kernel hardness and baking quality of wheat — a genetic analysis using chromosome substitution...
TRANSCRIPT

Euphytica 25 (1976) 565-576
K E R N E L H A R D N E S S A N D B A K I N G Q U A L I T Y O F W H E A T - A G E N E T I C A N A L Y S I S U S I N G
C H R O M O S O M E S U B S T I T U T I O N L I N E S
G. J. D O E K E S and B. B E L D E R O K
Institute for Cereals, Flour and Bread TNO, Wageningen, the Netherlands
Received 15 April 1976
INDEX WORDS
Triticum aestivum, wheat, chromosome substitution lines, kernel hardness, baking absorption, dough stability, baking quality, genetic analysis.
SUMMARY
An attempt was made to identify the chromosomal location of genetic control of a few components of wheat quality, using chromosome substitution lines of Cappelle Desprez, Cheyenne, Hope, and Timstein into the recipient variety Chinese Spring.
Major factors for kernel hardness and increased baking absorption were found on chromosomes 5D of Cheyenne and Hope, and on 3B, 5D and 7D of Timstein. In Timstein, the presence of one of these chromosomes sufficed to make the wheat kernels hard.
Factors for favourable dough properties were identified on a few other chromosomes, different in various varieties. These were 1A of Cappelle Desprez and Cheyenne, 3B of Hope, and 2D of Timstein. All but one of these chromosomes showed an increase in loaf volume to a level in-between those of the recipient variety Chinese Spring and the donor varieties. No relationship was found between kernel hardness and dough-making and baking properties.
It was assumed that wheat quality is due to a combination of kernel hardness and favourable dough-making properties. As the genes for these factors are located on different chromosomes, it should not be too difficult to introduce both factors in existing varieties with poor baking properties. In a wheat breeding programme, the quality of new lines can be assessed in a rather simple way by determining kernel hardness and dough stability.
INTRODUCTION
Factors of wheat flour quality The quality of bread wheats (Triticum aestivum L.) cannot be expressed in terms of a single property, but depends on several milling, dough-making and baking character- istics. Besides flour yield, the most important factors involved are:
- Kernel hardness. Large differences in kernel hardness exist between various wheat varieties. Hardness is an important character for the miller, because hard wheats must be tempered to a higher moisture level and conditioned for a longer period of time before milling than soft wheats. Also the setting of the mill is different for both types of wheat. Moreover, hard wheat flours generally require more water ( baking absorption) to make a dough of proper consistency than soft wheat flours.
Protein content. This is an important flour property, because various dough-making and bread-baking properties are in one way or another related to protein content within a variety of wheat.
Dough properties. Flours of good baking quality form during mixing a tenacious
565

G . J . D O E K E S A N D B. B E L D E R O K
elastic dough with good gas retention properties. Such a dough has excellent handling qualities, and is not critical in its mixing and fermentation requirements. For this reason, it yields good bread over a wide range of baking conditions. Flours of poor baking quality form a soft, relatively non-elastic dough with poor gas retention. They yield doughs of inferior handling quality (which give trouble in machine baking) and are critical in their mixing and fermentation requirements so that they are more likely to fail in baking.
Loaf volume. A flour of good quality for bread-making has a good loaf volume potentiality (considering protein content). Also, it yields a loaf having a good outer appearance and a good internal crumb grain and colour.
Chromosomal location of genetic control of ~1,heat quality Substitution lines have proved to be a useful tool in determining the control of mor- phological and agricultural characteristics of wheat. However, only a few studies have been made of the chromosomal location of control of wheat quality character- istics. In fact, the reports have mainly come from two groups of workers : MORRIS and co-workers at Lincoln, Nebraska, USA, and WELSH and co-workers at Bozeman, Montana, USA.
MORRIS et al. (1966, 1968) developed a set of substitution lines of the donor variety Cheyenne into the recipient Chinese Spring. Cheyenne is a hard red winter wheat with desirable milling, dough-handling and baking properties, whereas Chinese Spring is a soft red spring wheat with poor milling, dough-handling and baking qual- ity. The chromosomes 7B and 5D were identified as carriers of control of high flour yield, and 5D was the only chromosome on which factors for kernel hardness were located (MATTERN et al., 1973). Major factors for the strong dough-mixing character- istics of Cheyenne were located on chromosomes 4B, 7B and 5D (MORRIS et al., 1966, 1968). Important contributions to loaf volume, crust appearance and crumb tex- ture were made by chromosome 1B, 4B, 7B, and possibly 1A. A depression of general quality characteristics was observed in the 1D substitution line.
