isolation of a reticuloendotheliosis‐like virus from naturally occurring lymphoreticular tumours...
TRANSCRIPT

This article was downloaded by: [Southern Taiwan University of Science and Technology]On: 27 October 2014, At: 02:01Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Avian PathologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/cavp20
Isolation of a reticuloendotheliosis‐like virus fromnaturally occurring lymphoreticular tumours ofdomestic gooseCs. N. Drén a , I. Németh a , I. Sári b , F. Rátz c , R. Glávits c & P. Somogyi da Veterinary Medical Research Institute, Hungarian Academy of Sciences , P.O. Box 18,Budapest, H‐1581, Hungaryb Veterinary Institute of Debrecen , Debrecen, H‐4031, Hungaryc Central Veterinary Institute , Budapest, H‐1158, Hungaryd Microbiological Research Group of the National Institute of Hygiene , Budapest, H‐1529,HungaryPublished online: 02 Jan 2008.
To cite this article: Cs. N. Drén , I. Németh , I. Sári , F. Rátz , R. Glávits & P. Somogyi (1988) Isolation of areticuloendotheliosis‐like virus from naturally occurring lymphoreticular tumours of domestic goose, Avian Pathology, 17:2,259-277, DOI: 10.1080/03079458808436446
To link to this article: http://dx.doi.org/10.1080/03079458808436446
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in thepublications on our platform. However, Taylor & Francis, our agents, and our licensors make no representationsor warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Anyopinions and views expressed in this publication are the opinions and views of the authors, and are not theviews of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should beindependently verified with primary sources of information. Taylor and Francis shall not be liable for any losses,actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoevercaused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Avian Pathology, 17: 259-277,1988
ISOLATION OF A RETICULOENDOTHELIOSIS-LIKE VIRUSFROM NATURALLY OCCURRING LYMPHORETICULAR
TUMOURS OF DOMESTIC GOOSE
Cs. N. DRÉN1, I. NÉMETH1, I. SARI2, F. RÁTZ3,R. GLÁVITS3 and P. SOMOGYI4
1 Veterinary Medical Research Institute, Hungarian Academy of Sciences,H-1581 Budapest, P.O. Box 18, Hungary
2 Veterinary Institute of Debrecen, H-4031 Debrecen, Hungary
3 Central Veterinary Institute, H-1158 Budapest, Hungary
4Microbiological Research Group of the National Institute of Hygiene,H-1529 Budapest, Hungary
SUMMARY
Naturally occurring lymphoreticular tumours were found in an adultgoose flock. Diffuse and/or nodular lymphoma-like tumours occurredin the spleen, liver, pancreas, intestines and rarely in other internalorgans. Osteopetrosis-like lesions and signs of other diseases such asamidostomosis, enteritis, arthritis and air sacculitis were also observed.The tumourous lesions consisted of proliferations of undifferentiatedlymphoblasts and slightly differentiated plasmablasts of the bursalsystem of lymphocytes. Tumour cell proliferation was usually associat-ed with parenchymal atrophy and occasionally with amyloid depo-sitions along the walls of capillaries of the spleen and the liver. In tissuecultures, chloroform sensitive, typical C-type virus particles of about100 nm were isolated. The isolate incorporated 3H-uridine, banded atabout 1.19 to 1.20 g/cm3 in sucrose gradient ultracentrifugation andwas shown to be antigenically related to the reticuloendotheliosisgroup of avian retroviruses. In transmission experiments with chickensand geese, one of the four tested reticuloendotheliosis virus-positivecellular field materials caused a highly lethal neoplastic disease similarin cytology to that seen in the natural disease outbreak within fourweeks after infection. The other three cellular field materials and thetissue-culture-propagated virus isolates induced either a runting diseasesyndrome or a similar chronic neoplasia to that found in the naturallydiseased geese during an experimental period of 30 weeks. Reticuloen-dotheliosis virus was reisolated from the experimentally inoculated
Received 7 April 1987Accepted 26 August 1987
Dow
nloa
ded
by [
Sout
hern
Tai
wan
Uni
vers
ity o
f Sc
ienc
e an
d T
echn
olog
y] a
t 02:
01 2
7 O
ctob
er 2
014

260 Cs.N.DiénefcZ.
birds, and antibodies to the T strain of reticuloendotheliosis viruswere also demonstrated in sera of surviving chickens. Neither Marek'sdisease virus nor avian leukosis-sarcoma viruses could be demonstratedin tissue cultures or in chickens of transmission experiments. Thereported transmissible lymphoreticular tumours of domestic gooseseem to be associated with infection of a new isolate of the reticu-loendotheliosis group of viruses designated as ReticuloendotheliosisVirus Hungary-2 (REV-H-2).
INTRODUCTION
Naturally occurring transmissible neoplasms of domestic and free-ranging birds arefrequently associated with infections of either Marek's disease virus or avian C-typeretroviruses belonging to the avian leukosis-sarcoma group (ALSV) or to the reticu-loendotheliosis group (REV) of viruses (Calnek, 1984). Diseases associated with theC-type retroviruses are also called collectively, leukosis (Biggs, 1980). It is uncertainwhether the domestic goose (Anser anser domesticus) is susceptible to infectionwith Marek's disease virus (Ianconescu and Samberg, 1971)and to the developmentof disease (Calnek and Witter, 1984) or to infections of ALSVs (Tooze, 1973).This species is, however, known to be susceptible to both natural (Purchase etal.,1973) and experimental infections and to tumour development induced by the Tstrain of REV (Taylor and Olson, 1972). On the basis of sero-epidemiologicalstudies, the goose is considered as one of the natural hosts of REVs (Witter, 1984).However, the prevalence of infection in this species has not been studied in detail(Purchase et al., 1973; Neumann et al., 1981). Bauer and Temin (1979) demon-strated that tissue culture cells from normal goose embryo released sedimentableparticles with RNA-directed DNA polymerase activity. These particles were foundnot to be infectious viruses and shown to be chemically and physically differentfrom known avian retroviruses, though the polymerases of these particles and thespleen necrosis virus, a representative member of REV group (Purchase et ah,1973), were similar but not identical. The relationship between these particlesand REVs is enigmatic. Thus, neither true REV isolation nor REV-associateddisease has so far been reported from goose. In this report, we describe a naturallyoccurring transmissible lymphoproliferative disease of domestic goose, associatedwith a C-type retrovirus antigenically related to the REV group of avian retro-viruses.
MATERIALS AND METHODS
Experimental animalsFor transmission experiments one-day-old specifíc-pathogen-free (SPF) WhiteLeghorn chickens were obtained from the flock of Phylaxia Veterinary BiologicalsCo. (Budapest, Hungary) and one-day-old goose poults were purchased froma goose breeding farm. The birds were housed in deep-litter pens in animal roomsand were fed on commercial diets. Feed and water was available ad libitum.
Immune seraAnti-T cell and anti-B cell rabbit immune sera were obtained through the courtesyof Dr P.C. Powell (Houghton Poultry Research Station, Houghton, England) orwere produced as described elsewhere (Dren and Németh, 1985). Preparation ofanti-u heavy chain rabbit immune serum has been described previously (Dren and
Dow
nloa
ded
by [
Sout
hern
Tai
wan
Uni
vers
ity o
f Sc
ienc
e an
d T
echn
olog
y] a
t 02:
01 2
7 O
ctob
er 2
014

