isolation, characterization and genome sequencing of a
TRANSCRIPT

Isolation characterization and genome sequencing of a soil-borne
Citrobacter freundii strain capable of detoxifying trichothecene mycotoxins
by
Rafiqul Islam
A Thesis
Presented to
The University of Guelph
In Partial Fulfilment of Requirements
for the Degree of
Doctor of Philosophy
in
Plant Agriculture
Guelph Ontario Canada
copy Rafiqul Islam April 2012
ABSTRACT
ISOLATION CHARACTERIZATION AND GENOME SEQUENCING OF A SOIL-
BORNE CITROBACTER FREUNDII STRAIN CAPABLE OF DETOXIFIYING
TRICHOTHECENE MYCOTOXINS
Rafiqul Islam Advisors
University of Guelph 2012 Dr K Peter Pauls
Dr Ting Zhou
Cereals are frequently contaminated with tricthothecene mycotoxins like
deoxynivalenol (DON vomitoxin) which are toxic to humans animals and plants The goals
of the research were to discover and characterize microbes capable of detoxifying DON under
aerobic conditions and moderate temperatures To identify microbes capable of detoxifying
DON five soil samples collected from Southern Ontario crop fields were tested for the ability
to convert DON to a de-epoxidized derivative One soil sample showed DON de-epoxidation
activity under aerobic conditions at 22-24degC To isolate the microbes responsible for DON
detoxification (de-epoxidation) activity the mixed culture was grown with antibiotics at 50ordmC
for 15 h and high concentrations of DON The treatments resulted in the isolation of a pure
DON de-epoxidating bacterial strain ADS47 and phenotypic and molecular analyses
identified the bacterium as Citrobacter freundii The bacterium was also able to de-epoxidize
andor de-acetylate 10 other food-contaminating trichothecene mycotoxins
A fosmid genomic DNA library of strain ADS47 was prepared in E coli and screened
for DON detoxification activity However no library clone was found with DON
detoxification activity The whole genome of ADS47 was also sequenced and from
comparative genome analyses 10 genes (characterized as reductases oxidoreductases and
deacetylase) were identified as potential candidates for DONtrichothecene detoxifying
enzymes Bioinformatic analyses showed that the major animal pathogenic locus of
enterocyte effacement (LEE) operon genes absent from the ADS47 genome Strain ADS47
has the potential to be used for feed detoxification and the development of mycotoxin
resistance cereal cultivars
iii
DEDICATIONS
The PhD thesis is dedicated to the departed souls of my mother MOZIBUN NESSA and
father-in-law DR MA AHAD MIAH
iv
ACKNOWLEDGEMENTS
Foremost I am deeply indebted to my advisors Prof K Peter Pauls and Dr Ting Zhou
for the continuous support of my PhD study for their endurance inspiration enthusiasm and
mentoring Their continued guidance has helped me in all aspects of my research and in
writing of this dissertation I sincerely express my gratitude to them for their co-operation
from the beginning to conclusion of the thesis and many other areas This will guide me in my
future professional career
My thesis committee guided me through all these years I would like to express my
gratitude to Prof Paul Goodwin for his insightful comments and advice on my thesis research
Special thanks to Dr Chris Young for teaching me chemistry chemical structures and
interpreting liquid chromatography ultraviolet mass spectra data I am sincerely grateful to
Dion Lepp for helping in analyzing the bacterial genome and sharing his knowledge and
valuable suggestions on writing the genomic chapter
I would like to express my appreciation to staff members of Dr Ting Zhou and Prof
K Peter Pauls who supported me in many aspects during my PhD study I am thankful to
Jianwei He Xiu-Zhen Li Honghui Zhu for collections and preparation of soil samples and
LC-UV-MS analyses of numerous samples Special thanks to Dr David Morris Johnston
Monje for sharing his knowledge discussions and camaraderie at coffee breaks
I wish to acknowledge the Natural Sciences and Engineering Research Council of
Canada (NSERC) for awarding me a prestigious scholarship for my PhD program and
Deanrsquos Tri-Council Scholarship (University of Guelph) which made my life easier during this
study I also would like to convey my thanks to Agriculture and Agri-Food CanadaBinational
v
Agricultural Research and Development Fund (AAFCBARD) project for providing me
financial support
Mike Bryan a very supportive person helped provide solutions to my software issues
that improved my exposure and caliber at conference awards competitions Several student
competition awards honored and inspired me to work hard and lead to the successful
completion of my thesis research I would like to convey my appreciation to the judges of
different student award committees who recognized my work They are Society for Industrial
Microbiology and Biotechnology conference 2010 (USA) Food Safety Research Forum-
Ontario Ministry of Agriculture and Rural Affairs annual meeting 2010 (Canada)
International Mycological Conference 2009 (UK) and Southwestern Ontario Regional
Association of Canadian Phytopathological Society annual meeting 2009-2011 (Canada)
I would like to express my very special thanks to Prof Golam Hafeez Kennedy my
brother-in-law who is like my elder brother In many aspects I am indebt to him for his
cooperations which aided me to achieve my current status
Lastly and most importantly I wish to give my special thanks to my wife Mimi and
lovely daughter Racy whose endless love sacrifice support and encouragement enabled me
to cross the tough long road and made possible to achieve my goal and aspirations Racy
once you told me that one of your desires for me is to finish my PhD which is done now I
know you want to be a scientist which is a great ambition I wish you all the best in your
future aims Racy
vi
TABLE OF CONTENTS
DEDICATIONShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipiii
ACKNOWLEDGEMENTShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipiv
TABLE OF CONTENTShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipvi
LIST OF TABLEShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipix
LIST OF FIGUREShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipx
LIST OF ABBREVIATIONShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipxiii
10 GENERAL INTRODUCTION AND LITERATURE REVIEW helliphelliphelliphelliphelliphelliphelliphellip1
11 Backgroundhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip1
12 Plant pathogenesis and trichothecene productionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip3
13 Biosynthesis of food contaminating TChelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip6
14 Occurrence of TC mycotoxins in food and feed helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip11
15 Significance of deoxynivalenol and trichotheceneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip16
16 Toxic effects of deoxynivalenol and trichotheceneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip19
17 Cellular and molecular mechanisms of DON toxicity in animals and plantshelliphelliphelliphellip22
18 Strategies to control Fusarium head blight and trichotheceneshelliphelliphelliphelliphelliphelliphelliphelliphellip26
19 Isolation of DONTC de-epoxidating micro-organisms and geneshelliphelliphelliphelliphelliphelliphelliphellip28
110 The bacterial species Citrobacter freundiihelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip30
111 Hypotheses and objectiveshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip31
vii
20 AEROBIC AND ANAEROBIC DE-EPOXIDATION OF MYCOTOXIN
DEOXYNIVALENOL BY BACTERIA ORIGINATING FROM AGRICULTURAL
SOIL33
21 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip33
22 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
23 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip37
24 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip45
25 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip62
26 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip64
30 ISOALTION AND CHARACTERIZATION OF A NOVEL SOIL BACTERIUM
CAPABLE OF DETOXIFYING ELEVEN TRICHOTHECENE MYCOTOXINS65
31 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip65
32 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip66
33 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip68
34 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip75
35 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip96
36 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip100
40 CONSTRUCTION AND FUNCTIONAL SCREENING OF GENOMIC DNA
LIBRARY FOR ISOLATION OF DON DE-EPOXYDIZING GENE(S) FROM
BACTERIAL STRAIN ADS47helliphelliphelliphelliphelliphelliphelliphellip102
41 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip102
42 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip103
43 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip104
viii
44 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip112
45 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip118
46 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip121
50 GENOME SEQUENCING CHARACTERIZATION AND COMPARATIVE
ANALYSIS OF A TRICHOTHECENE MYCOTOXIN DETOXIFYING
CITROBACTER FREUNDII STRAIN ADS47helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip122
51 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip122
52 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip123
53 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip125
54 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip129
55 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip162
56 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip168
60 CONCLUSIONhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip171
61 Summary helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip171
62 Research contributions helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip174
63 Future workhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip175
70 REFERENCEShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip176
ix
LIST OF TABLES
Table 1-1 Fusaria produced type-A and type-B trichothecene mycotoxinshelliphelliphelliphelliphelliphelliphellip9
Table 1-2 Worldwide occurrence of deoxynivalenol (DON) in animal feedshelliphelliphelliphelliphellip12
Table 1-3 Occurrence of DON in Canadian grains grains foods and beershelliphelliphelliphelliphelliphelliphellip13
Table 1-4 Legislative maximum tolerated levels of trichothecene mycotoxins in Canadahellip14
Table 1-5 In vitro and in vivo synergistic effects of different food contaminating
mycotoxins helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip18
Table 3-1 Microbial transformation of type-A and type-B tricthothecene mycotoxins
after 72 h of incubation under aerobic conditions and at room temperaturehelliphelliphelliphelliphelliphellip95
Table 4-1 Analysis of Fosmid DNA library for deoxynivalenol (DON) de-epoxidation
capability helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip117
Table 5-1 General features of C freundii strain ADS47 genomehelliphelliphelliphelliphelliphelliphelliphelliphellip132
Table 5-2 Genome comparisons between strain ADS47 and five Citrobacter species141
Table 5-3 Functional categorized gene abundances among the six Citrobacter genomeshellip145
Table 5-4 Unique genes of C freundii strain ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip156
Supplemental Table S5-1 List of unique genes of strain ADS47 gene IDs and their
functional roleshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip169
x
LIST OF FIGURES
Figure 1-1 Structures of trichothecene mycotoxins produced by Fusariahelliphelliphelliphelliphelliphelliphelliphellip7
Figure 1-2 Cellular and molecular mechanisms of deoxynivalenol toxicity in
eukaryotic organismshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip23
Figure 2-1 Mechanism of microbial DON de-epoxidationhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip36
Figure 2-2 LC-UV-MS chromatographic analysis of DON and its de-epoxidated product
dE-DON obtained by culturing with selected soil microbeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip46
Figure 2-3 Time course of relative biotransformation of DON to dE-DON by the initial
soil suspension culture in MSB mediumhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip49
Figure 2-4 Microbial de-epoxidation of DON after five days of incubationhelliphelliphelliphelliphelliphellip51
Figure 2-5 Analysis of microbial population reduction by terminal restriction fragment
length polymorphism (T-RFLP)helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip53
Figure 2-6 Microbial genera identified in the enriched DON de-epoxidizing microbial
culturehelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip56
Figure 2-7 Effects of culture conditions on DON to dE-DON transformation by the
enriched microbial culturehelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip58
Figure 2-8 Microbial DON to dE (de-epoxy)-DON transformation activity under aerobic
and anaerobic conditionshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip61
Figure 3-1 Colony color shape and size of the mycotoxin degrading bacterial strain
ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip76
Figure 3-2 Dendrogram based on 16S-rDNA sequences for strain ADS47 and 11
reference Citrobacter specieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip77
xi
Figure 3-3 Effects of DON concentration on bacterial growth and de-epoxidation
activitieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip79
Figure 3-4 Bacterial growth and DON de-epoxidation capabilities under aerobic and
anaerobic conditions helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip82
Figure 3-5 Effects of media on bacterial growth and DON de-epoxidation activityhellip85
Figure3-6 Effects of temperature on microbial growth and DON de-epoxydation helliphelliphellip87
Figure 3-7 Influence of pH on microbial growth and DON de-epoxydation capabilityhellip89
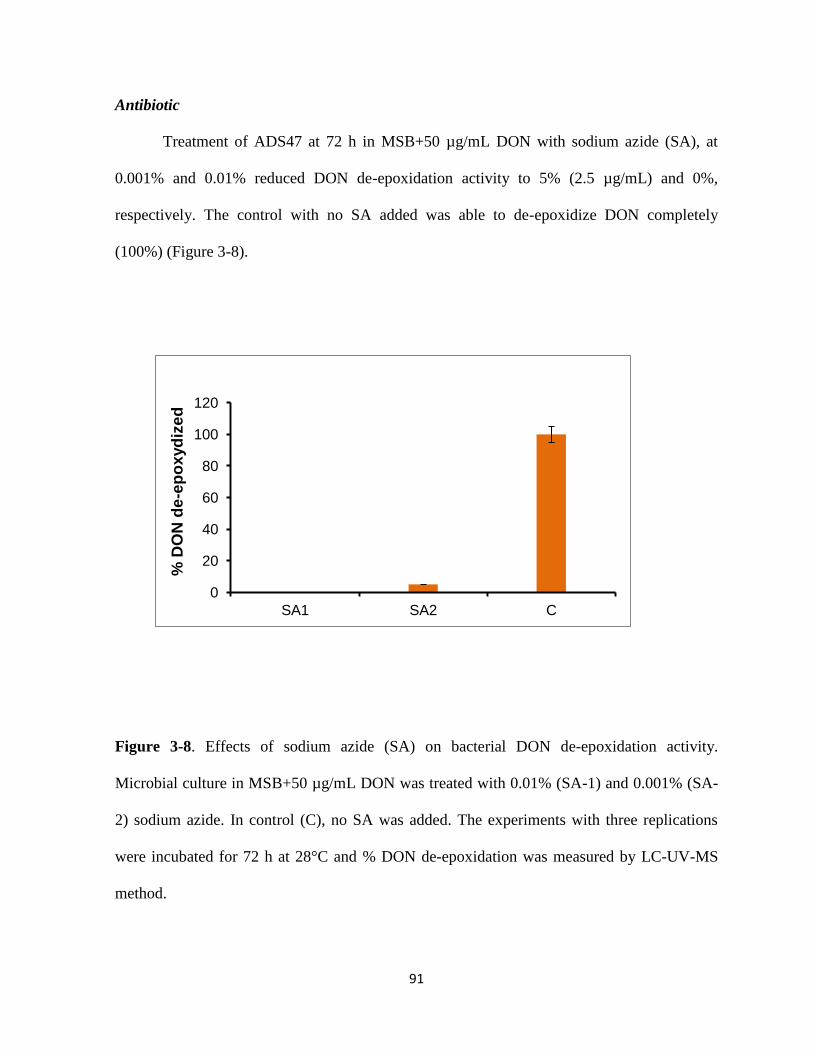
Figure 3-8 Effects of sodium azide (SA) on bacterial DON de-epoxidation activityhelliphelliphellip91
Figure 3-9 Determination of DON to deepoxy-DON transformation by bacterial culture
filtrate and cell extracthelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip92
Figure 3-10 Determination of microbial de-epoxidation and de-acetylation of a type-A
trichothecene mycotoxin (HT-2 toxin) by LC-UV-MShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip94
Supplemental Figure S3-1 Flow chart depicted the procedures of enrichment and isolation
of DON de-epoxidation bacterial strain ADS47helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip101
Figure 4-1 Schematic overview of the construction and functional screening of Fosmid
DNA libraryhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip105
Figure 4-2 Genetic map of copy controlled Fosmid vector (pCC1FOS)helliphelliphelliphelliphelliphelliphellip108
Figure 4-3 Preparation of 40 kb size insert DNA from a DON de-epoxidizing bacterium113
Figure 4-4 Production of genomic DNA library in E coli EPI300-T1 host strainhelliphelliphelliphellip114
Figure 4-5 Confirmation of the Fosmid DNA library construction in E coli helliphelliphelliphelliphellip 116
Figure 5-1 Co-linearity between C freundii strain ADS47 genome and C koseri helliphelliphellip129
Figure 5-2 Circular genome map of C freundii strain ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphellip133
xii
Figure 5-3 Size distribution of the predicted genes identified in C freundii strain
ADS47helliphelliphellip 135
Figure 5-4 Functional classification of the predicted coding sequences (CDS) of
C freundii strain ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip137
Figure 5-5 Phylogeny of the trichothecene detoxifying C freundii strain ADS47 139
Figure 5-6 Comparisons of coding sequences between C freundii strain ADS47 and
five Citrobacter specieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip143
Figure 5-7 Determination of the locus enterocyte effacement (LEE) operons in
Citrobacter species by comparing C rodentium helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip151
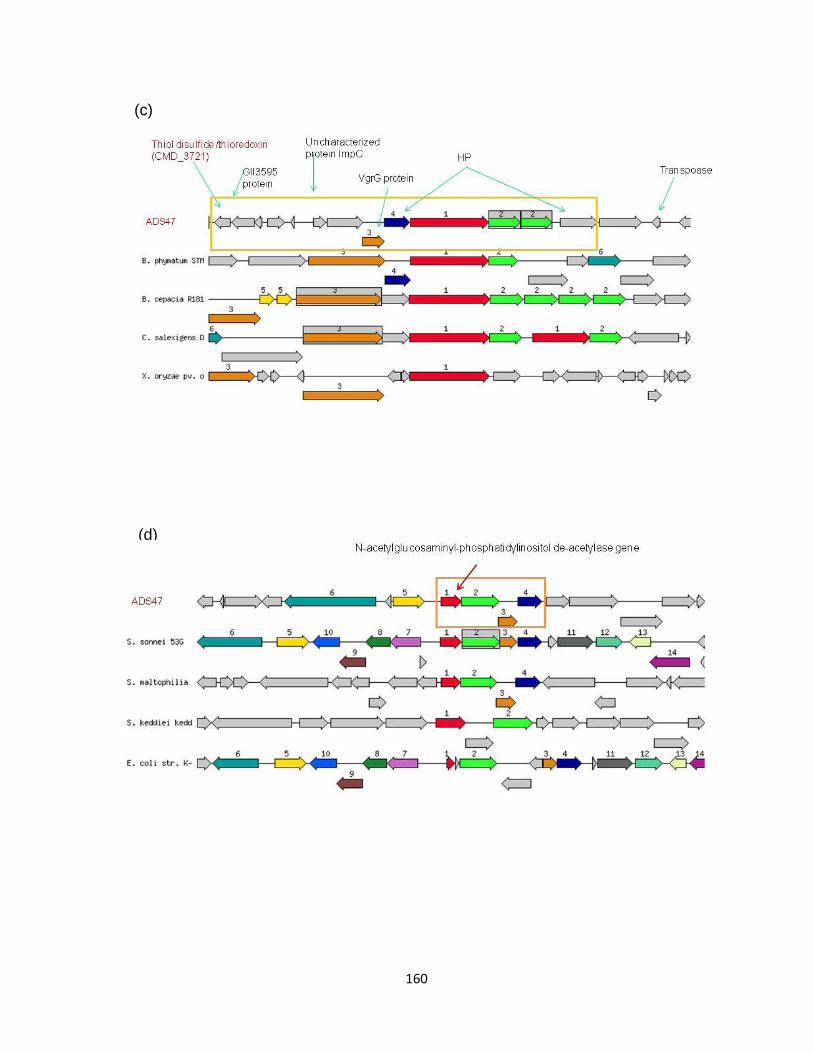
Figure 5-8 Locations of unique genes of C freundii strain ADS467helliphelliphelliphelliphelliphelliphelliphellip157
Figure 5-9 Unique genomic regions and associated genes in strain ADS47 compared
to five Citrobacter specieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip159
xiii
LIST OF ABBREVIATIONS
APCI= atmospheric pressure chemical ionization
AE= attaching and effacing
A= aerobic conditions
ABC= ATP-binding cassette
ACT= artemis comparative tools
AN= anaerobic conditions
3-ADON= 3-acetyl-deoxynivalenol
15-ADON= 15-acetyl-deoxynivalenol
ATA= alimentary toxic aleukia
bp= base pair
BGI= Beijing genomics institute
BLAST= basic local alignment search tool
BRIG= blast ring image generator
Bw= body weight
C= carbon
CDS= coding sequence
CFIA= Canadian food inspection agency
CHEF= clamped homogeneous electric fields
CMB= corn meal broth
CVTree= composition vector tree
DAS= diacetoxyscirpenol
dH2O= distilled water
DOGT1= UDP-glucosyltransferase
DON= deoxynivalenol
dsRNA= double stranded RNA
dE= de-epoxy
dE-DON= de-epoxy DON
E coli= Escherichia coli
ER=endoplasmic reticulum
FAO= Food amp Agriculture Organization
FM= full medium
FHB= Fusarium head blight
FUS= fusarenonX
gDNA= genomic DNA
GC= guanine cytosine
GER= Gebberella ear rot
GPI= glycosylphosphatidylinositol
h= hour
Hck= hematopoietic cell kinase
Kb= kilobase
xiv
Kg= kilogram
LC-UV-MS= liquid chromatography-ultraviolet-
mass spectrometry
LEE= locus of enterocyte effacement
LD50= lethal dose of 50 animal
LB= Luria Bertani
LMP= low melting point
MAPK= mitogen-activated protein kinase
microg= microgram
mg= milligram
mL= milliliter
microL= microliter
microM= micromole
min= minute
Mb= megabases
MCS=multiple cloning site
MSA= MSB+ agar
MSB= mineral salts broth
MM= minimal medium
MSC= mineral salts + 03 Bacto Peptone
MS-DON= mineral salts + 200 microgmL DON
mRNA= messenger RNA
MSY= mineral salts + yeast extract
mz= mass ion ratio
N= nitrogen
Na-Cl= sodiumndashclorine
NADH= (reduced) nicotinamide adenine
dinucleotide
NAD(P)= nicotinamide adenine
dinucleotide phosphate
NCBI= national center for biotechnology
information
NEO= neosolaniol
ng= nanogram
NIV= nivalenol
OD600nm= optical density at 600
nanometer wavelength
OMAFRA= Ontario Ministry of
Agriculture and Rural Affairs
Paa= phenylacetic acid
PCR= polymerase chain reaction
PDB= potato dextrose broth
ppm= parts per million
PFGE= pulsed field gel electrophoresis
PKR= protein kinase
xv
PNDO= pyridine nucleotide-disulfide
oxidoreductase
QTL= quantitative trait locus
RAST= rapid annotation subsystem technology
RDP= ribosomal database project
Rpl3= 60S ribosomal protein L3
rRNA= ribosomal RNA
s= second
SA= sodium azide
SDR= short chain dehydrogenasereductase
SE= soil extract
SIM= selected ion monitoring
SOAP= short oligonucleotide analysis package
sp= species
sRNA= small RNA
TC= trichothecene
TCA= tricarboxylic acid cycle
TSB= tryptic soy broth
TF= transcription factor
tRNA= transfer RNA
TIC= total ion count
TRAP= tripartite ATP-independent
periplasmic
T-RFLP=terminal restriction fragment
length polymorphism
TRI101= trichothecene 3-O-
acetyltransferase
YEB= yeast extract broth
VER= verrucarol
VKOR= vitamin K epoxide reductase
wv= weight by volume
1
CHAPTER 1
10 GENERAL INTRODUCTION AND LITERATURE REVIEW
11 Background
According to the Food and Agriculture Organization (FAO) of the United Nations
more than 25 of the worldrsquos food crops are contaminated with mycotoxins every year
causing serious adverse effects on livestock industry crop production and international trade
(Boutrif and Canet 1998 Desjardins 2006 Binder et al 2007 Surai et al 2010) The negative
effects of mycotoxins on the consumers are diverse including organ toxicity mutagenicity
carcinogenicity neurotoxicity teratogenicity modulation of the immune system reproductive
system effects and growth retardation Mycotoxins are toxic secondary metabolites produced
by various fungal species during crop colonization in field andor post harvest conditions
(FAO 1991 Barrett 2000) Aspergillus Penicillum and Fusarium are the predominant fungal
genera that accumulate economically important mycotoxins such as aflatoxins zearalenone
trichothecenes fumonisins ochratoxins and ergot alkaloids in agricultural commodities
Humans and animals are principally exposed to mycotoxins through consumption of
contaminated foodfeed and animal products such as milk meat and eggs (Fink-Gremmels
1989 Hollinger and Ekperigin 1999)
Trichothecenes (TCs) produced by a variety of toxigenic Fusarium species are
probably the most food and feed contaminating mycotoxins in the temperate regions of
America Europe and Asia (Creppy 2002 Yazar and Omurtag 2008) The fungal pathogen
accumulates various type-A (eg T-2 toxin HT-2 toxin) and type-B trichothecenes (eg
2
deoxynivalenol nivalenol and their derivatives) in cereals Among them type-B
trichothecenes particularly deoxynivalenol (DON) andor its derivatives appear to be the
most commonly found mycotoxins in grain and grain products worldwide impairing human
health and productivity of farm animals (Placinta et al 1999 Sherif et al 2009) A number of
human and animal disease outbreaks associated with TC contaminated grain food and feed
consumption have been reported in Asia (Japan India and central Asia) Europe New
Zealand and United States of America (USA) (Pestka and Smolinski 2005 Desjardins 2006)
A severe outbreak of Alimentary Toxic Aleukia (ATA) mycotoxicosis killed 100000 people
in the Orenburg region of Russia (Eriksen 2003) In 1996 losses of over $100 million were
estimated in Ontario because of costs related to managing DON (Schaafsma 2002)
Historically T-2 toxin DON and nivalenol (NIV) have likely been used as a biological
warfare in Laos Kampuchea and Afghanistan and there is concern because of the potential
use of these compounds for bioterrorism (Wannemacher et al 2000 Tucker 2001)
Despite the extensive efforts that have been made to prevent TC contamination with
the current varieties and production practices its presence in agricultural commodities is
likely unavoidable There are no crop cultivars currently available that are completely
resistant to Fusarium head blightGibberella ear rot (FHBGER) diseases Chemical or
physical methods cannot safely remove TC mycotoxins from grain completely or they are not
economically viable (Varga and Toacuteth 2005) Therefore an alternative strategy to address the
issue is of great importance (Awad et al 2010 Karlovsky 2011)
3
12 Plant pathogenesis and trichothecene production
Cereals infested by the ascomyceteous fungi may cause Gibberella ear rot (GER) in
maize (Zea mays L) and Fusarium head blight (FHB also known as lsquoscabrsquo) of small grain
cereals [wheat (Triticum aestivum L) barley (Hordeum vulgare L)] and other small grain
cereals (Goswami and Kistler 2004) As many as 17 fungal species have been reported to be
associated with FHBGER which include F graminearum Schwabe [telemorph Gibberella
zeae (Schwein) Petch] F culmorum (WG Smith) Sacc F sporotrichioides Scherb F
poae (Peck) Wollenw Microdochium nivale (Fr) Samuels amp IC Hallett and F avenaceum
(Fr) Sacc (Osborne and Stein 2007 Audenaert et al 2009) The two species F
graminearum and F culmorum are the principal species causing crop damage Among them
F graminearum is the widely adapted and most damaging species across the world including
North America (Dyer et al 2006) F culmorum tend to be the main species in the cooler parts
of North Central and Western Europe (Wagacha and Muthomi 2007)
The fungi overwinter mainly on crop residues as saprophytes where they produce
asexual (micro- and macro-conidia) andor sexual (ascospores) fruiting structures which act
as the initial source of inoculums in the subsequent crop season (Sutton 1982 Wagacha and
Muthomi 2007) Ascospores are primarily dispersed by air (Trail et al 2002) and conidia
move by rain-splash (Paul et al 2004) After they land on maize silks and florets of small
grain cereals infection occurs through natural openings (eg stomata) or occasionally by
direct penetration The pathogens can invade maize kernels through wounds caused in the
husks by insects or birds Infection may occur throughout the crop growing season but
disease incidence and severity is facilitated by warm temperatures (gt20degC) high humidity
(gt80) and frequent precipitation that coincides with anthesis (Osborne and Stein 2007
4
Wagacha and Muthomi 2007) The Fusarium pathogen initially infects as biotroph which
becomes a necrotroph as the disease progresses After penetration Fusaria produce different
extracellular cell wall degrading enzymes that trigger infection and hyphal progression
towards the developing earkernel where further colonization occurs and development of
FHBGER diseases (Voigt et al 2005 Miller et al 2007 Kikot et al 2009) FHB disease is
characterized by a bleached appearance around the middle of the head Severely infected
grains have a pinkish discoloration and can be shriveled In maize the disease produces a
reddishndashbrown discoloration of the kernels and progresses from the tip to the bottom of
affected ears
The fungi associated with FHB and GER contaminate cereals by accumulating various
host-nonspecific TC mycotoxins (Brennen et al 2007) TCs have typically been associated
with cereals grown in temperate climatic conditions including Canada (Doohan et al 2003)
DON production by F graminearum and F culmorum is maximal at warm temperatures (22-
28˚C) high grain moisture contents (gt17) and humid conditions (gt70 RH) (Martins and
Martins 2002 Hope et al 2005 Llorens et al 2006) Grain can be contaminated with TCs in
the field as well as in storage if conditions for fungal growth are optimal and grain contains
high moisture (gt17) (Larsen et al 2004 Birzele et al 2002)
Populations of FHBGER causing pathogens are dynamic Genetic variability between
and within Fusarium species appears to have occurred to allow for their adaptation to new
agro-climates (Desjardins 2006 Fernando et al 2011) Three strain specific Fusarium
chemotypes NIV 3-ADON (3-acetyl-deoxynivalenol) and 15-ADON (15- acetyl-
deoxynivalenol) have been identified which correlate with the geographical distributions
(Starkey et al 2007 Zhang et al 2007 Audenaert et al 2009) The NIV chemotype mainly
5
produced by F culmorum accumulates NIV and 4-NIV (Fusarinon X) is predominant in
Asia Africa and Europe (Lee et al 2001) The 3-ADON chemotype which synthesizes DON
and 3-ADON is the dominant strain in EuropeFar East (Larsen et al 2004) In North
America 15-ADON is the main chemotype which produces DON and 15-ADON (Goswami
and Kistler 2004) Significant Fusarium population shifting was observed in the past and in
recent years (Kant et al 2011) A dynamic change from F culmorum to F gramninearum
and DON to NIV producing chemotyes occurred in North West European countries The 3-
ADON chemotype which is uncommon in North America was found in North Dakota and
Minnesota (Goswami and Kistler 2004) F pseudograminearum which historically occurs in
Australia Africa and North West Pacific America has been found in Western Canada (Larsen
et al 2004 Kant et al 2011) In Manitoba a replacement of 15-ADON chemotype by the
more phytoxic 3-ADON type was observed in recent years (Fernando et al 2011) All
together the FHBGER associated pathogen complex is highly dynamic which not only
reduces crop yield but also decreases grain quality by accumulating various mycotoxins
6
13 Biosynthesis of food contaminating TC
TCs are fungal secondary metabolites that were initially identified in Trichotheceium
roseum Thus the name of this class of compounds follows the name of the genus
Trichotheceium (Freeman and Morrison 1949) There have been 180 TCs identified in various
fungal species belonging to the genera Fusarium Myrothecium Stachybotrys Trichoderma
and Thrichothecium (Eriksen 2003 Eriksen et al 2004) The FHBGER associated TCs are
mainly synthesized by different Fusaria and are characterized as tricyclic sesquiterpenoids
with a double bond at position C9-10 and a C12-13 epoxide ring
7
Figure 1-1 Structures of trichothecene mycotoxins produced by Fusaria (a) General
structure of trichothecenes R1 = H OH OiValeryl keto R2 = H OH R3= OH OAc R4 =
H OH OAc R5= OH OAc (b) Type-A trichothecene mycotoxin (c) Type-B trichothecene
mycotoxin which contain a carbonyl functional group at C8 position (Young et al 2007)
(a) (b)
(c)
8
The fungal metabolites are small in size (molecular weight 250-550) heat stable and
non-volatile molecules (Pestka 2007 Yazar and Omurtag 2008) Food contaminating TCs are
divided into two groups type-A and type-B (Figure 1-1b and 1c) Type-A is characterized by
the lack of a ketone function at C8 position and include T-2 toxin HT-2 toxin
diacetoxyscirpenol (DAS) and neosolaniol (NEO) which are mainly produced by F
sporotrichioides and F poae (Creppy 2002 Quarta et al 2006) The most frequently found
TC belongs to type-B which contains carbonyl functions at C8 positions DON NIV and
their acetylated derivatives The two types are further differentiated by the presence or
absence of functional hydroxyl (-OH) and acetyl (CH3-CO-) groups at C-3 C-4 C-7 C-8 and
C-15 positions (Table 1-1)
The syntheses of various TCs are speciesstrain specific and are genetically controlled
but influenced by environmental conditions (Desjardins 2006 Paterson and Lima 2010) Until
now 15 Fusaria catalytic genes and regulators involved in TC productions have been
identified (Merhej et al 2011) These biosynthetic genes are located at three loci on different
chromosomes The core trichothecene (TRI) gene cluster is comprised of 12 genes localized
in a 25-kb genomic region (Kimura et al 2001) The remaining three genes are encoded by
the TRI1-TRI16 and TRI101 loci (Gale et al 2005 Alexander et al 2009) Fusaria produce
TC following a number of oxygenation isomerization acetylation hydroxylation and
esterification reaction steps (Desjardins 2006 Desjardins and Proctor 2007 Kimura et al
2007 Merhej et al 2011) In brief farnesyl pyrophosphate is initially cyclized into
trichodiene by TRI5 which codes for trichodiene synthase In the next four steps
isotrichotriol is produced by TRI4 (a multifunctional cytochrome P450) via C-2 hydroxylation
and 1213 epoxidation followed by two hydroxylation reactions
9
Table 1-1 Fusaria produced type-A and type-B trichothecene mycotoxins Molecular weight
(MW) and acetyl (-R) hydroxyl (-OH) and ketone (=O) functions at different carbon (C)
position of the tricyclic rings A-types T-2 toxin HT-2 toxin DAS (diacetoxyscirpenol)
NEO (neosolaniol) VER (verrucarol) B-type DON (deoxynivalenol) 3-ADON (3-acetyl-
deoxynivalenol) 15-ADON (15-acetyl-deoxynivalenol) NIV (nivalenol) and FusX
(fusarenon X) (Young et al 2007)
Trichothecenes MW R1C3 R2C4 R4C7 R3C15 R5C8
Type-A
T-2 toxin 466 OH OCOCH3 H OCOCH3 OCOCH2CH(CH3)2
HT-2 toxin 424 OH OH H OCOCH3 OCOCH2CH(CH3)2
DAS 366 OH OCOCH3 H OCOCH3 H
NEO 382 OH OCOCH3 OH OCOCH3 H
VER 266 H OH H OH H
Type-B
DON 296 OH H OH OH =O
3-ADON 338 OCOCH3 H OH OH =O
15-ADON 338 OH H OH OCOCH3 =O
NIV 312 OH OH OH OH =O
FusX 354 OH OCOCH3 OH OH =O
10
The next two non-enzymatic reactions results in isotrichodemol Eventually the
trichothecene skeleton is produced via the formation of a C-O bond between the C-2 oxygen
and C-11 Subsequent acetylation at C-3 by TRI101 hydroxylation at C-15 by TRI1 and
catalytic activity of TRI3 leads to calonectrin production The pathway described above is
common for both type-A (eg T-2 toxin HT-2 toxin) and type-B (eg DON NIV and their
acetylated derivatives) TCs
The following steps are speciesstrain specific In DON producing F graminearum
strains calonectrin is converted to either 3-ADON or 15-ADON followed by DON production
by the catabolic activities of TRI1 and TRI8 The biosynthesis of 3-ADON or 15-ADON is
controlled by variation in the esterase coding sequences of TRI8 F graminearum produces
NIV from calonectrin via synthesis of 4-NIV followed by NIV by TRI1 and TRI8 gene
actions NIV can be produced either from DON by hydroxylation at C-4 or via
transformation of calonectrin to 315-diacetoxyscirpenol (315-DAS) followed by addition of
a ketone at C-8 position by the activity of the TRI7 and TRI13 genes 315-DAS is the
principal substrate for synthesis of type-A TCs by F sporotrichioides the main type-A
producer The biosynthetic pathway of T-2 toxin involves C-4 acetylation of 315-DAS by the
TRI7 gene product and TRI8 triggered by TRI1 315-DAS is also the substrate for HT-2
toxin synthesis In the absence of TRI7 expression HT-2 toxin is produced through
hydroxylation of 315-DAS and isovalerate addition to C-8 position (Foroud and Eudes 2009)
11
14 Occurrence of TC mycotoxins in food and feed
TC producing Fusaria are adapted to variable climatic conditions and agricultural
ecosystems As a consequence this class of mycotoxins particularly DON is frequently
detected in foods and feeds including breakfast cereals bakery products snack foods beer
and animal feed derived from cereals in most part of the world (Pittet 2001 Desjardins 2006
Lancova et al 2008) A two-year survey during 2003-2005 conducted by Romer Lab Inc
revealed unacceptable levels of DON in animal feedsfeed raw materials (such as maize
wheat wheat bran) in Asia and Oceania (1062 microgg) central Europe (802 microgg) Mid-West
USA (55 microgg) northern Europe (551 microgg) and southern Europe (303 microgg) (Binder et al
2007) (Table 1-2)
Quite high levels of DON contamination (19 microgg) were detected in north Asian and
Chinese feedfeed materials Recently five-year on-farm studies in Switzerland showed that
74 of the wheat grain and straw samples were contaminated with high levels of DON
(average 5 microgg) The amount is higher than the European maximum limit of 125 microgg
(Vogelgsang et al 2011)
The occurrences of TCs in Canadian agricultural commodities are indicated in Table
1-3 Seventeen surveys conducted during the 1980s and 1990s showed up to 100 DON
incidence in Ontario wheat and maize samples with an 18 average incidence (Scott 1997) A
three-year survey recently (2006-08) conducted by the Ontario Ministry of Agriculture and
Rural Affairs (OMAFRA) detected high levels of DON in spring (maximum 26 microgg) and
winter wheat (maximum 29 microgg) samples (Ana Matu personal communication) The DON
levels were much higher than Health Canadarsquos maximum limit of 2 microgg for uncleaned soft
wheat for staple foods (Table 1-4)
12
Table 1-2 Worldwide occurrence of DON in animal feed Feed and feed raw material
samples were collected from animal farmsfeed production sites during 2003-2005 The
survey was conducted by Romer Labs Inc (httpwwwromerlabscom)
Country
region
Sample
type
Incidence
()
Maximum
DON content
(microgg)
Mean DON
content
(microgg)
Survey
period
Source
North
AsiaChina
Animal
feed
071 1899 092
Binder et
al 2007
South East
Asia
014 152 016
South Asia 070 1062 049
Oceania 2003-05
Australia 041 186 030
Northern
Europe
070 551 059
Central
Europe
066 802 057
Southern
Europe
052 303 030
13
Table 1-3 Occurrence of DON in Canadian grains grain foods and beers Infant cereal foods
oat barley soy and multi-grain based foods
Province Sample
type
Incidence
()
Maximum
DON
contents
(microgg)
Multiple
mycotoxins
samples ()
Survey
period
Sources
Ontario Spring
Wheat
70 260 Yes 2006-08 OMAFRA1
Ontario Winter
wheat
49 290 Yes 2006-08 OMAFRA
Ontario Maize 78 140 Yes 2009-10 Limay-Rios
and
Schaafsma
2010
Ontario Maize - 1000 Yes 2011 AampL
Biologicals
Canada Breakfast
cereals
46 094 430 1999-01 Roscoe et al
2008
Canada Animal
feeds
73 425 Yes 1990-10 Zeibdawi et
al 2011
Canada Beers 58 0052 50 1990s Scott 1997
Canada Infant
Cereals
63 098 Yes 1997-99 Lombaert et
al 2003
1
Ontario Ministry of Agriculture and Rural Affairs
2 microgmL
14
Table 1-4 Legislative maximum tolerated levels of trichothecene mycotoxins in Canada
according to Canadian Food Inspection Fact Sheet April 23 2009 DON- deoxynivalenol
DAS- diacetoxyscirpenol
Products DON
(microgg)
T-2 toxin
(microgg)
HT-2 toxin
(microgg)
DAS
(microgg)
Uncleaned soft wheat for human
consumption
2 - - -
Cattlepoultry diets 5 lt1 01 lt1
Swinedairy cattle feeds 1 lt1 0025 lt2
15
High DON incidence (78) was observed in maize samples collected from
southwestern Ontario fields with moderate levels (lt1 microgg) of contamination during 2009 and
2010 (Limay-Rios and Schaafsma 2010) Some samples had high DON levels (14 microgg) In
the 2011 crop season extremely high levels of DON (gt100 microgg) were found in some
southern Ontario maize samples (AampL Biologicals London ON Personal communication)
In a Canadian Food Inspection Agency (CFIA) 20 year survey (1990-2010) 73 of
animal feed samples collected from feed mills farms and retail stores across Canada were
contaminated with DON and for more than 25 of the samples the mycotoxin levels were
above the maximum limit (ge 1 microgg) (Zeibdawi et al 2011) The highest level of DON (15
microgg) detected in the 2006-2007 samples were much lower than the levels found in the 1999-
2000 samples which contained a maximum of 425 microgg DON Importantly this study
reported a significant number of samples contaminated with multiple mycotoxins DON NIV
3-ADON 15-ADON T-2 toxin HT-2 toxin DAS and NEO
Earlier studies in the 1990s detected high levels of DON content in Eastern Canadian
cereals maize (maximum 175 microgg) wheat (maximum 916 microgg) barley (maximum 911
microgg) and oats (maximum 12 microgg) (Campbell et al 2002) The study identified a significant
number of samples (15-367) that contained multiple mycotoxins and up to 33 of the
wheat samples exceeded 1 microgg DON Low amounts but high incidences of TCs were found
in cereal-based foods collected from Canadian retail stores Sixty three percent of infant
cereal foods 46 of breakfast cereals and 58 of beers were contaminated with one or
multiple mycotoxins (Scott 1997 Lombaert et al 2003 Roscoe et al 2008) The maximum
amount of DON was found in infant foods (098 microgg) followed by breakfast cereals (094
microgg) and beers (005 microgmL)
16
15 Significance of deoxynivalenol and trichothecenes
For most cereals including wheat maize rice barley oats and rye the risk of
contamination with TCs resulted in grain quality reduction and FHBGER disease progression
(Bai et al 2002 Jansen et al 2005) The impacts of TC contamination in agricultural
commodities are diverse including reduced livestock production induced human illnesses
and enhanced FHB disease severity which may result in the reduction of crop yield Exposure
of farm animals (particularly pigs) to DON is a continuing threat to the livestock industries in
Canada USA and Europe (DrsquoMello et al 1999) Ingestion of TC-contaminated feeds
generally causes reduction of feed intake feed refusal poor feed conversion retardation of
weight gain increased disease susceptibility and reduced reproductive capacities (Binder et al
2007) High doses of DON result in acute toxic effects in pigs (eg emesis) while low doses
(ge1 microgmL) cause chronic effects (eg growth retardation in pigs) (Rotter et al 1996
Accensi et al 2006) Reduction of growth and reproduction ability was observed in
experimental rats exposured to much higher doses of TC (Sprando et al 2005 Collins et al
2006) Several animal disease outbreaks that occurred in different continents have been
attributed to consumption of feed contaminated with DON NIV and DAS (Desjardins 2006)
The consequences of human exposure to TCs include liver damage gastricintestinal
lesions and central nervous system toxicity resulting in anorexia nausea hypotensionshock
vomiting diarrhea and death (Pestka 2007) Human mycotoxicosis (illnesses and outbreaks)
by consumption of type-A and type-B TC mycotoxins in Europe New Zealand Russia
Japan South America India and Vietnam have been reported (Bhat et al 1989 Yoshizawa
1993 Luo 1994 Desjardins 2006) Severe outbreaks of Alimentary Toxic Aleukia (ATA)
associated with TC killed thousands of people in the Orenburg region of Russia during the
17
1930rsquos and 1940rsquos (Eriksen 2003) TCs (T-2 DON and NIV) were likely used for biological
warfare in Laos Kampuchea and Afghanistan and its potential application for bioterrorism is
an additional concern associated with TC (Tucker 2001 Bennett and Klich 2003)
TCs are phytotoxic and may contribute to FHB disease severity in cereals (Proctor et
al 1995 Maier et al 2006 Masuda et al 2007) Studies demonstrated that DON caused
wilting chlorosis inhibition of seedling growth and necrosis in cereals (Cossette and Miller
1995 Mitterbauer et al 2004 Rocha et al 2005) DON-mediated host cell damage and
subsequent necrotrophic fungal spread has been shown to lead to FHBGER disease
development and higher kernel damage (Procter et al 1995 Mitterbauer et al 2004 Maier et
al 2005) This suggests that DON contributes to yield losses as well as to reductions in the
market value of cereals (Osborne and Stein 2007) Severely infected grain may contain more
than 60 microgg DON which is unacceptable to the milling and brewing industries (Okubara et
al 2002) Significant crop losses due to FHB disease epidemics and mycotoxin contamination
have been reported in Ontario (Schaafsma 2002) FHB disease epidemics during 1998 to 2000
caused huge crop losses resulting in farm failures in the northern Great Plains in the USA
(Windels 2000) In that period FHB associated losses were estimated at US $ 27 billion in
the region (Nganje et al 2002)
Importantly Fusaria infested grains are often co-contaminated with multiple
mycotoxins which further increase health risks as interactions between the mycotoxins may
cause greater toxic effects than expected (Schollenberger et al 2006) This implies that an
apparently low level of contamination of a single mycotoxin can have severe effects on
consumers because of synergistic effects among multiple co-occuring mycotoxins (Surai et al
2010)
18
A number of studies demonstrated synergistic toxic effects among Fusarium
mycotoxins on pigs turkeys lamb chicks as well as in in vitro Caco-2 cell and yeast
bioassays (Table 1-5)
Table 1-5 In vitro and in vivo synergistic effects of different food contaminating mycotoxins
DON-deoxynivalenol DAS-diacetoxyscirpenol FB1-fumonisins 1 NIV-nivalenol ZEN-
zearelenone FA-fusaric acid Afl-aflatoxin
Mycotoxins source Interacting
mycotoxins
Experimental
animalcells
Effects Reference
Naturally contaminated
cereals
DON FA Pigs Inhibit growth Smith et al
1997
DON contaminated
wheat and FB1
DON FB1 Pigs Weight loss Harvey et al
1996
Culture materials
contained mycotoxins
T-2 FB1 Turkey poults Weight loss oral
lesion
Kubena et al
1997
Inoculated rice and
pure mycotoxin
DAS Afl Lamb Weight reduction Harvey et al
1995
Naturally DON
contaminated wheat
and pure T-2 toxin
DON T-2 Chicks Reduced body
weight significantly
Kubena et al
1989
Pure mycotoxins DAS Afl Chicks Reduced body
weight
Kubena et al
1993
Pure mycotoxins T-2 Afl Chicks Reduced weight
oral lesion
Kubena et al
1990
Pure mycotoxins DON
ZENFB1
Human intestinal
Caco-2 cells
Increased DNA
fragmentations
Kouadio et al
2007
Pure mycotoxins DON NIV Yeast bioassay Synergistic effects Madhyastha et
al 1994
Pure mycotoxins T-2 HT-2 Yeast bioassay Synergistic effects Madhyastha et
al 1994
19
16 Toxic effects of deoxynivalnol and trichothecenes
As small molecules TCs simply enter into cells and interact with different cell
components resulting in disruption of cellular functions and causing various toxic effects that
include inhibition of protein synthesis and ribosomal fragmentation (Eudes et al 2000
Minervini et al 2004 Pestka 2007) Simple absorption of DON into human intestinal Caco-2
cells and crossing human placenta barriers through passive diffusion has been reported
(Sergent et al 2006 Nielsen et al 2011)
TC may cause various toxicities in animals and plants including apoptosis
immunotoxicity chromatin condensation disruption of cellularmitochondrion membranes
disruption of gut tissues inhibition of protein and ribosome synthesis breakage of ribosome
and cell damage The toxicity of TC is influenced by structural variability (ie position
number and type of functional groups attached to the tricyclic nucleus and C-C double bond at
C9-10 position) (Desjardins 2006) Nevertheless epoxide function at the C12-13 position is
the key for TC cytotoxicity (Swanson et al 1988 Eriksen et al 2004)
Different in vitro and in vivo studies revealed that the method and period of exposure
amount of TC and types of celltissue and host species have significant effects on TCrsquos
cytotoxicity (Scientific Committee for Food 2002 Rocha et al 2005 Nielsen et al 2009)
Administration of 10 mgkg body weight (bw) NIV caused apoptosis in mice thymus Peyerrsquos
patch and spleen tissues (Poapolathep et al 2002) DON and NIV at 887 mgkg bw for 3 days
a week for four weeks induced immunotoxicity in experimental mice determined by
estimating increased immunoglobulins (IgA) and decreased IgM levels (Gouze et al 2007)
Exposure of mice to 1 mgkg bw of DON induced skeletal malformations and reduced mating
tendency while significant resorption of embryos observed at the dose of gt10 mgkg bw
20
(Khera et al 1982) At high acute doses (4972 mgkg bw) DONNIV and type-A
trichothecenes caused mice death because of intestinal haemorrhage necrosis of bone
marrowlymphoid tissues and kidneyheart lesions (Yoshizawa and Morooka 1973
Yoshizawa et al 1994 Forsell et al 1987 Ryu et al 1988) In a separate study 15-ADON
showed more toxicity than DON when the mycotoxins were orally administered to mice
(Forsell et al 1987)
Studies suggested that type-A TCs are more toxic in mammals than type-B which
means that functional variability at the C-8 position of TC has a significant role in toxicity
The toxicity of various TC mycotoxins in animals can be ranked in the order of T-2
toxingtHT-2 toxingtNIVgtDONge3-ADONgt15-ADON according to the results from human cell
lines and mice model systems bioassays (Ueno et al 1983 Pestka et al 1987 Nielsen et al
2009) Moreover animal feeding assays demonstrated that monogastric animals are most
sensitive to DON The order of sensitivity of animals to DON is
pigsgtrodentsgtdogsgtcatsgtpoultrygtruminants (Pestka 2005) The reasons for the variations in
DON toxicity species might be due to differences in metabolism absorption distribution and
elimination of the mycotoxin among animal species (Pestka 2007)
To my knowledge the mechanism of TC uptake by plant cells has not been studied
Nonetheless cereals are exposed to TC by toxigenic Fusaria infection which may cause
various toxicities such as cell damage wilting chlorosis necrosis program cell death and
inhibition of seedling growth in sensitive plants (eg wheat maize and barley) (Cossette and
Miller 1995 Rocha et al 2005 Boddu et al 2007) As animals TC inhibits protein synthesis
in non-host A thaliana (Nishiuchi et al 2006) Immunogold assays showed that the cellular
targets of DON in wheat and maize were cell membrane ribosome endoplasmic reticulum
21
vacuoles and mitochondria Modulation of cell membrane integrity and electrolyte losses
were observed by direct DON interference in maize and wheat cell assays (Cossette and
Miller 1995 Mitterbauer et al 2004) DON-mediated mitochondrion membrane alterations
disruption of host cells enzymatic activity apoptosis and necrosis have been observed in
cereals (Mitterbauer et al 2004 Rocha et al 2005) In contrast to animal type-B is more toxic
to plants than type-A DON showed 8- and 13-fold phytotoxicity to wheat coleoptiles than T-
2 and HT-2 toxins respectively (Eudes et al 2000) By compilation of different studies TC
phytoxicity can be ordered as DONge3-ADONgtT-2 toxingtDAS (Wakulinski 1989 McLean
1996 Eudes et al 2000)
22
17 Cellular and molecular mechanisms of DON toxicity in animals and plants
The underlying cellular and molecular mechanisms of DONTC toxicity are evolving
Exposure to DON proceeds with the up and down regulation of numerous critical genes of
eukaryotic organisms which causes various negative effects on animal and plant organisms
(Figure 1-2)
One of the early events of DON toxicity occurs when the mycotoxin binds to the
active ribosome resulting in blocking protein translation through different routes by
damaging the ribosome (Shifrin and Anderson 1999) degrading or altering conformational
changes of 28S-rRNA (Alisi et al 2008 Li and Pestka 2008) and inducing expression of
miRNAs This results in down regulation of the ribosome associated genes (He et al 2010a
Pestka 2010) Both animal and plant cell assays demonstrated that DON bound to the peptidyl
transferase (Rpl3) domain of the 60S subunit of ribosome and blocked protein chain
elongation and termination at high DON concentrations while peptide chain termination and
ribosome breakage occurred at a low level of DON exposure (Zhou et al 2003a)
23
Figure 1-2 Cellular and molecular mechanisms of deoxynivalenol toxicity in eukaryotic
organisms DON is simply absorbed by the cell and interacts with the cytoplasmic membrane
(CM) mitochondrion (MT) membrane and ribosome Disruption of the CM and alteration of
the MT membrane causes a loss of electrolytes and affects the function of the MT
respectively DON binds to the active ribosomes which leads to ribosome damage and
induces micro RNAs (miRNA) Expression of miRNA impairs the synthesis of ribosome
associated protein The subsequent transduction and phosphorylation of the RNA-activated
protein kinase (PKR) and hematopoietic cell kinase (Hck) activate the mitogen-activated
protein kinase (MAPK) cascade Phosphorylation of the MAPKs induces targeted
transcription factors (TF) which results in ribotoxis stress response upregulation of
proinflammatory cytokines and dysregulation of neuroendocrine signaling (derived from
Pestka 2007 and 2010)
24
The consequence of DON interaction with ribosomes leads to cellular and molecular
changes which are complex and not fully understood However it is well-demonstrated that
the activation of the mitogen-activated protein kinase (MAPK) and subsequent molecular
actions play key roles in the cytotoxicity of TC Upon binding to the ribosome DON rapidly
induced interactions between the protein kinases and the ribosomal 60S subunit as observed in
mononuclear phagocytes (Bae and Pestka 2008) Ribosomal binding to DON activated the
ribosome associated kinases for instances protein kinase R (PKR) hematopoietic cell kinase
(Hck) and P38
mitogen-activated protein kinase The protein kinases are induced by double-
stranded RNA (dsRNA) or possibly by the damaged ribosomal peptides (Iordanov et al 1997
Shifrin and Anderson 1999 Zhou et al 2003b Pestka 2007) Subsequent phosphorylation of
kinases (eg PKRHckP38
) activates the MAPK cascade resulting in ribotoxis stress
response upregulation of proinflammatory cytokines and dysregulation of neuroendocrine
signaling (Pestka and Smolinki 2005 Pestka 2010) Studies suggested that high doses of
DON induces ribotoxis stress response (apoptosis) preceded by immunosuppression and
disruption of intestinal permeabilitytissue functions (Yang et al 2000 Pestka 2008) Further
study observed stabilization of the mRNA and expression of transcription factors for critical
genes when cells were exposed to a low concentration of DON (Chung et al 2003)
The MAPK pathway proceeds with upregulation of a broad range of proinflammatory
cytokine genes and dysregulation of neuroendocrine signaling within the central and enteric
nervous systems Modulation of neuroendocrine signaling by DON increased the level of
serotonin resulted in vomiting reduced food intake and growth retardation in experimental
animalscell lines (Prelusky and Trenholm 1993 Li 2007) Upregulation of proinflammatory
cytokines and chemokines genes was determined in mice liver spleen kidney and lung
25
following in 1 h of exposure to DON A subsequent shock-like effect was observed in mice
because of a ldquocytokine stormrdquo (Pestka 2008) Induction of proinflammatory cytokines also led
to various cellular effects on the central nervous system interference of the innate immune
system and growth hormone signaling These contribute to anorexia growth retardation
aberrant immune response and tissue dysfunction as demonstrated in experimental animal
systems (Pestka 2010)
DON-mediated growth retardation in mice via suppression of growth hormones
(insulin-like growth factor 1 and insulin-like growth factor acid-labile subunit) has been
reported by Amuzie et al (2009) Disruption of gut tissues by DONNIV and subsequent
impairment of intestinal transporter activity was demonstrated in a human epithelial cell
model (Marseca et al 2002) The study proposed that alteration of gut tissue permeability
may cause gastroenteritis induction and growth retardation in livestock animals Growth
retardation of livestock may also occur via dysregulation of neuroendocrine signaling-
mediated induction of emesis and anorexia (Pestka et al 2010)
The molecular mechanism of TC toxicity in plant systems has not been well
documented A recent study showed the upregulation of numerous defense related genes in
wheat upon exposure to DON (Desmond et al 2008) The study further suggested that DON-
induced program cell death in wheat occurred through a hydrogen peroxide signaling
pathway This mechanism had not been observed in mammals
26
18 Strategies to control FHB and trichothecenes
During the past decades many pre- and post-harvest strategies have been applied to
protect cereal crops from Fusarium infestation and to eliminate TC from contaminated
agricultural commodities (FAO 2001 Munkvold 2003 Varga and Toth 2005 Kabak and
Dobson 2009) Some of the above crop protection techniques have been patented (He and
Zhou 2010) In spite of those efforts TC contamination of cereals appears to be unavoidable
using the currently available methods
Pre-harvest control strategies involving cultural practices (eg crop rotation tillage
early planting) and fungicide applications partially reduced Fusarium infection and DON
contamination in cereals (Dill-Macky and Jones 2000 Schaafsma et al 2001) However
repeated application of fungicides may lead to the evolution of Fusarium strains resistant to
fungicides with greater mycotoxin production ability (DrsquoMello et al 1998) At post-harvest
reduction of grain moisture by artificial drying storage at low temperatures (1-4degC) and
proper ventilation of the storage room have shown to be effective in inhibiting fungal growth
and mycotoxin production in infected grain (Midwest Plan Service 1987 Wilcke and Morey
1995) This technology has become less applicable particularly in developing countries
because of the frequent observation required to maintain proper storage conditions (Hell et al
2000)
Removal of Fusarium infected grains by physical methods (eg mechanical
separation density segregation colour sorting etc) was unable to eliminate TC completely
(Abbas et al 1985 He et al 2010b) This is partly because FHB symptomless kernels may
contain a considerable amount of DON which are not eliminated by physical separation
procedures (Seitz and Bechtel 1985) Several studies showed that milling and food processing
27
steps were unable to completely remove DON from grain-based foods (Jackson and
Bullerman 1999 Placinta et al 1999 Kushiro 2008) Detoxification of TC by chemical
treatments (eg ozone ammonia chlorine hydrogen peroxide and sodium metabisulfite) has
been demonstrated (Young et al 1986 Young et al 1987 Young et al 2006) The problem
with these techniques is that the application of such chemicals is not economically viable and
it also has a negative impact on grain quality (Karlovsky 2011) Although the technique has
been popular and showed effectiveness to certain mycotoxins utilization of absorbents to
inhibit mycotoxin absorption by the livestock gastrointestinal tract appeared to be ineffective
for TC (Huwig et al 2001)
The introduction of Fusarium resistant cultivars would be the best strategy to address
the FHB and associated mycotoxin problems in cereals However breeding for cultivars
resistant to FHBGER is difficult and time consuming as natural resistance against Fusarium
is quantitative in nature FHB resistant quantitative trait loci (QTLs) have been identified in
several cereal genotypes (eg Sumai 3 Ning 7840 Bussard) (Bernando et al 2007 Golkari
et al 2009 Haumlberle et al 2009 Yang et al 2010) The 3B 5A and 6B QTLs for type-I
(resistant to initial infection) and type-II (resistant to fungal spread in the kernel) resistances
have been identified on different wheat chromosomes (Buerstmayr et al 2009) Type-I
(Qfhsifa-5A) and type-II (Fhb1Fhb2-3B) QTLs from Sumai 3 are being used in many
breeding programs and pyramiding of the QTLs in spring wheat showed additional effects
Nonetheless the key genes involved in FHB resistance in cereals are yet to be identified This
makes it difficult to integrate the resistance genes into elite cultivars
28
Cultivar development by direct incorporation of TCDON resistance genes may be an
alternative way of protecting grains from these mycotoxins contaminations Many genes from
distantly related species have been integrated into crop cultivars using a transgenic approach
with great economic benefits (Brookes and Barfoot 2006) DON transforming trichothecene
3-O-acetyltransferase (TRI101 from Fusarium sporotrichioides) and UDP-
glucosyltransferase (from Arabidopsis thaliana) genes expressed in cereal cultivars showed
some DONDAS detoxifying ability under in vitro and controlled conditions (Kimura et al
1998 Okubara et al 2002 Poppenberger et al 2003 Ohsato et al 2007) By contrast no
reduction of mycotoxins in either of the above cases was observed under field conditions
(Manoharan et al 2006) Importantly the transformed acetylated and glycosylated
compounds could revert to the original substances or convert to even more toxic products
(Gareis et al 1990 Alexander 2008)
19 Isolation of DONTC de-epoxidating micro-organisms and genes
An enzyme with the capability of de-epoxidizing DONTC appears to be a potential
source for completely removing TC from contaminated cereals (Karlovsky 2011) However
such trichothecene detoxifying genes do not exist in cereals and other plants (Boutigny et al
2008) Because of this several research groups have attempted to obtain microorganisms that
encode DONTC de-epoxidation genes Mixed microbial culture capable of de-epoxydizing
trichothecenes have been isolated from chicken gut (He et al 1992) cow ruminal fluid (King
et al 1984 Swanson et al 1987 Fuchs et al 2002) pig gut (Kollarczik et al 1994) and
digesta of chicken (Young et al 2007) rats (Yoshizawa et al 1983) sheep (Westlake et al
1989) and fish (Guan et al 2009) All of the microbial cultures cited above showed de-
29
epoxydation capabilities only under anaerobic conditions realtively high temperatures and at
lowhigh pH except microbes from fish intestine that showed efficient de-epoxidation at low
temperatures and wide range of temperatures (Guan et al 2009) Such enzymes active under
anaerobic conditions high temperatues and low pH may have potential applications as feed
additives and the eubacterial strain BBSH 797 is an animal feed additive product with the
brand name Mycofixreg by BIOMIN (He et al 2010b) Biotechnological applications of
anaerobically-active enzymes from the above microbial sources seem to be inappropriate
under the aerobic and moderate conditions at which cereals plants are grown
Most of these efforts have failed to isolate single DONTC degrading microbes
effective under aerobic conditions because of inappropriate microbial growth conditions
under laboratory conditions Earlier attempts were made to obtain TC de-epoxidizing aerobic
bacteria but the culture did not retain DONNIV de-epoxidation activity after reculturing
(Ueno 1983 He et al 1992) In addition the co-metabolic nature of the microbial TC de-
epoxidation reaction has limited the use of mycotoxins as the sole nutrient source for selective
isolation of the target microorganisms Several DONTC de-epoxidation microbial strains
including Bacillus strain LS100 and Eubacterial strain BBSH 797 have been isolated from
chicken gut and rumen fluids through multi-step enrichment procedures (Fuchs et al 2002
Yu et al 2010) A DON to 3-keto DON transforming bactial strain was islated from soil by
prolonged incubation with high concentration DON (Shima et al 1997)
Microbial genesenzymes with TC de-epoxidation activity are not known Based on
the microbial TC de-epoxidation activity of a mixed microbial culture it has been proposed
that ceratin reductases or oxidoreductases may catalyze such a biochemical reaction (Islam et
al 2012) With the advent of molecular techniques many novel genes with
30
industrialagricultural value have been cloned by reverse genetics (genomic DNA library
preparation and functional screening of the library clones) forward genetics (transposon
mutagenesis) and comparative genomics methods (Frey et al 1998 Koonin et al 2001
Suenaga et al 2007 Chung et al 2008 Ansorge 2009) The tremendous advancement of next
generation sequencing technology and bioinformatics tools may facilitate the identification of
the microbial genes responsible for DONTC detoxification (Ansorge 2009) Isolation of TC
de-epoxidation genes may play a major role in producing safe foods and feeds derived from
grains
110 The bacterial species Citrobacter freundii
Citrobacter freundii is a bacterial species belonging to the family Enterobacteriace It
is a facultatively anaerobic and Gram-negative bacillus (Wang et al 2000) The bacterium
survives almost everywhere including animalhuman intestines blood water and soils
(Wang et al 2000 Whalen et al 2007) C freundii is known as an opportunistic human
pathogen and causes a variety of respiratory tract urinary tract and blood infections and
responsible for serious neonatal meningistis disease (Badger et al 1999 Whalen et al 2007)
Interestingly this bacterium plays a positive environmental role through nitrogen recycling
(Puchenkova 1996) Strains of C freundii also contribute to the environment by sequenstering
heavy metals and degrading xenobiotic compounds (Kumar et al 1999 Sharma and Fulekar
2009)
31
111 Hypotheses and objectives
The study was based on the following hypotheses
i Certain microorganisms in DON contaminated agricultural soils are able to transform DON
to de-epoxy DON (de-epoxidation) under aerobic conditions and moderate temperatures
ii DON de-epoxidation may be achieved by a single microbial strain
iii Microbial DON detoxifying genes and associated regulators are able to express and
function in a closely related host organism
iv DON detoxifying microbial strains carry genes for de-epoxidation activity that are absent
from related species that have not evolved under DON selection pressure
The objectives of this research were to
i Select microbial cultures from soil having DON de-epoxidation activity under aerobic
conditions and moderate temperatures by analyzing potential DON contaminated soils in
various growth conditions namely aerobicanaerobic conditions different growth media
temperature and pH
ii Select for organisms with DON de-epoxidation activity from the mixed microbial culture
by treating with antibiotics heating subculturing on agar medium and incubating for a
prolonged period in media with a high DON concentration
iii Identify microbial de-epoxidation genes by producing a microbial genomic DNA library
from the DON de-epoxidizing strain followed by screening of the clones for DON de-
epoxidation
32
iv Determine microbial de-epoxidation genes by sequencing of the genome and comparative
analysis of the genome with closely related species and identification of putative DON
detoxification genes
33
CHAPTER 2
20 AEROBIC AND ANAEROBIC DE-EPOXIDATION OF MYCOTOXIN
DEOXYNIVALENOL BY BACTERIA ORIGINATING FROM AGRICULTURAL
SOIL
Published in World Journal of Microbiology and Biotechnology 2012 287-13
21 Abstract
Soil samples collected from different cereal crop fields in southern Ontario Canada
were screened to obtain microorganisms capable of transforming deoxynivalenol (DON) to
de-epoxy DON (dE-DON) Microbial DON to dE-DON transformation (ie de-epoxidation)
was monitored by using liquid chromatography-ultraviolet-mass spectrometry (LC-UV-MS)
The effects of growth substrates temperature pH incubation time and aerobic versus
anaerobic conditions on the ability of the microbes to de-epoxidize DON were evaluated A
soil microbial culture showed 100 DON to dE-DON biotransformation in mineral salts
broth (MSB) after 144 h of incubation Treatments of the culture with selective antibiotics
followed an elevated temperature (50ordmC) for 15 h and subculturing on agar medium
considerably reduced the microbial diversity Partial 16S-rRNA gene sequence analysis of the
bacteria in the enriched culture indicated the presence of at least Serratia Citrobacter
Clostridium Enterococcus Stenotrophomonas and Streptomyces The enriched culture
completely de-epoxidized DON after 60 h of incubation Bacterial de-epoxidation of DON
occurred at pH 60 to 75 and a wide array of temperatures (12 to 40ordmC) The culture showed
34
rapid de-epoxidation activity under aerobic conditions compared to anaerobic conditions This
is the first report on microbial DON to dE-DON transformation under aerobic conditions and
moderate temperatures The culture could be used to detoxify DON contaminated feed and
might be a potential source for gene(s) for DON de-epoxidation
Keywords biotransformation de-epoxidation deoxynivalenol soil bacteria vomitoxin
22 Introduction
Deoxynivalenol (DON commonly known as lsquovomitoxinrsquo) 3α 7α 15-trihydroxy-
1213-epoxytrichothec-9-en-8-one is a trichothecene produced by toxigenic Fusarium species
causing Fusarium head blight (FHB) or Gibberella ear rot (GER) disease on small grain
cereals and maize respectively (Starkey et al 2007 Foroud and Eudes 2009) DON
contamination in cereals and maize is widespread and can lead to tremendous losses to the
growers It has also detrimental effects on human and livestock health (Pestka and Smolinski
2005 Rocha et al 2005) The toxin interferes with the eukaryotic ribosomal peptidyl
transferase (Rpl3) activity resulting in human and animal mycotoxicosis (Ueno 1985 Bae
and Pestka 2008) Common effects of DON toxicity in animals include diarrhea vomiting
feed refusal growth retardation immunosuppression reduced ovarian function and
reproductive disorders (Pestka and Smolinski 2005 Pestka 2007) In plants DON causes a
variety of effects viz inhibition of seed germination seedling growth green plant
regeneration root and coleoptile growth (cited in Rocha et al 2005) In addition DON
enhances disease severity in sensitive host plants (Maier et al 2006)
35
Avoidance of DON contamination or reduction of toxin levels in contaminated grains
remains a challenging task Physical and chemical processes are either inappropriate or
prohibitively expensive to use to decrease the DON content of agricultural commodities
(Kushiro 2008 He et al 2010b) The use of Fusarium resistant crop varieties might be the
best method to reduce DON contamination However breeding for FHB or GER resistant
cultivars appears to be very difficult and time consuming as natural sources of resistance
against Fusarium-caused diseases is quantitative in nature (Ali et al 2005 Buerstmayr et al
2009) As an alternative expressing genes for enzymatic detoxification of DON in elite
cultivars may be an effective strategy for protecting the value of feeds and foods (Karlovsky
1999) Heterologous expression of eukaryotic trichothecene 3-O-acetyltransferase and UDP-
glucosyltransferase genes in cereals and Arabidopsis have resulted in reduced toxicity via
acetylation and glycosylation of the DON (Poppenberger et al 2003 Ohsato et al 2007)
It has been proposed that enzymatic de-epoxidation of trichothecene toxins could be
used to limit DON contamination (Zhou et al 2008 Foroud and Eudes 2009) DON belongs
to type B-trichothecenes which contain an epoxide ring at C12-13 and the epoxide is highly
reactive and mainly responsible for toxicity (Figure 2-1) (Swanson et al 1988 Eriksen et al
2004)
36
Figure 2-1 Mechanism of microbial DON de-epoxidation Removal of the oxygen atom from
the epoxide ring results in formation of a C=C double bond at C-12 position which reduces
cytotoxicity (Swanson et al 1988 Eriksen et al 2004)
Microbial DON de-epoxidation by single bacterial strains and by mixed cultures has
been reported (King et al 1984 Swanson et al 1987 He et al 1992 Kollarczik et al 1994
Fuchs et al 2002 Young et al 2007 Guan et al 2009 Yu et al 2010) However all of these
microbes originated from fish and other animals and were strictly anaerobic The recovered
microbial cultures and single strains only showed de-epoxidation of trichothecenes at low
(le15ordmC) and high (gt30degC) temperatures Such enzymes might have limited applications for
toxin decontamination in cereal grains The aim of the current study was to isolate the
microbes from soil capable of transforming DON to a de-epoxy metabolite under aerobic
conditions and moderate temperatures
De-epoxydation
Deoxynivalenol (DON)
De-epoxyDeoxynivalenol (dE-DON)
37
23 Materials and methods
231 Soil samples
The upper layer (0-30 cm) of soils along with crop debris were randomly collected
from cereal (eg maize wheat barley) crop fields during 2004-2006 in Southern Ontario
Canada Detailed of the crops and locations of the fields have been described in Jianwei Hersquos
Master Degree thesis (He 2007) Briefly soil samples were collected in sterile plastic bags
and transported to the laboratory Samples were sieved to remove large roots stones and
stored at 4ordmC until used The samples were mixed with grounded mouldy corn (containing 95
microgg DON) + F graminearum and incubated at 22-24degC for six weeks Five samples were
prepared for DON de-epoxidation activity analysis by dispersing 5 g soil in 50 mL distilled
water and storing at4degC until further use
232 Culture media
Media used in the study were nutrient broth (NB) Luria Bertani (LB) minimal
medium (MM) (He 2007) six mineral salts + 05 bacto peptone (MSB) (He 2007) tryptic
soy broth (TSB) corn meal broth (CMB) (Guan et al 2009) potato dextrose broth (PDB) six
mineral salts + 05 yeast extract broth (MSY) and soil extract (SE) MM was composed of
sucrose-100 g K2HPO4-25 g KH2PO4-25 g (NH4)2HPO4-10 g MgSO47H2O-02 g
FeSO4-001 g MnSO47H2O- 0007 g water- 985 mL and MSB was composed of the same
salts as MM but was supplemented with Bactotrade Peptone (05 wv) as the sole C source
instead of sucrose To prepare soil extract 100 g of the composite soil was homogenized in 1
L water The soil particles and plant debris were removed by passing the soil suspension
through Whatmanreg grade 3 filter paper (Sigma-Aldrich Oakville Ontario Canada) The
38
extract was autoclaved at 121degC for 30 min Media and ingredients were purchased either
from Fisher Scientific Inc (Fair Lawn New Jersey USA) Sigma-Aldrich (Oakvile Ontario
Canada) or Difco (Sparks Maryland USA) Prior to autoclaving pH of the broth media was
adjusted to 70 Solid media were prepared by adding 15 agar
233 Screening of soil samples
Frozen soil suspensions were thawed and vortexed to homogenize the microbial
population An aliquot (100 microL) of the suspension was immediately transferred to MSB PDB
and MM broth media DON at 50 microgmL was added to the microbial suspension cultures The
cultures were incubated at 22-24degC with continuous shaking at 180 revolutions per minute
(rpm) The extent of DON de-epoxidation was determined at 24 h 48 h 72 h 96 h 120 h and
144 h of incubation
234 Determination of de-epoxidation product
Microbial DON de-epoxidation was measured according to Guan et al (2009)
Samples were prepared by mixing equal volume of microbial culture and methanol and
allowed to stand for 30 min at room temperature Microorganisms in the mixture were
removed by passing them through a 045 microm Millipore filter (Whatmanreg Florham Park New
Jersey USA) The filtrates were used for de-epoxidation analysis by a liquid chromatography-
ultraviolet-mass spectrometry (LC-UV-MS) Identification and quantification of DON and its
transformation product was achieved using a Finnigan LCQ DECA ion trap LC-MS
instrument (ThermoFinnigan San Jose CA USA) The equipment detects 30-2000 mz
(molecular mass ion ratio) compounds and able to determine 001 micromL DON The HPLC
39
system was equipped with a Finnigan Spectra System UV6000LP ultraviolet (UV) detector
and an Agilent Zorbax Eclipse XDB C18 column (46times150 mm 35 μm) The binary mobile
phase consisted of solvent A (methanol) and solvent B (water) The system was run with a
gradient program 25 A at 0 min 25ndash41 A over 5 min isocratically at 41 A for 2 min
and 41ndash25 A over 1 min There was a 3 min post-run at its starting condition to allow for
the equilibration of the column The flow-rate was set at 10 mLmin UV absorbance was
measured at 218 nm Atmospheric pressure chemical ionization (APCI) positive ion mode
was used for MS detection A DON standard was used to adjust the instrument to its
maximum response The system was operated as follows shear gas and auxiliary flow-rates
were set at 95 and 43 (arbitrary units) voltages on the capillary tube lens offset multipole 1
offset multipole 2 offset and entrance lens were set at 3000 minus1700 minus370 minus850 minus3500
and minus4200 V respectively capillary and vaporizer temperatures were set at 200 and 450degC
respectively and the discharge needle current was set at 7 μA Identification of mycotoxin
was achieved by comparing their retention times UV spectra and mass spectrum with DON
and de-epoxy DON (Biopure Union MO USA) as standards Quantification of DON and
product was based on peak areas related to their UV and MS calibration curves
40
235 Biological DON de-epoxidation
To help identify the types of microorganisms causing the DON de-epoxidation MSB
medium was supplemented with either 100 microgmL penicillin G or streptomycin sulfate for
suppressing gram positive or negative bacterial growth and activities respectively Three
replications of antibiotic-treated media were inoculated with 100 microL of microbial culture
capable of de-epoxidizing DON The antibiotic-treated cultures were incubated at 27degC
spiked with 50 microgmL DON Controls were prepared without adding antibiotics in the
microbial culture Inhibition of DON de-epoxidation was analyzed after five day of
incubation
236 Reduction of microbial diversity by antibiotic treatments heating and
subculturing on agar medium
In order to reduce non-DON degrading microbial populations the DON de-
epoxidation capable cultures were sequentially treated with antibiotics having diverse modes
of action including i) dichloramphenicol that kills various gram positive Gram-negative and
anaerobic bacteria ii) trimethoprim that kills Gram-positive and negative bacteria iii)
penicillin that is lethal to Gram-positive bacteria and iv) cyclohexamide to kill filamentous
fungi and yeasts Antibiotics were purchased from Sigma-Aldrich Antibiotic stock solutions
(10 mgmL) were prepared with sterile water or methanol Microbial cultures in MSB
medium were treated with 100 microgmL of each antibiotic followed by incubation of the culture
at 50ordmC for 15 hours to kill undesirded microbial species After heating cultures were treated
with 50 microgmL DON and incubated at 27ordmC for five days DON de-epoxidation activity of the
microbial cultures was determined following the procedure described above Cultures that
41
showed gt99 de-epoxidation were considered for subsequent antibiotic and heat treatments
Aliquots of the achieved culture were spread on MSB agar plates and grown at room
temperature for few days Colonies appeared on the plates were collected and incubated in
MSB medium supplemented with 50 microgmL DON De-epoxidation ability of the subcultured
microbial cultures was determined after five days of incubation at 27degC by LC-UV-MS
method
237 Analysis of microbial population diversity
Terminal Restriction Fragment Length Polymorphism (T-RFLP) analysis
Initially microbial diversity was determined by terminal restriction fragment length
polymorphism (T-RFLP) fingerprint analysis (Bruce and Hughes 2000) Genomic DNA from
microbial cultures with and without enrichment was extracted using DNeasy Plant DNA
Extraction Mini Kit (Qiagen Inc Mississauga Ontario Canada) 16S-rDNA was amplified
by PCR using the HEX-labeled forward primer Bac8f
(5-agagtttgatcctggctcag-3) and
unlabeled reverse primer Univ1492r (5-ggttaccttgttacgactt-3) (Fierer and Jackson 2006)
Fifty microlitter PCR mixture for each sample was prepared having 1x HotStarTaq Master
Mix (Qiagen) 05 microM of each primer 50 microg of BSA and 50 ng of DNA The 35 PCR cycles
followed 60 s at 94degC 30 s at 50degC and 60 s at 72degC After size verification by agarose-gel
electrophoresis 50 ng of purified PCR products was digested for 3 h in a 20 microL reaction
mixture with 5 U of HhaI restriction enzyme (New England Biolabs Ipswich MA USA)
The size and relative abundance of the labeled T-RFLP fragments were separated by capillary
electrophoresis in an automated DNA sequencer equipped with fluorescence detector
(Applied Biosystem 3730 DNA Analyzer) The DNA fragments were analyzed with the Peak
42
Scannertrade Software v10 to create an electropherogram with the genetic fingerprints of the
microbial community
Analysis of V3-V5 region of the 16S-rDNA
To define the bacterial genera present in the enrichment culture the 16S-rDNA was
amplified using two sets of bacterial universal primers Primarily 16S-rDNA was PCR
amplified using the primers 46f-5rsquogcytaacacatgcaagtcga3rsquo (Kaplan and Kitts 2004) and
1371r-5rsquogtgtgtacaagncccgggaa3rsquo (Zhao et al 2007) PCR was performed in a total volume of
25 μL which contained 06 μM of each primer 1x of HotStarTaqreg Master Mix (Qiagen Inc)
and 20-40 ng of DNA The PCR cycles included the following steps 95ordmC (15 min) 35 cycles
of 94ordmC (20 s) 52ordmC (15 s) 72ordmC (1 min 30 s) and a 72ordmC (7 min) extension step The second
pair of universal bacterial primers were 27f-5rsquoagagtttgatcmtggctcag3rsquoand 1492r-
5rsquoggttaccttgttacgactt3rsquo (Lane 1991) PCR was conducted with 25 cycles following the
procedures of Tian et al (2009) Amplification was carried out in a GeneAmpreg PCR System
9700 (Applied Biosystems) Furthermore the PCR products were used as templates to
amplify the hypervariable V3-V5 region of the 16S-rDNA using primers 357f-
5rsquocctacgggaggcagcag3rsquo and 907r7 5rsquoccgtcaattcctttgagttt3rsquo (Yu and Morrison 2004)
After purification of the PCR products by QIAquick PCR purification kit two
amplicon libraries were prepared by ligating to pCR-TOPO vector (Invitrogen) and the library
clones were transformed into TOPO10 competent cells according to Invitrogenrsquos TOPO
cloning manual The clone libraries were plated on LB plates containing 25 μgmL
kanamycin Fifty colonies from each library were randomly picked up from the overnight
plates and inserts were sequenced as described by Geng et al (2006) PCR products were
43
sequenced with an automated ABI 3730 DNA analyzer (Applied Biosystems) The sequence
similarities of the amplified V3-V5 region were compared with the ribosomal database project
(RDP) (httprdpcmemsuedu) Sequences with greater than 95 identity were assigned to
the same bacterial genus
238 Factors influencing DON de-epoxidation
DON de-epoxidation ability of the enriched microbial culture was investigated under
different conditions as described below
Maintaince of active culture
To retain microbial de-epoxidation activity the active culture has been maintained in
MSB medium containing 50 microgmL DON
Media growth temperature and pH
Overnight microbial cultures (100 μL) were added to 2 mL of NB LB MM TSB
MSB CMB MSY PDB or SE broths The pH of the media was adjusted to 70 and the
experiments were conducted at 27degC temperature The effect of temperature was studied by
incubating the culture in MSB medium (pH 7) at 12 20 25 30 and 40ordmC To determine the
effects of pH MSB medium pH was adjusted to 55 60 65 70 75 and 80 Media with
different pHs were inoculated with aliquots of the overnight active culture and then incubated
at 27ordmC The cultures were supplemented with 50 μgmL DON before incubation Three
replications for each medium temperature and pH condition were evaluated
44
De-epoxidation under aerobic and anaerobic conditions
Three replicate microbial cultures of five mL each were grown in sterile 15 mL
centrifuge tubes under aerobic conditions and in an anaerobic chamber filled with 95 CO2
and 5 H2 (COY Laboratory Products Inc Ann Arbor MI USA) Three days after initiation
half (25 mL) of the cultures from anaerobic conditions was transferred to new centrifuge
tubes and brought to aerobic conditions Similarly half of the aerobically grown cultures were
placed into an anaerobic chamber Cultures moved from anaerobic to aerobic conditions were
shaken after opening the lids of the tubes in a laminar flow every eight hours intervals to
maintain aerobic conditions in the culture Lids of the culture tubes transferred to anaerobic
conditions were kept loose to allow oxygen to escape from the culture Two days after
transferring to the new conditions the cultures in both conditions were treated with 50 microgmL
DON and incubated at 27degC Every 12 h 500 microL were removed from each tube and analyzed
for microbial DON de-epoxidation activity by LC-UV-MS
239 Data analysis
Data were processed using Microsoft Excel Program 2007 Statistical analysis was
done using General Linear Model procedures in SAS version 91 (SAS Institute Inc Cary
NC USA 2002-03) The results were the average of three replications stated as mean plusmn
standard error Differences between means were considered significant at plt 005 according
to Tukeyrsquos-HSD test
45
24 Results
241 Analyses of DON to dE-DON biotransformation
A typical LC-UV-MS analysis where there is incomplete de-epoxidation of DON by a
particular soil suspension culture is shown in Figure 2-2 Analysis by LC with UV detection
and integration of peak areas showed 662 conversion of DON to dE-DON (Figure 2-2a)
Analysis LC with MS detection (expressed as total ion counts) indicated 643 conversion
(Figure 2-2b) LC analyses with UV and TIC (total ion count) detection are subject to
interferences due to co-eluting contaminants and therefore integration of small peaks was
very difficult making quantification inaccurate MS detection by SIM (selected ion
monitoring) can eliminate the influence of co-eluting contaminants For example the SIM LC
chromatogram for DON (Figure 2-2c) was generated by using the major ions resulting from
the APCI positive ion MS of DON (Figure 2-2e)
Similarly the SIM LC chromatogram for dE-DON (Figure 2-2d) was derived from the
MS ions of dE-DON (Figure 2-2f) The signal to noise ratios (SN) of peaks observed by SIM
were at least 20 times greater than those detected by TIC because there is reduced likelihood
of co-eluting peaks having the same ions Integration of the SIM peaks revealed a 616
conversion The three methods gave similar results but the SIM LC results were considered
to be the most accurate Using that method the conversion of DON to dE-DON by the mixed
culture occurred most quickly at 48 h and was completed by 144 h (Figure 2-3)
46
47
Figure 2-2 LC-UV-MS chromatographic analysis of DON and its de-epoxidated product dE-
DON obtained by culturing with selected soil microbes (a) LC-UV chromatogram of product
mixture monitored at 218 nm (b) LC-MS total ion chromatogram (TIC) of product mixture
(c) selected ion monitoring (SIM) chromatogram of cultured DON at mz 231 249 267 279
and 297 (d) SIM chromatogram of cultured dE-DON at mz 215 233 245 251 263 and 281
(e) atmospheric pressure chemical ionization (APCI) positive ion mass spectrum of DON (f)
APCI positive ion mass spectrum of dE-DON
48
242 Screening of soil samples
Initially samples originating from soils were analyzed in three types of culture media
to support the growth of fungi yeast and bacteria However cultures growing in PDB which
is a suitable substrate for many fungi and yeasts did not permit transform DON to dE-DON
(data not shown by microorganisms) A define medium (ie MM) was also used in the
sample screening assay None of the soil samples showed de-epoxidation activity in MM
(data not shown) From five soil samples one soil sample culture showed de-epoxidation of
DON in MSB The culture in MSB converted gt99 DON to dE-DON within 144 h (Figure 2-
3) Conversion began after a lag of 48 h and was linear thereafter with a rate of 1 per hour
49
Figure 2-3 Time course of relative biotransformation of DON to dE-DON by the initial soil
suspension culture in MSB medium The culture contained 50 μgmL DON at time 0 Culture
with three replications was incubated at room temperature with continuous shaking at 180
rpm The conversion of DON to dE-DON was analysed by LC-UV-MS at 24 h intervals Blue
and red lines are the percent conversion of DON and recovery of dE-DON respectively
50
243 Evidence of biological DON de-epoxidation
Treatment of active microbial culture with 100 microgmL of streptomycin completely
inhibited DON de-epoxidation In contrast the same concentration of penicillin G did not
affect activity of the culture (Figure 2-4) Streptomycin is effective against Gram-negative
bacteria while penicillin G is lethal to Gram-positive bacteria These experiments provided
evidence that DON de-epoxidation may be caused by certain Gram-negative bacteria
51
Figure 2-4 Microbial de-epoxidation of DON after five days of incubation The active DON
de-epoxidation culture was treated with either 100 microgmL penicillin G or streptomycin
(Strepto) in MSB Cultures having three replications were added 50 microgmL DON and
incubated at room temperatures with continuous shaking at 180 rpm For controls cultures
were without antibiotics DON to dE-DON transformation was monitored after five day of
incubation by LC-UV-MS method
0
20
40
60
80
100
120
Penicillin Strepto Control
DO
N d
e-e
po
xyd
ati
on
(
)
Antibiotics
52
244 Reduction of the microbial diversity
Plating the suspension cultures onto MSB agar revealed colonies with a variety of
sizes morphologies and colours reflecting a mixed culture (data not shown) To reduce the
complexity of the culture three steps of selection pressures were applied namely treatment
with antibiotics heat shocks and subculturing on solid medium The above indicated
treatments did not significantly affect DON de-epoxidation of the microbial cultures (data not
shown) T-RFLP profiles of the mixed culture before treatments showed six distinct peaks of
about 110 150 200 220 340 and 420 bp restriction fragments with the 110 bp peak being
the predominant one (Figure 2-5a)
After treatment with antibiotics and heat only three peaks at 110 150 and 325 bp
remained (Figure 2-5b) suggesting that there was a reduction in the microbial diversity
without affecting DON de-epoxidation After enrichments the bacteria species in the culture
changed their growth patterns as the electropherogram produced different sizes of pick
lengths compared to untreated culture
53
(b)
(a)
Rela
tive flu
ore
scen
ce
32000
0
0 500
16S-rDNA fragment size (bp)
100 200 300 400
Rela
tive flu
ore
scen
ce
32000
0
500
16S-rDNA fragment size (bp)
0 300200100 400
54
Figure 2-5 Analysis of microbial population reduction by terminal restriction fragment
length polymorphism (T-RFLP) (a) Restriction fragments of the 16S-rDNA of the untreated
culture and (b) fragmentation patterns of the enriched culture Picks along the X-axis showed
the fragmentation patterns of the 16S-rDNA which is related to the bacterial species diversity
The length of picks (along the Y-axis) showed abundance of terminal restriction fragments
related to the abundance of particular bacterial speciesgenera
55
245 Microbial population structures in the enriched culture
Microbial diversity in the enriched culture was determined by analyzing the sequences
of the V3-V5 region (~586 bp) amplified from the 16S-rDNAs of the microorganisms it
contained The sequence analysis showed the presence of Serratia Citrobacter Clostridium
Enterococcus Stenotrophomonas and Streptomyces in the enriched culture (Figure 2-6)
Out of 100 V3-V5 sequences 54 sequences were found to be belonging to Serratia
the predominant species of the culture The remaining sequences matched with 12
Citrobacter 8 Clostridium 14 Enterococcus 8 Stenotrophomonas and 4 Streptomyces
species
56
Figure 2-6 Microbial genera identified in the enriched DON de-epoxidizing microbial
culture The bacterial genera were determined by comparing the hypervariable regions (V3-
V5) of the 16S-rDNA using ribosomal database project (httprdpcmemsuedu) X-axis
showed the identified bacterial genera and Y-axis demonstrated the abundance of the V3-V5
sequences belonging to different bacterial genera
54
12 8
14
8 4
0
20
40
60 S
eq
uen
ce
fre
qu
en
cy
Microbial genera
57
246 Effects of media growth temperature pH and oxygen requirements on DON de-
epoxidation
Of the nine media that were tested DON de-epoxidation by the enriched microbial
culture only occurred in NB and MSB (Figure 2-7a) It appeared that C andor N
concentrations in the media were crucial for the enzymatic activity because less or more than
05 Bacto Peptone in MSB reduced the microbial DON transformation ability
DON de-epoxidation activity occurred over a wide range of temperatures from 12 to
40ordmC (Figure 2-7b) The fastest rates of DON de-epoxidation occurred between 25 to 40ordmC
De-epoxidation was observed within a relatively narrow range of pH (60 to 75) with
the most rapid DON biotransformation activity observed at pH 70 (Figure 2-7c) At pH 70
the enriched culture showed DON de-epoxidation activity 125 times faster than at pH 60 or
75 At pH 55 and 80 there was no observable DON de-epoxidation
58
0
20
40
60
80
100
120
12degC
20degC
25degC
30degC
40degC
DO
N d
e-e
po
xyd
ati
on
(
)
Temperature
0
20
40
60
80
100
120
DO
N d
e-e
pxyd
ati
on
(
)
Media
(a)
(b)
59
Figure 2-7 Effects of culture conditions on DON to de-epoxy DON transformation by the
enriched microbial culture For the media experiment (a) all the media were adjusted to pH 7
and the incubation temperature was 27degC For the temperature experiment (b) MSB pH 7 was
used For the pH experiment (c) MSB at 27degC was used Microbial cultures were inoculated
with 50 μgmL DON De-epoxidation was determined after 72 h of incubation Media MSY
(mineral salts + 05 yeast extract) LB (Luria Bertani) TSB (tryptic soy broth) MSB
(mineral salts + 05 bacto peptone) SE (soil extract) CMB (corn meal broth) PDB (potato
dextrose broth) and MM (minimal medium)
0
20
40
60
80
100
120
55
60
65
70
75
80
DO
N d
e-e
po
xyd
ati
on
(
)
pH
(c)
60
The enriched culture showed DON de-epoxidation activity under both aerobic and
anaerobic conditions (Figure 2-8) The fastest transformation (100 de-epoxidation after 60 h
of incubation) occurred in cultures (A) that were grown under aerobic conditions throughout
the culture growth period A medium rate of de-epoxidation (100 de-epoxidation after 72 h)
was observed in cultures that were initially grown in either aerobic or anaerobic conditions
followed by transferring to anaerobic or aerobic conditions (A+AN and AN+A) respectively
The lowest rate occurred (100 de-epoxidation after 96 h) in cultures grown in continual
anaerobic conditions (AN) The means comparison of microbial DON de-epoxidation
efficiencies at 60 h of incubation showed that cultures continually grown in conditions A and
AN were statistically different Biotransformation abilities of the cultures that were switched
from one conditions to another (A+AN and AN+A) were statistically similar among each
other but different from cultures that were only grown in conditions A or AN (plt005)
61
Figure 2-8 Microbial DON to dE (de-epoxy DON)-DON transformation activity under
aerobic and anaerobic conditions A = culture incubated and examined the activity only under
aerobic conditions A+AN = culture initially was grown in aerobic conditions and then
transferred to anaerobic conditions AN = cultures grown throughout the assay under
anaerobic conditions AN+A = culture initially grown under anaerobic conditions and then
transferred to aerobic conditions Differences of lower case letters are significantly different
in de-epoxidation capability (after 60 h) among incubation conditions according to Tukeyrsquos
HSD test (p lt 005) The experiment was conducted with three replications
62
25 Discussion
The current study describes a novel bacterial culture isolated from agricultural soil that
was capable of transforming DON to de-epoxy DON under aerobic and anaerobic conditions
at moderate temperatures Because it has been speculated that DON translocates to soil from
contaminated crops and DON that enters into the soil could be metabolized by microbes
(Voumllkl et al 2004) soil samples for the current study were mainly collected from fields where
cereal crops contaminated with DON were grown
In our initial sample screening steps upwards of 144 h of incubation were required to
achieve gt99 DON de-epoxidation Such a long period may have been due to the presence of
fast growing microorganisms that had suppressive effects on the growth or activity of the
DON transforming bacteria (Davis et al 2005) The sequential use of different antibiotics
with different modes of actions followed by thermal treatments and subculturing on solid
medium resulted in a reduction in the number of bacterial species as determined by T-RFLP
analysis Analysis of V3-V5 region of the 16S-rDNA showed that five bacterial genera were
present in the enriched culture The culture showed higher DON de-epoxidation activity (only
60 h required for gt99 biotransformation) compared to initial soil suspension culture that
required 144 h to complete the de-epoxidation This microbial culture could be used to isolate
the DON de-epoxidizing gene(s) applying metagenomics approaches (Daniel 2005)
The culture showed complete de-epoxidation activity both in MSB and NB media
However we have found that the use of nutritionally rich NB medium enhanced non-target
bacterial (eg Serratia lequifaciences) growth but this did not occur in MSB which is a
comparably poor medium To avoid the possible suppressive and detrimental effects of the
fast growing bacteria on the target bacteria MSB was used for enrichment and
63
characterization of the culture An important characteristic of the DON de-epoxidation
activity observed for the microbial culture in the current study is its activity at moderate
temperature (25-30degC) In contrast previously isolated cultures showed optimal activities at
low (le15degC) or high temperatures (37degC) (Guan et al 2009) Also the culture showed DON
de-epoxidation activity at neutral pH range (pH 60-75) in contrast to previous reports which
found that microbial trichothecene de-epoxidation occurred at acidic and basic pHs (Swanson
et al 1987 He et al 1992 Kollarczik et al 1994 Young et al 2007) However a microbial
culture from fish gut showed de-epoxidation under wide range of pHs (Guan et al 2009)
Probably the pH range reflects the environment in which the bacteria naturally grow which in
this case is similar to the soil pH of a typical crop field in Southern Ontario of Canada The
advantage of the present finding is that DON de-epoxidation activity of the culture was
optimal in a temperature range that matches the temperature range of activity for most arable
plants Therefore it is reasonable to suggest that the gene(s)enzyme(s) responsible for the
activity have potential application for enzymatic detoxification of DON in cereals (Karlovsky
1999)
The current observation that bacterial DON de-epoxidation activity occurred under
both aerobic and anaerobic conditions was interesting because DON de-epoxidation is a
reductive chemical reaction that might be expected to be promoted by anaerobic conditions
However there are previous reports of aerobic de-epoxidation of eldrindieldrin by soil
bacteria (Matsumoto et al 2008) and Goodstadt and Ponting (2004) described aerobic
reductive de-epoxidation by vitamin K epoxide reductasephylloquinone reductase A
definitive determination of the oxygen sensitivity of the enzyme responsible for DON de-
epoxidation will ultimately come from experiments with the purified protein
64
In conclusion this study reports on a novel bacterial source from soil that efficiently
de-epoxidizes DON at moderate temperatures under both aerobic and anaerobic conditions
This research may lead to the isolation of microbes and gene(s) encoding enzyme(s) capable
of de-epoxidizing DON that could create transgenic plants with reduced DON contamination
thus improving food and feed safety and reducing negative effects of the toxin on public
health
26 Acknowledgements
The authors gratefully acknowledged the Natural Sciences and Engineering Research
Council of Canada (NSERC) for awarding a scholarship to Rafiqul Islam for PhD program
The research was supported by the Agriculture and Agri-Food Canada Binational
Agricultural Research and Development Fund (AAFCBARD) project We are thankful for
the contributions of Jianwei He Xiu-Zhen Li and Honghui Zhu
65
CHAPTER 3
30 ISOLATION AND CHARACTERIZATION OF A NOVEL SOIL BACTERIUM
CAPABLE OF DETOXIFYING ELEVEN TRICHOTHECENE MYCOTOXINS
31 Abstract
Deoxynivalenol (DON vomitoxin) is one of the most commonly detected food and
feed contaminating mycotoxins worldwide In order to develop a control method effective
under aerobic conditions and moderate temperatures a bacterial strain ADS47 was isolated
capable of transforming DON to the less toxic de-epoxy DON The bacterial strain is
facultatively anaerobic Gram-negative rod shaped with an average cell size of 150 X 072
microm and has peritrichous (multiple) flagella The 16S-rRNA gene sequence had gt99
nucleotide identity to Citrobacter freundii a bacterium that belongs to the gamma subclass of
proteobacteria Strain ADS47 showed near 100 de-epoxidation of DON after 42 h of
incubation in nutrient broth (NB) under aerobic conditions moderate temperature (28˚C) and
neutral pH (70) However de-epoxidation could occur at 10-45˚C and pH 60-75 Under
anaerobic conditions near 100 de-epoxidation was observed after 72 h mainly during the
stationary growth phase Microbial DON de-epoxidation ability increased with increasing
DON concentrations in the culture media Inhibition of bacterial de-epoxidation by sodium
azide suggested that an active electron transport chain was required In addition to DON
strain ADS47 was able to de-epoxidize andor de-acetylate five type-A and five type-B
trichothecene mycotoxins This is the first report of a bacterial isolate capable of de-
epoxidizing DON and trichothecene mycotoxins under aerobic conditions and moderate
66
temperatures Strain ADS47 could potentially be used in bioremediation to eliminate DON
and other trichothecene mycotoxins from contaminated food and feed
Keywords aerobic de-epoxidation Citrobacter freundii isolation soil bacterium
trichothecenes
32 Introduction
Cereals are often contaminated with a variety of type-A and type-B trichothecene
mycotoxins produced by a number of toxigenic Fusarium species (Placinta et al 1999
Binder et al 2007 Foroud and Eudes 2009) Trichothecenes are tricyclic sesquiterpenes with
a double bond at the C9 10 position and a C12 13 epoxy group (Desjardins 2006) The two
types are distinguished by the presence of a ketone at position C8 in type-B which is absent
in type-A (Yazar and Omurtag 2008) The type-B toxin deoxynivalenol (DON) is the most
frequently found mycotoxin in food and feed around the world (Larsen et al 2004 Desjardins
2006 Starkey et al 2007) Different Fusarium species associated with Fusarium head blight
(FHB) or Gibberella ear rot (GER) disease of small grain cereals and maize are mainly
responsible for contaminating grains with DON and other type-A and type-B trichothecenes
These can cause serious mycotoxicosis in humans animals and plants (Ueno 1985 DrsquoMello
et al 1999 Bae and Pestka 2008) Common effects of DONTC toxicity in animals include
diarrhea vomiting feed refusal growth retardation immunosuppression reduced ovarian
functionsreproductive disorders and even death (Pestka and Smolinski 2005 Pestka 2007)
DON enhances FHB disease severity in wheat and maize (Proctor et al 1995 Desjardins
1996 Maier et al 2006)
67
Reduction of mycotoxin levels in contaminated agricultural products remains a
challenging task Current physical and chemical processes to decrease DON content are either
ineffective or prohibitively expensive (Kushiro 2008 He et al 2010b) However the use of
detoxifying microbes to biodegrade mycotoxins is an alternative (Awad et al 2010
Karlovsky 2011) The toxicity of DON is mainly due to the epoxide and there is a significant
reduction of DON toxicity (54) when it is reduced to produce de-epoxy DON Related
mycotoxins also show reductions in toxicity when they are de-epoxidized such as NIV (55)
and T-2 toxin (400) (Swanson et al 1988 Eriksen et al 2004) Bacillus arbutinivorans
strain LS100 and Eubacterial strain BBSH 797 originating from chicken intestine and rumen
fluid respectively are capable of cleaving the epoxide ring of DON under anaerobic
conditions and high temperature (37degC) (Fuchs et al 2002 Yu et al 2010) Strain BBSH 797
which can de-epoxidize DON scirpentriol and T-2 triol trichothecene mycotoxins has been
used as an additive for poultry and swine diets (Karlovsky 2011) However there are no
reports of microbes able to de-epoxidze DON and other trichothecenes under aerobic
conditions and at moderate temperatures
To develop an effective method for eliminating DON and trichothecene mycotoxins
from contaminated grains soils from fields with FHBGER infected crops were screened for
DON de-epoxidizing microbes (refer to chapter 2 Islam et al 2012) One soil sample yielded
a mixed microbial culture was partially enriched for bacteria with DON de-epoxidizing
activity (chapter 2) The current study describes the purification and characterization of a
novel DON de-epoxidizing bacterium from this culture
68
33 Materials and methods
331 Microbial source
A mixed microbial culture originating from a screen of more than 150 separate soil
samples collected from maize and wheat fields near Guelph Ontario Canada (Islam et al
2012) was used for enrichment purification and characterization of the DON de-epoxidizing
bacterium
332 Media and chemicals
The media used were nutrient broth (NB) 110 strength NB (110 NB) minimal
medium (MM) consisting of six mineral salts (MS) + 100g per liter sucrose (Islam et al
2012) MS + 05 Bacto Peptone (MSB) MS + 03 Bacto Peptone (MSC) MS + 05
yeast extract (MSY) MS + 200 microgmL DON (MS-DON) sterile water-soluble soil extract
(SE) (Islam et al 2012) potato dextrose broth (PDB) and Luria Bertani (LB) Prepared media
and media components were obtained from Fisher Scientific (Fair Lawn New Jersey USA)
Sigma-Aldrich (Oakville ON Canada) andor Difco (Sparks Maryland USA) Media pH
was adjusted as required prior to autoclaving For solid media preparation 15 agar was
added DON and all trichothecene mycotoxins were purchased from Biomin (Romer Lab
Union MO USA)
333 Isolation of the DON de-epoxidizing bacterial strain
Aliquots (20 microL) of the microbial culture partially enriched for DON de-epoxidizing
ability (Islam et al 2012) was transferred to 05 mL MSB (pH 70) in sterile 15 mL
Eppendorf tubes After addition of 200 microgmL pure DON the culture was incubated at room
69
temperature with continuous shaking at 220 revolutions per minute (rpm) Seven to ten days
later 20 microL of the culture was transferred to fresh MSB supplemented with 200 microgmL DON
The culture was then incubated for 7 to 10 days under the above conditions This procedure
was repeated 12 times
After three months of repeated transfers in broth with 200 microgmL DON 100 microL of the
culture was serially diluted in 09 mL MSB containing 05 Tweenreg20 (Sigma Aldrich) to
reduce bacterial cell aggregation (Mabagala 1997) The dilutions were vigorously shaken for
ten minutes A 100 microL aliquot from each dilution was spread on MSB + agar (MSA) The
MSA plates were incubated at room temperature until single colonies appeared
Single colonies from plates were transferred to 1 mL MSB+ 50 microgmL DON and
incubated at room temperature with continuous shaking at 180 rpm under aerobic conditions
After three days the DON de-epoxidation ability was examined by mixing an equal volume
of the bacterial culture and methanol and incubating for 30 min at room temperature to lyse
the cells Insoluble materials in the mixture were removed by passing it through a 045 microm
Millipore filter (Whatmanreg Florham Park New Jersey USA) The level of de-epoxy DON
(dE-DON also known as DOM-1) in the filtrate was determined by liquid chromatography-
ultraviolet-mass spectrometry (LC-UV-MS) (Guan et al 2009)
Individual colonies that showed de-epoxidation activity were serially diluted in
MSB+05 Tweenreg20 and plated onto MSA Single colonies that appeared on the agar plate
were incubated in MSB as described above and retested for DON de-epoxidation ability To
ensure that pure cultures were obtained this was repeated 10 times and individual colonies
were selected on MSA each time
70
334 Identification of a DON de-epoxidizing bacterium
Strain ADS47 with DON de-epoxidation ability was obtained from a single colony
after 10 serial transfers and was grown on MSA at room temperature under aerobic
conditions for 72 h Colony color shape edge and elevation were observed The cell
morphology of the bacterium was examined with a transmission electron microscope at the
University of Guelph Guelph Ontario Canada Whole bacterial cells were placed directly on
a formvar and carbon-coated 200-mesh copper grid They were washed two times by touching
the grids to a drop of clean dH2O and the excess water was blotted off The samples were
negatively stained using 1 Uranyl Acetate (aq) for five seconds Electron microscopy was
performed on a Philips CM10 transmission electron microscope at 60kV under standard
operating conditions (Philips Optics Eindhoven Netherlands) Gram staining was performed
using the procedure described in the commercial gram stain kit from BD Canada
(Mississauga Ontario Canada)
Total bacterial DNA was extracted using a DNeasy Plant DNA Extraction Mini Kit
(Qiagen Inc) The 16S-rDNA was PCR amplified using the primers 46f-
5rsquogcytaacacatgcaagtcga3rsquo (Kaplan and Kitts 2004) and 1371r-5rsquogtgtgtacaagncccgggaa3rsquo
(Zhao et al 2007) PCR was performed in 25 microL containing 06 microM of each primer 1x of
HotStarTaqreg Master Mix (Qiagen Inc) and ~40 ng of DNA The PCR cycles were 95ordmC (15
min) 35 cycles of 94ordmC (20 s) 52ordmC (15 s) 72ordmC (1 min 30 s) and a 72ordmC (7 min) extension
step Amplification was carried out in a GeneAmpreg PCR System 9700 (Applied Biosystems)
PCR products were sequenced with an automated ABI 3730 DNA analyzer (Applied
Biosystems) The 16S-rRNA gene sequence similarity was performed by BlastN using the
Ribosomal Database Project (RDP) (httprdpcmemsueduseqmatch)
71
A phylogenetic tree was constructed by comparing the partial 16S-rDNA sequence of
strain ADS47 with the 11 known Citrobacter species (Bornstein and Schafer 2006 Brenner et
al 1993) The Citrobacter strains were C freundii [FN997616] C koseri [AF025372] C
amalonaticus [AF025370] C farmeri [AF025371] C youngae [AB273741] C braakii
[AF025368] C werkmanii [AF025373] C sedlakii [AF025364] C rodentium [AF025363]
C gillenii [AF025367] and C murliniae [AF025369] Escherichia coli O157H7 [AE005174]
was used as an out group strain The analysis was performed using distance matrix with a gap
opening of 100 and a gap extension of 01 by a server based ClustalW2 program
(httpwwwebiacukToolsmsaclustalw2) A dissimilarity tree was created and viewed
using a TreeView X software (Page 1996)
335 Effects of DON concentration aeration substrate temperature pH and antibiotic
To determine if DON concentration can affect the growth of the isolated bacteria
tubes containing 5 mL of NB or MSB with 20 200 or 500 microgmL of DON were inoculated
with strain ADS47 that had been grown for 16 h with continuous shaking at 220 rpm and then
adjusted to cell density at 025 OD600nm (107 cellsmL) The cultures were incubated at 28degC
with continuous shaking at 180 rpm and 05 mL was removed every 12 h to measure
OD600nm The culture (05 mL) was then analysed for DON de-epoxidation activity by LC-
UV-MS (Guan et al 2009) as described above Three replicate cultures were tested
To examine the effect of aeration five mL of NB or MSB containing 50 microgmL DON
were inoculated with an overnight culture of strain ADS47 prepared as described above and
incubated at 28˚C with continuous shaking at 180 rpm One mL of culture was removed every
six hours and bacterial growth was determined by measuring cell density at OD600nm At the
72
same time points culture samples were analysed for DON to dE-DON transformation ability
(Guan et al 2009) The experiment was repeated with MSB medium containing 50 microgmL
DON under anaerobic conditions as described by Islam et al (2012) with DON to dE-DON
transformation ability being determined every 6 h Three replicate cultures were prepared for
each assay
The effect of substrate on DON de-epoxidation ability was tested after growing
ADS47 for 72 h in NB 110 NB MM MS-DON MSB MSC MSY SE PDB or LB
containing 50 microgmL DON For this assay the media were maintained at pH70plusmn02 and 28degC
for 72 h The effect of temperature was examined with MSB containing 50 microgmL DON at 10
15 20 25 30 35 40 45 or 50degC The effect of pH on growth of strain ADS47 was examined
with MSB adjusted to pH 55 60 65 70 75 or 80 and the cultures were incubated at 28degC
for 72 h Three replicate cultures were used for each assay
Overnight cultures of ADS47 in MSB were treated with 001 and 0001 (wv) of
an antibiotic sodium azide (SA) (Sigma-Aldrich) Controls were prepared in MSB without
SA Three replicate cultures were spiked with 50 microgmL DON DON to dE-DON
transformation ability of the bacterial culture was analysed (Guan et al 2009) after 72 h at
28degC with continuous shaking at 180 rpm Three replicate cultures were tested
336 Localization of DON to dE-DON transformation ability in the bacterium
Bacterial cell extracts were prepared using B-PER II Bacterial Protein Extraction
Reagent following the manufacturerrsquos instructions (PI 78260 Thermo Scientific Rockford
IL USA) ADS47 was grown in 15 mL MSB for 48 h under the growth conditions described
above A culture (05 mL) was centrifuged at 5000 x g for 10 minutes and the pellet was
73
collected B-PER II reagent (2 mL) was added to the pellet and the cells were lysed by
pipetting the suspension up and down several times The cell lysate was treated with 20 microL
Halt Protease Inhibitor Cocktail (Thermo Scientific Rockford IL USA) Culture filtrate was
prepared by passing 05 mL of the culture through a 022 microm Millipore filter (Fisher
Scientific) The remaining 05 mL culture was used as control Cell lysate culture filtrate and
control were incubated with 50 microgmL DON and incubated at 28degC DON to dE-DON
transformation ability of the cell lysate culture filtrate and control were analysed after 72 h
incubation (Guan et al 2009) Three replicate cultures were tested
337 Detoxification of additional trichothecene mycotoxins
Strain ADS47 was evaluated for its ability to degrade five type-A trichothecene
mycotoxins (HT-2 toxin T-2 toxin T2-triol diacetoxyscirpenol and neosolaniol) and five
type-B trichothecene mycotoxins (nivalenol verrucarol fusarenon X 3-acetyl-
deoxynivalenol and 15-acetyl-deoxynivalenol) One and a half mL NB was added with 100
microgmL of each mycotoxin and then inoculated with 100 microL of ADS47 culture grown
overnight as previously described The culture of NB + mycotoxin + ADS47 was incubated at
28degC with continuous shaking at 180 rpm and DON to dE-DON transformation ability of the
culture was analysed by LC-UV-MS (Guan et al 2009) Five replicate cultures were tested for
each trichothecene mycotoxin
74
338 Statistical analysis
Data were processed using Microsoft Excel Program 2007 Statistical analysis was
performed using General Linear Model procedure in SAS version 91 (SAS Institute Cary
NC USA 2002-03) Replications were averaged and stated as mean plusmn standard error The
mean differences were considered significant at plt 005 according to Tukeyrsquos-HSD test
75
34 Results
341 Isolation and identification of a DON de-epoxidizing bacterium
A DON de-epoxidizing bacterium designated strain ADS47 was isolated after 12
transfers over three months from cultures growing in MSB containing 200 microgmL DON
Strain ADS47 produced small to medium size circular convex and white colonies on MSA at
room temperature under aerobic conditions (Figure 3-1a) The bacterium is rod shaped and
150 x 072 microm in size (Figure 3-1b) It has numerous flagella that cover the surface of the cell
(peritrichous flagella) and is Gram-negative
BlastN analysis of the 16S-rDNA sequence of strain ADS47 showed the most
similarity to the eleven known Citrobacter species in the gamma proteobacteria of the
Enterobacteriaceae In particular the 16S-rDNA nucleotide sequence of ADS47 was more
than 99 identical to C fruendii (Figure 3-2) The next most similar sequences were a cluster
of C werkmanii and C murliniae followed by more distantly related C braakii C gilleni
and C youngae A separate cluster was formed by C rodentium C farmeri C sedlakii C
koseri and C amalonaticus
76
Figure 3-1 (a) Colony color shape and size of the mycotoxin degrading bacterial strain
ADS47 on mineral salts agar (MSA) medium (b) Transmission electron microscopic (TEM)
image of the Citrobacter freundii strain ADS47 The bacterium showed multiple flagella
(peritrichous flagella) and numerous fimbriae around the cell surface
(a) (b)
77
Figure 3-2 Dendrogram based on 16S-rDNA sequences for strain ADS47 and 11 reference
Citrobacter species An E coli O157H7 strain was considered as an out group species The
tree was constructed using a genetic distance matrix by ClustalW 2 and viewed by
TreeViewX software The distance was converted to scale and shown by branch lengths (scale
bar 001 which indicated the 16S-rDNA nucleotide sequence variability between the bacterial
species) GenBank accession numbers for 16S-rDNA sequences of the species used for
comparison are shown in materials and methods section (334)
78
342 Effects of DON concentrations on bacterial DON de-epoxidation
Growth of ADS47 aerobically in MSB with 20 200 and 500 microgmL DON was not
significantly different from that observed in control cultures without DON (Figure 3-3a)
Similarly increasing DON concentrations did not affect bacterial growth in NB medium
(Figure 3-3b)
DON de-epoxidation activity determined as microgmL DON convertedcell density
(OD600) was measured lowest in MSB+20 microgmL DON and highest in MSB+ 500 microgmL
DON containing cultures (Figure 3-3c) The highest rate of dE-DON formation was 115 microgh
in ADS47 cultures in MSB with 500 microgmL DON between 48 to 72 h All of the DON was
converted in the cultures containing 20 microgmL DON and 200 microgmL DON between 48-60 h
and 60-72 h respectively In 200 microgmL and 500 microgmL of DON cultures the rate of de-
epoxidation increased significantly after 36 h
79
00
04
08
12
16
0 24 36 48 60 72 96
OD
600n
m
Incubation time (h)
Control
20 microgmL
200 microgmL
500 microgmL
(b)
(a)
80
Figure 3-3 Effects of DON concentration on bacterial growth and de-epoxidation activities
Strain ADS47 was grown in MSB and NB media with three different DON concentrations 20
microgmL 200 microgmL and 500 microgmL Control cultures were prepared without DON Cultures
were grown at 28degC with continuous shaking at 180 rpm (a) Microbial density (OD600 nm) in
MSB with different DON concentrations was measured at 24 36 48 60 72 and 96 h (b)
Microbial growth (OD600 nm) in NB having different DON concentration was measured at 24
48 72 and 96 h (c) Bacterial DON de-epoxidation activity in 20 200 and 500 microgmL DON
containing cultures at different time points DON de-epoxidation was measured in microgmL per
OD600 nm Three replicate cultures for each assay were analysed
(C)
81
343 Influence of growth conditions and antibiotic on microbial DON de-epoxydation
Aerobic and anaerobic conditions
ADS47 grew more quickly uner aerobic conditions under MSB+50 microgmL DON than
in MSB+50 microgmL DON under anaerobic conditions (Figure 3-4a) Aerobic growth was
faster in NB+50 microgmL DON compared to MSB+50 microgmL DON
However the DON de-epoxidation activities on a cell density basis were very similar
in NB+50 microgmL DON compared to MSB+50 microgmL DON (Figure 3-4b) Also DON de-
epoxidation activity of ADS47 under aerobic conditions in MSB+50 microgmL DON was higher
than in the same medium under anaerobic conditions (Figure 3-4b)
82
(b)
(a)
83
Figure 3-4 Bacterial growth and DON de-epoxidation capabilities under aerobic and
anaerobic conditions (a) Microbial growth under aerobic conditions in MSB and NB medium
and anaerobic conditions in MSB medium (b) Microbial DON de-epoxidation ability under
aerobic conditions in MSB and NB media and anaerobic conditions in MSB medium
Cultures media having 50 microgmL DON were inoculated with overnight culture of strain
ADS47 Initial microbial concentration in the cultures was adjusted to 025 OD600nm Cultures
were incubated at 28degC having three replications Bacterial growth at OD600nm and DON de-
epoxidation in microgmL per OD600nm were measured at six hour interval
84
Media temperature and pH
Growth of ADS47 in media containing 50 microgmL DON under aerobic conditions at 72
h was three-fold higher in NB and LB than in SE two-fold higher than in 110 NB and MSC
and did not occur in MS-DON (Figure 3-5a) Growth in MM MSB and MSY was
intermediate
Over the 72 h DON de-epoxidation per cell density was highest in MSB and MSY and
approximately 25 lower in NB and LB and lower by 75 and 87 in 110 NB and MSC
respectively (Figure 3-5b) No DON de-epoxidation was observed in MS-DON in which
DON was used as a carbon source Bacteria grown in MM broth which contains sucrose as a
sole carbon source were unable to de-epoxidize DON No DON de-epoxidation was observed
in SE and PDB grown cultures
Growth of ADS47 under aerobic conditions for 72 h in MSB+50 microgmL DON was
greatest between 25-40degC but no growth occurred at 50degC (Figure 3-6a) DON de-
epoxidation activity was greatest and equivalent between 20-40degC and lower at 10 15 and
45degC temperatures (Figure 3-6b)
ADS47 growth under aerobic conditions for 72 h in MSB+50 microgmL DON was
limited to neutral pHs (ie 60 - 75) (Figure 3-7a) The transformation of DON to dE-DON
occurred at the same pH range and DON de-epoxidation activity was not significantly
different between pH 60 to 75 (Figure 3-7b)
85
b
c
a
b
a
d c
b b
(a)
(b)
b
c
b
a a
d
86
Figure 3-5 Effects of media on bacterial growth and DON de-epoxidation activity (a)
Microbial growth (at OD600nm) on different culture media Media pHs were adjusted to
70plusmn02 (b) Bacterial DON to dE-DON transformation in microgmL by OD600nm in different
media after 72 h The cultures having three replications were incubated for 72 h at 28degC and
DON de-epoxidation was measured The Media NB = nutrient broth 1NB = 110 strength
NB MM = minimal medium MS-DON = six mineral salts + 200 microgmL DON as sole C
source MSB = six mineral salts + 05 bacto peptone MSC = six mineral salts broth + 03
bacto peptone MSY = six mineral salts with 05 yeast extract SE = soil extract PDB =
potato dextrose broth and LB = Luria Bertani Means with the same label are not significantly
different according to Tukeyrsquos HSD test (p lt 005)
87
(a)
(b)
88
Figure 3-6 Effects of temperature on microbial growth and DON de-epoxydation capability
(a) Microbial growth (at OD600nm) at different temperatures after 72 h of incubation (b)
Bacteria DON de-epoxidation in microgmL by OD600nm in different temperatures The most
efficient temperature range (ge40 microgmL DON de-epoxidation) is marked by dot lines The
experiment was conducted in MSB medium at pH 70plusmn02 with three replications
89
(a)
(b)
90
Figure 3-7 Influence of pH on microbial growth and DON de-epoxydation (a) Microbial
growth (OD600nm) in MSB at different pH (b) Bacterial DON to de-epoxy-DON
transformation in microgmL by OD600nm in different pH Dot lines indicated the efficient de-
epoxidation (ge40 microgmL DON conversion) pH range Experiment was conducted at 28degC
with three replications
91
Antibiotic
Treatment of ADS47 at 72 h in MSB+50 microgmL DON with sodium azide (SA) at
0001 and 001 reduced DON de-epoxidation activity to 5 (25 microgmL) and 0
respectively The control with no SA added was able to de-epoxidize DON completely
(100) (Figure 3-8)
Figure 3-8 Effects of sodium azide (SA) on bacterial DON de-epoxidation activity
Microbial culture in MSB+50 microgmL DON was treated with 001 (SA-1) and 0001 (SA-
2) sodium azide In control (C) no SA was added The experiments with three replications
were incubated for 72 h at 28degC and DON de-epoxidation was measured by LC-UV-MS
method
0
20
40
60
80
100
120
SA1 SA2 C
D
ON
de
-ep
oxyd
ize
d
92
344 Localization of bacterial DON de-epoxidation
A cell lysate prepared from ADS47 grown for 48 h in MSB transformed 20 (10
microgmL) of DON added at 50 microgmL to dE-DON within 72 h (Figure 3-9) In contrast a cell-
free culture filtrate did not show DON de-epoxidation activity
Figure 3-9 Determination of DON to de-epoxy DON transformation by bacterial culture
filtrate and cell extract The treatments having three replications were spiked with 50 microgmL
DON and incubated at 28degC for 72 h In control strain ADS47 was incubated in MSB broth
with 50 microgmL DON DON de-epoxidation activity in percentage was examined
0
20
40
60
80
100
120
Filtrate Extract Control
D
ON
de
-ep
oxyd
ize
d
93
345 Bacterial detoxification (de-epoxidtaion and de-acetylation) of other trichothecene
mycotoxins
Aerobic cultures of ADS47 grown for 72 h in NB+100 microgmL HT-2 converted 74 of
the toxin to de-epoxy HT-2 toxin and 26 to a de-acetylated product (Figure 3-10 and Table
3-1)
In a culture treated with T2-toxin 46 was converted to de-epoxy T-2 16 was
converted to an unknown compound and 38 was not converted In ADS47 culture with
diacetoxyscirpenol (DAS) 61 was transformed into de-epoxy DAS and 39 was de-
acetylated DAS (39) Neosolaniol (NEO) was transformed to de-epoxy NEO (55) and
de-acetylated NEO (45) T2-triol was transformed in to de-epoxy T2-triol (47) and de-
acetylated T2-triol (53)
Aerobic cultures of ADS47 were also grown for 72 h in NB+100 microgmL of one of five
type-B trichothecenes For nivalenol (NIV) 92 was de-epoxydized while the remaining 8
NIV was transformed to DON 3-acetyl-deoxynivalenol (3-ADON) was transformed into
three products de-epoxy NIV (5) de-epoxy DON (54) and DON (41) For 15-
acetyldeoxynivalenol (15-ADON) 8 was transformed to de-epoxy 15-ADON and 92 to
de-epoxy DON (dE-DON) FusarenonX (FUS) had 95 transformed into de-epoxy FUS
while 97 of verrucarol (VER) was transformed to de-epoxy VER The remaining 5 FUS
and 3 VER were not transformed
94
Figure 3-10 Determination of microbial de-epoxidation and de-acetylation of HT-2 toxin by
LC-UV-MS Bacterial culture in NB was added with 100 microgmL HT-2 toxin Bacterial HT-2
toxin transformation ability was examined after 72 h of incubation at 28degC The upper panel
illustrated mass spectrometry chromatograph of the de-acetylated (red peak) and de-
epoxydized (blue peak) products Arrows indicated the targeted acetyl (red) and epoxy (blue)
functional groups of HT-toxin The middle and lower panels showed the mass spectra of the
de-acetylated (dA) and de-epoxidized (dE) compounds Arrows on the upper panel indicated
the targeted acetyl (red) and epoxy (blue) functional groups of HT-2 toxin Red and blue
arrows on the middle and lower panels pointed at the degradation sites of HT-2 toxin
95
Table 3-1 Microbial transformation of type-A and type-B tricthothecene mycotoxins after 72
h of incubation under aerobic conditions at room temperature DAS=diacetoxyscirpenol
DON=deoxynivalenol FUS=FusarenonX 3AON=3-acetyl-deoxynivalenol and
15ADON=15-acetyl-deoxynivalenol
1 Convertion of trichothecene mycotoxins to either DON or an unknown compound
Trichothecene
mycotoxin
Untransformed
trichothecenes ()
Transformed products of
trichothecenes ()
De-
epoxy
De-
acetylate
Other1
Type-A
HT-2 toxin 0 74 26 -
T-2 toxin 38 46 - 16
(unknown)
DAS 0 61 39 -
Neosolanol 0 55 45 -
T2-triol 0 47 53 -
Type-B
DON 0 100 - -
Nivalenol 0 92 - 8 (DON)
3-ADON 0 59 41 (DON)
15-ADON 0 100 - -
FUS 5 95 - -
Varrucarol 3 97 - -
96
35 Discussion
The ADS47 strain of C freundii was isolated in the present study by a prolonged
incubation of a mixed microbial culture with a high concentration DON since the bacterium
efficiently de-epoxidized (detoxified) DON and number of other trichothecene mycotoxins
under aerobic and anaerobic conditions at moderate temperatures and neutral pH The total
procedures for enrichment and isolation of the strain ADS47 is depicted in supplemental
Figure S3-1 The reductive de-epoxidation activity may be due to a cytoplasmic enzyme
which would be in agreement with a previous study that showed de-epoxidation of
epoxyalkanes was catalyzed by a bacterial cytoplasmic flavoprotein (Westphal et al 1998)
The ability of the currently isolated strain of C freundii to biodegrade environmental
pollutants such as aminoaromatic acids and heavy metals (copper) is also likely mediated by
cytoplasmic enzymes since these compounds need to be taken up by the cells before they are
metabolized (Savelieva et al 2004 Sharma and Fulekar 2009) In contrast oxidative
transformation of DON to 3-keto DON by microbial extracellular enzyme has been reported
(Shima et al 1997)
Isolation of a trichothecene de-epoxidizing microorganism from a mixed microbial
community is very challenging Although trichothecenes are toxic to eukaryotes prokaryotes
appear to be resistant to these mycotoxins (He et al 1992 Rocha et al 2005) The present
study showed that the bacteria grew well in media with high concentrations of DON (500
microgmL) Bacterial de-epoxidation of DON involves the removal of an oxygen atom from the
functional epoxy ring of the tricyclic DON structure (Yoshizawa et al 1983) Such co-
metabolic degradation does not provide the bacteria with any nutrients from the reacting
compounds (Horvath 1972 Fedorak and Grabic-Galic 1991) This is supported by the results
97
of the current study which showed that strain ADS47 was unable to grow in media with DON
as a sole carbon source and increasing the concentration of DON did not increase ADS47
growth in MSB Hence DON could not be used as sole nutrient source in the mixed culture
enrichment process
Many previous attempts to obtain DON de-epoxidizing microbes have failed
particularly under aerobic conditions because media or laboratory conditions were
inappropriate to support growth or initially active cultures lost de-epoxidizing function after
serial transfer (Karlovsky 2011)
Media composition appears to be a crucial factor for microbial trichothecene de-
epoxidation activity (Voumllkl et al 2004) Effective biotransformation of trichothecene
mycotoxins by the fish digesta microbial culture C133 occurred in full media (FM) but a low
level of de-epoxidation occurred in media having a single nitrogen source and no
transformation activity was observed in the common bacterial medium LB (Guan et al
2009) Zhou et al (2010) also showed that chitin de-acetylation by soil bacterial strains
occurred more rapidly in yeast extract compared to common bacterial media Therefore a
screening strategy for microbial DON de-epoxidation ability should use different media in
parallel as was done in this study
This study observed efficient de-epoxidation in common bacterial media (NB and LB)
as well as in mineral salts medium containing yeast extract (MSY) or 05 Bacto Peptone
(MSB) However reduction of Bacto-Peptone to 03 (MSC) significantly reduced the
bacterial de-epoxidation activity suggesting that the activity is sensitive to the availability of
protein and amino acids The study found that medium containing sucrose as carbon source
(MM) and a commonly used medium PDB blocked microbial DON de-epoxidation even
98
though bacteria grew in those growth media These suggest that media containing high sugar
contents had inhibitory effects on microbial de-epoxidation enzymatic activity Previously
sugars have been found to have negative effects on the production andor activity of bacterial
enzymes (Saier and Roseman 1971)
Losses of microbial functions under laboratory conditions limited researchers to obtain
economically important microbial genetic sources The soil borne Curtobacterium sp strain
114-2 efficiently de-epoxydized nivalenol but unfortunately the strain lost its activity in
latter generations (Ueno et al 1983) Instability of de-epoxidation activity of ADS47
observed in cultures grown on agar media without DON resulted in the loss of DON de-
epoxidation activity by more than 80 of the colonies after one generation However strain
ADS47 retained de-epoxidation activity after ten subcultures in MSB (data not shown)
The non-specific de-epoxidation of DON made it impossible to use DON as a sole
substrate C source for selective isolation of ADS47 Thus a number of techniques were
utilized to select ADS47 and suppress other bacteria including the use of a variety of
antibiotics and culture at elevated temperatures (Islam et al 2012) Selection of the isolate
ADS47 required repeated subculturing and prolonged exposure to media containing 200
microgmL DON After three months of selection with DON culture that was obtained had a
DON de-epoxidation activity that was up to 30-fold higher than the mixed culture As growth
of ADS47 was not affected by 200 microgmL DON the prolonged incubation may have reduced
populations of other bacteria that are more sensitive to DON Enrichment and isolation of a
DON to 3-keto DON transforming bacterial strain E3-39 by prolonged incubation of a soil
microbial culture with 200 microgmL DON was reported by Shima et al (1997)
99
This study demonstrated that microbial DON de-epoxidation occurred mainly at the
stationary growth phase A DON degrading strain of Citrobacter may have gained energy
from DON by co-metabolic cleavage of the epoxy ring under less nutrient conditions
Microbial use of energy from co-metabolic reaction has been suggested by Chiu et al (2005)
The rate of de-epoxidation activity increased when ADS47 grew at higher concentrations of
DON Taken together a prolonged exposure to high concentration DON may have resulted in
non-DON metabolizing microbial populations declining by DON toxicity while facilitated
the population of DON metabolizing bacterium by providing energy
DON is a small molecule and is simply absorbed by cells (Sergent et al 2006)
Microbial DON de-epoxidation occurred in intact and lysed bacterial cells In addition
treating the ADS47 culture with SA which blocks gram negative bacterial electron transport
and activity inhibited DON de-epoxidation activity (Lichstein and Malcolm 1943 Erecinska
and Wilson 1981) implying that an active electron transport system required for microbial de-
epoxidation reaction (He et al 1992) The data suggest that a cytoplasmic epoxy reductase is
involved in the DON de-epoxidation reaction in the ADS47 microbial strain that was isolated
in the present study
Since agricultural commodities are often contaminated with multiple fungal
mycotoxins the ability of bacteria like ADS47 to degrade mycotoxins by several
mechanisms may be an advantage De-acetylation might be another mechanism of microbial
mycotoxin detoxification since toxicity of trichothecenes also relies on the acetyl functional
groups (Desjardins 2006) Strain ADS47 showed efficient de-epoxidation and de-acetylation
of several type-A and type-B trichothecene mycotoxins that are commonly detected in cereals
in association with DON In conclusion the soil-borne gram-negative bacterium C freundii
100
strain ADS47 is capable of detoxifying trichothecene mycotoxins by both de-epoxidation and
de-acetylation under aerobic and anaerobic conditions neutral pH and a range of
temperatures ADS47 has great potential to detoxify DON contaminated animal feed
Furthermore isolation of novel bacterial de-epoxidation genes might be used to generate
transgenic plants that can detoxify DON in contaminated seed Alternatively contaminated
seed might be treated with manufacture de-epoxidation enzyme(s) These applications can
improve the quality and safety of agricultural commodities that are prone to trichothecene
mycotoxin contamination
36 Acknowledgements
Thank to the Natural Sciences and Engineering Research Council of Canada (NSERC) for a
scholarship to Rafiqul Islam The work was funded by the Agriculture and Agri-Food Canada
Binational Agricultural Research and Development Fund (AAFCBARD) project We
appreciate Honghui Zhu for technical assistance
101
Supplemental Figure S3-1 Flow chart depicts the procedures of enrichment and isolation of
DON de-epoxidation bacterial strain ADS47 Steps 1 to 4 describe the optimization of
microbial DON to dE-DON transformation activity and removal of undesired microbial
species Step 5 to 7 shows the culture enrichment by incubation with high concentration DON
and isolation of the single strain ADS47
Step 1 Screening of soil samples
Step 2 Optimization of cultural conditions
Step 3 Elimination of undesired microbial species
Step 4 Removal of untargeted microbial species
Step 5 High concentration DON enrichment
Step 6 Separation of bacterial cells and dilution plating
Step 7 Single colony analysis
Active culture was sequentially treated with different mode of actions antibiotics heating cultures at 50degC and subculturing on agar media
Colonies were tested for their DON de-epoxidation ability Purified the single colony via several times subculturing on agar medium
Optimized the culture conditions ie growth substrates temperatures pH and oxygen requirements
Obtained DON de-epoxidation capable microbial culture obtained by analyzing soil samples collected from cereal fields
Serially diluted active culture was grown on agar media Collected washed colonies having DON de-epoxidation activity
The partially enriched culture that retained DON de-epoxidation activity was incubated with 200 microgmL DON in MSB media with continuous shaking at 220 microgmL for 7 d The procedure was repeated 12 times
The enriched culture was serially diluted and treated dilutes with 05 Tween 20 After vigorously vortex for ten min aliquot cultures were spread on different agar media plates to produce single colonies
102
CHAPTER 4
40 CONSTRUCTION AND FUNCTIONAL SCREENING OF GENOMIC DNA
LIBRARY FOR ISOLATION OF DON DE-EPOXYDIZING GENE(S) FROM
CITROBACTER FREUNDII STRAIN ADS47
41 Abstract
Enzymatic activity-based screening of a genomic DNA library is a robust strategy for
isolating novel microbial genes A genomic DNA library of a DON detoxifying bacterium
(Citrobacter freundii strain ADS47) was constructed using a copy control Fosmid vector
(pCC1FOSTM
) and E coli EPI300-T1R as host strain A total of 1250 E coli clones containing
large DNA inserts (~40 kb) were generated which resulted in about 10-fold theoretical
coverage of the ADS47 genome The efficiency of E coli transformation with the
recombinant Fosmid vector was examined by restriction digestion of Fosmids extracted from
15 randomly selected library clones The fragmentation patterns showed that all of the clones
contained an 81 kb Fosmid backbone and ~40 kb of insert DNA except one that had a small
insert This suggests that a DNA library was prepared with expected DNA inserts The de-
epoxidation ability of the library was analysed by incubating clones in Luria Bertani (LB)
broth medium supplemented with 20 microgmL DON with induction (by addition of L-arabinose)
or without induction None of the 1000 clones showed DON de-epoxidation activity The
potential DON de-epoxidation gene(s) of ADS47 may not have been expressed in E coli or
might have been expressed but lacked additional factors required for DON de-epoxidation
103
Keywords DNA library fosmid vector functional library screening DON de-epoxidation
activity
42 Introduction
Microorganisms contain numerous valuable genes and biocatalysts which may have
significant roles in agricultural productivity (Jube and Borthakur 2007) Various techniques
have been used to isolate novel genes from different organisms including transposon
mutagenesis (Frey et al 1998 Tsuge et al 2001) gene knockout (Winzeler et al 1999)
whole genome sequencing (Krause et al 2006) comparative genomics (Koonin et al 2001)
and suppression subtractive hybridization (Dai et al 2010)
The genomic DNA library preparation and activity-based screening of the library is a
relatively new technique which has been efficiently used for isolation of microbial novel
enzymes and compounds such as lipases proteases oxidoreductases antibiotic resistance
genes membrane proteins and antibiotics (Henne et al 2000 Gupta et al 2002 Knietsch et
al 2003 Gabor 2004 Suenaga et al 2007 van Elsas et al 2008) The strength of the
technique is that genes from microbial sources could be obtained without prior sequence
information and not requiring culturing the organisms under laboratory conditions Using this
technique an unknown gene or a set of genes could be obtained by determining particular
phenotypes of the library clones Eventually the gene of interest from the positive clones is
isolated by using a forward (eg mutation) or reverse genetics method (eg subcloning the
positive clones into a microbial expression vector) followed by functional analysis of the
subclones
104
Although this strategy has been used effectively for isolating novel microbial genes
several factors need to be considered in order for the method to be successful These factors
include DNA quality vector type (eg plasmid cosmid fosmid bacterial artificial
chromosome etc) vector copy number and host strains (Daniel 2005 Singh et al 2009) The
Fosmid vector-based technique has been used successfully to clone bacterial genes including
xenobiotics degradation genes antifungal gene clusters and antibiotic resistance genes from
soil metagenomes (Hall 2004 Suenaga et al 2007 Chung et al 2008) This study reports the
construction and activity-based screening of a Fosmid DNA library in an E coli host with the
aim of isolating a novel DON de-epoxidation gene from Citrobacter freundii strain ADS47
(Chapter 3)
43 Materials and methods
431 Genomic DNA library construction
The genomic DNA library was prepared using a Copy Control Fosmid Library
Production Kit (Epicentre Biotechnologies Madison Wisconsin USA) following the
manufacturerrsquos instructions as depicted in Figure 4-1
105
Figure 4-1 Schematic overview of the construction and functional screening of a Fosmid
DNA library
106
Citrobacter freundii strain ADS47 was grown in NB medium at 28degC with continuous
shaking at 220 rpm The high molecular weight bacterial genomic DNA was extracted from
the overnight culture using the Gentra Puregene YeastBacteria Kit (Qiagen Inc Mississauga
Ontario Canada) The purity of the DNA was confirmed by determining OD260280 at ~190 by
a NanoDrop spectrophotometer A 5 microL sample having ~25 microg high molecular weight DNA
was sheared by passing through a 200-microL pipette tip and achieved a sufficient amount of
DNA about 40 kb in size Blunt ended 5rsquo-phosphorylated insert DNA was generated by
thoroughly mixing it with End-Repair Enzyme reagents on ice The 20 microL mixture was
incubated for 45 min at room temperature followed by incubation at 70degC for 10 min to
inactivate the End-Repair Enzyme activity
The treated DNA was size fractionated by a Low Melting Point-Pulsed Field Gel
Electrophoresis (LMP-PFGE) on the clamped homogeneous electric fields (CHEF-DR III)
system (Bio-RAD Laboratories) using 1 UltraPure Low Melting Point Agarose (Invitrogen)
The PFGE was conditioned according to manufacturerrsquos instructions as follows initial switch
time-42 s final switch time-761 s run time 16 h 50 min angle-120deg gradient- 6 Vcm
temperature-14degC and running buffer-05 TBE DNA size markers for the 42 kb Fosmid
control DNA (Epicentre Biotechnologies) and low range Pulse Field Gel (PFG) markers (New
England Biolabs Inc) were loaded adjacent to the DNA sample slots After electrophoresis a
portion of the gel that contained DNA size markers was cut off The gel was stained in SYBR
Safe DNA gel stain (Invitrogen) for 30 min visualized and marked on the 23 42 and 485 kb
marker bands A slice of PFGE gel containing 40 plusmn 10 kb size DNA was cut off with a sterile
razor blade by comparing the marker sizes DNA from the gel was recovered by digesting it
with GELase enzyme ( 1 U100 microL melted agarose) in 1X GELase Buffer followed by a
107
sodium acetate-ethanol precipitation The DNA with the desired size was stored at 4degC for
next dayrsquos ligation to a cloning-ready Fosmid vector (pCC1FOSTM
)
Ligation was performed in a 10 microL reaction mixture by thoroughly mixing 10X Fast-
Link Ligation Buffer 10mM ATP 025 microg of ~40 kb insert DNA 05 microg Fosmid Copy
Control Vector (pCC1FOS) (Figure 4-2) and 2 U Fast-Link DNA lipase The mixture was
incubated for 4 h at room temperature preceded by inactivating the ligation reaction by
heating at 70degC for 15 min The recombinant Fosmid vector was stored at -20degC for later use
Ten microL of the ligated DNA was thawed on ice and transferred to a pre-thawed
MaxPlax Lambda Packaging Extract tube containing 25 microL extract By pipetting several
times the mixture was incubated at 30degC for lamda packaging After 2 h of packaging an
additional 25 microL packaging extract was added into the mixture and incubated for a further 2 h
at 30degC The packaged phage particles were volumed to 05 mL by adding ~400 microL Phage
Dilution Buffer and 25 microL chloramphenicol
Transformation was performed via transduction by mixing the packaged particles with
the overnight cultured E coli EPI300-T1R cells (1 10 vv ratio) in LB supplemented with 10
mM MgSO4 and 02 Maltose Transformation was completed by incubation at 37degC for 1 h
The transformed E coli cells were spread on an LB agar plate containing 125 microgmL
chloramphenicol The plates were incubated overnight at 37degC Colonies that appeared on the
plates were individually transferred to 96-well microtiter plates having 200 microL LB medium
supplemented with 10 glycerol and 125 microgmL chloramphenicol The microtiter plates
were incubated at 37degC for 24 h with continuous shaking at 180 rpm The prepared library
clones were stored at -80degC for further analysis and functional screening
108
109
Figure 4-2 Genetic map of copy controlled Fosmid vector (pCC1FOSTM
) The 813 kb
Fosmid vector backbone shows a multiple cloning site (MCS) chloramphenicol resistance
selectable marker (CmR) replication initiation fragment (redF) inducible high copy number
origin (oriV) replication initiation control (repE) and F-factor-based partitioning genes (ParA
ParB ParC) The vector contains a cos site for lamda packaging (not shown) and lacZ gene
for bluewhite colony selection The vector map was provided by the Epicentre
Biotechnologies (httpwwwepibiocomitemaspid=385)
110
432 Confirmation of library construction
The quality of the DNA library prepared was determined by examining 15 E coli
clones randomly selected A 15 mL LB broth supplemented with 125 microgmL
chloramphenicol was inoculated by each of the transformed clones To obtain higher Fosmid
DNA yield 1 microL autoinduction solution (L-arabinose) to increase the copy number of the
Fosmid vector (Epicentre) was added to the cultures After an overnight incubation of the
cultures at 37degC with continuous shaking at 220 rpm Fosmid DNA was extracted using a
Qiagen Large-Construct Kit followed by restriction digestion with Not I enzyme A 20 microL
digestion mixture containing 05 microg Fosmid DNA 1X digestion buffer and 05 U Not I was
incubated at 37degC for 15 h The partially digested DNA and a 42 kb DNA size marker
(Fosmid control DNA) were ran on 08 agrose gel for 15 h at 100 V The linear Fosmid
vector backbone (813 kb) and insert DNA fragments were visualized on the gel and the
image was recorded
433 Functional screening of the clones
Eppendorf tubes (15 mL) containing 500 microL LB supplemented with 125 microgmL
chloramphenicol were inoculated with a single clone (400 clones tested) or a combination of
12 library clones (600 tested) from microtiter plates stored at -80degC One half of the cultures
in each group (single or combined clones) were induced to increase the Fosmid copy number
by adding a 1 microL (500X) autoinduction solution (contained L-arabinose) to the culture tubes
L-arabinose induces the expression of a mutant trfA gene of the E coli host strain which
leads to the initiation of Fosmid replication from a low-copy to a high-copy number origin of
replication (oriV) The cultures were spiked with 20 microgmL DON and incubated at 37degC for
111
72 h with continuous shaking at 180 rpm The DON to de-epoxy DON transformation ability
of the clone cultures was examined by a liquid chromatography-ultraviolet-mass spectrometry
(LC-UV-MS) method as described by Islam et al (2012)
112
44 Results
441 Production of the DNA library
High molecular weight intact (non-degraded) and good quality (OD600280 ~190)
genomic DNA was obtained from the DON de-epoxidizing bacterial strain ADS47 (data not
shown) Pipetting of the DNA extract resulted in an adequate amount of insert DNA (gt025
microg) of approximately 40 kb size as showed on a LMP-PFGE (Figure 4-3)
A DNA library in E coli host was constructed using LMP-PFGE purified DNA The
library was comprised of 1250 E coli transformants which were selected on LB agar plates
supplemented with chloramphenicol (Figure 4-4)
113
Figure 4-3 Preparation of a 40 kb size insert DNA from a DON de-epoxidizing bacterium
Genomic DNA was extracted from the overnight culture of a DON de-epoxidizing
Citrobacter freundii strain ADS47 After fragmentation the DNA was size fractionated by
Low Melting Point- Pulse Field Gel Electrophoresis (LMP-PFGE) with markers for the PFGE
(M1) and 42 kb Fosmid control DNA (M2) S1 S2 S3= different DNA samples extracted
from bacterial strain ADS47 The red dots area indicates the section of the gel containing
genomic DNA of the expected size (~40 kb)
114
Figure 4-4 Production of the genomic DNA library in E coli EPI300-T1 host strain
Approximately 40 kb long blunt ended genomic DNA from a DON de-epoxidizing bacterial
strain ADS47 was ligated to a Copy Control Fosmid vector pCC1FOS The construct was
packaged into lamda bacteriophage followed by a transduction-mediated transformation of E
coli host cells The clones which appeared on the LB agar plate supplemented with 125
microgmL chloramphenicol are expected to be transformed by the recombinant Fosmid vector
containing insert DNA
115
442 Conformation of the library construction
All of the clones tested acquired the Fosmid vector Out of 15 clones examined 14
received the recombinant Fosmid vector that contain the expected size insert DNA as
determined by an 81 kb vector backbone and at least one high molecular weight insert DNA
band (~40 kb) on agarose gel (Figure 4-5) In contrast one clone (lane 4) showed an 81 kb
vector backbone and a low molecular weight band which is also a fragment of the Fosmid
vector The result indicated that approximately 94 of the library clones were transformed
with the recombinant vector
443 Activity-based screening of the library
Out of 500 single copy number clones (200 individual and 300 combined clones)
none showed DON to de-epoxy DON transformation activity (Table 4-1) An additional 500
high copy number clones (200 single and 300 combined clones) were unable to de-epoxidize
DON as well
116
Figure 4-5 Confirmation of the Fosmid DNA library production in E coli Fosmid DNA
collected from 15 clones was digested with Not I and run on 08 agarose gel at 100 V for 15
h The presence of the vector backbone (81 kb) and the insert DNA fragment (~40 kb) in the
clones was determined by comparing a 42 kb molecular size marker (M) Lanes (1-15) denote
the recombinant Fosmid DNA extracted from 15 E coli clones
117
Table 4-1 Analysis of Fosmid DNA library clones for deoxynivalenol (DON) de-epoxidation
capability
Type of clones Analysis of
singlecombined
clones
Number of
clones
examined
Number of clones
showed DON de-
epoxidation activity
Without induction
(single copy number clones)
Singly 200 0
Combination of 12 300 0
With induction (multi-copy
number clones)
Singly 200 0
Combination of 12 300 0
De-epoxidation activity of the clones was examined alone or in combination of 12 clones
without or with induction by adding L-arabinose The clones grown in LB supplemented with
125 microgmL and 20 microgmL DON were analysed for DON to de-epoxy DON transformation
ability by a liquid chromatography-ultra violet- mass spectrum (LC-UV-MS)
118
45 Discussion
While a Fosmid DNA library with the expected genome coverage of a DON de-
epoxidizing bacterial genome was produced the function-derived screening of the library
clones did not identify any clones that showed DON de-epoxidation enzymatic activity This
is unlikely due to lack of coverage of the bacterial genome as 1000 clones analyzed in the
study should have provided eight fold coverage of the genome Therefore it was likely due to
a lack of expression of the enzyme(s) for DON de-epoxidizing activity under the conditions
used
Thus far there is only one report of heterologous expression of bacterial epoxide
reductase gene in another bacterium A high level of heterologous expression of a
Synechococcus sp vitamin K epoxide reductase gene was achieved in E coli with codon
optimization and placing the gene under the control of a prokaryotic T7 promoter in an
expression vector (Li et al 2010) Therefore heterologous expression of bacterial epoxide
reducatse gene using its own promoter has not yet been demonstrated The success of the
method used in this chapter relies on an intact copy of the gene or operon being cloned the
selection of an appropriate cloning vector a host organism that can recognize the cloned C
fruendii promoter and a suitable library screening method for DON degradation by E coli
(Mocali and Benedetti 2010)
The unbiased cloning efficiency of the Fosmid vector its ability to contain large insert
DNA high stability in E coli as single copy number and autoinduction to increase copy
number in host cells made it appear to be a suitable method for the present work (Kim et al
1992 Lee et al 2004) Library production with large insertions is a major advantage as gene
cluster involved in particular pathway could be trapped into single clone and their functions
119
could be analysed The pathway of microbial DON de-epoxidation activity and the genes
encoding the activity are not known Therefore it was reasonable to prepare a library with
large inserts to ensure the possibility of cloning single as well as whole pathway genes of
interest for functional screening A further advantage of the method is that Fosmid clones are
inducible which allows detecting poorly expressed genes in a heterologous host by inducing
high copy numbers (Daniel 2005)
One of the limitations of the method is that the functions of the target genes rely
mostly on the host organismrsquos transcription and translation machineries Therefore selection
of the host organism is critical for activity-based screening of library clones in heterologous
hosts A number of host strains including E coli Streptomyces lividans Rhizobium
leguminosarum and Pseudomonas aeruginosa have been used for functional screening
activities from diverse organisms (Daniel 2005) The present work attempted to isolate C
freundii strain ADS47 genes encoding DON de-epoxidation enzymes Both strain ADS47 and
E coli belong to gamma proteobacteria which suggest that E coli would be an appropriate
host organism for heterologous expression of ADS47 genes Poor expression of heterologous
genes could be another limitation of the method which may be overcome by overexpression
to increase the catalytic activity of less expressed genes However clones with DON de-
epoxidation activity were not identified in this study even the Fosmid copy number was
increased by induction
In general a large number of clones must be analysed to obtain the gene of interest
using an activity-based screening technique (Lee et al 2004 van Elsas et al 2008) In this
study the constructed DNA library comprised of 1160 clones having approximately ~40 kb
insert DNA suggesting that the prepared library achieved ~10 fold coverage of the ADS47
120
genome (48 Mb) which ensured gt99 probability of the presence of all DNA sequence in
the library In the produced library three clones (lanes 3 10 and 12) showed one intact band
greater than 42 kb in size on the agarose gel It is presumed that the Fosmid DNA of the three
samples was not digested because of insufficient amount of restriction enzyme added to the
digestion mixture About half of the samples demonstrated multiple bands suggesting that the
insert DNA might have contained multiple Not I restriction sites
However not every heterologous gene may necessarily express or function in the E
coli host (Gabor et al 2004) The functionality of the gene requires expression of the genes
and proper folding of the protein which may be influenced by environmental conditions (eg
substrate temperature and pH) and genetic factors In this study library clones were screened
in growth conditions at which strain ADS47 performed the best de-epoxidation activity The
expression of genes may be controlled by the promoter suitable ribosome binding site and
transndashacting factors (eg special transcription factors inducers chaperones cofactors proper
modifying enzymes or an appropriate secretion system) (Gabor et al 2004) Those factors
have to be supplied by the cloned DNA fragments or host cells to allow any heterologous
gene to be functional Since DON de-epoxidation genes of strain ADS47 evolved in a distinct
environmental condition the expression and activity of the gene(s) might have required
specific factorsregulators that were absent in E coli Even though the Fosmid based library
screening technique facilitated isolation of many microbial genes clones having de-
epoxidation activity were not identified using such technique in this study
121
46 Acknowledgements
We appreciate the Agriculture and Agri-Food CanadaBinational Agricultural
Research and Development Fund (AAFCBARD) project for providing the financial support
for this work The authors thank Honghui Zhu for analyzing numerous samples by liquid
chromatography ultraviolet mass spectrum method
122
CHAPTER 5
50 GENOME SEQUENCING CHARACTERIZATION AND COMPARATIVE
ANALYSIS OF CITROBACTER FREUNDII STRAIN ADS47
51 Abstract
The genome of a trichothecene mycotoxin detoxifying C freundii strain ADS47
originating from soil was sequenced and analyzed Strain ADS47 contains a circular
chromosome of 4878242 bp that encodes 4610 predicted protein coding sequences (CDSs)
with an average gene size of 893 bp Rapid Annotation Subsystems Technology (RAST)
analysis assigned functions to 2821 (62) of the CDSs while 1781 (38) CDSs were
assigned as uncharacterized or hypothetical proteins Comparative bioinformatics analyses
were done for strain ADS47 C freundii Ballerup 785139 Citrobacter sp 30_2 C youngae
ATCC 29220 C koseri ATCC BAA-895 and C rodentium ICC168 There were a significant
number of common orthologous genes (2821) were found in the six Citrobacter genomes
implying a common evolutionary relationship ADS47 is most closely related to the
opportunistic human pathogen C freundii Ballerup The human and mouse pathogenic
species C rodentium and C koseri are the most distantly related to ADS47 and non-
humananimal pathogens C youngae and Citrobacter sp 30_2 were closely related ADS47
C koseri C youngae and Citrobacter sp 30_2 lacked the clinically important locus for
enterocyte effacement (LEE)-encoded type-III secretion systems which are present in C
rodentium and C freundii Ballerup Strain ADS47 had 270 unique CDSs many of which are
located in the large unique genomic regions potentially indicating horizontal gene transfer
123
Among the unique CDSs nine were reductasesoxidoreductases and a de-acetylase that may
be involved in trichothecene mycotoxin de-epoxidation and de-acetylation reactions
Keywords Citrobacter freundii comparative genomics genome sequencing unique genes
reductases
52 Introduction
According to the Food and Agriculture Organization (FAO) at least 25 of the
worldrsquos food crops are contaminated with mycotoxins causing serious negative impacts on
livestock and international trade (Boutrif and Canet 1998 Desjardins 2006 Binder et al
2007) Trichothecenes (TCs) produced by a variety of toxigenic Fusarium species are
probably the most prevalent food and feed contaminating mycotoxins It is estimated that
infections by the fungus caused more than $1 billion in crop yield losses in USA and Canada
alone (Windels 2000 Schaafsma et al 2002 Yazar and Omurtag 2008) TC mycotoxins can
have adverse affects on human health including organ toxicity mutagenicity carcinogenicity
neurotoxicity modulation of the immune system negative reproductive system affects and
growth retardation (DrsquoMello et al 1999 Pestka et al 2007) A strain of Citrobacter freundii
ADS47 was isolated from soil in southern Ontario that is capable of detoxifying (ie de-
epoxidation and de-acetylation) several TC mycotoxins including deoxynivalenol nivalenol
and T2-toxin (Chapters 2-3)
Citrobacter freundii is a gram negative motile rod shaped facultatively anaerobic
bacterium belonging to the family of Enterobacteriaceae (Wang et al 2000 Whalen et al
2007) The opportunistic human pathogen is responsible for a number of infections including
124
nosocomial infections of respiratory tract urinay tract blood and causes neonatal meningitis
(Badger et al 1999 Wang et al 2000) C freundii exists almost ubiquitously in soil water
wastewater feces food and the human intestine Biodegradations of tannery effluents copper
aromatic amino acids by C freundii strains isolated from water mine tailings and soil
respectively have been previously reported (Kumar et al 1999 Savelieva et al 2004 Sharma
and Fulekar 2009) Because of their biochemical capabilities bacteria have played important
roles in industrial biotechnology including de-epoxidation and de-acetylation activities
(Demain and Adrio 2008 Plank et al 2009 Zhao et al 2010) However the identification of
genes for de-epoxidation activity against trichothecene mycotoxins is limited (Boutigny et al
2008)
The advent of molecular techniques such as next generation sequencing technologies
and the related bioinformatics tools has enabled the discovery of novel genes and the
enzymes they encode in pathogenic and beneficial bacteria (van Lanen and Shen 2006
Ansorge 2009 Zhao 2011) In the present study the genome of mycotoxin-detoxifying strain
C freundii ADS47 was sequenced and subjected to comparative genomic analyses with other
Citrobacter species to identify putative mycotoxin-detoxifying genes
125
53 Materials and methods
531 Sequencing quality control and assemble of ADS47 genome
Genomic DNA of the strain ADS47 was extracted using Gentra Puregene
YeastBacteria Kit (Qiagen Inc Mississauga Ontario Canada) and purity of the DNA was
confirmed by determining OD 260280 at ~190 using a Nanodrop spectrophotometer
The genomic DNA was sent to be sequenced by the Illumina HiSeq 2000 sequencing
analyzer at Beijing Genomics Institute (BGI) Hong Kong
(httpwwwbgisequencecomhomeservicessequencing-servicesde-novo-sequencing) The
sequencing workflow involved short read library preparation cluster generation sequencing
and data analysis Briefly 3-5 ug of bacterial gDNA was fragmented to produce a 500 bp
insert library by preparative electrophoresis through a 08 agarose gel The fragmented
DNAs were blunt-ended by treating with a mixture of enzymes (T4 DNA polymerase klenow
fragment) and subsequently adapters were ligated to the blunt ends Each of the DNA
fragments was amplified to a cluster which was performed in the flow cell by template
hybridization and bridge amplification Millions of clusters were then sequenced by
sequencing-by-synthesis technology leading to base calls The raw data was filtered to remove
low abundant sequences from the assembly according to 15-mer frequencies The low
complexity reads low quality (leQ20) base calls and adapter contaminations (24 bp overlap
between adapter and reads) were eliminated The quality of the sequences was analysed by
GC content and depth correlative analysis (for GC biasness) K-mer analysis was performed
using 15 bp of the short reads for heterologous sequence identification
126
The cleaned short reads (approximately 90 bp) were self-assembled using the Short
Oligonucleotide Analysis Package (SOAP) denovo (httpsoapgenomicsorgcn) (Li et al
2008) The assembly errors were corrected by filling gaps and proofreading single base pairs
using mapping information The assembly produced 441 contigs with an N50 contig size of
29222 bp (the largest contig size 114839 bp) and 80 scaffolds with an N50 scaffold size of
325726 bp (largest scaffold 662452 bp) with an total sequence length of 4878242 bp which
constitutes 33-fold genome coverage The scaffolds were ordered and oriented according to a
reference genome C koseri ATCC BAA-895 the closest completely sequenced species of
the newly sequenced bacterial genome by alignment with the MUMmer program v322
(httpmummersourceforgenet) (Delcher et al 2002) Genome coverage was determined by
mapping the reads to the assembled genome with Bowtie software (Langmead et al 2009) A
preliminary genome annotation was performed to define the origin of replication by
identifying colocalization of multiple genes ie gyrB recF dnaN and dnaA (usually found
near the origin of replication) and comparison with the C koseri reference genome Synteny
analysis between the reference bacterium and assembled scaffolds was performed with the
MUMmer program
532 Gene function annotation and classification
The genome of ADS47 was annotated by Rapid Annotation using Subsystems
Technology v40 (RAST) a server-based genome analysis system (httprastnmpdrorg)
(Aziz et al 2008) A circular genome map was constructed using CGView program
(httpstothardafnsualbertacacgview_server) (Stothard and Wishart 2005)
127
533 Molecular phylogenetic analysis
A robust phylogenetic tree was produced using Composition Vector Tree (CVTree)
web-based program v20 (httpcvtreecbipkueducn) (Qi and Hao 2004) by comparing
whole genome amino acid sequences of strain ADS47 two completely sequenced Citrobacter
genomes [C rodentium ICC168 (NC_013716) and C koseri ATCC BAA-8959 NC_009792)]
and three partially sequenced Citrobacter genomes [C freundii strain Ballerup 785139
(CACD00000000) Citrobacter sp 30-2 (NZ_ACDJ00000000) and C youngae ATCC 29220
(NZ_ABWL00000000)] E coli O157H7 strain EDL933 (NC_002655) was used as an
outgroup species The tree was constructed from the generated pairwise distance matrix data
using Neighbor-Joining Program in PHYLIP v369 package (Saitou and Nei 1987 Felsenstein
1989)
534 Comparative genomic analysis
Genome-wide comparative analyses between C freundii ADS47 C freundii strain
Ballerup 785139 Citrobacter sp 30-2 C youngae ATCC 29220 C Koseri ATCC BAA-895
and C rodentium ICC168 was carried out The nucleotide sequence idendities were
determined by pairwise alignments of the ADS47 genome with the five other Citrobacter
genomes using MUMmer software (Delcher et al 2002) and GC content total CDSs genome
coverage by CDSs and number of stable RNAs were determined by RAST program
Orthologous CDS groups among the total set of 29079 CDSs derived from the six
Citrobacter genomes were determined using OrthoMCL software v202
(httporthomclsourceforgenet) based on all-against-all reciprocal BLASTP alignment of
amino acid sequences (Li et al 2003) Genes unique to each strain were identified as those
128
CDSs not included in any orthologous group of the five other Citrobacter genomes BLASTP
comparisons were visualized with Blast Ring Image Generator (BRIG) software v095
(httpsourceforgenetprojectsbrig) (Alikhan et al 2011)
Protein sequences of the five Citrobacter genomes pooled from the GenBank data
bases were compared with ADS47 A circular map was developed by BRIG software that
showed the unique genomic regionsgenes of ADS47 The unique genes were grouped
according to their functional similarities previously identified by RAST software and
genomic regions possibly acquired by horizontal transfer were identified using RAST
software
129
54 Results
541 Assembly and organization of the ADS47 bacterial genome
Scaffolds derived from Citrobacter freundii strain ADS47 were ordered and oriented
by alignment to Citrobacter koseri ATCC BAA-895 the most closely related genome that has
been completely sequenced The genomes of ADS47 and C koseri displayed close synteny
(Figure 5-1)
Figure 5-1 Co-linearity between C freundii strain ADS47 genome and C koseri Scaffolds
derived from ADS47 genome sequences (Y- axis) were aligned and assembled by comparison
with the closest completely sequenced genome C koseri ATCC BAA-875 (X- axis)
130
542 An overview of the ADS47 genome
General features of the C freundii strain ADS47 are presented in Table 5-1 Some
notable features are that strain ADS47 contains 4881122 bp circular chromosome with an
average GC content of 5202 The genome contains 4610 CDSs which comprise 8784 of
the whole genome Annotation by RAST assigned 2821 (62) CDSs to known biological
functions and remaining 1789 (38) genes were classified to unknown or hypothetical
proteins
The GC content in the coding regions is significantly higher (5318) than in the non-
coding intergenic region (4379) which covers 1215 (592781 bp) of the genome The
intergenic region is comprised of 018 repeat sequences (total 8999 bp) 5 (009)
transposons (4806 bp) 025 stable RNA (12091 bp) and the remaining ~115 is
classified as regulatory and other functions The ADS47 genome contains 62 tRNAs 3
rRNAs and 34 copies of sRNAs The average lengths of the tRNAs rRNAs and sRNAs are
785 885 and 13441 bp respectively There are also 42 phageprophage-related genes
identified in the genome
Figure 5-2 shows the distribution of the 26 categorized CDSs rRNA tRNA GC
content and GC skew It appears that ADS47 genome contains equal quantity of genes in
leading and lagging strands and showed occasional clustering (likely operons) of genes in the
two strands The genome contains three rRNAs (two large subunit and a small subunit) and 62
tRNAs which are distributed across the genome A consistent GC content was observed in
the ADS47 genome however 7-10 low GC content (high black color peaks) genomic regions
were found at locations origin of replication 600 2000 3000 3300 3550 3850 4300 and