WELSH et al. (1968) subjected three sets of hard red spring substitution lines to mill- ing and baking tests. The variety Chinese Spring was again used as the common genetic background; the three donors were Thatcher, Hope, and Timstein, strong quality wheat varieties with satisfactory flour yields.
Minor flour yield changes were effected by different chromosomes with 5D giving the greatest increase in the Hope set. In the Thatcher substitution set, only chromo- some 2A increased baking absorption and loaf volume. Several chromosomes pro- duced changes in the Hope set: chromosome 2B increased the dough stability, baking absorption was increased by different chromosomes with 5D giving the largest change over Chinese Spring, and loaf volume was increased by 2B and 3A. For Tim- stein, the authors reported quality differences in the case of 16 different chromosome substitutions with the most important effects coming from substituted chromosomes 3B and 5D (baking absorption), 1B, 3B and 6B (dough-handling properties) and 3B (loaf volume). Chromosome 1D produced a loaf volume significantly smaller than Chinese Spring.
A lack of agreement exists between the results of MORRIS et al. and those of WELSH et al. ; however, different wheat varieties were involved.
566 Euphytica 25 (1976)

GENE L O C A L I Z A T I O N OF W H E A T BREAD C H A R A C T E R S
Aim of the investigation In this paper, an at tempt has been made to find the chromosomal location of genetic control of a few components of wheat quality, using three of the chromosome sub- stitution sets already described in literature and one set not yet investigated for flour quality.
MATERIAL
Four substitution sets, Cappelle Desprez, Cheyenne, Hope, and Timstein, in Chinese Spring were used. Chinese Spring is a soft red spring wheat with poor milling and baking characteristics. The four donor varieties have satisfactory milling and baking properties. Cappelle Desprez is a soft red winter wheat; Cheyenne is a hard red winter wheat; Hope and Timstein are hard red spring wheats.
The substitution lines of Cappelle Desprez were made by the Plant Breeding In- stitute, Cambridge, England (LAw, 1968), those of Cheyenne originated from MORRIS et al. (1966) and those of Hope and Timstein were developed by SEARS et al. (1957).
The samples used in this study were obtained from Dr A. C. Zeven, Institute of Plant Breeding, Agricultural University, Wageningen, the Netherlands. The sub- stitution sets and parental varieties of Hope and Timstein were grown near Wage- ningen in 1972, those of Cappelle Desprez and Cheyenne in 1973. All sets were grown again in 1974. Of Cheyenne, the 2B and 1D substitutions were not available.
It should be mentioned that some of the lines have only had four backcrosses (not- ably in the Cheyenne and Timstein sets). As a consequence of this, we cannot be sure that a certain gene is not a contaminant from the donor in the recipient background.
METHODS
Milling The wheat was tempered to 15.5 % moisture and milled on a Brabender Quadrumat Jr. experimental mill with sieves removed. The milled stock was sieved on a Jel laboratory shaker with 150pro sieves.
Protein content Protein content was determined by a micro Kjeldahl method. Flour protein content
Kjeldahl nitrogen × 5.7.
Kernel hardness In this investigation, the starch damage content of the flour was used as a measure of kernel hardness. It is normal for flour to contain some starch granules that were dam- aged mechanically during milling. In a soft wheat flour, the starch damage is low, 5 7 % of dry matter ; in a hard wheat flour, more starch damage occurs, 8-12 % of dry matter. The starch damage content was determined by Method 76-30A of the Ap- proved Methods of the American Association of Cereal Chemists, St. Paul, 1969.
