theseobservations assist the interpretation of and provide a con-text to previous reports on the role of PRL in modulatinghypothalamic circuits important in regulating mammalianreproduction.
Acknowledgments
We thank Florence Boutillon (INSERM, Paris, France) for over-seeing the synthesis of �1-9-G129R-hPRL, Rebecca Gresham(King’s College London, London, United Kingdom) for help withthe sulpiride administration, Sheng Yun Li and Ming Han Hu(King’s College London) for assistance with the surgeries and theLH RIA, Dave Grattan (University of Otago, Dunedin, NewZealand) for the PRL RIA protocol, Domingo Tortonese (Uni-versity of Bristol, Bristol, United Kingdom) for helpful commentson the figures, and Al Parlow (National Hormone and PeptideProgram, Torrance, California) for providing the LH and PRLRIA reagents.
Address all correspondence and requests for reprints to:Dr. Pasha Grachev, West Virginia University, Robert C. ByrdHealth Sciences Center, Room 113-BMRF, 1 Medical CenterDrive, PO Box 9229, Morgantown, WV 26506-9229. E-mail:[email protected].
P.G. is a recipient of the Medical Research Council (UnitedKingdom) Centenary Early Career Award. The supporter wasnot involved in the study design, the acquisition and interpreta-tion of data, the decision to publish the work, or the preparationof the manuscript.
Disclosure Summary: The authors have nothing to disclose.
References
1. Tyson JE, Friesen HG, Anderson MS. Human lactational and ovar-ian response to endogenous prolactin release. Science. 1972;177:897–900.
2. McNeilly AS. Prolactin and the control of gonadotrophin secretionin the female. J Reprod Fertil. 1980;58:537–549.
3. Walsh RJ, Slaby FJ, Posner BI. A receptor-mediated mechanism forthe transport of prolactin from blood to cerebrospinal fluid.Endocrinology. 1987;120:1846–1850.
4. Thompson SA. Localization of immunoreactive prolactin inependyma and circumventricular organs of rat brain. Cell TissueRes. 1982;225:79–93.
5. Binart N, Bachelot A, Bouilly J. Impact of prolactin receptor iso-forms on reproduction. Trends Endocrinol Metab. 2010;21:362–368.
6. Ferraris J, Bernichtein S, Pisera D, Goffin V. Use of prolactin re-ceptor antagonist to better understand prolactin regulation of pi-tuitary homeostasis. Neuroendocrinology. 2013;98:171–179.
7. Pi XJ, Grattan DR. Distribution of prolactin receptor immunore-activity in the brain of estrogen-treated, ovariectomized rats.J Comp Neurol. 1998;394:462–474.
8. Pi XJ, Grattan DR. Differential expression of the two forms ofprolactin receptor mRNA within microdissected hypothalamic nu-clei of the rat. Brain Res Mol Brain Res. 1998;59:1–12.
9. Pi XJ, Grattan DR. Increased expression of both short and longforms of prolactin receptor mRNA in hypothalamic nuclei of lac-tating rats. J Mol Endocrinol. 1999;23:13–22.
10. Pi XJ, Grattan DR. Increased prolactin receptor immunoreactivityin the hypothalamus of lactating rats. J Neuroendocrinol. 1999;11:693–705.
11. Di Carlo R, Muccioli G, Papotti M, Bussolati G. Characterizationof prolactin receptor in human brain and choroid plexus. BrainRes. 1992;570:341–346.
12. Kokay IC, Petersen SL, Grattan DR. Identification of prolactin-sensitive GABA and kisspeptin neurons in regions of the rat hypo-thalamus involved in the control of fertility. Endocrinology. 2011;152:526–535.
13. Wise PM. Effects of hyperprolactinemia on estrous cyclicity, serumluteinizing hormone, prolactin, estradiol, and progesterone con-centrations, and catecholamine activity in microdissected brain ar-eas. Endocrinology. 1986;118:1237–1245.
14. Pasqualini C, El Abed A, Kerdelhue B. Strain differences in neu-roendocrine responses to exogenous prolactin in the cycling femalerat. Neurosci Lett. 1981;22:271–277.