4800 kbp This is indicative of possible horizontally transferred genes or insertions in those
131
genomic locations GC skew denotes the excess of C over G in certain genomic locations
Positive GC skew (+) (green color) indicates the skew value in the leading strand while
negative skew (-) (purple color) in the lagging strand As in many bacterial genomes the
ADS47 genome map shows the separation of positive and negative GC skew at the origin of
replication (base pair position 1) The GC skew in the leading strand is highly skewed (high
green color peak) indicating that possible horizontal gene transfer occurred into the ADS47
genome
The CDSs ranged from ~100 bp to ~2000 bp and have an average size of 893 bp
(Figure 5-3) The highest numbers of CDS (770) were 600-800 bp long A considerable
number of genes (220) were gt2000 bp in size Some of the biological functions for the
smallest genes (100-200 bp) are sulfur carrier protein methionine sulfoxide reductase
ribonuclease P protein component while the largest genes (gt2000 bp) are ferrous iron
transport protein B translation elongation factor G pyruvate flavodoxin oxidoreductase and
tetrathionate reductase
132
Table 5-1 General features of the C freundii strain ADS47 genome
Feature Chromosome
Genome size 4881122 bp
Overall GC content 5202
Number of genes 4610
Genes with assigned function (number and ) 2821 (62 )
Genes without assigned function (number and ) 1789 (38 )
Total length of protein coding sequences 4285461 bp
Genome coverage by coding sequences () 8784
Average length of coding sequences 893 bp
GC content in the coding region 5318
GC content in the non-coding region 4379
Length and percentage of intergenic region 592781 bp and 1215
Total repeat sequences and genome coverage 8999 bp and 018
Total transposons sequences and genome coverage 4806 and 009
Number average length and genome coverage of tRNA 62 copies 785 bp and 0099
Number average length and genome coverage of rRNA 3 copies 885 bp and 0067
Number average length and genome coverage of sRNA 34 copies 13441 bp and 0093
133
134
Figure 5-2 Circular genome map of C freundii strain ADS47 Map was created with the
CGView software From outside to inside the first circle indicates the genomic position in kb
Different functional categories of predicted coding sequences (CDS) are distinguished by
colours with clockwise transcribed CDSs in the second circle and anticlockwise transcribed
CDSs in the third circle Bars on the fourth and fifth circles indicate the rRNAs and tRNAs
respectively Sixth and seventh circles show the coverage and GC content respectively The
innermost circle demonstrates the GC skew + (green) and GC skew- (purple)
135
Figure 5-3 Size distribution of the predicted genes identified in C freundii strain ADS47
The X axis identifies the gene size categories (bp) and Y axis the abundance of genes
136
543 Annotation and functional classification of ADS47 genome
RAST analysis identified predicted CDSs in the genome of Citrobacter strain ADS47
assigned functions and grouped the annotated genes into 26 functional subsystem categories
out of 27 No CDS was found belonging to the photosynthesis category A graphical
presentation of the distribution of the CDSs into each of the 26 categories is given in
Figure 5-4
A large portion (38) of the CDSs belong to hypothetical or unknown functions
Among the 62 known or predicted proteins the highest numbers (835) belong to the
carbohydrate category followed by amino acids and derivatives (468) and
cofactorsvitaminsprosthetic groups (324) A noticeable number of stress response (236) and
respiration (198) categories genes were identified in the bacterial genome The ADS47
genome encoded by 171 membrane transport genes 113 virulencedefense genes and 126
genes involved in regulationcell signaling categories Two hundred and one CDSs were
grouped into the miscellaneous category whose functions are different than the indicated
RAST categories The RAST annotations further divided the categorized CDSs into
subcategories based on similar functional roles which is discussed in section 545
137
Figure 5-4 Functional classification of the predicted coding sequences (CDS) of C freundii
strain ADS47 The annotation was performed by Rapid Annotation using Subsystems
Technology (RAST) program
138
544 Molecular phylogenetic relationships
Genome-wide phylogenetic analysis based on comparisons of whole genome amino
acid sequences of two completely sequenced Citrobacter genomes (C koseri and C
rodentium) and three partially sequenced Citrobacter genome sequences (C freundii
Citrobacter sp and C youngae) illustrates the relationships of ADS47 to five Citrobacter
species (Figure 5-5)
C youngae ATCC 29220 originating from human feces and two pathogenic species
(C koseri ATCC BAA-895 and C rodentium strain ICC168) are progressively more distantly
related to ADS47 The present analysis is consistent with the 16S-rRNA sequence based
phylogenetic analysis (Chapter 3) which showed that the closest bacterial strain to ADS47 is
C freundii (Figure 3-2) This confirmed the identification of strain ADS47 as the gamma
proteobacterial species C freundii made in Chapter 3
139
Figure 5-5 Phylogeny of the trichothecene detoxifying C freundii strain ADS47 The
dendrogram was constructed by comparing the ADS47 genome (amino acid sequences) with
five Citrobacter genomes and a E coli strain (out group species) using the CVTree software
The calculated pairwise distances matrix by Protdist program was transformed into a tree by
neighbor-joining program The dissimilarity tree was drawn to scale with branch lengths
(scale bar 002 indicated the amino acid substitutions among the genomes) The C koseri C
rodentium C freundii Citrobacter sp C youngae and E coli protein sequences were
uploaded from GenBank (Benson et al 2009)
140
545 Comparative genomics
Genome-wide comparative analyses between the genome of strain ADS47 and the five
previously sequenced Citrobacter genomes showed that 872 of the total nucleotides
between ADS47 and C freundii Ballerup were identical The values for the comparisons
between C freundii strain ADS47 were 8212 for Citrobacter sp 30-2 7637 for C
youngae 5981 for C koseri and 4213 for C rodentium (Table 5-2)
The two human and mouse pathogenic species namely C rodentium and C koseri
and the bacteria originally isolated from human feces (C youngae) have higher GC contents
(546 and 532 respectively) compared to C freundii strain ADS47 C freundii strain
Ballerup and Citrobacter sp 30_2 which have slightly lower GC contents (~52) The two
pathogenic species contain 22 copies of rRNA which is significantly higher than the 9 found
in C freundii strain Ballerup 6 in Citrobacter sp 30_2 and 3 in C freundii strain ADS47 but
less than the 25 copies in C youngae The genome of C freundii strain ADS47 C freundii
strain Ballerup C youngae and Citrobacter sp 30-2 had a similar percentage of the genome
comprising coding sequences (865-88) C koseri and C rodentium genomes contained the
highest (90) and lowest (84) genome coverage by coding sequences respectively
141
Table 5-2 Genome comparisons between strain ADS47 and five Citrobacter species The
base nucleotide sequence identities were performed using MUMmer DNAdff script program
(Delcher et al 2002) The features (GC content total CDS percentage CDS and number of
ribosomal RNA) were determined using RAST program (Aziz et al 2008)
Genome feature Bacterial species
C
freundii
ADS47
C freundii
Ballerup
Citrobacter
sp 30_2
C
youngae
C koseri C
rodentium
Genome length
(bp)
4878242
4904641
5125150
5154159
4720462
5346659
Base similarity to
ADS47 ()
NA 8715 8212 7637 5981 4213
GC () 5202 5218 516 5265 530 546
Number of coding
sequences
4610 4685 4728 5278 4978 4800
Genome coverage
with coding
sequences ()
880 865 866 872 900 840
rRNA copies 3 9 6 25 22 22
Total stable RNAs 99 81 78 105 120 142
142
The number of orthologs shared between ADS47 and other Citrobacter species is
illustrated in Figure 5-6 ADS47 shares the smallest number of orthologous CDSs with C
rodentium and C koseri 3250 and 3341 respectively The highest number of shared CDS
were found with C freundii Ballerup (3706) followed by C youngae (3693) and Citrobacter
sp 30_2 (3676) A total of 2821 CDSs were found to be common to all of the six Citrobacter
genomes analysed (data not shown) Nonetheless significant diversity exists between C
freundii ADS47 and the other species as a considerable number of unshared genes were
identified from the comparisons with C freundii Ballerup [979 (20)] Citrobacter sp 30_2
[1064 (23)] C youngae [1654 (31)] C koseri [1693 (34)] and C rodentium [1634
(34)]
143
Figure 5-6 Comparisons of coding sequences between strain C freundii ADS47 and five
Citrobacter species Coding sequences of ADS47 were compared with C freundii Ballerup
Citrobacter sp 30_2 C youngae C koseri and C rodentium genomes based on all-versus-all
reciprocal FASTA comparison of at least 30 amino acid identity and gt70 coverage of the
total length using OrthoMCL program A Venn diagram demonstrates the shared and
unshared (indicated in the brackets) coding sequences
144
Table 5-3 shows differences in the 26 RAST categorized genes between the six
Citrobacter genomes Even though a low abundance of virulencediseasedefense category
genes were found in the animalhuman pathogenic bacteria of C rodentium (85) and C koseri
(92) significantly higher mycobacterium virulence genes (13) were identified in this two
species The genomes of the two closely related strains C freundii ADS47 and C freundii
Ballerup lack this type of virulence gene Instead elevated numbers of genes for resistance to
antibiotic and toxic compounds were found in C freundii ADS47 (94) C freundii Ballerup
(88) and Citrobacter sp (110) compared to other three species Copper homeostasis and
cobalt-zinc-cadmium resistance genes were identified in strain C freundii ADS47 (13 and 21
respectively) and closely related C freundii Ballerup (12 and 22) and Citobacter sp (18 and
22) Significantly lower numbers of these genes were present in C rodentium (6 and 4) and C
koseri (7 and 8) C rodentium and C koseri lack mercuric reductase mercuric resistance and
arsenic resistance genes perhaps because being animalhuman pathogens but these genes
were found in the remaining four genomes The C koseri genome is richer in betandashlactamase
genes (5) compared to the remaining genomes which encodes 1-3 genes
145
Table 5-3 Functional categorized gene abundances among the genomes of C freundii
ADS47 C freundii Ballerup Citrobacter sp 30_2 C youngae C koseri and C rodentium
The genomes were classified into different functional categories with the RAST program
which classified the total CDSs
RAST subsystem category Citrobacter species
C
freundii
ADS47
C
freundii
Ballerup
C
sp
C
youngae
C
koseri
C
rodentium
Cofactorsvitaminsprosthetic
groupspigments
323
317
310
311
325
304
Cell wall and capsule 279 272 248 262 247 262
Virulence disease and defense 113 107 134 100 92 85
Potassium metabolism 50 53 33 32 30 28
Miscellaneous 210 228 200 185 245 185
Phageprophagetransposable
element plasmid
24 75 25 4 6 49
Membrane transport 170 156 171 195 171 172
Iron acquisition and metabolism 80 81 86 62 113 27
RNA metabolism 220 222 217 215 238 212
Nucleosides and nucleotides 119 112 110 111 120 114
Protein metabolism 249 280 269 298 287 254
Cell division and cell cycle 30 29 32 29 38 29
Motility and chemotaxis 133 124 90 147 143 133
Regulation and cell signaling 126 130 124 135 132 123
Secondary metabolism 21 22 20 21 0 0
DNA metabolism 145 167 165 155 147 152
Fatty acids lipids and isoprenoids 121 149 122 121 134 133
Nitrogen metabolism 50 52 53 49 56 58
Dormancy and sporulation 3 3 3 4 3 3
146
Table 5-3 (continued)
RAST subsystem category
Citrobacter species
C
freundii
ADS47
C
freundii
Ballerup
C
sp
C
youngae
C
koseri
C
rodentium
Respiration 198 184 180 191 166 162
Stress response 236 235 170 177 179 174
Metabolism of aromatic
compounds
20 20 28 25 17 48
Amino acids and derivatives 482 476 450 466 428 416
Sulfur metabolism 48 50 52 57 56 56
Phosphorus metabolism 50 49 43 42 43 28
Carbohydrates 837 803 708 731 733 737
147
Strains C freundii ASD47 and C freundii Ballerup have much more potassium
metabolism category genes (50 and 53 respectively) than the other bacterial species which
code for 28-33 proteins Larger numbers of genes associated with the glutathione regulated
potassium efflux system were also identified in C freundii ASD47 with 23 and C freundii
Ballerup with 26 compared to the remaining four species that encode only five genes All of
the six genomes contained phageprophage-related genes with the maximum identified in C
freundii Ballerup (75) followed by C rodentium (49) Citrobacter sp (25) and C freundii
ADS47 (24) Lower numbers of phageprophage-related genes were found in C koseri (6) and
C youngae (4)
The six Citrobacter genomes showed remarkable variability in their secretion systems
The type-III secretion operon genes are only found in C rodentium and C freundii Ballerup
Type-II secretion genes are predominant in C rodentium (12) followed by ADS47 (9)
Type-IV secretion system is uncommon in Citrobacter species since it was only
identified in Citrobacter sp where it is represented by 19 genes Variable numbers of type-VI
secretion system genes are present in the five genomes except C koseri which lacks such
genes C youngae has the highest number (43) of type-VI secretion proteins The other strains
have a range of type-VI secretion proteins in particular 27 were identified in C freundii
Ballerup 15 in C freundii ADS47 and 8 in Citrobacter sp Type-VII (chaperoneusher)
secretion system genes were found in all of the six Citrobacter species with the largest (42)
and smallest (13) numbers of these genes occurring in C koseri and C freundii Ballerup
respectively
Although a relatively similar number of ABC transporters (37-42) were identified in
the six genomes C freundii ADS47 contain the largest number of tripartite ATP-independent
148
periplasmic (TRAP) transporter genes (7) By contrast the closely related C freundii
Ballerup lacks such transporter system An identical number of iron acquisition and
metabolism category genes were found in C freundii ADS47 (80) and C freundii Ballerup
(81) while the two pathogenic species C rodentium and C koseri code for the lowest (27)
and highest (113) numbers of such genes C koseri is rich in siderophore related genes (64)
but the remaining five genomes have smaller numbers of siderophore-associated genes (20-
22)
Variable numbers of motility and chemotaxis related genes were found in the
Citrobacter genomes with the largest number occurring in C youngae (147) followed by C
koseri (143) C freundii ADS47 (133) C rodentium (133) C freundii Ballerup (124) and
Citrobacter sp (90) The two pathogenic species (C rodentium and C koseri) encode a
remarkably low number of quorum sensing genes (2) In contrast C youngae codes for the
highest number of quorum sensing genes (19) A common and distinctive feature of the two
pathogenic species is the lack of genes required for secondary metabolism In contrast the
non-pathogenic species contain a common set of genes for degrading a range of
phenylpropanoids and cinnamic acid (20-22)
The lowest number of genes related to phospholipidsfatty acids biosynthesis and
metabolism were found in C freundii ADS47 (66) compared to C freundii Balllerup (94) and
C koseri (98) The smallest numbers of nitratenitrite ammonification genes were found in
ADS47 (31) and its closest genome C freundii Ballerup (32) In contrast the two pathogens
encode the highest number of such proteins (39-40)
C rodentium and C koseri code for significantly low numbers of respiratory category
genes (162-165) while C freundii ADS47 contains the highest number of such genes (198)
149
Further analysis of the subcategory genes determined that the two gut species (C youngae and
Citrobacter sp 30_2) contain the largest numbers of genes encoding anaerobic reductases
(22) and identical numbers of formate dehydrogenases (17) The largest numbers of formate
dehydrogenase genes were identified in C freundii Ballerup (32) and C freundii ADS47 (30)
but the two pathogenic species differed from the other species by encoding the lowest number
(13) of c-type cytochrome biogenesis genes
A higher number of aromatic compound metabolism category genes were found in the
mouse pathogen C rodentium (48) compared to the human pathogen (C koseri) which
contain 17 genes A significantly higher number of genes involved in central aromatic
intermediate metabolism (26) were found in C rodentium compared to the mouse pathogen
(C koseri) and other genomes which only contain 6-8 such genes The two C freundii strains
ADS47 and Ballerup lacked the anaerobic degradation of aromatic compound
(hydroxyaromatic decarboxylase family) genes
In spite of the abundance of stress response category genes strain ADS47 has only a
small number of desiccation stress genes (5) compared to the other species which ranged
from 9 to10 However ADS47 and Ballerup code for higher number of genes related to
oxidative stress (94 and 91 respectively) and cold shock (8 and 7 respectively) than the other
species
The smallest numbers of amino acids and its derivatives category genes were found in
the two pathogens C rodentium and C koseri (416 and 428 respectively) while the largest
number of these genes were identified in strain ADS47 (482) and C freundii Ballerup (476)
Despite the similar number of sulfur metabolism category genes determined across the six
species only ADS47 lacks taurine utilization genes Strains ADS47 and Ballerup were found
150
to be rich in the phosphorous metabolism gene category (49-50) while C rodentium contains
a significantly decreased number of these category genes (28)
The bioinformatics analyses found the largest number of carbohydrate category genes
in ADS47 (837) followed by Ballerup (803) The gut bacteria (Citrobacter sp 30_2) codes for
the least number of genes assigned to the carbohydrate category (708) while C koseri C
rodentium and C youngae contained 731-737 in this category genes The analyses also
determined that there are relatively small numbers of TCA and Entner Doudoroff pathway
genes in C rodentium and C youngae compared to the other four species and relatively large
numbers of fermentation-related genes in ADS47 (81) and C freundii (85) compared to the
other four species
A comparison of the pathogenicity operons [locus of enterocyte effacement (LEE)] of
C rodentium with other five Citrobacter genomes revealed the absence of this virulence
determinant gene region in the other four Citrobacter species including ADS47 (Figure 5-7)
The analysis showed the presence of LEE operon genes in C freundii strain Ballerup as no
gap was observed in that genomic location while C freundii ADS47 C koseri C youngae
and Citrobacter sp 30_2 lacked that loci (7a) Further ananlysis showed alignments between
C rodentium and E coli LEE operon genes (Figure 7b) In contrast no alignment of such
genes was observed in ADS47 genome
151
(a)
152
Figure 5-7 (a) Blast Ring Image Generator (BRIG) output based of Citrobacter rodentium
(NC_013716) compared against ADS47 C freundii C koseri C youngae E coli and
Citrobacter sp genomes The targeted comparative genome analysis was performed based on
locus of enterocyte effacement (LEE) operons of C rodentium (Petty et al 2011) The
innermost rings show GC content (black) and GC skew (purple and green) The remaining
rings show BLAST comparisons of the other Citrobacter and E coli genomes against C
rodentium The region shown in red is LEE operon genes (b) The absence of LEE operons in
the ADS47 genome comparing to C rodentium and E coli Analysis was performed and
visualized by Artemis Comparison Tool software v132
(httpwwwsangeracukresourcessoftwareact) (Tatusov et al 2000 Craver et al 2005)
LEE operons
(b)
153
546 Analysis of the unique genes in C freundii ADS47
Two hundred and seventy genes unique to ADS47 were identified from a comparison
of its genome to the other five Citrobacter species In general the unique genes are
distributed throughout the ADS47 genome but some clusters of unique genes occurred
including eight unique genomic regions (Figure 5-8) Six large unique regions are located at
767204-775090 (~8 kb) 1025572-1042510 (17 kb) 2798289-2807217 (~9 kb)
3045220-3055912 (~11 kb) 3930366-3943075 (12 kb) and 4022195-4032444 (10 kb)
Out of 270 unique genes 147 (54 ) are hypothetical proteins The remaining 123
(46) genes are grouped into reductasesoxidoreductases (9 genes) de-acetylase (1)
hydrolases (2) transferases (4) sulfatasesarylsulfatases (3) bacteriocins (3) type VI
secretion proteins (2) kinases (4) transporters (11) membraneexported proteins (8)
regulators (12) integraserecombinases (5) phagesprophagestransposaes related genes (27)
and miscellaneous (32) that includes DNA helicases ligases fimbrial proteins and
chromosome partitioning genes (Table 5-4)
The unique regions encode various functional genes that may cause trichothecene de-
epoxidation or de-acetylation reactions The 8 kb unique regions contains six genes encoding
glycosyltransferases and two copies of oxidoreductase-like coding sequences (CMD_0711
and CMD_0712) that showed 39 identity to the Marinobacter aquaeolei F420
hydrogenasedehydrogenase beta subunit The largest (17 kb) unique genomic region codes
for 16 genes including dehydrogenase hydrolase bacteriocin (pyocin) integrase and a short
chain dehydrogenasereductase-like (SDR) (CMD_0979) gene (Figure 5-9a)
154
The SDR like gene is 49 amino acid similarity to an oxidoreductase of the
phytopathogenic Dickeya dadantii and the enzyme contains a domain with 77 homology to
the Dickeya zeae NAD(P) binding site The 9 kb unique genomic section contains 8 genes
including a NaCl symporter a dehydratase (CMD_2679) with 86 homology to
Enterobacteriaceae bacterium dihydroxy-acid dehydratase and a putative oxidoreductase
(CMD_2683) having 37 similarity with oxopropionate reductase of Bacillus sp The unique
region also encodes a short chain dehydrogenasereductase (SDR) (CMD_2684) that shows
85 identity to Enterobacteriaceae bacterium (Figure 5-9b) A unique thiol-disulfide and
thioredoxin like reductase (CMD_3721) was identified in a 10 kb unique region that contains
13 genes (Figure 5-9c) This gene is 59 identical to Oligotropha carboxidovorans thiol-
disulfide isomerase and thioredoxin In addition several reductasesoxidoreductase-like
unique genes were identified in the ADS47 genome including an oxidoreductase
(CMD_1024) with 27 identity to Canadidatus nitrospira NADH-quinone oxidoreductase
membrane L Another oxidoreductase-like gene identified in a different location of the
ADS47 genome (CMD_1393) shows 87 homology to Klebsiella pneumoniae
deoxygluconate dehydrogenase contains a SDR active site A putative nitrate reductase gene
(CMD_3184) has a low level of identity (27) with an uncultured bacterial respiratory nitrate
reductase The ~11 kb unique region encodes sulfatasesarylsulfatses
(CMD_2898CMD_2905) and related genes Furthermore ADS47 contains a unique
glycosylphosphatidylinositol (GPI) anchor biosynthesis operon comprised of four genes
including a N-acetylglucosaminyl-phosphatidylinositol de-acetylase (CMD_2431) which
showed 78 similarity to E coli de-acetylase (Figure 5-9e)
155
Other unique genespredicted genes in the ADS47 genome include
polysaccharidephenazine biosynthesis genes (CMD_0710CMD_1406) heavy metal kinase
(CMD_0989) two component heavy metal regulator (CMD_0990) nitrate reductase regulator
(CMD_3477) putative potassiumzinc transporters (CMD_0973CMD_4600) sodium
symporter (CMD_2678) (tripartite ATP-independent periplasmic transporters) TRAP
transporters (CMD_3693 CMD_3694 CMD_3695 CMD_4337) glutamineribose ABC
transporters (CMD_3176CMD_3544) toxin transcription regulator (CMD_3464) fimbrial
proteinchaperone (CMD_1695 CMD_2434 CMD_3277 CMD_3278) and ParB
chromosomeplasmid partitioning proteins (CMD_4033CMD_4034)
Majority of the unique genes and their distributions are shown in comparative circular
genome maps (Figure 5-8)
156
Table 5-4 Unique genes of C freundii strain ADS47 Genes were categorized according to
the functional similarity The unique genes were determined by comparing ADS47 with five
sequenced Citrobacter species using OrthoMCL software
Unique gene classes Number
Total unique genes
270
Reductases oxidoreductases 9
De-acetylase 1
Hydrolase 2
Transferase 4
Sulfatase 3
Bacteriocin 2
Type VI secretion 2
Kinase 4
Transporter 9
Membraneexported protein 8
Regulator 12
Integraserecombinase 5
Phageprophagetransposae 27
Miscellaneous 34
Hypothetical protein 147
157
158
Figure 5-8 Locations of the unique genes of C freundii strain ADS47 The unique genes
were determined by comparing the total CDS of strain C freundii ADS47 with C freundii
Ballerup Citrobacter sp C youngae C koseri and C rodentium CDSs by OrthoMCL
program The map was developed by Blast Ring Generator (BRIG) software First second
and third circles from inside to out side show the genome size GC and GC skew of strain
ADS47 Forward (Fwd) and reversed (Rev) transcribed CDSs of ADS47 are shown in green
(forth circle) and red (fifth circle) respectively The next five rings represent the circular
genome maps of five Citrobacter species distinguished by different ring colours (C
rodentium as sky blue C koseri as dark blue C youngae as light blue C freundii Ballerup as
yellow and Citrobacter sp as green) The degree of orthologous gene identity between
ADS47 and other five genomes are shown by color intensity (light color= 75 gene identity
ash color= 50 gene identity) Gaps across five genome rings indicate the absence of
genesregions compared to ADS47 Unique genes are shown surrounding the map rings and
the identities of some of the genes are given Arrows indicate putative
reductasesoxidoreductases and de-acetylase
159
(a)
(b)
160
(c)
(d)
161
Figure 5-9 Unique genomic regions and associated genes in C freundii ADS47 compared to
five Citrobacter species RAST server based analysis compared the unique genesregions of
ADS47 with the publicly available databases The upper panels demonstrate the large unique
regions defined by yellow boxes and genes located in the genomic regions are indicated by
arrows The lower panels are closely related genesoperons found in distantly related species
The unique regions that contain potential trichothecene de-epoxidation genes are shown in (a)
short chain dehydrogenasereductase (SDR) and dehydrogenase (b) putative oxidoreductase
dehydratase and SDR (c) thiol-disulfide isomerase and thioredoxin oxidoreductase and (d)
unique glycosylphosphatidylinositol (GPI) biosynthesis operon having N-acetylglucosaminyl-
phosphatidylinositol de-acetylase gene
162
55 Discussion
The de novo sequenced ADS47 genome was oriented and assembled using a
completely sequenced C koseri genome as reference strain The synteny analysis between
ADS47 and C koseri showed very similar overall organization of the two genomes The high
preservation of genes among five Citrobacter species and ADS47 were emphasized by the
close alignments observed by Blast Ring Image Generator (BRIG) outputs obtained from
these genomes
Genome-wide phylogenetic analyses based on a large number of common CDSs
(2821) observed within the six Citrobacter genomes confirmed the species identification
based on 16S-rDNA sequencing of the trichothecene mycotoxin-detoxifying C freundii strain
ADS47 The base sequence idendity was 8715 between these two genomes They shared
~80 CDS and had a similar GC content (52) Citrobacter are Gram-negative rod shaped
and facultatively anaerobic bacteria belonging to the family of Enterobacteriaceae (Wang et
al 2000 Whalen et al 2007) Strains ADS47 and Ballerup of C freundii are both aerobic
gram-negative bacilli that have numerous flagella and inhabit in soil (Cong et al 2011 Islam
et al 2012) C freundii is considered to have a positive environmental role by reducing nitrate
to nitrite (Rehr and Klemme 1989) which is an important step in the nitrogen cycle
C freundii C rodentium and C koseri are animalhuman pathogenic species C
rodentium is a natural mice pathogen causing hemorrhagic colitis and hemolytic uremic
syndrome (Gareau et al 2010) C koseri causes human disease (Badger et al 1999 Kariholu
et al 2009) particularly neonatal meningitis and C freundii is an opportunistic human
pathogen responsible for a various types of infections like respiratory tract urinary tract and
blood (Whalen et al 2007) C freundii is very common representing approximately 29 of
163
all opportunistic human infections (Hodges et al 1978) Outside of animals they can be found
in soil water sewages and different organs of diseased animals (Wang et al 2000) These
pathogens code for large numbers of mycobacterium virulence operonsgenes which were
absent in the three other genomes However they have fewer genes involved in environmental
stress such as copper homeostasis genes and no mercuricarsenic resistance and secondary
metabolism category genes unlike the other Citrobacter genomes Nonetheless considerable
variability exists between the two pathogens correlates with C rodentium causing disease in
mice C koseri causing meningistis in human and C freundii causing infections in different
human organsThere were attaching and effacing (AE) lesion forming (LEE) operons in C
rodentium and C freundii but not in C koseri Those genes have shown to be acquired by
lateral gene transfer playing major roles in virulence of enteropathogenic and
enterohemorrhagic E coli and C rodentium (Mundy et al 2005 Petty et al 2010 Petty et al
2011 The ~356 kb pathogenic LEE gene island is comprised of five operons (LEE1 to
LEE5) that code for ~40 virulence genes that include type-III secretion genes and associated
effectors (Berdichevsky et al 2005) In addition an elevated number of type-II and -VI
secretion proteins identified in C rodentium which may act as additional virulence factors
(Petty et al 2010) Because it lacks type-III secretion apparatus genes it is likely that C
koseri uses a different strategy to cause human neonatal meningitis (Badger et al 1999 Ong
et al 2010)
To adapt to different environmental niches or hosts bacteria like Citrobacter species
frequently gainlose genes increase repeat sequences and recombine genomic regions using
different mechanisms (Parkhill et al 2001 Petty et al 2011) The existence of large numbers
of phageprophagetransposae related genes in the Citrobacter genomes particularly in C
164
freundii Ballerup suggests that their genomes are unstable For example C freundii Ballerup
and C koseri might have acquired LEE operons by horizontal transfer via phage mediated
transpositions Interestingly although ADS47 is closely related to C freundii Ballerup it
lacks the pathogen-related LEE gene island and mycobacterium virulence operons required
for invasion and intracellular survival in macrophages (Gao et al 2006) Instead ADS47 has a
larger number and variety of stress-responsive genes such as efflux genes and bacteriocins
(pyocin hemolysin) genes that provide protection against heavy metals and antibiotics and
increase their competitiveness against other bacteria These findings suggest that the strain
ADS47 is not an opportunistic animal pathogen but has evolved to be highly competitive in
the soil micro-environment (Arnold 1998 Bengoechea and Skurnik 2000) Perhaps ADS47
evolved trichothecene degrading genes for adaptation to soils containing trichothecene
mycotoxins contaminated soils (Gadd and Griffiths 1978 Sharma and Fulekar 2009 Han et
al 2011) Conversely the absence or reduced number of stress-responsive genes in C
rodentium and C koseri may imply that those two pathogenic species are not exposed to such
stressful and xenobiotic-contaminated environments as ADS47
This interpretation is supported by the observation that the two of the three
animalhuman pathogenic species contained high of rRNA copy number but lower numbers
were found in ADS47 C freundii Ballerup and Citrobacter sp 30_2 Bacteria maintain large
number of rRNA for higher production of ribosome during high growth rates (Stevenson et al
1997 Klappenbach et al 2000) such as might occur during the establishment of an infection
However a large copy number of rRNAs may not be metabolically favourable for bacteria
such as ADS47 that evolved in a highly competitive environment as that costs higher cellular
energy (Hsu et al 1994)
165
C freundii Ballerup seems to be an exception in that it is similar to the other animal
pathogens for containing virulent factors (ie LEE-operon genes) but more like the
environmental species for encoding high number genes for stress defence heavy metal and
toxic compounds resistances
The comparison of the genes in C freundii ADS47 with the unique ability to detoxify
trichothecenes with genes in other Citrobacter species allowed approximately 270 unique
genes to be identified in ADS47 Some of those genes may be involved in trichothecene de-
epoxidation or de-acetylation Currently the pathways and microbial enzymes leading to
trichothecene de-epoxidation and de-acetylation are unknown but based on the required
reaction ie the elimination of an oxygen atom from epoxide moiety it is likely that epoxide
reductases or epoxide oxidoreductases are responsible for such biodegradation activity (He et
al 1992)
From database searches three enzymes having epoxide reductase activity were found
These are a vitamin K epoxide reductase (VKOR) a monooxygenase PaaABCE and pyridine
nucleotide-disulfide oxidoreductase (PNDO) which reduce vitamin K epoxide phenylacetyl-
CoA and epoxyalkanes respectively (Swaving et al 1996 Li et al 2004 Teufel et al 2012)
In mammals VKOR plays important roles in vitamin K cycle by reducing vitamin K epoxide
to vitamin K and its homologues that are found in plants bacteria and archaea (Goodstadt
2004 Li et al 2010) The microbial multi-component PaaABCE monooxygenase catalyzes
degradation of variety of aromatic compounds or environmental contaminants via epoxidation
of phenylacetic acid (Paa) and NADPH-dependant de-epoxidation of epoxy-Paa (Luengo et
al 2001 Teufel et al 2012) PNDO is a unique type of NADP-dependent oxidoreductase
identified in Xanthobacter strain Py2 that was obtained by propane enrichment procedure
166
(van Ginkel and de Bont 1986) However no orthologs of these genes were found in ADS47
genome
Nine ADS47-unique reductaseoxidoreductase-like genes are potential candidates for
the trichothecene de-epoxidation enzyme Genes belong to short chain
dehydrogenasereductase (SDR) family are structurally and functionally diverse and cause
different enzymatic activities including xenobiotic degradation (Kavanagh et al 2008) This is
a promising candidate for the trichothecene de-epoxidation gene Genes belong to dehydratase
family catalyze reduction reactions by cleaving carbon-oxygen bonds of organic compounds
that result in removal of oxygen in various biosynthetic pathways (Flint et al 1993) This
mechanism may be relevant to microbial trichothecene de-epoxidation reaction since the latter
involves removal of the oxygen atom forming the epoxide in trichothecene mycotoxins
(Young et al 2007)
Another ADS47-unique gene was for a nitrate reductase that may be a candidate de-
epoxidation gene Under stress conditions such an enzyme causes reductive removal of an
oxygen molecule (Morozkina and Zvyagilskaya 2007)
A fourth type of ADS47-unique gene is for a thiol-disulfide isomerasethioredoxin like
(Trx) oxidoreductase The thiol-disulfide isomerase and thioredoxin oxidoreductases contain a
thioredoxin like domain with a CXXC catalytic motif which catalyzes various biological
processes that allow the organism to adapt to environmental stress and develop competence to
sporulate (Scharf et al 1998 Petersohn et al 2001 Smits et al 2005) The membrane-
anchored Trxndashlike domain initiates the vitamin K de-epoxidation reaction by reducing the
active motif (CXXC) of VKOR (Schulman et al 2010)
167
The present study did not find any homologues of VKOR in ADS47 genome but we
identified two copies of structurally similar quinone reductase protein disulfide bond
formation (DsbB) gene (CMD_1044 and CMD_2641) Both VKOR and DsbB contain four
transmembrane segments an active site CXXC motif and cofactor quinone and use a
common electron transfer pathway for reducing substrates (Li et al 2010 Malojcic et al
2008) As VKOR bacterial DsbB requires a Trx-like partner (eg DsbA) for activity Further
experiments are required to examine if DON de-epoxidation occurs by the catalytic activity of
DsbB coupled with a unique thiol-disulfide isomerase and thioredoxin oxidoreductase present
in ADS47 genome
The analysis also identified an ADS47-unique glycosylphosphatidylinositol (GPI)
biosynthetic operon that is comprised of four genes including an N-
acetylglucosaminylphosphatidylinositol de-acetylase The GPI membrane anchor protein
synthesis is initiated by transferring N-acetylglucosamine from uridine diphosphate N-
acetylglucosamine to phosphatidylinositol by N-acetylglucosaminyltransferase an
endoplasmic reticulum (ER) membrane-bound protein complex (Hong and Kinoshita 2009)
In the second step of the pathway N-acetylglucosamine-phosphatidylinositol is de-acetylated
to produce glucosaminyl-phosphatidylinositol by the catalytic activity of a zinc
metalloenzyme N-acetylglucosaminylphosphatidylinositol de-acetylase which is localized on
the cytoplasmic side of the ER (Nakamura et al 1997 Hong and Kinoshita 2009) It is
possible that the identified unique N-acetylglucosaminylphosphatidylinositol de-acetylase
gene of ADS47 may be involved in trichothecene de-acetylation activity
168
Further analysis is needed to determine which of these candidate gene or genes may be
involved in tricthothecene mycotoxin de-epoxidation and de-acetylation activities
Identification of gene responsible for the trichothecene detoxification activities of C freundii
ADS47 opens many potential applications for its use in agri-based industries to contribute to
cropanimal production and food safety (Karlovsky 2011)
56 Acknowledgement
We gratefully acknowledge Agriculture and Agri-Food CanadaBinational
Agricultural Research and Development Fund (AAFCBARD) project for providing the
financial supports of the work
169
Supplemental Table S5-1 List of unique genes of strain ADS47 gene IDs and their
functional roles The unique genes were determined by comparing the ADS47 genome with
five sequenced Citrobacter species (C freundii Citrobacter sp C youngae C koseri and C
rodentium) using OrthoMCL (Li et al 2003)
Unique gene
classes
Number Gene ID and functions
Total unique
genes
270 123 known functions 147 hypothetical proteins
Reductases
oxidoreductases
9 CMD_0711 coenzyme F420 hydrogenasedehydrogenase beta subunit
CMD_0979 Short chain dehydrogenase reductase (SDR) Oxidoreductase
SDR dehydrogenase CMD_1024 NADH-quinone oxidoreductase
membrane L CMD_1393 oxidoreductase SDR putative deoxygluconate
dehydrogenase (NAD+ 3-oxidoreductase) CMD_2679
Dehydratasedihydroxy acid dehydratase CMD_2683 oxidoreductase
oxopropionate reductase CMD_2684 short-chain dehydrogenasereductase
SDR CMD_3184 nitrate reductase alfa subunit CMD_3721 thiol-disulfide
isomerase and thioredoxin oxidoreductase
Deacetylase 1 CMD_2431 N-acetylglucosaminylphosphatidylinositol deacetylase
superfamily
Hydrolase 2 CMD_0991 Dienelactone hydrolase CMD_2895 Putative glycosyl
hydrolase
Transferase 4 CMD_1640 D12 class N6 adenine-specific DNA methyltransferase
CMD_2432 Glycosyltransferase CMD_2433 putative acetyltransferase
CMD_3023 adenine-specific DNA modification methyltransferase
Sulfatase 3 CMD_2898 Possible sulfatase arylsulfatasearylsulfatransferase
CMD_2902 Arylsulfatasesulfatase CMD_2905 Putative
sulfatasearylsulfatase
Bacteriocin 3 CMD_0982 Thermosatble hemolysin superfamily CMD_0993 Pyocin S2
superfamily (bacteriocin that kills other bacterial strain) CMD_3705 S-type
pyocin containing domain protein
Type VI secretion 2 CMD_1802 EvpB family type VI secretion protein CMD_2364 type VI
effector Hcp1
Kinase 4 CMD_0989 Heavy metal Sensor kinase CMD_1794 Polo kinase kinase
(PKK) superfamily CMD_3124 predicted diacylglycerol kinase CMD_4266
putative signal transduction kinase
Transporter 11 CMD_0973 TrkA-N domain-containing protein (K uptake transporter)
CMD_1407 Branched-chain amino acid transport system carrier protein
CMD_2678 Sodium Symporter CMD_3176 glutamine ABC transporter
CMD_3544 Ribose ABC transport system CMD_3693 TRAP-type C4-
dicarboxylate transport system binding protein CMD_3694 TRAP-type C4-
dicarboxylate transport system binding protein small permease component
(predicted N-acetylneuraminate transporter) CMD_3695 TRAP-type C4-
dicarboxylate transport system large permease CMD_4337 23-diketo-L-
gulonate TRAP transporter small permease protein yiaM component
CMD_4338 23-diketo-L-gulonate TRAP transporter large permease protein
yiaN CMD_4600 putative transporter involved in Zinc uptake (YciA
protein)
170
Supplmental Table S5-1 (continued)
Unique gene
classes
Number Gene ID and functions
Membrane
exported protein
8 CMD_0713 putative membrane protein wzy CMD_0983 Putative exported
protein precursor CMD_0986 Putative transmembrane protein
CMD_0987 putative exported protein precursor CMD_1458 putative
exported protein CMD_2903 Putative exported protein CMD_2904
putative secreted 5-nucleotidase CMD_4291 (S) putative integral
membrane export protein
Regulator 12 CMD_0977 probable transcriptional regulator protein CMD_0980 Putative
transcription activator TenA family CMD_0990 Two component heavy
metal response transcription regulator CMD_1766 Leucine responsive
regulatory protein CMD_2677 transcriptional regulator GntR CMD_2909
putative arylsulfatase regulator CMD_3279 Uncharacterized outer
membrane usher protein yqiG precursor CMD_3464 toxin transcription
regulatory protein CMD_3465 EAL regulator protein (signaling)
CMD_3475 putative transcription regulator CMD_3477 Fumarate and
nitrate reduction regulatory protein (for anaerobic adaptation) CMD_3812
putative regulator from phage
Integrase
recombinase
5 CMD_0785 CMD_0992 CMD_0994 CMD_4032 CMD_4424
Phage prophage
transposae related
genes
27 Phages CMD_1608 CMD_1609 CMD_1610 CMD_1611 CMD_1619
CMD_1631 CMD_1913 CMD_1914 CMD_1915 CMD_1917
CMD_1920 CMD_1921 CMD_1936 CMD_3351 CMD_3804
CMD_3805 CMD_3807 CMD_3808 CMD_4581 CMD_4609
CMD_4610 Transposae CMD_0252 CMD_0398 Prophage CMD_1645
CMD_3806 CMD_4425 CMD_4597
Miscellaneous 32 CMD_0095 ATP-dependent DNA helicase Rep CMD_0656 LysA protein
CMD_0710 Polysaccharide biosynthesis family protein CMD_0978 TPR
repeat containing protein CMD_0981 AMP dependant synthatase and
ligase CMD_0985 Cupin superfamily CMD_0987 flavodoxin
CMD_1404 Putative endoribonuclease CMD_1405 Putative PAC domain
protein CMD_1406 Phenazine (PhzF) biosynthesis protein CMD_1695
Putative fimbrial protein CMD_1793 HPChromosome segregation ATPase
domain protein CMD_1922 FRG uncharacterized protein domain
CMD_2206 L-fucose isomerase like protein CMD_2434 CS12 fimbria
outer membrane usher protein CMD_2680 HpcHHpaI aldosecitrate lyase
protein family CMD_2681 Aldose -1 epimerase CMD_2682
Gluconolaconase LRE like region CMD_2900 Putative cytoplasmic
protein CMD_3009 putative restriction endonuclease CMD_3277 Putative
yqi fimbrial adhesion CMD_3278 Hypothetical fimbrial chaperone yqiH
CMD_3340 Pea-VEAacid family CMD_3518 3-hydroxy-3-methylglutaryl
CoA synthase CMD_3801 putative reverse transcriptase CMD_3840 beta-
D-galactosidase alpha subunit CMD_4033 ParB family chromosome
partitioning protein CMD_4034 ParBrepB chromosomeplasmid
partitioning protein CMD_4035 RelASpoT protein CMD_4532 putative
DNA ligase -like protein CMD_4541 Antitermination protein Q and
CMD_4585 putative ATP dependent helicase
171
CHAPTER 6
60 CONCLUSION
61 Summary
Contamination of grain with mycotoxins including deoxynivalenol (DON) and other
trichothecenes is becoming a limiting factor for cereal and animal production (Binder et al
2007) To address the issue this study obtained a novel microbial source for DON
detoxification and determined candidate genes for the function which may have potential
applications to detoxify grain and grain-derived foodfeed contaminated with DON and TC
mycotoxins
From a screen of soil samples from crop field microorganism was isolated that is able
to de-epoxidize the most frequently detected foodfeed contaminating mycotoxin DON The
effects of media compositions temperature pH and aerobicanaerobic conditions on
microbial detoxification activity were assessed From 150 analyzed soil samples a mixed soil
microbial culture that showed DON de-epoxidation activity under aerobic conditions and
moderate temperature (~24degC) was obtained This culture exhibited a slow rate of microbial
DON to de-epoxyDON transformation (de-epoxidation) possibly because of competition
between de-epoxidizing and other microorganisms in the culture
The mixed microbial culture was enriched for DON detoxifying bacteria by reducing
the non-targeted microbial species through antibiotic treatments heating and subculturing on
agar medium and selecting colonies The enriched culture had higher DON de-epoxidation
activity than the initial culture Because of low abundance of DON detoxifying
microorganisms further culture enrichments were required to increase the target microbial
172
population Reculture of the partially enriched microbial community in nutritonally poor MSB
medium with high level of DON increased the DON de-epoxidizing bacterial population
Serial dilution plating and selecting single colonies eventually led to isolation of a DON-
detoxifying bacterial strain named ADS47
Phenotypic and 16S-rDNA sequence analyses provided evidence that the bacterium is
a strain of C freundii Strain ADS47 is a gram negative rod-shaped facultative anaerobe and
has peritrichous flagella Strain ADS47 was unable to use DON as sole carbon source
however increasing DON concentrations in growth media enhanced its DON metabolism
efficiency The pure bacterial culture was shown to be able to rapidly de-epoxidize DON
under aerobic conditions moderate temperatures and neutral pH Strain ADS47 more quickly
de-epoxidizes DON in aerobic conditions than the anaerobic conditions In addition to DON
strain ADS47 showed capability to de-epoxidize andor de-acetylate 10 different type-A and
type-B TC mycotoxins which often co-contaminant grain with DON
To clone the DON detoxifying genes a DON detoxifying bacterial DNA library in E
coli host was constructed followed by screening of the library clones for DON detoxification
activity The study produced a library giving approximately 10 fold coverage of the genome
of strain ADS47 Restriction digestion analysis determined that most of the clones contained
ADS47 DNA inserts This indicates that a good DNA library was produced which had gt99
probability of containing all the DNA of the ADS47 genome However activity-based
screening of the library could not identify any clones with DON de-epoxidation ability It is
assumed that bacterial DON de-epoxidation genes evolved in a certain environmental niche
which may be required for the DON de-epoxidation to be expressed and active but was not
supplied by the host E coli cells
173
An attempt was made to identify the bacterial genes encoding DON de-epoxidation
and de-acetylation enzymes by sequencing the genome of strain ADS47 and comparing its
sequence to the genomes of related bacteria that are not known to posses DON de-epoxidation
activity A RAST server based bioinformatics analysis determined that the 48 Mb circular
genome of ADS47 codes for 4610 predicted CDS A genome wide phylogenetic analysis
determined that strain ADS47 was gt87 identical to the pathogen C freundii strain Ballerup
The relationship to two other animalhuman pathogenic species C rodentium and C koseri
was more distant This result is consistent with the 16S-rDNA based phylogenetic analysis
It is assumed that the microbial DON detoxifying genes were acquired by horizontal
gene transfer or other means during the adaptation of ADS47 in mycotoxin contaminated soil
If it is correct then this means that those newly acquired genes are not common in other
related microbial species and no other Citrobacter species is known to degrade DON Hence
the study identified 270 unique genes to ASDS47 with a subset of 10 genes for reductases
oxidoreductases and a de-acetylase which were considered to be candidate genes for the
enzymes that have DON detoxifying activity Furthermore a targeted comparative genome
analysis showed that the mycotoxin detoxifying ADS47 genome does not harbour the
clinically important LEE encoded typendashIII secretion system and mycobacterium virulence
genes This suggests that strain ADS47 is not likely to be an animal pathogenic bacterium
unlike C freundii Ballerup which encodes the pathogenic relevance LEE operons
174
62 Research contributions
The research made four major contributions which may aid in reducing the risks of
animals and humans from DONTC exposure Firstly the study identified a novel bacterial
strain ADS47 that is able to detoxify DON and 10 related mycotoxins over a wide range of
temperatures under aerobic and anaerobic conditions The discovery has led to a patent
application for strain ADS47 supporting the novelty and importance of the finding This
bacterial strain could be used to detoxify trichothecene mycotoxins in contaminated animal
feed thus reducing food safety risks
Secondly the study resulted in the development of a method for isolating DON de-
epoxidizing microbes from soil as well as optimizing the conditions for bacterial DONTC
detoxification activity to increase the effectiveness usefulness of the strain
Thirdly the thesis work resulted in availability of the genome sequence of a
trichothecene detoxifying microbe which may ultimately lead to an understanding of the
cellularmolecular mechanisms of DONTC de-epoxidation and de-acetylation
genespathways The genome analysis resulted in identification of several candidate genes for
DONTC de-epoxidation and de-acetylation activities Genes which show de-epoxidation
activity could be used to develop DONTC resistant cereal cultivars Fourthly strain ADS47
lacks major animal pathogenicity genes (in particular the LEE-operons) as well as does not
encodes mycobacterium virulence genes Thus strain ADS47 has a great potential to be
utilized to detoxify cereal grain and may be used as a feed additive for livestockfishery All
together the reported work may have significant applications to produce and supply
DONTC-free food and feed
175
63 Further research
This research provides the basis for developing biological detoxification of DON and
related mycotoxins This goal might be achieved by using DON de-epoxidizing gene to
develop resistant plants enzymatic detoxification of contaminated feed and use of the bacteria
as a feed additive Follow-up studies may reach the above indicated goals Because the
bacterium showed de-epoxidation activity under aerobicanaerobic conditions and under wide
range of temperatures it is likely to have an application for enzymatic detoxification of
contaminated animal feed Therefore additional studies on bacterial efficiency to detoxify
contaminated cereal and grain-derived feed need to be conducted Although strain ADS47
does not encode major animal pathogenicity genes the bacterium must be directly tested on
animals to see if it could negatively affect animal digestive systems If not then the potential
for ADS47 is considerable for its use as a feed additive for pig poultry and fish Animal
feeding trials will also need to be performed to determine if DON detoxification is sufficient
to be cost effective as a feed additive In order to develop DONTC resistant cereal cultivars
the gene responsible to DON degrading activity among the presently identified candidate
genes (reductases oxidoreductases and de-acetylase) will need to be expressed in other
organisms like E coli and a codon modified version adapted for expression in plants like
corn and wheat
176
70 REFERENCES
Abbas HK Mirocha CJ Pawlosky RJ Pusch DJ 1985 Effect of cleaning milling and
baking on deoxynivalenol in wheat Applied and Environmental Microbiology 50482ndash
486
Accensi F Pinton P Callu P Abella-Bourges N Guel WJF Grosjean F Oswald IP 2006
Ingestion of low doses of deoxynivalenol does not affect haematological biochemical or
immune responses of piglets Journal of Animal Science 841935ndash1942
Agrawal D Mahapatra AK 2005 Vertically acquired neonatal Citrobacter brain abscessndash
case report and review of the literature Journal of Clinical Neuroscience 12188ndash190
Alexander NJ 2008 The TRI101 story engineering wheat and barley to resist Fusarium head
blight World Mycotoxin Journal 131-37
Alexander NJ Proctor RH McCormick SP 2009 Genes gene clusters and biosynthesis of
trichothecenes and fumonisins in Fusarium Toxin Reviews 28198ndash215
Ali ML Taylor JH Jie L Sun G William M Kasha KJ Reid LM Pauls KP 2005
Molecular mapping of QTLs for resistance to Gibberella ear rot in corn caused by
Fusarium graminearum Genome 48521-533
Alikhan N-F Petty NK Zakour NLB Beatson SA 2011 BLAST Ring Image Generator
(BRIG) simple prokaryote genome comparisons BMC Genomics 12402
Alisi A Spaziani A Anticoli S Ghidinelli M Balsano C 2008 PKR is a novel functional
direct player that coordinates skeletal muscle differentiation via p38MAPKAKT
pathways Cellular Signalling 20534ndash542
177
Amuzie CJ Shinozuka J Pestka JJ 2009 Induction of suppressors of cytokine signaling by
the trichothecene deoxynivalenol in the mouse Toxicological Science 111277ndash287
Ansorge WJ 2009 Next generation DNA sequencing techniques New Biotechnology
25195-203
Arnold JW Senter SD 1998 Use of digital aroma technology and SPME GC-MS to compare
volatile compounds produced by bacteria isolated from processed poultry Journal of the
Science of Food and Agriculture 78343ndash348
Audenaert K Van Broeck R Bekaert B De Witte F Heremans B Messens K Hofte
M Haesaert G 2009 Fusarium head blight (FHB) in Flanders population
diversity inter-species associations and DON contamination in commercial winter
wheat varieties European Journal of Plant Pathology 125445-458
Awad WA Ghareeb K Bohm J Zentek J 2010 Decontamination and detoxification
strategies for the Fusarium mycotoxin deoxynivalenol in animal feed and the
effectiveness of microbial biodegradation Food Additives and Contaminants 27510-520
Aziz K Bartels D Best AA DeJongh M Disz T Edwards RA Formsma K Gerdes S Glass
EM Kubal M Meyer F Olsen GJ Olson R Osterman AL Overbeek RA McNeil LK
Paarmann D Paczian T Parrello1 B Pusch GD Reich C Stevens R Vassieva O
Vonstein V Wilke A Zagnitko O 2008 The RAST Server Rapid Annotations using
Subsystems Technology BMC Genomics 975
Badger JL Stins MF Kim KS 1999 Citrobacter freundii invades and replicates in human
brain microvascular endothelial cells Infection and Immunology 674208ndash4215
Bae HK Pestka JJ 2008 Deoxynivalenol induces p38 interaction with the ribosome in
monocytes and macrophages Toxicological Science 10559-66
178
Bai GH Desjardins AE Plattner RD 2002 Deoxynivalenol-nonproducing Fusarium
graminearum causes initial infection but does not cause disease spread in wheat spikes
Mycopathologia 15391ndash98
Barrett JR 2000 Mycotoxins of molds and maladies Environmental Health Perspectives
108A20ndashA23
Bengoechea JA Skurnik M 2000 Temperature-regulated efflux pumppotassium antiporter
system mediates resistance to cationic antimicrobial peptides in Yersinia Molecular
Microbiology 3767ndash80
Bennett JW Klich M 2003 Mycotoxins Clinical Microbiology Reviews 16497ndash516
Benson DA Karsch-Mizrachi I Lipman DJ Ostell J Sayers EW 2009 GenBank Nucleic
Acids Research 37D26-31
Berdichevsky T Friedberg D Nadler C Rokney A Oppenheim A Rosenshine I 2005 Ler is
a negative autoregulator of the LEE1 operon in neuropathogenic Escherichia coli Journal
of Bacteriology 187349ndash357
Bernardo A Bai GH Guo PG Xiao K Guenzi AC Ayoubi P 2007 Fusarium graminearum-
induced changes in gene expression between Fusarium head blight-resistant and
susceptible wheat cultivars Functional and Integrative Genomics 769ndash77
Bhat RV Beedu SR Ramakrishna Y Munshi KL 1989 Outbreak of trichothecene
mycotoxicosis associated with consumption of mould-damaged wheat production in
Kashmir Valley India The Lancet 33335ndash37
Binder EM Tan LM Chin LJ Handl J Richard J 2007 Worldwide occurrence of
mycotoxins in commodities feeds and feed ingredients Animal Feed Science and
Technology 137265-282
179
Birzele B Meier A Hindorf H Kraumlmer J Dehne H-W 2002 Epidemiology of Fusarium
infection and deoxynivalenol content in winter wheat in the Rhineland Germany
European Journal of Plant Pathology 108667ndash673
Boddu J Cho S Muehlbauer GJ 2007 Transcriptome analysis of trichothecene-induced gene
expression in barley Molecular PlantndashMicrobe Interactions 201364ndash1375
Borenshtein D Schauer DB 2006 The Genus Citrobacter Prokaryotes 690ndash98
Boutigny A-L Richard-Forget F Barreau C 2008 Natural mechanisms for cereal resistance
to the accumulation of Fusarium trichothecenes European Journal of Plant Pathology
121411ndash423
Boutrif E Canet C 1998 Mycotoxin prevention and control FAO programmes Revue de
Meacutedecine Veacuteterinaire 149681-694
Brennan JM Leonard G Fagan B Cooke BM Ritieni A Ferracane R et al 2007
Comparison of commercial European wheat cultivars to Fusarium infection of head and
seedling tissue Plant Pathology 5655ndash64
Brenner DJ Grimont PA Steigerwalt AG Fanning GR Ageron E Riddle CF 1993
Classification of Citrobacteria by DNA hybridization designation of Citrobacter farmeri
sp nov Citrobacter youngae sp nov Citrobacter braakii sp nov Citrobacter
werkmanii sp nov Citrobacter sedlakii sp nov and three unnamed Citrobacter
genomospecies International Journal of Systematic Bacteriology 43645-58
Brookes G Barfoot P 2006 GM Crops The first ten years-global socio-economic and
environmental Impacts no 36 ISAAA (International Service for the Acquisition of Agri-
Biotech Applications) Ithaca New York
180
Bruce KD Hughes MR 2000 Terminal restriction fragment length polymorphism monitoring
of genes amplified directly from bacterial communities in soils and sediments Molecular
Biotechnology 16261ndash269
Buerstmayr H Ban T Anderson JA 2009 QTL mapping and marker-assisted selection for
Fusarium head blight resistance in wheat A review Plant Breeding 1281ndash26
Campbell H Choo TM Vigier B Underhill L 2002 Comparison of mycotoxin profiles
among cereal samples from eastern Canada Canadian Journal of Botany 80526ndash532
Carver et al 2005 ACT the Artemis Comparison Tool Bioinformatics 213422-3423
Carver et al 2009 DNAPlotter circular and linear interactive genome visualization
Bioinformatics 25119-120
Chen CY et al 2003 Comparative genome analysis of Vibrio vulnificus a marine pathogen
Genome Research 132577ndash2587
Chiu TC Yen JH Hsieh YN Wang YS 2005 Reductive transformation of dieldrin under
anaerobic sediment culture Chemosphere 601182ndash1189
Chung YJ Zhou HR Pestka JJ 2003 Transcriptional and posttranscriptional roles for p38
mitogen-activated protein kinase in upregulation of TNF-alpha expression by
deoxynivalenol (vomitoxin) Toxicology and Applied Pharmacology 193188ndash201
Chung EJ Lim HK Kim J-C Choi GJ Park EJ Lee MH Chung YR Lee S-W 2008 Forest
soil metagenome gene cluster involved in antifungal activity expression in Escherichia
coli Applied and Environmental Microbiology 74723-730
Collins TFX Sprando RL Black TN Olejnik N Eppley RM Hines FA Rorie J Ruggles DI
2006 Effects of deoxynivalenol (DON vomitoxin) on in utero development in rats Food
and Chemical Toxicology 44747ndash757
181
Cong Y Wang J Chen Z Xiong K Xu Q Hu F 2011 Characterization of swarming motility
in Citrobacter freundii FEMS Microbiology Letters 371160-171
Cossette F Miller JD 1995 Phytotoxic effect of deoxynivalenol and Gibberella ear rot
resistance of corn Natural Toxins 3383ndash388
Creppy EE 2002 Update of survey regulation and toxic effects of mycotoxins in Europe
Toxicological Letters 12719-28
Dai JS Wang D Guerlebeck C Laturnus S Guenther Z Lu SC Ewers C 2010 Suppression
subtractive hybridization identifies an autotransporter adhesin gene of E coli IMT5155
specifically associated with avian pathogenic Escherichia coli (APEC) BMC
Microbiology 10236
Daniel R 2005 The metagenomics of soil Nature Review Microbiology 3470-478
Davis KER Joseph SJ Janssen PH 2005 Effects of growth medium inoculum size and
incubation time on culturability and isolation of soil bacteria Applied and Environmental
Microbiology 71826-834
Delcher AL Phillippy A Carlton J Salzberg SL 2002 MUMmer 21 NUCmer and
PROmer are described in Fast Algorithms for Large-scale Genome Alignment and
Comparisonrdquo Nucleic Acids Research 302478-2483
Demain AL Adrio JL 2008 Contributions of microorganisms to industrial biology
Molecular Biotechnology 3841ndash455
Deng W Li Y Vallance BA Finlay BB 2001 Locus of enterocyte efffacement from
Citrobacter rodentium sequence analysis and evidence for horizontal transfer among
attaching and effacing pathogens Infection and Immunity 696323ndash6335
182
Desjardins AE Proctor RH Bai G McCormick SP Shaner G Beuchley G Hohn TM 1996
Reduced virulence of trichothecene-nonproducing mutants of Gibberella zeae in wheat
field tests Molecular Plant-Microbe Interactions 9775-781
Desjardins AE 2006 Fusarium mycotoxins chemistry genetics and biology The American
Phytopathological Society St Paul Minnesota USA p 260
Desjardins AE Proctor RH 2007 Molecular biology of Fusarium mycotoxins International
Journal of Food Microbiology 11947-50
Desmond OJ Manners JM Stephens AE MacLean DJ Schenk PM Gardiner DM Munn
AL Kazan K 2008 The Fusarium mycotoxin deoxynivalenol elicits hydrogen peroxide
production programmed cell death and defence responses in wheat Molecular Plant
Pathology 9435-445
Dill-Macky R Jones RK 2000 The effect of previous crop residues and tillage on Fusarium
head blight of wheat Plant Disease 8471ndash76
DrsquoMello JPF Macdonald AMC Dijksma WTP 1998 3-Acetyl deoxynivalenol and esterase
production in a fungicide-insensitive strain of Fusarium culmorum Mycotoxin Research
149ndash18
DrsquoMello JPF Placinta CM Macdonald AMC 1999 Fusarium mycotoxins a review of
global implications of animal health welfare and productivity Animal Feed Science and
Technology 80183-205
Doohan FM Brennan J Cooke BM 2003 Influence of climatic factors on Fusarium species
pathogenic to cereals European Journal of Plant Pathology 109755ndash768
183
Dyer RB Kendra DF Brown DW 2006 Real-time PCR assay to quantify Fusarium
graminearum wild-type and recombinant mutant DNA in plant material Journal of
Microbiological Methods 67534ndash542
Erecinska M Wilson DF 1981 Inhibitors of cytochrome c oxidase In inhibitors of
mitochondrial function edited by M Erecinska and DF Wilson Pergamon Press Ltd
Oxford pp 145-160
Eriksen GS 2003 Metabolism and toxicity of trichothecene Doctoral thesis Swedish
University of Agricultural Sciences Uppsala Sweden
Eriksen GS Pettersson H Lundh T 2004 Comparative cytotoxicity of deoxynivalenol
nivalenol their acetylated derivatives and de-epoxy metabolites Food Chemistry and
Toxicology 42619-624
Eudes F Comeau A Rioux S Collin J 2000 Phytotoxicity of eight mycotoxins associated
with Fusarium in wheat head blight Canadian Journal of Plant Pathology 22286ndash292
FAO (Food and Agriculture Organization) 1991 Food Nutrition and Agriculture Food for
the Future volume 1 FAO Rome
FAO (Food and Agriculture Organization) 2001 FNP73 Manual of the application of
HACCP system in mycotoxin prevention and control FAO Food and Nutrition paper no
73 FAO Rome
Fedorak PM Grbic-Galic D 1991 Aerobic microbial cometabolism of benzothiophene and
3-methylbenzothiophene Applied and Environmental Microbiology 57932-940
Felsenstein J 1989 PHYLIP - Phylogeny Inference Package (Version 32) Cladistics 5164-
166
184
Fernando WGD Amarasinghe C Demoz B Surujdeo‐Maharaj S Gauthier V Guerrieri G
Al‐Taweel K Brucircleacute‐Babel A Tamburic‐Illincic L Gilbert J 2011 What do we know of
the Fusarium graminearum chemotypes their epidemiology genetics and infection
process in Canada In Proc 7th
Canadian Workshop on Fusarium Head Blight edited by
J Gilbert A Tekauz N Sweetland and K Slusarenko Winnipeg Manitoba Canada p 45
Fierer N Jackson RB 2006 The diversity and biogeography of soil bacterial community
Proceedings of the National Academy of Sciences 103626-631
Fink-Gremmels J 1989 The significance of mycotoxin assimilation for meat animals
Deutsche Tierarztliche Wochenschrift 96360-363
Flint DH Emptage MH Finnegan MG Fu W Johnson MK 1993 The role and properties of
the iron-sulfur cluster in Escherichia coli dihydroxy-acid dehydratase Journal of
Biological Chemistry 26814732ndash14742
Foroud NA Eudes F 2009 Trichothecenes in cereal grains International Journal of
Molecular Science 10147-173
Forsell JH Jensen R Tai JH Witt M Lin WS Pestka JJ 1987 Comparison of acute
toxicities of deoxynivalenol (vomitoxin) and 15-acetyldeoxynivalenol in the B6C3F1
mouse Food and Chemical Toxicology 25155ndash162
Frey M Stettner C Gierl A 1998 A general method for gene isolation in tagging approaches
amplification of insertion mutagenised sites (AIMS) The Plant Journal 13717-721
Freeman GG Morrison RI 1949 The isolation and chemical properties of trichothecin an
antifungal substance from Trichothecium roseum Link Biochemical Journal 441ndash5
185
Fuchs E Binder EM Heidler D Krska R 2002 Structural characterization of metabolites
after the microbial degradation of type A trichothecenes by the bacterial strain BBSH 797
Food Additives and Contaminants 19379ndash386
Gabor EM Alkema WBL Janssen DB 2004 Quantifying the accessibility of the
metagenome by random expression cloning techniques Environmental Microbiology
6879ndash886
Gadd GM Griffiths AJ 1978 Microorganisms and heavy metal toxicity Microbial Ecology
4303ndash317
Gale LR Bryant JD Calvo S Giese H 2005 Chromosome complement of the fungal plant
pathogen Fusarium graminearum based on genetic and physical mapping and cytological
observations Genetics 171985ndash1001
Gao LY Pak M Kish R Kajihara K Brown EJ 2006 A mycobacterial operon essential for
virulence in vivo and invasion and intracellular persistence in macrophages Infection and
Immunity 741757ndash1767
Gareau MG Wine E Reardon C et al 2010 Probiotics prevent death caused by Citrobacter
rodentium infection in neonatal mice Journal of Infectious Diseases 20181e91
Gareis M Bauer J Thiem J Plank G Grabley S Gedek B 1990 Cleavage of
zearalenoneglycoside a masked mycotoxin during digestion in swine Journal of
Veterinary Medicine 37236-240
Geng L Xin W Haung DW Feng G 2006 A universal cloning vector using vaccinia
topoisomerase I Molecular Biotechnology 3323-28
186
Golkari S Gilbert J Ban T Procunier JD 2009 QTLspecific microarray gene expression
analysis of wheat resistance to Fusarium head blight in Sumai-3 and two susceptible
NILs Genome 52409ndash418
Goodstadt L Ponting CP 2004 Vitamin K epoxide reductase homology active site and
catalytic mechanism Trends in Biochemical Sciences 29289-292
Goswami RS Kistler HC 2004 Heading for disaster Fusarium graminearum on cereal
crops Molecular Plant Pathology 5515-525
Gouze ME Laffitte J Printon P Dedieux G Galinier A Thouvenot JP Loiseau N Oswald
IP Galtier P 2007 Effect of subacute oral doses of nivalenol on immune and metabolic
defence systems in mice Veterinary Research 38635ndash646
Guan S He J Young JC Zhu H Li X Ji C Zhou T 2009 Transformation of trichothecene
mycotoxins by microorganisms from fish digesta Aquaculture 290290ndash295
Gupta R Beg QK Lorenz P 2002 Bacterial alkaline proteases molecular approaches and
industrial applications Applied Microbiology and Biotechnology 5915ndash32
Haberle J Schweizer G Schondelmaier J Zimmermann G Hart L 2009 Mapping of QTL
for resistance against Fusarium head blight in the winter wheat population
PelikanBussardNing8026 Plant Breeding 12827ndash35
Hall BG 2004 Predicting the evolution of antibiotic resistance genes Nature Reviews
Microbiology 2430-435
Han JI et al 2011 Complete genome sequence of the metabolically versatile plant growth-
promoting endophyte Variovorax paradoxus S110 Journal of Bacteriology 1931183ndash
1190
187
Harvey RB Edrington TS Kubena LF Elissalde MH Corrier DE Rottinghaus GE 1995
Effect of aflatoxin and diacetoxyscirpenol in ewe lambs Bulletin of Environmental
Contamination and Toxicology 54325-330
Harvey RB Edrington TS Kubena LF Elissalde MH Casper HH Rottinghaus GE Turk JR
1996 Effects of dietary fumonisin B1-containing culture material deoxynivalenol-
contaminated wheat or their combination on growing barrows American Journal of
Veterinary Research 571790-1794
He J 2007 Microbial strategies for management of Fusarium graminearum and
transformation of deoxynivalenol Master of Science Thesis University of Guelph
Guelph Ontario Canada
He K Vines L Pestka JJ 2010a Deoxynivalenol-induced modulation of microRNA
expression in RAW 2647 macrophages-A potential novel mechanism for translational
inhibition Toxicologist 114310
He J Zhou T Young JC Boland GJ Scott PM 2010b Chemical and biological
transformations for trichothecene mycotoxins in human and animal food chains A review
Trends in Food Science and Technology 2167-76
He J Zhou T 2010 Patented techniques for detoxification of mycotoxins in feeds and food
matrices Rec Pat Food Nutr Agric 296ndash104
He P Young LG Forsberg C 1992 Microbial transformation of deoxynivalenol (vomitoxin)
Applied and Environmental Microbiology 583857-3863
Hell K Cardwell KF Setamou M Poehling H-M 2000 The influence of storage practices on
aflatoxins contamination in maize in four agroecological zones in Benin West Africa
Journal of Stored Product Research 36365ndash82
188
Henne A Schmitz RA Bomeke M Gottschalk G Daniel R 2000 Screening of
environmental DNA libraries for the presence of genes conferring lipolytic activity on
Escherichia coli Applied and Environmental Microbiology 663113ndash3116
Hodges GR Degener CE Barnes WG 1978 Clinical significance of Citrobacter isolates
American Journal of Clinical Pathology 7037ndash40
Hollinger K Ekperigin HE 1999 Mycotoxicosis in food producing animals The Veterinary
Clinics of North America Food Animal Practice 15133-165
Hong Y Kinoshita T 2009 Trypanosome glycosylphosphatidylinositol biosynthesis Korean
Journal of Parasitology 47197ndash204
Hope RJ Aldred D Magan N 2005 Comparison of environmental profiles for growth and
deoxynivalenol production by Fusarium culmorum and F graminearum on wheat grain
Letters in Applied Microbiology 40295ndash300
Horvath RS 1972 Microbial co-metabolism and the degradation of organic compounds in
nature Bacteriological Reviews 36146ndash155
Hsu D Shih LM Zee YC 1994 Degradation of rRNA in Salmonella strains a novel
mechanism to regulate the concentrations of rRNA and ribosomes Journal of
Bacteriology 1764761ndash4765
Huwig A Freimund S Kappeli O Dutler H 2001 Mycotoxin detoxification of animal feed
by different adsorbents Toxicological Letters 122179ndash188
Iordanov MS Pribnow D Magun JL Dinh TH Pearson JA Magun BE 1997 Ribotoxic
stress response activation of the stress activated protein kinase JNK1 by inhibitors of the
peptidyl transferase reaction and by sequence-specific RNA damage to the sarcinricin
loop in the 28S rRNA Molecular and Cellular Biology 173373ndash3381
189
Islam R Zhou T Young JC Goodwin PH Pauls KP 2012 Aerobic and anaerobic de-
epoxidation of mycotoxin deoxynivalenol by bacteria originating from agricultural soil
World Journal of Microbiology and Biotechnology 287-13
Jackson LS Bullerman LB 1999 Effect of processing on Fusarium mycotoxins Advances in
Experimental Medicine and Biology 459243-261
Jansen C von Wettstein D Schafer W Kogel K-H Felk A Maier FJ 2005 Infection patterns
in barley and wheat spikes inoculated with wild-type and trichodiene synthase gene
disrupted Fusarium graminearum Proceedings of the National Academy of Sciences
10216892ndash16897
Jube S Borthakur D 2007 Expression of bacterial genes in transgenic tobacco methods
applications and future prospects Electronic Journal of Biotechnology 10452-467
Kabak B Dobson AD 2009 Biological strategies to counteract the effects of mycotoxins
Journal of Food Protection 722006-2016
Kant P Reinprecht Y Martin CJ Islam R Pauls KP 2011 Integration of biotechnologies
disease resistance pathology and Fusarium In Comprehensive Biotechnology edited by
Moo-Young M Second Edition 4729-743
Kaplan CW Kitts CL 2004 Bacterial succession in a petroleum land treatment unit Applied
and Environmental Microbiology 701777-1786
Kaprelyants AS Kell DB 1996 Do bacteria need to communicate with each other for
growth Trends in Microbiology 4237-42
190
Kariholu U Rawal J Namnyak S 2009 Neonatal Citrobacter koseri meningitis and brain
abscess The Internet Journal of Pediatrics and Neonatology 10
(httpwwwispubcomjournalthe-internet-journal-of-pediatrics-and-
neonatologyvolume-10-number-1neonatal-citrobacter-koseri-meningitis-and-brain-
abscesshtml) (accessed on April 19 2012)
Karlovsky P 1999 Biological detoxification of fungal toxins and its use in plant breeding
feed and food production Natural Toxins 71ndash23
Karlovsky P 2011 Biological detoxification of the mycotoxin deoxynivalenol and its use in
genetically engineered crops and feed additives Applied Microbiology and
Biotechnology 91491-504
Kavanagh KL Jornvall H Persson B Oppermann U 2008 Medium and short-chain
dehydrogenasereductase gene and protein families the SDR superfamily functional and
structural diversity within a family of metabolic and regulatory enzymes Cellular and
Molecular Life Sciences 653895ndash3906
Khera KS Whalen C Angers G Vesonder RF Kuiper-Goodman T 1982 Embryotoxicity of
4-deoxynivalenol (vomitoxin) in mice Bulletin of Environmental Contamination and
Toxicology 29487ndash491
Kikot GE Hours RA Alconada TM 2009 Contribution of cell wall degrading enzymes to
pathogenesis of Fusarium graminearum a review Journal of Basic Microbiology 49
231ndash241
Kim UJ Shizuya H Jong PJ Birren B Simon MI 1992 Stable propogation of cosmid sized
human DNA inserts in a F factor based vector Nucleic Acids Research 201083-1065
191
Kimura M Kaneko I Komiyama M Takatsuki A Koshino H Yoneyama K Yamaguchi I
1998 Trichothecene 3-O-acetyltransferase protects both the producing organism and
transformed yeast from related mycotoxins cloning and characterization of Tri101
Journal of Biological Chemistry 2731654-1661
Kimura M Anzai H Yamaguchi I 2001 Microbial toxins in plantndashpathogen interactions
biosynthesis resistance mechanisms and significance The Journal of General and
Applied Microbiology 47149ndash160
Kimura M Tokai T Takahashi-Ando N Ohsato S Fujimura M 2007 Molecular and genetics
studies of Fusarium trichothecene biosynthesis pathways genes and evolution
Bioscience Biotechnology and Biochemistry 712105-2123
King RR McQueen RE Levesque D Greenhalgh R 1984 Transformation of deoxynivalenol
(vomitoxin) by rumen microorganisms Journal of Agricultural and Food Chemistry
32181ndash1183
Klappenbach JA Dunbar JM Schmidt TM 2000 rRNA operon copy number reflects
ecological strategies of bacteria Applied and Environmental Microbiology 661328ndash
1333
Knietsch A Waschkowitz T Bowien S Henne A Daniel R 2003 Metagenomes of complex
microbial consortia derived from different soils as sources for novel genes conferring
formation of carbonyls from short-chain polyols on Escherichia coli Journal of Molecular
Microbiology and Biotechnology 546ndash56
Kollarczik B Gareis M Hanelt M 1994 In vitro transformation of the Fusarium mycotoxins
deoxynivalenol and zearalenone by the normal gut microflora of pigs Natural Toxins
2105ndash110
192
Koonin EV Wolf YI Aravind L 2001 Prediction of the archaeal exosome and its
connections with the proteasome and the translation and transcription machineries by a
comparative-genomic approach Genome Research 11240ndash252
Kouadio JH Dano SD Moukha S Mobio TA Creppy EE 2007 Effects of combinations of
Fusarium mycotoxins on the inhibition of macromolecular synthesis malondialdehyde
levels DNA methylation and fragmentation and viability in Caco-2 cells Toxicon
49306ndash317
Krause L Diaz NN Bartels D Edwards RA Puumlhler A et al 2006 Finding novel genes in
bacterial communities isolated from the environment Bioinformatics 22281ndash289
Kubena LF Huff WE Harvey RB Phillips TD Rottinghaus GE 1989 Individual and
combined toxicity of deoxynivalenol and T-2 toxin in broiler chicks Poultry Science
68622ndash626
Kubena LF Harvey RB Huff WE Corrier DE Phillips TD Rottinghaus GE 1990 Efficacy
of hydrated sodium calcium aluminosilicate to reduce the toxicity of aflatoxin and T-2
toxin Poultry Science 691078ndash1086
Kubena LF Harvey RB Huff WE Elissalde MH Yersin AG Phillips TD Rottinghaus GE
1993 Efficacy of hydrated sodium calcium aluminosilicate to reduce the toxicity of
aflatoxin and diacetoxyscirpenol Poultry Science 7251ndash59
Kubena LF Edrington TS Harvey RB Buckley SA Phillips TD Rottinghaus GE Casper
HH 1997 Individual and combined effects of fumonisin B1 present in Fusarium
moniliforme culture material and T-2 toxin or deoxynivalenol in broiler chicks Poultry
Science 761239ndash1247
193
Kumar RA Gunasekaran P Lakshmanan M 1999 Biodegradation of tannic acid by
Citrobacter freundii isolated from a tannery effluent Journal of Basic Microbiology
39161-168
Kushiro M 2008 Effects of milling and cooking processes on the deoxynivalenol content in
wheat International Journal Molecular Science 92127-2145
Lane DJ 1991 16S23S rRNA sequencing In Nucleic acid techniques in bacterial
systematic edited by Stackebrandt E and Goodfellow M Wiley J and Sons Inc New
York pp 115-175
Lancova K Hajslova J Poustka J Krplova A Zachariasova M Dostalek P Saschambula L
2008 Transfer of Fusarium mycotoxins and ldquomaskedrdquo deoxynivalenol (deoxynivalenol-3-
glucoside) from field barley through malt to beer Food Additives and Contaminants
25732ndash744
Langmead B Trapnell C Pop M Salzberg SL 2009 Ultrafast and memory-efficient
alignment of short DNA sequences to the human genome Genome Biology 10R25
Larsen JC Hunt J Perrin I Ruckenbauer P 2004 Workshop on trichothecenes with a focus
on DON summary report Toxicological Letters 1531-22
Lee T Oh DW Kim HS Lee J Kim YH Yun SH Lee YW 2001 Identification of
deoxynivalenol- and nivalenol-producing chemotypes of Gibberella zeae by using PCR
Applied and Environmental Microbiology 672966ndash2972
Lee S-W Won K Lim HK Kim J-C Choi GJ Cho KY 2004 Screening for novel lipolytic
enzymes from uncultured soil microorganisms Applied and Environmental Microbiology
65720-726
194
Li L Christian J Stoeckert J David SR 2003 OrthoMCL Identification of Ortholog Groups
for Eukaryotic Genomes Genome Research 132178-2189
Li T Chang CY Jin DY Lin PJ Khvorova A Stafford DW 2004 Identification of the gene
for vitamin K epoxide reductase Nature 427541ndash544
Li Y 2007 Sensory signal transduction in the vagal primary afferent neurons Current
Medicinal Chemistry 142554ndash2563
Li MX Pestka JJ 2008 Comparative induction of 28S ribosomal RNA cleavage by ricin and
the trichothecenes deoxynivalenol and T-2 toxin in the macrophage Toxicological
Science 10567ndash78
Li R Li Y Kristiansen K Wang J 2008 SOAP short oligonucleotide alignment program
Bioinformatics 24713-4
Li W Schulman S Dutton RJ Boyd D Beckwith J Rapoport TA 2010 Structure of a
bacterial homologue of vitamin K epoxide reductase Nature 463507-512
Lichstein HC Malcolm HS 1943 Studies of the effect of sodium azide on microbic growth
and respiration Journal of Bacteriology 47221ndash230
Limay-Rios V Schaafsma AW 2011 2010 Update on the development of an integrated
mycotoxin management system in Ontario corn 30th
Centralia Swine Research Update
Kirkton Ontario Canada
Llorens A Hinojo MJ Mateo R Gonzalez-Jaen MT Valle- Algarra FM Logrieco A Jimenez
M 2006 Characterization of Fusarium spp isolates by PCR-RFLP analysis of the
intergenic spacer region of the rRNA gene (rDNA) International Journal of Food
Microbiology 106287ndash306
195
Lombaert GA Pellaers P Roscoe V Mankotia M Neil R Scott PM 2003 Mycotoxins in
infant cereal foods from the Canadian retail market Food Additives and Contaminants
20494ndash504
Luengo JM Garciacutea JL Olivera ER 2001 The phenylacetyl-CoA catabolon a complex
catabolic unit with broad biotechnological applications Molecular Microbiology
391434ndash1442
Luo X 1994 Food poisoning caused by Fusarium toxins In Proceedings of the Second
Asian Conference on Food Safety ILSI Thailand pp 129ndash136
Mabagala RB 1997 The effect of populations of Xanthomonas campestris pv phaseoli in
bean reproductive tissues on seed infection of resistant and susceptible bean genotypes
European Journal of Plant Pathology 103175ndash181
Madhyastha MS Marquardt RR Abramson D 1994 Structurendashactivity relationships and
interactions among trichothecene mycotoxins as assessed by yeast bioassay Toxicon
321147ndash1152
Maier FJ Miedaner T Hadeler B Felk A Salomon S Lemmens M Kassner H Schaumlfer W
2006 Involvement of trichothecenes in fusarioses of wheat barley and maize evaluated
by gene disruption of the trichodiene synthase (Tri5) gene in three field isolates of
different chemotype and virulence Molecular Plant Pathology 7449ndash461
Malojcic G Owen RL Grimshaw JP Glockshuber R 2008 Preparation and structure of the
charge-transfer intermediate of the transmembrane redox catalyst DsbB FEBS Letters
5823301ndash3307
196
Manoharan M Dahleen LS Hohn TM Neate SM Yu XH Alexander NJ et al 2006
Expression of 3-OH trichothecene acetyltransferase in barley (Hordeum vulgare L) and
effects on deoxynivalenol Plant Science 171699ndash706
Maresca M Mahfoud R Garmy N Fantini J 2002 The mycotoxin deoxynivalenol affects
nutrient absorption in human intestinal epithelial cells The Journal of Nutrition
1322723ndash2731
Martins ML Martins HM 2002 Influence of water activity temperature and incubation time
on the simultaneous production of deoxynivalenol and zearalenone in corn (Zea mays) by
Fusarium graminearum Food Chemistry 79315ndash318
Masuda D Ishida M Yamaguchi K Yamaguchi I Kimura M Nishiuchi T 2007 Phytotoxic
effects of trichothecenes on the growth and morphology of Arabidopsis thaliana Journal
of Experimental Botany 581617ndash1626
Matsumoto E Kawanaka Y Yun SJ Oyaizu H 2008 Isolation of dieldrin- and endrin-
degrading bacteria using 12-epoxycyclohexane as a structural analog of both compounds
Applied Microbiology and Biotechnology 801095-1103
McLean M 1996 The phytotoxicity of Fusarium metabolites an update since 1989
Mycopathologia 133163ndash179
Merhej J Richard-Forget F Barreau C 2011 Regulation of trichothecene biosynthesis in
Fusarium recent advances and new insights Applied Microbiology and Biotechnology
91519ndash528
Midwest Plan Service 1987 Grain Drying Handling Storage Handbook Ames Iowa Iowa
State University 2nd edition
197
Miller SS Reid LM Harris LJ 2007 Colonization of maize silks by Fusarium graminearum
the causative organism of Gibberella ear rot Canadian Journal of Botany 85369-376
Minervini F Fornelli F Flynn KM 2004 Toxicity and apoptosis induced by mycotoxins
nivalenol deoxynivalenol and fumonisin B1 in a human erythroleukemia cell line
Toxicology In Vitro 1821ndash28
Mitterbauer R Poppenberger B RaditschnigA Lucyshyn D Lemmens M Glossl J Adam G
2004 Toxin-dependent utilization of engineered ribosomal protein L3 limits trichothecene
resistance in transgenic plants Plant Biotechnology Journal 2329ndash340
Mocali S Benedetti A 2010 Exploring research frontiers in microbiology the challenge of
metagenomics in soil microbiology Research Microbiology 161497-505
Morozkina EV Zvyagilskaya RA 2007 Nitrate reductases structure functions and effect of
stress factors Biochemistry 721151ndash1160
Munkvold GP 2003 Cultural and genetic approaches to managing mycotoxins in maize
Annual Review of Phytopathology 4199-116
Mundy R MacDonald TT Dougan G Frankel G Wiles S 2005 Citrobacter rodentium of
mice and man Cellular Microbiology 71697ndash1706
Nakamura N Inoue N Watanabe R Takahashi M Takeda J Stevens VL Kinoshita T 1997
Expression cloning of PIG-L a candidate N-acetylglucosaminyl-phosphatidylinositol
deacetylase The Journal of Biological Chemistry 27215834ndash15840
Nganje WE Bangsund DA Leistritz FL Wilson WW Tiapo NM 2002 Estimating the
economic impact of a crop disease the case of Fusarium head blight in US wheat and
barley National Fusarium Head Blight Forum Proceedings East Lansing Michigan State
University pp 275ndash281
198
Nielsen C Lippke H Didier A Dietrich R Martlbauer E 2009 Potential of deoxynivalenol
to induce transcription factors in humanhepatoma cells Molecular Nutrition and Food
Research 53479ndash491
Nielsen JKS Vikstroumlm AC Turner P Knudsen LE 2011 Deoxynivalenol transport across
the human placental barrier Food and Chemical Toxicology 492046-2052
Nishiuchi T Masuda D Nakashita H Ichimura K Shinozaki K Yoshida S et al 2006
Fusarium phytotoxin trichothecenes have an elicitor-like activity in Arabidopsis thaliana
but the activity differed significantly among their molecular species Molecular Plantndash
Microbe Interactions 19512ndash520
Ohsato S Ochiai-Fukuda T Nishiuchi T Takahashi-Ando N Koizumi S Hamamoto H
Kudo T Yamaguchi I Kimura M 2007 Transgenic rice plants expressing trichothecene
3-O-acetyltransferase show resistance to the Fusarium phytotoxin deoxynivalenol Plant
Cell Reports 26531-538
Okubara PA Blechl AE McCormick SP Alexander NJ Dill-Macky R Hohn TM 2002
Engineering deoxynivalenol metabolism in wheat through the expression of a fungal
trichothecene acetyltransferase gene Theoritical and Applied Genetics 10674-83
Ong CL Beatson SA Totsika M Forestier C McEwan AG Schembri MA 2010 Molecular
analysis of type 3 fimbrial genes from Escherichia coli Klebsiella and Citrobacter
species BMC Microbiology 10183
Osborne LE Stein JM 2007 Epidemiology of Fusarium head blight on small-grain cereals
International Journal of Food Microbiology 119103-108
Page RDM 1996 TREEVIEW An application to display phylogenetic trees on personal
computers Computer Applications in the Biosciences 12357-358
199
Parkhill J Dougan G James KD Thomson NR Pickard D Wain J Churcher C Mungall KL
Bentley SD Holden MT et al 2001 Complete genome sequence of a multiple drug
resistant Salmonella enterica serovar Typhi CT18 Nature 413848ndash852
Paterson RRM Lima N 2010 How will climate change affect mycotoxins in food Food
Research International 431902ndash14
Paul PA El-Allaf SM Lipps PE Madden LV 2004 Rain splash dispersal of Gibberella zeae
within wheat canopies in Ohio Phytopathology 941342-1349
Pestka JJ Lin WS Miller ER 1987 Emetic activity of the trichothecene 15-
acetyldeoxynivalenol in pigs Food and Chemical Toxicology 25855ndash858
Pestka JJ Smolinski AT 2005 Deoxynivalenol Toxicology and potential effects on humans
Journal of Toxicology Environmental Health 839ndash69
Pestka JJ 2007 Deoxynivalenol toxicity mechanisms and animal health risks Animal Feed
Science and Technology 137283-298
Pestka JJ 2008 Mechanisms of deoxynivalenol-induced gene expression and apoptosis Food
Additives and Contaminants 251128-1140
Pestka J2010 Toxicological mechanisms and potential health effects of deoxynivalenol and
nivalenol World Mycotoxin Journal 3323ndash347
Petersohn A Brigulla M Haas S Hoheisel JD Voumllker U Hecker M 2001 Global Analysis
of the General Stress Response of Bacillus subtilis Journal of Bacteriology 1835617-
5631
200
Petty NK Bulgin R Crepin VF Cerdeno-Tarraga AM Schroeder GN Quail MA Lennard N
Corton C Barron A Clark L Toribio AL Parkhill J Dougan G Frankel G Thomson NR
2010 The Citrobacter rodentium genome sequence reveals convergent evolution with
human pathogenic Escherichia coli Journal of Bacteriology 192525ndash538
Petty NK Feltwell T Pickard D Clare S Toribio AL et al 2011 Citrobacter rodentium is
an unstable pathogen showing evidence of significant genomic flux PLoS Pathology
7e1002018 doi101371journalppat1002018
Pittet A 2001 Natural occurrence of mycotoxins in foods and feeds A decade in review In
Mycotoxins and phytotoxins in perspective at the turn of the millennium edited by De
Koe WJ Samson RA Van Egmond HP Gilbert J Sabino M W J de Koe Wageningen
Netherlands pp 153ndash172
Placinta CM DrsquoMello JPF Macdonald AMC 1999 A review of worldwide contamination of
cereal grains and animal feed with Fusarium mycotoxins Animal Feed Science and
Technology 7821ndash37
Plank B Schuh M Binder EM 2009 Investigations on the effect of two feed additives
Biomin BBSH 797 and Mycofix Plus (R) 3E as detoxificants of DON contaminated feed
of piglets Wien Tierarztl Monatssch 9655ndash71
Poapolathep A Ohtsuka R Kiatipattanasakul W Ishigami N Nakayama H Doi K 2002
Nivalenol-induced apoptosis in thymus spleen and Peyerrsquos patches of mice Experimental
and Toxicological Pathology 53441ndash446
201
Poppenberger B Berthiller F Lucyshyn D Sieberer T Schuhmacher R Krska R Kuchler K
Gloumlssl J Luschnig C Adam G 2003 Detoxification of the Fusarium mycotoxin
deoxynivalenol by a UDP-glucosyltransferase from Arabidopsis thaliana Journal of
Biological Chemistry 27847905ndash47914
Prelusky DB Trenholm HL 1993 The efficacy of various classes of anti-emetics in
preventing deoxynivalenol-induced vomiting in swine Natural Toxins 1296ndash302
Proctor RH Hohn TM McCormick SP 1995 Reduced virulence of Gibberella zeae caused
by disruption of a trichothecene toxin biosynthetic gene Molecular Plant-Microbe
Interactions 8593-601
Puchenkova SG 1996 Enterobacteria in areas of water along the Crimean Coast
Mikrobiolohichnyĭ zhurnal 58 3-7
Qi BW Hao BL 2004 Whole-proteome prokaryote phylogeny without sequence alignment
a K-string composition approach Journal of Molecular Evolution 581ndash11
Quarta A Mita G Haidukowski M Logrieco A Mule G Visconti A 2006 Multiplex PCR
assay for the identification of nivalenol 3- and 15-acetyl-deoxynivalenol chemotypes in
Fusarium FEMS Microbiological Letters 2597ndash13
Rehr B Klemme JH 1989 Formate dependent nitrate and nitrite reduction to ammonia by
Citrobacter freundii and competition with denitrifying bacteria Antonie van Leeuwenhoek
Journal of Microbiology 56311ndash321
Rocha O Ansari K Doohan FM 2005 Effects of trichothecene mycotoxins on eukaryotic
cells A review Food Additives and Contaminants 22369ndash378
Roscoe V Lombaert GA Huzel V et al 2008 Mycotoxins in breakfast cereals from the
Canadian retail market a 3-year survey Food Additives and Contaminants 25347ndash355
202
Rotter BA Prelusky DB Pestka JJ 1996 Toxicology of deoxynivalenol (vomitoxin) Journal
of Toxicology and Environment Health 481ndash34
Ryu JC Ohtsubo K Izurniyarna N Nakamura K Tanaka T Yamarnura H Ueno Y 1988
The acute and chronic toxicities of nivalenol in mice Fundamental and Applied
Toxicology 1138ndash47
Saier MH Roseman S 1972 Inducer exclusion and repression of enzyme synthesis in
mutants of Salmonella typhimurium defective in enzyme I of the phosphoenolpyruvate
sugar phosphotransferase system The Journal of Biological Chemistry 247972ndash975
Saier MHJ Tran CV Barabote RD 2006 TCDB the transporter classification database for
membrane transport protein analyses and information Nucleic Acids Research 34D181ndash
186
Saitou N Nei M 1987 The neighbor-joining method a new method for reconstructing
phylogenetic trees Molecular Biology and Evolution 4406ndash425
Savelieva O Kotova I Roelofsen W Stams AJM Netrusov A 2004 Utilization of
aminoaromatic acids by a methanogenic enrichment culture and by a novel Citrobacter
freundii strain Archives of Microbiology 181163ndash170
Schaafsma AW Tamburic-Ilinic L Miller JD Hooker DC 2001 Agronomic considerations
for reducing deoxynivalenol in wheat grain Canadian Journal of Plant Pathology 23279ndash
285
Schaafsma AW 2002 Economic changes imposed by mycotoxins in food grains case study
of deoxynivalenol in winter wheat Advances in Experimental Medicine and Biology
504271ndash276
203
Scharf C Riethdorf S Ernst H Engelmann S Voumllker U Hecker M 1998 Theoredoxin is an
essential protein induced by heat shock and other stresses in Bacillus subtilis Journal of
Bacteriology 1801869-1877
Schatzmayr G Zehner F Taubel M Schatzmayr D Klimitsch A Loibner AP Binder EM
2006 Microbiologicals for deactivating mycotoxins Molecular Nutrition and Food
Research 50543-551
Schollenberger M Muumlller H-M Ruumlfle R Suchy S Plank S Drochner W 2006 Natural
occurrence of 16 Fusarium toxins in grains and feedstuffs of plant origin from Germany
Mycopathologia 16143ndash52
Scientific Committee for Food 2002 Opinion on Fusarium toxins Part 6 Group evaluation
of T-2 toxin HT-2 toxin nivalenol and deoxynivalenol (available at http
wwweuropaeuintcommfoodfsscscfout123_enpdf)
Schulman S Wang B Li W Rapoport TA 2010 Vitamin K epoxide reductase prefers ER
membrane-anchored thioredoxin-like redox partners Proceedings of the National
Academy of Sciences 10715027ndash15032
Scott PM 1997 Multi-year monitoring of Canadian grains and grain-based foods for
trichothecenes and zearalenone Food Additives and Contaminants 14333-339
Seitz LM Bechtel DB 1985 Chemical physical and microscopal studies of scab-infected
hard red winter wheat Journal of Agricultural and Food Chemistry 33373-7
Sergent T Parys M Garsou S Pussemier L Schneider YJ Larondelle Y 2006
Deoxynivalenol transport across human intestinal Caco-2 cells and its effects on cellular
metabolism at realistic intestinal concentrations Toxicological Letters 164167ndash176
204
Sharma J Fulekar MH 2009 Potential of Citrobacter freundii for bioaccumulation of
heavy metalndashcopper Biology and Medicine 17-14
Sherif SO Salama EE Abdel-Wahhab MA 2009 Mycotoxins and child health the need for
health risk assessment International Journal of Hygiene and Environmental Health
212347ndash368
Shifrin VI Anderson P 1999 Trichothecene mycotoxins trigger a ribotoxic stress response
that activates c-Jun N-terminal kinase and p38 mitogen-activated protein kinase and
induces apoptosis The Journal of Biological Chemistry 27413985ndash13992
Shima J Takase S Takahashi Y Iwai Y Fujimoto H Yamazaki M Ochi K 1997 Novel
detoxification of the trichothecene mycotoxin deoxynivalenol by a soil bacterium isolated
by enrichment culture Applied and Environmental Microbiology 633825ndash3830
Singh J Behal A Singla N Joshi A Birbian N et al 2009 Metagenomics Concept
methodology ecological inference and recent advances Biotechnology Journal 4480ndash
494
Smith TK McMillan EG Castillo JB 1997 Effect of feeding blends of Fusarium mycotoxin-
contaminated grains containing deoxynivalenol and fusaric acid on growth and feed
consumption of immature swine Journal of Animal Science 752184ndash2191
Smits WK Dubois JYF Bron S van Dijl JM Kuipers OP 2005 Tricksy business
transcriptome analysis reveals the involvement of thioredoxin in redox homeostasis
oxidative stress sulfur metabolism and cellular differentiation in Bacillus subtilis Journal
of Bacteriology 1873921ndash3930
205
Sprando RL Collins TFX Black TN Olejnik N Rorie JI Eppley RM Ruggles DI 2005
Characterization of the effect of deoxynivalenol on selected male reproductive endpoints
Food Chemistry and Toxicology 43623ndash635
Starkey DE Ward TJ Aoki T Gale LR Kistler HC Geiser DM Suga H Toacuteth B Varga J
ODonnell K 2007 Global molecular surveillance reveals novel Fusarium head blight
species and trichothecene toxin diversity Fungal Genetics and Biology 441191ndash1204
Stevenson BS Schmidt TM 1997 Growth rate-dependent expression of RNA from plasmid-
borne rRNA operons in Escherichia coli Journal of Bacteriology 1801970ndash1972
Stothard P Wishart DS 2005 Circular genome visualization and exploration using
CGView Bioinformatics 21537ndash539
Suenaga H Ohnuki T Miyazaki K 2007 Functional screening of a metagenomic library for
genes involved in microbial degradation of aromatic compounds Environmental
Microbiology 92289-97
Surai P Mezes M Fotina TI Denev SD 2010 Mycotoxins in Human Diet A Hidden
Danger In Modern Dietary Fat Intakes in Disease Promotion Edited by F De Meester et
al Nutrition and Health DOI 101007978-1-60327-571-2_18
Sutton JC 1982 Epidemiology of wheat head blight and maize ear rot caused by Fusarium
graminearum Canadian Journal of Plant Pathology 4195
Swaving J De Bont JAM Westphal A De Kok A 1996 A novel type pyridine nucleotide-
disulfide oxidoreductase is essential for NAD+- and NADPH-dependent degradation of
epoxyalkanes by Xanthobacter strain Py2 Journal of Bacteriology 1786644ndash6
206
Swanson SP Rood JHD Behrens JC Sanders PE 1987 Preparation and characterization of
the deepoxy trichothecenes deepoxy HT-2 deepoxy T-2 triol deepoxy T-2 tetraol
deepoxy 15-monoacetoxyscirpenol and deepoxy scirpentriol Applied and Environmental
Microbiology 532821ndash2826
Swanson SP Helaszek C Buck WB Rood JHD Haschek WM 1988 The role of intestinal
microflora in the metabolism of trichothecene mycotoxins Food Chemistry and
Toxicology 26823ndash829
Tian F Yu Y Chen B Li H Yao Y-F Guo X-K 2009 Bacterial archaeal and eukaryotic
diversity in Arctic sediment as revealed by 16S rRNA and 18S rRNA gene clone libraries
analysis Polar Biology 3293-103
Tatusov RL Galperin MY Natale DA Koonin EV et al 2000 Artemis sequence
visualization and annotation Bioinformatics 16944-945
Trail F Haixin X Loranger R Gadoury D 2002 Physiological and environmental aspects of
ascospore discharge in Gibberella zeae (anamorph Fusarium graminearum) Mycologia
94181-189
Tsuge K Ohata Y Shoda M 2001 Gene yerP involved in surfactin self-resistance in
Bacillus subtilis Antimicrobial Agents and Chemotherapy 453566ndash3573
Tucker 2001 The ldquoyellow rainrdquo controversy Lesson for arms control compliance The
Nonproliferation Review 825-42
Ueno Y 1985 The toxicology of mycotoxins Critical Reviews in Toxicology 1499-132
Ueno Y Nakayama K Ishii K Tashiro F Minoda Y Omori T Komagata K 1983
Metabolism of T-2 toxin in Curtobacterium sp strain 114-2 Applied and Environmental
Microbiology 46120ndash127
207
van Elsas JD Speksnijder AJ van Overbeek LS 2008 A procedure for the metagenomics
exploration of disease suppressive soils Journal of Microbiological Methods 75515ndash522
van Ginkel CG de Bont JAM 1986 Isolation and characterization of alkene-utilizing
Xanthobacter spp Archives of Microbiology 145403ndash407
van Lanen SG Shen B 2006 Microbial genomics for the improvement of natural product
discovery Current Opinion in Microbiology 9252ndash260
Varga J Toacuteth B 2005 Novel strategies to control mycotoxins in feeds a review Acta
Veterinaria Hungarica 53189-203
Voigt CA Schafer W Salomon S 2005 A secreted lipase of Fusarium graminearum is a
virulence factor required for infection of cereals The Plant Journal 42364ndash375
Vogelgsang S Hecker A Musa T Dorn B Forrer HR 2011 On farm experiments over five
years in a grain maizewinter wheat rotation Effect of maize residue treatments on
Fusarium graminearum infection and deoxynivalenol contamination in wheat In Proc
7th
Canadian Workshop on Fusarium Head Blight edited by J Gilbert A Tekauz N
Sweetland and K Slusarenko Winnipeg Manitoba Canada p 41
Voumllkl A Vogler B Schollenberger M Karlovsky P 2004 Microbial detoxification of
mycotoxin deoxynivalenol Journal of Basic Microbiology 44147ndash156
Wagacha JM Muthomi JW 2007 Fusarium culmorum Infection process mechanisms of
mycotoxin production and their role in pathogenesis in wheat Crop Protection 26877-
885
Wakulinski W 1989 Phytotoxicity of the secondary metabolites of fungi causing wheat head
fusariosis (head blight) Acta Physiologiae Plantarum 11301ndash306
208
Wang JT Chang SC Chen YC Luh KT 2000 Comparison of antimicrobial susceptibility of
Citrobacter freundii isolates in two different time periods The Journal of Microbiology
Immunology and Infection 33258-62
Wannemacher RJ Stanley L Wiener MD 2000 Trichothecenes mycotoxins In Medical
aspects of chemical and biological warfare edited by Zajtchuk R Washington DC
Department of the Army pp 656-676
Westphal AH Swaving J Jacobs L de Kok A 1998 Purification and characterization of a
flavoprotein involved in the degradation of epoxyalkanes by Xanthobacter Py2 European
Journal of Biochemistry 257160ndash168
Whalen JG Mully TW Enlgish JC 2007 Spontaneous Citrobacter freundii infection in an
immunocompetent patient Archives of Dermatology 143124-5
White 2007 The physiology and biochemistry of prokaryotes Third edition Oxford
University Press New York Oxford p 628
Wilcke WF Morey RV 1995 Natural-air corn drying in the upper Midwest University of
Minnesota Minnesota Extension Service Publication BU-6577-E pp 20
Windels CE 2000 Economic and social impacts of Fusarium headblight changing farms and
rural communities in the Northern Great Plains Phytopathology 9017ndash21
Winzeler EA Shoemaker DD Astromoff A Liang H Anderson K Andre B Bangham R
Benito R Boeke JD Bussey H et al 1999 Functional characterization of the S
cerevisiae genome by gene deletion and parallel analysis Science 285901ndash906
Yang GH Jarvis BB Chung YJ Pestka JJ 2000 Apoptosis induction by the satratoxins and
other trichothecene mycotoxins relationship to ERK p38 MAPK and SAPKJNK
activation Toxicology and Applied Pharmacology 164149ndash160
209
Yang Q Yin GM Guo YL Zhang DF Chen SJ Xu ML 2010 A major QTL for resistance
to Gibberella stalk rot in maize Theoretical and Applied Genetics 121673ndash687
Yazar S Omurtag GZ 2008 Fumonisins trichothecenes and zearalenone in cereals
International Journal of Molecular Science 92062ndash2090
Yoshizawa T Morooka N 1973 Deoxynivalenol and its monoacetate new mycotoxins from
Fusaruium roseum and moldy barley Agricultural and Biological Chemistry 372933ndash
2934
Yoshizawa T Takeda H Ohi T 1983 Structure of a novel metabolite from deoxynivalenol a
trichothecene mycotoxin in animals Agricultural and Biological Chemistry 472133ndash
2135
Yoshizawa T 1993 Red-mold diseases and natural occurrence in Japan In Trichothecenes-
Chemistry Biology and Toxicological Aspects edited by Y Ueno Elsevier Amsterdam
Netherlands p 195-209
Yoshizawa T Sakamoto T Okamoto K 1984 In vitro formation of 3rsquo-hydroxy T-2 and 3rsquo-
hydroxy HT-2 toxins from T-2 by liver homogenates from mice and monkeys Applied
and Environmental Microbiology 47130-134
Young JC Subryan LM Potts D McLaren ME Gobran FH 1986 Reduction in levels of
deoxynivalenol in contaminated wheat by chemical and physical treatment Journal of
Agricultural and Food Chemistry 34461ndash465
Young JC Trenholm HL Friend DW Prelusky DB 1987 Detoxification of deoxynivalenol
with sodium bisulfite and evaluation of the effects when pure mycotoxin or contaminated
corn was treated and given to pigs Journal of Agricultural and Food Chemistry 35259ndash
261
210
Young JC Zhu H Zhou T 2006 Degradation of trichothecene mycotoxins by aqueous
ozone Food Chemistry and Toxicology 44417ndash424
Young JC Zhou T Yu H Zhu H Gong J 2007 Degradation of trichothecene mycotoxins by
chicken intestinal microbes Food Chemistry and Toxicology 45136-143
Yu Z Morrison M 2004 Comparisons of different hypervariable regions of rrs genes for use
in fingerprinting of microbial communities by PCR-denaturing gradient gel
electrophoresis Applied and Environmental Microbiology 704800-4806
Yu H Zhou T Gong J Young C Su X Li X Zhu H Tsao R Yang R 2010 Isolation of
deoxynivalenol-transforming bacteria from the chicken intestines using the approach of
PCR-DGGE guided microbial selection BMC Microbiology 10182
Zeibdawi A Steele M Savoie A 2011 An overview of CFIAs mycotoxin monitoring
program in livestock feeds In Proc 7th
Canadian Workshop on Fusarium Head Blight
edited by J Gilbert A Tekauz N Sweetland and K Slusarenko Winnipeg Manitoba
Canada p 25
Zhang J-B Li H-P Dang F-J Qu B Xu Y-B Zhao C-S Lia Y-C 2007 Determination of the
trichothecene mycotoxins chemotypes and associated geographical distribution and
phylogenetic species of the Fusarium graminearum clade from China Mycological
Research 111967ndash975
Zhao JS Manno D Thiboutot S Ampleman G Hawari J 2007 Shewanella canadensis sp
nov and Shewanella atlantica sp nov manganese dioxide- and hexahydro-135-trinitro-
135-triazine-reducing psychrophilic marine bacteria International Journal of Systemic
Evolution and Microbiology 572155ndash2162
211
Zhao X-Q 2011 Genome-based studies of marine microorganisms to maximize the diversity
of natural products discovery for medical treatments Evidence-Based Complementary
and Alternative Medicine Article ID 384572 doi1011552011384572
ZhaoY Park RD Muzzarelli RA 2010 Chitin deacetylases properties and applications
Marine Drugs 824ndash46
Zhou HR Lau AS Pestka JJ 2003a Role of double-stranded RNA-activated protein kinase R
(PKR) in deoxynivalenol-induced ribotoxic stress response Toxicological Science
74335ndash344
Zhou HR Islam Z Pestka JJ 2003b Rapid sequential activation of mitogen-activated protein
kinases and transcription factors precedes proinflammatory cytokine mRNA expression in
spleen of mice exposed to the trichothecene vomitoxin Toxicological Sciences 72130ndash
142
Zhou T He J Gong J 2008 Microbial transformation of trichothecene mycotoxins World
Mycotoxin Journal 123-30
Zhou G Zhang H He Y He L 2010 Identification of a chitin deacetylase producing bacteria
isolated from soil and its fermentation optimization African Journal of Microbiology
Research 42597-2603

ABSTRACT
ISOLATION CHARACTERIZATION AND GENOME SEQUENCING OF A SOIL-
BORNE CITROBACTER FREUNDII STRAIN CAPABLE OF DETOXIFIYING
TRICHOTHECENE MYCOTOXINS
Rafiqul Islam Advisors
University of Guelph 2012 Dr K Peter Pauls
Dr Ting Zhou
Cereals are frequently contaminated with tricthothecene mycotoxins like
deoxynivalenol (DON vomitoxin) which are toxic to humans animals and plants The goals
of the research were to discover and characterize microbes capable of detoxifying DON under
aerobic conditions and moderate temperatures To identify microbes capable of detoxifying
DON five soil samples collected from Southern Ontario crop fields were tested for the ability
to convert DON to a de-epoxidized derivative One soil sample showed DON de-epoxidation
activity under aerobic conditions at 22-24degC To isolate the microbes responsible for DON
detoxification (de-epoxidation) activity the mixed culture was grown with antibiotics at 50ordmC
for 15 h and high concentrations of DON The treatments resulted in the isolation of a pure
DON de-epoxidating bacterial strain ADS47 and phenotypic and molecular analyses
identified the bacterium as Citrobacter freundii The bacterium was also able to de-epoxidize
andor de-acetylate 10 other food-contaminating trichothecene mycotoxins
A fosmid genomic DNA library of strain ADS47 was prepared in E coli and screened
for DON detoxification activity However no library clone was found with DON
detoxification activity The whole genome of ADS47 was also sequenced and from
comparative genome analyses 10 genes (characterized as reductases oxidoreductases and
deacetylase) were identified as potential candidates for DONtrichothecene detoxifying
enzymes Bioinformatic analyses showed that the major animal pathogenic locus of
enterocyte effacement (LEE) operon genes absent from the ADS47 genome Strain ADS47
has the potential to be used for feed detoxification and the development of mycotoxin
resistance cereal cultivars
iii
DEDICATIONS
The PhD thesis is dedicated to the departed souls of my mother MOZIBUN NESSA and
father-in-law DR MA AHAD MIAH
iv
ACKNOWLEDGEMENTS
Foremost I am deeply indebted to my advisors Prof K Peter Pauls and Dr Ting Zhou
for the continuous support of my PhD study for their endurance inspiration enthusiasm and
mentoring Their continued guidance has helped me in all aspects of my research and in
writing of this dissertation I sincerely express my gratitude to them for their co-operation
from the beginning to conclusion of the thesis and many other areas This will guide me in my
future professional career
My thesis committee guided me through all these years I would like to express my
gratitude to Prof Paul Goodwin for his insightful comments and advice on my thesis research
Special thanks to Dr Chris Young for teaching me chemistry chemical structures and
interpreting liquid chromatography ultraviolet mass spectra data I am sincerely grateful to
Dion Lepp for helping in analyzing the bacterial genome and sharing his knowledge and
valuable suggestions on writing the genomic chapter
I would like to express my appreciation to staff members of Dr Ting Zhou and Prof
K Peter Pauls who supported me in many aspects during my PhD study I am thankful to
Jianwei He Xiu-Zhen Li Honghui Zhu for collections and preparation of soil samples and
LC-UV-MS analyses of numerous samples Special thanks to Dr David Morris Johnston
Monje for sharing his knowledge discussions and camaraderie at coffee breaks
I wish to acknowledge the Natural Sciences and Engineering Research Council of
Canada (NSERC) for awarding me a prestigious scholarship for my PhD program and
Deanrsquos Tri-Council Scholarship (University of Guelph) which made my life easier during this
study I also would like to convey my thanks to Agriculture and Agri-Food CanadaBinational
v
Agricultural Research and Development Fund (AAFCBARD) project for providing me
financial support
Mike Bryan a very supportive person helped provide solutions to my software issues
that improved my exposure and caliber at conference awards competitions Several student
competition awards honored and inspired me to work hard and lead to the successful
completion of my thesis research I would like to convey my appreciation to the judges of
different student award committees who recognized my work They are Society for Industrial
Microbiology and Biotechnology conference 2010 (USA) Food Safety Research Forum-
Ontario Ministry of Agriculture and Rural Affairs annual meeting 2010 (Canada)
International Mycological Conference 2009 (UK) and Southwestern Ontario Regional
Association of Canadian Phytopathological Society annual meeting 2009-2011 (Canada)
I would like to express my very special thanks to Prof Golam Hafeez Kennedy my
brother-in-law who is like my elder brother In many aspects I am indebt to him for his
cooperations which aided me to achieve my current status
Lastly and most importantly I wish to give my special thanks to my wife Mimi and
lovely daughter Racy whose endless love sacrifice support and encouragement enabled me
to cross the tough long road and made possible to achieve my goal and aspirations Racy
once you told me that one of your desires for me is to finish my PhD which is done now I
know you want to be a scientist which is a great ambition I wish you all the best in your
future aims Racy
vi
TABLE OF CONTENTS
DEDICATIONShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipiii
ACKNOWLEDGEMENTShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipiv
TABLE OF CONTENTShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipvi
LIST OF TABLEShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipix
LIST OF FIGUREShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipx
LIST OF ABBREVIATIONShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipxiii
10 GENERAL INTRODUCTION AND LITERATURE REVIEW helliphelliphelliphelliphelliphelliphelliphellip1
11 Backgroundhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip1
12 Plant pathogenesis and trichothecene productionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip3
13 Biosynthesis of food contaminating TChelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip6
14 Occurrence of TC mycotoxins in food and feed helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip11
15 Significance of deoxynivalenol and trichotheceneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip16
16 Toxic effects of deoxynivalenol and trichotheceneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip19
17 Cellular and molecular mechanisms of DON toxicity in animals and plantshelliphelliphelliphellip22
18 Strategies to control Fusarium head blight and trichotheceneshelliphelliphelliphelliphelliphelliphelliphelliphellip26
19 Isolation of DONTC de-epoxidating micro-organisms and geneshelliphelliphelliphelliphelliphelliphelliphellip28
110 The bacterial species Citrobacter freundiihelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip30
111 Hypotheses and objectiveshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip31
vii
20 AEROBIC AND ANAEROBIC DE-EPOXIDATION OF MYCOTOXIN
DEOXYNIVALENOL BY BACTERIA ORIGINATING FROM AGRICULTURAL
SOIL33
21 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip33
22 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
23 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip37
24 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip45
25 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip62
26 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip64
30 ISOALTION AND CHARACTERIZATION OF A NOVEL SOIL BACTERIUM
CAPABLE OF DETOXIFYING ELEVEN TRICHOTHECENE MYCOTOXINS65
31 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip65
32 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip66
33 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip68
34 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip75
35 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip96
36 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip100
40 CONSTRUCTION AND FUNCTIONAL SCREENING OF GENOMIC DNA
LIBRARY FOR ISOLATION OF DON DE-EPOXYDIZING GENE(S) FROM
BACTERIAL STRAIN ADS47helliphelliphelliphelliphelliphelliphelliphellip102
41 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip102
42 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip103
43 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip104
viii
44 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip112
45 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip118
46 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip121
50 GENOME SEQUENCING CHARACTERIZATION AND COMPARATIVE
ANALYSIS OF A TRICHOTHECENE MYCOTOXIN DETOXIFYING
CITROBACTER FREUNDII STRAIN ADS47helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip122
51 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip122
52 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip123
53 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip125
54 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip129
55 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip162
56 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip168
60 CONCLUSIONhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip171
61 Summary helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip171
62 Research contributions helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip174
63 Future workhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip175
70 REFERENCEShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip176
ix
LIST OF TABLES
Table 1-1 Fusaria produced type-A and type-B trichothecene mycotoxinshelliphelliphelliphelliphelliphelliphellip9
Table 1-2 Worldwide occurrence of deoxynivalenol (DON) in animal feedshelliphelliphelliphelliphellip12
Table 1-3 Occurrence of DON in Canadian grains grains foods and beershelliphelliphelliphelliphelliphelliphellip13
Table 1-4 Legislative maximum tolerated levels of trichothecene mycotoxins in Canadahellip14
Table 1-5 In vitro and in vivo synergistic effects of different food contaminating
mycotoxins helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip18
Table 3-1 Microbial transformation of type-A and type-B tricthothecene mycotoxins
after 72 h of incubation under aerobic conditions and at room temperaturehelliphelliphelliphelliphelliphellip95
Table 4-1 Analysis of Fosmid DNA library for deoxynivalenol (DON) de-epoxidation
capability helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip117
Table 5-1 General features of C freundii strain ADS47 genomehelliphelliphelliphelliphelliphelliphelliphelliphellip132
Table 5-2 Genome comparisons between strain ADS47 and five Citrobacter species141
Table 5-3 Functional categorized gene abundances among the six Citrobacter genomeshellip145
Table 5-4 Unique genes of C freundii strain ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip156
Supplemental Table S5-1 List of unique genes of strain ADS47 gene IDs and their
functional roleshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip169
x
LIST OF FIGURES
Figure 1-1 Structures of trichothecene mycotoxins produced by Fusariahelliphelliphelliphelliphelliphelliphelliphellip7
Figure 1-2 Cellular and molecular mechanisms of deoxynivalenol toxicity in
eukaryotic organismshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip23
Figure 2-1 Mechanism of microbial DON de-epoxidationhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip36
Figure 2-2 LC-UV-MS chromatographic analysis of DON and its de-epoxidated product
dE-DON obtained by culturing with selected soil microbeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip46
Figure 2-3 Time course of relative biotransformation of DON to dE-DON by the initial
soil suspension culture in MSB mediumhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip49
Figure 2-4 Microbial de-epoxidation of DON after five days of incubationhelliphelliphelliphelliphelliphellip51
Figure 2-5 Analysis of microbial population reduction by terminal restriction fragment
length polymorphism (T-RFLP)helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip53
Figure 2-6 Microbial genera identified in the enriched DON de-epoxidizing microbial
culturehelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip56
Figure 2-7 Effects of culture conditions on DON to dE-DON transformation by the
enriched microbial culturehelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip58
Figure 2-8 Microbial DON to dE (de-epoxy)-DON transformation activity under aerobic
and anaerobic conditionshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip61
Figure 3-1 Colony color shape and size of the mycotoxin degrading bacterial strain
ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip76
Figure 3-2 Dendrogram based on 16S-rDNA sequences for strain ADS47 and 11
reference Citrobacter specieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip77
xi
Figure 3-3 Effects of DON concentration on bacterial growth and de-epoxidation
activitieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip79
Figure 3-4 Bacterial growth and DON de-epoxidation capabilities under aerobic and
anaerobic conditions helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip82
Figure 3-5 Effects of media on bacterial growth and DON de-epoxidation activityhellip85
Figure3-6 Effects of temperature on microbial growth and DON de-epoxydation helliphelliphellip87
Figure 3-7 Influence of pH on microbial growth and DON de-epoxydation capabilityhellip89
Figure 3-8 Effects of sodium azide (SA) on bacterial DON de-epoxidation activityhelliphelliphellip91
Figure 3-9 Determination of DON to deepoxy-DON transformation by bacterial culture
filtrate and cell extracthelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip92
Figure 3-10 Determination of microbial de-epoxidation and de-acetylation of a type-A
trichothecene mycotoxin (HT-2 toxin) by LC-UV-MShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip94
Supplemental Figure S3-1 Flow chart depicted the procedures of enrichment and isolation
of DON de-epoxidation bacterial strain ADS47helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip101
Figure 4-1 Schematic overview of the construction and functional screening of Fosmid
DNA libraryhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip105
Figure 4-2 Genetic map of copy controlled Fosmid vector (pCC1FOS)helliphelliphelliphelliphelliphelliphellip108
Figure 4-3 Preparation of 40 kb size insert DNA from a DON de-epoxidizing bacterium113
Figure 4-4 Production of genomic DNA library in E coli EPI300-T1 host strainhelliphelliphelliphellip114
Figure 4-5 Confirmation of the Fosmid DNA library construction in E coli helliphelliphelliphelliphellip 116
Figure 5-1 Co-linearity between C freundii strain ADS47 genome and C koseri helliphelliphellip129
Figure 5-2 Circular genome map of C freundii strain ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphellip133
xii
Figure 5-3 Size distribution of the predicted genes identified in C freundii strain
ADS47helliphelliphellip 135
Figure 5-4 Functional classification of the predicted coding sequences (CDS) of
C freundii strain ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip137
Figure 5-5 Phylogeny of the trichothecene detoxifying C freundii strain ADS47 139
Figure 5-6 Comparisons of coding sequences between C freundii strain ADS47 and
five Citrobacter specieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip143
Figure 5-7 Determination of the locus enterocyte effacement (LEE) operons in
Citrobacter species by comparing C rodentium helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip151
Figure 5-8 Locations of unique genes of C freundii strain ADS467helliphelliphelliphelliphelliphelliphelliphellip157
Figure 5-9 Unique genomic regions and associated genes in strain ADS47 compared
to five Citrobacter specieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip159
xiii
LIST OF ABBREVIATIONS
APCI= atmospheric pressure chemical ionization
AE= attaching and effacing
A= aerobic conditions
ABC= ATP-binding cassette
ACT= artemis comparative tools
AN= anaerobic conditions
3-ADON= 3-acetyl-deoxynivalenol
15-ADON= 15-acetyl-deoxynivalenol
ATA= alimentary toxic aleukia
bp= base pair
BGI= Beijing genomics institute
BLAST= basic local alignment search tool
BRIG= blast ring image generator
Bw= body weight
C= carbon
CDS= coding sequence
CFIA= Canadian food inspection agency
CHEF= clamped homogeneous electric fields
CMB= corn meal broth
CVTree= composition vector tree
DAS= diacetoxyscirpenol
dH2O= distilled water
DOGT1= UDP-glucosyltransferase
DON= deoxynivalenol
dsRNA= double stranded RNA
dE= de-epoxy
dE-DON= de-epoxy DON
E coli= Escherichia coli
ER=endoplasmic reticulum
FAO= Food amp Agriculture Organization
FM= full medium
FHB= Fusarium head blight
FUS= fusarenonX
gDNA= genomic DNA
GC= guanine cytosine
GER= Gebberella ear rot
GPI= glycosylphosphatidylinositol
h= hour
Hck= hematopoietic cell kinase
Kb= kilobase
xiv
Kg= kilogram
LC-UV-MS= liquid chromatography-ultraviolet-
mass spectrometry
LEE= locus of enterocyte effacement
LD50= lethal dose of 50 animal
LB= Luria Bertani
LMP= low melting point
MAPK= mitogen-activated protein kinase
microg= microgram
mg= milligram
mL= milliliter
microL= microliter
microM= micromole
min= minute
Mb= megabases
MCS=multiple cloning site
MSA= MSB+ agar
MSB= mineral salts broth
MM= minimal medium
MSC= mineral salts + 03 Bacto Peptone
MS-DON= mineral salts + 200 microgmL DON
mRNA= messenger RNA
MSY= mineral salts + yeast extract
mz= mass ion ratio
N= nitrogen
Na-Cl= sodiumndashclorine
NADH= (reduced) nicotinamide adenine
dinucleotide
NAD(P)= nicotinamide adenine
dinucleotide phosphate
NCBI= national center for biotechnology
information
NEO= neosolaniol
ng= nanogram
NIV= nivalenol
OD600nm= optical density at 600
nanometer wavelength
OMAFRA= Ontario Ministry of
Agriculture and Rural Affairs
Paa= phenylacetic acid
PCR= polymerase chain reaction
PDB= potato dextrose broth
ppm= parts per million
PFGE= pulsed field gel electrophoresis
PKR= protein kinase
xv
PNDO= pyridine nucleotide-disulfide
oxidoreductase
QTL= quantitative trait locus
RAST= rapid annotation subsystem technology
RDP= ribosomal database project
Rpl3= 60S ribosomal protein L3
rRNA= ribosomal RNA
s= second
SA= sodium azide
SDR= short chain dehydrogenasereductase
SE= soil extract
SIM= selected ion monitoring
SOAP= short oligonucleotide analysis package
sp= species
sRNA= small RNA
TC= trichothecene
TCA= tricarboxylic acid cycle
TSB= tryptic soy broth
TF= transcription factor
tRNA= transfer RNA
TIC= total ion count
TRAP= tripartite ATP-independent
periplasmic
T-RFLP=terminal restriction fragment
length polymorphism
TRI101= trichothecene 3-O-
acetyltransferase
YEB= yeast extract broth
VER= verrucarol
VKOR= vitamin K epoxide reductase
wv= weight by volume
1
CHAPTER 1
10 GENERAL INTRODUCTION AND LITERATURE REVIEW
11 Background
According to the Food and Agriculture Organization (FAO) of the United Nations
more than 25 of the worldrsquos food crops are contaminated with mycotoxins every year
causing serious adverse effects on livestock industry crop production and international trade
(Boutrif and Canet 1998 Desjardins 2006 Binder et al 2007 Surai et al 2010) The negative
effects of mycotoxins on the consumers are diverse including organ toxicity mutagenicity
carcinogenicity neurotoxicity teratogenicity modulation of the immune system reproductive
system effects and growth retardation Mycotoxins are toxic secondary metabolites produced
by various fungal species during crop colonization in field andor post harvest conditions
(FAO 1991 Barrett 2000) Aspergillus Penicillum and Fusarium are the predominant fungal
genera that accumulate economically important mycotoxins such as aflatoxins zearalenone
trichothecenes fumonisins ochratoxins and ergot alkaloids in agricultural commodities
Humans and animals are principally exposed to mycotoxins through consumption of
contaminated foodfeed and animal products such as milk meat and eggs (Fink-Gremmels
1989 Hollinger and Ekperigin 1999)
Trichothecenes (TCs) produced by a variety of toxigenic Fusarium species are
probably the most food and feed contaminating mycotoxins in the temperate regions of
America Europe and Asia (Creppy 2002 Yazar and Omurtag 2008) The fungal pathogen
accumulates various type-A (eg T-2 toxin HT-2 toxin) and type-B trichothecenes (eg
2
deoxynivalenol nivalenol and their derivatives) in cereals Among them type-B
trichothecenes particularly deoxynivalenol (DON) andor its derivatives appear to be the
most commonly found mycotoxins in grain and grain products worldwide impairing human
health and productivity of farm animals (Placinta et al 1999 Sherif et al 2009) A number of
human and animal disease outbreaks associated with TC contaminated grain food and feed
consumption have been reported in Asia (Japan India and central Asia) Europe New
Zealand and United States of America (USA) (Pestka and Smolinski 2005 Desjardins 2006)
A severe outbreak of Alimentary Toxic Aleukia (ATA) mycotoxicosis killed 100000 people
in the Orenburg region of Russia (Eriksen 2003) In 1996 losses of over $100 million were
estimated in Ontario because of costs related to managing DON (Schaafsma 2002)
Historically T-2 toxin DON and nivalenol (NIV) have likely been used as a biological
warfare in Laos Kampuchea and Afghanistan and there is concern because of the potential
use of these compounds for bioterrorism (Wannemacher et al 2000 Tucker 2001)
Despite the extensive efforts that have been made to prevent TC contamination with
the current varieties and production practices its presence in agricultural commodities is
likely unavoidable There are no crop cultivars currently available that are completely
resistant to Fusarium head blightGibberella ear rot (FHBGER) diseases Chemical or
physical methods cannot safely remove TC mycotoxins from grain completely or they are not
economically viable (Varga and Toacuteth 2005) Therefore an alternative strategy to address the
issue is of great importance (Awad et al 2010 Karlovsky 2011)
3
12 Plant pathogenesis and trichothecene production
Cereals infested by the ascomyceteous fungi may cause Gibberella ear rot (GER) in
maize (Zea mays L) and Fusarium head blight (FHB also known as lsquoscabrsquo) of small grain
cereals [wheat (Triticum aestivum L) barley (Hordeum vulgare L)] and other small grain
cereals (Goswami and Kistler 2004) As many as 17 fungal species have been reported to be
associated with FHBGER which include F graminearum Schwabe [telemorph Gibberella
zeae (Schwein) Petch] F culmorum (WG Smith) Sacc F sporotrichioides Scherb F
poae (Peck) Wollenw Microdochium nivale (Fr) Samuels amp IC Hallett and F avenaceum
(Fr) Sacc (Osborne and Stein 2007 Audenaert et al 2009) The two species F
graminearum and F culmorum are the principal species causing crop damage Among them
F graminearum is the widely adapted and most damaging species across the world including
North America (Dyer et al 2006) F culmorum tend to be the main species in the cooler parts
of North Central and Western Europe (Wagacha and Muthomi 2007)
The fungi overwinter mainly on crop residues as saprophytes where they produce
asexual (micro- and macro-conidia) andor sexual (ascospores) fruiting structures which act
as the initial source of inoculums in the subsequent crop season (Sutton 1982 Wagacha and
Muthomi 2007) Ascospores are primarily dispersed by air (Trail et al 2002) and conidia
move by rain-splash (Paul et al 2004) After they land on maize silks and florets of small
grain cereals infection occurs through natural openings (eg stomata) or occasionally by
direct penetration The pathogens can invade maize kernels through wounds caused in the
husks by insects or birds Infection may occur throughout the crop growing season but
disease incidence and severity is facilitated by warm temperatures (gt20degC) high humidity
(gt80) and frequent precipitation that coincides with anthesis (Osborne and Stein 2007
4
Wagacha and Muthomi 2007) The Fusarium pathogen initially infects as biotroph which
becomes a necrotroph as the disease progresses After penetration Fusaria produce different
extracellular cell wall degrading enzymes that trigger infection and hyphal progression
towards the developing earkernel where further colonization occurs and development of
FHBGER diseases (Voigt et al 2005 Miller et al 2007 Kikot et al 2009) FHB disease is
characterized by a bleached appearance around the middle of the head Severely infected
grains have a pinkish discoloration and can be shriveled In maize the disease produces a
reddishndashbrown discoloration of the kernels and progresses from the tip to the bottom of
affected ears
The fungi associated with FHB and GER contaminate cereals by accumulating various
host-nonspecific TC mycotoxins (Brennen et al 2007) TCs have typically been associated
with cereals grown in temperate climatic conditions including Canada (Doohan et al 2003)
DON production by F graminearum and F culmorum is maximal at warm temperatures (22-
28˚C) high grain moisture contents (gt17) and humid conditions (gt70 RH) (Martins and
Martins 2002 Hope et al 2005 Llorens et al 2006) Grain can be contaminated with TCs in
the field as well as in storage if conditions for fungal growth are optimal and grain contains
high moisture (gt17) (Larsen et al 2004 Birzele et al 2002)
Populations of FHBGER causing pathogens are dynamic Genetic variability between
and within Fusarium species appears to have occurred to allow for their adaptation to new
agro-climates (Desjardins 2006 Fernando et al 2011) Three strain specific Fusarium
chemotypes NIV 3-ADON (3-acetyl-deoxynivalenol) and 15-ADON (15- acetyl-
deoxynivalenol) have been identified which correlate with the geographical distributions
(Starkey et al 2007 Zhang et al 2007 Audenaert et al 2009) The NIV chemotype mainly
5
produced by F culmorum accumulates NIV and 4-NIV (Fusarinon X) is predominant in
Asia Africa and Europe (Lee et al 2001) The 3-ADON chemotype which synthesizes DON
and 3-ADON is the dominant strain in EuropeFar East (Larsen et al 2004) In North
America 15-ADON is the main chemotype which produces DON and 15-ADON (Goswami
and Kistler 2004) Significant Fusarium population shifting was observed in the past and in
recent years (Kant et al 2011) A dynamic change from F culmorum to F gramninearum
and DON to NIV producing chemotyes occurred in North West European countries The 3-
ADON chemotype which is uncommon in North America was found in North Dakota and
Minnesota (Goswami and Kistler 2004) F pseudograminearum which historically occurs in
Australia Africa and North West Pacific America has been found in Western Canada (Larsen
et al 2004 Kant et al 2011) In Manitoba a replacement of 15-ADON chemotype by the
more phytoxic 3-ADON type was observed in recent years (Fernando et al 2011) All
together the FHBGER associated pathogen complex is highly dynamic which not only
reduces crop yield but also decreases grain quality by accumulating various mycotoxins
6
13 Biosynthesis of food contaminating TC
TCs are fungal secondary metabolites that were initially identified in Trichotheceium
roseum Thus the name of this class of compounds follows the name of the genus
Trichotheceium (Freeman and Morrison 1949) There have been 180 TCs identified in various
fungal species belonging to the genera Fusarium Myrothecium Stachybotrys Trichoderma
and Thrichothecium (Eriksen 2003 Eriksen et al 2004) The FHBGER associated TCs are
mainly synthesized by different Fusaria and are characterized as tricyclic sesquiterpenoids
with a double bond at position C9-10 and a C12-13 epoxide ring
7
Figure 1-1 Structures of trichothecene mycotoxins produced by Fusaria (a) General
structure of trichothecenes R1 = H OH OiValeryl keto R2 = H OH R3= OH OAc R4 =
H OH OAc R5= OH OAc (b) Type-A trichothecene mycotoxin (c) Type-B trichothecene
mycotoxin which contain a carbonyl functional group at C8 position (Young et al 2007)
(a) (b)
(c)
8
The fungal metabolites are small in size (molecular weight 250-550) heat stable and
non-volatile molecules (Pestka 2007 Yazar and Omurtag 2008) Food contaminating TCs are
divided into two groups type-A and type-B (Figure 1-1b and 1c) Type-A is characterized by
the lack of a ketone function at C8 position and include T-2 toxin HT-2 toxin
diacetoxyscirpenol (DAS) and neosolaniol (NEO) which are mainly produced by F
sporotrichioides and F poae (Creppy 2002 Quarta et al 2006) The most frequently found
TC belongs to type-B which contains carbonyl functions at C8 positions DON NIV and
their acetylated derivatives The two types are further differentiated by the presence or
absence of functional hydroxyl (-OH) and acetyl (CH3-CO-) groups at C-3 C-4 C-7 C-8 and
C-15 positions (Table 1-1)
The syntheses of various TCs are speciesstrain specific and are genetically controlled
but influenced by environmental conditions (Desjardins 2006 Paterson and Lima 2010) Until
now 15 Fusaria catalytic genes and regulators involved in TC productions have been
identified (Merhej et al 2011) These biosynthetic genes are located at three loci on different
chromosomes The core trichothecene (TRI) gene cluster is comprised of 12 genes localized
in a 25-kb genomic region (Kimura et al 2001) The remaining three genes are encoded by
the TRI1-TRI16 and TRI101 loci (Gale et al 2005 Alexander et al 2009) Fusaria produce
TC following a number of oxygenation isomerization acetylation hydroxylation and
esterification reaction steps (Desjardins 2006 Desjardins and Proctor 2007 Kimura et al
2007 Merhej et al 2011) In brief farnesyl pyrophosphate is initially cyclized into
trichodiene by TRI5 which codes for trichodiene synthase In the next four steps
isotrichotriol is produced by TRI4 (a multifunctional cytochrome P450) via C-2 hydroxylation
and 1213 epoxidation followed by two hydroxylation reactions
9
Table 1-1 Fusaria produced type-A and type-B trichothecene mycotoxins Molecular weight
(MW) and acetyl (-R) hydroxyl (-OH) and ketone (=O) functions at different carbon (C)
position of the tricyclic rings A-types T-2 toxin HT-2 toxin DAS (diacetoxyscirpenol)
NEO (neosolaniol) VER (verrucarol) B-type DON (deoxynivalenol) 3-ADON (3-acetyl-
deoxynivalenol) 15-ADON (15-acetyl-deoxynivalenol) NIV (nivalenol) and FusX
(fusarenon X) (Young et al 2007)
Trichothecenes MW R1C3 R2C4 R4C7 R3C15 R5C8
Type-A
T-2 toxin 466 OH OCOCH3 H OCOCH3 OCOCH2CH(CH3)2
HT-2 toxin 424 OH OH H OCOCH3 OCOCH2CH(CH3)2
DAS 366 OH OCOCH3 H OCOCH3 H
NEO 382 OH OCOCH3 OH OCOCH3 H
VER 266 H OH H OH H
Type-B
DON 296 OH H OH OH =O
3-ADON 338 OCOCH3 H OH OH =O
15-ADON 338 OH H OH OCOCH3 =O
NIV 312 OH OH OH OH =O
FusX 354 OH OCOCH3 OH OH =O
10
The next two non-enzymatic reactions results in isotrichodemol Eventually the
trichothecene skeleton is produced via the formation of a C-O bond between the C-2 oxygen
and C-11 Subsequent acetylation at C-3 by TRI101 hydroxylation at C-15 by TRI1 and
catalytic activity of TRI3 leads to calonectrin production The pathway described above is
common for both type-A (eg T-2 toxin HT-2 toxin) and type-B (eg DON NIV and their
acetylated derivatives) TCs
The following steps are speciesstrain specific In DON producing F graminearum
strains calonectrin is converted to either 3-ADON or 15-ADON followed by DON production
by the catabolic activities of TRI1 and TRI8 The biosynthesis of 3-ADON or 15-ADON is
controlled by variation in the esterase coding sequences of TRI8 F graminearum produces
NIV from calonectrin via synthesis of 4-NIV followed by NIV by TRI1 and TRI8 gene
actions NIV can be produced either from DON by hydroxylation at C-4 or via
transformation of calonectrin to 315-diacetoxyscirpenol (315-DAS) followed by addition of
a ketone at C-8 position by the activity of the TRI7 and TRI13 genes 315-DAS is the
principal substrate for synthesis of type-A TCs by F sporotrichioides the main type-A
producer The biosynthetic pathway of T-2 toxin involves C-4 acetylation of 315-DAS by the
TRI7 gene product and TRI8 triggered by TRI1 315-DAS is also the substrate for HT-2
toxin synthesis In the absence of TRI7 expression HT-2 toxin is produced through
hydroxylation of 315-DAS and isovalerate addition to C-8 position (Foroud and Eudes 2009)
11
14 Occurrence of TC mycotoxins in food and feed
TC producing Fusaria are adapted to variable climatic conditions and agricultural
ecosystems As a consequence this class of mycotoxins particularly DON is frequently
detected in foods and feeds including breakfast cereals bakery products snack foods beer
and animal feed derived from cereals in most part of the world (Pittet 2001 Desjardins 2006
Lancova et al 2008) A two-year survey during 2003-2005 conducted by Romer Lab Inc
revealed unacceptable levels of DON in animal feedsfeed raw materials (such as maize
wheat wheat bran) in Asia and Oceania (1062 microgg) central Europe (802 microgg) Mid-West
USA (55 microgg) northern Europe (551 microgg) and southern Europe (303 microgg) (Binder et al
2007) (Table 1-2)
Quite high levels of DON contamination (19 microgg) were detected in north Asian and
Chinese feedfeed materials Recently five-year on-farm studies in Switzerland showed that
74 of the wheat grain and straw samples were contaminated with high levels of DON
(average 5 microgg) The amount is higher than the European maximum limit of 125 microgg
(Vogelgsang et al 2011)
The occurrences of TCs in Canadian agricultural commodities are indicated in Table
1-3 Seventeen surveys conducted during the 1980s and 1990s showed up to 100 DON
incidence in Ontario wheat and maize samples with an 18 average incidence (Scott 1997) A
three-year survey recently (2006-08) conducted by the Ontario Ministry of Agriculture and
Rural Affairs (OMAFRA) detected high levels of DON in spring (maximum 26 microgg) and
winter wheat (maximum 29 microgg) samples (Ana Matu personal communication) The DON
levels were much higher than Health Canadarsquos maximum limit of 2 microgg for uncleaned soft
wheat for staple foods (Table 1-4)
12
Table 1-2 Worldwide occurrence of DON in animal feed Feed and feed raw material
samples were collected from animal farmsfeed production sites during 2003-2005 The
survey was conducted by Romer Labs Inc (httpwwwromerlabscom)
Country
region
Sample
type
Incidence
()
Maximum
DON content
(microgg)
Mean DON
content
(microgg)
Survey
period
Source
North
AsiaChina
Animal
feed
071 1899 092
Binder et
al 2007
South East
Asia
014 152 016
South Asia 070 1062 049
Oceania 2003-05
Australia 041 186 030
Northern
Europe
070 551 059
Central
Europe
066 802 057
Southern
Europe
052 303 030
13
Table 1-3 Occurrence of DON in Canadian grains grain foods and beers Infant cereal foods
oat barley soy and multi-grain based foods
Province Sample
type
Incidence
()
Maximum
DON
contents
(microgg)
Multiple
mycotoxins
samples ()
Survey
period
Sources
Ontario Spring
Wheat
70 260 Yes 2006-08 OMAFRA1
Ontario Winter
wheat
49 290 Yes 2006-08 OMAFRA
Ontario Maize 78 140 Yes 2009-10 Limay-Rios
and
Schaafsma
2010
Ontario Maize - 1000 Yes 2011 AampL
Biologicals
Canada Breakfast
cereals
46 094 430 1999-01 Roscoe et al
2008
Canada Animal
feeds
73 425 Yes 1990-10 Zeibdawi et
al 2011
Canada Beers 58 0052 50 1990s Scott 1997
Canada Infant
Cereals
63 098 Yes 1997-99 Lombaert et
al 2003
1
Ontario Ministry of Agriculture and Rural Affairs
2 microgmL
14
Table 1-4 Legislative maximum tolerated levels of trichothecene mycotoxins in Canada
according to Canadian Food Inspection Fact Sheet April 23 2009 DON- deoxynivalenol
DAS- diacetoxyscirpenol
Products DON
(microgg)
T-2 toxin
(microgg)
HT-2 toxin
(microgg)
DAS
(microgg)
Uncleaned soft wheat for human
consumption
2 - - -
Cattlepoultry diets 5 lt1 01 lt1
Swinedairy cattle feeds 1 lt1 0025 lt2
15
High DON incidence (78) was observed in maize samples collected from
southwestern Ontario fields with moderate levels (lt1 microgg) of contamination during 2009 and
2010 (Limay-Rios and Schaafsma 2010) Some samples had high DON levels (14 microgg) In
the 2011 crop season extremely high levels of DON (gt100 microgg) were found in some
southern Ontario maize samples (AampL Biologicals London ON Personal communication)
In a Canadian Food Inspection Agency (CFIA) 20 year survey (1990-2010) 73 of
animal feed samples collected from feed mills farms and retail stores across Canada were
contaminated with DON and for more than 25 of the samples the mycotoxin levels were
above the maximum limit (ge 1 microgg) (Zeibdawi et al 2011) The highest level of DON (15
microgg) detected in the 2006-2007 samples were much lower than the levels found in the 1999-
2000 samples which contained a maximum of 425 microgg DON Importantly this study
reported a significant number of samples contaminated with multiple mycotoxins DON NIV
3-ADON 15-ADON T-2 toxin HT-2 toxin DAS and NEO
Earlier studies in the 1990s detected high levels of DON content in Eastern Canadian
cereals maize (maximum 175 microgg) wheat (maximum 916 microgg) barley (maximum 911
microgg) and oats (maximum 12 microgg) (Campbell et al 2002) The study identified a significant
number of samples (15-367) that contained multiple mycotoxins and up to 33 of the
wheat samples exceeded 1 microgg DON Low amounts but high incidences of TCs were found
in cereal-based foods collected from Canadian retail stores Sixty three percent of infant
cereal foods 46 of breakfast cereals and 58 of beers were contaminated with one or
multiple mycotoxins (Scott 1997 Lombaert et al 2003 Roscoe et al 2008) The maximum
amount of DON was found in infant foods (098 microgg) followed by breakfast cereals (094
microgg) and beers (005 microgmL)
16
15 Significance of deoxynivalenol and trichothecenes
For most cereals including wheat maize rice barley oats and rye the risk of
contamination with TCs resulted in grain quality reduction and FHBGER disease progression
(Bai et al 2002 Jansen et al 2005) The impacts of TC contamination in agricultural
commodities are diverse including reduced livestock production induced human illnesses
and enhanced FHB disease severity which may result in the reduction of crop yield Exposure
of farm animals (particularly pigs) to DON is a continuing threat to the livestock industries in
Canada USA and Europe (DrsquoMello et al 1999) Ingestion of TC-contaminated feeds
generally causes reduction of feed intake feed refusal poor feed conversion retardation of
weight gain increased disease susceptibility and reduced reproductive capacities (Binder et al
2007) High doses of DON result in acute toxic effects in pigs (eg emesis) while low doses
(ge1 microgmL) cause chronic effects (eg growth retardation in pigs) (Rotter et al 1996
Accensi et al 2006) Reduction of growth and reproduction ability was observed in
experimental rats exposured to much higher doses of TC (Sprando et al 2005 Collins et al
2006) Several animal disease outbreaks that occurred in different continents have been
attributed to consumption of feed contaminated with DON NIV and DAS (Desjardins 2006)
The consequences of human exposure to TCs include liver damage gastricintestinal
lesions and central nervous system toxicity resulting in anorexia nausea hypotensionshock
vomiting diarrhea and death (Pestka 2007) Human mycotoxicosis (illnesses and outbreaks)
by consumption of type-A and type-B TC mycotoxins in Europe New Zealand Russia
Japan South America India and Vietnam have been reported (Bhat et al 1989 Yoshizawa
1993 Luo 1994 Desjardins 2006) Severe outbreaks of Alimentary Toxic Aleukia (ATA)
associated with TC killed thousands of people in the Orenburg region of Russia during the
17
1930rsquos and 1940rsquos (Eriksen 2003) TCs (T-2 DON and NIV) were likely used for biological
warfare in Laos Kampuchea and Afghanistan and its potential application for bioterrorism is
an additional concern associated with TC (Tucker 2001 Bennett and Klich 2003)
TCs are phytotoxic and may contribute to FHB disease severity in cereals (Proctor et
al 1995 Maier et al 2006 Masuda et al 2007) Studies demonstrated that DON caused
wilting chlorosis inhibition of seedling growth and necrosis in cereals (Cossette and Miller
1995 Mitterbauer et al 2004 Rocha et al 2005) DON-mediated host cell damage and
subsequent necrotrophic fungal spread has been shown to lead to FHBGER disease
development and higher kernel damage (Procter et al 1995 Mitterbauer et al 2004 Maier et
al 2005) This suggests that DON contributes to yield losses as well as to reductions in the
market value of cereals (Osborne and Stein 2007) Severely infected grain may contain more
than 60 microgg DON which is unacceptable to the milling and brewing industries (Okubara et
al 2002) Significant crop losses due to FHB disease epidemics and mycotoxin contamination
have been reported in Ontario (Schaafsma 2002) FHB disease epidemics during 1998 to 2000
caused huge crop losses resulting in farm failures in the northern Great Plains in the USA
(Windels 2000) In that period FHB associated losses were estimated at US $ 27 billion in
the region (Nganje et al 2002)
Importantly Fusaria infested grains are often co-contaminated with multiple
mycotoxins which further increase health risks as interactions between the mycotoxins may
cause greater toxic effects than expected (Schollenberger et al 2006) This implies that an
apparently low level of contamination of a single mycotoxin can have severe effects on
consumers because of synergistic effects among multiple co-occuring mycotoxins (Surai et al
2010)
18
A number of studies demonstrated synergistic toxic effects among Fusarium
mycotoxins on pigs turkeys lamb chicks as well as in in vitro Caco-2 cell and yeast
bioassays (Table 1-5)
Table 1-5 In vitro and in vivo synergistic effects of different food contaminating mycotoxins
DON-deoxynivalenol DAS-diacetoxyscirpenol FB1-fumonisins 1 NIV-nivalenol ZEN-
zearelenone FA-fusaric acid Afl-aflatoxin
Mycotoxins source Interacting
mycotoxins
Experimental
animalcells
Effects Reference
Naturally contaminated
cereals
DON FA Pigs Inhibit growth Smith et al
1997
DON contaminated
wheat and FB1
DON FB1 Pigs Weight loss Harvey et al
1996
Culture materials
contained mycotoxins
T-2 FB1 Turkey poults Weight loss oral
lesion
Kubena et al
1997
Inoculated rice and
pure mycotoxin
DAS Afl Lamb Weight reduction Harvey et al
1995
Naturally DON
contaminated wheat
and pure T-2 toxin
DON T-2 Chicks Reduced body
weight significantly
Kubena et al
1989
Pure mycotoxins DAS Afl Chicks Reduced body
weight
Kubena et al
1993
Pure mycotoxins T-2 Afl Chicks Reduced weight
oral lesion
Kubena et al
1990
Pure mycotoxins DON
ZENFB1
Human intestinal
Caco-2 cells
Increased DNA
fragmentations
Kouadio et al
2007
Pure mycotoxins DON NIV Yeast bioassay Synergistic effects Madhyastha et
al 1994
Pure mycotoxins T-2 HT-2 Yeast bioassay Synergistic effects Madhyastha et
al 1994
19
16 Toxic effects of deoxynivalnol and trichothecenes
As small molecules TCs simply enter into cells and interact with different cell
components resulting in disruption of cellular functions and causing various toxic effects that
include inhibition of protein synthesis and ribosomal fragmentation (Eudes et al 2000
Minervini et al 2004 Pestka 2007) Simple absorption of DON into human intestinal Caco-2
cells and crossing human placenta barriers through passive diffusion has been reported
(Sergent et al 2006 Nielsen et al 2011)
TC may cause various toxicities in animals and plants including apoptosis
immunotoxicity chromatin condensation disruption of cellularmitochondrion membranes
disruption of gut tissues inhibition of protein and ribosome synthesis breakage of ribosome
and cell damage The toxicity of TC is influenced by structural variability (ie position
number and type of functional groups attached to the tricyclic nucleus and C-C double bond at
C9-10 position) (Desjardins 2006) Nevertheless epoxide function at the C12-13 position is
the key for TC cytotoxicity (Swanson et al 1988 Eriksen et al 2004)
Different in vitro and in vivo studies revealed that the method and period of exposure
amount of TC and types of celltissue and host species have significant effects on TCrsquos
cytotoxicity (Scientific Committee for Food 2002 Rocha et al 2005 Nielsen et al 2009)
Administration of 10 mgkg body weight (bw) NIV caused apoptosis in mice thymus Peyerrsquos
patch and spleen tissues (Poapolathep et al 2002) DON and NIV at 887 mgkg bw for 3 days
a week for four weeks induced immunotoxicity in experimental mice determined by
estimating increased immunoglobulins (IgA) and decreased IgM levels (Gouze et al 2007)
Exposure of mice to 1 mgkg bw of DON induced skeletal malformations and reduced mating
tendency while significant resorption of embryos observed at the dose of gt10 mgkg bw
20
(Khera et al 1982) At high acute doses (4972 mgkg bw) DONNIV and type-A
trichothecenes caused mice death because of intestinal haemorrhage necrosis of bone
marrowlymphoid tissues and kidneyheart lesions (Yoshizawa and Morooka 1973
Yoshizawa et al 1994 Forsell et al 1987 Ryu et al 1988) In a separate study 15-ADON
showed more toxicity than DON when the mycotoxins were orally administered to mice
(Forsell et al 1987)
Studies suggested that type-A TCs are more toxic in mammals than type-B which
means that functional variability at the C-8 position of TC has a significant role in toxicity
The toxicity of various TC mycotoxins in animals can be ranked in the order of T-2
toxingtHT-2 toxingtNIVgtDONge3-ADONgt15-ADON according to the results from human cell
lines and mice model systems bioassays (Ueno et al 1983 Pestka et al 1987 Nielsen et al
2009) Moreover animal feeding assays demonstrated that monogastric animals are most
sensitive to DON The order of sensitivity of animals to DON is
pigsgtrodentsgtdogsgtcatsgtpoultrygtruminants (Pestka 2005) The reasons for the variations in
DON toxicity species might be due to differences in metabolism absorption distribution and
elimination of the mycotoxin among animal species (Pestka 2007)
To my knowledge the mechanism of TC uptake by plant cells has not been studied
Nonetheless cereals are exposed to TC by toxigenic Fusaria infection which may cause
various toxicities such as cell damage wilting chlorosis necrosis program cell death and
inhibition of seedling growth in sensitive plants (eg wheat maize and barley) (Cossette and
Miller 1995 Rocha et al 2005 Boddu et al 2007) As animals TC inhibits protein synthesis
in non-host A thaliana (Nishiuchi et al 2006) Immunogold assays showed that the cellular
targets of DON in wheat and maize were cell membrane ribosome endoplasmic reticulum
21
vacuoles and mitochondria Modulation of cell membrane integrity and electrolyte losses
were observed by direct DON interference in maize and wheat cell assays (Cossette and
Miller 1995 Mitterbauer et al 2004) DON-mediated mitochondrion membrane alterations
disruption of host cells enzymatic activity apoptosis and necrosis have been observed in
cereals (Mitterbauer et al 2004 Rocha et al 2005) In contrast to animal type-B is more toxic
to plants than type-A DON showed 8- and 13-fold phytotoxicity to wheat coleoptiles than T-
2 and HT-2 toxins respectively (Eudes et al 2000) By compilation of different studies TC
phytoxicity can be ordered as DONge3-ADONgtT-2 toxingtDAS (Wakulinski 1989 McLean
1996 Eudes et al 2000)
22
17 Cellular and molecular mechanisms of DON toxicity in animals and plants
The underlying cellular and molecular mechanisms of DONTC toxicity are evolving
Exposure to DON proceeds with the up and down regulation of numerous critical genes of
eukaryotic organisms which causes various negative effects on animal and plant organisms
(Figure 1-2)
One of the early events of DON toxicity occurs when the mycotoxin binds to the
active ribosome resulting in blocking protein translation through different routes by
damaging the ribosome (Shifrin and Anderson 1999) degrading or altering conformational
changes of 28S-rRNA (Alisi et al 2008 Li and Pestka 2008) and inducing expression of
miRNAs This results in down regulation of the ribosome associated genes (He et al 2010a
Pestka 2010) Both animal and plant cell assays demonstrated that DON bound to the peptidyl
transferase (Rpl3) domain of the 60S subunit of ribosome and blocked protein chain
elongation and termination at high DON concentrations while peptide chain termination and
ribosome breakage occurred at a low level of DON exposure (Zhou et al 2003a)
23
Figure 1-2 Cellular and molecular mechanisms of deoxynivalenol toxicity in eukaryotic
organisms DON is simply absorbed by the cell and interacts with the cytoplasmic membrane
(CM) mitochondrion (MT) membrane and ribosome Disruption of the CM and alteration of
the MT membrane causes a loss of electrolytes and affects the function of the MT
respectively DON binds to the active ribosomes which leads to ribosome damage and
induces micro RNAs (miRNA) Expression of miRNA impairs the synthesis of ribosome
associated protein The subsequent transduction and phosphorylation of the RNA-activated
protein kinase (PKR) and hematopoietic cell kinase (Hck) activate the mitogen-activated
protein kinase (MAPK) cascade Phosphorylation of the MAPKs induces targeted
transcription factors (TF) which results in ribotoxis stress response upregulation of
proinflammatory cytokines and dysregulation of neuroendocrine signaling (derived from
Pestka 2007 and 2010)
24
The consequence of DON interaction with ribosomes leads to cellular and molecular
changes which are complex and not fully understood However it is well-demonstrated that
the activation of the mitogen-activated protein kinase (MAPK) and subsequent molecular
actions play key roles in the cytotoxicity of TC Upon binding to the ribosome DON rapidly
induced interactions between the protein kinases and the ribosomal 60S subunit as observed in
mononuclear phagocytes (Bae and Pestka 2008) Ribosomal binding to DON activated the
ribosome associated kinases for instances protein kinase R (PKR) hematopoietic cell kinase
(Hck) and P38
mitogen-activated protein kinase The protein kinases are induced by double-
stranded RNA (dsRNA) or possibly by the damaged ribosomal peptides (Iordanov et al 1997
Shifrin and Anderson 1999 Zhou et al 2003b Pestka 2007) Subsequent phosphorylation of
kinases (eg PKRHckP38
) activates the MAPK cascade resulting in ribotoxis stress
response upregulation of proinflammatory cytokines and dysregulation of neuroendocrine
signaling (Pestka and Smolinki 2005 Pestka 2010) Studies suggested that high doses of
DON induces ribotoxis stress response (apoptosis) preceded by immunosuppression and
disruption of intestinal permeabilitytissue functions (Yang et al 2000 Pestka 2008) Further
study observed stabilization of the mRNA and expression of transcription factors for critical
genes when cells were exposed to a low concentration of DON (Chung et al 2003)
The MAPK pathway proceeds with upregulation of a broad range of proinflammatory
cytokine genes and dysregulation of neuroendocrine signaling within the central and enteric
nervous systems Modulation of neuroendocrine signaling by DON increased the level of
serotonin resulted in vomiting reduced food intake and growth retardation in experimental
animalscell lines (Prelusky and Trenholm 1993 Li 2007) Upregulation of proinflammatory
cytokines and chemokines genes was determined in mice liver spleen kidney and lung
25
following in 1 h of exposure to DON A subsequent shock-like effect was observed in mice
because of a ldquocytokine stormrdquo (Pestka 2008) Induction of proinflammatory cytokines also led
to various cellular effects on the central nervous system interference of the innate immune
system and growth hormone signaling These contribute to anorexia growth retardation
aberrant immune response and tissue dysfunction as demonstrated in experimental animal
systems (Pestka 2010)
DON-mediated growth retardation in mice via suppression of growth hormones
(insulin-like growth factor 1 and insulin-like growth factor acid-labile subunit) has been
reported by Amuzie et al (2009) Disruption of gut tissues by DONNIV and subsequent
impairment of intestinal transporter activity was demonstrated in a human epithelial cell
model (Marseca et al 2002) The study proposed that alteration of gut tissue permeability
may cause gastroenteritis induction and growth retardation in livestock animals Growth
retardation of livestock may also occur via dysregulation of neuroendocrine signaling-
mediated induction of emesis and anorexia (Pestka et al 2010)
The molecular mechanism of TC toxicity in plant systems has not been well
documented A recent study showed the upregulation of numerous defense related genes in
wheat upon exposure to DON (Desmond et al 2008) The study further suggested that DON-
induced program cell death in wheat occurred through a hydrogen peroxide signaling
pathway This mechanism had not been observed in mammals
26
18 Strategies to control FHB and trichothecenes
During the past decades many pre- and post-harvest strategies have been applied to
protect cereal crops from Fusarium infestation and to eliminate TC from contaminated
agricultural commodities (FAO 2001 Munkvold 2003 Varga and Toth 2005 Kabak and
Dobson 2009) Some of the above crop protection techniques have been patented (He and
Zhou 2010) In spite of those efforts TC contamination of cereals appears to be unavoidable
using the currently available methods
Pre-harvest control strategies involving cultural practices (eg crop rotation tillage
early planting) and fungicide applications partially reduced Fusarium infection and DON
contamination in cereals (Dill-Macky and Jones 2000 Schaafsma et al 2001) However
repeated application of fungicides may lead to the evolution of Fusarium strains resistant to
fungicides with greater mycotoxin production ability (DrsquoMello et al 1998) At post-harvest
reduction of grain moisture by artificial drying storage at low temperatures (1-4degC) and
proper ventilation of the storage room have shown to be effective in inhibiting fungal growth
and mycotoxin production in infected grain (Midwest Plan Service 1987 Wilcke and Morey
1995) This technology has become less applicable particularly in developing countries
because of the frequent observation required to maintain proper storage conditions (Hell et al
2000)
Removal of Fusarium infected grains by physical methods (eg mechanical
separation density segregation colour sorting etc) was unable to eliminate TC completely
(Abbas et al 1985 He et al 2010b) This is partly because FHB symptomless kernels may
contain a considerable amount of DON which are not eliminated by physical separation
procedures (Seitz and Bechtel 1985) Several studies showed that milling and food processing
27
steps were unable to completely remove DON from grain-based foods (Jackson and
Bullerman 1999 Placinta et al 1999 Kushiro 2008) Detoxification of TC by chemical
treatments (eg ozone ammonia chlorine hydrogen peroxide and sodium metabisulfite) has
been demonstrated (Young et al 1986 Young et al 1987 Young et al 2006) The problem
with these techniques is that the application of such chemicals is not economically viable and
it also has a negative impact on grain quality (Karlovsky 2011) Although the technique has
been popular and showed effectiveness to certain mycotoxins utilization of absorbents to
inhibit mycotoxin absorption by the livestock gastrointestinal tract appeared to be ineffective
for TC (Huwig et al 2001)
The introduction of Fusarium resistant cultivars would be the best strategy to address
the FHB and associated mycotoxin problems in cereals However breeding for cultivars
resistant to FHBGER is difficult and time consuming as natural resistance against Fusarium
is quantitative in nature FHB resistant quantitative trait loci (QTLs) have been identified in
several cereal genotypes (eg Sumai 3 Ning 7840 Bussard) (Bernando et al 2007 Golkari
et al 2009 Haumlberle et al 2009 Yang et al 2010) The 3B 5A and 6B QTLs for type-I
(resistant to initial infection) and type-II (resistant to fungal spread in the kernel) resistances
have been identified on different wheat chromosomes (Buerstmayr et al 2009) Type-I
(Qfhsifa-5A) and type-II (Fhb1Fhb2-3B) QTLs from Sumai 3 are being used in many
breeding programs and pyramiding of the QTLs in spring wheat showed additional effects
Nonetheless the key genes involved in FHB resistance in cereals are yet to be identified This
makes it difficult to integrate the resistance genes into elite cultivars
28
Cultivar development by direct incorporation of TCDON resistance genes may be an
alternative way of protecting grains from these mycotoxins contaminations Many genes from
distantly related species have been integrated into crop cultivars using a transgenic approach
with great economic benefits (Brookes and Barfoot 2006) DON transforming trichothecene
3-O-acetyltransferase (TRI101 from Fusarium sporotrichioides) and UDP-
glucosyltransferase (from Arabidopsis thaliana) genes expressed in cereal cultivars showed
some DONDAS detoxifying ability under in vitro and controlled conditions (Kimura et al
1998 Okubara et al 2002 Poppenberger et al 2003 Ohsato et al 2007) By contrast no
reduction of mycotoxins in either of the above cases was observed under field conditions
(Manoharan et al 2006) Importantly the transformed acetylated and glycosylated
compounds could revert to the original substances or convert to even more toxic products
(Gareis et al 1990 Alexander 2008)
19 Isolation of DONTC de-epoxidating micro-organisms and genes
An enzyme with the capability of de-epoxidizing DONTC appears to be a potential
source for completely removing TC from contaminated cereals (Karlovsky 2011) However
such trichothecene detoxifying genes do not exist in cereals and other plants (Boutigny et al
2008) Because of this several research groups have attempted to obtain microorganisms that
encode DONTC de-epoxidation genes Mixed microbial culture capable of de-epoxydizing
trichothecenes have been isolated from chicken gut (He et al 1992) cow ruminal fluid (King
et al 1984 Swanson et al 1987 Fuchs et al 2002) pig gut (Kollarczik et al 1994) and
digesta of chicken (Young et al 2007) rats (Yoshizawa et al 1983) sheep (Westlake et al
1989) and fish (Guan et al 2009) All of the microbial cultures cited above showed de-
29
epoxydation capabilities only under anaerobic conditions realtively high temperatures and at
lowhigh pH except microbes from fish intestine that showed efficient de-epoxidation at low
temperatures and wide range of temperatures (Guan et al 2009) Such enzymes active under
anaerobic conditions high temperatues and low pH may have potential applications as feed
additives and the eubacterial strain BBSH 797 is an animal feed additive product with the
brand name Mycofixreg by BIOMIN (He et al 2010b) Biotechnological applications of
anaerobically-active enzymes from the above microbial sources seem to be inappropriate
under the aerobic and moderate conditions at which cereals plants are grown
Most of these efforts have failed to isolate single DONTC degrading microbes
effective under aerobic conditions because of inappropriate microbial growth conditions
under laboratory conditions Earlier attempts were made to obtain TC de-epoxidizing aerobic
bacteria but the culture did not retain DONNIV de-epoxidation activity after reculturing
(Ueno 1983 He et al 1992) In addition the co-metabolic nature of the microbial TC de-
epoxidation reaction has limited the use of mycotoxins as the sole nutrient source for selective
isolation of the target microorganisms Several DONTC de-epoxidation microbial strains
including Bacillus strain LS100 and Eubacterial strain BBSH 797 have been isolated from
chicken gut and rumen fluids through multi-step enrichment procedures (Fuchs et al 2002
Yu et al 2010) A DON to 3-keto DON transforming bactial strain was islated from soil by
prolonged incubation with high concentration DON (Shima et al 1997)
Microbial genesenzymes with TC de-epoxidation activity are not known Based on
the microbial TC de-epoxidation activity of a mixed microbial culture it has been proposed
that ceratin reductases or oxidoreductases may catalyze such a biochemical reaction (Islam et
al 2012) With the advent of molecular techniques many novel genes with
30
industrialagricultural value have been cloned by reverse genetics (genomic DNA library
preparation and functional screening of the library clones) forward genetics (transposon
mutagenesis) and comparative genomics methods (Frey et al 1998 Koonin et al 2001
Suenaga et al 2007 Chung et al 2008 Ansorge 2009) The tremendous advancement of next
generation sequencing technology and bioinformatics tools may facilitate the identification of
the microbial genes responsible for DONTC detoxification (Ansorge 2009) Isolation of TC
de-epoxidation genes may play a major role in producing safe foods and feeds derived from
grains
110 The bacterial species Citrobacter freundii
Citrobacter freundii is a bacterial species belonging to the family Enterobacteriace It
is a facultatively anaerobic and Gram-negative bacillus (Wang et al 2000) The bacterium
survives almost everywhere including animalhuman intestines blood water and soils
(Wang et al 2000 Whalen et al 2007) C freundii is known as an opportunistic human
pathogen and causes a variety of respiratory tract urinary tract and blood infections and
responsible for serious neonatal meningistis disease (Badger et al 1999 Whalen et al 2007)
Interestingly this bacterium plays a positive environmental role through nitrogen recycling
(Puchenkova 1996) Strains of C freundii also contribute to the environment by sequenstering
heavy metals and degrading xenobiotic compounds (Kumar et al 1999 Sharma and Fulekar
2009)
31
111 Hypotheses and objectives
The study was based on the following hypotheses
i Certain microorganisms in DON contaminated agricultural soils are able to transform DON
to de-epoxy DON (de-epoxidation) under aerobic conditions and moderate temperatures
ii DON de-epoxidation may be achieved by a single microbial strain
iii Microbial DON detoxifying genes and associated regulators are able to express and