Baking absorption A modified MAES & PIROTTE ( 1 9 5 5 ) centrifuge method was used. In a tared centrifuge
Euphytica 25 (1976) 567

G. J, DOEKES AND B. BELDEROK
tube 1.5 g of flour was weighed out, an excess of water was added, and the suspension stirred in a mechanical stirrer with a glass rod for 6 min and centrifuged (750 g) for 30 min. Next, the supernatant was decanted and the weight increase of the sediment determined. Water absorptions found by this method show a linear relationship with baking absorptions determined by means of the farinograph (460 F.U. 1, with salt and yeast). The amount of water to be added to the flour may be calculated from the re- gression line to obtain doughs having the same consistency as in the standard baking test with 800-g loaves.
Dough characteristics Doughs (without salt and yeast) were mixed in a 50-g bowl of the Brabender Farino- graph at 30 °C. For most of the samples, insufficient quantities of flour were avail- able to determine the quantity of water required for a maximum consistency of 500 F,U. The farinograph water absorption of composite samples of hard and soft sub- stitution lines were 73 and 67 ~ , respectively. These water additions were used for the farinograph tests with all samples. Dough development time is defined in ICC Standard No 115, and stability is defined in Method 54-21 of the Approved Methods of the American Association of Cereal Chemists, St. Paul, 1969. A horizontal line through the centre of the band at its maximum was substituted for the 500 F.U. line.
Micro baking The equipment used in the micro baking test was designed and constructed at the in- stitute. The procedure of the test was as follows: 10-g flour samples were mixed with a micro pin-mixer (125 rev/min) with the appropriate amount of water, 2 ~ salt and
lo 2 2 ~ yeast for 3 min. The doughs were fermented at 30°C. First fermentation took 45 min. Then, dough pieces containing 8 g of flour were weighed and mechanically punched between a pair of rolls. Second fermentation took 15 min. The doughs were moulded by hand and panned. Final p roof took 60 min. Baking was performed in a small rotating reel-type oven at 210°C. In the oven, a wooden frame soaked with water was placed round the baking pan during the first 15 min of baking. The aim of the wooden frame was to shield off the oven heat and to prevent a too rapid forma- tion of the crust. After removal of the frame, the loaf was baked of f for a further 5 min.
Next, the loaves were coated with paraffin, and their volume measured via the buoyant force on the coated loaves in water (Archimedian principle). The volume was expressed in ml/100 g flour. A good correlation exists between the loaf volume determined in the micro baking-test and the loaf volume determined in the standard baking-test for 800-g loaves. In micro baking-tests, a reliable estimation of crust and crumb characteristics is hardly possible. Therefore, we confine ourselves to pres- enting the loaf volumes obtained.
The substitution lines and parental varieties were grown and harvested under fav- ourable weather conditions. Consequently, the diastatic activity of the flour was low. In baking, 0.5 ~ of barley malt was added to the flour to bring the diastatic activity to an opt imum level.
The micro baking-tests were made both without and with several additions of po-
1F.U. Far inograph Unit (defined in ICC Standard No. 115).
568 Euphytica 25 (1976)

G E N E L O C A L I Z A T I O N O F W H E A T B R E A D C H A R A C T E R S
tassium bromate to the flour. In Table 5 only the results of baking tests at the optimum bromate level (i.e. the level at which maximum loaf volumes are obtained) are pres- ented.
R E S U L T S A N D D I S C U S S I O N
All material grown in 1972 and 1973 was submitted to determinations of flour yield, protein content, kernel hardness and baking absorption, and to baking tests. As in- sufficient quantities of flour were left for rheological examination, the parental vari- ieties and the chromosome substitution lines were sown again in 1974, and later mill- ed and submitted to farinograph tests.
Flour yield As only a small volume of grain (100-200 g) was available, the Brabender Quadrumat Jr. experimental mill was used in our experiments. We doubt whether such a small mill is a suitable means of determining flour yield. Therefore, we present no detailed results and confine ourselves to the following comments.
Milling yield varied from 46 to 59 ~ . The highest values were obtained with the parental varieties Cappelle Desprez, Cheyenne, and Hope, the lowest values with the varieties Chinese Spring and Timstein. Most substitution lines yielded values com- parable to the poor milling recipient variety Chinese Spring. The exceptions were in the Cheyenne substitution set where the flour yields of 7B and 5D were at the same level of the high-yielding Cheyenne variety. This corroborates the finding of MAT- TERN et al. (1973) who also identified chromosomes 7B and 5D as major carriers of a high flour yield in the variety Cheyenne. No relationship was found between kernel hardness (see below) and flour yield, except for chromosome 5D of Cheyenne.