Reticuloendotheliosis virus-associated lymphoma in goose 261
Németh, 1985). Chicken anti-REV-T immune serum was kindly provided byIntervet (Netherlands) and rabbit anti-p27 of ALSV immune serum by Dr E.J.Smith (Regional Poultry Research Laboratory, East Lansing, USA). Fluorescein-isothiocyanate-conjugated swine anti-rabbit globulin was from SEVAC (Prague,Czechoslovakia) and was used at 1:20 dilution for indirect immunofluorescence(IF) test."Fluorescein-ldbelled rabbit antiserum to chicken immunoglobulinsChicken immunoglobulins were purified as described elsewhere (Dren and Németh,1985). Three groups of rabbits were injected intradermally each with chicken IgG,IgM and IgA in a dose of 50 ¿tg/kg body mass emulsified in incomplete Freund'sadjuvant six times fortnightly. Five weeks after the last inoculation, rabbits wereboosted with the same antigen dose and bled out 10 to 14 days later. The antibodytitre of the obtained antisera was estimated by rocket immunoelectrophoresis(Harboe and Ingild, 1973), and they were appropriately mixed to give a poly-specific immune serum to chicken immunoglobulins. The IgG fraction of thisimmune serum was isolated (Harboe and Ingild, 1973) and conjugated with fluores-cein-isothiocyanate (Isomer 1, Serva Feinbiochemica, Heidelberg, Germany) at arate of 16 jug dye/mg IgG according to Goldman (1968). The conjugate was charac-terised as described by Holborow and Johnson (1967). It contained 21.6 mg/mlof protein, had a dye-protein molar ratio of 3.9 and had a standard gel diffusionprecipitation titre of >64, 64 and 64 against chicken IgG, IgM and IgA, respec-tively. It was used at 1:40 dilution for indirect IF test.
Cell culturesCultures of chicken embryo fibroblasts (CEF) and goose embryo fibroblasts (GEF)were prepared and maintained as described by Temin and Rubin (1958), from10-day-old SPF White Leghorn chicken embryos and 13-day-old goose embryosderived from commercial flocks. Chicken kidney cell (CKC) cultures were preparedby the method described by Churchill and Biggs (1967) from 3-week-old SPF WhiteLeghorn chickens.
VirusesThe nondefective REV-T strain was obtained from Dr R.L Witter (Regional PoultryResearch Laboratory, East Lansing, USA) and the HPRS-F42 strain of avian lym-phoid leukosis virus from Dr L.N. Payne (Houghton Poultry Research Station,Houghton, England), and were propagated in CEF cultures.
Virus assaysFor virus isolation cell suspensions were prepared aseptically from suspectedtumourous organs and/or the spleens in Ham's F-10 medium (Flow LaboratoriesLtd, Irvine, Scotland) containing 10% (v/v) newborn calf serum, 10%(w/v)tryptosephosphate broth (Difco Laboratories, Detroit, USA) and 7.5% (v/v) dimethylsulphoxide, and stored in the gaseous phase over liquid nitrogen for further pro-cessing. Cell viability was determined by the trypan-blue exclusion test. Dupli-cate 18- to 24-hour monolayers of CEF, GEF and CKC in 60 mm Petri disheswere inoculated into the medium with about 106 cells/0.2 ml. Control cultureswere either inoculated with the same dose of cells from normal healthy birds orremained uninoculated. CKC were used for Marek's disease virus assay by observingthe cells through 14 days post-inoculation for typical herpesvirus cytopathiceffect (Churchill and Biggs, 1967). Inoculated and control cultures were then
Dow
nloa
ded
by [
Sout
hern
Tai
wan
Uni
vers
ity o
f Sc
ienc
e an
d T
echn
olog
y] a
t 02:
01 2
7 O
ctob
er 2
014

262 Cs.N. Dren etal.
subcultured and their supernatant fluids were transferred to fresh CKC mono-layers. The cultures were observed for another 14 days and examined for inclusionbodies by acridine orange staining (Mayor and Diwan, 1961) at the end of the4-week test period. CEF and GEF served a dual purpose as the growth cells forboth REV and ALSV. Inoculated and control culture cells were subcultured on thethird day after inoculation and cultivated for seven days. At the end of this growthcycle, culture supernatant fluid was transferred to fresh primary monolayers, whichwere cultivated again for a week. At the end of each passage, or when a suspectedcytopathic effect appeared, cell cultures were examined for REV and ALSV group-specific antigens by the indirect IF technique (Payne et al, 1966; Witter et al,1970) using respectively chicken antiserum to REV-T strain and rabbit antiserum top27 of ALSV and the appropriate fluorescein-labelled rabbit anti-chicken immuno-globulins or swine anti-rabbit globulins. Uninoculated cultures served as negativecontrols and those infected with nondefective REV-T or with the HPRS-F42 strainof avian lymphoid leukosis virus were used as positive controls for the indirect IFassays. Diffuse and/or granular, bright-green cytoplasmic fluorescence, in theabsence of reactions in uninfected culture cells, was considered specific for viralantigens. CEF and GEF cultures were also tested for the presence of inclusionbodies by acridine orange staining (Mayor and Diwan, 1961) at the end of the assayperiod.
Test for sensitivity to chloroformSensitivity of the new REV isolates to 5% (v/v) chloroform was tested according toLukert (1980). Residual infectivity of the chloroform-treated virus isolates wastested in CEF cultures as described in the preceding paragraph.
Sucrose gradient ultracentrifugationPrimary, drained CEF monolayers were infected with 0.2 ml virus under test, andafter one hour of adsorption at 37°C 5 ml culture fluid containing 10% (v/v)newborn calf serum and 15 /LtCi/ml 3H-uridine (specific activity 750 GBq/mmol;Radiochemie, Czechoslovakia) was added to the cultures. Six hours later 75 juCicold isotope was added so that the final concentration of isotope in the mediumamounted to 30 /xCi/ml. Culture fluids were harvested 96 hours after inoculationand clarified by centrifugation at 10,000g for 20 min. After further centrifugationof the supernatant at 100,000 g for 90 min in an MSE Superspeed 65 ultracentri-fuge the resulting virus pellet was homogenised in 200 jul lOmM Tris/hydroxy-methyl/aminomethane-HCl + 0.1 M NaCl + 1 mM EDTANa2, pH 7.6, buffer,immediately layered on to a 5-ml, 10 to 50% (w/w) linear sucrose gradient in thesame buffer, and centrifuged at 200,000 g for 4 hours at 4°C. The gradient wascollected into 20 fractions, which were counted for radioactivity in Bray's scintilla-tion fluid using a Packard 3330 liquid scintillation counter. The concentration ofsucrose in each fraction was determined by reading a companion gradient in anAbbe refractometer. Simultaneously processed culture fluids from uninfected andnondefective REV strain T-infected CEF cultures were used as controls.
Pathological studiesDead and/or culled ailing geese were dissected, and portions from organs with grosslesions and other internal organs including the brain, spinal cord, brachial andsciatic nerves and the bone marrow, were fixed in 10% (v/v) neutral formaldehyde,embedded in paraffin and 5 /urn sections were stained with haematoxylin and eosin.
Dow
nloa
ded
by [
Sout
hern
Tai
wan
Uni
vers
ity o
f Sc
ienc
e an
d T
echn
olog
y] a
t 02:
01 2
7 O
ctob
er 2
014