function in a closely related host organism
iv DON detoxifying microbial strains carry genes for de-epoxidation activity that are absent
from related species that have not evolved under DON selection pressure
The objectives of this research were to
i Select microbial cultures from soil having DON de-epoxidation activity under aerobic
conditions and moderate temperatures by analyzing potential DON contaminated soils in
various growth conditions namely aerobicanaerobic conditions different growth media
temperature and pH
ii Select for organisms with DON de-epoxidation activity from the mixed microbial culture
by treating with antibiotics heating subculturing on agar medium and incubating for a
prolonged period in media with a high DON concentration
iii Identify microbial de-epoxidation genes by producing a microbial genomic DNA library
from the DON de-epoxidizing strain followed by screening of the clones for DON de-
epoxidation
32
iv Determine microbial de-epoxidation genes by sequencing of the genome and comparative
analysis of the genome with closely related species and identification of putative DON
detoxification genes
33
CHAPTER 2
20 AEROBIC AND ANAEROBIC DE-EPOXIDATION OF MYCOTOXIN
DEOXYNIVALENOL BY BACTERIA ORIGINATING FROM AGRICULTURAL
SOIL
Published in World Journal of Microbiology and Biotechnology 2012 287-13
21 Abstract
Soil samples collected from different cereal crop fields in southern Ontario Canada
were screened to obtain microorganisms capable of transforming deoxynivalenol (DON) to
de-epoxy DON (dE-DON) Microbial DON to dE-DON transformation (ie de-epoxidation)
was monitored by using liquid chromatography-ultraviolet-mass spectrometry (LC-UV-MS)
The effects of growth substrates temperature pH incubation time and aerobic versus
anaerobic conditions on the ability of the microbes to de-epoxidize DON were evaluated A
soil microbial culture showed 100 DON to dE-DON biotransformation in mineral salts
broth (MSB) after 144 h of incubation Treatments of the culture with selective antibiotics
followed an elevated temperature (50ordmC) for 15 h and subculturing on agar medium
considerably reduced the microbial diversity Partial 16S-rRNA gene sequence analysis of the
bacteria in the enriched culture indicated the presence of at least Serratia Citrobacter
Clostridium Enterococcus Stenotrophomonas and Streptomyces The enriched culture
completely de-epoxidized DON after 60 h of incubation Bacterial de-epoxidation of DON
occurred at pH 60 to 75 and a wide array of temperatures (12 to 40ordmC) The culture showed
34
rapid de-epoxidation activity under aerobic conditions compared to anaerobic conditions This
is the first report on microbial DON to dE-DON transformation under aerobic conditions and
moderate temperatures The culture could be used to detoxify DON contaminated feed and
might be a potential source for gene(s) for DON de-epoxidation
Keywords biotransformation de-epoxidation deoxynivalenol soil bacteria vomitoxin
22 Introduction
Deoxynivalenol (DON commonly known as lsquovomitoxinrsquo) 3α 7α 15-trihydroxy-
1213-epoxytrichothec-9-en-8-one is a trichothecene produced by toxigenic Fusarium species
causing Fusarium head blight (FHB) or Gibberella ear rot (GER) disease on small grain
cereals and maize respectively (Starkey et al 2007 Foroud and Eudes 2009) DON
contamination in cereals and maize is widespread and can lead to tremendous losses to the
growers It has also detrimental effects on human and livestock health (Pestka and Smolinski
2005 Rocha et al 2005) The toxin interferes with the eukaryotic ribosomal peptidyl
transferase (Rpl3) activity resulting in human and animal mycotoxicosis (Ueno 1985 Bae
and Pestka 2008) Common effects of DON toxicity in animals include diarrhea vomiting
feed refusal growth retardation immunosuppression reduced ovarian function and
reproductive disorders (Pestka and Smolinski 2005 Pestka 2007) In plants DON causes a
variety of effects viz inhibition of seed germination seedling growth green plant
regeneration root and coleoptile growth (cited in Rocha et al 2005) In addition DON
enhances disease severity in sensitive host plants (Maier et al 2006)
35
Avoidance of DON contamination or reduction of toxin levels in contaminated grains
remains a challenging task Physical and chemical processes are either inappropriate or
prohibitively expensive to use to decrease the DON content of agricultural commodities
(Kushiro 2008 He et al 2010b) The use of Fusarium resistant crop varieties might be the
best method to reduce DON contamination However breeding for FHB or GER resistant
cultivars appears to be very difficult and time consuming as natural sources of resistance
against Fusarium-caused diseases is quantitative in nature (Ali et al 2005 Buerstmayr et al
2009) As an alternative expressing genes for enzymatic detoxification of DON in elite
cultivars may be an effective strategy for protecting the value of feeds and foods (Karlovsky
1999) Heterologous expression of eukaryotic trichothecene 3-O-acetyltransferase and UDP-
glucosyltransferase genes in cereals and Arabidopsis have resulted in reduced toxicity via
acetylation and glycosylation of the DON (Poppenberger et al 2003 Ohsato et al 2007)
It has been proposed that enzymatic de-epoxidation of trichothecene toxins could be
used to limit DON contamination (Zhou et al 2008 Foroud and Eudes 2009) DON belongs
to type B-trichothecenes which contain an epoxide ring at C12-13 and the epoxide is highly
reactive and mainly responsible for toxicity (Figure 2-1) (Swanson et al 1988 Eriksen et al
2004)
36
Figure 2-1 Mechanism of microbial DON de-epoxidation Removal of the oxygen atom from
the epoxide ring results in formation of a C=C double bond at C-12 position which reduces
cytotoxicity (Swanson et al 1988 Eriksen et al 2004)
Microbial DON de-epoxidation by single bacterial strains and by mixed cultures has
been reported (King et al 1984 Swanson et al 1987 He et al 1992 Kollarczik et al 1994
Fuchs et al 2002 Young et al 2007 Guan et al 2009 Yu et al 2010) However all of these
microbes originated from fish and other animals and were strictly anaerobic The recovered
microbial cultures and single strains only showed de-epoxidation of trichothecenes at low
(le15ordmC) and high (gt30degC) temperatures Such enzymes might have limited applications for
toxin decontamination in cereal grains The aim of the current study was to isolate the
microbes from soil capable of transforming DON to a de-epoxy metabolite under aerobic
conditions and moderate temperatures
De-epoxydation
Deoxynivalenol (DON)
De-epoxyDeoxynivalenol (dE-DON)
37
23 Materials and methods
231 Soil samples
The upper layer (0-30 cm) of soils along with crop debris were randomly collected
from cereal (eg maize wheat barley) crop fields during 2004-2006 in Southern Ontario
Canada Detailed of the crops and locations of the fields have been described in Jianwei Hersquos
Master Degree thesis (He 2007) Briefly soil samples were collected in sterile plastic bags
and transported to the laboratory Samples were sieved to remove large roots stones and
stored at 4ordmC until used The samples were mixed with grounded mouldy corn (containing 95
microgg DON) + F graminearum and incubated at 22-24degC for six weeks Five samples were
prepared for DON de-epoxidation activity analysis by dispersing 5 g soil in 50 mL distilled
water and storing at4degC until further use
232 Culture media
Media used in the study were nutrient broth (NB) Luria Bertani (LB) minimal
medium (MM) (He 2007) six mineral salts + 05 bacto peptone (MSB) (He 2007) tryptic
soy broth (TSB) corn meal broth (CMB) (Guan et al 2009) potato dextrose broth (PDB) six
mineral salts + 05 yeast extract broth (MSY) and soil extract (SE) MM was composed of
sucrose-100 g K2HPO4-25 g KH2PO4-25 g (NH4)2HPO4-10 g MgSO47H2O-02 g
FeSO4-001 g MnSO47H2O- 0007 g water- 985 mL and MSB was composed of the same
salts as MM but was supplemented with Bactotrade Peptone (05 wv) as the sole C source
instead of sucrose To prepare soil extract 100 g of the composite soil was homogenized in 1
L water The soil particles and plant debris were removed by passing the soil suspension
through Whatmanreg grade 3 filter paper (Sigma-Aldrich Oakville Ontario Canada) The
38
extract was autoclaved at 121degC for 30 min Media and ingredients were purchased either
from Fisher Scientific Inc (Fair Lawn New Jersey USA) Sigma-Aldrich (Oakvile Ontario
Canada) or Difco (Sparks Maryland USA) Prior to autoclaving pH of the broth media was
adjusted to 70 Solid media were prepared by adding 15 agar
233 Screening of soil samples
Frozen soil suspensions were thawed and vortexed to homogenize the microbial
population An aliquot (100 microL) of the suspension was immediately transferred to MSB PDB
and MM broth media DON at 50 microgmL was added to the microbial suspension cultures The
cultures were incubated at 22-24degC with continuous shaking at 180 revolutions per minute
(rpm) The extent of DON de-epoxidation was determined at 24 h 48 h 72 h 96 h 120 h and
144 h of incubation
234 Determination of de-epoxidation product
Microbial DON de-epoxidation was measured according to Guan et al (2009)
Samples were prepared by mixing equal volume of microbial culture and methanol and
allowed to stand for 30 min at room temperature Microorganisms in the mixture were
removed by passing them through a 045 microm Millipore filter (Whatmanreg Florham Park New
Jersey USA) The filtrates were used for de-epoxidation analysis by a liquid chromatography-
ultraviolet-mass spectrometry (LC-UV-MS) Identification and quantification of DON and its
transformation product was achieved using a Finnigan LCQ DECA ion trap LC-MS
instrument (ThermoFinnigan San Jose CA USA) The equipment detects 30-2000 mz
(molecular mass ion ratio) compounds and able to determine 001 micromL DON The HPLC
39
system was equipped with a Finnigan Spectra System UV6000LP ultraviolet (UV) detector
and an Agilent Zorbax Eclipse XDB C18 column (46times150 mm 35 μm) The binary mobile
phase consisted of solvent A (methanol) and solvent B (water) The system was run with a
gradient program 25 A at 0 min 25ndash41 A over 5 min isocratically at 41 A for 2 min
and 41ndash25 A over 1 min There was a 3 min post-run at its starting condition to allow for
the equilibration of the column The flow-rate was set at 10 mLmin UV absorbance was
measured at 218 nm Atmospheric pressure chemical ionization (APCI) positive ion mode
was used for MS detection A DON standard was used to adjust the instrument to its
maximum response The system was operated as follows shear gas and auxiliary flow-rates
were set at 95 and 43 (arbitrary units) voltages on the capillary tube lens offset multipole 1
offset multipole 2 offset and entrance lens were set at 3000 minus1700 minus370 minus850 minus3500
and minus4200 V respectively capillary and vaporizer temperatures were set at 200 and 450degC
respectively and the discharge needle current was set at 7 μA Identification of mycotoxin
was achieved by comparing their retention times UV spectra and mass spectrum with DON
and de-epoxy DON (Biopure Union MO USA) as standards Quantification of DON and
product was based on peak areas related to their UV and MS calibration curves
40
235 Biological DON de-epoxidation
To help identify the types of microorganisms causing the DON de-epoxidation MSB
medium was supplemented with either 100 microgmL penicillin G or streptomycin sulfate for
suppressing gram positive or negative bacterial growth and activities respectively Three
replications of antibiotic-treated media were inoculated with 100 microL of microbial culture
capable of de-epoxidizing DON The antibiotic-treated cultures were incubated at 27degC
spiked with 50 microgmL DON Controls were prepared without adding antibiotics in the
microbial culture Inhibition of DON de-epoxidation was analyzed after five day of
incubation
236 Reduction of microbial diversity by antibiotic treatments heating and
subculturing on agar medium
In order to reduce non-DON degrading microbial populations the DON de-
epoxidation capable cultures were sequentially treated with antibiotics having diverse modes
of action including i) dichloramphenicol that kills various gram positive Gram-negative and
anaerobic bacteria ii) trimethoprim that kills Gram-positive and negative bacteria iii)
penicillin that is lethal to Gram-positive bacteria and iv) cyclohexamide to kill filamentous
fungi and yeasts Antibiotics were purchased from Sigma-Aldrich Antibiotic stock solutions
(10 mgmL) were prepared with sterile water or methanol Microbial cultures in MSB
medium were treated with 100 microgmL of each antibiotic followed by incubation of the culture
at 50ordmC for 15 hours to kill undesirded microbial species After heating cultures were treated
with 50 microgmL DON and incubated at 27ordmC for five days DON de-epoxidation activity of the
microbial cultures was determined following the procedure described above Cultures that
41
showed gt99 de-epoxidation were considered for subsequent antibiotic and heat treatments
Aliquots of the achieved culture were spread on MSB agar plates and grown at room
temperature for few days Colonies appeared on the plates were collected and incubated in
MSB medium supplemented with 50 microgmL DON De-epoxidation ability of the subcultured
microbial cultures was determined after five days of incubation at 27degC by LC-UV-MS
method
237 Analysis of microbial population diversity
Terminal Restriction Fragment Length Polymorphism (T-RFLP) analysis
Initially microbial diversity was determined by terminal restriction fragment length
polymorphism (T-RFLP) fingerprint analysis (Bruce and Hughes 2000) Genomic DNA from
microbial cultures with and without enrichment was extracted using DNeasy Plant DNA
Extraction Mini Kit (Qiagen Inc Mississauga Ontario Canada) 16S-rDNA was amplified
by PCR using the HEX-labeled forward primer Bac8f
(5-agagtttgatcctggctcag-3) and
unlabeled reverse primer Univ1492r (5-ggttaccttgttacgactt-3) (Fierer and Jackson 2006)
Fifty microlitter PCR mixture for each sample was prepared having 1x HotStarTaq Master
Mix (Qiagen) 05 microM of each primer 50 microg of BSA and 50 ng of DNA The 35 PCR cycles
followed 60 s at 94degC 30 s at 50degC and 60 s at 72degC After size verification by agarose-gel
electrophoresis 50 ng of purified PCR products was digested for 3 h in a 20 microL reaction
mixture with 5 U of HhaI restriction enzyme (New England Biolabs Ipswich MA USA)
The size and relative abundance of the labeled T-RFLP fragments were separated by capillary
electrophoresis in an automated DNA sequencer equipped with fluorescence detector
(Applied Biosystem 3730 DNA Analyzer) The DNA fragments were analyzed with the Peak
42
Scannertrade Software v10 to create an electropherogram with the genetic fingerprints of the
microbial community
Analysis of V3-V5 region of the 16S-rDNA
To define the bacterial genera present in the enrichment culture the 16S-rDNA was
amplified using two sets of bacterial universal primers Primarily 16S-rDNA was PCR
amplified using the primers 46f-5rsquogcytaacacatgcaagtcga3rsquo (Kaplan and Kitts 2004) and
1371r-5rsquogtgtgtacaagncccgggaa3rsquo (Zhao et al 2007) PCR was performed in a total volume of
25 μL which contained 06 μM of each primer 1x of HotStarTaqreg Master Mix (Qiagen Inc)
and 20-40 ng of DNA The PCR cycles included the following steps 95ordmC (15 min) 35 cycles
of 94ordmC (20 s) 52ordmC (15 s) 72ordmC (1 min 30 s) and a 72ordmC (7 min) extension step The second
pair of universal bacterial primers were 27f-5rsquoagagtttgatcmtggctcag3rsquoand 1492r-
5rsquoggttaccttgttacgactt3rsquo (Lane 1991) PCR was conducted with 25 cycles following the
procedures of Tian et al (2009) Amplification was carried out in a GeneAmpreg PCR System
9700 (Applied Biosystems) Furthermore the PCR products were used as templates to
amplify the hypervariable V3-V5 region of the 16S-rDNA using primers 357f-
5rsquocctacgggaggcagcag3rsquo and 907r7 5rsquoccgtcaattcctttgagttt3rsquo (Yu and Morrison 2004)
After purification of the PCR products by QIAquick PCR purification kit two
amplicon libraries were prepared by ligating to pCR-TOPO vector (Invitrogen) and the library
clones were transformed into TOPO10 competent cells according to Invitrogenrsquos TOPO
cloning manual The clone libraries were plated on LB plates containing 25 μgmL
kanamycin Fifty colonies from each library were randomly picked up from the overnight
plates and inserts were sequenced as described by Geng et al (2006) PCR products were
43
sequenced with an automated ABI 3730 DNA analyzer (Applied Biosystems) The sequence
similarities of the amplified V3-V5 region were compared with the ribosomal database project
(RDP) (httprdpcmemsuedu) Sequences with greater than 95 identity were assigned to
the same bacterial genus
238 Factors influencing DON de-epoxidation
DON de-epoxidation ability of the enriched microbial culture was investigated under
different conditions as described below
Maintaince of active culture
To retain microbial de-epoxidation activity the active culture has been maintained in
MSB medium containing 50 microgmL DON
Media growth temperature and pH
Overnight microbial cultures (100 μL) were added to 2 mL of NB LB MM TSB
MSB CMB MSY PDB or SE broths The pH of the media was adjusted to 70 and the
experiments were conducted at 27degC temperature The effect of temperature was studied by
incubating the culture in MSB medium (pH 7) at 12 20 25 30 and 40ordmC To determine the
effects of pH MSB medium pH was adjusted to 55 60 65 70 75 and 80 Media with
different pHs were inoculated with aliquots of the overnight active culture and then incubated
at 27ordmC The cultures were supplemented with 50 μgmL DON before incubation Three
replications for each medium temperature and pH condition were evaluated
44
De-epoxidation under aerobic and anaerobic conditions
Three replicate microbial cultures of five mL each were grown in sterile 15 mL
centrifuge tubes under aerobic conditions and in an anaerobic chamber filled with 95 CO2
and 5 H2 (COY Laboratory Products Inc Ann Arbor MI USA) Three days after initiation
half (25 mL) of the cultures from anaerobic conditions was transferred to new centrifuge
tubes and brought to aerobic conditions Similarly half of the aerobically grown cultures were
placed into an anaerobic chamber Cultures moved from anaerobic to aerobic conditions were
shaken after opening the lids of the tubes in a laminar flow every eight hours intervals to
maintain aerobic conditions in the culture Lids of the culture tubes transferred to anaerobic
conditions were kept loose to allow oxygen to escape from the culture Two days after
transferring to the new conditions the cultures in both conditions were treated with 50 microgmL
DON and incubated at 27degC Every 12 h 500 microL were removed from each tube and analyzed
for microbial DON de-epoxidation activity by LC-UV-MS
239 Data analysis
Data were processed using Microsoft Excel Program 2007 Statistical analysis was
done using General Linear Model procedures in SAS version 91 (SAS Institute Inc Cary
NC USA 2002-03) The results were the average of three replications stated as mean plusmn
standard error Differences between means were considered significant at plt 005 according
to Tukeyrsquos-HSD test
45
24 Results
241 Analyses of DON to dE-DON biotransformation
A typical LC-UV-MS analysis where there is incomplete de-epoxidation of DON by a
particular soil suspension culture is shown in Figure 2-2 Analysis by LC with UV detection
and integration of peak areas showed 662 conversion of DON to dE-DON (Figure 2-2a)
Analysis LC with MS detection (expressed as total ion counts) indicated 643 conversion
(Figure 2-2b) LC analyses with UV and TIC (total ion count) detection are subject to
interferences due to co-eluting contaminants and therefore integration of small peaks was
very difficult making quantification inaccurate MS detection by SIM (selected ion
monitoring) can eliminate the influence of co-eluting contaminants For example the SIM LC
chromatogram for DON (Figure 2-2c) was generated by using the major ions resulting from
the APCI positive ion MS of DON (Figure 2-2e)
Similarly the SIM LC chromatogram for dE-DON (Figure 2-2d) was derived from the
MS ions of dE-DON (Figure 2-2f) The signal to noise ratios (SN) of peaks observed by SIM
were at least 20 times greater than those detected by TIC because there is reduced likelihood
of co-eluting peaks having the same ions Integration of the SIM peaks revealed a 616
conversion The three methods gave similar results but the SIM LC results were considered
to be the most accurate Using that method the conversion of DON to dE-DON by the mixed
culture occurred most quickly at 48 h and was completed by 144 h (Figure 2-3)
46
47
Figure 2-2 LC-UV-MS chromatographic analysis of DON and its de-epoxidated product dE-
DON obtained by culturing with selected soil microbes (a) LC-UV chromatogram of product
mixture monitored at 218 nm (b) LC-MS total ion chromatogram (TIC) of product mixture
(c) selected ion monitoring (SIM) chromatogram of cultured DON at mz 231 249 267 279
and 297 (d) SIM chromatogram of cultured dE-DON at mz 215 233 245 251 263 and 281
(e) atmospheric pressure chemical ionization (APCI) positive ion mass spectrum of DON (f)
APCI positive ion mass spectrum of dE-DON
48
242 Screening of soil samples
Initially samples originating from soils were analyzed in three types of culture media
to support the growth of fungi yeast and bacteria However cultures growing in PDB which
is a suitable substrate for many fungi and yeasts did not permit transform DON to dE-DON
(data not shown by microorganisms) A define medium (ie MM) was also used in the
sample screening assay None of the soil samples showed de-epoxidation activity in MM
(data not shown) From five soil samples one soil sample culture showed de-epoxidation of
DON in MSB The culture in MSB converted gt99 DON to dE-DON within 144 h (Figure 2-
3) Conversion began after a lag of 48 h and was linear thereafter with a rate of 1 per hour
49
Figure 2-3 Time course of relative biotransformation of DON to dE-DON by the initial soil
suspension culture in MSB medium The culture contained 50 μgmL DON at time 0 Culture
with three replications was incubated at room temperature with continuous shaking at 180
rpm The conversion of DON to dE-DON was analysed by LC-UV-MS at 24 h intervals Blue
and red lines are the percent conversion of DON and recovery of dE-DON respectively
50
243 Evidence of biological DON de-epoxidation
Treatment of active microbial culture with 100 microgmL of streptomycin completely
inhibited DON de-epoxidation In contrast the same concentration of penicillin G did not
affect activity of the culture (Figure 2-4) Streptomycin is effective against Gram-negative
bacteria while penicillin G is lethal to Gram-positive bacteria These experiments provided
evidence that DON de-epoxidation may be caused by certain Gram-negative bacteria
51
Figure 2-4 Microbial de-epoxidation of DON after five days of incubation The active DON
de-epoxidation culture was treated with either 100 microgmL penicillin G or streptomycin
(Strepto) in MSB Cultures having three replications were added 50 microgmL DON and
incubated at room temperatures with continuous shaking at 180 rpm For controls cultures
were without antibiotics DON to dE-DON transformation was monitored after five day of
incubation by LC-UV-MS method
0
20
40
60
80
100
120
Penicillin Strepto Control
DO
N d
e-e
po
xyd
ati
on
(
)
Antibiotics
52
244 Reduction of the microbial diversity
Plating the suspension cultures onto MSB agar revealed colonies with a variety of
sizes morphologies and colours reflecting a mixed culture (data not shown) To reduce the
complexity of the culture three steps of selection pressures were applied namely treatment
with antibiotics heat shocks and subculturing on solid medium The above indicated
treatments did not significantly affect DON de-epoxidation of the microbial cultures (data not
shown) T-RFLP profiles of the mixed culture before treatments showed six distinct peaks of
about 110 150 200 220 340 and 420 bp restriction fragments with the 110 bp peak being
the predominant one (Figure 2-5a)
After treatment with antibiotics and heat only three peaks at 110 150 and 325 bp
remained (Figure 2-5b) suggesting that there was a reduction in the microbial diversity
without affecting DON de-epoxidation After enrichments the bacteria species in the culture
changed their growth patterns as the electropherogram produced different sizes of pick
lengths compared to untreated culture
53
(b)
(a)
Rela
tive flu
ore
scen
ce
32000
0
0 500
16S-rDNA fragment size (bp)
100 200 300 400
Rela
tive flu
ore
scen
ce
32000
0
500
16S-rDNA fragment size (bp)
0 300200100 400
54
Figure 2-5 Analysis of microbial population reduction by terminal restriction fragment
length polymorphism (T-RFLP) (a) Restriction fragments of the 16S-rDNA of the untreated
culture and (b) fragmentation patterns of the enriched culture Picks along the X-axis showed
the fragmentation patterns of the 16S-rDNA which is related to the bacterial species diversity
The length of picks (along the Y-axis) showed abundance of terminal restriction fragments
related to the abundance of particular bacterial speciesgenera
55
245 Microbial population structures in the enriched culture
Microbial diversity in the enriched culture was determined by analyzing the sequences
of the V3-V5 region (~586 bp) amplified from the 16S-rDNAs of the microorganisms it
contained The sequence analysis showed the presence of Serratia Citrobacter Clostridium
Enterococcus Stenotrophomonas and Streptomyces in the enriched culture (Figure 2-6)
Out of 100 V3-V5 sequences 54 sequences were found to be belonging to Serratia
the predominant species of the culture The remaining sequences matched with 12
Citrobacter 8 Clostridium 14 Enterococcus 8 Stenotrophomonas and 4 Streptomyces
species
56
Figure 2-6 Microbial genera identified in the enriched DON de-epoxidizing microbial
culture The bacterial genera were determined by comparing the hypervariable regions (V3-
V5) of the 16S-rDNA using ribosomal database project (httprdpcmemsuedu) X-axis
showed the identified bacterial genera and Y-axis demonstrated the abundance of the V3-V5
sequences belonging to different bacterial genera
54
12 8
14
8 4
0
20
40
60 S
eq
uen
ce
fre
qu
en
cy
Microbial genera
57
246 Effects of media growth temperature pH and oxygen requirements on DON de-
epoxidation
Of the nine media that were tested DON de-epoxidation by the enriched microbial
culture only occurred in NB and MSB (Figure 2-7a) It appeared that C andor N
concentrations in the media were crucial for the enzymatic activity because less or more than
05 Bacto Peptone in MSB reduced the microbial DON transformation ability
DON de-epoxidation activity occurred over a wide range of temperatures from 12 to
40ordmC (Figure 2-7b) The fastest rates of DON de-epoxidation occurred between 25 to 40ordmC
De-epoxidation was observed within a relatively narrow range of pH (60 to 75) with
the most rapid DON biotransformation activity observed at pH 70 (Figure 2-7c) At pH 70
the enriched culture showed DON de-epoxidation activity 125 times faster than at pH 60 or
75 At pH 55 and 80 there was no observable DON de-epoxidation
58
0
20
40
60
80
100
120
12degC
20degC
25degC
30degC
40degC
DO
N d
e-e
po
xyd
ati
on
(
)
Temperature
0
20
40
60
80
100
120
DO
N d
e-e
pxyd
ati
on
(
)
Media
(a)
(b)
59
Figure 2-7 Effects of culture conditions on DON to de-epoxy DON transformation by the
enriched microbial culture For the media experiment (a) all the media were adjusted to pH 7
and the incubation temperature was 27degC For the temperature experiment (b) MSB pH 7 was
used For the pH experiment (c) MSB at 27degC was used Microbial cultures were inoculated
with 50 μgmL DON De-epoxidation was determined after 72 h of incubation Media MSY
(mineral salts + 05 yeast extract) LB (Luria Bertani) TSB (tryptic soy broth) MSB
(mineral salts + 05 bacto peptone) SE (soil extract) CMB (corn meal broth) PDB (potato
dextrose broth) and MM (minimal medium)
0
20
40
60
80
100
120
55
60
65
70
75
80
DO
N d
e-e
po
xyd
ati
on
(
)
pH
(c)
60
The enriched culture showed DON de-epoxidation activity under both aerobic and
anaerobic conditions (Figure 2-8) The fastest transformation (100 de-epoxidation after 60 h
of incubation) occurred in cultures (A) that were grown under aerobic conditions throughout
the culture growth period A medium rate of de-epoxidation (100 de-epoxidation after 72 h)
was observed in cultures that were initially grown in either aerobic or anaerobic conditions
followed by transferring to anaerobic or aerobic conditions (A+AN and AN+A) respectively
The lowest rate occurred (100 de-epoxidation after 96 h) in cultures grown in continual
anaerobic conditions (AN) The means comparison of microbial DON de-epoxidation
efficiencies at 60 h of incubation showed that cultures continually grown in conditions A and
AN were statistically different Biotransformation abilities of the cultures that were switched
from one conditions to another (A+AN and AN+A) were statistically similar among each
other but different from cultures that were only grown in conditions A or AN (plt005)
61
Figure 2-8 Microbial DON to dE (de-epoxy DON)-DON transformation activity under
aerobic and anaerobic conditions A = culture incubated and examined the activity only under
aerobic conditions A+AN = culture initially was grown in aerobic conditions and then
transferred to anaerobic conditions AN = cultures grown throughout the assay under
anaerobic conditions AN+A = culture initially grown under anaerobic conditions and then
transferred to aerobic conditions Differences of lower case letters are significantly different
in de-epoxidation capability (after 60 h) among incubation conditions according to Tukeyrsquos
HSD test (p lt 005) The experiment was conducted with three replications
62
25 Discussion
The current study describes a novel bacterial culture isolated from agricultural soil that
was capable of transforming DON to de-epoxy DON under aerobic and anaerobic conditions
at moderate temperatures Because it has been speculated that DON translocates to soil from
contaminated crops and DON that enters into the soil could be metabolized by microbes
(Voumllkl et al 2004) soil samples for the current study were mainly collected from fields where
cereal crops contaminated with DON were grown
In our initial sample screening steps upwards of 144 h of incubation were required to
achieve gt99 DON de-epoxidation Such a long period may have been due to the presence of
fast growing microorganisms that had suppressive effects on the growth or activity of the
DON transforming bacteria (Davis et al 2005) The sequential use of different antibiotics
with different modes of actions followed by thermal treatments and subculturing on solid
medium resulted in a reduction in the number of bacterial species as determined by T-RFLP
analysis Analysis of V3-V5 region of the 16S-rDNA showed that five bacterial genera were
present in the enriched culture The culture showed higher DON de-epoxidation activity (only
60 h required for gt99 biotransformation) compared to initial soil suspension culture that
required 144 h to complete the de-epoxidation This microbial culture could be used to isolate
the DON de-epoxidizing gene(s) applying metagenomics approaches (Daniel 2005)
The culture showed complete de-epoxidation activity both in MSB and NB media
However we have found that the use of nutritionally rich NB medium enhanced non-target
bacterial (eg Serratia lequifaciences) growth but this did not occur in MSB which is a
comparably poor medium To avoid the possible suppressive and detrimental effects of the
fast growing bacteria on the target bacteria MSB was used for enrichment and
63
characterization of the culture An important characteristic of the DON de-epoxidation
activity observed for the microbial culture in the current study is its activity at moderate
temperature (25-30degC) In contrast previously isolated cultures showed optimal activities at
low (le15degC) or high temperatures (37degC) (Guan et al 2009) Also the culture showed DON
de-epoxidation activity at neutral pH range (pH 60-75) in contrast to previous reports which
found that microbial trichothecene de-epoxidation occurred at acidic and basic pHs (Swanson
et al 1987 He et al 1992 Kollarczik et al 1994 Young et al 2007) However a microbial
culture from fish gut showed de-epoxidation under wide range of pHs (Guan et al 2009)
Probably the pH range reflects the environment in which the bacteria naturally grow which in
this case is similar to the soil pH of a typical crop field in Southern Ontario of Canada The
advantage of the present finding is that DON de-epoxidation activity of the culture was
optimal in a temperature range that matches the temperature range of activity for most arable
plants Therefore it is reasonable to suggest that the gene(s)enzyme(s) responsible for the
activity have potential application for enzymatic detoxification of DON in cereals (Karlovsky
1999)
The current observation that bacterial DON de-epoxidation activity occurred under
both aerobic and anaerobic conditions was interesting because DON de-epoxidation is a
reductive chemical reaction that might be expected to be promoted by anaerobic conditions
However there are previous reports of aerobic de-epoxidation of eldrindieldrin by soil
bacteria (Matsumoto et al 2008) and Goodstadt and Ponting (2004) described aerobic
reductive de-epoxidation by vitamin K epoxide reductasephylloquinone reductase A
definitive determination of the oxygen sensitivity of the enzyme responsible for DON de-
epoxidation will ultimately come from experiments with the purified protein
64
In conclusion this study reports on a novel bacterial source from soil that efficiently
de-epoxidizes DON at moderate temperatures under both aerobic and anaerobic conditions
This research may lead to the isolation of microbes and gene(s) encoding enzyme(s) capable
of de-epoxidizing DON that could create transgenic plants with reduced DON contamination
thus improving food and feed safety and reducing negative effects of the toxin on public
health
26 Acknowledgements
The authors gratefully acknowledged the Natural Sciences and Engineering Research
Council of Canada (NSERC) for awarding a scholarship to Rafiqul Islam for PhD program
The research was supported by the Agriculture and Agri-Food Canada Binational
Agricultural Research and Development Fund (AAFCBARD) project We are thankful for
the contributions of Jianwei He Xiu-Zhen Li and Honghui Zhu
65
CHAPTER 3
30 ISOLATION AND CHARACTERIZATION OF A NOVEL SOIL BACTERIUM
CAPABLE OF DETOXIFYING ELEVEN TRICHOTHECENE MYCOTOXINS
31 Abstract
Deoxynivalenol (DON vomitoxin) is one of the most commonly detected food and
feed contaminating mycotoxins worldwide In order to develop a control method effective
under aerobic conditions and moderate temperatures a bacterial strain ADS47 was isolated
capable of transforming DON to the less toxic de-epoxy DON The bacterial strain is
facultatively anaerobic Gram-negative rod shaped with an average cell size of 150 X 072
microm and has peritrichous (multiple) flagella The 16S-rRNA gene sequence had gt99
nucleotide identity to Citrobacter freundii a bacterium that belongs to the gamma subclass of
proteobacteria Strain ADS47 showed near 100 de-epoxidation of DON after 42 h of
incubation in nutrient broth (NB) under aerobic conditions moderate temperature (28˚C) and
neutral pH (70) However de-epoxidation could occur at 10-45˚C and pH 60-75 Under
anaerobic conditions near 100 de-epoxidation was observed after 72 h mainly during the
stationary growth phase Microbial DON de-epoxidation ability increased with increasing
DON concentrations in the culture media Inhibition of bacterial de-epoxidation by sodium
azide suggested that an active electron transport chain was required In addition to DON
strain ADS47 was able to de-epoxidize andor de-acetylate five type-A and five type-B
trichothecene mycotoxins This is the first report of a bacterial isolate capable of de-
epoxidizing DON and trichothecene mycotoxins under aerobic conditions and moderate
66
temperatures Strain ADS47 could potentially be used in bioremediation to eliminate DON
and other trichothecene mycotoxins from contaminated food and feed
Keywords aerobic de-epoxidation Citrobacter freundii isolation soil bacterium
trichothecenes
32 Introduction
Cereals are often contaminated with a variety of type-A and type-B trichothecene
mycotoxins produced by a number of toxigenic Fusarium species (Placinta et al 1999
Binder et al 2007 Foroud and Eudes 2009) Trichothecenes are tricyclic sesquiterpenes with
a double bond at the C9 10 position and a C12 13 epoxy group (Desjardins 2006) The two
types are distinguished by the presence of a ketone at position C8 in type-B which is absent
in type-A (Yazar and Omurtag 2008) The type-B toxin deoxynivalenol (DON) is the most
frequently found mycotoxin in food and feed around the world (Larsen et al 2004 Desjardins
2006 Starkey et al 2007) Different Fusarium species associated with Fusarium head blight
(FHB) or Gibberella ear rot (GER) disease of small grain cereals and maize are mainly
responsible for contaminating grains with DON and other type-A and type-B trichothecenes
These can cause serious mycotoxicosis in humans animals and plants (Ueno 1985 DrsquoMello
et al 1999 Bae and Pestka 2008) Common effects of DONTC toxicity in animals include
diarrhea vomiting feed refusal growth retardation immunosuppression reduced ovarian
functionsreproductive disorders and even death (Pestka and Smolinski 2005 Pestka 2007)
DON enhances FHB disease severity in wheat and maize (Proctor et al 1995 Desjardins
1996 Maier et al 2006)
67
Reduction of mycotoxin levels in contaminated agricultural products remains a
challenging task Current physical and chemical processes to decrease DON content are either
ineffective or prohibitively expensive (Kushiro 2008 He et al 2010b) However the use of
detoxifying microbes to biodegrade mycotoxins is an alternative (Awad et al 2010
Karlovsky 2011) The toxicity of DON is mainly due to the epoxide and there is a significant
reduction of DON toxicity (54) when it is reduced to produce de-epoxy DON Related
mycotoxins also show reductions in toxicity when they are de-epoxidized such as NIV (55)
and T-2 toxin (400) (Swanson et al 1988 Eriksen et al 2004) Bacillus arbutinivorans
strain LS100 and Eubacterial strain BBSH 797 originating from chicken intestine and rumen
fluid respectively are capable of cleaving the epoxide ring of DON under anaerobic
conditions and high temperature (37degC) (Fuchs et al 2002 Yu et al 2010) Strain BBSH 797
which can de-epoxidize DON scirpentriol and T-2 triol trichothecene mycotoxins has been
used as an additive for poultry and swine diets (Karlovsky 2011) However there are no
reports of microbes able to de-epoxidze DON and other trichothecenes under aerobic
conditions and at moderate temperatures
To develop an effective method for eliminating DON and trichothecene mycotoxins
from contaminated grains soils from fields with FHBGER infected crops were screened for
DON de-epoxidizing microbes (refer to chapter 2 Islam et al 2012) One soil sample yielded
a mixed microbial culture was partially enriched for bacteria with DON de-epoxidizing
activity (chapter 2) The current study describes the purification and characterization of a
novel DON de-epoxidizing bacterium from this culture
68
33 Materials and methods
331 Microbial source
A mixed microbial culture originating from a screen of more than 150 separate soil
samples collected from maize and wheat fields near Guelph Ontario Canada (Islam et al
2012) was used for enrichment purification and characterization of the DON de-epoxidizing
bacterium
332 Media and chemicals
The media used were nutrient broth (NB) 110 strength NB (110 NB) minimal
medium (MM) consisting of six mineral salts (MS) + 100g per liter sucrose (Islam et al
2012) MS + 05 Bacto Peptone (MSB) MS + 03 Bacto Peptone (MSC) MS + 05
yeast extract (MSY) MS + 200 microgmL DON (MS-DON) sterile water-soluble soil extract
(SE) (Islam et al 2012) potato dextrose broth (PDB) and Luria Bertani (LB) Prepared media
and media components were obtained from Fisher Scientific (Fair Lawn New Jersey USA)
Sigma-Aldrich (Oakville ON Canada) andor Difco (Sparks Maryland USA) Media pH
was adjusted as required prior to autoclaving For solid media preparation 15 agar was
added DON and all trichothecene mycotoxins were purchased from Biomin (Romer Lab
Union MO USA)
333 Isolation of the DON de-epoxidizing bacterial strain
Aliquots (20 microL) of the microbial culture partially enriched for DON de-epoxidizing
ability (Islam et al 2012) was transferred to 05 mL MSB (pH 70) in sterile 15 mL
Eppendorf tubes After addition of 200 microgmL pure DON the culture was incubated at room
69
temperature with continuous shaking at 220 revolutions per minute (rpm) Seven to ten days
later 20 microL of the culture was transferred to fresh MSB supplemented with 200 microgmL DON
The culture was then incubated for 7 to 10 days under the above conditions This procedure
was repeated 12 times
After three months of repeated transfers in broth with 200 microgmL DON 100 microL of the
culture was serially diluted in 09 mL MSB containing 05 Tweenreg20 (Sigma Aldrich) to
reduce bacterial cell aggregation (Mabagala 1997) The dilutions were vigorously shaken for
ten minutes A 100 microL aliquot from each dilution was spread on MSB + agar (MSA) The
MSA plates were incubated at room temperature until single colonies appeared
Single colonies from plates were transferred to 1 mL MSB+ 50 microgmL DON and
incubated at room temperature with continuous shaking at 180 rpm under aerobic conditions
After three days the DON de-epoxidation ability was examined by mixing an equal volume
of the bacterial culture and methanol and incubating for 30 min at room temperature to lyse
the cells Insoluble materials in the mixture were removed by passing it through a 045 microm
Millipore filter (Whatmanreg Florham Park New Jersey USA) The level of de-epoxy DON
(dE-DON also known as DOM-1) in the filtrate was determined by liquid chromatography-
ultraviolet-mass spectrometry (LC-UV-MS) (Guan et al 2009)
Individual colonies that showed de-epoxidation activity were serially diluted in
MSB+05 Tweenreg20 and plated onto MSA Single colonies that appeared on the agar plate
were incubated in MSB as described above and retested for DON de-epoxidation ability To
ensure that pure cultures were obtained this was repeated 10 times and individual colonies
were selected on MSA each time
70
334 Identification of a DON de-epoxidizing bacterium
Strain ADS47 with DON de-epoxidation ability was obtained from a single colony
after 10 serial transfers and was grown on MSA at room temperature under aerobic
conditions for 72 h Colony color shape edge and elevation were observed The cell
morphology of the bacterium was examined with a transmission electron microscope at the
University of Guelph Guelph Ontario Canada Whole bacterial cells were placed directly on
a formvar and carbon-coated 200-mesh copper grid They were washed two times by touching
the grids to a drop of clean dH2O and the excess water was blotted off The samples were
negatively stained using 1 Uranyl Acetate (aq) for five seconds Electron microscopy was
performed on a Philips CM10 transmission electron microscope at 60kV under standard
operating conditions (Philips Optics Eindhoven Netherlands) Gram staining was performed
using the procedure described in the commercial gram stain kit from BD Canada
(Mississauga Ontario Canada)
Total bacterial DNA was extracted using a DNeasy Plant DNA Extraction Mini Kit
(Qiagen Inc) The 16S-rDNA was PCR amplified using the primers 46f-
5rsquogcytaacacatgcaagtcga3rsquo (Kaplan and Kitts 2004) and 1371r-5rsquogtgtgtacaagncccgggaa3rsquo
(Zhao et al 2007) PCR was performed in 25 microL containing 06 microM of each primer 1x of
HotStarTaqreg Master Mix (Qiagen Inc) and ~40 ng of DNA The PCR cycles were 95ordmC (15
min) 35 cycles of 94ordmC (20 s) 52ordmC (15 s) 72ordmC (1 min 30 s) and a 72ordmC (7 min) extension
step Amplification was carried out in a GeneAmpreg PCR System 9700 (Applied Biosystems)
PCR products were sequenced with an automated ABI 3730 DNA analyzer (Applied
Biosystems) The 16S-rRNA gene sequence similarity was performed by BlastN using the
Ribosomal Database Project (RDP) (httprdpcmemsueduseqmatch)
71
A phylogenetic tree was constructed by comparing the partial 16S-rDNA sequence of
strain ADS47 with the 11 known Citrobacter species (Bornstein and Schafer 2006 Brenner et
al 1993) The Citrobacter strains were C freundii [FN997616] C koseri [AF025372] C
amalonaticus [AF025370] C farmeri [AF025371] C youngae [AB273741] C braakii
[AF025368] C werkmanii [AF025373] C sedlakii [AF025364] C rodentium [AF025363]
C gillenii [AF025367] and C murliniae [AF025369] Escherichia coli O157H7 [AE005174]
was used as an out group strain The analysis was performed using distance matrix with a gap
opening of 100 and a gap extension of 01 by a server based ClustalW2 program
(httpwwwebiacukToolsmsaclustalw2) A dissimilarity tree was created and viewed
using a TreeView X software (Page 1996)
335 Effects of DON concentration aeration substrate temperature pH and antibiotic
To determine if DON concentration can affect the growth of the isolated bacteria
tubes containing 5 mL of NB or MSB with 20 200 or 500 microgmL of DON were inoculated
with strain ADS47 that had been grown for 16 h with continuous shaking at 220 rpm and then
adjusted to cell density at 025 OD600nm (107 cellsmL) The cultures were incubated at 28degC
with continuous shaking at 180 rpm and 05 mL was removed every 12 h to measure
OD600nm The culture (05 mL) was then analysed for DON de-epoxidation activity by LC-
UV-MS (Guan et al 2009) as described above Three replicate cultures were tested
To examine the effect of aeration five mL of NB or MSB containing 50 microgmL DON
were inoculated with an overnight culture of strain ADS47 prepared as described above and
incubated at 28˚C with continuous shaking at 180 rpm One mL of culture was removed every
six hours and bacterial growth was determined by measuring cell density at OD600nm At the
72
same time points culture samples were analysed for DON to dE-DON transformation ability
(Guan et al 2009) The experiment was repeated with MSB medium containing 50 microgmL
DON under anaerobic conditions as described by Islam et al (2012) with DON to dE-DON
transformation ability being determined every 6 h Three replicate cultures were prepared for
each assay
The effect of substrate on DON de-epoxidation ability was tested after growing
ADS47 for 72 h in NB 110 NB MM MS-DON MSB MSC MSY SE PDB or LB
containing 50 microgmL DON For this assay the media were maintained at pH70plusmn02 and 28degC
for 72 h The effect of temperature was examined with MSB containing 50 microgmL DON at 10
15 20 25 30 35 40 45 or 50degC The effect of pH on growth of strain ADS47 was examined
with MSB adjusted to pH 55 60 65 70 75 or 80 and the cultures were incubated at 28degC
for 72 h Three replicate cultures were used for each assay
Overnight cultures of ADS47 in MSB were treated with 001 and 0001 (wv) of
an antibiotic sodium azide (SA) (Sigma-Aldrich) Controls were prepared in MSB without
SA Three replicate cultures were spiked with 50 microgmL DON DON to dE-DON
transformation ability of the bacterial culture was analysed (Guan et al 2009) after 72 h at
28degC with continuous shaking at 180 rpm Three replicate cultures were tested
336 Localization of DON to dE-DON transformation ability in the bacterium
Bacterial cell extracts were prepared using B-PER II Bacterial Protein Extraction
Reagent following the manufacturerrsquos instructions (PI 78260 Thermo Scientific Rockford
IL USA) ADS47 was grown in 15 mL MSB for 48 h under the growth conditions described
above A culture (05 mL) was centrifuged at 5000 x g for 10 minutes and the pellet was
73
collected B-PER II reagent (2 mL) was added to the pellet and the cells were lysed by
pipetting the suspension up and down several times The cell lysate was treated with 20 microL
Halt Protease Inhibitor Cocktail (Thermo Scientific Rockford IL USA) Culture filtrate was
prepared by passing 05 mL of the culture through a 022 microm Millipore filter (Fisher
Scientific) The remaining 05 mL culture was used as control Cell lysate culture filtrate and
control were incubated with 50 microgmL DON and incubated at 28degC DON to dE-DON
transformation ability of the cell lysate culture filtrate and control were analysed after 72 h
incubation (Guan et al 2009) Three replicate cultures were tested
337 Detoxification of additional trichothecene mycotoxins
Strain ADS47 was evaluated for its ability to degrade five type-A trichothecene
mycotoxins (HT-2 toxin T-2 toxin T2-triol diacetoxyscirpenol and neosolaniol) and five
type-B trichothecene mycotoxins (nivalenol verrucarol fusarenon X 3-acetyl-
deoxynivalenol and 15-acetyl-deoxynivalenol) One and a half mL NB was added with 100
microgmL of each mycotoxin and then inoculated with 100 microL of ADS47 culture grown
overnight as previously described The culture of NB + mycotoxin + ADS47 was incubated at
28degC with continuous shaking at 180 rpm and DON to dE-DON transformation ability of the
culture was analysed by LC-UV-MS (Guan et al 2009) Five replicate cultures were tested for
each trichothecene mycotoxin
74
338 Statistical analysis
Data were processed using Microsoft Excel Program 2007 Statistical analysis was
performed using General Linear Model procedure in SAS version 91 (SAS Institute Cary
NC USA 2002-03) Replications were averaged and stated as mean plusmn standard error The
mean differences were considered significant at plt 005 according to Tukeyrsquos-HSD test
75
34 Results
341 Isolation and identification of a DON de-epoxidizing bacterium
A DON de-epoxidizing bacterium designated strain ADS47 was isolated after 12
transfers over three months from cultures growing in MSB containing 200 microgmL DON
Strain ADS47 produced small to medium size circular convex and white colonies on MSA at
room temperature under aerobic conditions (Figure 3-1a) The bacterium is rod shaped and
150 x 072 microm in size (Figure 3-1b) It has numerous flagella that cover the surface of the cell
(peritrichous flagella) and is Gram-negative
BlastN analysis of the 16S-rDNA sequence of strain ADS47 showed the most
similarity to the eleven known Citrobacter species in the gamma proteobacteria of the
Enterobacteriaceae In particular the 16S-rDNA nucleotide sequence of ADS47 was more
than 99 identical to C fruendii (Figure 3-2) The next most similar sequences were a cluster
of C werkmanii and C murliniae followed by more distantly related C braakii C gilleni
and C youngae A separate cluster was formed by C rodentium C farmeri C sedlakii C
koseri and C amalonaticus
76
Figure 3-1 (a) Colony color shape and size of the mycotoxin degrading bacterial strain
ADS47 on mineral salts agar (MSA) medium (b) Transmission electron microscopic (TEM)
image of the Citrobacter freundii strain ADS47 The bacterium showed multiple flagella
(peritrichous flagella) and numerous fimbriae around the cell surface
(a) (b)
77
Figure 3-2 Dendrogram based on 16S-rDNA sequences for strain ADS47 and 11 reference
Citrobacter species An E coli O157H7 strain was considered as an out group species The
tree was constructed using a genetic distance matrix by ClustalW 2 and viewed by
TreeViewX software The distance was converted to scale and shown by branch lengths (scale
bar 001 which indicated the 16S-rDNA nucleotide sequence variability between the bacterial
species) GenBank accession numbers for 16S-rDNA sequences of the species used for
comparison are shown in materials and methods section (334)
78
342 Effects of DON concentrations on bacterial DON de-epoxidation
Growth of ADS47 aerobically in MSB with 20 200 and 500 microgmL DON was not
significantly different from that observed in control cultures without DON (Figure 3-3a)
Similarly increasing DON concentrations did not affect bacterial growth in NB medium
(Figure 3-3b)
DON de-epoxidation activity determined as microgmL DON convertedcell density
(OD600) was measured lowest in MSB+20 microgmL DON and highest in MSB+ 500 microgmL
DON containing cultures (Figure 3-3c) The highest rate of dE-DON formation was 115 microgh
in ADS47 cultures in MSB with 500 microgmL DON between 48 to 72 h All of the DON was
converted in the cultures containing 20 microgmL DON and 200 microgmL DON between 48-60 h
and 60-72 h respectively In 200 microgmL and 500 microgmL of DON cultures the rate of de-
epoxidation increased significantly after 36 h
79
00
04
08
12
16
0 24 36 48 60 72 96
OD
600n
m
Incubation time (h)
Control
20 microgmL
200 microgmL
500 microgmL
(b)
(a)
80
Figure 3-3 Effects of DON concentration on bacterial growth and de-epoxidation activities
Strain ADS47 was grown in MSB and NB media with three different DON concentrations 20
microgmL 200 microgmL and 500 microgmL Control cultures were prepared without DON Cultures
were grown at 28degC with continuous shaking at 180 rpm (a) Microbial density (OD600 nm) in
MSB with different DON concentrations was measured at 24 36 48 60 72 and 96 h (b)
Microbial growth (OD600 nm) in NB having different DON concentration was measured at 24
48 72 and 96 h (c) Bacterial DON de-epoxidation activity in 20 200 and 500 microgmL DON
containing cultures at different time points DON de-epoxidation was measured in microgmL per
OD600 nm Three replicate cultures for each assay were analysed
(C)
81
343 Influence of growth conditions and antibiotic on microbial DON de-epoxydation
Aerobic and anaerobic conditions
ADS47 grew more quickly uner aerobic conditions under MSB+50 microgmL DON than
in MSB+50 microgmL DON under anaerobic conditions (Figure 3-4a) Aerobic growth was
faster in NB+50 microgmL DON compared to MSB+50 microgmL DON
However the DON de-epoxidation activities on a cell density basis were very similar
in NB+50 microgmL DON compared to MSB+50 microgmL DON (Figure 3-4b) Also DON de-
epoxidation activity of ADS47 under aerobic conditions in MSB+50 microgmL DON was higher
than in the same medium under anaerobic conditions (Figure 3-4b)
82
(b)
(a)
83
Figure 3-4 Bacterial growth and DON de-epoxidation capabilities under aerobic and
anaerobic conditions (a) Microbial growth under aerobic conditions in MSB and NB medium
and anaerobic conditions in MSB medium (b) Microbial DON de-epoxidation ability under
aerobic conditions in MSB and NB media and anaerobic conditions in MSB medium
Cultures media having 50 microgmL DON were inoculated with overnight culture of strain
ADS47 Initial microbial concentration in the cultures was adjusted to 025 OD600nm Cultures
were incubated at 28degC having three replications Bacterial growth at OD600nm and DON de-
epoxidation in microgmL per OD600nm were measured at six hour interval
84
Media temperature and pH
Growth of ADS47 in media containing 50 microgmL DON under aerobic conditions at 72
h was three-fold higher in NB and LB than in SE two-fold higher than in 110 NB and MSC
and did not occur in MS-DON (Figure 3-5a) Growth in MM MSB and MSY was
intermediate
Over the 72 h DON de-epoxidation per cell density was highest in MSB and MSY and
approximately 25 lower in NB and LB and lower by 75 and 87 in 110 NB and MSC
respectively (Figure 3-5b) No DON de-epoxidation was observed in MS-DON in which
DON was used as a carbon source Bacteria grown in MM broth which contains sucrose as a
sole carbon source were unable to de-epoxidize DON No DON de-epoxidation was observed
in SE and PDB grown cultures
Growth of ADS47 under aerobic conditions for 72 h in MSB+50 microgmL DON was
greatest between 25-40degC but no growth occurred at 50degC (Figure 3-6a) DON de-
epoxidation activity was greatest and equivalent between 20-40degC and lower at 10 15 and
45degC temperatures (Figure 3-6b)
ADS47 growth under aerobic conditions for 72 h in MSB+50 microgmL DON was
limited to neutral pHs (ie 60 - 75) (Figure 3-7a) The transformation of DON to dE-DON
occurred at the same pH range and DON de-epoxidation activity was not significantly
different between pH 60 to 75 (Figure 3-7b)
85
b
c
a
b
a
d c
b b
(a)
(b)
b
c
b
a a
d
86
Figure 3-5 Effects of media on bacterial growth and DON de-epoxidation activity (a)
Microbial growth (at OD600nm) on different culture media Media pHs were adjusted to
70plusmn02 (b) Bacterial DON to dE-DON transformation in microgmL by OD600nm in different
media after 72 h The cultures having three replications were incubated for 72 h at 28degC and
DON de-epoxidation was measured The Media NB = nutrient broth 1NB = 110 strength
NB MM = minimal medium MS-DON = six mineral salts + 200 microgmL DON as sole C
source MSB = six mineral salts + 05 bacto peptone MSC = six mineral salts broth + 03
bacto peptone MSY = six mineral salts with 05 yeast extract SE = soil extract PDB =
potato dextrose broth and LB = Luria Bertani Means with the same label are not significantly
different according to Tukeyrsquos HSD test (p lt 005)
87
(a)
(b)
88
Figure 3-6 Effects of temperature on microbial growth and DON de-epoxydation capability
(a) Microbial growth (at OD600nm) at different temperatures after 72 h of incubation (b)
Bacteria DON de-epoxidation in microgmL by OD600nm in different temperatures The most
efficient temperature range (ge40 microgmL DON de-epoxidation) is marked by dot lines The
experiment was conducted in MSB medium at pH 70plusmn02 with three replications
89
(a)
(b)
90
Figure 3-7 Influence of pH on microbial growth and DON de-epoxydation (a) Microbial
growth (OD600nm) in MSB at different pH (b) Bacterial DON to de-epoxy-DON
transformation in microgmL by OD600nm in different pH Dot lines indicated the efficient de-
epoxidation (ge40 microgmL DON conversion) pH range Experiment was conducted at 28degC
with three replications
91
Antibiotic
Treatment of ADS47 at 72 h in MSB+50 microgmL DON with sodium azide (SA) at
0001 and 001 reduced DON de-epoxidation activity to 5 (25 microgmL) and 0
respectively The control with no SA added was able to de-epoxidize DON completely
(100) (Figure 3-8)
Figure 3-8 Effects of sodium azide (SA) on bacterial DON de-epoxidation activity
Microbial culture in MSB+50 microgmL DON was treated with 001 (SA-1) and 0001 (SA-
2) sodium azide In control (C) no SA was added The experiments with three replications
were incubated for 72 h at 28degC and DON de-epoxidation was measured by LC-UV-MS
method
0
20
40
60
80
100
120
SA1 SA2 C
D
ON
de
-ep
oxyd
ize
d
92
344 Localization of bacterial DON de-epoxidation
A cell lysate prepared from ADS47 grown for 48 h in MSB transformed 20 (10
microgmL) of DON added at 50 microgmL to dE-DON within 72 h (Figure 3-9) In contrast a cell-
free culture filtrate did not show DON de-epoxidation activity
Figure 3-9 Determination of DON to de-epoxy DON transformation by bacterial culture
filtrate and cell extract The treatments having three replications were spiked with 50 microgmL
DON and incubated at 28degC for 72 h In control strain ADS47 was incubated in MSB broth
with 50 microgmL DON DON de-epoxidation activity in percentage was examined
0
20
40
60
80
100
120
Filtrate Extract Control
D
ON
de
-ep
oxyd
ize
d
93
345 Bacterial detoxification (de-epoxidtaion and de-acetylation) of other trichothecene
mycotoxins
Aerobic cultures of ADS47 grown for 72 h in NB+100 microgmL HT-2 converted 74 of
the toxin to de-epoxy HT-2 toxin and 26 to a de-acetylated product (Figure 3-10 and Table
3-1)
In a culture treated with T2-toxin 46 was converted to de-epoxy T-2 16 was
converted to an unknown compound and 38 was not converted In ADS47 culture with
diacetoxyscirpenol (DAS) 61 was transformed into de-epoxy DAS and 39 was de-
acetylated DAS (39) Neosolaniol (NEO) was transformed to de-epoxy NEO (55) and
de-acetylated NEO (45) T2-triol was transformed in to de-epoxy T2-triol (47) and de-
acetylated T2-triol (53)
Aerobic cultures of ADS47 were also grown for 72 h in NB+100 microgmL of one of five
type-B trichothecenes For nivalenol (NIV) 92 was de-epoxydized while the remaining 8
NIV was transformed to DON 3-acetyl-deoxynivalenol (3-ADON) was transformed into
three products de-epoxy NIV (5) de-epoxy DON (54) and DON (41) For 15-
acetyldeoxynivalenol (15-ADON) 8 was transformed to de-epoxy 15-ADON and 92 to
de-epoxy DON (dE-DON) FusarenonX (FUS) had 95 transformed into de-epoxy FUS
while 97 of verrucarol (VER) was transformed to de-epoxy VER The remaining 5 FUS
and 3 VER were not transformed
94
Figure 3-10 Determination of microbial de-epoxidation and de-acetylation of HT-2 toxin by
LC-UV-MS Bacterial culture in NB was added with 100 microgmL HT-2 toxin Bacterial HT-2
toxin transformation ability was examined after 72 h of incubation at 28degC The upper panel
illustrated mass spectrometry chromatograph of the de-acetylated (red peak) and de-
epoxydized (blue peak) products Arrows indicated the targeted acetyl (red) and epoxy (blue)
functional groups of HT-toxin The middle and lower panels showed the mass spectra of the
de-acetylated (dA) and de-epoxidized (dE) compounds Arrows on the upper panel indicated
the targeted acetyl (red) and epoxy (blue) functional groups of HT-2 toxin Red and blue
arrows on the middle and lower panels pointed at the degradation sites of HT-2 toxin
95
Table 3-1 Microbial transformation of type-A and type-B tricthothecene mycotoxins after 72
h of incubation under aerobic conditions at room temperature DAS=diacetoxyscirpenol
DON=deoxynivalenol FUS=FusarenonX 3AON=3-acetyl-deoxynivalenol and
15ADON=15-acetyl-deoxynivalenol
1 Convertion of trichothecene mycotoxins to either DON or an unknown compound
Trichothecene
mycotoxin
Untransformed
trichothecenes ()
Transformed products of
trichothecenes ()
De-
epoxy
De-
acetylate
Other1
Type-A
HT-2 toxin 0 74 26 -
T-2 toxin 38 46 - 16
(unknown)
DAS 0 61 39 -
Neosolanol 0 55 45 -
T2-triol 0 47 53 -
Type-B
DON 0 100 - -
Nivalenol 0 92 - 8 (DON)
3-ADON 0 59 41 (DON)
15-ADON 0 100 - -
FUS 5 95 - -
Varrucarol 3 97 - -
96
35 Discussion
The ADS47 strain of C freundii was isolated in the present study by a prolonged
incubation of a mixed microbial culture with a high concentration DON since the bacterium
efficiently de-epoxidized (detoxified) DON and number of other trichothecene mycotoxins
under aerobic and anaerobic conditions at moderate temperatures and neutral pH The total
procedures for enrichment and isolation of the strain ADS47 is depicted in supplemental
Figure S3-1 The reductive de-epoxidation activity may be due to a cytoplasmic enzyme
which would be in agreement with a previous study that showed de-epoxidation of
epoxyalkanes was catalyzed by a bacterial cytoplasmic flavoprotein (Westphal et al 1998)
The ability of the currently isolated strain of C freundii to biodegrade environmental
pollutants such as aminoaromatic acids and heavy metals (copper) is also likely mediated by
cytoplasmic enzymes since these compounds need to be taken up by the cells before they are
metabolized (Savelieva et al 2004 Sharma and Fulekar 2009) In contrast oxidative
transformation of DON to 3-keto DON by microbial extracellular enzyme has been reported
(Shima et al 1997)
Isolation of a trichothecene de-epoxidizing microorganism from a mixed microbial
community is very challenging Although trichothecenes are toxic to eukaryotes prokaryotes
appear to be resistant to these mycotoxins (He et al 1992 Rocha et al 2005) The present
study showed that the bacteria grew well in media with high concentrations of DON (500
microgmL) Bacterial de-epoxidation of DON involves the removal of an oxygen atom from the
functional epoxy ring of the tricyclic DON structure (Yoshizawa et al 1983) Such co-
metabolic degradation does not provide the bacteria with any nutrients from the reacting
compounds (Horvath 1972 Fedorak and Grabic-Galic 1991) This is supported by the results
97
of the current study which showed that strain ADS47 was unable to grow in media with DON
as a sole carbon source and increasing the concentration of DON did not increase ADS47
growth in MSB Hence DON could not be used as sole nutrient source in the mixed culture
enrichment process
Many previous attempts to obtain DON de-epoxidizing microbes have failed
particularly under aerobic conditions because media or laboratory conditions were
inappropriate to support growth or initially active cultures lost de-epoxidizing function after
serial transfer (Karlovsky 2011)
Media composition appears to be a crucial factor for microbial trichothecene de-
epoxidation activity (Voumllkl et al 2004) Effective biotransformation of trichothecene
mycotoxins by the fish digesta microbial culture C133 occurred in full media (FM) but a low
level of de-epoxidation occurred in media having a single nitrogen source and no
transformation activity was observed in the common bacterial medium LB (Guan et al
2009) Zhou et al (2010) also showed that chitin de-acetylation by soil bacterial strains
occurred more rapidly in yeast extract compared to common bacterial media Therefore a
screening strategy for microbial DON de-epoxidation ability should use different media in
parallel as was done in this study
This study observed efficient de-epoxidation in common bacterial media (NB and LB)
as well as in mineral salts medium containing yeast extract (MSY) or 05 Bacto Peptone
(MSB) However reduction of Bacto-Peptone to 03 (MSC) significantly reduced the
bacterial de-epoxidation activity suggesting that the activity is sensitive to the availability of
protein and amino acids The study found that medium containing sucrose as carbon source
(MM) and a commonly used medium PDB blocked microbial DON de-epoxidation even
98
though bacteria grew in those growth media These suggest that media containing high sugar
contents had inhibitory effects on microbial de-epoxidation enzymatic activity Previously
sugars have been found to have negative effects on the production andor activity of bacterial
enzymes (Saier and Roseman 1971)
Losses of microbial functions under laboratory conditions limited researchers to obtain
economically important microbial genetic sources The soil borne Curtobacterium sp strain
114-2 efficiently de-epoxydized nivalenol but unfortunately the strain lost its activity in
latter generations (Ueno et al 1983) Instability of de-epoxidation activity of ADS47
observed in cultures grown on agar media without DON resulted in the loss of DON de-
epoxidation activity by more than 80 of the colonies after one generation However strain
ADS47 retained de-epoxidation activity after ten subcultures in MSB (data not shown)
The non-specific de-epoxidation of DON made it impossible to use DON as a sole
substrate C source for selective isolation of ADS47 Thus a number of techniques were
utilized to select ADS47 and suppress other bacteria including the use of a variety of
antibiotics and culture at elevated temperatures (Islam et al 2012) Selection of the isolate
ADS47 required repeated subculturing and prolonged exposure to media containing 200
microgmL DON After three months of selection with DON culture that was obtained had a
DON de-epoxidation activity that was up to 30-fold higher than the mixed culture As growth
of ADS47 was not affected by 200 microgmL DON the prolonged incubation may have reduced
populations of other bacteria that are more sensitive to DON Enrichment and isolation of a
DON to 3-keto DON transforming bacterial strain E3-39 by prolonged incubation of a soil
microbial culture with 200 microgmL DON was reported by Shima et al (1997)
99
This study demonstrated that microbial DON de-epoxidation occurred mainly at the
stationary growth phase A DON degrading strain of Citrobacter may have gained energy
from DON by co-metabolic cleavage of the epoxy ring under less nutrient conditions
Microbial use of energy from co-metabolic reaction has been suggested by Chiu et al (2005)
The rate of de-epoxidation activity increased when ADS47 grew at higher concentrations of
DON Taken together a prolonged exposure to high concentration DON may have resulted in
non-DON metabolizing microbial populations declining by DON toxicity while facilitated
the population of DON metabolizing bacterium by providing energy
DON is a small molecule and is simply absorbed by cells (Sergent et al 2006)
Microbial DON de-epoxidation occurred in intact and lysed bacterial cells In addition
treating the ADS47 culture with SA which blocks gram negative bacterial electron transport
and activity inhibited DON de-epoxidation activity (Lichstein and Malcolm 1943 Erecinska
and Wilson 1981) implying that an active electron transport system required for microbial de-
epoxidation reaction (He et al 1992) The data suggest that a cytoplasmic epoxy reductase is
involved in the DON de-epoxidation reaction in the ADS47 microbial strain that was isolated
in the present study
Since agricultural commodities are often contaminated with multiple fungal
mycotoxins the ability of bacteria like ADS47 to degrade mycotoxins by several
mechanisms may be an advantage De-acetylation might be another mechanism of microbial
mycotoxin detoxification since toxicity of trichothecenes also relies on the acetyl functional
groups (Desjardins 2006) Strain ADS47 showed efficient de-epoxidation and de-acetylation
of several type-A and type-B trichothecene mycotoxins that are commonly detected in cereals
in association with DON In conclusion the soil-borne gram-negative bacterium C freundii
100
strain ADS47 is capable of detoxifying trichothecene mycotoxins by both de-epoxidation and
de-acetylation under aerobic and anaerobic conditions neutral pH and a range of
temperatures ADS47 has great potential to detoxify DON contaminated animal feed
Furthermore isolation of novel bacterial de-epoxidation genes might be used to generate
transgenic plants that can detoxify DON in contaminated seed Alternatively contaminated
seed might be treated with manufacture de-epoxidation enzyme(s) These applications can
improve the quality and safety of agricultural commodities that are prone to trichothecene
mycotoxin contamination
36 Acknowledgements
Thank to the Natural Sciences and Engineering Research Council of Canada (NSERC) for a
scholarship to Rafiqul Islam The work was funded by the Agriculture and Agri-Food Canada
Binational Agricultural Research and Development Fund (AAFCBARD) project We
appreciate Honghui Zhu for technical assistance
101
Supplemental Figure S3-1 Flow chart depicts the procedures of enrichment and isolation of
DON de-epoxidation bacterial strain ADS47 Steps 1 to 4 describe the optimization of
microbial DON to dE-DON transformation activity and removal of undesired microbial
species Step 5 to 7 shows the culture enrichment by incubation with high concentration DON
and isolation of the single strain ADS47
Step 1 Screening of soil samples
Step 2 Optimization of cultural conditions
Step 3 Elimination of undesired microbial species
Step 4 Removal of untargeted microbial species
Step 5 High concentration DON enrichment
Step 6 Separation of bacterial cells and dilution plating
Step 7 Single colony analysis
Active culture was sequentially treated with different mode of actions antibiotics heating cultures at 50degC and subculturing on agar media
Colonies were tested for their DON de-epoxidation ability Purified the single colony via several times subculturing on agar medium
Optimized the culture conditions ie growth substrates temperatures pH and oxygen requirements
Obtained DON de-epoxidation capable microbial culture obtained by analyzing soil samples collected from cereal fields
Serially diluted active culture was grown on agar media Collected washed colonies having DON de-epoxidation activity
The partially enriched culture that retained DON de-epoxidation activity was incubated with 200 microgmL DON in MSB media with continuous shaking at 220 microgmL for 7 d The procedure was repeated 12 times
The enriched culture was serially diluted and treated dilutes with 05 Tween 20 After vigorously vortex for ten min aliquot cultures were spread on different agar media plates to produce single colonies
102
CHAPTER 4
40 CONSTRUCTION AND FUNCTIONAL SCREENING OF GENOMIC DNA
LIBRARY FOR ISOLATION OF DON DE-EPOXYDIZING GENE(S) FROM
CITROBACTER FREUNDII STRAIN ADS47
41 Abstract
Enzymatic activity-based screening of a genomic DNA library is a robust strategy for
isolating novel microbial genes A genomic DNA library of a DON detoxifying bacterium
(Citrobacter freundii strain ADS47) was constructed using a copy control Fosmid vector
(pCC1FOSTM
) and E coli EPI300-T1R as host strain A total of 1250 E coli clones containing
large DNA inserts (~40 kb) were generated which resulted in about 10-fold theoretical
coverage of the ADS47 genome The efficiency of E coli transformation with the
recombinant Fosmid vector was examined by restriction digestion of Fosmids extracted from
15 randomly selected library clones The fragmentation patterns showed that all of the clones
contained an 81 kb Fosmid backbone and ~40 kb of insert DNA except one that had a small
insert This suggests that a DNA library was prepared with expected DNA inserts The de-
epoxidation ability of the library was analysed by incubating clones in Luria Bertani (LB)
broth medium supplemented with 20 microgmL DON with induction (by addition of L-arabinose)
or without induction None of the 1000 clones showed DON de-epoxidation activity The
potential DON de-epoxidation gene(s) of ADS47 may not have been expressed in E coli or
might have been expressed but lacked additional factors required for DON de-epoxidation
103
Keywords DNA library fosmid vector functional library screening DON de-epoxidation
activity
42 Introduction
Microorganisms contain numerous valuable genes and biocatalysts which may have
significant roles in agricultural productivity (Jube and Borthakur 2007) Various techniques
have been used to isolate novel genes from different organisms including transposon
mutagenesis (Frey et al 1998 Tsuge et al 2001) gene knockout (Winzeler et al 1999)
whole genome sequencing (Krause et al 2006) comparative genomics (Koonin et al 2001)
and suppression subtractive hybridization (Dai et al 2010)
The genomic DNA library preparation and activity-based screening of the library is a
relatively new technique which has been efficiently used for isolation of microbial novel
enzymes and compounds such as lipases proteases oxidoreductases antibiotic resistance
genes membrane proteins and antibiotics (Henne et al 2000 Gupta et al 2002 Knietsch et
al 2003 Gabor 2004 Suenaga et al 2007 van Elsas et al 2008) The strength of the
technique is that genes from microbial sources could be obtained without prior sequence
information and not requiring culturing the organisms under laboratory conditions Using this
technique an unknown gene or a set of genes could be obtained by determining particular
phenotypes of the library clones Eventually the gene of interest from the positive clones is
isolated by using a forward (eg mutation) or reverse genetics method (eg subcloning the
positive clones into a microbial expression vector) followed by functional analysis of the
subclones
104
Although this strategy has been used effectively for isolating novel microbial genes
several factors need to be considered in order for the method to be successful These factors
include DNA quality vector type (eg plasmid cosmid fosmid bacterial artificial
chromosome etc) vector copy number and host strains (Daniel 2005 Singh et al 2009) The
Fosmid vector-based technique has been used successfully to clone bacterial genes including
xenobiotics degradation genes antifungal gene clusters and antibiotic resistance genes from
soil metagenomes (Hall 2004 Suenaga et al 2007 Chung et al 2008) This study reports the
construction and activity-based screening of a Fosmid DNA library in an E coli host with the
aim of isolating a novel DON de-epoxidation gene from Citrobacter freundii strain ADS47
(Chapter 3)
43 Materials and methods
431 Genomic DNA library construction
The genomic DNA library was prepared using a Copy Control Fosmid Library
Production Kit (Epicentre Biotechnologies Madison Wisconsin USA) following the
manufacturerrsquos instructions as depicted in Figure 4-1
105
Figure 4-1 Schematic overview of the construction and functional screening of a Fosmid
DNA library
106
Citrobacter freundii strain ADS47 was grown in NB medium at 28degC with continuous
shaking at 220 rpm The high molecular weight bacterial genomic DNA was extracted from
the overnight culture using the Gentra Puregene YeastBacteria Kit (Qiagen Inc Mississauga
Ontario Canada) The purity of the DNA was confirmed by determining OD260280 at ~190 by
a NanoDrop spectrophotometer A 5 microL sample having ~25 microg high molecular weight DNA
was sheared by passing through a 200-microL pipette tip and achieved a sufficient amount of
DNA about 40 kb in size Blunt ended 5rsquo-phosphorylated insert DNA was generated by
thoroughly mixing it with End-Repair Enzyme reagents on ice The 20 microL mixture was
incubated for 45 min at room temperature followed by incubation at 70degC for 10 min to
inactivate the End-Repair Enzyme activity
The treated DNA was size fractionated by a Low Melting Point-Pulsed Field Gel
Electrophoresis (LMP-PFGE) on the clamped homogeneous electric fields (CHEF-DR III)
system (Bio-RAD Laboratories) using 1 UltraPure Low Melting Point Agarose (Invitrogen)
The PFGE was conditioned according to manufacturerrsquos instructions as follows initial switch
time-42 s final switch time-761 s run time 16 h 50 min angle-120deg gradient- 6 Vcm
temperature-14degC and running buffer-05 TBE DNA size markers for the 42 kb Fosmid
control DNA (Epicentre Biotechnologies) and low range Pulse Field Gel (PFG) markers (New
England Biolabs Inc) were loaded adjacent to the DNA sample slots After electrophoresis a
portion of the gel that contained DNA size markers was cut off The gel was stained in SYBR
Safe DNA gel stain (Invitrogen) for 30 min visualized and marked on the 23 42 and 485 kb
marker bands A slice of PFGE gel containing 40 plusmn 10 kb size DNA was cut off with a sterile
razor blade by comparing the marker sizes DNA from the gel was recovered by digesting it
with GELase enzyme ( 1 U100 microL melted agarose) in 1X GELase Buffer followed by a
107
sodium acetate-ethanol precipitation The DNA with the desired size was stored at 4degC for
next dayrsquos ligation to a cloning-ready Fosmid vector (pCC1FOSTM
)
Ligation was performed in a 10 microL reaction mixture by thoroughly mixing 10X Fast-
Link Ligation Buffer 10mM ATP 025 microg of ~40 kb insert DNA 05 microg Fosmid Copy
Control Vector (pCC1FOS) (Figure 4-2) and 2 U Fast-Link DNA lipase The mixture was
incubated for 4 h at room temperature preceded by inactivating the ligation reaction by
heating at 70degC for 15 min The recombinant Fosmid vector was stored at -20degC for later use
Ten microL of the ligated DNA was thawed on ice and transferred to a pre-thawed
MaxPlax Lambda Packaging Extract tube containing 25 microL extract By pipetting several
times the mixture was incubated at 30degC for lamda packaging After 2 h of packaging an
additional 25 microL packaging extract was added into the mixture and incubated for a further 2 h
at 30degC The packaged phage particles were volumed to 05 mL by adding ~400 microL Phage
Dilution Buffer and 25 microL chloramphenicol
Transformation was performed via transduction by mixing the packaged particles with
the overnight cultured E coli EPI300-T1R cells (1 10 vv ratio) in LB supplemented with 10
mM MgSO4 and 02 Maltose Transformation was completed by incubation at 37degC for 1 h
The transformed E coli cells were spread on an LB agar plate containing 125 microgmL
chloramphenicol The plates were incubated overnight at 37degC Colonies that appeared on the
plates were individually transferred to 96-well microtiter plates having 200 microL LB medium
supplemented with 10 glycerol and 125 microgmL chloramphenicol The microtiter plates
were incubated at 37degC for 24 h with continuous shaking at 180 rpm The prepared library
clones were stored at -80degC for further analysis and functional screening
108
109
Figure 4-2 Genetic map of copy controlled Fosmid vector (pCC1FOSTM
) The 813 kb
Fosmid vector backbone shows a multiple cloning site (MCS) chloramphenicol resistance
selectable marker (CmR) replication initiation fragment (redF) inducible high copy number
origin (oriV) replication initiation control (repE) and F-factor-based partitioning genes (ParA
ParB ParC) The vector contains a cos site for lamda packaging (not shown) and lacZ gene
for bluewhite colony selection The vector map was provided by the Epicentre
Biotechnologies (httpwwwepibiocomitemaspid=385)
110
432 Confirmation of library construction
The quality of the DNA library prepared was determined by examining 15 E coli
clones randomly selected A 15 mL LB broth supplemented with 125 microgmL
chloramphenicol was inoculated by each of the transformed clones To obtain higher Fosmid
DNA yield 1 microL autoinduction solution (L-arabinose) to increase the copy number of the
Fosmid vector (Epicentre) was added to the cultures After an overnight incubation of the
cultures at 37degC with continuous shaking at 220 rpm Fosmid DNA was extracted using a
Qiagen Large-Construct Kit followed by restriction digestion with Not I enzyme A 20 microL
digestion mixture containing 05 microg Fosmid DNA 1X digestion buffer and 05 U Not I was
incubated at 37degC for 15 h The partially digested DNA and a 42 kb DNA size marker
(Fosmid control DNA) were ran on 08 agrose gel for 15 h at 100 V The linear Fosmid
vector backbone (813 kb) and insert DNA fragments were visualized on the gel and the
image was recorded
433 Functional screening of the clones
Eppendorf tubes (15 mL) containing 500 microL LB supplemented with 125 microgmL
chloramphenicol were inoculated with a single clone (400 clones tested) or a combination of
12 library clones (600 tested) from microtiter plates stored at -80degC One half of the cultures
in each group (single or combined clones) were induced to increase the Fosmid copy number
by adding a 1 microL (500X) autoinduction solution (contained L-arabinose) to the culture tubes
L-arabinose induces the expression of a mutant trfA gene of the E coli host strain which
leads to the initiation of Fosmid replication from a low-copy to a high-copy number origin of
replication (oriV) The cultures were spiked with 20 microgmL DON and incubated at 37degC for
111
72 h with continuous shaking at 180 rpm The DON to de-epoxy DON transformation ability
of the clone cultures was examined by a liquid chromatography-ultraviolet-mass spectrometry
(LC-UV-MS) method as described by Islam et al (2012)
112
44 Results
441 Production of the DNA library
High molecular weight intact (non-degraded) and good quality (OD600280 ~190)
genomic DNA was obtained from the DON de-epoxidizing bacterial strain ADS47 (data not
shown) Pipetting of the DNA extract resulted in an adequate amount of insert DNA (gt025
microg) of approximately 40 kb size as showed on a LMP-PFGE (Figure 4-3)
A DNA library in E coli host was constructed using LMP-PFGE purified DNA The
library was comprised of 1250 E coli transformants which were selected on LB agar plates
supplemented with chloramphenicol (Figure 4-4)
113
Figure 4-3 Preparation of a 40 kb size insert DNA from a DON de-epoxidizing bacterium
Genomic DNA was extracted from the overnight culture of a DON de-epoxidizing
Citrobacter freundii strain ADS47 After fragmentation the DNA was size fractionated by
Low Melting Point- Pulse Field Gel Electrophoresis (LMP-PFGE) with markers for the PFGE
(M1) and 42 kb Fosmid control DNA (M2) S1 S2 S3= different DNA samples extracted
from bacterial strain ADS47 The red dots area indicates the section of the gel containing
genomic DNA of the expected size (~40 kb)
114
Figure 4-4 Production of the genomic DNA library in E coli EPI300-T1 host strain
Approximately 40 kb long blunt ended genomic DNA from a DON de-epoxidizing bacterial
strain ADS47 was ligated to a Copy Control Fosmid vector pCC1FOS The construct was
packaged into lamda bacteriophage followed by a transduction-mediated transformation of E
coli host cells The clones which appeared on the LB agar plate supplemented with 125
microgmL chloramphenicol are expected to be transformed by the recombinant Fosmid vector
containing insert DNA
115
442 Conformation of the library construction
All of the clones tested acquired the Fosmid vector Out of 15 clones examined 14
received the recombinant Fosmid vector that contain the expected size insert DNA as
determined by an 81 kb vector backbone and at least one high molecular weight insert DNA
band (~40 kb) on agarose gel (Figure 4-5) In contrast one clone (lane 4) showed an 81 kb
vector backbone and a low molecular weight band which is also a fragment of the Fosmid
vector The result indicated that approximately 94 of the library clones were transformed
with the recombinant vector
443 Activity-based screening of the library
Out of 500 single copy number clones (200 individual and 300 combined clones)
none showed DON to de-epoxy DON transformation activity (Table 4-1) An additional 500
high copy number clones (200 single and 300 combined clones) were unable to de-epoxidize
DON as well
116
Figure 4-5 Confirmation of the Fosmid DNA library production in E coli Fosmid DNA
collected from 15 clones was digested with Not I and run on 08 agarose gel at 100 V for 15
h The presence of the vector backbone (81 kb) and the insert DNA fragment (~40 kb) in the
clones was determined by comparing a 42 kb molecular size marker (M) Lanes (1-15) denote
the recombinant Fosmid DNA extracted from 15 E coli clones
117
Table 4-1 Analysis of Fosmid DNA library clones for deoxynivalenol (DON) de-epoxidation
capability
Type of clones Analysis of
singlecombined
clones
Number of
clones
examined
Number of clones
showed DON de-
epoxidation activity
Without induction
(single copy number clones)
Singly 200 0
Combination of 12 300 0
With induction (multi-copy
number clones)
Singly 200 0
Combination of 12 300 0
De-epoxidation activity of the clones was examined alone or in combination of 12 clones
without or with induction by adding L-arabinose The clones grown in LB supplemented with
125 microgmL and 20 microgmL DON were analysed for DON to de-epoxy DON transformation
ability by a liquid chromatography-ultra violet- mass spectrum (LC-UV-MS)
118
45 Discussion
While a Fosmid DNA library with the expected genome coverage of a DON de-
epoxidizing bacterial genome was produced the function-derived screening of the library
clones did not identify any clones that showed DON de-epoxidation enzymatic activity This
is unlikely due to lack of coverage of the bacterial genome as 1000 clones analyzed in the
study should have provided eight fold coverage of the genome Therefore it was likely due to
a lack of expression of the enzyme(s) for DON de-epoxidizing activity under the conditions
used
Thus far there is only one report of heterologous expression of bacterial epoxide
reductase gene in another bacterium A high level of heterologous expression of a
Synechococcus sp vitamin K epoxide reductase gene was achieved in E coli with codon
optimization and placing the gene under the control of a prokaryotic T7 promoter in an
expression vector (Li et al 2010) Therefore heterologous expression of bacterial epoxide
reducatse gene using its own promoter has not yet been demonstrated The success of the
method used in this chapter relies on an intact copy of the gene or operon being cloned the
selection of an appropriate cloning vector a host organism that can recognize the cloned C
fruendii promoter and a suitable library screening method for DON degradation by E coli
(Mocali and Benedetti 2010)
The unbiased cloning efficiency of the Fosmid vector its ability to contain large insert
DNA high stability in E coli as single copy number and autoinduction to increase copy
number in host cells made it appear to be a suitable method for the present work (Kim et al
1992 Lee et al 2004) Library production with large insertions is a major advantage as gene
cluster involved in particular pathway could be trapped into single clone and their functions
119
could be analysed The pathway of microbial DON de-epoxidation activity and the genes
encoding the activity are not known Therefore it was reasonable to prepare a library with
large inserts to ensure the possibility of cloning single as well as whole pathway genes of
interest for functional screening A further advantage of the method is that Fosmid clones are
inducible which allows detecting poorly expressed genes in a heterologous host by inducing
high copy numbers (Daniel 2005)
One of the limitations of the method is that the functions of the target genes rely
mostly on the host organismrsquos transcription and translation machineries Therefore selection
of the host organism is critical for activity-based screening of library clones in heterologous
hosts A number of host strains including E coli Streptomyces lividans Rhizobium
leguminosarum and Pseudomonas aeruginosa have been used for functional screening
activities from diverse organisms (Daniel 2005) The present work attempted to isolate C
freundii strain ADS47 genes encoding DON de-epoxidation enzymes Both strain ADS47 and
E coli belong to gamma proteobacteria which suggest that E coli would be an appropriate
host organism for heterologous expression of ADS47 genes Poor expression of heterologous
genes could be another limitation of the method which may be overcome by overexpression
to increase the catalytic activity of less expressed genes However clones with DON de-
epoxidation activity were not identified in this study even the Fosmid copy number was
increased by induction
In general a large number of clones must be analysed to obtain the gene of interest
using an activity-based screening technique (Lee et al 2004 van Elsas et al 2008) In this
study the constructed DNA library comprised of 1160 clones having approximately ~40 kb
insert DNA suggesting that the prepared library achieved ~10 fold coverage of the ADS47
120
genome (48 Mb) which ensured gt99 probability of the presence of all DNA sequence in
the library In the produced library three clones (lanes 3 10 and 12) showed one intact band
greater than 42 kb in size on the agarose gel It is presumed that the Fosmid DNA of the three
samples was not digested because of insufficient amount of restriction enzyme added to the
digestion mixture About half of the samples demonstrated multiple bands suggesting that the
insert DNA might have contained multiple Not I restriction sites
However not every heterologous gene may necessarily express or function in the E
coli host (Gabor et al 2004) The functionality of the gene requires expression of the genes
and proper folding of the protein which may be influenced by environmental conditions (eg
substrate temperature and pH) and genetic factors In this study library clones were screened
in growth conditions at which strain ADS47 performed the best de-epoxidation activity The
expression of genes may be controlled by the promoter suitable ribosome binding site and
transndashacting factors (eg special transcription factors inducers chaperones cofactors proper
modifying enzymes or an appropriate secretion system) (Gabor et al 2004) Those factors
have to be supplied by the cloned DNA fragments or host cells to allow any heterologous
gene to be functional Since DON de-epoxidation genes of strain ADS47 evolved in a distinct
environmental condition the expression and activity of the gene(s) might have required
specific factorsregulators that were absent in E coli Even though the Fosmid based library
screening technique facilitated isolation of many microbial genes clones having de-
epoxidation activity were not identified using such technique in this study
121
46 Acknowledgements
We appreciate the Agriculture and Agri-Food CanadaBinational Agricultural
Research and Development Fund (AAFCBARD) project for providing the financial support
for this work The authors thank Honghui Zhu for analyzing numerous samples by liquid
chromatography ultraviolet mass spectrum method
122
CHAPTER 5
50 GENOME SEQUENCING CHARACTERIZATION AND COMPARATIVE
ANALYSIS OF CITROBACTER FREUNDII STRAIN ADS47
51 Abstract
The genome of a trichothecene mycotoxin detoxifying C freundii strain ADS47
originating from soil was sequenced and analyzed Strain ADS47 contains a circular
chromosome of 4878242 bp that encodes 4610 predicted protein coding sequences (CDSs)
with an average gene size of 893 bp Rapid Annotation Subsystems Technology (RAST)
analysis assigned functions to 2821 (62) of the CDSs while 1781 (38) CDSs were
assigned as uncharacterized or hypothetical proteins Comparative bioinformatics analyses
were done for strain ADS47 C freundii Ballerup 785139 Citrobacter sp 30_2 C youngae
ATCC 29220 C koseri ATCC BAA-895 and C rodentium ICC168 There were a significant
number of common orthologous genes (2821) were found in the six Citrobacter genomes
implying a common evolutionary relationship ADS47 is most closely related to the
opportunistic human pathogen C freundii Ballerup The human and mouse pathogenic
species C rodentium and C koseri are the most distantly related to ADS47 and non-
humananimal pathogens C youngae and Citrobacter sp 30_2 were closely related ADS47
C koseri C youngae and Citrobacter sp 30_2 lacked the clinically important locus for
enterocyte effacement (LEE)-encoded type-III secretion systems which are present in C
rodentium and C freundii Ballerup Strain ADS47 had 270 unique CDSs many of which are
located in the large unique genomic regions potentially indicating horizontal gene transfer
123
Among the unique CDSs nine were reductasesoxidoreductases and a de-acetylase that may
be involved in trichothecene mycotoxin de-epoxidation and de-acetylation reactions
Keywords Citrobacter freundii comparative genomics genome sequencing unique genes
reductases
52 Introduction
According to the Food and Agriculture Organization (FAO) at least 25 of the
worldrsquos food crops are contaminated with mycotoxins causing serious negative impacts on
livestock and international trade (Boutrif and Canet 1998 Desjardins 2006 Binder et al
2007) Trichothecenes (TCs) produced by a variety of toxigenic Fusarium species are
probably the most prevalent food and feed contaminating mycotoxins It is estimated that
infections by the fungus caused more than $1 billion in crop yield losses in USA and Canada
alone (Windels 2000 Schaafsma et al 2002 Yazar and Omurtag 2008) TC mycotoxins can
have adverse affects on human health including organ toxicity mutagenicity carcinogenicity
neurotoxicity modulation of the immune system negative reproductive system affects and
growth retardation (DrsquoMello et al 1999 Pestka et al 2007) A strain of Citrobacter freundii
ADS47 was isolated from soil in southern Ontario that is capable of detoxifying (ie de-
epoxidation and de-acetylation) several TC mycotoxins including deoxynivalenol nivalenol
and T2-toxin (Chapters 2-3)
Citrobacter freundii is a gram negative motile rod shaped facultatively anaerobic
bacterium belonging to the family of Enterobacteriaceae (Wang et al 2000 Whalen et al
2007) The opportunistic human pathogen is responsible for a number of infections including
124
nosocomial infections of respiratory tract urinay tract blood and causes neonatal meningitis
(Badger et al 1999 Wang et al 2000) C freundii exists almost ubiquitously in soil water
wastewater feces food and the human intestine Biodegradations of tannery effluents copper
aromatic amino acids by C freundii strains isolated from water mine tailings and soil
respectively have been previously reported (Kumar et al 1999 Savelieva et al 2004 Sharma
and Fulekar 2009) Because of their biochemical capabilities bacteria have played important
roles in industrial biotechnology including de-epoxidation and de-acetylation activities
(Demain and Adrio 2008 Plank et al 2009 Zhao et al 2010) However the identification of
genes for de-epoxidation activity against trichothecene mycotoxins is limited (Boutigny et al
2008)
The advent of molecular techniques such as next generation sequencing technologies
and the related bioinformatics tools has enabled the discovery of novel genes and the
enzymes they encode in pathogenic and beneficial bacteria (van Lanen and Shen 2006
Ansorge 2009 Zhao 2011) In the present study the genome of mycotoxin-detoxifying strain
C freundii ADS47 was sequenced and subjected to comparative genomic analyses with other
Citrobacter species to identify putative mycotoxin-detoxifying genes
125
53 Materials and methods
531 Sequencing quality control and assemble of ADS47 genome
Genomic DNA of the strain ADS47 was extracted using Gentra Puregene
YeastBacteria Kit (Qiagen Inc Mississauga Ontario Canada) and purity of the DNA was
confirmed by determining OD 260280 at ~190 using a Nanodrop spectrophotometer
The genomic DNA was sent to be sequenced by the Illumina HiSeq 2000 sequencing
analyzer at Beijing Genomics Institute (BGI) Hong Kong
(httpwwwbgisequencecomhomeservicessequencing-servicesde-novo-sequencing) The
sequencing workflow involved short read library preparation cluster generation sequencing
and data analysis Briefly 3-5 ug of bacterial gDNA was fragmented to produce a 500 bp
insert library by preparative electrophoresis through a 08 agarose gel The fragmented
DNAs were blunt-ended by treating with a mixture of enzymes (T4 DNA polymerase klenow
fragment) and subsequently adapters were ligated to the blunt ends Each of the DNA
fragments was amplified to a cluster which was performed in the flow cell by template
hybridization and bridge amplification Millions of clusters were then sequenced by
sequencing-by-synthesis technology leading to base calls The raw data was filtered to remove
low abundant sequences from the assembly according to 15-mer frequencies The low
complexity reads low quality (leQ20) base calls and adapter contaminations (24 bp overlap
between adapter and reads) were eliminated The quality of the sequences was analysed by
GC content and depth correlative analysis (for GC biasness) K-mer analysis was performed
using 15 bp of the short reads for heterologous sequence identification
126
The cleaned short reads (approximately 90 bp) were self-assembled using the Short
Oligonucleotide Analysis Package (SOAP) denovo (httpsoapgenomicsorgcn) (Li et al
2008) The assembly errors were corrected by filling gaps and proofreading single base pairs
using mapping information The assembly produced 441 contigs with an N50 contig size of
29222 bp (the largest contig size 114839 bp) and 80 scaffolds with an N50 scaffold size of
325726 bp (largest scaffold 662452 bp) with an total sequence length of 4878242 bp which
constitutes 33-fold genome coverage The scaffolds were ordered and oriented according to a
reference genome C koseri ATCC BAA-895 the closest completely sequenced species of
the newly sequenced bacterial genome by alignment with the MUMmer program v322
(httpmummersourceforgenet) (Delcher et al 2002) Genome coverage was determined by
mapping the reads to the assembled genome with Bowtie software (Langmead et al 2009) A
preliminary genome annotation was performed to define the origin of replication by
identifying colocalization of multiple genes ie gyrB recF dnaN and dnaA (usually found
near the origin of replication) and comparison with the C koseri reference genome Synteny
analysis between the reference bacterium and assembled scaffolds was performed with the
MUMmer program
532 Gene function annotation and classification
The genome of ADS47 was annotated by Rapid Annotation using Subsystems
Technology v40 (RAST) a server-based genome analysis system (httprastnmpdrorg)
(Aziz et al 2008) A circular genome map was constructed using CGView program
(httpstothardafnsualbertacacgview_server) (Stothard and Wishart 2005)
127
533 Molecular phylogenetic analysis
A robust phylogenetic tree was produced using Composition Vector Tree (CVTree)
web-based program v20 (httpcvtreecbipkueducn) (Qi and Hao 2004) by comparing
whole genome amino acid sequences of strain ADS47 two completely sequenced Citrobacter
genomes [C rodentium ICC168 (NC_013716) and C koseri ATCC BAA-8959 NC_009792)]
and three partially sequenced Citrobacter genomes [C freundii strain Ballerup 785139
(CACD00000000) Citrobacter sp 30-2 (NZ_ACDJ00000000) and C youngae ATCC 29220
(NZ_ABWL00000000)] E coli O157H7 strain EDL933 (NC_002655) was used as an
outgroup species The tree was constructed from the generated pairwise distance matrix data
using Neighbor-Joining Program in PHYLIP v369 package (Saitou and Nei 1987 Felsenstein
1989)
534 Comparative genomic analysis
Genome-wide comparative analyses between C freundii ADS47 C freundii strain
Ballerup 785139 Citrobacter sp 30-2 C youngae ATCC 29220 C Koseri ATCC BAA-895
and C rodentium ICC168 was carried out The nucleotide sequence idendities were
determined by pairwise alignments of the ADS47 genome with the five other Citrobacter
genomes using MUMmer software (Delcher et al 2002) and GC content total CDSs genome
coverage by CDSs and number of stable RNAs were determined by RAST program
Orthologous CDS groups among the total set of 29079 CDSs derived from the six
Citrobacter genomes were determined using OrthoMCL software v202
(httporthomclsourceforgenet) based on all-against-all reciprocal BLASTP alignment of
amino acid sequences (Li et al 2003) Genes unique to each strain were identified as those
128
CDSs not included in any orthologous group of the five other Citrobacter genomes BLASTP
comparisons were visualized with Blast Ring Image Generator (BRIG) software v095
(httpsourceforgenetprojectsbrig) (Alikhan et al 2011)
Protein sequences of the five Citrobacter genomes pooled from the GenBank data
bases were compared with ADS47 A circular map was developed by BRIG software that
showed the unique genomic regionsgenes of ADS47 The unique genes were grouped
according to their functional similarities previously identified by RAST software and
genomic regions possibly acquired by horizontal transfer were identified using RAST
software
129
54 Results
541 Assembly and organization of the ADS47 bacterial genome
Scaffolds derived from Citrobacter freundii strain ADS47 were ordered and oriented
by alignment to Citrobacter koseri ATCC BAA-895 the most closely related genome that has
been completely sequenced The genomes of ADS47 and C koseri displayed close synteny
(Figure 5-1)
Figure 5-1 Co-linearity between C freundii strain ADS47 genome and C koseri Scaffolds
derived from ADS47 genome sequences (Y- axis) were aligned and assembled by comparison
with the closest completely sequenced genome C koseri ATCC BAA-875 (X- axis)
130
542 An overview of the ADS47 genome
General features of the C freundii strain ADS47 are presented in Table 5-1 Some
notable features are that strain ADS47 contains 4881122 bp circular chromosome with an
average GC content of 5202 The genome contains 4610 CDSs which comprise 8784 of
the whole genome Annotation by RAST assigned 2821 (62) CDSs to known biological
functions and remaining 1789 (38) genes were classified to unknown or hypothetical
proteins
The GC content in the coding regions is significantly higher (5318) than in the non-
coding intergenic region (4379) which covers 1215 (592781 bp) of the genome The
intergenic region is comprised of 018 repeat sequences (total 8999 bp) 5 (009)
transposons (4806 bp) 025 stable RNA (12091 bp) and the remaining ~115 is
classified as regulatory and other functions The ADS47 genome contains 62 tRNAs 3
rRNAs and 34 copies of sRNAs The average lengths of the tRNAs rRNAs and sRNAs are
785 885 and 13441 bp respectively There are also 42 phageprophage-related genes
identified in the genome
Figure 5-2 shows the distribution of the 26 categorized CDSs rRNA tRNA GC
content and GC skew It appears that ADS47 genome contains equal quantity of genes in
leading and lagging strands and showed occasional clustering (likely operons) of genes in the
two strands The genome contains three rRNAs (two large subunit and a small subunit) and 62
tRNAs which are distributed across the genome A consistent GC content was observed in
the ADS47 genome however 7-10 low GC content (high black color peaks) genomic regions
were found at locations origin of replication 600 2000 3000 3300 3550 3850 4300 and
4800 kbp This is indicative of possible horizontally transferred genes or insertions in those
131
genomic locations GC skew denotes the excess of C over G in certain genomic locations
Positive GC skew (+) (green color) indicates the skew value in the leading strand while
negative skew (-) (purple color) in the lagging strand As in many bacterial genomes the
ADS47 genome map shows the separation of positive and negative GC skew at the origin of
replication (base pair position 1) The GC skew in the leading strand is highly skewed (high
green color peak) indicating that possible horizontal gene transfer occurred into the ADS47
genome
The CDSs ranged from ~100 bp to ~2000 bp and have an average size of 893 bp
(Figure 5-3) The highest numbers of CDS (770) were 600-800 bp long A considerable
number of genes (220) were gt2000 bp in size Some of the biological functions for the
smallest genes (100-200 bp) are sulfur carrier protein methionine sulfoxide reductase
ribonuclease P protein component while the largest genes (gt2000 bp) are ferrous iron
transport protein B translation elongation factor G pyruvate flavodoxin oxidoreductase and
tetrathionate reductase
132
Table 5-1 General features of the C freundii strain ADS47 genome
Feature Chromosome
Genome size 4881122 bp
Overall GC content 5202
Number of genes 4610
Genes with assigned function (number and ) 2821 (62 )
Genes without assigned function (number and ) 1789 (38 )
Total length of protein coding sequences 4285461 bp
Genome coverage by coding sequences () 8784
Average length of coding sequences 893 bp
GC content in the coding region 5318
GC content in the non-coding region 4379
Length and percentage of intergenic region 592781 bp and 1215
Total repeat sequences and genome coverage 8999 bp and 018
Total transposons sequences and genome coverage 4806 and 009
Number average length and genome coverage of tRNA 62 copies 785 bp and 0099
Number average length and genome coverage of rRNA 3 copies 885 bp and 0067
Number average length and genome coverage of sRNA 34 copies 13441 bp and 0093
133
134
Figure 5-2 Circular genome map of C freundii strain ADS47 Map was created with the
CGView software From outside to inside the first circle indicates the genomic position in kb
Different functional categories of predicted coding sequences (CDS) are distinguished by
colours with clockwise transcribed CDSs in the second circle and anticlockwise transcribed
CDSs in the third circle Bars on the fourth and fifth circles indicate the rRNAs and tRNAs
respectively Sixth and seventh circles show the coverage and GC content respectively The
innermost circle demonstrates the GC skew + (green) and GC skew- (purple)
135
Figure 5-3 Size distribution of the predicted genes identified in C freundii strain ADS47
The X axis identifies the gene size categories (bp) and Y axis the abundance of genes
136
543 Annotation and functional classification of ADS47 genome
RAST analysis identified predicted CDSs in the genome of Citrobacter strain ADS47
assigned functions and grouped the annotated genes into 26 functional subsystem categories
out of 27 No CDS was found belonging to the photosynthesis category A graphical
presentation of the distribution of the CDSs into each of the 26 categories is given in
Figure 5-4
A large portion (38) of the CDSs belong to hypothetical or unknown functions
Among the 62 known or predicted proteins the highest numbers (835) belong to the
carbohydrate category followed by amino acids and derivatives (468) and
cofactorsvitaminsprosthetic groups (324) A noticeable number of stress response (236) and
respiration (198) categories genes were identified in the bacterial genome The ADS47
genome encoded by 171 membrane transport genes 113 virulencedefense genes and 126
genes involved in regulationcell signaling categories Two hundred and one CDSs were
grouped into the miscellaneous category whose functions are different than the indicated
RAST categories The RAST annotations further divided the categorized CDSs into
subcategories based on similar functional roles which is discussed in section 545
137
Figure 5-4 Functional classification of the predicted coding sequences (CDS) of C freundii
strain ADS47 The annotation was performed by Rapid Annotation using Subsystems
Technology (RAST) program
138
544 Molecular phylogenetic relationships
Genome-wide phylogenetic analysis based on comparisons of whole genome amino
acid sequences of two completely sequenced Citrobacter genomes (C koseri and C
rodentium) and three partially sequenced Citrobacter genome sequences (C freundii
Citrobacter sp and C youngae) illustrates the relationships of ADS47 to five Citrobacter
species (Figure 5-5)
C youngae ATCC 29220 originating from human feces and two pathogenic species
(C koseri ATCC BAA-895 and C rodentium strain ICC168) are progressively more distantly
related to ADS47 The present analysis is consistent with the 16S-rRNA sequence based
phylogenetic analysis (Chapter 3) which showed that the closest bacterial strain to ADS47 is
C freundii (Figure 3-2) This confirmed the identification of strain ADS47 as the gamma
proteobacterial species C freundii made in Chapter 3
139
Figure 5-5 Phylogeny of the trichothecene detoxifying C freundii strain ADS47 The
dendrogram was constructed by comparing the ADS47 genome (amino acid sequences) with
five Citrobacter genomes and a E coli strain (out group species) using the CVTree software
The calculated pairwise distances matrix by Protdist program was transformed into a tree by
neighbor-joining program The dissimilarity tree was drawn to scale with branch lengths
(scale bar 002 indicated the amino acid substitutions among the genomes) The C koseri C
rodentium C freundii Citrobacter sp C youngae and E coli protein sequences were
uploaded from GenBank (Benson et al 2009)
140
545 Comparative genomics
Genome-wide comparative analyses between the genome of strain ADS47 and the five
previously sequenced Citrobacter genomes showed that 872 of the total nucleotides
between ADS47 and C freundii Ballerup were identical The values for the comparisons
between C freundii strain ADS47 were 8212 for Citrobacter sp 30-2 7637 for C
youngae 5981 for C koseri and 4213 for C rodentium (Table 5-2)
The two human and mouse pathogenic species namely C rodentium and C koseri
and the bacteria originally isolated from human feces (C youngae) have higher GC contents
(546 and 532 respectively) compared to C freundii strain ADS47 C freundii strain
Ballerup and Citrobacter sp 30_2 which have slightly lower GC contents (~52) The two
pathogenic species contain 22 copies of rRNA which is significantly higher than the 9 found
in C freundii strain Ballerup 6 in Citrobacter sp 30_2 and 3 in C freundii strain ADS47 but
less than the 25 copies in C youngae The genome of C freundii strain ADS47 C freundii
strain Ballerup C youngae and Citrobacter sp 30-2 had a similar percentage of the genome
comprising coding sequences (865-88) C koseri and C rodentium genomes contained the
highest (90) and lowest (84) genome coverage by coding sequences respectively
141
Table 5-2 Genome comparisons between strain ADS47 and five Citrobacter species The
base nucleotide sequence identities were performed using MUMmer DNAdff script program
(Delcher et al 2002) The features (GC content total CDS percentage CDS and number of
ribosomal RNA) were determined using RAST program (Aziz et al 2008)
Genome feature Bacterial species
C
freundii
ADS47
C freundii
Ballerup
Citrobacter
sp 30_2
C
youngae
C koseri C
rodentium
Genome length
(bp)
4878242
4904641
5125150
5154159
4720462
5346659
Base similarity to
ADS47 ()
NA 8715 8212 7637 5981 4213
GC () 5202 5218 516 5265 530 546
Number of coding
sequences
4610 4685 4728 5278 4978 4800
Genome coverage
with coding
sequences ()
880 865 866 872 900 840
rRNA copies 3 9 6 25 22 22
Total stable RNAs 99 81 78 105 120 142
142
The number of orthologs shared between ADS47 and other Citrobacter species is
illustrated in Figure 5-6 ADS47 shares the smallest number of orthologous CDSs with C
rodentium and C koseri 3250 and 3341 respectively The highest number of shared CDS
were found with C freundii Ballerup (3706) followed by C youngae (3693) and Citrobacter
sp 30_2 (3676) A total of 2821 CDSs were found to be common to all of the six Citrobacter
genomes analysed (data not shown) Nonetheless significant diversity exists between C
freundii ADS47 and the other species as a considerable number of unshared genes were
identified from the comparisons with C freundii Ballerup [979 (20)] Citrobacter sp 30_2
[1064 (23)] C youngae [1654 (31)] C koseri [1693 (34)] and C rodentium [1634
(34)]
143
Figure 5-6 Comparisons of coding sequences between strain C freundii ADS47 and five
Citrobacter species Coding sequences of ADS47 were compared with C freundii Ballerup
Citrobacter sp 30_2 C youngae C koseri and C rodentium genomes based on all-versus-all
reciprocal FASTA comparison of at least 30 amino acid identity and gt70 coverage of the
total length using OrthoMCL program A Venn diagram demonstrates the shared and
unshared (indicated in the brackets) coding sequences
144
Table 5-3 shows differences in the 26 RAST categorized genes between the six
Citrobacter genomes Even though a low abundance of virulencediseasedefense category
genes were found in the animalhuman pathogenic bacteria of C rodentium (85) and C koseri
(92) significantly higher mycobacterium virulence genes (13) were identified in this two
species The genomes of the two closely related strains C freundii ADS47 and C freundii
Ballerup lack this type of virulence gene Instead elevated numbers of genes for resistance to
antibiotic and toxic compounds were found in C freundii ADS47 (94) C freundii Ballerup
(88) and Citrobacter sp (110) compared to other three species Copper homeostasis and
cobalt-zinc-cadmium resistance genes were identified in strain C freundii ADS47 (13 and 21
respectively) and closely related C freundii Ballerup (12 and 22) and Citobacter sp (18 and
22) Significantly lower numbers of these genes were present in C rodentium (6 and 4) and C
koseri (7 and 8) C rodentium and C koseri lack mercuric reductase mercuric resistance and
arsenic resistance genes perhaps because being animalhuman pathogens but these genes
were found in the remaining four genomes The C koseri genome is richer in betandashlactamase
genes (5) compared to the remaining genomes which encodes 1-3 genes
145
Table 5-3 Functional categorized gene abundances among the genomes of C freundii
ADS47 C freundii Ballerup Citrobacter sp 30_2 C youngae C koseri and C rodentium
The genomes were classified into different functional categories with the RAST program
which classified the total CDSs
RAST subsystem category Citrobacter species
C
freundii
ADS47
C
freundii
Ballerup
C
sp
C
youngae
C
koseri
C
rodentium
Cofactorsvitaminsprosthetic
groupspigments
323
317
310
311
325
304
Cell wall and capsule 279 272 248 262 247 262
Virulence disease and defense 113 107 134 100 92 85
Potassium metabolism 50 53 33 32 30 28
Miscellaneous 210 228 200 185 245 185
Phageprophagetransposable
element plasmid
24 75 25 4 6 49
Membrane transport 170 156 171 195 171 172
Iron acquisition and metabolism 80 81 86 62 113 27
RNA metabolism 220 222 217 215 238 212
Nucleosides and nucleotides 119 112 110 111 120 114
Protein metabolism 249 280 269 298 287 254
Cell division and cell cycle 30 29 32 29 38 29
Motility and chemotaxis 133 124 90 147 143 133
Regulation and cell signaling 126 130 124 135 132 123
Secondary metabolism 21 22 20 21 0 0
DNA metabolism 145 167 165 155 147 152
Fatty acids lipids and isoprenoids 121 149 122 121 134 133
Nitrogen metabolism 50 52 53 49 56 58
Dormancy and sporulation 3 3 3 4 3 3
146
Table 5-3 (continued)
RAST subsystem category
Citrobacter species
C
freundii
ADS47
C
freundii
Ballerup
C
sp
C
youngae
C
koseri
C
rodentium
Respiration 198 184 180 191 166 162
Stress response 236 235 170 177 179 174
Metabolism of aromatic
compounds
20 20 28 25 17 48
Amino acids and derivatives 482 476 450 466 428 416
Sulfur metabolism 48 50 52 57 56 56
Phosphorus metabolism 50 49 43 42 43 28
Carbohydrates 837 803 708 731 733 737
147
Strains C freundii ASD47 and C freundii Ballerup have much more potassium
metabolism category genes (50 and 53 respectively) than the other bacterial species which
code for 28-33 proteins Larger numbers of genes associated with the glutathione regulated
potassium efflux system were also identified in C freundii ASD47 with 23 and C freundii
Ballerup with 26 compared to the remaining four species that encode only five genes All of
the six genomes contained phageprophage-related genes with the maximum identified in C
freundii Ballerup (75) followed by C rodentium (49) Citrobacter sp (25) and C freundii
ADS47 (24) Lower numbers of phageprophage-related genes were found in C koseri (6) and
C youngae (4)
The six Citrobacter genomes showed remarkable variability in their secretion systems
The type-III secretion operon genes are only found in C rodentium and C freundii Ballerup
Type-II secretion genes are predominant in C rodentium (12) followed by ADS47 (9)
Type-IV secretion system is uncommon in Citrobacter species since it was only
identified in Citrobacter sp where it is represented by 19 genes Variable numbers of type-VI
secretion system genes are present in the five genomes except C koseri which lacks such
genes C youngae has the highest number (43) of type-VI secretion proteins The other strains
have a range of type-VI secretion proteins in particular 27 were identified in C freundii
Ballerup 15 in C freundii ADS47 and 8 in Citrobacter sp Type-VII (chaperoneusher)
secretion system genes were found in all of the six Citrobacter species with the largest (42)
and smallest (13) numbers of these genes occurring in C koseri and C freundii Ballerup
respectively
Although a relatively similar number of ABC transporters (37-42) were identified in
the six genomes C freundii ADS47 contain the largest number of tripartite ATP-independent
148
periplasmic (TRAP) transporter genes (7) By contrast the closely related C freundii
Ballerup lacks such transporter system An identical number of iron acquisition and
metabolism category genes were found in C freundii ADS47 (80) and C freundii Ballerup
(81) while the two pathogenic species C rodentium and C koseri code for the lowest (27)
and highest (113) numbers of such genes C koseri is rich in siderophore related genes (64)
but the remaining five genomes have smaller numbers of siderophore-associated genes (20-
22)
Variable numbers of motility and chemotaxis related genes were found in the
Citrobacter genomes with the largest number occurring in C youngae (147) followed by C
koseri (143) C freundii ADS47 (133) C rodentium (133) C freundii Ballerup (124) and
Citrobacter sp (90) The two pathogenic species (C rodentium and C koseri) encode a
remarkably low number of quorum sensing genes (2) In contrast C youngae codes for the
highest number of quorum sensing genes (19) A common and distinctive feature of the two
pathogenic species is the lack of genes required for secondary metabolism In contrast the
non-pathogenic species contain a common set of genes for degrading a range of
phenylpropanoids and cinnamic acid (20-22)
The lowest number of genes related to phospholipidsfatty acids biosynthesis and
metabolism were found in C freundii ADS47 (66) compared to C freundii Balllerup (94) and
C koseri (98) The smallest numbers of nitratenitrite ammonification genes were found in
ADS47 (31) and its closest genome C freundii Ballerup (32) In contrast the two pathogens
encode the highest number of such proteins (39-40)
C rodentium and C koseri code for significantly low numbers of respiratory category
genes (162-165) while C freundii ADS47 contains the highest number of such genes (198)
149
Further analysis of the subcategory genes determined that the two gut species (C youngae and
Citrobacter sp 30_2) contain the largest numbers of genes encoding anaerobic reductases
(22) and identical numbers of formate dehydrogenases (17) The largest numbers of formate
dehydrogenase genes were identified in C freundii Ballerup (32) and C freundii ADS47 (30)
but the two pathogenic species differed from the other species by encoding the lowest number
(13) of c-type cytochrome biogenesis genes
A higher number of aromatic compound metabolism category genes were found in the
mouse pathogen C rodentium (48) compared to the human pathogen (C koseri) which
contain 17 genes A significantly higher number of genes involved in central aromatic
intermediate metabolism (26) were found in C rodentium compared to the mouse pathogen
(C koseri) and other genomes which only contain 6-8 such genes The two C freundii strains
ADS47 and Ballerup lacked the anaerobic degradation of aromatic compound
(hydroxyaromatic decarboxylase family) genes
In spite of the abundance of stress response category genes strain ADS47 has only a
small number of desiccation stress genes (5) compared to the other species which ranged
from 9 to10 However ADS47 and Ballerup code for higher number of genes related to
oxidative stress (94 and 91 respectively) and cold shock (8 and 7 respectively) than the other
species
The smallest numbers of amino acids and its derivatives category genes were found in
the two pathogens C rodentium and C koseri (416 and 428 respectively) while the largest
number of these genes were identified in strain ADS47 (482) and C freundii Ballerup (476)
Despite the similar number of sulfur metabolism category genes determined across the six
species only ADS47 lacks taurine utilization genes Strains ADS47 and Ballerup were found
150
to be rich in the phosphorous metabolism gene category (49-50) while C rodentium contains
a significantly decreased number of these category genes (28)
The bioinformatics analyses found the largest number of carbohydrate category genes
in ADS47 (837) followed by Ballerup (803) The gut bacteria (Citrobacter sp 30_2) codes for
the least number of genes assigned to the carbohydrate category (708) while C koseri C
rodentium and C youngae contained 731-737 in this category genes The analyses also
determined that there are relatively small numbers of TCA and Entner Doudoroff pathway
genes in C rodentium and C youngae compared to the other four species and relatively large
numbers of fermentation-related genes in ADS47 (81) and C freundii (85) compared to the
other four species
A comparison of the pathogenicity operons [locus of enterocyte effacement (LEE)] of
C rodentium with other five Citrobacter genomes revealed the absence of this virulence
determinant gene region in the other four Citrobacter species including ADS47 (Figure 5-7)
The analysis showed the presence of LEE operon genes in C freundii strain Ballerup as no
gap was observed in that genomic location while C freundii ADS47 C koseri C youngae
and Citrobacter sp 30_2 lacked that loci (7a) Further ananlysis showed alignments between
C rodentium and E coli LEE operon genes (Figure 7b) In contrast no alignment of such
genes was observed in ADS47 genome
151
(a)
152
Figure 5-7 (a) Blast Ring Image Generator (BRIG) output based of Citrobacter rodentium
(NC_013716) compared against ADS47 C freundii C koseri C youngae E coli and
Citrobacter sp genomes The targeted comparative genome analysis was performed based on
locus of enterocyte effacement (LEE) operons of C rodentium (Petty et al 2011) The
innermost rings show GC content (black) and GC skew (purple and green) The remaining
rings show BLAST comparisons of the other Citrobacter and E coli genomes against C
rodentium The region shown in red is LEE operon genes (b) The absence of LEE operons in
the ADS47 genome comparing to C rodentium and E coli Analysis was performed and
visualized by Artemis Comparison Tool software v132
(httpwwwsangeracukresourcessoftwareact) (Tatusov et al 2000 Craver et al 2005)
LEE operons
(b)
153
546 Analysis of the unique genes in C freundii ADS47
Two hundred and seventy genes unique to ADS47 were identified from a comparison
of its genome to the other five Citrobacter species In general the unique genes are
distributed throughout the ADS47 genome but some clusters of unique genes occurred
including eight unique genomic regions (Figure 5-8) Six large unique regions are located at
767204-775090 (~8 kb) 1025572-1042510 (17 kb) 2798289-2807217 (~9 kb)
3045220-3055912 (~11 kb) 3930366-3943075 (12 kb) and 4022195-4032444 (10 kb)
Out of 270 unique genes 147 (54 ) are hypothetical proteins The remaining 123
(46) genes are grouped into reductasesoxidoreductases (9 genes) de-acetylase (1)
hydrolases (2) transferases (4) sulfatasesarylsulfatases (3) bacteriocins (3) type VI
secretion proteins (2) kinases (4) transporters (11) membraneexported proteins (8)
regulators (12) integraserecombinases (5) phagesprophagestransposaes related genes (27)
and miscellaneous (32) that includes DNA helicases ligases fimbrial proteins and
chromosome partitioning genes (Table 5-4)
The unique regions encode various functional genes that may cause trichothecene de-
epoxidation or de-acetylation reactions The 8 kb unique regions contains six genes encoding
glycosyltransferases and two copies of oxidoreductase-like coding sequences (CMD_0711
and CMD_0712) that showed 39 identity to the Marinobacter aquaeolei F420
hydrogenasedehydrogenase beta subunit The largest (17 kb) unique genomic region codes
for 16 genes including dehydrogenase hydrolase bacteriocin (pyocin) integrase and a short
chain dehydrogenasereductase-like (SDR) (CMD_0979) gene (Figure 5-9a)
154
The SDR like gene is 49 amino acid similarity to an oxidoreductase of the
phytopathogenic Dickeya dadantii and the enzyme contains a domain with 77 homology to
the Dickeya zeae NAD(P) binding site The 9 kb unique genomic section contains 8 genes
including a NaCl symporter a dehydratase (CMD_2679) with 86 homology to
Enterobacteriaceae bacterium dihydroxy-acid dehydratase and a putative oxidoreductase
(CMD_2683) having 37 similarity with oxopropionate reductase of Bacillus sp The unique
region also encodes a short chain dehydrogenasereductase (SDR) (CMD_2684) that shows
85 identity to Enterobacteriaceae bacterium (Figure 5-9b) A unique thiol-disulfide and
thioredoxin like reductase (CMD_3721) was identified in a 10 kb unique region that contains
13 genes (Figure 5-9c) This gene is 59 identical to Oligotropha carboxidovorans thiol-
disulfide isomerase and thioredoxin In addition several reductasesoxidoreductase-like
unique genes were identified in the ADS47 genome including an oxidoreductase
(CMD_1024) with 27 identity to Canadidatus nitrospira NADH-quinone oxidoreductase
membrane L Another oxidoreductase-like gene identified in a different location of the
ADS47 genome (CMD_1393) shows 87 homology to Klebsiella pneumoniae
deoxygluconate dehydrogenase contains a SDR active site A putative nitrate reductase gene
(CMD_3184) has a low level of identity (27) with an uncultured bacterial respiratory nitrate
reductase The ~11 kb unique region encodes sulfatasesarylsulfatses
(CMD_2898CMD_2905) and related genes Furthermore ADS47 contains a unique
glycosylphosphatidylinositol (GPI) anchor biosynthesis operon comprised of four genes
including a N-acetylglucosaminyl-phosphatidylinositol de-acetylase (CMD_2431) which
showed 78 similarity to E coli de-acetylase (Figure 5-9e)
155
Other unique genespredicted genes in the ADS47 genome include
polysaccharidephenazine biosynthesis genes (CMD_0710CMD_1406) heavy metal kinase
(CMD_0989) two component heavy metal regulator (CMD_0990) nitrate reductase regulator
(CMD_3477) putative potassiumzinc transporters (CMD_0973CMD_4600) sodium
symporter (CMD_2678) (tripartite ATP-independent periplasmic transporters) TRAP
transporters (CMD_3693 CMD_3694 CMD_3695 CMD_4337) glutamineribose ABC
transporters (CMD_3176CMD_3544) toxin transcription regulator (CMD_3464) fimbrial
proteinchaperone (CMD_1695 CMD_2434 CMD_3277 CMD_3278) and ParB
chromosomeplasmid partitioning proteins (CMD_4033CMD_4034)
Majority of the unique genes and their distributions are shown in comparative circular
genome maps (Figure 5-8)
156
Table 5-4 Unique genes of C freundii strain ADS47 Genes were categorized according to
the functional similarity The unique genes were determined by comparing ADS47 with five
sequenced Citrobacter species using OrthoMCL software
Unique gene classes Number
Total unique genes
270
Reductases oxidoreductases 9
De-acetylase 1
Hydrolase 2
Transferase 4
Sulfatase 3
Bacteriocin 2
Type VI secretion 2
Kinase 4
Transporter 9
Membraneexported protein 8
Regulator 12
Integraserecombinase 5
Phageprophagetransposae 27
Miscellaneous 34
Hypothetical protein 147
157
158
Figure 5-8 Locations of the unique genes of C freundii strain ADS47 The unique genes
were determined by comparing the total CDS of strain C freundii ADS47 with C freundii
Ballerup Citrobacter sp C youngae C koseri and C rodentium CDSs by OrthoMCL
program The map was developed by Blast Ring Generator (BRIG) software First second
and third circles from inside to out side show the genome size GC and GC skew of strain
ADS47 Forward (Fwd) and reversed (Rev) transcribed CDSs of ADS47 are shown in green
(forth circle) and red (fifth circle) respectively The next five rings represent the circular
genome maps of five Citrobacter species distinguished by different ring colours (C
rodentium as sky blue C koseri as dark blue C youngae as light blue C freundii Ballerup as
yellow and Citrobacter sp as green) The degree of orthologous gene identity between
ADS47 and other five genomes are shown by color intensity (light color= 75 gene identity
ash color= 50 gene identity) Gaps across five genome rings indicate the absence of
genesregions compared to ADS47 Unique genes are shown surrounding the map rings and
the identities of some of the genes are given Arrows indicate putative
reductasesoxidoreductases and de-acetylase
159
(a)
(b)
160
(c)
(d)
161
Figure 5-9 Unique genomic regions and associated genes in C freundii ADS47 compared to
five Citrobacter species RAST server based analysis compared the unique genesregions of
ADS47 with the publicly available databases The upper panels demonstrate the large unique
regions defined by yellow boxes and genes located in the genomic regions are indicated by
arrows The lower panels are closely related genesoperons found in distantly related species
The unique regions that contain potential trichothecene de-epoxidation genes are shown in (a)
short chain dehydrogenasereductase (SDR) and dehydrogenase (b) putative oxidoreductase
dehydratase and SDR (c) thiol-disulfide isomerase and thioredoxin oxidoreductase and (d)
unique glycosylphosphatidylinositol (GPI) biosynthesis operon having N-acetylglucosaminyl-
phosphatidylinositol de-acetylase gene
162
55 Discussion
The de novo sequenced ADS47 genome was oriented and assembled using a
completely sequenced C koseri genome as reference strain The synteny analysis between
ADS47 and C koseri showed very similar overall organization of the two genomes The high
preservation of genes among five Citrobacter species and ADS47 were emphasized by the
close alignments observed by Blast Ring Image Generator (BRIG) outputs obtained from
these genomes
Genome-wide phylogenetic analyses based on a large number of common CDSs
(2821) observed within the six Citrobacter genomes confirmed the species identification
based on 16S-rDNA sequencing of the trichothecene mycotoxin-detoxifying C freundii strain
ADS47 The base sequence idendity was 8715 between these two genomes They shared
~80 CDS and had a similar GC content (52) Citrobacter are Gram-negative rod shaped
and facultatively anaerobic bacteria belonging to the family of Enterobacteriaceae (Wang et
al 2000 Whalen et al 2007) Strains ADS47 and Ballerup of C freundii are both aerobic
gram-negative bacilli that have numerous flagella and inhabit in soil (Cong et al 2011 Islam
et al 2012) C freundii is considered to have a positive environmental role by reducing nitrate
to nitrite (Rehr and Klemme 1989) which is an important step in the nitrogen cycle
C freundii C rodentium and C koseri are animalhuman pathogenic species C
rodentium is a natural mice pathogen causing hemorrhagic colitis and hemolytic uremic
syndrome (Gareau et al 2010) C koseri causes human disease (Badger et al 1999 Kariholu
et al 2009) particularly neonatal meningitis and C freundii is an opportunistic human
pathogen responsible for a various types of infections like respiratory tract urinary tract and
blood (Whalen et al 2007) C freundii is very common representing approximately 29 of
163
all opportunistic human infections (Hodges et al 1978) Outside of animals they can be found
in soil water sewages and different organs of diseased animals (Wang et al 2000) These
pathogens code for large numbers of mycobacterium virulence operonsgenes which were
absent in the three other genomes However they have fewer genes involved in environmental
stress such as copper homeostasis genes and no mercuricarsenic resistance and secondary
metabolism category genes unlike the other Citrobacter genomes Nonetheless considerable
variability exists between the two pathogens correlates with C rodentium causing disease in
mice C koseri causing meningistis in human and C freundii causing infections in different
human organsThere were attaching and effacing (AE) lesion forming (LEE) operons in C
rodentium and C freundii but not in C koseri Those genes have shown to be acquired by
lateral gene transfer playing major roles in virulence of enteropathogenic and
enterohemorrhagic E coli and C rodentium (Mundy et al 2005 Petty et al 2010 Petty et al
2011 The ~356 kb pathogenic LEE gene island is comprised of five operons (LEE1 to
LEE5) that code for ~40 virulence genes that include type-III secretion genes and associated
effectors (Berdichevsky et al 2005) In addition an elevated number of type-II and -VI
secretion proteins identified in C rodentium which may act as additional virulence factors
(Petty et al 2010) Because it lacks type-III secretion apparatus genes it is likely that C
koseri uses a different strategy to cause human neonatal meningitis (Badger et al 1999 Ong
et al 2010)
To adapt to different environmental niches or hosts bacteria like Citrobacter species
frequently gainlose genes increase repeat sequences and recombine genomic regions using
different mechanisms (Parkhill et al 2001 Petty et al 2011) The existence of large numbers
of phageprophagetransposae related genes in the Citrobacter genomes particularly in C
164
freundii Ballerup suggests that their genomes are unstable For example C freundii Ballerup
and C koseri might have acquired LEE operons by horizontal transfer via phage mediated
transpositions Interestingly although ADS47 is closely related to C freundii Ballerup it
lacks the pathogen-related LEE gene island and mycobacterium virulence operons required
for invasion and intracellular survival in macrophages (Gao et al 2006) Instead ADS47 has a
larger number and variety of stress-responsive genes such as efflux genes and bacteriocins
(pyocin hemolysin) genes that provide protection against heavy metals and antibiotics and
increase their competitiveness against other bacteria These findings suggest that the strain
ADS47 is not an opportunistic animal pathogen but has evolved to be highly competitive in
the soil micro-environment (Arnold 1998 Bengoechea and Skurnik 2000) Perhaps ADS47
evolved trichothecene degrading genes for adaptation to soils containing trichothecene
mycotoxins contaminated soils (Gadd and Griffiths 1978 Sharma and Fulekar 2009 Han et
al 2011) Conversely the absence or reduced number of stress-responsive genes in C
rodentium and C koseri may imply that those two pathogenic species are not exposed to such
stressful and xenobiotic-contaminated environments as ADS47
This interpretation is supported by the observation that the two of the three
animalhuman pathogenic species contained high of rRNA copy number but lower numbers
were found in ADS47 C freundii Ballerup and Citrobacter sp 30_2 Bacteria maintain large
number of rRNA for higher production of ribosome during high growth rates (Stevenson et al
1997 Klappenbach et al 2000) such as might occur during the establishment of an infection
However a large copy number of rRNAs may not be metabolically favourable for bacteria
such as ADS47 that evolved in a highly competitive environment as that costs higher cellular
energy (Hsu et al 1994)
165
C freundii Ballerup seems to be an exception in that it is similar to the other animal
pathogens for containing virulent factors (ie LEE-operon genes) but more like the
environmental species for encoding high number genes for stress defence heavy metal and
toxic compounds resistances
The comparison of the genes in C freundii ADS47 with the unique ability to detoxify
trichothecenes with genes in other Citrobacter species allowed approximately 270 unique
genes to be identified in ADS47 Some of those genes may be involved in trichothecene de-
epoxidation or de-acetylation Currently the pathways and microbial enzymes leading to
trichothecene de-epoxidation and de-acetylation are unknown but based on the required
reaction ie the elimination of an oxygen atom from epoxide moiety it is likely that epoxide
reductases or epoxide oxidoreductases are responsible for such biodegradation activity (He et
al 1992)
From database searches three enzymes having epoxide reductase activity were found
These are a vitamin K epoxide reductase (VKOR) a monooxygenase PaaABCE and pyridine
nucleotide-disulfide oxidoreductase (PNDO) which reduce vitamin K epoxide phenylacetyl-
CoA and epoxyalkanes respectively (Swaving et al 1996 Li et al 2004 Teufel et al 2012)
In mammals VKOR plays important roles in vitamin K cycle by reducing vitamin K epoxide
to vitamin K and its homologues that are found in plants bacteria and archaea (Goodstadt
2004 Li et al 2010) The microbial multi-component PaaABCE monooxygenase catalyzes
degradation of variety of aromatic compounds or environmental contaminants via epoxidation
of phenylacetic acid (Paa) and NADPH-dependant de-epoxidation of epoxy-Paa (Luengo et
al 2001 Teufel et al 2012) PNDO is a unique type of NADP-dependent oxidoreductase
identified in Xanthobacter strain Py2 that was obtained by propane enrichment procedure
166
(van Ginkel and de Bont 1986) However no orthologs of these genes were found in ADS47
genome
Nine ADS47-unique reductaseoxidoreductase-like genes are potential candidates for
the trichothecene de-epoxidation enzyme Genes belong to short chain
dehydrogenasereductase (SDR) family are structurally and functionally diverse and cause
different enzymatic activities including xenobiotic degradation (Kavanagh et al 2008) This is
a promising candidate for the trichothecene de-epoxidation gene Genes belong to dehydratase
family catalyze reduction reactions by cleaving carbon-oxygen bonds of organic compounds
that result in removal of oxygen in various biosynthetic pathways (Flint et al 1993) This
mechanism may be relevant to microbial trichothecene de-epoxidation reaction since the latter
involves removal of the oxygen atom forming the epoxide in trichothecene mycotoxins
(Young et al 2007)
Another ADS47-unique gene was for a nitrate reductase that may be a candidate de-
epoxidation gene Under stress conditions such an enzyme causes reductive removal of an
oxygen molecule (Morozkina and Zvyagilskaya 2007)
A fourth type of ADS47-unique gene is for a thiol-disulfide isomerasethioredoxin like
(Trx) oxidoreductase The thiol-disulfide isomerase and thioredoxin oxidoreductases contain a
thioredoxin like domain with a CXXC catalytic motif which catalyzes various biological
processes that allow the organism to adapt to environmental stress and develop competence to
sporulate (Scharf et al 1998 Petersohn et al 2001 Smits et al 2005) The membrane-
anchored Trxndashlike domain initiates the vitamin K de-epoxidation reaction by reducing the
active motif (CXXC) of VKOR (Schulman et al 2010)
167
The present study did not find any homologues of VKOR in ADS47 genome but we
identified two copies of structurally similar quinone reductase protein disulfide bond
formation (DsbB) gene (CMD_1044 and CMD_2641) Both VKOR and DsbB contain four
transmembrane segments an active site CXXC motif and cofactor quinone and use a
common electron transfer pathway for reducing substrates (Li et al 2010 Malojcic et al
2008) As VKOR bacterial DsbB requires a Trx-like partner (eg DsbA) for activity Further
experiments are required to examine if DON de-epoxidation occurs by the catalytic activity of
DsbB coupled with a unique thiol-disulfide isomerase and thioredoxin oxidoreductase present
in ADS47 genome
The analysis also identified an ADS47-unique glycosylphosphatidylinositol (GPI)
biosynthetic operon that is comprised of four genes including an N-
acetylglucosaminylphosphatidylinositol de-acetylase The GPI membrane anchor protein
synthesis is initiated by transferring N-acetylglucosamine from uridine diphosphate N-
acetylglucosamine to phosphatidylinositol by N-acetylglucosaminyltransferase an
endoplasmic reticulum (ER) membrane-bound protein complex (Hong and Kinoshita 2009)
In the second step of the pathway N-acetylglucosamine-phosphatidylinositol is de-acetylated
to produce glucosaminyl-phosphatidylinositol by the catalytic activity of a zinc
metalloenzyme N-acetylglucosaminylphosphatidylinositol de-acetylase which is localized on
the cytoplasmic side of the ER (Nakamura et al 1997 Hong and Kinoshita 2009) It is
possible that the identified unique N-acetylglucosaminylphosphatidylinositol de-acetylase
gene of ADS47 may be involved in trichothecene de-acetylation activity
168
Further analysis is needed to determine which of these candidate gene or genes may be
involved in tricthothecene mycotoxin de-epoxidation and de-acetylation activities
Identification of gene responsible for the trichothecene detoxification activities of C freundii
ADS47 opens many potential applications for its use in agri-based industries to contribute to
cropanimal production and food safety (Karlovsky 2011)
56 Acknowledgement
We gratefully acknowledge Agriculture and Agri-Food CanadaBinational
Agricultural Research and Development Fund (AAFCBARD) project for providing the
financial supports of the work
169
Supplemental Table S5-1 List of unique genes of strain ADS47 gene IDs and their
functional roles The unique genes were determined by comparing the ADS47 genome with
five sequenced Citrobacter species (C freundii Citrobacter sp C youngae C koseri and C
rodentium) using OrthoMCL (Li et al 2003)
Unique gene
classes
Number Gene ID and functions
Total unique
genes
270 123 known functions 147 hypothetical proteins
Reductases
oxidoreductases
9 CMD_0711 coenzyme F420 hydrogenasedehydrogenase beta subunit
CMD_0979 Short chain dehydrogenase reductase (SDR) Oxidoreductase
SDR dehydrogenase CMD_1024 NADH-quinone oxidoreductase
membrane L CMD_1393 oxidoreductase SDR putative deoxygluconate
dehydrogenase (NAD+ 3-oxidoreductase) CMD_2679
Dehydratasedihydroxy acid dehydratase CMD_2683 oxidoreductase
oxopropionate reductase CMD_2684 short-chain dehydrogenasereductase
SDR CMD_3184 nitrate reductase alfa subunit CMD_3721 thiol-disulfide
isomerase and thioredoxin oxidoreductase
Deacetylase 1 CMD_2431 N-acetylglucosaminylphosphatidylinositol deacetylase
superfamily
Hydrolase 2 CMD_0991 Dienelactone hydrolase CMD_2895 Putative glycosyl
hydrolase
Transferase 4 CMD_1640 D12 class N6 adenine-specific DNA methyltransferase
CMD_2432 Glycosyltransferase CMD_2433 putative acetyltransferase
CMD_3023 adenine-specific DNA modification methyltransferase
Sulfatase 3 CMD_2898 Possible sulfatase arylsulfatasearylsulfatransferase
CMD_2902 Arylsulfatasesulfatase CMD_2905 Putative
sulfatasearylsulfatase
Bacteriocin 3 CMD_0982 Thermosatble hemolysin superfamily CMD_0993 Pyocin S2
superfamily (bacteriocin that kills other bacterial strain) CMD_3705 S-type
pyocin containing domain protein
Type VI secretion 2 CMD_1802 EvpB family type VI secretion protein CMD_2364 type VI
effector Hcp1
Kinase 4 CMD_0989 Heavy metal Sensor kinase CMD_1794 Polo kinase kinase
(PKK) superfamily CMD_3124 predicted diacylglycerol kinase CMD_4266
putative signal transduction kinase
Transporter 11 CMD_0973 TrkA-N domain-containing protein (K uptake transporter)
CMD_1407 Branched-chain amino acid transport system carrier protein
CMD_2678 Sodium Symporter CMD_3176 glutamine ABC transporter
CMD_3544 Ribose ABC transport system CMD_3693 TRAP-type C4-
dicarboxylate transport system binding protein CMD_3694 TRAP-type C4-
dicarboxylate transport system binding protein small permease component
(predicted N-acetylneuraminate transporter) CMD_3695 TRAP-type C4-
dicarboxylate transport system large permease CMD_4337 23-diketo-L-
gulonate TRAP transporter small permease protein yiaM component
CMD_4338 23-diketo-L-gulonate TRAP transporter large permease protein
yiaN CMD_4600 putative transporter involved in Zinc uptake (YciA
protein)
170
Supplmental Table S5-1 (continued)
Unique gene
classes
Number Gene ID and functions
Membrane
exported protein
8 CMD_0713 putative membrane protein wzy CMD_0983 Putative exported
protein precursor CMD_0986 Putative transmembrane protein
CMD_0987 putative exported protein precursor CMD_1458 putative
exported protein CMD_2903 Putative exported protein CMD_2904
putative secreted 5-nucleotidase CMD_4291 (S) putative integral
membrane export protein
Regulator 12 CMD_0977 probable transcriptional regulator protein CMD_0980 Putative
transcription activator TenA family CMD_0990 Two component heavy
metal response transcription regulator CMD_1766 Leucine responsive
regulatory protein CMD_2677 transcriptional regulator GntR CMD_2909
putative arylsulfatase regulator CMD_3279 Uncharacterized outer
membrane usher protein yqiG precursor CMD_3464 toxin transcription
regulatory protein CMD_3465 EAL regulator protein (signaling)
CMD_3475 putative transcription regulator CMD_3477 Fumarate and
nitrate reduction regulatory protein (for anaerobic adaptation) CMD_3812
putative regulator from phage
Integrase
recombinase
5 CMD_0785 CMD_0992 CMD_0994 CMD_4032 CMD_4424
Phage prophage
transposae related
genes
27 Phages CMD_1608 CMD_1609 CMD_1610 CMD_1611 CMD_1619
CMD_1631 CMD_1913 CMD_1914 CMD_1915 CMD_1917
CMD_1920 CMD_1921 CMD_1936 CMD_3351 CMD_3804
CMD_3805 CMD_3807 CMD_3808 CMD_4581 CMD_4609
CMD_4610 Transposae CMD_0252 CMD_0398 Prophage CMD_1645
CMD_3806 CMD_4425 CMD_4597
Miscellaneous 32 CMD_0095 ATP-dependent DNA helicase Rep CMD_0656 LysA protein
CMD_0710 Polysaccharide biosynthesis family protein CMD_0978 TPR
repeat containing protein CMD_0981 AMP dependant synthatase and
ligase CMD_0985 Cupin superfamily CMD_0987 flavodoxin
CMD_1404 Putative endoribonuclease CMD_1405 Putative PAC domain
protein CMD_1406 Phenazine (PhzF) biosynthesis protein CMD_1695
Putative fimbrial protein CMD_1793 HPChromosome segregation ATPase
domain protein CMD_1922 FRG uncharacterized protein domain
CMD_2206 L-fucose isomerase like protein CMD_2434 CS12 fimbria
outer membrane usher protein CMD_2680 HpcHHpaI aldosecitrate lyase
protein family CMD_2681 Aldose -1 epimerase CMD_2682
Gluconolaconase LRE like region CMD_2900 Putative cytoplasmic
protein CMD_3009 putative restriction endonuclease CMD_3277 Putative
yqi fimbrial adhesion CMD_3278 Hypothetical fimbrial chaperone yqiH
CMD_3340 Pea-VEAacid family CMD_3518 3-hydroxy-3-methylglutaryl
CoA synthase CMD_3801 putative reverse transcriptase CMD_3840 beta-
D-galactosidase alpha subunit CMD_4033 ParB family chromosome
partitioning protein CMD_4034 ParBrepB chromosomeplasmid
partitioning protein CMD_4035 RelASpoT protein CMD_4532 putative
DNA ligase -like protein CMD_4541 Antitermination protein Q and
CMD_4585 putative ATP dependent helicase
171
CHAPTER 6
60 CONCLUSION
61 Summary
Contamination of grain with mycotoxins including deoxynivalenol (DON) and other
trichothecenes is becoming a limiting factor for cereal and animal production (Binder et al
2007) To address the issue this study obtained a novel microbial source for DON
detoxification and determined candidate genes for the function which may have potential
applications to detoxify grain and grain-derived foodfeed contaminated with DON and TC
mycotoxins
From a screen of soil samples from crop field microorganism was isolated that is able
to de-epoxidize the most frequently detected foodfeed contaminating mycotoxin DON The
effects of media compositions temperature pH and aerobicanaerobic conditions on
microbial detoxification activity were assessed From 150 analyzed soil samples a mixed soil
microbial culture that showed DON de-epoxidation activity under aerobic conditions and
moderate temperature (~24degC) was obtained This culture exhibited a slow rate of microbial
DON to de-epoxyDON transformation (de-epoxidation) possibly because of competition
between de-epoxidizing and other microorganisms in the culture
The mixed microbial culture was enriched for DON detoxifying bacteria by reducing
the non-targeted microbial species through antibiotic treatments heating and subculturing on
agar medium and selecting colonies The enriched culture had higher DON de-epoxidation
activity than the initial culture Because of low abundance of DON detoxifying
microorganisms further culture enrichments were required to increase the target microbial
172
population Reculture of the partially enriched microbial community in nutritonally poor MSB
medium with high level of DON increased the DON de-epoxidizing bacterial population
Serial dilution plating and selecting single colonies eventually led to isolation of a DON-
detoxifying bacterial strain named ADS47
Phenotypic and 16S-rDNA sequence analyses provided evidence that the bacterium is
a strain of C freundii Strain ADS47 is a gram negative rod-shaped facultative anaerobe and
has peritrichous flagella Strain ADS47 was unable to use DON as sole carbon source
however increasing DON concentrations in growth media enhanced its DON metabolism
efficiency The pure bacterial culture was shown to be able to rapidly de-epoxidize DON
under aerobic conditions moderate temperatures and neutral pH Strain ADS47 more quickly
de-epoxidizes DON in aerobic conditions than the anaerobic conditions In addition to DON
strain ADS47 showed capability to de-epoxidize andor de-acetylate 10 different type-A and
type-B TC mycotoxins which often co-contaminant grain with DON
To clone the DON detoxifying genes a DON detoxifying bacterial DNA library in E
coli host was constructed followed by screening of the library clones for DON detoxification
activity The study produced a library giving approximately 10 fold coverage of the genome
of strain ADS47 Restriction digestion analysis determined that most of the clones contained
ADS47 DNA inserts This indicates that a good DNA library was produced which had gt99
probability of containing all the DNA of the ADS47 genome However activity-based
screening of the library could not identify any clones with DON de-epoxidation ability It is
assumed that bacterial DON de-epoxidation genes evolved in a certain environmental niche
which may be required for the DON de-epoxidation to be expressed and active but was not
supplied by the host E coli cells
173
An attempt was made to identify the bacterial genes encoding DON de-epoxidation
and de-acetylation enzymes by sequencing the genome of strain ADS47 and comparing its
sequence to the genomes of related bacteria that are not known to posses DON de-epoxidation
activity A RAST server based bioinformatics analysis determined that the 48 Mb circular
genome of ADS47 codes for 4610 predicted CDS A genome wide phylogenetic analysis
determined that strain ADS47 was gt87 identical to the pathogen C freundii strain Ballerup
The relationship to two other animalhuman pathogenic species C rodentium and C koseri
was more distant This result is consistent with the 16S-rDNA based phylogenetic analysis
It is assumed that the microbial DON detoxifying genes were acquired by horizontal
gene transfer or other means during the adaptation of ADS47 in mycotoxin contaminated soil
If it is correct then this means that those newly acquired genes are not common in other
related microbial species and no other Citrobacter species is known to degrade DON Hence
the study identified 270 unique genes to ASDS47 with a subset of 10 genes for reductases
oxidoreductases and a de-acetylase which were considered to be candidate genes for the
enzymes that have DON detoxifying activity Furthermore a targeted comparative genome
analysis showed that the mycotoxin detoxifying ADS47 genome does not harbour the
clinically important LEE encoded typendashIII secretion system and mycobacterium virulence
genes This suggests that strain ADS47 is not likely to be an animal pathogenic bacterium
unlike C freundii Ballerup which encodes the pathogenic relevance LEE operons
174
62 Research contributions
The research made four major contributions which may aid in reducing the risks of
animals and humans from DONTC exposure Firstly the study identified a novel bacterial
strain ADS47 that is able to detoxify DON and 10 related mycotoxins over a wide range of
temperatures under aerobic and anaerobic conditions The discovery has led to a patent
application for strain ADS47 supporting the novelty and importance of the finding This
bacterial strain could be used to detoxify trichothecene mycotoxins in contaminated animal
feed thus reducing food safety risks
Secondly the study resulted in the development of a method for isolating DON de-
epoxidizing microbes from soil as well as optimizing the conditions for bacterial DONTC
detoxification activity to increase the effectiveness usefulness of the strain
Thirdly the thesis work resulted in availability of the genome sequence of a
trichothecene detoxifying microbe which may ultimately lead to an understanding of the
cellularmolecular mechanisms of DONTC de-epoxidation and de-acetylation
genespathways The genome analysis resulted in identification of several candidate genes for
DONTC de-epoxidation and de-acetylation activities Genes which show de-epoxidation
activity could be used to develop DONTC resistant cereal cultivars Fourthly strain ADS47
lacks major animal pathogenicity genes (in particular the LEE-operons) as well as does not
encodes mycobacterium virulence genes Thus strain ADS47 has a great potential to be
utilized to detoxify cereal grain and may be used as a feed additive for livestockfishery All
together the reported work may have significant applications to produce and supply
DONTC-free food and feed
175
63 Further research
This research provides the basis for developing biological detoxification of DON and
related mycotoxins This goal might be achieved by using DON de-epoxidizing gene to
develop resistant plants enzymatic detoxification of contaminated feed and use of the bacteria
as a feed additive Follow-up studies may reach the above indicated goals Because the
bacterium showed de-epoxidation activity under aerobicanaerobic conditions and under wide
range of temperatures it is likely to have an application for enzymatic detoxification of
contaminated animal feed Therefore additional studies on bacterial efficiency to detoxify
contaminated cereal and grain-derived feed need to be conducted Although strain ADS47
does not encode major animal pathogenicity genes the bacterium must be directly tested on
animals to see if it could negatively affect animal digestive systems If not then the potential
for ADS47 is considerable for its use as a feed additive for pig poultry and fish Animal
feeding trials will also need to be performed to determine if DON detoxification is sufficient
to be cost effective as a feed additive In order to develop DONTC resistant cereal cultivars
the gene responsible to DON degrading activity among the presently identified candidate
genes (reductases oxidoreductases and de-acetylase) will need to be expressed in other
organisms like E coli and a codon modified version adapted for expression in plants like
corn and wheat
176
70 REFERENCES
Abbas HK Mirocha CJ Pawlosky RJ Pusch DJ 1985 Effect of cleaning milling and
baking on deoxynivalenol in wheat Applied and Environmental Microbiology 50482ndash
486
Accensi F Pinton P Callu P Abella-Bourges N Guel WJF Grosjean F Oswald IP 2006
Ingestion of low doses of deoxynivalenol does not affect haematological biochemical or
immune responses of piglets Journal of Animal Science 841935ndash1942
Agrawal D Mahapatra AK 2005 Vertically acquired neonatal Citrobacter brain abscessndash
case report and review of the literature Journal of Clinical Neuroscience 12188ndash190
Alexander NJ 2008 The TRI101 story engineering wheat and barley to resist Fusarium head
blight World Mycotoxin Journal 131-37
Alexander NJ Proctor RH McCormick SP 2009 Genes gene clusters and biosynthesis of
trichothecenes and fumonisins in Fusarium Toxin Reviews 28198ndash215
Ali ML Taylor JH Jie L Sun G William M Kasha KJ Reid LM Pauls KP 2005
Molecular mapping of QTLs for resistance to Gibberella ear rot in corn caused by
Fusarium graminearum Genome 48521-533
Alikhan N-F Petty NK Zakour NLB Beatson SA 2011 BLAST Ring Image Generator
(BRIG) simple prokaryote genome comparisons BMC Genomics 12402
Alisi A Spaziani A Anticoli S Ghidinelli M Balsano C 2008 PKR is a novel functional
direct player that coordinates skeletal muscle differentiation via p38MAPKAKT
pathways Cellular Signalling 20534ndash542
177
Amuzie CJ Shinozuka J Pestka JJ 2009 Induction of suppressors of cytokine signaling by
the trichothecene deoxynivalenol in the mouse Toxicological Science 111277ndash287
Ansorge WJ 2009 Next generation DNA sequencing techniques New Biotechnology
25195-203
Arnold JW Senter SD 1998 Use of digital aroma technology and SPME GC-MS to compare
volatile compounds produced by bacteria isolated from processed poultry Journal of the
Science of Food and Agriculture 78343ndash348
Audenaert K Van Broeck R Bekaert B De Witte F Heremans B Messens K Hofte
M Haesaert G 2009 Fusarium head blight (FHB) in Flanders population
diversity inter-species associations and DON contamination in commercial winter
wheat varieties European Journal of Plant Pathology 125445-458
Awad WA Ghareeb K Bohm J Zentek J 2010 Decontamination and detoxification
strategies for the Fusarium mycotoxin deoxynivalenol in animal feed and the
effectiveness of microbial biodegradation Food Additives and Contaminants 27510-520
Aziz K Bartels D Best AA DeJongh M Disz T Edwards RA Formsma K Gerdes S Glass
EM Kubal M Meyer F Olsen GJ Olson R Osterman AL Overbeek RA McNeil LK
Paarmann D Paczian T Parrello1 B Pusch GD Reich C Stevens R Vassieva O
Vonstein V Wilke A Zagnitko O 2008 The RAST Server Rapid Annotations using
Subsystems Technology BMC Genomics 975
Badger JL Stins MF Kim KS 1999 Citrobacter freundii invades and replicates in human
brain microvascular endothelial cells Infection and Immunology 674208ndash4215
Bae HK Pestka JJ 2008 Deoxynivalenol induces p38 interaction with the ribosome in
monocytes and macrophages Toxicological Science 10559-66
178
Bai GH Desjardins AE Plattner RD 2002 Deoxynivalenol-nonproducing Fusarium
graminearum causes initial infection but does not cause disease spread in wheat spikes
Mycopathologia 15391ndash98
Barrett JR 2000 Mycotoxins of molds and maladies Environmental Health Perspectives
108A20ndashA23
Bengoechea JA Skurnik M 2000 Temperature-regulated efflux pumppotassium antiporter
system mediates resistance to cationic antimicrobial peptides in Yersinia Molecular
Microbiology 3767ndash80
Bennett JW Klich M 2003 Mycotoxins Clinical Microbiology Reviews 16497ndash516
Benson DA Karsch-Mizrachi I Lipman DJ Ostell J Sayers EW 2009 GenBank Nucleic
Acids Research 37D26-31
Berdichevsky T Friedberg D Nadler C Rokney A Oppenheim A Rosenshine I 2005 Ler is
a negative autoregulator of the LEE1 operon in neuropathogenic Escherichia coli Journal
of Bacteriology 187349ndash357
Bernardo A Bai GH Guo PG Xiao K Guenzi AC Ayoubi P 2007 Fusarium graminearum-
induced changes in gene expression between Fusarium head blight-resistant and
susceptible wheat cultivars Functional and Integrative Genomics 769ndash77
Bhat RV Beedu SR Ramakrishna Y Munshi KL 1989 Outbreak of trichothecene
mycotoxicosis associated with consumption of mould-damaged wheat production in
Kashmir Valley India The Lancet 33335ndash37
Binder EM Tan LM Chin LJ Handl J Richard J 2007 Worldwide occurrence of
mycotoxins in commodities feeds and feed ingredients Animal Feed Science and
Technology 137265-282
179
Birzele B Meier A Hindorf H Kraumlmer J Dehne H-W 2002 Epidemiology of Fusarium
infection and deoxynivalenol content in winter wheat in the Rhineland Germany
European Journal of Plant Pathology 108667ndash673
Boddu J Cho S Muehlbauer GJ 2007 Transcriptome analysis of trichothecene-induced gene
expression in barley Molecular PlantndashMicrobe Interactions 201364ndash1375
Borenshtein D Schauer DB 2006 The Genus Citrobacter Prokaryotes 690ndash98
Boutigny A-L Richard-Forget F Barreau C 2008 Natural mechanisms for cereal resistance
to the accumulation of Fusarium trichothecenes European Journal of Plant Pathology
121411ndash423
Boutrif E Canet C 1998 Mycotoxin prevention and control FAO programmes Revue de
Meacutedecine Veacuteterinaire 149681-694
Brennan JM Leonard G Fagan B Cooke BM Ritieni A Ferracane R et al 2007
Comparison of commercial European wheat cultivars to Fusarium infection of head and
seedling tissue Plant Pathology 5655ndash64
Brenner DJ Grimont PA Steigerwalt AG Fanning GR Ageron E Riddle CF 1993
Classification of Citrobacteria by DNA hybridization designation of Citrobacter farmeri
sp nov Citrobacter youngae sp nov Citrobacter braakii sp nov Citrobacter
werkmanii sp nov Citrobacter sedlakii sp nov and three unnamed Citrobacter
genomospecies International Journal of Systematic Bacteriology 43645-58
Brookes G Barfoot P 2006 GM Crops The first ten years-global socio-economic and
environmental Impacts no 36 ISAAA (International Service for the Acquisition of Agri-
Biotech Applications) Ithaca New York
180
Bruce KD Hughes MR 2000 Terminal restriction fragment length polymorphism monitoring
of genes amplified directly from bacterial communities in soils and sediments Molecular
Biotechnology 16261ndash269
Buerstmayr H Ban T Anderson JA 2009 QTL mapping and marker-assisted selection for
Fusarium head blight resistance in wheat A review Plant Breeding 1281ndash26
Campbell H Choo TM Vigier B Underhill L 2002 Comparison of mycotoxin profiles
among cereal samples from eastern Canada Canadian Journal of Botany 80526ndash532
Carver et al 2005 ACT the Artemis Comparison Tool Bioinformatics 213422-3423
Carver et al 2009 DNAPlotter circular and linear interactive genome visualization
Bioinformatics 25119-120
Chen CY et al 2003 Comparative genome analysis of Vibrio vulnificus a marine pathogen
Genome Research 132577ndash2587
Chiu TC Yen JH Hsieh YN Wang YS 2005 Reductive transformation of dieldrin under
anaerobic sediment culture Chemosphere 601182ndash1189
Chung YJ Zhou HR Pestka JJ 2003 Transcriptional and posttranscriptional roles for p38
mitogen-activated protein kinase in upregulation of TNF-alpha expression by
deoxynivalenol (vomitoxin) Toxicology and Applied Pharmacology 193188ndash201
Chung EJ Lim HK Kim J-C Choi GJ Park EJ Lee MH Chung YR Lee S-W 2008 Forest
soil metagenome gene cluster involved in antifungal activity expression in Escherichia
coli Applied and Environmental Microbiology 74723-730
Collins TFX Sprando RL Black TN Olejnik N Eppley RM Hines FA Rorie J Ruggles DI
2006 Effects of deoxynivalenol (DON vomitoxin) on in utero development in rats Food
and Chemical Toxicology 44747ndash757
181
Cong Y Wang J Chen Z Xiong K Xu Q Hu F 2011 Characterization of swarming motility
in Citrobacter freundii FEMS Microbiology Letters 371160-171
Cossette F Miller JD 1995 Phytotoxic effect of deoxynivalenol and Gibberella ear rot
resistance of corn Natural Toxins 3383ndash388
Creppy EE 2002 Update of survey regulation and toxic effects of mycotoxins in Europe
Toxicological Letters 12719-28
Dai JS Wang D Guerlebeck C Laturnus S Guenther Z Lu SC Ewers C 2010 Suppression
subtractive hybridization identifies an autotransporter adhesin gene of E coli IMT5155
specifically associated with avian pathogenic Escherichia coli (APEC) BMC
Microbiology 10236
Daniel R 2005 The metagenomics of soil Nature Review Microbiology 3470-478
Davis KER Joseph SJ Janssen PH 2005 Effects of growth medium inoculum size and
incubation time on culturability and isolation of soil bacteria Applied and Environmental
Microbiology 71826-834
Delcher AL Phillippy A Carlton J Salzberg SL 2002 MUMmer 21 NUCmer and
PROmer are described in Fast Algorithms for Large-scale Genome Alignment and
Comparisonrdquo Nucleic Acids Research 302478-2483
Demain AL Adrio JL 2008 Contributions of microorganisms to industrial biology
Molecular Biotechnology 3841ndash455
Deng W Li Y Vallance BA Finlay BB 2001 Locus of enterocyte efffacement from
Citrobacter rodentium sequence analysis and evidence for horizontal transfer among
attaching and effacing pathogens Infection and Immunity 696323ndash6335
182
Desjardins AE Proctor RH Bai G McCormick SP Shaner G Beuchley G Hohn TM 1996
Reduced virulence of trichothecene-nonproducing mutants of Gibberella zeae in wheat
field tests Molecular Plant-Microbe Interactions 9775-781
Desjardins AE 2006 Fusarium mycotoxins chemistry genetics and biology The American
Phytopathological Society St Paul Minnesota USA p 260
Desjardins AE Proctor RH 2007 Molecular biology of Fusarium mycotoxins International
Journal of Food Microbiology 11947-50
Desmond OJ Manners JM Stephens AE MacLean DJ Schenk PM Gardiner DM Munn
AL Kazan K 2008 The Fusarium mycotoxin deoxynivalenol elicits hydrogen peroxide
production programmed cell death and defence responses in wheat Molecular Plant
Pathology 9435-445
Dill-Macky R Jones RK 2000 The effect of previous crop residues and tillage on Fusarium
head blight of wheat Plant Disease 8471ndash76
DrsquoMello JPF Macdonald AMC Dijksma WTP 1998 3-Acetyl deoxynivalenol and esterase
production in a fungicide-insensitive strain of Fusarium culmorum Mycotoxin Research
149ndash18
DrsquoMello JPF Placinta CM Macdonald AMC 1999 Fusarium mycotoxins a review of
global implications of animal health welfare and productivity Animal Feed Science and
Technology 80183-205
Doohan FM Brennan J Cooke BM 2003 Influence of climatic factors on Fusarium species
pathogenic to cereals European Journal of Plant Pathology 109755ndash768
183