Protein content The protein contents of the flours of the parental varieties and the ranges and mean values found in the chromosome substitution lines are summarized in Table 1.
Only in one case, substitution line 5D of Cappelle Desprez, the protein content was significantly higher than that of the two parents, viz 19.5 ~ ; this could not be con- firmed in 1974. Therefore, the increased protein content of 5D of Cappelle Desprez grown in 1973 was assumed to be a chance variation.
It has long been accepted that the protein content of wheat largely depends on the nitrogen condititon of the soil and the length of the ripening period. The variation in protein content within each set of substitution lines as shown in Table 1 will have no genetical meaning and may be due to small variations in growing conditions in the small field plots.
Kernel hardness Table 2 shows that the starch damage content of the flours of the varieties Chinese Spring and Cappelle Desprez is low. This merely confirms the fact that these var- ieties are soft wheats. Accordingly, all chromosome substitution lines of Cappelle Desprez in Chinese Spring are soft wheats, having a low starch damage content.
The varieties Cheyenne, Hope and Timstein are hard wheats with a high degree
Euphytica 25 (1976) 569

G. J. D O E K E S A N D B. B E L D E R O K
Table 1. Protein content of flour ( ~ of dry matter) of chromosome substitution lines and parental varieties.
Parental Substitution lines varieties
range
Chinese Spring 15.6 - - Cappelle Desprez 14.4 13.8 19.5 16.5 Cheyenne 15.0 14.0-17.5 15.9 Hope 15.9 14.3-16.9 15.9 Timstein 14.5 14.718.0 16.1
Table 2. Starch damage of flour ( ~ of dry matter) of chromosome substitution lines and parental varieties.
Substitution Chinese Spring- Chinese Spring- lines and Cappelle Desp~z Cheyenne varieties 1973 1973
IA 6.2 5.8 8.0 2A 6.5 6.1 7.3 3A 6.5 6.3 6.9 4A 6.3 6.0 7.8 5A 5.8 6.1 7.5 6A 6.4 5.7 6.7 7A 6.5 6.0 7.1
1B 6.2 6.2 7.3 2B 6.7 - 6.7 3B 6.9 6.4 6.8 4B 6.5 6.0 7.3 5B 6.2 6.0 6.8 6B 6.0 6.2 7.5 7B 6.7 6.5 7.5
Chinese Spring- Chinese Spring- Hope Timstein 1972 1972
7.1 7.1 7.4 7.1 6.7 6.7 6.7
6.3 6.7
10.8 6.3 6.0 6.3 6.0
ID 6.5 7.7 8.0 2D 6.1 6.0 6.7 6.2 3D 6.3 5.4 6.9 7.4 4D 6.1 5.0 6.7 6.4 5D 6.5 8.8 9.9 11.0 6D 6.3 6.5 6.9 6.9 7D 6.4 6.4 7.1 11.8
Mean and con- fidence limits (P~0.05) 6.4 :£ 0.5 6.0 ± 0.8 7.2 ± 0.8
Chinese Spring 6.1 6.1 6.5 Cappelle Desprez 6.2 Cheyenne - 8.5 Hope 9.7 Timstein -
6.7 ± 1.2
6.5
10.6
570 Euphytica 25 (1976)

G E N E L O C A L I Z A T I O N OF W H E A T B R E A D C H A R A C T E R S
of starch damage in the flour. Table 2 shows clearly that in these varieties the factor kernel hardness, as measured by starch damage content, is located on chromosome 5D. Also MATTERN et al. (1973) found the control of kernel hardness located on chromosome 5D in Cheyenne.
An increased starch damage content was also found for chromosomes 3B and 7D of Timstein. However, the occurrence of only one hardness chromosome proved to be sufficient to make the recipient as hard as the donor. A slight, but significant in- crease was caused by chromosome 1D of Timstein. As already mentioned before, some lines of Timstein have only had four backcrosses and, in the absence of dupli- cates, we cannot be sure that a certain gene is not a contaminant from the donor in the recipient background. Thus, there is a possibility that the 3B, 1 D, 5D, and 7D effects are all caused by the same gene.