Reticuloendotheliosis virus-associated lymphoma in goose 263
Dead or culled birds from transmission experiments were processed likewise.
Electron microscopyPortions of organs with gross tumourous lesions and tissue culture cells were fixedin 2.5% (v/v) glutaraldehyde, post-fixed in 1% (w/v) osmium tetroxide and em-bedded in Durcupan (Fluka AG, Buchs, Switzerland). Ultrathin sections werestained with uranyl acetate/lead citrate and examined in a Philips 201 electronmicroscope.
Assay for REV antibodySera from experimentally infected chickens were assayed for antibody to REVby the indirect IF technique (Witter et al., 1970) using as antigen REV strainT-infected CEF cells deposited and fixed on to Multitest slides (Flow LaboratoriesLtd, Irvine, Scotland). Slides with the fixed cells were stored over silica gel at-20°C until used. Uninfected CEF cells, positive and negative serum controls and aserum-free phosphate-buffered saline (0.01 M, pH 7.2) conjugate control wereincluded in the assay. The specificity of the assay was proved by blocking thestaining with a surplus of unconjugated rabbit anti-chicken immunoglobulinsimmune serum before the addition of the conjugate. All samples were tested at1:20 dilution.
Demonstration of antigens on tumour cellsSurface antigens on tumour cells induced by the new REV isolate in experimentallyinfected chickens were studied by the peroxidase-antiperoxidase (PAP) stainingwith anti-T cell, anti-B cell and anti-/u heavy chain rabbit immune sera using formal-dehyde-fixed paraffin-embedded tissue sections. The technique used has beendescribed previously (Dren and Németh, 1987). Simultaneously processed sectionsfrom thymus and bursa of Fabricius of 3- to 6-week-old SPF White Leghorn chick-ens and from lymphoid leukosis virus strain HPRS-F42-induced tumours (Dren andNémeth, 1987) were used as reference preparations to assess the efficiency andspecificity of immunostaining.
Transmission experimentsExperiment 1. In the first transmission trial 81 one-day-old SPF White Leghornchickens were divided into eight groups (Table 1). Thirty-five chickens of groupsA, B, C and D were inoculated intraperitoneally each with 106 cells/0.2 ml pre-pared from the spleens of REV-positive tumour-bearing geese nos 8, 9, 10 and 14from the field outbreak of the disease, respectively. Ten chickens of groups A-land B-l were inoculated with 0.2 ml per chick of cell-free first passage supernatantfluids of CEF cultures infected with spleen cells from nos 8 and 9 field cases ofthe disease, respectively. Sixteen chickens were mixed with the inoculated groupsand formed a contact exposed group (group E). The different groups were accom-modated in separate pens in the same room. Twenty uninoculated chickens werekept separately and served as a control (group F). At 7 weeks of age, all survivorswere weighed, and ten from the control group and one to four from the othergroups were selected at random and killed for virus isolation, antibody and histo-pathological studies. The remaining chickens were observed until they were 29
. weeks of age when the experiment was terminated.
Experiment 2. In the second transmission trial 40 six-day-old commercial goosepoults were divided into seven groups (Table 2). Twenty-five poults of groups A, B,B-l, C and D were inoculated subcutaneously in the neck with the same dose of
Dow
nloa
ded
by [
Sout
hern
Tai
wan
Uni
vers
ity o
f Sc
ienc
e an
d T
echn
olog
y] a
t 02:
01 2
7 O
ctob
er 2
014

Table 1. Response of chickens to inoculation at 1-day-old with spleen cells from naturally diseased tumourous geese and with reticu-loendotheliosis virus isolates made in chick embryo fibroblast cultures
Group
AA-1
BB-1
CDEF
Inoculum
No. 8 REV+ cellsa
Supernatant of CEFculturese
No. 9 REV+ cellsb
Supernatant of CEFcultures'
No. 10REV+cellsc
No. 14 REV* cellsd
Contact exposedNone
No.chicks
10
510
55
101620
No.died
19
1141
No. withtumours
06
1030
Age atdeath(days)
18511-107
141123
111-176164
Isolationof REVS
2/2h
1/15/8
3/32/21/22/40/10
j
Antibodyto REV
0/2
o/i
2/30/20/21/40/10
fit 7 weeks
Feather-ing
defect
4/10
5/52/2
5/55/5
10/100/160/20
Bodymass(g)
356.01
298.0i
315.0*314.01
319.51
394.5393.6
At 29.weeksJ
2/8
0/3o/i
0/11/21/41/110/10
o
£7
a,b,c,d Spleen cells from nos 8, 9,10 and 14 field cases of the disease from which reticuloendotheliosis virus (REV) was isolated, respectively.e>f Cell-free first passage supernatant fluids of chicken embryo fibroblast cultures inoculated with spleen cells from field cases nos 8 and 9,
respectively.S Virus isolations were made from birds culled at 7 weeks of age and from those dying before that age.. No. positive/No, tested.?P?C0.05 compared with group F.J No. birds with gross tumours/No, killed at termination.
Dow
nloa
ded
by [
Sout
hern
Tai
wan
Uni
vers
ity o
f Sc
ienc
e an
d T
echn
olog
y] a
t 02:
01 2
7 O
ctob
er 2
014

Table 2. Response of geese to inoculation at 6-day-old with spleen cells from naturally diseased tumour-bearing geese and with aretículoendotheliosis virus isolate made in chick embryo fibroblast cultures
Group
ABB-1CDEF
Inoculum
No. 8 REV+ cellsa
No. 9 REV"1" cellsb
Supernatant of CEF culturese
No. 10 REV+cells0
No. 14 REV+ cellsd
Contact exposedNone
No.geese
55555
105
No.died
51232
No. witlitumours
30211
Day of-deathafter
infection
11-2212
177-18480-172
128-151
Bodymass(g)
1886.0f
2276.51888.0f
1886.0f
2078.03152.5
Isolationof REVS
2/2
1/11/32/23/34/40/5
At 30 weeks
No.killed
5
43285
No. withtumours
2
21100
a '">c '" Spleen cells from nos 8,9,10 and 14 field cases of the disease from which reticuloendotheliosis virus (REV) was isolated, respectively.e Cell-free first passage supernatant fluids of chicken embryo fibroblast cultures infected with spleen cells from no. 9 field case of the disease.
P<0.05 compared with group F five weeks after inoculation.*> Virus isolations were made from birds dying within 80 days and from heparinised blood samples of survivors collected five weeks after inoculation;
No. positive/No, tested.
D.
S'
1
to<
Dow
nloa
ded
by [
Sout
hern
Tai
wan
Uni
vers
ity o
f Sc
ienc
e an
d T
echn
olog
y] a
t 02:
01 2
7 O
ctob
er 2
014