Dyer RB Kendra DF Brown DW 2006 Real-time PCR assay to quantify Fusarium
graminearum wild-type and recombinant mutant DNA in plant material Journal of
Microbiological Methods 67534ndash542
Erecinska M Wilson DF 1981 Inhibitors of cytochrome c oxidase In inhibitors of
mitochondrial function edited by M Erecinska and DF Wilson Pergamon Press Ltd
Oxford pp 145-160
Eriksen GS 2003 Metabolism and toxicity of trichothecene Doctoral thesis Swedish
University of Agricultural Sciences Uppsala Sweden
Eriksen GS Pettersson H Lundh T 2004 Comparative cytotoxicity of deoxynivalenol
nivalenol their acetylated derivatives and de-epoxy metabolites Food Chemistry and
Toxicology 42619-624
Eudes F Comeau A Rioux S Collin J 2000 Phytotoxicity of eight mycotoxins associated
with Fusarium in wheat head blight Canadian Journal of Plant Pathology 22286ndash292
FAO (Food and Agriculture Organization) 1991 Food Nutrition and Agriculture Food for
the Future volume 1 FAO Rome
FAO (Food and Agriculture Organization) 2001 FNP73 Manual of the application of
HACCP system in mycotoxin prevention and control FAO Food and Nutrition paper no
73 FAO Rome
Fedorak PM Grbic-Galic D 1991 Aerobic microbial cometabolism of benzothiophene and
3-methylbenzothiophene Applied and Environmental Microbiology 57932-940
Felsenstein J 1989 PHYLIP - Phylogeny Inference Package (Version 32) Cladistics 5164-
166
184
Fernando WGD Amarasinghe C Demoz B Surujdeo‐Maharaj S Gauthier V Guerrieri G
Al‐Taweel K Brucircleacute‐Babel A Tamburic‐Illincic L Gilbert J 2011 What do we know of
the Fusarium graminearum chemotypes their epidemiology genetics and infection
process in Canada In Proc 7th
Canadian Workshop on Fusarium Head Blight edited by
J Gilbert A Tekauz N Sweetland and K Slusarenko Winnipeg Manitoba Canada p 45
Fierer N Jackson RB 2006 The diversity and biogeography of soil bacterial community
Proceedings of the National Academy of Sciences 103626-631
Fink-Gremmels J 1989 The significance of mycotoxin assimilation for meat animals
Deutsche Tierarztliche Wochenschrift 96360-363
Flint DH Emptage MH Finnegan MG Fu W Johnson MK 1993 The role and properties of
the iron-sulfur cluster in Escherichia coli dihydroxy-acid dehydratase Journal of
Biological Chemistry 26814732ndash14742
Foroud NA Eudes F 2009 Trichothecenes in cereal grains International Journal of
Molecular Science 10147-173
Forsell JH Jensen R Tai JH Witt M Lin WS Pestka JJ 1987 Comparison of acute
toxicities of deoxynivalenol (vomitoxin) and 15-acetyldeoxynivalenol in the B6C3F1
mouse Food and Chemical Toxicology 25155ndash162
Frey M Stettner C Gierl A 1998 A general method for gene isolation in tagging approaches
amplification of insertion mutagenised sites (AIMS) The Plant Journal 13717-721
Freeman GG Morrison RI 1949 The isolation and chemical properties of trichothecin an
antifungal substance from Trichothecium roseum Link Biochemical Journal 441ndash5
185
Fuchs E Binder EM Heidler D Krska R 2002 Structural characterization of metabolites
after the microbial degradation of type A trichothecenes by the bacterial strain BBSH 797
Food Additives and Contaminants 19379ndash386
Gabor EM Alkema WBL Janssen DB 2004 Quantifying the accessibility of the
metagenome by random expression cloning techniques Environmental Microbiology
6879ndash886
Gadd GM Griffiths AJ 1978 Microorganisms and heavy metal toxicity Microbial Ecology
4303ndash317
Gale LR Bryant JD Calvo S Giese H 2005 Chromosome complement of the fungal plant
pathogen Fusarium graminearum based on genetic and physical mapping and cytological
observations Genetics 171985ndash1001
Gao LY Pak M Kish R Kajihara K Brown EJ 2006 A mycobacterial operon essential for
virulence in vivo and invasion and intracellular persistence in macrophages Infection and
Immunity 741757ndash1767
Gareau MG Wine E Reardon C et al 2010 Probiotics prevent death caused by Citrobacter
rodentium infection in neonatal mice Journal of Infectious Diseases 20181e91
Gareis M Bauer J Thiem J Plank G Grabley S Gedek B 1990 Cleavage of
zearalenoneglycoside a masked mycotoxin during digestion in swine Journal of
Veterinary Medicine 37236-240
Geng L Xin W Haung DW Feng G 2006 A universal cloning vector using vaccinia
topoisomerase I Molecular Biotechnology 3323-28
186
Golkari S Gilbert J Ban T Procunier JD 2009 QTLspecific microarray gene expression
analysis of wheat resistance to Fusarium head blight in Sumai-3 and two susceptible
NILs Genome 52409ndash418
Goodstadt L Ponting CP 2004 Vitamin K epoxide reductase homology active site and
catalytic mechanism Trends in Biochemical Sciences 29289-292
Goswami RS Kistler HC 2004 Heading for disaster Fusarium graminearum on cereal
crops Molecular Plant Pathology 5515-525
Gouze ME Laffitte J Printon P Dedieux G Galinier A Thouvenot JP Loiseau N Oswald
IP Galtier P 2007 Effect of subacute oral doses of nivalenol on immune and metabolic
defence systems in mice Veterinary Research 38635ndash646
Guan S He J Young JC Zhu H Li X Ji C Zhou T 2009 Transformation of trichothecene
mycotoxins by microorganisms from fish digesta Aquaculture 290290ndash295
Gupta R Beg QK Lorenz P 2002 Bacterial alkaline proteases molecular approaches and
industrial applications Applied Microbiology and Biotechnology 5915ndash32
Haberle J Schweizer G Schondelmaier J Zimmermann G Hart L 2009 Mapping of QTL
for resistance against Fusarium head blight in the winter wheat population
PelikanBussardNing8026 Plant Breeding 12827ndash35
Hall BG 2004 Predicting the evolution of antibiotic resistance genes Nature Reviews
Microbiology 2430-435
Han JI et al 2011 Complete genome sequence of the metabolically versatile plant growth-
promoting endophyte Variovorax paradoxus S110 Journal of Bacteriology 1931183ndash
1190
187
Harvey RB Edrington TS Kubena LF Elissalde MH Corrier DE Rottinghaus GE 1995
Effect of aflatoxin and diacetoxyscirpenol in ewe lambs Bulletin of Environmental
Contamination and Toxicology 54325-330
Harvey RB Edrington TS Kubena LF Elissalde MH Casper HH Rottinghaus GE Turk JR
1996 Effects of dietary fumonisin B1-containing culture material deoxynivalenol-
contaminated wheat or their combination on growing barrows American Journal of
Veterinary Research 571790-1794
He J 2007 Microbial strategies for management of Fusarium graminearum and
transformation of deoxynivalenol Master of Science Thesis University of Guelph
Guelph Ontario Canada
He K Vines L Pestka JJ 2010a Deoxynivalenol-induced modulation of microRNA
expression in RAW 2647 macrophages-A potential novel mechanism for translational
inhibition Toxicologist 114310
He J Zhou T Young JC Boland GJ Scott PM 2010b Chemical and biological
transformations for trichothecene mycotoxins in human and animal food chains A review
Trends in Food Science and Technology 2167-76
He J Zhou T 2010 Patented techniques for detoxification of mycotoxins in feeds and food
matrices Rec Pat Food Nutr Agric 296ndash104
He P Young LG Forsberg C 1992 Microbial transformation of deoxynivalenol (vomitoxin)
Applied and Environmental Microbiology 583857-3863
Hell K Cardwell KF Setamou M Poehling H-M 2000 The influence of storage practices on
aflatoxins contamination in maize in four agroecological zones in Benin West Africa
Journal of Stored Product Research 36365ndash82
188
Henne A Schmitz RA Bomeke M Gottschalk G Daniel R 2000 Screening of
environmental DNA libraries for the presence of genes conferring lipolytic activity on
Escherichia coli Applied and Environmental Microbiology 663113ndash3116
Hodges GR Degener CE Barnes WG 1978 Clinical significance of Citrobacter isolates
American Journal of Clinical Pathology 7037ndash40
Hollinger K Ekperigin HE 1999 Mycotoxicosis in food producing animals The Veterinary
Clinics of North America Food Animal Practice 15133-165
Hong Y Kinoshita T 2009 Trypanosome glycosylphosphatidylinositol biosynthesis Korean
Journal of Parasitology 47197ndash204
Hope RJ Aldred D Magan N 2005 Comparison of environmental profiles for growth and
deoxynivalenol production by Fusarium culmorum and F graminearum on wheat grain
Letters in Applied Microbiology 40295ndash300
Horvath RS 1972 Microbial co-metabolism and the degradation of organic compounds in
nature Bacteriological Reviews 36146ndash155
Hsu D Shih LM Zee YC 1994 Degradation of rRNA in Salmonella strains a novel
mechanism to regulate the concentrations of rRNA and ribosomes Journal of
Bacteriology 1764761ndash4765
Huwig A Freimund S Kappeli O Dutler H 2001 Mycotoxin detoxification of animal feed
by different adsorbents Toxicological Letters 122179ndash188
Iordanov MS Pribnow D Magun JL Dinh TH Pearson JA Magun BE 1997 Ribotoxic
stress response activation of the stress activated protein kinase JNK1 by inhibitors of the
peptidyl transferase reaction and by sequence-specific RNA damage to the sarcinricin
loop in the 28S rRNA Molecular and Cellular Biology 173373ndash3381
189
Islam R Zhou T Young JC Goodwin PH Pauls KP 2012 Aerobic and anaerobic de-
epoxidation of mycotoxin deoxynivalenol by bacteria originating from agricultural soil
World Journal of Microbiology and Biotechnology 287-13
Jackson LS Bullerman LB 1999 Effect of processing on Fusarium mycotoxins Advances in
Experimental Medicine and Biology 459243-261
Jansen C von Wettstein D Schafer W Kogel K-H Felk A Maier FJ 2005 Infection patterns
in barley and wheat spikes inoculated with wild-type and trichodiene synthase gene
disrupted Fusarium graminearum Proceedings of the National Academy of Sciences
10216892ndash16897
Jube S Borthakur D 2007 Expression of bacterial genes in transgenic tobacco methods
applications and future prospects Electronic Journal of Biotechnology 10452-467
Kabak B Dobson AD 2009 Biological strategies to counteract the effects of mycotoxins
Journal of Food Protection 722006-2016
Kant P Reinprecht Y Martin CJ Islam R Pauls KP 2011 Integration of biotechnologies
disease resistance pathology and Fusarium In Comprehensive Biotechnology edited by
Moo-Young M Second Edition 4729-743
Kaplan CW Kitts CL 2004 Bacterial succession in a petroleum land treatment unit Applied
and Environmental Microbiology 701777-1786
Kaprelyants AS Kell DB 1996 Do bacteria need to communicate with each other for
growth Trends in Microbiology 4237-42
190
Kariholu U Rawal J Namnyak S 2009 Neonatal Citrobacter koseri meningitis and brain
abscess The Internet Journal of Pediatrics and Neonatology 10
(httpwwwispubcomjournalthe-internet-journal-of-pediatrics-and-
neonatologyvolume-10-number-1neonatal-citrobacter-koseri-meningitis-and-brain-
abscesshtml) (accessed on April 19 2012)
Karlovsky P 1999 Biological detoxification of fungal toxins and its use in plant breeding
feed and food production Natural Toxins 71ndash23
Karlovsky P 2011 Biological detoxification of the mycotoxin deoxynivalenol and its use in
genetically engineered crops and feed additives Applied Microbiology and
Biotechnology 91491-504
Kavanagh KL Jornvall H Persson B Oppermann U 2008 Medium and short-chain
dehydrogenasereductase gene and protein families the SDR superfamily functional and
structural diversity within a family of metabolic and regulatory enzymes Cellular and
Molecular Life Sciences 653895ndash3906
Khera KS Whalen C Angers G Vesonder RF Kuiper-Goodman T 1982 Embryotoxicity of
4-deoxynivalenol (vomitoxin) in mice Bulletin of Environmental Contamination and
Toxicology 29487ndash491
Kikot GE Hours RA Alconada TM 2009 Contribution of cell wall degrading enzymes to
pathogenesis of Fusarium graminearum a review Journal of Basic Microbiology 49
231ndash241
Kim UJ Shizuya H Jong PJ Birren B Simon MI 1992 Stable propogation of cosmid sized
human DNA inserts in a F factor based vector Nucleic Acids Research 201083-1065
191
Kimura M Kaneko I Komiyama M Takatsuki A Koshino H Yoneyama K Yamaguchi I
1998 Trichothecene 3-O-acetyltransferase protects both the producing organism and
transformed yeast from related mycotoxins cloning and characterization of Tri101
Journal of Biological Chemistry 2731654-1661
Kimura M Anzai H Yamaguchi I 2001 Microbial toxins in plantndashpathogen interactions
biosynthesis resistance mechanisms and significance The Journal of General and
Applied Microbiology 47149ndash160
Kimura M Tokai T Takahashi-Ando N Ohsato S Fujimura M 2007 Molecular and genetics
studies of Fusarium trichothecene biosynthesis pathways genes and evolution
Bioscience Biotechnology and Biochemistry 712105-2123
King RR McQueen RE Levesque D Greenhalgh R 1984 Transformation of deoxynivalenol
(vomitoxin) by rumen microorganisms Journal of Agricultural and Food Chemistry
32181ndash1183
Klappenbach JA Dunbar JM Schmidt TM 2000 rRNA operon copy number reflects
ecological strategies of bacteria Applied and Environmental Microbiology 661328ndash
1333
Knietsch A Waschkowitz T Bowien S Henne A Daniel R 2003 Metagenomes of complex
microbial consortia derived from different soils as sources for novel genes conferring
formation of carbonyls from short-chain polyols on Escherichia coli Journal of Molecular
Microbiology and Biotechnology 546ndash56
Kollarczik B Gareis M Hanelt M 1994 In vitro transformation of the Fusarium mycotoxins
deoxynivalenol and zearalenone by the normal gut microflora of pigs Natural Toxins
2105ndash110
192
Koonin EV Wolf YI Aravind L 2001 Prediction of the archaeal exosome and its
connections with the proteasome and the translation and transcription machineries by a
comparative-genomic approach Genome Research 11240ndash252
Kouadio JH Dano SD Moukha S Mobio TA Creppy EE 2007 Effects of combinations of
Fusarium mycotoxins on the inhibition of macromolecular synthesis malondialdehyde
levels DNA methylation and fragmentation and viability in Caco-2 cells Toxicon
49306ndash317
Krause L Diaz NN Bartels D Edwards RA Puumlhler A et al 2006 Finding novel genes in
bacterial communities isolated from the environment Bioinformatics 22281ndash289
Kubena LF Huff WE Harvey RB Phillips TD Rottinghaus GE 1989 Individual and
combined toxicity of deoxynivalenol and T-2 toxin in broiler chicks Poultry Science
68622ndash626
Kubena LF Harvey RB Huff WE Corrier DE Phillips TD Rottinghaus GE 1990 Efficacy
of hydrated sodium calcium aluminosilicate to reduce the toxicity of aflatoxin and T-2
toxin Poultry Science 691078ndash1086
Kubena LF Harvey RB Huff WE Elissalde MH Yersin AG Phillips TD Rottinghaus GE
1993 Efficacy of hydrated sodium calcium aluminosilicate to reduce the toxicity of
aflatoxin and diacetoxyscirpenol Poultry Science 7251ndash59
Kubena LF Edrington TS Harvey RB Buckley SA Phillips TD Rottinghaus GE Casper
HH 1997 Individual and combined effects of fumonisin B1 present in Fusarium
moniliforme culture material and T-2 toxin or deoxynivalenol in broiler chicks Poultry
Science 761239ndash1247
193
Kumar RA Gunasekaran P Lakshmanan M 1999 Biodegradation of tannic acid by
Citrobacter freundii isolated from a tannery effluent Journal of Basic Microbiology
39161-168
Kushiro M 2008 Effects of milling and cooking processes on the deoxynivalenol content in
wheat International Journal Molecular Science 92127-2145
Lane DJ 1991 16S23S rRNA sequencing In Nucleic acid techniques in bacterial
systematic edited by Stackebrandt E and Goodfellow M Wiley J and Sons Inc New
York pp 115-175
Lancova K Hajslova J Poustka J Krplova A Zachariasova M Dostalek P Saschambula L
2008 Transfer of Fusarium mycotoxins and ldquomaskedrdquo deoxynivalenol (deoxynivalenol-3-
glucoside) from field barley through malt to beer Food Additives and Contaminants
25732ndash744
Langmead B Trapnell C Pop M Salzberg SL 2009 Ultrafast and memory-efficient
alignment of short DNA sequences to the human genome Genome Biology 10R25
Larsen JC Hunt J Perrin I Ruckenbauer P 2004 Workshop on trichothecenes with a focus
on DON summary report Toxicological Letters 1531-22
Lee T Oh DW Kim HS Lee J Kim YH Yun SH Lee YW 2001 Identification of
deoxynivalenol- and nivalenol-producing chemotypes of Gibberella zeae by using PCR
Applied and Environmental Microbiology 672966ndash2972
Lee S-W Won K Lim HK Kim J-C Choi GJ Cho KY 2004 Screening for novel lipolytic
enzymes from uncultured soil microorganisms Applied and Environmental Microbiology
65720-726
194
Li L Christian J Stoeckert J David SR 2003 OrthoMCL Identification of Ortholog Groups
for Eukaryotic Genomes Genome Research 132178-2189
Li T Chang CY Jin DY Lin PJ Khvorova A Stafford DW 2004 Identification of the gene
for vitamin K epoxide reductase Nature 427541ndash544
Li Y 2007 Sensory signal transduction in the vagal primary afferent neurons Current
Medicinal Chemistry 142554ndash2563
Li MX Pestka JJ 2008 Comparative induction of 28S ribosomal RNA cleavage by ricin and
the trichothecenes deoxynivalenol and T-2 toxin in the macrophage Toxicological
Science 10567ndash78
Li R Li Y Kristiansen K Wang J 2008 SOAP short oligonucleotide alignment program
Bioinformatics 24713-4
Li W Schulman S Dutton RJ Boyd D Beckwith J Rapoport TA 2010 Structure of a
bacterial homologue of vitamin K epoxide reductase Nature 463507-512
Lichstein HC Malcolm HS 1943 Studies of the effect of sodium azide on microbic growth
and respiration Journal of Bacteriology 47221ndash230
Limay-Rios V Schaafsma AW 2011 2010 Update on the development of an integrated
mycotoxin management system in Ontario corn 30th
Centralia Swine Research Update
Kirkton Ontario Canada
Llorens A Hinojo MJ Mateo R Gonzalez-Jaen MT Valle- Algarra FM Logrieco A Jimenez
M 2006 Characterization of Fusarium spp isolates by PCR-RFLP analysis of the
intergenic spacer region of the rRNA gene (rDNA) International Journal of Food
Microbiology 106287ndash306
195
Lombaert GA Pellaers P Roscoe V Mankotia M Neil R Scott PM 2003 Mycotoxins in
infant cereal foods from the Canadian retail market Food Additives and Contaminants
20494ndash504
Luengo JM Garciacutea JL Olivera ER 2001 The phenylacetyl-CoA catabolon a complex
catabolic unit with broad biotechnological applications Molecular Microbiology
391434ndash1442
Luo X 1994 Food poisoning caused by Fusarium toxins In Proceedings of the Second
Asian Conference on Food Safety ILSI Thailand pp 129ndash136
Mabagala RB 1997 The effect of populations of Xanthomonas campestris pv phaseoli in
bean reproductive tissues on seed infection of resistant and susceptible bean genotypes
European Journal of Plant Pathology 103175ndash181
Madhyastha MS Marquardt RR Abramson D 1994 Structurendashactivity relationships and
interactions among trichothecene mycotoxins as assessed by yeast bioassay Toxicon
321147ndash1152
Maier FJ Miedaner T Hadeler B Felk A Salomon S Lemmens M Kassner H Schaumlfer W
2006 Involvement of trichothecenes in fusarioses of wheat barley and maize evaluated
by gene disruption of the trichodiene synthase (Tri5) gene in three field isolates of
different chemotype and virulence Molecular Plant Pathology 7449ndash461
Malojcic G Owen RL Grimshaw JP Glockshuber R 2008 Preparation and structure of the
charge-transfer intermediate of the transmembrane redox catalyst DsbB FEBS Letters
5823301ndash3307
196
Manoharan M Dahleen LS Hohn TM Neate SM Yu XH Alexander NJ et al 2006
Expression of 3-OH trichothecene acetyltransferase in barley (Hordeum vulgare L) and
effects on deoxynivalenol Plant Science 171699ndash706
Maresca M Mahfoud R Garmy N Fantini J 2002 The mycotoxin deoxynivalenol affects
nutrient absorption in human intestinal epithelial cells The Journal of Nutrition
1322723ndash2731
Martins ML Martins HM 2002 Influence of water activity temperature and incubation time
on the simultaneous production of deoxynivalenol and zearalenone in corn (Zea mays) by
Fusarium graminearum Food Chemistry 79315ndash318
Masuda D Ishida M Yamaguchi K Yamaguchi I Kimura M Nishiuchi T 2007 Phytotoxic
effects of trichothecenes on the growth and morphology of Arabidopsis thaliana Journal
of Experimental Botany 581617ndash1626
Matsumoto E Kawanaka Y Yun SJ Oyaizu H 2008 Isolation of dieldrin- and endrin-
degrading bacteria using 12-epoxycyclohexane as a structural analog of both compounds
Applied Microbiology and Biotechnology 801095-1103
McLean M 1996 The phytotoxicity of Fusarium metabolites an update since 1989
Mycopathologia 133163ndash179
Merhej J Richard-Forget F Barreau C 2011 Regulation of trichothecene biosynthesis in
Fusarium recent advances and new insights Applied Microbiology and Biotechnology
91519ndash528
Midwest Plan Service 1987 Grain Drying Handling Storage Handbook Ames Iowa Iowa
State University 2nd edition
197
Miller SS Reid LM Harris LJ 2007 Colonization of maize silks by Fusarium graminearum
the causative organism of Gibberella ear rot Canadian Journal of Botany 85369-376
Minervini F Fornelli F Flynn KM 2004 Toxicity and apoptosis induced by mycotoxins
nivalenol deoxynivalenol and fumonisin B1 in a human erythroleukemia cell line
Toxicology In Vitro 1821ndash28
Mitterbauer R Poppenberger B RaditschnigA Lucyshyn D Lemmens M Glossl J Adam G
2004 Toxin-dependent utilization of engineered ribosomal protein L3 limits trichothecene
resistance in transgenic plants Plant Biotechnology Journal 2329ndash340
Mocali S Benedetti A 2010 Exploring research frontiers in microbiology the challenge of
metagenomics in soil microbiology Research Microbiology 161497-505
Morozkina EV Zvyagilskaya RA 2007 Nitrate reductases structure functions and effect of
stress factors Biochemistry 721151ndash1160
Munkvold GP 2003 Cultural and genetic approaches to managing mycotoxins in maize
Annual Review of Phytopathology 4199-116
Mundy R MacDonald TT Dougan G Frankel G Wiles S 2005 Citrobacter rodentium of
mice and man Cellular Microbiology 71697ndash1706
Nakamura N Inoue N Watanabe R Takahashi M Takeda J Stevens VL Kinoshita T 1997
Expression cloning of PIG-L a candidate N-acetylglucosaminyl-phosphatidylinositol
deacetylase The Journal of Biological Chemistry 27215834ndash15840
Nganje WE Bangsund DA Leistritz FL Wilson WW Tiapo NM 2002 Estimating the
economic impact of a crop disease the case of Fusarium head blight in US wheat and
barley National Fusarium Head Blight Forum Proceedings East Lansing Michigan State
University pp 275ndash281
198
Nielsen C Lippke H Didier A Dietrich R Martlbauer E 2009 Potential of deoxynivalenol
to induce transcription factors in humanhepatoma cells Molecular Nutrition and Food
Research 53479ndash491
Nielsen JKS Vikstroumlm AC Turner P Knudsen LE 2011 Deoxynivalenol transport across
the human placental barrier Food and Chemical Toxicology 492046-2052
Nishiuchi T Masuda D Nakashita H Ichimura K Shinozaki K Yoshida S et al 2006
Fusarium phytotoxin trichothecenes have an elicitor-like activity in Arabidopsis thaliana
but the activity differed significantly among their molecular species Molecular Plantndash
Microbe Interactions 19512ndash520
Ohsato S Ochiai-Fukuda T Nishiuchi T Takahashi-Ando N Koizumi S Hamamoto H
Kudo T Yamaguchi I Kimura M 2007 Transgenic rice plants expressing trichothecene
3-O-acetyltransferase show resistance to the Fusarium phytotoxin deoxynivalenol Plant
Cell Reports 26531-538
Okubara PA Blechl AE McCormick SP Alexander NJ Dill-Macky R Hohn TM 2002
Engineering deoxynivalenol metabolism in wheat through the expression of a fungal
trichothecene acetyltransferase gene Theoritical and Applied Genetics 10674-83
Ong CL Beatson SA Totsika M Forestier C McEwan AG Schembri MA 2010 Molecular
analysis of type 3 fimbrial genes from Escherichia coli Klebsiella and Citrobacter
species BMC Microbiology 10183
Osborne LE Stein JM 2007 Epidemiology of Fusarium head blight on small-grain cereals
International Journal of Food Microbiology 119103-108
Page RDM 1996 TREEVIEW An application to display phylogenetic trees on personal
computers Computer Applications in the Biosciences 12357-358
199
Parkhill J Dougan G James KD Thomson NR Pickard D Wain J Churcher C Mungall KL
Bentley SD Holden MT et al 2001 Complete genome sequence of a multiple drug
resistant Salmonella enterica serovar Typhi CT18 Nature 413848ndash852
Paterson RRM Lima N 2010 How will climate change affect mycotoxins in food Food
Research International 431902ndash14
Paul PA El-Allaf SM Lipps PE Madden LV 2004 Rain splash dispersal of Gibberella zeae
within wheat canopies in Ohio Phytopathology 941342-1349
Pestka JJ Lin WS Miller ER 1987 Emetic activity of the trichothecene 15-
acetyldeoxynivalenol in pigs Food and Chemical Toxicology 25855ndash858
Pestka JJ Smolinski AT 2005 Deoxynivalenol Toxicology and potential effects on humans
Journal of Toxicology Environmental Health 839ndash69
Pestka JJ 2007 Deoxynivalenol toxicity mechanisms and animal health risks Animal Feed
Science and Technology 137283-298
Pestka JJ 2008 Mechanisms of deoxynivalenol-induced gene expression and apoptosis Food
Additives and Contaminants 251128-1140
Pestka J2010 Toxicological mechanisms and potential health effects of deoxynivalenol and
nivalenol World Mycotoxin Journal 3323ndash347
Petersohn A Brigulla M Haas S Hoheisel JD Voumllker U Hecker M 2001 Global Analysis
of the General Stress Response of Bacillus subtilis Journal of Bacteriology 1835617-
5631
200
Petty NK Bulgin R Crepin VF Cerdeno-Tarraga AM Schroeder GN Quail MA Lennard N
Corton C Barron A Clark L Toribio AL Parkhill J Dougan G Frankel G Thomson NR
2010 The Citrobacter rodentium genome sequence reveals convergent evolution with
human pathogenic Escherichia coli Journal of Bacteriology 192525ndash538
Petty NK Feltwell T Pickard D Clare S Toribio AL et al 2011 Citrobacter rodentium is
an unstable pathogen showing evidence of significant genomic flux PLoS Pathology
7e1002018 doi101371journalppat1002018
Pittet A 2001 Natural occurrence of mycotoxins in foods and feeds A decade in review In
Mycotoxins and phytotoxins in perspective at the turn of the millennium edited by De
Koe WJ Samson RA Van Egmond HP Gilbert J Sabino M W J de Koe Wageningen
Netherlands pp 153ndash172
Placinta CM DrsquoMello JPF Macdonald AMC 1999 A review of worldwide contamination of
cereal grains and animal feed with Fusarium mycotoxins Animal Feed Science and
Technology 7821ndash37
Plank B Schuh M Binder EM 2009 Investigations on the effect of two feed additives
Biomin BBSH 797 and Mycofix Plus (R) 3E as detoxificants of DON contaminated feed
of piglets Wien Tierarztl Monatssch 9655ndash71
Poapolathep A Ohtsuka R Kiatipattanasakul W Ishigami N Nakayama H Doi K 2002
Nivalenol-induced apoptosis in thymus spleen and Peyerrsquos patches of mice Experimental
and Toxicological Pathology 53441ndash446
201
Poppenberger B Berthiller F Lucyshyn D Sieberer T Schuhmacher R Krska R Kuchler K
Gloumlssl J Luschnig C Adam G 2003 Detoxification of the Fusarium mycotoxin
deoxynivalenol by a UDP-glucosyltransferase from Arabidopsis thaliana Journal of
Biological Chemistry 27847905ndash47914
Prelusky DB Trenholm HL 1993 The efficacy of various classes of anti-emetics in
preventing deoxynivalenol-induced vomiting in swine Natural Toxins 1296ndash302
Proctor RH Hohn TM McCormick SP 1995 Reduced virulence of Gibberella zeae caused
by disruption of a trichothecene toxin biosynthetic gene Molecular Plant-Microbe
Interactions 8593-601
Puchenkova SG 1996 Enterobacteria in areas of water along the Crimean Coast
Mikrobiolohichnyĭ zhurnal 58 3-7
Qi BW Hao BL 2004 Whole-proteome prokaryote phylogeny without sequence alignment
a K-string composition approach Journal of Molecular Evolution 581ndash11
Quarta A Mita G Haidukowski M Logrieco A Mule G Visconti A 2006 Multiplex PCR
assay for the identification of nivalenol 3- and 15-acetyl-deoxynivalenol chemotypes in
Fusarium FEMS Microbiological Letters 2597ndash13
Rehr B Klemme JH 1989 Formate dependent nitrate and nitrite reduction to ammonia by
Citrobacter freundii and competition with denitrifying bacteria Antonie van Leeuwenhoek
Journal of Microbiology 56311ndash321
Rocha O Ansari K Doohan FM 2005 Effects of trichothecene mycotoxins on eukaryotic
cells A review Food Additives and Contaminants 22369ndash378
Roscoe V Lombaert GA Huzel V et al 2008 Mycotoxins in breakfast cereals from the
Canadian retail market a 3-year survey Food Additives and Contaminants 25347ndash355
202
Rotter BA Prelusky DB Pestka JJ 1996 Toxicology of deoxynivalenol (vomitoxin) Journal
of Toxicology and Environment Health 481ndash34
Ryu JC Ohtsubo K Izurniyarna N Nakamura K Tanaka T Yamarnura H Ueno Y 1988
The acute and chronic toxicities of nivalenol in mice Fundamental and Applied
Toxicology 1138ndash47
Saier MH Roseman S 1972 Inducer exclusion and repression of enzyme synthesis in
mutants of Salmonella typhimurium defective in enzyme I of the phosphoenolpyruvate
sugar phosphotransferase system The Journal of Biological Chemistry 247972ndash975
Saier MHJ Tran CV Barabote RD 2006 TCDB the transporter classification database for
membrane transport protein analyses and information Nucleic Acids Research 34D181ndash
186
Saitou N Nei M 1987 The neighbor-joining method a new method for reconstructing
phylogenetic trees Molecular Biology and Evolution 4406ndash425
Savelieva O Kotova I Roelofsen W Stams AJM Netrusov A 2004 Utilization of
aminoaromatic acids by a methanogenic enrichment culture and by a novel Citrobacter
freundii strain Archives of Microbiology 181163ndash170
Schaafsma AW Tamburic-Ilinic L Miller JD Hooker DC 2001 Agronomic considerations
for reducing deoxynivalenol in wheat grain Canadian Journal of Plant Pathology 23279ndash
285
Schaafsma AW 2002 Economic changes imposed by mycotoxins in food grains case study
of deoxynivalenol in winter wheat Advances in Experimental Medicine and Biology
504271ndash276
203
Scharf C Riethdorf S Ernst H Engelmann S Voumllker U Hecker M 1998 Theoredoxin is an
essential protein induced by heat shock and other stresses in Bacillus subtilis Journal of
Bacteriology 1801869-1877
Schatzmayr G Zehner F Taubel M Schatzmayr D Klimitsch A Loibner AP Binder EM
2006 Microbiologicals for deactivating mycotoxins Molecular Nutrition and Food
Research 50543-551
Schollenberger M Muumlller H-M Ruumlfle R Suchy S Plank S Drochner W 2006 Natural
occurrence of 16 Fusarium toxins in grains and feedstuffs of plant origin from Germany
Mycopathologia 16143ndash52
Scientific Committee for Food 2002 Opinion on Fusarium toxins Part 6 Group evaluation
of T-2 toxin HT-2 toxin nivalenol and deoxynivalenol (available at http
wwweuropaeuintcommfoodfsscscfout123_enpdf)
Schulman S Wang B Li W Rapoport TA 2010 Vitamin K epoxide reductase prefers ER
membrane-anchored thioredoxin-like redox partners Proceedings of the National
Academy of Sciences 10715027ndash15032
Scott PM 1997 Multi-year monitoring of Canadian grains and grain-based foods for
trichothecenes and zearalenone Food Additives and Contaminants 14333-339
Seitz LM Bechtel DB 1985 Chemical physical and microscopal studies of scab-infected
hard red winter wheat Journal of Agricultural and Food Chemistry 33373-7
Sergent T Parys M Garsou S Pussemier L Schneider YJ Larondelle Y 2006
Deoxynivalenol transport across human intestinal Caco-2 cells and its effects on cellular
metabolism at realistic intestinal concentrations Toxicological Letters 164167ndash176
204
Sharma J Fulekar MH 2009 Potential of Citrobacter freundii for bioaccumulation of
heavy metalndashcopper Biology and Medicine 17-14
Sherif SO Salama EE Abdel-Wahhab MA 2009 Mycotoxins and child health the need for
health risk assessment International Journal of Hygiene and Environmental Health
212347ndash368
Shifrin VI Anderson P 1999 Trichothecene mycotoxins trigger a ribotoxic stress response
that activates c-Jun N-terminal kinase and p38 mitogen-activated protein kinase and
induces apoptosis The Journal of Biological Chemistry 27413985ndash13992
Shima J Takase S Takahashi Y Iwai Y Fujimoto H Yamazaki M Ochi K 1997 Novel
detoxification of the trichothecene mycotoxin deoxynivalenol by a soil bacterium isolated
by enrichment culture Applied and Environmental Microbiology 633825ndash3830
Singh J Behal A Singla N Joshi A Birbian N et al 2009 Metagenomics Concept
methodology ecological inference and recent advances Biotechnology Journal 4480ndash
494
Smith TK McMillan EG Castillo JB 1997 Effect of feeding blends of Fusarium mycotoxin-
contaminated grains containing deoxynivalenol and fusaric acid on growth and feed
consumption of immature swine Journal of Animal Science 752184ndash2191
Smits WK Dubois JYF Bron S van Dijl JM Kuipers OP 2005 Tricksy business
transcriptome analysis reveals the involvement of thioredoxin in redox homeostasis
oxidative stress sulfur metabolism and cellular differentiation in Bacillus subtilis Journal
of Bacteriology 1873921ndash3930
205
Sprando RL Collins TFX Black TN Olejnik N Rorie JI Eppley RM Ruggles DI 2005
Characterization of the effect of deoxynivalenol on selected male reproductive endpoints
Food Chemistry and Toxicology 43623ndash635
Starkey DE Ward TJ Aoki T Gale LR Kistler HC Geiser DM Suga H Toacuteth B Varga J
ODonnell K 2007 Global molecular surveillance reveals novel Fusarium head blight
species and trichothecene toxin diversity Fungal Genetics and Biology 441191ndash1204
Stevenson BS Schmidt TM 1997 Growth rate-dependent expression of RNA from plasmid-
borne rRNA operons in Escherichia coli Journal of Bacteriology 1801970ndash1972
Stothard P Wishart DS 2005 Circular genome visualization and exploration using
CGView Bioinformatics 21537ndash539
Suenaga H Ohnuki T Miyazaki K 2007 Functional screening of a metagenomic library for
genes involved in microbial degradation of aromatic compounds Environmental
Microbiology 92289-97
Surai P Mezes M Fotina TI Denev SD 2010 Mycotoxins in Human Diet A Hidden
Danger In Modern Dietary Fat Intakes in Disease Promotion Edited by F De Meester et
al Nutrition and Health DOI 101007978-1-60327-571-2_18
Sutton JC 1982 Epidemiology of wheat head blight and maize ear rot caused by Fusarium
graminearum Canadian Journal of Plant Pathology 4195
Swaving J De Bont JAM Westphal A De Kok A 1996 A novel type pyridine nucleotide-
disulfide oxidoreductase is essential for NAD+- and NADPH-dependent degradation of
epoxyalkanes by Xanthobacter strain Py2 Journal of Bacteriology 1786644ndash6
206
Swanson SP Rood JHD Behrens JC Sanders PE 1987 Preparation and characterization of
the deepoxy trichothecenes deepoxy HT-2 deepoxy T-2 triol deepoxy T-2 tetraol
deepoxy 15-monoacetoxyscirpenol and deepoxy scirpentriol Applied and Environmental
Microbiology 532821ndash2826
Swanson SP Helaszek C Buck WB Rood JHD Haschek WM 1988 The role of intestinal
microflora in the metabolism of trichothecene mycotoxins Food Chemistry and
Toxicology 26823ndash829
Tian F Yu Y Chen B Li H Yao Y-F Guo X-K 2009 Bacterial archaeal and eukaryotic
diversity in Arctic sediment as revealed by 16S rRNA and 18S rRNA gene clone libraries
analysis Polar Biology 3293-103
Tatusov RL Galperin MY Natale DA Koonin EV et al 2000 Artemis sequence
visualization and annotation Bioinformatics 16944-945
Trail F Haixin X Loranger R Gadoury D 2002 Physiological and environmental aspects of
ascospore discharge in Gibberella zeae (anamorph Fusarium graminearum) Mycologia
94181-189
Tsuge K Ohata Y Shoda M 2001 Gene yerP involved in surfactin self-resistance in
Bacillus subtilis Antimicrobial Agents and Chemotherapy 453566ndash3573
Tucker 2001 The ldquoyellow rainrdquo controversy Lesson for arms control compliance The
Nonproliferation Review 825-42
Ueno Y 1985 The toxicology of mycotoxins Critical Reviews in Toxicology 1499-132
Ueno Y Nakayama K Ishii K Tashiro F Minoda Y Omori T Komagata K 1983
Metabolism of T-2 toxin in Curtobacterium sp strain 114-2 Applied and Environmental
Microbiology 46120ndash127
207
van Elsas JD Speksnijder AJ van Overbeek LS 2008 A procedure for the metagenomics
exploration of disease suppressive soils Journal of Microbiological Methods 75515ndash522
van Ginkel CG de Bont JAM 1986 Isolation and characterization of alkene-utilizing
Xanthobacter spp Archives of Microbiology 145403ndash407
van Lanen SG Shen B 2006 Microbial genomics for the improvement of natural product
discovery Current Opinion in Microbiology 9252ndash260
Varga J Toacuteth B 2005 Novel strategies to control mycotoxins in feeds a review Acta
Veterinaria Hungarica 53189-203
Voigt CA Schafer W Salomon S 2005 A secreted lipase of Fusarium graminearum is a
virulence factor required for infection of cereals The Plant Journal 42364ndash375
Vogelgsang S Hecker A Musa T Dorn B Forrer HR 2011 On farm experiments over five
years in a grain maizewinter wheat rotation Effect of maize residue treatments on
Fusarium graminearum infection and deoxynivalenol contamination in wheat In Proc
7th
Canadian Workshop on Fusarium Head Blight edited by J Gilbert A Tekauz N
Sweetland and K Slusarenko Winnipeg Manitoba Canada p 41
Voumllkl A Vogler B Schollenberger M Karlovsky P 2004 Microbial detoxification of
mycotoxin deoxynivalenol Journal of Basic Microbiology 44147ndash156
Wagacha JM Muthomi JW 2007 Fusarium culmorum Infection process mechanisms of
mycotoxin production and their role in pathogenesis in wheat Crop Protection 26877-
885
Wakulinski W 1989 Phytotoxicity of the secondary metabolites of fungi causing wheat head
fusariosis (head blight) Acta Physiologiae Plantarum 11301ndash306
208
Wang JT Chang SC Chen YC Luh KT 2000 Comparison of antimicrobial susceptibility of
Citrobacter freundii isolates in two different time periods The Journal of Microbiology
Immunology and Infection 33258-62
Wannemacher RJ Stanley L Wiener MD 2000 Trichothecenes mycotoxins In Medical
aspects of chemical and biological warfare edited by Zajtchuk R Washington DC
Department of the Army pp 656-676
Westphal AH Swaving J Jacobs L de Kok A 1998 Purification and characterization of a
flavoprotein involved in the degradation of epoxyalkanes by Xanthobacter Py2 European
Journal of Biochemistry 257160ndash168
Whalen JG Mully TW Enlgish JC 2007 Spontaneous Citrobacter freundii infection in an
immunocompetent patient Archives of Dermatology 143124-5
White 2007 The physiology and biochemistry of prokaryotes Third edition Oxford
University Press New York Oxford p 628
Wilcke WF Morey RV 1995 Natural-air corn drying in the upper Midwest University of
Minnesota Minnesota Extension Service Publication BU-6577-E pp 20
Windels CE 2000 Economic and social impacts of Fusarium headblight changing farms and
rural communities in the Northern Great Plains Phytopathology 9017ndash21
Winzeler EA Shoemaker DD Astromoff A Liang H Anderson K Andre B Bangham R
Benito R Boeke JD Bussey H et al 1999 Functional characterization of the S
cerevisiae genome by gene deletion and parallel analysis Science 285901ndash906
Yang GH Jarvis BB Chung YJ Pestka JJ 2000 Apoptosis induction by the satratoxins and
other trichothecene mycotoxins relationship to ERK p38 MAPK and SAPKJNK
activation Toxicology and Applied Pharmacology 164149ndash160
209
Yang Q Yin GM Guo YL Zhang DF Chen SJ Xu ML 2010 A major QTL for resistance
to Gibberella stalk rot in maize Theoretical and Applied Genetics 121673ndash687
Yazar S Omurtag GZ 2008 Fumonisins trichothecenes and zearalenone in cereals
International Journal of Molecular Science 92062ndash2090
Yoshizawa T Morooka N 1973 Deoxynivalenol and its monoacetate new mycotoxins from
Fusaruium roseum and moldy barley Agricultural and Biological Chemistry 372933ndash
2934
Yoshizawa T Takeda H Ohi T 1983 Structure of a novel metabolite from deoxynivalenol a
trichothecene mycotoxin in animals Agricultural and Biological Chemistry 472133ndash
2135
Yoshizawa T 1993 Red-mold diseases and natural occurrence in Japan In Trichothecenes-
Chemistry Biology and Toxicological Aspects edited by Y Ueno Elsevier Amsterdam
Netherlands p 195-209
Yoshizawa T Sakamoto T Okamoto K 1984 In vitro formation of 3rsquo-hydroxy T-2 and 3rsquo-
hydroxy HT-2 toxins from T-2 by liver homogenates from mice and monkeys Applied
and Environmental Microbiology 47130-134
Young JC Subryan LM Potts D McLaren ME Gobran FH 1986 Reduction in levels of
deoxynivalenol in contaminated wheat by chemical and physical treatment Journal of
Agricultural and Food Chemistry 34461ndash465
Young JC Trenholm HL Friend DW Prelusky DB 1987 Detoxification of deoxynivalenol
with sodium bisulfite and evaluation of the effects when pure mycotoxin or contaminated
corn was treated and given to pigs Journal of Agricultural and Food Chemistry 35259ndash
261
210
Young JC Zhu H Zhou T 2006 Degradation of trichothecene mycotoxins by aqueous
ozone Food Chemistry and Toxicology 44417ndash424
Young JC Zhou T Yu H Zhu H Gong J 2007 Degradation of trichothecene mycotoxins by
chicken intestinal microbes Food Chemistry and Toxicology 45136-143
Yu Z Morrison M 2004 Comparisons of different hypervariable regions of rrs genes for use
in fingerprinting of microbial communities by PCR-denaturing gradient gel
electrophoresis Applied and Environmental Microbiology 704800-4806
Yu H Zhou T Gong J Young C Su X Li X Zhu H Tsao R Yang R 2010 Isolation of
deoxynivalenol-transforming bacteria from the chicken intestines using the approach of
PCR-DGGE guided microbial selection BMC Microbiology 10182
Zeibdawi A Steele M Savoie A 2011 An overview of CFIAs mycotoxin monitoring
program in livestock feeds In Proc 7th
Canadian Workshop on Fusarium Head Blight
edited by J Gilbert A Tekauz N Sweetland and K Slusarenko Winnipeg Manitoba
Canada p 25
Zhang J-B Li H-P Dang F-J Qu B Xu Y-B Zhao C-S Lia Y-C 2007 Determination of the
trichothecene mycotoxins chemotypes and associated geographical distribution and
phylogenetic species of the Fusarium graminearum clade from China Mycological
Research 111967ndash975
Zhao JS Manno D Thiboutot S Ampleman G Hawari J 2007 Shewanella canadensis sp
nov and Shewanella atlantica sp nov manganese dioxide- and hexahydro-135-trinitro-
135-triazine-reducing psychrophilic marine bacteria International Journal of Systemic
Evolution and Microbiology 572155ndash2162
211
Zhao X-Q 2011 Genome-based studies of marine microorganisms to maximize the diversity
of natural products discovery for medical treatments Evidence-Based Complementary
and Alternative Medicine Article ID 384572 doi1011552011384572
ZhaoY Park RD Muzzarelli RA 2010 Chitin deacetylases properties and applications
Marine Drugs 824ndash46
Zhou HR Lau AS Pestka JJ 2003a Role of double-stranded RNA-activated protein kinase R
(PKR) in deoxynivalenol-induced ribotoxic stress response Toxicological Science
74335ndash344
Zhou HR Islam Z Pestka JJ 2003b Rapid sequential activation of mitogen-activated protein
kinases and transcription factors precedes proinflammatory cytokine mRNA expression in
spleen of mice exposed to the trichothecene vomitoxin Toxicological Sciences 72130ndash
142
Zhou T He J Gong J 2008 Microbial transformation of trichothecene mycotoxins World
Mycotoxin Journal 123-30
Zhou G Zhang H He Y He L 2010 Identification of a chitin deacetylase producing bacteria
isolated from soil and its fermentation optimization African Journal of Microbiology
Research 42597-2603

A fosmid genomic DNA library of strain ADS47 was prepared in E coli and screened
for DON detoxification activity However no library clone was found with DON
detoxification activity The whole genome of ADS47 was also sequenced and from
comparative genome analyses 10 genes (characterized as reductases oxidoreductases and
deacetylase) were identified as potential candidates for DONtrichothecene detoxifying
enzymes Bioinformatic analyses showed that the major animal pathogenic locus of
enterocyte effacement (LEE) operon genes absent from the ADS47 genome Strain ADS47
has the potential to be used for feed detoxification and the development of mycotoxin
resistance cereal cultivars
iii
DEDICATIONS
The PhD thesis is dedicated to the departed souls of my mother MOZIBUN NESSA and
father-in-law DR MA AHAD MIAH
iv
ACKNOWLEDGEMENTS
Foremost I am deeply indebted to my advisors Prof K Peter Pauls and Dr Ting Zhou
for the continuous support of my PhD study for their endurance inspiration enthusiasm and
mentoring Their continued guidance has helped me in all aspects of my research and in
writing of this dissertation I sincerely express my gratitude to them for their co-operation
from the beginning to conclusion of the thesis and many other areas This will guide me in my
future professional career
My thesis committee guided me through all these years I would like to express my
gratitude to Prof Paul Goodwin for his insightful comments and advice on my thesis research
Special thanks to Dr Chris Young for teaching me chemistry chemical structures and
interpreting liquid chromatography ultraviolet mass spectra data I am sincerely grateful to
Dion Lepp for helping in analyzing the bacterial genome and sharing his knowledge and
valuable suggestions on writing the genomic chapter
I would like to express my appreciation to staff members of Dr Ting Zhou and Prof
K Peter Pauls who supported me in many aspects during my PhD study I am thankful to
Jianwei He Xiu-Zhen Li Honghui Zhu for collections and preparation of soil samples and
LC-UV-MS analyses of numerous samples Special thanks to Dr David Morris Johnston
Monje for sharing his knowledge discussions and camaraderie at coffee breaks
I wish to acknowledge the Natural Sciences and Engineering Research Council of
Canada (NSERC) for awarding me a prestigious scholarship for my PhD program and
Deanrsquos Tri-Council Scholarship (University of Guelph) which made my life easier during this
study I also would like to convey my thanks to Agriculture and Agri-Food CanadaBinational
v
Agricultural Research and Development Fund (AAFCBARD) project for providing me
financial support
Mike Bryan a very supportive person helped provide solutions to my software issues
that improved my exposure and caliber at conference awards competitions Several student
competition awards honored and inspired me to work hard and lead to the successful
completion of my thesis research I would like to convey my appreciation to the judges of
different student award committees who recognized my work They are Society for Industrial
Microbiology and Biotechnology conference 2010 (USA) Food Safety Research Forum-
Ontario Ministry of Agriculture and Rural Affairs annual meeting 2010 (Canada)
International Mycological Conference 2009 (UK) and Southwestern Ontario Regional
Association of Canadian Phytopathological Society annual meeting 2009-2011 (Canada)
I would like to express my very special thanks to Prof Golam Hafeez Kennedy my
brother-in-law who is like my elder brother In many aspects I am indebt to him for his
cooperations which aided me to achieve my current status
Lastly and most importantly I wish to give my special thanks to my wife Mimi and
lovely daughter Racy whose endless love sacrifice support and encouragement enabled me
to cross the tough long road and made possible to achieve my goal and aspirations Racy
once you told me that one of your desires for me is to finish my PhD which is done now I
know you want to be a scientist which is a great ambition I wish you all the best in your
future aims Racy
vi
TABLE OF CONTENTS
DEDICATIONShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipiii
ACKNOWLEDGEMENTShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipiv
TABLE OF CONTENTShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipvi
LIST OF TABLEShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipix
LIST OF FIGUREShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipx
LIST OF ABBREVIATIONShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipxiii
10 GENERAL INTRODUCTION AND LITERATURE REVIEW helliphelliphelliphelliphelliphelliphelliphellip1
11 Backgroundhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip1
12 Plant pathogenesis and trichothecene productionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip3
13 Biosynthesis of food contaminating TChelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip6
14 Occurrence of TC mycotoxins in food and feed helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip11
15 Significance of deoxynivalenol and trichotheceneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip16
16 Toxic effects of deoxynivalenol and trichotheceneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip19
17 Cellular and molecular mechanisms of DON toxicity in animals and plantshelliphelliphelliphellip22
18 Strategies to control Fusarium head blight and trichotheceneshelliphelliphelliphelliphelliphelliphelliphelliphellip26
19 Isolation of DONTC de-epoxidating micro-organisms and geneshelliphelliphelliphelliphelliphelliphelliphellip28
110 The bacterial species Citrobacter freundiihelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip30
111 Hypotheses and objectiveshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip31
vii
20 AEROBIC AND ANAEROBIC DE-EPOXIDATION OF MYCOTOXIN
DEOXYNIVALENOL BY BACTERIA ORIGINATING FROM AGRICULTURAL
SOIL33
21 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip33
22 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
23 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip37
24 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip45
25 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip62
26 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip64
30 ISOALTION AND CHARACTERIZATION OF A NOVEL SOIL BACTERIUM
CAPABLE OF DETOXIFYING ELEVEN TRICHOTHECENE MYCOTOXINS65
31 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip65
32 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip66
33 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip68
34 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip75
35 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip96
36 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip100
40 CONSTRUCTION AND FUNCTIONAL SCREENING OF GENOMIC DNA
LIBRARY FOR ISOLATION OF DON DE-EPOXYDIZING GENE(S) FROM
BACTERIAL STRAIN ADS47helliphelliphelliphelliphelliphelliphelliphellip102
41 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip102
42 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip103
43 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip104
viii
44 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip112
45 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip118
46 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip121
50 GENOME SEQUENCING CHARACTERIZATION AND COMPARATIVE
ANALYSIS OF A TRICHOTHECENE MYCOTOXIN DETOXIFYING
CITROBACTER FREUNDII STRAIN ADS47helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip122
51 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip122
52 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip123
53 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip125
54 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip129
55 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip162
56 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip168
60 CONCLUSIONhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip171
61 Summary helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip171
62 Research contributions helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip174
63 Future workhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip175
70 REFERENCEShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip176
ix
LIST OF TABLES
Table 1-1 Fusaria produced type-A and type-B trichothecene mycotoxinshelliphelliphelliphelliphelliphelliphellip9
Table 1-2 Worldwide occurrence of deoxynivalenol (DON) in animal feedshelliphelliphelliphelliphellip12
Table 1-3 Occurrence of DON in Canadian grains grains foods and beershelliphelliphelliphelliphelliphelliphellip13
Table 1-4 Legislative maximum tolerated levels of trichothecene mycotoxins in Canadahellip14
Table 1-5 In vitro and in vivo synergistic effects of different food contaminating
mycotoxins helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip18
Table 3-1 Microbial transformation of type-A and type-B tricthothecene mycotoxins
after 72 h of incubation under aerobic conditions and at room temperaturehelliphelliphelliphelliphelliphellip95
Table 4-1 Analysis of Fosmid DNA library for deoxynivalenol (DON) de-epoxidation
capability helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip117
Table 5-1 General features of C freundii strain ADS47 genomehelliphelliphelliphelliphelliphelliphelliphelliphellip132
Table 5-2 Genome comparisons between strain ADS47 and five Citrobacter species141
Table 5-3 Functional categorized gene abundances among the six Citrobacter genomeshellip145
Table 5-4 Unique genes of C freundii strain ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip156
Supplemental Table S5-1 List of unique genes of strain ADS47 gene IDs and their
functional roleshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip169
x
LIST OF FIGURES
Figure 1-1 Structures of trichothecene mycotoxins produced by Fusariahelliphelliphelliphelliphelliphelliphelliphellip7
Figure 1-2 Cellular and molecular mechanisms of deoxynivalenol toxicity in
eukaryotic organismshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip23
Figure 2-1 Mechanism of microbial DON de-epoxidationhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip36
Figure 2-2 LC-UV-MS chromatographic analysis of DON and its de-epoxidated product
dE-DON obtained by culturing with selected soil microbeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip46
Figure 2-3 Time course of relative biotransformation of DON to dE-DON by the initial
soil suspension culture in MSB mediumhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip49
Figure 2-4 Microbial de-epoxidation of DON after five days of incubationhelliphelliphelliphelliphelliphellip51
Figure 2-5 Analysis of microbial population reduction by terminal restriction fragment
length polymorphism (T-RFLP)helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip53
Figure 2-6 Microbial genera identified in the enriched DON de-epoxidizing microbial
culturehelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip56
Figure 2-7 Effects of culture conditions on DON to dE-DON transformation by the
enriched microbial culturehelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip58
Figure 2-8 Microbial DON to dE (de-epoxy)-DON transformation activity under aerobic
and anaerobic conditionshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip61
Figure 3-1 Colony color shape and size of the mycotoxin degrading bacterial strain
ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip76
Figure 3-2 Dendrogram based on 16S-rDNA sequences for strain ADS47 and 11
reference Citrobacter specieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip77
xi
Figure 3-3 Effects of DON concentration on bacterial growth and de-epoxidation
activitieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip79
Figure 3-4 Bacterial growth and DON de-epoxidation capabilities under aerobic and
anaerobic conditions helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip82
Figure 3-5 Effects of media on bacterial growth and DON de-epoxidation activityhellip85
Figure3-6 Effects of temperature on microbial growth and DON de-epoxydation helliphelliphellip87
Figure 3-7 Influence of pH on microbial growth and DON de-epoxydation capabilityhellip89
Figure 3-8 Effects of sodium azide (SA) on bacterial DON de-epoxidation activityhelliphelliphellip91
Figure 3-9 Determination of DON to deepoxy-DON transformation by bacterial culture
filtrate and cell extracthelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip92
Figure 3-10 Determination of microbial de-epoxidation and de-acetylation of a type-A
trichothecene mycotoxin (HT-2 toxin) by LC-UV-MShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip94
Supplemental Figure S3-1 Flow chart depicted the procedures of enrichment and isolation
of DON de-epoxidation bacterial strain ADS47helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip101
Figure 4-1 Schematic overview of the construction and functional screening of Fosmid
DNA libraryhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip105
Figure 4-2 Genetic map of copy controlled Fosmid vector (pCC1FOS)helliphelliphelliphelliphelliphelliphellip108
Figure 4-3 Preparation of 40 kb size insert DNA from a DON de-epoxidizing bacterium113
Figure 4-4 Production of genomic DNA library in E coli EPI300-T1 host strainhelliphelliphelliphellip114
Figure 4-5 Confirmation of the Fosmid DNA library construction in E coli helliphelliphelliphelliphellip 116
Figure 5-1 Co-linearity between C freundii strain ADS47 genome and C koseri helliphelliphellip129
Figure 5-2 Circular genome map of C freundii strain ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphellip133
xii
Figure 5-3 Size distribution of the predicted genes identified in C freundii strain
ADS47helliphelliphellip 135
Figure 5-4 Functional classification of the predicted coding sequences (CDS) of
C freundii strain ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip137
Figure 5-5 Phylogeny of the trichothecene detoxifying C freundii strain ADS47 139
Figure 5-6 Comparisons of coding sequences between C freundii strain ADS47 and
five Citrobacter specieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip143
Figure 5-7 Determination of the locus enterocyte effacement (LEE) operons in
Citrobacter species by comparing C rodentium helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip151
Figure 5-8 Locations of unique genes of C freundii strain ADS467helliphelliphelliphelliphelliphelliphelliphellip157
Figure 5-9 Unique genomic regions and associated genes in strain ADS47 compared
to five Citrobacter specieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip159
xiii
LIST OF ABBREVIATIONS
APCI= atmospheric pressure chemical ionization
AE= attaching and effacing
A= aerobic conditions
ABC= ATP-binding cassette
ACT= artemis comparative tools
AN= anaerobic conditions
3-ADON= 3-acetyl-deoxynivalenol
15-ADON= 15-acetyl-deoxynivalenol
ATA= alimentary toxic aleukia
bp= base pair
BGI= Beijing genomics institute
BLAST= basic local alignment search tool
BRIG= blast ring image generator
Bw= body weight
C= carbon
CDS= coding sequence
CFIA= Canadian food inspection agency
CHEF= clamped homogeneous electric fields
CMB= corn meal broth
CVTree= composition vector tree
DAS= diacetoxyscirpenol
dH2O= distilled water
DOGT1= UDP-glucosyltransferase
DON= deoxynivalenol
dsRNA= double stranded RNA
dE= de-epoxy
dE-DON= de-epoxy DON
E coli= Escherichia coli
ER=endoplasmic reticulum
FAO= Food amp Agriculture Organization
FM= full medium
FHB= Fusarium head blight
FUS= fusarenonX
gDNA= genomic DNA
GC= guanine cytosine
GER= Gebberella ear rot
GPI= glycosylphosphatidylinositol
h= hour
Hck= hematopoietic cell kinase
Kb= kilobase
xiv
Kg= kilogram
LC-UV-MS= liquid chromatography-ultraviolet-
mass spectrometry
LEE= locus of enterocyte effacement
LD50= lethal dose of 50 animal
LB= Luria Bertani
LMP= low melting point
MAPK= mitogen-activated protein kinase
microg= microgram
mg= milligram
mL= milliliter
microL= microliter
microM= micromole
min= minute
Mb= megabases
MCS=multiple cloning site
MSA= MSB+ agar
MSB= mineral salts broth
MM= minimal medium
MSC= mineral salts + 03 Bacto Peptone
MS-DON= mineral salts + 200 microgmL DON
mRNA= messenger RNA
MSY= mineral salts + yeast extract
mz= mass ion ratio
N= nitrogen
Na-Cl= sodiumndashclorine
NADH= (reduced) nicotinamide adenine
dinucleotide
NAD(P)= nicotinamide adenine
dinucleotide phosphate
NCBI= national center for biotechnology
information
NEO= neosolaniol
ng= nanogram
NIV= nivalenol
OD600nm= optical density at 600
nanometer wavelength
OMAFRA= Ontario Ministry of
Agriculture and Rural Affairs
Paa= phenylacetic acid
PCR= polymerase chain reaction
PDB= potato dextrose broth
ppm= parts per million
PFGE= pulsed field gel electrophoresis
PKR= protein kinase
xv
PNDO= pyridine nucleotide-disulfide
oxidoreductase
QTL= quantitative trait locus
RAST= rapid annotation subsystem technology
RDP= ribosomal database project
Rpl3= 60S ribosomal protein L3
rRNA= ribosomal RNA
s= second
SA= sodium azide
SDR= short chain dehydrogenasereductase
SE= soil extract
SIM= selected ion monitoring
SOAP= short oligonucleotide analysis package
sp= species
sRNA= small RNA
TC= trichothecene
TCA= tricarboxylic acid cycle
TSB= tryptic soy broth
TF= transcription factor
tRNA= transfer RNA
TIC= total ion count
TRAP= tripartite ATP-independent
periplasmic
T-RFLP=terminal restriction fragment
length polymorphism
TRI101= trichothecene 3-O-
acetyltransferase
YEB= yeast extract broth
VER= verrucarol
VKOR= vitamin K epoxide reductase
wv= weight by volume
1
CHAPTER 1
10 GENERAL INTRODUCTION AND LITERATURE REVIEW
11 Background
According to the Food and Agriculture Organization (FAO) of the United Nations
more than 25 of the worldrsquos food crops are contaminated with mycotoxins every year
causing serious adverse effects on livestock industry crop production and international trade
(Boutrif and Canet 1998 Desjardins 2006 Binder et al 2007 Surai et al 2010) The negative
effects of mycotoxins on the consumers are diverse including organ toxicity mutagenicity
carcinogenicity neurotoxicity teratogenicity modulation of the immune system reproductive
system effects and growth retardation Mycotoxins are toxic secondary metabolites produced
by various fungal species during crop colonization in field andor post harvest conditions
(FAO 1991 Barrett 2000) Aspergillus Penicillum and Fusarium are the predominant fungal
genera that accumulate economically important mycotoxins such as aflatoxins zearalenone
trichothecenes fumonisins ochratoxins and ergot alkaloids in agricultural commodities
Humans and animals are principally exposed to mycotoxins through consumption of
contaminated foodfeed and animal products such as milk meat and eggs (Fink-Gremmels
1989 Hollinger and Ekperigin 1999)
Trichothecenes (TCs) produced by a variety of toxigenic Fusarium species are
probably the most food and feed contaminating mycotoxins in the temperate regions of
America Europe and Asia (Creppy 2002 Yazar and Omurtag 2008) The fungal pathogen
accumulates various type-A (eg T-2 toxin HT-2 toxin) and type-B trichothecenes (eg
2
deoxynivalenol nivalenol and their derivatives) in cereals Among them type-B
trichothecenes particularly deoxynivalenol (DON) andor its derivatives appear to be the
most commonly found mycotoxins in grain and grain products worldwide impairing human
health and productivity of farm animals (Placinta et al 1999 Sherif et al 2009) A number of
human and animal disease outbreaks associated with TC contaminated grain food and feed
consumption have been reported in Asia (Japan India and central Asia) Europe New
Zealand and United States of America (USA) (Pestka and Smolinski 2005 Desjardins 2006)
A severe outbreak of Alimentary Toxic Aleukia (ATA) mycotoxicosis killed 100000 people
in the Orenburg region of Russia (Eriksen 2003) In 1996 losses of over $100 million were
estimated in Ontario because of costs related to managing DON (Schaafsma 2002)
Historically T-2 toxin DON and nivalenol (NIV) have likely been used as a biological
warfare in Laos Kampuchea and Afghanistan and there is concern because of the potential
use of these compounds for bioterrorism (Wannemacher et al 2000 Tucker 2001)
Despite the extensive efforts that have been made to prevent TC contamination with
the current varieties and production practices its presence in agricultural commodities is
likely unavoidable There are no crop cultivars currently available that are completely
resistant to Fusarium head blightGibberella ear rot (FHBGER) diseases Chemical or
physical methods cannot safely remove TC mycotoxins from grain completely or they are not
economically viable (Varga and Toacuteth 2005) Therefore an alternative strategy to address the
issue is of great importance (Awad et al 2010 Karlovsky 2011)
3
12 Plant pathogenesis and trichothecene production
Cereals infested by the ascomyceteous fungi may cause Gibberella ear rot (GER) in
maize (Zea mays L) and Fusarium head blight (FHB also known as lsquoscabrsquo) of small grain
cereals [wheat (Triticum aestivum L) barley (Hordeum vulgare L)] and other small grain
cereals (Goswami and Kistler 2004) As many as 17 fungal species have been reported to be
associated with FHBGER which include F graminearum Schwabe [telemorph Gibberella
zeae (Schwein) Petch] F culmorum (WG Smith) Sacc F sporotrichioides Scherb F
poae (Peck) Wollenw Microdochium nivale (Fr) Samuels amp IC Hallett and F avenaceum
(Fr) Sacc (Osborne and Stein 2007 Audenaert et al 2009) The two species F
graminearum and F culmorum are the principal species causing crop damage Among them
F graminearum is the widely adapted and most damaging species across the world including
North America (Dyer et al 2006) F culmorum tend to be the main species in the cooler parts
of North Central and Western Europe (Wagacha and Muthomi 2007)
The fungi overwinter mainly on crop residues as saprophytes where they produce
asexual (micro- and macro-conidia) andor sexual (ascospores) fruiting structures which act
as the initial source of inoculums in the subsequent crop season (Sutton 1982 Wagacha and
Muthomi 2007) Ascospores are primarily dispersed by air (Trail et al 2002) and conidia
move by rain-splash (Paul et al 2004) After they land on maize silks and florets of small
grain cereals infection occurs through natural openings (eg stomata) or occasionally by
direct penetration The pathogens can invade maize kernels through wounds caused in the
husks by insects or birds Infection may occur throughout the crop growing season but
disease incidence and severity is facilitated by warm temperatures (gt20degC) high humidity
(gt80) and frequent precipitation that coincides with anthesis (Osborne and Stein 2007
4
Wagacha and Muthomi 2007) The Fusarium pathogen initially infects as biotroph which
becomes a necrotroph as the disease progresses After penetration Fusaria produce different
extracellular cell wall degrading enzymes that trigger infection and hyphal progression
towards the developing earkernel where further colonization occurs and development of
FHBGER diseases (Voigt et al 2005 Miller et al 2007 Kikot et al 2009) FHB disease is
characterized by a bleached appearance around the middle of the head Severely infected
grains have a pinkish discoloration and can be shriveled In maize the disease produces a
reddishndashbrown discoloration of the kernels and progresses from the tip to the bottom of
affected ears
The fungi associated with FHB and GER contaminate cereals by accumulating various
host-nonspecific TC mycotoxins (Brennen et al 2007) TCs have typically been associated
with cereals grown in temperate climatic conditions including Canada (Doohan et al 2003)
DON production by F graminearum and F culmorum is maximal at warm temperatures (22-
28˚C) high grain moisture contents (gt17) and humid conditions (gt70 RH) (Martins and
Martins 2002 Hope et al 2005 Llorens et al 2006) Grain can be contaminated with TCs in
the field as well as in storage if conditions for fungal growth are optimal and grain contains
high moisture (gt17) (Larsen et al 2004 Birzele et al 2002)
Populations of FHBGER causing pathogens are dynamic Genetic variability between
and within Fusarium species appears to have occurred to allow for their adaptation to new
agro-climates (Desjardins 2006 Fernando et al 2011) Three strain specific Fusarium
chemotypes NIV 3-ADON (3-acetyl-deoxynivalenol) and 15-ADON (15- acetyl-
deoxynivalenol) have been identified which correlate with the geographical distributions
(Starkey et al 2007 Zhang et al 2007 Audenaert et al 2009) The NIV chemotype mainly
5
produced by F culmorum accumulates NIV and 4-NIV (Fusarinon X) is predominant in
Asia Africa and Europe (Lee et al 2001) The 3-ADON chemotype which synthesizes DON
and 3-ADON is the dominant strain in EuropeFar East (Larsen et al 2004) In North
America 15-ADON is the main chemotype which produces DON and 15-ADON (Goswami
and Kistler 2004) Significant Fusarium population shifting was observed in the past and in
recent years (Kant et al 2011) A dynamic change from F culmorum to F gramninearum
and DON to NIV producing chemotyes occurred in North West European countries The 3-
ADON chemotype which is uncommon in North America was found in North Dakota and
Minnesota (Goswami and Kistler 2004) F pseudograminearum which historically occurs in
Australia Africa and North West Pacific America has been found in Western Canada (Larsen
et al 2004 Kant et al 2011) In Manitoba a replacement of 15-ADON chemotype by the
more phytoxic 3-ADON type was observed in recent years (Fernando et al 2011) All
together the FHBGER associated pathogen complex is highly dynamic which not only
reduces crop yield but also decreases grain quality by accumulating various mycotoxins
6
13 Biosynthesis of food contaminating TC
TCs are fungal secondary metabolites that were initially identified in Trichotheceium
roseum Thus the name of this class of compounds follows the name of the genus
Trichotheceium (Freeman and Morrison 1949) There have been 180 TCs identified in various
fungal species belonging to the genera Fusarium Myrothecium Stachybotrys Trichoderma
and Thrichothecium (Eriksen 2003 Eriksen et al 2004) The FHBGER associated TCs are
mainly synthesized by different Fusaria and are characterized as tricyclic sesquiterpenoids
with a double bond at position C9-10 and a C12-13 epoxide ring
7
Figure 1-1 Structures of trichothecene mycotoxins produced by Fusaria (a) General
structure of trichothecenes R1 = H OH OiValeryl keto R2 = H OH R3= OH OAc R4 =
H OH OAc R5= OH OAc (b) Type-A trichothecene mycotoxin (c) Type-B trichothecene
mycotoxin which contain a carbonyl functional group at C8 position (Young et al 2007)
(a) (b)
(c)
8
The fungal metabolites are small in size (molecular weight 250-550) heat stable and
non-volatile molecules (Pestka 2007 Yazar and Omurtag 2008) Food contaminating TCs are
divided into two groups type-A and type-B (Figure 1-1b and 1c) Type-A is characterized by
the lack of a ketone function at C8 position and include T-2 toxin HT-2 toxin
diacetoxyscirpenol (DAS) and neosolaniol (NEO) which are mainly produced by F
sporotrichioides and F poae (Creppy 2002 Quarta et al 2006) The most frequently found
TC belongs to type-B which contains carbonyl functions at C8 positions DON NIV and
their acetylated derivatives The two types are further differentiated by the presence or
absence of functional hydroxyl (-OH) and acetyl (CH3-CO-) groups at C-3 C-4 C-7 C-8 and
C-15 positions (Table 1-1)
The syntheses of various TCs are speciesstrain specific and are genetically controlled
but influenced by environmental conditions (Desjardins 2006 Paterson and Lima 2010) Until
now 15 Fusaria catalytic genes and regulators involved in TC productions have been
identified (Merhej et al 2011) These biosynthetic genes are located at three loci on different
chromosomes The core trichothecene (TRI) gene cluster is comprised of 12 genes localized
in a 25-kb genomic region (Kimura et al 2001) The remaining three genes are encoded by
the TRI1-TRI16 and TRI101 loci (Gale et al 2005 Alexander et al 2009) Fusaria produce
TC following a number of oxygenation isomerization acetylation hydroxylation and
esterification reaction steps (Desjardins 2006 Desjardins and Proctor 2007 Kimura et al
2007 Merhej et al 2011) In brief farnesyl pyrophosphate is initially cyclized into
trichodiene by TRI5 which codes for trichodiene synthase In the next four steps
isotrichotriol is produced by TRI4 (a multifunctional cytochrome P450) via C-2 hydroxylation
and 1213 epoxidation followed by two hydroxylation reactions
9
Table 1-1 Fusaria produced type-A and type-B trichothecene mycotoxins Molecular weight
(MW) and acetyl (-R) hydroxyl (-OH) and ketone (=O) functions at different carbon (C)
position of the tricyclic rings A-types T-2 toxin HT-2 toxin DAS (diacetoxyscirpenol)
NEO (neosolaniol) VER (verrucarol) B-type DON (deoxynivalenol) 3-ADON (3-acetyl-
deoxynivalenol) 15-ADON (15-acetyl-deoxynivalenol) NIV (nivalenol) and FusX
(fusarenon X) (Young et al 2007)
Trichothecenes MW R1C3 R2C4 R4C7 R3C15 R5C8
Type-A
T-2 toxin 466 OH OCOCH3 H OCOCH3 OCOCH2CH(CH3)2
HT-2 toxin 424 OH OH H OCOCH3 OCOCH2CH(CH3)2
DAS 366 OH OCOCH3 H OCOCH3 H
NEO 382 OH OCOCH3 OH OCOCH3 H
VER 266 H OH H OH H
Type-B
DON 296 OH H OH OH =O
3-ADON 338 OCOCH3 H OH OH =O
15-ADON 338 OH H OH OCOCH3 =O
NIV 312 OH OH OH OH =O
FusX 354 OH OCOCH3 OH OH =O
10
The next two non-enzymatic reactions results in isotrichodemol Eventually the
trichothecene skeleton is produced via the formation of a C-O bond between the C-2 oxygen
and C-11 Subsequent acetylation at C-3 by TRI101 hydroxylation at C-15 by TRI1 and
catalytic activity of TRI3 leads to calonectrin production The pathway described above is
common for both type-A (eg T-2 toxin HT-2 toxin) and type-B (eg DON NIV and their
acetylated derivatives) TCs
The following steps are speciesstrain specific In DON producing F graminearum
strains calonectrin is converted to either 3-ADON or 15-ADON followed by DON production
by the catabolic activities of TRI1 and TRI8 The biosynthesis of 3-ADON or 15-ADON is
controlled by variation in the esterase coding sequences of TRI8 F graminearum produces
NIV from calonectrin via synthesis of 4-NIV followed by NIV by TRI1 and TRI8 gene
actions NIV can be produced either from DON by hydroxylation at C-4 or via
transformation of calonectrin to 315-diacetoxyscirpenol (315-DAS) followed by addition of
a ketone at C-8 position by the activity of the TRI7 and TRI13 genes 315-DAS is the
principal substrate for synthesis of type-A TCs by F sporotrichioides the main type-A
producer The biosynthetic pathway of T-2 toxin involves C-4 acetylation of 315-DAS by the
TRI7 gene product and TRI8 triggered by TRI1 315-DAS is also the substrate for HT-2
toxin synthesis In the absence of TRI7 expression HT-2 toxin is produced through
hydroxylation of 315-DAS and isovalerate addition to C-8 position (Foroud and Eudes 2009)
11
14 Occurrence of TC mycotoxins in food and feed
TC producing Fusaria are adapted to variable climatic conditions and agricultural
ecosystems As a consequence this class of mycotoxins particularly DON is frequently
detected in foods and feeds including breakfast cereals bakery products snack foods beer
and animal feed derived from cereals in most part of the world (Pittet 2001 Desjardins 2006
Lancova et al 2008) A two-year survey during 2003-2005 conducted by Romer Lab Inc
revealed unacceptable levels of DON in animal feedsfeed raw materials (such as maize
wheat wheat bran) in Asia and Oceania (1062 microgg) central Europe (802 microgg) Mid-West
USA (55 microgg) northern Europe (551 microgg) and southern Europe (303 microgg) (Binder et al
2007) (Table 1-2)
Quite high levels of DON contamination (19 microgg) were detected in north Asian and
Chinese feedfeed materials Recently five-year on-farm studies in Switzerland showed that
74 of the wheat grain and straw samples were contaminated with high levels of DON
(average 5 microgg) The amount is higher than the European maximum limit of 125 microgg
(Vogelgsang et al 2011)
The occurrences of TCs in Canadian agricultural commodities are indicated in Table
1-3 Seventeen surveys conducted during the 1980s and 1990s showed up to 100 DON
incidence in Ontario wheat and maize samples with an 18 average incidence (Scott 1997) A
three-year survey recently (2006-08) conducted by the Ontario Ministry of Agriculture and
Rural Affairs (OMAFRA) detected high levels of DON in spring (maximum 26 microgg) and
winter wheat (maximum 29 microgg) samples (Ana Matu personal communication) The DON
levels were much higher than Health Canadarsquos maximum limit of 2 microgg for uncleaned soft
wheat for staple foods (Table 1-4)
12
Table 1-2 Worldwide occurrence of DON in animal feed Feed and feed raw material
samples were collected from animal farmsfeed production sites during 2003-2005 The
survey was conducted by Romer Labs Inc (httpwwwromerlabscom)
Country
region
Sample
type
Incidence
()
Maximum
DON content
(microgg)
Mean DON
content
(microgg)
Survey
period
Source
North
AsiaChina
Animal
feed
071 1899 092
Binder et
al 2007
South East
Asia
014 152 016
South Asia 070 1062 049
Oceania 2003-05
Australia 041 186 030
Northern
Europe
070 551 059
Central
Europe
066 802 057
Southern
Europe
052 303 030
13
Table 1-3 Occurrence of DON in Canadian grains grain foods and beers Infant cereal foods
oat barley soy and multi-grain based foods
Province Sample
type
Incidence
()
Maximum
DON
contents
(microgg)
Multiple
mycotoxins
samples ()
Survey
period
Sources
Ontario Spring
Wheat
70 260 Yes 2006-08 OMAFRA1
Ontario Winter
wheat
49 290 Yes 2006-08 OMAFRA
Ontario Maize 78 140 Yes 2009-10 Limay-Rios
and
Schaafsma
2010
Ontario Maize - 1000 Yes 2011 AampL
Biologicals
Canada Breakfast
cereals
46 094 430 1999-01 Roscoe et al
2008
Canada Animal
feeds
73 425 Yes 1990-10 Zeibdawi et
al 2011
Canada Beers 58 0052 50 1990s Scott 1997
Canada Infant
Cereals
63 098 Yes 1997-99 Lombaert et
al 2003
1
Ontario Ministry of Agriculture and Rural Affairs
2 microgmL
14
Table 1-4 Legislative maximum tolerated levels of trichothecene mycotoxins in Canada
according to Canadian Food Inspection Fact Sheet April 23 2009 DON- deoxynivalenol
DAS- diacetoxyscirpenol
Products DON
(microgg)
T-2 toxin
(microgg)
HT-2 toxin
(microgg)
DAS
(microgg)
Uncleaned soft wheat for human
consumption
2 - - -
Cattlepoultry diets 5 lt1 01 lt1
Swinedairy cattle feeds 1 lt1 0025 lt2
15
High DON incidence (78) was observed in maize samples collected from
southwestern Ontario fields with moderate levels (lt1 microgg) of contamination during 2009 and
2010 (Limay-Rios and Schaafsma 2010) Some samples had high DON levels (14 microgg) In
the 2011 crop season extremely high levels of DON (gt100 microgg) were found in some
southern Ontario maize samples (AampL Biologicals London ON Personal communication)
In a Canadian Food Inspection Agency (CFIA) 20 year survey (1990-2010) 73 of
animal feed samples collected from feed mills farms and retail stores across Canada were
contaminated with DON and for more than 25 of the samples the mycotoxin levels were
above the maximum limit (ge 1 microgg) (Zeibdawi et al 2011) The highest level of DON (15
microgg) detected in the 2006-2007 samples were much lower than the levels found in the 1999-
2000 samples which contained a maximum of 425 microgg DON Importantly this study
reported a significant number of samples contaminated with multiple mycotoxins DON NIV
3-ADON 15-ADON T-2 toxin HT-2 toxin DAS and NEO
Earlier studies in the 1990s detected high levels of DON content in Eastern Canadian
cereals maize (maximum 175 microgg) wheat (maximum 916 microgg) barley (maximum 911
microgg) and oats (maximum 12 microgg) (Campbell et al 2002) The study identified a significant
number of samples (15-367) that contained multiple mycotoxins and up to 33 of the
wheat samples exceeded 1 microgg DON Low amounts but high incidences of TCs were found
in cereal-based foods collected from Canadian retail stores Sixty three percent of infant
cereal foods 46 of breakfast cereals and 58 of beers were contaminated with one or
multiple mycotoxins (Scott 1997 Lombaert et al 2003 Roscoe et al 2008) The maximum
amount of DON was found in infant foods (098 microgg) followed by breakfast cereals (094
microgg) and beers (005 microgmL)
16
15 Significance of deoxynivalenol and trichothecenes
For most cereals including wheat maize rice barley oats and rye the risk of
contamination with TCs resulted in grain quality reduction and FHBGER disease progression
(Bai et al 2002 Jansen et al 2005) The impacts of TC contamination in agricultural
commodities are diverse including reduced livestock production induced human illnesses
and enhanced FHB disease severity which may result in the reduction of crop yield Exposure
of farm animals (particularly pigs) to DON is a continuing threat to the livestock industries in
Canada USA and Europe (DrsquoMello et al 1999) Ingestion of TC-contaminated feeds
generally causes reduction of feed intake feed refusal poor feed conversion retardation of
weight gain increased disease susceptibility and reduced reproductive capacities (Binder et al
2007) High doses of DON result in acute toxic effects in pigs (eg emesis) while low doses
(ge1 microgmL) cause chronic effects (eg growth retardation in pigs) (Rotter et al 1996
Accensi et al 2006) Reduction of growth and reproduction ability was observed in
experimental rats exposured to much higher doses of TC (Sprando et al 2005 Collins et al
2006) Several animal disease outbreaks that occurred in different continents have been
attributed to consumption of feed contaminated with DON NIV and DAS (Desjardins 2006)
The consequences of human exposure to TCs include liver damage gastricintestinal
lesions and central nervous system toxicity resulting in anorexia nausea hypotensionshock
vomiting diarrhea and death (Pestka 2007) Human mycotoxicosis (illnesses and outbreaks)
by consumption of type-A and type-B TC mycotoxins in Europe New Zealand Russia
Japan South America India and Vietnam have been reported (Bhat et al 1989 Yoshizawa
1993 Luo 1994 Desjardins 2006) Severe outbreaks of Alimentary Toxic Aleukia (ATA)
associated with TC killed thousands of people in the Orenburg region of Russia during the
17
1930rsquos and 1940rsquos (Eriksen 2003) TCs (T-2 DON and NIV) were likely used for biological
warfare in Laos Kampuchea and Afghanistan and its potential application for bioterrorism is
an additional concern associated with TC (Tucker 2001 Bennett and Klich 2003)
TCs are phytotoxic and may contribute to FHB disease severity in cereals (Proctor et
al 1995 Maier et al 2006 Masuda et al 2007) Studies demonstrated that DON caused
wilting chlorosis inhibition of seedling growth and necrosis in cereals (Cossette and Miller
1995 Mitterbauer et al 2004 Rocha et al 2005) DON-mediated host cell damage and
subsequent necrotrophic fungal spread has been shown to lead to FHBGER disease
development and higher kernel damage (Procter et al 1995 Mitterbauer et al 2004 Maier et
al 2005) This suggests that DON contributes to yield losses as well as to reductions in the
market value of cereals (Osborne and Stein 2007) Severely infected grain may contain more
than 60 microgg DON which is unacceptable to the milling and brewing industries (Okubara et
al 2002) Significant crop losses due to FHB disease epidemics and mycotoxin contamination
have been reported in Ontario (Schaafsma 2002) FHB disease epidemics during 1998 to 2000
caused huge crop losses resulting in farm failures in the northern Great Plains in the USA
(Windels 2000) In that period FHB associated losses were estimated at US $ 27 billion in
the region (Nganje et al 2002)
Importantly Fusaria infested grains are often co-contaminated with multiple
mycotoxins which further increase health risks as interactions between the mycotoxins may
cause greater toxic effects than expected (Schollenberger et al 2006) This implies that an
apparently low level of contamination of a single mycotoxin can have severe effects on
consumers because of synergistic effects among multiple co-occuring mycotoxins (Surai et al
2010)
18
A number of studies demonstrated synergistic toxic effects among Fusarium
mycotoxins on pigs turkeys lamb chicks as well as in in vitro Caco-2 cell and yeast
bioassays (Table 1-5)
Table 1-5 In vitro and in vivo synergistic effects of different food contaminating mycotoxins
DON-deoxynivalenol DAS-diacetoxyscirpenol FB1-fumonisins 1 NIV-nivalenol ZEN-
zearelenone FA-fusaric acid Afl-aflatoxin
Mycotoxins source Interacting
mycotoxins
Experimental
animalcells
Effects Reference
Naturally contaminated
cereals
DON FA Pigs Inhibit growth Smith et al
1997
DON contaminated
wheat and FB1
DON FB1 Pigs Weight loss Harvey et al
1996
Culture materials
contained mycotoxins
T-2 FB1 Turkey poults Weight loss oral
lesion
Kubena et al
1997
Inoculated rice and
pure mycotoxin
DAS Afl Lamb Weight reduction Harvey et al
1995
Naturally DON
contaminated wheat
and pure T-2 toxin
DON T-2 Chicks Reduced body
weight significantly
Kubena et al
1989
Pure mycotoxins DAS Afl Chicks Reduced body
weight
Kubena et al
1993
Pure mycotoxins T-2 Afl Chicks Reduced weight
oral lesion
Kubena et al
1990
Pure mycotoxins DON
ZENFB1
Human intestinal
Caco-2 cells
Increased DNA
fragmentations
Kouadio et al
2007
Pure mycotoxins DON NIV Yeast bioassay Synergistic effects Madhyastha et
al 1994
Pure mycotoxins T-2 HT-2 Yeast bioassay Synergistic effects Madhyastha et
al 1994
19
16 Toxic effects of deoxynivalnol and trichothecenes
As small molecules TCs simply enter into cells and interact with different cell
components resulting in disruption of cellular functions and causing various toxic effects that
include inhibition of protein synthesis and ribosomal fragmentation (Eudes et al 2000
Minervini et al 2004 Pestka 2007) Simple absorption of DON into human intestinal Caco-2
cells and crossing human placenta barriers through passive diffusion has been reported
(Sergent et al 2006 Nielsen et al 2011)
TC may cause various toxicities in animals and plants including apoptosis
immunotoxicity chromatin condensation disruption of cellularmitochondrion membranes
disruption of gut tissues inhibition of protein and ribosome synthesis breakage of ribosome
and cell damage The toxicity of TC is influenced by structural variability (ie position
number and type of functional groups attached to the tricyclic nucleus and C-C double bond at
C9-10 position) (Desjardins 2006) Nevertheless epoxide function at the C12-13 position is
the key for TC cytotoxicity (Swanson et al 1988 Eriksen et al 2004)
Different in vitro and in vivo studies revealed that the method and period of exposure
amount of TC and types of celltissue and host species have significant effects on TCrsquos
cytotoxicity (Scientific Committee for Food 2002 Rocha et al 2005 Nielsen et al 2009)
Administration of 10 mgkg body weight (bw) NIV caused apoptosis in mice thymus Peyerrsquos
patch and spleen tissues (Poapolathep et al 2002) DON and NIV at 887 mgkg bw for 3 days
a week for four weeks induced immunotoxicity in experimental mice determined by
estimating increased immunoglobulins (IgA) and decreased IgM levels (Gouze et al 2007)
Exposure of mice to 1 mgkg bw of DON induced skeletal malformations and reduced mating
tendency while significant resorption of embryos observed at the dose of gt10 mgkg bw
20
(Khera et al 1982) At high acute doses (4972 mgkg bw) DONNIV and type-A
trichothecenes caused mice death because of intestinal haemorrhage necrosis of bone
marrowlymphoid tissues and kidneyheart lesions (Yoshizawa and Morooka 1973
Yoshizawa et al 1994 Forsell et al 1987 Ryu et al 1988) In a separate study 15-ADON
showed more toxicity than DON when the mycotoxins were orally administered to mice
(Forsell et al 1987)
Studies suggested that type-A TCs are more toxic in mammals than type-B which
means that functional variability at the C-8 position of TC has a significant role in toxicity
The toxicity of various TC mycotoxins in animals can be ranked in the order of T-2
toxingtHT-2 toxingtNIVgtDONge3-ADONgt15-ADON according to the results from human cell
lines and mice model systems bioassays (Ueno et al 1983 Pestka et al 1987 Nielsen et al
2009) Moreover animal feeding assays demonstrated that monogastric animals are most
sensitive to DON The order of sensitivity of animals to DON is
pigsgtrodentsgtdogsgtcatsgtpoultrygtruminants (Pestka 2005) The reasons for the variations in
DON toxicity species might be due to differences in metabolism absorption distribution and
elimination of the mycotoxin among animal species (Pestka 2007)
To my knowledge the mechanism of TC uptake by plant cells has not been studied
Nonetheless cereals are exposed to TC by toxigenic Fusaria infection which may cause
various toxicities such as cell damage wilting chlorosis necrosis program cell death and
inhibition of seedling growth in sensitive plants (eg wheat maize and barley) (Cossette and
Miller 1995 Rocha et al 2005 Boddu et al 2007) As animals TC inhibits protein synthesis
in non-host A thaliana (Nishiuchi et al 2006) Immunogold assays showed that the cellular
targets of DON in wheat and maize were cell membrane ribosome endoplasmic reticulum
21
vacuoles and mitochondria Modulation of cell membrane integrity and electrolyte losses
were observed by direct DON interference in maize and wheat cell assays (Cossette and
Miller 1995 Mitterbauer et al 2004) DON-mediated mitochondrion membrane alterations
disruption of host cells enzymatic activity apoptosis and necrosis have been observed in
cereals (Mitterbauer et al 2004 Rocha et al 2005) In contrast to animal type-B is more toxic
to plants than type-A DON showed 8- and 13-fold phytotoxicity to wheat coleoptiles than T-
2 and HT-2 toxins respectively (Eudes et al 2000) By compilation of different studies TC
phytoxicity can be ordered as DONge3-ADONgtT-2 toxingtDAS (Wakulinski 1989 McLean
1996 Eudes et al 2000)
22
17 Cellular and molecular mechanisms of DON toxicity in animals and plants
The underlying cellular and molecular mechanisms of DONTC toxicity are evolving
Exposure to DON proceeds with the up and down regulation of numerous critical genes of
eukaryotic organisms which causes various negative effects on animal and plant organisms
(Figure 1-2)
One of the early events of DON toxicity occurs when the mycotoxin binds to the
active ribosome resulting in blocking protein translation through different routes by
damaging the ribosome (Shifrin and Anderson 1999) degrading or altering conformational
changes of 28S-rRNA (Alisi et al 2008 Li and Pestka 2008) and inducing expression of
miRNAs This results in down regulation of the ribosome associated genes (He et al 2010a
Pestka 2010) Both animal and plant cell assays demonstrated that DON bound to the peptidyl
transferase (Rpl3) domain of the 60S subunit of ribosome and blocked protein chain
elongation and termination at high DON concentrations while peptide chain termination and
ribosome breakage occurred at a low level of DON exposure (Zhou et al 2003a)
23
Figure 1-2 Cellular and molecular mechanisms of deoxynivalenol toxicity in eukaryotic
organisms DON is simply absorbed by the cell and interacts with the cytoplasmic membrane
(CM) mitochondrion (MT) membrane and ribosome Disruption of the CM and alteration of
the MT membrane causes a loss of electrolytes and affects the function of the MT
respectively DON binds to the active ribosomes which leads to ribosome damage and
induces micro RNAs (miRNA) Expression of miRNA impairs the synthesis of ribosome
associated protein The subsequent transduction and phosphorylation of the RNA-activated
protein kinase (PKR) and hematopoietic cell kinase (Hck) activate the mitogen-activated
protein kinase (MAPK) cascade Phosphorylation of the MAPKs induces targeted
transcription factors (TF) which results in ribotoxis stress response upregulation of
proinflammatory cytokines and dysregulation of neuroendocrine signaling (derived from
Pestka 2007 and 2010)
24
The consequence of DON interaction with ribosomes leads to cellular and molecular
changes which are complex and not fully understood However it is well-demonstrated that
the activation of the mitogen-activated protein kinase (MAPK) and subsequent molecular
actions play key roles in the cytotoxicity of TC Upon binding to the ribosome DON rapidly
induced interactions between the protein kinases and the ribosomal 60S subunit as observed in
mononuclear phagocytes (Bae and Pestka 2008) Ribosomal binding to DON activated the
ribosome associated kinases for instances protein kinase R (PKR) hematopoietic cell kinase
(Hck) and P38
mitogen-activated protein kinase The protein kinases are induced by double-
stranded RNA (dsRNA) or possibly by the damaged ribosomal peptides (Iordanov et al 1997
Shifrin and Anderson 1999 Zhou et al 2003b Pestka 2007) Subsequent phosphorylation of
kinases (eg PKRHckP38
) activates the MAPK cascade resulting in ribotoxis stress
response upregulation of proinflammatory cytokines and dysregulation of neuroendocrine
signaling (Pestka and Smolinki 2005 Pestka 2010) Studies suggested that high doses of
DON induces ribotoxis stress response (apoptosis) preceded by immunosuppression and
disruption of intestinal permeabilitytissue functions (Yang et al 2000 Pestka 2008) Further
study observed stabilization of the mRNA and expression of transcription factors for critical
genes when cells were exposed to a low concentration of DON (Chung et al 2003)
The MAPK pathway proceeds with upregulation of a broad range of proinflammatory
cytokine genes and dysregulation of neuroendocrine signaling within the central and enteric
nervous systems Modulation of neuroendocrine signaling by DON increased the level of
serotonin resulted in vomiting reduced food intake and growth retardation in experimental
animalscell lines (Prelusky and Trenholm 1993 Li 2007) Upregulation of proinflammatory
cytokines and chemokines genes was determined in mice liver spleen kidney and lung
25
following in 1 h of exposure to DON A subsequent shock-like effect was observed in mice
because of a ldquocytokine stormrdquo (Pestka 2008) Induction of proinflammatory cytokines also led
to various cellular effects on the central nervous system interference of the innate immune
system and growth hormone signaling These contribute to anorexia growth retardation
aberrant immune response and tissue dysfunction as demonstrated in experimental animal
systems (Pestka 2010)
DON-mediated growth retardation in mice via suppression of growth hormones
(insulin-like growth factor 1 and insulin-like growth factor acid-labile subunit) has been
reported by Amuzie et al (2009) Disruption of gut tissues by DONNIV and subsequent
impairment of intestinal transporter activity was demonstrated in a human epithelial cell
model (Marseca et al 2002) The study proposed that alteration of gut tissue permeability
may cause gastroenteritis induction and growth retardation in livestock animals Growth
retardation of livestock may also occur via dysregulation of neuroendocrine signaling-
mediated induction of emesis and anorexia (Pestka et al 2010)
The molecular mechanism of TC toxicity in plant systems has not been well
documented A recent study showed the upregulation of numerous defense related genes in
wheat upon exposure to DON (Desmond et al 2008) The study further suggested that DON-
induced program cell death in wheat occurred through a hydrogen peroxide signaling
pathway This mechanism had not been observed in mammals
26
18 Strategies to control FHB and trichothecenes
During the past decades many pre- and post-harvest strategies have been applied to
protect cereal crops from Fusarium infestation and to eliminate TC from contaminated
agricultural commodities (FAO 2001 Munkvold 2003 Varga and Toth 2005 Kabak and
Dobson 2009) Some of the above crop protection techniques have been patented (He and
Zhou 2010) In spite of those efforts TC contamination of cereals appears to be unavoidable
using the currently available methods
Pre-harvest control strategies involving cultural practices (eg crop rotation tillage
early planting) and fungicide applications partially reduced Fusarium infection and DON
contamination in cereals (Dill-Macky and Jones 2000 Schaafsma et al 2001) However
repeated application of fungicides may lead to the evolution of Fusarium strains resistant to
fungicides with greater mycotoxin production ability (DrsquoMello et al 1998) At post-harvest
reduction of grain moisture by artificial drying storage at low temperatures (1-4degC) and
proper ventilation of the storage room have shown to be effective in inhibiting fungal growth
and mycotoxin production in infected grain (Midwest Plan Service 1987 Wilcke and Morey
1995) This technology has become less applicable particularly in developing countries
because of the frequent observation required to maintain proper storage conditions (Hell et al
2000)
Removal of Fusarium infected grains by physical methods (eg mechanical
separation density segregation colour sorting etc) was unable to eliminate TC completely
(Abbas et al 1985 He et al 2010b) This is partly because FHB symptomless kernels may
contain a considerable amount of DON which are not eliminated by physical separation
procedures (Seitz and Bechtel 1985) Several studies showed that milling and food processing
27
steps were unable to completely remove DON from grain-based foods (Jackson and
Bullerman 1999 Placinta et al 1999 Kushiro 2008) Detoxification of TC by chemical
treatments (eg ozone ammonia chlorine hydrogen peroxide and sodium metabisulfite) has
been demonstrated (Young et al 1986 Young et al 1987 Young et al 2006) The problem
with these techniques is that the application of such chemicals is not economically viable and
it also has a negative impact on grain quality (Karlovsky 2011) Although the technique has
been popular and showed effectiveness to certain mycotoxins utilization of absorbents to
inhibit mycotoxin absorption by the livestock gastrointestinal tract appeared to be ineffective
for TC (Huwig et al 2001)
The introduction of Fusarium resistant cultivars would be the best strategy to address
the FHB and associated mycotoxin problems in cereals However breeding for cultivars
resistant to FHBGER is difficult and time consuming as natural resistance against Fusarium
is quantitative in nature FHB resistant quantitative trait loci (QTLs) have been identified in
several cereal genotypes (eg Sumai 3 Ning 7840 Bussard) (Bernando et al 2007 Golkari
et al 2009 Haumlberle et al 2009 Yang et al 2010) The 3B 5A and 6B QTLs for type-I
(resistant to initial infection) and type-II (resistant to fungal spread in the kernel) resistances
have been identified on different wheat chromosomes (Buerstmayr et al 2009) Type-I
(Qfhsifa-5A) and type-II (Fhb1Fhb2-3B) QTLs from Sumai 3 are being used in many
breeding programs and pyramiding of the QTLs in spring wheat showed additional effects
Nonetheless the key genes involved in FHB resistance in cereals are yet to be identified This
makes it difficult to integrate the resistance genes into elite cultivars
28
Cultivar development by direct incorporation of TCDON resistance genes may be an
alternative way of protecting grains from these mycotoxins contaminations Many genes from
distantly related species have been integrated into crop cultivars using a transgenic approach
with great economic benefits (Brookes and Barfoot 2006) DON transforming trichothecene
3-O-acetyltransferase (TRI101 from Fusarium sporotrichioides) and UDP-
glucosyltransferase (from Arabidopsis thaliana) genes expressed in cereal cultivars showed
some DONDAS detoxifying ability under in vitro and controlled conditions (Kimura et al
1998 Okubara et al 2002 Poppenberger et al 2003 Ohsato et al 2007) By contrast no
reduction of mycotoxins in either of the above cases was observed under field conditions
(Manoharan et al 2006) Importantly the transformed acetylated and glycosylated
compounds could revert to the original substances or convert to even more toxic products
(Gareis et al 1990 Alexander 2008)
19 Isolation of DONTC de-epoxidating micro-organisms and genes
An enzyme with the capability of de-epoxidizing DONTC appears to be a potential
source for completely removing TC from contaminated cereals (Karlovsky 2011) However
such trichothecene detoxifying genes do not exist in cereals and other plants (Boutigny et al
2008) Because of this several research groups have attempted to obtain microorganisms that
encode DONTC de-epoxidation genes Mixed microbial culture capable of de-epoxydizing
trichothecenes have been isolated from chicken gut (He et al 1992) cow ruminal fluid (King
et al 1984 Swanson et al 1987 Fuchs et al 2002) pig gut (Kollarczik et al 1994) and
digesta of chicken (Young et al 2007) rats (Yoshizawa et al 1983) sheep (Westlake et al
1989) and fish (Guan et al 2009) All of the microbial cultures cited above showed de-
29
epoxydation capabilities only under anaerobic conditions realtively high temperatures and at
lowhigh pH except microbes from fish intestine that showed efficient de-epoxidation at low
temperatures and wide range of temperatures (Guan et al 2009) Such enzymes active under
anaerobic conditions high temperatues and low pH may have potential applications as feed
additives and the eubacterial strain BBSH 797 is an animal feed additive product with the
brand name Mycofixreg by BIOMIN (He et al 2010b) Biotechnological applications of
anaerobically-active enzymes from the above microbial sources seem to be inappropriate
under the aerobic and moderate conditions at which cereals plants are grown
Most of these efforts have failed to isolate single DONTC degrading microbes
effective under aerobic conditions because of inappropriate microbial growth conditions
under laboratory conditions Earlier attempts were made to obtain TC de-epoxidizing aerobic
bacteria but the culture did not retain DONNIV de-epoxidation activity after reculturing
(Ueno 1983 He et al 1992) In addition the co-metabolic nature of the microbial TC de-
epoxidation reaction has limited the use of mycotoxins as the sole nutrient source for selective
isolation of the target microorganisms Several DONTC de-epoxidation microbial strains
including Bacillus strain LS100 and Eubacterial strain BBSH 797 have been isolated from
chicken gut and rumen fluids through multi-step enrichment procedures (Fuchs et al 2002
Yu et al 2010) A DON to 3-keto DON transforming bactial strain was islated from soil by
prolonged incubation with high concentration DON (Shima et al 1997)
Microbial genesenzymes with TC de-epoxidation activity are not known Based on
the microbial TC de-epoxidation activity of a mixed microbial culture it has been proposed
that ceratin reductases or oxidoreductases may catalyze such a biochemical reaction (Islam et
al 2012) With the advent of molecular techniques many novel genes with
30
industrialagricultural value have been cloned by reverse genetics (genomic DNA library
preparation and functional screening of the library clones) forward genetics (transposon
mutagenesis) and comparative genomics methods (Frey et al 1998 Koonin et al 2001
Suenaga et al 2007 Chung et al 2008 Ansorge 2009) The tremendous advancement of next
generation sequencing technology and bioinformatics tools may facilitate the identification of
the microbial genes responsible for DONTC detoxification (Ansorge 2009) Isolation of TC
de-epoxidation genes may play a major role in producing safe foods and feeds derived from
grains
110 The bacterial species Citrobacter freundii
Citrobacter freundii is a bacterial species belonging to the family Enterobacteriace It
is a facultatively anaerobic and Gram-negative bacillus (Wang et al 2000) The bacterium
survives almost everywhere including animalhuman intestines blood water and soils
(Wang et al 2000 Whalen et al 2007) C freundii is known as an opportunistic human
pathogen and causes a variety of respiratory tract urinary tract and blood infections and
responsible for serious neonatal meningistis disease (Badger et al 1999 Whalen et al 2007)
Interestingly this bacterium plays a positive environmental role through nitrogen recycling
(Puchenkova 1996) Strains of C freundii also contribute to the environment by sequenstering
heavy metals and degrading xenobiotic compounds (Kumar et al 1999 Sharma and Fulekar
2009)
31
111 Hypotheses and objectives
The study was based on the following hypotheses
i Certain microorganisms in DON contaminated agricultural soils are able to transform DON
to de-epoxy DON (de-epoxidation) under aerobic conditions and moderate temperatures
ii DON de-epoxidation may be achieved by a single microbial strain
iii Microbial DON detoxifying genes and associated regulators are able to express and
function in a closely related host organism
iv DON detoxifying microbial strains carry genes for de-epoxidation activity that are absent
from related species that have not evolved under DON selection pressure
The objectives of this research were to
i Select microbial cultures from soil having DON de-epoxidation activity under aerobic
conditions and moderate temperatures by analyzing potential DON contaminated soils in
various growth conditions namely aerobicanaerobic conditions different growth media
temperature and pH
ii Select for organisms with DON de-epoxidation activity from the mixed microbial culture
by treating with antibiotics heating subculturing on agar medium and incubating for a
prolonged period in media with a high DON concentration
iii Identify microbial de-epoxidation genes by producing a microbial genomic DNA library
from the DON de-epoxidizing strain followed by screening of the clones for DON de-
epoxidation
32
iv Determine microbial de-epoxidation genes by sequencing of the genome and comparative
analysis of the genome with closely related species and identification of putative DON
detoxification genes
33
CHAPTER 2
20 AEROBIC AND ANAEROBIC DE-EPOXIDATION OF MYCOTOXIN
DEOXYNIVALENOL BY BACTERIA ORIGINATING FROM AGRICULTURAL
SOIL
Published in World Journal of Microbiology and Biotechnology 2012 287-13
21 Abstract
Soil samples collected from different cereal crop fields in southern Ontario Canada
were screened to obtain microorganisms capable of transforming deoxynivalenol (DON) to
de-epoxy DON (dE-DON) Microbial DON to dE-DON transformation (ie de-epoxidation)
was monitored by using liquid chromatography-ultraviolet-mass spectrometry (LC-UV-MS)
The effects of growth substrates temperature pH incubation time and aerobic versus
anaerobic conditions on the ability of the microbes to de-epoxidize DON were evaluated A
soil microbial culture showed 100 DON to dE-DON biotransformation in mineral salts
broth (MSB) after 144 h of incubation Treatments of the culture with selective antibiotics
followed an elevated temperature (50ordmC) for 15 h and subculturing on agar medium
considerably reduced the microbial diversity Partial 16S-rRNA gene sequence analysis of the
bacteria in the enriched culture indicated the presence of at least Serratia Citrobacter
Clostridium Enterococcus Stenotrophomonas and Streptomyces The enriched culture
completely de-epoxidized DON after 60 h of incubation Bacterial de-epoxidation of DON
occurred at pH 60 to 75 and a wide array of temperatures (12 to 40ordmC) The culture showed
34
rapid de-epoxidation activity under aerobic conditions compared to anaerobic conditions This
is the first report on microbial DON to dE-DON transformation under aerobic conditions and
moderate temperatures The culture could be used to detoxify DON contaminated feed and
might be a potential source for gene(s) for DON de-epoxidation
Keywords biotransformation de-epoxidation deoxynivalenol soil bacteria vomitoxin
22 Introduction
Deoxynivalenol (DON commonly known as lsquovomitoxinrsquo) 3α 7α 15-trihydroxy-
1213-epoxytrichothec-9-en-8-one is a trichothecene produced by toxigenic Fusarium species
causing Fusarium head blight (FHB) or Gibberella ear rot (GER) disease on small grain
cereals and maize respectively (Starkey et al 2007 Foroud and Eudes 2009) DON
contamination in cereals and maize is widespread and can lead to tremendous losses to the
growers It has also detrimental effects on human and livestock health (Pestka and Smolinski
2005 Rocha et al 2005) The toxin interferes with the eukaryotic ribosomal peptidyl
transferase (Rpl3) activity resulting in human and animal mycotoxicosis (Ueno 1985 Bae
and Pestka 2008) Common effects of DON toxicity in animals include diarrhea vomiting
feed refusal growth retardation immunosuppression reduced ovarian function and
reproductive disorders (Pestka and Smolinski 2005 Pestka 2007) In plants DON causes a
variety of effects viz inhibition of seed germination seedling growth green plant
regeneration root and coleoptile growth (cited in Rocha et al 2005) In addition DON
enhances disease severity in sensitive host plants (Maier et al 2006)
35
Avoidance of DON contamination or reduction of toxin levels in contaminated grains
remains a challenging task Physical and chemical processes are either inappropriate or
prohibitively expensive to use to decrease the DON content of agricultural commodities
(Kushiro 2008 He et al 2010b) The use of Fusarium resistant crop varieties might be the
best method to reduce DON contamination However breeding for FHB or GER resistant
cultivars appears to be very difficult and time consuming as natural sources of resistance
against Fusarium-caused diseases is quantitative in nature (Ali et al 2005 Buerstmayr et al
2009) As an alternative expressing genes for enzymatic detoxification of DON in elite
cultivars may be an effective strategy for protecting the value of feeds and foods (Karlovsky
1999) Heterologous expression of eukaryotic trichothecene 3-O-acetyltransferase and UDP-
glucosyltransferase genes in cereals and Arabidopsis have resulted in reduced toxicity via
acetylation and glycosylation of the DON (Poppenberger et al 2003 Ohsato et al 2007)
It has been proposed that enzymatic de-epoxidation of trichothecene toxins could be
used to limit DON contamination (Zhou et al 2008 Foroud and Eudes 2009) DON belongs
to type B-trichothecenes which contain an epoxide ring at C12-13 and the epoxide is highly
reactive and mainly responsible for toxicity (Figure 2-1) (Swanson et al 1988 Eriksen et al
2004)
36
Figure 2-1 Mechanism of microbial DON de-epoxidation Removal of the oxygen atom from
the epoxide ring results in formation of a C=C double bond at C-12 position which reduces
cytotoxicity (Swanson et al 1988 Eriksen et al 2004)
Microbial DON de-epoxidation by single bacterial strains and by mixed cultures has
been reported (King et al 1984 Swanson et al 1987 He et al 1992 Kollarczik et al 1994
Fuchs et al 2002 Young et al 2007 Guan et al 2009 Yu et al 2010) However all of these
microbes originated from fish and other animals and were strictly anaerobic The recovered
microbial cultures and single strains only showed de-epoxidation of trichothecenes at low
(le15ordmC) and high (gt30degC) temperatures Such enzymes might have limited applications for
toxin decontamination in cereal grains The aim of the current study was to isolate the
microbes from soil capable of transforming DON to a de-epoxy metabolite under aerobic
conditions and moderate temperatures
De-epoxydation
Deoxynivalenol (DON)
De-epoxyDeoxynivalenol (dE-DON)
37
23 Materials and methods
231 Soil samples
The upper layer (0-30 cm) of soils along with crop debris were randomly collected
from cereal (eg maize wheat barley) crop fields during 2004-2006 in Southern Ontario
Canada Detailed of the crops and locations of the fields have been described in Jianwei Hersquos
Master Degree thesis (He 2007) Briefly soil samples were collected in sterile plastic bags
and transported to the laboratory Samples were sieved to remove large roots stones and
stored at 4ordmC until used The samples were mixed with grounded mouldy corn (containing 95
microgg DON) + F graminearum and incubated at 22-24degC for six weeks Five samples were
prepared for DON de-epoxidation activity analysis by dispersing 5 g soil in 50 mL distilled
water and storing at4degC until further use
232 Culture media
Media used in the study were nutrient broth (NB) Luria Bertani (LB) minimal
medium (MM) (He 2007) six mineral salts + 05 bacto peptone (MSB) (He 2007) tryptic
soy broth (TSB) corn meal broth (CMB) (Guan et al 2009) potato dextrose broth (PDB) six
mineral salts + 05 yeast extract broth (MSY) and soil extract (SE) MM was composed of
sucrose-100 g K2HPO4-25 g KH2PO4-25 g (NH4)2HPO4-10 g MgSO47H2O-02 g
FeSO4-001 g MnSO47H2O- 0007 g water- 985 mL and MSB was composed of the same
salts as MM but was supplemented with Bactotrade Peptone (05 wv) as the sole C source
instead of sucrose To prepare soil extract 100 g of the composite soil was homogenized in 1
L water The soil particles and plant debris were removed by passing the soil suspension
through Whatmanreg grade 3 filter paper (Sigma-Aldrich Oakville Ontario Canada) The
38
extract was autoclaved at 121degC for 30 min Media and ingredients were purchased either
from Fisher Scientific Inc (Fair Lawn New Jersey USA) Sigma-Aldrich (Oakvile Ontario
Canada) or Difco (Sparks Maryland USA) Prior to autoclaving pH of the broth media was
adjusted to 70 Solid media were prepared by adding 15 agar
233 Screening of soil samples
Frozen soil suspensions were thawed and vortexed to homogenize the microbial
population An aliquot (100 microL) of the suspension was immediately transferred to MSB PDB
and MM broth media DON at 50 microgmL was added to the microbial suspension cultures The
cultures were incubated at 22-24degC with continuous shaking at 180 revolutions per minute
(rpm) The extent of DON de-epoxidation was determined at 24 h 48 h 72 h 96 h 120 h and
144 h of incubation
234 Determination of de-epoxidation product
Microbial DON de-epoxidation was measured according to Guan et al (2009)
Samples were prepared by mixing equal volume of microbial culture and methanol and
allowed to stand for 30 min at room temperature Microorganisms in the mixture were
removed by passing them through a 045 microm Millipore filter (Whatmanreg Florham Park New
Jersey USA) The filtrates were used for de-epoxidation analysis by a liquid chromatography-
ultraviolet-mass spectrometry (LC-UV-MS) Identification and quantification of DON and its
transformation product was achieved using a Finnigan LCQ DECA ion trap LC-MS
instrument (ThermoFinnigan San Jose CA USA) The equipment detects 30-2000 mz
(molecular mass ion ratio) compounds and able to determine 001 micromL DON The HPLC
39
system was equipped with a Finnigan Spectra System UV6000LP ultraviolet (UV) detector
and an Agilent Zorbax Eclipse XDB C18 column (46times150 mm 35 μm) The binary mobile
phase consisted of solvent A (methanol) and solvent B (water) The system was run with a
gradient program 25 A at 0 min 25ndash41 A over 5 min isocratically at 41 A for 2 min
and 41ndash25 A over 1 min There was a 3 min post-run at its starting condition to allow for
the equilibration of the column The flow-rate was set at 10 mLmin UV absorbance was
measured at 218 nm Atmospheric pressure chemical ionization (APCI) positive ion mode
was used for MS detection A DON standard was used to adjust the instrument to its
maximum response The system was operated as follows shear gas and auxiliary flow-rates
were set at 95 and 43 (arbitrary units) voltages on the capillary tube lens offset multipole 1
offset multipole 2 offset and entrance lens were set at 3000 minus1700 minus370 minus850 minus3500
and minus4200 V respectively capillary and vaporizer temperatures were set at 200 and 450degC
respectively and the discharge needle current was set at 7 μA Identification of mycotoxin
was achieved by comparing their retention times UV spectra and mass spectrum with DON
and de-epoxy DON (Biopure Union MO USA) as standards Quantification of DON and
product was based on peak areas related to their UV and MS calibration curves
40
235 Biological DON de-epoxidation
To help identify the types of microorganisms causing the DON de-epoxidation MSB
medium was supplemented with either 100 microgmL penicillin G or streptomycin sulfate for
suppressing gram positive or negative bacterial growth and activities respectively Three
replications of antibiotic-treated media were inoculated with 100 microL of microbial culture
capable of de-epoxidizing DON The antibiotic-treated cultures were incubated at 27degC
spiked with 50 microgmL DON Controls were prepared without adding antibiotics in the
microbial culture Inhibition of DON de-epoxidation was analyzed after five day of
incubation
236 Reduction of microbial diversity by antibiotic treatments heating and
subculturing on agar medium
In order to reduce non-DON degrading microbial populations the DON de-
epoxidation capable cultures were sequentially treated with antibiotics having diverse modes
of action including i) dichloramphenicol that kills various gram positive Gram-negative and
anaerobic bacteria ii) trimethoprim that kills Gram-positive and negative bacteria iii)
penicillin that is lethal to Gram-positive bacteria and iv) cyclohexamide to kill filamentous
fungi and yeasts Antibiotics were purchased from Sigma-Aldrich Antibiotic stock solutions
(10 mgmL) were prepared with sterile water or methanol Microbial cultures in MSB
medium were treated with 100 microgmL of each antibiotic followed by incubation of the culture
at 50ordmC for 15 hours to kill undesirded microbial species After heating cultures were treated
with 50 microgmL DON and incubated at 27ordmC for five days DON de-epoxidation activity of the
microbial cultures was determined following the procedure described above Cultures that
41
showed gt99 de-epoxidation were considered for subsequent antibiotic and heat treatments
Aliquots of the achieved culture were spread on MSB agar plates and grown at room
temperature for few days Colonies appeared on the plates were collected and incubated in
MSB medium supplemented with 50 microgmL DON De-epoxidation ability of the subcultured
microbial cultures was determined after five days of incubation at 27degC by LC-UV-MS
method
237 Analysis of microbial population diversity
Terminal Restriction Fragment Length Polymorphism (T-RFLP) analysis
Initially microbial diversity was determined by terminal restriction fragment length
polymorphism (T-RFLP) fingerprint analysis (Bruce and Hughes 2000) Genomic DNA from
microbial cultures with and without enrichment was extracted using DNeasy Plant DNA
Extraction Mini Kit (Qiagen Inc Mississauga Ontario Canada) 16S-rDNA was amplified
by PCR using the HEX-labeled forward primer Bac8f
(5-agagtttgatcctggctcag-3) and
unlabeled reverse primer Univ1492r (5-ggttaccttgttacgactt-3) (Fierer and Jackson 2006)
Fifty microlitter PCR mixture for each sample was prepared having 1x HotStarTaq Master
Mix (Qiagen) 05 microM of each primer 50 microg of BSA and 50 ng of DNA The 35 PCR cycles
followed 60 s at 94degC 30 s at 50degC and 60 s at 72degC After size verification by agarose-gel
electrophoresis 50 ng of purified PCR products was digested for 3 h in a 20 microL reaction
mixture with 5 U of HhaI restriction enzyme (New England Biolabs Ipswich MA USA)
The size and relative abundance of the labeled T-RFLP fragments were separated by capillary
electrophoresis in an automated DNA sequencer equipped with fluorescence detector
(Applied Biosystem 3730 DNA Analyzer) The DNA fragments were analyzed with the Peak
42
Scannertrade Software v10 to create an electropherogram with the genetic fingerprints of the
microbial community
Analysis of V3-V5 region of the 16S-rDNA
To define the bacterial genera present in the enrichment culture the 16S-rDNA was
amplified using two sets of bacterial universal primers Primarily 16S-rDNA was PCR
amplified using the primers 46f-5rsquogcytaacacatgcaagtcga3rsquo (Kaplan and Kitts 2004) and
1371r-5rsquogtgtgtacaagncccgggaa3rsquo (Zhao et al 2007) PCR was performed in a total volume of
25 μL which contained 06 μM of each primer 1x of HotStarTaqreg Master Mix (Qiagen Inc)
and 20-40 ng of DNA The PCR cycles included the following steps 95ordmC (15 min) 35 cycles
of 94ordmC (20 s) 52ordmC (15 s) 72ordmC (1 min 30 s) and a 72ordmC (7 min) extension step The second
pair of universal bacterial primers were 27f-5rsquoagagtttgatcmtggctcag3rsquoand 1492r-
5rsquoggttaccttgttacgactt3rsquo (Lane 1991) PCR was conducted with 25 cycles following the
procedures of Tian et al (2009) Amplification was carried out in a GeneAmpreg PCR System
9700 (Applied Biosystems) Furthermore the PCR products were used as templates to
amplify the hypervariable V3-V5 region of the 16S-rDNA using primers 357f-
5rsquocctacgggaggcagcag3rsquo and 907r7 5rsquoccgtcaattcctttgagttt3rsquo (Yu and Morrison 2004)
After purification of the PCR products by QIAquick PCR purification kit two
amplicon libraries were prepared by ligating to pCR-TOPO vector (Invitrogen) and the library
clones were transformed into TOPO10 competent cells according to Invitrogenrsquos TOPO
cloning manual The clone libraries were plated on LB plates containing 25 μgmL
kanamycin Fifty colonies from each library were randomly picked up from the overnight
plates and inserts were sequenced as described by Geng et al (2006) PCR products were
43
sequenced with an automated ABI 3730 DNA analyzer (Applied Biosystems) The sequence
similarities of the amplified V3-V5 region were compared with the ribosomal database project
(RDP) (httprdpcmemsuedu) Sequences with greater than 95 identity were assigned to
the same bacterial genus
238 Factors influencing DON de-epoxidation
DON de-epoxidation ability of the enriched microbial culture was investigated under
different conditions as described below
Maintaince of active culture
To retain microbial de-epoxidation activity the active culture has been maintained in
MSB medium containing 50 microgmL DON
Media growth temperature and pH
Overnight microbial cultures (100 μL) were added to 2 mL of NB LB MM TSB
MSB CMB MSY PDB or SE broths The pH of the media was adjusted to 70 and the
experiments were conducted at 27degC temperature The effect of temperature was studied by
incubating the culture in MSB medium (pH 7) at 12 20 25 30 and 40ordmC To determine the
effects of pH MSB medium pH was adjusted to 55 60 65 70 75 and 80 Media with
different pHs were inoculated with aliquots of the overnight active culture and then incubated
at 27ordmC The cultures were supplemented with 50 μgmL DON before incubation Three
replications for each medium temperature and pH condition were evaluated
44
De-epoxidation under aerobic and anaerobic conditions
Three replicate microbial cultures of five mL each were grown in sterile 15 mL
centrifuge tubes under aerobic conditions and in an anaerobic chamber filled with 95 CO2
and 5 H2 (COY Laboratory Products Inc Ann Arbor MI USA) Three days after initiation
half (25 mL) of the cultures from anaerobic conditions was transferred to new centrifuge
tubes and brought to aerobic conditions Similarly half of the aerobically grown cultures were
placed into an anaerobic chamber Cultures moved from anaerobic to aerobic conditions were
shaken after opening the lids of the tubes in a laminar flow every eight hours intervals to
maintain aerobic conditions in the culture Lids of the culture tubes transferred to anaerobic
conditions were kept loose to allow oxygen to escape from the culture Two days after
transferring to the new conditions the cultures in both conditions were treated with 50 microgmL
DON and incubated at 27degC Every 12 h 500 microL were removed from each tube and analyzed
for microbial DON de-epoxidation activity by LC-UV-MS
239 Data analysis
Data were processed using Microsoft Excel Program 2007 Statistical analysis was
done using General Linear Model procedures in SAS version 91 (SAS Institute Inc Cary
NC USA 2002-03) The results were the average of three replications stated as mean plusmn
standard error Differences between means were considered significant at plt 005 according
to Tukeyrsquos-HSD test
45
24 Results
241 Analyses of DON to dE-DON biotransformation
A typical LC-UV-MS analysis where there is incomplete de-epoxidation of DON by a
particular soil suspension culture is shown in Figure 2-2 Analysis by LC with UV detection
and integration of peak areas showed 662 conversion of DON to dE-DON (Figure 2-2a)
Analysis LC with MS detection (expressed as total ion counts) indicated 643 conversion
(Figure 2-2b) LC analyses with UV and TIC (total ion count) detection are subject to
interferences due to co-eluting contaminants and therefore integration of small peaks was
very difficult making quantification inaccurate MS detection by SIM (selected ion
monitoring) can eliminate the influence of co-eluting contaminants For example the SIM LC
chromatogram for DON (Figure 2-2c) was generated by using the major ions resulting from
the APCI positive ion MS of DON (Figure 2-2e)
Similarly the SIM LC chromatogram for dE-DON (Figure 2-2d) was derived from the
MS ions of dE-DON (Figure 2-2f) The signal to noise ratios (SN) of peaks observed by SIM
were at least 20 times greater than those detected by TIC because there is reduced likelihood
of co-eluting peaks having the same ions Integration of the SIM peaks revealed a 616
conversion The three methods gave similar results but the SIM LC results were considered
to be the most accurate Using that method the conversion of DON to dE-DON by the mixed
culture occurred most quickly at 48 h and was completed by 144 h (Figure 2-3)
46
47
Figure 2-2 LC-UV-MS chromatographic analysis of DON and its de-epoxidated product dE-
DON obtained by culturing with selected soil microbes (a) LC-UV chromatogram of product
mixture monitored at 218 nm (b) LC-MS total ion chromatogram (TIC) of product mixture
(c) selected ion monitoring (SIM) chromatogram of cultured DON at mz 231 249 267 279
and 297 (d) SIM chromatogram of cultured dE-DON at mz 215 233 245 251 263 and 281
(e) atmospheric pressure chemical ionization (APCI) positive ion mass spectrum of DON (f)
APCI positive ion mass spectrum of dE-DON
48
242 Screening of soil samples
Initially samples originating from soils were analyzed in three types of culture media
to support the growth of fungi yeast and bacteria However cultures growing in PDB which
is a suitable substrate for many fungi and yeasts did not permit transform DON to dE-DON
(data not shown by microorganisms) A define medium (ie MM) was also used in the
sample screening assay None of the soil samples showed de-epoxidation activity in MM
(data not shown) From five soil samples one soil sample culture showed de-epoxidation of
DON in MSB The culture in MSB converted gt99 DON to dE-DON within 144 h (Figure 2-
3) Conversion began after a lag of 48 h and was linear thereafter with a rate of 1 per hour
49
Figure 2-3 Time course of relative biotransformation of DON to dE-DON by the initial soil
suspension culture in MSB medium The culture contained 50 μgmL DON at time 0 Culture
with three replications was incubated at room temperature with continuous shaking at 180
rpm The conversion of DON to dE-DON was analysed by LC-UV-MS at 24 h intervals Blue
and red lines are the percent conversion of DON and recovery of dE-DON respectively
50
243 Evidence of biological DON de-epoxidation
Treatment of active microbial culture with 100 microgmL of streptomycin completely
inhibited DON de-epoxidation In contrast the same concentration of penicillin G did not
affect activity of the culture (Figure 2-4) Streptomycin is effective against Gram-negative
bacteria while penicillin G is lethal to Gram-positive bacteria These experiments provided
evidence that DON de-epoxidation may be caused by certain Gram-negative bacteria
51
Figure 2-4 Microbial de-epoxidation of DON after five days of incubation The active DON
de-epoxidation culture was treated with either 100 microgmL penicillin G or streptomycin
(Strepto) in MSB Cultures having three replications were added 50 microgmL DON and
incubated at room temperatures with continuous shaking at 180 rpm For controls cultures
were without antibiotics DON to dE-DON transformation was monitored after five day of
incubation by LC-UV-MS method
0
20
40
60
80
100
120
Penicillin Strepto Control
DO
N d
e-e
po
xyd
ati
on
(
)
Antibiotics
52
244 Reduction of the microbial diversity
Plating the suspension cultures onto MSB agar revealed colonies with a variety of
sizes morphologies and colours reflecting a mixed culture (data not shown) To reduce the
complexity of the culture three steps of selection pressures were applied namely treatment
with antibiotics heat shocks and subculturing on solid medium The above indicated
treatments did not significantly affect DON de-epoxidation of the microbial cultures (data not
shown) T-RFLP profiles of the mixed culture before treatments showed six distinct peaks of
about 110 150 200 220 340 and 420 bp restriction fragments with the 110 bp peak being
the predominant one (Figure 2-5a)
After treatment with antibiotics and heat only three peaks at 110 150 and 325 bp
remained (Figure 2-5b) suggesting that there was a reduction in the microbial diversity
without affecting DON de-epoxidation After enrichments the bacteria species in the culture
changed their growth patterns as the electropherogram produced different sizes of pick
lengths compared to untreated culture
53
(b)
(a)
Rela
tive flu
ore
scen
ce
32000
0
0 500
16S-rDNA fragment size (bp)
100 200 300 400
Rela
tive flu
ore
scen
ce
32000
0
500
16S-rDNA fragment size (bp)
0 300200100 400
54
Figure 2-5 Analysis of microbial population reduction by terminal restriction fragment
length polymorphism (T-RFLP) (a) Restriction fragments of the 16S-rDNA of the untreated
culture and (b) fragmentation patterns of the enriched culture Picks along the X-axis showed
the fragmentation patterns of the 16S-rDNA which is related to the bacterial species diversity
The length of picks (along the Y-axis) showed abundance of terminal restriction fragments
related to the abundance of particular bacterial speciesgenera
55
245 Microbial population structures in the enriched culture
Microbial diversity in the enriched culture was determined by analyzing the sequences
of the V3-V5 region (~586 bp) amplified from the 16S-rDNAs of the microorganisms it
contained The sequence analysis showed the presence of Serratia Citrobacter Clostridium
Enterococcus Stenotrophomonas and Streptomyces in the enriched culture (Figure 2-6)
Out of 100 V3-V5 sequences 54 sequences were found to be belonging to Serratia
the predominant species of the culture The remaining sequences matched with 12
Citrobacter 8 Clostridium 14 Enterococcus 8 Stenotrophomonas and 4 Streptomyces
species
56
Figure 2-6 Microbial genera identified in the enriched DON de-epoxidizing microbial
culture The bacterial genera were determined by comparing the hypervariable regions (V3-
V5) of the 16S-rDNA using ribosomal database project (httprdpcmemsuedu) X-axis
showed the identified bacterial genera and Y-axis demonstrated the abundance of the V3-V5
sequences belonging to different bacterial genera
54
12 8
14
8 4
0
20
40
60 S
eq
uen
ce
fre
qu
en
cy
Microbial genera
57
246 Effects of media growth temperature pH and oxygen requirements on DON de-
epoxidation
Of the nine media that were tested DON de-epoxidation by the enriched microbial
culture only occurred in NB and MSB (Figure 2-7a) It appeared that C andor N
concentrations in the media were crucial for the enzymatic activity because less or more than
05 Bacto Peptone in MSB reduced the microbial DON transformation ability
DON de-epoxidation activity occurred over a wide range of temperatures from 12 to
40ordmC (Figure 2-7b) The fastest rates of DON de-epoxidation occurred between 25 to 40ordmC
De-epoxidation was observed within a relatively narrow range of pH (60 to 75) with
the most rapid DON biotransformation activity observed at pH 70 (Figure 2-7c) At pH 70
the enriched culture showed DON de-epoxidation activity 125 times faster than at pH 60 or
75 At pH 55 and 80 there was no observable DON de-epoxidation
58
0
20
40
60
80
100
120
12degC
20degC
25degC
30degC
40degC
DO
N d
e-e
po
xyd
ati
on
(
)
Temperature
0
20
40
60
80
100
120
DO
N d
e-e
pxyd
ati
on
(
)
Media
(a)
(b)
59
Figure 2-7 Effects of culture conditions on DON to de-epoxy DON transformation by the
enriched microbial culture For the media experiment (a) all the media were adjusted to pH 7
and the incubation temperature was 27degC For the temperature experiment (b) MSB pH 7 was
used For the pH experiment (c) MSB at 27degC was used Microbial cultures were inoculated
with 50 μgmL DON De-epoxidation was determined after 72 h of incubation Media MSY
(mineral salts + 05 yeast extract) LB (Luria Bertani) TSB (tryptic soy broth) MSB
(mineral salts + 05 bacto peptone) SE (soil extract) CMB (corn meal broth) PDB (potato
dextrose broth) and MM (minimal medium)
0
20
40
60
80
100
120
55
60
65
70
75
80
DO
N d
e-e
po
xyd
ati
on
(
)
pH
(c)
60
The enriched culture showed DON de-epoxidation activity under both aerobic and
anaerobic conditions (Figure 2-8) The fastest transformation (100 de-epoxidation after 60 h
of incubation) occurred in cultures (A) that were grown under aerobic conditions throughout
the culture growth period A medium rate of de-epoxidation (100 de-epoxidation after 72 h)
was observed in cultures that were initially grown in either aerobic or anaerobic conditions
followed by transferring to anaerobic or aerobic conditions (A+AN and AN+A) respectively
The lowest rate occurred (100 de-epoxidation after 96 h) in cultures grown in continual
anaerobic conditions (AN) The means comparison of microbial DON de-epoxidation
efficiencies at 60 h of incubation showed that cultures continually grown in conditions A and
AN were statistically different Biotransformation abilities of the cultures that were switched
from one conditions to another (A+AN and AN+A) were statistically similar among each
other but different from cultures that were only grown in conditions A or AN (plt005)
61
Figure 2-8 Microbial DON to dE (de-epoxy DON)-DON transformation activity under
aerobic and anaerobic conditions A = culture incubated and examined the activity only under
aerobic conditions A+AN = culture initially was grown in aerobic conditions and then
transferred to anaerobic conditions AN = cultures grown throughout the assay under
anaerobic conditions AN+A = culture initially grown under anaerobic conditions and then
transferred to aerobic conditions Differences of lower case letters are significantly different
in de-epoxidation capability (after 60 h) among incubation conditions according to Tukeyrsquos
HSD test (p lt 005) The experiment was conducted with three replications
62
25 Discussion
The current study describes a novel bacterial culture isolated from agricultural soil that
was capable of transforming DON to de-epoxy DON under aerobic and anaerobic conditions
at moderate temperatures Because it has been speculated that DON translocates to soil from
contaminated crops and DON that enters into the soil could be metabolized by microbes
(Voumllkl et al 2004) soil samples for the current study were mainly collected from fields where
cereal crops contaminated with DON were grown
In our initial sample screening steps upwards of 144 h of incubation were required to
achieve gt99 DON de-epoxidation Such a long period may have been due to the presence of
fast growing microorganisms that had suppressive effects on the growth or activity of the
DON transforming bacteria (Davis et al 2005) The sequential use of different antibiotics
with different modes of actions followed by thermal treatments and subculturing on solid
medium resulted in a reduction in the number of bacterial species as determined by T-RFLP
analysis Analysis of V3-V5 region of the 16S-rDNA showed that five bacterial genera were
present in the enriched culture The culture showed higher DON de-epoxidation activity (only
60 h required for gt99 biotransformation) compared to initial soil suspension culture that
required 144 h to complete the de-epoxidation This microbial culture could be used to isolate
the DON de-epoxidizing gene(s) applying metagenomics approaches (Daniel 2005)
The culture showed complete de-epoxidation activity both in MSB and NB media
However we have found that the use of nutritionally rich NB medium enhanced non-target
bacterial (eg Serratia lequifaciences) growth but this did not occur in MSB which is a
comparably poor medium To avoid the possible suppressive and detrimental effects of the
fast growing bacteria on the target bacteria MSB was used for enrichment and
63
characterization of the culture An important characteristic of the DON de-epoxidation
activity observed for the microbial culture in the current study is its activity at moderate
temperature (25-30degC) In contrast previously isolated cultures showed optimal activities at
low (le15degC) or high temperatures (37degC) (Guan et al 2009) Also the culture showed DON
de-epoxidation activity at neutral pH range (pH 60-75) in contrast to previous reports which
found that microbial trichothecene de-epoxidation occurred at acidic and basic pHs (Swanson
et al 1987 He et al 1992 Kollarczik et al 1994 Young et al 2007) However a microbial
culture from fish gut showed de-epoxidation under wide range of pHs (Guan et al 2009)
Probably the pH range reflects the environment in which the bacteria naturally grow which in
this case is similar to the soil pH of a typical crop field in Southern Ontario of Canada The
advantage of the present finding is that DON de-epoxidation activity of the culture was
optimal in a temperature range that matches the temperature range of activity for most arable
plants Therefore it is reasonable to suggest that the gene(s)enzyme(s) responsible for the
activity have potential application for enzymatic detoxification of DON in cereals (Karlovsky
1999)
The current observation that bacterial DON de-epoxidation activity occurred under
both aerobic and anaerobic conditions was interesting because DON de-epoxidation is a
reductive chemical reaction that might be expected to be promoted by anaerobic conditions
However there are previous reports of aerobic de-epoxidation of eldrindieldrin by soil
bacteria (Matsumoto et al 2008) and Goodstadt and Ponting (2004) described aerobic
reductive de-epoxidation by vitamin K epoxide reductasephylloquinone reductase A
definitive determination of the oxygen sensitivity of the enzyme responsible for DON de-
epoxidation will ultimately come from experiments with the purified protein
64
In conclusion this study reports on a novel bacterial source from soil that efficiently
de-epoxidizes DON at moderate temperatures under both aerobic and anaerobic conditions
This research may lead to the isolation of microbes and gene(s) encoding enzyme(s) capable
of de-epoxidizing DON that could create transgenic plants with reduced DON contamination
thus improving food and feed safety and reducing negative effects of the toxin on public
health
26 Acknowledgements
The authors gratefully acknowledged the Natural Sciences and Engineering Research
Council of Canada (NSERC) for awarding a scholarship to Rafiqul Islam for PhD program
The research was supported by the Agriculture and Agri-Food Canada Binational
Agricultural Research and Development Fund (AAFCBARD) project We are thankful for
the contributions of Jianwei He Xiu-Zhen Li and Honghui Zhu
65
CHAPTER 3
30 ISOLATION AND CHARACTERIZATION OF A NOVEL SOIL BACTERIUM
CAPABLE OF DETOXIFYING ELEVEN TRICHOTHECENE MYCOTOXINS
31 Abstract
Deoxynivalenol (DON vomitoxin) is one of the most commonly detected food and
feed contaminating mycotoxins worldwide In order to develop a control method effective
under aerobic conditions and moderate temperatures a bacterial strain ADS47 was isolated
capable of transforming DON to the less toxic de-epoxy DON The bacterial strain is
facultatively anaerobic Gram-negative rod shaped with an average cell size of 150 X 072
microm and has peritrichous (multiple) flagella The 16S-rRNA gene sequence had gt99
nucleotide identity to Citrobacter freundii a bacterium that belongs to the gamma subclass of
proteobacteria Strain ADS47 showed near 100 de-epoxidation of DON after 42 h of
incubation in nutrient broth (NB) under aerobic conditions moderate temperature (28˚C) and
neutral pH (70) However de-epoxidation could occur at 10-45˚C and pH 60-75 Under
anaerobic conditions near 100 de-epoxidation was observed after 72 h mainly during the
stationary growth phase Microbial DON de-epoxidation ability increased with increasing
DON concentrations in the culture media Inhibition of bacterial de-epoxidation by sodium
azide suggested that an active electron transport chain was required In addition to DON
strain ADS47 was able to de-epoxidize andor de-acetylate five type-A and five type-B
trichothecene mycotoxins This is the first report of a bacterial isolate capable of de-
epoxidizing DON and trichothecene mycotoxins under aerobic conditions and moderate
66
temperatures Strain ADS47 could potentially be used in bioremediation to eliminate DON
and other trichothecene mycotoxins from contaminated food and feed
Keywords aerobic de-epoxidation Citrobacter freundii isolation soil bacterium
trichothecenes
32 Introduction
Cereals are often contaminated with a variety of type-A and type-B trichothecene
mycotoxins produced by a number of toxigenic Fusarium species (Placinta et al 1999
Binder et al 2007 Foroud and Eudes 2009) Trichothecenes are tricyclic sesquiterpenes with
a double bond at the C9 10 position and a C12 13 epoxy group (Desjardins 2006) The two
types are distinguished by the presence of a ketone at position C8 in type-B which is absent
in type-A (Yazar and Omurtag 2008) The type-B toxin deoxynivalenol (DON) is the most
frequently found mycotoxin in food and feed around the world (Larsen et al 2004 Desjardins
2006 Starkey et al 2007) Different Fusarium species associated with Fusarium head blight
(FHB) or Gibberella ear rot (GER) disease of small grain cereals and maize are mainly
responsible for contaminating grains with DON and other type-A and type-B trichothecenes
These can cause serious mycotoxicosis in humans animals and plants (Ueno 1985 DrsquoMello
et al 1999 Bae and Pestka 2008) Common effects of DONTC toxicity in animals include
diarrhea vomiting feed refusal growth retardation immunosuppression reduced ovarian
functionsreproductive disorders and even death (Pestka and Smolinski 2005 Pestka 2007)
DON enhances FHB disease severity in wheat and maize (Proctor et al 1995 Desjardins
1996 Maier et al 2006)
67
Reduction of mycotoxin levels in contaminated agricultural products remains a
challenging task Current physical and chemical processes to decrease DON content are either
ineffective or prohibitively expensive (Kushiro 2008 He et al 2010b) However the use of
detoxifying microbes to biodegrade mycotoxins is an alternative (Awad et al 2010
Karlovsky 2011) The toxicity of DON is mainly due to the epoxide and there is a significant
reduction of DON toxicity (54) when it is reduced to produce de-epoxy DON Related
mycotoxins also show reductions in toxicity when they are de-epoxidized such as NIV (55)
and T-2 toxin (400) (Swanson et al 1988 Eriksen et al 2004) Bacillus arbutinivorans
strain LS100 and Eubacterial strain BBSH 797 originating from chicken intestine and rumen
fluid respectively are capable of cleaving the epoxide ring of DON under anaerobic
conditions and high temperature (37degC) (Fuchs et al 2002 Yu et al 2010) Strain BBSH 797
which can de-epoxidize DON scirpentriol and T-2 triol trichothecene mycotoxins has been
used as an additive for poultry and swine diets (Karlovsky 2011) However there are no
reports of microbes able to de-epoxidze DON and other trichothecenes under aerobic
conditions and at moderate temperatures
To develop an effective method for eliminating DON and trichothecene mycotoxins
from contaminated grains soils from fields with FHBGER infected crops were screened for
DON de-epoxidizing microbes (refer to chapter 2 Islam et al 2012) One soil sample yielded
a mixed microbial culture was partially enriched for bacteria with DON de-epoxidizing
activity (chapter 2) The current study describes the purification and characterization of a
novel DON de-epoxidizing bacterium from this culture
68
33 Materials and methods
331 Microbial source
A mixed microbial culture originating from a screen of more than 150 separate soil
samples collected from maize and wheat fields near Guelph Ontario Canada (Islam et al
2012) was used for enrichment purification and characterization of the DON de-epoxidizing
bacterium
332 Media and chemicals
The media used were nutrient broth (NB) 110 strength NB (110 NB) minimal
medium (MM) consisting of six mineral salts (MS) + 100g per liter sucrose (Islam et al
2012) MS + 05 Bacto Peptone (MSB) MS + 03 Bacto Peptone (MSC) MS + 05
yeast extract (MSY) MS + 200 microgmL DON (MS-DON) sterile water-soluble soil extract
(SE) (Islam et al 2012) potato dextrose broth (PDB) and Luria Bertani (LB) Prepared media
and media components were obtained from Fisher Scientific (Fair Lawn New Jersey USA)
Sigma-Aldrich (Oakville ON Canada) andor Difco (Sparks Maryland USA) Media pH
was adjusted as required prior to autoclaving For solid media preparation 15 agar was
added DON and all trichothecene mycotoxins were purchased from Biomin (Romer Lab
Union MO USA)
333 Isolation of the DON de-epoxidizing bacterial strain
Aliquots (20 microL) of the microbial culture partially enriched for DON de-epoxidizing
ability (Islam et al 2012) was transferred to 05 mL MSB (pH 70) in sterile 15 mL
Eppendorf tubes After addition of 200 microgmL pure DON the culture was incubated at room
69
temperature with continuous shaking at 220 revolutions per minute (rpm) Seven to ten days
later 20 microL of the culture was transferred to fresh MSB supplemented with 200 microgmL DON
The culture was then incubated for 7 to 10 days under the above conditions This procedure
was repeated 12 times
After three months of repeated transfers in broth with 200 microgmL DON 100 microL of the
culture was serially diluted in 09 mL MSB containing 05 Tweenreg20 (Sigma Aldrich) to
reduce bacterial cell aggregation (Mabagala 1997) The dilutions were vigorously shaken for
ten minutes A 100 microL aliquot from each dilution was spread on MSB + agar (MSA) The
MSA plates were incubated at room temperature until single colonies appeared
Single colonies from plates were transferred to 1 mL MSB+ 50 microgmL DON and
incubated at room temperature with continuous shaking at 180 rpm under aerobic conditions
After three days the DON de-epoxidation ability was examined by mixing an equal volume
of the bacterial culture and methanol and incubating for 30 min at room temperature to lyse
the cells Insoluble materials in the mixture were removed by passing it through a 045 microm
Millipore filter (Whatmanreg Florham Park New Jersey USA) The level of de-epoxy DON
(dE-DON also known as DOM-1) in the filtrate was determined by liquid chromatography-
ultraviolet-mass spectrometry (LC-UV-MS) (Guan et al 2009)
Individual colonies that showed de-epoxidation activity were serially diluted in
MSB+05 Tweenreg20 and plated onto MSA Single colonies that appeared on the agar plate
were incubated in MSB as described above and retested for DON de-epoxidation ability To
ensure that pure cultures were obtained this was repeated 10 times and individual colonies
were selected on MSA each time
70
334 Identification of a DON de-epoxidizing bacterium
Strain ADS47 with DON de-epoxidation ability was obtained from a single colony
after 10 serial transfers and was grown on MSA at room temperature under aerobic
conditions for 72 h Colony color shape edge and elevation were observed The cell
morphology of the bacterium was examined with a transmission electron microscope at the
University of Guelph Guelph Ontario Canada Whole bacterial cells were placed directly on
a formvar and carbon-coated 200-mesh copper grid They were washed two times by touching
the grids to a drop of clean dH2O and the excess water was blotted off The samples were
negatively stained using 1 Uranyl Acetate (aq) for five seconds Electron microscopy was
performed on a Philips CM10 transmission electron microscope at 60kV under standard
operating conditions (Philips Optics Eindhoven Netherlands) Gram staining was performed
using the procedure described in the commercial gram stain kit from BD Canada
(Mississauga Ontario Canada)
Total bacterial DNA was extracted using a DNeasy Plant DNA Extraction Mini Kit
(Qiagen Inc) The 16S-rDNA was PCR amplified using the primers 46f-
5rsquogcytaacacatgcaagtcga3rsquo (Kaplan and Kitts 2004) and 1371r-5rsquogtgtgtacaagncccgggaa3rsquo
(Zhao et al 2007) PCR was performed in 25 microL containing 06 microM of each primer 1x of
HotStarTaqreg Master Mix (Qiagen Inc) and ~40 ng of DNA The PCR cycles were 95ordmC (15
min) 35 cycles of 94ordmC (20 s) 52ordmC (15 s) 72ordmC (1 min 30 s) and a 72ordmC (7 min) extension
step Amplification was carried out in a GeneAmpreg PCR System 9700 (Applied Biosystems)
PCR products were sequenced with an automated ABI 3730 DNA analyzer (Applied
Biosystems) The 16S-rRNA gene sequence similarity was performed by BlastN using the
Ribosomal Database Project (RDP) (httprdpcmemsueduseqmatch)
71
A phylogenetic tree was constructed by comparing the partial 16S-rDNA sequence of
strain ADS47 with the 11 known Citrobacter species (Bornstein and Schafer 2006 Brenner et
al 1993) The Citrobacter strains were C freundii [FN997616] C koseri [AF025372] C
amalonaticus [AF025370] C farmeri [AF025371] C youngae [AB273741] C braakii
[AF025368] C werkmanii [AF025373] C sedlakii [AF025364] C rodentium [AF025363]
C gillenii [AF025367] and C murliniae [AF025369] Escherichia coli O157H7 [AE005174]
was used as an out group strain The analysis was performed using distance matrix with a gap
opening of 100 and a gap extension of 01 by a server based ClustalW2 program
(httpwwwebiacukToolsmsaclustalw2) A dissimilarity tree was created and viewed
using a TreeView X software (Page 1996)
335 Effects of DON concentration aeration substrate temperature pH and antibiotic
To determine if DON concentration can affect the growth of the isolated bacteria
tubes containing 5 mL of NB or MSB with 20 200 or 500 microgmL of DON were inoculated
with strain ADS47 that had been grown for 16 h with continuous shaking at 220 rpm and then
adjusted to cell density at 025 OD600nm (107 cellsmL) The cultures were incubated at 28degC
with continuous shaking at 180 rpm and 05 mL was removed every 12 h to measure
OD600nm The culture (05 mL) was then analysed for DON de-epoxidation activity by LC-
UV-MS (Guan et al 2009) as described above Three replicate cultures were tested
To examine the effect of aeration five mL of NB or MSB containing 50 microgmL DON
were inoculated with an overnight culture of strain ADS47 prepared as described above and
incubated at 28˚C with continuous shaking at 180 rpm One mL of culture was removed every
six hours and bacterial growth was determined by measuring cell density at OD600nm At the
72
same time points culture samples were analysed for DON to dE-DON transformation ability
(Guan et al 2009) The experiment was repeated with MSB medium containing 50 microgmL
DON under anaerobic conditions as described by Islam et al (2012) with DON to dE-DON
transformation ability being determined every 6 h Three replicate cultures were prepared for
each assay
The effect of substrate on DON de-epoxidation ability was tested after growing
ADS47 for 72 h in NB 110 NB MM MS-DON MSB MSC MSY SE PDB or LB
containing 50 microgmL DON For this assay the media were maintained at pH70plusmn02 and 28degC
for 72 h The effect of temperature was examined with MSB containing 50 microgmL DON at 10
15 20 25 30 35 40 45 or 50degC The effect of pH on growth of strain ADS47 was examined
with MSB adjusted to pH 55 60 65 70 75 or 80 and the cultures were incubated at 28degC
for 72 h Three replicate cultures were used for each assay
Overnight cultures of ADS47 in MSB were treated with 001 and 0001 (wv) of
an antibiotic sodium azide (SA) (Sigma-Aldrich) Controls were prepared in MSB without
SA Three replicate cultures were spiked with 50 microgmL DON DON to dE-DON
transformation ability of the bacterial culture was analysed (Guan et al 2009) after 72 h at
28degC with continuous shaking at 180 rpm Three replicate cultures were tested
336 Localization of DON to dE-DON transformation ability in the bacterium
Bacterial cell extracts were prepared using B-PER II Bacterial Protein Extraction
Reagent following the manufacturerrsquos instructions (PI 78260 Thermo Scientific Rockford
IL USA) ADS47 was grown in 15 mL MSB for 48 h under the growth conditions described
above A culture (05 mL) was centrifuged at 5000 x g for 10 minutes and the pellet was
73
collected B-PER II reagent (2 mL) was added to the pellet and the cells were lysed by
pipetting the suspension up and down several times The cell lysate was treated with 20 microL
Halt Protease Inhibitor Cocktail (Thermo Scientific Rockford IL USA) Culture filtrate was
prepared by passing 05 mL of the culture through a 022 microm Millipore filter (Fisher
Scientific) The remaining 05 mL culture was used as control Cell lysate culture filtrate and
control were incubated with 50 microgmL DON and incubated at 28degC DON to dE-DON
transformation ability of the cell lysate culture filtrate and control were analysed after 72 h
incubation (Guan et al 2009) Three replicate cultures were tested
337 Detoxification of additional trichothecene mycotoxins
Strain ADS47 was evaluated for its ability to degrade five type-A trichothecene
mycotoxins (HT-2 toxin T-2 toxin T2-triol diacetoxyscirpenol and neosolaniol) and five
type-B trichothecene mycotoxins (nivalenol verrucarol fusarenon X 3-acetyl-
deoxynivalenol and 15-acetyl-deoxynivalenol) One and a half mL NB was added with 100
microgmL of each mycotoxin and then inoculated with 100 microL of ADS47 culture grown
overnight as previously described The culture of NB + mycotoxin + ADS47 was incubated at
28degC with continuous shaking at 180 rpm and DON to dE-DON transformation ability of the
culture was analysed by LC-UV-MS (Guan et al 2009) Five replicate cultures were tested for
each trichothecene mycotoxin
74
338 Statistical analysis
Data were processed using Microsoft Excel Program 2007 Statistical analysis was
performed using General Linear Model procedure in SAS version 91 (SAS Institute Cary
NC USA 2002-03) Replications were averaged and stated as mean plusmn standard error The
mean differences were considered significant at plt 005 according to Tukeyrsquos-HSD test
75
34 Results
341 Isolation and identification of a DON de-epoxidizing bacterium
A DON de-epoxidizing bacterium designated strain ADS47 was isolated after 12
transfers over three months from cultures growing in MSB containing 200 microgmL DON
Strain ADS47 produced small to medium size circular convex and white colonies on MSA at
room temperature under aerobic conditions (Figure 3-1a) The bacterium is rod shaped and
150 x 072 microm in size (Figure 3-1b) It has numerous flagella that cover the surface of the cell
(peritrichous flagella) and is Gram-negative
BlastN analysis of the 16S-rDNA sequence of strain ADS47 showed the most
similarity to the eleven known Citrobacter species in the gamma proteobacteria of the
Enterobacteriaceae In particular the 16S-rDNA nucleotide sequence of ADS47 was more
than 99 identical to C fruendii (Figure 3-2) The next most similar sequences were a cluster
of C werkmanii and C murliniae followed by more distantly related C braakii C gilleni
and C youngae A separate cluster was formed by C rodentium C farmeri C sedlakii C
koseri and C amalonaticus
76
Figure 3-1 (a) Colony color shape and size of the mycotoxin degrading bacterial strain
ADS47 on mineral salts agar (MSA) medium (b) Transmission electron microscopic (TEM)
image of the Citrobacter freundii strain ADS47 The bacterium showed multiple flagella
(peritrichous flagella) and numerous fimbriae around the cell surface
(a) (b)
77
Figure 3-2 Dendrogram based on 16S-rDNA sequences for strain ADS47 and 11 reference
Citrobacter species An E coli O157H7 strain was considered as an out group species The
tree was constructed using a genetic distance matrix by ClustalW 2 and viewed by
TreeViewX software The distance was converted to scale and shown by branch lengths (scale
bar 001 which indicated the 16S-rDNA nucleotide sequence variability between the bacterial
species) GenBank accession numbers for 16S-rDNA sequences of the species used for
comparison are shown in materials and methods section (334)
78
342 Effects of DON concentrations on bacterial DON de-epoxidation
Growth of ADS47 aerobically in MSB with 20 200 and 500 microgmL DON was not
significantly different from that observed in control cultures without DON (Figure 3-3a)
Similarly increasing DON concentrations did not affect bacterial growth in NB medium
(Figure 3-3b)
DON de-epoxidation activity determined as microgmL DON convertedcell density
(OD600) was measured lowest in MSB+20 microgmL DON and highest in MSB+ 500 microgmL
DON containing cultures (Figure 3-3c) The highest rate of dE-DON formation was 115 microgh
in ADS47 cultures in MSB with 500 microgmL DON between 48 to 72 h All of the DON was
converted in the cultures containing 20 microgmL DON and 200 microgmL DON between 48-60 h
and 60-72 h respectively In 200 microgmL and 500 microgmL of DON cultures the rate of de-
epoxidation increased significantly after 36 h
79
00
04
08
12
16
0 24 36 48 60 72 96
OD
600n
m
Incubation time (h)
Control
20 microgmL
200 microgmL
500 microgmL
(b)
(a)
80
Figure 3-3 Effects of DON concentration on bacterial growth and de-epoxidation activities
Strain ADS47 was grown in MSB and NB media with three different DON concentrations 20
microgmL 200 microgmL and 500 microgmL Control cultures were prepared without DON Cultures
were grown at 28degC with continuous shaking at 180 rpm (a) Microbial density (OD600 nm) in
MSB with different DON concentrations was measured at 24 36 48 60 72 and 96 h (b)
Microbial growth (OD600 nm) in NB having different DON concentration was measured at 24
48 72 and 96 h (c) Bacterial DON de-epoxidation activity in 20 200 and 500 microgmL DON
containing cultures at different time points DON de-epoxidation was measured in microgmL per
OD600 nm Three replicate cultures for each assay were analysed
(C)
81
343 Influence of growth conditions and antibiotic on microbial DON de-epoxydation
Aerobic and anaerobic conditions
ADS47 grew more quickly uner aerobic conditions under MSB+50 microgmL DON than
in MSB+50 microgmL DON under anaerobic conditions (Figure 3-4a) Aerobic growth was
faster in NB+50 microgmL DON compared to MSB+50 microgmL DON
However the DON de-epoxidation activities on a cell density basis were very similar
in NB+50 microgmL DON compared to MSB+50 microgmL DON (Figure 3-4b) Also DON de-
epoxidation activity of ADS47 under aerobic conditions in MSB+50 microgmL DON was higher
than in the same medium under anaerobic conditions (Figure 3-4b)
82
(b)
(a)
83
Figure 3-4 Bacterial growth and DON de-epoxidation capabilities under aerobic and
anaerobic conditions (a) Microbial growth under aerobic conditions in MSB and NB medium
and anaerobic conditions in MSB medium (b) Microbial DON de-epoxidation ability under
aerobic conditions in MSB and NB media and anaerobic conditions in MSB medium
Cultures media having 50 microgmL DON were inoculated with overnight culture of strain
ADS47 Initial microbial concentration in the cultures was adjusted to 025 OD600nm Cultures
were incubated at 28degC having three replications Bacterial growth at OD600nm and DON de-
epoxidation in microgmL per OD600nm were measured at six hour interval
84
Media temperature and pH
Growth of ADS47 in media containing 50 microgmL DON under aerobic conditions at 72
h was three-fold higher in NB and LB than in SE two-fold higher than in 110 NB and MSC
and did not occur in MS-DON (Figure 3-5a) Growth in MM MSB and MSY was
intermediate
Over the 72 h DON de-epoxidation per cell density was highest in MSB and MSY and
approximately 25 lower in NB and LB and lower by 75 and 87 in 110 NB and MSC
respectively (Figure 3-5b) No DON de-epoxidation was observed in MS-DON in which
DON was used as a carbon source Bacteria grown in MM broth which contains sucrose as a
sole carbon source were unable to de-epoxidize DON No DON de-epoxidation was observed
in SE and PDB grown cultures
Growth of ADS47 under aerobic conditions for 72 h in MSB+50 microgmL DON was
greatest between 25-40degC but no growth occurred at 50degC (Figure 3-6a) DON de-
epoxidation activity was greatest and equivalent between 20-40degC and lower at 10 15 and
45degC temperatures (Figure 3-6b)
ADS47 growth under aerobic conditions for 72 h in MSB+50 microgmL DON was
limited to neutral pHs (ie 60 - 75) (Figure 3-7a) The transformation of DON to dE-DON
occurred at the same pH range and DON de-epoxidation activity was not significantly
different between pH 60 to 75 (Figure 3-7b)
85
b
c
a
b
a
d c
b b
(a)
(b)
b
c
b
a a
d
86
Figure 3-5 Effects of media on bacterial growth and DON de-epoxidation activity (a)
Microbial growth (at OD600nm) on different culture media Media pHs were adjusted to
70plusmn02 (b) Bacterial DON to dE-DON transformation in microgmL by OD600nm in different
media after 72 h The cultures having three replications were incubated for 72 h at 28degC and
DON de-epoxidation was measured The Media NB = nutrient broth 1NB = 110 strength
NB MM = minimal medium MS-DON = six mineral salts + 200 microgmL DON as sole C
source MSB = six mineral salts + 05 bacto peptone MSC = six mineral salts broth + 03
bacto peptone MSY = six mineral salts with 05 yeast extract SE = soil extract PDB =
potato dextrose broth and LB = Luria Bertani Means with the same label are not significantly
different according to Tukeyrsquos HSD test (p lt 005)
87
(a)
(b)
88
Figure 3-6 Effects of temperature on microbial growth and DON de-epoxydation capability
(a) Microbial growth (at OD600nm) at different temperatures after 72 h of incubation (b)
Bacteria DON de-epoxidation in microgmL by OD600nm in different temperatures The most
efficient temperature range (ge40 microgmL DON de-epoxidation) is marked by dot lines The
experiment was conducted in MSB medium at pH 70plusmn02 with three replications
89
(a)
(b)
90
Figure 3-7 Influence of pH on microbial growth and DON de-epoxydation (a) Microbial
growth (OD600nm) in MSB at different pH (b) Bacterial DON to de-epoxy-DON
transformation in microgmL by OD600nm in different pH Dot lines indicated the efficient de-
epoxidation (ge40 microgmL DON conversion) pH range Experiment was conducted at 28degC
with three replications
91
Antibiotic
Treatment of ADS47 at 72 h in MSB+50 microgmL DON with sodium azide (SA) at
0001 and 001 reduced DON de-epoxidation activity to 5 (25 microgmL) and 0
respectively The control with no SA added was able to de-epoxidize DON completely
(100) (Figure 3-8)
Figure 3-8 Effects of sodium azide (SA) on bacterial DON de-epoxidation activity
Microbial culture in MSB+50 microgmL DON was treated with 001 (SA-1) and 0001 (SA-
2) sodium azide In control (C) no SA was added The experiments with three replications
were incubated for 72 h at 28degC and DON de-epoxidation was measured by LC-UV-MS
method
0
20
40
60
80
100
120
SA1 SA2 C
D
ON
de
-ep
oxyd
ize
d
92
344 Localization of bacterial DON de-epoxidation
A cell lysate prepared from ADS47 grown for 48 h in MSB transformed 20 (10
microgmL) of DON added at 50 microgmL to dE-DON within 72 h (Figure 3-9) In contrast a cell-
free culture filtrate did not show DON de-epoxidation activity
Figure 3-9 Determination of DON to de-epoxy DON transformation by bacterial culture
filtrate and cell extract The treatments having three replications were spiked with 50 microgmL
DON and incubated at 28degC for 72 h In control strain ADS47 was incubated in MSB broth
with 50 microgmL DON DON de-epoxidation activity in percentage was examined
0
20
40
60
80
100
120
Filtrate Extract Control
D
ON
de
-ep
oxyd
ize
d
93
345 Bacterial detoxification (de-epoxidtaion and de-acetylation) of other trichothecene
mycotoxins
Aerobic cultures of ADS47 grown for 72 h in NB+100 microgmL HT-2 converted 74 of
the toxin to de-epoxy HT-2 toxin and 26 to a de-acetylated product (Figure 3-10 and Table
3-1)
In a culture treated with T2-toxin 46 was converted to de-epoxy T-2 16 was
converted to an unknown compound and 38 was not converted In ADS47 culture with
diacetoxyscirpenol (DAS) 61 was transformed into de-epoxy DAS and 39 was de-
acetylated DAS (39) Neosolaniol (NEO) was transformed to de-epoxy NEO (55) and
de-acetylated NEO (45) T2-triol was transformed in to de-epoxy T2-triol (47) and de-
acetylated T2-triol (53)
Aerobic cultures of ADS47 were also grown for 72 h in NB+100 microgmL of one of five
type-B trichothecenes For nivalenol (NIV) 92 was de-epoxydized while the remaining 8
NIV was transformed to DON 3-acetyl-deoxynivalenol (3-ADON) was transformed into
three products de-epoxy NIV (5) de-epoxy DON (54) and DON (41) For 15-
acetyldeoxynivalenol (15-ADON) 8 was transformed to de-epoxy 15-ADON and 92 to
de-epoxy DON (dE-DON) FusarenonX (FUS) had 95 transformed into de-epoxy FUS
while 97 of verrucarol (VER) was transformed to de-epoxy VER The remaining 5 FUS
and 3 VER were not transformed
94
Figure 3-10 Determination of microbial de-epoxidation and de-acetylation of HT-2 toxin by
LC-UV-MS Bacterial culture in NB was added with 100 microgmL HT-2 toxin Bacterial HT-2
toxin transformation ability was examined after 72 h of incubation at 28degC The upper panel
illustrated mass spectrometry chromatograph of the de-acetylated (red peak) and de-
epoxydized (blue peak) products Arrows indicated the targeted acetyl (red) and epoxy (blue)
functional groups of HT-toxin The middle and lower panels showed the mass spectra of the
de-acetylated (dA) and de-epoxidized (dE) compounds Arrows on the upper panel indicated
the targeted acetyl (red) and epoxy (blue) functional groups of HT-2 toxin Red and blue
arrows on the middle and lower panels pointed at the degradation sites of HT-2 toxin
95
Table 3-1 Microbial transformation of type-A and type-B tricthothecene mycotoxins after 72
h of incubation under aerobic conditions at room temperature DAS=diacetoxyscirpenol
DON=deoxynivalenol FUS=FusarenonX 3AON=3-acetyl-deoxynivalenol and
15ADON=15-acetyl-deoxynivalenol
1 Convertion of trichothecene mycotoxins to either DON or an unknown compound
Trichothecene
mycotoxin
Untransformed
trichothecenes ()
Transformed products of
trichothecenes ()
De-
epoxy
De-
acetylate
Other1
Type-A
HT-2 toxin 0 74 26 -
T-2 toxin 38 46 - 16
(unknown)
DAS 0 61 39 -
Neosolanol 0 55 45 -
T2-triol 0 47 53 -
Type-B
DON 0 100 - -
Nivalenol 0 92 - 8 (DON)
3-ADON 0 59 41 (DON)
15-ADON 0 100 - -
FUS 5 95 - -
Varrucarol 3 97 - -
96
35 Discussion
The ADS47 strain of C freundii was isolated in the present study by a prolonged
incubation of a mixed microbial culture with a high concentration DON since the bacterium
efficiently de-epoxidized (detoxified) DON and number of other trichothecene mycotoxins
under aerobic and anaerobic conditions at moderate temperatures and neutral pH The total
procedures for enrichment and isolation of the strain ADS47 is depicted in supplemental
Figure S3-1 The reductive de-epoxidation activity may be due to a cytoplasmic enzyme
which would be in agreement with a previous study that showed de-epoxidation of
epoxyalkanes was catalyzed by a bacterial cytoplasmic flavoprotein (Westphal et al 1998)
The ability of the currently isolated strain of C freundii to biodegrade environmental
pollutants such as aminoaromatic acids and heavy metals (copper) is also likely mediated by
cytoplasmic enzymes since these compounds need to be taken up by the cells before they are
metabolized (Savelieva et al 2004 Sharma and Fulekar 2009) In contrast oxidative
transformation of DON to 3-keto DON by microbial extracellular enzyme has been reported
(Shima et al 1997)
Isolation of a trichothecene de-epoxidizing microorganism from a mixed microbial
community is very challenging Although trichothecenes are toxic to eukaryotes prokaryotes
appear to be resistant to these mycotoxins (He et al 1992 Rocha et al 2005) The present
study showed that the bacteria grew well in media with high concentrations of DON (500