The inheritance of kernel hardness has been dealt with by SVMES (1965), who in crosses between soft and hard wheats demonstrated that in most cases a single gene was responsible for the hardness differences between the two parents, although mod- ifying genes could play a role, too. This is an important conclusion for the practical wheat breeder, for it should be relatively simple to convert a soft wheat into a hard wheat, and vice versa, by crossing and backcrossing.
Baking absorption The baking absorptions of the flours are given in Table 3. Very high baking absorp- tions were effected by the chromosomes 5D of Cheyenne and Hope, and 3B, 5D and 7D of Timstein. Minor effects were caused by the chromosomes 3B of Cappelle Des- prez and 7B of Cheyenne. A decreased baking absorption was found in the flour of the substitution line 6A of Timstein.
Comparison of Tables 2 and 3 shows that the chromosome substitution lines with a very high baking absorption (more than 60 ~ ) are all hard-grained wheats with a high degree of starch damage in the flour. Surprisingly, the donor wheats Cheyenne, Hope, and Timstein behaved differently in this respect: here, kernel hardness went along with a normal baking absorption of the flour. A possible explanation for this will be given in the discussion on the baking quality (see below).
Dough characteristics Wheats of good baking quality are generally characterized by long mixing times (long development times) and stable doughs.
In the farinograph tests, all but one of the substitution lines had short dough development times (mean value for all lines: 4.6 minutes) not significantly different from Chinese Spring (4.0 min). Only chromosome 1D of Cheyenne appeared to carry factors that extended the dough development time beyond that of Chinese Spring, viz to 14.5 min.
Table 4 shows that the variation in farinograph dough stability within each set of chromosome substitution lines was large. Consequently, the ranges between the lower and upper confidence limits (P < 0.05) were large, too. Nevertheless, some of the substitution lines exceeded these limits. A long dough stability was found in the chromosome substitution lines 1A of Cappelle Desprez and Cheyenne, 3B of Hope, and 2D of Timstein, and in the four donor varieties.
Euphytica 25 (1976) 571

G . J . D O E K E S A N D B. B E L D E R O K
Table 3. Baking absorption of flour (%) of chromosome substitution lines and parental varieties.
Substitution Chinese Spring- Chinese Spring- Chinese Spring- Chinese Spring- lines and Cappelle Desprez Cheyenne Hope Timstein varieties 1973 1973 1972 1972
1A 55.0 55.5 59.5 57.5 2A 56.0 56.0 59.5 56.0 3A 56.0 55.5 57.0 56.0 4A 55.5 56.0 58.0 57.0 5A 55.0 55.5 56.5 57.0 6A 55.5 54.5 57.5 53.5 7A 54.5 55.0 56.0 56.0
1B 55.5 54.5 57.0 57.5 2B 56.0 - 57.5 57.0 3B 58.0 55.5 57.5 62.5 4B 55.5 54.0 58.5 56.5 5B 55.0 55.0 57.5 56.0 6B 54.5 55.0 55.5 55.5 7B 55.5 57.5 55.5 56.5
1D 56.5 57.0 56.0 2D 56.0 54.0 57.5 56.5 3D 57.5 55.0 57.0 57.5 4D 55.0 53.0 56.0 56.0 5D 57.0 62.8 61.0 65.2 6D 56.0 55.0 56.5 58.0 7D 55.5 55.0 57.0 65.0
Mean and con- fidence limits (P<0.05) 55.7 ± 1.9 55.1 ± 1.9 57.2 ± 2.2 56.4 ± 2.0
Chinese Spring 54.0 54.0 57.5 57.5 Cappelle Desprez 53.5 Cheyenne 56.5 .... Hope 57.0 Timstein 57.0
Comparison of Tables 2 and 4 shows that there is no relationship between in- creased kernel hardness and favourable dough properties.