15. Araujo-Lopes R, Crampton JR, Aquino NS, et al. Prolactin regu-lates kisspeptin neurons in the arcuate nucleus to suppress LHsecretion in female rats. Endocrinology. 2014;155:1010–1020.
16. Anderson GM, Kieser DC, Steyn FJ, Grattan DR. Hypothalamicprolactin receptor messenger ribonucleic acid levels, prolactin sig-naling, and hyperprolactinemic inhibition of pulsatile luteinizinghormone secretion are dependent on estradiol. Endocrinology.2008;149:1562–1570.
17. Goudreau JL, Falls WM, Lookingland KJ, Moore KE. Periven-tricular-hypophysial dopaminergic neurons innervate the interme-diate but not the neural lobe of the rat pituitary gland. Neuroen-docrinology. 1995;62:147–154.
18. Ben-Jonathan N, LaPensee CR, LaPensee EW. What can we learnfrom rodents about prolactin in humans? Endocr Rev. 2008;29:1–41.
19. Freeman ME, Kanyicska B, Lerant A, Nagy G. Prolactin: structure,function, and regulation of secretion. Physiol Rev. 2000;80:1523–1631.
20. Sawai N, Iijima N, Takumi K, Matsumoto K, Ozawa H. Immu-nofluorescent histochemical and ultrastructural studies on the in-nervation of kisspeptin/neurokinin B neurons to tuberoinfundibu-lar dopaminergic neurons in the arcuate nucleus of rats. NeurosciRes. 2012;74:10–16.
21. Sawai N, Iijima N, Ozawa H, Matsuzaki T. Neurokinin B- andkisspeptin-positive fibers as well as tuberoinfundibular dopa-minergic neurons directly innervate periventricular hypophysealdopaminergic neurons in rats and mice. Neurosci Res. 2014;84:10 –18.
22. Fitzsimmons MD, Olschowka JA, Wiegand SJ, Hoffman GE.Interaction of opioid peptide-containing terminals with dopa-minergic perikarya in the rat hypothalamus. Brain Res. 1992;581:10 –18.
23. Szawka RE, Ribeiro AB, Leite CM, et al. Kisspeptin regulatesprolactin release through hypothalamic dopaminergic neurons.Endocrinology. 2010;151:3247–3257.
24. Micevych P, Sinchak K. Estradiol regulation of progesterone syn-thesis in the brain. Mol Cell Endocrinol. 2008;290:44–50.
25. Micevych P, Soma KK, Sinchak K. Neuroprogesterone: key to es-trogen positive feedback? Brain Res Rev. 2008;57:470–480.
26. Kuo J, Micevych P. Neurosteroids, trigger of the LH surge. J SteroidBiochem Mol Biol. 2012;131:57–65.
tapraid4/zee-end/zee-end/zee99915/zee8070-15z xppws S�5 6/2/15 20:33 Art: EN-15-1040 Input-ss
10 Grachev et al Central PRL Signaling Regulates LH Secretion Endocrinology
AQ:10,AQ:11
AQ: 12

27. Smith JT. Sex steroid regulation of kisspeptin circuits. Adv ExpMed Biol. 2013;784:275–295.
28. Miller BH, Takahashi JS. Central circadian control of female re-productive function. Front Endocrinol. 2013;4:195.
29. McGarrigle HH, Sarris S, Little V, Lawrence D, Radwanska E,Swyer GI. Induction of ovulation with clomiphene and humanchorionic gonadotrophin in women with hyperprolactinaemicamenorrhoea. Br J Obstet Gynaecol. 1978;85:692–697.
30. Oseko F, Morikawa K, Motohashi T, Aso T. Effects of chronicsulpiride-induced hyperprolactinemia on menstrual cycles of nor-mal women. Obstet Gynecol. 1988;72:267–271.
31. Liu X, Brown RS, Herbison AE, Grattan DR. Lactational anovu-lation in mice results from a selective loss of kisspeptin input toGnRH neurons. Endocrinology. 2014;155:193–203.
32. Brown RS, Herbison AE, Grattan DR. Prolactin regulation of kiss-peptin neurones in the mouse brain and its role in the lactation-induced suppression of kisspeptin expression. J Neuroendocrinol.2014;26:898–908.