266 Cs. N. Drén et al.
the same materials used for inoculation of the respective groups of Expt. 1. Tenpoults were left untreated and formed a contact exposed group (group E). Thepoults of these groups were mixed together following inoculation and assignedat random to three pens in the same room. Five uninoculated poults were housedseparately and served as controls (group F). Five weeks after inoculation, all sur-vivor poults were weighed, and heparinised whole blood was collected at randomfor virus assays. The experiment was terminated when the geese were 31 weeksold.
All birds dying during the experiments or killed at the terminations were necrop-sied, and gross lesions were recorded. Some selected birds were examined histo-logically. Body mass differences between the infected and control groups of animalswere evaluated according to the Kruskall-Wallis test (Campbell, 1974).
RESULTS
Ginical features and pathologyIn an isolated goose flock, increasing mortality was noticed at about one week afterthe second plucking at 16 weeks of age. Mortality reached almost 40% by the 22ndweeks of age, when the flock was slaughtered. The affected birds were listless,usually emaciated, often showed lameness, and their feathers were ruffled. Atnecropsy of 23 dead or culled ailing geese of 20 weeks of age, gross neoplasticlesions were noted in 14 birds. The spleen and the liver were usually diffuselyenlarged and characteristically mottled, especially the spleen, from yellowish-white irregular areas throughout their substance. In the other affected organsmainly nodular lymphoma-like tumours were encountered (Fig. 1). Most oftenaffected organs were the spleen (14 birds), liver (8), pancreas (5), intestines (4)and occasionally solitary focal tumours occurred in the heart (1), kidney (1) andthe lung (1). Osteopetrosis-like lesions (1) and signs of other diseases such asamidostomosis (5), enteritis (1), arthritis (1) and air sacculitis (1) were also ob-served.
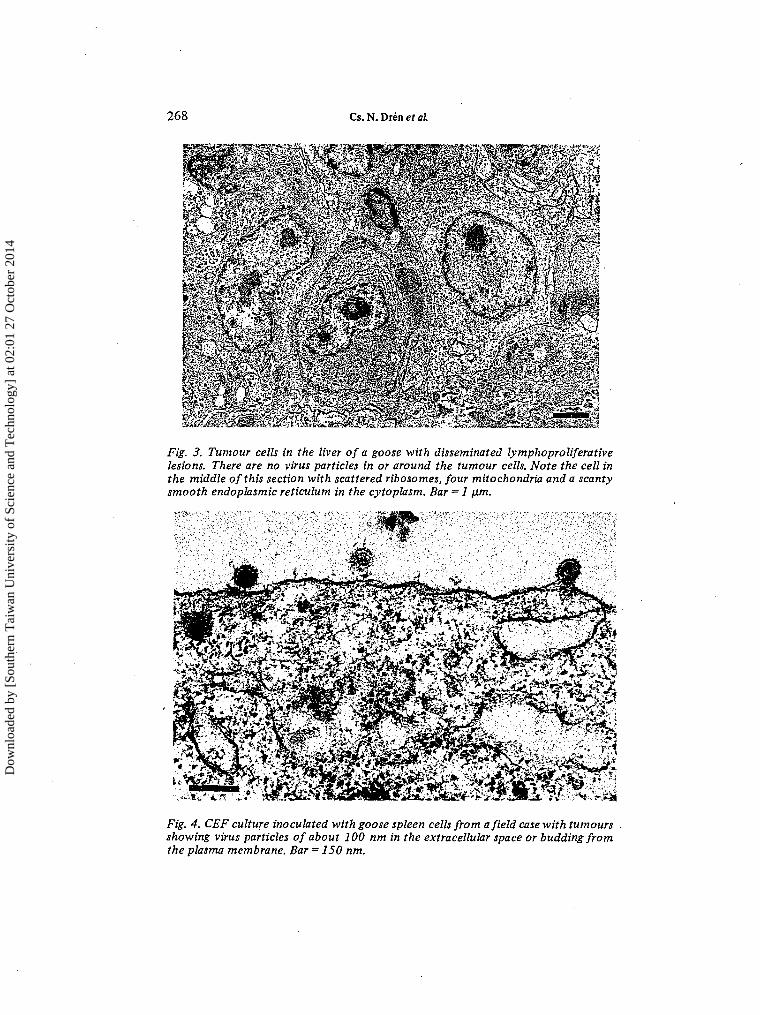
Detailed histological studies were performed on a total of seven dead and 10culled geese, of which 10 had gross neoplastic lesions at necropsy. No histologicallesions were found in four geese. In the remaining birds, diffuse and/or focalproliferative lesions consisting of large, slightly different, lymphoblast-type cells(Fig. 2) were seen in the spleen (13 birds), liver (13), pancreas (11), small intestines(10), proventriculus (8), kidney (8), heart (5), lung (4) and occasionally in adrenaland thyroid glands, bone-marrow, gonads, thymus and the bursa of Fabricius.The nervous system was unaffected. Tumour cell proliferation usually associatedwith paienchymal atrophy and occasionally with amyloid depositions along thewalls of capillaries in the spleen and the liver. Electron microscopic examinationsrevealed no virus particles in or around the tumour cells, which seemed to beprimitive undifferentiated lymphoblasts and slightly differentiated plasmablastsof the bursal system of lymphocytes. In the relatively large spherical or irregularnucleus of tumour cells one or two prominent dense nucleoli were seen, and thefinely granulated nuclear chromatin was located along the nuclear membrane.The cytoplasm contained either only scattered ribosomes and ribosomal aggre-gates or some mitochondria and a scanty smooth endoplasmic reticulum in addition(Fig. 3).
Dow
nloa
ded
by [
Sout
hern
Tai
wan
Uni
vers
ity o
f Sc
ienc
e an
d T
echn
olog
y] a
t 02:
01 2
7 O
ctob
er 2
014

Reticuloendotheliosis virus-associated lymphoma in goose 267
Fig. 1. Lymphoma-like tumours in various internal organs of a goose.
Fig. 2. Lymphoblastoid tumourcell proliferation in the liver of a goose.HE; bar = 100 urn.
Dow
nloa
ded
by [
Sout
hern
Tai
wan
Uni
vers
ity o
f Sc
ienc
e an
d T
echn
olog
y] a
t 02:
01 2
7 O
ctob
er 2
014
268 Cs. N. Drén et al.
Fig. 3. Tumour cells in the liver of a goose with disseminated lymphoproliferativelesions. There are no virus particles in or around the tumour cells. Note the cell inthe middle of this section with scattered ribosomes, four mitochondria and a scantysmooth endoplasmic reticulum in the cytoplasm. Bar = 1 ¡xm.
Fig. 4. CEF culture inoculated with goose spleen cells from afield case with tumoursshowing virus particles of about 100 nm in the extracellular space or budding fromthe plasma membrane. Bar = 150 nm.
Dow
nloa
ded
by [
Sout
hern
Tai
wan
Uni
vers
ity o
f Sc
ienc
e an
d T
echn
olog
y] a
t 02:
01 2
7 O
ctob
er 2
014

Reticuloendotheliosis virus-associated lymphoma in goose 269
Virus isolation and characterisationFor detailed virological examinations, 12 geese were selected from those studiedhistologically. With three exceptions, all birds had gross and/or histological proli-ferative lesions. In 60 to 66% of CEF and GEF cultures, but in no case in CKCcultures, a transient cytopathic effect was observed at about 5 to 7 days afterinoculation with spleen cell suspensions containing 50 to 80% living cells. Duringthe cytopathic effect the cell number tended to decrease in the infected cultures.Their cells became spindle shaped and granular, and sometimes they rounded upand detached from the wall of the culture dish. On further incubation, the cellsbegan to divide again and developed a monolayer indistinguishable from that of theuninfected control cultures. These cultures, however, became confluent 3 to 4 dayslater than the controls. This phenomenon was not observed in control cultures,which were inoculated with spleen cell suspension from healthy adult geese. Onindirect IF staining of cell cultures with anti-REV-T chicken serum, eight out of 12CEF, three out of five GEF and four out of 12 CKC cultures showed diffuse and/orgranular cytoplasmic fluorescence. Electron microscopic examination of ultrathinsections of the cells from these REV-positive cultures revealed typical C-type virusparticles of about 100 nm in diameter in the extracellular space or budding on theplasma membrane (Fig. 4) .
All cultures were negative for ALSV group-specific antigen by indirect IF stainingwith anti-p27 of ALSV rabbit serum, while the cells of lymphoid leukosis virusstrain HPRS-F42-infected CEF cultures, used as positive controls, gave intense cyto-plasmic fluorescence. In the 12 attempted isolations, no sign of Marek's diseaseherpesvirus cytopathic effect was noted in CKC cultures. Neither intracytoplasmicnor intranuclear inclusions were revealed by acridine orange staining in any CEF,GEF and CKC cultures at the end of the assay period.
Tissue-culture-propagated REV isolates from nos 8 and 9 field cases of the diseasewere selected for further characterisation studies. Both virus isolates grew in CEFcultures with only a mild and transient cytopathology. The fluorescence in infectedcells stained with the reference anti-REV-T immune serum by the indirect IFmethod was indistinguishable from that in nondefective REV strain T-infected cells.Both virus isolates were completely inactivated on 5% (v/v) chloroform treatment,incorporated 3H-uridine in CEF cultures and banded at about 1.19 to 1.20 g/cm3
in sucrose gradient ultracentrifugation.
Experimental transmissionTransmission experiments were carried out with spleen cells from REV-positivetumour-bearing geese nos 8, 9, 10 and 14 from the natural disease outbreak andwith tissue-culture-propagated viruses from field cases nos 8 and 9.
Experiment 1. Results of experiments with chickens are recorded in Table 1. Eightchickens from group B died between 11 and 28 days after inoculation. They wereseverely stunted and lethargic, but showed no other specific symptom before death.At necropsy, six birds were found to have gross neoplastic lesions. The spleen andthe liver were moderately enlarged and had small focal tumours throughout theirsubstance. Discrete nodular tumours occurred also at the site of inoculation. Thethymus and the bursa of Fabricius of these chicks were substantially smaller in sizethan those of age-matched normal controls. Histologically, hepatitis or hepaticdistrophy were found in'two birds, whereas in all the others diffuse and/or focal
Dow
nloa
ded
by [
Sout
hern
Tai
wan
Uni
vers
ity o
f Sc
ienc
e an
d T
echn
olog
y] a
t 02:
01 2
7 O
ctob
er 2
014