microgmL) Bacterial de-epoxidation of DON involves the removal of an oxygen atom from the
functional epoxy ring of the tricyclic DON structure (Yoshizawa et al 1983) Such co-
metabolic degradation does not provide the bacteria with any nutrients from the reacting
compounds (Horvath 1972 Fedorak and Grabic-Galic 1991) This is supported by the results
97
of the current study which showed that strain ADS47 was unable to grow in media with DON
as a sole carbon source and increasing the concentration of DON did not increase ADS47
growth in MSB Hence DON could not be used as sole nutrient source in the mixed culture
enrichment process
Many previous attempts to obtain DON de-epoxidizing microbes have failed
particularly under aerobic conditions because media or laboratory conditions were
inappropriate to support growth or initially active cultures lost de-epoxidizing function after
serial transfer (Karlovsky 2011)
Media composition appears to be a crucial factor for microbial trichothecene de-
epoxidation activity (Voumllkl et al 2004) Effective biotransformation of trichothecene
mycotoxins by the fish digesta microbial culture C133 occurred in full media (FM) but a low
level of de-epoxidation occurred in media having a single nitrogen source and no
transformation activity was observed in the common bacterial medium LB (Guan et al
2009) Zhou et al (2010) also showed that chitin de-acetylation by soil bacterial strains
occurred more rapidly in yeast extract compared to common bacterial media Therefore a
screening strategy for microbial DON de-epoxidation ability should use different media in
parallel as was done in this study
This study observed efficient de-epoxidation in common bacterial media (NB and LB)
as well as in mineral salts medium containing yeast extract (MSY) or 05 Bacto Peptone
(MSB) However reduction of Bacto-Peptone to 03 (MSC) significantly reduced the
bacterial de-epoxidation activity suggesting that the activity is sensitive to the availability of
protein and amino acids The study found that medium containing sucrose as carbon source
(MM) and a commonly used medium PDB blocked microbial DON de-epoxidation even
98
though bacteria grew in those growth media These suggest that media containing high sugar
contents had inhibitory effects on microbial de-epoxidation enzymatic activity Previously
sugars have been found to have negative effects on the production andor activity of bacterial
enzymes (Saier and Roseman 1971)
Losses of microbial functions under laboratory conditions limited researchers to obtain
economically important microbial genetic sources The soil borne Curtobacterium sp strain
114-2 efficiently de-epoxydized nivalenol but unfortunately the strain lost its activity in
latter generations (Ueno et al 1983) Instability of de-epoxidation activity of ADS47
observed in cultures grown on agar media without DON resulted in the loss of DON de-
epoxidation activity by more than 80 of the colonies after one generation However strain
ADS47 retained de-epoxidation activity after ten subcultures in MSB (data not shown)
The non-specific de-epoxidation of DON made it impossible to use DON as a sole
substrate C source for selective isolation of ADS47 Thus a number of techniques were
utilized to select ADS47 and suppress other bacteria including the use of a variety of
antibiotics and culture at elevated temperatures (Islam et al 2012) Selection of the isolate
ADS47 required repeated subculturing and prolonged exposure to media containing 200
microgmL DON After three months of selection with DON culture that was obtained had a
DON de-epoxidation activity that was up to 30-fold higher than the mixed culture As growth
of ADS47 was not affected by 200 microgmL DON the prolonged incubation may have reduced
populations of other bacteria that are more sensitive to DON Enrichment and isolation of a
DON to 3-keto DON transforming bacterial strain E3-39 by prolonged incubation of a soil
microbial culture with 200 microgmL DON was reported by Shima et al (1997)
99
This study demonstrated that microbial DON de-epoxidation occurred mainly at the
stationary growth phase A DON degrading strain of Citrobacter may have gained energy
from DON by co-metabolic cleavage of the epoxy ring under less nutrient conditions
Microbial use of energy from co-metabolic reaction has been suggested by Chiu et al (2005)
The rate of de-epoxidation activity increased when ADS47 grew at higher concentrations of
DON Taken together a prolonged exposure to high concentration DON may have resulted in
non-DON metabolizing microbial populations declining by DON toxicity while facilitated
the population of DON metabolizing bacterium by providing energy
DON is a small molecule and is simply absorbed by cells (Sergent et al 2006)
Microbial DON de-epoxidation occurred in intact and lysed bacterial cells In addition
treating the ADS47 culture with SA which blocks gram negative bacterial electron transport
and activity inhibited DON de-epoxidation activity (Lichstein and Malcolm 1943 Erecinska
and Wilson 1981) implying that an active electron transport system required for microbial de-
epoxidation reaction (He et al 1992) The data suggest that a cytoplasmic epoxy reductase is
involved in the DON de-epoxidation reaction in the ADS47 microbial strain that was isolated
in the present study
Since agricultural commodities are often contaminated with multiple fungal
mycotoxins the ability of bacteria like ADS47 to degrade mycotoxins by several
mechanisms may be an advantage De-acetylation might be another mechanism of microbial
mycotoxin detoxification since toxicity of trichothecenes also relies on the acetyl functional
groups (Desjardins 2006) Strain ADS47 showed efficient de-epoxidation and de-acetylation
of several type-A and type-B trichothecene mycotoxins that are commonly detected in cereals
in association with DON In conclusion the soil-borne gram-negative bacterium C freundii
100
strain ADS47 is capable of detoxifying trichothecene mycotoxins by both de-epoxidation and
de-acetylation under aerobic and anaerobic conditions neutral pH and a range of
temperatures ADS47 has great potential to detoxify DON contaminated animal feed
Furthermore isolation of novel bacterial de-epoxidation genes might be used to generate
transgenic plants that can detoxify DON in contaminated seed Alternatively contaminated
seed might be treated with manufacture de-epoxidation enzyme(s) These applications can
improve the quality and safety of agricultural commodities that are prone to trichothecene
mycotoxin contamination
36 Acknowledgements
Thank to the Natural Sciences and Engineering Research Council of Canada (NSERC) for a
scholarship to Rafiqul Islam The work was funded by the Agriculture and Agri-Food Canada
Binational Agricultural Research and Development Fund (AAFCBARD) project We
appreciate Honghui Zhu for technical assistance
101
Supplemental Figure S3-1 Flow chart depicts the procedures of enrichment and isolation of
DON de-epoxidation bacterial strain ADS47 Steps 1 to 4 describe the optimization of
microbial DON to dE-DON transformation activity and removal of undesired microbial
species Step 5 to 7 shows the culture enrichment by incubation with high concentration DON
and isolation of the single strain ADS47
Step 1 Screening of soil samples
Step 2 Optimization of cultural conditions
Step 3 Elimination of undesired microbial species
Step 4 Removal of untargeted microbial species
Step 5 High concentration DON enrichment
Step 6 Separation of bacterial cells and dilution plating
Step 7 Single colony analysis
Active culture was sequentially treated with different mode of actions antibiotics heating cultures at 50degC and subculturing on agar media
Colonies were tested for their DON de-epoxidation ability Purified the single colony via several times subculturing on agar medium
Optimized the culture conditions ie growth substrates temperatures pH and oxygen requirements
Obtained DON de-epoxidation capable microbial culture obtained by analyzing soil samples collected from cereal fields
Serially diluted active culture was grown on agar media Collected washed colonies having DON de-epoxidation activity
The partially enriched culture that retained DON de-epoxidation activity was incubated with 200 microgmL DON in MSB media with continuous shaking at 220 microgmL for 7 d The procedure was repeated 12 times
The enriched culture was serially diluted and treated dilutes with 05 Tween 20 After vigorously vortex for ten min aliquot cultures were spread on different agar media plates to produce single colonies
102
CHAPTER 4
40 CONSTRUCTION AND FUNCTIONAL SCREENING OF GENOMIC DNA
LIBRARY FOR ISOLATION OF DON DE-EPOXYDIZING GENE(S) FROM
CITROBACTER FREUNDII STRAIN ADS47
41 Abstract
Enzymatic activity-based screening of a genomic DNA library is a robust strategy for
isolating novel microbial genes A genomic DNA library of a DON detoxifying bacterium
(Citrobacter freundii strain ADS47) was constructed using a copy control Fosmid vector
(pCC1FOSTM
) and E coli EPI300-T1R as host strain A total of 1250 E coli clones containing
large DNA inserts (~40 kb) were generated which resulted in about 10-fold theoretical
coverage of the ADS47 genome The efficiency of E coli transformation with the
recombinant Fosmid vector was examined by restriction digestion of Fosmids extracted from
15 randomly selected library clones The fragmentation patterns showed that all of the clones
contained an 81 kb Fosmid backbone and ~40 kb of insert DNA except one that had a small
insert This suggests that a DNA library was prepared with expected DNA inserts The de-
epoxidation ability of the library was analysed by incubating clones in Luria Bertani (LB)
broth medium supplemented with 20 microgmL DON with induction (by addition of L-arabinose)
or without induction None of the 1000 clones showed DON de-epoxidation activity The
potential DON de-epoxidation gene(s) of ADS47 may not have been expressed in E coli or
might have been expressed but lacked additional factors required for DON de-epoxidation
103
Keywords DNA library fosmid vector functional library screening DON de-epoxidation
activity
42 Introduction
Microorganisms contain numerous valuable genes and biocatalysts which may have
significant roles in agricultural productivity (Jube and Borthakur 2007) Various techniques
have been used to isolate novel genes from different organisms including transposon
mutagenesis (Frey et al 1998 Tsuge et al 2001) gene knockout (Winzeler et al 1999)
whole genome sequencing (Krause et al 2006) comparative genomics (Koonin et al 2001)
and suppression subtractive hybridization (Dai et al 2010)
The genomic DNA library preparation and activity-based screening of the library is a
relatively new technique which has been efficiently used for isolation of microbial novel
enzymes and compounds such as lipases proteases oxidoreductases antibiotic resistance
genes membrane proteins and antibiotics (Henne et al 2000 Gupta et al 2002 Knietsch et
al 2003 Gabor 2004 Suenaga et al 2007 van Elsas et al 2008) The strength of the
technique is that genes from microbial sources could be obtained without prior sequence
information and not requiring culturing the organisms under laboratory conditions Using this
technique an unknown gene or a set of genes could be obtained by determining particular
phenotypes of the library clones Eventually the gene of interest from the positive clones is
isolated by using a forward (eg mutation) or reverse genetics method (eg subcloning the
positive clones into a microbial expression vector) followed by functional analysis of the
subclones
104
Although this strategy has been used effectively for isolating novel microbial genes
several factors need to be considered in order for the method to be successful These factors
include DNA quality vector type (eg plasmid cosmid fosmid bacterial artificial
chromosome etc) vector copy number and host strains (Daniel 2005 Singh et al 2009) The
Fosmid vector-based technique has been used successfully to clone bacterial genes including
xenobiotics degradation genes antifungal gene clusters and antibiotic resistance genes from
soil metagenomes (Hall 2004 Suenaga et al 2007 Chung et al 2008) This study reports the
construction and activity-based screening of a Fosmid DNA library in an E coli host with the
aim of isolating a novel DON de-epoxidation gene from Citrobacter freundii strain ADS47
(Chapter 3)
43 Materials and methods
431 Genomic DNA library construction
The genomic DNA library was prepared using a Copy Control Fosmid Library
Production Kit (Epicentre Biotechnologies Madison Wisconsin USA) following the
manufacturerrsquos instructions as depicted in Figure 4-1
105
Figure 4-1 Schematic overview of the construction and functional screening of a Fosmid
DNA library
106
Citrobacter freundii strain ADS47 was grown in NB medium at 28degC with continuous
shaking at 220 rpm The high molecular weight bacterial genomic DNA was extracted from
the overnight culture using the Gentra Puregene YeastBacteria Kit (Qiagen Inc Mississauga
Ontario Canada) The purity of the DNA was confirmed by determining OD260280 at ~190 by
a NanoDrop spectrophotometer A 5 microL sample having ~25 microg high molecular weight DNA
was sheared by passing through a 200-microL pipette tip and achieved a sufficient amount of
DNA about 40 kb in size Blunt ended 5rsquo-phosphorylated insert DNA was generated by
thoroughly mixing it with End-Repair Enzyme reagents on ice The 20 microL mixture was
incubated for 45 min at room temperature followed by incubation at 70degC for 10 min to
inactivate the End-Repair Enzyme activity
The treated DNA was size fractionated by a Low Melting Point-Pulsed Field Gel
Electrophoresis (LMP-PFGE) on the clamped homogeneous electric fields (CHEF-DR III)
system (Bio-RAD Laboratories) using 1 UltraPure Low Melting Point Agarose (Invitrogen)
The PFGE was conditioned according to manufacturerrsquos instructions as follows initial switch
time-42 s final switch time-761 s run time 16 h 50 min angle-120deg gradient- 6 Vcm
temperature-14degC and running buffer-05 TBE DNA size markers for the 42 kb Fosmid
control DNA (Epicentre Biotechnologies) and low range Pulse Field Gel (PFG) markers (New
England Biolabs Inc) were loaded adjacent to the DNA sample slots After electrophoresis a
portion of the gel that contained DNA size markers was cut off The gel was stained in SYBR
Safe DNA gel stain (Invitrogen) for 30 min visualized and marked on the 23 42 and 485 kb
marker bands A slice of PFGE gel containing 40 plusmn 10 kb size DNA was cut off with a sterile
razor blade by comparing the marker sizes DNA from the gel was recovered by digesting it
with GELase enzyme ( 1 U100 microL melted agarose) in 1X GELase Buffer followed by a
107
sodium acetate-ethanol precipitation The DNA with the desired size was stored at 4degC for
next dayrsquos ligation to a cloning-ready Fosmid vector (pCC1FOSTM
)
Ligation was performed in a 10 microL reaction mixture by thoroughly mixing 10X Fast-
Link Ligation Buffer 10mM ATP 025 microg of ~40 kb insert DNA 05 microg Fosmid Copy
Control Vector (pCC1FOS) (Figure 4-2) and 2 U Fast-Link DNA lipase The mixture was
incubated for 4 h at room temperature preceded by inactivating the ligation reaction by
heating at 70degC for 15 min The recombinant Fosmid vector was stored at -20degC for later use
Ten microL of the ligated DNA was thawed on ice and transferred to a pre-thawed
MaxPlax Lambda Packaging Extract tube containing 25 microL extract By pipetting several
times the mixture was incubated at 30degC for lamda packaging After 2 h of packaging an
additional 25 microL packaging extract was added into the mixture and incubated for a further 2 h
at 30degC The packaged phage particles were volumed to 05 mL by adding ~400 microL Phage
Dilution Buffer and 25 microL chloramphenicol
Transformation was performed via transduction by mixing the packaged particles with
the overnight cultured E coli EPI300-T1R cells (1 10 vv ratio) in LB supplemented with 10
mM MgSO4 and 02 Maltose Transformation was completed by incubation at 37degC for 1 h
The transformed E coli cells were spread on an LB agar plate containing 125 microgmL
chloramphenicol The plates were incubated overnight at 37degC Colonies that appeared on the
plates were individually transferred to 96-well microtiter plates having 200 microL LB medium
supplemented with 10 glycerol and 125 microgmL chloramphenicol The microtiter plates
were incubated at 37degC for 24 h with continuous shaking at 180 rpm The prepared library
clones were stored at -80degC for further analysis and functional screening
108
109
Figure 4-2 Genetic map of copy controlled Fosmid vector (pCC1FOSTM
) The 813 kb
Fosmid vector backbone shows a multiple cloning site (MCS) chloramphenicol resistance
selectable marker (CmR) replication initiation fragment (redF) inducible high copy number
origin (oriV) replication initiation control (repE) and F-factor-based partitioning genes (ParA
ParB ParC) The vector contains a cos site for lamda packaging (not shown) and lacZ gene
for bluewhite colony selection The vector map was provided by the Epicentre
Biotechnologies (httpwwwepibiocomitemaspid=385)
110
432 Confirmation of library construction
The quality of the DNA library prepared was determined by examining 15 E coli
clones randomly selected A 15 mL LB broth supplemented with 125 microgmL
chloramphenicol was inoculated by each of the transformed clones To obtain higher Fosmid
DNA yield 1 microL autoinduction solution (L-arabinose) to increase the copy number of the
Fosmid vector (Epicentre) was added to the cultures After an overnight incubation of the
cultures at 37degC with continuous shaking at 220 rpm Fosmid DNA was extracted using a
Qiagen Large-Construct Kit followed by restriction digestion with Not I enzyme A 20 microL
digestion mixture containing 05 microg Fosmid DNA 1X digestion buffer and 05 U Not I was
incubated at 37degC for 15 h The partially digested DNA and a 42 kb DNA size marker
(Fosmid control DNA) were ran on 08 agrose gel for 15 h at 100 V The linear Fosmid
vector backbone (813 kb) and insert DNA fragments were visualized on the gel and the
image was recorded
433 Functional screening of the clones
Eppendorf tubes (15 mL) containing 500 microL LB supplemented with 125 microgmL
chloramphenicol were inoculated with a single clone (400 clones tested) or a combination of
12 library clones (600 tested) from microtiter plates stored at -80degC One half of the cultures
in each group (single or combined clones) were induced to increase the Fosmid copy number
by adding a 1 microL (500X) autoinduction solution (contained L-arabinose) to the culture tubes
L-arabinose induces the expression of a mutant trfA gene of the E coli host strain which
leads to the initiation of Fosmid replication from a low-copy to a high-copy number origin of
replication (oriV) The cultures were spiked with 20 microgmL DON and incubated at 37degC for
111
72 h with continuous shaking at 180 rpm The DON to de-epoxy DON transformation ability
of the clone cultures was examined by a liquid chromatography-ultraviolet-mass spectrometry
(LC-UV-MS) method as described by Islam et al (2012)
112
44 Results
441 Production of the DNA library
High molecular weight intact (non-degraded) and good quality (OD600280 ~190)
genomic DNA was obtained from the DON de-epoxidizing bacterial strain ADS47 (data not
shown) Pipetting of the DNA extract resulted in an adequate amount of insert DNA (gt025
microg) of approximately 40 kb size as showed on a LMP-PFGE (Figure 4-3)
A DNA library in E coli host was constructed using LMP-PFGE purified DNA The
library was comprised of 1250 E coli transformants which were selected on LB agar plates
supplemented with chloramphenicol (Figure 4-4)
113
Figure 4-3 Preparation of a 40 kb size insert DNA from a DON de-epoxidizing bacterium
Genomic DNA was extracted from the overnight culture of a DON de-epoxidizing
Citrobacter freundii strain ADS47 After fragmentation the DNA was size fractionated by
Low Melting Point- Pulse Field Gel Electrophoresis (LMP-PFGE) with markers for the PFGE
(M1) and 42 kb Fosmid control DNA (M2) S1 S2 S3= different DNA samples extracted
from bacterial strain ADS47 The red dots area indicates the section of the gel containing
genomic DNA of the expected size (~40 kb)
114
Figure 4-4 Production of the genomic DNA library in E coli EPI300-T1 host strain
Approximately 40 kb long blunt ended genomic DNA from a DON de-epoxidizing bacterial
strain ADS47 was ligated to a Copy Control Fosmid vector pCC1FOS The construct was
packaged into lamda bacteriophage followed by a transduction-mediated transformation of E
coli host cells The clones which appeared on the LB agar plate supplemented with 125
microgmL chloramphenicol are expected to be transformed by the recombinant Fosmid vector
containing insert DNA
115
442 Conformation of the library construction
All of the clones tested acquired the Fosmid vector Out of 15 clones examined 14
received the recombinant Fosmid vector that contain the expected size insert DNA as
determined by an 81 kb vector backbone and at least one high molecular weight insert DNA
band (~40 kb) on agarose gel (Figure 4-5) In contrast one clone (lane 4) showed an 81 kb
vector backbone and a low molecular weight band which is also a fragment of the Fosmid
vector The result indicated that approximately 94 of the library clones were transformed
with the recombinant vector
443 Activity-based screening of the library
Out of 500 single copy number clones (200 individual and 300 combined clones)
none showed DON to de-epoxy DON transformation activity (Table 4-1) An additional 500
high copy number clones (200 single and 300 combined clones) were unable to de-epoxidize
DON as well
116
Figure 4-5 Confirmation of the Fosmid DNA library production in E coli Fosmid DNA
collected from 15 clones was digested with Not I and run on 08 agarose gel at 100 V for 15
h The presence of the vector backbone (81 kb) and the insert DNA fragment (~40 kb) in the
clones was determined by comparing a 42 kb molecular size marker (M) Lanes (1-15) denote
the recombinant Fosmid DNA extracted from 15 E coli clones
117
Table 4-1 Analysis of Fosmid DNA library clones for deoxynivalenol (DON) de-epoxidation
capability
Type of clones Analysis of
singlecombined
clones
Number of
clones
examined
Number of clones
showed DON de-
epoxidation activity
Without induction
(single copy number clones)
Singly 200 0
Combination of 12 300 0
With induction (multi-copy
number clones)
Singly 200 0
Combination of 12 300 0
De-epoxidation activity of the clones was examined alone or in combination of 12 clones
without or with induction by adding L-arabinose The clones grown in LB supplemented with
125 microgmL and 20 microgmL DON were analysed for DON to de-epoxy DON transformation
ability by a liquid chromatography-ultra violet- mass spectrum (LC-UV-MS)
118
45 Discussion
While a Fosmid DNA library with the expected genome coverage of a DON de-
epoxidizing bacterial genome was produced the function-derived screening of the library
clones did not identify any clones that showed DON de-epoxidation enzymatic activity This
is unlikely due to lack of coverage of the bacterial genome as 1000 clones analyzed in the
study should have provided eight fold coverage of the genome Therefore it was likely due to
a lack of expression of the enzyme(s) for DON de-epoxidizing activity under the conditions
used
Thus far there is only one report of heterologous expression of bacterial epoxide
reductase gene in another bacterium A high level of heterologous expression of a
Synechococcus sp vitamin K epoxide reductase gene was achieved in E coli with codon
optimization and placing the gene under the control of a prokaryotic T7 promoter in an
expression vector (Li et al 2010) Therefore heterologous expression of bacterial epoxide
reducatse gene using its own promoter has not yet been demonstrated The success of the
method used in this chapter relies on an intact copy of the gene or operon being cloned the
selection of an appropriate cloning vector a host organism that can recognize the cloned C
fruendii promoter and a suitable library screening method for DON degradation by E coli
(Mocali and Benedetti 2010)
The unbiased cloning efficiency of the Fosmid vector its ability to contain large insert
DNA high stability in E coli as single copy number and autoinduction to increase copy
number in host cells made it appear to be a suitable method for the present work (Kim et al
1992 Lee et al 2004) Library production with large insertions is a major advantage as gene
cluster involved in particular pathway could be trapped into single clone and their functions
119
could be analysed The pathway of microbial DON de-epoxidation activity and the genes
encoding the activity are not known Therefore it was reasonable to prepare a library with
large inserts to ensure the possibility of cloning single as well as whole pathway genes of
interest for functional screening A further advantage of the method is that Fosmid clones are
inducible which allows detecting poorly expressed genes in a heterologous host by inducing
high copy numbers (Daniel 2005)
One of the limitations of the method is that the functions of the target genes rely
mostly on the host organismrsquos transcription and translation machineries Therefore selection
of the host organism is critical for activity-based screening of library clones in heterologous
hosts A number of host strains including E coli Streptomyces lividans Rhizobium
leguminosarum and Pseudomonas aeruginosa have been used for functional screening
activities from diverse organisms (Daniel 2005) The present work attempted to isolate C
freundii strain ADS47 genes encoding DON de-epoxidation enzymes Both strain ADS47 and
E coli belong to gamma proteobacteria which suggest that E coli would be an appropriate
host organism for heterologous expression of ADS47 genes Poor expression of heterologous
genes could be another limitation of the method which may be overcome by overexpression
to increase the catalytic activity of less expressed genes However clones with DON de-
epoxidation activity were not identified in this study even the Fosmid copy number was
increased by induction
In general a large number of clones must be analysed to obtain the gene of interest
using an activity-based screening technique (Lee et al 2004 van Elsas et al 2008) In this
study the constructed DNA library comprised of 1160 clones having approximately ~40 kb
insert DNA suggesting that the prepared library achieved ~10 fold coverage of the ADS47
120
genome (48 Mb) which ensured gt99 probability of the presence of all DNA sequence in
the library In the produced library three clones (lanes 3 10 and 12) showed one intact band
greater than 42 kb in size on the agarose gel It is presumed that the Fosmid DNA of the three
samples was not digested because of insufficient amount of restriction enzyme added to the
digestion mixture About half of the samples demonstrated multiple bands suggesting that the
insert DNA might have contained multiple Not I restriction sites
However not every heterologous gene may necessarily express or function in the E
coli host (Gabor et al 2004) The functionality of the gene requires expression of the genes
and proper folding of the protein which may be influenced by environmental conditions (eg
substrate temperature and pH) and genetic factors In this study library clones were screened
in growth conditions at which strain ADS47 performed the best de-epoxidation activity The
expression of genes may be controlled by the promoter suitable ribosome binding site and
transndashacting factors (eg special transcription factors inducers chaperones cofactors proper
modifying enzymes or an appropriate secretion system) (Gabor et al 2004) Those factors
have to be supplied by the cloned DNA fragments or host cells to allow any heterologous
gene to be functional Since DON de-epoxidation genes of strain ADS47 evolved in a distinct
environmental condition the expression and activity of the gene(s) might have required
specific factorsregulators that were absent in E coli Even though the Fosmid based library
screening technique facilitated isolation of many microbial genes clones having de-
epoxidation activity were not identified using such technique in this study
121
46 Acknowledgements
We appreciate the Agriculture and Agri-Food CanadaBinational Agricultural
Research and Development Fund (AAFCBARD) project for providing the financial support
for this work The authors thank Honghui Zhu for analyzing numerous samples by liquid
chromatography ultraviolet mass spectrum method
122
CHAPTER 5
50 GENOME SEQUENCING CHARACTERIZATION AND COMPARATIVE
ANALYSIS OF CITROBACTER FREUNDII STRAIN ADS47
51 Abstract
The genome of a trichothecene mycotoxin detoxifying C freundii strain ADS47
originating from soil was sequenced and analyzed Strain ADS47 contains a circular
chromosome of 4878242 bp that encodes 4610 predicted protein coding sequences (CDSs)
with an average gene size of 893 bp Rapid Annotation Subsystems Technology (RAST)
analysis assigned functions to 2821 (62) of the CDSs while 1781 (38) CDSs were
assigned as uncharacterized or hypothetical proteins Comparative bioinformatics analyses
were done for strain ADS47 C freundii Ballerup 785139 Citrobacter sp 30_2 C youngae
ATCC 29220 C koseri ATCC BAA-895 and C rodentium ICC168 There were a significant
number of common orthologous genes (2821) were found in the six Citrobacter genomes
implying a common evolutionary relationship ADS47 is most closely related to the
opportunistic human pathogen C freundii Ballerup The human and mouse pathogenic
species C rodentium and C koseri are the most distantly related to ADS47 and non-
humananimal pathogens C youngae and Citrobacter sp 30_2 were closely related ADS47
C koseri C youngae and Citrobacter sp 30_2 lacked the clinically important locus for
enterocyte effacement (LEE)-encoded type-III secretion systems which are present in C
rodentium and C freundii Ballerup Strain ADS47 had 270 unique CDSs many of which are
located in the large unique genomic regions potentially indicating horizontal gene transfer
123
Among the unique CDSs nine were reductasesoxidoreductases and a de-acetylase that may
be involved in trichothecene mycotoxin de-epoxidation and de-acetylation reactions
Keywords Citrobacter freundii comparative genomics genome sequencing unique genes
reductases
52 Introduction
According to the Food and Agriculture Organization (FAO) at least 25 of the
worldrsquos food crops are contaminated with mycotoxins causing serious negative impacts on
livestock and international trade (Boutrif and Canet 1998 Desjardins 2006 Binder et al
2007) Trichothecenes (TCs) produced by a variety of toxigenic Fusarium species are
probably the most prevalent food and feed contaminating mycotoxins It is estimated that
infections by the fungus caused more than $1 billion in crop yield losses in USA and Canada
alone (Windels 2000 Schaafsma et al 2002 Yazar and Omurtag 2008) TC mycotoxins can
have adverse affects on human health including organ toxicity mutagenicity carcinogenicity
neurotoxicity modulation of the immune system negative reproductive system affects and
growth retardation (DrsquoMello et al 1999 Pestka et al 2007) A strain of Citrobacter freundii
ADS47 was isolated from soil in southern Ontario that is capable of detoxifying (ie de-
epoxidation and de-acetylation) several TC mycotoxins including deoxynivalenol nivalenol
and T2-toxin (Chapters 2-3)
Citrobacter freundii is a gram negative motile rod shaped facultatively anaerobic
bacterium belonging to the family of Enterobacteriaceae (Wang et al 2000 Whalen et al
2007) The opportunistic human pathogen is responsible for a number of infections including
124
nosocomial infections of respiratory tract urinay tract blood and causes neonatal meningitis
(Badger et al 1999 Wang et al 2000) C freundii exists almost ubiquitously in soil water
wastewater feces food and the human intestine Biodegradations of tannery effluents copper
aromatic amino acids by C freundii strains isolated from water mine tailings and soil
respectively have been previously reported (Kumar et al 1999 Savelieva et al 2004 Sharma
and Fulekar 2009) Because of their biochemical capabilities bacteria have played important
roles in industrial biotechnology including de-epoxidation and de-acetylation activities
(Demain and Adrio 2008 Plank et al 2009 Zhao et al 2010) However the identification of
genes for de-epoxidation activity against trichothecene mycotoxins is limited (Boutigny et al
2008)
The advent of molecular techniques such as next generation sequencing technologies
and the related bioinformatics tools has enabled the discovery of novel genes and the
enzymes they encode in pathogenic and beneficial bacteria (van Lanen and Shen 2006
Ansorge 2009 Zhao 2011) In the present study the genome of mycotoxin-detoxifying strain
C freundii ADS47 was sequenced and subjected to comparative genomic analyses with other
Citrobacter species to identify putative mycotoxin-detoxifying genes
125
53 Materials and methods
531 Sequencing quality control and assemble of ADS47 genome
Genomic DNA of the strain ADS47 was extracted using Gentra Puregene
YeastBacteria Kit (Qiagen Inc Mississauga Ontario Canada) and purity of the DNA was
confirmed by determining OD 260280 at ~190 using a Nanodrop spectrophotometer
The genomic DNA was sent to be sequenced by the Illumina HiSeq 2000 sequencing
analyzer at Beijing Genomics Institute (BGI) Hong Kong
(httpwwwbgisequencecomhomeservicessequencing-servicesde-novo-sequencing) The
sequencing workflow involved short read library preparation cluster generation sequencing
and data analysis Briefly 3-5 ug of bacterial gDNA was fragmented to produce a 500 bp
insert library by preparative electrophoresis through a 08 agarose gel The fragmented
DNAs were blunt-ended by treating with a mixture of enzymes (T4 DNA polymerase klenow
fragment) and subsequently adapters were ligated to the blunt ends Each of the DNA
fragments was amplified to a cluster which was performed in the flow cell by template
hybridization and bridge amplification Millions of clusters were then sequenced by
sequencing-by-synthesis technology leading to base calls The raw data was filtered to remove
low abundant sequences from the assembly according to 15-mer frequencies The low
complexity reads low quality (leQ20) base calls and adapter contaminations (24 bp overlap
between adapter and reads) were eliminated The quality of the sequences was analysed by
GC content and depth correlative analysis (for GC biasness) K-mer analysis was performed
using 15 bp of the short reads for heterologous sequence identification
126
The cleaned short reads (approximately 90 bp) were self-assembled using the Short
Oligonucleotide Analysis Package (SOAP) denovo (httpsoapgenomicsorgcn) (Li et al
2008) The assembly errors were corrected by filling gaps and proofreading single base pairs
using mapping information The assembly produced 441 contigs with an N50 contig size of
29222 bp (the largest contig size 114839 bp) and 80 scaffolds with an N50 scaffold size of
325726 bp (largest scaffold 662452 bp) with an total sequence length of 4878242 bp which
constitutes 33-fold genome coverage The scaffolds were ordered and oriented according to a
reference genome C koseri ATCC BAA-895 the closest completely sequenced species of
the newly sequenced bacterial genome by alignment with the MUMmer program v322
(httpmummersourceforgenet) (Delcher et al 2002) Genome coverage was determined by
mapping the reads to the assembled genome with Bowtie software (Langmead et al 2009) A
preliminary genome annotation was performed to define the origin of replication by
identifying colocalization of multiple genes ie gyrB recF dnaN and dnaA (usually found
near the origin of replication) and comparison with the C koseri reference genome Synteny
analysis between the reference bacterium and assembled scaffolds was performed with the
MUMmer program
532 Gene function annotation and classification
The genome of ADS47 was annotated by Rapid Annotation using Subsystems
Technology v40 (RAST) a server-based genome analysis system (httprastnmpdrorg)
(Aziz et al 2008) A circular genome map was constructed using CGView program
(httpstothardafnsualbertacacgview_server) (Stothard and Wishart 2005)
127
533 Molecular phylogenetic analysis
A robust phylogenetic tree was produced using Composition Vector Tree (CVTree)
web-based program v20 (httpcvtreecbipkueducn) (Qi and Hao 2004) by comparing
whole genome amino acid sequences of strain ADS47 two completely sequenced Citrobacter
genomes [C rodentium ICC168 (NC_013716) and C koseri ATCC BAA-8959 NC_009792)]
and three partially sequenced Citrobacter genomes [C freundii strain Ballerup 785139
(CACD00000000) Citrobacter sp 30-2 (NZ_ACDJ00000000) and C youngae ATCC 29220
(NZ_ABWL00000000)] E coli O157H7 strain EDL933 (NC_002655) was used as an
outgroup species The tree was constructed from the generated pairwise distance matrix data
using Neighbor-Joining Program in PHYLIP v369 package (Saitou and Nei 1987 Felsenstein
1989)
534 Comparative genomic analysis
Genome-wide comparative analyses between C freundii ADS47 C freundii strain
Ballerup 785139 Citrobacter sp 30-2 C youngae ATCC 29220 C Koseri ATCC BAA-895
and C rodentium ICC168 was carried out The nucleotide sequence idendities were
determined by pairwise alignments of the ADS47 genome with the five other Citrobacter
genomes using MUMmer software (Delcher et al 2002) and GC content total CDSs genome
coverage by CDSs and number of stable RNAs were determined by RAST program
Orthologous CDS groups among the total set of 29079 CDSs derived from the six
Citrobacter genomes were determined using OrthoMCL software v202
(httporthomclsourceforgenet) based on all-against-all reciprocal BLASTP alignment of
amino acid sequences (Li et al 2003) Genes unique to each strain were identified as those
128
CDSs not included in any orthologous group of the five other Citrobacter genomes BLASTP
comparisons were visualized with Blast Ring Image Generator (BRIG) software v095
(httpsourceforgenetprojectsbrig) (Alikhan et al 2011)
Protein sequences of the five Citrobacter genomes pooled from the GenBank data
bases were compared with ADS47 A circular map was developed by BRIG software that
showed the unique genomic regionsgenes of ADS47 The unique genes were grouped
according to their functional similarities previously identified by RAST software and
genomic regions possibly acquired by horizontal transfer were identified using RAST
software
129
54 Results
541 Assembly and organization of the ADS47 bacterial genome
Scaffolds derived from Citrobacter freundii strain ADS47 were ordered and oriented
by alignment to Citrobacter koseri ATCC BAA-895 the most closely related genome that has
been completely sequenced The genomes of ADS47 and C koseri displayed close synteny
(Figure 5-1)
Figure 5-1 Co-linearity between C freundii strain ADS47 genome and C koseri Scaffolds
derived from ADS47 genome sequences (Y- axis) were aligned and assembled by comparison
with the closest completely sequenced genome C koseri ATCC BAA-875 (X- axis)
130
542 An overview of the ADS47 genome
General features of the C freundii strain ADS47 are presented in Table 5-1 Some
notable features are that strain ADS47 contains 4881122 bp circular chromosome with an
average GC content of 5202 The genome contains 4610 CDSs which comprise 8784 of
the whole genome Annotation by RAST assigned 2821 (62) CDSs to known biological
functions and remaining 1789 (38) genes were classified to unknown or hypothetical
proteins
The GC content in the coding regions is significantly higher (5318) than in the non-
coding intergenic region (4379) which covers 1215 (592781 bp) of the genome The
intergenic region is comprised of 018 repeat sequences (total 8999 bp) 5 (009)
transposons (4806 bp) 025 stable RNA (12091 bp) and the remaining ~115 is
classified as regulatory and other functions The ADS47 genome contains 62 tRNAs 3
rRNAs and 34 copies of sRNAs The average lengths of the tRNAs rRNAs and sRNAs are
785 885 and 13441 bp respectively There are also 42 phageprophage-related genes
identified in the genome
Figure 5-2 shows the distribution of the 26 categorized CDSs rRNA tRNA GC
content and GC skew It appears that ADS47 genome contains equal quantity of genes in
leading and lagging strands and showed occasional clustering (likely operons) of genes in the
two strands The genome contains three rRNAs (two large subunit and a small subunit) and 62
tRNAs which are distributed across the genome A consistent GC content was observed in
the ADS47 genome however 7-10 low GC content (high black color peaks) genomic regions
were found at locations origin of replication 600 2000 3000 3300 3550 3850 4300 and
4800 kbp This is indicative of possible horizontally transferred genes or insertions in those
131
genomic locations GC skew denotes the excess of C over G in certain genomic locations
Positive GC skew (+) (green color) indicates the skew value in the leading strand while
negative skew (-) (purple color) in the lagging strand As in many bacterial genomes the
ADS47 genome map shows the separation of positive and negative GC skew at the origin of
replication (base pair position 1) The GC skew in the leading strand is highly skewed (high
green color peak) indicating that possible horizontal gene transfer occurred into the ADS47
genome
The CDSs ranged from ~100 bp to ~2000 bp and have an average size of 893 bp
(Figure 5-3) The highest numbers of CDS (770) were 600-800 bp long A considerable
number of genes (220) were gt2000 bp in size Some of the biological functions for the
smallest genes (100-200 bp) are sulfur carrier protein methionine sulfoxide reductase
ribonuclease P protein component while the largest genes (gt2000 bp) are ferrous iron
transport protein B translation elongation factor G pyruvate flavodoxin oxidoreductase and
tetrathionate reductase
132
Table 5-1 General features of the C freundii strain ADS47 genome
Feature Chromosome
Genome size 4881122 bp
Overall GC content 5202
Number of genes 4610
Genes with assigned function (number and ) 2821 (62 )
Genes without assigned function (number and ) 1789 (38 )
Total length of protein coding sequences 4285461 bp
Genome coverage by coding sequences () 8784
Average length of coding sequences 893 bp
GC content in the coding region 5318
GC content in the non-coding region 4379
Length and percentage of intergenic region 592781 bp and 1215
Total repeat sequences and genome coverage 8999 bp and 018
Total transposons sequences and genome coverage 4806 and 009
Number average length and genome coverage of tRNA 62 copies 785 bp and 0099
Number average length and genome coverage of rRNA 3 copies 885 bp and 0067
Number average length and genome coverage of sRNA 34 copies 13441 bp and 0093
133
134
Figure 5-2 Circular genome map of C freundii strain ADS47 Map was created with the
CGView software From outside to inside the first circle indicates the genomic position in kb
Different functional categories of predicted coding sequences (CDS) are distinguished by
colours with clockwise transcribed CDSs in the second circle and anticlockwise transcribed
CDSs in the third circle Bars on the fourth and fifth circles indicate the rRNAs and tRNAs
respectively Sixth and seventh circles show the coverage and GC content respectively The
innermost circle demonstrates the GC skew + (green) and GC skew- (purple)
135
Figure 5-3 Size distribution of the predicted genes identified in C freundii strain ADS47
The X axis identifies the gene size categories (bp) and Y axis the abundance of genes
136
543 Annotation and functional classification of ADS47 genome
RAST analysis identified predicted CDSs in the genome of Citrobacter strain ADS47
assigned functions and grouped the annotated genes into 26 functional subsystem categories
out of 27 No CDS was found belonging to the photosynthesis category A graphical
presentation of the distribution of the CDSs into each of the 26 categories is given in
Figure 5-4
A large portion (38) of the CDSs belong to hypothetical or unknown functions
Among the 62 known or predicted proteins the highest numbers (835) belong to the
carbohydrate category followed by amino acids and derivatives (468) and
cofactorsvitaminsprosthetic groups (324) A noticeable number of stress response (236) and
respiration (198) categories genes were identified in the bacterial genome The ADS47
genome encoded by 171 membrane transport genes 113 virulencedefense genes and 126
genes involved in regulationcell signaling categories Two hundred and one CDSs were
grouped into the miscellaneous category whose functions are different than the indicated
RAST categories The RAST annotations further divided the categorized CDSs into
subcategories based on similar functional roles which is discussed in section 545
137
Figure 5-4 Functional classification of the predicted coding sequences (CDS) of C freundii
strain ADS47 The annotation was performed by Rapid Annotation using Subsystems
Technology (RAST) program
138
544 Molecular phylogenetic relationships
Genome-wide phylogenetic analysis based on comparisons of whole genome amino
acid sequences of two completely sequenced Citrobacter genomes (C koseri and C
rodentium) and three partially sequenced Citrobacter genome sequences (C freundii
Citrobacter sp and C youngae) illustrates the relationships of ADS47 to five Citrobacter
species (Figure 5-5)
C youngae ATCC 29220 originating from human feces and two pathogenic species
(C koseri ATCC BAA-895 and C rodentium strain ICC168) are progressively more distantly
related to ADS47 The present analysis is consistent with the 16S-rRNA sequence based
phylogenetic analysis (Chapter 3) which showed that the closest bacterial strain to ADS47 is
C freundii (Figure 3-2) This confirmed the identification of strain ADS47 as the gamma
proteobacterial species C freundii made in Chapter 3
139
Figure 5-5 Phylogeny of the trichothecene detoxifying C freundii strain ADS47 The
dendrogram was constructed by comparing the ADS47 genome (amino acid sequences) with
five Citrobacter genomes and a E coli strain (out group species) using the CVTree software
The calculated pairwise distances matrix by Protdist program was transformed into a tree by
neighbor-joining program The dissimilarity tree was drawn to scale with branch lengths
(scale bar 002 indicated the amino acid substitutions among the genomes) The C koseri C
rodentium C freundii Citrobacter sp C youngae and E coli protein sequences were
uploaded from GenBank (Benson et al 2009)
140
545 Comparative genomics
Genome-wide comparative analyses between the genome of strain ADS47 and the five
previously sequenced Citrobacter genomes showed that 872 of the total nucleotides
between ADS47 and C freundii Ballerup were identical The values for the comparisons
between C freundii strain ADS47 were 8212 for Citrobacter sp 30-2 7637 for C
youngae 5981 for C koseri and 4213 for C rodentium (Table 5-2)
The two human and mouse pathogenic species namely C rodentium and C koseri
and the bacteria originally isolated from human feces (C youngae) have higher GC contents
(546 and 532 respectively) compared to C freundii strain ADS47 C freundii strain
Ballerup and Citrobacter sp 30_2 which have slightly lower GC contents (~52) The two
pathogenic species contain 22 copies of rRNA which is significantly higher than the 9 found
in C freundii strain Ballerup 6 in Citrobacter sp 30_2 and 3 in C freundii strain ADS47 but
less than the 25 copies in C youngae The genome of C freundii strain ADS47 C freundii
strain Ballerup C youngae and Citrobacter sp 30-2 had a similar percentage of the genome
comprising coding sequences (865-88) C koseri and C rodentium genomes contained the
highest (90) and lowest (84) genome coverage by coding sequences respectively
141
Table 5-2 Genome comparisons between strain ADS47 and five Citrobacter species The
base nucleotide sequence identities were performed using MUMmer DNAdff script program
(Delcher et al 2002) The features (GC content total CDS percentage CDS and number of
ribosomal RNA) were determined using RAST program (Aziz et al 2008)
Genome feature Bacterial species
C
freundii
ADS47
C freundii
Ballerup
Citrobacter
sp 30_2
C
youngae
C koseri C
rodentium
Genome length
(bp)
4878242
4904641
5125150
5154159
4720462
5346659
Base similarity to
ADS47 ()
NA 8715 8212 7637 5981 4213
GC () 5202 5218 516 5265 530 546
Number of coding
sequences
4610 4685 4728 5278 4978 4800
Genome coverage
with coding
sequences ()
880 865 866 872 900 840
rRNA copies 3 9 6 25 22 22
Total stable RNAs 99 81 78 105 120 142
142
The number of orthologs shared between ADS47 and other Citrobacter species is
illustrated in Figure 5-6 ADS47 shares the smallest number of orthologous CDSs with C
rodentium and C koseri 3250 and 3341 respectively The highest number of shared CDS
were found with C freundii Ballerup (3706) followed by C youngae (3693) and Citrobacter
sp 30_2 (3676) A total of 2821 CDSs were found to be common to all of the six Citrobacter
genomes analysed (data not shown) Nonetheless significant diversity exists between C
freundii ADS47 and the other species as a considerable number of unshared genes were
identified from the comparisons with C freundii Ballerup [979 (20)] Citrobacter sp 30_2
[1064 (23)] C youngae [1654 (31)] C koseri [1693 (34)] and C rodentium [1634
(34)]
143
Figure 5-6 Comparisons of coding sequences between strain C freundii ADS47 and five
Citrobacter species Coding sequences of ADS47 were compared with C freundii Ballerup
Citrobacter sp 30_2 C youngae C koseri and C rodentium genomes based on all-versus-all
reciprocal FASTA comparison of at least 30 amino acid identity and gt70 coverage of the
total length using OrthoMCL program A Venn diagram demonstrates the shared and
unshared (indicated in the brackets) coding sequences
144
Table 5-3 shows differences in the 26 RAST categorized genes between the six
Citrobacter genomes Even though a low abundance of virulencediseasedefense category
genes were found in the animalhuman pathogenic bacteria of C rodentium (85) and C koseri
(92) significantly higher mycobacterium virulence genes (13) were identified in this two
species The genomes of the two closely related strains C freundii ADS47 and C freundii
Ballerup lack this type of virulence gene Instead elevated numbers of genes for resistance to
antibiotic and toxic compounds were found in C freundii ADS47 (94) C freundii Ballerup
(88) and Citrobacter sp (110) compared to other three species Copper homeostasis and
cobalt-zinc-cadmium resistance genes were identified in strain C freundii ADS47 (13 and 21
respectively) and closely related C freundii Ballerup (12 and 22) and Citobacter sp (18 and
22) Significantly lower numbers of these genes were present in C rodentium (6 and 4) and C
koseri (7 and 8) C rodentium and C koseri lack mercuric reductase mercuric resistance and
arsenic resistance genes perhaps because being animalhuman pathogens but these genes
were found in the remaining four genomes The C koseri genome is richer in betandashlactamase
genes (5) compared to the remaining genomes which encodes 1-3 genes
145
Table 5-3 Functional categorized gene abundances among the genomes of C freundii
ADS47 C freundii Ballerup Citrobacter sp 30_2 C youngae C koseri and C rodentium
The genomes were classified into different functional categories with the RAST program
which classified the total CDSs
RAST subsystem category Citrobacter species
C
freundii
ADS47
C
freundii
Ballerup
C
sp
C
youngae
C
koseri
C
rodentium
Cofactorsvitaminsprosthetic
groupspigments
323
317
310
311
325
304
Cell wall and capsule 279 272 248 262 247 262
Virulence disease and defense 113 107 134 100 92 85
Potassium metabolism 50 53 33 32 30 28
Miscellaneous 210 228 200 185 245 185
Phageprophagetransposable
element plasmid
24 75 25 4 6 49
Membrane transport 170 156 171 195 171 172
Iron acquisition and metabolism 80 81 86 62 113 27
RNA metabolism 220 222 217 215 238 212
Nucleosides and nucleotides 119 112 110 111 120 114
Protein metabolism 249 280 269 298 287 254
Cell division and cell cycle 30 29 32 29 38 29
Motility and chemotaxis 133 124 90 147 143 133
Regulation and cell signaling 126 130 124 135 132 123
Secondary metabolism 21 22 20 21 0 0
DNA metabolism 145 167 165 155 147 152
Fatty acids lipids and isoprenoids 121 149 122 121 134 133
Nitrogen metabolism 50 52 53 49 56 58
Dormancy and sporulation 3 3 3 4 3 3
146
Table 5-3 (continued)
RAST subsystem category
Citrobacter species
C
freundii
ADS47
C
freundii
Ballerup
C
sp
C
youngae
C
koseri
C
rodentium
Respiration 198 184 180 191 166 162
Stress response 236 235 170 177 179 174
Metabolism of aromatic
compounds
20 20 28 25 17 48
Amino acids and derivatives 482 476 450 466 428 416
Sulfur metabolism 48 50 52 57 56 56
Phosphorus metabolism 50 49 43 42 43 28
Carbohydrates 837 803 708 731 733 737
147
Strains C freundii ASD47 and C freundii Ballerup have much more potassium
metabolism category genes (50 and 53 respectively) than the other bacterial species which
code for 28-33 proteins Larger numbers of genes associated with the glutathione regulated
potassium efflux system were also identified in C freundii ASD47 with 23 and C freundii
Ballerup with 26 compared to the remaining four species that encode only five genes All of
the six genomes contained phageprophage-related genes with the maximum identified in C
freundii Ballerup (75) followed by C rodentium (49) Citrobacter sp (25) and C freundii
ADS47 (24) Lower numbers of phageprophage-related genes were found in C koseri (6) and
C youngae (4)
The six Citrobacter genomes showed remarkable variability in their secretion systems
The type-III secretion operon genes are only found in C rodentium and C freundii Ballerup
Type-II secretion genes are predominant in C rodentium (12) followed by ADS47 (9)
Type-IV secretion system is uncommon in Citrobacter species since it was only
identified in Citrobacter sp where it is represented by 19 genes Variable numbers of type-VI
secretion system genes are present in the five genomes except C koseri which lacks such
genes C youngae has the highest number (43) of type-VI secretion proteins The other strains
have a range of type-VI secretion proteins in particular 27 were identified in C freundii
Ballerup 15 in C freundii ADS47 and 8 in Citrobacter sp Type-VII (chaperoneusher)
secretion system genes were found in all of the six Citrobacter species with the largest (42)
and smallest (13) numbers of these genes occurring in C koseri and C freundii Ballerup
respectively
Although a relatively similar number of ABC transporters (37-42) were identified in
the six genomes C freundii ADS47 contain the largest number of tripartite ATP-independent
148
periplasmic (TRAP) transporter genes (7) By contrast the closely related C freundii
Ballerup lacks such transporter system An identical number of iron acquisition and
metabolism category genes were found in C freundii ADS47 (80) and C freundii Ballerup
(81) while the two pathogenic species C rodentium and C koseri code for the lowest (27)
and highest (113) numbers of such genes C koseri is rich in siderophore related genes (64)
but the remaining five genomes have smaller numbers of siderophore-associated genes (20-
22)
Variable numbers of motility and chemotaxis related genes were found in the
Citrobacter genomes with the largest number occurring in C youngae (147) followed by C
koseri (143) C freundii ADS47 (133) C rodentium (133) C freundii Ballerup (124) and
Citrobacter sp (90) The two pathogenic species (C rodentium and C koseri) encode a
remarkably low number of quorum sensing genes (2) In contrast C youngae codes for the
highest number of quorum sensing genes (19) A common and distinctive feature of the two
pathogenic species is the lack of genes required for secondary metabolism In contrast the
non-pathogenic species contain a common set of genes for degrading a range of
phenylpropanoids and cinnamic acid (20-22)
The lowest number of genes related to phospholipidsfatty acids biosynthesis and
metabolism were found in C freundii ADS47 (66) compared to C freundii Balllerup (94) and
C koseri (98) The smallest numbers of nitratenitrite ammonification genes were found in
ADS47 (31) and its closest genome C freundii Ballerup (32) In contrast the two pathogens
encode the highest number of such proteins (39-40)
C rodentium and C koseri code for significantly low numbers of respiratory category
genes (162-165) while C freundii ADS47 contains the highest number of such genes (198)
149
Further analysis of the subcategory genes determined that the two gut species (C youngae and
Citrobacter sp 30_2) contain the largest numbers of genes encoding anaerobic reductases
(22) and identical numbers of formate dehydrogenases (17) The largest numbers of formate
dehydrogenase genes were identified in C freundii Ballerup (32) and C freundii ADS47 (30)
but the two pathogenic species differed from the other species by encoding the lowest number
(13) of c-type cytochrome biogenesis genes
A higher number of aromatic compound metabolism category genes were found in the
mouse pathogen C rodentium (48) compared to the human pathogen (C koseri) which
contain 17 genes A significantly higher number of genes involved in central aromatic
intermediate metabolism (26) were found in C rodentium compared to the mouse pathogen
(C koseri) and other genomes which only contain 6-8 such genes The two C freundii strains
ADS47 and Ballerup lacked the anaerobic degradation of aromatic compound
(hydroxyaromatic decarboxylase family) genes
In spite of the abundance of stress response category genes strain ADS47 has only a
small number of desiccation stress genes (5) compared to the other species which ranged
from 9 to10 However ADS47 and Ballerup code for higher number of genes related to
oxidative stress (94 and 91 respectively) and cold shock (8 and 7 respectively) than the other
species
The smallest numbers of amino acids and its derivatives category genes were found in
the two pathogens C rodentium and C koseri (416 and 428 respectively) while the largest
number of these genes were identified in strain ADS47 (482) and C freundii Ballerup (476)
Despite the similar number of sulfur metabolism category genes determined across the six
species only ADS47 lacks taurine utilization genes Strains ADS47 and Ballerup were found
150
to be rich in the phosphorous metabolism gene category (49-50) while C rodentium contains
a significantly decreased number of these category genes (28)
The bioinformatics analyses found the largest number of carbohydrate category genes
in ADS47 (837) followed by Ballerup (803) The gut bacteria (Citrobacter sp 30_2) codes for
the least number of genes assigned to the carbohydrate category (708) while C koseri C
rodentium and C youngae contained 731-737 in this category genes The analyses also
determined that there are relatively small numbers of TCA and Entner Doudoroff pathway
genes in C rodentium and C youngae compared to the other four species and relatively large
numbers of fermentation-related genes in ADS47 (81) and C freundii (85) compared to the
other four species
A comparison of the pathogenicity operons [locus of enterocyte effacement (LEE)] of
C rodentium with other five Citrobacter genomes revealed the absence of this virulence
determinant gene region in the other four Citrobacter species including ADS47 (Figure 5-7)
The analysis showed the presence of LEE operon genes in C freundii strain Ballerup as no
gap was observed in that genomic location while C freundii ADS47 C koseri C youngae
and Citrobacter sp 30_2 lacked that loci (7a) Further ananlysis showed alignments between
C rodentium and E coli LEE operon genes (Figure 7b) In contrast no alignment of such
genes was observed in ADS47 genome
151
(a)
152
Figure 5-7 (a) Blast Ring Image Generator (BRIG) output based of Citrobacter rodentium
(NC_013716) compared against ADS47 C freundii C koseri C youngae E coli and
Citrobacter sp genomes The targeted comparative genome analysis was performed based on
locus of enterocyte effacement (LEE) operons of C rodentium (Petty et al 2011) The
innermost rings show GC content (black) and GC skew (purple and green) The remaining
rings show BLAST comparisons of the other Citrobacter and E coli genomes against C
rodentium The region shown in red is LEE operon genes (b) The absence of LEE operons in
the ADS47 genome comparing to C rodentium and E coli Analysis was performed and
visualized by Artemis Comparison Tool software v132
(httpwwwsangeracukresourcessoftwareact) (Tatusov et al 2000 Craver et al 2005)
LEE operons
(b)
153
546 Analysis of the unique genes in C freundii ADS47
Two hundred and seventy genes unique to ADS47 were identified from a comparison
of its genome to the other five Citrobacter species In general the unique genes are
distributed throughout the ADS47 genome but some clusters of unique genes occurred
including eight unique genomic regions (Figure 5-8) Six large unique regions are located at
767204-775090 (~8 kb) 1025572-1042510 (17 kb) 2798289-2807217 (~9 kb)
3045220-3055912 (~11 kb) 3930366-3943075 (12 kb) and 4022195-4032444 (10 kb)
Out of 270 unique genes 147 (54 ) are hypothetical proteins The remaining 123
(46) genes are grouped into reductasesoxidoreductases (9 genes) de-acetylase (1)
hydrolases (2) transferases (4) sulfatasesarylsulfatases (3) bacteriocins (3) type VI
secretion proteins (2) kinases (4) transporters (11) membraneexported proteins (8)
regulators (12) integraserecombinases (5) phagesprophagestransposaes related genes (27)
and miscellaneous (32) that includes DNA helicases ligases fimbrial proteins and
chromosome partitioning genes (Table 5-4)
The unique regions encode various functional genes that may cause trichothecene de-
epoxidation or de-acetylation reactions The 8 kb unique regions contains six genes encoding
glycosyltransferases and two copies of oxidoreductase-like coding sequences (CMD_0711
and CMD_0712) that showed 39 identity to the Marinobacter aquaeolei F420
hydrogenasedehydrogenase beta subunit The largest (17 kb) unique genomic region codes
for 16 genes including dehydrogenase hydrolase bacteriocin (pyocin) integrase and a short
chain dehydrogenasereductase-like (SDR) (CMD_0979) gene (Figure 5-9a)
154
The SDR like gene is 49 amino acid similarity to an oxidoreductase of the
phytopathogenic Dickeya dadantii and the enzyme contains a domain with 77 homology to
the Dickeya zeae NAD(P) binding site The 9 kb unique genomic section contains 8 genes
including a NaCl symporter a dehydratase (CMD_2679) with 86 homology to
Enterobacteriaceae bacterium dihydroxy-acid dehydratase and a putative oxidoreductase
(CMD_2683) having 37 similarity with oxopropionate reductase of Bacillus sp The unique
region also encodes a short chain dehydrogenasereductase (SDR) (CMD_2684) that shows
85 identity to Enterobacteriaceae bacterium (Figure 5-9b) A unique thiol-disulfide and
thioredoxin like reductase (CMD_3721) was identified in a 10 kb unique region that contains
13 genes (Figure 5-9c) This gene is 59 identical to Oligotropha carboxidovorans thiol-
disulfide isomerase and thioredoxin In addition several reductasesoxidoreductase-like
unique genes were identified in the ADS47 genome including an oxidoreductase
(CMD_1024) with 27 identity to Canadidatus nitrospira NADH-quinone oxidoreductase
membrane L Another oxidoreductase-like gene identified in a different location of the
ADS47 genome (CMD_1393) shows 87 homology to Klebsiella pneumoniae
deoxygluconate dehydrogenase contains a SDR active site A putative nitrate reductase gene
(CMD_3184) has a low level of identity (27) with an uncultured bacterial respiratory nitrate
reductase The ~11 kb unique region encodes sulfatasesarylsulfatses
(CMD_2898CMD_2905) and related genes Furthermore ADS47 contains a unique
glycosylphosphatidylinositol (GPI) anchor biosynthesis operon comprised of four genes
including a N-acetylglucosaminyl-phosphatidylinositol de-acetylase (CMD_2431) which
showed 78 similarity to E coli de-acetylase (Figure 5-9e)
155
Other unique genespredicted genes in the ADS47 genome include
polysaccharidephenazine biosynthesis genes (CMD_0710CMD_1406) heavy metal kinase
(CMD_0989) two component heavy metal regulator (CMD_0990) nitrate reductase regulator
(CMD_3477) putative potassiumzinc transporters (CMD_0973CMD_4600) sodium
symporter (CMD_2678) (tripartite ATP-independent periplasmic transporters) TRAP
transporters (CMD_3693 CMD_3694 CMD_3695 CMD_4337) glutamineribose ABC
transporters (CMD_3176CMD_3544) toxin transcription regulator (CMD_3464) fimbrial
proteinchaperone (CMD_1695 CMD_2434 CMD_3277 CMD_3278) and ParB
chromosomeplasmid partitioning proteins (CMD_4033CMD_4034)
Majority of the unique genes and their distributions are shown in comparative circular
genome maps (Figure 5-8)
156
Table 5-4 Unique genes of C freundii strain ADS47 Genes were categorized according to
the functional similarity The unique genes were determined by comparing ADS47 with five
sequenced Citrobacter species using OrthoMCL software
Unique gene classes Number
Total unique genes
270
Reductases oxidoreductases 9
De-acetylase 1
Hydrolase 2
Transferase 4
Sulfatase 3
Bacteriocin 2
Type VI secretion 2
Kinase 4
Transporter 9
Membraneexported protein 8
Regulator 12
Integraserecombinase 5
Phageprophagetransposae 27
Miscellaneous 34
Hypothetical protein 147
157
158
Figure 5-8 Locations of the unique genes of C freundii strain ADS47 The unique genes
were determined by comparing the total CDS of strain C freundii ADS47 with C freundii
Ballerup Citrobacter sp C youngae C koseri and C rodentium CDSs by OrthoMCL
program The map was developed by Blast Ring Generator (BRIG) software First second
and third circles from inside to out side show the genome size GC and GC skew of strain
ADS47 Forward (Fwd) and reversed (Rev) transcribed CDSs of ADS47 are shown in green
(forth circle) and red (fifth circle) respectively The next five rings represent the circular
genome maps of five Citrobacter species distinguished by different ring colours (C
rodentium as sky blue C koseri as dark blue C youngae as light blue C freundii Ballerup as
yellow and Citrobacter sp as green) The degree of orthologous gene identity between
ADS47 and other five genomes are shown by color intensity (light color= 75 gene identity
ash color= 50 gene identity) Gaps across five genome rings indicate the absence of
genesregions compared to ADS47 Unique genes are shown surrounding the map rings and
the identities of some of the genes are given Arrows indicate putative
reductasesoxidoreductases and de-acetylase
159
(a)
(b)
160
(c)
(d)
161
Figure 5-9 Unique genomic regions and associated genes in C freundii ADS47 compared to
five Citrobacter species RAST server based analysis compared the unique genesregions of
ADS47 with the publicly available databases The upper panels demonstrate the large unique
regions defined by yellow boxes and genes located in the genomic regions are indicated by
arrows The lower panels are closely related genesoperons found in distantly related species
The unique regions that contain potential trichothecene de-epoxidation genes are shown in (a)
short chain dehydrogenasereductase (SDR) and dehydrogenase (b) putative oxidoreductase
dehydratase and SDR (c) thiol-disulfide isomerase and thioredoxin oxidoreductase and (d)
unique glycosylphosphatidylinositol (GPI) biosynthesis operon having N-acetylglucosaminyl-
phosphatidylinositol de-acetylase gene
162
55 Discussion
The de novo sequenced ADS47 genome was oriented and assembled using a
completely sequenced C koseri genome as reference strain The synteny analysis between
ADS47 and C koseri showed very similar overall organization of the two genomes The high
preservation of genes among five Citrobacter species and ADS47 were emphasized by the
close alignments observed by Blast Ring Image Generator (BRIG) outputs obtained from
these genomes
Genome-wide phylogenetic analyses based on a large number of common CDSs
(2821) observed within the six Citrobacter genomes confirmed the species identification
based on 16S-rDNA sequencing of the trichothecene mycotoxin-detoxifying C freundii strain
ADS47 The base sequence idendity was 8715 between these two genomes They shared
~80 CDS and had a similar GC content (52) Citrobacter are Gram-negative rod shaped
and facultatively anaerobic bacteria belonging to the family of Enterobacteriaceae (Wang et
al 2000 Whalen et al 2007) Strains ADS47 and Ballerup of C freundii are both aerobic
gram-negative bacilli that have numerous flagella and inhabit in soil (Cong et al 2011 Islam
et al 2012) C freundii is considered to have a positive environmental role by reducing nitrate
to nitrite (Rehr and Klemme 1989) which is an important step in the nitrogen cycle
C freundii C rodentium and C koseri are animalhuman pathogenic species C
rodentium is a natural mice pathogen causing hemorrhagic colitis and hemolytic uremic
syndrome (Gareau et al 2010) C koseri causes human disease (Badger et al 1999 Kariholu
et al 2009) particularly neonatal meningitis and C freundii is an opportunistic human
pathogen responsible for a various types of infections like respiratory tract urinary tract and
blood (Whalen et al 2007) C freundii is very common representing approximately 29 of
163
all opportunistic human infections (Hodges et al 1978) Outside of animals they can be found
in soil water sewages and different organs of diseased animals (Wang et al 2000) These
pathogens code for large numbers of mycobacterium virulence operonsgenes which were
absent in the three other genomes However they have fewer genes involved in environmental
stress such as copper homeostasis genes and no mercuricarsenic resistance and secondary
metabolism category genes unlike the other Citrobacter genomes Nonetheless considerable
variability exists between the two pathogens correlates with C rodentium causing disease in
mice C koseri causing meningistis in human and C freundii causing infections in different
human organsThere were attaching and effacing (AE) lesion forming (LEE) operons in C
rodentium and C freundii but not in C koseri Those genes have shown to be acquired by
lateral gene transfer playing major roles in virulence of enteropathogenic and
enterohemorrhagic E coli and C rodentium (Mundy et al 2005 Petty et al 2010 Petty et al
2011 The ~356 kb pathogenic LEE gene island is comprised of five operons (LEE1 to
LEE5) that code for ~40 virulence genes that include type-III secretion genes and associated
effectors (Berdichevsky et al 2005) In addition an elevated number of type-II and -VI
secretion proteins identified in C rodentium which may act as additional virulence factors
(Petty et al 2010) Because it lacks type-III secretion apparatus genes it is likely that C
koseri uses a different strategy to cause human neonatal meningitis (Badger et al 1999 Ong
et al 2010)
To adapt to different environmental niches or hosts bacteria like Citrobacter species
frequently gainlose genes increase repeat sequences and recombine genomic regions using
different mechanisms (Parkhill et al 2001 Petty et al 2011) The existence of large numbers
of phageprophagetransposae related genes in the Citrobacter genomes particularly in C
164
freundii Ballerup suggests that their genomes are unstable For example C freundii Ballerup
and C koseri might have acquired LEE operons by horizontal transfer via phage mediated
transpositions Interestingly although ADS47 is closely related to C freundii Ballerup it
lacks the pathogen-related LEE gene island and mycobacterium virulence operons required
for invasion and intracellular survival in macrophages (Gao et al 2006) Instead ADS47 has a
larger number and variety of stress-responsive genes such as efflux genes and bacteriocins
(pyocin hemolysin) genes that provide protection against heavy metals and antibiotics and
increase their competitiveness against other bacteria These findings suggest that the strain
ADS47 is not an opportunistic animal pathogen but has evolved to be highly competitive in
the soil micro-environment (Arnold 1998 Bengoechea and Skurnik 2000) Perhaps ADS47
evolved trichothecene degrading genes for adaptation to soils containing trichothecene
mycotoxins contaminated soils (Gadd and Griffiths 1978 Sharma and Fulekar 2009 Han et
al 2011) Conversely the absence or reduced number of stress-responsive genes in C
rodentium and C koseri may imply that those two pathogenic species are not exposed to such
stressful and xenobiotic-contaminated environments as ADS47
This interpretation is supported by the observation that the two of the three
animalhuman pathogenic species contained high of rRNA copy number but lower numbers
were found in ADS47 C freundii Ballerup and Citrobacter sp 30_2 Bacteria maintain large
number of rRNA for higher production of ribosome during high growth rates (Stevenson et al
1997 Klappenbach et al 2000) such as might occur during the establishment of an infection
However a large copy number of rRNAs may not be metabolically favourable for bacteria
such as ADS47 that evolved in a highly competitive environment as that costs higher cellular
energy (Hsu et al 1994)
165
C freundii Ballerup seems to be an exception in that it is similar to the other animal
pathogens for containing virulent factors (ie LEE-operon genes) but more like the
environmental species for encoding high number genes for stress defence heavy metal and
toxic compounds resistances
The comparison of the genes in C freundii ADS47 with the unique ability to detoxify
trichothecenes with genes in other Citrobacter species allowed approximately 270 unique
genes to be identified in ADS47 Some of those genes may be involved in trichothecene de-
epoxidation or de-acetylation Currently the pathways and microbial enzymes leading to
trichothecene de-epoxidation and de-acetylation are unknown but based on the required
reaction ie the elimination of an oxygen atom from epoxide moiety it is likely that epoxide
reductases or epoxide oxidoreductases are responsible for such biodegradation activity (He et
al 1992)
From database searches three enzymes having epoxide reductase activity were found
These are a vitamin K epoxide reductase (VKOR) a monooxygenase PaaABCE and pyridine
nucleotide-disulfide oxidoreductase (PNDO) which reduce vitamin K epoxide phenylacetyl-
CoA and epoxyalkanes respectively (Swaving et al 1996 Li et al 2004 Teufel et al 2012)
In mammals VKOR plays important roles in vitamin K cycle by reducing vitamin K epoxide
to vitamin K and its homologues that are found in plants bacteria and archaea (Goodstadt
2004 Li et al 2010) The microbial multi-component PaaABCE monooxygenase catalyzes
degradation of variety of aromatic compounds or environmental contaminants via epoxidation
of phenylacetic acid (Paa) and NADPH-dependant de-epoxidation of epoxy-Paa (Luengo et
al 2001 Teufel et al 2012) PNDO is a unique type of NADP-dependent oxidoreductase
identified in Xanthobacter strain Py2 that was obtained by propane enrichment procedure
166
(van Ginkel and de Bont 1986) However no orthologs of these genes were found in ADS47
genome
Nine ADS47-unique reductaseoxidoreductase-like genes are potential candidates for
the trichothecene de-epoxidation enzyme Genes belong to short chain
dehydrogenasereductase (SDR) family are structurally and functionally diverse and cause
different enzymatic activities including xenobiotic degradation (Kavanagh et al 2008) This is
a promising candidate for the trichothecene de-epoxidation gene Genes belong to dehydratase
family catalyze reduction reactions by cleaving carbon-oxygen bonds of organic compounds
that result in removal of oxygen in various biosynthetic pathways (Flint et al 1993) This
mechanism may be relevant to microbial trichothecene de-epoxidation reaction since the latter
involves removal of the oxygen atom forming the epoxide in trichothecene mycotoxins
(Young et al 2007)
Another ADS47-unique gene was for a nitrate reductase that may be a candidate de-
epoxidation gene Under stress conditions such an enzyme causes reductive removal of an
oxygen molecule (Morozkina and Zvyagilskaya 2007)
A fourth type of ADS47-unique gene is for a thiol-disulfide isomerasethioredoxin like
(Trx) oxidoreductase The thiol-disulfide isomerase and thioredoxin oxidoreductases contain a
thioredoxin like domain with a CXXC catalytic motif which catalyzes various biological
processes that allow the organism to adapt to environmental stress and develop competence to
sporulate (Scharf et al 1998 Petersohn et al 2001 Smits et al 2005) The membrane-
anchored Trxndashlike domain initiates the vitamin K de-epoxidation reaction by reducing the
active motif (CXXC) of VKOR (Schulman et al 2010)
167
The present study did not find any homologues of VKOR in ADS47 genome but we
identified two copies of structurally similar quinone reductase protein disulfide bond
formation (DsbB) gene (CMD_1044 and CMD_2641) Both VKOR and DsbB contain four
transmembrane segments an active site CXXC motif and cofactor quinone and use a
common electron transfer pathway for reducing substrates (Li et al 2010 Malojcic et al
2008) As VKOR bacterial DsbB requires a Trx-like partner (eg DsbA) for activity Further
experiments are required to examine if DON de-epoxidation occurs by the catalytic activity of
DsbB coupled with a unique thiol-disulfide isomerase and thioredoxin oxidoreductase present
in ADS47 genome
The analysis also identified an ADS47-unique glycosylphosphatidylinositol (GPI)
biosynthetic operon that is comprised of four genes including an N-
acetylglucosaminylphosphatidylinositol de-acetylase The GPI membrane anchor protein
synthesis is initiated by transferring N-acetylglucosamine from uridine diphosphate N-
acetylglucosamine to phosphatidylinositol by N-acetylglucosaminyltransferase an
endoplasmic reticulum (ER) membrane-bound protein complex (Hong and Kinoshita 2009)
In the second step of the pathway N-acetylglucosamine-phosphatidylinositol is de-acetylated
to produce glucosaminyl-phosphatidylinositol by the catalytic activity of a zinc
metalloenzyme N-acetylglucosaminylphosphatidylinositol de-acetylase which is localized on
the cytoplasmic side of the ER (Nakamura et al 1997 Hong and Kinoshita 2009) It is
possible that the identified unique N-acetylglucosaminylphosphatidylinositol de-acetylase
gene of ADS47 may be involved in trichothecene de-acetylation activity
168
Further analysis is needed to determine which of these candidate gene or genes may be
involved in tricthothecene mycotoxin de-epoxidation and de-acetylation activities
Identification of gene responsible for the trichothecene detoxification activities of C freundii
ADS47 opens many potential applications for its use in agri-based industries to contribute to
cropanimal production and food safety (Karlovsky 2011)
56 Acknowledgement
We gratefully acknowledge Agriculture and Agri-Food CanadaBinational
Agricultural Research and Development Fund (AAFCBARD) project for providing the
financial supports of the work
169
Supplemental Table S5-1 List of unique genes of strain ADS47 gene IDs and their
functional roles The unique genes were determined by comparing the ADS47 genome with
five sequenced Citrobacter species (C freundii Citrobacter sp C youngae C koseri and C
rodentium) using OrthoMCL (Li et al 2003)
Unique gene
classes
Number Gene ID and functions
Total unique
genes
270 123 known functions 147 hypothetical proteins
Reductases
oxidoreductases
9 CMD_0711 coenzyme F420 hydrogenasedehydrogenase beta subunit
CMD_0979 Short chain dehydrogenase reductase (SDR) Oxidoreductase
SDR dehydrogenase CMD_1024 NADH-quinone oxidoreductase
membrane L CMD_1393 oxidoreductase SDR putative deoxygluconate
dehydrogenase (NAD+ 3-oxidoreductase) CMD_2679
Dehydratasedihydroxy acid dehydratase CMD_2683 oxidoreductase
oxopropionate reductase CMD_2684 short-chain dehydrogenasereductase
SDR CMD_3184 nitrate reductase alfa subunit CMD_3721 thiol-disulfide
isomerase and thioredoxin oxidoreductase
Deacetylase 1 CMD_2431 N-acetylglucosaminylphosphatidylinositol deacetylase
superfamily
Hydrolase 2 CMD_0991 Dienelactone hydrolase CMD_2895 Putative glycosyl
hydrolase
Transferase 4 CMD_1640 D12 class N6 adenine-specific DNA methyltransferase
CMD_2432 Glycosyltransferase CMD_2433 putative acetyltransferase
CMD_3023 adenine-specific DNA modification methyltransferase
Sulfatase 3 CMD_2898 Possible sulfatase arylsulfatasearylsulfatransferase
CMD_2902 Arylsulfatasesulfatase CMD_2905 Putative
sulfatasearylsulfatase
Bacteriocin 3 CMD_0982 Thermosatble hemolysin superfamily CMD_0993 Pyocin S2
superfamily (bacteriocin that kills other bacterial strain) CMD_3705 S-type
pyocin containing domain protein
Type VI secretion 2 CMD_1802 EvpB family type VI secretion protein CMD_2364 type VI
effector Hcp1
Kinase 4 CMD_0989 Heavy metal Sensor kinase CMD_1794 Polo kinase kinase
(PKK) superfamily CMD_3124 predicted diacylglycerol kinase CMD_4266
putative signal transduction kinase
Transporter 11 CMD_0973 TrkA-N domain-containing protein (K uptake transporter)
CMD_1407 Branched-chain amino acid transport system carrier protein
CMD_2678 Sodium Symporter CMD_3176 glutamine ABC transporter
CMD_3544 Ribose ABC transport system CMD_3693 TRAP-type C4-
dicarboxylate transport system binding protein CMD_3694 TRAP-type C4-
dicarboxylate transport system binding protein small permease component
(predicted N-acetylneuraminate transporter) CMD_3695 TRAP-type C4-
dicarboxylate transport system large permease CMD_4337 23-diketo-L-
gulonate TRAP transporter small permease protein yiaM component
CMD_4338 23-diketo-L-gulonate TRAP transporter large permease protein
yiaN CMD_4600 putative transporter involved in Zinc uptake (YciA
protein)
170
Supplmental Table S5-1 (continued)
Unique gene
classes
Number Gene ID and functions
Membrane
exported protein
8 CMD_0713 putative membrane protein wzy CMD_0983 Putative exported
protein precursor CMD_0986 Putative transmembrane protein
CMD_0987 putative exported protein precursor CMD_1458 putative
exported protein CMD_2903 Putative exported protein CMD_2904
putative secreted 5-nucleotidase CMD_4291 (S) putative integral
membrane export protein
Regulator 12 CMD_0977 probable transcriptional regulator protein CMD_0980 Putative
transcription activator TenA family CMD_0990 Two component heavy
metal response transcription regulator CMD_1766 Leucine responsive
regulatory protein CMD_2677 transcriptional regulator GntR CMD_2909
putative arylsulfatase regulator CMD_3279 Uncharacterized outer
membrane usher protein yqiG precursor CMD_3464 toxin transcription
regulatory protein CMD_3465 EAL regulator protein (signaling)
CMD_3475 putative transcription regulator CMD_3477 Fumarate and
nitrate reduction regulatory protein (for anaerobic adaptation) CMD_3812
putative regulator from phage
Integrase
recombinase
5 CMD_0785 CMD_0992 CMD_0994 CMD_4032 CMD_4424
Phage prophage
transposae related
genes
27 Phages CMD_1608 CMD_1609 CMD_1610 CMD_1611 CMD_1619
CMD_1631 CMD_1913 CMD_1914 CMD_1915 CMD_1917
CMD_1920 CMD_1921 CMD_1936 CMD_3351 CMD_3804
CMD_3805 CMD_3807 CMD_3808 CMD_4581 CMD_4609
CMD_4610 Transposae CMD_0252 CMD_0398 Prophage CMD_1645
CMD_3806 CMD_4425 CMD_4597
Miscellaneous 32 CMD_0095 ATP-dependent DNA helicase Rep CMD_0656 LysA protein
CMD_0710 Polysaccharide biosynthesis family protein CMD_0978 TPR
repeat containing protein CMD_0981 AMP dependant synthatase and
ligase CMD_0985 Cupin superfamily CMD_0987 flavodoxin
CMD_1404 Putative endoribonuclease CMD_1405 Putative PAC domain
protein CMD_1406 Phenazine (PhzF) biosynthesis protein CMD_1695
Putative fimbrial protein CMD_1793 HPChromosome segregation ATPase
domain protein CMD_1922 FRG uncharacterized protein domain
CMD_2206 L-fucose isomerase like protein CMD_2434 CS12 fimbria
outer membrane usher protein CMD_2680 HpcHHpaI aldosecitrate lyase
protein family CMD_2681 Aldose -1 epimerase CMD_2682
Gluconolaconase LRE like region CMD_2900 Putative cytoplasmic
protein CMD_3009 putative restriction endonuclease CMD_3277 Putative
yqi fimbrial adhesion CMD_3278 Hypothetical fimbrial chaperone yqiH
CMD_3340 Pea-VEAacid family CMD_3518 3-hydroxy-3-methylglutaryl
CoA synthase CMD_3801 putative reverse transcriptase CMD_3840 beta-
D-galactosidase alpha subunit CMD_4033 ParB family chromosome
partitioning protein CMD_4034 ParBrepB chromosomeplasmid
partitioning protein CMD_4035 RelASpoT protein CMD_4532 putative
DNA ligase -like protein CMD_4541 Antitermination protein Q and
CMD_4585 putative ATP dependent helicase
171
CHAPTER 6
60 CONCLUSION
61 Summary
Contamination of grain with mycotoxins including deoxynivalenol (DON) and other
trichothecenes is becoming a limiting factor for cereal and animal production (Binder et al
2007) To address the issue this study obtained a novel microbial source for DON
detoxification and determined candidate genes for the function which may have potential
applications to detoxify grain and grain-derived foodfeed contaminated with DON and TC
mycotoxins
From a screen of soil samples from crop field microorganism was isolated that is able
to de-epoxidize the most frequently detected foodfeed contaminating mycotoxin DON The
effects of media compositions temperature pH and aerobicanaerobic conditions on
microbial detoxification activity were assessed From 150 analyzed soil samples a mixed soil
microbial culture that showed DON de-epoxidation activity under aerobic conditions and
moderate temperature (~24degC) was obtained This culture exhibited a slow rate of microbial
DON to de-epoxyDON transformation (de-epoxidation) possibly because of competition
between de-epoxidizing and other microorganisms in the culture
The mixed microbial culture was enriched for DON detoxifying bacteria by reducing
the non-targeted microbial species through antibiotic treatments heating and subculturing on
agar medium and selecting colonies The enriched culture had higher DON de-epoxidation
activity than the initial culture Because of low abundance of DON detoxifying
microorganisms further culture enrichments were required to increase the target microbial
172
population Reculture of the partially enriched microbial community in nutritonally poor MSB
medium with high level of DON increased the DON de-epoxidizing bacterial population
Serial dilution plating and selecting single colonies eventually led to isolation of a DON-
detoxifying bacterial strain named ADS47
Phenotypic and 16S-rDNA sequence analyses provided evidence that the bacterium is
a strain of C freundii Strain ADS47 is a gram negative rod-shaped facultative anaerobe and
has peritrichous flagella Strain ADS47 was unable to use DON as sole carbon source
however increasing DON concentrations in growth media enhanced its DON metabolism
efficiency The pure bacterial culture was shown to be able to rapidly de-epoxidize DON
under aerobic conditions moderate temperatures and neutral pH Strain ADS47 more quickly
de-epoxidizes DON in aerobic conditions than the anaerobic conditions In addition to DON
strain ADS47 showed capability to de-epoxidize andor de-acetylate 10 different type-A and
type-B TC mycotoxins which often co-contaminant grain with DON
To clone the DON detoxifying genes a DON detoxifying bacterial DNA library in E
coli host was constructed followed by screening of the library clones for DON detoxification
activity The study produced a library giving approximately 10 fold coverage of the genome
of strain ADS47 Restriction digestion analysis determined that most of the clones contained
ADS47 DNA inserts This indicates that a good DNA library was produced which had gt99
probability of containing all the DNA of the ADS47 genome However activity-based
screening of the library could not identify any clones with DON de-epoxidation ability It is
assumed that bacterial DON de-epoxidation genes evolved in a certain environmental niche
which may be required for the DON de-epoxidation to be expressed and active but was not
supplied by the host E coli cells
173
An attempt was made to identify the bacterial genes encoding DON de-epoxidation
and de-acetylation enzymes by sequencing the genome of strain ADS47 and comparing its
sequence to the genomes of related bacteria that are not known to posses DON de-epoxidation
activity A RAST server based bioinformatics analysis determined that the 48 Mb circular
genome of ADS47 codes for 4610 predicted CDS A genome wide phylogenetic analysis
determined that strain ADS47 was gt87 identical to the pathogen C freundii strain Ballerup
The relationship to two other animalhuman pathogenic species C rodentium and C koseri
was more distant This result is consistent with the 16S-rDNA based phylogenetic analysis
It is assumed that the microbial DON detoxifying genes were acquired by horizontal
gene transfer or other means during the adaptation of ADS47 in mycotoxin contaminated soil
If it is correct then this means that those newly acquired genes are not common in other
related microbial species and no other Citrobacter species is known to degrade DON Hence
the study identified 270 unique genes to ASDS47 with a subset of 10 genes for reductases
oxidoreductases and a de-acetylase which were considered to be candidate genes for the
enzymes that have DON detoxifying activity Furthermore a targeted comparative genome
analysis showed that the mycotoxin detoxifying ADS47 genome does not harbour the
clinically important LEE encoded typendashIII secretion system and mycobacterium virulence
genes This suggests that strain ADS47 is not likely to be an animal pathogenic bacterium
unlike C freundii Ballerup which encodes the pathogenic relevance LEE operons
174
62 Research contributions
The research made four major contributions which may aid in reducing the risks of
animals and humans from DONTC exposure Firstly the study identified a novel bacterial
strain ADS47 that is able to detoxify DON and 10 related mycotoxins over a wide range of
temperatures under aerobic and anaerobic conditions The discovery has led to a patent
application for strain ADS47 supporting the novelty and importance of the finding This
bacterial strain could be used to detoxify trichothecene mycotoxins in contaminated animal
feed thus reducing food safety risks
Secondly the study resulted in the development of a method for isolating DON de-
epoxidizing microbes from soil as well as optimizing the conditions for bacterial DONTC
detoxification activity to increase the effectiveness usefulness of the strain
Thirdly the thesis work resulted in availability of the genome sequence of a
trichothecene detoxifying microbe which may ultimately lead to an understanding of the
cellularmolecular mechanisms of DONTC de-epoxidation and de-acetylation
genespathways The genome analysis resulted in identification of several candidate genes for
DONTC de-epoxidation and de-acetylation activities Genes which show de-epoxidation
activity could be used to develop DONTC resistant cereal cultivars Fourthly strain ADS47
lacks major animal pathogenicity genes (in particular the LEE-operons) as well as does not
encodes mycobacterium virulence genes Thus strain ADS47 has a great potential to be
utilized to detoxify cereal grain and may be used as a feed additive for livestockfishery All
together the reported work may have significant applications to produce and supply
DONTC-free food and feed
175
63 Further research
This research provides the basis for developing biological detoxification of DON and
related mycotoxins This goal might be achieved by using DON de-epoxidizing gene to
develop resistant plants enzymatic detoxification of contaminated feed and use of the bacteria
as a feed additive Follow-up studies may reach the above indicated goals Because the
bacterium showed de-epoxidation activity under aerobicanaerobic conditions and under wide
range of temperatures it is likely to have an application for enzymatic detoxification of
contaminated animal feed Therefore additional studies on bacterial efficiency to detoxify
contaminated cereal and grain-derived feed need to be conducted Although strain ADS47
does not encode major animal pathogenicity genes the bacterium must be directly tested on
animals to see if it could negatively affect animal digestive systems If not then the potential
for ADS47 is considerable for its use as a feed additive for pig poultry and fish Animal
feeding trials will also need to be performed to determine if DON detoxification is sufficient
to be cost effective as a feed additive In order to develop DONTC resistant cereal cultivars
the gene responsible to DON degrading activity among the presently identified candidate
genes (reductases oxidoreductases and de-acetylase) will need to be expressed in other
organisms like E coli and a codon modified version adapted for expression in plants like
corn and wheat
176
70 REFERENCES
Abbas HK Mirocha CJ Pawlosky RJ Pusch DJ 1985 Effect of cleaning milling and
baking on deoxynivalenol in wheat Applied and Environmental Microbiology 50482ndash
486
Accensi F Pinton P Callu P Abella-Bourges N Guel WJF Grosjean F Oswald IP 2006
Ingestion of low doses of deoxynivalenol does not affect haematological biochemical or
immune responses of piglets Journal of Animal Science 841935ndash1942
Agrawal D Mahapatra AK 2005 Vertically acquired neonatal Citrobacter brain abscessndash
case report and review of the literature Journal of Clinical Neuroscience 12188ndash190
Alexander NJ 2008 The TRI101 story engineering wheat and barley to resist Fusarium head
blight World Mycotoxin Journal 131-37
Alexander NJ Proctor RH McCormick SP 2009 Genes gene clusters and biosynthesis of
trichothecenes and fumonisins in Fusarium Toxin Reviews 28198ndash215
Ali ML Taylor JH Jie L Sun G William M Kasha KJ Reid LM Pauls KP 2005
Molecular mapping of QTLs for resistance to Gibberella ear rot in corn caused by
Fusarium graminearum Genome 48521-533
Alikhan N-F Petty NK Zakour NLB Beatson SA 2011 BLAST Ring Image Generator
(BRIG) simple prokaryote genome comparisons BMC Genomics 12402
Alisi A Spaziani A Anticoli S Ghidinelli M Balsano C 2008 PKR is a novel functional
direct player that coordinates skeletal muscle differentiation via p38MAPKAKT
pathways Cellular Signalling 20534ndash542
177
Amuzie CJ Shinozuka J Pestka JJ 2009 Induction of suppressors of cytokine signaling by
the trichothecene deoxynivalenol in the mouse Toxicological Science 111277ndash287
Ansorge WJ 2009 Next generation DNA sequencing techniques New Biotechnology
25195-203
Arnold JW Senter SD 1998 Use of digital aroma technology and SPME GC-MS to compare
volatile compounds produced by bacteria isolated from processed poultry Journal of the
Science of Food and Agriculture 78343ndash348
Audenaert K Van Broeck R Bekaert B De Witte F Heremans B Messens K Hofte
M Haesaert G 2009 Fusarium head blight (FHB) in Flanders population
diversity inter-species associations and DON contamination in commercial winter
wheat varieties European Journal of Plant Pathology 125445-458
Awad WA Ghareeb K Bohm J Zentek J 2010 Decontamination and detoxification
strategies for the Fusarium mycotoxin deoxynivalenol in animal feed and the
effectiveness of microbial biodegradation Food Additives and Contaminants 27510-520
Aziz K Bartels D Best AA DeJongh M Disz T Edwards RA Formsma K Gerdes S Glass
EM Kubal M Meyer F Olsen GJ Olson R Osterman AL Overbeek RA McNeil LK
Paarmann D Paczian T Parrello1 B Pusch GD Reich C Stevens R Vassieva O
Vonstein V Wilke A Zagnitko O 2008 The RAST Server Rapid Annotations using
Subsystems Technology BMC Genomics 975
Badger JL Stins MF Kim KS 1999 Citrobacter freundii invades and replicates in human
brain microvascular endothelial cells Infection and Immunology 674208ndash4215
Bae HK Pestka JJ 2008 Deoxynivalenol induces p38 interaction with the ribosome in
monocytes and macrophages Toxicological Science 10559-66
178
Bai GH Desjardins AE Plattner RD 2002 Deoxynivalenol-nonproducing Fusarium
graminearum causes initial infection but does not cause disease spread in wheat spikes
Mycopathologia 15391ndash98
Barrett JR 2000 Mycotoxins of molds and maladies Environmental Health Perspectives
108A20ndashA23
Bengoechea JA Skurnik M 2000 Temperature-regulated efflux pumppotassium antiporter
system mediates resistance to cationic antimicrobial peptides in Yersinia Molecular
Microbiology 3767ndash80
Bennett JW Klich M 2003 Mycotoxins Clinical Microbiology Reviews 16497ndash516
Benson DA Karsch-Mizrachi I Lipman DJ Ostell J Sayers EW 2009 GenBank Nucleic
Acids Research 37D26-31
Berdichevsky T Friedberg D Nadler C Rokney A Oppenheim A Rosenshine I 2005 Ler is
a negative autoregulator of the LEE1 operon in neuropathogenic Escherichia coli Journal
of Bacteriology 187349ndash357
Bernardo A Bai GH Guo PG Xiao K Guenzi AC Ayoubi P 2007 Fusarium graminearum-
induced changes in gene expression between Fusarium head blight-resistant and
susceptible wheat cultivars Functional and Integrative Genomics 769ndash77
Bhat RV Beedu SR Ramakrishna Y Munshi KL 1989 Outbreak of trichothecene
mycotoxicosis associated with consumption of mould-damaged wheat production in
Kashmir Valley India The Lancet 33335ndash37
Binder EM Tan LM Chin LJ Handl J Richard J 2007 Worldwide occurrence of
mycotoxins in commodities feeds and feed ingredients Animal Feed Science and
Technology 137265-282
179
Birzele B Meier A Hindorf H Kraumlmer J Dehne H-W 2002 Epidemiology of Fusarium
infection and deoxynivalenol content in winter wheat in the Rhineland Germany
European Journal of Plant Pathology 108667ndash673
Boddu J Cho S Muehlbauer GJ 2007 Transcriptome analysis of trichothecene-induced gene
expression in barley Molecular PlantndashMicrobe Interactions 201364ndash1375
Borenshtein D Schauer DB 2006 The Genus Citrobacter Prokaryotes 690ndash98
Boutigny A-L Richard-Forget F Barreau C 2008 Natural mechanisms for cereal resistance
to the accumulation of Fusarium trichothecenes European Journal of Plant Pathology
121411ndash423
Boutrif E Canet C 1998 Mycotoxin prevention and control FAO programmes Revue de
Meacutedecine Veacuteterinaire 149681-694
Brennan JM Leonard G Fagan B Cooke BM Ritieni A Ferracane R et al 2007
Comparison of commercial European wheat cultivars to Fusarium infection of head and
seedling tissue Plant Pathology 5655ndash64
Brenner DJ Grimont PA Steigerwalt AG Fanning GR Ageron E Riddle CF 1993
Classification of Citrobacteria by DNA hybridization designation of Citrobacter farmeri
sp nov Citrobacter youngae sp nov Citrobacter braakii sp nov Citrobacter
werkmanii sp nov Citrobacter sedlakii sp nov and three unnamed Citrobacter
genomospecies International Journal of Systematic Bacteriology 43645-58
Brookes G Barfoot P 2006 GM Crops The first ten years-global socio-economic and
environmental Impacts no 36 ISAAA (International Service for the Acquisition of Agri-
Biotech Applications) Ithaca New York
180
Bruce KD Hughes MR 2000 Terminal restriction fragment length polymorphism monitoring
of genes amplified directly from bacterial communities in soils and sediments Molecular
Biotechnology 16261ndash269
Buerstmayr H Ban T Anderson JA 2009 QTL mapping and marker-assisted selection for
Fusarium head blight resistance in wheat A review Plant Breeding 1281ndash26
Campbell H Choo TM Vigier B Underhill L 2002 Comparison of mycotoxin profiles
among cereal samples from eastern Canada Canadian Journal of Botany 80526ndash532
Carver et al 2005 ACT the Artemis Comparison Tool Bioinformatics 213422-3423
Carver et al 2009 DNAPlotter circular and linear interactive genome visualization
Bioinformatics 25119-120
Chen CY et al 2003 Comparative genome analysis of Vibrio vulnificus a marine pathogen
Genome Research 132577ndash2587
Chiu TC Yen JH Hsieh YN Wang YS 2005 Reductive transformation of dieldrin under
anaerobic sediment culture Chemosphere 601182ndash1189
Chung YJ Zhou HR Pestka JJ 2003 Transcriptional and posttranscriptional roles for p38
mitogen-activated protein kinase in upregulation of TNF-alpha expression by
deoxynivalenol (vomitoxin) Toxicology and Applied Pharmacology 193188ndash201
Chung EJ Lim HK Kim J-C Choi GJ Park EJ Lee MH Chung YR Lee S-W 2008 Forest
soil metagenome gene cluster involved in antifungal activity expression in Escherichia
coli Applied and Environmental Microbiology 74723-730
Collins TFX Sprando RL Black TN Olejnik N Eppley RM Hines FA Rorie J Ruggles DI
2006 Effects of deoxynivalenol (DON vomitoxin) on in utero development in rats Food
and Chemical Toxicology 44747ndash757
181
Cong Y Wang J Chen Z Xiong K Xu Q Hu F 2011 Characterization of swarming motility
in Citrobacter freundii FEMS Microbiology Letters 371160-171
Cossette F Miller JD 1995 Phytotoxic effect of deoxynivalenol and Gibberella ear rot
resistance of corn Natural Toxins 3383ndash388
Creppy EE 2002 Update of survey regulation and toxic effects of mycotoxins in Europe
Toxicological Letters 12719-28
Dai JS Wang D Guerlebeck C Laturnus S Guenther Z Lu SC Ewers C 2010 Suppression
subtractive hybridization identifies an autotransporter adhesin gene of E coli IMT5155
specifically associated with avian pathogenic Escherichia coli (APEC) BMC
Microbiology 10236
Daniel R 2005 The metagenomics of soil Nature Review Microbiology 3470-478
Davis KER Joseph SJ Janssen PH 2005 Effects of growth medium inoculum size and
incubation time on culturability and isolation of soil bacteria Applied and Environmental
Microbiology 71826-834
Delcher AL Phillippy A Carlton J Salzberg SL 2002 MUMmer 21 NUCmer and
PROmer are described in Fast Algorithms for Large-scale Genome Alignment and
Comparisonrdquo Nucleic Acids Research 302478-2483
Demain AL Adrio JL 2008 Contributions of microorganisms to industrial biology
Molecular Biotechnology 3841ndash455
Deng W Li Y Vallance BA Finlay BB 2001 Locus of enterocyte efffacement from
Citrobacter rodentium sequence analysis and evidence for horizontal transfer among
attaching and effacing pathogens Infection and Immunity 696323ndash6335
182
Desjardins AE Proctor RH Bai G McCormick SP Shaner G Beuchley G Hohn TM 1996
Reduced virulence of trichothecene-nonproducing mutants of Gibberella zeae in wheat
field tests Molecular Plant-Microbe Interactions 9775-781
Desjardins AE 2006 Fusarium mycotoxins chemistry genetics and biology The American
Phytopathological Society St Paul Minnesota USA p 260
Desjardins AE Proctor RH 2007 Molecular biology of Fusarium mycotoxins International
Journal of Food Microbiology 11947-50
Desmond OJ Manners JM Stephens AE MacLean DJ Schenk PM Gardiner DM Munn
AL Kazan K 2008 The Fusarium mycotoxin deoxynivalenol elicits hydrogen peroxide
production programmed cell death and defence responses in wheat Molecular Plant
Pathology 9435-445
Dill-Macky R Jones RK 2000 The effect of previous crop residues and tillage on Fusarium
head blight of wheat Plant Disease 8471ndash76
DrsquoMello JPF Macdonald AMC Dijksma WTP 1998 3-Acetyl deoxynivalenol and esterase
production in a fungicide-insensitive strain of Fusarium culmorum Mycotoxin Research
149ndash18
DrsquoMello JPF Placinta CM Macdonald AMC 1999 Fusarium mycotoxins a review of
global implications of animal health welfare and productivity Animal Feed Science and
Technology 80183-205
Doohan FM Brennan J Cooke BM 2003 Influence of climatic factors on Fusarium species
pathogenic to cereals European Journal of Plant Pathology 109755ndash768
183
Dyer RB Kendra DF Brown DW 2006 Real-time PCR assay to quantify Fusarium
graminearum wild-type and recombinant mutant DNA in plant material Journal of
Microbiological Methods 67534ndash542
Erecinska M Wilson DF 1981 Inhibitors of cytochrome c oxidase In inhibitors of
mitochondrial function edited by M Erecinska and DF Wilson Pergamon Press Ltd
Oxford pp 145-160
Eriksen GS 2003 Metabolism and toxicity of trichothecene Doctoral thesis Swedish
University of Agricultural Sciences Uppsala Sweden
Eriksen GS Pettersson H Lundh T 2004 Comparative cytotoxicity of deoxynivalenol
nivalenol their acetylated derivatives and de-epoxy metabolites Food Chemistry and
Toxicology 42619-624
Eudes F Comeau A Rioux S Collin J 2000 Phytotoxicity of eight mycotoxins associated
with Fusarium in wheat head blight Canadian Journal of Plant Pathology 22286ndash292
FAO (Food and Agriculture Organization) 1991 Food Nutrition and Agriculture Food for
the Future volume 1 FAO Rome
FAO (Food and Agriculture Organization) 2001 FNP73 Manual of the application of
HACCP system in mycotoxin prevention and control FAO Food and Nutrition paper no
73 FAO Rome
Fedorak PM Grbic-Galic D 1991 Aerobic microbial cometabolism of benzothiophene and
3-methylbenzothiophene Applied and Environmental Microbiology 57932-940
Felsenstein J 1989 PHYLIP - Phylogeny Inference Package (Version 32) Cladistics 5164-
166
184
Fernando WGD Amarasinghe C Demoz B Surujdeo‐Maharaj S Gauthier V Guerrieri G
Al‐Taweel K Brucircleacute‐Babel A Tamburic‐Illincic L Gilbert J 2011 What do we know of
the Fusarium graminearum chemotypes their epidemiology genetics and infection
process in Canada In Proc 7th
Canadian Workshop on Fusarium Head Blight edited by
J Gilbert A Tekauz N Sweetland and K Slusarenko Winnipeg Manitoba Canada p 45
Fierer N Jackson RB 2006 The diversity and biogeography of soil bacterial community
Proceedings of the National Academy of Sciences 103626-631
Fink-Gremmels J 1989 The significance of mycotoxin assimilation for meat animals
Deutsche Tierarztliche Wochenschrift 96360-363
Flint DH Emptage MH Finnegan MG Fu W Johnson MK 1993 The role and properties of
the iron-sulfur cluster in Escherichia coli dihydroxy-acid dehydratase Journal of
Biological Chemistry 26814732ndash14742
Foroud NA Eudes F 2009 Trichothecenes in cereal grains International Journal of
Molecular Science 10147-173
Forsell JH Jensen R Tai JH Witt M Lin WS Pestka JJ 1987 Comparison of acute
toxicities of deoxynivalenol (vomitoxin) and 15-acetyldeoxynivalenol in the B6C3F1
mouse Food and Chemical Toxicology 25155ndash162
Frey M Stettner C Gierl A 1998 A general method for gene isolation in tagging approaches
amplification of insertion mutagenised sites (AIMS) The Plant Journal 13717-721
Freeman GG Morrison RI 1949 The isolation and chemical properties of trichothecin an
antifungal substance from Trichothecium roseum Link Biochemical Journal 441ndash5
185
Fuchs E Binder EM Heidler D Krska R 2002 Structural characterization of metabolites
after the microbial degradation of type A trichothecenes by the bacterial strain BBSH 797
Food Additives and Contaminants 19379ndash386
Gabor EM Alkema WBL Janssen DB 2004 Quantifying the accessibility of the
metagenome by random expression cloning techniques Environmental Microbiology
6879ndash886
Gadd GM Griffiths AJ 1978 Microorganisms and heavy metal toxicity Microbial Ecology
4303ndash317
Gale LR Bryant JD Calvo S Giese H 2005 Chromosome complement of the fungal plant
pathogen Fusarium graminearum based on genetic and physical mapping and cytological
observations Genetics 171985ndash1001
Gao LY Pak M Kish R Kajihara K Brown EJ 2006 A mycobacterial operon essential for
virulence in vivo and invasion and intracellular persistence in macrophages Infection and
Immunity 741757ndash1767
Gareau MG Wine E Reardon C et al 2010 Probiotics prevent death caused by Citrobacter
rodentium infection in neonatal mice Journal of Infectious Diseases 20181e91
Gareis M Bauer J Thiem J Plank G Grabley S Gedek B 1990 Cleavage of
zearalenoneglycoside a masked mycotoxin during digestion in swine Journal of
Veterinary Medicine 37236-240
Geng L Xin W Haung DW Feng G 2006 A universal cloning vector using vaccinia
topoisomerase I Molecular Biotechnology 3323-28
186
Golkari S Gilbert J Ban T Procunier JD 2009 QTLspecific microarray gene expression
analysis of wheat resistance to Fusarium head blight in Sumai-3 and two susceptible
NILs Genome 52409ndash418
Goodstadt L Ponting CP 2004 Vitamin K epoxide reductase homology active site and
catalytic mechanism Trends in Biochemical Sciences 29289-292
Goswami RS Kistler HC 2004 Heading for disaster Fusarium graminearum on cereal
crops Molecular Plant Pathology 5515-525
Gouze ME Laffitte J Printon P Dedieux G Galinier A Thouvenot JP Loiseau N Oswald
IP Galtier P 2007 Effect of subacute oral doses of nivalenol on immune and metabolic
defence systems in mice Veterinary Research 38635ndash646
Guan S He J Young JC Zhu H Li X Ji C Zhou T 2009 Transformation of trichothecene
mycotoxins by microorganisms from fish digesta Aquaculture 290290ndash295
Gupta R Beg QK Lorenz P 2002 Bacterial alkaline proteases molecular approaches and
industrial applications Applied Microbiology and Biotechnology 5915ndash32
Haberle J Schweizer G Schondelmaier J Zimmermann G Hart L 2009 Mapping of QTL
for resistance against Fusarium head blight in the winter wheat population
PelikanBussardNing8026 Plant Breeding 12827ndash35
Hall BG 2004 Predicting the evolution of antibiotic resistance genes Nature Reviews
Microbiology 2430-435
Han JI et al 2011 Complete genome sequence of the metabolically versatile plant growth-
promoting endophyte Variovorax paradoxus S110 Journal of Bacteriology 1931183ndash
1190
187
Harvey RB Edrington TS Kubena LF Elissalde MH Corrier DE Rottinghaus GE 1995
Effect of aflatoxin and diacetoxyscirpenol in ewe lambs Bulletin of Environmental
Contamination and Toxicology 54325-330
Harvey RB Edrington TS Kubena LF Elissalde MH Casper HH Rottinghaus GE Turk JR
1996 Effects of dietary fumonisin B1-containing culture material deoxynivalenol-
contaminated wheat or their combination on growing barrows American Journal of
Veterinary Research 571790-1794
He J 2007 Microbial strategies for management of Fusarium graminearum and
transformation of deoxynivalenol Master of Science Thesis University of Guelph
Guelph Ontario Canada
He K Vines L Pestka JJ 2010a Deoxynivalenol-induced modulation of microRNA
expression in RAW 2647 macrophages-A potential novel mechanism for translational
inhibition Toxicologist 114310
He J Zhou T Young JC Boland GJ Scott PM 2010b Chemical and biological
transformations for trichothecene mycotoxins in human and animal food chains A review
Trends in Food Science and Technology 2167-76
He J Zhou T 2010 Patented techniques for detoxification of mycotoxins in feeds and food
matrices Rec Pat Food Nutr Agric 296ndash104
He P Young LG Forsberg C 1992 Microbial transformation of deoxynivalenol (vomitoxin)
Applied and Environmental Microbiology 583857-3863
Hell K Cardwell KF Setamou M Poehling H-M 2000 The influence of storage practices on
aflatoxins contamination in maize in four agroecological zones in Benin West Africa
Journal of Stored Product Research 36365ndash82
188
Henne A Schmitz RA Bomeke M Gottschalk G Daniel R 2000 Screening of
environmental DNA libraries for the presence of genes conferring lipolytic activity on
Escherichia coli Applied and Environmental Microbiology 663113ndash3116
Hodges GR Degener CE Barnes WG 1978 Clinical significance of Citrobacter isolates
American Journal of Clinical Pathology 7037ndash40
Hollinger K Ekperigin HE 1999 Mycotoxicosis in food producing animals The Veterinary
Clinics of North America Food Animal Practice 15133-165
Hong Y Kinoshita T 2009 Trypanosome glycosylphosphatidylinositol biosynthesis Korean
Journal of Parasitology 47197ndash204
Hope RJ Aldred D Magan N 2005 Comparison of environmental profiles for growth and
deoxynivalenol production by Fusarium culmorum and F graminearum on wheat grain
Letters in Applied Microbiology 40295ndash300
Horvath RS 1972 Microbial co-metabolism and the degradation of organic compounds in
nature Bacteriological Reviews 36146ndash155
Hsu D Shih LM Zee YC 1994 Degradation of rRNA in Salmonella strains a novel
mechanism to regulate the concentrations of rRNA and ribosomes Journal of
Bacteriology 1764761ndash4765
Huwig A Freimund S Kappeli O Dutler H 2001 Mycotoxin detoxification of animal feed
by different adsorbents Toxicological Letters 122179ndash188
Iordanov MS Pribnow D Magun JL Dinh TH Pearson JA Magun BE 1997 Ribotoxic
stress response activation of the stress activated protein kinase JNK1 by inhibitors of the
peptidyl transferase reaction and by sequence-specific RNA damage to the sarcinricin
loop in the 28S rRNA Molecular and Cellular Biology 173373ndash3381
189
Islam R Zhou T Young JC Goodwin PH Pauls KP 2012 Aerobic and anaerobic de-
epoxidation of mycotoxin deoxynivalenol by bacteria originating from agricultural soil
World Journal of Microbiology and Biotechnology 287-13
Jackson LS Bullerman LB 1999 Effect of processing on Fusarium mycotoxins Advances in
Experimental Medicine and Biology 459243-261
Jansen C von Wettstein D Schafer W Kogel K-H Felk A Maier FJ 2005 Infection patterns
in barley and wheat spikes inoculated with wild-type and trichodiene synthase gene
disrupted Fusarium graminearum Proceedings of the National Academy of Sciences
10216892ndash16897
Jube S Borthakur D 2007 Expression of bacterial genes in transgenic tobacco methods
applications and future prospects Electronic Journal of Biotechnology 10452-467
Kabak B Dobson AD 2009 Biological strategies to counteract the effects of mycotoxins
Journal of Food Protection 722006-2016
Kant P Reinprecht Y Martin CJ Islam R Pauls KP 2011 Integration of biotechnologies
disease resistance pathology and Fusarium In Comprehensive Biotechnology edited by
Moo-Young M Second Edition 4729-743
Kaplan CW Kitts CL 2004 Bacterial succession in a petroleum land treatment unit Applied
and Environmental Microbiology 701777-1786
Kaprelyants AS Kell DB 1996 Do bacteria need to communicate with each other for
growth Trends in Microbiology 4237-42
190
Kariholu U Rawal J Namnyak S 2009 Neonatal Citrobacter koseri meningitis and brain
abscess The Internet Journal of Pediatrics and Neonatology 10
(httpwwwispubcomjournalthe-internet-journal-of-pediatrics-and-
neonatologyvolume-10-number-1neonatal-citrobacter-koseri-meningitis-and-brain-
abscesshtml) (accessed on April 19 2012)
Karlovsky P 1999 Biological detoxification of fungal toxins and its use in plant breeding
feed and food production Natural Toxins 71ndash23
Karlovsky P 2011 Biological detoxification of the mycotoxin deoxynivalenol and its use in
genetically engineered crops and feed additives Applied Microbiology and
Biotechnology 91491-504
Kavanagh KL Jornvall H Persson B Oppermann U 2008 Medium and short-chain
dehydrogenasereductase gene and protein families the SDR superfamily functional and
structural diversity within a family of metabolic and regulatory enzymes Cellular and
Molecular Life Sciences 653895ndash3906
Khera KS Whalen C Angers G Vesonder RF Kuiper-Goodman T 1982 Embryotoxicity of
4-deoxynivalenol (vomitoxin) in mice Bulletin of Environmental Contamination and
Toxicology 29487ndash491
Kikot GE Hours RA Alconada TM 2009 Contribution of cell wall degrading enzymes to
pathogenesis of Fusarium graminearum a review Journal of Basic Microbiology 49
231ndash241
Kim UJ Shizuya H Jong PJ Birren B Simon MI 1992 Stable propogation of cosmid sized
human DNA inserts in a F factor based vector Nucleic Acids Research 201083-1065
191
Kimura M Kaneko I Komiyama M Takatsuki A Koshino H Yoneyama K Yamaguchi I
1998 Trichothecene 3-O-acetyltransferase protects both the producing organism and
transformed yeast from related mycotoxins cloning and characterization of Tri101
Journal of Biological Chemistry 2731654-1661
Kimura M Anzai H Yamaguchi I 2001 Microbial toxins in plantndashpathogen interactions
biosynthesis resistance mechanisms and significance The Journal of General and
Applied Microbiology 47149ndash160
Kimura M Tokai T Takahashi-Ando N Ohsato S Fujimura M 2007 Molecular and genetics
studies of Fusarium trichothecene biosynthesis pathways genes and evolution
Bioscience Biotechnology and Biochemistry 712105-2123
King RR McQueen RE Levesque D Greenhalgh R 1984 Transformation of deoxynivalenol
(vomitoxin) by rumen microorganisms Journal of Agricultural and Food Chemistry
32181ndash1183
Klappenbach JA Dunbar JM Schmidt TM 2000 rRNA operon copy number reflects
ecological strategies of bacteria Applied and Environmental Microbiology 661328ndash
1333
Knietsch A Waschkowitz T Bowien S Henne A Daniel R 2003 Metagenomes of complex
microbial consortia derived from different soils as sources for novel genes conferring
formation of carbonyls from short-chain polyols on Escherichia coli Journal of Molecular
Microbiology and Biotechnology 546ndash56
Kollarczik B Gareis M Hanelt M 1994 In vitro transformation of the Fusarium mycotoxins
deoxynivalenol and zearalenone by the normal gut microflora of pigs Natural Toxins
2105ndash110
192
Koonin EV Wolf YI Aravind L 2001 Prediction of the archaeal exosome and its
connections with the proteasome and the translation and transcription machineries by a
comparative-genomic approach Genome Research 11240ndash252
Kouadio JH Dano SD Moukha S Mobio TA Creppy EE 2007 Effects of combinations of
Fusarium mycotoxins on the inhibition of macromolecular synthesis malondialdehyde
levels DNA methylation and fragmentation and viability in Caco-2 cells Toxicon
49306ndash317
Krause L Diaz NN Bartels D Edwards RA Puumlhler A et al 2006 Finding novel genes in
bacterial communities isolated from the environment Bioinformatics 22281ndash289
Kubena LF Huff WE Harvey RB Phillips TD Rottinghaus GE 1989 Individual and
combined toxicity of deoxynivalenol and T-2 toxin in broiler chicks Poultry Science
68622ndash626
Kubena LF Harvey RB Huff WE Corrier DE Phillips TD Rottinghaus GE 1990 Efficacy
of hydrated sodium calcium aluminosilicate to reduce the toxicity of aflatoxin and T-2
toxin Poultry Science 691078ndash1086
Kubena LF Harvey RB Huff WE Elissalde MH Yersin AG Phillips TD Rottinghaus GE
1993 Efficacy of hydrated sodium calcium aluminosilicate to reduce the toxicity of
aflatoxin and diacetoxyscirpenol Poultry Science 7251ndash59
Kubena LF Edrington TS Harvey RB Buckley SA Phillips TD Rottinghaus GE Casper
HH 1997 Individual and combined effects of fumonisin B1 present in Fusarium
moniliforme culture material and T-2 toxin or deoxynivalenol in broiler chicks Poultry
Science 761239ndash1247
193
Kumar RA Gunasekaran P Lakshmanan M 1999 Biodegradation of tannic acid by
Citrobacter freundii isolated from a tannery effluent Journal of Basic Microbiology
39161-168
Kushiro M 2008 Effects of milling and cooking processes on the deoxynivalenol content in
wheat International Journal Molecular Science 92127-2145
Lane DJ 1991 16S23S rRNA sequencing In Nucleic acid techniques in bacterial
systematic edited by Stackebrandt E and Goodfellow M Wiley J and Sons Inc New
York pp 115-175
Lancova K Hajslova J Poustka J Krplova A Zachariasova M Dostalek P Saschambula L
2008 Transfer of Fusarium mycotoxins and ldquomaskedrdquo deoxynivalenol (deoxynivalenol-3-
glucoside) from field barley through malt to beer Food Additives and Contaminants
25732ndash744
Langmead B Trapnell C Pop M Salzberg SL 2009 Ultrafast and memory-efficient
alignment of short DNA sequences to the human genome Genome Biology 10R25
Larsen JC Hunt J Perrin I Ruckenbauer P 2004 Workshop on trichothecenes with a focus
on DON summary report Toxicological Letters 1531-22
Lee T Oh DW Kim HS Lee J Kim YH Yun SH Lee YW 2001 Identification of
deoxynivalenol- and nivalenol-producing chemotypes of Gibberella zeae by using PCR
Applied and Environmental Microbiology 672966ndash2972
Lee S-W Won K Lim HK Kim J-C Choi GJ Cho KY 2004 Screening for novel lipolytic
enzymes from uncultured soil microorganisms Applied and Environmental Microbiology
65720-726
194
Li L Christian J Stoeckert J David SR 2003 OrthoMCL Identification of Ortholog Groups
for Eukaryotic Genomes Genome Research 132178-2189
Li T Chang CY Jin DY Lin PJ Khvorova A Stafford DW 2004 Identification of the gene
for vitamin K epoxide reductase Nature 427541ndash544
Li Y 2007 Sensory signal transduction in the vagal primary afferent neurons Current
Medicinal Chemistry 142554ndash2563
Li MX Pestka JJ 2008 Comparative induction of 28S ribosomal RNA cleavage by ricin and
the trichothecenes deoxynivalenol and T-2 toxin in the macrophage Toxicological
Science 10567ndash78
Li R Li Y Kristiansen K Wang J 2008 SOAP short oligonucleotide alignment program
Bioinformatics 24713-4
Li W Schulman S Dutton RJ Boyd D Beckwith J Rapoport TA 2010 Structure of a
bacterial homologue of vitamin K epoxide reductase Nature 463507-512
Lichstein HC Malcolm HS 1943 Studies of the effect of sodium azide on microbic growth
and respiration Journal of Bacteriology 47221ndash230
Limay-Rios V Schaafsma AW 2011 2010 Update on the development of an integrated
mycotoxin management system in Ontario corn 30th
Centralia Swine Research Update
Kirkton Ontario Canada
Llorens A Hinojo MJ Mateo R Gonzalez-Jaen MT Valle- Algarra FM Logrieco A Jimenez
M 2006 Characterization of Fusarium spp isolates by PCR-RFLP analysis of the
intergenic spacer region of the rRNA gene (rDNA) International Journal of Food
Microbiology 106287ndash306
195
Lombaert GA Pellaers P Roscoe V Mankotia M Neil R Scott PM 2003 Mycotoxins in
infant cereal foods from the Canadian retail market Food Additives and Contaminants
20494ndash504
Luengo JM Garciacutea JL Olivera ER 2001 The phenylacetyl-CoA catabolon a complex
catabolic unit with broad biotechnological applications Molecular Microbiology
391434ndash1442
Luo X 1994 Food poisoning caused by Fusarium toxins In Proceedings of the Second
Asian Conference on Food Safety ILSI Thailand pp 129ndash136
Mabagala RB 1997 The effect of populations of Xanthomonas campestris pv phaseoli in
bean reproductive tissues on seed infection of resistant and susceptible bean genotypes
European Journal of Plant Pathology 103175ndash181
Madhyastha MS Marquardt RR Abramson D 1994 Structurendashactivity relationships and
interactions among trichothecene mycotoxins as assessed by yeast bioassay Toxicon
321147ndash1152
Maier FJ Miedaner T Hadeler B Felk A Salomon S Lemmens M Kassner H Schaumlfer W
2006 Involvement of trichothecenes in fusarioses of wheat barley and maize evaluated
by gene disruption of the trichodiene synthase (Tri5) gene in three field isolates of
different chemotype and virulence Molecular Plant Pathology 7449ndash461
Malojcic G Owen RL Grimshaw JP Glockshuber R 2008 Preparation and structure of the
charge-transfer intermediate of the transmembrane redox catalyst DsbB FEBS Letters
5823301ndash3307
196
Manoharan M Dahleen LS Hohn TM Neate SM Yu XH Alexander NJ et al 2006
Expression of 3-OH trichothecene acetyltransferase in barley (Hordeum vulgare L) and
effects on deoxynivalenol Plant Science 171699ndash706
Maresca M Mahfoud R Garmy N Fantini J 2002 The mycotoxin deoxynivalenol affects
nutrient absorption in human intestinal epithelial cells The Journal of Nutrition
1322723ndash2731
Martins ML Martins HM 2002 Influence of water activity temperature and incubation time
on the simultaneous production of deoxynivalenol and zearalenone in corn (Zea mays) by
Fusarium graminearum Food Chemistry 79315ndash318
Masuda D Ishida M Yamaguchi K Yamaguchi I Kimura M Nishiuchi T 2007 Phytotoxic
effects of trichothecenes on the growth and morphology of Arabidopsis thaliana Journal
of Experimental Botany 581617ndash1626
Matsumoto E Kawanaka Y Yun SJ Oyaizu H 2008 Isolation of dieldrin- and endrin-
degrading bacteria using 12-epoxycyclohexane as a structural analog of both compounds
Applied Microbiology and Biotechnology 801095-1103
McLean M 1996 The phytotoxicity of Fusarium metabolites an update since 1989
Mycopathologia 133163ndash179
Merhej J Richard-Forget F Barreau C 2011 Regulation of trichothecene biosynthesis in
Fusarium recent advances and new insights Applied Microbiology and Biotechnology
91519ndash528
Midwest Plan Service 1987 Grain Drying Handling Storage Handbook Ames Iowa Iowa
State University 2nd edition
197
Miller SS Reid LM Harris LJ 2007 Colonization of maize silks by Fusarium graminearum
the causative organism of Gibberella ear rot Canadian Journal of Botany 85369-376
Minervini F Fornelli F Flynn KM 2004 Toxicity and apoptosis induced by mycotoxins
nivalenol deoxynivalenol and fumonisin B1 in a human erythroleukemia cell line
Toxicology In Vitro 1821ndash28
Mitterbauer R Poppenberger B RaditschnigA Lucyshyn D Lemmens M Glossl J Adam G
2004 Toxin-dependent utilization of engineered ribosomal protein L3 limits trichothecene
resistance in transgenic plants Plant Biotechnology Journal 2329ndash340
Mocali S Benedetti A 2010 Exploring research frontiers in microbiology the challenge of
metagenomics in soil microbiology Research Microbiology 161497-505
Morozkina EV Zvyagilskaya RA 2007 Nitrate reductases structure functions and effect of
stress factors Biochemistry 721151ndash1160
Munkvold GP 2003 Cultural and genetic approaches to managing mycotoxins in maize
Annual Review of Phytopathology 4199-116
Mundy R MacDonald TT Dougan G Frankel G Wiles S 2005 Citrobacter rodentium of
mice and man Cellular Microbiology 71697ndash1706
Nakamura N Inoue N Watanabe R Takahashi M Takeda J Stevens VL Kinoshita T 1997
Expression cloning of PIG-L a candidate N-acetylglucosaminyl-phosphatidylinositol
deacetylase The Journal of Biological Chemistry 27215834ndash15840
Nganje WE Bangsund DA Leistritz FL Wilson WW Tiapo NM 2002 Estimating the
economic impact of a crop disease the case of Fusarium head blight in US wheat and
barley National Fusarium Head Blight Forum Proceedings East Lansing Michigan State
University pp 275ndash281
198
Nielsen C Lippke H Didier A Dietrich R Martlbauer E 2009 Potential of deoxynivalenol
to induce transcription factors in humanhepatoma cells Molecular Nutrition and Food
Research 53479ndash491
Nielsen JKS Vikstroumlm AC Turner P Knudsen LE 2011 Deoxynivalenol transport across
the human placental barrier Food and Chemical Toxicology 492046-2052
Nishiuchi T Masuda D Nakashita H Ichimura K Shinozaki K Yoshida S et al 2006
Fusarium phytotoxin trichothecenes have an elicitor-like activity in Arabidopsis thaliana
but the activity differed significantly among their molecular species Molecular Plantndash
Microbe Interactions 19512ndash520
Ohsato S Ochiai-Fukuda T Nishiuchi T Takahashi-Ando N Koizumi S Hamamoto H
Kudo T Yamaguchi I Kimura M 2007 Transgenic rice plants expressing trichothecene
3-O-acetyltransferase show resistance to the Fusarium phytotoxin deoxynivalenol Plant
Cell Reports 26531-538
Okubara PA Blechl AE McCormick SP Alexander NJ Dill-Macky R Hohn TM 2002
Engineering deoxynivalenol metabolism in wheat through the expression of a fungal
trichothecene acetyltransferase gene Theoritical and Applied Genetics 10674-83
Ong CL Beatson SA Totsika M Forestier C McEwan AG Schembri MA 2010 Molecular
analysis of type 3 fimbrial genes from Escherichia coli Klebsiella and Citrobacter
species BMC Microbiology 10183
Osborne LE Stein JM 2007 Epidemiology of Fusarium head blight on small-grain cereals
International Journal of Food Microbiology 119103-108
Page RDM 1996 TREEVIEW An application to display phylogenetic trees on personal
computers Computer Applications in the Biosciences 12357-358
199
Parkhill J Dougan G James KD Thomson NR Pickard D Wain J Churcher C Mungall KL
Bentley SD Holden MT et al 2001 Complete genome sequence of a multiple drug
resistant Salmonella enterica serovar Typhi CT18 Nature 413848ndash852
Paterson RRM Lima N 2010 How will climate change affect mycotoxins in food Food
Research International 431902ndash14
Paul PA El-Allaf SM Lipps PE Madden LV 2004 Rain splash dispersal of Gibberella zeae
within wheat canopies in Ohio Phytopathology 941342-1349
Pestka JJ Lin WS Miller ER 1987 Emetic activity of the trichothecene 15-
acetyldeoxynivalenol in pigs Food and Chemical Toxicology 25855ndash858
Pestka JJ Smolinski AT 2005 Deoxynivalenol Toxicology and potential effects on humans
Journal of Toxicology Environmental Health 839ndash69
Pestka JJ 2007 Deoxynivalenol toxicity mechanisms and animal health risks Animal Feed
Science and Technology 137283-298
Pestka JJ 2008 Mechanisms of deoxynivalenol-induced gene expression and apoptosis Food
Additives and Contaminants 251128-1140
Pestka J2010 Toxicological mechanisms and potential health effects of deoxynivalenol and
nivalenol World Mycotoxin Journal 3323ndash347
Petersohn A Brigulla M Haas S Hoheisel JD Voumllker U Hecker M 2001 Global Analysis
of the General Stress Response of Bacillus subtilis Journal of Bacteriology 1835617-
5631
200
Petty NK Bulgin R Crepin VF Cerdeno-Tarraga AM Schroeder GN Quail MA Lennard N
Corton C Barron A Clark L Toribio AL Parkhill J Dougan G Frankel G Thomson NR
2010 The Citrobacter rodentium genome sequence reveals convergent evolution with
human pathogenic Escherichia coli Journal of Bacteriology 192525ndash538
Petty NK Feltwell T Pickard D Clare S Toribio AL et al 2011 Citrobacter rodentium is
an unstable pathogen showing evidence of significant genomic flux PLoS Pathology
7e1002018 doi101371journalppat1002018
Pittet A 2001 Natural occurrence of mycotoxins in foods and feeds A decade in review In
Mycotoxins and phytotoxins in perspective at the turn of the millennium edited by De
Koe WJ Samson RA Van Egmond HP Gilbert J Sabino M W J de Koe Wageningen
Netherlands pp 153ndash172
Placinta CM DrsquoMello JPF Macdonald AMC 1999 A review of worldwide contamination of
cereal grains and animal feed with Fusarium mycotoxins Animal Feed Science and
Technology 7821ndash37
Plank B Schuh M Binder EM 2009 Investigations on the effect of two feed additives
Biomin BBSH 797 and Mycofix Plus (R) 3E as detoxificants of DON contaminated feed
of piglets Wien Tierarztl Monatssch 9655ndash71
Poapolathep A Ohtsuka R Kiatipattanasakul W Ishigami N Nakayama H Doi K 2002
Nivalenol-induced apoptosis in thymus spleen and Peyerrsquos patches of mice Experimental
and Toxicological Pathology 53441ndash446
201
Poppenberger B Berthiller F Lucyshyn D Sieberer T Schuhmacher R Krska R Kuchler K
Gloumlssl J Luschnig C Adam G 2003 Detoxification of the Fusarium mycotoxin
deoxynivalenol by a UDP-glucosyltransferase from Arabidopsis thaliana Journal of
Biological Chemistry 27847905ndash47914
Prelusky DB Trenholm HL 1993 The efficacy of various classes of anti-emetics in
preventing deoxynivalenol-induced vomiting in swine Natural Toxins 1296ndash302
Proctor RH Hohn TM McCormick SP 1995 Reduced virulence of Gibberella zeae caused
by disruption of a trichothecene toxin biosynthetic gene Molecular Plant-Microbe
Interactions 8593-601
Puchenkova SG 1996 Enterobacteria in areas of water along the Crimean Coast
Mikrobiolohichnyĭ zhurnal 58 3-7
Qi BW Hao BL 2004 Whole-proteome prokaryote phylogeny without sequence alignment
a K-string composition approach Journal of Molecular Evolution 581ndash11
Quarta A Mita G Haidukowski M Logrieco A Mule G Visconti A 2006 Multiplex PCR
assay for the identification of nivalenol 3- and 15-acetyl-deoxynivalenol chemotypes in
Fusarium FEMS Microbiological Letters 2597ndash13
Rehr B Klemme JH 1989 Formate dependent nitrate and nitrite reduction to ammonia by
Citrobacter freundii and competition with denitrifying bacteria Antonie van Leeuwenhoek
Journal of Microbiology 56311ndash321
Rocha O Ansari K Doohan FM 2005 Effects of trichothecene mycotoxins on eukaryotic
cells A review Food Additives and Contaminants 22369ndash378
Roscoe V Lombaert GA Huzel V et al 2008 Mycotoxins in breakfast cereals from the
Canadian retail market a 3-year survey Food Additives and Contaminants 25347ndash355
202
Rotter BA Prelusky DB Pestka JJ 1996 Toxicology of deoxynivalenol (vomitoxin) Journal
of Toxicology and Environment Health 481ndash34
Ryu JC Ohtsubo K Izurniyarna N Nakamura K Tanaka T Yamarnura H Ueno Y 1988
The acute and chronic toxicities of nivalenol in mice Fundamental and Applied
Toxicology 1138ndash47
Saier MH Roseman S 1972 Inducer exclusion and repression of enzyme synthesis in
mutants of Salmonella typhimurium defective in enzyme I of the phosphoenolpyruvate
sugar phosphotransferase system The Journal of Biological Chemistry 247972ndash975
Saier MHJ Tran CV Barabote RD 2006 TCDB the transporter classification database for
membrane transport protein analyses and information Nucleic Acids Research 34D181ndash
186
Saitou N Nei M 1987 The neighbor-joining method a new method for reconstructing
phylogenetic trees Molecular Biology and Evolution 4406ndash425
Savelieva O Kotova I Roelofsen W Stams AJM Netrusov A 2004 Utilization of
aminoaromatic acids by a methanogenic enrichment culture and by a novel Citrobacter
freundii strain Archives of Microbiology 181163ndash170
Schaafsma AW Tamburic-Ilinic L Miller JD Hooker DC 2001 Agronomic considerations
for reducing deoxynivalenol in wheat grain Canadian Journal of Plant Pathology 23279ndash
285
Schaafsma AW 2002 Economic changes imposed by mycotoxins in food grains case study
of deoxynivalenol in winter wheat Advances in Experimental Medicine and Biology
504271ndash276
203
Scharf C Riethdorf S Ernst H Engelmann S Voumllker U Hecker M 1998 Theoredoxin is an
essential protein induced by heat shock and other stresses in Bacillus subtilis Journal of
Bacteriology 1801869-1877
Schatzmayr G Zehner F Taubel M Schatzmayr D Klimitsch A Loibner AP Binder EM
2006 Microbiologicals for deactivating mycotoxins Molecular Nutrition and Food
Research 50543-551
Schollenberger M Muumlller H-M Ruumlfle R Suchy S Plank S Drochner W 2006 Natural
occurrence of 16 Fusarium toxins in grains and feedstuffs of plant origin from Germany
Mycopathologia 16143ndash52
Scientific Committee for Food 2002 Opinion on Fusarium toxins Part 6 Group evaluation
of T-2 toxin HT-2 toxin nivalenol and deoxynivalenol (available at http
wwweuropaeuintcommfoodfsscscfout123_enpdf)
Schulman S Wang B Li W Rapoport TA 2010 Vitamin K epoxide reductase prefers ER
membrane-anchored thioredoxin-like redox partners Proceedings of the National
Academy of Sciences 10715027ndash15032
Scott PM 1997 Multi-year monitoring of Canadian grains and grain-based foods for
trichothecenes and zearalenone Food Additives and Contaminants 14333-339
Seitz LM Bechtel DB 1985 Chemical physical and microscopal studies of scab-infected
hard red winter wheat Journal of Agricultural and Food Chemistry 33373-7
Sergent T Parys M Garsou S Pussemier L Schneider YJ Larondelle Y 2006
Deoxynivalenol transport across human intestinal Caco-2 cells and its effects on cellular
metabolism at realistic intestinal concentrations Toxicological Letters 164167ndash176
204
Sharma J Fulekar MH 2009 Potential of Citrobacter freundii for bioaccumulation of
heavy metalndashcopper Biology and Medicine 17-14
Sherif SO Salama EE Abdel-Wahhab MA 2009 Mycotoxins and child health the need for
health risk assessment International Journal of Hygiene and Environmental Health
212347ndash368
Shifrin VI Anderson P 1999 Trichothecene mycotoxins trigger a ribotoxic stress response
that activates c-Jun N-terminal kinase and p38 mitogen-activated protein kinase and
induces apoptosis The Journal of Biological Chemistry 27413985ndash13992
Shima J Takase S Takahashi Y Iwai Y Fujimoto H Yamazaki M Ochi K 1997 Novel
detoxification of the trichothecene mycotoxin deoxynivalenol by a soil bacterium isolated
by enrichment culture Applied and Environmental Microbiology 633825ndash3830
Singh J Behal A Singla N Joshi A Birbian N et al 2009 Metagenomics Concept
methodology ecological inference and recent advances Biotechnology Journal 4480ndash
494
Smith TK McMillan EG Castillo JB 1997 Effect of feeding blends of Fusarium mycotoxin-
contaminated grains containing deoxynivalenol and fusaric acid on growth and feed
consumption of immature swine Journal of Animal Science 752184ndash2191
Smits WK Dubois JYF Bron S van Dijl JM Kuipers OP 2005 Tricksy business
transcriptome analysis reveals the involvement of thioredoxin in redox homeostasis
oxidative stress sulfur metabolism and cellular differentiation in Bacillus subtilis Journal
of Bacteriology 1873921ndash3930
205
Sprando RL Collins TFX Black TN Olejnik N Rorie JI Eppley RM Ruggles DI 2005
Characterization of the effect of deoxynivalenol on selected male reproductive endpoints
Food Chemistry and Toxicology 43623ndash635
Starkey DE Ward TJ Aoki T Gale LR Kistler HC Geiser DM Suga H Toacuteth B Varga J
ODonnell K 2007 Global molecular surveillance reveals novel Fusarium head blight
species and trichothecene toxin diversity Fungal Genetics and Biology 441191ndash1204
Stevenson BS Schmidt TM 1997 Growth rate-dependent expression of RNA from plasmid-
borne rRNA operons in Escherichia coli Journal of Bacteriology 1801970ndash1972
Stothard P Wishart DS 2005 Circular genome visualization and exploration using
CGView Bioinformatics 21537ndash539
Suenaga H Ohnuki T Miyazaki K 2007 Functional screening of a metagenomic library for
genes involved in microbial degradation of aromatic compounds Environmental
Microbiology 92289-97
Surai P Mezes M Fotina TI Denev SD 2010 Mycotoxins in Human Diet A Hidden
Danger In Modern Dietary Fat Intakes in Disease Promotion Edited by F De Meester et
al Nutrition and Health DOI 101007978-1-60327-571-2_18
Sutton JC 1982 Epidemiology of wheat head blight and maize ear rot caused by Fusarium
graminearum Canadian Journal of Plant Pathology 4195
Swaving J De Bont JAM Westphal A De Kok A 1996 A novel type pyridine nucleotide-
disulfide oxidoreductase is essential for NAD+- and NADPH-dependent degradation of
epoxyalkanes by Xanthobacter strain Py2 Journal of Bacteriology 1786644ndash6
206
Swanson SP Rood JHD Behrens JC Sanders PE 1987 Preparation and characterization of
the deepoxy trichothecenes deepoxy HT-2 deepoxy T-2 triol deepoxy T-2 tetraol
deepoxy 15-monoacetoxyscirpenol and deepoxy scirpentriol Applied and Environmental
Microbiology 532821ndash2826
Swanson SP Helaszek C Buck WB Rood JHD Haschek WM 1988 The role of intestinal
microflora in the metabolism of trichothecene mycotoxins Food Chemistry and
Toxicology 26823ndash829
Tian F Yu Y Chen B Li H Yao Y-F Guo X-K 2009 Bacterial archaeal and eukaryotic
diversity in Arctic sediment as revealed by 16S rRNA and 18S rRNA gene clone libraries
analysis Polar Biology 3293-103
Tatusov RL Galperin MY Natale DA Koonin EV et al 2000 Artemis sequence
visualization and annotation Bioinformatics 16944-945
Trail F Haixin X Loranger R Gadoury D 2002 Physiological and environmental aspects of
ascospore discharge in Gibberella zeae (anamorph Fusarium graminearum) Mycologia
94181-189
Tsuge K Ohata Y Shoda M 2001 Gene yerP involved in surfactin self-resistance in
Bacillus subtilis Antimicrobial Agents and Chemotherapy 453566ndash3573
Tucker 2001 The ldquoyellow rainrdquo controversy Lesson for arms control compliance The
Nonproliferation Review 825-42
Ueno Y 1985 The toxicology of mycotoxins Critical Reviews in Toxicology 1499-132
Ueno Y Nakayama K Ishii K Tashiro F Minoda Y Omori T Komagata K 1983
Metabolism of T-2 toxin in Curtobacterium sp strain 114-2 Applied and Environmental
Microbiology 46120ndash127
207
van Elsas JD Speksnijder AJ van Overbeek LS 2008 A procedure for the metagenomics
exploration of disease suppressive soils Journal of Microbiological Methods 75515ndash522
van Ginkel CG de Bont JAM 1986 Isolation and characterization of alkene-utilizing
Xanthobacter spp Archives of Microbiology 145403ndash407
van Lanen SG Shen B 2006 Microbial genomics for the improvement of natural product
discovery Current Opinion in Microbiology 9252ndash260
Varga J Toacuteth B 2005 Novel strategies to control mycotoxins in feeds a review Acta
Veterinaria Hungarica 53189-203
Voigt CA Schafer W Salomon S 2005 A secreted lipase of Fusarium graminearum is a
virulence factor required for infection of cereals The Plant Journal 42364ndash375
Vogelgsang S Hecker A Musa T Dorn B Forrer HR 2011 On farm experiments over five
years in a grain maizewinter wheat rotation Effect of maize residue treatments on
Fusarium graminearum infection and deoxynivalenol contamination in wheat In Proc
7th
Canadian Workshop on Fusarium Head Blight edited by J Gilbert A Tekauz N
Sweetland and K Slusarenko Winnipeg Manitoba Canada p 41
Voumllkl A Vogler B Schollenberger M Karlovsky P 2004 Microbial detoxification of
mycotoxin deoxynivalenol Journal of Basic Microbiology 44147ndash156
Wagacha JM Muthomi JW 2007 Fusarium culmorum Infection process mechanisms of
mycotoxin production and their role in pathogenesis in wheat Crop Protection 26877-
885
Wakulinski W 1989 Phytotoxicity of the secondary metabolites of fungi causing wheat head
fusariosis (head blight) Acta Physiologiae Plantarum 11301ndash306
208
Wang JT Chang SC Chen YC Luh KT 2000 Comparison of antimicrobial susceptibility of
Citrobacter freundii isolates in two different time periods The Journal of Microbiology
Immunology and Infection 33258-62
Wannemacher RJ Stanley L Wiener MD 2000 Trichothecenes mycotoxins In Medical
aspects of chemical and biological warfare edited by Zajtchuk R Washington DC
Department of the Army pp 656-676
Westphal AH Swaving J Jacobs L de Kok A 1998 Purification and characterization of a
flavoprotein involved in the degradation of epoxyalkanes by Xanthobacter Py2 European
Journal of Biochemistry 257160ndash168
Whalen JG Mully TW Enlgish JC 2007 Spontaneous Citrobacter freundii infection in an
immunocompetent patient Archives of Dermatology 143124-5
White 2007 The physiology and biochemistry of prokaryotes Third edition Oxford
University Press New York Oxford p 628
Wilcke WF Morey RV 1995 Natural-air corn drying in the upper Midwest University of
Minnesota Minnesota Extension Service Publication BU-6577-E pp 20
Windels CE 2000 Economic and social impacts of Fusarium headblight changing farms and
rural communities in the Northern Great Plains Phytopathology 9017ndash21
Winzeler EA Shoemaker DD Astromoff A Liang H Anderson K Andre B Bangham R
Benito R Boeke JD Bussey H et al 1999 Functional characterization of the S
cerevisiae genome by gene deletion and parallel analysis Science 285901ndash906
Yang GH Jarvis BB Chung YJ Pestka JJ 2000 Apoptosis induction by the satratoxins and
other trichothecene mycotoxins relationship to ERK p38 MAPK and SAPKJNK
activation Toxicology and Applied Pharmacology 164149ndash160
209
Yang Q Yin GM Guo YL Zhang DF Chen SJ Xu ML 2010 A major QTL for resistance
to Gibberella stalk rot in maize Theoretical and Applied Genetics 121673ndash687
Yazar S Omurtag GZ 2008 Fumonisins trichothecenes and zearalenone in cereals
International Journal of Molecular Science 92062ndash2090
Yoshizawa T Morooka N 1973 Deoxynivalenol and its monoacetate new mycotoxins from
Fusaruium roseum and moldy barley Agricultural and Biological Chemistry 372933ndash
2934
Yoshizawa T Takeda H Ohi T 1983 Structure of a novel metabolite from deoxynivalenol a
trichothecene mycotoxin in animals Agricultural and Biological Chemistry 472133ndash
2135
Yoshizawa T 1993 Red-mold diseases and natural occurrence in Japan In Trichothecenes-
Chemistry Biology and Toxicological Aspects edited by Y Ueno Elsevier Amsterdam
Netherlands p 195-209
Yoshizawa T Sakamoto T Okamoto K 1984 In vitro formation of 3rsquo-hydroxy T-2 and 3rsquo-
hydroxy HT-2 toxins from T-2 by liver homogenates from mice and monkeys Applied
and Environmental Microbiology 47130-134
Young JC Subryan LM Potts D McLaren ME Gobran FH 1986 Reduction in levels of
deoxynivalenol in contaminated wheat by chemical and physical treatment Journal of
Agricultural and Food Chemistry 34461ndash465
Young JC Trenholm HL Friend DW Prelusky DB 1987 Detoxification of deoxynivalenol
with sodium bisulfite and evaluation of the effects when pure mycotoxin or contaminated
corn was treated and given to pigs Journal of Agricultural and Food Chemistry 35259ndash
261
210
Young JC Zhu H Zhou T 2006 Degradation of trichothecene mycotoxins by aqueous
ozone Food Chemistry and Toxicology 44417ndash424
Young JC Zhou T Yu H Zhu H Gong J 2007 Degradation of trichothecene mycotoxins by
chicken intestinal microbes Food Chemistry and Toxicology 45136-143
Yu Z Morrison M 2004 Comparisons of different hypervariable regions of rrs genes for use
in fingerprinting of microbial communities by PCR-denaturing gradient gel
electrophoresis Applied and Environmental Microbiology 704800-4806
Yu H Zhou T Gong J Young C Su X Li X Zhu H Tsao R Yang R 2010 Isolation of
deoxynivalenol-transforming bacteria from the chicken intestines using the approach of
PCR-DGGE guided microbial selection BMC Microbiology 10182
Zeibdawi A Steele M Savoie A 2011 An overview of CFIAs mycotoxin monitoring
program in livestock feeds In Proc 7th
Canadian Workshop on Fusarium Head Blight
edited by J Gilbert A Tekauz N Sweetland and K Slusarenko Winnipeg Manitoba
Canada p 25
Zhang J-B Li H-P Dang F-J Qu B Xu Y-B Zhao C-S Lia Y-C 2007 Determination of the
trichothecene mycotoxins chemotypes and associated geographical distribution and
phylogenetic species of the Fusarium graminearum clade from China Mycological
Research 111967ndash975
Zhao JS Manno D Thiboutot S Ampleman G Hawari J 2007 Shewanella canadensis sp
nov and Shewanella atlantica sp nov manganese dioxide- and hexahydro-135-trinitro-
135-triazine-reducing psychrophilic marine bacteria International Journal of Systemic
Evolution and Microbiology 572155ndash2162
211
Zhao X-Q 2011 Genome-based studies of marine microorganisms to maximize the diversity
of natural products discovery for medical treatments Evidence-Based Complementary
and Alternative Medicine Article ID 384572 doi1011552011384572
ZhaoY Park RD Muzzarelli RA 2010 Chitin deacetylases properties and applications
Marine Drugs 824ndash46
Zhou HR Lau AS Pestka JJ 2003a Role of double-stranded RNA-activated protein kinase R
(PKR) in deoxynivalenol-induced ribotoxic stress response Toxicological Science
74335ndash344
Zhou HR Islam Z Pestka JJ 2003b Rapid sequential activation of mitogen-activated protein
kinases and transcription factors precedes proinflammatory cytokine mRNA expression in
spleen of mice exposed to the trichothecene vomitoxin Toxicological Sciences 72130ndash
142
Zhou T He J Gong J 2008 Microbial transformation of trichothecene mycotoxins World
Mycotoxin Journal 123-30
Zhou G Zhang H He Y He L 2010 Identification of a chitin deacetylase producing bacteria
isolated from soil and its fermentation optimization African Journal of Microbiology
Research 42597-2603

iii
DEDICATIONS
The PhD thesis is dedicated to the departed souls of my mother MOZIBUN NESSA and
father-in-law DR MA AHAD MIAH
iv
ACKNOWLEDGEMENTS
Foremost I am deeply indebted to my advisors Prof K Peter Pauls and Dr Ting Zhou
for the continuous support of my PhD study for their endurance inspiration enthusiasm and
mentoring Their continued guidance has helped me in all aspects of my research and in
writing of this dissertation I sincerely express my gratitude to them for their co-operation
from the beginning to conclusion of the thesis and many other areas This will guide me in my
future professional career
My thesis committee guided me through all these years I would like to express my
gratitude to Prof Paul Goodwin for his insightful comments and advice on my thesis research
Special thanks to Dr Chris Young for teaching me chemistry chemical structures and
interpreting liquid chromatography ultraviolet mass spectra data I am sincerely grateful to
Dion Lepp for helping in analyzing the bacterial genome and sharing his knowledge and
valuable suggestions on writing the genomic chapter
I would like to express my appreciation to staff members of Dr Ting Zhou and Prof
K Peter Pauls who supported me in many aspects during my PhD study I am thankful to
Jianwei He Xiu-Zhen Li Honghui Zhu for collections and preparation of soil samples and
LC-UV-MS analyses of numerous samples Special thanks to Dr David Morris Johnston
Monje for sharing his knowledge discussions and camaraderie at coffee breaks
I wish to acknowledge the Natural Sciences and Engineering Research Council of
Canada (NSERC) for awarding me a prestigious scholarship for my PhD program and
Deanrsquos Tri-Council Scholarship (University of Guelph) which made my life easier during this
study I also would like to convey my thanks to Agriculture and Agri-Food CanadaBinational
v
Agricultural Research and Development Fund (AAFCBARD) project for providing me
financial support
Mike Bryan a very supportive person helped provide solutions to my software issues
that improved my exposure and caliber at conference awards competitions Several student
competition awards honored and inspired me to work hard and lead to the successful
completion of my thesis research I would like to convey my appreciation to the judges of
different student award committees who recognized my work They are Society for Industrial
Microbiology and Biotechnology conference 2010 (USA) Food Safety Research Forum-
Ontario Ministry of Agriculture and Rural Affairs annual meeting 2010 (Canada)
International Mycological Conference 2009 (UK) and Southwestern Ontario Regional
Association of Canadian Phytopathological Society annual meeting 2009-2011 (Canada)
I would like to express my very special thanks to Prof Golam Hafeez Kennedy my
brother-in-law who is like my elder brother In many aspects I am indebt to him for his
cooperations which aided me to achieve my current status
Lastly and most importantly I wish to give my special thanks to my wife Mimi and
lovely daughter Racy whose endless love sacrifice support and encouragement enabled me
to cross the tough long road and made possible to achieve my goal and aspirations Racy
once you told me that one of your desires for me is to finish my PhD which is done now I
know you want to be a scientist which is a great ambition I wish you all the best in your
future aims Racy
vi
TABLE OF CONTENTS
DEDICATIONShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipiii
ACKNOWLEDGEMENTShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipiv
TABLE OF CONTENTShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipvi
LIST OF TABLEShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipix
LIST OF FIGUREShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipx
LIST OF ABBREVIATIONShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipxiii
10 GENERAL INTRODUCTION AND LITERATURE REVIEW helliphelliphelliphelliphelliphelliphelliphellip1
11 Backgroundhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip1
12 Plant pathogenesis and trichothecene productionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip3
13 Biosynthesis of food contaminating TChelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip6
14 Occurrence of TC mycotoxins in food and feed helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip11
15 Significance of deoxynivalenol and trichotheceneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip16
16 Toxic effects of deoxynivalenol and trichotheceneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip19
17 Cellular and molecular mechanisms of DON toxicity in animals and plantshelliphelliphelliphellip22
18 Strategies to control Fusarium head blight and trichotheceneshelliphelliphelliphelliphelliphelliphelliphelliphellip26
19 Isolation of DONTC de-epoxidating micro-organisms and geneshelliphelliphelliphelliphelliphelliphelliphellip28
110 The bacterial species Citrobacter freundiihelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip30
111 Hypotheses and objectiveshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip31
vii
20 AEROBIC AND ANAEROBIC DE-EPOXIDATION OF MYCOTOXIN
DEOXYNIVALENOL BY BACTERIA ORIGINATING FROM AGRICULTURAL
SOIL33
21 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip33
22 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
23 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip37
24 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip45
25 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip62
26 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip64
30 ISOALTION AND CHARACTERIZATION OF A NOVEL SOIL BACTERIUM
CAPABLE OF DETOXIFYING ELEVEN TRICHOTHECENE MYCOTOXINS65
31 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip65
32 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip66
33 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip68
34 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip75
35 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip96
36 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip100
40 CONSTRUCTION AND FUNCTIONAL SCREENING OF GENOMIC DNA
LIBRARY FOR ISOLATION OF DON DE-EPOXYDIZING GENE(S) FROM
BACTERIAL STRAIN ADS47helliphelliphelliphelliphelliphelliphelliphellip102
41 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip102
42 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip103
43 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip104
viii
44 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip112
45 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip118
46 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip121
50 GENOME SEQUENCING CHARACTERIZATION AND COMPARATIVE
ANALYSIS OF A TRICHOTHECENE MYCOTOXIN DETOXIFYING
CITROBACTER FREUNDII STRAIN ADS47helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip122
51 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip122
52 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip123
53 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip125
54 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip129
55 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip162
56 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip168
60 CONCLUSIONhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip171
61 Summary helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip171
62 Research contributions helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip174
63 Future workhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip175
70 REFERENCEShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip176
ix
LIST OF TABLES
Table 1-1 Fusaria produced type-A and type-B trichothecene mycotoxinshelliphelliphelliphelliphelliphelliphellip9
Table 1-2 Worldwide occurrence of deoxynivalenol (DON) in animal feedshelliphelliphelliphelliphellip12
Table 1-3 Occurrence of DON in Canadian grains grains foods and beershelliphelliphelliphelliphelliphelliphellip13
Table 1-4 Legislative maximum tolerated levels of trichothecene mycotoxins in Canadahellip14
Table 1-5 In vitro and in vivo synergistic effects of different food contaminating
mycotoxins helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip18
Table 3-1 Microbial transformation of type-A and type-B tricthothecene mycotoxins
after 72 h of incubation under aerobic conditions and at room temperaturehelliphelliphelliphelliphelliphellip95
Table 4-1 Analysis of Fosmid DNA library for deoxynivalenol (DON) de-epoxidation
capability helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip117
Table 5-1 General features of C freundii strain ADS47 genomehelliphelliphelliphelliphelliphelliphelliphelliphellip132
Table 5-2 Genome comparisons between strain ADS47 and five Citrobacter species141
Table 5-3 Functional categorized gene abundances among the six Citrobacter genomeshellip145
Table 5-4 Unique genes of C freundii strain ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip156
Supplemental Table S5-1 List of unique genes of strain ADS47 gene IDs and their
functional roleshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip169
x
LIST OF FIGURES
Figure 1-1 Structures of trichothecene mycotoxins produced by Fusariahelliphelliphelliphelliphelliphelliphelliphellip7
Figure 1-2 Cellular and molecular mechanisms of deoxynivalenol toxicity in
eukaryotic organismshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip23
Figure 2-1 Mechanism of microbial DON de-epoxidationhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip36
Figure 2-2 LC-UV-MS chromatographic analysis of DON and its de-epoxidated product
dE-DON obtained by culturing with selected soil microbeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip46
Figure 2-3 Time course of relative biotransformation of DON to dE-DON by the initial
soil suspension culture in MSB mediumhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip49
Figure 2-4 Microbial de-epoxidation of DON after five days of incubationhelliphelliphelliphelliphelliphellip51
Figure 2-5 Analysis of microbial population reduction by terminal restriction fragment
length polymorphism (T-RFLP)helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip53
Figure 2-6 Microbial genera identified in the enriched DON de-epoxidizing microbial
culturehelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip56
Figure 2-7 Effects of culture conditions on DON to dE-DON transformation by the
enriched microbial culturehelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip58
Figure 2-8 Microbial DON to dE (de-epoxy)-DON transformation activity under aerobic
and anaerobic conditionshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip61
Figure 3-1 Colony color shape and size of the mycotoxin degrading bacterial strain
ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip76
Figure 3-2 Dendrogram based on 16S-rDNA sequences for strain ADS47 and 11
reference Citrobacter specieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip77
xi
Figure 3-3 Effects of DON concentration on bacterial growth and de-epoxidation
activitieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip79
Figure 3-4 Bacterial growth and DON de-epoxidation capabilities under aerobic and
anaerobic conditions helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip82
Figure 3-5 Effects of media on bacterial growth and DON de-epoxidation activityhellip85
Figure3-6 Effects of temperature on microbial growth and DON de-epoxydation helliphelliphellip87
Figure 3-7 Influence of pH on microbial growth and DON de-epoxydation capabilityhellip89
Figure 3-8 Effects of sodium azide (SA) on bacterial DON de-epoxidation activityhelliphelliphellip91
Figure 3-9 Determination of DON to deepoxy-DON transformation by bacterial culture
filtrate and cell extracthelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip92
Figure 3-10 Determination of microbial de-epoxidation and de-acetylation of a type-A
trichothecene mycotoxin (HT-2 toxin) by LC-UV-MShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip94
Supplemental Figure S3-1 Flow chart depicted the procedures of enrichment and isolation
of DON de-epoxidation bacterial strain ADS47helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip101
Figure 4-1 Schematic overview of the construction and functional screening of Fosmid
DNA libraryhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip105
Figure 4-2 Genetic map of copy controlled Fosmid vector (pCC1FOS)helliphelliphelliphelliphelliphelliphellip108
Figure 4-3 Preparation of 40 kb size insert DNA from a DON de-epoxidizing bacterium113
Figure 4-4 Production of genomic DNA library in E coli EPI300-T1 host strainhelliphelliphelliphellip114
Figure 4-5 Confirmation of the Fosmid DNA library construction in E coli helliphelliphelliphelliphellip 116
Figure 5-1 Co-linearity between C freundii strain ADS47 genome and C koseri helliphelliphellip129
Figure 5-2 Circular genome map of C freundii strain ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphellip133
xii
Figure 5-3 Size distribution of the predicted genes identified in C freundii strain
ADS47helliphelliphellip 135
Figure 5-4 Functional classification of the predicted coding sequences (CDS) of
C freundii strain ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip137
Figure 5-5 Phylogeny of the trichothecene detoxifying C freundii strain ADS47 139
Figure 5-6 Comparisons of coding sequences between C freundii strain ADS47 and
five Citrobacter specieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip143
Figure 5-7 Determination of the locus enterocyte effacement (LEE) operons in
Citrobacter species by comparing C rodentium helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip151
Figure 5-8 Locations of unique genes of C freundii strain ADS467helliphelliphelliphelliphelliphelliphelliphellip157
Figure 5-9 Unique genomic regions and associated genes in strain ADS47 compared
to five Citrobacter specieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip159
xiii
LIST OF ABBREVIATIONS
APCI= atmospheric pressure chemical ionization
AE= attaching and effacing
A= aerobic conditions
ABC= ATP-binding cassette
ACT= artemis comparative tools
AN= anaerobic conditions
3-ADON= 3-acetyl-deoxynivalenol
15-ADON= 15-acetyl-deoxynivalenol
ATA= alimentary toxic aleukia
bp= base pair
BGI= Beijing genomics institute
BLAST= basic local alignment search tool
BRIG= blast ring image generator
Bw= body weight
C= carbon
CDS= coding sequence
CFIA= Canadian food inspection agency
CHEF= clamped homogeneous electric fields
CMB= corn meal broth
CVTree= composition vector tree
DAS= diacetoxyscirpenol
dH2O= distilled water
DOGT1= UDP-glucosyltransferase
DON= deoxynivalenol
dsRNA= double stranded RNA
dE= de-epoxy
dE-DON= de-epoxy DON
E coli= Escherichia coli
ER=endoplasmic reticulum
FAO= Food amp Agriculture Organization
FM= full medium
FHB= Fusarium head blight
FUS= fusarenonX
gDNA= genomic DNA
GC= guanine cytosine
GER= Gebberella ear rot
GPI= glycosylphosphatidylinositol
h= hour
Hck= hematopoietic cell kinase
Kb= kilobase
xiv
Kg= kilogram
LC-UV-MS= liquid chromatography-ultraviolet-
mass spectrometry
LEE= locus of enterocyte effacement
LD50= lethal dose of 50 animal
LB= Luria Bertani
LMP= low melting point
MAPK= mitogen-activated protein kinase
microg= microgram
mg= milligram
mL= milliliter
microL= microliter
microM= micromole
min= minute
Mb= megabases
MCS=multiple cloning site
MSA= MSB+ agar
MSB= mineral salts broth
MM= minimal medium
MSC= mineral salts + 03 Bacto Peptone
MS-DON= mineral salts + 200 microgmL DON
mRNA= messenger RNA
MSY= mineral salts + yeast extract
mz= mass ion ratio
N= nitrogen
Na-Cl= sodiumndashclorine
NADH= (reduced) nicotinamide adenine
dinucleotide
NAD(P)= nicotinamide adenine
dinucleotide phosphate
NCBI= national center for biotechnology
information
NEO= neosolaniol
ng= nanogram
NIV= nivalenol
OD600nm= optical density at 600
nanometer wavelength
OMAFRA= Ontario Ministry of
Agriculture and Rural Affairs
Paa= phenylacetic acid
PCR= polymerase chain reaction
PDB= potato dextrose broth
ppm= parts per million
PFGE= pulsed field gel electrophoresis
PKR= protein kinase
xv
PNDO= pyridine nucleotide-disulfide
oxidoreductase
QTL= quantitative trait locus
RAST= rapid annotation subsystem technology
RDP= ribosomal database project
Rpl3= 60S ribosomal protein L3
rRNA= ribosomal RNA
s= second
SA= sodium azide
SDR= short chain dehydrogenasereductase
SE= soil extract
SIM= selected ion monitoring
SOAP= short oligonucleotide analysis package
sp= species
sRNA= small RNA
TC= trichothecene
TCA= tricarboxylic acid cycle
TSB= tryptic soy broth
TF= transcription factor
tRNA= transfer RNA
TIC= total ion count
TRAP= tripartite ATP-independent
periplasmic
T-RFLP=terminal restriction fragment
length polymorphism
TRI101= trichothecene 3-O-
acetyltransferase
YEB= yeast extract broth
VER= verrucarol
VKOR= vitamin K epoxide reductase
wv= weight by volume
1
CHAPTER 1
10 GENERAL INTRODUCTION AND LITERATURE REVIEW
11 Background
According to the Food and Agriculture Organization (FAO) of the United Nations
more than 25 of the worldrsquos food crops are contaminated with mycotoxins every year
causing serious adverse effects on livestock industry crop production and international trade
(Boutrif and Canet 1998 Desjardins 2006 Binder et al 2007 Surai et al 2010) The negative
effects of mycotoxins on the consumers are diverse including organ toxicity mutagenicity
carcinogenicity neurotoxicity teratogenicity modulation of the immune system reproductive
system effects and growth retardation Mycotoxins are toxic secondary metabolites produced
by various fungal species during crop colonization in field andor post harvest conditions
(FAO 1991 Barrett 2000) Aspergillus Penicillum and Fusarium are the predominant fungal
genera that accumulate economically important mycotoxins such as aflatoxins zearalenone
trichothecenes fumonisins ochratoxins and ergot alkaloids in agricultural commodities
Humans and animals are principally exposed to mycotoxins through consumption of
contaminated foodfeed and animal products such as milk meat and eggs (Fink-Gremmels
1989 Hollinger and Ekperigin 1999)
Trichothecenes (TCs) produced by a variety of toxigenic Fusarium species are
probably the most food and feed contaminating mycotoxins in the temperate regions of
America Europe and Asia (Creppy 2002 Yazar and Omurtag 2008) The fungal pathogen
accumulates various type-A (eg T-2 toxin HT-2 toxin) and type-B trichothecenes (eg
2
deoxynivalenol nivalenol and their derivatives) in cereals Among them type-B
trichothecenes particularly deoxynivalenol (DON) andor its derivatives appear to be the
most commonly found mycotoxins in grain and grain products worldwide impairing human
health and productivity of farm animals (Placinta et al 1999 Sherif et al 2009) A number of
human and animal disease outbreaks associated with TC contaminated grain food and feed
consumption have been reported in Asia (Japan India and central Asia) Europe New
Zealand and United States of America (USA) (Pestka and Smolinski 2005 Desjardins 2006)
A severe outbreak of Alimentary Toxic Aleukia (ATA) mycotoxicosis killed 100000 people
in the Orenburg region of Russia (Eriksen 2003) In 1996 losses of over $100 million were
estimated in Ontario because of costs related to managing DON (Schaafsma 2002)
Historically T-2 toxin DON and nivalenol (NIV) have likely been used as a biological
warfare in Laos Kampuchea and Afghanistan and there is concern because of the potential
use of these compounds for bioterrorism (Wannemacher et al 2000 Tucker 2001)
Despite the extensive efforts that have been made to prevent TC contamination with
the current varieties and production practices its presence in agricultural commodities is
likely unavoidable There are no crop cultivars currently available that are completely
resistant to Fusarium head blightGibberella ear rot (FHBGER) diseases Chemical or
physical methods cannot safely remove TC mycotoxins from grain completely or they are not
economically viable (Varga and Toacuteth 2005) Therefore an alternative strategy to address the
issue is of great importance (Awad et al 2010 Karlovsky 2011)
3
12 Plant pathogenesis and trichothecene production
Cereals infested by the ascomyceteous fungi may cause Gibberella ear rot (GER) in
maize (Zea mays L) and Fusarium head blight (FHB also known as lsquoscabrsquo) of small grain
cereals [wheat (Triticum aestivum L) barley (Hordeum vulgare L)] and other small grain
cereals (Goswami and Kistler 2004) As many as 17 fungal species have been reported to be
associated with FHBGER which include F graminearum Schwabe [telemorph Gibberella
zeae (Schwein) Petch] F culmorum (WG Smith) Sacc F sporotrichioides Scherb F
poae (Peck) Wollenw Microdochium nivale (Fr) Samuels amp IC Hallett and F avenaceum
(Fr) Sacc (Osborne and Stein 2007 Audenaert et al 2009) The two species F
graminearum and F culmorum are the principal species causing crop damage Among them
F graminearum is the widely adapted and most damaging species across the world including
North America (Dyer et al 2006) F culmorum tend to be the main species in the cooler parts
of North Central and Western Europe (Wagacha and Muthomi 2007)