33. Arbogast LA, Ben-Jonathan N. The preovulatory prolactin surge:an evaluation of the role of dopamine. Endocrinology. 1988;123:2690–2695.
34. Campbell BK, Mann GE, McNeilly AS, Baird DT. The pattern ofovarian inhibin, estradiol, and androstenedione secretion duringthe estrous cycle of the ewe. Endocrinology. 1990;127:227–235.
35. Djahanbakhch O, McNeilly AS, Warner PM, Swanston IA, BairdDT. Changes in plasma levels of prolactin, in relation to those ofFSH, oestradiol, androstenedione and progesterone around thepreovulatory surge of LH in women. Clin Endocrinol (Oxf). 1984;20:463–472.
36. Gaytan F, Bellido C, Morales C, Sanchez-Criado JE. Both prolactinand progesterone in proestrus are necessary for the induction ofapoptosis in the regressing corpus luteum of the rat. Biol Reprod.1998;59:1200–1206.
37. Freeman MC, Dupke KC, Croteau CM. Extinction of the estrogen-induced daily signal for LH release in the rat: a role for the proes-trous surge of progesterone. Endocrinology. 1976;99:223–229.
38. Cagampang FR, Maeda KI, Tsukamura H, Ohkura S, Ota K. In-volvement of ovarian steroids and endogenous opioids in the fast-ing-induced suppression of pulsatile LH release in ovariectomizedrats. J Endocrinol. 1991;129:321–328.
39. Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordinates.2nd ed. Sydney, Orlando, FL: Academic Press; 1986.
40. Li XF, Mitchell JC, Wood S, Coen CW, Lightman SL, O’Byrne KT.The effect of oestradiol and progesterone on hypoglycaemic stress-induced suppression of pulsatile luteinizing hormone release andon corticotropin-releasing hormone mRNA expression in the rat.J Neuroendocrinol. 2003;15:468–476.
41. Bernichtein S, Kayser C, Dillner K, et al. Development of pureprolactin receptor antagonists. J Biol Chem. 2003;278:35988–35999.
42. Paris N, Rentier-Delrue F, Defontaine A, et al. Bacterial productionand purification of recombinant human prolactin. Biotechnol ApplBiochem. 1990;12:436–449.
43. Neal-Perry GS, Zeevalk GD, Santoro NF, Etgen AM. Attenuationof preoptic area glutamate release correlates with reduced lutein-izing hormone secretion in middle-aged female rats. Endocrinol-ogy. 2005;146:4331–4339.
44. Van Cauter E. Estimating false-positive and false-negative errors inanalyses of hormonal pulsatility. Am J Physiol. 1988;254:E786–E794.
45. Cave BJ, Wakerley JB, Luckman SM, Tortonese DJ. Hypothalamictargets for prolactin: assessment of c-Fos induction in tyrosine hy-droxylase- and proopiomelanocortin-containing neurones in therat arcuate nucleus following acute central prolactin administra-tion. Neuroendocrinology. 2001;74:386–395.
46. Sapsford TJ, Kokay IC, Ostberg L, Bridges RS, Grattan DR. Dif-ferential sensitivity of specific neuronal populations of the rat hy-
pothalamus to prolactin action. J Comp Neurol. 2012;520:1062–1077.
47. Everitt BJ, Hokfelt T, Wu JY, Goldstein M. Coexistence of tyrosinehydroxylase-like and �-aminobutyric acid-like immunoreactivitiesin neurons of the arcuate nucleus. Neuroendocrinology. 1984;39:189–191.
48. Meister B, Hokfelt T. Peptide- and transmitter-containing neuronsin the mediobasal hypothalamus and their relation to GABAergicsystems: possible roles in control of prolactin and growth hormonesecretion. Synapse. 1988;2:585–605.
49. Meister B, Ceccatelli S, Hokfelt T, Anden NE, Anden M, The-odorsson E. Neurotransmitters, neuropeptides and binding sites inthe rat mediobasal hypothalamus: effects of monosodium gluta-mate (MSG) lesions. Exp Brain Res. 1989;76:343–368.
50. Kosaka T, Kosaka K, Hataguchi Y, et al. Catecholaminergic neu-rons containing GABA-like and/or glutamic acid decarboxylase-like immunoreactivities in various brain regions of the rat. ExpBrain Res. 1987;66:191–210.