270 Cs.N. Dren era/.
proliferation of relatively uniform, large, lymphoblastoid cells were seen in everyinternal organ examined except the nervous system. There was no additional morta-lity among chickens of the inoculated groups until 107 days of age. At 7 weeks ofage, however, all chicks, with six exceptions in group A, showed feathering defectssimilar to 'nakanuke' (Koyama et al., 1976), and the mean body mass of chicks inthese groups was significantly lower than that of 20 chicks in the control group.Birds that were culled at this age for virological and antibody assays had no detect-able gross and microscopic lymphoproliferative lesions. From the 27 survivors, eightchickens died between 107 and 185 days of age, of these four had gross neoplasticlesions and four appeared to have the ranting syndrome. Of the remaining 19chickens, gross tumours were noted in four birds when killed at the end of theexperiment. Among the eight chickens with lymphomas that died late in theexperiment (141 to 176 days) or were killed at the end, diffuse and/or nodularneoplasms occurred in the liver (4 birds), heart (4), thymus (4), kidney (3), spleen(2), lung (2), ovary (2), muscles (1), pericardium (1) and the bursa of Fabricius (1).Histologically, the tumours were composed of moderately pleomorphic lympho-blastoid cells. There were no lesions in the nervous system of the affected chickens.
Contact exposed chickens (group E) did not show feathering defects and growthretardation, and neither gross nor microscopic lesions were noted in those fourchicks that were culled at 7 weeks of age. One bird died at 164 days because ofcannibalism with no detectable neoplastic lesions, and one of the 11 survivors haddiscrete tumours in the liver and kidney when killed at the end of the experiment.
REV-related viruses having the same cultural, morphological and antigeniccharacteristics as the field isolated were reisolated in CEF cultures from 14 out of18 dead or culled chickens from the inoculated groups and from two out of fourbirds from the contact exposed group. In the attempted 22 viras isolations frombirds of the infected groups (groups A to E) all CEF cultures gave negative reactionby indirect IF staining with anti-p27 of ALSV immune serum, and at no time wereinclusion bodies evident with acridine orange staining.
Antibodies to REV were demonstrated in two of 10 chickens from the inoculatedgroups and in one of four birds from the contact exposed group when tested at7 weeks of age.
Control chickens (group F) appeared clinically and pathologically normal, andexperienced no mortality during the experiment. Also, no antibody to REV wasdetected, and REV could not be isolated from 10 selected chickens when examinedat 7 weeks of age.
Experiment 2. Results of transmission experiments with geese are given in Table 2.AU goose poults of group B died between 11 and 22 days after inoculation. Birdswere lethargic and showed growth retardation compared with the uninoculatedcontrols, but no other specific sign before death. At necropsy, gross tumourouslesions were noted in three birds. The liver and particularly the spleen werediffusely enlarged up to three times normal size and congested with small irregulartumours throughout their substance. Discrete focal tumours were also present inthe lung, heart, kidney, small intestine, pancreas and at the site of inoculation. Themucosa often ulcerated above the tumour nodules in the wall of the small intestineand was covered with yellowish caseous necrotic debris. Histologically, acuteinterstitial hepatitis and necrotic enteritis in the small intestine or hepatitis were
Dow
nloa
ded
by [
Sout
hern
Tai
wan
Uni
vers
ity o
f Sc
ienc
e an
d T
echn
olog
y] a
t 02:
01 2
7 O
ctob
er 2
014

Reticuloendotheliosis virus-associated lymphoma in goose 271
found in two birds, while in the other three tumour-bearing poults diffuse and/orfocal proliferative lesions similar in cytology to those seen in the natural diseasewere present in all abdominal and thoracic viscera. One poult from group B-l diedof thymus and bursa atrophy, pericarditis and hepatic dystrophy 12 days afterinoculation.
Most survivors of the inoculated groups showed 'nakanuke' 5 weeks post-inocula-tion, and abnormalities of the primary wing feathers could be observed on somegeese up to their death or to the end of the experiment. These birds also grewpoorly: mean body mass.of the inoculated groups, excepting group B-l, was signifi-cantly lower than that of the control group. Of the 19 survivors, five geese diedbetween 80 and 184 days after inoculation, and at necropsy three were found tohave gross neoplasms with accompanying air sacculitis, pericarditis, enteritis in theupper small intestine and peritonitis in two birds. The two birds died from groupD 80 days post-inoculation, which lacked definite gross proliferative lesions, hadair sacculitis, thymus and bursa atrophy, moderately enlarged liver and greatlyenlarged spleen with some evidence of grey mottling. Histological examinationrevealed acute hepatitis or increased activity of reticulo-histiocytic cells in the liverand marked reticulum cell proliferation with accompanying amyloidosis in thespleen. Of the remaining 14 geese, gross tumours were observed in six birds whenkilled at the end of the experiment. In the nine tumourous geese that died late inthe experiment (172 to 184 days) or were killed at the end, the tumours werenodular and the organs affected included thymus (7 birds), spleen (5), liver (4),lung (2), heart (1), kidney (1), pancreas (1), ovary (1), muscles (1), peritoneum (1)and bursa of Fabricius (1). The cellular morphology of these neoplasms did notdiffer apparently from that of the experimentally induced lymphomas in chickensof Expt 1 and from that of the tumours from naturally diseased geese.
In the contact exposed group (group E), two geese died 128 and 151 days after con-tact exposure: one from chronic air sacculitis, peritonitis and diffuse neoplastic les-ions in the spleen and liver, and the other because of cannibalism with no sign ofneoplasia. The remaining eight geese showed no lesions at the end of the experiment.
REV with the same features as the field isolates were reisolated in CEF culturesfrom nine of 11 birds from the inoculated groups and also from four of four birdsfrom the contact exposed group. In the attempted 15 virus isolation from birdsof the infected groups (groups A to E), ALSV could not be demonstrated byindirect IF staining with anti-p27 of ALSV immune serum in CEF cultures, andCKC cultures showed no evidence of herpes-type virus plaques. Also, no inclusionbodies were noted in these cultures after acridine orange staining at the end of theassay period.
There was no loss among the control geese (group F) during the experiment, andthey had no detectable lesions at termination. Also, REV could not be isolatedfrom these birds at 6 weeks of age.
Examination of antigens on experimentally induced tumour cellsDeparaffinised histological sections of gross tumours developed in chickens ofExpt 1 were tested for the presence of specific T-cell, B-cell and IgM antigenicmarkers by the PAP technique. Table 3 summarises the result of these studies.None of the sections of the 10 examined tumorous organs from five chickensshowed positive reaction with anti-T cell or anti-B cell serum, though both immunesera stained brightly the homologous cells on sections of normal thymus and
Dow
nloa
ded
by [
Sout
hern
Tai
wan
Uni
vers
ity o
f Sc
ienc
e an
d T
echn
olog
y] a
t 02:
01 2
7 O
ctob
er 2
014