The fungi overwinter mainly on crop residues as saprophytes where they produce
asexual (micro- and macro-conidia) andor sexual (ascospores) fruiting structures which act
as the initial source of inoculums in the subsequent crop season (Sutton 1982 Wagacha and
Muthomi 2007) Ascospores are primarily dispersed by air (Trail et al 2002) and conidia
move by rain-splash (Paul et al 2004) After they land on maize silks and florets of small
grain cereals infection occurs through natural openings (eg stomata) or occasionally by
direct penetration The pathogens can invade maize kernels through wounds caused in the
husks by insects or birds Infection may occur throughout the crop growing season but
disease incidence and severity is facilitated by warm temperatures (gt20degC) high humidity
(gt80) and frequent precipitation that coincides with anthesis (Osborne and Stein 2007
4
Wagacha and Muthomi 2007) The Fusarium pathogen initially infects as biotroph which
becomes a necrotroph as the disease progresses After penetration Fusaria produce different
extracellular cell wall degrading enzymes that trigger infection and hyphal progression
towards the developing earkernel where further colonization occurs and development of
FHBGER diseases (Voigt et al 2005 Miller et al 2007 Kikot et al 2009) FHB disease is
characterized by a bleached appearance around the middle of the head Severely infected
grains have a pinkish discoloration and can be shriveled In maize the disease produces a
reddishndashbrown discoloration of the kernels and progresses from the tip to the bottom of
affected ears
The fungi associated with FHB and GER contaminate cereals by accumulating various
host-nonspecific TC mycotoxins (Brennen et al 2007) TCs have typically been associated
with cereals grown in temperate climatic conditions including Canada (Doohan et al 2003)
DON production by F graminearum and F culmorum is maximal at warm temperatures (22-
28˚C) high grain moisture contents (gt17) and humid conditions (gt70 RH) (Martins and
Martins 2002 Hope et al 2005 Llorens et al 2006) Grain can be contaminated with TCs in
the field as well as in storage if conditions for fungal growth are optimal and grain contains
high moisture (gt17) (Larsen et al 2004 Birzele et al 2002)
Populations of FHBGER causing pathogens are dynamic Genetic variability between
and within Fusarium species appears to have occurred to allow for their adaptation to new
agro-climates (Desjardins 2006 Fernando et al 2011) Three strain specific Fusarium
chemotypes NIV 3-ADON (3-acetyl-deoxynivalenol) and 15-ADON (15- acetyl-
deoxynivalenol) have been identified which correlate with the geographical distributions
(Starkey et al 2007 Zhang et al 2007 Audenaert et al 2009) The NIV chemotype mainly
5
produced by F culmorum accumulates NIV and 4-NIV (Fusarinon X) is predominant in
Asia Africa and Europe (Lee et al 2001) The 3-ADON chemotype which synthesizes DON
and 3-ADON is the dominant strain in EuropeFar East (Larsen et al 2004) In North
America 15-ADON is the main chemotype which produces DON and 15-ADON (Goswami
and Kistler 2004) Significant Fusarium population shifting was observed in the past and in
recent years (Kant et al 2011) A dynamic change from F culmorum to F gramninearum
and DON to NIV producing chemotyes occurred in North West European countries The 3-
ADON chemotype which is uncommon in North America was found in North Dakota and
Minnesota (Goswami and Kistler 2004) F pseudograminearum which historically occurs in
Australia Africa and North West Pacific America has been found in Western Canada (Larsen
et al 2004 Kant et al 2011) In Manitoba a replacement of 15-ADON chemotype by the
more phytoxic 3-ADON type was observed in recent years (Fernando et al 2011) All
together the FHBGER associated pathogen complex is highly dynamic which not only
reduces crop yield but also decreases grain quality by accumulating various mycotoxins
6
13 Biosynthesis of food contaminating TC
TCs are fungal secondary metabolites that were initially identified in Trichotheceium
roseum Thus the name of this class of compounds follows the name of the genus
Trichotheceium (Freeman and Morrison 1949) There have been 180 TCs identified in various
fungal species belonging to the genera Fusarium Myrothecium Stachybotrys Trichoderma
and Thrichothecium (Eriksen 2003 Eriksen et al 2004) The FHBGER associated TCs are
mainly synthesized by different Fusaria and are characterized as tricyclic sesquiterpenoids
with a double bond at position C9-10 and a C12-13 epoxide ring
7
Figure 1-1 Structures of trichothecene mycotoxins produced by Fusaria (a) General
structure of trichothecenes R1 = H OH OiValeryl keto R2 = H OH R3= OH OAc R4 =
H OH OAc R5= OH OAc (b) Type-A trichothecene mycotoxin (c) Type-B trichothecene
mycotoxin which contain a carbonyl functional group at C8 position (Young et al 2007)
(a) (b)
(c)
8
The fungal metabolites are small in size (molecular weight 250-550) heat stable and
non-volatile molecules (Pestka 2007 Yazar and Omurtag 2008) Food contaminating TCs are
divided into two groups type-A and type-B (Figure 1-1b and 1c) Type-A is characterized by
the lack of a ketone function at C8 position and include T-2 toxin HT-2 toxin
diacetoxyscirpenol (DAS) and neosolaniol (NEO) which are mainly produced by F
sporotrichioides and F poae (Creppy 2002 Quarta et al 2006) The most frequently found
TC belongs to type-B which contains carbonyl functions at C8 positions DON NIV and
their acetylated derivatives The two types are further differentiated by the presence or
absence of functional hydroxyl (-OH) and acetyl (CH3-CO-) groups at C-3 C-4 C-7 C-8 and
C-15 positions (Table 1-1)
The syntheses of various TCs are speciesstrain specific and are genetically controlled
but influenced by environmental conditions (Desjardins 2006 Paterson and Lima 2010) Until
now 15 Fusaria catalytic genes and regulators involved in TC productions have been
identified (Merhej et al 2011) These biosynthetic genes are located at three loci on different
chromosomes The core trichothecene (TRI) gene cluster is comprised of 12 genes localized
in a 25-kb genomic region (Kimura et al 2001) The remaining three genes are encoded by
the TRI1-TRI16 and TRI101 loci (Gale et al 2005 Alexander et al 2009) Fusaria produce
TC following a number of oxygenation isomerization acetylation hydroxylation and
esterification reaction steps (Desjardins 2006 Desjardins and Proctor 2007 Kimura et al
2007 Merhej et al 2011) In brief farnesyl pyrophosphate is initially cyclized into
trichodiene by TRI5 which codes for trichodiene synthase In the next four steps
isotrichotriol is produced by TRI4 (a multifunctional cytochrome P450) via C-2 hydroxylation
and 1213 epoxidation followed by two hydroxylation reactions
9
Table 1-1 Fusaria produced type-A and type-B trichothecene mycotoxins Molecular weight
(MW) and acetyl (-R) hydroxyl (-OH) and ketone (=O) functions at different carbon (C)
position of the tricyclic rings A-types T-2 toxin HT-2 toxin DAS (diacetoxyscirpenol)
NEO (neosolaniol) VER (verrucarol) B-type DON (deoxynivalenol) 3-ADON (3-acetyl-
deoxynivalenol) 15-ADON (15-acetyl-deoxynivalenol) NIV (nivalenol) and FusX
(fusarenon X) (Young et al 2007)
Trichothecenes MW R1C3 R2C4 R4C7 R3C15 R5C8
Type-A
T-2 toxin 466 OH OCOCH3 H OCOCH3 OCOCH2CH(CH3)2
HT-2 toxin 424 OH OH H OCOCH3 OCOCH2CH(CH3)2
DAS 366 OH OCOCH3 H OCOCH3 H
NEO 382 OH OCOCH3 OH OCOCH3 H
VER 266 H OH H OH H
Type-B
DON 296 OH H OH OH =O
3-ADON 338 OCOCH3 H OH OH =O
15-ADON 338 OH H OH OCOCH3 =O
NIV 312 OH OH OH OH =O
FusX 354 OH OCOCH3 OH OH =O
10
The next two non-enzymatic reactions results in isotrichodemol Eventually the
trichothecene skeleton is produced via the formation of a C-O bond between the C-2 oxygen
and C-11 Subsequent acetylation at C-3 by TRI101 hydroxylation at C-15 by TRI1 and
catalytic activity of TRI3 leads to calonectrin production The pathway described above is
common for both type-A (eg T-2 toxin HT-2 toxin) and type-B (eg DON NIV and their
acetylated derivatives) TCs
The following steps are speciesstrain specific In DON producing F graminearum
strains calonectrin is converted to either 3-ADON or 15-ADON followed by DON production
by the catabolic activities of TRI1 and TRI8 The biosynthesis of 3-ADON or 15-ADON is
controlled by variation in the esterase coding sequences of TRI8 F graminearum produces
NIV from calonectrin via synthesis of 4-NIV followed by NIV by TRI1 and TRI8 gene
actions NIV can be produced either from DON by hydroxylation at C-4 or via
transformation of calonectrin to 315-diacetoxyscirpenol (315-DAS) followed by addition of
a ketone at C-8 position by the activity of the TRI7 and TRI13 genes 315-DAS is the
principal substrate for synthesis of type-A TCs by F sporotrichioides the main type-A
producer The biosynthetic pathway of T-2 toxin involves C-4 acetylation of 315-DAS by the
TRI7 gene product and TRI8 triggered by TRI1 315-DAS is also the substrate for HT-2
toxin synthesis In the absence of TRI7 expression HT-2 toxin is produced through
hydroxylation of 315-DAS and isovalerate addition to C-8 position (Foroud and Eudes 2009)
11
14 Occurrence of TC mycotoxins in food and feed
TC producing Fusaria are adapted to variable climatic conditions and agricultural
ecosystems As a consequence this class of mycotoxins particularly DON is frequently
detected in foods and feeds including breakfast cereals bakery products snack foods beer
and animal feed derived from cereals in most part of the world (Pittet 2001 Desjardins 2006
Lancova et al 2008) A two-year survey during 2003-2005 conducted by Romer Lab Inc
revealed unacceptable levels of DON in animal feedsfeed raw materials (such as maize
wheat wheat bran) in Asia and Oceania (1062 microgg) central Europe (802 microgg) Mid-West
USA (55 microgg) northern Europe (551 microgg) and southern Europe (303 microgg) (Binder et al
2007) (Table 1-2)
Quite high levels of DON contamination (19 microgg) were detected in north Asian and
Chinese feedfeed materials Recently five-year on-farm studies in Switzerland showed that
74 of the wheat grain and straw samples were contaminated with high levels of DON
(average 5 microgg) The amount is higher than the European maximum limit of 125 microgg
(Vogelgsang et al 2011)
The occurrences of TCs in Canadian agricultural commodities are indicated in Table
1-3 Seventeen surveys conducted during the 1980s and 1990s showed up to 100 DON
incidence in Ontario wheat and maize samples with an 18 average incidence (Scott 1997) A
three-year survey recently (2006-08) conducted by the Ontario Ministry of Agriculture and
Rural Affairs (OMAFRA) detected high levels of DON in spring (maximum 26 microgg) and
winter wheat (maximum 29 microgg) samples (Ana Matu personal communication) The DON
levels were much higher than Health Canadarsquos maximum limit of 2 microgg for uncleaned soft
wheat for staple foods (Table 1-4)
12
Table 1-2 Worldwide occurrence of DON in animal feed Feed and feed raw material
samples were collected from animal farmsfeed production sites during 2003-2005 The
survey was conducted by Romer Labs Inc (httpwwwromerlabscom)
Country
region
Sample
type
Incidence
()
Maximum
DON content
(microgg)
Mean DON
content
(microgg)
Survey
period
Source
North
AsiaChina
Animal
feed
071 1899 092
Binder et
al 2007
South East
Asia
014 152 016
South Asia 070 1062 049
Oceania 2003-05
Australia 041 186 030
Northern
Europe
070 551 059
Central
Europe
066 802 057
Southern
Europe
052 303 030
13
Table 1-3 Occurrence of DON in Canadian grains grain foods and beers Infant cereal foods
oat barley soy and multi-grain based foods
Province Sample
type
Incidence
()
Maximum
DON
contents
(microgg)
Multiple
mycotoxins
samples ()
Survey
period
Sources
Ontario Spring
Wheat
70 260 Yes 2006-08 OMAFRA1
Ontario Winter
wheat
49 290 Yes 2006-08 OMAFRA
Ontario Maize 78 140 Yes 2009-10 Limay-Rios
and
Schaafsma
2010
Ontario Maize - 1000 Yes 2011 AampL
Biologicals
Canada Breakfast
cereals
46 094 430 1999-01 Roscoe et al
2008
Canada Animal
feeds
73 425 Yes 1990-10 Zeibdawi et
al 2011
Canada Beers 58 0052 50 1990s Scott 1997
Canada Infant
Cereals
63 098 Yes 1997-99 Lombaert et
al 2003
1
Ontario Ministry of Agriculture and Rural Affairs
2 microgmL
14
Table 1-4 Legislative maximum tolerated levels of trichothecene mycotoxins in Canada
according to Canadian Food Inspection Fact Sheet April 23 2009 DON- deoxynivalenol
DAS- diacetoxyscirpenol
Products DON
(microgg)
T-2 toxin
(microgg)
HT-2 toxin
(microgg)
DAS
(microgg)
Uncleaned soft wheat for human
consumption
2 - - -
Cattlepoultry diets 5 lt1 01 lt1
Swinedairy cattle feeds 1 lt1 0025 lt2
15
High DON incidence (78) was observed in maize samples collected from
southwestern Ontario fields with moderate levels (lt1 microgg) of contamination during 2009 and
2010 (Limay-Rios and Schaafsma 2010) Some samples had high DON levels (14 microgg) In
the 2011 crop season extremely high levels of DON (gt100 microgg) were found in some
southern Ontario maize samples (AampL Biologicals London ON Personal communication)
In a Canadian Food Inspection Agency (CFIA) 20 year survey (1990-2010) 73 of
animal feed samples collected from feed mills farms and retail stores across Canada were
contaminated with DON and for more than 25 of the samples the mycotoxin levels were
above the maximum limit (ge 1 microgg) (Zeibdawi et al 2011) The highest level of DON (15
microgg) detected in the 2006-2007 samples were much lower than the levels found in the 1999-
2000 samples which contained a maximum of 425 microgg DON Importantly this study
reported a significant number of samples contaminated with multiple mycotoxins DON NIV
3-ADON 15-ADON T-2 toxin HT-2 toxin DAS and NEO
Earlier studies in the 1990s detected high levels of DON content in Eastern Canadian
cereals maize (maximum 175 microgg) wheat (maximum 916 microgg) barley (maximum 911
microgg) and oats (maximum 12 microgg) (Campbell et al 2002) The study identified a significant
number of samples (15-367) that contained multiple mycotoxins and up to 33 of the
wheat samples exceeded 1 microgg DON Low amounts but high incidences of TCs were found
in cereal-based foods collected from Canadian retail stores Sixty three percent of infant
cereal foods 46 of breakfast cereals and 58 of beers were contaminated with one or
multiple mycotoxins (Scott 1997 Lombaert et al 2003 Roscoe et al 2008) The maximum
amount of DON was found in infant foods (098 microgg) followed by breakfast cereals (094
microgg) and beers (005 microgmL)
16
15 Significance of deoxynivalenol and trichothecenes
For most cereals including wheat maize rice barley oats and rye the risk of
contamination with TCs resulted in grain quality reduction and FHBGER disease progression
(Bai et al 2002 Jansen et al 2005) The impacts of TC contamination in agricultural
commodities are diverse including reduced livestock production induced human illnesses
and enhanced FHB disease severity which may result in the reduction of crop yield Exposure
of farm animals (particularly pigs) to DON is a continuing threat to the livestock industries in
Canada USA and Europe (DrsquoMello et al 1999) Ingestion of TC-contaminated feeds
generally causes reduction of feed intake feed refusal poor feed conversion retardation of
weight gain increased disease susceptibility and reduced reproductive capacities (Binder et al
2007) High doses of DON result in acute toxic effects in pigs (eg emesis) while low doses
(ge1 microgmL) cause chronic effects (eg growth retardation in pigs) (Rotter et al 1996
Accensi et al 2006) Reduction of growth and reproduction ability was observed in
experimental rats exposured to much higher doses of TC (Sprando et al 2005 Collins et al
2006) Several animal disease outbreaks that occurred in different continents have been
attributed to consumption of feed contaminated with DON NIV and DAS (Desjardins 2006)
The consequences of human exposure to TCs include liver damage gastricintestinal
lesions and central nervous system toxicity resulting in anorexia nausea hypotensionshock
vomiting diarrhea and death (Pestka 2007) Human mycotoxicosis (illnesses and outbreaks)
by consumption of type-A and type-B TC mycotoxins in Europe New Zealand Russia
Japan South America India and Vietnam have been reported (Bhat et al 1989 Yoshizawa
1993 Luo 1994 Desjardins 2006) Severe outbreaks of Alimentary Toxic Aleukia (ATA)
associated with TC killed thousands of people in the Orenburg region of Russia during the
17
1930rsquos and 1940rsquos (Eriksen 2003) TCs (T-2 DON and NIV) were likely used for biological
warfare in Laos Kampuchea and Afghanistan and its potential application for bioterrorism is
an additional concern associated with TC (Tucker 2001 Bennett and Klich 2003)
TCs are phytotoxic and may contribute to FHB disease severity in cereals (Proctor et
al 1995 Maier et al 2006 Masuda et al 2007) Studies demonstrated that DON caused
wilting chlorosis inhibition of seedling growth and necrosis in cereals (Cossette and Miller
1995 Mitterbauer et al 2004 Rocha et al 2005) DON-mediated host cell damage and
subsequent necrotrophic fungal spread has been shown to lead to FHBGER disease
development and higher kernel damage (Procter et al 1995 Mitterbauer et al 2004 Maier et
al 2005) This suggests that DON contributes to yield losses as well as to reductions in the
market value of cereals (Osborne and Stein 2007) Severely infected grain may contain more
than 60 microgg DON which is unacceptable to the milling and brewing industries (Okubara et
al 2002) Significant crop losses due to FHB disease epidemics and mycotoxin contamination
have been reported in Ontario (Schaafsma 2002) FHB disease epidemics during 1998 to 2000
caused huge crop losses resulting in farm failures in the northern Great Plains in the USA
(Windels 2000) In that period FHB associated losses were estimated at US $ 27 billion in
the region (Nganje et al 2002)
Importantly Fusaria infested grains are often co-contaminated with multiple
mycotoxins which further increase health risks as interactions between the mycotoxins may
cause greater toxic effects than expected (Schollenberger et al 2006) This implies that an
apparently low level of contamination of a single mycotoxin can have severe effects on
consumers because of synergistic effects among multiple co-occuring mycotoxins (Surai et al
2010)
18
A number of studies demonstrated synergistic toxic effects among Fusarium
mycotoxins on pigs turkeys lamb chicks as well as in in vitro Caco-2 cell and yeast
bioassays (Table 1-5)
Table 1-5 In vitro and in vivo synergistic effects of different food contaminating mycotoxins
DON-deoxynivalenol DAS-diacetoxyscirpenol FB1-fumonisins 1 NIV-nivalenol ZEN-
zearelenone FA-fusaric acid Afl-aflatoxin
Mycotoxins source Interacting
mycotoxins
Experimental
animalcells
Effects Reference
Naturally contaminated
cereals
DON FA Pigs Inhibit growth Smith et al
1997
DON contaminated
wheat and FB1
DON FB1 Pigs Weight loss Harvey et al
1996
Culture materials
contained mycotoxins
T-2 FB1 Turkey poults Weight loss oral
lesion
Kubena et al
1997
Inoculated rice and
pure mycotoxin
DAS Afl Lamb Weight reduction Harvey et al
1995
Naturally DON
contaminated wheat
and pure T-2 toxin
DON T-2 Chicks Reduced body
weight significantly
Kubena et al
1989
Pure mycotoxins DAS Afl Chicks Reduced body
weight
Kubena et al
1993
Pure mycotoxins T-2 Afl Chicks Reduced weight
oral lesion
Kubena et al
1990
Pure mycotoxins DON
ZENFB1
Human intestinal
Caco-2 cells
Increased DNA
fragmentations
Kouadio et al
2007
Pure mycotoxins DON NIV Yeast bioassay Synergistic effects Madhyastha et
al 1994
Pure mycotoxins T-2 HT-2 Yeast bioassay Synergistic effects Madhyastha et
al 1994
19
16 Toxic effects of deoxynivalnol and trichothecenes
As small molecules TCs simply enter into cells and interact with different cell
components resulting in disruption of cellular functions and causing various toxic effects that
include inhibition of protein synthesis and ribosomal fragmentation (Eudes et al 2000
Minervini et al 2004 Pestka 2007) Simple absorption of DON into human intestinal Caco-2
cells and crossing human placenta barriers through passive diffusion has been reported
(Sergent et al 2006 Nielsen et al 2011)
TC may cause various toxicities in animals and plants including apoptosis
immunotoxicity chromatin condensation disruption of cellularmitochondrion membranes
disruption of gut tissues inhibition of protein and ribosome synthesis breakage of ribosome
and cell damage The toxicity of TC is influenced by structural variability (ie position
number and type of functional groups attached to the tricyclic nucleus and C-C double bond at
C9-10 position) (Desjardins 2006) Nevertheless epoxide function at the C12-13 position is
the key for TC cytotoxicity (Swanson et al 1988 Eriksen et al 2004)
Different in vitro and in vivo studies revealed that the method and period of exposure
amount of TC and types of celltissue and host species have significant effects on TCrsquos
cytotoxicity (Scientific Committee for Food 2002 Rocha et al 2005 Nielsen et al 2009)
Administration of 10 mgkg body weight (bw) NIV caused apoptosis in mice thymus Peyerrsquos
patch and spleen tissues (Poapolathep et al 2002) DON and NIV at 887 mgkg bw for 3 days
a week for four weeks induced immunotoxicity in experimental mice determined by
estimating increased immunoglobulins (IgA) and decreased IgM levels (Gouze et al 2007)
Exposure of mice to 1 mgkg bw of DON induced skeletal malformations and reduced mating
tendency while significant resorption of embryos observed at the dose of gt10 mgkg bw
20
(Khera et al 1982) At high acute doses (4972 mgkg bw) DONNIV and type-A
trichothecenes caused mice death because of intestinal haemorrhage necrosis of bone
marrowlymphoid tissues and kidneyheart lesions (Yoshizawa and Morooka 1973
Yoshizawa et al 1994 Forsell et al 1987 Ryu et al 1988) In a separate study 15-ADON
showed more toxicity than DON when the mycotoxins were orally administered to mice
(Forsell et al 1987)
Studies suggested that type-A TCs are more toxic in mammals than type-B which
means that functional variability at the C-8 position of TC has a significant role in toxicity
The toxicity of various TC mycotoxins in animals can be ranked in the order of T-2
toxingtHT-2 toxingtNIVgtDONge3-ADONgt15-ADON according to the results from human cell
lines and mice model systems bioassays (Ueno et al 1983 Pestka et al 1987 Nielsen et al
2009) Moreover animal feeding assays demonstrated that monogastric animals are most
sensitive to DON The order of sensitivity of animals to DON is
pigsgtrodentsgtdogsgtcatsgtpoultrygtruminants (Pestka 2005) The reasons for the variations in
DON toxicity species might be due to differences in metabolism absorption distribution and
elimination of the mycotoxin among animal species (Pestka 2007)
To my knowledge the mechanism of TC uptake by plant cells has not been studied
Nonetheless cereals are exposed to TC by toxigenic Fusaria infection which may cause
various toxicities such as cell damage wilting chlorosis necrosis program cell death and
inhibition of seedling growth in sensitive plants (eg wheat maize and barley) (Cossette and
Miller 1995 Rocha et al 2005 Boddu et al 2007) As animals TC inhibits protein synthesis
in non-host A thaliana (Nishiuchi et al 2006) Immunogold assays showed that the cellular
targets of DON in wheat and maize were cell membrane ribosome endoplasmic reticulum
21
vacuoles and mitochondria Modulation of cell membrane integrity and electrolyte losses
were observed by direct DON interference in maize and wheat cell assays (Cossette and
Miller 1995 Mitterbauer et al 2004) DON-mediated mitochondrion membrane alterations
disruption of host cells enzymatic activity apoptosis and necrosis have been observed in
cereals (Mitterbauer et al 2004 Rocha et al 2005) In contrast to animal type-B is more toxic
to plants than type-A DON showed 8- and 13-fold phytotoxicity to wheat coleoptiles than T-
2 and HT-2 toxins respectively (Eudes et al 2000) By compilation of different studies TC
phytoxicity can be ordered as DONge3-ADONgtT-2 toxingtDAS (Wakulinski 1989 McLean
1996 Eudes et al 2000)
22
17 Cellular and molecular mechanisms of DON toxicity in animals and plants
The underlying cellular and molecular mechanisms of DONTC toxicity are evolving
Exposure to DON proceeds with the up and down regulation of numerous critical genes of
eukaryotic organisms which causes various negative effects on animal and plant organisms
(Figure 1-2)
One of the early events of DON toxicity occurs when the mycotoxin binds to the
active ribosome resulting in blocking protein translation through different routes by
damaging the ribosome (Shifrin and Anderson 1999) degrading or altering conformational
changes of 28S-rRNA (Alisi et al 2008 Li and Pestka 2008) and inducing expression of
miRNAs This results in down regulation of the ribosome associated genes (He et al 2010a
Pestka 2010) Both animal and plant cell assays demonstrated that DON bound to the peptidyl
transferase (Rpl3) domain of the 60S subunit of ribosome and blocked protein chain
elongation and termination at high DON concentrations while peptide chain termination and
ribosome breakage occurred at a low level of DON exposure (Zhou et al 2003a)
23
Figure 1-2 Cellular and molecular mechanisms of deoxynivalenol toxicity in eukaryotic
organisms DON is simply absorbed by the cell and interacts with the cytoplasmic membrane
(CM) mitochondrion (MT) membrane and ribosome Disruption of the CM and alteration of
the MT membrane causes a loss of electrolytes and affects the function of the MT
respectively DON binds to the active ribosomes which leads to ribosome damage and
induces micro RNAs (miRNA) Expression of miRNA impairs the synthesis of ribosome
associated protein The subsequent transduction and phosphorylation of the RNA-activated
protein kinase (PKR) and hematopoietic cell kinase (Hck) activate the mitogen-activated
protein kinase (MAPK) cascade Phosphorylation of the MAPKs induces targeted
transcription factors (TF) which results in ribotoxis stress response upregulation of
proinflammatory cytokines and dysregulation of neuroendocrine signaling (derived from
Pestka 2007 and 2010)
24
The consequence of DON interaction with ribosomes leads to cellular and molecular
changes which are complex and not fully understood However it is well-demonstrated that
the activation of the mitogen-activated protein kinase (MAPK) and subsequent molecular
actions play key roles in the cytotoxicity of TC Upon binding to the ribosome DON rapidly
induced interactions between the protein kinases and the ribosomal 60S subunit as observed in
mononuclear phagocytes (Bae and Pestka 2008) Ribosomal binding to DON activated the
ribosome associated kinases for instances protein kinase R (PKR) hematopoietic cell kinase
(Hck) and P38
mitogen-activated protein kinase The protein kinases are induced by double-
stranded RNA (dsRNA) or possibly by the damaged ribosomal peptides (Iordanov et al 1997
Shifrin and Anderson 1999 Zhou et al 2003b Pestka 2007) Subsequent phosphorylation of
kinases (eg PKRHckP38
) activates the MAPK cascade resulting in ribotoxis stress
response upregulation of proinflammatory cytokines and dysregulation of neuroendocrine
signaling (Pestka and Smolinki 2005 Pestka 2010) Studies suggested that high doses of
DON induces ribotoxis stress response (apoptosis) preceded by immunosuppression and
disruption of intestinal permeabilitytissue functions (Yang et al 2000 Pestka 2008) Further
study observed stabilization of the mRNA and expression of transcription factors for critical
genes when cells were exposed to a low concentration of DON (Chung et al 2003)
The MAPK pathway proceeds with upregulation of a broad range of proinflammatory
cytokine genes and dysregulation of neuroendocrine signaling within the central and enteric
nervous systems Modulation of neuroendocrine signaling by DON increased the level of
serotonin resulted in vomiting reduced food intake and growth retardation in experimental
animalscell lines (Prelusky and Trenholm 1993 Li 2007) Upregulation of proinflammatory
cytokines and chemokines genes was determined in mice liver spleen kidney and lung
25
following in 1 h of exposure to DON A subsequent shock-like effect was observed in mice
because of a ldquocytokine stormrdquo (Pestka 2008) Induction of proinflammatory cytokines also led
to various cellular effects on the central nervous system interference of the innate immune
system and growth hormone signaling These contribute to anorexia growth retardation
aberrant immune response and tissue dysfunction as demonstrated in experimental animal
systems (Pestka 2010)
DON-mediated growth retardation in mice via suppression of growth hormones
(insulin-like growth factor 1 and insulin-like growth factor acid-labile subunit) has been
reported by Amuzie et al (2009) Disruption of gut tissues by DONNIV and subsequent
impairment of intestinal transporter activity was demonstrated in a human epithelial cell
model (Marseca et al 2002) The study proposed that alteration of gut tissue permeability
may cause gastroenteritis induction and growth retardation in livestock animals Growth
retardation of livestock may also occur via dysregulation of neuroendocrine signaling-
mediated induction of emesis and anorexia (Pestka et al 2010)
The molecular mechanism of TC toxicity in plant systems has not been well
documented A recent study showed the upregulation of numerous defense related genes in
wheat upon exposure to DON (Desmond et al 2008) The study further suggested that DON-
induced program cell death in wheat occurred through a hydrogen peroxide signaling
pathway This mechanism had not been observed in mammals
26
18 Strategies to control FHB and trichothecenes
During the past decades many pre- and post-harvest strategies have been applied to
protect cereal crops from Fusarium infestation and to eliminate TC from contaminated
agricultural commodities (FAO 2001 Munkvold 2003 Varga and Toth 2005 Kabak and
Dobson 2009) Some of the above crop protection techniques have been patented (He and
Zhou 2010) In spite of those efforts TC contamination of cereals appears to be unavoidable
using the currently available methods
Pre-harvest control strategies involving cultural practices (eg crop rotation tillage
early planting) and fungicide applications partially reduced Fusarium infection and DON
contamination in cereals (Dill-Macky and Jones 2000 Schaafsma et al 2001) However
repeated application of fungicides may lead to the evolution of Fusarium strains resistant to
fungicides with greater mycotoxin production ability (DrsquoMello et al 1998) At post-harvest
reduction of grain moisture by artificial drying storage at low temperatures (1-4degC) and
proper ventilation of the storage room have shown to be effective in inhibiting fungal growth
and mycotoxin production in infected grain (Midwest Plan Service 1987 Wilcke and Morey
1995) This technology has become less applicable particularly in developing countries
because of the frequent observation required to maintain proper storage conditions (Hell et al
2000)
Removal of Fusarium infected grains by physical methods (eg mechanical
separation density segregation colour sorting etc) was unable to eliminate TC completely
(Abbas et al 1985 He et al 2010b) This is partly because FHB symptomless kernels may
contain a considerable amount of DON which are not eliminated by physical separation
procedures (Seitz and Bechtel 1985) Several studies showed that milling and food processing
27
steps were unable to completely remove DON from grain-based foods (Jackson and
Bullerman 1999 Placinta et al 1999 Kushiro 2008) Detoxification of TC by chemical
treatments (eg ozone ammonia chlorine hydrogen peroxide and sodium metabisulfite) has
been demonstrated (Young et al 1986 Young et al 1987 Young et al 2006) The problem
with these techniques is that the application of such chemicals is not economically viable and
it also has a negative impact on grain quality (Karlovsky 2011) Although the technique has
been popular and showed effectiveness to certain mycotoxins utilization of absorbents to
inhibit mycotoxin absorption by the livestock gastrointestinal tract appeared to be ineffective
for TC (Huwig et al 2001)
The introduction of Fusarium resistant cultivars would be the best strategy to address
the FHB and associated mycotoxin problems in cereals However breeding for cultivars
resistant to FHBGER is difficult and time consuming as natural resistance against Fusarium
is quantitative in nature FHB resistant quantitative trait loci (QTLs) have been identified in
several cereal genotypes (eg Sumai 3 Ning 7840 Bussard) (Bernando et al 2007 Golkari
et al 2009 Haumlberle et al 2009 Yang et al 2010) The 3B 5A and 6B QTLs for type-I
(resistant to initial infection) and type-II (resistant to fungal spread in the kernel) resistances
have been identified on different wheat chromosomes (Buerstmayr et al 2009) Type-I
(Qfhsifa-5A) and type-II (Fhb1Fhb2-3B) QTLs from Sumai 3 are being used in many
breeding programs and pyramiding of the QTLs in spring wheat showed additional effects
Nonetheless the key genes involved in FHB resistance in cereals are yet to be identified This
makes it difficult to integrate the resistance genes into elite cultivars
28
Cultivar development by direct incorporation of TCDON resistance genes may be an
alternative way of protecting grains from these mycotoxins contaminations Many genes from
distantly related species have been integrated into crop cultivars using a transgenic approach
with great economic benefits (Brookes and Barfoot 2006) DON transforming trichothecene
3-O-acetyltransferase (TRI101 from Fusarium sporotrichioides) and UDP-
glucosyltransferase (from Arabidopsis thaliana) genes expressed in cereal cultivars showed
some DONDAS detoxifying ability under in vitro and controlled conditions (Kimura et al
1998 Okubara et al 2002 Poppenberger et al 2003 Ohsato et al 2007) By contrast no
reduction of mycotoxins in either of the above cases was observed under field conditions
(Manoharan et al 2006) Importantly the transformed acetylated and glycosylated
compounds could revert to the original substances or convert to even more toxic products
(Gareis et al 1990 Alexander 2008)
19 Isolation of DONTC de-epoxidating micro-organisms and genes
An enzyme with the capability of de-epoxidizing DONTC appears to be a potential
source for completely removing TC from contaminated cereals (Karlovsky 2011) However
such trichothecene detoxifying genes do not exist in cereals and other plants (Boutigny et al
2008) Because of this several research groups have attempted to obtain microorganisms that
encode DONTC de-epoxidation genes Mixed microbial culture capable of de-epoxydizing
trichothecenes have been isolated from chicken gut (He et al 1992) cow ruminal fluid (King
et al 1984 Swanson et al 1987 Fuchs et al 2002) pig gut (Kollarczik et al 1994) and
digesta of chicken (Young et al 2007) rats (Yoshizawa et al 1983) sheep (Westlake et al
1989) and fish (Guan et al 2009) All of the microbial cultures cited above showed de-
29
epoxydation capabilities only under anaerobic conditions realtively high temperatures and at
lowhigh pH except microbes from fish intestine that showed efficient de-epoxidation at low
temperatures and wide range of temperatures (Guan et al 2009) Such enzymes active under
anaerobic conditions high temperatues and low pH may have potential applications as feed
additives and the eubacterial strain BBSH 797 is an animal feed additive product with the
brand name Mycofixreg by BIOMIN (He et al 2010b) Biotechnological applications of
anaerobically-active enzymes from the above microbial sources seem to be inappropriate
under the aerobic and moderate conditions at which cereals plants are grown
Most of these efforts have failed to isolate single DONTC degrading microbes
effective under aerobic conditions because of inappropriate microbial growth conditions
under laboratory conditions Earlier attempts were made to obtain TC de-epoxidizing aerobic
bacteria but the culture did not retain DONNIV de-epoxidation activity after reculturing
(Ueno 1983 He et al 1992) In addition the co-metabolic nature of the microbial TC de-
epoxidation reaction has limited the use of mycotoxins as the sole nutrient source for selective
isolation of the target microorganisms Several DONTC de-epoxidation microbial strains
including Bacillus strain LS100 and Eubacterial strain BBSH 797 have been isolated from
chicken gut and rumen fluids through multi-step enrichment procedures (Fuchs et al 2002
Yu et al 2010) A DON to 3-keto DON transforming bactial strain was islated from soil by
prolonged incubation with high concentration DON (Shima et al 1997)
Microbial genesenzymes with TC de-epoxidation activity are not known Based on
the microbial TC de-epoxidation activity of a mixed microbial culture it has been proposed
that ceratin reductases or oxidoreductases may catalyze such a biochemical reaction (Islam et
al 2012) With the advent of molecular techniques many novel genes with
30
industrialagricultural value have been cloned by reverse genetics (genomic DNA library
preparation and functional screening of the library clones) forward genetics (transposon
mutagenesis) and comparative genomics methods (Frey et al 1998 Koonin et al 2001
Suenaga et al 2007 Chung et al 2008 Ansorge 2009) The tremendous advancement of next
generation sequencing technology and bioinformatics tools may facilitate the identification of
the microbial genes responsible for DONTC detoxification (Ansorge 2009) Isolation of TC
de-epoxidation genes may play a major role in producing safe foods and feeds derived from
grains
110 The bacterial species Citrobacter freundii
Citrobacter freundii is a bacterial species belonging to the family Enterobacteriace It
is a facultatively anaerobic and Gram-negative bacillus (Wang et al 2000) The bacterium
survives almost everywhere including animalhuman intestines blood water and soils
(Wang et al 2000 Whalen et al 2007) C freundii is known as an opportunistic human
pathogen and causes a variety of respiratory tract urinary tract and blood infections and
responsible for serious neonatal meningistis disease (Badger et al 1999 Whalen et al 2007)
Interestingly this bacterium plays a positive environmental role through nitrogen recycling
(Puchenkova 1996) Strains of C freundii also contribute to the environment by sequenstering
heavy metals and degrading xenobiotic compounds (Kumar et al 1999 Sharma and Fulekar
2009)
31
111 Hypotheses and objectives
The study was based on the following hypotheses
i Certain microorganisms in DON contaminated agricultural soils are able to transform DON
to de-epoxy DON (de-epoxidation) under aerobic conditions and moderate temperatures
ii DON de-epoxidation may be achieved by a single microbial strain
iii Microbial DON detoxifying genes and associated regulators are able to express and
function in a closely related host organism
iv DON detoxifying microbial strains carry genes for de-epoxidation activity that are absent
from related species that have not evolved under DON selection pressure
The objectives of this research were to
i Select microbial cultures from soil having DON de-epoxidation activity under aerobic
conditions and moderate temperatures by analyzing potential DON contaminated soils in
various growth conditions namely aerobicanaerobic conditions different growth media
temperature and pH
ii Select for organisms with DON de-epoxidation activity from the mixed microbial culture
by treating with antibiotics heating subculturing on agar medium and incubating for a
prolonged period in media with a high DON concentration
iii Identify microbial de-epoxidation genes by producing a microbial genomic DNA library
from the DON de-epoxidizing strain followed by screening of the clones for DON de-
epoxidation
32
iv Determine microbial de-epoxidation genes by sequencing of the genome and comparative
analysis of the genome with closely related species and identification of putative DON
detoxification genes
33
CHAPTER 2
20 AEROBIC AND ANAEROBIC DE-EPOXIDATION OF MYCOTOXIN
DEOXYNIVALENOL BY BACTERIA ORIGINATING FROM AGRICULTURAL
SOIL
Published in World Journal of Microbiology and Biotechnology 2012 287-13
21 Abstract
Soil samples collected from different cereal crop fields in southern Ontario Canada
were screened to obtain microorganisms capable of transforming deoxynivalenol (DON) to
de-epoxy DON (dE-DON) Microbial DON to dE-DON transformation (ie de-epoxidation)
was monitored by using liquid chromatography-ultraviolet-mass spectrometry (LC-UV-MS)
The effects of growth substrates temperature pH incubation time and aerobic versus
anaerobic conditions on the ability of the microbes to de-epoxidize DON were evaluated A
soil microbial culture showed 100 DON to dE-DON biotransformation in mineral salts
broth (MSB) after 144 h of incubation Treatments of the culture with selective antibiotics
followed an elevated temperature (50ordmC) for 15 h and subculturing on agar medium
considerably reduced the microbial diversity Partial 16S-rRNA gene sequence analysis of the
bacteria in the enriched culture indicated the presence of at least Serratia Citrobacter
Clostridium Enterococcus Stenotrophomonas and Streptomyces The enriched culture
completely de-epoxidized DON after 60 h of incubation Bacterial de-epoxidation of DON
occurred at pH 60 to 75 and a wide array of temperatures (12 to 40ordmC) The culture showed
34
rapid de-epoxidation activity under aerobic conditions compared to anaerobic conditions This
is the first report on microbial DON to dE-DON transformation under aerobic conditions and
moderate temperatures The culture could be used to detoxify DON contaminated feed and
might be a potential source for gene(s) for DON de-epoxidation
Keywords biotransformation de-epoxidation deoxynivalenol soil bacteria vomitoxin
22 Introduction
Deoxynivalenol (DON commonly known as lsquovomitoxinrsquo) 3α 7α 15-trihydroxy-
1213-epoxytrichothec-9-en-8-one is a trichothecene produced by toxigenic Fusarium species
causing Fusarium head blight (FHB) or Gibberella ear rot (GER) disease on small grain
cereals and maize respectively (Starkey et al 2007 Foroud and Eudes 2009) DON
contamination in cereals and maize is widespread and can lead to tremendous losses to the
growers It has also detrimental effects on human and livestock health (Pestka and Smolinski
2005 Rocha et al 2005) The toxin interferes with the eukaryotic ribosomal peptidyl
transferase (Rpl3) activity resulting in human and animal mycotoxicosis (Ueno 1985 Bae
and Pestka 2008) Common effects of DON toxicity in animals include diarrhea vomiting
feed refusal growth retardation immunosuppression reduced ovarian function and
reproductive disorders (Pestka and Smolinski 2005 Pestka 2007) In plants DON causes a
variety of effects viz inhibition of seed germination seedling growth green plant
regeneration root and coleoptile growth (cited in Rocha et al 2005) In addition DON
enhances disease severity in sensitive host plants (Maier et al 2006)
35
Avoidance of DON contamination or reduction of toxin levels in contaminated grains
remains a challenging task Physical and chemical processes are either inappropriate or
prohibitively expensive to use to decrease the DON content of agricultural commodities
(Kushiro 2008 He et al 2010b) The use of Fusarium resistant crop varieties might be the
best method to reduce DON contamination However breeding for FHB or GER resistant
cultivars appears to be very difficult and time consuming as natural sources of resistance
against Fusarium-caused diseases is quantitative in nature (Ali et al 2005 Buerstmayr et al
2009) As an alternative expressing genes for enzymatic detoxification of DON in elite
cultivars may be an effective strategy for protecting the value of feeds and foods (Karlovsky
1999) Heterologous expression of eukaryotic trichothecene 3-O-acetyltransferase and UDP-
glucosyltransferase genes in cereals and Arabidopsis have resulted in reduced toxicity via
acetylation and glycosylation of the DON (Poppenberger et al 2003 Ohsato et al 2007)
It has been proposed that enzymatic de-epoxidation of trichothecene toxins could be
used to limit DON contamination (Zhou et al 2008 Foroud and Eudes 2009) DON belongs
to type B-trichothecenes which contain an epoxide ring at C12-13 and the epoxide is highly
reactive and mainly responsible for toxicity (Figure 2-1) (Swanson et al 1988 Eriksen et al
2004)
36
Figure 2-1 Mechanism of microbial DON de-epoxidation Removal of the oxygen atom from
the epoxide ring results in formation of a C=C double bond at C-12 position which reduces
cytotoxicity (Swanson et al 1988 Eriksen et al 2004)
Microbial DON de-epoxidation by single bacterial strains and by mixed cultures has
been reported (King et al 1984 Swanson et al 1987 He et al 1992 Kollarczik et al 1994
Fuchs et al 2002 Young et al 2007 Guan et al 2009 Yu et al 2010) However all of these
microbes originated from fish and other animals and were strictly anaerobic The recovered
microbial cultures and single strains only showed de-epoxidation of trichothecenes at low
(le15ordmC) and high (gt30degC) temperatures Such enzymes might have limited applications for
toxin decontamination in cereal grains The aim of the current study was to isolate the
microbes from soil capable of transforming DON to a de-epoxy metabolite under aerobic
conditions and moderate temperatures
De-epoxydation
Deoxynivalenol (DON)
De-epoxyDeoxynivalenol (dE-DON)
37
23 Materials and methods
231 Soil samples
The upper layer (0-30 cm) of soils along with crop debris were randomly collected
from cereal (eg maize wheat barley) crop fields during 2004-2006 in Southern Ontario
Canada Detailed of the crops and locations of the fields have been described in Jianwei Hersquos
Master Degree thesis (He 2007) Briefly soil samples were collected in sterile plastic bags
and transported to the laboratory Samples were sieved to remove large roots stones and
stored at 4ordmC until used The samples were mixed with grounded mouldy corn (containing 95
microgg DON) + F graminearum and incubated at 22-24degC for six weeks Five samples were
prepared for DON de-epoxidation activity analysis by dispersing 5 g soil in 50 mL distilled
water and storing at4degC until further use
232 Culture media
Media used in the study were nutrient broth (NB) Luria Bertani (LB) minimal
medium (MM) (He 2007) six mineral salts + 05 bacto peptone (MSB) (He 2007) tryptic
soy broth (TSB) corn meal broth (CMB) (Guan et al 2009) potato dextrose broth (PDB) six
mineral salts + 05 yeast extract broth (MSY) and soil extract (SE) MM was composed of
sucrose-100 g K2HPO4-25 g KH2PO4-25 g (NH4)2HPO4-10 g MgSO47H2O-02 g
FeSO4-001 g MnSO47H2O- 0007 g water- 985 mL and MSB was composed of the same
salts as MM but was supplemented with Bactotrade Peptone (05 wv) as the sole C source
instead of sucrose To prepare soil extract 100 g of the composite soil was homogenized in 1
L water The soil particles and plant debris were removed by passing the soil suspension
through Whatmanreg grade 3 filter paper (Sigma-Aldrich Oakville Ontario Canada) The
38
extract was autoclaved at 121degC for 30 min Media and ingredients were purchased either
from Fisher Scientific Inc (Fair Lawn New Jersey USA) Sigma-Aldrich (Oakvile Ontario
Canada) or Difco (Sparks Maryland USA) Prior to autoclaving pH of the broth media was
adjusted to 70 Solid media were prepared by adding 15 agar
233 Screening of soil samples
Frozen soil suspensions were thawed and vortexed to homogenize the microbial
population An aliquot (100 microL) of the suspension was immediately transferred to MSB PDB
and MM broth media DON at 50 microgmL was added to the microbial suspension cultures The
cultures were incubated at 22-24degC with continuous shaking at 180 revolutions per minute
(rpm) The extent of DON de-epoxidation was determined at 24 h 48 h 72 h 96 h 120 h and
144 h of incubation
234 Determination of de-epoxidation product
Microbial DON de-epoxidation was measured according to Guan et al (2009)
Samples were prepared by mixing equal volume of microbial culture and methanol and
allowed to stand for 30 min at room temperature Microorganisms in the mixture were
removed by passing them through a 045 microm Millipore filter (Whatmanreg Florham Park New
Jersey USA) The filtrates were used for de-epoxidation analysis by a liquid chromatography-
ultraviolet-mass spectrometry (LC-UV-MS) Identification and quantification of DON and its
transformation product was achieved using a Finnigan LCQ DECA ion trap LC-MS
instrument (ThermoFinnigan San Jose CA USA) The equipment detects 30-2000 mz
(molecular mass ion ratio) compounds and able to determine 001 micromL DON The HPLC
39
system was equipped with a Finnigan Spectra System UV6000LP ultraviolet (UV) detector
and an Agilent Zorbax Eclipse XDB C18 column (46times150 mm 35 μm) The binary mobile
phase consisted of solvent A (methanol) and solvent B (water) The system was run with a
gradient program 25 A at 0 min 25ndash41 A over 5 min isocratically at 41 A for 2 min
and 41ndash25 A over 1 min There was a 3 min post-run at its starting condition to allow for
the equilibration of the column The flow-rate was set at 10 mLmin UV absorbance was
measured at 218 nm Atmospheric pressure chemical ionization (APCI) positive ion mode
was used for MS detection A DON standard was used to adjust the instrument to its
maximum response The system was operated as follows shear gas and auxiliary flow-rates
were set at 95 and 43 (arbitrary units) voltages on the capillary tube lens offset multipole 1
offset multipole 2 offset and entrance lens were set at 3000 minus1700 minus370 minus850 minus3500
and minus4200 V respectively capillary and vaporizer temperatures were set at 200 and 450degC
respectively and the discharge needle current was set at 7 μA Identification of mycotoxin
was achieved by comparing their retention times UV spectra and mass spectrum with DON
and de-epoxy DON (Biopure Union MO USA) as standards Quantification of DON and
product was based on peak areas related to their UV and MS calibration curves
40
235 Biological DON de-epoxidation
To help identify the types of microorganisms causing the DON de-epoxidation MSB
medium was supplemented with either 100 microgmL penicillin G or streptomycin sulfate for
suppressing gram positive or negative bacterial growth and activities respectively Three
replications of antibiotic-treated media were inoculated with 100 microL of microbial culture
capable of de-epoxidizing DON The antibiotic-treated cultures were incubated at 27degC
spiked with 50 microgmL DON Controls were prepared without adding antibiotics in the
microbial culture Inhibition of DON de-epoxidation was analyzed after five day of
incubation
236 Reduction of microbial diversity by antibiotic treatments heating and
subculturing on agar medium
In order to reduce non-DON degrading microbial populations the DON de-
epoxidation capable cultures were sequentially treated with antibiotics having diverse modes
of action including i) dichloramphenicol that kills various gram positive Gram-negative and
anaerobic bacteria ii) trimethoprim that kills Gram-positive and negative bacteria iii)
penicillin that is lethal to Gram-positive bacteria and iv) cyclohexamide to kill filamentous
fungi and yeasts Antibiotics were purchased from Sigma-Aldrich Antibiotic stock solutions
(10 mgmL) were prepared with sterile water or methanol Microbial cultures in MSB
medium were treated with 100 microgmL of each antibiotic followed by incubation of the culture
at 50ordmC for 15 hours to kill undesirded microbial species After heating cultures were treated
with 50 microgmL DON and incubated at 27ordmC for five days DON de-epoxidation activity of the
microbial cultures was determined following the procedure described above Cultures that
41
showed gt99 de-epoxidation were considered for subsequent antibiotic and heat treatments
Aliquots of the achieved culture were spread on MSB agar plates and grown at room
temperature for few days Colonies appeared on the plates were collected and incubated in
MSB medium supplemented with 50 microgmL DON De-epoxidation ability of the subcultured
microbial cultures was determined after five days of incubation at 27degC by LC-UV-MS
method
237 Analysis of microbial population diversity
Terminal Restriction Fragment Length Polymorphism (T-RFLP) analysis
Initially microbial diversity was determined by terminal restriction fragment length
polymorphism (T-RFLP) fingerprint analysis (Bruce and Hughes 2000) Genomic DNA from
microbial cultures with and without enrichment was extracted using DNeasy Plant DNA
Extraction Mini Kit (Qiagen Inc Mississauga Ontario Canada) 16S-rDNA was amplified
by PCR using the HEX-labeled forward primer Bac8f
(5-agagtttgatcctggctcag-3) and
unlabeled reverse primer Univ1492r (5-ggttaccttgttacgactt-3) (Fierer and Jackson 2006)
Fifty microlitter PCR mixture for each sample was prepared having 1x HotStarTaq Master
Mix (Qiagen) 05 microM of each primer 50 microg of BSA and 50 ng of DNA The 35 PCR cycles
followed 60 s at 94degC 30 s at 50degC and 60 s at 72degC After size verification by agarose-gel
electrophoresis 50 ng of purified PCR products was digested for 3 h in a 20 microL reaction
mixture with 5 U of HhaI restriction enzyme (New England Biolabs Ipswich MA USA)
The size and relative abundance of the labeled T-RFLP fragments were separated by capillary
electrophoresis in an automated DNA sequencer equipped with fluorescence detector
(Applied Biosystem 3730 DNA Analyzer) The DNA fragments were analyzed with the Peak
42
Scannertrade Software v10 to create an electropherogram with the genetic fingerprints of the
microbial community
Analysis of V3-V5 region of the 16S-rDNA
To define the bacterial genera present in the enrichment culture the 16S-rDNA was
amplified using two sets of bacterial universal primers Primarily 16S-rDNA was PCR
amplified using the primers 46f-5rsquogcytaacacatgcaagtcga3rsquo (Kaplan and Kitts 2004) and
1371r-5rsquogtgtgtacaagncccgggaa3rsquo (Zhao et al 2007) PCR was performed in a total volume of
25 μL which contained 06 μM of each primer 1x of HotStarTaqreg Master Mix (Qiagen Inc)
and 20-40 ng of DNA The PCR cycles included the following steps 95ordmC (15 min) 35 cycles
of 94ordmC (20 s) 52ordmC (15 s) 72ordmC (1 min 30 s) and a 72ordmC (7 min) extension step The second
pair of universal bacterial primers were 27f-5rsquoagagtttgatcmtggctcag3rsquoand 1492r-
5rsquoggttaccttgttacgactt3rsquo (Lane 1991) PCR was conducted with 25 cycles following the
procedures of Tian et al (2009) Amplification was carried out in a GeneAmpreg PCR System
9700 (Applied Biosystems) Furthermore the PCR products were used as templates to
amplify the hypervariable V3-V5 region of the 16S-rDNA using primers 357f-
5rsquocctacgggaggcagcag3rsquo and 907r7 5rsquoccgtcaattcctttgagttt3rsquo (Yu and Morrison 2004)
After purification of the PCR products by QIAquick PCR purification kit two
amplicon libraries were prepared by ligating to pCR-TOPO vector (Invitrogen) and the library
clones were transformed into TOPO10 competent cells according to Invitrogenrsquos TOPO
cloning manual The clone libraries were plated on LB plates containing 25 μgmL
kanamycin Fifty colonies from each library were randomly picked up from the overnight
plates and inserts were sequenced as described by Geng et al (2006) PCR products were
43
sequenced with an automated ABI 3730 DNA analyzer (Applied Biosystems) The sequence
similarities of the amplified V3-V5 region were compared with the ribosomal database project
(RDP) (httprdpcmemsuedu) Sequences with greater than 95 identity were assigned to
the same bacterial genus
238 Factors influencing DON de-epoxidation
DON de-epoxidation ability of the enriched microbial culture was investigated under
different conditions as described below
Maintaince of active culture
To retain microbial de-epoxidation activity the active culture has been maintained in
MSB medium containing 50 microgmL DON
Media growth temperature and pH
Overnight microbial cultures (100 μL) were added to 2 mL of NB LB MM TSB
MSB CMB MSY PDB or SE broths The pH of the media was adjusted to 70 and the
experiments were conducted at 27degC temperature The effect of temperature was studied by
incubating the culture in MSB medium (pH 7) at 12 20 25 30 and 40ordmC To determine the
effects of pH MSB medium pH was adjusted to 55 60 65 70 75 and 80 Media with
different pHs were inoculated with aliquots of the overnight active culture and then incubated
at 27ordmC The cultures were supplemented with 50 μgmL DON before incubation Three
replications for each medium temperature and pH condition were evaluated
44
De-epoxidation under aerobic and anaerobic conditions
Three replicate microbial cultures of five mL each were grown in sterile 15 mL
centrifuge tubes under aerobic conditions and in an anaerobic chamber filled with 95 CO2
and 5 H2 (COY Laboratory Products Inc Ann Arbor MI USA) Three days after initiation
half (25 mL) of the cultures from anaerobic conditions was transferred to new centrifuge
tubes and brought to aerobic conditions Similarly half of the aerobically grown cultures were
placed into an anaerobic chamber Cultures moved from anaerobic to aerobic conditions were
shaken after opening the lids of the tubes in a laminar flow every eight hours intervals to
maintain aerobic conditions in the culture Lids of the culture tubes transferred to anaerobic
conditions were kept loose to allow oxygen to escape from the culture Two days after
transferring to the new conditions the cultures in both conditions were treated with 50 microgmL
DON and incubated at 27degC Every 12 h 500 microL were removed from each tube and analyzed
for microbial DON de-epoxidation activity by LC-UV-MS
239 Data analysis
Data were processed using Microsoft Excel Program 2007 Statistical analysis was
done using General Linear Model procedures in SAS version 91 (SAS Institute Inc Cary
NC USA 2002-03) The results were the average of three replications stated as mean plusmn
standard error Differences between means were considered significant at plt 005 according
to Tukeyrsquos-HSD test
45
24 Results
241 Analyses of DON to dE-DON biotransformation
A typical LC-UV-MS analysis where there is incomplete de-epoxidation of DON by a
particular soil suspension culture is shown in Figure 2-2 Analysis by LC with UV detection
and integration of peak areas showed 662 conversion of DON to dE-DON (Figure 2-2a)
Analysis LC with MS detection (expressed as total ion counts) indicated 643 conversion
(Figure 2-2b) LC analyses with UV and TIC (total ion count) detection are subject to
interferences due to co-eluting contaminants and therefore integration of small peaks was
very difficult making quantification inaccurate MS detection by SIM (selected ion
monitoring) can eliminate the influence of co-eluting contaminants For example the SIM LC
chromatogram for DON (Figure 2-2c) was generated by using the major ions resulting from
the APCI positive ion MS of DON (Figure 2-2e)
Similarly the SIM LC chromatogram for dE-DON (Figure 2-2d) was derived from the
MS ions of dE-DON (Figure 2-2f) The signal to noise ratios (SN) of peaks observed by SIM
were at least 20 times greater than those detected by TIC because there is reduced likelihood
of co-eluting peaks having the same ions Integration of the SIM peaks revealed a 616
conversion The three methods gave similar results but the SIM LC results were considered
to be the most accurate Using that method the conversion of DON to dE-DON by the mixed
culture occurred most quickly at 48 h and was completed by 144 h (Figure 2-3)
46
47
Figure 2-2 LC-UV-MS chromatographic analysis of DON and its de-epoxidated product dE-
DON obtained by culturing with selected soil microbes (a) LC-UV chromatogram of product
mixture monitored at 218 nm (b) LC-MS total ion chromatogram (TIC) of product mixture
(c) selected ion monitoring (SIM) chromatogram of cultured DON at mz 231 249 267 279
and 297 (d) SIM chromatogram of cultured dE-DON at mz 215 233 245 251 263 and 281
(e) atmospheric pressure chemical ionization (APCI) positive ion mass spectrum of DON (f)
APCI positive ion mass spectrum of dE-DON
48
242 Screening of soil samples
Initially samples originating from soils were analyzed in three types of culture media
to support the growth of fungi yeast and bacteria However cultures growing in PDB which
is a suitable substrate for many fungi and yeasts did not permit transform DON to dE-DON
(data not shown by microorganisms) A define medium (ie MM) was also used in the
sample screening assay None of the soil samples showed de-epoxidation activity in MM
(data not shown) From five soil samples one soil sample culture showed de-epoxidation of
DON in MSB The culture in MSB converted gt99 DON to dE-DON within 144 h (Figure 2-
3) Conversion began after a lag of 48 h and was linear thereafter with a rate of 1 per hour
49
Figure 2-3 Time course of relative biotransformation of DON to dE-DON by the initial soil
suspension culture in MSB medium The culture contained 50 μgmL DON at time 0 Culture
with three replications was incubated at room temperature with continuous shaking at 180
rpm The conversion of DON to dE-DON was analysed by LC-UV-MS at 24 h intervals Blue
and red lines are the percent conversion of DON and recovery of dE-DON respectively
50
243 Evidence of biological DON de-epoxidation
Treatment of active microbial culture with 100 microgmL of streptomycin completely
inhibited DON de-epoxidation In contrast the same concentration of penicillin G did not
affect activity of the culture (Figure 2-4) Streptomycin is effective against Gram-negative
bacteria while penicillin G is lethal to Gram-positive bacteria These experiments provided
evidence that DON de-epoxidation may be caused by certain Gram-negative bacteria
51
Figure 2-4 Microbial de-epoxidation of DON after five days of incubation The active DON
de-epoxidation culture was treated with either 100 microgmL penicillin G or streptomycin
(Strepto) in MSB Cultures having three replications were added 50 microgmL DON and
incubated at room temperatures with continuous shaking at 180 rpm For controls cultures
were without antibiotics DON to dE-DON transformation was monitored after five day of
incubation by LC-UV-MS method
0
20
40
60
80
100
120
Penicillin Strepto Control
DO
N d
e-e
po
xyd
ati
on
(
)
Antibiotics
52
244 Reduction of the microbial diversity
Plating the suspension cultures onto MSB agar revealed colonies with a variety of
sizes morphologies and colours reflecting a mixed culture (data not shown) To reduce the
complexity of the culture three steps of selection pressures were applied namely treatment
with antibiotics heat shocks and subculturing on solid medium The above indicated
treatments did not significantly affect DON de-epoxidation of the microbial cultures (data not
shown) T-RFLP profiles of the mixed culture before treatments showed six distinct peaks of
about 110 150 200 220 340 and 420 bp restriction fragments with the 110 bp peak being
the predominant one (Figure 2-5a)
After treatment with antibiotics and heat only three peaks at 110 150 and 325 bp
remained (Figure 2-5b) suggesting that there was a reduction in the microbial diversity
without affecting DON de-epoxidation After enrichments the bacteria species in the culture
changed their growth patterns as the electropherogram produced different sizes of pick
lengths compared to untreated culture
53
(b)
(a)
Rela
tive flu
ore
scen
ce
32000
0
0 500
16S-rDNA fragment size (bp)
100 200 300 400
Rela
tive flu
ore
scen
ce
32000
0
500
16S-rDNA fragment size (bp)
0 300200100 400
54
Figure 2-5 Analysis of microbial population reduction by terminal restriction fragment
length polymorphism (T-RFLP) (a) Restriction fragments of the 16S-rDNA of the untreated
culture and (b) fragmentation patterns of the enriched culture Picks along the X-axis showed
the fragmentation patterns of the 16S-rDNA which is related to the bacterial species diversity
The length of picks (along the Y-axis) showed abundance of terminal restriction fragments
related to the abundance of particular bacterial speciesgenera
55
245 Microbial population structures in the enriched culture
Microbial diversity in the enriched culture was determined by analyzing the sequences
of the V3-V5 region (~586 bp) amplified from the 16S-rDNAs of the microorganisms it
contained The sequence analysis showed the presence of Serratia Citrobacter Clostridium
Enterococcus Stenotrophomonas and Streptomyces in the enriched culture (Figure 2-6)
Out of 100 V3-V5 sequences 54 sequences were found to be belonging to Serratia
the predominant species of the culture The remaining sequences matched with 12
Citrobacter 8 Clostridium 14 Enterococcus 8 Stenotrophomonas and 4 Streptomyces
species
56
Figure 2-6 Microbial genera identified in the enriched DON de-epoxidizing microbial
culture The bacterial genera were determined by comparing the hypervariable regions (V3-
V5) of the 16S-rDNA using ribosomal database project (httprdpcmemsuedu) X-axis
showed the identified bacterial genera and Y-axis demonstrated the abundance of the V3-V5
sequences belonging to different bacterial genera
54
12 8
14
8 4
0
20
40
60 S
eq
uen
ce
fre
qu
en
cy
Microbial genera
57
246 Effects of media growth temperature pH and oxygen requirements on DON de-
epoxidation
Of the nine media that were tested DON de-epoxidation by the enriched microbial
culture only occurred in NB and MSB (Figure 2-7a) It appeared that C andor N
concentrations in the media were crucial for the enzymatic activity because less or more than
05 Bacto Peptone in MSB reduced the microbial DON transformation ability
DON de-epoxidation activity occurred over a wide range of temperatures from 12 to
40ordmC (Figure 2-7b) The fastest rates of DON de-epoxidation occurred between 25 to 40ordmC
De-epoxidation was observed within a relatively narrow range of pH (60 to 75) with
the most rapid DON biotransformation activity observed at pH 70 (Figure 2-7c) At pH 70
the enriched culture showed DON de-epoxidation activity 125 times faster than at pH 60 or
75 At pH 55 and 80 there was no observable DON de-epoxidation
58
0
20
40
60
80
100
120
12degC
20degC
25degC
30degC
40degC
DO
N d
e-e
po
xyd
ati
on
(
)
Temperature
0
20
40
60
80
100
120
DO
N d
e-e
pxyd
ati
on
(
)
Media
(a)
(b)
59
Figure 2-7 Effects of culture conditions on DON to de-epoxy DON transformation by the
enriched microbial culture For the media experiment (a) all the media were adjusted to pH 7
and the incubation temperature was 27degC For the temperature experiment (b) MSB pH 7 was
used For the pH experiment (c) MSB at 27degC was used Microbial cultures were inoculated
with 50 μgmL DON De-epoxidation was determined after 72 h of incubation Media MSY
(mineral salts + 05 yeast extract) LB (Luria Bertani) TSB (tryptic soy broth) MSB
(mineral salts + 05 bacto peptone) SE (soil extract) CMB (corn meal broth) PDB (potato
dextrose broth) and MM (minimal medium)
0
20
40
60
80
100
120
55
60
65
70
75
80
DO
N d
e-e
po
xyd
ati
on
(
)
pH
(c)
60
The enriched culture showed DON de-epoxidation activity under both aerobic and
anaerobic conditions (Figure 2-8) The fastest transformation (100 de-epoxidation after 60 h
of incubation) occurred in cultures (A) that were grown under aerobic conditions throughout
the culture growth period A medium rate of de-epoxidation (100 de-epoxidation after 72 h)
was observed in cultures that were initially grown in either aerobic or anaerobic conditions
followed by transferring to anaerobic or aerobic conditions (A+AN and AN+A) respectively
The lowest rate occurred (100 de-epoxidation after 96 h) in cultures grown in continual
anaerobic conditions (AN) The means comparison of microbial DON de-epoxidation
efficiencies at 60 h of incubation showed that cultures continually grown in conditions A and
AN were statistically different Biotransformation abilities of the cultures that were switched
from one conditions to another (A+AN and AN+A) were statistically similar among each
other but different from cultures that were only grown in conditions A or AN (plt005)
61
Figure 2-8 Microbial DON to dE (de-epoxy DON)-DON transformation activity under
aerobic and anaerobic conditions A = culture incubated and examined the activity only under
aerobic conditions A+AN = culture initially was grown in aerobic conditions and then
transferred to anaerobic conditions AN = cultures grown throughout the assay under
anaerobic conditions AN+A = culture initially grown under anaerobic conditions and then
transferred to aerobic conditions Differences of lower case letters are significantly different
in de-epoxidation capability (after 60 h) among incubation conditions according to Tukeyrsquos
HSD test (p lt 005) The experiment was conducted with three replications
62
25 Discussion
The current study describes a novel bacterial culture isolated from agricultural soil that
was capable of transforming DON to de-epoxy DON under aerobic and anaerobic conditions
at moderate temperatures Because it has been speculated that DON translocates to soil from
contaminated crops and DON that enters into the soil could be metabolized by microbes
(Voumllkl et al 2004) soil samples for the current study were mainly collected from fields where
cereal crops contaminated with DON were grown
In our initial sample screening steps upwards of 144 h of incubation were required to
achieve gt99 DON de-epoxidation Such a long period may have been due to the presence of
fast growing microorganisms that had suppressive effects on the growth or activity of the
DON transforming bacteria (Davis et al 2005) The sequential use of different antibiotics
with different modes of actions followed by thermal treatments and subculturing on solid
medium resulted in a reduction in the number of bacterial species as determined by T-RFLP
analysis Analysis of V3-V5 region of the 16S-rDNA showed that five bacterial genera were
present in the enriched culture The culture showed higher DON de-epoxidation activity (only
60 h required for gt99 biotransformation) compared to initial soil suspension culture that
required 144 h to complete the de-epoxidation This microbial culture could be used to isolate
the DON de-epoxidizing gene(s) applying metagenomics approaches (Daniel 2005)
The culture showed complete de-epoxidation activity both in MSB and NB media
However we have found that the use of nutritionally rich NB medium enhanced non-target
bacterial (eg Serratia lequifaciences) growth but this did not occur in MSB which is a
comparably poor medium To avoid the possible suppressive and detrimental effects of the
fast growing bacteria on the target bacteria MSB was used for enrichment and
63
characterization of the culture An important characteristic of the DON de-epoxidation
activity observed for the microbial culture in the current study is its activity at moderate
temperature (25-30degC) In contrast previously isolated cultures showed optimal activities at
low (le15degC) or high temperatures (37degC) (Guan et al 2009) Also the culture showed DON
de-epoxidation activity at neutral pH range (pH 60-75) in contrast to previous reports which
found that microbial trichothecene de-epoxidation occurred at acidic and basic pHs (Swanson
et al 1987 He et al 1992 Kollarczik et al 1994 Young et al 2007) However a microbial
culture from fish gut showed de-epoxidation under wide range of pHs (Guan et al 2009)
Probably the pH range reflects the environment in which the bacteria naturally grow which in
this case is similar to the soil pH of a typical crop field in Southern Ontario of Canada The
advantage of the present finding is that DON de-epoxidation activity of the culture was
optimal in a temperature range that matches the temperature range of activity for most arable
plants Therefore it is reasonable to suggest that the gene(s)enzyme(s) responsible for the
activity have potential application for enzymatic detoxification of DON in cereals (Karlovsky
1999)
The current observation that bacterial DON de-epoxidation activity occurred under
both aerobic and anaerobic conditions was interesting because DON de-epoxidation is a
reductive chemical reaction that might be expected to be promoted by anaerobic conditions
However there are previous reports of aerobic de-epoxidation of eldrindieldrin by soil
bacteria (Matsumoto et al 2008) and Goodstadt and Ponting (2004) described aerobic
reductive de-epoxidation by vitamin K epoxide reductasephylloquinone reductase A
definitive determination of the oxygen sensitivity of the enzyme responsible for DON de-
epoxidation will ultimately come from experiments with the purified protein
64
In conclusion this study reports on a novel bacterial source from soil that efficiently
de-epoxidizes DON at moderate temperatures under both aerobic and anaerobic conditions
This research may lead to the isolation of microbes and gene(s) encoding enzyme(s) capable
of de-epoxidizing DON that could create transgenic plants with reduced DON contamination
thus improving food and feed safety and reducing negative effects of the toxin on public
health
26 Acknowledgements
The authors gratefully acknowledged the Natural Sciences and Engineering Research
Council of Canada (NSERC) for awarding a scholarship to Rafiqul Islam for PhD program
The research was supported by the Agriculture and Agri-Food Canada Binational
Agricultural Research and Development Fund (AAFCBARD) project We are thankful for
the contributions of Jianwei He Xiu-Zhen Li and Honghui Zhu
65
CHAPTER 3
30 ISOLATION AND CHARACTERIZATION OF A NOVEL SOIL BACTERIUM
CAPABLE OF DETOXIFYING ELEVEN TRICHOTHECENE MYCOTOXINS
31 Abstract
Deoxynivalenol (DON vomitoxin) is one of the most commonly detected food and
feed contaminating mycotoxins worldwide In order to develop a control method effective
under aerobic conditions and moderate temperatures a bacterial strain ADS47 was isolated
capable of transforming DON to the less toxic de-epoxy DON The bacterial strain is
facultatively anaerobic Gram-negative rod shaped with an average cell size of 150 X 072
microm and has peritrichous (multiple) flagella The 16S-rRNA gene sequence had gt99
nucleotide identity to Citrobacter freundii a bacterium that belongs to the gamma subclass of
proteobacteria Strain ADS47 showed near 100 de-epoxidation of DON after 42 h of
incubation in nutrient broth (NB) under aerobic conditions moderate temperature (28˚C) and
neutral pH (70) However de-epoxidation could occur at 10-45˚C and pH 60-75 Under
anaerobic conditions near 100 de-epoxidation was observed after 72 h mainly during the
stationary growth phase Microbial DON de-epoxidation ability increased with increasing
DON concentrations in the culture media Inhibition of bacterial de-epoxidation by sodium
azide suggested that an active electron transport chain was required In addition to DON
strain ADS47 was able to de-epoxidize andor de-acetylate five type-A and five type-B
trichothecene mycotoxins This is the first report of a bacterial isolate capable of de-
epoxidizing DON and trichothecene mycotoxins under aerobic conditions and moderate
66
temperatures Strain ADS47 could potentially be used in bioremediation to eliminate DON
and other trichothecene mycotoxins from contaminated food and feed
Keywords aerobic de-epoxidation Citrobacter freundii isolation soil bacterium
trichothecenes
32 Introduction
Cereals are often contaminated with a variety of type-A and type-B trichothecene
mycotoxins produced by a number of toxigenic Fusarium species (Placinta et al 1999
Binder et al 2007 Foroud and Eudes 2009) Trichothecenes are tricyclic sesquiterpenes with
a double bond at the C9 10 position and a C12 13 epoxy group (Desjardins 2006) The two
types are distinguished by the presence of a ketone at position C8 in type-B which is absent
in type-A (Yazar and Omurtag 2008) The type-B toxin deoxynivalenol (DON) is the most
frequently found mycotoxin in food and feed around the world (Larsen et al 2004 Desjardins
2006 Starkey et al 2007) Different Fusarium species associated with Fusarium head blight
(FHB) or Gibberella ear rot (GER) disease of small grain cereals and maize are mainly
responsible for contaminating grains with DON and other type-A and type-B trichothecenes
These can cause serious mycotoxicosis in humans animals and plants (Ueno 1985 DrsquoMello
et al 1999 Bae and Pestka 2008) Common effects of DONTC toxicity in animals include
diarrhea vomiting feed refusal growth retardation immunosuppression reduced ovarian
functionsreproductive disorders and even death (Pestka and Smolinski 2005 Pestka 2007)
DON enhances FHB disease severity in wheat and maize (Proctor et al 1995 Desjardins
1996 Maier et al 2006)
67
Reduction of mycotoxin levels in contaminated agricultural products remains a
challenging task Current physical and chemical processes to decrease DON content are either
ineffective or prohibitively expensive (Kushiro 2008 He et al 2010b) However the use of
detoxifying microbes to biodegrade mycotoxins is an alternative (Awad et al 2010
Karlovsky 2011) The toxicity of DON is mainly due to the epoxide and there is a significant
reduction of DON toxicity (54) when it is reduced to produce de-epoxy DON Related
mycotoxins also show reductions in toxicity when they are de-epoxidized such as NIV (55)
and T-2 toxin (400) (Swanson et al 1988 Eriksen et al 2004) Bacillus arbutinivorans
strain LS100 and Eubacterial strain BBSH 797 originating from chicken intestine and rumen
fluid respectively are capable of cleaving the epoxide ring of DON under anaerobic
conditions and high temperature (37degC) (Fuchs et al 2002 Yu et al 2010) Strain BBSH 797
which can de-epoxidize DON scirpentriol and T-2 triol trichothecene mycotoxins has been
used as an additive for poultry and swine diets (Karlovsky 2011) However there are no
reports of microbes able to de-epoxidze DON and other trichothecenes under aerobic
conditions and at moderate temperatures
To develop an effective method for eliminating DON and trichothecene mycotoxins
from contaminated grains soils from fields with FHBGER infected crops were screened for
DON de-epoxidizing microbes (refer to chapter 2 Islam et al 2012) One soil sample yielded
a mixed microbial culture was partially enriched for bacteria with DON de-epoxidizing
activity (chapter 2) The current study describes the purification and characterization of a
novel DON de-epoxidizing bacterium from this culture
68
33 Materials and methods
331 Microbial source
A mixed microbial culture originating from a screen of more than 150 separate soil
samples collected from maize and wheat fields near Guelph Ontario Canada (Islam et al
2012) was used for enrichment purification and characterization of the DON de-epoxidizing
bacterium
332 Media and chemicals
The media used were nutrient broth (NB) 110 strength NB (110 NB) minimal
medium (MM) consisting of six mineral salts (MS) + 100g per liter sucrose (Islam et al
2012) MS + 05 Bacto Peptone (MSB) MS + 03 Bacto Peptone (MSC) MS + 05
yeast extract (MSY) MS + 200 microgmL DON (MS-DON) sterile water-soluble soil extract
(SE) (Islam et al 2012) potato dextrose broth (PDB) and Luria Bertani (LB) Prepared media
and media components were obtained from Fisher Scientific (Fair Lawn New Jersey USA)
Sigma-Aldrich (Oakville ON Canada) andor Difco (Sparks Maryland USA) Media pH
was adjusted as required prior to autoclaving For solid media preparation 15 agar was
added DON and all trichothecene mycotoxins were purchased from Biomin (Romer Lab
Union MO USA)
333 Isolation of the DON de-epoxidizing bacterial strain
Aliquots (20 microL) of the microbial culture partially enriched for DON de-epoxidizing
ability (Islam et al 2012) was transferred to 05 mL MSB (pH 70) in sterile 15 mL
Eppendorf tubes After addition of 200 microgmL pure DON the culture was incubated at room
69
temperature with continuous shaking at 220 revolutions per minute (rpm) Seven to ten days
later 20 microL of the culture was transferred to fresh MSB supplemented with 200 microgmL DON
The culture was then incubated for 7 to 10 days under the above conditions This procedure
was repeated 12 times
After three months of repeated transfers in broth with 200 microgmL DON 100 microL of the
culture was serially diluted in 09 mL MSB containing 05 Tweenreg20 (Sigma Aldrich) to
reduce bacterial cell aggregation (Mabagala 1997) The dilutions were vigorously shaken for
ten minutes A 100 microL aliquot from each dilution was spread on MSB + agar (MSA) The
MSA plates were incubated at room temperature until single colonies appeared
Single colonies from plates were transferred to 1 mL MSB+ 50 microgmL DON and
incubated at room temperature with continuous shaking at 180 rpm under aerobic conditions
After three days the DON de-epoxidation ability was examined by mixing an equal volume
of the bacterial culture and methanol and incubating for 30 min at room temperature to lyse
the cells Insoluble materials in the mixture were removed by passing it through a 045 microm
Millipore filter (Whatmanreg Florham Park New Jersey USA) The level of de-epoxy DON
(dE-DON also known as DOM-1) in the filtrate was determined by liquid chromatography-
ultraviolet-mass spectrometry (LC-UV-MS) (Guan et al 2009)
Individual colonies that showed de-epoxidation activity were serially diluted in
MSB+05 Tweenreg20 and plated onto MSA Single colonies that appeared on the agar plate
were incubated in MSB as described above and retested for DON de-epoxidation ability To
ensure that pure cultures were obtained this was repeated 10 times and individual colonies
were selected on MSA each time
70
334 Identification of a DON de-epoxidizing bacterium
Strain ADS47 with DON de-epoxidation ability was obtained from a single colony
after 10 serial transfers and was grown on MSA at room temperature under aerobic
conditions for 72 h Colony color shape edge and elevation were observed The cell
morphology of the bacterium was examined with a transmission electron microscope at the
University of Guelph Guelph Ontario Canada Whole bacterial cells were placed directly on
a formvar and carbon-coated 200-mesh copper grid They were washed two times by touching
the grids to a drop of clean dH2O and the excess water was blotted off The samples were
negatively stained using 1 Uranyl Acetate (aq) for five seconds Electron microscopy was
performed on a Philips CM10 transmission electron microscope at 60kV under standard
operating conditions (Philips Optics Eindhoven Netherlands) Gram staining was performed
using the procedure described in the commercial gram stain kit from BD Canada
(Mississauga Ontario Canada)
Total bacterial DNA was extracted using a DNeasy Plant DNA Extraction Mini Kit
(Qiagen Inc) The 16S-rDNA was PCR amplified using the primers 46f-
5rsquogcytaacacatgcaagtcga3rsquo (Kaplan and Kitts 2004) and 1371r-5rsquogtgtgtacaagncccgggaa3rsquo
(Zhao et al 2007) PCR was performed in 25 microL containing 06 microM of each primer 1x of
HotStarTaqreg Master Mix (Qiagen Inc) and ~40 ng of DNA The PCR cycles were 95ordmC (15
min) 35 cycles of 94ordmC (20 s) 52ordmC (15 s) 72ordmC (1 min 30 s) and a 72ordmC (7 min) extension
step Amplification was carried out in a GeneAmpreg PCR System 9700 (Applied Biosystems)
PCR products were sequenced with an automated ABI 3730 DNA analyzer (Applied
Biosystems) The 16S-rRNA gene sequence similarity was performed by BlastN using the
Ribosomal Database Project (RDP) (httprdpcmemsueduseqmatch)
71
A phylogenetic tree was constructed by comparing the partial 16S-rDNA sequence of
strain ADS47 with the 11 known Citrobacter species (Bornstein and Schafer 2006 Brenner et
al 1993) The Citrobacter strains were C freundii [FN997616] C koseri [AF025372] C
amalonaticus [AF025370] C farmeri [AF025371] C youngae [AB273741] C braakii
[AF025368] C werkmanii [AF025373] C sedlakii [AF025364] C rodentium [AF025363]
C gillenii [AF025367] and C murliniae [AF025369] Escherichia coli O157H7 [AE005174]
was used as an out group strain The analysis was performed using distance matrix with a gap
opening of 100 and a gap extension of 01 by a server based ClustalW2 program
(httpwwwebiacukToolsmsaclustalw2) A dissimilarity tree was created and viewed
using a TreeView X software (Page 1996)
335 Effects of DON concentration aeration substrate temperature pH and antibiotic
To determine if DON concentration can affect the growth of the isolated bacteria
tubes containing 5 mL of NB or MSB with 20 200 or 500 microgmL of DON were inoculated
with strain ADS47 that had been grown for 16 h with continuous shaking at 220 rpm and then
adjusted to cell density at 025 OD600nm (107 cellsmL) The cultures were incubated at 28degC
with continuous shaking at 180 rpm and 05 mL was removed every 12 h to measure
OD600nm The culture (05 mL) was then analysed for DON de-epoxidation activity by LC-
UV-MS (Guan et al 2009) as described above Three replicate cultures were tested
To examine the effect of aeration five mL of NB or MSB containing 50 microgmL DON
were inoculated with an overnight culture of strain ADS47 prepared as described above and
incubated at 28˚C with continuous shaking at 180 rpm One mL of culture was removed every
six hours and bacterial growth was determined by measuring cell density at OD600nm At the
72
same time points culture samples were analysed for DON to dE-DON transformation ability
(Guan et al 2009) The experiment was repeated with MSB medium containing 50 microgmL
DON under anaerobic conditions as described by Islam et al (2012) with DON to dE-DON
transformation ability being determined every 6 h Three replicate cultures were prepared for
each assay
The effect of substrate on DON de-epoxidation ability was tested after growing
ADS47 for 72 h in NB 110 NB MM MS-DON MSB MSC MSY SE PDB or LB
containing 50 microgmL DON For this assay the media were maintained at pH70plusmn02 and 28degC
for 72 h The effect of temperature was examined with MSB containing 50 microgmL DON at 10
15 20 25 30 35 40 45 or 50degC The effect of pH on growth of strain ADS47 was examined
with MSB adjusted to pH 55 60 65 70 75 or 80 and the cultures were incubated at 28degC
for 72 h Three replicate cultures were used for each assay
Overnight cultures of ADS47 in MSB were treated with 001 and 0001 (wv) of
an antibiotic sodium azide (SA) (Sigma-Aldrich) Controls were prepared in MSB without
SA Three replicate cultures were spiked with 50 microgmL DON DON to dE-DON
transformation ability of the bacterial culture was analysed (Guan et al 2009) after 72 h at
28degC with continuous shaking at 180 rpm Three replicate cultures were tested
336 Localization of DON to dE-DON transformation ability in the bacterium
Bacterial cell extracts were prepared using B-PER II Bacterial Protein Extraction
Reagent following the manufacturerrsquos instructions (PI 78260 Thermo Scientific Rockford
IL USA) ADS47 was grown in 15 mL MSB for 48 h under the growth conditions described
above A culture (05 mL) was centrifuged at 5000 x g for 10 minutes and the pellet was
73
collected B-PER II reagent (2 mL) was added to the pellet and the cells were lysed by
pipetting the suspension up and down several times The cell lysate was treated with 20 microL
Halt Protease Inhibitor Cocktail (Thermo Scientific Rockford IL USA) Culture filtrate was
prepared by passing 05 mL of the culture through a 022 microm Millipore filter (Fisher
Scientific) The remaining 05 mL culture was used as control Cell lysate culture filtrate and
control were incubated with 50 microgmL DON and incubated at 28degC DON to dE-DON
transformation ability of the cell lysate culture filtrate and control were analysed after 72 h
incubation (Guan et al 2009) Three replicate cultures were tested
337 Detoxification of additional trichothecene mycotoxins
Strain ADS47 was evaluated for its ability to degrade five type-A trichothecene
mycotoxins (HT-2 toxin T-2 toxin T2-triol diacetoxyscirpenol and neosolaniol) and five
type-B trichothecene mycotoxins (nivalenol verrucarol fusarenon X 3-acetyl-
deoxynivalenol and 15-acetyl-deoxynivalenol) One and a half mL NB was added with 100
microgmL of each mycotoxin and then inoculated with 100 microL of ADS47 culture grown
overnight as previously described The culture of NB + mycotoxin + ADS47 was incubated at
28degC with continuous shaking at 180 rpm and DON to dE-DON transformation ability of the
culture was analysed by LC-UV-MS (Guan et al 2009) Five replicate cultures were tested for
each trichothecene mycotoxin
74
338 Statistical analysis
Data were processed using Microsoft Excel Program 2007 Statistical analysis was
performed using General Linear Model procedure in SAS version 91 (SAS Institute Cary
NC USA 2002-03) Replications were averaged and stated as mean plusmn standard error The
mean differences were considered significant at plt 005 according to Tukeyrsquos-HSD test
75
34 Results
341 Isolation and identification of a DON de-epoxidizing bacterium
A DON de-epoxidizing bacterium designated strain ADS47 was isolated after 12
transfers over three months from cultures growing in MSB containing 200 microgmL DON
Strain ADS47 produced small to medium size circular convex and white colonies on MSA at
room temperature under aerobic conditions (Figure 3-1a) The bacterium is rod shaped and
150 x 072 microm in size (Figure 3-1b) It has numerous flagella that cover the surface of the cell
(peritrichous flagella) and is Gram-negative
BlastN analysis of the 16S-rDNA sequence of strain ADS47 showed the most
similarity to the eleven known Citrobacter species in the gamma proteobacteria of the
Enterobacteriaceae In particular the 16S-rDNA nucleotide sequence of ADS47 was more
than 99 identical to C fruendii (Figure 3-2) The next most similar sequences were a cluster
of C werkmanii and C murliniae followed by more distantly related C braakii C gilleni
and C youngae A separate cluster was formed by C rodentium C farmeri C sedlakii C
koseri and C amalonaticus
76
Figure 3-1 (a) Colony color shape and size of the mycotoxin degrading bacterial strain
ADS47 on mineral salts agar (MSA) medium (b) Transmission electron microscopic (TEM)
image of the Citrobacter freundii strain ADS47 The bacterium showed multiple flagella
(peritrichous flagella) and numerous fimbriae around the cell surface
(a) (b)
77
Figure 3-2 Dendrogram based on 16S-rDNA sequences for strain ADS47 and 11 reference
Citrobacter species An E coli O157H7 strain was considered as an out group species The
tree was constructed using a genetic distance matrix by ClustalW 2 and viewed by
TreeViewX software The distance was converted to scale and shown by branch lengths (scale
bar 001 which indicated the 16S-rDNA nucleotide sequence variability between the bacterial
species) GenBank accession numbers for 16S-rDNA sequences of the species used for
comparison are shown in materials and methods section (334)
78
342 Effects of DON concentrations on bacterial DON de-epoxidation
Growth of ADS47 aerobically in MSB with 20 200 and 500 microgmL DON was not
significantly different from that observed in control cultures without DON (Figure 3-3a)
Similarly increasing DON concentrations did not affect bacterial growth in NB medium
(Figure 3-3b)
DON de-epoxidation activity determined as microgmL DON convertedcell density
(OD600) was measured lowest in MSB+20 microgmL DON and highest in MSB+ 500 microgmL
DON containing cultures (Figure 3-3c) The highest rate of dE-DON formation was 115 microgh
in ADS47 cultures in MSB with 500 microgmL DON between 48 to 72 h All of the DON was
converted in the cultures containing 20 microgmL DON and 200 microgmL DON between 48-60 h
and 60-72 h respectively In 200 microgmL and 500 microgmL of DON cultures the rate of de-
epoxidation increased significantly after 36 h
79
00
04
08
12
16
0 24 36 48 60 72 96
OD
600n
m
Incubation time (h)
Control
20 microgmL
200 microgmL
500 microgmL
(b)
(a)
80
Figure 3-3 Effects of DON concentration on bacterial growth and de-epoxidation activities
Strain ADS47 was grown in MSB and NB media with three different DON concentrations 20
microgmL 200 microgmL and 500 microgmL Control cultures were prepared without DON Cultures
were grown at 28degC with continuous shaking at 180 rpm (a) Microbial density (OD600 nm) in
MSB with different DON concentrations was measured at 24 36 48 60 72 and 96 h (b)
Microbial growth (OD600 nm) in NB having different DON concentration was measured at 24
48 72 and 96 h (c) Bacterial DON de-epoxidation activity in 20 200 and 500 microgmL DON
containing cultures at different time points DON de-epoxidation was measured in microgmL per
OD600 nm Three replicate cultures for each assay were analysed
(C)
81
343 Influence of growth conditions and antibiotic on microbial DON de-epoxydation
Aerobic and anaerobic conditions
ADS47 grew more quickly uner aerobic conditions under MSB+50 microgmL DON than
in MSB+50 microgmL DON under anaerobic conditions (Figure 3-4a) Aerobic growth was
faster in NB+50 microgmL DON compared to MSB+50 microgmL DON
However the DON de-epoxidation activities on a cell density basis were very similar
in NB+50 microgmL DON compared to MSB+50 microgmL DON (Figure 3-4b) Also DON de-
epoxidation activity of ADS47 under aerobic conditions in MSB+50 microgmL DON was higher
than in the same medium under anaerobic conditions (Figure 3-4b)
82
(b)
(a)
83
Figure 3-4 Bacterial growth and DON de-epoxidation capabilities under aerobic and
anaerobic conditions (a) Microbial growth under aerobic conditions in MSB and NB medium
and anaerobic conditions in MSB medium (b) Microbial DON de-epoxidation ability under
aerobic conditions in MSB and NB media and anaerobic conditions in MSB medium
Cultures media having 50 microgmL DON were inoculated with overnight culture of strain
ADS47 Initial microbial concentration in the cultures was adjusted to 025 OD600nm Cultures
were incubated at 28degC having three replications Bacterial growth at OD600nm and DON de-
epoxidation in microgmL per OD600nm were measured at six hour interval
84
Media temperature and pH
Growth of ADS47 in media containing 50 microgmL DON under aerobic conditions at 72
h was three-fold higher in NB and LB than in SE two-fold higher than in 110 NB and MSC
and did not occur in MS-DON (Figure 3-5a) Growth in MM MSB and MSY was
intermediate
Over the 72 h DON de-epoxidation per cell density was highest in MSB and MSY and
approximately 25 lower in NB and LB and lower by 75 and 87 in 110 NB and MSC
respectively (Figure 3-5b) No DON de-epoxidation was observed in MS-DON in which
DON was used as a carbon source Bacteria grown in MM broth which contains sucrose as a
sole carbon source were unable to de-epoxidize DON No DON de-epoxidation was observed
in SE and PDB grown cultures
Growth of ADS47 under aerobic conditions for 72 h in MSB+50 microgmL DON was
greatest between 25-40degC but no growth occurred at 50degC (Figure 3-6a) DON de-
epoxidation activity was greatest and equivalent between 20-40degC and lower at 10 15 and
45degC temperatures (Figure 3-6b)
ADS47 growth under aerobic conditions for 72 h in MSB+50 microgmL DON was
limited to neutral pHs (ie 60 - 75) (Figure 3-7a) The transformation of DON to dE-DON
occurred at the same pH range and DON de-epoxidation activity was not significantly
different between pH 60 to 75 (Figure 3-7b)
85
b
c
a
b
a
d c
b b
(a)
(b)
b
c
b
a a
d
86
Figure 3-5 Effects of media on bacterial growth and DON de-epoxidation activity (a)
Microbial growth (at OD600nm) on different culture media Media pHs were adjusted to
70plusmn02 (b) Bacterial DON to dE-DON transformation in microgmL by OD600nm in different
media after 72 h The cultures having three replications were incubated for 72 h at 28degC and
DON de-epoxidation was measured The Media NB = nutrient broth 1NB = 110 strength
NB MM = minimal medium MS-DON = six mineral salts + 200 microgmL DON as sole C
source MSB = six mineral salts + 05 bacto peptone MSC = six mineral salts broth + 03
bacto peptone MSY = six mineral salts with 05 yeast extract SE = soil extract PDB =
potato dextrose broth and LB = Luria Bertani Means with the same label are not significantly
different according to Tukeyrsquos HSD test (p lt 005)
87
(a)
(b)
88
Figure 3-6 Effects of temperature on microbial growth and DON de-epoxydation capability
(a) Microbial growth (at OD600nm) at different temperatures after 72 h of incubation (b)
Bacteria DON de-epoxidation in microgmL by OD600nm in different temperatures The most
efficient temperature range (ge40 microgmL DON de-epoxidation) is marked by dot lines The
experiment was conducted in MSB medium at pH 70plusmn02 with three replications
89
(a)
(b)
90
Figure 3-7 Influence of pH on microbial growth and DON de-epoxydation (a) Microbial
growth (OD600nm) in MSB at different pH (b) Bacterial DON to de-epoxy-DON
transformation in microgmL by OD600nm in different pH Dot lines indicated the efficient de-
epoxidation (ge40 microgmL DON conversion) pH range Experiment was conducted at 28degC
with three replications
91
Antibiotic
Treatment of ADS47 at 72 h in MSB+50 microgmL DON with sodium azide (SA) at
0001 and 001 reduced DON de-epoxidation activity to 5 (25 microgmL) and 0
respectively The control with no SA added was able to de-epoxidize DON completely
(100) (Figure 3-8)
Figure 3-8 Effects of sodium azide (SA) on bacterial DON de-epoxidation activity
Microbial culture in MSB+50 microgmL DON was treated with 001 (SA-1) and 0001 (SA-
2) sodium azide In control (C) no SA was added The experiments with three replications
were incubated for 72 h at 28degC and DON de-epoxidation was measured by LC-UV-MS
method
0
20
40
60
80
100
120
SA1 SA2 C
D
ON
de
-ep
oxyd
ize
d
92
344 Localization of bacterial DON de-epoxidation
A cell lysate prepared from ADS47 grown for 48 h in MSB transformed 20 (10
microgmL) of DON added at 50 microgmL to dE-DON within 72 h (Figure 3-9) In contrast a cell-
free culture filtrate did not show DON de-epoxidation activity
Figure 3-9 Determination of DON to de-epoxy DON transformation by bacterial culture
filtrate and cell extract The treatments having three replications were spiked with 50 microgmL
DON and incubated at 28degC for 72 h In control strain ADS47 was incubated in MSB broth
with 50 microgmL DON DON de-epoxidation activity in percentage was examined
0
20
40
60
80
100
120
Filtrate Extract Control
D
ON
de
-ep
oxyd
ize
d
93
345 Bacterial detoxification (de-epoxidtaion and de-acetylation) of other trichothecene
mycotoxins
Aerobic cultures of ADS47 grown for 72 h in NB+100 microgmL HT-2 converted 74 of
the toxin to de-epoxy HT-2 toxin and 26 to a de-acetylated product (Figure 3-10 and Table
3-1)
In a culture treated with T2-toxin 46 was converted to de-epoxy T-2 16 was
converted to an unknown compound and 38 was not converted In ADS47 culture with
diacetoxyscirpenol (DAS) 61 was transformed into de-epoxy DAS and 39 was de-
acetylated DAS (39) Neosolaniol (NEO) was transformed to de-epoxy NEO (55) and
de-acetylated NEO (45) T2-triol was transformed in to de-epoxy T2-triol (47) and de-
acetylated T2-triol (53)
Aerobic cultures of ADS47 were also grown for 72 h in NB+100 microgmL of one of five
type-B trichothecenes For nivalenol (NIV) 92 was de-epoxydized while the remaining 8
NIV was transformed to DON 3-acetyl-deoxynivalenol (3-ADON) was transformed into
three products de-epoxy NIV (5) de-epoxy DON (54) and DON (41) For 15-
acetyldeoxynivalenol (15-ADON) 8 was transformed to de-epoxy 15-ADON and 92 to
de-epoxy DON (dE-DON) FusarenonX (FUS) had 95 transformed into de-epoxy FUS
while 97 of verrucarol (VER) was transformed to de-epoxy VER The remaining 5 FUS
and 3 VER were not transformed
94
Figure 3-10 Determination of microbial de-epoxidation and de-acetylation of HT-2 toxin by
LC-UV-MS Bacterial culture in NB was added with 100 microgmL HT-2 toxin Bacterial HT-2
toxin transformation ability was examined after 72 h of incubation at 28degC The upper panel
illustrated mass spectrometry chromatograph of the de-acetylated (red peak) and de-
epoxydized (blue peak) products Arrows indicated the targeted acetyl (red) and epoxy (blue)
functional groups of HT-toxin The middle and lower panels showed the mass spectra of the
de-acetylated (dA) and de-epoxidized (dE) compounds Arrows on the upper panel indicated
the targeted acetyl (red) and epoxy (blue) functional groups of HT-2 toxin Red and blue
arrows on the middle and lower panels pointed at the degradation sites of HT-2 toxin
95
Table 3-1 Microbial transformation of type-A and type-B tricthothecene mycotoxins after 72
h of incubation under aerobic conditions at room temperature DAS=diacetoxyscirpenol
DON=deoxynivalenol FUS=FusarenonX 3AON=3-acetyl-deoxynivalenol and
15ADON=15-acetyl-deoxynivalenol
1 Convertion of trichothecene mycotoxins to either DON or an unknown compound
Trichothecene
mycotoxin
Untransformed
trichothecenes ()
Transformed products of
trichothecenes ()
De-
epoxy
De-
acetylate
Other1
Type-A
HT-2 toxin 0 74 26 -
T-2 toxin 38 46 - 16
(unknown)
DAS 0 61 39 -
Neosolanol 0 55 45 -
T2-triol 0 47 53 -
Type-B
DON 0 100 - -
Nivalenol 0 92 - 8 (DON)
3-ADON 0 59 41 (DON)
15-ADON 0 100 - -
FUS 5 95 - -
Varrucarol 3 97 - -
96
35 Discussion
The ADS47 strain of C freundii was isolated in the present study by a prolonged
incubation of a mixed microbial culture with a high concentration DON since the bacterium
efficiently de-epoxidized (detoxified) DON and number of other trichothecene mycotoxins
under aerobic and anaerobic conditions at moderate temperatures and neutral pH The total
procedures for enrichment and isolation of the strain ADS47 is depicted in supplemental
Figure S3-1 The reductive de-epoxidation activity may be due to a cytoplasmic enzyme
which would be in agreement with a previous study that showed de-epoxidation of
epoxyalkanes was catalyzed by a bacterial cytoplasmic flavoprotein (Westphal et al 1998)
The ability of the currently isolated strain of C freundii to biodegrade environmental
pollutants such as aminoaromatic acids and heavy metals (copper) is also likely mediated by
cytoplasmic enzymes since these compounds need to be taken up by the cells before they are
metabolized (Savelieva et al 2004 Sharma and Fulekar 2009) In contrast oxidative
transformation of DON to 3-keto DON by microbial extracellular enzyme has been reported
(Shima et al 1997)
Isolation of a trichothecene de-epoxidizing microorganism from a mixed microbial
community is very challenging Although trichothecenes are toxic to eukaryotes prokaryotes
appear to be resistant to these mycotoxins (He et al 1992 Rocha et al 2005) The present
study showed that the bacteria grew well in media with high concentrations of DON (500
microgmL) Bacterial de-epoxidation of DON involves the removal of an oxygen atom from the
functional epoxy ring of the tricyclic DON structure (Yoshizawa et al 1983) Such co-
metabolic degradation does not provide the bacteria with any nutrients from the reacting
compounds (Horvath 1972 Fedorak and Grabic-Galic 1991) This is supported by the results
97
of the current study which showed that strain ADS47 was unable to grow in media with DON
as a sole carbon source and increasing the concentration of DON did not increase ADS47
growth in MSB Hence DON could not be used as sole nutrient source in the mixed culture
enrichment process
Many previous attempts to obtain DON de-epoxidizing microbes have failed
particularly under aerobic conditions because media or laboratory conditions were
inappropriate to support growth or initially active cultures lost de-epoxidizing function after
serial transfer (Karlovsky 2011)
Media composition appears to be a crucial factor for microbial trichothecene de-
epoxidation activity (Voumllkl et al 2004) Effective biotransformation of trichothecene
mycotoxins by the fish digesta microbial culture C133 occurred in full media (FM) but a low
level of de-epoxidation occurred in media having a single nitrogen source and no
transformation activity was observed in the common bacterial medium LB (Guan et al
2009) Zhou et al (2010) also showed that chitin de-acetylation by soil bacterial strains
occurred more rapidly in yeast extract compared to common bacterial media Therefore a
screening strategy for microbial DON de-epoxidation ability should use different media in
parallel as was done in this study
This study observed efficient de-epoxidation in common bacterial media (NB and LB)
as well as in mineral salts medium containing yeast extract (MSY) or 05 Bacto Peptone
(MSB) However reduction of Bacto-Peptone to 03 (MSC) significantly reduced the
bacterial de-epoxidation activity suggesting that the activity is sensitive to the availability of
protein and amino acids The study found that medium containing sucrose as carbon source
(MM) and a commonly used medium PDB blocked microbial DON de-epoxidation even
98
though bacteria grew in those growth media These suggest that media containing high sugar
contents had inhibitory effects on microbial de-epoxidation enzymatic activity Previously
sugars have been found to have negative effects on the production andor activity of bacterial
enzymes (Saier and Roseman 1971)
Losses of microbial functions under laboratory conditions limited researchers to obtain
economically important microbial genetic sources The soil borne Curtobacterium sp strain
114-2 efficiently de-epoxydized nivalenol but unfortunately the strain lost its activity in
latter generations (Ueno et al 1983) Instability of de-epoxidation activity of ADS47
observed in cultures grown on agar media without DON resulted in the loss of DON de-
epoxidation activity by more than 80 of the colonies after one generation However strain
ADS47 retained de-epoxidation activity after ten subcultures in MSB (data not shown)
The non-specific de-epoxidation of DON made it impossible to use DON as a sole
substrate C source for selective isolation of ADS47 Thus a number of techniques were
utilized to select ADS47 and suppress other bacteria including the use of a variety of
antibiotics and culture at elevated temperatures (Islam et al 2012) Selection of the isolate
ADS47 required repeated subculturing and prolonged exposure to media containing 200
microgmL DON After three months of selection with DON culture that was obtained had a
DON de-epoxidation activity that was up to 30-fold higher than the mixed culture As growth
of ADS47 was not affected by 200 microgmL DON the prolonged incubation may have reduced
populations of other bacteria that are more sensitive to DON Enrichment and isolation of a
DON to 3-keto DON transforming bacterial strain E3-39 by prolonged incubation of a soil
microbial culture with 200 microgmL DON was reported by Shima et al (1997)
99
This study demonstrated that microbial DON de-epoxidation occurred mainly at the
stationary growth phase A DON degrading strain of Citrobacter may have gained energy
from DON by co-metabolic cleavage of the epoxy ring under less nutrient conditions
Microbial use of energy from co-metabolic reaction has been suggested by Chiu et al (2005)
The rate of de-epoxidation activity increased when ADS47 grew at higher concentrations of
DON Taken together a prolonged exposure to high concentration DON may have resulted in
non-DON metabolizing microbial populations declining by DON toxicity while facilitated
the population of DON metabolizing bacterium by providing energy
DON is a small molecule and is simply absorbed by cells (Sergent et al 2006)
Microbial DON de-epoxidation occurred in intact and lysed bacterial cells In addition
treating the ADS47 culture with SA which blocks gram negative bacterial electron transport
and activity inhibited DON de-epoxidation activity (Lichstein and Malcolm 1943 Erecinska
and Wilson 1981) implying that an active electron transport system required for microbial de-
epoxidation reaction (He et al 1992) The data suggest that a cytoplasmic epoxy reductase is
involved in the DON de-epoxidation reaction in the ADS47 microbial strain that was isolated
in the present study
Since agricultural commodities are often contaminated with multiple fungal
mycotoxins the ability of bacteria like ADS47 to degrade mycotoxins by several
mechanisms may be an advantage De-acetylation might be another mechanism of microbial
mycotoxin detoxification since toxicity of trichothecenes also relies on the acetyl functional
groups (Desjardins 2006) Strain ADS47 showed efficient de-epoxidation and de-acetylation
of several type-A and type-B trichothecene mycotoxins that are commonly detected in cereals
in association with DON In conclusion the soil-borne gram-negative bacterium C freundii
100
strain ADS47 is capable of detoxifying trichothecene mycotoxins by both de-epoxidation and
de-acetylation under aerobic and anaerobic conditions neutral pH and a range of
temperatures ADS47 has great potential to detoxify DON contaminated animal feed
Furthermore isolation of novel bacterial de-epoxidation genes might be used to generate
transgenic plants that can detoxify DON in contaminated seed Alternatively contaminated
seed might be treated with manufacture de-epoxidation enzyme(s) These applications can
improve the quality and safety of agricultural commodities that are prone to trichothecene
mycotoxin contamination
36 Acknowledgements
Thank to the Natural Sciences and Engineering Research Council of Canada (NSERC) for a
scholarship to Rafiqul Islam The work was funded by the Agriculture and Agri-Food Canada
Binational Agricultural Research and Development Fund (AAFCBARD) project We
appreciate Honghui Zhu for technical assistance
101
Supplemental Figure S3-1 Flow chart depicts the procedures of enrichment and isolation of
DON de-epoxidation bacterial strain ADS47 Steps 1 to 4 describe the optimization of
microbial DON to dE-DON transformation activity and removal of undesired microbial
species Step 5 to 7 shows the culture enrichment by incubation with high concentration DON
and isolation of the single strain ADS47
Step 1 Screening of soil samples
Step 2 Optimization of cultural conditions
Step 3 Elimination of undesired microbial species
Step 4 Removal of untargeted microbial species
Step 5 High concentration DON enrichment
Step 6 Separation of bacterial cells and dilution plating
Step 7 Single colony analysis
Active culture was sequentially treated with different mode of actions antibiotics heating cultures at 50degC and subculturing on agar media
Colonies were tested for their DON de-epoxidation ability Purified the single colony via several times subculturing on agar medium
Optimized the culture conditions ie growth substrates temperatures pH and oxygen requirements
Obtained DON de-epoxidation capable microbial culture obtained by analyzing soil samples collected from cereal fields
Serially diluted active culture was grown on agar media Collected washed colonies having DON de-epoxidation activity
The partially enriched culture that retained DON de-epoxidation activity was incubated with 200 microgmL DON in MSB media with continuous shaking at 220 microgmL for 7 d The procedure was repeated 12 times
The enriched culture was serially diluted and treated dilutes with 05 Tween 20 After vigorously vortex for ten min aliquot cultures were spread on different agar media plates to produce single colonies
102
CHAPTER 4
40 CONSTRUCTION AND FUNCTIONAL SCREENING OF GENOMIC DNA
LIBRARY FOR ISOLATION OF DON DE-EPOXYDIZING GENE(S) FROM
CITROBACTER FREUNDII STRAIN ADS47
41 Abstract
Enzymatic activity-based screening of a genomic DNA library is a robust strategy for
isolating novel microbial genes A genomic DNA library of a DON detoxifying bacterium
(Citrobacter freundii strain ADS47) was constructed using a copy control Fosmid vector
(pCC1FOSTM
) and E coli EPI300-T1R as host strain A total of 1250 E coli clones containing
large DNA inserts (~40 kb) were generated which resulted in about 10-fold theoretical
coverage of the ADS47 genome The efficiency of E coli transformation with the
recombinant Fosmid vector was examined by restriction digestion of Fosmids extracted from
15 randomly selected library clones The fragmentation patterns showed that all of the clones
contained an 81 kb Fosmid backbone and ~40 kb of insert DNA except one that had a small
insert This suggests that a DNA library was prepared with expected DNA inserts The de-
epoxidation ability of the library was analysed by incubating clones in Luria Bertani (LB)
broth medium supplemented with 20 microgmL DON with induction (by addition of L-arabinose)
or without induction None of the 1000 clones showed DON de-epoxidation activity The
potential DON de-epoxidation gene(s) of ADS47 may not have been expressed in E coli or
might have been expressed but lacked additional factors required for DON de-epoxidation
103
Keywords DNA library fosmid vector functional library screening DON de-epoxidation
activity
42 Introduction
Microorganisms contain numerous valuable genes and biocatalysts which may have
significant roles in agricultural productivity (Jube and Borthakur 2007) Various techniques
have been used to isolate novel genes from different organisms including transposon
mutagenesis (Frey et al 1998 Tsuge et al 2001) gene knockout (Winzeler et al 1999)
whole genome sequencing (Krause et al 2006) comparative genomics (Koonin et al 2001)
and suppression subtractive hybridization (Dai et al 2010)
The genomic DNA library preparation and activity-based screening of the library is a
relatively new technique which has been efficiently used for isolation of microbial novel
enzymes and compounds such as lipases proteases oxidoreductases antibiotic resistance
genes membrane proteins and antibiotics (Henne et al 2000 Gupta et al 2002 Knietsch et
al 2003 Gabor 2004 Suenaga et al 2007 van Elsas et al 2008) The strength of the
technique is that genes from microbial sources could be obtained without prior sequence
information and not requiring culturing the organisms under laboratory conditions Using this
technique an unknown gene or a set of genes could be obtained by determining particular
phenotypes of the library clones Eventually the gene of interest from the positive clones is
isolated by using a forward (eg mutation) or reverse genetics method (eg subcloning the
positive clones into a microbial expression vector) followed by functional analysis of the
subclones
104
Although this strategy has been used effectively for isolating novel microbial genes
several factors need to be considered in order for the method to be successful These factors
include DNA quality vector type (eg plasmid cosmid fosmid bacterial artificial
chromosome etc) vector copy number and host strains (Daniel 2005 Singh et al 2009) The
Fosmid vector-based technique has been used successfully to clone bacterial genes including
xenobiotics degradation genes antifungal gene clusters and antibiotic resistance genes from
soil metagenomes (Hall 2004 Suenaga et al 2007 Chung et al 2008) This study reports the
construction and activity-based screening of a Fosmid DNA library in an E coli host with the
aim of isolating a novel DON de-epoxidation gene from Citrobacter freundii strain ADS47
(Chapter 3)
43 Materials and methods
431 Genomic DNA library construction
The genomic DNA library was prepared using a Copy Control Fosmid Library
Production Kit (Epicentre Biotechnologies Madison Wisconsin USA) following the
manufacturerrsquos instructions as depicted in Figure 4-1
105
Figure 4-1 Schematic overview of the construction and functional screening of a Fosmid
DNA library
106
Citrobacter freundii strain ADS47 was grown in NB medium at 28degC with continuous
shaking at 220 rpm The high molecular weight bacterial genomic DNA was extracted from
the overnight culture using the Gentra Puregene YeastBacteria Kit (Qiagen Inc Mississauga
Ontario Canada) The purity of the DNA was confirmed by determining OD260280 at ~190 by
a NanoDrop spectrophotometer A 5 microL sample having ~25 microg high molecular weight DNA
was sheared by passing through a 200-microL pipette tip and achieved a sufficient amount of
DNA about 40 kb in size Blunt ended 5rsquo-phosphorylated insert DNA was generated by
thoroughly mixing it with End-Repair Enzyme reagents on ice The 20 microL mixture was
incubated for 45 min at room temperature followed by incubation at 70degC for 10 min to
inactivate the End-Repair Enzyme activity
The treated DNA was size fractionated by a Low Melting Point-Pulsed Field Gel
Electrophoresis (LMP-PFGE) on the clamped homogeneous electric fields (CHEF-DR III)
system (Bio-RAD Laboratories) using 1 UltraPure Low Melting Point Agarose (Invitrogen)
The PFGE was conditioned according to manufacturerrsquos instructions as follows initial switch
time-42 s final switch time-761 s run time 16 h 50 min angle-120deg gradient- 6 Vcm
temperature-14degC and running buffer-05 TBE DNA size markers for the 42 kb Fosmid
control DNA (Epicentre Biotechnologies) and low range Pulse Field Gel (PFG) markers (New
England Biolabs Inc) were loaded adjacent to the DNA sample slots After electrophoresis a
portion of the gel that contained DNA size markers was cut off The gel was stained in SYBR
Safe DNA gel stain (Invitrogen) for 30 min visualized and marked on the 23 42 and 485 kb
marker bands A slice of PFGE gel containing 40 plusmn 10 kb size DNA was cut off with a sterile
razor blade by comparing the marker sizes DNA from the gel was recovered by digesting it
with GELase enzyme ( 1 U100 microL melted agarose) in 1X GELase Buffer followed by a
107
sodium acetate-ethanol precipitation The DNA with the desired size was stored at 4degC for
next dayrsquos ligation to a cloning-ready Fosmid vector (pCC1FOSTM
)
Ligation was performed in a 10 microL reaction mixture by thoroughly mixing 10X Fast-
Link Ligation Buffer 10mM ATP 025 microg of ~40 kb insert DNA 05 microg Fosmid Copy
Control Vector (pCC1FOS) (Figure 4-2) and 2 U Fast-Link DNA lipase The mixture was
incubated for 4 h at room temperature preceded by inactivating the ligation reaction by
heating at 70degC for 15 min The recombinant Fosmid vector was stored at -20degC for later use
Ten microL of the ligated DNA was thawed on ice and transferred to a pre-thawed
MaxPlax Lambda Packaging Extract tube containing 25 microL extract By pipetting several
times the mixture was incubated at 30degC for lamda packaging After 2 h of packaging an
additional 25 microL packaging extract was added into the mixture and incubated for a further 2 h
at 30degC The packaged phage particles were volumed to 05 mL by adding ~400 microL Phage
Dilution Buffer and 25 microL chloramphenicol
Transformation was performed via transduction by mixing the packaged particles with
the overnight cultured E coli EPI300-T1R cells (1 10 vv ratio) in LB supplemented with 10
mM MgSO4 and 02 Maltose Transformation was completed by incubation at 37degC for 1 h
The transformed E coli cells were spread on an LB agar plate containing 125 microgmL
chloramphenicol The plates were incubated overnight at 37degC Colonies that appeared on the
plates were individually transferred to 96-well microtiter plates having 200 microL LB medium
supplemented with 10 glycerol and 125 microgmL chloramphenicol The microtiter plates
were incubated at 37degC for 24 h with continuous shaking at 180 rpm The prepared library
clones were stored at -80degC for further analysis and functional screening
108
109
Figure 4-2 Genetic map of copy controlled Fosmid vector (pCC1FOSTM
) The 813 kb
Fosmid vector backbone shows a multiple cloning site (MCS) chloramphenicol resistance
selectable marker (CmR) replication initiation fragment (redF) inducible high copy number
origin (oriV) replication initiation control (repE) and F-factor-based partitioning genes (ParA
ParB ParC) The vector contains a cos site for lamda packaging (not shown) and lacZ gene
for bluewhite colony selection The vector map was provided by the Epicentre
Biotechnologies (httpwwwepibiocomitemaspid=385)
110
432 Confirmation of library construction
The quality of the DNA library prepared was determined by examining 15 E coli
clones randomly selected A 15 mL LB broth supplemented with 125 microgmL
chloramphenicol was inoculated by each of the transformed clones To obtain higher Fosmid
DNA yield 1 microL autoinduction solution (L-arabinose) to increase the copy number of the
Fosmid vector (Epicentre) was added to the cultures After an overnight incubation of the
cultures at 37degC with continuous shaking at 220 rpm Fosmid DNA was extracted using a
Qiagen Large-Construct Kit followed by restriction digestion with Not I enzyme A 20 microL
digestion mixture containing 05 microg Fosmid DNA 1X digestion buffer and 05 U Not I was
incubated at 37degC for 15 h The partially digested DNA and a 42 kb DNA size marker
(Fosmid control DNA) were ran on 08 agrose gel for 15 h at 100 V The linear Fosmid
vector backbone (813 kb) and insert DNA fragments were visualized on the gel and the
image was recorded
433 Functional screening of the clones
Eppendorf tubes (15 mL) containing 500 microL LB supplemented with 125 microgmL
chloramphenicol were inoculated with a single clone (400 clones tested) or a combination of
12 library clones (600 tested) from microtiter plates stored at -80degC One half of the cultures
in each group (single or combined clones) were induced to increase the Fosmid copy number
by adding a 1 microL (500X) autoinduction solution (contained L-arabinose) to the culture tubes
L-arabinose induces the expression of a mutant trfA gene of the E coli host strain which
leads to the initiation of Fosmid replication from a low-copy to a high-copy number origin of
replication (oriV) The cultures were spiked with 20 microgmL DON and incubated at 37degC for
111
72 h with continuous shaking at 180 rpm The DON to de-epoxy DON transformation ability
of the clone cultures was examined by a liquid chromatography-ultraviolet-mass spectrometry
(LC-UV-MS) method as described by Islam et al (2012)
112
44 Results
441 Production of the DNA library
High molecular weight intact (non-degraded) and good quality (OD600280 ~190)
genomic DNA was obtained from the DON de-epoxidizing bacterial strain ADS47 (data not
shown) Pipetting of the DNA extract resulted in an adequate amount of insert DNA (gt025
microg) of approximately 40 kb size as showed on a LMP-PFGE (Figure 4-3)
A DNA library in E coli host was constructed using LMP-PFGE purified DNA The
library was comprised of 1250 E coli transformants which were selected on LB agar plates
supplemented with chloramphenicol (Figure 4-4)
113
Figure 4-3 Preparation of a 40 kb size insert DNA from a DON de-epoxidizing bacterium
Genomic DNA was extracted from the overnight culture of a DON de-epoxidizing
Citrobacter freundii strain ADS47 After fragmentation the DNA was size fractionated by
Low Melting Point- Pulse Field Gel Electrophoresis (LMP-PFGE) with markers for the PFGE
(M1) and 42 kb Fosmid control DNA (M2) S1 S2 S3= different DNA samples extracted
from bacterial strain ADS47 The red dots area indicates the section of the gel containing
genomic DNA of the expected size (~40 kb)
114
Figure 4-4 Production of the genomic DNA library in E coli EPI300-T1 host strain
Approximately 40 kb long blunt ended genomic DNA from a DON de-epoxidizing bacterial
strain ADS47 was ligated to a Copy Control Fosmid vector pCC1FOS The construct was
packaged into lamda bacteriophage followed by a transduction-mediated transformation of E
coli host cells The clones which appeared on the LB agar plate supplemented with 125
microgmL chloramphenicol are expected to be transformed by the recombinant Fosmid vector
containing insert DNA
115
442 Conformation of the library construction
All of the clones tested acquired the Fosmid vector Out of 15 clones examined 14
received the recombinant Fosmid vector that contain the expected size insert DNA as
determined by an 81 kb vector backbone and at least one high molecular weight insert DNA
band (~40 kb) on agarose gel (Figure 4-5) In contrast one clone (lane 4) showed an 81 kb
vector backbone and a low molecular weight band which is also a fragment of the Fosmid
vector The result indicated that approximately 94 of the library clones were transformed
with the recombinant vector
443 Activity-based screening of the library
Out of 500 single copy number clones (200 individual and 300 combined clones)
none showed DON to de-epoxy DON transformation activity (Table 4-1) An additional 500
high copy number clones (200 single and 300 combined clones) were unable to de-epoxidize
DON as well
116
Figure 4-5 Confirmation of the Fosmid DNA library production in E coli Fosmid DNA
collected from 15 clones was digested with Not I and run on 08 agarose gel at 100 V for 15
h The presence of the vector backbone (81 kb) and the insert DNA fragment (~40 kb) in the
clones was determined by comparing a 42 kb molecular size marker (M) Lanes (1-15) denote
the recombinant Fosmid DNA extracted from 15 E coli clones
117
Table 4-1 Analysis of Fosmid DNA library clones for deoxynivalenol (DON) de-epoxidation
capability
Type of clones Analysis of
singlecombined
clones
Number of
clones
examined
Number of clones
showed DON de-
epoxidation activity
Without induction
(single copy number clones)
Singly 200 0
Combination of 12 300 0
With induction (multi-copy
number clones)
Singly 200 0
Combination of 12 300 0
De-epoxidation activity of the clones was examined alone or in combination of 12 clones
without or with induction by adding L-arabinose The clones grown in LB supplemented with
125 microgmL and 20 microgmL DON were analysed for DON to de-epoxy DON transformation
ability by a liquid chromatography-ultra violet- mass spectrum (LC-UV-MS)
118
45 Discussion
While a Fosmid DNA library with the expected genome coverage of a DON de-
epoxidizing bacterial genome was produced the function-derived screening of the library
clones did not identify any clones that showed DON de-epoxidation enzymatic activity This
is unlikely due to lack of coverage of the bacterial genome as 1000 clones analyzed in the
study should have provided eight fold coverage of the genome Therefore it was likely due to
a lack of expression of the enzyme(s) for DON de-epoxidizing activity under the conditions
used
Thus far there is only one report of heterologous expression of bacterial epoxide
reductase gene in another bacterium A high level of heterologous expression of a
Synechococcus sp vitamin K epoxide reductase gene was achieved in E coli with codon
optimization and placing the gene under the control of a prokaryotic T7 promoter in an
expression vector (Li et al 2010) Therefore heterologous expression of bacterial epoxide
reducatse gene using its own promoter has not yet been demonstrated The success of the
method used in this chapter relies on an intact copy of the gene or operon being cloned the
selection of an appropriate cloning vector a host organism that can recognize the cloned C
fruendii promoter and a suitable library screening method for DON degradation by E coli
(Mocali and Benedetti 2010)
The unbiased cloning efficiency of the Fosmid vector its ability to contain large insert
DNA high stability in E coli as single copy number and autoinduction to increase copy
number in host cells made it appear to be a suitable method for the present work (Kim et al
1992 Lee et al 2004) Library production with large insertions is a major advantage as gene
cluster involved in particular pathway could be trapped into single clone and their functions
119
could be analysed The pathway of microbial DON de-epoxidation activity and the genes
encoding the activity are not known Therefore it was reasonable to prepare a library with
large inserts to ensure the possibility of cloning single as well as whole pathway genes of
interest for functional screening A further advantage of the method is that Fosmid clones are
inducible which allows detecting poorly expressed genes in a heterologous host by inducing
high copy numbers (Daniel 2005)
One of the limitations of the method is that the functions of the target genes rely
mostly on the host organismrsquos transcription and translation machineries Therefore selection
of the host organism is critical for activity-based screening of library clones in heterologous
hosts A number of host strains including E coli Streptomyces lividans Rhizobium
leguminosarum and Pseudomonas aeruginosa have been used for functional screening
activities from diverse organisms (Daniel 2005) The present work attempted to isolate C
freundii strain ADS47 genes encoding DON de-epoxidation enzymes Both strain ADS47 and
E coli belong to gamma proteobacteria which suggest that E coli would be an appropriate
host organism for heterologous expression of ADS47 genes Poor expression of heterologous
genes could be another limitation of the method which may be overcome by overexpression
to increase the catalytic activity of less expressed genes However clones with DON de-
epoxidation activity were not identified in this study even the Fosmid copy number was
increased by induction
In general a large number of clones must be analysed to obtain the gene of interest
using an activity-based screening technique (Lee et al 2004 van Elsas et al 2008) In this
study the constructed DNA library comprised of 1160 clones having approximately ~40 kb
insert DNA suggesting that the prepared library achieved ~10 fold coverage of the ADS47
120
genome (48 Mb) which ensured gt99 probability of the presence of all DNA sequence in
the library In the produced library three clones (lanes 3 10 and 12) showed one intact band
greater than 42 kb in size on the agarose gel It is presumed that the Fosmid DNA of the three
samples was not digested because of insufficient amount of restriction enzyme added to the
digestion mixture About half of the samples demonstrated multiple bands suggesting that the
insert DNA might have contained multiple Not I restriction sites
However not every heterologous gene may necessarily express or function in the E
coli host (Gabor et al 2004) The functionality of the gene requires expression of the genes
and proper folding of the protein which may be influenced by environmental conditions (eg
substrate temperature and pH) and genetic factors In this study library clones were screened
in growth conditions at which strain ADS47 performed the best de-epoxidation activity The
expression of genes may be controlled by the promoter suitable ribosome binding site and
transndashacting factors (eg special transcription factors inducers chaperones cofactors proper
modifying enzymes or an appropriate secretion system) (Gabor et al 2004) Those factors
have to be supplied by the cloned DNA fragments or host cells to allow any heterologous
gene to be functional Since DON de-epoxidation genes of strain ADS47 evolved in a distinct
environmental condition the expression and activity of the gene(s) might have required
specific factorsregulators that were absent in E coli Even though the Fosmid based library
screening technique facilitated isolation of many microbial genes clones having de-
epoxidation activity were not identified using such technique in this study
121
46 Acknowledgements
We appreciate the Agriculture and Agri-Food CanadaBinational Agricultural
Research and Development Fund (AAFCBARD) project for providing the financial support
for this work The authors thank Honghui Zhu for analyzing numerous samples by liquid
chromatography ultraviolet mass spectrum method
122
CHAPTER 5
50 GENOME SEQUENCING CHARACTERIZATION AND COMPARATIVE
ANALYSIS OF CITROBACTER FREUNDII STRAIN ADS47
51 Abstract
The genome of a trichothecene mycotoxin detoxifying C freundii strain ADS47
originating from soil was sequenced and analyzed Strain ADS47 contains a circular
chromosome of 4878242 bp that encodes 4610 predicted protein coding sequences (CDSs)
with an average gene size of 893 bp Rapid Annotation Subsystems Technology (RAST)
analysis assigned functions to 2821 (62) of the CDSs while 1781 (38) CDSs were
assigned as uncharacterized or hypothetical proteins Comparative bioinformatics analyses
were done for strain ADS47 C freundii Ballerup 785139 Citrobacter sp 30_2 C youngae
ATCC 29220 C koseri ATCC BAA-895 and C rodentium ICC168 There were a significant
number of common orthologous genes (2821) were found in the six Citrobacter genomes
implying a common evolutionary relationship ADS47 is most closely related to the
opportunistic human pathogen C freundii Ballerup The human and mouse pathogenic
species C rodentium and C koseri are the most distantly related to ADS47 and non-
humananimal pathogens C youngae and Citrobacter sp 30_2 were closely related ADS47
C koseri C youngae and Citrobacter sp 30_2 lacked the clinically important locus for
enterocyte effacement (LEE)-encoded type-III secretion systems which are present in C
rodentium and C freundii Ballerup Strain ADS47 had 270 unique CDSs many of which are
located in the large unique genomic regions potentially indicating horizontal gene transfer
123
Among the unique CDSs nine were reductasesoxidoreductases and a de-acetylase that may
be involved in trichothecene mycotoxin de-epoxidation and de-acetylation reactions
Keywords Citrobacter freundii comparative genomics genome sequencing unique genes
reductases
52 Introduction
According to the Food and Agriculture Organization (FAO) at least 25 of the
worldrsquos food crops are contaminated with mycotoxins causing serious negative impacts on
livestock and international trade (Boutrif and Canet 1998 Desjardins 2006 Binder et al
2007) Trichothecenes (TCs) produced by a variety of toxigenic Fusarium species are
probably the most prevalent food and feed contaminating mycotoxins It is estimated that
infections by the fungus caused more than $1 billion in crop yield losses in USA and Canada
alone (Windels 2000 Schaafsma et al 2002 Yazar and Omurtag 2008) TC mycotoxins can
have adverse affects on human health including organ toxicity mutagenicity carcinogenicity
neurotoxicity modulation of the immune system negative reproductive system affects and
growth retardation (DrsquoMello et al 1999 Pestka et al 2007) A strain of Citrobacter freundii
ADS47 was isolated from soil in southern Ontario that is capable of detoxifying (ie de-
epoxidation and de-acetylation) several TC mycotoxins including deoxynivalenol nivalenol
and T2-toxin (Chapters 2-3)
Citrobacter freundii is a gram negative motile rod shaped facultatively anaerobic
bacterium belonging to the family of Enterobacteriaceae (Wang et al 2000 Whalen et al
2007) The opportunistic human pathogen is responsible for a number of infections including
124
nosocomial infections of respiratory tract urinay tract blood and causes neonatal meningitis
(Badger et al 1999 Wang et al 2000) C freundii exists almost ubiquitously in soil water
wastewater feces food and the human intestine Biodegradations of tannery effluents copper
aromatic amino acids by C freundii strains isolated from water mine tailings and soil
respectively have been previously reported (Kumar et al 1999 Savelieva et al 2004 Sharma
and Fulekar 2009) Because of their biochemical capabilities bacteria have played important
roles in industrial biotechnology including de-epoxidation and de-acetylation activities
(Demain and Adrio 2008 Plank et al 2009 Zhao et al 2010) However the identification of
genes for de-epoxidation activity against trichothecene mycotoxins is limited (Boutigny et al
2008)
The advent of molecular techniques such as next generation sequencing technologies
and the related bioinformatics tools has enabled the discovery of novel genes and the
enzymes they encode in pathogenic and beneficial bacteria (van Lanen and Shen 2006
Ansorge 2009 Zhao 2011) In the present study the genome of mycotoxin-detoxifying strain
C freundii ADS47 was sequenced and subjected to comparative genomic analyses with other
Citrobacter species to identify putative mycotoxin-detoxifying genes
125
53 Materials and methods
531 Sequencing quality control and assemble of ADS47 genome
Genomic DNA of the strain ADS47 was extracted using Gentra Puregene
YeastBacteria Kit (Qiagen Inc Mississauga Ontario Canada) and purity of the DNA was
confirmed by determining OD 260280 at ~190 using a Nanodrop spectrophotometer
The genomic DNA was sent to be sequenced by the Illumina HiSeq 2000 sequencing
analyzer at Beijing Genomics Institute (BGI) Hong Kong
(httpwwwbgisequencecomhomeservicessequencing-servicesde-novo-sequencing) The
sequencing workflow involved short read library preparation cluster generation sequencing
and data analysis Briefly 3-5 ug of bacterial gDNA was fragmented to produce a 500 bp
insert library by preparative electrophoresis through a 08 agarose gel The fragmented
DNAs were blunt-ended by treating with a mixture of enzymes (T4 DNA polymerase klenow
fragment) and subsequently adapters were ligated to the blunt ends Each of the DNA
fragments was amplified to a cluster which was performed in the flow cell by template
hybridization and bridge amplification Millions of clusters were then sequenced by
sequencing-by-synthesis technology leading to base calls The raw data was filtered to remove
low abundant sequences from the assembly according to 15-mer frequencies The low
complexity reads low quality (leQ20) base calls and adapter contaminations (24 bp overlap
between adapter and reads) were eliminated The quality of the sequences was analysed by
GC content and depth correlative analysis (for GC biasness) K-mer analysis was performed
using 15 bp of the short reads for heterologous sequence identification
126
The cleaned short reads (approximately 90 bp) were self-assembled using the Short
Oligonucleotide Analysis Package (SOAP) denovo (httpsoapgenomicsorgcn) (Li et al
2008) The assembly errors were corrected by filling gaps and proofreading single base pairs
using mapping information The assembly produced 441 contigs with an N50 contig size of
29222 bp (the largest contig size 114839 bp) and 80 scaffolds with an N50 scaffold size of
325726 bp (largest scaffold 662452 bp) with an total sequence length of 4878242 bp which
constitutes 33-fold genome coverage The scaffolds were ordered and oriented according to a
reference genome C koseri ATCC BAA-895 the closest completely sequenced species of
the newly sequenced bacterial genome by alignment with the MUMmer program v322
(httpmummersourceforgenet) (Delcher et al 2002) Genome coverage was determined by
mapping the reads to the assembled genome with Bowtie software (Langmead et al 2009) A
preliminary genome annotation was performed to define the origin of replication by
identifying colocalization of multiple genes ie gyrB recF dnaN and dnaA (usually found
near the origin of replication) and comparison with the C koseri reference genome Synteny
analysis between the reference bacterium and assembled scaffolds was performed with the
MUMmer program
532 Gene function annotation and classification
The genome of ADS47 was annotated by Rapid Annotation using Subsystems
Technology v40 (RAST) a server-based genome analysis system (httprastnmpdrorg)
(Aziz et al 2008) A circular genome map was constructed using CGView program
(httpstothardafnsualbertacacgview_server) (Stothard and Wishart 2005)
127
533 Molecular phylogenetic analysis
A robust phylogenetic tree was produced using Composition Vector Tree (CVTree)
web-based program v20 (httpcvtreecbipkueducn) (Qi and Hao 2004) by comparing
whole genome amino acid sequences of strain ADS47 two completely sequenced Citrobacter
genomes [C rodentium ICC168 (NC_013716) and C koseri ATCC BAA-8959 NC_009792)]
and three partially sequenced Citrobacter genomes [C freundii strain Ballerup 785139
(CACD00000000) Citrobacter sp 30-2 (NZ_ACDJ00000000) and C youngae ATCC 29220
(NZ_ABWL00000000)] E coli O157H7 strain EDL933 (NC_002655) was used as an
outgroup species The tree was constructed from the generated pairwise distance matrix data
using Neighbor-Joining Program in PHYLIP v369 package (Saitou and Nei 1987 Felsenstein
1989)
534 Comparative genomic analysis
Genome-wide comparative analyses between C freundii ADS47 C freundii strain
Ballerup 785139 Citrobacter sp 30-2 C youngae ATCC 29220 C Koseri ATCC BAA-895
and C rodentium ICC168 was carried out The nucleotide sequence idendities were
determined by pairwise alignments of the ADS47 genome with the five other Citrobacter
genomes using MUMmer software (Delcher et al 2002) and GC content total CDSs genome
coverage by CDSs and number of stable RNAs were determined by RAST program
Orthologous CDS groups among the total set of 29079 CDSs derived from the six
Citrobacter genomes were determined using OrthoMCL software v202
(httporthomclsourceforgenet) based on all-against-all reciprocal BLASTP alignment of
amino acid sequences (Li et al 2003) Genes unique to each strain were identified as those
128
CDSs not included in any orthologous group of the five other Citrobacter genomes BLASTP
comparisons were visualized with Blast Ring Image Generator (BRIG) software v095
(httpsourceforgenetprojectsbrig) (Alikhan et al 2011)
Protein sequences of the five Citrobacter genomes pooled from the GenBank data
bases were compared with ADS47 A circular map was developed by BRIG software that
showed the unique genomic regionsgenes of ADS47 The unique genes were grouped
according to their functional similarities previously identified by RAST software and
genomic regions possibly acquired by horizontal transfer were identified using RAST
software
129
54 Results
541 Assembly and organization of the ADS47 bacterial genome
Scaffolds derived from Citrobacter freundii strain ADS47 were ordered and oriented
by alignment to Citrobacter koseri ATCC BAA-895 the most closely related genome that has
been completely sequenced The genomes of ADS47 and C koseri displayed close synteny
(Figure 5-1)
Figure 5-1 Co-linearity between C freundii strain ADS47 genome and C koseri Scaffolds
derived from ADS47 genome sequences (Y- axis) were aligned and assembled by comparison
with the closest completely sequenced genome C koseri ATCC BAA-875 (X- axis)
130
542 An overview of the ADS47 genome
General features of the C freundii strain ADS47 are presented in Table 5-1 Some
notable features are that strain ADS47 contains 4881122 bp circular chromosome with an
average GC content of 5202 The genome contains 4610 CDSs which comprise 8784 of
the whole genome Annotation by RAST assigned 2821 (62) CDSs to known biological
functions and remaining 1789 (38) genes were classified to unknown or hypothetical
proteins
The GC content in the coding regions is significantly higher (5318) than in the non-
coding intergenic region (4379) which covers 1215 (592781 bp) of the genome The
intergenic region is comprised of 018 repeat sequences (total 8999 bp) 5 (009)
transposons (4806 bp) 025 stable RNA (12091 bp) and the remaining ~115 is
classified as regulatory and other functions The ADS47 genome contains 62 tRNAs 3
rRNAs and 34 copies of sRNAs The average lengths of the tRNAs rRNAs and sRNAs are
785 885 and 13441 bp respectively There are also 42 phageprophage-related genes
identified in the genome
Figure 5-2 shows the distribution of the 26 categorized CDSs rRNA tRNA GC
content and GC skew It appears that ADS47 genome contains equal quantity of genes in
leading and lagging strands and showed occasional clustering (likely operons) of genes in the
two strands The genome contains three rRNAs (two large subunit and a small subunit) and 62
tRNAs which are distributed across the genome A consistent GC content was observed in
the ADS47 genome however 7-10 low GC content (high black color peaks) genomic regions
were found at locations origin of replication 600 2000 3000 3300 3550 3850 4300 and
4800 kbp This is indicative of possible horizontally transferred genes or insertions in those
131
genomic locations GC skew denotes the excess of C over G in certain genomic locations
Positive GC skew (+) (green color) indicates the skew value in the leading strand while
negative skew (-) (purple color) in the lagging strand As in many bacterial genomes the
ADS47 genome map shows the separation of positive and negative GC skew at the origin of
replication (base pair position 1) The GC skew in the leading strand is highly skewed (high
green color peak) indicating that possible horizontal gene transfer occurred into the ADS47
genome
The CDSs ranged from ~100 bp to ~2000 bp and have an average size of 893 bp
(Figure 5-3) The highest numbers of CDS (770) were 600-800 bp long A considerable
number of genes (220) were gt2000 bp in size Some of the biological functions for the
smallest genes (100-200 bp) are sulfur carrier protein methionine sulfoxide reductase
ribonuclease P protein component while the largest genes (gt2000 bp) are ferrous iron
transport protein B translation elongation factor G pyruvate flavodoxin oxidoreductase and
tetrathionate reductase
132
Table 5-1 General features of the C freundii strain ADS47 genome
Feature Chromosome
Genome size 4881122 bp
Overall GC content 5202
Number of genes 4610
Genes with assigned function (number and ) 2821 (62 )
Genes without assigned function (number and ) 1789 (38 )
Total length of protein coding sequences 4285461 bp
Genome coverage by coding sequences () 8784
Average length of coding sequences 893 bp
GC content in the coding region 5318
GC content in the non-coding region 4379
Length and percentage of intergenic region 592781 bp and 1215
Total repeat sequences and genome coverage 8999 bp and 018
Total transposons sequences and genome coverage 4806 and 009
Number average length and genome coverage of tRNA 62 copies 785 bp and 0099
Number average length and genome coverage of rRNA 3 copies 885 bp and 0067
Number average length and genome coverage of sRNA 34 copies 13441 bp and 0093
133
134
Figure 5-2 Circular genome map of C freundii strain ADS47 Map was created with the
CGView software From outside to inside the first circle indicates the genomic position in kb
Different functional categories of predicted coding sequences (CDS) are distinguished by
colours with clockwise transcribed CDSs in the second circle and anticlockwise transcribed
CDSs in the third circle Bars on the fourth and fifth circles indicate the rRNAs and tRNAs
respectively Sixth and seventh circles show the coverage and GC content respectively The
innermost circle demonstrates the GC skew + (green) and GC skew- (purple)
135
Figure 5-3 Size distribution of the predicted genes identified in C freundii strain ADS47
The X axis identifies the gene size categories (bp) and Y axis the abundance of genes
136
543 Annotation and functional classification of ADS47 genome
RAST analysis identified predicted CDSs in the genome of Citrobacter strain ADS47
assigned functions and grouped the annotated genes into 26 functional subsystem categories
out of 27 No CDS was found belonging to the photosynthesis category A graphical
presentation of the distribution of the CDSs into each of the 26 categories is given in
Figure 5-4
A large portion (38) of the CDSs belong to hypothetical or unknown functions
Among the 62 known or predicted proteins the highest numbers (835) belong to the
carbohydrate category followed by amino acids and derivatives (468) and
cofactorsvitaminsprosthetic groups (324) A noticeable number of stress response (236) and
respiration (198) categories genes were identified in the bacterial genome The ADS47
genome encoded by 171 membrane transport genes 113 virulencedefense genes and 126
genes involved in regulationcell signaling categories Two hundred and one CDSs were
grouped into the miscellaneous category whose functions are different than the indicated
RAST categories The RAST annotations further divided the categorized CDSs into
subcategories based on similar functional roles which is discussed in section 545
137
Figure 5-4 Functional classification of the predicted coding sequences (CDS) of C freundii
strain ADS47 The annotation was performed by Rapid Annotation using Subsystems
Technology (RAST) program
138
544 Molecular phylogenetic relationships
Genome-wide phylogenetic analysis based on comparisons of whole genome amino
acid sequences of two completely sequenced Citrobacter genomes (C koseri and C
rodentium) and three partially sequenced Citrobacter genome sequences (C freundii
Citrobacter sp and C youngae) illustrates the relationships of ADS47 to five Citrobacter
species (Figure 5-5)
C youngae ATCC 29220 originating from human feces and two pathogenic species
(C koseri ATCC BAA-895 and C rodentium strain ICC168) are progressively more distantly
related to ADS47 The present analysis is consistent with the 16S-rRNA sequence based
phylogenetic analysis (Chapter 3) which showed that the closest bacterial strain to ADS47 is
C freundii (Figure 3-2) This confirmed the identification of strain ADS47 as the gamma
proteobacterial species C freundii made in Chapter 3
139
Figure 5-5 Phylogeny of the trichothecene detoxifying C freundii strain ADS47 The
dendrogram was constructed by comparing the ADS47 genome (amino acid sequences) with
five Citrobacter genomes and a E coli strain (out group species) using the CVTree software
The calculated pairwise distances matrix by Protdist program was transformed into a tree by
neighbor-joining program The dissimilarity tree was drawn to scale with branch lengths
(scale bar 002 indicated the amino acid substitutions among the genomes) The C koseri C
rodentium C freundii Citrobacter sp C youngae and E coli protein sequences were
uploaded from GenBank (Benson et al 2009)
140
545 Comparative genomics
Genome-wide comparative analyses between the genome of strain ADS47 and the five
previously sequenced Citrobacter genomes showed that 872 of the total nucleotides
between ADS47 and C freundii Ballerup were identical The values for the comparisons
between C freundii strain ADS47 were 8212 for Citrobacter sp 30-2 7637 for C
youngae 5981 for C koseri and 4213 for C rodentium (Table 5-2)
The two human and mouse pathogenic species namely C rodentium and C koseri
and the bacteria originally isolated from human feces (C youngae) have higher GC contents
(546 and 532 respectively) compared to C freundii strain ADS47 C freundii strain
Ballerup and Citrobacter sp 30_2 which have slightly lower GC contents (~52) The two
pathogenic species contain 22 copies of rRNA which is significantly higher than the 9 found
in C freundii strain Ballerup 6 in Citrobacter sp 30_2 and 3 in C freundii strain ADS47 but
less than the 25 copies in C youngae The genome of C freundii strain ADS47 C freundii
strain Ballerup C youngae and Citrobacter sp 30-2 had a similar percentage of the genome
comprising coding sequences (865-88) C koseri and C rodentium genomes contained the
highest (90) and lowest (84) genome coverage by coding sequences respectively
141
Table 5-2 Genome comparisons between strain ADS47 and five Citrobacter species The
base nucleotide sequence identities were performed using MUMmer DNAdff script program
(Delcher et al 2002) The features (GC content total CDS percentage CDS and number of
ribosomal RNA) were determined using RAST program (Aziz et al 2008)
Genome feature Bacterial species
C
freundii
ADS47
C freundii
Ballerup
Citrobacter
sp 30_2
C
youngae
C koseri C
rodentium
Genome length
(bp)
4878242
4904641
5125150
5154159
4720462
5346659
Base similarity to
ADS47 ()
NA 8715 8212 7637 5981 4213
GC () 5202 5218 516 5265 530 546
Number of coding
sequences
4610 4685 4728 5278 4978 4800
Genome coverage
with coding
sequences ()
880 865 866 872 900 840
rRNA copies 3 9 6 25 22 22
Total stable RNAs 99 81 78 105 120 142
142
The number of orthologs shared between ADS47 and other Citrobacter species is
illustrated in Figure 5-6 ADS47 shares the smallest number of orthologous CDSs with C
rodentium and C koseri 3250 and 3341 respectively The highest number of shared CDS
were found with C freundii Ballerup (3706) followed by C youngae (3693) and Citrobacter
sp 30_2 (3676) A total of 2821 CDSs were found to be common to all of the six Citrobacter
genomes analysed (data not shown) Nonetheless significant diversity exists between C
freundii ADS47 and the other species as a considerable number of unshared genes were
identified from the comparisons with C freundii Ballerup [979 (20)] Citrobacter sp 30_2
[1064 (23)] C youngae [1654 (31)] C koseri [1693 (34)] and C rodentium [1634
(34)]
143
Figure 5-6 Comparisons of coding sequences between strain C freundii ADS47 and five
Citrobacter species Coding sequences of ADS47 were compared with C freundii Ballerup
Citrobacter sp 30_2 C youngae C koseri and C rodentium genomes based on all-versus-all
reciprocal FASTA comparison of at least 30 amino acid identity and gt70 coverage of the
total length using OrthoMCL program A Venn diagram demonstrates the shared and
unshared (indicated in the brackets) coding sequences
144
Table 5-3 shows differences in the 26 RAST categorized genes between the six
Citrobacter genomes Even though a low abundance of virulencediseasedefense category
genes were found in the animalhuman pathogenic bacteria of C rodentium (85) and C koseri
(92) significantly higher mycobacterium virulence genes (13) were identified in this two
species The genomes of the two closely related strains C freundii ADS47 and C freundii
Ballerup lack this type of virulence gene Instead elevated numbers of genes for resistance to
antibiotic and toxic compounds were found in C freundii ADS47 (94) C freundii Ballerup
(88) and Citrobacter sp (110) compared to other three species Copper homeostasis and
cobalt-zinc-cadmium resistance genes were identified in strain C freundii ADS47 (13 and 21
respectively) and closely related C freundii Ballerup (12 and 22) and Citobacter sp (18 and
22) Significantly lower numbers of these genes were present in C rodentium (6 and 4) and C
koseri (7 and 8) C rodentium and C koseri lack mercuric reductase mercuric resistance and
arsenic resistance genes perhaps because being animalhuman pathogens but these genes
were found in the remaining four genomes The C koseri genome is richer in betandashlactamase
genes (5) compared to the remaining genomes which encodes 1-3 genes
145
Table 5-3 Functional categorized gene abundances among the genomes of C freundii
ADS47 C freundii Ballerup Citrobacter sp 30_2 C youngae C koseri and C rodentium
The genomes were classified into different functional categories with the RAST program
which classified the total CDSs
RAST subsystem category Citrobacter species
C
freundii
ADS47
C
freundii
Ballerup
C
sp
C
youngae
C
koseri
C
rodentium
Cofactorsvitaminsprosthetic
groupspigments
323
317
310
311
325
304
Cell wall and capsule 279 272 248 262 247 262
Virulence disease and defense 113 107 134 100 92 85
Potassium metabolism 50 53 33 32 30 28
Miscellaneous 210 228 200 185 245 185
Phageprophagetransposable
element plasmid
24 75 25 4 6 49
Membrane transport 170 156 171 195 171 172
Iron acquisition and metabolism 80 81 86 62 113 27
RNA metabolism 220 222 217 215 238 212
Nucleosides and nucleotides 119 112 110 111 120 114
Protein metabolism 249 280 269 298 287 254
Cell division and cell cycle 30 29 32 29 38 29
Motility and chemotaxis 133 124 90 147 143 133
Regulation and cell signaling 126 130 124 135 132 123
Secondary metabolism 21 22 20 21 0 0
DNA metabolism 145 167 165 155 147 152
Fatty acids lipids and isoprenoids 121 149 122 121 134 133
Nitrogen metabolism 50 52 53 49 56 58
Dormancy and sporulation 3 3 3 4 3 3
146
Table 5-3 (continued)
RAST subsystem category
Citrobacter species
C
freundii
ADS47
C
freundii
Ballerup
C
sp
C
youngae
C
koseri
C
rodentium
Respiration 198 184 180 191 166 162
Stress response 236 235 170 177 179 174
Metabolism of aromatic
compounds
20 20 28 25 17 48
Amino acids and derivatives 482 476 450 466 428 416
Sulfur metabolism 48 50 52 57 56 56
Phosphorus metabolism 50 49 43 42 43 28
Carbohydrates 837 803 708 731 733 737
147
Strains C freundii ASD47 and C freundii Ballerup have much more potassium
metabolism category genes (50 and 53 respectively) than the other bacterial species which
code for 28-33 proteins Larger numbers of genes associated with the glutathione regulated
potassium efflux system were also identified in C freundii ASD47 with 23 and C freundii
Ballerup with 26 compared to the remaining four species that encode only five genes All of
the six genomes contained phageprophage-related genes with the maximum identified in C
freundii Ballerup (75) followed by C rodentium (49) Citrobacter sp (25) and C freundii
ADS47 (24) Lower numbers of phageprophage-related genes were found in C koseri (6) and
C youngae (4)
The six Citrobacter genomes showed remarkable variability in their secretion systems
The type-III secretion operon genes are only found in C rodentium and C freundii Ballerup
Type-II secretion genes are predominant in C rodentium (12) followed by ADS47 (9)
Type-IV secretion system is uncommon in Citrobacter species since it was only
identified in Citrobacter sp where it is represented by 19 genes Variable numbers of type-VI
secretion system genes are present in the five genomes except C koseri which lacks such
genes C youngae has the highest number (43) of type-VI secretion proteins The other strains
have a range of type-VI secretion proteins in particular 27 were identified in C freundii
Ballerup 15 in C freundii ADS47 and 8 in Citrobacter sp Type-VII (chaperoneusher)
secretion system genes were found in all of the six Citrobacter species with the largest (42)
and smallest (13) numbers of these genes occurring in C koseri and C freundii Ballerup
respectively
Although a relatively similar number of ABC transporters (37-42) were identified in
the six genomes C freundii ADS47 contain the largest number of tripartite ATP-independent
148
periplasmic (TRAP) transporter genes (7) By contrast the closely related C freundii
Ballerup lacks such transporter system An identical number of iron acquisition and
metabolism category genes were found in C freundii ADS47 (80) and C freundii Ballerup
(81) while the two pathogenic species C rodentium and C koseri code for the lowest (27)
and highest (113) numbers of such genes C koseri is rich in siderophore related genes (64)
but the remaining five genomes have smaller numbers of siderophore-associated genes (20-
22)
Variable numbers of motility and chemotaxis related genes were found in the
Citrobacter genomes with the largest number occurring in C youngae (147) followed by C
koseri (143) C freundii ADS47 (133) C rodentium (133) C freundii Ballerup (124) and
Citrobacter sp (90) The two pathogenic species (C rodentium and C koseri) encode a
remarkably low number of quorum sensing genes (2) In contrast C youngae codes for the
highest number of quorum sensing genes (19) A common and distinctive feature of the two
pathogenic species is the lack of genes required for secondary metabolism In contrast the
non-pathogenic species contain a common set of genes for degrading a range of
phenylpropanoids and cinnamic acid (20-22)
The lowest number of genes related to phospholipidsfatty acids biosynthesis and
metabolism were found in C freundii ADS47 (66) compared to C freundii Balllerup (94) and
C koseri (98) The smallest numbers of nitratenitrite ammonification genes were found in
ADS47 (31) and its closest genome C freundii Ballerup (32) In contrast the two pathogens
encode the highest number of such proteins (39-40)
C rodentium and C koseri code for significantly low numbers of respiratory category
genes (162-165) while C freundii ADS47 contains the highest number of such genes (198)
149
Further analysis of the subcategory genes determined that the two gut species (C youngae and
Citrobacter sp 30_2) contain the largest numbers of genes encoding anaerobic reductases
(22) and identical numbers of formate dehydrogenases (17) The largest numbers of formate
dehydrogenase genes were identified in C freundii Ballerup (32) and C freundii ADS47 (30)
but the two pathogenic species differed from the other species by encoding the lowest number
(13) of c-type cytochrome biogenesis genes
A higher number of aromatic compound metabolism category genes were found in the
mouse pathogen C rodentium (48) compared to the human pathogen (C koseri) which
contain 17 genes A significantly higher number of genes involved in central aromatic
intermediate metabolism (26) were found in C rodentium compared to the mouse pathogen
(C koseri) and other genomes which only contain 6-8 such genes The two C freundii strains
ADS47 and Ballerup lacked the anaerobic degradation of aromatic compound
(hydroxyaromatic decarboxylase family) genes
In spite of the abundance of stress response category genes strain ADS47 has only a
small number of desiccation stress genes (5) compared to the other species which ranged
from 9 to10 However ADS47 and Ballerup code for higher number of genes related to
oxidative stress (94 and 91 respectively) and cold shock (8 and 7 respectively) than the other
species
The smallest numbers of amino acids and its derivatives category genes were found in
the two pathogens C rodentium and C koseri (416 and 428 respectively) while the largest
number of these genes were identified in strain ADS47 (482) and C freundii Ballerup (476)
Despite the similar number of sulfur metabolism category genes determined across the six
species only ADS47 lacks taurine utilization genes Strains ADS47 and Ballerup were found
150
to be rich in the phosphorous metabolism gene category (49-50) while C rodentium contains
a significantly decreased number of these category genes (28)
The bioinformatics analyses found the largest number of carbohydrate category genes
in ADS47 (837) followed by Ballerup (803) The gut bacteria (Citrobacter sp 30_2) codes for
the least number of genes assigned to the carbohydrate category (708) while C koseri C
rodentium and C youngae contained 731-737 in this category genes The analyses also
determined that there are relatively small numbers of TCA and Entner Doudoroff pathway
genes in C rodentium and C youngae compared to the other four species and relatively large
numbers of fermentation-related genes in ADS47 (81) and C freundii (85) compared to the
other four species
A comparison of the pathogenicity operons [locus of enterocyte effacement (LEE)] of
C rodentium with other five Citrobacter genomes revealed the absence of this virulence
determinant gene region in the other four Citrobacter species including ADS47 (Figure 5-7)
The analysis showed the presence of LEE operon genes in C freundii strain Ballerup as no
gap was observed in that genomic location while C freundii ADS47 C koseri C youngae
and Citrobacter sp 30_2 lacked that loci (7a) Further ananlysis showed alignments between
C rodentium and E coli LEE operon genes (Figure 7b) In contrast no alignment of such
genes was observed in ADS47 genome
151
(a)
152
Figure 5-7 (a) Blast Ring Image Generator (BRIG) output based of Citrobacter rodentium
(NC_013716) compared against ADS47 C freundii C koseri C youngae E coli and
Citrobacter sp genomes The targeted comparative genome analysis was performed based on
locus of enterocyte effacement (LEE) operons of C rodentium (Petty et al 2011) The
innermost rings show GC content (black) and GC skew (purple and green) The remaining
rings show BLAST comparisons of the other Citrobacter and E coli genomes against C
rodentium The region shown in red is LEE operon genes (b) The absence of LEE operons in
the ADS47 genome comparing to C rodentium and E coli Analysis was performed and
visualized by Artemis Comparison Tool software v132
(httpwwwsangeracukresourcessoftwareact) (Tatusov et al 2000 Craver et al 2005)
LEE operons
(b)
153
546 Analysis of the unique genes in C freundii ADS47
Two hundred and seventy genes unique to ADS47 were identified from a comparison
of its genome to the other five Citrobacter species In general the unique genes are
distributed throughout the ADS47 genome but some clusters of unique genes occurred
including eight unique genomic regions (Figure 5-8) Six large unique regions are located at
767204-775090 (~8 kb) 1025572-1042510 (17 kb) 2798289-2807217 (~9 kb)
3045220-3055912 (~11 kb) 3930366-3943075 (12 kb) and 4022195-4032444 (10 kb)
Out of 270 unique genes 147 (54 ) are hypothetical proteins The remaining 123
(46) genes are grouped into reductasesoxidoreductases (9 genes) de-acetylase (1)
hydrolases (2) transferases (4) sulfatasesarylsulfatases (3) bacteriocins (3) type VI
secretion proteins (2) kinases (4) transporters (11) membraneexported proteins (8)
regulators (12) integraserecombinases (5) phagesprophagestransposaes related genes (27)
and miscellaneous (32) that includes DNA helicases ligases fimbrial proteins and
chromosome partitioning genes (Table 5-4)
The unique regions encode various functional genes that may cause trichothecene de-
epoxidation or de-acetylation reactions The 8 kb unique regions contains six genes encoding
glycosyltransferases and two copies of oxidoreductase-like coding sequences (CMD_0711
and CMD_0712) that showed 39 identity to the Marinobacter aquaeolei F420
hydrogenasedehydrogenase beta subunit The largest (17 kb) unique genomic region codes
for 16 genes including dehydrogenase hydrolase bacteriocin (pyocin) integrase and a short
chain dehydrogenasereductase-like (SDR) (CMD_0979) gene (Figure 5-9a)
154
The SDR like gene is 49 amino acid similarity to an oxidoreductase of the
phytopathogenic Dickeya dadantii and the enzyme contains a domain with 77 homology to
the Dickeya zeae NAD(P) binding site The 9 kb unique genomic section contains 8 genes
including a NaCl symporter a dehydratase (CMD_2679) with 86 homology to
Enterobacteriaceae bacterium dihydroxy-acid dehydratase and a putative oxidoreductase
(CMD_2683) having 37 similarity with oxopropionate reductase of Bacillus sp The unique
region also encodes a short chain dehydrogenasereductase (SDR) (CMD_2684) that shows
85 identity to Enterobacteriaceae bacterium (Figure 5-9b) A unique thiol-disulfide and
thioredoxin like reductase (CMD_3721) was identified in a 10 kb unique region that contains
13 genes (Figure 5-9c) This gene is 59 identical to Oligotropha carboxidovorans thiol-
disulfide isomerase and thioredoxin In addition several reductasesoxidoreductase-like
unique genes were identified in the ADS47 genome including an oxidoreductase
(CMD_1024) with 27 identity to Canadidatus nitrospira NADH-quinone oxidoreductase
membrane L Another oxidoreductase-like gene identified in a different location of the
ADS47 genome (CMD_1393) shows 87 homology to Klebsiella pneumoniae
deoxygluconate dehydrogenase contains a SDR active site A putative nitrate reductase gene
(CMD_3184) has a low level of identity (27) with an uncultured bacterial respiratory nitrate
reductase The ~11 kb unique region encodes sulfatasesarylsulfatses
(CMD_2898CMD_2905) and related genes Furthermore ADS47 contains a unique
glycosylphosphatidylinositol (GPI) anchor biosynthesis operon comprised of four genes
including a N-acetylglucosaminyl-phosphatidylinositol de-acetylase (CMD_2431) which
showed 78 similarity to E coli de-acetylase (Figure 5-9e)
155
Other unique genespredicted genes in the ADS47 genome include
polysaccharidephenazine biosynthesis genes (CMD_0710CMD_1406) heavy metal kinase
(CMD_0989) two component heavy metal regulator (CMD_0990) nitrate reductase regulator