51. Schimchowitsch S, Vuillez P, Tappaz ML, Klein MJ, Stoeckel ME.Systematic presence of GABA-immunoreactivity in the tubero-in-fundibular and tubero-hypophyseal dopaminergic axonal systems:an ultrastructural immunogold study on several mammals. ExpBrain Res. 1991;83:575–586.
52. Nishihara M, Kimura F. Roles of �-aminobutyric acid and sero-tonin in the arcuate nucleus in the control of prolactin and lutein-izing hormone secretion. Jap J Physiol. 1987;37:955–961.
53. Hiruma H, Sano A, Kimura F. Injection of bicuculline elicits firingof luteinizing hormone releasing hormone pulse generator in mus-cimol-treated ovariectomized rats. Brain Res. 1994;641:191–197.
54. Gottsch ML, Popa SM, Lawhorn JK, et al. Molecular properties ofKiss1 neurons in the arcuate nucleus of the mouse. Endocrinology.2011;152:4298–4309.
55. Garcia-Galiano D, Pineda R, Roa J, et al.Differential modulationof gonadotropin responses to kisspeptin by aminoacidergic, pep-tidergic, and nitric oxide neurotransmission. Am J Physiol Endo-crinol Metab. 2012;303:E1252–E1263.
56. Yamada S, Uenoyama Y, Kinoshita M, et al. Inhibition of metastin(kisspeptin-54)-GPR54 signaling in the arcuate nucleus-medianeminence region during lactation in rats. Endocrinology. 2007;148:2226–2232.
57. Yamada S, Uenoyama Y, Deura C, et al. Oestrogen-dependentsuppression of pulsatile luteinising hormone secretion and kiss1mRNA expression in the arcuate nucleus during late lactation inrats. J Neuroendocrinol. 2012;24:1234–1242.
58. Smith MS. The relative contribution of suckling and prolactin tothe inhibition of gonadotropin secretion during lactation in the rat.Biol Reprod. 1978;19:77–83.
59. Qureshi GA, Hansen S, Sodersten P. Offspring control of cerebro-spinal fluid GABA concentrations in lactating rats. Neurosci Lett.1987;75:85–88.
60. Kornblatt JJ, Grattan DR. Lactation alters �-aminobutyric acidneuronal activity in the hypothalamus and cerebral cortex in therat. Neuroendocrinology. 2001;73:175–184.
61. Liu X, Porteous R, d’Anglemont de Tassigny X, et al. Frequency-dependent recruitment of fast amino acid and slow neuropeptideneurotransmitter release controls gonadotropin-releasing hor-mone neuron excitability. J Neurosci. 2011;31:2421–2430.
62. Petersen SL, McCrone S, Coy D, Adelman JP, Mahan LC. GABA-Areceptor subunit mRNAs in cells of the preoptic area: colocaliza-tion with LHRH mRNA using dual-label in situ hybridization his-tochemistry. Endocrinol J. 1993;1:29–34.
63. Sliwowska JH, Billings HJ, Goodman RL, Lehman MN. Immu-nocytochemical colocalization of GABA-B receptor subunits in go-nadotropin-releasing hormone neurons of the sheep. Neurosci-ence. 2006;141:311–319.
64. Zhang C, Bosch MA, Ronnekleiv OK, Kelly MJ. �-Aminobutyricacid B receptor mediated inhibition of gonadotropin-releasing hor-
tapraid4/zee-end/zee-end/zee99915/zee8070-15z xppws S�5 6/2/15 20:33 Art: EN-15-1040 Input-ss
doi: 10.1210/en.2015-1040 press.endocrine.org/journal/endo 11

mone neurons is suppressed by kisspeptin-G protein-coupled re-ceptor 54 signaling. Endocrinology. 2009;150:2388–2394.
65. Gudelsky GA, Porter JC. Release of dopamine from tuberoin-fundibular neurons into pituitary stalk blood after prolactin orhaloperidol administration. Endocrinology. 1980;106:526–529.
66. Milenkovic L, Parlow AF, McCann SM. Physiological significanceof the negative short-loop feedback of prolactin. Neuroendocri-nology. 1990;52:389–392.