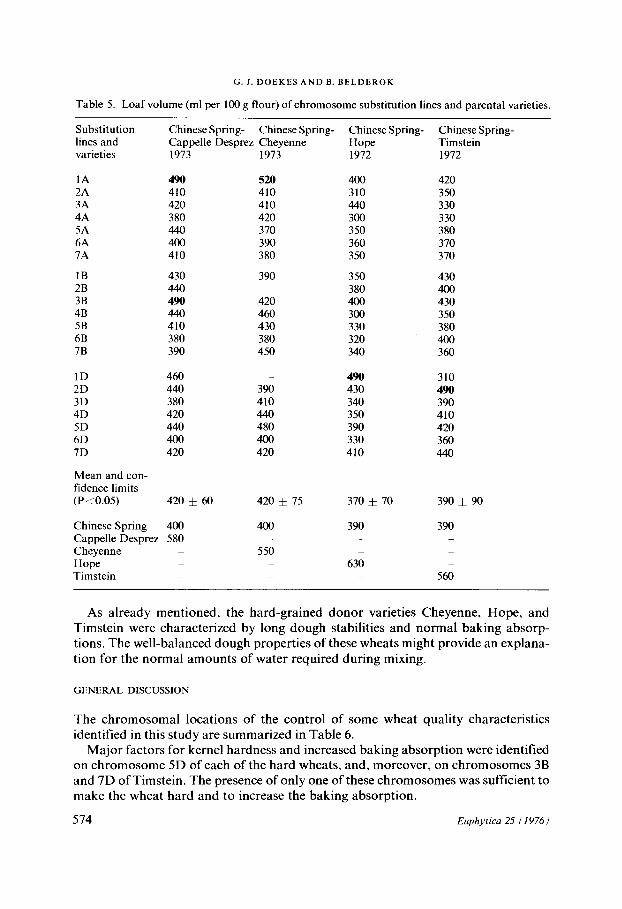
Baking quality The loaf volumes of the substitution lines and parental varieties are presented in Table 5.
All four donor varieties had a good baking quality; the loaf volumes obtained were considerably greater than the volume o f the recipient variety Chinese Spring. N o n e of the substitution lines reached a level o f baking quality as high as that of the donor varieties. Some had a loaf volume between those of the two parents, caused by dif- ferent chromosomes in the various sets o f substitution lines. In Cappelle Desprez, good baking quality was located on chromosomes 1A and 3B. In Cheyenne, only
572 Euphytica 25 (1976)
G E N E L O C A L I Z A T I O N OF W H E A T B R E A D C H A R A C T E R S
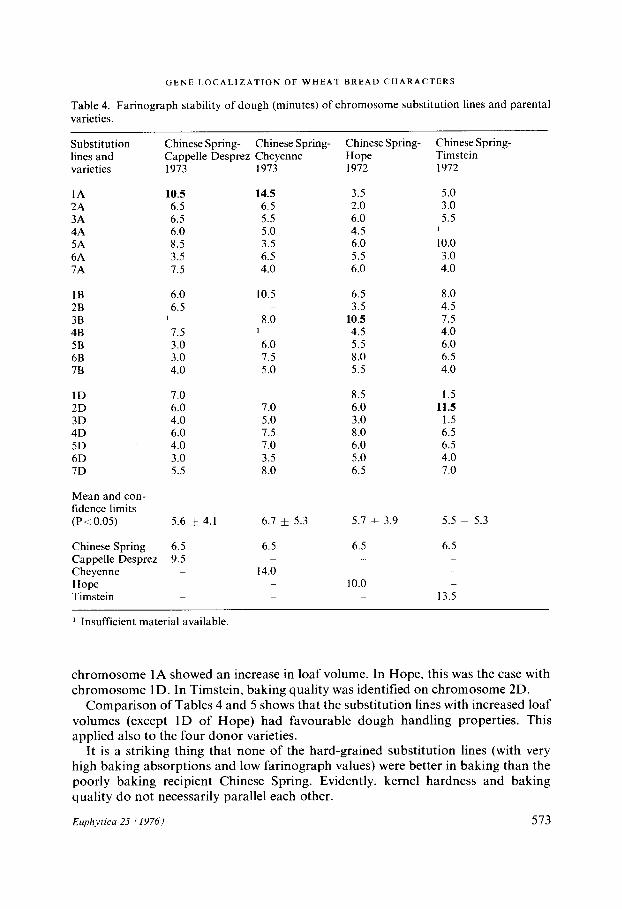
Table 4. Farinograph stability of dough (minutes) of chromosome substitution lines and parental varieties.