(CMD_3477) putative potassiumzinc transporters (CMD_0973CMD_4600) sodium
symporter (CMD_2678) (tripartite ATP-independent periplasmic transporters) TRAP
transporters (CMD_3693 CMD_3694 CMD_3695 CMD_4337) glutamineribose ABC
transporters (CMD_3176CMD_3544) toxin transcription regulator (CMD_3464) fimbrial
proteinchaperone (CMD_1695 CMD_2434 CMD_3277 CMD_3278) and ParB
chromosomeplasmid partitioning proteins (CMD_4033CMD_4034)
Majority of the unique genes and their distributions are shown in comparative circular
genome maps (Figure 5-8)
156
Table 5-4 Unique genes of C freundii strain ADS47 Genes were categorized according to
the functional similarity The unique genes were determined by comparing ADS47 with five
sequenced Citrobacter species using OrthoMCL software
Unique gene classes Number
Total unique genes
270
Reductases oxidoreductases 9
De-acetylase 1
Hydrolase 2
Transferase 4
Sulfatase 3
Bacteriocin 2
Type VI secretion 2
Kinase 4
Transporter 9
Membraneexported protein 8
Regulator 12
Integraserecombinase 5
Phageprophagetransposae 27
Miscellaneous 34
Hypothetical protein 147
157
158
Figure 5-8 Locations of the unique genes of C freundii strain ADS47 The unique genes
were determined by comparing the total CDS of strain C freundii ADS47 with C freundii
Ballerup Citrobacter sp C youngae C koseri and C rodentium CDSs by OrthoMCL
program The map was developed by Blast Ring Generator (BRIG) software First second
and third circles from inside to out side show the genome size GC and GC skew of strain
ADS47 Forward (Fwd) and reversed (Rev) transcribed CDSs of ADS47 are shown in green
(forth circle) and red (fifth circle) respectively The next five rings represent the circular
genome maps of five Citrobacter species distinguished by different ring colours (C
rodentium as sky blue C koseri as dark blue C youngae as light blue C freundii Ballerup as
yellow and Citrobacter sp as green) The degree of orthologous gene identity between
ADS47 and other five genomes are shown by color intensity (light color= 75 gene identity
ash color= 50 gene identity) Gaps across five genome rings indicate the absence of
genesregions compared to ADS47 Unique genes are shown surrounding the map rings and
the identities of some of the genes are given Arrows indicate putative
reductasesoxidoreductases and de-acetylase
159
(a)
(b)
160
(c)
(d)
161
Figure 5-9 Unique genomic regions and associated genes in C freundii ADS47 compared to
five Citrobacter species RAST server based analysis compared the unique genesregions of
ADS47 with the publicly available databases The upper panels demonstrate the large unique
regions defined by yellow boxes and genes located in the genomic regions are indicated by
arrows The lower panels are closely related genesoperons found in distantly related species
The unique regions that contain potential trichothecene de-epoxidation genes are shown in (a)
short chain dehydrogenasereductase (SDR) and dehydrogenase (b) putative oxidoreductase
dehydratase and SDR (c) thiol-disulfide isomerase and thioredoxin oxidoreductase and (d)
unique glycosylphosphatidylinositol (GPI) biosynthesis operon having N-acetylglucosaminyl-
phosphatidylinositol de-acetylase gene
162
55 Discussion
The de novo sequenced ADS47 genome was oriented and assembled using a
completely sequenced C koseri genome as reference strain The synteny analysis between
ADS47 and C koseri showed very similar overall organization of the two genomes The high
preservation of genes among five Citrobacter species and ADS47 were emphasized by the
close alignments observed by Blast Ring Image Generator (BRIG) outputs obtained from
these genomes
Genome-wide phylogenetic analyses based on a large number of common CDSs
(2821) observed within the six Citrobacter genomes confirmed the species identification
based on 16S-rDNA sequencing of the trichothecene mycotoxin-detoxifying C freundii strain
ADS47 The base sequence idendity was 8715 between these two genomes They shared
~80 CDS and had a similar GC content (52) Citrobacter are Gram-negative rod shaped
and facultatively anaerobic bacteria belonging to the family of Enterobacteriaceae (Wang et
al 2000 Whalen et al 2007) Strains ADS47 and Ballerup of C freundii are both aerobic
gram-negative bacilli that have numerous flagella and inhabit in soil (Cong et al 2011 Islam
et al 2012) C freundii is considered to have a positive environmental role by reducing nitrate
to nitrite (Rehr and Klemme 1989) which is an important step in the nitrogen cycle
C freundii C rodentium and C koseri are animalhuman pathogenic species C
rodentium is a natural mice pathogen causing hemorrhagic colitis and hemolytic uremic
syndrome (Gareau et al 2010) C koseri causes human disease (Badger et al 1999 Kariholu
et al 2009) particularly neonatal meningitis and C freundii is an opportunistic human
pathogen responsible for a various types of infections like respiratory tract urinary tract and
blood (Whalen et al 2007) C freundii is very common representing approximately 29 of
163
all opportunistic human infections (Hodges et al 1978) Outside of animals they can be found
in soil water sewages and different organs of diseased animals (Wang et al 2000) These
pathogens code for large numbers of mycobacterium virulence operonsgenes which were
absent in the three other genomes However they have fewer genes involved in environmental
stress such as copper homeostasis genes and no mercuricarsenic resistance and secondary
metabolism category genes unlike the other Citrobacter genomes Nonetheless considerable
variability exists between the two pathogens correlates with C rodentium causing disease in
mice C koseri causing meningistis in human and C freundii causing infections in different
human organsThere were attaching and effacing (AE) lesion forming (LEE) operons in C
rodentium and C freundii but not in C koseri Those genes have shown to be acquired by
lateral gene transfer playing major roles in virulence of enteropathogenic and
enterohemorrhagic E coli and C rodentium (Mundy et al 2005 Petty et al 2010 Petty et al
2011 The ~356 kb pathogenic LEE gene island is comprised of five operons (LEE1 to
LEE5) that code for ~40 virulence genes that include type-III secretion genes and associated
effectors (Berdichevsky et al 2005) In addition an elevated number of type-II and -VI
secretion proteins identified in C rodentium which may act as additional virulence factors
(Petty et al 2010) Because it lacks type-III secretion apparatus genes it is likely that C
koseri uses a different strategy to cause human neonatal meningitis (Badger et al 1999 Ong
et al 2010)
To adapt to different environmental niches or hosts bacteria like Citrobacter species
frequently gainlose genes increase repeat sequences and recombine genomic regions using
different mechanisms (Parkhill et al 2001 Petty et al 2011) The existence of large numbers
of phageprophagetransposae related genes in the Citrobacter genomes particularly in C
164
freundii Ballerup suggests that their genomes are unstable For example C freundii Ballerup
and C koseri might have acquired LEE operons by horizontal transfer via phage mediated
transpositions Interestingly although ADS47 is closely related to C freundii Ballerup it
lacks the pathogen-related LEE gene island and mycobacterium virulence operons required
for invasion and intracellular survival in macrophages (Gao et al 2006) Instead ADS47 has a
larger number and variety of stress-responsive genes such as efflux genes and bacteriocins
(pyocin hemolysin) genes that provide protection against heavy metals and antibiotics and
increase their competitiveness against other bacteria These findings suggest that the strain
ADS47 is not an opportunistic animal pathogen but has evolved to be highly competitive in
the soil micro-environment (Arnold 1998 Bengoechea and Skurnik 2000) Perhaps ADS47
evolved trichothecene degrading genes for adaptation to soils containing trichothecene
mycotoxins contaminated soils (Gadd and Griffiths 1978 Sharma and Fulekar 2009 Han et
al 2011) Conversely the absence or reduced number of stress-responsive genes in C
rodentium and C koseri may imply that those two pathogenic species are not exposed to such
stressful and xenobiotic-contaminated environments as ADS47
This interpretation is supported by the observation that the two of the three
animalhuman pathogenic species contained high of rRNA copy number but lower numbers
were found in ADS47 C freundii Ballerup and Citrobacter sp 30_2 Bacteria maintain large
number of rRNA for higher production of ribosome during high growth rates (Stevenson et al
1997 Klappenbach et al 2000) such as might occur during the establishment of an infection
However a large copy number of rRNAs may not be metabolically favourable for bacteria
such as ADS47 that evolved in a highly competitive environment as that costs higher cellular
energy (Hsu et al 1994)
165
C freundii Ballerup seems to be an exception in that it is similar to the other animal
pathogens for containing virulent factors (ie LEE-operon genes) but more like the
environmental species for encoding high number genes for stress defence heavy metal and
toxic compounds resistances
The comparison of the genes in C freundii ADS47 with the unique ability to detoxify
trichothecenes with genes in other Citrobacter species allowed approximately 270 unique
genes to be identified in ADS47 Some of those genes may be involved in trichothecene de-
epoxidation or de-acetylation Currently the pathways and microbial enzymes leading to
trichothecene de-epoxidation and de-acetylation are unknown but based on the required
reaction ie the elimination of an oxygen atom from epoxide moiety it is likely that epoxide
reductases or epoxide oxidoreductases are responsible for such biodegradation activity (He et
al 1992)
From database searches three enzymes having epoxide reductase activity were found
These are a vitamin K epoxide reductase (VKOR) a monooxygenase PaaABCE and pyridine
nucleotide-disulfide oxidoreductase (PNDO) which reduce vitamin K epoxide phenylacetyl-
CoA and epoxyalkanes respectively (Swaving et al 1996 Li et al 2004 Teufel et al 2012)
In mammals VKOR plays important roles in vitamin K cycle by reducing vitamin K epoxide
to vitamin K and its homologues that are found in plants bacteria and archaea (Goodstadt
2004 Li et al 2010) The microbial multi-component PaaABCE monooxygenase catalyzes
degradation of variety of aromatic compounds or environmental contaminants via epoxidation
of phenylacetic acid (Paa) and NADPH-dependant de-epoxidation of epoxy-Paa (Luengo et
al 2001 Teufel et al 2012) PNDO is a unique type of NADP-dependent oxidoreductase
identified in Xanthobacter strain Py2 that was obtained by propane enrichment procedure
166
(van Ginkel and de Bont 1986) However no orthologs of these genes were found in ADS47
genome
Nine ADS47-unique reductaseoxidoreductase-like genes are potential candidates for
the trichothecene de-epoxidation enzyme Genes belong to short chain
dehydrogenasereductase (SDR) family are structurally and functionally diverse and cause
different enzymatic activities including xenobiotic degradation (Kavanagh et al 2008) This is
a promising candidate for the trichothecene de-epoxidation gene Genes belong to dehydratase
family catalyze reduction reactions by cleaving carbon-oxygen bonds of organic compounds
that result in removal of oxygen in various biosynthetic pathways (Flint et al 1993) This
mechanism may be relevant to microbial trichothecene de-epoxidation reaction since the latter
involves removal of the oxygen atom forming the epoxide in trichothecene mycotoxins
(Young et al 2007)
Another ADS47-unique gene was for a nitrate reductase that may be a candidate de-
epoxidation gene Under stress conditions such an enzyme causes reductive removal of an
oxygen molecule (Morozkina and Zvyagilskaya 2007)
A fourth type of ADS47-unique gene is for a thiol-disulfide isomerasethioredoxin like
(Trx) oxidoreductase The thiol-disulfide isomerase and thioredoxin oxidoreductases contain a
thioredoxin like domain with a CXXC catalytic motif which catalyzes various biological
processes that allow the organism to adapt to environmental stress and develop competence to
sporulate (Scharf et al 1998 Petersohn et al 2001 Smits et al 2005) The membrane-
anchored Trxndashlike domain initiates the vitamin K de-epoxidation reaction by reducing the
active motif (CXXC) of VKOR (Schulman et al 2010)
167
The present study did not find any homologues of VKOR in ADS47 genome but we
identified two copies of structurally similar quinone reductase protein disulfide bond
formation (DsbB) gene (CMD_1044 and CMD_2641) Both VKOR and DsbB contain four
transmembrane segments an active site CXXC motif and cofactor quinone and use a
common electron transfer pathway for reducing substrates (Li et al 2010 Malojcic et al
2008) As VKOR bacterial DsbB requires a Trx-like partner (eg DsbA) for activity Further
experiments are required to examine if DON de-epoxidation occurs by the catalytic activity of
DsbB coupled with a unique thiol-disulfide isomerase and thioredoxin oxidoreductase present
in ADS47 genome
The analysis also identified an ADS47-unique glycosylphosphatidylinositol (GPI)
biosynthetic operon that is comprised of four genes including an N-
acetylglucosaminylphosphatidylinositol de-acetylase The GPI membrane anchor protein
synthesis is initiated by transferring N-acetylglucosamine from uridine diphosphate N-
acetylglucosamine to phosphatidylinositol by N-acetylglucosaminyltransferase an
endoplasmic reticulum (ER) membrane-bound protein complex (Hong and Kinoshita 2009)
In the second step of the pathway N-acetylglucosamine-phosphatidylinositol is de-acetylated
to produce glucosaminyl-phosphatidylinositol by the catalytic activity of a zinc
metalloenzyme N-acetylglucosaminylphosphatidylinositol de-acetylase which is localized on
the cytoplasmic side of the ER (Nakamura et al 1997 Hong and Kinoshita 2009) It is
possible that the identified unique N-acetylglucosaminylphosphatidylinositol de-acetylase
gene of ADS47 may be involved in trichothecene de-acetylation activity
168
Further analysis is needed to determine which of these candidate gene or genes may be
involved in tricthothecene mycotoxin de-epoxidation and de-acetylation activities
Identification of gene responsible for the trichothecene detoxification activities of C freundii
ADS47 opens many potential applications for its use in agri-based industries to contribute to
cropanimal production and food safety (Karlovsky 2011)
56 Acknowledgement
We gratefully acknowledge Agriculture and Agri-Food CanadaBinational
Agricultural Research and Development Fund (AAFCBARD) project for providing the
financial supports of the work
169
Supplemental Table S5-1 List of unique genes of strain ADS47 gene IDs and their
functional roles The unique genes were determined by comparing the ADS47 genome with
five sequenced Citrobacter species (C freundii Citrobacter sp C youngae C koseri and C
rodentium) using OrthoMCL (Li et al 2003)
Unique gene
classes
Number Gene ID and functions
Total unique
genes
270 123 known functions 147 hypothetical proteins
Reductases
oxidoreductases
9 CMD_0711 coenzyme F420 hydrogenasedehydrogenase beta subunit
CMD_0979 Short chain dehydrogenase reductase (SDR) Oxidoreductase
SDR dehydrogenase CMD_1024 NADH-quinone oxidoreductase
membrane L CMD_1393 oxidoreductase SDR putative deoxygluconate
dehydrogenase (NAD+ 3-oxidoreductase) CMD_2679
Dehydratasedihydroxy acid dehydratase CMD_2683 oxidoreductase
oxopropionate reductase CMD_2684 short-chain dehydrogenasereductase
SDR CMD_3184 nitrate reductase alfa subunit CMD_3721 thiol-disulfide
isomerase and thioredoxin oxidoreductase
Deacetylase 1 CMD_2431 N-acetylglucosaminylphosphatidylinositol deacetylase
superfamily
Hydrolase 2 CMD_0991 Dienelactone hydrolase CMD_2895 Putative glycosyl
hydrolase
Transferase 4 CMD_1640 D12 class N6 adenine-specific DNA methyltransferase
CMD_2432 Glycosyltransferase CMD_2433 putative acetyltransferase
CMD_3023 adenine-specific DNA modification methyltransferase
Sulfatase 3 CMD_2898 Possible sulfatase arylsulfatasearylsulfatransferase
CMD_2902 Arylsulfatasesulfatase CMD_2905 Putative
sulfatasearylsulfatase
Bacteriocin 3 CMD_0982 Thermosatble hemolysin superfamily CMD_0993 Pyocin S2
superfamily (bacteriocin that kills other bacterial strain) CMD_3705 S-type
pyocin containing domain protein
Type VI secretion 2 CMD_1802 EvpB family type VI secretion protein CMD_2364 type VI
effector Hcp1
Kinase 4 CMD_0989 Heavy metal Sensor kinase CMD_1794 Polo kinase kinase
(PKK) superfamily CMD_3124 predicted diacylglycerol kinase CMD_4266
putative signal transduction kinase
Transporter 11 CMD_0973 TrkA-N domain-containing protein (K uptake transporter)
CMD_1407 Branched-chain amino acid transport system carrier protein
CMD_2678 Sodium Symporter CMD_3176 glutamine ABC transporter
CMD_3544 Ribose ABC transport system CMD_3693 TRAP-type C4-
dicarboxylate transport system binding protein CMD_3694 TRAP-type C4-
dicarboxylate transport system binding protein small permease component
(predicted N-acetylneuraminate transporter) CMD_3695 TRAP-type C4-
dicarboxylate transport system large permease CMD_4337 23-diketo-L-
gulonate TRAP transporter small permease protein yiaM component
CMD_4338 23-diketo-L-gulonate TRAP transporter large permease protein
yiaN CMD_4600 putative transporter involved in Zinc uptake (YciA
protein)
170
Supplmental Table S5-1 (continued)
Unique gene
classes
Number Gene ID and functions
Membrane
exported protein
8 CMD_0713 putative membrane protein wzy CMD_0983 Putative exported
protein precursor CMD_0986 Putative transmembrane protein
CMD_0987 putative exported protein precursor CMD_1458 putative
exported protein CMD_2903 Putative exported protein CMD_2904
putative secreted 5-nucleotidase CMD_4291 (S) putative integral
membrane export protein
Regulator 12 CMD_0977 probable transcriptional regulator protein CMD_0980 Putative
transcription activator TenA family CMD_0990 Two component heavy
metal response transcription regulator CMD_1766 Leucine responsive
regulatory protein CMD_2677 transcriptional regulator GntR CMD_2909
putative arylsulfatase regulator CMD_3279 Uncharacterized outer
membrane usher protein yqiG precursor CMD_3464 toxin transcription
regulatory protein CMD_3465 EAL regulator protein (signaling)
CMD_3475 putative transcription regulator CMD_3477 Fumarate and
nitrate reduction regulatory protein (for anaerobic adaptation) CMD_3812
putative regulator from phage
Integrase
recombinase
5 CMD_0785 CMD_0992 CMD_0994 CMD_4032 CMD_4424
Phage prophage
transposae related
genes
27 Phages CMD_1608 CMD_1609 CMD_1610 CMD_1611 CMD_1619
CMD_1631 CMD_1913 CMD_1914 CMD_1915 CMD_1917
CMD_1920 CMD_1921 CMD_1936 CMD_3351 CMD_3804
CMD_3805 CMD_3807 CMD_3808 CMD_4581 CMD_4609
CMD_4610 Transposae CMD_0252 CMD_0398 Prophage CMD_1645
CMD_3806 CMD_4425 CMD_4597
Miscellaneous 32 CMD_0095 ATP-dependent DNA helicase Rep CMD_0656 LysA protein
CMD_0710 Polysaccharide biosynthesis family protein CMD_0978 TPR
repeat containing protein CMD_0981 AMP dependant synthatase and
ligase CMD_0985 Cupin superfamily CMD_0987 flavodoxin
CMD_1404 Putative endoribonuclease CMD_1405 Putative PAC domain
protein CMD_1406 Phenazine (PhzF) biosynthesis protein CMD_1695
Putative fimbrial protein CMD_1793 HPChromosome segregation ATPase
domain protein CMD_1922 FRG uncharacterized protein domain
CMD_2206 L-fucose isomerase like protein CMD_2434 CS12 fimbria
outer membrane usher protein CMD_2680 HpcHHpaI aldosecitrate lyase
protein family CMD_2681 Aldose -1 epimerase CMD_2682
Gluconolaconase LRE like region CMD_2900 Putative cytoplasmic
protein CMD_3009 putative restriction endonuclease CMD_3277 Putative
yqi fimbrial adhesion CMD_3278 Hypothetical fimbrial chaperone yqiH
CMD_3340 Pea-VEAacid family CMD_3518 3-hydroxy-3-methylglutaryl
CoA synthase CMD_3801 putative reverse transcriptase CMD_3840 beta-
D-galactosidase alpha subunit CMD_4033 ParB family chromosome
partitioning protein CMD_4034 ParBrepB chromosomeplasmid
partitioning protein CMD_4035 RelASpoT protein CMD_4532 putative
DNA ligase -like protein CMD_4541 Antitermination protein Q and
CMD_4585 putative ATP dependent helicase
171
CHAPTER 6
60 CONCLUSION
61 Summary
Contamination of grain with mycotoxins including deoxynivalenol (DON) and other
trichothecenes is becoming a limiting factor for cereal and animal production (Binder et al
2007) To address the issue this study obtained a novel microbial source for DON
detoxification and determined candidate genes for the function which may have potential
applications to detoxify grain and grain-derived foodfeed contaminated with DON and TC
mycotoxins
From a screen of soil samples from crop field microorganism was isolated that is able
to de-epoxidize the most frequently detected foodfeed contaminating mycotoxin DON The
effects of media compositions temperature pH and aerobicanaerobic conditions on
microbial detoxification activity were assessed From 150 analyzed soil samples a mixed soil
microbial culture that showed DON de-epoxidation activity under aerobic conditions and
moderate temperature (~24degC) was obtained This culture exhibited a slow rate of microbial
DON to de-epoxyDON transformation (de-epoxidation) possibly because of competition
between de-epoxidizing and other microorganisms in the culture
The mixed microbial culture was enriched for DON detoxifying bacteria by reducing
the non-targeted microbial species through antibiotic treatments heating and subculturing on
agar medium and selecting colonies The enriched culture had higher DON de-epoxidation
activity than the initial culture Because of low abundance of DON detoxifying
microorganisms further culture enrichments were required to increase the target microbial
172
population Reculture of the partially enriched microbial community in nutritonally poor MSB
medium with high level of DON increased the DON de-epoxidizing bacterial population
Serial dilution plating and selecting single colonies eventually led to isolation of a DON-
detoxifying bacterial strain named ADS47
Phenotypic and 16S-rDNA sequence analyses provided evidence that the bacterium is
a strain of C freundii Strain ADS47 is a gram negative rod-shaped facultative anaerobe and
has peritrichous flagella Strain ADS47 was unable to use DON as sole carbon source
however increasing DON concentrations in growth media enhanced its DON metabolism
efficiency The pure bacterial culture was shown to be able to rapidly de-epoxidize DON
under aerobic conditions moderate temperatures and neutral pH Strain ADS47 more quickly
de-epoxidizes DON in aerobic conditions than the anaerobic conditions In addition to DON
strain ADS47 showed capability to de-epoxidize andor de-acetylate 10 different type-A and
type-B TC mycotoxins which often co-contaminant grain with DON
To clone the DON detoxifying genes a DON detoxifying bacterial DNA library in E
coli host was constructed followed by screening of the library clones for DON detoxification
activity The study produced a library giving approximately 10 fold coverage of the genome
of strain ADS47 Restriction digestion analysis determined that most of the clones contained
ADS47 DNA inserts This indicates that a good DNA library was produced which had gt99
probability of containing all the DNA of the ADS47 genome However activity-based
screening of the library could not identify any clones with DON de-epoxidation ability It is
assumed that bacterial DON de-epoxidation genes evolved in a certain environmental niche
which may be required for the DON de-epoxidation to be expressed and active but was not
supplied by the host E coli cells
173
An attempt was made to identify the bacterial genes encoding DON de-epoxidation
and de-acetylation enzymes by sequencing the genome of strain ADS47 and comparing its
sequence to the genomes of related bacteria that are not known to posses DON de-epoxidation
activity A RAST server based bioinformatics analysis determined that the 48 Mb circular
genome of ADS47 codes for 4610 predicted CDS A genome wide phylogenetic analysis
determined that strain ADS47 was gt87 identical to the pathogen C freundii strain Ballerup
The relationship to two other animalhuman pathogenic species C rodentium and C koseri
was more distant This result is consistent with the 16S-rDNA based phylogenetic analysis
It is assumed that the microbial DON detoxifying genes were acquired by horizontal
gene transfer or other means during the adaptation of ADS47 in mycotoxin contaminated soil
If it is correct then this means that those newly acquired genes are not common in other
related microbial species and no other Citrobacter species is known to degrade DON Hence
the study identified 270 unique genes to ASDS47 with a subset of 10 genes for reductases
oxidoreductases and a de-acetylase which were considered to be candidate genes for the
enzymes that have DON detoxifying activity Furthermore a targeted comparative genome
analysis showed that the mycotoxin detoxifying ADS47 genome does not harbour the
clinically important LEE encoded typendashIII secretion system and mycobacterium virulence
genes This suggests that strain ADS47 is not likely to be an animal pathogenic bacterium
unlike C freundii Ballerup which encodes the pathogenic relevance LEE operons
174
62 Research contributions
The research made four major contributions which may aid in reducing the risks of
animals and humans from DONTC exposure Firstly the study identified a novel bacterial
strain ADS47 that is able to detoxify DON and 10 related mycotoxins over a wide range of
temperatures under aerobic and anaerobic conditions The discovery has led to a patent
application for strain ADS47 supporting the novelty and importance of the finding This
bacterial strain could be used to detoxify trichothecene mycotoxins in contaminated animal
feed thus reducing food safety risks
Secondly the study resulted in the development of a method for isolating DON de-
epoxidizing microbes from soil as well as optimizing the conditions for bacterial DONTC
detoxification activity to increase the effectiveness usefulness of the strain
Thirdly the thesis work resulted in availability of the genome sequence of a
trichothecene detoxifying microbe which may ultimately lead to an understanding of the
cellularmolecular mechanisms of DONTC de-epoxidation and de-acetylation
genespathways The genome analysis resulted in identification of several candidate genes for
DONTC de-epoxidation and de-acetylation activities Genes which show de-epoxidation
activity could be used to develop DONTC resistant cereal cultivars Fourthly strain ADS47
lacks major animal pathogenicity genes (in particular the LEE-operons) as well as does not
encodes mycobacterium virulence genes Thus strain ADS47 has a great potential to be
utilized to detoxify cereal grain and may be used as a feed additive for livestockfishery All
together the reported work may have significant applications to produce and supply
DONTC-free food and feed
175
63 Further research
This research provides the basis for developing biological detoxification of DON and
related mycotoxins This goal might be achieved by using DON de-epoxidizing gene to
develop resistant plants enzymatic detoxification of contaminated feed and use of the bacteria
as a feed additive Follow-up studies may reach the above indicated goals Because the
bacterium showed de-epoxidation activity under aerobicanaerobic conditions and under wide
range of temperatures it is likely to have an application for enzymatic detoxification of
contaminated animal feed Therefore additional studies on bacterial efficiency to detoxify
contaminated cereal and grain-derived feed need to be conducted Although strain ADS47
does not encode major animal pathogenicity genes the bacterium must be directly tested on
animals to see if it could negatively affect animal digestive systems If not then the potential
for ADS47 is considerable for its use as a feed additive for pig poultry and fish Animal
feeding trials will also need to be performed to determine if DON detoxification is sufficient
to be cost effective as a feed additive In order to develop DONTC resistant cereal cultivars
the gene responsible to DON degrading activity among the presently identified candidate
genes (reductases oxidoreductases and de-acetylase) will need to be expressed in other
organisms like E coli and a codon modified version adapted for expression in plants like
corn and wheat
176
70 REFERENCES
Abbas HK Mirocha CJ Pawlosky RJ Pusch DJ 1985 Effect of cleaning milling and
baking on deoxynivalenol in wheat Applied and Environmental Microbiology 50482ndash
486
Accensi F Pinton P Callu P Abella-Bourges N Guel WJF Grosjean F Oswald IP 2006
Ingestion of low doses of deoxynivalenol does not affect haematological biochemical or
immune responses of piglets Journal of Animal Science 841935ndash1942
Agrawal D Mahapatra AK 2005 Vertically acquired neonatal Citrobacter brain abscessndash
case report and review of the literature Journal of Clinical Neuroscience 12188ndash190
Alexander NJ 2008 The TRI101 story engineering wheat and barley to resist Fusarium head
blight World Mycotoxin Journal 131-37
Alexander NJ Proctor RH McCormick SP 2009 Genes gene clusters and biosynthesis of
trichothecenes and fumonisins in Fusarium Toxin Reviews 28198ndash215
Ali ML Taylor JH Jie L Sun G William M Kasha KJ Reid LM Pauls KP 2005
Molecular mapping of QTLs for resistance to Gibberella ear rot in corn caused by
Fusarium graminearum Genome 48521-533
Alikhan N-F Petty NK Zakour NLB Beatson SA 2011 BLAST Ring Image Generator
(BRIG) simple prokaryote genome comparisons BMC Genomics 12402
Alisi A Spaziani A Anticoli S Ghidinelli M Balsano C 2008 PKR is a novel functional
direct player that coordinates skeletal muscle differentiation via p38MAPKAKT
pathways Cellular Signalling 20534ndash542
177
Amuzie CJ Shinozuka J Pestka JJ 2009 Induction of suppressors of cytokine signaling by
the trichothecene deoxynivalenol in the mouse Toxicological Science 111277ndash287
Ansorge WJ 2009 Next generation DNA sequencing techniques New Biotechnology
25195-203
Arnold JW Senter SD 1998 Use of digital aroma technology and SPME GC-MS to compare
volatile compounds produced by bacteria isolated from processed poultry Journal of the
Science of Food and Agriculture 78343ndash348
Audenaert K Van Broeck R Bekaert B De Witte F Heremans B Messens K Hofte
M Haesaert G 2009 Fusarium head blight (FHB) in Flanders population
diversity inter-species associations and DON contamination in commercial winter
wheat varieties European Journal of Plant Pathology 125445-458
Awad WA Ghareeb K Bohm J Zentek J 2010 Decontamination and detoxification
strategies for the Fusarium mycotoxin deoxynivalenol in animal feed and the
effectiveness of microbial biodegradation Food Additives and Contaminants 27510-520
Aziz K Bartels D Best AA DeJongh M Disz T Edwards RA Formsma K Gerdes S Glass
EM Kubal M Meyer F Olsen GJ Olson R Osterman AL Overbeek RA McNeil LK
Paarmann D Paczian T Parrello1 B Pusch GD Reich C Stevens R Vassieva O
Vonstein V Wilke A Zagnitko O 2008 The RAST Server Rapid Annotations using
Subsystems Technology BMC Genomics 975
Badger JL Stins MF Kim KS 1999 Citrobacter freundii invades and replicates in human
brain microvascular endothelial cells Infection and Immunology 674208ndash4215
Bae HK Pestka JJ 2008 Deoxynivalenol induces p38 interaction with the ribosome in
monocytes and macrophages Toxicological Science 10559-66
178
Bai GH Desjardins AE Plattner RD 2002 Deoxynivalenol-nonproducing Fusarium
graminearum causes initial infection but does not cause disease spread in wheat spikes
Mycopathologia 15391ndash98
Barrett JR 2000 Mycotoxins of molds and maladies Environmental Health Perspectives
108A20ndashA23
Bengoechea JA Skurnik M 2000 Temperature-regulated efflux pumppotassium antiporter
system mediates resistance to cationic antimicrobial peptides in Yersinia Molecular
Microbiology 3767ndash80
Bennett JW Klich M 2003 Mycotoxins Clinical Microbiology Reviews 16497ndash516
Benson DA Karsch-Mizrachi I Lipman DJ Ostell J Sayers EW 2009 GenBank Nucleic
Acids Research 37D26-31
Berdichevsky T Friedberg D Nadler C Rokney A Oppenheim A Rosenshine I 2005 Ler is
a negative autoregulator of the LEE1 operon in neuropathogenic Escherichia coli Journal
of Bacteriology 187349ndash357
Bernardo A Bai GH Guo PG Xiao K Guenzi AC Ayoubi P 2007 Fusarium graminearum-
induced changes in gene expression between Fusarium head blight-resistant and
susceptible wheat cultivars Functional and Integrative Genomics 769ndash77
Bhat RV Beedu SR Ramakrishna Y Munshi KL 1989 Outbreak of trichothecene
mycotoxicosis associated with consumption of mould-damaged wheat production in
Kashmir Valley India The Lancet 33335ndash37
Binder EM Tan LM Chin LJ Handl J Richard J 2007 Worldwide occurrence of
mycotoxins in commodities feeds and feed ingredients Animal Feed Science and
Technology 137265-282
179
Birzele B Meier A Hindorf H Kraumlmer J Dehne H-W 2002 Epidemiology of Fusarium
infection and deoxynivalenol content in winter wheat in the Rhineland Germany
European Journal of Plant Pathology 108667ndash673
Boddu J Cho S Muehlbauer GJ 2007 Transcriptome analysis of trichothecene-induced gene
expression in barley Molecular PlantndashMicrobe Interactions 201364ndash1375
Borenshtein D Schauer DB 2006 The Genus Citrobacter Prokaryotes 690ndash98
Boutigny A-L Richard-Forget F Barreau C 2008 Natural mechanisms for cereal resistance
to the accumulation of Fusarium trichothecenes European Journal of Plant Pathology
121411ndash423
Boutrif E Canet C 1998 Mycotoxin prevention and control FAO programmes Revue de
Meacutedecine Veacuteterinaire 149681-694
Brennan JM Leonard G Fagan B Cooke BM Ritieni A Ferracane R et al 2007
Comparison of commercial European wheat cultivars to Fusarium infection of head and
seedling tissue Plant Pathology 5655ndash64
Brenner DJ Grimont PA Steigerwalt AG Fanning GR Ageron E Riddle CF 1993
Classification of Citrobacteria by DNA hybridization designation of Citrobacter farmeri
sp nov Citrobacter youngae sp nov Citrobacter braakii sp nov Citrobacter
werkmanii sp nov Citrobacter sedlakii sp nov and three unnamed Citrobacter
genomospecies International Journal of Systematic Bacteriology 43645-58
Brookes G Barfoot P 2006 GM Crops The first ten years-global socio-economic and
environmental Impacts no 36 ISAAA (International Service for the Acquisition of Agri-
Biotech Applications) Ithaca New York
180
Bruce KD Hughes MR 2000 Terminal restriction fragment length polymorphism monitoring
of genes amplified directly from bacterial communities in soils and sediments Molecular
Biotechnology 16261ndash269
Buerstmayr H Ban T Anderson JA 2009 QTL mapping and marker-assisted selection for
Fusarium head blight resistance in wheat A review Plant Breeding 1281ndash26
Campbell H Choo TM Vigier B Underhill L 2002 Comparison of mycotoxin profiles
among cereal samples from eastern Canada Canadian Journal of Botany 80526ndash532
Carver et al 2005 ACT the Artemis Comparison Tool Bioinformatics 213422-3423
Carver et al 2009 DNAPlotter circular and linear interactive genome visualization
Bioinformatics 25119-120
Chen CY et al 2003 Comparative genome analysis of Vibrio vulnificus a marine pathogen
Genome Research 132577ndash2587
Chiu TC Yen JH Hsieh YN Wang YS 2005 Reductive transformation of dieldrin under
anaerobic sediment culture Chemosphere 601182ndash1189
Chung YJ Zhou HR Pestka JJ 2003 Transcriptional and posttranscriptional roles for p38
mitogen-activated protein kinase in upregulation of TNF-alpha expression by
deoxynivalenol (vomitoxin) Toxicology and Applied Pharmacology 193188ndash201
Chung EJ Lim HK Kim J-C Choi GJ Park EJ Lee MH Chung YR Lee S-W 2008 Forest
soil metagenome gene cluster involved in antifungal activity expression in Escherichia
coli Applied and Environmental Microbiology 74723-730
Collins TFX Sprando RL Black TN Olejnik N Eppley RM Hines FA Rorie J Ruggles DI
2006 Effects of deoxynivalenol (DON vomitoxin) on in utero development in rats Food
and Chemical Toxicology 44747ndash757
181
Cong Y Wang J Chen Z Xiong K Xu Q Hu F 2011 Characterization of swarming motility
in Citrobacter freundii FEMS Microbiology Letters 371160-171
Cossette F Miller JD 1995 Phytotoxic effect of deoxynivalenol and Gibberella ear rot
resistance of corn Natural Toxins 3383ndash388
Creppy EE 2002 Update of survey regulation and toxic effects of mycotoxins in Europe
Toxicological Letters 12719-28
Dai JS Wang D Guerlebeck C Laturnus S Guenther Z Lu SC Ewers C 2010 Suppression
subtractive hybridization identifies an autotransporter adhesin gene of E coli IMT5155
specifically associated with avian pathogenic Escherichia coli (APEC) BMC
Microbiology 10236
Daniel R 2005 The metagenomics of soil Nature Review Microbiology 3470-478
Davis KER Joseph SJ Janssen PH 2005 Effects of growth medium inoculum size and
incubation time on culturability and isolation of soil bacteria Applied and Environmental
Microbiology 71826-834
Delcher AL Phillippy A Carlton J Salzberg SL 2002 MUMmer 21 NUCmer and
PROmer are described in Fast Algorithms for Large-scale Genome Alignment and
Comparisonrdquo Nucleic Acids Research 302478-2483
Demain AL Adrio JL 2008 Contributions of microorganisms to industrial biology
Molecular Biotechnology 3841ndash455
Deng W Li Y Vallance BA Finlay BB 2001 Locus of enterocyte efffacement from
Citrobacter rodentium sequence analysis and evidence for horizontal transfer among
attaching and effacing pathogens Infection and Immunity 696323ndash6335
182
Desjardins AE Proctor RH Bai G McCormick SP Shaner G Beuchley G Hohn TM 1996
Reduced virulence of trichothecene-nonproducing mutants of Gibberella zeae in wheat
field tests Molecular Plant-Microbe Interactions 9775-781
Desjardins AE 2006 Fusarium mycotoxins chemistry genetics and biology The American
Phytopathological Society St Paul Minnesota USA p 260
Desjardins AE Proctor RH 2007 Molecular biology of Fusarium mycotoxins International
Journal of Food Microbiology 11947-50
Desmond OJ Manners JM Stephens AE MacLean DJ Schenk PM Gardiner DM Munn
AL Kazan K 2008 The Fusarium mycotoxin deoxynivalenol elicits hydrogen peroxide
production programmed cell death and defence responses in wheat Molecular Plant
Pathology 9435-445
Dill-Macky R Jones RK 2000 The effect of previous crop residues and tillage on Fusarium
head blight of wheat Plant Disease 8471ndash76
DrsquoMello JPF Macdonald AMC Dijksma WTP 1998 3-Acetyl deoxynivalenol and esterase
production in a fungicide-insensitive strain of Fusarium culmorum Mycotoxin Research
149ndash18
DrsquoMello JPF Placinta CM Macdonald AMC 1999 Fusarium mycotoxins a review of
global implications of animal health welfare and productivity Animal Feed Science and
Technology 80183-205
Doohan FM Brennan J Cooke BM 2003 Influence of climatic factors on Fusarium species
pathogenic to cereals European Journal of Plant Pathology 109755ndash768
183
Dyer RB Kendra DF Brown DW 2006 Real-time PCR assay to quantify Fusarium
graminearum wild-type and recombinant mutant DNA in plant material Journal of
Microbiological Methods 67534ndash542
Erecinska M Wilson DF 1981 Inhibitors of cytochrome c oxidase In inhibitors of
mitochondrial function edited by M Erecinska and DF Wilson Pergamon Press Ltd
Oxford pp 145-160
Eriksen GS 2003 Metabolism and toxicity of trichothecene Doctoral thesis Swedish
University of Agricultural Sciences Uppsala Sweden
Eriksen GS Pettersson H Lundh T 2004 Comparative cytotoxicity of deoxynivalenol
nivalenol their acetylated derivatives and de-epoxy metabolites Food Chemistry and
Toxicology 42619-624
Eudes F Comeau A Rioux S Collin J 2000 Phytotoxicity of eight mycotoxins associated
with Fusarium in wheat head blight Canadian Journal of Plant Pathology 22286ndash292
FAO (Food and Agriculture Organization) 1991 Food Nutrition and Agriculture Food for
the Future volume 1 FAO Rome
FAO (Food and Agriculture Organization) 2001 FNP73 Manual of the application of
HACCP system in mycotoxin prevention and control FAO Food and Nutrition paper no
73 FAO Rome
Fedorak PM Grbic-Galic D 1991 Aerobic microbial cometabolism of benzothiophene and
3-methylbenzothiophene Applied and Environmental Microbiology 57932-940
Felsenstein J 1989 PHYLIP - Phylogeny Inference Package (Version 32) Cladistics 5164-
166
184
Fernando WGD Amarasinghe C Demoz B Surujdeo‐Maharaj S Gauthier V Guerrieri G
Al‐Taweel K Brucircleacute‐Babel A Tamburic‐Illincic L Gilbert J 2011 What do we know of
the Fusarium graminearum chemotypes their epidemiology genetics and infection
process in Canada In Proc 7th
Canadian Workshop on Fusarium Head Blight edited by
J Gilbert A Tekauz N Sweetland and K Slusarenko Winnipeg Manitoba Canada p 45
Fierer N Jackson RB 2006 The diversity and biogeography of soil bacterial community
Proceedings of the National Academy of Sciences 103626-631
Fink-Gremmels J 1989 The significance of mycotoxin assimilation for meat animals
Deutsche Tierarztliche Wochenschrift 96360-363
Flint DH Emptage MH Finnegan MG Fu W Johnson MK 1993 The role and properties of
the iron-sulfur cluster in Escherichia coli dihydroxy-acid dehydratase Journal of
Biological Chemistry 26814732ndash14742
Foroud NA Eudes F 2009 Trichothecenes in cereal grains International Journal of
Molecular Science 10147-173
Forsell JH Jensen R Tai JH Witt M Lin WS Pestka JJ 1987 Comparison of acute
toxicities of deoxynivalenol (vomitoxin) and 15-acetyldeoxynivalenol in the B6C3F1
mouse Food and Chemical Toxicology 25155ndash162
Frey M Stettner C Gierl A 1998 A general method for gene isolation in tagging approaches
amplification of insertion mutagenised sites (AIMS) The Plant Journal 13717-721
Freeman GG Morrison RI 1949 The isolation and chemical properties of trichothecin an
antifungal substance from Trichothecium roseum Link Biochemical Journal 441ndash5
185
Fuchs E Binder EM Heidler D Krska R 2002 Structural characterization of metabolites
after the microbial degradation of type A trichothecenes by the bacterial strain BBSH 797
Food Additives and Contaminants 19379ndash386
Gabor EM Alkema WBL Janssen DB 2004 Quantifying the accessibility of the
metagenome by random expression cloning techniques Environmental Microbiology
6879ndash886
Gadd GM Griffiths AJ 1978 Microorganisms and heavy metal toxicity Microbial Ecology
4303ndash317
Gale LR Bryant JD Calvo S Giese H 2005 Chromosome complement of the fungal plant
pathogen Fusarium graminearum based on genetic and physical mapping and cytological
observations Genetics 171985ndash1001
Gao LY Pak M Kish R Kajihara K Brown EJ 2006 A mycobacterial operon essential for
virulence in vivo and invasion and intracellular persistence in macrophages Infection and
Immunity 741757ndash1767
Gareau MG Wine E Reardon C et al 2010 Probiotics prevent death caused by Citrobacter
rodentium infection in neonatal mice Journal of Infectious Diseases 20181e91
Gareis M Bauer J Thiem J Plank G Grabley S Gedek B 1990 Cleavage of
zearalenoneglycoside a masked mycotoxin during digestion in swine Journal of
Veterinary Medicine 37236-240
Geng L Xin W Haung DW Feng G 2006 A universal cloning vector using vaccinia
topoisomerase I Molecular Biotechnology 3323-28
186
Golkari S Gilbert J Ban T Procunier JD 2009 QTLspecific microarray gene expression
analysis of wheat resistance to Fusarium head blight in Sumai-3 and two susceptible
NILs Genome 52409ndash418
Goodstadt L Ponting CP 2004 Vitamin K epoxide reductase homology active site and
catalytic mechanism Trends in Biochemical Sciences 29289-292
Goswami RS Kistler HC 2004 Heading for disaster Fusarium graminearum on cereal
crops Molecular Plant Pathology 5515-525
Gouze ME Laffitte J Printon P Dedieux G Galinier A Thouvenot JP Loiseau N Oswald
IP Galtier P 2007 Effect of subacute oral doses of nivalenol on immune and metabolic
defence systems in mice Veterinary Research 38635ndash646
Guan S He J Young JC Zhu H Li X Ji C Zhou T 2009 Transformation of trichothecene
mycotoxins by microorganisms from fish digesta Aquaculture 290290ndash295
Gupta R Beg QK Lorenz P 2002 Bacterial alkaline proteases molecular approaches and
industrial applications Applied Microbiology and Biotechnology 5915ndash32
Haberle J Schweizer G Schondelmaier J Zimmermann G Hart L 2009 Mapping of QTL
for resistance against Fusarium head blight in the winter wheat population
PelikanBussardNing8026 Plant Breeding 12827ndash35
Hall BG 2004 Predicting the evolution of antibiotic resistance genes Nature Reviews
Microbiology 2430-435
Han JI et al 2011 Complete genome sequence of the metabolically versatile plant growth-
promoting endophyte Variovorax paradoxus S110 Journal of Bacteriology 1931183ndash
1190
187
Harvey RB Edrington TS Kubena LF Elissalde MH Corrier DE Rottinghaus GE 1995
Effect of aflatoxin and diacetoxyscirpenol in ewe lambs Bulletin of Environmental
Contamination and Toxicology 54325-330
Harvey RB Edrington TS Kubena LF Elissalde MH Casper HH Rottinghaus GE Turk JR
1996 Effects of dietary fumonisin B1-containing culture material deoxynivalenol-
contaminated wheat or their combination on growing barrows American Journal of
Veterinary Research 571790-1794
He J 2007 Microbial strategies for management of Fusarium graminearum and
transformation of deoxynivalenol Master of Science Thesis University of Guelph
Guelph Ontario Canada
He K Vines L Pestka JJ 2010a Deoxynivalenol-induced modulation of microRNA
expression in RAW 2647 macrophages-A potential novel mechanism for translational
inhibition Toxicologist 114310
He J Zhou T Young JC Boland GJ Scott PM 2010b Chemical and biological
transformations for trichothecene mycotoxins in human and animal food chains A review
Trends in Food Science and Technology 2167-76
He J Zhou T 2010 Patented techniques for detoxification of mycotoxins in feeds and food
matrices Rec Pat Food Nutr Agric 296ndash104
He P Young LG Forsberg C 1992 Microbial transformation of deoxynivalenol (vomitoxin)
Applied and Environmental Microbiology 583857-3863
Hell K Cardwell KF Setamou M Poehling H-M 2000 The influence of storage practices on
aflatoxins contamination in maize in four agroecological zones in Benin West Africa
Journal of Stored Product Research 36365ndash82
188
Henne A Schmitz RA Bomeke M Gottschalk G Daniel R 2000 Screening of
environmental DNA libraries for the presence of genes conferring lipolytic activity on
Escherichia coli Applied and Environmental Microbiology 663113ndash3116
Hodges GR Degener CE Barnes WG 1978 Clinical significance of Citrobacter isolates
American Journal of Clinical Pathology 7037ndash40
Hollinger K Ekperigin HE 1999 Mycotoxicosis in food producing animals The Veterinary
Clinics of North America Food Animal Practice 15133-165
Hong Y Kinoshita T 2009 Trypanosome glycosylphosphatidylinositol biosynthesis Korean
Journal of Parasitology 47197ndash204
Hope RJ Aldred D Magan N 2005 Comparison of environmental profiles for growth and
deoxynivalenol production by Fusarium culmorum and F graminearum on wheat grain
Letters in Applied Microbiology 40295ndash300
Horvath RS 1972 Microbial co-metabolism and the degradation of organic compounds in
nature Bacteriological Reviews 36146ndash155
Hsu D Shih LM Zee YC 1994 Degradation of rRNA in Salmonella strains a novel
mechanism to regulate the concentrations of rRNA and ribosomes Journal of
Bacteriology 1764761ndash4765
Huwig A Freimund S Kappeli O Dutler H 2001 Mycotoxin detoxification of animal feed
by different adsorbents Toxicological Letters 122179ndash188
Iordanov MS Pribnow D Magun JL Dinh TH Pearson JA Magun BE 1997 Ribotoxic
stress response activation of the stress activated protein kinase JNK1 by inhibitors of the
peptidyl transferase reaction and by sequence-specific RNA damage to the sarcinricin
loop in the 28S rRNA Molecular and Cellular Biology 173373ndash3381
189
Islam R Zhou T Young JC Goodwin PH Pauls KP 2012 Aerobic and anaerobic de-
epoxidation of mycotoxin deoxynivalenol by bacteria originating from agricultural soil
World Journal of Microbiology and Biotechnology 287-13
Jackson LS Bullerman LB 1999 Effect of processing on Fusarium mycotoxins Advances in
Experimental Medicine and Biology 459243-261
Jansen C von Wettstein D Schafer W Kogel K-H Felk A Maier FJ 2005 Infection patterns
in barley and wheat spikes inoculated with wild-type and trichodiene synthase gene
disrupted Fusarium graminearum Proceedings of the National Academy of Sciences
10216892ndash16897
Jube S Borthakur D 2007 Expression of bacterial genes in transgenic tobacco methods
applications and future prospects Electronic Journal of Biotechnology 10452-467
Kabak B Dobson AD 2009 Biological strategies to counteract the effects of mycotoxins
Journal of Food Protection 722006-2016
Kant P Reinprecht Y Martin CJ Islam R Pauls KP 2011 Integration of biotechnologies
disease resistance pathology and Fusarium In Comprehensive Biotechnology edited by
Moo-Young M Second Edition 4729-743
Kaplan CW Kitts CL 2004 Bacterial succession in a petroleum land treatment unit Applied
and Environmental Microbiology 701777-1786
Kaprelyants AS Kell DB 1996 Do bacteria need to communicate with each other for
growth Trends in Microbiology 4237-42
190
Kariholu U Rawal J Namnyak S 2009 Neonatal Citrobacter koseri meningitis and brain
abscess The Internet Journal of Pediatrics and Neonatology 10
(httpwwwispubcomjournalthe-internet-journal-of-pediatrics-and-
neonatologyvolume-10-number-1neonatal-citrobacter-koseri-meningitis-and-brain-
abscesshtml) (accessed on April 19 2012)
Karlovsky P 1999 Biological detoxification of fungal toxins and its use in plant breeding
feed and food production Natural Toxins 71ndash23
Karlovsky P 2011 Biological detoxification of the mycotoxin deoxynivalenol and its use in
genetically engineered crops and feed additives Applied Microbiology and
Biotechnology 91491-504
Kavanagh KL Jornvall H Persson B Oppermann U 2008 Medium and short-chain
dehydrogenasereductase gene and protein families the SDR superfamily functional and
structural diversity within a family of metabolic and regulatory enzymes Cellular and
Molecular Life Sciences 653895ndash3906
Khera KS Whalen C Angers G Vesonder RF Kuiper-Goodman T 1982 Embryotoxicity of
4-deoxynivalenol (vomitoxin) in mice Bulletin of Environmental Contamination and
Toxicology 29487ndash491
Kikot GE Hours RA Alconada TM 2009 Contribution of cell wall degrading enzymes to
pathogenesis of Fusarium graminearum a review Journal of Basic Microbiology 49
231ndash241
Kim UJ Shizuya H Jong PJ Birren B Simon MI 1992 Stable propogation of cosmid sized
human DNA inserts in a F factor based vector Nucleic Acids Research 201083-1065
191
Kimura M Kaneko I Komiyama M Takatsuki A Koshino H Yoneyama K Yamaguchi I
1998 Trichothecene 3-O-acetyltransferase protects both the producing organism and
transformed yeast from related mycotoxins cloning and characterization of Tri101
Journal of Biological Chemistry 2731654-1661
Kimura M Anzai H Yamaguchi I 2001 Microbial toxins in plantndashpathogen interactions
biosynthesis resistance mechanisms and significance The Journal of General and
Applied Microbiology 47149ndash160
Kimura M Tokai T Takahashi-Ando N Ohsato S Fujimura M 2007 Molecular and genetics
studies of Fusarium trichothecene biosynthesis pathways genes and evolution
Bioscience Biotechnology and Biochemistry 712105-2123
King RR McQueen RE Levesque D Greenhalgh R 1984 Transformation of deoxynivalenol
(vomitoxin) by rumen microorganisms Journal of Agricultural and Food Chemistry
32181ndash1183
Klappenbach JA Dunbar JM Schmidt TM 2000 rRNA operon copy number reflects
ecological strategies of bacteria Applied and Environmental Microbiology 661328ndash
1333
Knietsch A Waschkowitz T Bowien S Henne A Daniel R 2003 Metagenomes of complex
microbial consortia derived from different soils as sources for novel genes conferring
formation of carbonyls from short-chain polyols on Escherichia coli Journal of Molecular
Microbiology and Biotechnology 546ndash56
Kollarczik B Gareis M Hanelt M 1994 In vitro transformation of the Fusarium mycotoxins
deoxynivalenol and zearalenone by the normal gut microflora of pigs Natural Toxins
2105ndash110
192
Koonin EV Wolf YI Aravind L 2001 Prediction of the archaeal exosome and its
connections with the proteasome and the translation and transcription machineries by a
comparative-genomic approach Genome Research 11240ndash252
Kouadio JH Dano SD Moukha S Mobio TA Creppy EE 2007 Effects of combinations of
Fusarium mycotoxins on the inhibition of macromolecular synthesis malondialdehyde
levels DNA methylation and fragmentation and viability in Caco-2 cells Toxicon
49306ndash317
Krause L Diaz NN Bartels D Edwards RA Puumlhler A et al 2006 Finding novel genes in
bacterial communities isolated from the environment Bioinformatics 22281ndash289
Kubena LF Huff WE Harvey RB Phillips TD Rottinghaus GE 1989 Individual and
combined toxicity of deoxynivalenol and T-2 toxin in broiler chicks Poultry Science
68622ndash626
Kubena LF Harvey RB Huff WE Corrier DE Phillips TD Rottinghaus GE 1990 Efficacy
of hydrated sodium calcium aluminosilicate to reduce the toxicity of aflatoxin and T-2
toxin Poultry Science 691078ndash1086
Kubena LF Harvey RB Huff WE Elissalde MH Yersin AG Phillips TD Rottinghaus GE
1993 Efficacy of hydrated sodium calcium aluminosilicate to reduce the toxicity of
aflatoxin and diacetoxyscirpenol Poultry Science 7251ndash59
Kubena LF Edrington TS Harvey RB Buckley SA Phillips TD Rottinghaus GE Casper
HH 1997 Individual and combined effects of fumonisin B1 present in Fusarium
moniliforme culture material and T-2 toxin or deoxynivalenol in broiler chicks Poultry
Science 761239ndash1247
193
Kumar RA Gunasekaran P Lakshmanan M 1999 Biodegradation of tannic acid by
Citrobacter freundii isolated from a tannery effluent Journal of Basic Microbiology
39161-168
Kushiro M 2008 Effects of milling and cooking processes on the deoxynivalenol content in
wheat International Journal Molecular Science 92127-2145
Lane DJ 1991 16S23S rRNA sequencing In Nucleic acid techniques in bacterial
systematic edited by Stackebrandt E and Goodfellow M Wiley J and Sons Inc New
York pp 115-175
Lancova K Hajslova J Poustka J Krplova A Zachariasova M Dostalek P Saschambula L
2008 Transfer of Fusarium mycotoxins and ldquomaskedrdquo deoxynivalenol (deoxynivalenol-3-
glucoside) from field barley through malt to beer Food Additives and Contaminants
25732ndash744
Langmead B Trapnell C Pop M Salzberg SL 2009 Ultrafast and memory-efficient
alignment of short DNA sequences to the human genome Genome Biology 10R25
Larsen JC Hunt J Perrin I Ruckenbauer P 2004 Workshop on trichothecenes with a focus
on DON summary report Toxicological Letters 1531-22
Lee T Oh DW Kim HS Lee J Kim YH Yun SH Lee YW 2001 Identification of
deoxynivalenol- and nivalenol-producing chemotypes of Gibberella zeae by using PCR
Applied and Environmental Microbiology 672966ndash2972
Lee S-W Won K Lim HK Kim J-C Choi GJ Cho KY 2004 Screening for novel lipolytic
enzymes from uncultured soil microorganisms Applied and Environmental Microbiology
65720-726
194
Li L Christian J Stoeckert J David SR 2003 OrthoMCL Identification of Ortholog Groups
for Eukaryotic Genomes Genome Research 132178-2189
Li T Chang CY Jin DY Lin PJ Khvorova A Stafford DW 2004 Identification of the gene
for vitamin K epoxide reductase Nature 427541ndash544
Li Y 2007 Sensory signal transduction in the vagal primary afferent neurons Current
Medicinal Chemistry 142554ndash2563
Li MX Pestka JJ 2008 Comparative induction of 28S ribosomal RNA cleavage by ricin and
the trichothecenes deoxynivalenol and T-2 toxin in the macrophage Toxicological
Science 10567ndash78
Li R Li Y Kristiansen K Wang J 2008 SOAP short oligonucleotide alignment program
Bioinformatics 24713-4
Li W Schulman S Dutton RJ Boyd D Beckwith J Rapoport TA 2010 Structure of a
bacterial homologue of vitamin K epoxide reductase Nature 463507-512
Lichstein HC Malcolm HS 1943 Studies of the effect of sodium azide on microbic growth
and respiration Journal of Bacteriology 47221ndash230
Limay-Rios V Schaafsma AW 2011 2010 Update on the development of an integrated
mycotoxin management system in Ontario corn 30th
Centralia Swine Research Update
Kirkton Ontario Canada
Llorens A Hinojo MJ Mateo R Gonzalez-Jaen MT Valle- Algarra FM Logrieco A Jimenez
M 2006 Characterization of Fusarium spp isolates by PCR-RFLP analysis of the
intergenic spacer region of the rRNA gene (rDNA) International Journal of Food
Microbiology 106287ndash306
195
Lombaert GA Pellaers P Roscoe V Mankotia M Neil R Scott PM 2003 Mycotoxins in
infant cereal foods from the Canadian retail market Food Additives and Contaminants
20494ndash504
Luengo JM Garciacutea JL Olivera ER 2001 The phenylacetyl-CoA catabolon a complex
catabolic unit with broad biotechnological applications Molecular Microbiology
391434ndash1442
Luo X 1994 Food poisoning caused by Fusarium toxins In Proceedings of the Second
Asian Conference on Food Safety ILSI Thailand pp 129ndash136
Mabagala RB 1997 The effect of populations of Xanthomonas campestris pv phaseoli in
bean reproductive tissues on seed infection of resistant and susceptible bean genotypes
European Journal of Plant Pathology 103175ndash181
Madhyastha MS Marquardt RR Abramson D 1994 Structurendashactivity relationships and
interactions among trichothecene mycotoxins as assessed by yeast bioassay Toxicon
321147ndash1152
Maier FJ Miedaner T Hadeler B Felk A Salomon S Lemmens M Kassner H Schaumlfer W
2006 Involvement of trichothecenes in fusarioses of wheat barley and maize evaluated
by gene disruption of the trichodiene synthase (Tri5) gene in three field isolates of
different chemotype and virulence Molecular Plant Pathology 7449ndash461
Malojcic G Owen RL Grimshaw JP Glockshuber R 2008 Preparation and structure of the
charge-transfer intermediate of the transmembrane redox catalyst DsbB FEBS Letters
5823301ndash3307
196
Manoharan M Dahleen LS Hohn TM Neate SM Yu XH Alexander NJ et al 2006
Expression of 3-OH trichothecene acetyltransferase in barley (Hordeum vulgare L) and
effects on deoxynivalenol Plant Science 171699ndash706
Maresca M Mahfoud R Garmy N Fantini J 2002 The mycotoxin deoxynivalenol affects
nutrient absorption in human intestinal epithelial cells The Journal of Nutrition
1322723ndash2731
Martins ML Martins HM 2002 Influence of water activity temperature and incubation time
on the simultaneous production of deoxynivalenol and zearalenone in corn (Zea mays) by
Fusarium graminearum Food Chemistry 79315ndash318
Masuda D Ishida M Yamaguchi K Yamaguchi I Kimura M Nishiuchi T 2007 Phytotoxic
effects of trichothecenes on the growth and morphology of Arabidopsis thaliana Journal
of Experimental Botany 581617ndash1626
Matsumoto E Kawanaka Y Yun SJ Oyaizu H 2008 Isolation of dieldrin- and endrin-
degrading bacteria using 12-epoxycyclohexane as a structural analog of both compounds
Applied Microbiology and Biotechnology 801095-1103
McLean M 1996 The phytotoxicity of Fusarium metabolites an update since 1989
Mycopathologia 133163ndash179
Merhej J Richard-Forget F Barreau C 2011 Regulation of trichothecene biosynthesis in
Fusarium recent advances and new insights Applied Microbiology and Biotechnology
91519ndash528
Midwest Plan Service 1987 Grain Drying Handling Storage Handbook Ames Iowa Iowa
State University 2nd edition
197
Miller SS Reid LM Harris LJ 2007 Colonization of maize silks by Fusarium graminearum
the causative organism of Gibberella ear rot Canadian Journal of Botany 85369-376
Minervini F Fornelli F Flynn KM 2004 Toxicity and apoptosis induced by mycotoxins
nivalenol deoxynivalenol and fumonisin B1 in a human erythroleukemia cell line
Toxicology In Vitro 1821ndash28
Mitterbauer R Poppenberger B RaditschnigA Lucyshyn D Lemmens M Glossl J Adam G
2004 Toxin-dependent utilization of engineered ribosomal protein L3 limits trichothecene
resistance in transgenic plants Plant Biotechnology Journal 2329ndash340
Mocali S Benedetti A 2010 Exploring research frontiers in microbiology the challenge of
metagenomics in soil microbiology Research Microbiology 161497-505
Morozkina EV Zvyagilskaya RA 2007 Nitrate reductases structure functions and effect of
stress factors Biochemistry 721151ndash1160
Munkvold GP 2003 Cultural and genetic approaches to managing mycotoxins in maize
Annual Review of Phytopathology 4199-116
Mundy R MacDonald TT Dougan G Frankel G Wiles S 2005 Citrobacter rodentium of
mice and man Cellular Microbiology 71697ndash1706
Nakamura N Inoue N Watanabe R Takahashi M Takeda J Stevens VL Kinoshita T 1997
Expression cloning of PIG-L a candidate N-acetylglucosaminyl-phosphatidylinositol
deacetylase The Journal of Biological Chemistry 27215834ndash15840
Nganje WE Bangsund DA Leistritz FL Wilson WW Tiapo NM 2002 Estimating the
economic impact of a crop disease the case of Fusarium head blight in US wheat and
barley National Fusarium Head Blight Forum Proceedings East Lansing Michigan State
University pp 275ndash281
198
Nielsen C Lippke H Didier A Dietrich R Martlbauer E 2009 Potential of deoxynivalenol
to induce transcription factors in humanhepatoma cells Molecular Nutrition and Food
Research 53479ndash491
Nielsen JKS Vikstroumlm AC Turner P Knudsen LE 2011 Deoxynivalenol transport across
the human placental barrier Food and Chemical Toxicology 492046-2052
Nishiuchi T Masuda D Nakashita H Ichimura K Shinozaki K Yoshida S et al 2006
Fusarium phytotoxin trichothecenes have an elicitor-like activity in Arabidopsis thaliana
but the activity differed significantly among their molecular species Molecular Plantndash
Microbe Interactions 19512ndash520
Ohsato S Ochiai-Fukuda T Nishiuchi T Takahashi-Ando N Koizumi S Hamamoto H
Kudo T Yamaguchi I Kimura M 2007 Transgenic rice plants expressing trichothecene
3-O-acetyltransferase show resistance to the Fusarium phytotoxin deoxynivalenol Plant
Cell Reports 26531-538
Okubara PA Blechl AE McCormick SP Alexander NJ Dill-Macky R Hohn TM 2002
Engineering deoxynivalenol metabolism in wheat through the expression of a fungal
trichothecene acetyltransferase gene Theoritical and Applied Genetics 10674-83
Ong CL Beatson SA Totsika M Forestier C McEwan AG Schembri MA 2010 Molecular
analysis of type 3 fimbrial genes from Escherichia coli Klebsiella and Citrobacter
species BMC Microbiology 10183
Osborne LE Stein JM 2007 Epidemiology of Fusarium head blight on small-grain cereals
International Journal of Food Microbiology 119103-108
Page RDM 1996 TREEVIEW An application to display phylogenetic trees on personal
computers Computer Applications in the Biosciences 12357-358
199
Parkhill J Dougan G James KD Thomson NR Pickard D Wain J Churcher C Mungall KL
Bentley SD Holden MT et al 2001 Complete genome sequence of a multiple drug
resistant Salmonella enterica serovar Typhi CT18 Nature 413848ndash852
Paterson RRM Lima N 2010 How will climate change affect mycotoxins in food Food
Research International 431902ndash14
Paul PA El-Allaf SM Lipps PE Madden LV 2004 Rain splash dispersal of Gibberella zeae
within wheat canopies in Ohio Phytopathology 941342-1349
Pestka JJ Lin WS Miller ER 1987 Emetic activity of the trichothecene 15-
acetyldeoxynivalenol in pigs Food and Chemical Toxicology 25855ndash858
Pestka JJ Smolinski AT 2005 Deoxynivalenol Toxicology and potential effects on humans
Journal of Toxicology Environmental Health 839ndash69
Pestka JJ 2007 Deoxynivalenol toxicity mechanisms and animal health risks Animal Feed
Science and Technology 137283-298
Pestka JJ 2008 Mechanisms of deoxynivalenol-induced gene expression and apoptosis Food
Additives and Contaminants 251128-1140
Pestka J2010 Toxicological mechanisms and potential health effects of deoxynivalenol and
nivalenol World Mycotoxin Journal 3323ndash347
Petersohn A Brigulla M Haas S Hoheisel JD Voumllker U Hecker M 2001 Global Analysis
of the General Stress Response of Bacillus subtilis Journal of Bacteriology 1835617-
5631
200
Petty NK Bulgin R Crepin VF Cerdeno-Tarraga AM Schroeder GN Quail MA Lennard N
Corton C Barron A Clark L Toribio AL Parkhill J Dougan G Frankel G Thomson NR
2010 The Citrobacter rodentium genome sequence reveals convergent evolution with
human pathogenic Escherichia coli Journal of Bacteriology 192525ndash538
Petty NK Feltwell T Pickard D Clare S Toribio AL et al 2011 Citrobacter rodentium is
an unstable pathogen showing evidence of significant genomic flux PLoS Pathology
7e1002018 doi101371journalppat1002018
Pittet A 2001 Natural occurrence of mycotoxins in foods and feeds A decade in review In
Mycotoxins and phytotoxins in perspective at the turn of the millennium edited by De
Koe WJ Samson RA Van Egmond HP Gilbert J Sabino M W J de Koe Wageningen
Netherlands pp 153ndash172
Placinta CM DrsquoMello JPF Macdonald AMC 1999 A review of worldwide contamination of
cereal grains and animal feed with Fusarium mycotoxins Animal Feed Science and
Technology 7821ndash37
Plank B Schuh M Binder EM 2009 Investigations on the effect of two feed additives
Biomin BBSH 797 and Mycofix Plus (R) 3E as detoxificants of DON contaminated feed
of piglets Wien Tierarztl Monatssch 9655ndash71
Poapolathep A Ohtsuka R Kiatipattanasakul W Ishigami N Nakayama H Doi K 2002
Nivalenol-induced apoptosis in thymus spleen and Peyerrsquos patches of mice Experimental
and Toxicological Pathology 53441ndash446
201
Poppenberger B Berthiller F Lucyshyn D Sieberer T Schuhmacher R Krska R Kuchler K
Gloumlssl J Luschnig C Adam G 2003 Detoxification of the Fusarium mycotoxin
deoxynivalenol by a UDP-glucosyltransferase from Arabidopsis thaliana Journal of
Biological Chemistry 27847905ndash47914
Prelusky DB Trenholm HL 1993 The efficacy of various classes of anti-emetics in
preventing deoxynivalenol-induced vomiting in swine Natural Toxins 1296ndash302
Proctor RH Hohn TM McCormick SP 1995 Reduced virulence of Gibberella zeae caused
by disruption of a trichothecene toxin biosynthetic gene Molecular Plant-Microbe
Interactions 8593-601
Puchenkova SG 1996 Enterobacteria in areas of water along the Crimean Coast
Mikrobiolohichnyĭ zhurnal 58 3-7
Qi BW Hao BL 2004 Whole-proteome prokaryote phylogeny without sequence alignment
a K-string composition approach Journal of Molecular Evolution 581ndash11
Quarta A Mita G Haidukowski M Logrieco A Mule G Visconti A 2006 Multiplex PCR
assay for the identification of nivalenol 3- and 15-acetyl-deoxynivalenol chemotypes in
Fusarium FEMS Microbiological Letters 2597ndash13
Rehr B Klemme JH 1989 Formate dependent nitrate and nitrite reduction to ammonia by
Citrobacter freundii and competition with denitrifying bacteria Antonie van Leeuwenhoek
Journal of Microbiology 56311ndash321
Rocha O Ansari K Doohan FM 2005 Effects of trichothecene mycotoxins on eukaryotic
cells A review Food Additives and Contaminants 22369ndash378
Roscoe V Lombaert GA Huzel V et al 2008 Mycotoxins in breakfast cereals from the
Canadian retail market a 3-year survey Food Additives and Contaminants 25347ndash355
202
Rotter BA Prelusky DB Pestka JJ 1996 Toxicology of deoxynivalenol (vomitoxin) Journal
of Toxicology and Environment Health 481ndash34
Ryu JC Ohtsubo K Izurniyarna N Nakamura K Tanaka T Yamarnura H Ueno Y 1988
The acute and chronic toxicities of nivalenol in mice Fundamental and Applied
Toxicology 1138ndash47
Saier MH Roseman S 1972 Inducer exclusion and repression of enzyme synthesis in
mutants of Salmonella typhimurium defective in enzyme I of the phosphoenolpyruvate
sugar phosphotransferase system The Journal of Biological Chemistry 247972ndash975
Saier MHJ Tran CV Barabote RD 2006 TCDB the transporter classification database for
membrane transport protein analyses and information Nucleic Acids Research 34D181ndash
186
Saitou N Nei M 1987 The neighbor-joining method a new method for reconstructing
phylogenetic trees Molecular Biology and Evolution 4406ndash425
Savelieva O Kotova I Roelofsen W Stams AJM Netrusov A 2004 Utilization of
aminoaromatic acids by a methanogenic enrichment culture and by a novel Citrobacter
freundii strain Archives of Microbiology 181163ndash170
Schaafsma AW Tamburic-Ilinic L Miller JD Hooker DC 2001 Agronomic considerations
for reducing deoxynivalenol in wheat grain Canadian Journal of Plant Pathology 23279ndash
285
Schaafsma AW 2002 Economic changes imposed by mycotoxins in food grains case study
of deoxynivalenol in winter wheat Advances in Experimental Medicine and Biology
504271ndash276
203
Scharf C Riethdorf S Ernst H Engelmann S Voumllker U Hecker M 1998 Theoredoxin is an
essential protein induced by heat shock and other stresses in Bacillus subtilis Journal of
Bacteriology 1801869-1877
Schatzmayr G Zehner F Taubel M Schatzmayr D Klimitsch A Loibner AP Binder EM
2006 Microbiologicals for deactivating mycotoxins Molecular Nutrition and Food
Research 50543-551
Schollenberger M Muumlller H-M Ruumlfle R Suchy S Plank S Drochner W 2006 Natural
occurrence of 16 Fusarium toxins in grains and feedstuffs of plant origin from Germany
Mycopathologia 16143ndash52
Scientific Committee for Food 2002 Opinion on Fusarium toxins Part 6 Group evaluation
of T-2 toxin HT-2 toxin nivalenol and deoxynivalenol (available at http
wwweuropaeuintcommfoodfsscscfout123_enpdf)
Schulman S Wang B Li W Rapoport TA 2010 Vitamin K epoxide reductase prefers ER
membrane-anchored thioredoxin-like redox partners Proceedings of the National
Academy of Sciences 10715027ndash15032
Scott PM 1997 Multi-year monitoring of Canadian grains and grain-based foods for
trichothecenes and zearalenone Food Additives and Contaminants 14333-339
Seitz LM Bechtel DB 1985 Chemical physical and microscopal studies of scab-infected
hard red winter wheat Journal of Agricultural and Food Chemistry 33373-7
Sergent T Parys M Garsou S Pussemier L Schneider YJ Larondelle Y 2006
Deoxynivalenol transport across human intestinal Caco-2 cells and its effects on cellular
metabolism at realistic intestinal concentrations Toxicological Letters 164167ndash176
204
Sharma J Fulekar MH 2009 Potential of Citrobacter freundii for bioaccumulation of
heavy metalndashcopper Biology and Medicine 17-14
Sherif SO Salama EE Abdel-Wahhab MA 2009 Mycotoxins and child health the need for
health risk assessment International Journal of Hygiene and Environmental Health
212347ndash368
Shifrin VI Anderson P 1999 Trichothecene mycotoxins trigger a ribotoxic stress response
that activates c-Jun N-terminal kinase and p38 mitogen-activated protein kinase and
induces apoptosis The Journal of Biological Chemistry 27413985ndash13992
Shima J Takase S Takahashi Y Iwai Y Fujimoto H Yamazaki M Ochi K 1997 Novel
detoxification of the trichothecene mycotoxin deoxynivalenol by a soil bacterium isolated
by enrichment culture Applied and Environmental Microbiology 633825ndash3830
Singh J Behal A Singla N Joshi A Birbian N et al 2009 Metagenomics Concept
methodology ecological inference and recent advances Biotechnology Journal 4480ndash
494
Smith TK McMillan EG Castillo JB 1997 Effect of feeding blends of Fusarium mycotoxin-
contaminated grains containing deoxynivalenol and fusaric acid on growth and feed
consumption of immature swine Journal of Animal Science 752184ndash2191
Smits WK Dubois JYF Bron S van Dijl JM Kuipers OP 2005 Tricksy business
transcriptome analysis reveals the involvement of thioredoxin in redox homeostasis
oxidative stress sulfur metabolism and cellular differentiation in Bacillus subtilis Journal
of Bacteriology 1873921ndash3930
205
Sprando RL Collins TFX Black TN Olejnik N Rorie JI Eppley RM Ruggles DI 2005
Characterization of the effect of deoxynivalenol on selected male reproductive endpoints
Food Chemistry and Toxicology 43623ndash635
Starkey DE Ward TJ Aoki T Gale LR Kistler HC Geiser DM Suga H Toacuteth B Varga J
ODonnell K 2007 Global molecular surveillance reveals novel Fusarium head blight
species and trichothecene toxin diversity Fungal Genetics and Biology 441191ndash1204
Stevenson BS Schmidt TM 1997 Growth rate-dependent expression of RNA from plasmid-
borne rRNA operons in Escherichia coli Journal of Bacteriology 1801970ndash1972
Stothard P Wishart DS 2005 Circular genome visualization and exploration using
CGView Bioinformatics 21537ndash539
Suenaga H Ohnuki T Miyazaki K 2007 Functional screening of a metagenomic library for
genes involved in microbial degradation of aromatic compounds Environmental
Microbiology 92289-97
Surai P Mezes M Fotina TI Denev SD 2010 Mycotoxins in Human Diet A Hidden
Danger In Modern Dietary Fat Intakes in Disease Promotion Edited by F De Meester et
al Nutrition and Health DOI 101007978-1-60327-571-2_18
Sutton JC 1982 Epidemiology of wheat head blight and maize ear rot caused by Fusarium
graminearum Canadian Journal of Plant Pathology 4195
Swaving J De Bont JAM Westphal A De Kok A 1996 A novel type pyridine nucleotide-
disulfide oxidoreductase is essential for NAD+- and NADPH-dependent degradation of
epoxyalkanes by Xanthobacter strain Py2 Journal of Bacteriology 1786644ndash6
206
Swanson SP Rood JHD Behrens JC Sanders PE 1987 Preparation and characterization of
the deepoxy trichothecenes deepoxy HT-2 deepoxy T-2 triol deepoxy T-2 tetraol
deepoxy 15-monoacetoxyscirpenol and deepoxy scirpentriol Applied and Environmental
Microbiology 532821ndash2826
Swanson SP Helaszek C Buck WB Rood JHD Haschek WM 1988 The role of intestinal
microflora in the metabolism of trichothecene mycotoxins Food Chemistry and
Toxicology 26823ndash829
Tian F Yu Y Chen B Li H Yao Y-F Guo X-K 2009 Bacterial archaeal and eukaryotic
diversity in Arctic sediment as revealed by 16S rRNA and 18S rRNA gene clone libraries
analysis Polar Biology 3293-103
Tatusov RL Galperin MY Natale DA Koonin EV et al 2000 Artemis sequence
visualization and annotation Bioinformatics 16944-945
Trail F Haixin X Loranger R Gadoury D 2002 Physiological and environmental aspects of
ascospore discharge in Gibberella zeae (anamorph Fusarium graminearum) Mycologia
94181-189
Tsuge K Ohata Y Shoda M 2001 Gene yerP involved in surfactin self-resistance in
Bacillus subtilis Antimicrobial Agents and Chemotherapy 453566ndash3573
Tucker 2001 The ldquoyellow rainrdquo controversy Lesson for arms control compliance The
Nonproliferation Review 825-42
Ueno Y 1985 The toxicology of mycotoxins Critical Reviews in Toxicology 1499-132
Ueno Y Nakayama K Ishii K Tashiro F Minoda Y Omori T Komagata K 1983
Metabolism of T-2 toxin in Curtobacterium sp strain 114-2 Applied and Environmental
Microbiology 46120ndash127
207
van Elsas JD Speksnijder AJ van Overbeek LS 2008 A procedure for the metagenomics
exploration of disease suppressive soils Journal of Microbiological Methods 75515ndash522
van Ginkel CG de Bont JAM 1986 Isolation and characterization of alkene-utilizing
Xanthobacter spp Archives of Microbiology 145403ndash407
van Lanen SG Shen B 2006 Microbial genomics for the improvement of natural product
discovery Current Opinion in Microbiology 9252ndash260
Varga J Toacuteth B 2005 Novel strategies to control mycotoxins in feeds a review Acta
Veterinaria Hungarica 53189-203
Voigt CA Schafer W Salomon S 2005 A secreted lipase of Fusarium graminearum is a
virulence factor required for infection of cereals The Plant Journal 42364ndash375
Vogelgsang S Hecker A Musa T Dorn B Forrer HR 2011 On farm experiments over five
years in a grain maizewinter wheat rotation Effect of maize residue treatments on
Fusarium graminearum infection and deoxynivalenol contamination in wheat In Proc
7th
Canadian Workshop on Fusarium Head Blight edited by J Gilbert A Tekauz N
Sweetland and K Slusarenko Winnipeg Manitoba Canada p 41
Voumllkl A Vogler B Schollenberger M Karlovsky P 2004 Microbial detoxification of
mycotoxin deoxynivalenol Journal of Basic Microbiology 44147ndash156
Wagacha JM Muthomi JW 2007 Fusarium culmorum Infection process mechanisms of
mycotoxin production and their role in pathogenesis in wheat Crop Protection 26877-
885
Wakulinski W 1989 Phytotoxicity of the secondary metabolites of fungi causing wheat head
fusariosis (head blight) Acta Physiologiae Plantarum 11301ndash306
208
Wang JT Chang SC Chen YC Luh KT 2000 Comparison of antimicrobial susceptibility of
Citrobacter freundii isolates in two different time periods The Journal of Microbiology
Immunology and Infection 33258-62
Wannemacher RJ Stanley L Wiener MD 2000 Trichothecenes mycotoxins In Medical
aspects of chemical and biological warfare edited by Zajtchuk R Washington DC
Department of the Army pp 656-676
Westphal AH Swaving J Jacobs L de Kok A 1998 Purification and characterization of a
flavoprotein involved in the degradation of epoxyalkanes by Xanthobacter Py2 European
Journal of Biochemistry 257160ndash168
Whalen JG Mully TW Enlgish JC 2007 Spontaneous Citrobacter freundii infection in an
immunocompetent patient Archives of Dermatology 143124-5
White 2007 The physiology and biochemistry of prokaryotes Third edition Oxford
University Press New York Oxford p 628
Wilcke WF Morey RV 1995 Natural-air corn drying in the upper Midwest University of
Minnesota Minnesota Extension Service Publication BU-6577-E pp 20
Windels CE 2000 Economic and social impacts of Fusarium headblight changing farms and
rural communities in the Northern Great Plains Phytopathology 9017ndash21
Winzeler EA Shoemaker DD Astromoff A Liang H Anderson K Andre B Bangham R
Benito R Boeke JD Bussey H et al 1999 Functional characterization of the S
cerevisiae genome by gene deletion and parallel analysis Science 285901ndash906
Yang GH Jarvis BB Chung YJ Pestka JJ 2000 Apoptosis induction by the satratoxins and
other trichothecene mycotoxins relationship to ERK p38 MAPK and SAPKJNK
activation Toxicology and Applied Pharmacology 164149ndash160
209
Yang Q Yin GM Guo YL Zhang DF Chen SJ Xu ML 2010 A major QTL for resistance
to Gibberella stalk rot in maize Theoretical and Applied Genetics 121673ndash687
Yazar S Omurtag GZ 2008 Fumonisins trichothecenes and zearalenone in cereals
International Journal of Molecular Science 92062ndash2090
Yoshizawa T Morooka N 1973 Deoxynivalenol and its monoacetate new mycotoxins from
Fusaruium roseum and moldy barley Agricultural and Biological Chemistry 372933ndash
2934
Yoshizawa T Takeda H Ohi T 1983 Structure of a novel metabolite from deoxynivalenol a
trichothecene mycotoxin in animals Agricultural and Biological Chemistry 472133ndash
2135
Yoshizawa T 1993 Red-mold diseases and natural occurrence in Japan In Trichothecenes-
Chemistry Biology and Toxicological Aspects edited by Y Ueno Elsevier Amsterdam
Netherlands p 195-209
Yoshizawa T Sakamoto T Okamoto K 1984 In vitro formation of 3rsquo-hydroxy T-2 and 3rsquo-
hydroxy HT-2 toxins from T-2 by liver homogenates from mice and monkeys Applied
and Environmental Microbiology 47130-134
Young JC Subryan LM Potts D McLaren ME Gobran FH 1986 Reduction in levels of
deoxynivalenol in contaminated wheat by chemical and physical treatment Journal of
Agricultural and Food Chemistry 34461ndash465
Young JC Trenholm HL Friend DW Prelusky DB 1987 Detoxification of deoxynivalenol
with sodium bisulfite and evaluation of the effects when pure mycotoxin or contaminated
corn was treated and given to pigs Journal of Agricultural and Food Chemistry 35259ndash
261
210
Young JC Zhu H Zhou T 2006 Degradation of trichothecene mycotoxins by aqueous
ozone Food Chemistry and Toxicology 44417ndash424
Young JC Zhou T Yu H Zhu H Gong J 2007 Degradation of trichothecene mycotoxins by
chicken intestinal microbes Food Chemistry and Toxicology 45136-143
Yu Z Morrison M 2004 Comparisons of different hypervariable regions of rrs genes for use
in fingerprinting of microbial communities by PCR-denaturing gradient gel
electrophoresis Applied and Environmental Microbiology 704800-4806
Yu H Zhou T Gong J Young C Su X Li X Zhu H Tsao R Yang R 2010 Isolation of
deoxynivalenol-transforming bacteria from the chicken intestines using the approach of
PCR-DGGE guided microbial selection BMC Microbiology 10182
Zeibdawi A Steele M Savoie A 2011 An overview of CFIAs mycotoxin monitoring
program in livestock feeds In Proc 7th
Canadian Workshop on Fusarium Head Blight
edited by J Gilbert A Tekauz N Sweetland and K Slusarenko Winnipeg Manitoba
Canada p 25
Zhang J-B Li H-P Dang F-J Qu B Xu Y-B Zhao C-S Lia Y-C 2007 Determination of the
trichothecene mycotoxins chemotypes and associated geographical distribution and
phylogenetic species of the Fusarium graminearum clade from China Mycological
Research 111967ndash975
Zhao JS Manno D Thiboutot S Ampleman G Hawari J 2007 Shewanella canadensis sp
nov and Shewanella atlantica sp nov manganese dioxide- and hexahydro-135-trinitro-
135-triazine-reducing psychrophilic marine bacteria International Journal of Systemic
Evolution and Microbiology 572155ndash2162
211
Zhao X-Q 2011 Genome-based studies of marine microorganisms to maximize the diversity
of natural products discovery for medical treatments Evidence-Based Complementary
and Alternative Medicine Article ID 384572 doi1011552011384572
ZhaoY Park RD Muzzarelli RA 2010 Chitin deacetylases properties and applications
Marine Drugs 824ndash46
Zhou HR Lau AS Pestka JJ 2003a Role of double-stranded RNA-activated protein kinase R
(PKR) in deoxynivalenol-induced ribotoxic stress response Toxicological Science
74335ndash344
Zhou HR Islam Z Pestka JJ 2003b Rapid sequential activation of mitogen-activated protein
kinases and transcription factors precedes proinflammatory cytokine mRNA expression in
spleen of mice exposed to the trichothecene vomitoxin Toxicological Sciences 72130ndash
142
Zhou T He J Gong J 2008 Microbial transformation of trichothecene mycotoxins World
Mycotoxin Journal 123-30
Zhou G Zhang H He Y He L 2010 Identification of a chitin deacetylase producing bacteria
isolated from soil and its fermentation optimization African Journal of Microbiology
Research 42597-2603

iv
ACKNOWLEDGEMENTS
Foremost I am deeply indebted to my advisors Prof K Peter Pauls and Dr Ting Zhou
for the continuous support of my PhD study for their endurance inspiration enthusiasm and
mentoring Their continued guidance has helped me in all aspects of my research and in
writing of this dissertation I sincerely express my gratitude to them for their co-operation
from the beginning to conclusion of the thesis and many other areas This will guide me in my
future professional career
My thesis committee guided me through all these years I would like to express my
gratitude to Prof Paul Goodwin for his insightful comments and advice on my thesis research
Special thanks to Dr Chris Young for teaching me chemistry chemical structures and
interpreting liquid chromatography ultraviolet mass spectra data I am sincerely grateful to
Dion Lepp for helping in analyzing the bacterial genome and sharing his knowledge and
valuable suggestions on writing the genomic chapter
I would like to express my appreciation to staff members of Dr Ting Zhou and Prof
K Peter Pauls who supported me in many aspects during my PhD study I am thankful to
Jianwei He Xiu-Zhen Li Honghui Zhu for collections and preparation of soil samples and
LC-UV-MS analyses of numerous samples Special thanks to Dr David Morris Johnston
Monje for sharing his knowledge discussions and camaraderie at coffee breaks
I wish to acknowledge the Natural Sciences and Engineering Research Council of
Canada (NSERC) for awarding me a prestigious scholarship for my PhD program and
Deanrsquos Tri-Council Scholarship (University of Guelph) which made my life easier during this
study I also would like to convey my thanks to Agriculture and Agri-Food CanadaBinational
v
Agricultural Research and Development Fund (AAFCBARD) project for providing me
financial support
Mike Bryan a very supportive person helped provide solutions to my software issues
that improved my exposure and caliber at conference awards competitions Several student
competition awards honored and inspired me to work hard and lead to the successful
completion of my thesis research I would like to convey my appreciation to the judges of
different student award committees who recognized my work They are Society for Industrial
Microbiology and Biotechnology conference 2010 (USA) Food Safety Research Forum-
Ontario Ministry of Agriculture and Rural Affairs annual meeting 2010 (Canada)
International Mycological Conference 2009 (UK) and Southwestern Ontario Regional
Association of Canadian Phytopathological Society annual meeting 2009-2011 (Canada)
I would like to express my very special thanks to Prof Golam Hafeez Kennedy my
brother-in-law who is like my elder brother In many aspects I am indebt to him for his
cooperations which aided me to achieve my current status
Lastly and most importantly I wish to give my special thanks to my wife Mimi and
lovely daughter Racy whose endless love sacrifice support and encouragement enabled me
to cross the tough long road and made possible to achieve my goal and aspirations Racy
once you told me that one of your desires for me is to finish my PhD which is done now I
know you want to be a scientist which is a great ambition I wish you all the best in your
future aims Racy
vi
TABLE OF CONTENTS
DEDICATIONShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipiii
ACKNOWLEDGEMENTShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipiv
TABLE OF CONTENTShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipvi
LIST OF TABLEShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipix
LIST OF FIGUREShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipx
LIST OF ABBREVIATIONShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipxiii
10 GENERAL INTRODUCTION AND LITERATURE REVIEW helliphelliphelliphelliphelliphelliphelliphellip1
11 Backgroundhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip1
12 Plant pathogenesis and trichothecene productionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip3
13 Biosynthesis of food contaminating TChelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip6
14 Occurrence of TC mycotoxins in food and feed helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip11
15 Significance of deoxynivalenol and trichotheceneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip16
16 Toxic effects of deoxynivalenol and trichotheceneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip19
17 Cellular and molecular mechanisms of DON toxicity in animals and plantshelliphelliphelliphellip22
18 Strategies to control Fusarium head blight and trichotheceneshelliphelliphelliphelliphelliphelliphelliphelliphellip26
19 Isolation of DONTC de-epoxidating micro-organisms and geneshelliphelliphelliphelliphelliphelliphelliphellip28
110 The bacterial species Citrobacter freundiihelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip30
111 Hypotheses and objectiveshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip31
vii
20 AEROBIC AND ANAEROBIC DE-EPOXIDATION OF MYCOTOXIN
DEOXYNIVALENOL BY BACTERIA ORIGINATING FROM AGRICULTURAL
SOIL33
21 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip33
22 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
23 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip37
24 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip45
25 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip62
26 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip64
30 ISOALTION AND CHARACTERIZATION OF A NOVEL SOIL BACTERIUM
CAPABLE OF DETOXIFYING ELEVEN TRICHOTHECENE MYCOTOXINS65
31 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip65
32 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip66
33 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip68
34 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip75
35 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip96
36 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip100
40 CONSTRUCTION AND FUNCTIONAL SCREENING OF GENOMIC DNA
LIBRARY FOR ISOLATION OF DON DE-EPOXYDIZING GENE(S) FROM
BACTERIAL STRAIN ADS47helliphelliphelliphelliphelliphelliphelliphellip102
41 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip102
42 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip103
43 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip104
viii
44 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip112
45 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip118
46 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip121
50 GENOME SEQUENCING CHARACTERIZATION AND COMPARATIVE
ANALYSIS OF A TRICHOTHECENE MYCOTOXIN DETOXIFYING
CITROBACTER FREUNDII STRAIN ADS47helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip122
51 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip122
52 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip123
53 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip125
54 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip129
55 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip162
56 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip168
60 CONCLUSIONhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip171
61 Summary helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip171
62 Research contributions helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip174
63 Future workhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip175
70 REFERENCEShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip176
ix
LIST OF TABLES
Table 1-1 Fusaria produced type-A and type-B trichothecene mycotoxinshelliphelliphelliphelliphelliphelliphellip9
Table 1-2 Worldwide occurrence of deoxynivalenol (DON) in animal feedshelliphelliphelliphelliphellip12
Table 1-3 Occurrence of DON in Canadian grains grains foods and beershelliphelliphelliphelliphelliphelliphellip13
Table 1-4 Legislative maximum tolerated levels of trichothecene mycotoxins in Canadahellip14
Table 1-5 In vitro and in vivo synergistic effects of different food contaminating
mycotoxins helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip18
Table 3-1 Microbial transformation of type-A and type-B tricthothecene mycotoxins
after 72 h of incubation under aerobic conditions and at room temperaturehelliphelliphelliphelliphelliphellip95
Table 4-1 Analysis of Fosmid DNA library for deoxynivalenol (DON) de-epoxidation
capability helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip117
Table 5-1 General features of C freundii strain ADS47 genomehelliphelliphelliphelliphelliphelliphelliphelliphellip132
Table 5-2 Genome comparisons between strain ADS47 and five Citrobacter species141
Table 5-3 Functional categorized gene abundances among the six Citrobacter genomeshellip145
Table 5-4 Unique genes of C freundii strain ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip156
Supplemental Table S5-1 List of unique genes of strain ADS47 gene IDs and their
functional roleshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip169
x
LIST OF FIGURES
Figure 1-1 Structures of trichothecene mycotoxins produced by Fusariahelliphelliphelliphelliphelliphelliphelliphellip7
Figure 1-2 Cellular and molecular mechanisms of deoxynivalenol toxicity in
eukaryotic organismshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip23
Figure 2-1 Mechanism of microbial DON de-epoxidationhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip36
Figure 2-2 LC-UV-MS chromatographic analysis of DON and its de-epoxidated product
dE-DON obtained by culturing with selected soil microbeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip46
Figure 2-3 Time course of relative biotransformation of DON to dE-DON by the initial
soil suspension culture in MSB mediumhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip49
Figure 2-4 Microbial de-epoxidation of DON after five days of incubationhelliphelliphelliphelliphelliphellip51
Figure 2-5 Analysis of microbial population reduction by terminal restriction fragment
length polymorphism (T-RFLP)helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip53
Figure 2-6 Microbial genera identified in the enriched DON de-epoxidizing microbial
culturehelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip56
Figure 2-7 Effects of culture conditions on DON to dE-DON transformation by the
enriched microbial culturehelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip58
Figure 2-8 Microbial DON to dE (de-epoxy)-DON transformation activity under aerobic
and anaerobic conditionshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip61
Figure 3-1 Colony color shape and size of the mycotoxin degrading bacterial strain
ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip76
Figure 3-2 Dendrogram based on 16S-rDNA sequences for strain ADS47 and 11
reference Citrobacter specieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip77
xi
Figure 3-3 Effects of DON concentration on bacterial growth and de-epoxidation
activitieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip79
Figure 3-4 Bacterial growth and DON de-epoxidation capabilities under aerobic and
anaerobic conditions helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip82
Figure 3-5 Effects of media on bacterial growth and DON de-epoxidation activityhellip85
Figure3-6 Effects of temperature on microbial growth and DON de-epoxydation helliphelliphellip87
Figure 3-7 Influence of pH on microbial growth and DON de-epoxydation capabilityhellip89
Figure 3-8 Effects of sodium azide (SA) on bacterial DON de-epoxidation activityhelliphelliphellip91
Figure 3-9 Determination of DON to deepoxy-DON transformation by bacterial culture
filtrate and cell extracthelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip92
Figure 3-10 Determination of microbial de-epoxidation and de-acetylation of a type-A
trichothecene mycotoxin (HT-2 toxin) by LC-UV-MShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip94
Supplemental Figure S3-1 Flow chart depicted the procedures of enrichment and isolation
of DON de-epoxidation bacterial strain ADS47helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip101
Figure 4-1 Schematic overview of the construction and functional screening of Fosmid
DNA libraryhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip105
Figure 4-2 Genetic map of copy controlled Fosmid vector (pCC1FOS)helliphelliphelliphelliphelliphelliphellip108
Figure 4-3 Preparation of 40 kb size insert DNA from a DON de-epoxidizing bacterium113
Figure 4-4 Production of genomic DNA library in E coli EPI300-T1 host strainhelliphelliphelliphellip114
Figure 4-5 Confirmation of the Fosmid DNA library construction in E coli helliphelliphelliphelliphellip 116
Figure 5-1 Co-linearity between C freundii strain ADS47 genome and C koseri helliphelliphellip129
Figure 5-2 Circular genome map of C freundii strain ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphellip133
xii
Figure 5-3 Size distribution of the predicted genes identified in C freundii strain
ADS47helliphelliphellip 135
Figure 5-4 Functional classification of the predicted coding sequences (CDS) of
C freundii strain ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip137
Figure 5-5 Phylogeny of the trichothecene detoxifying C freundii strain ADS47 139
Figure 5-6 Comparisons of coding sequences between C freundii strain ADS47 and
five Citrobacter specieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip143
Figure 5-7 Determination of the locus enterocyte effacement (LEE) operons in
Citrobacter species by comparing C rodentium helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip151
Figure 5-8 Locations of unique genes of C freundii strain ADS467helliphelliphelliphelliphelliphelliphelliphellip157
Figure 5-9 Unique genomic regions and associated genes in strain ADS47 compared
to five Citrobacter specieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip159
xiii
LIST OF ABBREVIATIONS
APCI= atmospheric pressure chemical ionization
AE= attaching and effacing
A= aerobic conditions
ABC= ATP-binding cassette
ACT= artemis comparative tools
AN= anaerobic conditions
3-ADON= 3-acetyl-deoxynivalenol
15-ADON= 15-acetyl-deoxynivalenol
ATA= alimentary toxic aleukia
bp= base pair
BGI= Beijing genomics institute
BLAST= basic local alignment search tool
BRIG= blast ring image generator
Bw= body weight
C= carbon
CDS= coding sequence
CFIA= Canadian food inspection agency
CHEF= clamped homogeneous electric fields
CMB= corn meal broth
CVTree= composition vector tree
DAS= diacetoxyscirpenol
dH2O= distilled water
DOGT1= UDP-glucosyltransferase
DON= deoxynivalenol
dsRNA= double stranded RNA
dE= de-epoxy
dE-DON= de-epoxy DON
E coli= Escherichia coli
ER=endoplasmic reticulum
FAO= Food amp Agriculture Organization
FM= full medium
FHB= Fusarium head blight
FUS= fusarenonX
gDNA= genomic DNA
GC= guanine cytosine
GER= Gebberella ear rot
GPI= glycosylphosphatidylinositol
h= hour
Hck= hematopoietic cell kinase
Kb= kilobase
xiv
Kg= kilogram
LC-UV-MS= liquid chromatography-ultraviolet-
mass spectrometry
LEE= locus of enterocyte effacement
LD50= lethal dose of 50 animal
LB= Luria Bertani
LMP= low melting point
MAPK= mitogen-activated protein kinase
microg= microgram
mg= milligram
mL= milliliter
microL= microliter
microM= micromole
min= minute
Mb= megabases
MCS=multiple cloning site
MSA= MSB+ agar
MSB= mineral salts broth
MM= minimal medium
MSC= mineral salts + 03 Bacto Peptone
MS-DON= mineral salts + 200 microgmL DON
mRNA= messenger RNA
MSY= mineral salts + yeast extract
mz= mass ion ratio
N= nitrogen
Na-Cl= sodiumndashclorine
NADH= (reduced) nicotinamide adenine
dinucleotide
NAD(P)= nicotinamide adenine
dinucleotide phosphate
NCBI= national center for biotechnology
information
NEO= neosolaniol
ng= nanogram
NIV= nivalenol
OD600nm= optical density at 600
nanometer wavelength
OMAFRA= Ontario Ministry of
Agriculture and Rural Affairs
Paa= phenylacetic acid
PCR= polymerase chain reaction
PDB= potato dextrose broth
ppm= parts per million
PFGE= pulsed field gel electrophoresis
PKR= protein kinase
xv
PNDO= pyridine nucleotide-disulfide
oxidoreductase
QTL= quantitative trait locus
RAST= rapid annotation subsystem technology
RDP= ribosomal database project
Rpl3= 60S ribosomal protein L3
rRNA= ribosomal RNA
s= second
SA= sodium azide
SDR= short chain dehydrogenasereductase
SE= soil extract
SIM= selected ion monitoring
SOAP= short oligonucleotide analysis package
sp= species
sRNA= small RNA
TC= trichothecene
TCA= tricarboxylic acid cycle
TSB= tryptic soy broth
TF= transcription factor
tRNA= transfer RNA
TIC= total ion count
TRAP= tripartite ATP-independent
periplasmic
T-RFLP=terminal restriction fragment
length polymorphism
TRI101= trichothecene 3-O-
acetyltransferase
YEB= yeast extract broth
VER= verrucarol
VKOR= vitamin K epoxide reductase
wv= weight by volume
1
CHAPTER 1
10 GENERAL INTRODUCTION AND LITERATURE REVIEW
11 Background
According to the Food and Agriculture Organization (FAO) of the United Nations
more than 25 of the worldrsquos food crops are contaminated with mycotoxins every year
causing serious adverse effects on livestock industry crop production and international trade
(Boutrif and Canet 1998 Desjardins 2006 Binder et al 2007 Surai et al 2010) The negative
effects of mycotoxins on the consumers are diverse including organ toxicity mutagenicity
carcinogenicity neurotoxicity teratogenicity modulation of the immune system reproductive
system effects and growth retardation Mycotoxins are toxic secondary metabolites produced
by various fungal species during crop colonization in field andor post harvest conditions
(FAO 1991 Barrett 2000) Aspergillus Penicillum and Fusarium are the predominant fungal
genera that accumulate economically important mycotoxins such as aflatoxins zearalenone
trichothecenes fumonisins ochratoxins and ergot alkaloids in agricultural commodities
Humans and animals are principally exposed to mycotoxins through consumption of
contaminated foodfeed and animal products such as milk meat and eggs (Fink-Gremmels
1989 Hollinger and Ekperigin 1999)
Trichothecenes (TCs) produced by a variety of toxigenic Fusarium species are
probably the most food and feed contaminating mycotoxins in the temperate regions of
America Europe and Asia (Creppy 2002 Yazar and Omurtag 2008) The fungal pathogen
accumulates various type-A (eg T-2 toxin HT-2 toxin) and type-B trichothecenes (eg
2
deoxynivalenol nivalenol and their derivatives) in cereals Among them type-B
trichothecenes particularly deoxynivalenol (DON) andor its derivatives appear to be the
most commonly found mycotoxins in grain and grain products worldwide impairing human
health and productivity of farm animals (Placinta et al 1999 Sherif et al 2009) A number of
human and animal disease outbreaks associated with TC contaminated grain food and feed
consumption have been reported in Asia (Japan India and central Asia) Europe New
Zealand and United States of America (USA) (Pestka and Smolinski 2005 Desjardins 2006)
A severe outbreak of Alimentary Toxic Aleukia (ATA) mycotoxicosis killed 100000 people
in the Orenburg region of Russia (Eriksen 2003) In 1996 losses of over $100 million were
estimated in Ontario because of costs related to managing DON (Schaafsma 2002)
Historically T-2 toxin DON and nivalenol (NIV) have likely been used as a biological
warfare in Laos Kampuchea and Afghanistan and there is concern because of the potential
use of these compounds for bioterrorism (Wannemacher et al 2000 Tucker 2001)
Despite the extensive efforts that have been made to prevent TC contamination with
the current varieties and production practices its presence in agricultural commodities is
likely unavoidable There are no crop cultivars currently available that are completely
resistant to Fusarium head blightGibberella ear rot (FHBGER) diseases Chemical or
physical methods cannot safely remove TC mycotoxins from grain completely or they are not
economically viable (Varga and Toacuteth 2005) Therefore an alternative strategy to address the
issue is of great importance (Awad et al 2010 Karlovsky 2011)
3
12 Plant pathogenesis and trichothecene production
Cereals infested by the ascomyceteous fungi may cause Gibberella ear rot (GER) in
maize (Zea mays L) and Fusarium head blight (FHB also known as lsquoscabrsquo) of small grain
cereals [wheat (Triticum aestivum L) barley (Hordeum vulgare L)] and other small grain
cereals (Goswami and Kistler 2004) As many as 17 fungal species have been reported to be
associated with FHBGER which include F graminearum Schwabe [telemorph Gibberella
zeae (Schwein) Petch] F culmorum (WG Smith) Sacc F sporotrichioides Scherb F
poae (Peck) Wollenw Microdochium nivale (Fr) Samuels amp IC Hallett and F avenaceum
(Fr) Sacc (Osborne and Stein 2007 Audenaert et al 2009) The two species F
graminearum and F culmorum are the principal species causing crop damage Among them
F graminearum is the widely adapted and most damaging species across the world including
North America (Dyer et al 2006) F culmorum tend to be the main species in the cooler parts
of North Central and Western Europe (Wagacha and Muthomi 2007)
The fungi overwinter mainly on crop residues as saprophytes where they produce
asexual (micro- and macro-conidia) andor sexual (ascospores) fruiting structures which act
as the initial source of inoculums in the subsequent crop season (Sutton 1982 Wagacha and
Muthomi 2007) Ascospores are primarily dispersed by air (Trail et al 2002) and conidia
move by rain-splash (Paul et al 2004) After they land on maize silks and florets of small
grain cereals infection occurs through natural openings (eg stomata) or occasionally by
direct penetration The pathogens can invade maize kernels through wounds caused in the
husks by insects or birds Infection may occur throughout the crop growing season but
disease incidence and severity is facilitated by warm temperatures (gt20degC) high humidity
(gt80) and frequent precipitation that coincides with anthesis (Osborne and Stein 2007
4
Wagacha and Muthomi 2007) The Fusarium pathogen initially infects as biotroph which
becomes a necrotroph as the disease progresses After penetration Fusaria produce different
extracellular cell wall degrading enzymes that trigger infection and hyphal progression
towards the developing earkernel where further colonization occurs and development of
FHBGER diseases (Voigt et al 2005 Miller et al 2007 Kikot et al 2009) FHB disease is
characterized by a bleached appearance around the middle of the head Severely infected
grains have a pinkish discoloration and can be shriveled In maize the disease produces a
reddishndashbrown discoloration of the kernels and progresses from the tip to the bottom of
affected ears
The fungi associated with FHB and GER contaminate cereals by accumulating various
host-nonspecific TC mycotoxins (Brennen et al 2007) TCs have typically been associated
with cereals grown in temperate climatic conditions including Canada (Doohan et al 2003)
DON production by F graminearum and F culmorum is maximal at warm temperatures (22-
28˚C) high grain moisture contents (gt17) and humid conditions (gt70 RH) (Martins and
Martins 2002 Hope et al 2005 Llorens et al 2006) Grain can be contaminated with TCs in
the field as well as in storage if conditions for fungal growth are optimal and grain contains
high moisture (gt17) (Larsen et al 2004 Birzele et al 2002)
Populations of FHBGER causing pathogens are dynamic Genetic variability between
and within Fusarium species appears to have occurred to allow for their adaptation to new
agro-climates (Desjardins 2006 Fernando et al 2011) Three strain specific Fusarium
chemotypes NIV 3-ADON (3-acetyl-deoxynivalenol) and 15-ADON (15- acetyl-
deoxynivalenol) have been identified which correlate with the geographical distributions
(Starkey et al 2007 Zhang et al 2007 Audenaert et al 2009) The NIV chemotype mainly
5
produced by F culmorum accumulates NIV and 4-NIV (Fusarinon X) is predominant in
Asia Africa and Europe (Lee et al 2001) The 3-ADON chemotype which synthesizes DON
and 3-ADON is the dominant strain in EuropeFar East (Larsen et al 2004) In North
America 15-ADON is the main chemotype which produces DON and 15-ADON (Goswami
and Kistler 2004) Significant Fusarium population shifting was observed in the past and in
recent years (Kant et al 2011) A dynamic change from F culmorum to F gramninearum
and DON to NIV producing chemotyes occurred in North West European countries The 3-
ADON chemotype which is uncommon in North America was found in North Dakota and
Minnesota (Goswami and Kistler 2004) F pseudograminearum which historically occurs in
Australia Africa and North West Pacific America has been found in Western Canada (Larsen
et al 2004 Kant et al 2011) In Manitoba a replacement of 15-ADON chemotype by the
more phytoxic 3-ADON type was observed in recent years (Fernando et al 2011) All
together the FHBGER associated pathogen complex is highly dynamic which not only
reduces crop yield but also decreases grain quality by accumulating various mycotoxins
6
13 Biosynthesis of food contaminating TC
TCs are fungal secondary metabolites that were initially identified in Trichotheceium
roseum Thus the name of this class of compounds follows the name of the genus
Trichotheceium (Freeman and Morrison 1949) There have been 180 TCs identified in various
fungal species belonging to the genera Fusarium Myrothecium Stachybotrys Trichoderma
and Thrichothecium (Eriksen 2003 Eriksen et al 2004) The FHBGER associated TCs are
mainly synthesized by different Fusaria and are characterized as tricyclic sesquiterpenoids
with a double bond at position C9-10 and a C12-13 epoxide ring
7
Figure 1-1 Structures of trichothecene mycotoxins produced by Fusaria (a) General
structure of trichothecenes R1 = H OH OiValeryl keto R2 = H OH R3= OH OAc R4 =
H OH OAc R5= OH OAc (b) Type-A trichothecene mycotoxin (c) Type-B trichothecene
mycotoxin which contain a carbonyl functional group at C8 position (Young et al 2007)
(a) (b)
(c)
8
The fungal metabolites are small in size (molecular weight 250-550) heat stable and
non-volatile molecules (Pestka 2007 Yazar and Omurtag 2008) Food contaminating TCs are
divided into two groups type-A and type-B (Figure 1-1b and 1c) Type-A is characterized by
the lack of a ketone function at C8 position and include T-2 toxin HT-2 toxin
diacetoxyscirpenol (DAS) and neosolaniol (NEO) which are mainly produced by F
sporotrichioides and F poae (Creppy 2002 Quarta et al 2006) The most frequently found
TC belongs to type-B which contains carbonyl functions at C8 positions DON NIV and
their acetylated derivatives The two types are further differentiated by the presence or
absence of functional hydroxyl (-OH) and acetyl (CH3-CO-) groups at C-3 C-4 C-7 C-8 and
C-15 positions (Table 1-1)
The syntheses of various TCs are speciesstrain specific and are genetically controlled
but influenced by environmental conditions (Desjardins 2006 Paterson and Lima 2010) Until
now 15 Fusaria catalytic genes and regulators involved in TC productions have been
identified (Merhej et al 2011) These biosynthetic genes are located at three loci on different
chromosomes The core trichothecene (TRI) gene cluster is comprised of 12 genes localized
in a 25-kb genomic region (Kimura et al 2001) The remaining three genes are encoded by
the TRI1-TRI16 and TRI101 loci (Gale et al 2005 Alexander et al 2009) Fusaria produce
TC following a number of oxygenation isomerization acetylation hydroxylation and
esterification reaction steps (Desjardins 2006 Desjardins and Proctor 2007 Kimura et al
2007 Merhej et al 2011) In brief farnesyl pyrophosphate is initially cyclized into
trichodiene by TRI5 which codes for trichodiene synthase In the next four steps
isotrichotriol is produced by TRI4 (a multifunctional cytochrome P450) via C-2 hydroxylation
and 1213 epoxidation followed by two hydroxylation reactions
9
Table 1-1 Fusaria produced type-A and type-B trichothecene mycotoxins Molecular weight
(MW) and acetyl (-R) hydroxyl (-OH) and ketone (=O) functions at different carbon (C)
position of the tricyclic rings A-types T-2 toxin HT-2 toxin DAS (diacetoxyscirpenol)
NEO (neosolaniol) VER (verrucarol) B-type DON (deoxynivalenol) 3-ADON (3-acetyl-
deoxynivalenol) 15-ADON (15-acetyl-deoxynivalenol) NIV (nivalenol) and FusX
(fusarenon X) (Young et al 2007)
Trichothecenes MW R1C3 R2C4 R4C7 R3C15 R5C8
Type-A
T-2 toxin 466 OH OCOCH3 H OCOCH3 OCOCH2CH(CH3)2
HT-2 toxin 424 OH OH H OCOCH3 OCOCH2CH(CH3)2
DAS 366 OH OCOCH3 H OCOCH3 H
NEO 382 OH OCOCH3 OH OCOCH3 H
VER 266 H OH H OH H
Type-B
DON 296 OH H OH OH =O
3-ADON 338 OCOCH3 H OH OH =O
15-ADON 338 OH H OH OCOCH3 =O
NIV 312 OH OH OH OH =O
FusX 354 OH OCOCH3 OH OH =O
10
The next two non-enzymatic reactions results in isotrichodemol Eventually the
trichothecene skeleton is produced via the formation of a C-O bond between the C-2 oxygen
and C-11 Subsequent acetylation at C-3 by TRI101 hydroxylation at C-15 by TRI1 and
catalytic activity of TRI3 leads to calonectrin production The pathway described above is
common for both type-A (eg T-2 toxin HT-2 toxin) and type-B (eg DON NIV and their
acetylated derivatives) TCs
The following steps are speciesstrain specific In DON producing F graminearum
strains calonectrin is converted to either 3-ADON or 15-ADON followed by DON production
by the catabolic activities of TRI1 and TRI8 The biosynthesis of 3-ADON or 15-ADON is
controlled by variation in the esterase coding sequences of TRI8 F graminearum produces
NIV from calonectrin via synthesis of 4-NIV followed by NIV by TRI1 and TRI8 gene
actions NIV can be produced either from DON by hydroxylation at C-4 or via
transformation of calonectrin to 315-diacetoxyscirpenol (315-DAS) followed by addition of
a ketone at C-8 position by the activity of the TRI7 and TRI13 genes 315-DAS is the
principal substrate for synthesis of type-A TCs by F sporotrichioides the main type-A
producer The biosynthetic pathway of T-2 toxin involves C-4 acetylation of 315-DAS by the
TRI7 gene product and TRI8 triggered by TRI1 315-DAS is also the substrate for HT-2
toxin synthesis In the absence of TRI7 expression HT-2 toxin is produced through
hydroxylation of 315-DAS and isovalerate addition to C-8 position (Foroud and Eudes 2009)
11
14 Occurrence of TC mycotoxins in food and feed
TC producing Fusaria are adapted to variable climatic conditions and agricultural
ecosystems As a consequence this class of mycotoxins particularly DON is frequently
detected in foods and feeds including breakfast cereals bakery products snack foods beer
and animal feed derived from cereals in most part of the world (Pittet 2001 Desjardins 2006
Lancova et al 2008) A two-year survey during 2003-2005 conducted by Romer Lab Inc
revealed unacceptable levels of DON in animal feedsfeed raw materials (such as maize
wheat wheat bran) in Asia and Oceania (1062 microgg) central Europe (802 microgg) Mid-West
USA (55 microgg) northern Europe (551 microgg) and southern Europe (303 microgg) (Binder et al
2007) (Table 1-2)
Quite high levels of DON contamination (19 microgg) were detected in north Asian and
Chinese feedfeed materials Recently five-year on-farm studies in Switzerland showed that
74 of the wheat grain and straw samples were contaminated with high levels of DON
(average 5 microgg) The amount is higher than the European maximum limit of 125 microgg
(Vogelgsang et al 2011)
The occurrences of TCs in Canadian agricultural commodities are indicated in Table
1-3 Seventeen surveys conducted during the 1980s and 1990s showed up to 100 DON
incidence in Ontario wheat and maize samples with an 18 average incidence (Scott 1997) A
three-year survey recently (2006-08) conducted by the Ontario Ministry of Agriculture and
Rural Affairs (OMAFRA) detected high levels of DON in spring (maximum 26 microgg) and
winter wheat (maximum 29 microgg) samples (Ana Matu personal communication) The DON
levels were much higher than Health Canadarsquos maximum limit of 2 microgg for uncleaned soft
wheat for staple foods (Table 1-4)
12
Table 1-2 Worldwide occurrence of DON in animal feed Feed and feed raw material
samples were collected from animal farmsfeed production sites during 2003-2005 The
survey was conducted by Romer Labs Inc (httpwwwromerlabscom)
Country
region
Sample
type
Incidence
()
Maximum
DON content
(microgg)
Mean DON
content
(microgg)
Survey
period
Source
North
AsiaChina
Animal
feed
071 1899 092
Binder et
al 2007
South East
Asia
014 152 016
South Asia 070 1062 049
Oceania 2003-05
Australia 041 186 030
Northern
Europe
070 551 059
Central
Europe
066 802 057
Southern
Europe
052 303 030
13
Table 1-3 Occurrence of DON in Canadian grains grain foods and beers Infant cereal foods
oat barley soy and multi-grain based foods
Province Sample
type
Incidence
()
Maximum
DON
contents
(microgg)
Multiple
mycotoxins
samples ()
Survey
period
Sources
Ontario Spring
Wheat
70 260 Yes 2006-08 OMAFRA1
Ontario Winter
wheat
49 290 Yes 2006-08 OMAFRA
Ontario Maize 78 140 Yes 2009-10 Limay-Rios
and
Schaafsma
2010
Ontario Maize - 1000 Yes 2011 AampL
Biologicals
Canada Breakfast
cereals
46 094 430 1999-01 Roscoe et al
2008
Canada Animal
feeds
73 425 Yes 1990-10 Zeibdawi et
al 2011
Canada Beers 58 0052 50 1990s Scott 1997
Canada Infant
Cereals
63 098 Yes 1997-99 Lombaert et
al 2003
1
Ontario Ministry of Agriculture and Rural Affairs
2 microgmL
14
Table 1-4 Legislative maximum tolerated levels of trichothecene mycotoxins in Canada
according to Canadian Food Inspection Fact Sheet April 23 2009 DON- deoxynivalenol
DAS- diacetoxyscirpenol
Products DON
(microgg)
T-2 toxin
(microgg)
HT-2 toxin
(microgg)
DAS
(microgg)
Uncleaned soft wheat for human
consumption
2 - - -
Cattlepoultry diets 5 lt1 01 lt1
Swinedairy cattle feeds 1 lt1 0025 lt2
15
High DON incidence (78) was observed in maize samples collected from
southwestern Ontario fields with moderate levels (lt1 microgg) of contamination during 2009 and
2010 (Limay-Rios and Schaafsma 2010) Some samples had high DON levels (14 microgg) In
the 2011 crop season extremely high levels of DON (gt100 microgg) were found in some
southern Ontario maize samples (AampL Biologicals London ON Personal communication)
In a Canadian Food Inspection Agency (CFIA) 20 year survey (1990-2010) 73 of
animal feed samples collected from feed mills farms and retail stores across Canada were
contaminated with DON and for more than 25 of the samples the mycotoxin levels were
above the maximum limit (ge 1 microgg) (Zeibdawi et al 2011) The highest level of DON (15
microgg) detected in the 2006-2007 samples were much lower than the levels found in the 1999-
2000 samples which contained a maximum of 425 microgg DON Importantly this study
reported a significant number of samples contaminated with multiple mycotoxins DON NIV
3-ADON 15-ADON T-2 toxin HT-2 toxin DAS and NEO
Earlier studies in the 1990s detected high levels of DON content in Eastern Canadian
cereals maize (maximum 175 microgg) wheat (maximum 916 microgg) barley (maximum 911
microgg) and oats (maximum 12 microgg) (Campbell et al 2002) The study identified a significant
number of samples (15-367) that contained multiple mycotoxins and up to 33 of the
wheat samples exceeded 1 microgg DON Low amounts but high incidences of TCs were found
in cereal-based foods collected from Canadian retail stores Sixty three percent of infant
cereal foods 46 of breakfast cereals and 58 of beers were contaminated with one or
multiple mycotoxins (Scott 1997 Lombaert et al 2003 Roscoe et al 2008) The maximum
amount of DON was found in infant foods (098 microgg) followed by breakfast cereals (094
microgg) and beers (005 microgmL)
16
15 Significance of deoxynivalenol and trichothecenes
For most cereals including wheat maize rice barley oats and rye the risk of
contamination with TCs resulted in grain quality reduction and FHBGER disease progression
(Bai et al 2002 Jansen et al 2005) The impacts of TC contamination in agricultural
commodities are diverse including reduced livestock production induced human illnesses
and enhanced FHB disease severity which may result in the reduction of crop yield Exposure
of farm animals (particularly pigs) to DON is a continuing threat to the livestock industries in
Canada USA and Europe (DrsquoMello et al 1999) Ingestion of TC-contaminated feeds
generally causes reduction of feed intake feed refusal poor feed conversion retardation of
weight gain increased disease susceptibility and reduced reproductive capacities (Binder et al
2007) High doses of DON result in acute toxic effects in pigs (eg emesis) while low doses
(ge1 microgmL) cause chronic effects (eg growth retardation in pigs) (Rotter et al 1996
Accensi et al 2006) Reduction of growth and reproduction ability was observed in
experimental rats exposured to much higher doses of TC (Sprando et al 2005 Collins et al
2006) Several animal disease outbreaks that occurred in different continents have been
attributed to consumption of feed contaminated with DON NIV and DAS (Desjardins 2006)
The consequences of human exposure to TCs include liver damage gastricintestinal
lesions and central nervous system toxicity resulting in anorexia nausea hypotensionshock
vomiting diarrhea and death (Pestka 2007) Human mycotoxicosis (illnesses and outbreaks)
by consumption of type-A and type-B TC mycotoxins in Europe New Zealand Russia
Japan South America India and Vietnam have been reported (Bhat et al 1989 Yoshizawa
1993 Luo 1994 Desjardins 2006) Severe outbreaks of Alimentary Toxic Aleukia (ATA)
associated with TC killed thousands of people in the Orenburg region of Russia during the
17
1930rsquos and 1940rsquos (Eriksen 2003) TCs (T-2 DON and NIV) were likely used for biological
warfare in Laos Kampuchea and Afghanistan and its potential application for bioterrorism is
an additional concern associated with TC (Tucker 2001 Bennett and Klich 2003)
TCs are phytotoxic and may contribute to FHB disease severity in cereals (Proctor et
al 1995 Maier et al 2006 Masuda et al 2007) Studies demonstrated that DON caused
wilting chlorosis inhibition of seedling growth and necrosis in cereals (Cossette and Miller
1995 Mitterbauer et al 2004 Rocha et al 2005) DON-mediated host cell damage and
subsequent necrotrophic fungal spread has been shown to lead to FHBGER disease
development and higher kernel damage (Procter et al 1995 Mitterbauer et al 2004 Maier et
al 2005) This suggests that DON contributes to yield losses as well as to reductions in the
market value of cereals (Osborne and Stein 2007) Severely infected grain may contain more
than 60 microgg DON which is unacceptable to the milling and brewing industries (Okubara et
al 2002) Significant crop losses due to FHB disease epidemics and mycotoxin contamination
have been reported in Ontario (Schaafsma 2002) FHB disease epidemics during 1998 to 2000
caused huge crop losses resulting in farm failures in the northern Great Plains in the USA
(Windels 2000) In that period FHB associated losses were estimated at US $ 27 billion in
the region (Nganje et al 2002)
Importantly Fusaria infested grains are often co-contaminated with multiple
mycotoxins which further increase health risks as interactions between the mycotoxins may
cause greater toxic effects than expected (Schollenberger et al 2006) This implies that an
apparently low level of contamination of a single mycotoxin can have severe effects on
consumers because of synergistic effects among multiple co-occuring mycotoxins (Surai et al
2010)
18
A number of studies demonstrated synergistic toxic effects among Fusarium
mycotoxins on pigs turkeys lamb chicks as well as in in vitro Caco-2 cell and yeast
bioassays (Table 1-5)
Table 1-5 In vitro and in vivo synergistic effects of different food contaminating mycotoxins
DON-deoxynivalenol DAS-diacetoxyscirpenol FB1-fumonisins 1 NIV-nivalenol ZEN-
zearelenone FA-fusaric acid Afl-aflatoxin
Mycotoxins source Interacting
mycotoxins
Experimental
animalcells
Effects Reference
Naturally contaminated
cereals
DON FA Pigs Inhibit growth Smith et al
1997
DON contaminated
wheat and FB1
DON FB1 Pigs Weight loss Harvey et al
1996
Culture materials
contained mycotoxins
T-2 FB1 Turkey poults Weight loss oral
lesion
Kubena et al
1997
Inoculated rice and
pure mycotoxin
DAS Afl Lamb Weight reduction Harvey et al
1995
Naturally DON
contaminated wheat
and pure T-2 toxin
DON T-2 Chicks Reduced body
weight significantly
Kubena et al
1989
Pure mycotoxins DAS Afl Chicks Reduced body
weight
Kubena et al
1993
Pure mycotoxins T-2 Afl Chicks Reduced weight
oral lesion
Kubena et al
1990
Pure mycotoxins DON
ZENFB1
Human intestinal
Caco-2 cells
Increased DNA
fragmentations
Kouadio et al
2007
Pure mycotoxins DON NIV Yeast bioassay Synergistic effects Madhyastha et
al 1994
Pure mycotoxins T-2 HT-2 Yeast bioassay Synergistic effects Madhyastha et
al 1994
19
16 Toxic effects of deoxynivalnol and trichothecenes
As small molecules TCs simply enter into cells and interact with different cell
components resulting in disruption of cellular functions and causing various toxic effects that
include inhibition of protein synthesis and ribosomal fragmentation (Eudes et al 2000
Minervini et al 2004 Pestka 2007) Simple absorption of DON into human intestinal Caco-2
cells and crossing human placenta barriers through passive diffusion has been reported
(Sergent et al 2006 Nielsen et al 2011)
TC may cause various toxicities in animals and plants including apoptosis
immunotoxicity chromatin condensation disruption of cellularmitochondrion membranes
disruption of gut tissues inhibition of protein and ribosome synthesis breakage of ribosome
and cell damage The toxicity of TC is influenced by structural variability (ie position
number and type of functional groups attached to the tricyclic nucleus and C-C double bond at
C9-10 position) (Desjardins 2006) Nevertheless epoxide function at the C12-13 position is
the key for TC cytotoxicity (Swanson et al 1988 Eriksen et al 2004)
Different in vitro and in vivo studies revealed that the method and period of exposure
amount of TC and types of celltissue and host species have significant effects on TCrsquos
cytotoxicity (Scientific Committee for Food 2002 Rocha et al 2005 Nielsen et al 2009)
Administration of 10 mgkg body weight (bw) NIV caused apoptosis in mice thymus Peyerrsquos
patch and spleen tissues (Poapolathep et al 2002) DON and NIV at 887 mgkg bw for 3 days
a week for four weeks induced immunotoxicity in experimental mice determined by
estimating increased immunoglobulins (IgA) and decreased IgM levels (Gouze et al 2007)
Exposure of mice to 1 mgkg bw of DON induced skeletal malformations and reduced mating
tendency while significant resorption of embryos observed at the dose of gt10 mgkg bw
20
(Khera et al 1982) At high acute doses (4972 mgkg bw) DONNIV and type-A
trichothecenes caused mice death because of intestinal haemorrhage necrosis of bone
marrowlymphoid tissues and kidneyheart lesions (Yoshizawa and Morooka 1973
Yoshizawa et al 1994 Forsell et al 1987 Ryu et al 1988) In a separate study 15-ADON
showed more toxicity than DON when the mycotoxins were orally administered to mice
(Forsell et al 1987)
Studies suggested that type-A TCs are more toxic in mammals than type-B which
means that functional variability at the C-8 position of TC has a significant role in toxicity
The toxicity of various TC mycotoxins in animals can be ranked in the order of T-2
toxingtHT-2 toxingtNIVgtDONge3-ADONgt15-ADON according to the results from human cell
lines and mice model systems bioassays (Ueno et al 1983 Pestka et al 1987 Nielsen et al
2009) Moreover animal feeding assays demonstrated that monogastric animals are most
sensitive to DON The order of sensitivity of animals to DON is
pigsgtrodentsgtdogsgtcatsgtpoultrygtruminants (Pestka 2005) The reasons for the variations in
DON toxicity species might be due to differences in metabolism absorption distribution and
elimination of the mycotoxin among animal species (Pestka 2007)
To my knowledge the mechanism of TC uptake by plant cells has not been studied
Nonetheless cereals are exposed to TC by toxigenic Fusaria infection which may cause
various toxicities such as cell damage wilting chlorosis necrosis program cell death and
inhibition of seedling growth in sensitive plants (eg wheat maize and barley) (Cossette and
Miller 1995 Rocha et al 2005 Boddu et al 2007) As animals TC inhibits protein synthesis
in non-host A thaliana (Nishiuchi et al 2006) Immunogold assays showed that the cellular
targets of DON in wheat and maize were cell membrane ribosome endoplasmic reticulum
21
vacuoles and mitochondria Modulation of cell membrane integrity and electrolyte losses
were observed by direct DON interference in maize and wheat cell assays (Cossette and
Miller 1995 Mitterbauer et al 2004) DON-mediated mitochondrion membrane alterations
disruption of host cells enzymatic activity apoptosis and necrosis have been observed in
cereals (Mitterbauer et al 2004 Rocha et al 2005) In contrast to animal type-B is more toxic
to plants than type-A DON showed 8- and 13-fold phytotoxicity to wheat coleoptiles than T-
2 and HT-2 toxins respectively (Eudes et al 2000) By compilation of different studies TC
phytoxicity can be ordered as DONge3-ADONgtT-2 toxingtDAS (Wakulinski 1989 McLean
1996 Eudes et al 2000)
22
17 Cellular and molecular mechanisms of DON toxicity in animals and plants
The underlying cellular and molecular mechanisms of DONTC toxicity are evolving
Exposure to DON proceeds with the up and down regulation of numerous critical genes of
eukaryotic organisms which causes various negative effects on animal and plant organisms
(Figure 1-2)
One of the early events of DON toxicity occurs when the mycotoxin binds to the
active ribosome resulting in blocking protein translation through different routes by
damaging the ribosome (Shifrin and Anderson 1999) degrading or altering conformational
changes of 28S-rRNA (Alisi et al 2008 Li and Pestka 2008) and inducing expression of
miRNAs This results in down regulation of the ribosome associated genes (He et al 2010a
Pestka 2010) Both animal and plant cell assays demonstrated that DON bound to the peptidyl
transferase (Rpl3) domain of the 60S subunit of ribosome and blocked protein chain
elongation and termination at high DON concentrations while peptide chain termination and
ribosome breakage occurred at a low level of DON exposure (Zhou et al 2003a)
23
Figure 1-2 Cellular and molecular mechanisms of deoxynivalenol toxicity in eukaryotic
organisms DON is simply absorbed by the cell and interacts with the cytoplasmic membrane
(CM) mitochondrion (MT) membrane and ribosome Disruption of the CM and alteration of
the MT membrane causes a loss of electrolytes and affects the function of the MT
respectively DON binds to the active ribosomes which leads to ribosome damage and
induces micro RNAs (miRNA) Expression of miRNA impairs the synthesis of ribosome
associated protein The subsequent transduction and phosphorylation of the RNA-activated
protein kinase (PKR) and hematopoietic cell kinase (Hck) activate the mitogen-activated
protein kinase (MAPK) cascade Phosphorylation of the MAPKs induces targeted
transcription factors (TF) which results in ribotoxis stress response upregulation of
proinflammatory cytokines and dysregulation of neuroendocrine signaling (derived from
Pestka 2007 and 2010)
24
The consequence of DON interaction with ribosomes leads to cellular and molecular
changes which are complex and not fully understood However it is well-demonstrated that
the activation of the mitogen-activated protein kinase (MAPK) and subsequent molecular
actions play key roles in the cytotoxicity of TC Upon binding to the ribosome DON rapidly
induced interactions between the protein kinases and the ribosomal 60S subunit as observed in
mononuclear phagocytes (Bae and Pestka 2008) Ribosomal binding to DON activated the
ribosome associated kinases for instances protein kinase R (PKR) hematopoietic cell kinase
(Hck) and P38
mitogen-activated protein kinase The protein kinases are induced by double-
stranded RNA (dsRNA) or possibly by the damaged ribosomal peptides (Iordanov et al 1997
Shifrin and Anderson 1999 Zhou et al 2003b Pestka 2007) Subsequent phosphorylation of
kinases (eg PKRHckP38
) activates the MAPK cascade resulting in ribotoxis stress
response upregulation of proinflammatory cytokines and dysregulation of neuroendocrine
signaling (Pestka and Smolinki 2005 Pestka 2010) Studies suggested that high doses of
DON induces ribotoxis stress response (apoptosis) preceded by immunosuppression and
disruption of intestinal permeabilitytissue functions (Yang et al 2000 Pestka 2008) Further
study observed stabilization of the mRNA and expression of transcription factors for critical
genes when cells were exposed to a low concentration of DON (Chung et al 2003)
The MAPK pathway proceeds with upregulation of a broad range of proinflammatory
cytokine genes and dysregulation of neuroendocrine signaling within the central and enteric
nervous systems Modulation of neuroendocrine signaling by DON increased the level of
serotonin resulted in vomiting reduced food intake and growth retardation in experimental
animalscell lines (Prelusky and Trenholm 1993 Li 2007) Upregulation of proinflammatory
cytokines and chemokines genes was determined in mice liver spleen kidney and lung
25
following in 1 h of exposure to DON A subsequent shock-like effect was observed in mice
because of a ldquocytokine stormrdquo (Pestka 2008) Induction of proinflammatory cytokines also led
to various cellular effects on the central nervous system interference of the innate immune
system and growth hormone signaling These contribute to anorexia growth retardation
aberrant immune response and tissue dysfunction as demonstrated in experimental animal
systems (Pestka 2010)
DON-mediated growth retardation in mice via suppression of growth hormones
(insulin-like growth factor 1 and insulin-like growth factor acid-labile subunit) has been
reported by Amuzie et al (2009) Disruption of gut tissues by DONNIV and subsequent
impairment of intestinal transporter activity was demonstrated in a human epithelial cell
model (Marseca et al 2002) The study proposed that alteration of gut tissue permeability
may cause gastroenteritis induction and growth retardation in livestock animals Growth
retardation of livestock may also occur via dysregulation of neuroendocrine signaling-
mediated induction of emesis and anorexia (Pestka et al 2010)
The molecular mechanism of TC toxicity in plant systems has not been well
documented A recent study showed the upregulation of numerous defense related genes in
wheat upon exposure to DON (Desmond et al 2008) The study further suggested that DON-
induced program cell death in wheat occurred through a hydrogen peroxide signaling
pathway This mechanism had not been observed in mammals
26
18 Strategies to control FHB and trichothecenes
During the past decades many pre- and post-harvest strategies have been applied to
protect cereal crops from Fusarium infestation and to eliminate TC from contaminated
agricultural commodities (FAO 2001 Munkvold 2003 Varga and Toth 2005 Kabak and
Dobson 2009) Some of the above crop protection techniques have been patented (He and
Zhou 2010) In spite of those efforts TC contamination of cereals appears to be unavoidable
using the currently available methods
Pre-harvest control strategies involving cultural practices (eg crop rotation tillage
early planting) and fungicide applications partially reduced Fusarium infection and DON
contamination in cereals (Dill-Macky and Jones 2000 Schaafsma et al 2001) However
repeated application of fungicides may lead to the evolution of Fusarium strains resistant to
fungicides with greater mycotoxin production ability (DrsquoMello et al 1998) At post-harvest
reduction of grain moisture by artificial drying storage at low temperatures (1-4degC) and
proper ventilation of the storage room have shown to be effective in inhibiting fungal growth
and mycotoxin production in infected grain (Midwest Plan Service 1987 Wilcke and Morey
1995) This technology has become less applicable particularly in developing countries
because of the frequent observation required to maintain proper storage conditions (Hell et al
2000)
Removal of Fusarium infected grains by physical methods (eg mechanical
separation density segregation colour sorting etc) was unable to eliminate TC completely
(Abbas et al 1985 He et al 2010b) This is partly because FHB symptomless kernels may
contain a considerable amount of DON which are not eliminated by physical separation
procedures (Seitz and Bechtel 1985) Several studies showed that milling and food processing
27
steps were unable to completely remove DON from grain-based foods (Jackson and
Bullerman 1999 Placinta et al 1999 Kushiro 2008) Detoxification of TC by chemical
treatments (eg ozone ammonia chlorine hydrogen peroxide and sodium metabisulfite) has
been demonstrated (Young et al 1986 Young et al 1987 Young et al 2006) The problem
with these techniques is that the application of such chemicals is not economically viable and
it also has a negative impact on grain quality (Karlovsky 2011) Although the technique has
been popular and showed effectiveness to certain mycotoxins utilization of absorbents to
inhibit mycotoxin absorption by the livestock gastrointestinal tract appeared to be ineffective
for TC (Huwig et al 2001)
The introduction of Fusarium resistant cultivars would be the best strategy to address
the FHB and associated mycotoxin problems in cereals However breeding for cultivars
resistant to FHBGER is difficult and time consuming as natural resistance against Fusarium
is quantitative in nature FHB resistant quantitative trait loci (QTLs) have been identified in
several cereal genotypes (eg Sumai 3 Ning 7840 Bussard) (Bernando et al 2007 Golkari
et al 2009 Haumlberle et al 2009 Yang et al 2010) The 3B 5A and 6B QTLs for type-I
(resistant to initial infection) and type-II (resistant to fungal spread in the kernel) resistances
have been identified on different wheat chromosomes (Buerstmayr et al 2009) Type-I
(Qfhsifa-5A) and type-II (Fhb1Fhb2-3B) QTLs from Sumai 3 are being used in many
breeding programs and pyramiding of the QTLs in spring wheat showed additional effects
Nonetheless the key genes involved in FHB resistance in cereals are yet to be identified This
makes it difficult to integrate the resistance genes into elite cultivars
28
Cultivar development by direct incorporation of TCDON resistance genes may be an
alternative way of protecting grains from these mycotoxins contaminations Many genes from
distantly related species have been integrated into crop cultivars using a transgenic approach
with great economic benefits (Brookes and Barfoot 2006) DON transforming trichothecene
3-O-acetyltransferase (TRI101 from Fusarium sporotrichioides) and UDP-
glucosyltransferase (from Arabidopsis thaliana) genes expressed in cereal cultivars showed
some DONDAS detoxifying ability under in vitro and controlled conditions (Kimura et al
1998 Okubara et al 2002 Poppenberger et al 2003 Ohsato et al 2007) By contrast no
reduction of mycotoxins in either of the above cases was observed under field conditions
(Manoharan et al 2006) Importantly the transformed acetylated and glycosylated
compounds could revert to the original substances or convert to even more toxic products
(Gareis et al 1990 Alexander 2008)
19 Isolation of DONTC de-epoxidating micro-organisms and genes
An enzyme with the capability of de-epoxidizing DONTC appears to be a potential
source for completely removing TC from contaminated cereals (Karlovsky 2011) However
such trichothecene detoxifying genes do not exist in cereals and other plants (Boutigny et al
2008) Because of this several research groups have attempted to obtain microorganisms that
encode DONTC de-epoxidation genes Mixed microbial culture capable of de-epoxydizing
trichothecenes have been isolated from chicken gut (He et al 1992) cow ruminal fluid (King
et al 1984 Swanson et al 1987 Fuchs et al 2002) pig gut (Kollarczik et al 1994) and
digesta of chicken (Young et al 2007) rats (Yoshizawa et al 1983) sheep (Westlake et al
1989) and fish (Guan et al 2009) All of the microbial cultures cited above showed de-
29
epoxydation capabilities only under anaerobic conditions realtively high temperatures and at
lowhigh pH except microbes from fish intestine that showed efficient de-epoxidation at low
temperatures and wide range of temperatures (Guan et al 2009) Such enzymes active under
anaerobic conditions high temperatues and low pH may have potential applications as feed
additives and the eubacterial strain BBSH 797 is an animal feed additive product with the
brand name Mycofixreg by BIOMIN (He et al 2010b) Biotechnological applications of
anaerobically-active enzymes from the above microbial sources seem to be inappropriate
under the aerobic and moderate conditions at which cereals plants are grown
Most of these efforts have failed to isolate single DONTC degrading microbes
effective under aerobic conditions because of inappropriate microbial growth conditions
under laboratory conditions Earlier attempts were made to obtain TC de-epoxidizing aerobic
bacteria but the culture did not retain DONNIV de-epoxidation activity after reculturing
(Ueno 1983 He et al 1992) In addition the co-metabolic nature of the microbial TC de-
epoxidation reaction has limited the use of mycotoxins as the sole nutrient source for selective
isolation of the target microorganisms Several DONTC de-epoxidation microbial strains
including Bacillus strain LS100 and Eubacterial strain BBSH 797 have been isolated from
chicken gut and rumen fluids through multi-step enrichment procedures (Fuchs et al 2002
Yu et al 2010) A DON to 3-keto DON transforming bactial strain was islated from soil by
prolonged incubation with high concentration DON (Shima et al 1997)
Microbial genesenzymes with TC de-epoxidation activity are not known Based on
the microbial TC de-epoxidation activity of a mixed microbial culture it has been proposed
that ceratin reductases or oxidoreductases may catalyze such a biochemical reaction (Islam et
al 2012) With the advent of molecular techniques many novel genes with
30
industrialagricultural value have been cloned by reverse genetics (genomic DNA library
preparation and functional screening of the library clones) forward genetics (transposon
mutagenesis) and comparative genomics methods (Frey et al 1998 Koonin et al 2001
Suenaga et al 2007 Chung et al 2008 Ansorge 2009) The tremendous advancement of next
generation sequencing technology and bioinformatics tools may facilitate the identification of
the microbial genes responsible for DONTC detoxification (Ansorge 2009) Isolation of TC
de-epoxidation genes may play a major role in producing safe foods and feeds derived from
grains
110 The bacterial species Citrobacter freundii
Citrobacter freundii is a bacterial species belonging to the family Enterobacteriace It
is a facultatively anaerobic and Gram-negative bacillus (Wang et al 2000) The bacterium
survives almost everywhere including animalhuman intestines blood water and soils
(Wang et al 2000 Whalen et al 2007) C freundii is known as an opportunistic human
pathogen and causes a variety of respiratory tract urinary tract and blood infections and
responsible for serious neonatal meningistis disease (Badger et al 1999 Whalen et al 2007)
Interestingly this bacterium plays a positive environmental role through nitrogen recycling
(Puchenkova 1996) Strains of C freundii also contribute to the environment by sequenstering
heavy metals and degrading xenobiotic compounds (Kumar et al 1999 Sharma and Fulekar
2009)
31
111 Hypotheses and objectives
The study was based on the following hypotheses
i Certain microorganisms in DON contaminated agricultural soils are able to transform DON
to de-epoxy DON (de-epoxidation) under aerobic conditions and moderate temperatures
ii DON de-epoxidation may be achieved by a single microbial strain
iii Microbial DON detoxifying genes and associated regulators are able to express and
function in a closely related host organism
iv DON detoxifying microbial strains carry genes for de-epoxidation activity that are absent
from related species that have not evolved under DON selection pressure
The objectives of this research were to
i Select microbial cultures from soil having DON de-epoxidation activity under aerobic
conditions and moderate temperatures by analyzing potential DON contaminated soils in
various growth conditions namely aerobicanaerobic conditions different growth media
temperature and pH
ii Select for organisms with DON de-epoxidation activity from the mixed microbial culture
by treating with antibiotics heating subculturing on agar medium and incubating for a
prolonged period in media with a high DON concentration
iii Identify microbial de-epoxidation genes by producing a microbial genomic DNA library
from the DON de-epoxidizing strain followed by screening of the clones for DON de-
epoxidation
32
iv Determine microbial de-epoxidation genes by sequencing of the genome and comparative
analysis of the genome with closely related species and identification of putative DON
detoxification genes
33
CHAPTER 2
20 AEROBIC AND ANAEROBIC DE-EPOXIDATION OF MYCOTOXIN
DEOXYNIVALENOL BY BACTERIA ORIGINATING FROM AGRICULTURAL
SOIL
Published in World Journal of Microbiology and Biotechnology 2012 287-13
21 Abstract
Soil samples collected from different cereal crop fields in southern Ontario Canada
were screened to obtain microorganisms capable of transforming deoxynivalenol (DON) to
de-epoxy DON (dE-DON) Microbial DON to dE-DON transformation (ie de-epoxidation)
was monitored by using liquid chromatography-ultraviolet-mass spectrometry (LC-UV-MS)
The effects of growth substrates temperature pH incubation time and aerobic versus
anaerobic conditions on the ability of the microbes to de-epoxidize DON were evaluated A
soil microbial culture showed 100 DON to dE-DON biotransformation in mineral salts
broth (MSB) after 144 h of incubation Treatments of the culture with selective antibiotics
followed an elevated temperature (50ordmC) for 15 h and subculturing on agar medium
considerably reduced the microbial diversity Partial 16S-rRNA gene sequence analysis of the
bacteria in the enriched culture indicated the presence of at least Serratia Citrobacter
Clostridium Enterococcus Stenotrophomonas and Streptomyces The enriched culture
completely de-epoxidized DON after 60 h of incubation Bacterial de-epoxidation of DON
occurred at pH 60 to 75 and a wide array of temperatures (12 to 40ordmC) The culture showed
34
rapid de-epoxidation activity under aerobic conditions compared to anaerobic conditions This
is the first report on microbial DON to dE-DON transformation under aerobic conditions and
moderate temperatures The culture could be used to detoxify DON contaminated feed and
might be a potential source for gene(s) for DON de-epoxidation
Keywords biotransformation de-epoxidation deoxynivalenol soil bacteria vomitoxin
22 Introduction
Deoxynivalenol (DON commonly known as lsquovomitoxinrsquo) 3α 7α 15-trihydroxy-
1213-epoxytrichothec-9-en-8-one is a trichothecene produced by toxigenic Fusarium species
causing Fusarium head blight (FHB) or Gibberella ear rot (GER) disease on small grain
cereals and maize respectively (Starkey et al 2007 Foroud and Eudes 2009) DON
contamination in cereals and maize is widespread and can lead to tremendous losses to the
growers It has also detrimental effects on human and livestock health (Pestka and Smolinski
2005 Rocha et al 2005) The toxin interferes with the eukaryotic ribosomal peptidyl
transferase (Rpl3) activity resulting in human and animal mycotoxicosis (Ueno 1985 Bae
and Pestka 2008) Common effects of DON toxicity in animals include diarrhea vomiting
feed refusal growth retardation immunosuppression reduced ovarian function and
reproductive disorders (Pestka and Smolinski 2005 Pestka 2007) In plants DON causes a
variety of effects viz inhibition of seed germination seedling growth green plant
regeneration root and coleoptile growth (cited in Rocha et al 2005) In addition DON
enhances disease severity in sensitive host plants (Maier et al 2006)
35
Avoidance of DON contamination or reduction of toxin levels in contaminated grains
remains a challenging task Physical and chemical processes are either inappropriate or
prohibitively expensive to use to decrease the DON content of agricultural commodities
(Kushiro 2008 He et al 2010b) The use of Fusarium resistant crop varieties might be the
best method to reduce DON contamination However breeding for FHB or GER resistant
cultivars appears to be very difficult and time consuming as natural sources of resistance
against Fusarium-caused diseases is quantitative in nature (Ali et al 2005 Buerstmayr et al
2009) As an alternative expressing genes for enzymatic detoxification of DON in elite
cultivars may be an effective strategy for protecting the value of feeds and foods (Karlovsky
1999) Heterologous expression of eukaryotic trichothecene 3-O-acetyltransferase and UDP-
glucosyltransferase genes in cereals and Arabidopsis have resulted in reduced toxicity via
acetylation and glycosylation of the DON (Poppenberger et al 2003 Ohsato et al 2007)
It has been proposed that enzymatic de-epoxidation of trichothecene toxins could be
used to limit DON contamination (Zhou et al 2008 Foroud and Eudes 2009) DON belongs
to type B-trichothecenes which contain an epoxide ring at C12-13 and the epoxide is highly
reactive and mainly responsible for toxicity (Figure 2-1) (Swanson et al 1988 Eriksen et al
2004)
36
Figure 2-1 Mechanism of microbial DON de-epoxidation Removal of the oxygen atom from
the epoxide ring results in formation of a C=C double bond at C-12 position which reduces
cytotoxicity (Swanson et al 1988 Eriksen et al 2004)
Microbial DON de-epoxidation by single bacterial strains and by mixed cultures has
been reported (King et al 1984 Swanson et al 1987 He et al 1992 Kollarczik et al 1994
Fuchs et al 2002 Young et al 2007 Guan et al 2009 Yu et al 2010) However all of these
microbes originated from fish and other animals and were strictly anaerobic The recovered
microbial cultures and single strains only showed de-epoxidation of trichothecenes at low
(le15ordmC) and high (gt30degC) temperatures Such enzymes might have limited applications for
toxin decontamination in cereal grains The aim of the current study was to isolate the
microbes from soil capable of transforming DON to a de-epoxy metabolite under aerobic
conditions and moderate temperatures
De-epoxydation
Deoxynivalenol (DON)
De-epoxyDeoxynivalenol (dE-DON)
37
23 Materials and methods
231 Soil samples
The upper layer (0-30 cm) of soils along with crop debris were randomly collected
from cereal (eg maize wheat barley) crop fields during 2004-2006 in Southern Ontario
Canada Detailed of the crops and locations of the fields have been described in Jianwei Hersquos
Master Degree thesis (He 2007) Briefly soil samples were collected in sterile plastic bags
and transported to the laboratory Samples were sieved to remove large roots stones and
stored at 4ordmC until used The samples were mixed with grounded mouldy corn (containing 95
microgg DON) + F graminearum and incubated at 22-24degC for six weeks Five samples were
prepared for DON de-epoxidation activity analysis by dispersing 5 g soil in 50 mL distilled
water and storing at4degC until further use
232 Culture media
Media used in the study were nutrient broth (NB) Luria Bertani (LB) minimal
medium (MM) (He 2007) six mineral salts + 05 bacto peptone (MSB) (He 2007) tryptic
soy broth (TSB) corn meal broth (CMB) (Guan et al 2009) potato dextrose broth (PDB) six
mineral salts + 05 yeast extract broth (MSY) and soil extract (SE) MM was composed of
sucrose-100 g K2HPO4-25 g KH2PO4-25 g (NH4)2HPO4-10 g MgSO47H2O-02 g
FeSO4-001 g MnSO47H2O- 0007 g water- 985 mL and MSB was composed of the same
salts as MM but was supplemented with Bactotrade Peptone (05 wv) as the sole C source
instead of sucrose To prepare soil extract 100 g of the composite soil was homogenized in 1
L water The soil particles and plant debris were removed by passing the soil suspension
through Whatmanreg grade 3 filter paper (Sigma-Aldrich Oakville Ontario Canada) The
38
extract was autoclaved at 121degC for 30 min Media and ingredients were purchased either
from Fisher Scientific Inc (Fair Lawn New Jersey USA) Sigma-Aldrich (Oakvile Ontario
Canada) or Difco (Sparks Maryland USA) Prior to autoclaving pH of the broth media was
adjusted to 70 Solid media were prepared by adding 15 agar
233 Screening of soil samples
Frozen soil suspensions were thawed and vortexed to homogenize the microbial
population An aliquot (100 microL) of the suspension was immediately transferred to MSB PDB
and MM broth media DON at 50 microgmL was added to the microbial suspension cultures The
cultures were incubated at 22-24degC with continuous shaking at 180 revolutions per minute
(rpm) The extent of DON de-epoxidation was determined at 24 h 48 h 72 h 96 h 120 h and
144 h of incubation
234 Determination of de-epoxidation product
Microbial DON de-epoxidation was measured according to Guan et al (2009)
Samples were prepared by mixing equal volume of microbial culture and methanol and
allowed to stand for 30 min at room temperature Microorganisms in the mixture were
removed by passing them through a 045 microm Millipore filter (Whatmanreg Florham Park New
Jersey USA) The filtrates were used for de-epoxidation analysis by a liquid chromatography-
ultraviolet-mass spectrometry (LC-UV-MS) Identification and quantification of DON and its
transformation product was achieved using a Finnigan LCQ DECA ion trap LC-MS
instrument (ThermoFinnigan San Jose CA USA) The equipment detects 30-2000 mz
(molecular mass ion ratio) compounds and able to determine 001 micromL DON The HPLC
39
system was equipped with a Finnigan Spectra System UV6000LP ultraviolet (UV) detector
and an Agilent Zorbax Eclipse XDB C18 column (46times150 mm 35 μm) The binary mobile
phase consisted of solvent A (methanol) and solvent B (water) The system was run with a
gradient program 25 A at 0 min 25ndash41 A over 5 min isocratically at 41 A for 2 min
and 41ndash25 A over 1 min There was a 3 min post-run at its starting condition to allow for
the equilibration of the column The flow-rate was set at 10 mLmin UV absorbance was
measured at 218 nm Atmospheric pressure chemical ionization (APCI) positive ion mode
was used for MS detection A DON standard was used to adjust the instrument to its
maximum response The system was operated as follows shear gas and auxiliary flow-rates
were set at 95 and 43 (arbitrary units) voltages on the capillary tube lens offset multipole 1
offset multipole 2 offset and entrance lens were set at 3000 minus1700 minus370 minus850 minus3500
and minus4200 V respectively capillary and vaporizer temperatures were set at 200 and 450degC
respectively and the discharge needle current was set at 7 μA Identification of mycotoxin
was achieved by comparing their retention times UV spectra and mass spectrum with DON
and de-epoxy DON (Biopure Union MO USA) as standards Quantification of DON and
product was based on peak areas related to their UV and MS calibration curves
40
235 Biological DON de-epoxidation
To help identify the types of microorganisms causing the DON de-epoxidation MSB
medium was supplemented with either 100 microgmL penicillin G or streptomycin sulfate for
suppressing gram positive or negative bacterial growth and activities respectively Three
replications of antibiotic-treated media were inoculated with 100 microL of microbial culture
capable of de-epoxidizing DON The antibiotic-treated cultures were incubated at 27degC
spiked with 50 microgmL DON Controls were prepared without adding antibiotics in the
microbial culture Inhibition of DON de-epoxidation was analyzed after five day of
incubation
236 Reduction of microbial diversity by antibiotic treatments heating and
subculturing on agar medium
In order to reduce non-DON degrading microbial populations the DON de-
epoxidation capable cultures were sequentially treated with antibiotics having diverse modes
of action including i) dichloramphenicol that kills various gram positive Gram-negative and
anaerobic bacteria ii) trimethoprim that kills Gram-positive and negative bacteria iii)
penicillin that is lethal to Gram-positive bacteria and iv) cyclohexamide to kill filamentous
fungi and yeasts Antibiotics were purchased from Sigma-Aldrich Antibiotic stock solutions
(10 mgmL) were prepared with sterile water or methanol Microbial cultures in MSB
medium were treated with 100 microgmL of each antibiotic followed by incubation of the culture
at 50ordmC for 15 hours to kill undesirded microbial species After heating cultures were treated
with 50 microgmL DON and incubated at 27ordmC for five days DON de-epoxidation activity of the
microbial cultures was determined following the procedure described above Cultures that
41
showed gt99 de-epoxidation were considered for subsequent antibiotic and heat treatments
Aliquots of the achieved culture were spread on MSB agar plates and grown at room
temperature for few days Colonies appeared on the plates were collected and incubated in
MSB medium supplemented with 50 microgmL DON De-epoxidation ability of the subcultured
microbial cultures was determined after five days of incubation at 27degC by LC-UV-MS
method
237 Analysis of microbial population diversity
Terminal Restriction Fragment Length Polymorphism (T-RFLP) analysis
Initially microbial diversity was determined by terminal restriction fragment length
polymorphism (T-RFLP) fingerprint analysis (Bruce and Hughes 2000) Genomic DNA from
microbial cultures with and without enrichment was extracted using DNeasy Plant DNA
Extraction Mini Kit (Qiagen Inc Mississauga Ontario Canada) 16S-rDNA was amplified
by PCR using the HEX-labeled forward primer Bac8f
(5-agagtttgatcctggctcag-3) and
unlabeled reverse primer Univ1492r (5-ggttaccttgttacgactt-3) (Fierer and Jackson 2006)
Fifty microlitter PCR mixture for each sample was prepared having 1x HotStarTaq Master
Mix (Qiagen) 05 microM of each primer 50 microg of BSA and 50 ng of DNA The 35 PCR cycles
followed 60 s at 94degC 30 s at 50degC and 60 s at 72degC After size verification by agarose-gel
electrophoresis 50 ng of purified PCR products was digested for 3 h in a 20 microL reaction
mixture with 5 U of HhaI restriction enzyme (New England Biolabs Ipswich MA USA)
The size and relative abundance of the labeled T-RFLP fragments were separated by capillary
electrophoresis in an automated DNA sequencer equipped with fluorescence detector
(Applied Biosystem 3730 DNA Analyzer) The DNA fragments were analyzed with the Peak
42
Scannertrade Software v10 to create an electropherogram with the genetic fingerprints of the
microbial community
Analysis of V3-V5 region of the 16S-rDNA
To define the bacterial genera present in the enrichment culture the 16S-rDNA was
amplified using two sets of bacterial universal primers Primarily 16S-rDNA was PCR
amplified using the primers 46f-5rsquogcytaacacatgcaagtcga3rsquo (Kaplan and Kitts 2004) and
1371r-5rsquogtgtgtacaagncccgggaa3rsquo (Zhao et al 2007) PCR was performed in a total volume of
25 μL which contained 06 μM of each primer 1x of HotStarTaqreg Master Mix (Qiagen Inc)
and 20-40 ng of DNA The PCR cycles included the following steps 95ordmC (15 min) 35 cycles
of 94ordmC (20 s) 52ordmC (15 s) 72ordmC (1 min 30 s) and a 72ordmC (7 min) extension step The second
pair of universal bacterial primers were 27f-5rsquoagagtttgatcmtggctcag3rsquoand 1492r-
5rsquoggttaccttgttacgactt3rsquo (Lane 1991) PCR was conducted with 25 cycles following the
procedures of Tian et al (2009) Amplification was carried out in a GeneAmpreg PCR System
9700 (Applied Biosystems) Furthermore the PCR products were used as templates to
amplify the hypervariable V3-V5 region of the 16S-rDNA using primers 357f-
5rsquocctacgggaggcagcag3rsquo and 907r7 5rsquoccgtcaattcctttgagttt3rsquo (Yu and Morrison 2004)
After purification of the PCR products by QIAquick PCR purification kit two
amplicon libraries were prepared by ligating to pCR-TOPO vector (Invitrogen) and the library
clones were transformed into TOPO10 competent cells according to Invitrogenrsquos TOPO
cloning manual The clone libraries were plated on LB plates containing 25 μgmL
kanamycin Fifty colonies from each library were randomly picked up from the overnight
plates and inserts were sequenced as described by Geng et al (2006) PCR products were
43
sequenced with an automated ABI 3730 DNA analyzer (Applied Biosystems) The sequence
similarities of the amplified V3-V5 region were compared with the ribosomal database project
(RDP) (httprdpcmemsuedu) Sequences with greater than 95 identity were assigned to
the same bacterial genus
238 Factors influencing DON de-epoxidation
DON de-epoxidation ability of the enriched microbial culture was investigated under
different conditions as described below
Maintaince of active culture
To retain microbial de-epoxidation activity the active culture has been maintained in
MSB medium containing 50 microgmL DON
Media growth temperature and pH
Overnight microbial cultures (100 μL) were added to 2 mL of NB LB MM TSB
MSB CMB MSY PDB or SE broths The pH of the media was adjusted to 70 and the
experiments were conducted at 27degC temperature The effect of temperature was studied by
incubating the culture in MSB medium (pH 7) at 12 20 25 30 and 40ordmC To determine the
effects of pH MSB medium pH was adjusted to 55 60 65 70 75 and 80 Media with
different pHs were inoculated with aliquots of the overnight active culture and then incubated
at 27ordmC The cultures were supplemented with 50 μgmL DON before incubation Three
replications for each medium temperature and pH condition were evaluated
44
De-epoxidation under aerobic and anaerobic conditions
Three replicate microbial cultures of five mL each were grown in sterile 15 mL
centrifuge tubes under aerobic conditions and in an anaerobic chamber filled with 95 CO2
and 5 H2 (COY Laboratory Products Inc Ann Arbor MI USA) Three days after initiation
half (25 mL) of the cultures from anaerobic conditions was transferred to new centrifuge
tubes and brought to aerobic conditions Similarly half of the aerobically grown cultures were
placed into an anaerobic chamber Cultures moved from anaerobic to aerobic conditions were
shaken after opening the lids of the tubes in a laminar flow every eight hours intervals to
maintain aerobic conditions in the culture Lids of the culture tubes transferred to anaerobic
conditions were kept loose to allow oxygen to escape from the culture Two days after
transferring to the new conditions the cultures in both conditions were treated with 50 microgmL
DON and incubated at 27degC Every 12 h 500 microL were removed from each tube and analyzed
for microbial DON de-epoxidation activity by LC-UV-MS
239 Data analysis
Data were processed using Microsoft Excel Program 2007 Statistical analysis was
done using General Linear Model procedures in SAS version 91 (SAS Institute Inc Cary
NC USA 2002-03) The results were the average of three replications stated as mean plusmn
standard error Differences between means were considered significant at plt 005 according
to Tukeyrsquos-HSD test
45
24 Results
241 Analyses of DON to dE-DON biotransformation
A typical LC-UV-MS analysis where there is incomplete de-epoxidation of DON by a
particular soil suspension culture is shown in Figure 2-2 Analysis by LC with UV detection
and integration of peak areas showed 662 conversion of DON to dE-DON (Figure 2-2a)
Analysis LC with MS detection (expressed as total ion counts) indicated 643 conversion
(Figure 2-2b) LC analyses with UV and TIC (total ion count) detection are subject to
interferences due to co-eluting contaminants and therefore integration of small peaks was
very difficult making quantification inaccurate MS detection by SIM (selected ion
monitoring) can eliminate the influence of co-eluting contaminants For example the SIM LC
chromatogram for DON (Figure 2-2c) was generated by using the major ions resulting from
the APCI positive ion MS of DON (Figure 2-2e)
Similarly the SIM LC chromatogram for dE-DON (Figure 2-2d) was derived from the
MS ions of dE-DON (Figure 2-2f) The signal to noise ratios (SN) of peaks observed by SIM
were at least 20 times greater than those detected by TIC because there is reduced likelihood
of co-eluting peaks having the same ions Integration of the SIM peaks revealed a 616
conversion The three methods gave similar results but the SIM LC results were considered
to be the most accurate Using that method the conversion of DON to dE-DON by the mixed
culture occurred most quickly at 48 h and was completed by 144 h (Figure 2-3)
46
47
Figure 2-2 LC-UV-MS chromatographic analysis of DON and its de-epoxidated product dE-
DON obtained by culturing with selected soil microbes (a) LC-UV chromatogram of product
mixture monitored at 218 nm (b) LC-MS total ion chromatogram (TIC) of product mixture
(c) selected ion monitoring (SIM) chromatogram of cultured DON at mz 231 249 267 279
and 297 (d) SIM chromatogram of cultured dE-DON at mz 215 233 245 251 263 and 281
(e) atmospheric pressure chemical ionization (APCI) positive ion mass spectrum of DON (f)
APCI positive ion mass spectrum of dE-DON
48
242 Screening of soil samples
Initially samples originating from soils were analyzed in three types of culture media
to support the growth of fungi yeast and bacteria However cultures growing in PDB which
is a suitable substrate for many fungi and yeasts did not permit transform DON to dE-DON
(data not shown by microorganisms) A define medium (ie MM) was also used in the
sample screening assay None of the soil samples showed de-epoxidation activity in MM
(data not shown) From five soil samples one soil sample culture showed de-epoxidation of
DON in MSB The culture in MSB converted gt99 DON to dE-DON within 144 h (Figure 2-
3) Conversion began after a lag of 48 h and was linear thereafter with a rate of 1 per hour
49
Figure 2-3 Time course of relative biotransformation of DON to dE-DON by the initial soil
suspension culture in MSB medium The culture contained 50 μgmL DON at time 0 Culture
with three replications was incubated at room temperature with continuous shaking at 180
rpm The conversion of DON to dE-DON was analysed by LC-UV-MS at 24 h intervals Blue
and red lines are the percent conversion of DON and recovery of dE-DON respectively
50
243 Evidence of biological DON de-epoxidation
Treatment of active microbial culture with 100 microgmL of streptomycin completely
inhibited DON de-epoxidation In contrast the same concentration of penicillin G did not
affect activity of the culture (Figure 2-4) Streptomycin is effective against Gram-negative
bacteria while penicillin G is lethal to Gram-positive bacteria These experiments provided
evidence that DON de-epoxidation may be caused by certain Gram-negative bacteria
51
Figure 2-4 Microbial de-epoxidation of DON after five days of incubation The active DON
de-epoxidation culture was treated with either 100 microgmL penicillin G or streptomycin
(Strepto) in MSB Cultures having three replications were added 50 microgmL DON and
incubated at room temperatures with continuous shaking at 180 rpm For controls cultures
were without antibiotics DON to dE-DON transformation was monitored after five day of
incubation by LC-UV-MS method
0
20
40
60
80
100
120
Penicillin Strepto Control
DO
N d
e-e
po
xyd
ati
on
(
)
Antibiotics
52
244 Reduction of the microbial diversity
Plating the suspension cultures onto MSB agar revealed colonies with a variety of
sizes morphologies and colours reflecting a mixed culture (data not shown) To reduce the
complexity of the culture three steps of selection pressures were applied namely treatment
with antibiotics heat shocks and subculturing on solid medium The above indicated
treatments did not significantly affect DON de-epoxidation of the microbial cultures (data not
shown) T-RFLP profiles of the mixed culture before treatments showed six distinct peaks of
about 110 150 200 220 340 and 420 bp restriction fragments with the 110 bp peak being
the predominant one (Figure 2-5a)
After treatment with antibiotics and heat only three peaks at 110 150 and 325 bp
remained (Figure 2-5b) suggesting that there was a reduction in the microbial diversity
without affecting DON de-epoxidation After enrichments the bacteria species in the culture
changed their growth patterns as the electropherogram produced different sizes of pick
lengths compared to untreated culture
53
(b)
(a)
Rela
tive flu
ore
scen
ce
32000
0
0 500
16S-rDNA fragment size (bp)
100 200 300 400
Rela
tive flu
ore
scen
ce
32000
0
500
16S-rDNA fragment size (bp)
0 300200100 400
54
Figure 2-5 Analysis of microbial population reduction by terminal restriction fragment
length polymorphism (T-RFLP) (a) Restriction fragments of the 16S-rDNA of the untreated
culture and (b) fragmentation patterns of the enriched culture Picks along the X-axis showed
the fragmentation patterns of the 16S-rDNA which is related to the bacterial species diversity
The length of picks (along the Y-axis) showed abundance of terminal restriction fragments
related to the abundance of particular bacterial speciesgenera
55
245 Microbial population structures in the enriched culture
Microbial diversity in the enriched culture was determined by analyzing the sequences
of the V3-V5 region (~586 bp) amplified from the 16S-rDNAs of the microorganisms it
contained The sequence analysis showed the presence of Serratia Citrobacter Clostridium