67. Curlewis JD, McNeilly AS. Prolactin short-loop feedback and pro-lactin inhibition of luteinizing hormone secretion during the breed-ing season and seasonal anoestrus in the ewe. Neuroendocrinology.1991;54:279–285.
68. Arbogast LA, Voogt JL. Prolactin (PRL) receptors are colocalizedin dopaminergic neurons in fetal hypothalamic cell cultures: effectof PRL on tyrosine hydroxylase activity. Endocrinology. 1997;138:3016–3023.
69. Annunziato L, Moore KE. Prolactin in CSF selectively increasesdopamine turnover in the median eminence. Life Sci. 1978;22:2037–2041.
70. Demarest KT, Moore KE. Sexual differences in the sensitivity oftuberoinfundibular dopamine neurons to the actions of prolactin.Neuroendocrinology. 1981;33:230–234.
71. Gudelsky GA, Simpkins J, Mueller GP, Meites J, Moore KE. Se-lective actions of prolactin on catecholamine turnover in the hy-pothalamus and on serum LH and FSH. Neuroendocrinology.1976;22:206–215.
72. Demarest KT, Riegle GD, Moore KE. Prolactin-induced activationof tuberoinfundibular dopaminergic neurons: evidence for both arapid ‘tonic’ and a delayed ‘induction’ component. Neuroendocri-nology. 1984;38:467–475.
73. Demarest KT, Riegle GD, Moore KE. The rapid ‘tonic’ and thedelayed ‘induction’ components of the prolactin-induced activa-tion of tuberoinfundibular dopaminergic neurons following thesystemic administration of prolactin. Neuroendocrinology. 1986;43:291–299.
74. Cohen-Becker IR, Selmanoff M, Wise PM. Inhibitory effects ofexogenously induced hyperprolactinemia on the endogenous cyclicrelease of luteinizing hormone and prolactin in the estrogen-primedovariectomized rat. Endocrinology. 1986;119:1718–1725.
75. Misztal T, Romanowicz K, Wankowska M, Wojcik-Gladysz A,Polkowska J. Does prolactin influence the hypothalamo-pituitaryGnRH-LH system in preovulatory-phase ewes? Reprod Biol. 2005;5:31–49.
76. Rao IM, Mahesh VB. Role of progesterone in the modulation of thepreovulatory surge of gonadotropins and ovulation in the pregnantmare’s serum gonadotropin-primed immature rat and the adult rat.Biol Reprod. 1986;35:1154–1161.
77. Witcher JA, Freeman ME. The proestrous surge of prolactin en-hances sexual receptivity in the rat. Biol Reprod. 1985;32:834–839.
78. Liu X, Herbison AE. Dopamine regulation of gonadotropin-re-leasing hormone neuron excitability in male and female mice.Endocrinology. 2013;154:340–350.
79. Clarkson J, Herbison AE. Dual phenotype kisspeptin-dopamineneurones of the rostral periventricular area of the third ventricleproject to gonadotrophin-releasing hormone neurones. J Neuroen-docrinol. 2011;23:293–301.
80. Pasqualini C, Bojda F, Gaudoux F, et al. Changes in tuberoin-fundibular dopaminergic neuron activity during the rat estrouscycle in relation to the prolactin surge: alteration by a mammarycarcinogen. Neuroendocrinology. 1988;48:320–327.
81. Mitchell V, Loyens A, Spergel DJ, et al. A confocal microscopicstudy of gonadotropin-releasing hormone (GnRH) neuron inputsto dopaminergic neurons containing estrogen receptor � in thearcuate nucleus of GnRH-green fluorescent protein transgenicmice. Neuroendocrinology. 2003;77:198–207.
82. Kuljis RO, Advis JP. Immunocytochemical and physiological evi-
dence of a synapse between dopamine- and luteinizing hormonereleasing hormone-containing neurons in the ewe median emi-nence. Endocrinology. 1989;124:1579–1581.
83. Horvath TL, Naftolin F, Leranth C. Luteinizing hormone-releasinghormone and gamma-aminobutyric acid neurons in the medial pre-optic area are synaptic targets of dopamine axons originating inanterior periventricular areas. J Neuroendocrinol. 1993;5:71–79.