Substitution Chinese Spring- Chinese Spring- Chinese Spring- Chinese Spring- lines and Cappelle Desprez Cheyenne Hope Timstein varieties 1973 1973 1972 1972
1A 10.5 14.5 3.5 5.0 2A 6.5 6.5 2.0 3.0 3A 6.5 5.5 6.0 5.5 4A 6.0 5.0 4.5 1 5A 8.5 3.5 6.0 10.0 6A 3.5 6.5 5.5 3.0 7A 7.5 4.0 6.0 4.0
1B 6.0 10.5 6.5 8.0 2B 6.5 3.5 4.5 3B 1 8,0 10.5 7.5 4B 7.5 1 4.5 4.0 5B 3.0 6.0 5.5 6.0 6B 3.0 7.5 8.0 6.5 7B 4.0 5.0 5.5 4.0
ID 7.0 8.5 1.5 2D 6.0 7.0 6.0 11.5 3D 4.0 5.0 3.0 1.5 4D 6.0 7.5 8.0 6.5 5D 4.0 7.0 6.0 6.5 6D 3.0 3.5 5.0 4.0 7D 5.5 8.0 6.5 7.0
Mean and con- fidence limits (P~:.0.05) 5.6 ± 4.1 6.7 ± 5.3 5.7 ÷ 3.9 5.5 ± 5.3
Chinese Spring 6.5 6.5 6.5 6.5 Cappelle Desprez 9.5 --- Cheyenne 14.0 - Hope - 10.0 Timstein 13.5
1 Insufficient material available.
chromosome 1A showed an increase in loaf volume. In Hope, this was the case with chromosome 1D. In Timstein, baking quality was identified on chromosome 2D.
Comparison of Tables 4 and 5 shows that the substitution lines with increased loaf volumes (except 1D of Hope) had favourable dough handling properties. This applied also to the four donor varieties.
It is a striking thing that none of the hard-grained substitution lines (with very high baking absorptions and low farinograph values) were better in baking than the poorly baking recipient Chinese Spring. Evidently, kernel hardness and baking quality do not necessarily parallel each other.
Euphytica 25 (1976) 573
G. J. D O E K E S A N D B . B E L D E R O K
Table 5. Loaf volume (ml per 100 g flour) of chromosome substitution lines and parental varieties.
Substitution Chinese Spring- Chinese Spring- Chinese Spring- Chinese Spring- lines and Cappelle Desprez Cheyenne Hope Timstein varieties 1973 1973 1972 1972
1 A 4 9 0 5 2 0 400 420 2A 410 410 310 350 3A 420 410 440 330 4A 380 420 300 330 5A 440 370 350 380 6A 400 390 360 370 7A 410 380 350 370
1B 430 390 350 430 2B 440 380 400 3 B 490 420 400 430 4B 440 460 300 350 5B 410 430 330 380 6B 380 380 320 400 7B 390 450 340 360
1D 460 - 4 9 0 310 2D 440 390 430 4 9 0
3D 380 410 340 390 4D 420 440 350 410 5D 440 480 390 420 6D 400 400 330 360 7D 420 420 410 440
Mean and con- fidence limits (P<0.05) 420 + 60 420 ± 75 370 ± 70 390 ± 90
Chinese Spring 400 400 390 390 Cappetle Desprez 580 - - - Cheyenne - 550 Hope 630 Timstein 560
As already mentioned, the hard-grained donor varieties Cheyenne, Hope, and Timstein were characterized by long dough stabilities and normal baking absorp- tions. The well-balanced dough properties of these wheats might provide an explana- tion for the normal amounts of water required during mixing.
GENERAL DISCUSSION
The chromosomal locations of the control of some wheat quality characteristics identified in this study are summarized in Table 6.
Major factors for kernel hardness and increased baking absorption were identified on chromosome 5D of each of the hard wheats, and, moreover, on chromosomes 3B and 7D of Timstein. The presence of only one of these chromosomes was sufficient to make the wheat hard and to increase the baking absorption.
574 Euphytica 25 (1976)

GENE LOCALIZATION OF WHEAT BREAD CHARACTERS
Table 6. C h r o m o s o m a l locat ion o f whea t qual i ty characteris t ics .