272 Cs. N. Drén et al.
Table 3. Demonstration of cell surface antigens by the peroxidase-antiperoxidase(PAP) method in tumours of chickens experimentally infected with thenew reticulcendotheliosis virus isolate
Inoculum
REV+ cellsa
HPRS-F42b
None
Birdc
no.
1
2
3
4
56
78
Organsd
with withouttumours
Si
SiLi
SpLi
MsThyBF
LiLiSpKiBF
ThyBF
ThyBF
ThyBF
ThyBF
PAP staining with serae
anti-T
++
+
++
—
1 1
1 1
—++
anti-B
+
++
++
++
—
++++++•H-
++
anti-jit
+
+ 1
11
+ 1
i+ 1
—
±
++++++++
+
normal
-
1 1
1 1
1 1
1 1
-
1 1
1 1
—
—
a Goose spleen cells from which leticuloendotheliosis virus was isolated.b HPRS-F42 strain of lymphoid leukosis virus.c Chicks nos. 1, 2 and 3 died or were killed when moribund between 11 and 28 days and nos.
4 and 5 were killed 29 weeks after inoculation at day-old. Birds nos. 6 and 7 were inoculatedas 8-day embryos with 104 tissue culture dose of HPRS-F42 strain of lymphoid leukosis virusand died 21 and 27 weeks after hatching, respectively.
d Organs with or without gross and microscopic proliferative lesions: BF = bursa of Fabricius,Ki = kidney, Li = liver, Ms = muscles, Si = site of inoculation, Sp = spleen, Thy = thymus,
e The result of staining was scored as follows:++ = strong diffuse positive reaction.+ = strong focal positive reaction.± = questionable reaction, - = negative reaction.
bursa of Fabricius. IgM-bearing cells also could not be demonstrated on the sectionsof neoplasms. Only on liver tumour sections from birds 3 and 5 occurred occasion-ally some solitary islands of mononuclear cells showing a weak staining with anti-fzserum. In contrast, most of the cells in the affected areas stained strongly on thereference tissue sections of lymphoid leukosis virus strain HPRS-F42-inducedlymphomas (birds 6 and 7), which are made up almost entirely of IgM-producingB-lymphoblasts. A varying number of cells also stained distinctly with anti-/u
Dow
nloa
ded
by [
Sout
hern
Tai
wan
Uni
vers
ity o
f Sc
ienc
e an
d T
echn
olog
y] a
t 02:
01 2
7 O
ctob
er 2
014

Reticuloendotheliosis virus-associated lymphoma in goose 273
serum in the medulla of several normal bursal follicles. Control preparations, inwhich normal rabbit serum was substituted for the specific primary antisera, wereconsistently negative whatever the sample under study.
DISCUSSION
Very little is known about the naturally occurring transmissible neoplasms ofdomestic goose. Thus far, no convincing experimental evidence exists as to whetherthis avian species is susceptible to infection with Marek's disease herpesvirus and tothe development of the disease (Ianconescu and Samberg, 1971 ; Calnek and Witter,1984) or to infection with ALSVs (Tooze, 1973). This bird is, however, susceptibleto infection and tumour development induced by the T strain of REV (Taylor andOlson, 1972). Although antibodies to REV have been demonstrated to occur ingoose flocks under natural conditions (Purchase et al, 1973), neither isolation ofREV nor REV-associated disease seems to have been reported previously fromgoose.
In the present study a typical C-type retrovirus was isolated from a flock of adultgeese suffering from a generalised lymphomatous disease. The isolated virus wasshown to have close physical and chemical, morphological and antigenical relation-ship to the T strain of REV. This strongly suggests that it is a member of the REVgroup of avian retroviruses and it has therefore been designated REV-Hungary-2(REV-H-2).
In transmission studies with chickens and geese, one of the four tested REV-containing cellular materials from naturally diseased tumourous geese produced ahighly lethal neoplasia similar in cytology to that seen in the field outbreak of thedisease within a short latent period of 11 to 28 days. Whether the high patho-genicity of tlüs material was due to the transplantation of tumour cells (Biilow andWeiland, 1980; Purchase and Payne, 1984) or to the presence of viral oncogenetransduced by REV as a result of recombination with host cell genome (Bishop,1983) remains to be elucidated. The other three tested REV-positive cellular fieldmaterials and the tissue-culture-propagated virus isolates induced either non-neoplastic disorders including runting, bursal and thymic atrophy and abnormalfeather development or a chronic lymphoid neoplasia similar in cellular morphologyto that found in the natural disease after a latent period of 20 to 30 weeks. Thesepathological syndromes strongly resemble those observed by others in chickensafter experimental infection with nondefective strains of REV (Mussman andTwiehaus, 1971; Witter and Crittenden, 1979; Witter et al, 1970, 1981, 1986).It is interesting that infiltration of the peripheral nerves, a feature reported to occurin REV infections (Purchase et al, 1973; Witter et al, 1970,1986), was not notedin the present investigations.
It is highly unlikely that Marek's disease virus was involved in the aetiology of thislymphoproliferative disease of goose since repeated efforts to isolate the virus wereunsuccessful, though the materials used for virus isolation contained a high propor-tion of living cells. In support of this view is the observation that the proliferativehistological lesions of both naturally diseased and experimentally infected birdslacked the pleomorphic lymphocyte populations typical of Marek's disease.Furthermore, chickens inoculated at one day of age with a large number of livingtumour cells from field cases of the disease did not show any evidence of Marek's
Dow
nloa
ded
by [
Sout
hern
Tai
wan
Uni
vers
ity o
f Sc
ienc
e an
d T
echn
olog
y] a
t 02:
01 2
7 O
ctob
er 2
014

274 Cs. N. Drén et al.
disease. The participation of exogenous ALSVs in the induction of this tumourouscondition can also be excluded with high probability, because we failed to demon-strate such viruses by cell-culture assay in any of the 49 attempted isolations fromnaturally diseased and experimentally infected birds. The indirect IF method weapplied in these studies to detect ALSV group-specific antigen in CEF cultures hasbeen reported to be analogous to the complement-fixation test for avian leukosisviruses (Kelloff and Vogt, 1966; Payne et al, 1966). As mentioned above, suscept-ibility of goose to infection with ALSVs is rather uncertain at present.
During the transmission experiments, REV with the" same features as the field iso-lates was reisolated from a number of inoculated and contact exposed birds, andantibodies to the T strain of REV were also demonstrated in sera of survivingchickens seven weeks after infection. The results reported in this paper fulfil Koch'spostulates. Therefore the foregoing experimental evidence allows the conclusionthat the generalised lymphomatous disease of domestic goose described in this studyis caused by a new isolate of the REV group. The origin of this virus is unknown.
The lineage of tumour cells induced by the new REV isolate could not be preciselydetermined, as PAP staining failed to demonstrate the presence of either T-cell orB-cell or IgM antigenic markers on the cells of histological sections of tumours fromexperimentally infected chickens, though the appropriate reference cells gave con-sistently unquestionable positive reaction. This failure is probably due to theabsence or inproper expression of antigen(s) on the surface of immature tumourcells. Similar results were published by others (Beug et al, 1981 ¡Lewis et al, 1981;Witter et al, 1986). In the present study, however, the cellular composition ofneoplasms was relatively uniform in both the naturally diseased and the experi-mentally infected birds. The proliferating cells were only moderately pleomorphicand lymphoid in morphology. By ultrastructural studies, a proportion of tumourcells of the naturally occurring goose neoplasms possessed some of the features ofprimitive plasmablasts, indicating that they belong to the B-cell series. Preciseidentification of the target cell needs, however, further studies.
AcknowledgementsWe wish to thank Dr L.N. Payne, Dr P.C. Powell, Dr E.J. Smith and Dr R.L. Witterfor kindly providing the reference virus strains and some of the immune sera usedin this study, and Mrs I. Császár for competent technical assistance.
REFERENCES
Bauer, G. and Temin, H.M. (1979). RNA-directed DNA polymerase from particles released bynormal goose cells. Journal of Virology, 29: 1006-1013.
Beug, H., Muller, H., Grieser, S., Doderlein, G., and Graft, T. (1981). Hematopoietic cellstransformed in vitro by REV-T avian reticuloendotheliosis virus express characteristicsof very immature lymphoid cells. Virology, 115: 295-309.
Biggs, P.M. (1980). The biology of avian leukosis. In: Leukemias, Lymphomas and Papillomas:Comparative Aspects, pp. 233-246. Edited by Bachmann, P.A. London: Taylor andFrancis.
Bishop, J.M. (1983). Cellular oncogenes and retroviruses. In: Annual Review of Biochemistry,pp. 301-354. Edited by Snell, E.E., Boyer, P.D., Meister, A. and Richardson, C.C. PaloAlto: Annual Reviews Inc.
Billow, V.v. and Weiland, F. (1980). Stimulation of local solid tumour development of thenonproducer Marek's disease tumour transplant JMV by virus-induced immunosuppres-sion. Avian Pathology, 9: 93-108.
Dow
nloa
ded
by [
Sout
hern
Tai
wan
Uni
vers
ity o
f Sc
ienc
e an
d T
echn
olog
y] a
t 02:
01 2
7 O
ctob
er 2
014