84. Majewska MD. Neurosteroids: endogenous bimodal modulatorsof the GABA-A receptor. Mechanism of action and physiologicalsignificance. Prog Neurobiol. 1992;38:379–395.
85. Haraguchi S, Koyama T, Hasunuma I, Vaudry H, Tsutsui K. Pro-lactin increases the synthesis of 7�-hydroxypregnenolone, a keyfactor for induction of locomotor activity, in breeding male newts.Endocrinology. 2010;151:2211–2222.
86. Barbaccia ML, Roscetti G, Trabucchi M, et al. Isoniazid-inducedinhibition of GABAergic transmission enhances neurosteroid con-tent in the rat brain. Neuropharmacology. 1996;35:1299–1305.
87. Do Rego JL, Seong JY, Burel D, et al. Regulation of neurosteroidbiosynthesis by neurotransmitters and neuropeptides. Front En-docrinol. 2012;3:4.
88. Helena CV, de Oliveira Poletini M, Sanvitto GL, Hayashi S, FranciCR, Anselmo-Franci JA. Changes in �-estradiol receptor and pro-gesterone receptor expression in the locus coeruleus and preopticarea throughout the rat estrous cycle. J Endocrinol. 2006;188:155–165.
89. Martins-Afferri MP, Ferreira-Silva IA, Franci CR, Anselmo-FranciJA. LHRH release depends on locus coeruleus noradrenergic inputsto the medial preoptic area and median eminence. Brain Res Bull.2003;61:521–527.
90. Anselmo-Franci JA, Franci CR, Krulich L, Antunes-Rodrigues J,McCann SM. Locus coeruleus lesions decrease norepinephrine in-put into the medial preoptic area and medial basal hypothalamusand block the LH, FSH and prolactin preovulatory surge. BrainRes. 1997;767:289–296.
91. Helena CV, Franci CR, Anselmo-Franci JA. Luteinizing hormoneand luteinizing hormone-releasing hormone secretion is under lo-cus coeruleus control in female rats. Brain Res. 2002;955:245–252.
92. Szawka RE, Poletini MO, Leite CM, et al. Release of norepineph-rine in the preoptic area activates anteroventral periventricular nu-cleus neurons and stimulates the surge of luteinizing hormone.Endocrinology. 2013;154:363–374.
93. Sarkar M, Dutta Borah BK, Prakash BS. Efficacy of Ovsynch pro-tocol with antiprolactin treatment for timed artificial inseminationduring non-breeding seasons in yaks (Poephagus grunniens L.).Reprod Domest Anim. 2010;45:393–398.
94. Christian CA, Moenter SM. The neurobiology of preovulatory andestradiol-induced gonadotropin-releasing hormone surges. En-docr Rev. 2010.31:544–577.
95. Williams WP 3rd, Jarjisian SG, Mikkelsen JD, Kriegsfeld LJ. Cir-cadian control of kisspeptin and a gated GnRH response mediatethe preovulatory luteinizing hormone surge. Endocrinology. 2011;152:595–606.
96. Brown RS, Kokay IC, Herbison AE, Grattan DR. Distribution ofprolactin-responsive neurons in the mouse forebrain. J Comp Neu-rol. 2010;518:92–102.
97. Kennett JE, McKee DT. Oxytocin: an emerging regulator of pro-lactin secretion in the female rat. J Neuroendocrinol. 2012;24:403–412.
98. Arbogast LA, Voogt JL. Progesterone suppresses tyrosine hydrox-ylase messenger ribonucleic acid levels in the arcuate nucleus onproestrus. Endocrinology. 1994;135:343–350.
99. Arbogast LA, Voogt JL. Progesterone induces dephosphorylationand inactivation of tyrosine hydroxylase in rat hypothalamic do-paminergic neurons. Neuroendocrinology. 2002;75:273–281.
100. Yen SH, Pan JT. Progesterone advances the diurnal rhythm oftuberoinfundibular dopaminergic neuronal activity and the pro-
tapraid4/zee-end/zee-end/zee99915/zee8070-15z xppws S�5 6/2/15 20:33 Art: EN-15-1040 Input-ss
12 Grachev et al Central PRL Signaling Regulates LH Secretion Endocrinology

lactin surge in ovariectomized, estrogen-primed rats and in intactproestrous rats. Endocrinology. 1998;139:1602–1609.