Kerne l ha rdnes s Baking D o u g h L o a f v o l u m e absorp t ion stabil i ty
C a p p e l l e D e s p r e z 3B IA 1 _ 1A 3B C h e y e n n e 5 D - - 7B 5D 1A - 1A H o p e 5D 5D - 3B 1D Tims te in 3B 1D 5D 7D 3B 5D 7D - 2D 2D
1 Insuff ic ient mater ia l avai lable o f c h r o m o s o m e subs t i tu t ion line 3B.
Factors for dough stability were identified on the chromosomes 1A of Cappelle Desprez and Cheyenne, 3B of Hope, and 2D of Timstein. All but one of these chro- mosomes effected an increase in loaf volume to a level in-between those of the poor- baking recipient variety Chinese Spring and the well-baking donor varieties. Ap- parently, more than one factor is needed to reach the baking quality level of the donors.
Table 6 shows clearly that factors for dough-making and baking properties are located on other chromosomes than factors for kernel hardness and baking absorp- tion. It means, that these properties inherit independently.
The objection could be raised that a chromosomal effect observed may not be caused by the introduction of a donor chromosome, but by the mere removal of the Chinese Spring chromosome which might have a repressing effect on the character studied (A. C. Zeven, personal communication). This idea is contradicted by our results. The removal of chromosome 5D, e.g., wo~ld have increased the kernel hardness of the relevant substitution lines of Cappelle Desprez, Cheyenne, Hope, and Timstein all to the same hardness level since the same repressing factor would have been removed. However, we found a large variation in kernel hardness of the four 5D substitution lines, each level corresponding to that of the donor variety (see Table 2).
We assume that wheat quality is due to a combination of kernel hardness and favour- able dough properties. As the genes for these factors are located on different chromo- somes, it should not be too difficult to introduce both factors in existing varieties with poor baking properties. In a wheat breeding programme, the quality of new lines can be assessed in a rather simple way by determining kernel hardness and dough stability.
ACKNOWLEDGMENTS
The assistance of Dr A. C. Zeven (Institute of Plant Breeding (IvP), Agricultural University, Wageningen) and Dr W. Lange (Institute De Haaff, Wageningen) is gratefully acknowledged.
REFERENCES
LAW, C. N., 1968. Gene t ic analys is us ing inter-varietal c h r o m o s o m e subs t i tu t ions . Proc. 3rd int. W h e a t Gene t . Symp. (Canber ra ) : 331-342.
Euphytica 25 (1976) 5 7 5

G. J. DOEKES AND B. BELDEROK
MAES, E. & A. PIROTTE, 1955. La d6termination du pouvoir d 'absorpt ion d'eau des farines par centrifugation. Meunerie belge 51" 15-17.
MATTERN, P. J., R. MORRIS, J. W. SCHMIDT & V. A. JOHNSON, 1973. Locations of genes for kernel properties in the wheat variety 'Cheyenne' using chromosome substitution lines. Proc. 4th int. Wheat Genet. Symp. (Columbia, Missouri): 703-707.
MORRIS, R., J. W. SCHMIDT, P. J. MATTERN & V. A. JOHNSON, 1966. Chromosomal location of genes for flour quality in the wheat variety 'Cheyenne' using substitution lines. Crop Sci. 6:119-122.
MORRIS, R., J. W. SCHMIDT, P. J. MATTERN & V. A. JOHNSON, 1968. Quality tests on six substitution lines involving 'Cheyenne' wheat chromosomes. Crop Sci. 8 : 121 123.
SEARS, E. R., W. Q. LOEGERING • H. m. RODENHISER, 1957. Identification of chromosomes carrying genes for stem rust resistance in four varieties of wheat. Agron. J. 49:208-212.
SYMES, K. J., 1965. The inheritance of grain hardness in wheat as measured by the particle size index. Aust. J. agric. Res. 16: 113-123.
WELSH, J. R., C. A. WATSON t~£ C. W. GREEN, 1968. Chromosomal control of flour properties in three substitution sets of common wheat (Triticum aestivum L.). Crop Sci. 8 : 81 82.
576 Euphytica 25 (1976)