Reticuloendotheliosis virus-associated lymphoma in goose 275
Calnek, B.W. (1984). Neoplastic diseases. In: Diseases of Poultry, 8th Edition, pp. 323-325.Edited by Hofstad, M.S., Calnek, B.W., Helmboldt, C.F., Reid, W.M. and Yoder, H.W.Ames: Iowa State University Press.
Calnek, B.W. and Witter, R.L. (1984). Marek's disease. In: Diseases of Poultry, 8th Edition,pp. 325-360. Edited by Hofstad, M.S., Calnek, B.W., Helmboldt, C.F., Reid, W.M. andYoder, H. W. Ames: Iowa State University Press.
Campbell, R.C. (1974). Statistics for Biologists. London: Cambridge University Press.Churchill, A.E. and Biggs, P.M. (1967). Agent of Marek's disease in tissue culture. Nature,
London, 215: 528-530.Drén, Cs.N. and Nèmeth, I. (1985). Demonstration of cell surface antigens of normal and virus-
transformed chicken lymphocytes by the peroxidase-antiperoxidase method. ActaVeterinaria Hungarica, 33: 149-162.
Drén, Cs.N. and Nèmeth, I. (1987). Demonstration of immunoglobulin M on avian lymphoidleukosis lymphoma cells by the unlabelled antibody peroxidase-antiperoxidase method.Avian Pathology, 16: 253-268.
Goldman, M. (1968). Fluorescent Antibody Methods. New York and London: AcademicPress.
Harboe, N. and Ingild, A. (1973). Immunisation, isolation of immunoglobulins, estimation ofantibody titre. Scandinavian Journal of Immunology, 2(Suppl. 1): 161-164.
Holborow, E.J. and Johnson, G.D. (1967). Immunofluorescence. In: Handbook of Experi-mental Immunology, pp. 571-596. Edited by Weir, D.M. Oxford and Edinburgh: Black-well Scientific Publications.
Ianconescu, M. and Samberg, Y. (1971). Etiological and immunological studies in Marek'sdisease. II. Incidence of Marek's disease precipitating antibodies in commercial flocksand in eggs. Avian Diseases, 15: 177-186.
Kelloff, G. and Vogt, P.K. (1966). Localization of avian tumor virus group-specific antigen incell and virus. Virology, 29: 377-384.
Koyama, H., Suzuki, Y., Ohvada, Y. and Saito, A. (1976). Reticuloendotheliosis group viruspathogenic to chicken isolated from material infected with turkey herpesvirus (HVT).Avian Diseases, 20: 429-434.
Lewis, R.B., McClure, J.M., Rup, B., Niesel, D.W., Garry, R.F., Hoelzer, J.D., Nazerian, K. andBose, H.R. (1981). Avian reticuloendotheliosis virus: Identification of the hematopoietictarget cell for transformation. Cell, 25: 421-431.
Lukert, P.D. (1980). Virus identification and classification. In: Isolation and Identification ofAvian Pathogens, 2nd Edition, pp. 122-123. Edited by Hitchner, S.B., Domermuth, C.H.Purchase, H.G. and Williams, J.E. New York: Creative Printing Company.
Mayor, H.D. and Diwan, A.R. (1961). Studies on the acridine orange staining of two purifiedRNA viruses: Poliovirus and tobacco mosaic virus. Virology, 14: 74-82.
Mussman, H.C. and Twiehaus, M..J. (1971). Pathogenesis of reticuloendothelial virus disease inchicks - an acute runting syndrome. Avian Diseases, 15: 483-502.
Neumann, U., Mikami, T., Kaleta, E.F., Busche, H.-J. and Heffels, U. (1981). SerologischeUntersuchungen zum Infektionsstatus mit Retikuloendotheliose-Virus beim Wirtschafts-geflügel in Norddeutschland. Deutsche Tierärztliche Wochenschrift, 88: 104-108.
Payne, F.E., Solomon, J.J. and Purchase, H.G. (1966). Immunofluorescent studies of group-specific antigen of the avian sarcoma-leukosis viruses. Proceedings of the NationalAcademy of Sciences, USA, 55: 341-349.
Purchase, H.G. and Payne, L.N. (1984). Leukosis/sarcoma group. In: Diseases of Poultry, 8thEdition, pp. 360-405. Edited by Hofstad, M.S., Calnek, B.W., Helmboldt, C.F., Reid,W.M. and Yoder, H.M. Ames: Iowa State University Press.
Purchase, H.G., Ludford, C., Nazerian, K. and Cox, H.W. (1973). A new group of oncogenicviruses: Reticuloendotheliosis, chick syncytial, duck infectious anemia and spleennecrosis viruses. Journal of the National Cancer Institute, 51: 489-499.
Taylor, H.W. and Olson, L.D. (1972). Spectrum of infectivity and transmission of the T virus.Avian Diseases, 16: 330-335.
Temin, H.M. and Rubin, H. (1958). Characterization of an assay for Rous sarcoma virus andRous sarcoma cells in tissue culture. Virology, 6: 669-688.
Tooze, J. (1973). The Molecular Biology of Tumour Viruses, pp. 531. Cold Spring Harbor:Cold Spring Harbor Laboratory.
Dow
nloa
ded
by [
Sout
hern
Tai
wan
Uni
vers
ity o
f Sc
ienc
e an
d T
echn
olog
y] a
t 02:
01 2
7 O
ctob
er 2
014