101. Babu GN, Vijayan E. Hypothalamic tyrosine hydroxylase activityand plasma gonadotropin and prolactin levels in ovariectomized-steroid treated rats. Brain Res Bull. 1984;12:555–558.
102. Kokay IC, Bull PM, Davis RL, Ludwig M, Grattan DR. Expressionof the long form of the prolactin receptor in magnocellular oxy-tocin neurons is associated with specific prolactin regulation ofoxytocin neurons. Am J Physiol Regul Integr Comp Physiol. 2006;290:R1216–R1225.
103. Sarkar DK, Gibbs DM. Cyclic variation of oxytocin in the blood ofpituitary portal vessels of rats. Neuroendocrinology. 1984;39:481–483.
104. Yamaguchi K, Akaishi T, Negoro H. Effect of estrogen treatmenton plasma oxytocin and vasopressin in ovariectomized rats. En-docrinol Japon. 1979;26:197–205.
105. Donner N, Neumann ID. Effects of chronic intracerebral prolactinon the oxytocinergic and vasopressinergic system of virgin ovari-ectomized rats. Neuroendocrinology. 2009;90:315–322.
106. Egli M, Bertram R, Sellix MT, Freeman ME. Rhythmic secretionof prolactin in rats: action of oxytocin coordinated by vasoactiveintestinal polypeptide of suprachiasmatic nucleus origin. Endocri-nology. 2004;145:3386–3394.
107. Gonzalez-IglesiasAE,FletcherPA,Arias-CristanchoJA,etal.Directstim-ulatoryeffectsofoxytocin infemaleratgonadotrophsandsomatotrophsin vitro: comparison with lactotrophs. Endocrinology. 2015;156:600–612.
108. Tabak J, Gonzalez-Iglesias AE, Toporikova N, Bertram R, Free-man ME. Variations in the response of pituitary lactotrophs to
oxytocin during the rat estrous cycle. Endocrinology. 2010;151:1806–1813.
109. Samson WK, Lumpkin MD, McCann SM. Evidence for a physio-logical role for oxytocin in the control of prolactin secretion.Endocrinology. 1986;119:554–560.
110. Kennett JE, Poletini MO, Fitch CA, Freeman ME. Antagonism ofoxytocin prevents suckling- and estradiol-induced, but not pro-gesterone-induced, secretion of prolactin. Endocrinology. 2009;150:2292–2299.
111. Johnston CA, Negro-Vilar A. Role of oxytocin on prolactin secre-tion during proestrus and in different physiological or pharmaco-logical paradigms. Endocrinology. 1988;122:341–350.
112. Lee TY, Pan JT. Involvement of central GABAergic neurons inbasal and diurnal changes of tuberoinfundibular dopaminergicneuronal activity and prolactin secretion. Life Sci. 2001;68:1965–1975.
113. Neal-Perry GS, Zeevalk GD, Shu J, Etgen AM. Restoration of theluteinizing hormone surge in middle-aged female rats by alteringthe balance of GABA and glutamate transmission in the medialpreoptic area. Biol Reprod. 2008;79:878–888.
114. Perovich RM, Lieberman JA, Fleischhacker WW, Alvir J. The be-havioral toxicity of bromocriptine in patients with psychiatric ill-ness. J Clin Psychopharmacol. 1989;9:417–422.
115. Smith S. Neuroleptic-associated hyperprolactinemia. Can it betreated with bromocriptine? J Reprod Med. 1992;37:737–740.
116. Miller KK. Management of hyperprolactinemia in patients receiv-ing antipsychotics. CNS Spect. 2004;9:28–32.
117. Mohankumar PS, Mohankumar SM, Quadri SK, Voogt JL.Chronic hyperprolactinemia and changes in dopamine neurons.Brain Res Bull. 1997;42:435–441.
tapraid4/zee-end/zee-end/zee99915/zee8070-15z xppws S�5 6/2/15 20:33 Art: EN-15-1040 Input-ss
doi: 10.1210/en.2015-1040 press.endocrine.org/journal/endo 13