276 Cs.N.DréneíJ/.
Witter, R.L. (1984). Reticuloendotheliosis. In: Diseases of Poultry, 8th Edition, pp. 406-417.Edited by Hofstad, M.S., Calnek, B.W., Helmboldt, C.F., Reid, W.M. and Yoder, H.W.Ames: Iowa State University Press.
Witter, R.L. and Crittenden, L.B. (1979). Lymphomas resembling lymphoid leukosis in chick-ens inoculated with leticuloendotheliosis virus. International Journal of Cancer, 23:673-678.
Witter, R.L., Purchase, H.G. and Burgoyne, G.H. (1970). Peripheral nerve lesions similar tothose of Marek's disease in chickens inoculated with reticuloendotheliosis virus. Journalof the National Cancer Institute, 45: 567-577.
Witter, R.L., Smith, E.J. and Crittenden, L.B. (1981). Tolerance, viral shedding, and neoplasiain chickens infected with non-defective reticuloendotheliosis viruses. Avian Diseases,25: 374-394.
Witter, R.L., Sharma, J.M. and Fadly, A.M. (1986). Nonbursal lymphomas induced by non-defective reticuloendotheliosis virus. Avian Pathology, 15: 467-486.
RESUME
Isolement d'un pseudo-virus de la réticulo-endothéliose à partir detumeurs lymphoréticulaires survenant naturellement chez des oies domestiques
Des tumeurs lymphoréticulaires survenant naturellement ont été observées dansun troupeau d'oies adultes. Ces tumeurs, ressemblant à des lymphomes diffuset/ou nodulaires, sont observées dans la rate, le foie, le pancréas, les intestins etrarement dans d'autres organes internes. Des lésions resemblant à de l'ostéopétroseainsi que les symptômes d'autres maladies, telles que l'amidostomose, l'entérite,l'arthrite et l'aérosacculite sont également observés. Les lésions tumorales consis-tent en la prolifération de lymphoblastes non différenciés et de plasmoblastespeu différenciés du système bursal des lymphocytes. La prolifération des cellulestumorales est habituellement associée à une atrophie du parenchyme et occasion-nellement à des dépôts amyloi'des le long des parois des capillaires de la rate et dufoie. En culture de tissus, des particules de virus, typiques de type C, sensibles auchloroforme, d'une taille d'environ 100 nm ont été isolées. Ces particules viralesont incorporé la 3H-uridine, se situent à environ 1,19 à 1,20 g/cm3 en gradient desucrose par ultracentnfugation et se révèlent antigéniquement appartenir au groupede la réticulo-endothéliose des rétrovirus aviaires. Les expérimentations, pourtransmettre le virus à des poulets et des oies, ont parmis, avec l'un des quatreprélèvements du terrain contenant des cellules positives pour le virus de la réticulo-endothéliose, de reproduire une maladie néoplasique hautement létale, semblabledu point de vue cytologique à celle observée dans la maladie naturelle au coursdes quatre semaines suivant l'infection. Les trois autres prélèvements du terrainet les isolements de virus multipliés sur cultures cellulaires ont entraîné soit unsyndrome de dépérissement soit une néoplasie chronique similaire à celle observéenaturellement chez des oies pendant une période d'observation de 30 semaines.Le virus de la réticulo-endothéliose a également été réisolé à partir des oiseauxinoculés expérimentalement et des anticorps vis-à-vis de la souche T du virus de laréticulo-endothéliose ont été également démontrés dans le sérum des pouletssurvivants. Ni le virus de la maladie de Marek, ni les virus du groupe leucoses-sarcome aviaires n'ont été mis en évidence dans les cultures de tissus ou dans lesessais de transmission au poulet. Ces tumeurs lymphoréticulaires transmissiblesde l'oie domestique semblent être associées à l'infection par un nouveau virus dugroupe de la réticulo-endothéliose désigné sous le nom de virus de la réticulo-endothéliose Hungrie-2 (REV-H-2).
Dow
nloa
ded
by [
Sout
hern
Tai
wan
Uni
vers
ity o
f Sc
ienc
e an
d T
echn
olog
y] a
t 02:
01 2
7 O
ctob
er 2
014

Reticuloendotheliosis virus-associated lymphoma in goose 277
ZUSAMMENFASSUNGIsolation eines reticuloendothelioseähnlichen Virus aus natürlich
vorkommenden lymphoreticulären Tumoren von HausgänsenIn einer Herde erwachsener Gänse wurden natürlich vorkommende lymphoreti-culäre Tumoren festgestellt. Diffuse oder knötchenförmige lymphomähnlicheTumoren traten in Milz, Leber, Pankreas, Darm aber seltener in adderen innerenOrganen auf. Osteopetroseähnliche Veränderungen und Zeichen anderer Krank-heiten, wie z.B. Amidostomiasis, Enteritis, Arthritis und Luftsackentzündungwurden gleichzeitig beobachtet.
Die tumorösen Veränderungen bestanden aus Proliferationen undifferenzierterLymphoblasten und wenig differenzierter Plasmablasten des Bursalymphozyten-systems. Die Tumorzellproliferation war gewöhnlich von einer Atrophie des Paren-chyms und gelegentlich von Amyloidablagerungen entlang der Kapillarwände vonMilz und Leber begleitet. In Gewebekulturen wurden chloroformempfindliche,typische C-Typ Viruspartikel von ca. 100 nm isoliert. Die Isolate inkorporierten3H-Uridine, bildeten eine Bande von ca. 1.19 bis 1.20 g/cm3 bei der Sucrose-gardienten-Ultrazentrifugation und zeigten Antigenverwandtschaft mit der Reti-culoendotheliosevirus Gruppe der aviaren Retroviren.
Bei Übertragungsversuchen mit Hühnern und Gänsen verursachte eines der vieruntersuchten reticuloendotheliosevirus-positiven Zellmaterialien aus Feldfälleninnerhalb von 4 Wochen nach Infektion eine tödliche neoplastische Krankheitmit einer ähnlichen Zytologie wie die der natürlichen Krankheit. Die anderen dreiZellmaterialien und die in Gewebekulturen vermehrten Virusisolate induziertenentweder ein Verzwergungssyndrom oder eine ähnlich chronisch verlaufendeNeoplasie wie bei der natürlichen Gänsekrankheit während des Versauchszeitraumesvon 30 Wochen. Das Reticuloendotheliosevirus wurde aus den experimentellinfizierten Tieren reisoliert. In Seren überlebender Tiere konnten Antikörpergegen den T-Stamm des Reticuloendotheliosevirus nachgewiesen werden. In denGewebekulturen und in den experimentell infizierten Tieren konnte weder dasMark Herpesyirus noch aviäres Leukose-Sarkomvirus nachgewiesen werden. Diebeschriebenen übertragbaren lymphoreticulären Tumoren von Hausgänsen scheinenmit der Infektion eines neuen Isolates der Reticuloendotheliosevirus-Gruppe,das als Reticuloendotheliosevirus Ungarn-2 (REV-H-2) bezeichnet wird, in Zusam-menhang zu stehen.
RESUMEN
Aislamiento de virus semejantes al virus de la reticuloendoteliosis a partir detumores linforeticulares de ocurrencia natural en gansos domesticos
Se encontraron tumores linforeticulares de presentación natural en una parvadade gansos adultos. Se observaron tumores de tipo linfoma difusos o nodularesen bazo, hígado, páncreas, intestinos y raramente en otras órganos internos. Tam-bién se observaron lesiones tipo osteopetrosas y signos de otras enfermedadestales como amidostomosis, enteritis, artritis y aerosoculitis.
Las lesiones tumorales consistieron en proliferaciones de linfoblastos indiferencia-dos y de plasmablastos ligeramente diferenciados de linfocitos del sistema bursal.La prolifereación de células tumorales fue usualmente asociada con atrofia paren-quimatosa y ocasionalmente con deposiciones amiloides a lo largo de las paredesde los capilares del bazo y del hígado. Se aislaron partículas víricas del tipo C deaproximadamente 100 nm en cultivo de tejidos sensibles a cloroformo.
Dow
nloa
ded
by [
Sout
hern
Tai
wan
Uni
vers
ity o
f Sc
ienc
e an
d T
echn
olog
y] a
t 02:
01 2
7 O
ctob
er 2
014