Enterococcus Stenotrophomonas and Streptomyces in the enriched culture (Figure 2-6)
Out of 100 V3-V5 sequences 54 sequences were found to be belonging to Serratia
the predominant species of the culture The remaining sequences matched with 12
Citrobacter 8 Clostridium 14 Enterococcus 8 Stenotrophomonas and 4 Streptomyces
species
56
Figure 2-6 Microbial genera identified in the enriched DON de-epoxidizing microbial
culture The bacterial genera were determined by comparing the hypervariable regions (V3-
V5) of the 16S-rDNA using ribosomal database project (httprdpcmemsuedu) X-axis
showed the identified bacterial genera and Y-axis demonstrated the abundance of the V3-V5
sequences belonging to different bacterial genera
54
12 8
14
8 4
0
20
40
60 S
eq
uen
ce
fre
qu
en
cy
Microbial genera
57
246 Effects of media growth temperature pH and oxygen requirements on DON de-
epoxidation
Of the nine media that were tested DON de-epoxidation by the enriched microbial
culture only occurred in NB and MSB (Figure 2-7a) It appeared that C andor N
concentrations in the media were crucial for the enzymatic activity because less or more than
05 Bacto Peptone in MSB reduced the microbial DON transformation ability
DON de-epoxidation activity occurred over a wide range of temperatures from 12 to
40ordmC (Figure 2-7b) The fastest rates of DON de-epoxidation occurred between 25 to 40ordmC
De-epoxidation was observed within a relatively narrow range of pH (60 to 75) with
the most rapid DON biotransformation activity observed at pH 70 (Figure 2-7c) At pH 70
the enriched culture showed DON de-epoxidation activity 125 times faster than at pH 60 or
75 At pH 55 and 80 there was no observable DON de-epoxidation
58
0
20
40
60
80
100
120
12degC
20degC
25degC
30degC
40degC
DO
N d
e-e
po
xyd
ati
on
(
)
Temperature
0
20
40
60
80
100
120
DO
N d
e-e
pxyd
ati
on
(
)
Media
(a)
(b)
59
Figure 2-7 Effects of culture conditions on DON to de-epoxy DON transformation by the
enriched microbial culture For the media experiment (a) all the media were adjusted to pH 7
and the incubation temperature was 27degC For the temperature experiment (b) MSB pH 7 was
used For the pH experiment (c) MSB at 27degC was used Microbial cultures were inoculated
with 50 μgmL DON De-epoxidation was determined after 72 h of incubation Media MSY
(mineral salts + 05 yeast extract) LB (Luria Bertani) TSB (tryptic soy broth) MSB
(mineral salts + 05 bacto peptone) SE (soil extract) CMB (corn meal broth) PDB (potato
dextrose broth) and MM (minimal medium)
0
20
40
60
80
100
120
55
60
65
70
75
80
DO
N d
e-e
po
xyd
ati
on
(
)
pH
(c)
60
The enriched culture showed DON de-epoxidation activity under both aerobic and
anaerobic conditions (Figure 2-8) The fastest transformation (100 de-epoxidation after 60 h
of incubation) occurred in cultures (A) that were grown under aerobic conditions throughout
the culture growth period A medium rate of de-epoxidation (100 de-epoxidation after 72 h)
was observed in cultures that were initially grown in either aerobic or anaerobic conditions
followed by transferring to anaerobic or aerobic conditions (A+AN and AN+A) respectively
The lowest rate occurred (100 de-epoxidation after 96 h) in cultures grown in continual
anaerobic conditions (AN) The means comparison of microbial DON de-epoxidation
efficiencies at 60 h of incubation showed that cultures continually grown in conditions A and
AN were statistically different Biotransformation abilities of the cultures that were switched
from one conditions to another (A+AN and AN+A) were statistically similar among each
other but different from cultures that were only grown in conditions A or AN (plt005)
61
Figure 2-8 Microbial DON to dE (de-epoxy DON)-DON transformation activity under
aerobic and anaerobic conditions A = culture incubated and examined the activity only under
aerobic conditions A+AN = culture initially was grown in aerobic conditions and then
transferred to anaerobic conditions AN = cultures grown throughout the assay under
anaerobic conditions AN+A = culture initially grown under anaerobic conditions and then
transferred to aerobic conditions Differences of lower case letters are significantly different
in de-epoxidation capability (after 60 h) among incubation conditions according to Tukeyrsquos
HSD test (p lt 005) The experiment was conducted with three replications
62
25 Discussion
The current study describes a novel bacterial culture isolated from agricultural soil that
was capable of transforming DON to de-epoxy DON under aerobic and anaerobic conditions
at moderate temperatures Because it has been speculated that DON translocates to soil from
contaminated crops and DON that enters into the soil could be metabolized by microbes
(Voumllkl et al 2004) soil samples for the current study were mainly collected from fields where
cereal crops contaminated with DON were grown
In our initial sample screening steps upwards of 144 h of incubation were required to
achieve gt99 DON de-epoxidation Such a long period may have been due to the presence of
fast growing microorganisms that had suppressive effects on the growth or activity of the
DON transforming bacteria (Davis et al 2005) The sequential use of different antibiotics
with different modes of actions followed by thermal treatments and subculturing on solid
medium resulted in a reduction in the number of bacterial species as determined by T-RFLP
analysis Analysis of V3-V5 region of the 16S-rDNA showed that five bacterial genera were
present in the enriched culture The culture showed higher DON de-epoxidation activity (only
60 h required for gt99 biotransformation) compared to initial soil suspension culture that
required 144 h to complete the de-epoxidation This microbial culture could be used to isolate
the DON de-epoxidizing gene(s) applying metagenomics approaches (Daniel 2005)
The culture showed complete de-epoxidation activity both in MSB and NB media
However we have found that the use of nutritionally rich NB medium enhanced non-target
bacterial (eg Serratia lequifaciences) growth but this did not occur in MSB which is a
comparably poor medium To avoid the possible suppressive and detrimental effects of the
fast growing bacteria on the target bacteria MSB was used for enrichment and
63
characterization of the culture An important characteristic of the DON de-epoxidation
activity observed for the microbial culture in the current study is its activity at moderate
temperature (25-30degC) In contrast previously isolated cultures showed optimal activities at
low (le15degC) or high temperatures (37degC) (Guan et al 2009) Also the culture showed DON
de-epoxidation activity at neutral pH range (pH 60-75) in contrast to previous reports which
found that microbial trichothecene de-epoxidation occurred at acidic and basic pHs (Swanson
et al 1987 He et al 1992 Kollarczik et al 1994 Young et al 2007) However a microbial
culture from fish gut showed de-epoxidation under wide range of pHs (Guan et al 2009)
Probably the pH range reflects the environment in which the bacteria naturally grow which in
this case is similar to the soil pH of a typical crop field in Southern Ontario of Canada The
advantage of the present finding is that DON de-epoxidation activity of the culture was
optimal in a temperature range that matches the temperature range of activity for most arable
plants Therefore it is reasonable to suggest that the gene(s)enzyme(s) responsible for the
activity have potential application for enzymatic detoxification of DON in cereals (Karlovsky
1999)
The current observation that bacterial DON de-epoxidation activity occurred under
both aerobic and anaerobic conditions was interesting because DON de-epoxidation is a
reductive chemical reaction that might be expected to be promoted by anaerobic conditions
However there are previous reports of aerobic de-epoxidation of eldrindieldrin by soil
bacteria (Matsumoto et al 2008) and Goodstadt and Ponting (2004) described aerobic
reductive de-epoxidation by vitamin K epoxide reductasephylloquinone reductase A
definitive determination of the oxygen sensitivity of the enzyme responsible for DON de-
epoxidation will ultimately come from experiments with the purified protein
64
In conclusion this study reports on a novel bacterial source from soil that efficiently
de-epoxidizes DON at moderate temperatures under both aerobic and anaerobic conditions
This research may lead to the isolation of microbes and gene(s) encoding enzyme(s) capable
of de-epoxidizing DON that could create transgenic plants with reduced DON contamination
thus improving food and feed safety and reducing negative effects of the toxin on public
health
26 Acknowledgements
The authors gratefully acknowledged the Natural Sciences and Engineering Research
Council of Canada (NSERC) for awarding a scholarship to Rafiqul Islam for PhD program
The research was supported by the Agriculture and Agri-Food Canada Binational
Agricultural Research and Development Fund (AAFCBARD) project We are thankful for
the contributions of Jianwei He Xiu-Zhen Li and Honghui Zhu
65
CHAPTER 3
30 ISOLATION AND CHARACTERIZATION OF A NOVEL SOIL BACTERIUM
CAPABLE OF DETOXIFYING ELEVEN TRICHOTHECENE MYCOTOXINS
31 Abstract
Deoxynivalenol (DON vomitoxin) is one of the most commonly detected food and
feed contaminating mycotoxins worldwide In order to develop a control method effective
under aerobic conditions and moderate temperatures a bacterial strain ADS47 was isolated
capable of transforming DON to the less toxic de-epoxy DON The bacterial strain is
facultatively anaerobic Gram-negative rod shaped with an average cell size of 150 X 072
microm and has peritrichous (multiple) flagella The 16S-rRNA gene sequence had gt99
nucleotide identity to Citrobacter freundii a bacterium that belongs to the gamma subclass of
proteobacteria Strain ADS47 showed near 100 de-epoxidation of DON after 42 h of
incubation in nutrient broth (NB) under aerobic conditions moderate temperature (28˚C) and
neutral pH (70) However de-epoxidation could occur at 10-45˚C and pH 60-75 Under
anaerobic conditions near 100 de-epoxidation was observed after 72 h mainly during the
stationary growth phase Microbial DON de-epoxidation ability increased with increasing
DON concentrations in the culture media Inhibition of bacterial de-epoxidation by sodium
azide suggested that an active electron transport chain was required In addition to DON
strain ADS47 was able to de-epoxidize andor de-acetylate five type-A and five type-B
trichothecene mycotoxins This is the first report of a bacterial isolate capable of de-
epoxidizing DON and trichothecene mycotoxins under aerobic conditions and moderate
66
temperatures Strain ADS47 could potentially be used in bioremediation to eliminate DON
and other trichothecene mycotoxins from contaminated food and feed
Keywords aerobic de-epoxidation Citrobacter freundii isolation soil bacterium
trichothecenes
32 Introduction
Cereals are often contaminated with a variety of type-A and type-B trichothecene
mycotoxins produced by a number of toxigenic Fusarium species (Placinta et al 1999
Binder et al 2007 Foroud and Eudes 2009) Trichothecenes are tricyclic sesquiterpenes with
a double bond at the C9 10 position and a C12 13 epoxy group (Desjardins 2006) The two
types are distinguished by the presence of a ketone at position C8 in type-B which is absent
in type-A (Yazar and Omurtag 2008) The type-B toxin deoxynivalenol (DON) is the most
frequently found mycotoxin in food and feed around the world (Larsen et al 2004 Desjardins
2006 Starkey et al 2007) Different Fusarium species associated with Fusarium head blight
(FHB) or Gibberella ear rot (GER) disease of small grain cereals and maize are mainly
responsible for contaminating grains with DON and other type-A and type-B trichothecenes
These can cause serious mycotoxicosis in humans animals and plants (Ueno 1985 DrsquoMello
et al 1999 Bae and Pestka 2008) Common effects of DONTC toxicity in animals include
diarrhea vomiting feed refusal growth retardation immunosuppression reduced ovarian
functionsreproductive disorders and even death (Pestka and Smolinski 2005 Pestka 2007)
DON enhances FHB disease severity in wheat and maize (Proctor et al 1995 Desjardins
1996 Maier et al 2006)
67
Reduction of mycotoxin levels in contaminated agricultural products remains a
challenging task Current physical and chemical processes to decrease DON content are either
ineffective or prohibitively expensive (Kushiro 2008 He et al 2010b) However the use of
detoxifying microbes to biodegrade mycotoxins is an alternative (Awad et al 2010
Karlovsky 2011) The toxicity of DON is mainly due to the epoxide and there is a significant
reduction of DON toxicity (54) when it is reduced to produce de-epoxy DON Related
mycotoxins also show reductions in toxicity when they are de-epoxidized such as NIV (55)
and T-2 toxin (400) (Swanson et al 1988 Eriksen et al 2004) Bacillus arbutinivorans
strain LS100 and Eubacterial strain BBSH 797 originating from chicken intestine and rumen
fluid respectively are capable of cleaving the epoxide ring of DON under anaerobic
conditions and high temperature (37degC) (Fuchs et al 2002 Yu et al 2010) Strain BBSH 797
which can de-epoxidize DON scirpentriol and T-2 triol trichothecene mycotoxins has been
used as an additive for poultry and swine diets (Karlovsky 2011) However there are no
reports of microbes able to de-epoxidze DON and other trichothecenes under aerobic
conditions and at moderate temperatures
To develop an effective method for eliminating DON and trichothecene mycotoxins
from contaminated grains soils from fields with FHBGER infected crops were screened for
DON de-epoxidizing microbes (refer to chapter 2 Islam et al 2012) One soil sample yielded
a mixed microbial culture was partially enriched for bacteria with DON de-epoxidizing
activity (chapter 2) The current study describes the purification and characterization of a
novel DON de-epoxidizing bacterium from this culture
68
33 Materials and methods
331 Microbial source
A mixed microbial culture originating from a screen of more than 150 separate soil
samples collected from maize and wheat fields near Guelph Ontario Canada (Islam et al
2012) was used for enrichment purification and characterization of the DON de-epoxidizing
bacterium
332 Media and chemicals
The media used were nutrient broth (NB) 110 strength NB (110 NB) minimal
medium (MM) consisting of six mineral salts (MS) + 100g per liter sucrose (Islam et al
2012) MS + 05 Bacto Peptone (MSB) MS + 03 Bacto Peptone (MSC) MS + 05
yeast extract (MSY) MS + 200 microgmL DON (MS-DON) sterile water-soluble soil extract
(SE) (Islam et al 2012) potato dextrose broth (PDB) and Luria Bertani (LB) Prepared media
and media components were obtained from Fisher Scientific (Fair Lawn New Jersey USA)
Sigma-Aldrich (Oakville ON Canada) andor Difco (Sparks Maryland USA) Media pH
was adjusted as required prior to autoclaving For solid media preparation 15 agar was
added DON and all trichothecene mycotoxins were purchased from Biomin (Romer Lab
Union MO USA)
333 Isolation of the DON de-epoxidizing bacterial strain
Aliquots (20 microL) of the microbial culture partially enriched for DON de-epoxidizing
ability (Islam et al 2012) was transferred to 05 mL MSB (pH 70) in sterile 15 mL
Eppendorf tubes After addition of 200 microgmL pure DON the culture was incubated at room
69
temperature with continuous shaking at 220 revolutions per minute (rpm) Seven to ten days
later 20 microL of the culture was transferred to fresh MSB supplemented with 200 microgmL DON
The culture was then incubated for 7 to 10 days under the above conditions This procedure
was repeated 12 times
After three months of repeated transfers in broth with 200 microgmL DON 100 microL of the
culture was serially diluted in 09 mL MSB containing 05 Tweenreg20 (Sigma Aldrich) to
reduce bacterial cell aggregation (Mabagala 1997) The dilutions were vigorously shaken for
ten minutes A 100 microL aliquot from each dilution was spread on MSB + agar (MSA) The
MSA plates were incubated at room temperature until single colonies appeared
Single colonies from plates were transferred to 1 mL MSB+ 50 microgmL DON and
incubated at room temperature with continuous shaking at 180 rpm under aerobic conditions
After three days the DON de-epoxidation ability was examined by mixing an equal volume
of the bacterial culture and methanol and incubating for 30 min at room temperature to lyse
the cells Insoluble materials in the mixture were removed by passing it through a 045 microm
Millipore filter (Whatmanreg Florham Park New Jersey USA) The level of de-epoxy DON
(dE-DON also known as DOM-1) in the filtrate was determined by liquid chromatography-
ultraviolet-mass spectrometry (LC-UV-MS) (Guan et al 2009)
Individual colonies that showed de-epoxidation activity were serially diluted in
MSB+05 Tweenreg20 and plated onto MSA Single colonies that appeared on the agar plate
were incubated in MSB as described above and retested for DON de-epoxidation ability To
ensure that pure cultures were obtained this was repeated 10 times and individual colonies
were selected on MSA each time
70
334 Identification of a DON de-epoxidizing bacterium
Strain ADS47 with DON de-epoxidation ability was obtained from a single colony
after 10 serial transfers and was grown on MSA at room temperature under aerobic
conditions for 72 h Colony color shape edge and elevation were observed The cell
morphology of the bacterium was examined with a transmission electron microscope at the
University of Guelph Guelph Ontario Canada Whole bacterial cells were placed directly on
a formvar and carbon-coated 200-mesh copper grid They were washed two times by touching
the grids to a drop of clean dH2O and the excess water was blotted off The samples were
negatively stained using 1 Uranyl Acetate (aq) for five seconds Electron microscopy was
performed on a Philips CM10 transmission electron microscope at 60kV under standard
operating conditions (Philips Optics Eindhoven Netherlands) Gram staining was performed
using the procedure described in the commercial gram stain kit from BD Canada
(Mississauga Ontario Canada)
Total bacterial DNA was extracted using a DNeasy Plant DNA Extraction Mini Kit
(Qiagen Inc) The 16S-rDNA was PCR amplified using the primers 46f-
5rsquogcytaacacatgcaagtcga3rsquo (Kaplan and Kitts 2004) and 1371r-5rsquogtgtgtacaagncccgggaa3rsquo
(Zhao et al 2007) PCR was performed in 25 microL containing 06 microM of each primer 1x of
HotStarTaqreg Master Mix (Qiagen Inc) and ~40 ng of DNA The PCR cycles were 95ordmC (15
min) 35 cycles of 94ordmC (20 s) 52ordmC (15 s) 72ordmC (1 min 30 s) and a 72ordmC (7 min) extension
step Amplification was carried out in a GeneAmpreg PCR System 9700 (Applied Biosystems)
PCR products were sequenced with an automated ABI 3730 DNA analyzer (Applied
Biosystems) The 16S-rRNA gene sequence similarity was performed by BlastN using the
Ribosomal Database Project (RDP) (httprdpcmemsueduseqmatch)
71
A phylogenetic tree was constructed by comparing the partial 16S-rDNA sequence of
strain ADS47 with the 11 known Citrobacter species (Bornstein and Schafer 2006 Brenner et
al 1993) The Citrobacter strains were C freundii [FN997616] C koseri [AF025372] C
amalonaticus [AF025370] C farmeri [AF025371] C youngae [AB273741] C braakii
[AF025368] C werkmanii [AF025373] C sedlakii [AF025364] C rodentium [AF025363]
C gillenii [AF025367] and C murliniae [AF025369] Escherichia coli O157H7 [AE005174]
was used as an out group strain The analysis was performed using distance matrix with a gap
opening of 100 and a gap extension of 01 by a server based ClustalW2 program
(httpwwwebiacukToolsmsaclustalw2) A dissimilarity tree was created and viewed
using a TreeView X software (Page 1996)
335 Effects of DON concentration aeration substrate temperature pH and antibiotic
To determine if DON concentration can affect the growth of the isolated bacteria
tubes containing 5 mL of NB or MSB with 20 200 or 500 microgmL of DON were inoculated
with strain ADS47 that had been grown for 16 h with continuous shaking at 220 rpm and then
adjusted to cell density at 025 OD600nm (107 cellsmL) The cultures were incubated at 28degC
with continuous shaking at 180 rpm and 05 mL was removed every 12 h to measure
OD600nm The culture (05 mL) was then analysed for DON de-epoxidation activity by LC-
UV-MS (Guan et al 2009) as described above Three replicate cultures were tested
To examine the effect of aeration five mL of NB or MSB containing 50 microgmL DON
were inoculated with an overnight culture of strain ADS47 prepared as described above and
incubated at 28˚C with continuous shaking at 180 rpm One mL of culture was removed every
six hours and bacterial growth was determined by measuring cell density at OD600nm At the
72
same time points culture samples were analysed for DON to dE-DON transformation ability
(Guan et al 2009) The experiment was repeated with MSB medium containing 50 microgmL
DON under anaerobic conditions as described by Islam et al (2012) with DON to dE-DON
transformation ability being determined every 6 h Three replicate cultures were prepared for
each assay
The effect of substrate on DON de-epoxidation ability was tested after growing
ADS47 for 72 h in NB 110 NB MM MS-DON MSB MSC MSY SE PDB or LB
containing 50 microgmL DON For this assay the media were maintained at pH70plusmn02 and 28degC
for 72 h The effect of temperature was examined with MSB containing 50 microgmL DON at 10
15 20 25 30 35 40 45 or 50degC The effect of pH on growth of strain ADS47 was examined
with MSB adjusted to pH 55 60 65 70 75 or 80 and the cultures were incubated at 28degC
for 72 h Three replicate cultures were used for each assay
Overnight cultures of ADS47 in MSB were treated with 001 and 0001 (wv) of
an antibiotic sodium azide (SA) (Sigma-Aldrich) Controls were prepared in MSB without
SA Three replicate cultures were spiked with 50 microgmL DON DON to dE-DON
transformation ability of the bacterial culture was analysed (Guan et al 2009) after 72 h at
28degC with continuous shaking at 180 rpm Three replicate cultures were tested
336 Localization of DON to dE-DON transformation ability in the bacterium
Bacterial cell extracts were prepared using B-PER II Bacterial Protein Extraction
Reagent following the manufacturerrsquos instructions (PI 78260 Thermo Scientific Rockford
IL USA) ADS47 was grown in 15 mL MSB for 48 h under the growth conditions described
above A culture (05 mL) was centrifuged at 5000 x g for 10 minutes and the pellet was
73
collected B-PER II reagent (2 mL) was added to the pellet and the cells were lysed by
pipetting the suspension up and down several times The cell lysate was treated with 20 microL
Halt Protease Inhibitor Cocktail (Thermo Scientific Rockford IL USA) Culture filtrate was
prepared by passing 05 mL of the culture through a 022 microm Millipore filter (Fisher
Scientific) The remaining 05 mL culture was used as control Cell lysate culture filtrate and
control were incubated with 50 microgmL DON and incubated at 28degC DON to dE-DON
transformation ability of the cell lysate culture filtrate and control were analysed after 72 h
incubation (Guan et al 2009) Three replicate cultures were tested
337 Detoxification of additional trichothecene mycotoxins
Strain ADS47 was evaluated for its ability to degrade five type-A trichothecene
mycotoxins (HT-2 toxin T-2 toxin T2-triol diacetoxyscirpenol and neosolaniol) and five
type-B trichothecene mycotoxins (nivalenol verrucarol fusarenon X 3-acetyl-
deoxynivalenol and 15-acetyl-deoxynivalenol) One and a half mL NB was added with 100
microgmL of each mycotoxin and then inoculated with 100 microL of ADS47 culture grown
overnight as previously described The culture of NB + mycotoxin + ADS47 was incubated at
28degC with continuous shaking at 180 rpm and DON to dE-DON transformation ability of the
culture was analysed by LC-UV-MS (Guan et al 2009) Five replicate cultures were tested for
each trichothecene mycotoxin
74
338 Statistical analysis
Data were processed using Microsoft Excel Program 2007 Statistical analysis was
performed using General Linear Model procedure in SAS version 91 (SAS Institute Cary
NC USA 2002-03) Replications were averaged and stated as mean plusmn standard error The
mean differences were considered significant at plt 005 according to Tukeyrsquos-HSD test
75
34 Results
341 Isolation and identification of a DON de-epoxidizing bacterium
A DON de-epoxidizing bacterium designated strain ADS47 was isolated after 12
transfers over three months from cultures growing in MSB containing 200 microgmL DON
Strain ADS47 produced small to medium size circular convex and white colonies on MSA at
room temperature under aerobic conditions (Figure 3-1a) The bacterium is rod shaped and
150 x 072 microm in size (Figure 3-1b) It has numerous flagella that cover the surface of the cell
(peritrichous flagella) and is Gram-negative
BlastN analysis of the 16S-rDNA sequence of strain ADS47 showed the most
similarity to the eleven known Citrobacter species in the gamma proteobacteria of the
Enterobacteriaceae In particular the 16S-rDNA nucleotide sequence of ADS47 was more
than 99 identical to C fruendii (Figure 3-2) The next most similar sequences were a cluster
of C werkmanii and C murliniae followed by more distantly related C braakii C gilleni
and C youngae A separate cluster was formed by C rodentium C farmeri C sedlakii C
koseri and C amalonaticus
76
Figure 3-1 (a) Colony color shape and size of the mycotoxin degrading bacterial strain
ADS47 on mineral salts agar (MSA) medium (b) Transmission electron microscopic (TEM)
image of the Citrobacter freundii strain ADS47 The bacterium showed multiple flagella
(peritrichous flagella) and numerous fimbriae around the cell surface
(a) (b)
77
Figure 3-2 Dendrogram based on 16S-rDNA sequences for strain ADS47 and 11 reference
Citrobacter species An E coli O157H7 strain was considered as an out group species The
tree was constructed using a genetic distance matrix by ClustalW 2 and viewed by
TreeViewX software The distance was converted to scale and shown by branch lengths (scale
bar 001 which indicated the 16S-rDNA nucleotide sequence variability between the bacterial
species) GenBank accession numbers for 16S-rDNA sequences of the species used for
comparison are shown in materials and methods section (334)
78
342 Effects of DON concentrations on bacterial DON de-epoxidation
Growth of ADS47 aerobically in MSB with 20 200 and 500 microgmL DON was not
significantly different from that observed in control cultures without DON (Figure 3-3a)
Similarly increasing DON concentrations did not affect bacterial growth in NB medium
(Figure 3-3b)
DON de-epoxidation activity determined as microgmL DON convertedcell density
(OD600) was measured lowest in MSB+20 microgmL DON and highest in MSB+ 500 microgmL
DON containing cultures (Figure 3-3c) The highest rate of dE-DON formation was 115 microgh
in ADS47 cultures in MSB with 500 microgmL DON between 48 to 72 h All of the DON was
converted in the cultures containing 20 microgmL DON and 200 microgmL DON between 48-60 h
and 60-72 h respectively In 200 microgmL and 500 microgmL of DON cultures the rate of de-
epoxidation increased significantly after 36 h
79
00
04
08
12
16
0 24 36 48 60 72 96
OD
600n
m
Incubation time (h)
Control
20 microgmL
200 microgmL
500 microgmL
(b)
(a)
80
Figure 3-3 Effects of DON concentration on bacterial growth and de-epoxidation activities
Strain ADS47 was grown in MSB and NB media with three different DON concentrations 20
microgmL 200 microgmL and 500 microgmL Control cultures were prepared without DON Cultures
were grown at 28degC with continuous shaking at 180 rpm (a) Microbial density (OD600 nm) in
MSB with different DON concentrations was measured at 24 36 48 60 72 and 96 h (b)
Microbial growth (OD600 nm) in NB having different DON concentration was measured at 24
48 72 and 96 h (c) Bacterial DON de-epoxidation activity in 20 200 and 500 microgmL DON
containing cultures at different time points DON de-epoxidation was measured in microgmL per
OD600 nm Three replicate cultures for each assay were analysed
(C)
81
343 Influence of growth conditions and antibiotic on microbial DON de-epoxydation
Aerobic and anaerobic conditions
ADS47 grew more quickly uner aerobic conditions under MSB+50 microgmL DON than
in MSB+50 microgmL DON under anaerobic conditions (Figure 3-4a) Aerobic growth was
faster in NB+50 microgmL DON compared to MSB+50 microgmL DON
However the DON de-epoxidation activities on a cell density basis were very similar
in NB+50 microgmL DON compared to MSB+50 microgmL DON (Figure 3-4b) Also DON de-
epoxidation activity of ADS47 under aerobic conditions in MSB+50 microgmL DON was higher
than in the same medium under anaerobic conditions (Figure 3-4b)
82
(b)
(a)
83
Figure 3-4 Bacterial growth and DON de-epoxidation capabilities under aerobic and
anaerobic conditions (a) Microbial growth under aerobic conditions in MSB and NB medium
and anaerobic conditions in MSB medium (b) Microbial DON de-epoxidation ability under
aerobic conditions in MSB and NB media and anaerobic conditions in MSB medium
Cultures media having 50 microgmL DON were inoculated with overnight culture of strain
ADS47 Initial microbial concentration in the cultures was adjusted to 025 OD600nm Cultures
were incubated at 28degC having three replications Bacterial growth at OD600nm and DON de-
epoxidation in microgmL per OD600nm were measured at six hour interval
84
Media temperature and pH
Growth of ADS47 in media containing 50 microgmL DON under aerobic conditions at 72
h was three-fold higher in NB and LB than in SE two-fold higher than in 110 NB and MSC
and did not occur in MS-DON (Figure 3-5a) Growth in MM MSB and MSY was
intermediate
Over the 72 h DON de-epoxidation per cell density was highest in MSB and MSY and
approximately 25 lower in NB and LB and lower by 75 and 87 in 110 NB and MSC
respectively (Figure 3-5b) No DON de-epoxidation was observed in MS-DON in which
DON was used as a carbon source Bacteria grown in MM broth which contains sucrose as a
sole carbon source were unable to de-epoxidize DON No DON de-epoxidation was observed
in SE and PDB grown cultures
Growth of ADS47 under aerobic conditions for 72 h in MSB+50 microgmL DON was
greatest between 25-40degC but no growth occurred at 50degC (Figure 3-6a) DON de-
epoxidation activity was greatest and equivalent between 20-40degC and lower at 10 15 and
45degC temperatures (Figure 3-6b)
ADS47 growth under aerobic conditions for 72 h in MSB+50 microgmL DON was
limited to neutral pHs (ie 60 - 75) (Figure 3-7a) The transformation of DON to dE-DON
occurred at the same pH range and DON de-epoxidation activity was not significantly
different between pH 60 to 75 (Figure 3-7b)
85
b
c
a
b
a
d c
b b
(a)
(b)
b
c
b
a a
d
86
Figure 3-5 Effects of media on bacterial growth and DON de-epoxidation activity (a)
Microbial growth (at OD600nm) on different culture media Media pHs were adjusted to
70plusmn02 (b) Bacterial DON to dE-DON transformation in microgmL by OD600nm in different
media after 72 h The cultures having three replications were incubated for 72 h at 28degC and
DON de-epoxidation was measured The Media NB = nutrient broth 1NB = 110 strength
NB MM = minimal medium MS-DON = six mineral salts + 200 microgmL DON as sole C
source MSB = six mineral salts + 05 bacto peptone MSC = six mineral salts broth + 03
bacto peptone MSY = six mineral salts with 05 yeast extract SE = soil extract PDB =
potato dextrose broth and LB = Luria Bertani Means with the same label are not significantly
different according to Tukeyrsquos HSD test (p lt 005)
87
(a)
(b)
88
Figure 3-6 Effects of temperature on microbial growth and DON de-epoxydation capability
(a) Microbial growth (at OD600nm) at different temperatures after 72 h of incubation (b)
Bacteria DON de-epoxidation in microgmL by OD600nm in different temperatures The most
efficient temperature range (ge40 microgmL DON de-epoxidation) is marked by dot lines The
experiment was conducted in MSB medium at pH 70plusmn02 with three replications
89
(a)
(b)
90
Figure 3-7 Influence of pH on microbial growth and DON de-epoxydation (a) Microbial
growth (OD600nm) in MSB at different pH (b) Bacterial DON to de-epoxy-DON
transformation in microgmL by OD600nm in different pH Dot lines indicated the efficient de-
epoxidation (ge40 microgmL DON conversion) pH range Experiment was conducted at 28degC
with three replications
91
Antibiotic
Treatment of ADS47 at 72 h in MSB+50 microgmL DON with sodium azide (SA) at
0001 and 001 reduced DON de-epoxidation activity to 5 (25 microgmL) and 0
respectively The control with no SA added was able to de-epoxidize DON completely
(100) (Figure 3-8)
Figure 3-8 Effects of sodium azide (SA) on bacterial DON de-epoxidation activity
Microbial culture in MSB+50 microgmL DON was treated with 001 (SA-1) and 0001 (SA-
2) sodium azide In control (C) no SA was added The experiments with three replications
were incubated for 72 h at 28degC and DON de-epoxidation was measured by LC-UV-MS
method
0
20
40
60
80
100
120
SA1 SA2 C
D
ON
de
-ep
oxyd
ize
d
92
344 Localization of bacterial DON de-epoxidation
A cell lysate prepared from ADS47 grown for 48 h in MSB transformed 20 (10
microgmL) of DON added at 50 microgmL to dE-DON within 72 h (Figure 3-9) In contrast a cell-
free culture filtrate did not show DON de-epoxidation activity
Figure 3-9 Determination of DON to de-epoxy DON transformation by bacterial culture
filtrate and cell extract The treatments having three replications were spiked with 50 microgmL
DON and incubated at 28degC for 72 h In control strain ADS47 was incubated in MSB broth
with 50 microgmL DON DON de-epoxidation activity in percentage was examined
0
20
40
60
80
100
120
Filtrate Extract Control
D
ON
de
-ep
oxyd
ize
d
93
345 Bacterial detoxification (de-epoxidtaion and de-acetylation) of other trichothecene
mycotoxins
Aerobic cultures of ADS47 grown for 72 h in NB+100 microgmL HT-2 converted 74 of
the toxin to de-epoxy HT-2 toxin and 26 to a de-acetylated product (Figure 3-10 and Table
3-1)
In a culture treated with T2-toxin 46 was converted to de-epoxy T-2 16 was
converted to an unknown compound and 38 was not converted In ADS47 culture with
diacetoxyscirpenol (DAS) 61 was transformed into de-epoxy DAS and 39 was de-
acetylated DAS (39) Neosolaniol (NEO) was transformed to de-epoxy NEO (55) and
de-acetylated NEO (45) T2-triol was transformed in to de-epoxy T2-triol (47) and de-
acetylated T2-triol (53)
Aerobic cultures of ADS47 were also grown for 72 h in NB+100 microgmL of one of five
type-B trichothecenes For nivalenol (NIV) 92 was de-epoxydized while the remaining 8
NIV was transformed to DON 3-acetyl-deoxynivalenol (3-ADON) was transformed into
three products de-epoxy NIV (5) de-epoxy DON (54) and DON (41) For 15-
acetyldeoxynivalenol (15-ADON) 8 was transformed to de-epoxy 15-ADON and 92 to
de-epoxy DON (dE-DON) FusarenonX (FUS) had 95 transformed into de-epoxy FUS
while 97 of verrucarol (VER) was transformed to de-epoxy VER The remaining 5 FUS
and 3 VER were not transformed
94
Figure 3-10 Determination of microbial de-epoxidation and de-acetylation of HT-2 toxin by
LC-UV-MS Bacterial culture in NB was added with 100 microgmL HT-2 toxin Bacterial HT-2
toxin transformation ability was examined after 72 h of incubation at 28degC The upper panel
illustrated mass spectrometry chromatograph of the de-acetylated (red peak) and de-
epoxydized (blue peak) products Arrows indicated the targeted acetyl (red) and epoxy (blue)
functional groups of HT-toxin The middle and lower panels showed the mass spectra of the
de-acetylated (dA) and de-epoxidized (dE) compounds Arrows on the upper panel indicated
the targeted acetyl (red) and epoxy (blue) functional groups of HT-2 toxin Red and blue
arrows on the middle and lower panels pointed at the degradation sites of HT-2 toxin
95
Table 3-1 Microbial transformation of type-A and type-B tricthothecene mycotoxins after 72
h of incubation under aerobic conditions at room temperature DAS=diacetoxyscirpenol
DON=deoxynivalenol FUS=FusarenonX 3AON=3-acetyl-deoxynivalenol and
15ADON=15-acetyl-deoxynivalenol
1 Convertion of trichothecene mycotoxins to either DON or an unknown compound
Trichothecene
mycotoxin
Untransformed
trichothecenes ()
Transformed products of
trichothecenes ()
De-
epoxy
De-
acetylate
Other1
Type-A
HT-2 toxin 0 74 26 -
T-2 toxin 38 46 - 16
(unknown)
DAS 0 61 39 -
Neosolanol 0 55 45 -
T2-triol 0 47 53 -
Type-B
DON 0 100 - -
Nivalenol 0 92 - 8 (DON)
3-ADON 0 59 41 (DON)
15-ADON 0 100 - -
FUS 5 95 - -
Varrucarol 3 97 - -
96
35 Discussion
The ADS47 strain of C freundii was isolated in the present study by a prolonged
incubation of a mixed microbial culture with a high concentration DON since the bacterium
efficiently de-epoxidized (detoxified) DON and number of other trichothecene mycotoxins
under aerobic and anaerobic conditions at moderate temperatures and neutral pH The total
procedures for enrichment and isolation of the strain ADS47 is depicted in supplemental
Figure S3-1 The reductive de-epoxidation activity may be due to a cytoplasmic enzyme
which would be in agreement with a previous study that showed de-epoxidation of
epoxyalkanes was catalyzed by a bacterial cytoplasmic flavoprotein (Westphal et al 1998)
The ability of the currently isolated strain of C freundii to biodegrade environmental
pollutants such as aminoaromatic acids and heavy metals (copper) is also likely mediated by
cytoplasmic enzymes since these compounds need to be taken up by the cells before they are
metabolized (Savelieva et al 2004 Sharma and Fulekar 2009) In contrast oxidative
transformation of DON to 3-keto DON by microbial extracellular enzyme has been reported
(Shima et al 1997)
Isolation of a trichothecene de-epoxidizing microorganism from a mixed microbial
community is very challenging Although trichothecenes are toxic to eukaryotes prokaryotes
appear to be resistant to these mycotoxins (He et al 1992 Rocha et al 2005) The present
study showed that the bacteria grew well in media with high concentrations of DON (500
microgmL) Bacterial de-epoxidation of DON involves the removal of an oxygen atom from the
functional epoxy ring of the tricyclic DON structure (Yoshizawa et al 1983) Such co-
metabolic degradation does not provide the bacteria with any nutrients from the reacting
compounds (Horvath 1972 Fedorak and Grabic-Galic 1991) This is supported by the results
97
of the current study which showed that strain ADS47 was unable to grow in media with DON
as a sole carbon source and increasing the concentration of DON did not increase ADS47
growth in MSB Hence DON could not be used as sole nutrient source in the mixed culture
enrichment process
Many previous attempts to obtain DON de-epoxidizing microbes have failed
particularly under aerobic conditions because media or laboratory conditions were
inappropriate to support growth or initially active cultures lost de-epoxidizing function after
serial transfer (Karlovsky 2011)
Media composition appears to be a crucial factor for microbial trichothecene de-
epoxidation activity (Voumllkl et al 2004) Effective biotransformation of trichothecene
mycotoxins by the fish digesta microbial culture C133 occurred in full media (FM) but a low
level of de-epoxidation occurred in media having a single nitrogen source and no
transformation activity was observed in the common bacterial medium LB (Guan et al
2009) Zhou et al (2010) also showed that chitin de-acetylation by soil bacterial strains
occurred more rapidly in yeast extract compared to common bacterial media Therefore a
screening strategy for microbial DON de-epoxidation ability should use different media in
parallel as was done in this study
This study observed efficient de-epoxidation in common bacterial media (NB and LB)
as well as in mineral salts medium containing yeast extract (MSY) or 05 Bacto Peptone
(MSB) However reduction of Bacto-Peptone to 03 (MSC) significantly reduced the
bacterial de-epoxidation activity suggesting that the activity is sensitive to the availability of
protein and amino acids The study found that medium containing sucrose as carbon source
(MM) and a commonly used medium PDB blocked microbial DON de-epoxidation even
98
though bacteria grew in those growth media These suggest that media containing high sugar
contents had inhibitory effects on microbial de-epoxidation enzymatic activity Previously
sugars have been found to have negative effects on the production andor activity of bacterial
enzymes (Saier and Roseman 1971)
Losses of microbial functions under laboratory conditions limited researchers to obtain
economically important microbial genetic sources The soil borne Curtobacterium sp strain
114-2 efficiently de-epoxydized nivalenol but unfortunately the strain lost its activity in
latter generations (Ueno et al 1983) Instability of de-epoxidation activity of ADS47
observed in cultures grown on agar media without DON resulted in the loss of DON de-
epoxidation activity by more than 80 of the colonies after one generation However strain
ADS47 retained de-epoxidation activity after ten subcultures in MSB (data not shown)
The non-specific de-epoxidation of DON made it impossible to use DON as a sole
substrate C source for selective isolation of ADS47 Thus a number of techniques were
utilized to select ADS47 and suppress other bacteria including the use of a variety of
antibiotics and culture at elevated temperatures (Islam et al 2012) Selection of the isolate
ADS47 required repeated subculturing and prolonged exposure to media containing 200
microgmL DON After three months of selection with DON culture that was obtained had a
DON de-epoxidation activity that was up to 30-fold higher than the mixed culture As growth
of ADS47 was not affected by 200 microgmL DON the prolonged incubation may have reduced
populations of other bacteria that are more sensitive to DON Enrichment and isolation of a
DON to 3-keto DON transforming bacterial strain E3-39 by prolonged incubation of a soil
microbial culture with 200 microgmL DON was reported by Shima et al (1997)
99
This study demonstrated that microbial DON de-epoxidation occurred mainly at the
stationary growth phase A DON degrading strain of Citrobacter may have gained energy
from DON by co-metabolic cleavage of the epoxy ring under less nutrient conditions
Microbial use of energy from co-metabolic reaction has been suggested by Chiu et al (2005)
The rate of de-epoxidation activity increased when ADS47 grew at higher concentrations of
DON Taken together a prolonged exposure to high concentration DON may have resulted in
non-DON metabolizing microbial populations declining by DON toxicity while facilitated
the population of DON metabolizing bacterium by providing energy
DON is a small molecule and is simply absorbed by cells (Sergent et al 2006)
Microbial DON de-epoxidation occurred in intact and lysed bacterial cells In addition
treating the ADS47 culture with SA which blocks gram negative bacterial electron transport
and activity inhibited DON de-epoxidation activity (Lichstein and Malcolm 1943 Erecinska
and Wilson 1981) implying that an active electron transport system required for microbial de-
epoxidation reaction (He et al 1992) The data suggest that a cytoplasmic epoxy reductase is
involved in the DON de-epoxidation reaction in the ADS47 microbial strain that was isolated
in the present study
Since agricultural commodities are often contaminated with multiple fungal
mycotoxins the ability of bacteria like ADS47 to degrade mycotoxins by several
mechanisms may be an advantage De-acetylation might be another mechanism of microbial
mycotoxin detoxification since toxicity of trichothecenes also relies on the acetyl functional
groups (Desjardins 2006) Strain ADS47 showed efficient de-epoxidation and de-acetylation
of several type-A and type-B trichothecene mycotoxins that are commonly detected in cereals
in association with DON In conclusion the soil-borne gram-negative bacterium C freundii
100
strain ADS47 is capable of detoxifying trichothecene mycotoxins by both de-epoxidation and
de-acetylation under aerobic and anaerobic conditions neutral pH and a range of
temperatures ADS47 has great potential to detoxify DON contaminated animal feed
Furthermore isolation of novel bacterial de-epoxidation genes might be used to generate
transgenic plants that can detoxify DON in contaminated seed Alternatively contaminated
seed might be treated with manufacture de-epoxidation enzyme(s) These applications can
improve the quality and safety of agricultural commodities that are prone to trichothecene
mycotoxin contamination
36 Acknowledgements
Thank to the Natural Sciences and Engineering Research Council of Canada (NSERC) for a
scholarship to Rafiqul Islam The work was funded by the Agriculture and Agri-Food Canada
Binational Agricultural Research and Development Fund (AAFCBARD) project We
appreciate Honghui Zhu for technical assistance
101
Supplemental Figure S3-1 Flow chart depicts the procedures of enrichment and isolation of
DON de-epoxidation bacterial strain ADS47 Steps 1 to 4 describe the optimization of
microbial DON to dE-DON transformation activity and removal of undesired microbial
species Step 5 to 7 shows the culture enrichment by incubation with high concentration DON
and isolation of the single strain ADS47
Step 1 Screening of soil samples
Step 2 Optimization of cultural conditions
Step 3 Elimination of undesired microbial species
Step 4 Removal of untargeted microbial species
Step 5 High concentration DON enrichment
Step 6 Separation of bacterial cells and dilution plating
Step 7 Single colony analysis
Active culture was sequentially treated with different mode of actions antibiotics heating cultures at 50degC and subculturing on agar media
Colonies were tested for their DON de-epoxidation ability Purified the single colony via several times subculturing on agar medium
Optimized the culture conditions ie growth substrates temperatures pH and oxygen requirements
Obtained DON de-epoxidation capable microbial culture obtained by analyzing soil samples collected from cereal fields
Serially diluted active culture was grown on agar media Collected washed colonies having DON de-epoxidation activity
The partially enriched culture that retained DON de-epoxidation activity was incubated with 200 microgmL DON in MSB media with continuous shaking at 220 microgmL for 7 d The procedure was repeated 12 times
The enriched culture was serially diluted and treated dilutes with 05 Tween 20 After vigorously vortex for ten min aliquot cultures were spread on different agar media plates to produce single colonies
102
CHAPTER 4
40 CONSTRUCTION AND FUNCTIONAL SCREENING OF GENOMIC DNA
LIBRARY FOR ISOLATION OF DON DE-EPOXYDIZING GENE(S) FROM
CITROBACTER FREUNDII STRAIN ADS47
41 Abstract
Enzymatic activity-based screening of a genomic DNA library is a robust strategy for
isolating novel microbial genes A genomic DNA library of a DON detoxifying bacterium
(Citrobacter freundii strain ADS47) was constructed using a copy control Fosmid vector
(pCC1FOSTM
) and E coli EPI300-T1R as host strain A total of 1250 E coli clones containing
large DNA inserts (~40 kb) were generated which resulted in about 10-fold theoretical
coverage of the ADS47 genome The efficiency of E coli transformation with the
recombinant Fosmid vector was examined by restriction digestion of Fosmids extracted from
15 randomly selected library clones The fragmentation patterns showed that all of the clones
contained an 81 kb Fosmid backbone and ~40 kb of insert DNA except one that had a small
insert This suggests that a DNA library was prepared with expected DNA inserts The de-
epoxidation ability of the library was analysed by incubating clones in Luria Bertani (LB)
broth medium supplemented with 20 microgmL DON with induction (by addition of L-arabinose)
or without induction None of the 1000 clones showed DON de-epoxidation activity The
potential DON de-epoxidation gene(s) of ADS47 may not have been expressed in E coli or
might have been expressed but lacked additional factors required for DON de-epoxidation
103
Keywords DNA library fosmid vector functional library screening DON de-epoxidation
activity
42 Introduction
Microorganisms contain numerous valuable genes and biocatalysts which may have
significant roles in agricultural productivity (Jube and Borthakur 2007) Various techniques
have been used to isolate novel genes from different organisms including transposon
mutagenesis (Frey et al 1998 Tsuge et al 2001) gene knockout (Winzeler et al 1999)
whole genome sequencing (Krause et al 2006) comparative genomics (Koonin et al 2001)
and suppression subtractive hybridization (Dai et al 2010)
The genomic DNA library preparation and activity-based screening of the library is a
relatively new technique which has been efficiently used for isolation of microbial novel
enzymes and compounds such as lipases proteases oxidoreductases antibiotic resistance
genes membrane proteins and antibiotics (Henne et al 2000 Gupta et al 2002 Knietsch et
al 2003 Gabor 2004 Suenaga et al 2007 van Elsas et al 2008) The strength of the
technique is that genes from microbial sources could be obtained without prior sequence
information and not requiring culturing the organisms under laboratory conditions Using this
technique an unknown gene or a set of genes could be obtained by determining particular
phenotypes of the library clones Eventually the gene of interest from the positive clones is
isolated by using a forward (eg mutation) or reverse genetics method (eg subcloning the
positive clones into a microbial expression vector) followed by functional analysis of the
subclones
104
Although this strategy has been used effectively for isolating novel microbial genes
several factors need to be considered in order for the method to be successful These factors
include DNA quality vector type (eg plasmid cosmid fosmid bacterial artificial
chromosome etc) vector copy number and host strains (Daniel 2005 Singh et al 2009) The
Fosmid vector-based technique has been used successfully to clone bacterial genes including
xenobiotics degradation genes antifungal gene clusters and antibiotic resistance genes from
soil metagenomes (Hall 2004 Suenaga et al 2007 Chung et al 2008) This study reports the
construction and activity-based screening of a Fosmid DNA library in an E coli host with the
aim of isolating a novel DON de-epoxidation gene from Citrobacter freundii strain ADS47
(Chapter 3)
43 Materials and methods
431 Genomic DNA library construction
The genomic DNA library was prepared using a Copy Control Fosmid Library
Production Kit (Epicentre Biotechnologies Madison Wisconsin USA) following the
manufacturerrsquos instructions as depicted in Figure 4-1
105
Figure 4-1 Schematic overview of the construction and functional screening of a Fosmid
DNA library
106
Citrobacter freundii strain ADS47 was grown in NB medium at 28degC with continuous
shaking at 220 rpm The high molecular weight bacterial genomic DNA was extracted from
the overnight culture using the Gentra Puregene YeastBacteria Kit (Qiagen Inc Mississauga
Ontario Canada) The purity of the DNA was confirmed by determining OD260280 at ~190 by
a NanoDrop spectrophotometer A 5 microL sample having ~25 microg high molecular weight DNA
was sheared by passing through a 200-microL pipette tip and achieved a sufficient amount of
DNA about 40 kb in size Blunt ended 5rsquo-phosphorylated insert DNA was generated by
thoroughly mixing it with End-Repair Enzyme reagents on ice The 20 microL mixture was
incubated for 45 min at room temperature followed by incubation at 70degC for 10 min to
inactivate the End-Repair Enzyme activity
The treated DNA was size fractionated by a Low Melting Point-Pulsed Field Gel
Electrophoresis (LMP-PFGE) on the clamped homogeneous electric fields (CHEF-DR III)
system (Bio-RAD Laboratories) using 1 UltraPure Low Melting Point Agarose (Invitrogen)
The PFGE was conditioned according to manufacturerrsquos instructions as follows initial switch
time-42 s final switch time-761 s run time 16 h 50 min angle-120deg gradient- 6 Vcm
temperature-14degC and running buffer-05 TBE DNA size markers for the 42 kb Fosmid
control DNA (Epicentre Biotechnologies) and low range Pulse Field Gel (PFG) markers (New
England Biolabs Inc) were loaded adjacent to the DNA sample slots After electrophoresis a
portion of the gel that contained DNA size markers was cut off The gel was stained in SYBR
Safe DNA gel stain (Invitrogen) for 30 min visualized and marked on the 23 42 and 485 kb
marker bands A slice of PFGE gel containing 40 plusmn 10 kb size DNA was cut off with a sterile
razor blade by comparing the marker sizes DNA from the gel was recovered by digesting it
with GELase enzyme ( 1 U100 microL melted agarose) in 1X GELase Buffer followed by a
107
sodium acetate-ethanol precipitation The DNA with the desired size was stored at 4degC for
next dayrsquos ligation to a cloning-ready Fosmid vector (pCC1FOSTM
)
Ligation was performed in a 10 microL reaction mixture by thoroughly mixing 10X Fast-
Link Ligation Buffer 10mM ATP 025 microg of ~40 kb insert DNA 05 microg Fosmid Copy
Control Vector (pCC1FOS) (Figure 4-2) and 2 U Fast-Link DNA lipase The mixture was
incubated for 4 h at room temperature preceded by inactivating the ligation reaction by
heating at 70degC for 15 min The recombinant Fosmid vector was stored at -20degC for later use
Ten microL of the ligated DNA was thawed on ice and transferred to a pre-thawed
MaxPlax Lambda Packaging Extract tube containing 25 microL extract By pipetting several
times the mixture was incubated at 30degC for lamda packaging After 2 h of packaging an
additional 25 microL packaging extract was added into the mixture and incubated for a further 2 h
at 30degC The packaged phage particles were volumed to 05 mL by adding ~400 microL Phage
Dilution Buffer and 25 microL chloramphenicol
Transformation was performed via transduction by mixing the packaged particles with
the overnight cultured E coli EPI300-T1R cells (1 10 vv ratio) in LB supplemented with 10
mM MgSO4 and 02 Maltose Transformation was completed by incubation at 37degC for 1 h
The transformed E coli cells were spread on an LB agar plate containing 125 microgmL
chloramphenicol The plates were incubated overnight at 37degC Colonies that appeared on the
plates were individually transferred to 96-well microtiter plates having 200 microL LB medium
supplemented with 10 glycerol and 125 microgmL chloramphenicol The microtiter plates
were incubated at 37degC for 24 h with continuous shaking at 180 rpm The prepared library
clones were stored at -80degC for further analysis and functional screening
108
109
Figure 4-2 Genetic map of copy controlled Fosmid vector (pCC1FOSTM
) The 813 kb
Fosmid vector backbone shows a multiple cloning site (MCS) chloramphenicol resistance
selectable marker (CmR) replication initiation fragment (redF) inducible high copy number
origin (oriV) replication initiation control (repE) and F-factor-based partitioning genes (ParA
ParB ParC) The vector contains a cos site for lamda packaging (not shown) and lacZ gene
for bluewhite colony selection The vector map was provided by the Epicentre
Biotechnologies (httpwwwepibiocomitemaspid=385)
110
432 Confirmation of library construction
The quality of the DNA library prepared was determined by examining 15 E coli
clones randomly selected A 15 mL LB broth supplemented with 125 microgmL
chloramphenicol was inoculated by each of the transformed clones To obtain higher Fosmid
DNA yield 1 microL autoinduction solution (L-arabinose) to increase the copy number of the
Fosmid vector (Epicentre) was added to the cultures After an overnight incubation of the
cultures at 37degC with continuous shaking at 220 rpm Fosmid DNA was extracted using a
Qiagen Large-Construct Kit followed by restriction digestion with Not I enzyme A 20 microL
digestion mixture containing 05 microg Fosmid DNA 1X digestion buffer and 05 U Not I was
incubated at 37degC for 15 h The partially digested DNA and a 42 kb DNA size marker
(Fosmid control DNA) were ran on 08 agrose gel for 15 h at 100 V The linear Fosmid
vector backbone (813 kb) and insert DNA fragments were visualized on the gel and the
image was recorded
433 Functional screening of the clones
Eppendorf tubes (15 mL) containing 500 microL LB supplemented with 125 microgmL
chloramphenicol were inoculated with a single clone (400 clones tested) or a combination of
12 library clones (600 tested) from microtiter plates stored at -80degC One half of the cultures
in each group (single or combined clones) were induced to increase the Fosmid copy number
by adding a 1 microL (500X) autoinduction solution (contained L-arabinose) to the culture tubes
L-arabinose induces the expression of a mutant trfA gene of the E coli host strain which
leads to the initiation of Fosmid replication from a low-copy to a high-copy number origin of
replication (oriV) The cultures were spiked with 20 microgmL DON and incubated at 37degC for
111
72 h with continuous shaking at 180 rpm The DON to de-epoxy DON transformation ability
of the clone cultures was examined by a liquid chromatography-ultraviolet-mass spectrometry
(LC-UV-MS) method as described by Islam et al (2012)
112
44 Results
441 Production of the DNA library
High molecular weight intact (non-degraded) and good quality (OD600280 ~190)
genomic DNA was obtained from the DON de-epoxidizing bacterial strain ADS47 (data not
shown) Pipetting of the DNA extract resulted in an adequate amount of insert DNA (gt025
microg) of approximately 40 kb size as showed on a LMP-PFGE (Figure 4-3)
A DNA library in E coli host was constructed using LMP-PFGE purified DNA The
library was comprised of 1250 E coli transformants which were selected on LB agar plates
supplemented with chloramphenicol (Figure 4-4)
113
Figure 4-3 Preparation of a 40 kb size insert DNA from a DON de-epoxidizing bacterium
Genomic DNA was extracted from the overnight culture of a DON de-epoxidizing
Citrobacter freundii strain ADS47 After fragmentation the DNA was size fractionated by
Low Melting Point- Pulse Field Gel Electrophoresis (LMP-PFGE) with markers for the PFGE
(M1) and 42 kb Fosmid control DNA (M2) S1 S2 S3= different DNA samples extracted
from bacterial strain ADS47 The red dots area indicates the section of the gel containing
genomic DNA of the expected size (~40 kb)
114
Figure 4-4 Production of the genomic DNA library in E coli EPI300-T1 host strain
Approximately 40 kb long blunt ended genomic DNA from a DON de-epoxidizing bacterial
strain ADS47 was ligated to a Copy Control Fosmid vector pCC1FOS The construct was
packaged into lamda bacteriophage followed by a transduction-mediated transformation of E
coli host cells The clones which appeared on the LB agar plate supplemented with 125
microgmL chloramphenicol are expected to be transformed by the recombinant Fosmid vector
containing insert DNA
115
442 Conformation of the library construction
All of the clones tested acquired the Fosmid vector Out of 15 clones examined 14
received the recombinant Fosmid vector that contain the expected size insert DNA as
determined by an 81 kb vector backbone and at least one high molecular weight insert DNA
band (~40 kb) on agarose gel (Figure 4-5) In contrast one clone (lane 4) showed an 81 kb
vector backbone and a low molecular weight band which is also a fragment of the Fosmid
vector The result indicated that approximately 94 of the library clones were transformed
with the recombinant vector
443 Activity-based screening of the library
Out of 500 single copy number clones (200 individual and 300 combined clones)
none showed DON to de-epoxy DON transformation activity (Table 4-1) An additional 500
high copy number clones (200 single and 300 combined clones) were unable to de-epoxidize
DON as well
116
Figure 4-5 Confirmation of the Fosmid DNA library production in E coli Fosmid DNA
collected from 15 clones was digested with Not I and run on 08 agarose gel at 100 V for 15
h The presence of the vector backbone (81 kb) and the insert DNA fragment (~40 kb) in the
clones was determined by comparing a 42 kb molecular size marker (M) Lanes (1-15) denote
the recombinant Fosmid DNA extracted from 15 E coli clones
117
Table 4-1 Analysis of Fosmid DNA library clones for deoxynivalenol (DON) de-epoxidation
capability
Type of clones Analysis of
singlecombined
clones
Number of
clones
examined
Number of clones
showed DON de-
epoxidation activity
Without induction
(single copy number clones)
Singly 200 0
Combination of 12 300 0
With induction (multi-copy
number clones)
Singly 200 0
Combination of 12 300 0
De-epoxidation activity of the clones was examined alone or in combination of 12 clones
without or with induction by adding L-arabinose The clones grown in LB supplemented with
125 microgmL and 20 microgmL DON were analysed for DON to de-epoxy DON transformation
ability by a liquid chromatography-ultra violet- mass spectrum (LC-UV-MS)
118
45 Discussion
While a Fosmid DNA library with the expected genome coverage of a DON de-
epoxidizing bacterial genome was produced the function-derived screening of the library
clones did not identify any clones that showed DON de-epoxidation enzymatic activity This
is unlikely due to lack of coverage of the bacterial genome as 1000 clones analyzed in the
study should have provided eight fold coverage of the genome Therefore it was likely due to
a lack of expression of the enzyme(s) for DON de-epoxidizing activity under the conditions
used
Thus far there is only one report of heterologous expression of bacterial epoxide
reductase gene in another bacterium A high level of heterologous expression of a
Synechococcus sp vitamin K epoxide reductase gene was achieved in E coli with codon
optimization and placing the gene under the control of a prokaryotic T7 promoter in an
expression vector (Li et al 2010) Therefore heterologous expression of bacterial epoxide
reducatse gene using its own promoter has not yet been demonstrated The success of the
method used in this chapter relies on an intact copy of the gene or operon being cloned the
selection of an appropriate cloning vector a host organism that can recognize the cloned C
fruendii promoter and a suitable library screening method for DON degradation by E coli
(Mocali and Benedetti 2010)
The unbiased cloning efficiency of the Fosmid vector its ability to contain large insert
DNA high stability in E coli as single copy number and autoinduction to increase copy
number in host cells made it appear to be a suitable method for the present work (Kim et al
1992 Lee et al 2004) Library production with large insertions is a major advantage as gene
cluster involved in particular pathway could be trapped into single clone and their functions
119
could be analysed The pathway of microbial DON de-epoxidation activity and the genes
encoding the activity are not known Therefore it was reasonable to prepare a library with
large inserts to ensure the possibility of cloning single as well as whole pathway genes of
interest for functional screening A further advantage of the method is that Fosmid clones are
inducible which allows detecting poorly expressed genes in a heterologous host by inducing
high copy numbers (Daniel 2005)
One of the limitations of the method is that the functions of the target genes rely
mostly on the host organismrsquos transcription and translation machineries Therefore selection
of the host organism is critical for activity-based screening of library clones in heterologous
hosts A number of host strains including E coli Streptomyces lividans Rhizobium
leguminosarum and Pseudomonas aeruginosa have been used for functional screening
activities from diverse organisms (Daniel 2005) The present work attempted to isolate C
freundii strain ADS47 genes encoding DON de-epoxidation enzymes Both strain ADS47 and
E coli belong to gamma proteobacteria which suggest that E coli would be an appropriate
host organism for heterologous expression of ADS47 genes Poor expression of heterologous
genes could be another limitation of the method which may be overcome by overexpression
to increase the catalytic activity of less expressed genes However clones with DON de-
epoxidation activity were not identified in this study even the Fosmid copy number was
increased by induction
In general a large number of clones must be analysed to obtain the gene of interest
using an activity-based screening technique (Lee et al 2004 van Elsas et al 2008) In this
study the constructed DNA library comprised of 1160 clones having approximately ~40 kb
insert DNA suggesting that the prepared library achieved ~10 fold coverage of the ADS47
120
genome (48 Mb) which ensured gt99 probability of the presence of all DNA sequence in
the library In the produced library three clones (lanes 3 10 and 12) showed one intact band
greater than 42 kb in size on the agarose gel It is presumed that the Fosmid DNA of the three
samples was not digested because of insufficient amount of restriction enzyme added to the
digestion mixture About half of the samples demonstrated multiple bands suggesting that the
insert DNA might have contained multiple Not I restriction sites
However not every heterologous gene may necessarily express or function in the E
coli host (Gabor et al 2004) The functionality of the gene requires expression of the genes
and proper folding of the protein which may be influenced by environmental conditions (eg
substrate temperature and pH) and genetic factors In this study library clones were screened
in growth conditions at which strain ADS47 performed the best de-epoxidation activity The
expression of genes may be controlled by the promoter suitable ribosome binding site and
transndashacting factors (eg special transcription factors inducers chaperones cofactors proper
modifying enzymes or an appropriate secretion system) (Gabor et al 2004) Those factors
have to be supplied by the cloned DNA fragments or host cells to allow any heterologous
gene to be functional Since DON de-epoxidation genes of strain ADS47 evolved in a distinct
environmental condition the expression and activity of the gene(s) might have required
specific factorsregulators that were absent in E coli Even though the Fosmid based library
screening technique facilitated isolation of many microbial genes clones having de-
epoxidation activity were not identified using such technique in this study
121
46 Acknowledgements
We appreciate the Agriculture and Agri-Food CanadaBinational Agricultural
Research and Development Fund (AAFCBARD) project for providing the financial support
for this work The authors thank Honghui Zhu for analyzing numerous samples by liquid
chromatography ultraviolet mass spectrum method
122
CHAPTER 5
50 GENOME SEQUENCING CHARACTERIZATION AND COMPARATIVE
ANALYSIS OF CITROBACTER FREUNDII STRAIN ADS47
51 Abstract
The genome of a trichothecene mycotoxin detoxifying C freundii strain ADS47
originating from soil was sequenced and analyzed Strain ADS47 contains a circular
chromosome of 4878242 bp that encodes 4610 predicted protein coding sequences (CDSs)
with an average gene size of 893 bp Rapid Annotation Subsystems Technology (RAST)
analysis assigned functions to 2821 (62) of the CDSs while 1781 (38) CDSs were
assigned as uncharacterized or hypothetical proteins Comparative bioinformatics analyses
were done for strain ADS47 C freundii Ballerup 785139 Citrobacter sp 30_2 C youngae
ATCC 29220 C koseri ATCC BAA-895 and C rodentium ICC168 There were a significant
number of common orthologous genes (2821) were found in the six Citrobacter genomes
implying a common evolutionary relationship ADS47 is most closely related to the
opportunistic human pathogen C freundii Ballerup The human and mouse pathogenic
species C rodentium and C koseri are the most distantly related to ADS47 and non-
humananimal pathogens C youngae and Citrobacter sp 30_2 were closely related ADS47
C koseri C youngae and Citrobacter sp 30_2 lacked the clinically important locus for
enterocyte effacement (LEE)-encoded type-III secretion systems which are present in C
rodentium and C freundii Ballerup Strain ADS47 had 270 unique CDSs many of which are
located in the large unique genomic regions potentially indicating horizontal gene transfer
123
Among the unique CDSs nine were reductasesoxidoreductases and a de-acetylase that may
be involved in trichothecene mycotoxin de-epoxidation and de-acetylation reactions
Keywords Citrobacter freundii comparative genomics genome sequencing unique genes
reductases
52 Introduction
According to the Food and Agriculture Organization (FAO) at least 25 of the
worldrsquos food crops are contaminated with mycotoxins causing serious negative impacts on
livestock and international trade (Boutrif and Canet 1998 Desjardins 2006 Binder et al
2007) Trichothecenes (TCs) produced by a variety of toxigenic Fusarium species are
probably the most prevalent food and feed contaminating mycotoxins It is estimated that
infections by the fungus caused more than $1 billion in crop yield losses in USA and Canada
alone (Windels 2000 Schaafsma et al 2002 Yazar and Omurtag 2008) TC mycotoxins can
have adverse affects on human health including organ toxicity mutagenicity carcinogenicity
neurotoxicity modulation of the immune system negative reproductive system affects and
growth retardation (DrsquoMello et al 1999 Pestka et al 2007) A strain of Citrobacter freundii
ADS47 was isolated from soil in southern Ontario that is capable of detoxifying (ie de-
epoxidation and de-acetylation) several TC mycotoxins including deoxynivalenol nivalenol
and T2-toxin (Chapters 2-3)
Citrobacter freundii is a gram negative motile rod shaped facultatively anaerobic
bacterium belonging to the family of Enterobacteriaceae (Wang et al 2000 Whalen et al
2007) The opportunistic human pathogen is responsible for a number of infections including
124
nosocomial infections of respiratory tract urinay tract blood and causes neonatal meningitis
(Badger et al 1999 Wang et al 2000) C freundii exists almost ubiquitously in soil water
wastewater feces food and the human intestine Biodegradations of tannery effluents copper
aromatic amino acids by C freundii strains isolated from water mine tailings and soil
respectively have been previously reported (Kumar et al 1999 Savelieva et al 2004 Sharma
and Fulekar 2009) Because of their biochemical capabilities bacteria have played important
roles in industrial biotechnology including de-epoxidation and de-acetylation activities
(Demain and Adrio 2008 Plank et al 2009 Zhao et al 2010) However the identification of
genes for de-epoxidation activity against trichothecene mycotoxins is limited (Boutigny et al
2008)
The advent of molecular techniques such as next generation sequencing technologies
and the related bioinformatics tools has enabled the discovery of novel genes and the
enzymes they encode in pathogenic and beneficial bacteria (van Lanen and Shen 2006
Ansorge 2009 Zhao 2011) In the present study the genome of mycotoxin-detoxifying strain
C freundii ADS47 was sequenced and subjected to comparative genomic analyses with other
Citrobacter species to identify putative mycotoxin-detoxifying genes
125
53 Materials and methods
531 Sequencing quality control and assemble of ADS47 genome
Genomic DNA of the strain ADS47 was extracted using Gentra Puregene
YeastBacteria Kit (Qiagen Inc Mississauga Ontario Canada) and purity of the DNA was
confirmed by determining OD 260280 at ~190 using a Nanodrop spectrophotometer
The genomic DNA was sent to be sequenced by the Illumina HiSeq 2000 sequencing
analyzer at Beijing Genomics Institute (BGI) Hong Kong
(httpwwwbgisequencecomhomeservicessequencing-servicesde-novo-sequencing) The
sequencing workflow involved short read library preparation cluster generation sequencing
and data analysis Briefly 3-5 ug of bacterial gDNA was fragmented to produce a 500 bp
insert library by preparative electrophoresis through a 08 agarose gel The fragmented
DNAs were blunt-ended by treating with a mixture of enzymes (T4 DNA polymerase klenow
fragment) and subsequently adapters were ligated to the blunt ends Each of the DNA
fragments was amplified to a cluster which was performed in the flow cell by template
hybridization and bridge amplification Millions of clusters were then sequenced by
sequencing-by-synthesis technology leading to base calls The raw data was filtered to remove
low abundant sequences from the assembly according to 15-mer frequencies The low
complexity reads low quality (leQ20) base calls and adapter contaminations (24 bp overlap
between adapter and reads) were eliminated The quality of the sequences was analysed by
GC content and depth correlative analysis (for GC biasness) K-mer analysis was performed
using 15 bp of the short reads for heterologous sequence identification
126
The cleaned short reads (approximately 90 bp) were self-assembled using the Short
Oligonucleotide Analysis Package (SOAP) denovo (httpsoapgenomicsorgcn) (Li et al
2008) The assembly errors were corrected by filling gaps and proofreading single base pairs
using mapping information The assembly produced 441 contigs with an N50 contig size of
29222 bp (the largest contig size 114839 bp) and 80 scaffolds with an N50 scaffold size of
325726 bp (largest scaffold 662452 bp) with an total sequence length of 4878242 bp which
constitutes 33-fold genome coverage The scaffolds were ordered and oriented according to a
reference genome C koseri ATCC BAA-895 the closest completely sequenced species of
the newly sequenced bacterial genome by alignment with the MUMmer program v322
(httpmummersourceforgenet) (Delcher et al 2002) Genome coverage was determined by
mapping the reads to the assembled genome with Bowtie software (Langmead et al 2009) A
preliminary genome annotation was performed to define the origin of replication by
identifying colocalization of multiple genes ie gyrB recF dnaN and dnaA (usually found
near the origin of replication) and comparison with the C koseri reference genome Synteny
analysis between the reference bacterium and assembled scaffolds was performed with the
MUMmer program
532 Gene function annotation and classification
The genome of ADS47 was annotated by Rapid Annotation using Subsystems
Technology v40 (RAST) a server-based genome analysis system (httprastnmpdrorg)
(Aziz et al 2008) A circular genome map was constructed using CGView program
(httpstothardafnsualbertacacgview_server) (Stothard and Wishart 2005)
127
533 Molecular phylogenetic analysis
A robust phylogenetic tree was produced using Composition Vector Tree (CVTree)
web-based program v20 (httpcvtreecbipkueducn) (Qi and Hao 2004) by comparing
whole genome amino acid sequences of strain ADS47 two completely sequenced Citrobacter
genomes [C rodentium ICC168 (NC_013716) and C koseri ATCC BAA-8959 NC_009792)]
and three partially sequenced Citrobacter genomes [C freundii strain Ballerup 785139
(CACD00000000) Citrobacter sp 30-2 (NZ_ACDJ00000000) and C youngae ATCC 29220
(NZ_ABWL00000000)] E coli O157H7 strain EDL933 (NC_002655) was used as an
outgroup species The tree was constructed from the generated pairwise distance matrix data
using Neighbor-Joining Program in PHYLIP v369 package (Saitou and Nei 1987 Felsenstein
1989)
534 Comparative genomic analysis
Genome-wide comparative analyses between C freundii ADS47 C freundii strain
Ballerup 785139 Citrobacter sp 30-2 C youngae ATCC 29220 C Koseri ATCC BAA-895
and C rodentium ICC168 was carried out The nucleotide sequence idendities were
determined by pairwise alignments of the ADS47 genome with the five other Citrobacter
genomes using MUMmer software (Delcher et al 2002) and GC content total CDSs genome
coverage by CDSs and number of stable RNAs were determined by RAST program
Orthologous CDS groups among the total set of 29079 CDSs derived from the six
Citrobacter genomes were determined using OrthoMCL software v202
(httporthomclsourceforgenet) based on all-against-all reciprocal BLASTP alignment of
amino acid sequences (Li et al 2003) Genes unique to each strain were identified as those
128
CDSs not included in any orthologous group of the five other Citrobacter genomes BLASTP
comparisons were visualized with Blast Ring Image Generator (BRIG) software v095
(httpsourceforgenetprojectsbrig) (Alikhan et al 2011)
Protein sequences of the five Citrobacter genomes pooled from the GenBank data
bases were compared with ADS47 A circular map was developed by BRIG software that
showed the unique genomic regionsgenes of ADS47 The unique genes were grouped
according to their functional similarities previously identified by RAST software and
genomic regions possibly acquired by horizontal transfer were identified using RAST
software
129
54 Results
541 Assembly and organization of the ADS47 bacterial genome
Scaffolds derived from Citrobacter freundii strain ADS47 were ordered and oriented
by alignment to Citrobacter koseri ATCC BAA-895 the most closely related genome that has
been completely sequenced The genomes of ADS47 and C koseri displayed close synteny
(Figure 5-1)
Figure 5-1 Co-linearity between C freundii strain ADS47 genome and C koseri Scaffolds
derived from ADS47 genome sequences (Y- axis) were aligned and assembled by comparison
with the closest completely sequenced genome C koseri ATCC BAA-875 (X- axis)
130
542 An overview of the ADS47 genome
General features of the C freundii strain ADS47 are presented in Table 5-1 Some
notable features are that strain ADS47 contains 4881122 bp circular chromosome with an
average GC content of 5202 The genome contains 4610 CDSs which comprise 8784 of
the whole genome Annotation by RAST assigned 2821 (62) CDSs to known biological
functions and remaining 1789 (38) genes were classified to unknown or hypothetical
proteins
The GC content in the coding regions is significantly higher (5318) than in the non-
coding intergenic region (4379) which covers 1215 (592781 bp) of the genome The
intergenic region is comprised of 018 repeat sequences (total 8999 bp) 5 (009)
transposons (4806 bp) 025 stable RNA (12091 bp) and the remaining ~115 is
classified as regulatory and other functions The ADS47 genome contains 62 tRNAs 3
rRNAs and 34 copies of sRNAs The average lengths of the tRNAs rRNAs and sRNAs are
785 885 and 13441 bp respectively There are also 42 phageprophage-related genes
identified in the genome
Figure 5-2 shows the distribution of the 26 categorized CDSs rRNA tRNA GC
content and GC skew It appears that ADS47 genome contains equal quantity of genes in
leading and lagging strands and showed occasional clustering (likely operons) of genes in the
two strands The genome contains three rRNAs (two large subunit and a small subunit) and 62
tRNAs which are distributed across the genome A consistent GC content was observed in
the ADS47 genome however 7-10 low GC content (high black color peaks) genomic regions
were found at locations origin of replication 600 2000 3000 3300 3550 3850 4300 and
4800 kbp This is indicative of possible horizontally transferred genes or insertions in those
131
genomic locations GC skew denotes the excess of C over G in certain genomic locations
Positive GC skew (+) (green color) indicates the skew value in the leading strand while
negative skew (-) (purple color) in the lagging strand As in many bacterial genomes the
ADS47 genome map shows the separation of positive and negative GC skew at the origin of
replication (base pair position 1) The GC skew in the leading strand is highly skewed (high
green color peak) indicating that possible horizontal gene transfer occurred into the ADS47
genome
The CDSs ranged from ~100 bp to ~2000 bp and have an average size of 893 bp
(Figure 5-3) The highest numbers of CDS (770) were 600-800 bp long A considerable
number of genes (220) were gt2000 bp in size Some of the biological functions for the
smallest genes (100-200 bp) are sulfur carrier protein methionine sulfoxide reductase
ribonuclease P protein component while the largest genes (gt2000 bp) are ferrous iron
transport protein B translation elongation factor G pyruvate flavodoxin oxidoreductase and
tetrathionate reductase
132
Table 5-1 General features of the C freundii strain ADS47 genome
Feature Chromosome
Genome size 4881122 bp
Overall GC content 5202
Number of genes 4610
Genes with assigned function (number and ) 2821 (62 )
Genes without assigned function (number and ) 1789 (38 )
Total length of protein coding sequences 4285461 bp
Genome coverage by coding sequences () 8784
Average length of coding sequences 893 bp
GC content in the coding region 5318
GC content in the non-coding region 4379
Length and percentage of intergenic region 592781 bp and 1215
Total repeat sequences and genome coverage 8999 bp and 018
Total transposons sequences and genome coverage 4806 and 009
Number average length and genome coverage of tRNA 62 copies 785 bp and 0099
Number average length and genome coverage of rRNA 3 copies 885 bp and 0067
Number average length and genome coverage of sRNA 34 copies 13441 bp and 0093
133
134
Figure 5-2 Circular genome map of C freundii strain ADS47 Map was created with the
CGView software From outside to inside the first circle indicates the genomic position in kb
Different functional categories of predicted coding sequences (CDS) are distinguished by
colours with clockwise transcribed CDSs in the second circle and anticlockwise transcribed
CDSs in the third circle Bars on the fourth and fifth circles indicate the rRNAs and tRNAs
respectively Sixth and seventh circles show the coverage and GC content respectively The
innermost circle demonstrates the GC skew + (green) and GC skew- (purple)
135
Figure 5-3 Size distribution of the predicted genes identified in C freundii strain ADS47
The X axis identifies the gene size categories (bp) and Y axis the abundance of genes
136
543 Annotation and functional classification of ADS47 genome
RAST analysis identified predicted CDSs in the genome of Citrobacter strain ADS47
assigned functions and grouped the annotated genes into 26 functional subsystem categories
out of 27 No CDS was found belonging to the photosynthesis category A graphical
presentation of the distribution of the CDSs into each of the 26 categories is given in
Figure 5-4
A large portion (38) of the CDSs belong to hypothetical or unknown functions
Among the 62 known or predicted proteins the highest numbers (835) belong to the
carbohydrate category followed by amino acids and derivatives (468) and
cofactorsvitaminsprosthetic groups (324) A noticeable number of stress response (236) and
respiration (198) categories genes were identified in the bacterial genome The ADS47
genome encoded by 171 membrane transport genes 113 virulencedefense genes and 126
genes involved in regulationcell signaling categories Two hundred and one CDSs were
grouped into the miscellaneous category whose functions are different than the indicated
RAST categories The RAST annotations further divided the categorized CDSs into
subcategories based on similar functional roles which is discussed in section 545
137
Figure 5-4 Functional classification of the predicted coding sequences (CDS) of C freundii
strain ADS47 The annotation was performed by Rapid Annotation using Subsystems
Technology (RAST) program
138
544 Molecular phylogenetic relationships
Genome-wide phylogenetic analysis based on comparisons of whole genome amino
acid sequences of two completely sequenced Citrobacter genomes (C koseri and C
rodentium) and three partially sequenced Citrobacter genome sequences (C freundii
Citrobacter sp and C youngae) illustrates the relationships of ADS47 to five Citrobacter
species (Figure 5-5)
C youngae ATCC 29220 originating from human feces and two pathogenic species
(C koseri ATCC BAA-895 and C rodentium strain ICC168) are progressively more distantly
related to ADS47 The present analysis is consistent with the 16S-rRNA sequence based
phylogenetic analysis (Chapter 3) which showed that the closest bacterial strain to ADS47 is
C freundii (Figure 3-2) This confirmed the identification of strain ADS47 as the gamma
proteobacterial species C freundii made in Chapter 3
139
Figure 5-5 Phylogeny of the trichothecene detoxifying C freundii strain ADS47 The
dendrogram was constructed by comparing the ADS47 genome (amino acid sequences) with
five Citrobacter genomes and a E coli strain (out group species) using the CVTree software
The calculated pairwise distances matrix by Protdist program was transformed into a tree by
neighbor-joining program The dissimilarity tree was drawn to scale with branch lengths
(scale bar 002 indicated the amino acid substitutions among the genomes) The C koseri C
rodentium C freundii Citrobacter sp C youngae and E coli protein sequences were
uploaded from GenBank (Benson et al 2009)
140
545 Comparative genomics
Genome-wide comparative analyses between the genome of strain ADS47 and the five
previously sequenced Citrobacter genomes showed that 872 of the total nucleotides
between ADS47 and C freundii Ballerup were identical The values for the comparisons
between C freundii strain ADS47 were 8212 for Citrobacter sp 30-2 7637 for C
youngae 5981 for C koseri and 4213 for C rodentium (Table 5-2)
The two human and mouse pathogenic species namely C rodentium and C koseri
and the bacteria originally isolated from human feces (C youngae) have higher GC contents
(546 and 532 respectively) compared to C freundii strain ADS47 C freundii strain
Ballerup and Citrobacter sp 30_2 which have slightly lower GC contents (~52) The two
pathogenic species contain 22 copies of rRNA which is significantly higher than the 9 found
in C freundii strain Ballerup 6 in Citrobacter sp 30_2 and 3 in C freundii strain ADS47 but
less than the 25 copies in C youngae The genome of C freundii strain ADS47 C freundii
strain Ballerup C youngae and Citrobacter sp 30-2 had a similar percentage of the genome
comprising coding sequences (865-88) C koseri and C rodentium genomes contained the
highest (90) and lowest (84) genome coverage by coding sequences respectively
141
Table 5-2 Genome comparisons between strain ADS47 and five Citrobacter species The
base nucleotide sequence identities were performed using MUMmer DNAdff script program
(Delcher et al 2002) The features (GC content total CDS percentage CDS and number of
ribosomal RNA) were determined using RAST program (Aziz et al 2008)
Genome feature Bacterial species
C
freundii
ADS47
C freundii
Ballerup
Citrobacter
sp 30_2
C
youngae
C koseri C
rodentium
Genome length
(bp)
4878242
4904641
5125150
5154159
4720462
5346659
Base similarity to
ADS47 ()
NA 8715 8212 7637 5981 4213
GC () 5202 5218 516 5265 530 546
Number of coding
sequences
4610 4685 4728 5278 4978 4800
Genome coverage
with coding
sequences ()
880 865 866 872 900 840
rRNA copies 3 9 6 25 22 22
Total stable RNAs 99 81 78 105 120 142
142
The number of orthologs shared between ADS47 and other Citrobacter species is
illustrated in Figure 5-6 ADS47 shares the smallest number of orthologous CDSs with C
rodentium and C koseri 3250 and 3341 respectively The highest number of shared CDS
were found with C freundii Ballerup (3706) followed by C youngae (3693) and Citrobacter
sp 30_2 (3676) A total of 2821 CDSs were found to be common to all of the six Citrobacter
genomes analysed (data not shown) Nonetheless significant diversity exists between C
freundii ADS47 and the other species as a considerable number of unshared genes were
identified from the comparisons with C freundii Ballerup [979 (20)] Citrobacter sp 30_2
[1064 (23)] C youngae [1654 (31)] C koseri [1693 (34)] and C rodentium [1634
(34)]
143
Figure 5-6 Comparisons of coding sequences between strain C freundii ADS47 and five
Citrobacter species Coding sequences of ADS47 were compared with C freundii Ballerup
Citrobacter sp 30_2 C youngae C koseri and C rodentium genomes based on all-versus-all
reciprocal FASTA comparison of at least 30 amino acid identity and gt70 coverage of the
total length using OrthoMCL program A Venn diagram demonstrates the shared and
unshared (indicated in the brackets) coding sequences
144
Table 5-3 shows differences in the 26 RAST categorized genes between the six
Citrobacter genomes Even though a low abundance of virulencediseasedefense category
genes were found in the animalhuman pathogenic bacteria of C rodentium (85) and C koseri
(92) significantly higher mycobacterium virulence genes (13) were identified in this two
species The genomes of the two closely related strains C freundii ADS47 and C freundii
Ballerup lack this type of virulence gene Instead elevated numbers of genes for resistance to
antibiotic and toxic compounds were found in C freundii ADS47 (94) C freundii Ballerup
(88) and Citrobacter sp (110) compared to other three species Copper homeostasis and
cobalt-zinc-cadmium resistance genes were identified in strain C freundii ADS47 (13 and 21
respectively) and closely related C freundii Ballerup (12 and 22) and Citobacter sp (18 and
22) Significantly lower numbers of these genes were present in C rodentium (6 and 4) and C
koseri (7 and 8) C rodentium and C koseri lack mercuric reductase mercuric resistance and
arsenic resistance genes perhaps because being animalhuman pathogens but these genes
were found in the remaining four genomes The C koseri genome is richer in betandashlactamase
genes (5) compared to the remaining genomes which encodes 1-3 genes
145
Table 5-3 Functional categorized gene abundances among the genomes of C freundii
ADS47 C freundii Ballerup Citrobacter sp 30_2 C youngae C koseri and C rodentium
The genomes were classified into different functional categories with the RAST program
which classified the total CDSs
RAST subsystem category Citrobacter species
C
freundii
ADS47
C
freundii
Ballerup
C
sp
C
youngae
C
koseri
C
rodentium
Cofactorsvitaminsprosthetic
groupspigments
323
317
310
311
325
304
Cell wall and capsule 279 272 248 262 247 262
Virulence disease and defense 113 107 134 100 92 85
Potassium metabolism 50 53 33 32 30 28
Miscellaneous 210 228 200 185 245 185
Phageprophagetransposable
element plasmid
24 75 25 4 6 49
Membrane transport 170 156 171 195 171 172
Iron acquisition and metabolism 80 81 86 62 113 27
RNA metabolism 220 222 217 215 238 212
Nucleosides and nucleotides 119 112 110 111 120 114
Protein metabolism 249 280 269 298 287 254
Cell division and cell cycle 30 29 32 29 38 29
Motility and chemotaxis 133 124 90 147 143 133
Regulation and cell signaling 126 130 124 135 132 123
Secondary metabolism 21 22 20 21 0 0
DNA metabolism 145 167 165 155 147 152
Fatty acids lipids and isoprenoids 121 149 122 121 134 133
Nitrogen metabolism 50 52 53 49 56 58
Dormancy and sporulation 3 3 3 4 3 3
146
Table 5-3 (continued)
RAST subsystem category
Citrobacter species
C
freundii
ADS47
C
freundii
Ballerup
C
sp
C
youngae
C
koseri
C
rodentium
Respiration 198 184 180 191 166 162
Stress response 236 235 170 177 179 174
Metabolism of aromatic
compounds
20 20 28 25 17 48
Amino acids and derivatives 482 476 450 466 428 416
Sulfur metabolism 48 50 52 57 56 56
Phosphorus metabolism 50 49 43 42 43 28
Carbohydrates 837 803 708 731 733 737
147
Strains C freundii ASD47 and C freundii Ballerup have much more potassium
metabolism category genes (50 and 53 respectively) than the other bacterial species which
code for 28-33 proteins Larger numbers of genes associated with the glutathione regulated
potassium efflux system were also identified in C freundii ASD47 with 23 and C freundii
Ballerup with 26 compared to the remaining four species that encode only five genes All of
the six genomes contained phageprophage-related genes with the maximum identified in C
freundii Ballerup (75) followed by C rodentium (49) Citrobacter sp (25) and C freundii
ADS47 (24) Lower numbers of phageprophage-related genes were found in C koseri (6) and
C youngae (4)
The six Citrobacter genomes showed remarkable variability in their secretion systems
The type-III secretion operon genes are only found in C rodentium and C freundii Ballerup
Type-II secretion genes are predominant in C rodentium (12) followed by ADS47 (9)
Type-IV secretion system is uncommon in Citrobacter species since it was only
identified in Citrobacter sp where it is represented by 19 genes Variable numbers of type-VI
secretion system genes are present in the five genomes except C koseri which lacks such
genes C youngae has the highest number (43) of type-VI secretion proteins The other strains
have a range of type-VI secretion proteins in particular 27 were identified in C freundii
Ballerup 15 in C freundii ADS47 and 8 in Citrobacter sp Type-VII (chaperoneusher)
secretion system genes were found in all of the six Citrobacter species with the largest (42)
and smallest (13) numbers of these genes occurring in C koseri and C freundii Ballerup
respectively
Although a relatively similar number of ABC transporters (37-42) were identified in
the six genomes C freundii ADS47 contain the largest number of tripartite ATP-independent
148
periplasmic (TRAP) transporter genes (7) By contrast the closely related C freundii
Ballerup lacks such transporter system An identical number of iron acquisition and
metabolism category genes were found in C freundii ADS47 (80) and C freundii Ballerup
(81) while the two pathogenic species C rodentium and C koseri code for the lowest (27)
and highest (113) numbers of such genes C koseri is rich in siderophore related genes (64)
but the remaining five genomes have smaller numbers of siderophore-associated genes (20-
22)
Variable numbers of motility and chemotaxis related genes were found in the
Citrobacter genomes with the largest number occurring in C youngae (147) followed by C
koseri (143) C freundii ADS47 (133) C rodentium (133) C freundii Ballerup (124) and
Citrobacter sp (90) The two pathogenic species (C rodentium and C koseri) encode a
remarkably low number of quorum sensing genes (2) In contrast C youngae codes for the
highest number of quorum sensing genes (19) A common and distinctive feature of the two
pathogenic species is the lack of genes required for secondary metabolism In contrast the
non-pathogenic species contain a common set of genes for degrading a range of
phenylpropanoids and cinnamic acid (20-22)
The lowest number of genes related to phospholipidsfatty acids biosynthesis and
metabolism were found in C freundii ADS47 (66) compared to C freundii Balllerup (94) and
C koseri (98) The smallest numbers of nitratenitrite ammonification genes were found in
ADS47 (31) and its closest genome C freundii Ballerup (32) In contrast the two pathogens
encode the highest number of such proteins (39-40)
C rodentium and C koseri code for significantly low numbers of respiratory category
genes (162-165) while C freundii ADS47 contains the highest number of such genes (198)
149
Further analysis of the subcategory genes determined that the two gut species (C youngae and
Citrobacter sp 30_2) contain the largest numbers of genes encoding anaerobic reductases
(22) and identical numbers of formate dehydrogenases (17) The largest numbers of formate
dehydrogenase genes were identified in C freundii Ballerup (32) and C freundii ADS47 (30)
but the two pathogenic species differed from the other species by encoding the lowest number
(13) of c-type cytochrome biogenesis genes
A higher number of aromatic compound metabolism category genes were found in the
mouse pathogen C rodentium (48) compared to the human pathogen (C koseri) which
contain 17 genes A significantly higher number of genes involved in central aromatic
intermediate metabolism (26) were found in C rodentium compared to the mouse pathogen
(C koseri) and other genomes which only contain 6-8 such genes The two C freundii strains
ADS47 and Ballerup lacked the anaerobic degradation of aromatic compound
(hydroxyaromatic decarboxylase family) genes
In spite of the abundance of stress response category genes strain ADS47 has only a
small number of desiccation stress genes (5) compared to the other species which ranged
from 9 to10 However ADS47 and Ballerup code for higher number of genes related to
oxidative stress (94 and 91 respectively) and cold shock (8 and 7 respectively) than the other
species
The smallest numbers of amino acids and its derivatives category genes were found in
the two pathogens C rodentium and C koseri (416 and 428 respectively) while the largest
number of these genes were identified in strain ADS47 (482) and C freundii Ballerup (476)
Despite the similar number of sulfur metabolism category genes determined across the six
species only ADS47 lacks taurine utilization genes Strains ADS47 and Ballerup were found
150
to be rich in the phosphorous metabolism gene category (49-50) while C rodentium contains
a significantly decreased number of these category genes (28)
The bioinformatics analyses found the largest number of carbohydrate category genes
in ADS47 (837) followed by Ballerup (803) The gut bacteria (Citrobacter sp 30_2) codes for
the least number of genes assigned to the carbohydrate category (708) while C koseri C
rodentium and C youngae contained 731-737 in this category genes The analyses also
determined that there are relatively small numbers of TCA and Entner Doudoroff pathway
genes in C rodentium and C youngae compared to the other four species and relatively large
numbers of fermentation-related genes in ADS47 (81) and C freundii (85) compared to the
other four species
A comparison of the pathogenicity operons [locus of enterocyte effacement (LEE)] of
C rodentium with other five Citrobacter genomes revealed the absence of this virulence
determinant gene region in the other four Citrobacter species including ADS47 (Figure 5-7)
The analysis showed the presence of LEE operon genes in C freundii strain Ballerup as no
gap was observed in that genomic location while C freundii ADS47 C koseri C youngae
and Citrobacter sp 30_2 lacked that loci (7a) Further ananlysis showed alignments between
C rodentium and E coli LEE operon genes (Figure 7b) In contrast no alignment of such
genes was observed in ADS47 genome
151
(a)
152
Figure 5-7 (a) Blast Ring Image Generator (BRIG) output based of Citrobacter rodentium
(NC_013716) compared against ADS47 C freundii C koseri C youngae E coli and
Citrobacter sp genomes The targeted comparative genome analysis was performed based on
locus of enterocyte effacement (LEE) operons of C rodentium (Petty et al 2011) The
innermost rings show GC content (black) and GC skew (purple and green) The remaining
rings show BLAST comparisons of the other Citrobacter and E coli genomes against C
rodentium The region shown in red is LEE operon genes (b) The absence of LEE operons in
the ADS47 genome comparing to C rodentium and E coli Analysis was performed and
visualized by Artemis Comparison Tool software v132
(httpwwwsangeracukresourcessoftwareact) (Tatusov et al 2000 Craver et al 2005)
LEE operons
(b)
153
546 Analysis of the unique genes in C freundii ADS47
Two hundred and seventy genes unique to ADS47 were identified from a comparison
of its genome to the other five Citrobacter species In general the unique genes are
distributed throughout the ADS47 genome but some clusters of unique genes occurred
including eight unique genomic regions (Figure 5-8) Six large unique regions are located at
767204-775090 (~8 kb) 1025572-1042510 (17 kb) 2798289-2807217 (~9 kb)
3045220-3055912 (~11 kb) 3930366-3943075 (12 kb) and 4022195-4032444 (10 kb)
Out of 270 unique genes 147 (54 ) are hypothetical proteins The remaining 123
(46) genes are grouped into reductasesoxidoreductases (9 genes) de-acetylase (1)
hydrolases (2) transferases (4) sulfatasesarylsulfatases (3) bacteriocins (3) type VI
secretion proteins (2) kinases (4) transporters (11) membraneexported proteins (8)
regulators (12) integraserecombinases (5) phagesprophagestransposaes related genes (27)
and miscellaneous (32) that includes DNA helicases ligases fimbrial proteins and
chromosome partitioning genes (Table 5-4)
The unique regions encode various functional genes that may cause trichothecene de-
epoxidation or de-acetylation reactions The 8 kb unique regions contains six genes encoding
glycosyltransferases and two copies of oxidoreductase-like coding sequences (CMD_0711
and CMD_0712) that showed 39 identity to the Marinobacter aquaeolei F420
hydrogenasedehydrogenase beta subunit The largest (17 kb) unique genomic region codes
for 16 genes including dehydrogenase hydrolase bacteriocin (pyocin) integrase and a short
chain dehydrogenasereductase-like (SDR) (CMD_0979) gene (Figure 5-9a)
154
The SDR like gene is 49 amino acid similarity to an oxidoreductase of the
phytopathogenic Dickeya dadantii and the enzyme contains a domain with 77 homology to
the Dickeya zeae NAD(P) binding site The 9 kb unique genomic section contains 8 genes
including a NaCl symporter a dehydratase (CMD_2679) with 86 homology to
Enterobacteriaceae bacterium dihydroxy-acid dehydratase and a putative oxidoreductase
(CMD_2683) having 37 similarity with oxopropionate reductase of Bacillus sp The unique
region also encodes a short chain dehydrogenasereductase (SDR) (CMD_2684) that shows
85 identity to Enterobacteriaceae bacterium (Figure 5-9b) A unique thiol-disulfide and
thioredoxin like reductase (CMD_3721) was identified in a 10 kb unique region that contains
13 genes (Figure 5-9c) This gene is 59 identical to Oligotropha carboxidovorans thiol-
disulfide isomerase and thioredoxin In addition several reductasesoxidoreductase-like
unique genes were identified in the ADS47 genome including an oxidoreductase
(CMD_1024) with 27 identity to Canadidatus nitrospira NADH-quinone oxidoreductase
membrane L Another oxidoreductase-like gene identified in a different location of the
ADS47 genome (CMD_1393) shows 87 homology to Klebsiella pneumoniae
deoxygluconate dehydrogenase contains a SDR active site A putative nitrate reductase gene
(CMD_3184) has a low level of identity (27) with an uncultured bacterial respiratory nitrate
reductase The ~11 kb unique region encodes sulfatasesarylsulfatses
(CMD_2898CMD_2905) and related genes Furthermore ADS47 contains a unique
glycosylphosphatidylinositol (GPI) anchor biosynthesis operon comprised of four genes
including a N-acetylglucosaminyl-phosphatidylinositol de-acetylase (CMD_2431) which
showed 78 similarity to E coli de-acetylase (Figure 5-9e)
155
Other unique genespredicted genes in the ADS47 genome include
polysaccharidephenazine biosynthesis genes (CMD_0710CMD_1406) heavy metal kinase
(CMD_0989) two component heavy metal regulator (CMD_0990) nitrate reductase regulator
(CMD_3477) putative potassiumzinc transporters (CMD_0973CMD_4600) sodium
symporter (CMD_2678) (tripartite ATP-independent periplasmic transporters) TRAP
transporters (CMD_3693 CMD_3694 CMD_3695 CMD_4337) glutamineribose ABC
transporters (CMD_3176CMD_3544) toxin transcription regulator (CMD_3464) fimbrial
proteinchaperone (CMD_1695 CMD_2434 CMD_3277 CMD_3278) and ParB
chromosomeplasmid partitioning proteins (CMD_4033CMD_4034)
Majority of the unique genes and their distributions are shown in comparative circular
genome maps (Figure 5-8)
156
Table 5-4 Unique genes of C freundii strain ADS47 Genes were categorized according to
the functional similarity The unique genes were determined by comparing ADS47 with five
sequenced Citrobacter species using OrthoMCL software
Unique gene classes Number
Total unique genes
270
Reductases oxidoreductases 9
De-acetylase 1
Hydrolase 2
Transferase 4
Sulfatase 3
Bacteriocin 2
Type VI secretion 2
Kinase 4
Transporter 9
Membraneexported protein 8
Regulator 12
Integraserecombinase 5
Phageprophagetransposae 27
Miscellaneous 34
Hypothetical protein 147
157
158
Figure 5-8 Locations of the unique genes of C freundii strain ADS47 The unique genes
were determined by comparing the total CDS of strain C freundii ADS47 with C freundii
Ballerup Citrobacter sp C youngae C koseri and C rodentium CDSs by OrthoMCL
program The map was developed by Blast Ring Generator (BRIG) software First second
and third circles from inside to out side show the genome size GC and GC skew of strain
ADS47 Forward (Fwd) and reversed (Rev) transcribed CDSs of ADS47 are shown in green
(forth circle) and red (fifth circle) respectively The next five rings represent the circular
genome maps of five Citrobacter species distinguished by different ring colours (C
rodentium as sky blue C koseri as dark blue C youngae as light blue C freundii Ballerup as
yellow and Citrobacter sp as green) The degree of orthologous gene identity between
ADS47 and other five genomes are shown by color intensity (light color= 75 gene identity
ash color= 50 gene identity) Gaps across five genome rings indicate the absence of
genesregions compared to ADS47 Unique genes are shown surrounding the map rings and
the identities of some of the genes are given Arrows indicate putative
reductasesoxidoreductases and de-acetylase
159
(a)
(b)
160
(c)
(d)
161
Figure 5-9 Unique genomic regions and associated genes in C freundii ADS47 compared to
five Citrobacter species RAST server based analysis compared the unique genesregions of
ADS47 with the publicly available databases The upper panels demonstrate the large unique
regions defined by yellow boxes and genes located in the genomic regions are indicated by
arrows The lower panels are closely related genesoperons found in distantly related species
The unique regions that contain potential trichothecene de-epoxidation genes are shown in (a)
short chain dehydrogenasereductase (SDR) and dehydrogenase (b) putative oxidoreductase
dehydratase and SDR (c) thiol-disulfide isomerase and thioredoxin oxidoreductase and (d)
unique glycosylphosphatidylinositol (GPI) biosynthesis operon having N-acetylglucosaminyl-
phosphatidylinositol de-acetylase gene
162
55 Discussion
The de novo sequenced ADS47 genome was oriented and assembled using a
completely sequenced C koseri genome as reference strain The synteny analysis between
ADS47 and C koseri showed very similar overall organization of the two genomes The high
preservation of genes among five Citrobacter species and ADS47 were emphasized by the
close alignments observed by Blast Ring Image Generator (BRIG) outputs obtained from
these genomes
Genome-wide phylogenetic analyses based on a large number of common CDSs
(2821) observed within the six Citrobacter genomes confirmed the species identification
based on 16S-rDNA sequencing of the trichothecene mycotoxin-detoxifying C freundii strain
ADS47 The base sequence idendity was 8715 between these two genomes They shared
~80 CDS and had a similar GC content (52) Citrobacter are Gram-negative rod shaped
and facultatively anaerobic bacteria belonging to the family of Enterobacteriaceae (Wang et
al 2000 Whalen et al 2007) Strains ADS47 and Ballerup of C freundii are both aerobic
gram-negative bacilli that have numerous flagella and inhabit in soil (Cong et al 2011 Islam
et al 2012) C freundii is considered to have a positive environmental role by reducing nitrate
to nitrite (Rehr and Klemme 1989) which is an important step in the nitrogen cycle
C freundii C rodentium and C koseri are animalhuman pathogenic species C
rodentium is a natural mice pathogen causing hemorrhagic colitis and hemolytic uremic
syndrome (Gareau et al 2010) C koseri causes human disease (Badger et al 1999 Kariholu
et al 2009) particularly neonatal meningitis and C freundii is an opportunistic human
pathogen responsible for a various types of infections like respiratory tract urinary tract and
blood (Whalen et al 2007) C freundii is very common representing approximately 29 of
163
all opportunistic human infections (Hodges et al 1978) Outside of animals they can be found
in soil water sewages and different organs of diseased animals (Wang et al 2000) These
pathogens code for large numbers of mycobacterium virulence operonsgenes which were
absent in the three other genomes However they have fewer genes involved in environmental
stress such as copper homeostasis genes and no mercuricarsenic resistance and secondary
metabolism category genes unlike the other Citrobacter genomes Nonetheless considerable
variability exists between the two pathogens correlates with C rodentium causing disease in
mice C koseri causing meningistis in human and C freundii causing infections in different
human organsThere were attaching and effacing (AE) lesion forming (LEE) operons in C
rodentium and C freundii but not in C koseri Those genes have shown to be acquired by
lateral gene transfer playing major roles in virulence of enteropathogenic and
enterohemorrhagic E coli and C rodentium (Mundy et al 2005 Petty et al 2010 Petty et al
2011 The ~356 kb pathogenic LEE gene island is comprised of five operons (LEE1 to
LEE5) that code for ~40 virulence genes that include type-III secretion genes and associated
effectors (Berdichevsky et al 2005) In addition an elevated number of type-II and -VI
secretion proteins identified in C rodentium which may act as additional virulence factors
(Petty et al 2010) Because it lacks type-III secretion apparatus genes it is likely that C
koseri uses a different strategy to cause human neonatal meningitis (Badger et al 1999 Ong
et al 2010)
To adapt to different environmental niches or hosts bacteria like Citrobacter species
frequently gainlose genes increase repeat sequences and recombine genomic regions using
different mechanisms (Parkhill et al 2001 Petty et al 2011) The existence of large numbers
of phageprophagetransposae related genes in the Citrobacter genomes particularly in C
164
freundii Ballerup suggests that their genomes are unstable For example C freundii Ballerup
and C koseri might have acquired LEE operons by horizontal transfer via phage mediated
transpositions Interestingly although ADS47 is closely related to C freundii Ballerup it
lacks the pathogen-related LEE gene island and mycobacterium virulence operons required
for invasion and intracellular survival in macrophages (Gao et al 2006) Instead ADS47 has a
larger number and variety of stress-responsive genes such as efflux genes and bacteriocins
(pyocin hemolysin) genes that provide protection against heavy metals and antibiotics and
increase their competitiveness against other bacteria These findings suggest that the strain
ADS47 is not an opportunistic animal pathogen but has evolved to be highly competitive in
the soil micro-environment (Arnold 1998 Bengoechea and Skurnik 2000) Perhaps ADS47
evolved trichothecene degrading genes for adaptation to soils containing trichothecene
mycotoxins contaminated soils (Gadd and Griffiths 1978 Sharma and Fulekar 2009 Han et
al 2011) Conversely the absence or reduced number of stress-responsive genes in C
rodentium and C koseri may imply that those two pathogenic species are not exposed to such
stressful and xenobiotic-contaminated environments as ADS47
This interpretation is supported by the observation that the two of the three
animalhuman pathogenic species contained high of rRNA copy number but lower numbers
were found in ADS47 C freundii Ballerup and Citrobacter sp 30_2 Bacteria maintain large
number of rRNA for higher production of ribosome during high growth rates (Stevenson et al
1997 Klappenbach et al 2000) such as might occur during the establishment of an infection
However a large copy number of rRNAs may not be metabolically favourable for bacteria
such as ADS47 that evolved in a highly competitive environment as that costs higher cellular
energy (Hsu et al 1994)
165
C freundii Ballerup seems to be an exception in that it is similar to the other animal
pathogens for containing virulent factors (ie LEE-operon genes) but more like the
environmental species for encoding high number genes for stress defence heavy metal and
toxic compounds resistances
The comparison of the genes in C freundii ADS47 with the unique ability to detoxify
trichothecenes with genes in other Citrobacter species allowed approximately 270 unique
genes to be identified in ADS47 Some of those genes may be involved in trichothecene de-
epoxidation or de-acetylation Currently the pathways and microbial enzymes leading to
trichothecene de-epoxidation and de-acetylation are unknown but based on the required
reaction ie the elimination of an oxygen atom from epoxide moiety it is likely that epoxide
reductases or epoxide oxidoreductases are responsible for such biodegradation activity (He et
al 1992)
From database searches three enzymes having epoxide reductase activity were found
These are a vitamin K epoxide reductase (VKOR) a monooxygenase PaaABCE and pyridine
nucleotide-disulfide oxidoreductase (PNDO) which reduce vitamin K epoxide phenylacetyl-
CoA and epoxyalkanes respectively (Swaving et al 1996 Li et al 2004 Teufel et al 2012)
In mammals VKOR plays important roles in vitamin K cycle by reducing vitamin K epoxide
to vitamin K and its homologues that are found in plants bacteria and archaea (Goodstadt
2004 Li et al 2010) The microbial multi-component PaaABCE monooxygenase catalyzes
degradation of variety of aromatic compounds or environmental contaminants via epoxidation
of phenylacetic acid (Paa) and NADPH-dependant de-epoxidation of epoxy-Paa (Luengo et
al 2001 Teufel et al 2012) PNDO is a unique type of NADP-dependent oxidoreductase
identified in Xanthobacter strain Py2 that was obtained by propane enrichment procedure
166
(van Ginkel and de Bont 1986) However no orthologs of these genes were found in ADS47
genome
Nine ADS47-unique reductaseoxidoreductase-like genes are potential candidates for
the trichothecene de-epoxidation enzyme Genes belong to short chain
dehydrogenasereductase (SDR) family are structurally and functionally diverse and cause
different enzymatic activities including xenobiotic degradation (Kavanagh et al 2008) This is
a promising candidate for the trichothecene de-epoxidation gene Genes belong to dehydratase
family catalyze reduction reactions by cleaving carbon-oxygen bonds of organic compounds
that result in removal of oxygen in various biosynthetic pathways (Flint et al 1993) This
mechanism may be relevant to microbial trichothecene de-epoxidation reaction since the latter
involves removal of the oxygen atom forming the epoxide in trichothecene mycotoxins
(Young et al 2007)
Another ADS47-unique gene was for a nitrate reductase that may be a candidate de-
epoxidation gene Under stress conditions such an enzyme causes reductive removal of an
oxygen molecule (Morozkina and Zvyagilskaya 2007)
A fourth type of ADS47-unique gene is for a thiol-disulfide isomerasethioredoxin like
(Trx) oxidoreductase The thiol-disulfide isomerase and thioredoxin oxidoreductases contain a
thioredoxin like domain with a CXXC catalytic motif which catalyzes various biological
processes that allow the organism to adapt to environmental stress and develop competence to
sporulate (Scharf et al 1998 Petersohn et al 2001 Smits et al 2005) The membrane-
anchored Trxndashlike domain initiates the vitamin K de-epoxidation reaction by reducing the
active motif (CXXC) of VKOR (Schulman et al 2010)
167
The present study did not find any homologues of VKOR in ADS47 genome but we
identified two copies of structurally similar quinone reductase protein disulfide bond
formation (DsbB) gene (CMD_1044 and CMD_2641) Both VKOR and DsbB contain four
transmembrane segments an active site CXXC motif and cofactor quinone and use a
common electron transfer pathway for reducing substrates (Li et al 2010 Malojcic et al
2008) As VKOR bacterial DsbB requires a Trx-like partner (eg DsbA) for activity Further
experiments are required to examine if DON de-epoxidation occurs by the catalytic activity of
DsbB coupled with a unique thiol-disulfide isomerase and thioredoxin oxidoreductase present
in ADS47 genome
The analysis also identified an ADS47-unique glycosylphosphatidylinositol (GPI)
biosynthetic operon that is comprised of four genes including an N-
acetylglucosaminylphosphatidylinositol de-acetylase The GPI membrane anchor protein
synthesis is initiated by transferring N-acetylglucosamine from uridine diphosphate N-
acetylglucosamine to phosphatidylinositol by N-acetylglucosaminyltransferase an
endoplasmic reticulum (ER) membrane-bound protein complex (Hong and Kinoshita 2009)
In the second step of the pathway N-acetylglucosamine-phosphatidylinositol is de-acetylated
to produce glucosaminyl-phosphatidylinositol by the catalytic activity of a zinc
metalloenzyme N-acetylglucosaminylphosphatidylinositol de-acetylase which is localized on
the cytoplasmic side of the ER (Nakamura et al 1997 Hong and Kinoshita 2009) It is
possible that the identified unique N-acetylglucosaminylphosphatidylinositol de-acetylase
gene of ADS47 may be involved in trichothecene de-acetylation activity
168
Further analysis is needed to determine which of these candidate gene or genes may be
involved in tricthothecene mycotoxin de-epoxidation and de-acetylation activities
Identification of gene responsible for the trichothecene detoxification activities of C freundii
ADS47 opens many potential applications for its use in agri-based industries to contribute to
cropanimal production and food safety (Karlovsky 2011)
56 Acknowledgement
We gratefully acknowledge Agriculture and Agri-Food CanadaBinational
Agricultural Research and Development Fund (AAFCBARD) project for providing the
financial supports of the work
169
Supplemental Table S5-1 List of unique genes of strain ADS47 gene IDs and their
functional roles The unique genes were determined by comparing the ADS47 genome with
five sequenced Citrobacter species (C freundii Citrobacter sp C youngae C koseri and C
rodentium) using OrthoMCL (Li et al 2003)
Unique gene
classes
Number Gene ID and functions
Total unique
genes
270 123 known functions 147 hypothetical proteins
Reductases
oxidoreductases
9 CMD_0711 coenzyme F420 hydrogenasedehydrogenase beta subunit
CMD_0979 Short chain dehydrogenase reductase (SDR) Oxidoreductase
SDR dehydrogenase CMD_1024 NADH-quinone oxidoreductase
membrane L CMD_1393 oxidoreductase SDR putative deoxygluconate
dehydrogenase (NAD+ 3-oxidoreductase) CMD_2679
Dehydratasedihydroxy acid dehydratase CMD_2683 oxidoreductase
oxopropionate reductase CMD_2684 short-chain dehydrogenasereductase
SDR CMD_3184 nitrate reductase alfa subunit CMD_3721 thiol-disulfide
isomerase and thioredoxin oxidoreductase
Deacetylase 1 CMD_2431 N-acetylglucosaminylphosphatidylinositol deacetylase
superfamily
Hydrolase 2 CMD_0991 Dienelactone hydrolase CMD_2895 Putative glycosyl
hydrolase
Transferase 4 CMD_1640 D12 class N6 adenine-specific DNA methyltransferase
CMD_2432 Glycosyltransferase CMD_2433 putative acetyltransferase
CMD_3023 adenine-specific DNA modification methyltransferase
Sulfatase 3 CMD_2898 Possible sulfatase arylsulfatasearylsulfatransferase
CMD_2902 Arylsulfatasesulfatase CMD_2905 Putative
sulfatasearylsulfatase
Bacteriocin 3 CMD_0982 Thermosatble hemolysin superfamily CMD_0993 Pyocin S2
superfamily (bacteriocin that kills other bacterial strain) CMD_3705 S-type
pyocin containing domain protein
Type VI secretion 2 CMD_1802 EvpB family type VI secretion protein CMD_2364 type VI
effector Hcp1
Kinase 4 CMD_0989 Heavy metal Sensor kinase CMD_1794 Polo kinase kinase
(PKK) superfamily CMD_3124 predicted diacylglycerol kinase CMD_4266
putative signal transduction kinase
Transporter 11 CMD_0973 TrkA-N domain-containing protein (K uptake transporter)
CMD_1407 Branched-chain amino acid transport system carrier protein
CMD_2678 Sodium Symporter CMD_3176 glutamine ABC transporter
CMD_3544 Ribose ABC transport system CMD_3693 TRAP-type C4-
dicarboxylate transport system binding protein CMD_3694 TRAP-type C4-
dicarboxylate transport system binding protein small permease component
(predicted N-acetylneuraminate transporter) CMD_3695 TRAP-type C4-
dicarboxylate transport system large permease CMD_4337 23-diketo-L-
gulonate TRAP transporter small permease protein yiaM component
CMD_4338 23-diketo-L-gulonate TRAP transporter large permease protein
yiaN CMD_4600 putative transporter involved in Zinc uptake (YciA
protein)
170
Supplmental Table S5-1 (continued)
Unique gene
classes
Number Gene ID and functions
Membrane
exported protein
8 CMD_0713 putative membrane protein wzy CMD_0983 Putative exported
protein precursor CMD_0986 Putative transmembrane protein
CMD_0987 putative exported protein precursor CMD_1458 putative
exported protein CMD_2903 Putative exported protein CMD_2904
putative secreted 5-nucleotidase CMD_4291 (S) putative integral
membrane export protein
Regulator 12 CMD_0977 probable transcriptional regulator protein CMD_0980 Putative
transcription activator TenA family CMD_0990 Two component heavy
metal response transcription regulator CMD_1766 Leucine responsive
regulatory protein CMD_2677 transcriptional regulator GntR CMD_2909
putative arylsulfatase regulator CMD_3279 Uncharacterized outer
membrane usher protein yqiG precursor CMD_3464 toxin transcription
regulatory protein CMD_3465 EAL regulator protein (signaling)
CMD_3475 putative transcription regulator CMD_3477 Fumarate and
nitrate reduction regulatory protein (for anaerobic adaptation) CMD_3812
putative regulator from phage
Integrase
recombinase
5 CMD_0785 CMD_0992 CMD_0994 CMD_4032 CMD_4424
Phage prophage
transposae related
genes
27 Phages CMD_1608 CMD_1609 CMD_1610 CMD_1611 CMD_1619
CMD_1631 CMD_1913 CMD_1914 CMD_1915 CMD_1917
CMD_1920 CMD_1921 CMD_1936 CMD_3351 CMD_3804
CMD_3805 CMD_3807 CMD_3808 CMD_4581 CMD_4609
CMD_4610 Transposae CMD_0252 CMD_0398 Prophage CMD_1645
CMD_3806 CMD_4425 CMD_4597
Miscellaneous 32 CMD_0095 ATP-dependent DNA helicase Rep CMD_0656 LysA protein
CMD_0710 Polysaccharide biosynthesis family protein CMD_0978 TPR
repeat containing protein CMD_0981 AMP dependant synthatase and
ligase CMD_0985 Cupin superfamily CMD_0987 flavodoxin
CMD_1404 Putative endoribonuclease CMD_1405 Putative PAC domain
protein CMD_1406 Phenazine (PhzF) biosynthesis protein CMD_1695
Putative fimbrial protein CMD_1793 HPChromosome segregation ATPase
domain protein CMD_1922 FRG uncharacterized protein domain
CMD_2206 L-fucose isomerase like protein CMD_2434 CS12 fimbria
outer membrane usher protein CMD_2680 HpcHHpaI aldosecitrate lyase
protein family CMD_2681 Aldose -1 epimerase CMD_2682
Gluconolaconase LRE like region CMD_2900 Putative cytoplasmic
protein CMD_3009 putative restriction endonuclease CMD_3277 Putative
yqi fimbrial adhesion CMD_3278 Hypothetical fimbrial chaperone yqiH
CMD_3340 Pea-VEAacid family CMD_3518 3-hydroxy-3-methylglutaryl
CoA synthase CMD_3801 putative reverse transcriptase CMD_3840 beta-
D-galactosidase alpha subunit CMD_4033 ParB family chromosome
partitioning protein CMD_4034 ParBrepB chromosomeplasmid
partitioning protein CMD_4035 RelASpoT protein CMD_4532 putative
DNA ligase -like protein CMD_4541 Antitermination protein Q and
CMD_4585 putative ATP dependent helicase
171
CHAPTER 6
60 CONCLUSION
61 Summary
Contamination of grain with mycotoxins including deoxynivalenol (DON) and other
trichothecenes is becoming a limiting factor for cereal and animal production (Binder et al
2007) To address the issue this study obtained a novel microbial source for DON
detoxification and determined candidate genes for the function which may have potential
applications to detoxify grain and grain-derived foodfeed contaminated with DON and TC
mycotoxins
From a screen of soil samples from crop field microorganism was isolated that is able
to de-epoxidize the most frequently detected foodfeed contaminating mycotoxin DON The
effects of media compositions temperature pH and aerobicanaerobic conditions on
microbial detoxification activity were assessed From 150 analyzed soil samples a mixed soil
microbial culture that showed DON de-epoxidation activity under aerobic conditions and
moderate temperature (~24degC) was obtained This culture exhibited a slow rate of microbial
DON to de-epoxyDON transformation (de-epoxidation) possibly because of competition
between de-epoxidizing and other microorganisms in the culture
The mixed microbial culture was enriched for DON detoxifying bacteria by reducing
the non-targeted microbial species through antibiotic treatments heating and subculturing on
agar medium and selecting colonies The enriched culture had higher DON de-epoxidation
activity than the initial culture Because of low abundance of DON detoxifying
microorganisms further culture enrichments were required to increase the target microbial
172
population Reculture of the partially enriched microbial community in nutritonally poor MSB
medium with high level of DON increased the DON de-epoxidizing bacterial population
Serial dilution plating and selecting single colonies eventually led to isolation of a DON-
detoxifying bacterial strain named ADS47
Phenotypic and 16S-rDNA sequence analyses provided evidence that the bacterium is
a strain of C freundii Strain ADS47 is a gram negative rod-shaped facultative anaerobe and
has peritrichous flagella Strain ADS47 was unable to use DON as sole carbon source
however increasing DON concentrations in growth media enhanced its DON metabolism
efficiency The pure bacterial culture was shown to be able to rapidly de-epoxidize DON
under aerobic conditions moderate temperatures and neutral pH Strain ADS47 more quickly
de-epoxidizes DON in aerobic conditions than the anaerobic conditions In addition to DON
strain ADS47 showed capability to de-epoxidize andor de-acetylate 10 different type-A and
type-B TC mycotoxins which often co-contaminant grain with DON
To clone the DON detoxifying genes a DON detoxifying bacterial DNA library in E
coli host was constructed followed by screening of the library clones for DON detoxification
activity The study produced a library giving approximately 10 fold coverage of the genome
of strain ADS47 Restriction digestion analysis determined that most of the clones contained
ADS47 DNA inserts This indicates that a good DNA library was produced which had gt99
probability of containing all the DNA of the ADS47 genome However activity-based
screening of the library could not identify any clones with DON de-epoxidation ability It is
assumed that bacterial DON de-epoxidation genes evolved in a certain environmental niche
which may be required for the DON de-epoxidation to be expressed and active but was not
supplied by the host E coli cells
173
An attempt was made to identify the bacterial genes encoding DON de-epoxidation
and de-acetylation enzymes by sequencing the genome of strain ADS47 and comparing its
sequence to the genomes of related bacteria that are not known to posses DON de-epoxidation
activity A RAST server based bioinformatics analysis determined that the 48 Mb circular
genome of ADS47 codes for 4610 predicted CDS A genome wide phylogenetic analysis
determined that strain ADS47 was gt87 identical to the pathogen C freundii strain Ballerup
The relationship to two other animalhuman pathogenic species C rodentium and C koseri
was more distant This result is consistent with the 16S-rDNA based phylogenetic analysis
It is assumed that the microbial DON detoxifying genes were acquired by horizontal
gene transfer or other means during the adaptation of ADS47 in mycotoxin contaminated soil
If it is correct then this means that those newly acquired genes are not common in other
related microbial species and no other Citrobacter species is known to degrade DON Hence
the study identified 270 unique genes to ASDS47 with a subset of 10 genes for reductases
oxidoreductases and a de-acetylase which were considered to be candidate genes for the
enzymes that have DON detoxifying activity Furthermore a targeted comparative genome
analysis showed that the mycotoxin detoxifying ADS47 genome does not harbour the
clinically important LEE encoded typendashIII secretion system and mycobacterium virulence
genes This suggests that strain ADS47 is not likely to be an animal pathogenic bacterium
unlike C freundii Ballerup which encodes the pathogenic relevance LEE operons
174
62 Research contributions
The research made four major contributions which may aid in reducing the risks of
animals and humans from DONTC exposure Firstly the study identified a novel bacterial
strain ADS47 that is able to detoxify DON and 10 related mycotoxins over a wide range of
temperatures under aerobic and anaerobic conditions The discovery has led to a patent
application for strain ADS47 supporting the novelty and importance of the finding This
bacterial strain could be used to detoxify trichothecene mycotoxins in contaminated animal
feed thus reducing food safety risks
Secondly the study resulted in the development of a method for isolating DON de-
epoxidizing microbes from soil as well as optimizing the conditions for bacterial DONTC
detoxification activity to increase the effectiveness usefulness of the strain
Thirdly the thesis work resulted in availability of the genome sequence of a
trichothecene detoxifying microbe which may ultimately lead to an understanding of the
cellularmolecular mechanisms of DONTC de-epoxidation and de-acetylation
genespathways The genome analysis resulted in identification of several candidate genes for
DONTC de-epoxidation and de-acetylation activities Genes which show de-epoxidation
activity could be used to develop DONTC resistant cereal cultivars Fourthly strain ADS47
lacks major animal pathogenicity genes (in particular the LEE-operons) as well as does not
encodes mycobacterium virulence genes Thus strain ADS47 has a great potential to be
utilized to detoxify cereal grain and may be used as a feed additive for livestockfishery All
together the reported work may have significant applications to produce and supply
DONTC-free food and feed
175
63 Further research
This research provides the basis for developing biological detoxification of DON and
related mycotoxins This goal might be achieved by using DON de-epoxidizing gene to
develop resistant plants enzymatic detoxification of contaminated feed and use of the bacteria
as a feed additive Follow-up studies may reach the above indicated goals Because the
bacterium showed de-epoxidation activity under aerobicanaerobic conditions and under wide
range of temperatures it is likely to have an application for enzymatic detoxification of
contaminated animal feed Therefore additional studies on bacterial efficiency to detoxify
contaminated cereal and grain-derived feed need to be conducted Although strain ADS47
does not encode major animal pathogenicity genes the bacterium must be directly tested on
animals to see if it could negatively affect animal digestive systems If not then the potential
for ADS47 is considerable for its use as a feed additive for pig poultry and fish Animal
feeding trials will also need to be performed to determine if DON detoxification is sufficient
to be cost effective as a feed additive In order to develop DONTC resistant cereal cultivars
the gene responsible to DON degrading activity among the presently identified candidate
genes (reductases oxidoreductases and de-acetylase) will need to be expressed in other
organisms like E coli and a codon modified version adapted for expression in plants like
corn and wheat
176
70 REFERENCES
Abbas HK Mirocha CJ Pawlosky RJ Pusch DJ 1985 Effect of cleaning milling and
baking on deoxynivalenol in wheat Applied and Environmental Microbiology 50482ndash
486
Accensi F Pinton P Callu P Abella-Bourges N Guel WJF Grosjean F Oswald IP 2006
Ingestion of low doses of deoxynivalenol does not affect haematological biochemical or
immune responses of piglets Journal of Animal Science 841935ndash1942
Agrawal D Mahapatra AK 2005 Vertically acquired neonatal Citrobacter brain abscessndash
case report and review of the literature Journal of Clinical Neuroscience 12188ndash190
Alexander NJ 2008 The TRI101 story engineering wheat and barley to resist Fusarium head
blight World Mycotoxin Journal 131-37
Alexander NJ Proctor RH McCormick SP 2009 Genes gene clusters and biosynthesis of
trichothecenes and fumonisins in Fusarium Toxin Reviews 28198ndash215
Ali ML Taylor JH Jie L Sun G William M Kasha KJ Reid LM Pauls KP 2005
Molecular mapping of QTLs for resistance to Gibberella ear rot in corn caused by
Fusarium graminearum Genome 48521-533
Alikhan N-F Petty NK Zakour NLB Beatson SA 2011 BLAST Ring Image Generator
(BRIG) simple prokaryote genome comparisons BMC Genomics 12402
Alisi A Spaziani A Anticoli S Ghidinelli M Balsano C 2008 PKR is a novel functional
direct player that coordinates skeletal muscle differentiation via p38MAPKAKT
pathways Cellular Signalling 20534ndash542
177
Amuzie CJ Shinozuka J Pestka JJ 2009 Induction of suppressors of cytokine signaling by
the trichothecene deoxynivalenol in the mouse Toxicological Science 111277ndash287
Ansorge WJ 2009 Next generation DNA sequencing techniques New Biotechnology
25195-203
Arnold JW Senter SD 1998 Use of digital aroma technology and SPME GC-MS to compare
volatile compounds produced by bacteria isolated from processed poultry Journal of the
Science of Food and Agriculture 78343ndash348
Audenaert K Van Broeck R Bekaert B De Witte F Heremans B Messens K Hofte
M Haesaert G 2009 Fusarium head blight (FHB) in Flanders population
diversity inter-species associations and DON contamination in commercial winter
wheat varieties European Journal of Plant Pathology 125445-458
Awad WA Ghareeb K Bohm J Zentek J 2010 Decontamination and detoxification
strategies for the Fusarium mycotoxin deoxynivalenol in animal feed and the
effectiveness of microbial biodegradation Food Additives and Contaminants 27510-520
Aziz K Bartels D Best AA DeJongh M Disz T Edwards RA Formsma K Gerdes S Glass
EM Kubal M Meyer F Olsen GJ Olson R Osterman AL Overbeek RA McNeil LK
Paarmann D Paczian T Parrello1 B Pusch GD Reich C Stevens R Vassieva O
Vonstein V Wilke A Zagnitko O 2008 The RAST Server Rapid Annotations using
Subsystems Technology BMC Genomics 975
Badger JL Stins MF Kim KS 1999 Citrobacter freundii invades and replicates in human
brain microvascular endothelial cells Infection and Immunology 674208ndash4215
Bae HK Pestka JJ 2008 Deoxynivalenol induces p38 interaction with the ribosome in
monocytes and macrophages Toxicological Science 10559-66
178
Bai GH Desjardins AE Plattner RD 2002 Deoxynivalenol-nonproducing Fusarium
graminearum causes initial infection but does not cause disease spread in wheat spikes
Mycopathologia 15391ndash98
Barrett JR 2000 Mycotoxins of molds and maladies Environmental Health Perspectives
108A20ndashA23
Bengoechea JA Skurnik M 2000 Temperature-regulated efflux pumppotassium antiporter
system mediates resistance to cationic antimicrobial peptides in Yersinia Molecular
Microbiology 3767ndash80
Bennett JW Klich M 2003 Mycotoxins Clinical Microbiology Reviews 16497ndash516
Benson DA Karsch-Mizrachi I Lipman DJ Ostell J Sayers EW 2009 GenBank Nucleic
Acids Research 37D26-31
Berdichevsky T Friedberg D Nadler C Rokney A Oppenheim A Rosenshine I 2005 Ler is
a negative autoregulator of the LEE1 operon in neuropathogenic Escherichia coli Journal
of Bacteriology 187349ndash357
Bernardo A Bai GH Guo PG Xiao K Guenzi AC Ayoubi P 2007 Fusarium graminearum-
induced changes in gene expression between Fusarium head blight-resistant and
susceptible wheat cultivars Functional and Integrative Genomics 769ndash77
Bhat RV Beedu SR Ramakrishna Y Munshi KL 1989 Outbreak of trichothecene
mycotoxicosis associated with consumption of mould-damaged wheat production in
Kashmir Valley India The Lancet 33335ndash37
Binder EM Tan LM Chin LJ Handl J Richard J 2007 Worldwide occurrence of
mycotoxins in commodities feeds and feed ingredients Animal Feed Science and
Technology 137265-282
179
Birzele B Meier A Hindorf H Kraumlmer J Dehne H-W 2002 Epidemiology of Fusarium
infection and deoxynivalenol content in winter wheat in the Rhineland Germany
European Journal of Plant Pathology 108667ndash673
Boddu J Cho S Muehlbauer GJ 2007 Transcriptome analysis of trichothecene-induced gene
expression in barley Molecular PlantndashMicrobe Interactions 201364ndash1375
Borenshtein D Schauer DB 2006 The Genus Citrobacter Prokaryotes 690ndash98
Boutigny A-L Richard-Forget F Barreau C 2008 Natural mechanisms for cereal resistance
to the accumulation of Fusarium trichothecenes European Journal of Plant Pathology
121411ndash423
Boutrif E Canet C 1998 Mycotoxin prevention and control FAO programmes Revue de
Meacutedecine Veacuteterinaire 149681-694
Brennan JM Leonard G Fagan B Cooke BM Ritieni A Ferracane R et al 2007
Comparison of commercial European wheat cultivars to Fusarium infection of head and
seedling tissue Plant Pathology 5655ndash64
Brenner DJ Grimont PA Steigerwalt AG Fanning GR Ageron E Riddle CF 1993
Classification of Citrobacteria by DNA hybridization designation of Citrobacter farmeri
sp nov Citrobacter youngae sp nov Citrobacter braakii sp nov Citrobacter
werkmanii sp nov Citrobacter sedlakii sp nov and three unnamed Citrobacter
genomospecies International Journal of Systematic Bacteriology 43645-58
Brookes G Barfoot P 2006 GM Crops The first ten years-global socio-economic and
environmental Impacts no 36 ISAAA (International Service for the Acquisition of Agri-
Biotech Applications) Ithaca New York
180
Bruce KD Hughes MR 2000 Terminal restriction fragment length polymorphism monitoring
of genes amplified directly from bacterial communities in soils and sediments Molecular
Biotechnology 16261ndash269
Buerstmayr H Ban T Anderson JA 2009 QTL mapping and marker-assisted selection for
Fusarium head blight resistance in wheat A review Plant Breeding 1281ndash26
Campbell H Choo TM Vigier B Underhill L 2002 Comparison of mycotoxin profiles
among cereal samples from eastern Canada Canadian Journal of Botany 80526ndash532
Carver et al 2005 ACT the Artemis Comparison Tool Bioinformatics 213422-3423
Carver et al 2009 DNAPlotter circular and linear interactive genome visualization
Bioinformatics 25119-120
Chen CY et al 2003 Comparative genome analysis of Vibrio vulnificus a marine pathogen
Genome Research 132577ndash2587
Chiu TC Yen JH Hsieh YN Wang YS 2005 Reductive transformation of dieldrin under
anaerobic sediment culture Chemosphere 601182ndash1189
Chung YJ Zhou HR Pestka JJ 2003 Transcriptional and posttranscriptional roles for p38
mitogen-activated protein kinase in upregulation of TNF-alpha expression by
deoxynivalenol (vomitoxin) Toxicology and Applied Pharmacology 193188ndash201
Chung EJ Lim HK Kim J-C Choi GJ Park EJ Lee MH Chung YR Lee S-W 2008 Forest
soil metagenome gene cluster involved in antifungal activity expression in Escherichia
coli Applied and Environmental Microbiology 74723-730
Collins TFX Sprando RL Black TN Olejnik N Eppley RM Hines FA Rorie J Ruggles DI
2006 Effects of deoxynivalenol (DON vomitoxin) on in utero development in rats Food
and Chemical Toxicology 44747ndash757
181
Cong Y Wang J Chen Z Xiong K Xu Q Hu F 2011 Characterization of swarming motility
in Citrobacter freundii FEMS Microbiology Letters 371160-171
Cossette F Miller JD 1995 Phytotoxic effect of deoxynivalenol and Gibberella ear rot
resistance of corn Natural Toxins 3383ndash388
Creppy EE 2002 Update of survey regulation and toxic effects of mycotoxins in Europe
Toxicological Letters 12719-28
Dai JS Wang D Guerlebeck C Laturnus S Guenther Z Lu SC Ewers C 2010 Suppression
subtractive hybridization identifies an autotransporter adhesin gene of E coli IMT5155
specifically associated with avian pathogenic Escherichia coli (APEC) BMC
Microbiology 10236
Daniel R 2005 The metagenomics of soil Nature Review Microbiology 3470-478
Davis KER Joseph SJ Janssen PH 2005 Effects of growth medium inoculum size and
incubation time on culturability and isolation of soil bacteria Applied and Environmental
Microbiology 71826-834
Delcher AL Phillippy A Carlton J Salzberg SL 2002 MUMmer 21 NUCmer and
PROmer are described in Fast Algorithms for Large-scale Genome Alignment and
Comparisonrdquo Nucleic Acids Research 302478-2483
Demain AL Adrio JL 2008 Contributions of microorganisms to industrial biology
Molecular Biotechnology 3841ndash455
Deng W Li Y Vallance BA Finlay BB 2001 Locus of enterocyte efffacement from
Citrobacter rodentium sequence analysis and evidence for horizontal transfer among
attaching and effacing pathogens Infection and Immunity 696323ndash6335
182
Desjardins AE Proctor RH Bai G McCormick SP Shaner G Beuchley G Hohn TM 1996
Reduced virulence of trichothecene-nonproducing mutants of Gibberella zeae in wheat
field tests Molecular Plant-Microbe Interactions 9775-781
Desjardins AE 2006 Fusarium mycotoxins chemistry genetics and biology The American
Phytopathological Society St Paul Minnesota USA p 260
Desjardins AE Proctor RH 2007 Molecular biology of Fusarium mycotoxins International
Journal of Food Microbiology 11947-50
Desmond OJ Manners JM Stephens AE MacLean DJ Schenk PM Gardiner DM Munn
AL Kazan K 2008 The Fusarium mycotoxin deoxynivalenol elicits hydrogen peroxide
production programmed cell death and defence responses in wheat Molecular Plant
Pathology 9435-445
Dill-Macky R Jones RK 2000 The effect of previous crop residues and tillage on Fusarium
head blight of wheat Plant Disease 8471ndash76
DrsquoMello JPF Macdonald AMC Dijksma WTP 1998 3-Acetyl deoxynivalenol and esterase
production in a fungicide-insensitive strain of Fusarium culmorum Mycotoxin Research
149ndash18
DrsquoMello JPF Placinta CM Macdonald AMC 1999 Fusarium mycotoxins a review of
global implications of animal health welfare and productivity Animal Feed Science and
Technology 80183-205
Doohan FM Brennan J Cooke BM 2003 Influence of climatic factors on Fusarium species
pathogenic to cereals European Journal of Plant Pathology 109755ndash768
183
Dyer RB Kendra DF Brown DW 2006 Real-time PCR assay to quantify Fusarium
graminearum wild-type and recombinant mutant DNA in plant material Journal of
Microbiological Methods 67534ndash542
Erecinska M Wilson DF 1981 Inhibitors of cytochrome c oxidase In inhibitors of
mitochondrial function edited by M Erecinska and DF Wilson Pergamon Press Ltd
Oxford pp 145-160
Eriksen GS 2003 Metabolism and toxicity of trichothecene Doctoral thesis Swedish
University of Agricultural Sciences Uppsala Sweden
Eriksen GS Pettersson H Lundh T 2004 Comparative cytotoxicity of deoxynivalenol
nivalenol their acetylated derivatives and de-epoxy metabolites Food Chemistry and
Toxicology 42619-624
Eudes F Comeau A Rioux S Collin J 2000 Phytotoxicity of eight mycotoxins associated
with Fusarium in wheat head blight Canadian Journal of Plant Pathology 22286ndash292
FAO (Food and Agriculture Organization) 1991 Food Nutrition and Agriculture Food for
the Future volume 1 FAO Rome
FAO (Food and Agriculture Organization) 2001 FNP73 Manual of the application of
HACCP system in mycotoxin prevention and control FAO Food and Nutrition paper no
73 FAO Rome
Fedorak PM Grbic-Galic D 1991 Aerobic microbial cometabolism of benzothiophene and
3-methylbenzothiophene Applied and Environmental Microbiology 57932-940
Felsenstein J 1989 PHYLIP - Phylogeny Inference Package (Version 32) Cladistics 5164-
166
184
Fernando WGD Amarasinghe C Demoz B Surujdeo‐Maharaj S Gauthier V Guerrieri G
Al‐Taweel K Brucircleacute‐Babel A Tamburic‐Illincic L Gilbert J 2011 What do we know of
the Fusarium graminearum chemotypes their epidemiology genetics and infection
process in Canada In Proc 7th
Canadian Workshop on Fusarium Head Blight edited by
J Gilbert A Tekauz N Sweetland and K Slusarenko Winnipeg Manitoba Canada p 45
Fierer N Jackson RB 2006 The diversity and biogeography of soil bacterial community
Proceedings of the National Academy of Sciences 103626-631
Fink-Gremmels J 1989 The significance of mycotoxin assimilation for meat animals
Deutsche Tierarztliche Wochenschrift 96360-363
Flint DH Emptage MH Finnegan MG Fu W Johnson MK 1993 The role and properties of
the iron-sulfur cluster in Escherichia coli dihydroxy-acid dehydratase Journal of
Biological Chemistry 26814732ndash14742
Foroud NA Eudes F 2009 Trichothecenes in cereal grains International Journal of
Molecular Science 10147-173
Forsell JH Jensen R Tai JH Witt M Lin WS Pestka JJ 1987 Comparison of acute
toxicities of deoxynivalenol (vomitoxin) and 15-acetyldeoxynivalenol in the B6C3F1
mouse Food and Chemical Toxicology 25155ndash162
Frey M Stettner C Gierl A 1998 A general method for gene isolation in tagging approaches
amplification of insertion mutagenised sites (AIMS) The Plant Journal 13717-721
Freeman GG Morrison RI 1949 The isolation and chemical properties of trichothecin an
antifungal substance from Trichothecium roseum Link Biochemical Journal 441ndash5
185
Fuchs E Binder EM Heidler D Krska R 2002 Structural characterization of metabolites
after the microbial degradation of type A trichothecenes by the bacterial strain BBSH 797
Food Additives and Contaminants 19379ndash386
Gabor EM Alkema WBL Janssen DB 2004 Quantifying the accessibility of the
metagenome by random expression cloning techniques Environmental Microbiology
6879ndash886
Gadd GM Griffiths AJ 1978 Microorganisms and heavy metal toxicity Microbial Ecology
4303ndash317
Gale LR Bryant JD Calvo S Giese H 2005 Chromosome complement of the fungal plant
pathogen Fusarium graminearum based on genetic and physical mapping and cytological
observations Genetics 171985ndash1001
Gao LY Pak M Kish R Kajihara K Brown EJ 2006 A mycobacterial operon essential for
virulence in vivo and invasion and intracellular persistence in macrophages Infection and
Immunity 741757ndash1767
Gareau MG Wine E Reardon C et al 2010 Probiotics prevent death caused by Citrobacter
rodentium infection in neonatal mice Journal of Infectious Diseases 20181e91
Gareis M Bauer J Thiem J Plank G Grabley S Gedek B 1990 Cleavage of
zearalenoneglycoside a masked mycotoxin during digestion in swine Journal of
Veterinary Medicine 37236-240
Geng L Xin W Haung DW Feng G 2006 A universal cloning vector using vaccinia
topoisomerase I Molecular Biotechnology 3323-28
186
Golkari S Gilbert J Ban T Procunier JD 2009 QTLspecific microarray gene expression
analysis of wheat resistance to Fusarium head blight in Sumai-3 and two susceptible
NILs Genome 52409ndash418
Goodstadt L Ponting CP 2004 Vitamin K epoxide reductase homology active site and
catalytic mechanism Trends in Biochemical Sciences 29289-292
Goswami RS Kistler HC 2004 Heading for disaster Fusarium graminearum on cereal
crops Molecular Plant Pathology 5515-525
Gouze ME Laffitte J Printon P Dedieux G Galinier A Thouvenot JP Loiseau N Oswald
IP Galtier P 2007 Effect of subacute oral doses of nivalenol on immune and metabolic
defence systems in mice Veterinary Research 38635ndash646
Guan S He J Young JC Zhu H Li X Ji C Zhou T 2009 Transformation of trichothecene
mycotoxins by microorganisms from fish digesta Aquaculture 290290ndash295
Gupta R Beg QK Lorenz P 2002 Bacterial alkaline proteases molecular approaches and
industrial applications Applied Microbiology and Biotechnology 5915ndash32
Haberle J Schweizer G Schondelmaier J Zimmermann G Hart L 2009 Mapping of QTL
for resistance against Fusarium head blight in the winter wheat population
PelikanBussardNing8026 Plant Breeding 12827ndash35
Hall BG 2004 Predicting the evolution of antibiotic resistance genes Nature Reviews
Microbiology 2430-435
Han JI et al 2011 Complete genome sequence of the metabolically versatile plant growth-
promoting endophyte Variovorax paradoxus S110 Journal of Bacteriology 1931183ndash
1190
187
Harvey RB Edrington TS Kubena LF Elissalde MH Corrier DE Rottinghaus GE 1995
Effect of aflatoxin and diacetoxyscirpenol in ewe lambs Bulletin of Environmental
Contamination and Toxicology 54325-330
Harvey RB Edrington TS Kubena LF Elissalde MH Casper HH Rottinghaus GE Turk JR
1996 Effects of dietary fumonisin B1-containing culture material deoxynivalenol-
contaminated wheat or their combination on growing barrows American Journal of
Veterinary Research 571790-1794
He J 2007 Microbial strategies for management of Fusarium graminearum and
transformation of deoxynivalenol Master of Science Thesis University of Guelph
Guelph Ontario Canada
He K Vines L Pestka JJ 2010a Deoxynivalenol-induced modulation of microRNA
expression in RAW 2647 macrophages-A potential novel mechanism for translational
inhibition Toxicologist 114310
He J Zhou T Young JC Boland GJ Scott PM 2010b Chemical and biological
transformations for trichothecene mycotoxins in human and animal food chains A review
Trends in Food Science and Technology 2167-76
He J Zhou T 2010 Patented techniques for detoxification of mycotoxins in feeds and food
matrices Rec Pat Food Nutr Agric 296ndash104
He P Young LG Forsberg C 1992 Microbial transformation of deoxynivalenol (vomitoxin)
Applied and Environmental Microbiology 583857-3863
Hell K Cardwell KF Setamou M Poehling H-M 2000 The influence of storage practices on
aflatoxins contamination in maize in four agroecological zones in Benin West Africa
Journal of Stored Product Research 36365ndash82
188
Henne A Schmitz RA Bomeke M Gottschalk G Daniel R 2000 Screening of
environmental DNA libraries for the presence of genes conferring lipolytic activity on
Escherichia coli Applied and Environmental Microbiology 663113ndash3116
Hodges GR Degener CE Barnes WG 1978 Clinical significance of Citrobacter isolates
American Journal of Clinical Pathology 7037ndash40
Hollinger K Ekperigin HE 1999 Mycotoxicosis in food producing animals The Veterinary
Clinics of North America Food Animal Practice 15133-165
Hong Y Kinoshita T 2009 Trypanosome glycosylphosphatidylinositol biosynthesis Korean
Journal of Parasitology 47197ndash204
Hope RJ Aldred D Magan N 2005 Comparison of environmental profiles for growth and
deoxynivalenol production by Fusarium culmorum and F graminearum on wheat grain
Letters in Applied Microbiology 40295ndash300
Horvath RS 1972 Microbial co-metabolism and the degradation of organic compounds in
nature Bacteriological Reviews 36146ndash155
Hsu D Shih LM Zee YC 1994 Degradation of rRNA in Salmonella strains a novel
mechanism to regulate the concentrations of rRNA and ribosomes Journal of
Bacteriology 1764761ndash4765
Huwig A Freimund S Kappeli O Dutler H 2001 Mycotoxin detoxification of animal feed
by different adsorbents Toxicological Letters 122179ndash188
Iordanov MS Pribnow D Magun JL Dinh TH Pearson JA Magun BE 1997 Ribotoxic
stress response activation of the stress activated protein kinase JNK1 by inhibitors of the
peptidyl transferase reaction and by sequence-specific RNA damage to the sarcinricin
loop in the 28S rRNA Molecular and Cellular Biology 173373ndash3381
189
Islam R Zhou T Young JC Goodwin PH Pauls KP 2012 Aerobic and anaerobic de-
epoxidation of mycotoxin deoxynivalenol by bacteria originating from agricultural soil
World Journal of Microbiology and Biotechnology 287-13
Jackson LS Bullerman LB 1999 Effect of processing on Fusarium mycotoxins Advances in
Experimental Medicine and Biology 459243-261
Jansen C von Wettstein D Schafer W Kogel K-H Felk A Maier FJ 2005 Infection patterns
in barley and wheat spikes inoculated with wild-type and trichodiene synthase gene
disrupted Fusarium graminearum Proceedings of the National Academy of Sciences
10216892ndash16897
Jube S Borthakur D 2007 Expression of bacterial genes in transgenic tobacco methods
applications and future prospects Electronic Journal of Biotechnology 10452-467
Kabak B Dobson AD 2009 Biological strategies to counteract the effects of mycotoxins
Journal of Food Protection 722006-2016
Kant P Reinprecht Y Martin CJ Islam R Pauls KP 2011 Integration of biotechnologies
disease resistance pathology and Fusarium In Comprehensive Biotechnology edited by
Moo-Young M Second Edition 4729-743
Kaplan CW Kitts CL 2004 Bacterial succession in a petroleum land treatment unit Applied
and Environmental Microbiology 701777-1786
Kaprelyants AS Kell DB 1996 Do bacteria need to communicate with each other for
growth Trends in Microbiology 4237-42
190
Kariholu U Rawal J Namnyak S 2009 Neonatal Citrobacter koseri meningitis and brain
abscess The Internet Journal of Pediatrics and Neonatology 10
(httpwwwispubcomjournalthe-internet-journal-of-pediatrics-and-
neonatologyvolume-10-number-1neonatal-citrobacter-koseri-meningitis-and-brain-
abscesshtml) (accessed on April 19 2012)
Karlovsky P 1999 Biological detoxification of fungal toxins and its use in plant breeding
feed and food production Natural Toxins 71ndash23
Karlovsky P 2011 Biological detoxification of the mycotoxin deoxynivalenol and its use in
genetically engineered crops and feed additives Applied Microbiology and
Biotechnology 91491-504
Kavanagh KL Jornvall H Persson B Oppermann U 2008 Medium and short-chain
dehydrogenasereductase gene and protein families the SDR superfamily functional and
structural diversity within a family of metabolic and regulatory enzymes Cellular and
Molecular Life Sciences 653895ndash3906
Khera KS Whalen C Angers G Vesonder RF Kuiper-Goodman T 1982 Embryotoxicity of
4-deoxynivalenol (vomitoxin) in mice Bulletin of Environmental Contamination and
Toxicology 29487ndash491
Kikot GE Hours RA Alconada TM 2009 Contribution of cell wall degrading enzymes to
pathogenesis of Fusarium graminearum a review Journal of Basic Microbiology 49
231ndash241
Kim UJ Shizuya H Jong PJ Birren B Simon MI 1992 Stable propogation of cosmid sized
human DNA inserts in a F factor based vector Nucleic Acids Research 201083-1065
191
Kimura M Kaneko I Komiyama M Takatsuki A Koshino H Yoneyama K Yamaguchi I
1998 Trichothecene 3-O-acetyltransferase protects both the producing organism and
transformed yeast from related mycotoxins cloning and characterization of Tri101
Journal of Biological Chemistry 2731654-1661
Kimura M Anzai H Yamaguchi I 2001 Microbial toxins in plantndashpathogen interactions
biosynthesis resistance mechanisms and significance The Journal of General and
Applied Microbiology 47149ndash160
Kimura M Tokai T Takahashi-Ando N Ohsato S Fujimura M 2007 Molecular and genetics
studies of Fusarium trichothecene biosynthesis pathways genes and evolution
Bioscience Biotechnology and Biochemistry 712105-2123
King RR McQueen RE Levesque D Greenhalgh R 1984 Transformation of deoxynivalenol
(vomitoxin) by rumen microorganisms Journal of Agricultural and Food Chemistry
32181ndash1183
Klappenbach JA Dunbar JM Schmidt TM 2000 rRNA operon copy number reflects
ecological strategies of bacteria Applied and Environmental Microbiology 661328ndash
1333
Knietsch A Waschkowitz T Bowien S Henne A Daniel R 2003 Metagenomes of complex
microbial consortia derived from different soils as sources for novel genes conferring
formation of carbonyls from short-chain polyols on Escherichia coli Journal of Molecular
Microbiology and Biotechnology 546ndash56
Kollarczik B Gareis M Hanelt M 1994 In vitro transformation of the Fusarium mycotoxins
deoxynivalenol and zearalenone by the normal gut microflora of pigs Natural Toxins
2105ndash110
192
Koonin EV Wolf YI Aravind L 2001 Prediction of the archaeal exosome and its
connections with the proteasome and the translation and transcription machineries by a
comparative-genomic approach Genome Research 11240ndash252
Kouadio JH Dano SD Moukha S Mobio TA Creppy EE 2007 Effects of combinations of
Fusarium mycotoxins on the inhibition of macromolecular synthesis malondialdehyde
levels DNA methylation and fragmentation and viability in Caco-2 cells Toxicon
49306ndash317
Krause L Diaz NN Bartels D Edwards RA Puumlhler A et al 2006 Finding novel genes in
bacterial communities isolated from the environment Bioinformatics 22281ndash289
Kubena LF Huff WE Harvey RB Phillips TD Rottinghaus GE 1989 Individual and
combined toxicity of deoxynivalenol and T-2 toxin in broiler chicks Poultry Science
68622ndash626
Kubena LF Harvey RB Huff WE Corrier DE Phillips TD Rottinghaus GE 1990 Efficacy
of hydrated sodium calcium aluminosilicate to reduce the toxicity of aflatoxin and T-2
toxin Poultry Science 691078ndash1086
Kubena LF Harvey RB Huff WE Elissalde MH Yersin AG Phillips TD Rottinghaus GE
1993 Efficacy of hydrated sodium calcium aluminosilicate to reduce the toxicity of
aflatoxin and diacetoxyscirpenol Poultry Science 7251ndash59
Kubena LF Edrington TS Harvey RB Buckley SA Phillips TD Rottinghaus GE Casper
HH 1997 Individual and combined effects of fumonisin B1 present in Fusarium
moniliforme culture material and T-2 toxin or deoxynivalenol in broiler chicks Poultry
Science 761239ndash1247
193
Kumar RA Gunasekaran P Lakshmanan M 1999 Biodegradation of tannic acid by
Citrobacter freundii isolated from a tannery effluent Journal of Basic Microbiology
39161-168
Kushiro M 2008 Effects of milling and cooking processes on the deoxynivalenol content in
wheat International Journal Molecular Science 92127-2145
Lane DJ 1991 16S23S rRNA sequencing In Nucleic acid techniques in bacterial
systematic edited by Stackebrandt E and Goodfellow M Wiley J and Sons Inc New
York pp 115-175
Lancova K Hajslova J Poustka J Krplova A Zachariasova M Dostalek P Saschambula L
2008 Transfer of Fusarium mycotoxins and ldquomaskedrdquo deoxynivalenol (deoxynivalenol-3-
glucoside) from field barley through malt to beer Food Additives and Contaminants
25732ndash744
Langmead B Trapnell C Pop M Salzberg SL 2009 Ultrafast and memory-efficient
alignment of short DNA sequences to the human genome Genome Biology 10R25
Larsen JC Hunt J Perrin I Ruckenbauer P 2004 Workshop on trichothecenes with a focus
on DON summary report Toxicological Letters 1531-22
Lee T Oh DW Kim HS Lee J Kim YH Yun SH Lee YW 2001 Identification of
deoxynivalenol- and nivalenol-producing chemotypes of Gibberella zeae by using PCR
Applied and Environmental Microbiology 672966ndash2972
Lee S-W Won K Lim HK Kim J-C Choi GJ Cho KY 2004 Screening for novel lipolytic
enzymes from uncultured soil microorganisms Applied and Environmental Microbiology
65720-726
194
Li L Christian J Stoeckert J David SR 2003 OrthoMCL Identification of Ortholog Groups
for Eukaryotic Genomes Genome Research 132178-2189
Li T Chang CY Jin DY Lin PJ Khvorova A Stafford DW 2004 Identification of the gene
for vitamin K epoxide reductase Nature 427541ndash544
Li Y 2007 Sensory signal transduction in the vagal primary afferent neurons Current
Medicinal Chemistry 142554ndash2563
Li MX Pestka JJ 2008 Comparative induction of 28S ribosomal RNA cleavage by ricin and
the trichothecenes deoxynivalenol and T-2 toxin in the macrophage Toxicological
Science 10567ndash78
Li R Li Y Kristiansen K Wang J 2008 SOAP short oligonucleotide alignment program
Bioinformatics 24713-4
Li W Schulman S Dutton RJ Boyd D Beckwith J Rapoport TA 2010 Structure of a
bacterial homologue of vitamin K epoxide reductase Nature 463507-512
Lichstein HC Malcolm HS 1943 Studies of the effect of sodium azide on microbic growth
and respiration Journal of Bacteriology 47221ndash230
Limay-Rios V Schaafsma AW 2011 2010 Update on the development of an integrated
mycotoxin management system in Ontario corn 30th
Centralia Swine Research Update
Kirkton Ontario Canada
Llorens A Hinojo MJ Mateo R Gonzalez-Jaen MT Valle- Algarra FM Logrieco A Jimenez
M 2006 Characterization of Fusarium spp isolates by PCR-RFLP analysis of the
intergenic spacer region of the rRNA gene (rDNA) International Journal of Food
Microbiology 106287ndash306
195
Lombaert GA Pellaers P Roscoe V Mankotia M Neil R Scott PM 2003 Mycotoxins in
infant cereal foods from the Canadian retail market Food Additives and Contaminants
20494ndash504
Luengo JM Garciacutea JL Olivera ER 2001 The phenylacetyl-CoA catabolon a complex
catabolic unit with broad biotechnological applications Molecular Microbiology
391434ndash1442
Luo X 1994 Food poisoning caused by Fusarium toxins In Proceedings of the Second
Asian Conference on Food Safety ILSI Thailand pp 129ndash136
Mabagala RB 1997 The effect of populations of Xanthomonas campestris pv phaseoli in
bean reproductive tissues on seed infection of resistant and susceptible bean genotypes
European Journal of Plant Pathology 103175ndash181
Madhyastha MS Marquardt RR Abramson D 1994 Structurendashactivity relationships and
interactions among trichothecene mycotoxins as assessed by yeast bioassay Toxicon
321147ndash1152
Maier FJ Miedaner T Hadeler B Felk A Salomon S Lemmens M Kassner H Schaumlfer W
2006 Involvement of trichothecenes in fusarioses of wheat barley and maize evaluated
by gene disruption of the trichodiene synthase (Tri5) gene in three field isolates of
different chemotype and virulence Molecular Plant Pathology 7449ndash461
Malojcic G Owen RL Grimshaw JP Glockshuber R 2008 Preparation and structure of the
charge-transfer intermediate of the transmembrane redox catalyst DsbB FEBS Letters
5823301ndash3307
196
Manoharan M Dahleen LS Hohn TM Neate SM Yu XH Alexander NJ et al 2006
Expression of 3-OH trichothecene acetyltransferase in barley (Hordeum vulgare L) and
effects on deoxynivalenol Plant Science 171699ndash706
Maresca M Mahfoud R Garmy N Fantini J 2002 The mycotoxin deoxynivalenol affects
nutrient absorption in human intestinal epithelial cells The Journal of Nutrition
1322723ndash2731
Martins ML Martins HM 2002 Influence of water activity temperature and incubation time
on the simultaneous production of deoxynivalenol and zearalenone in corn (Zea mays) by
Fusarium graminearum Food Chemistry 79315ndash318
Masuda D Ishida M Yamaguchi K Yamaguchi I Kimura M Nishiuchi T 2007 Phytotoxic
effects of trichothecenes on the growth and morphology of Arabidopsis thaliana Journal
of Experimental Botany 581617ndash1626
Matsumoto E Kawanaka Y Yun SJ Oyaizu H 2008 Isolation of dieldrin- and endrin-
degrading bacteria using 12-epoxycyclohexane as a structural analog of both compounds
Applied Microbiology and Biotechnology 801095-1103
McLean M 1996 The phytotoxicity of Fusarium metabolites an update since 1989
Mycopathologia 133163ndash179
Merhej J Richard-Forget F Barreau C 2011 Regulation of trichothecene biosynthesis in
Fusarium recent advances and new insights Applied Microbiology and Biotechnology
91519ndash528
Midwest Plan Service 1987 Grain Drying Handling Storage Handbook Ames Iowa Iowa
State University 2nd edition
197
Miller SS Reid LM Harris LJ 2007 Colonization of maize silks by Fusarium graminearum
the causative organism of Gibberella ear rot Canadian Journal of Botany 85369-376
Minervini F Fornelli F Flynn KM 2004 Toxicity and apoptosis induced by mycotoxins
nivalenol deoxynivalenol and fumonisin B1 in a human erythroleukemia cell line
Toxicology In Vitro 1821ndash28
Mitterbauer R Poppenberger B RaditschnigA Lucyshyn D Lemmens M Glossl J Adam G
2004 Toxin-dependent utilization of engineered ribosomal protein L3 limits trichothecene
resistance in transgenic plants Plant Biotechnology Journal 2329ndash340
Mocali S Benedetti A 2010 Exploring research frontiers in microbiology the challenge of
metagenomics in soil microbiology Research Microbiology 161497-505
Morozkina EV Zvyagilskaya RA 2007 Nitrate reductases structure functions and effect of
stress factors Biochemistry 721151ndash1160
Munkvold GP 2003 Cultural and genetic approaches to managing mycotoxins in maize
Annual Review of Phytopathology 4199-116
Mundy R MacDonald TT Dougan G Frankel G Wiles S 2005 Citrobacter rodentium of
mice and man Cellular Microbiology 71697ndash1706
Nakamura N Inoue N Watanabe R Takahashi M Takeda J Stevens VL Kinoshita T 1997
Expression cloning of PIG-L a candidate N-acetylglucosaminyl-phosphatidylinositol
deacetylase The Journal of Biological Chemistry 27215834ndash15840
Nganje WE Bangsund DA Leistritz FL Wilson WW Tiapo NM 2002 Estimating the
economic impact of a crop disease the case of Fusarium head blight in US wheat and
barley National Fusarium Head Blight Forum Proceedings East Lansing Michigan State
University pp 275ndash281
198
Nielsen C Lippke H Didier A Dietrich R Martlbauer E 2009 Potential of deoxynivalenol
to induce transcription factors in humanhepatoma cells Molecular Nutrition and Food
Research 53479ndash491
Nielsen JKS Vikstroumlm AC Turner P Knudsen LE 2011 Deoxynivalenol transport across
the human placental barrier Food and Chemical Toxicology 492046-2052
Nishiuchi T Masuda D Nakashita H Ichimura K Shinozaki K Yoshida S et al 2006
Fusarium phytotoxin trichothecenes have an elicitor-like activity in Arabidopsis thaliana
but the activity differed significantly among their molecular species Molecular Plantndash
Microbe Interactions 19512ndash520
Ohsato S Ochiai-Fukuda T Nishiuchi T Takahashi-Ando N Koizumi S Hamamoto H
Kudo T Yamaguchi I Kimura M 2007 Transgenic rice plants expressing trichothecene
3-O-acetyltransferase show resistance to the Fusarium phytotoxin deoxynivalenol Plant
Cell Reports 26531-538
Okubara PA Blechl AE McCormick SP Alexander NJ Dill-Macky R Hohn TM 2002
Engineering deoxynivalenol metabolism in wheat through the expression of a fungal
trichothecene acetyltransferase gene Theoritical and Applied Genetics 10674-83
Ong CL Beatson SA Totsika M Forestier C McEwan AG Schembri MA 2010 Molecular
analysis of type 3 fimbrial genes from Escherichia coli Klebsiella and Citrobacter
species BMC Microbiology 10183
Osborne LE Stein JM 2007 Epidemiology of Fusarium head blight on small-grain cereals
International Journal of Food Microbiology 119103-108
Page RDM 1996 TREEVIEW An application to display phylogenetic trees on personal
computers Computer Applications in the Biosciences 12357-358
199
Parkhill J Dougan G James KD Thomson NR Pickard D Wain J Churcher C Mungall KL
Bentley SD Holden MT et al 2001 Complete genome sequence of a multiple drug
resistant Salmonella enterica serovar Typhi CT18 Nature 413848ndash852
Paterson RRM Lima N 2010 How will climate change affect mycotoxins in food Food
Research International 431902ndash14
Paul PA El-Allaf SM Lipps PE Madden LV 2004 Rain splash dispersal of Gibberella zeae
within wheat canopies in Ohio Phytopathology 941342-1349
Pestka JJ Lin WS Miller ER 1987 Emetic activity of the trichothecene 15-
acetyldeoxynivalenol in pigs Food and Chemical Toxicology 25855ndash858
Pestka JJ Smolinski AT 2005 Deoxynivalenol Toxicology and potential effects on humans
Journal of Toxicology Environmental Health 839ndash69
Pestka JJ 2007 Deoxynivalenol toxicity mechanisms and animal health risks Animal Feed
Science and Technology 137283-298
Pestka JJ 2008 Mechanisms of deoxynivalenol-induced gene expression and apoptosis Food
Additives and Contaminants 251128-1140
Pestka J2010 Toxicological mechanisms and potential health effects of deoxynivalenol and
nivalenol World Mycotoxin Journal 3323ndash347
Petersohn A Brigulla M Haas S Hoheisel JD Voumllker U Hecker M 2001 Global Analysis
of the General Stress Response of Bacillus subtilis Journal of Bacteriology 1835617-
5631
200
Petty NK Bulgin R Crepin VF Cerdeno-Tarraga AM Schroeder GN Quail MA Lennard N
Corton C Barron A Clark L Toribio AL Parkhill J Dougan G Frankel G Thomson NR
2010 The Citrobacter rodentium genome sequence reveals convergent evolution with
human pathogenic Escherichia coli Journal of Bacteriology 192525ndash538
Petty NK Feltwell T Pickard D Clare S Toribio AL et al 2011 Citrobacter rodentium is
an unstable pathogen showing evidence of significant genomic flux PLoS Pathology
7e1002018 doi101371journalppat1002018
Pittet A 2001 Natural occurrence of mycotoxins in foods and feeds A decade in review In
Mycotoxins and phytotoxins in perspective at the turn of the millennium edited by De
Koe WJ Samson RA Van Egmond HP Gilbert J Sabino M W J de Koe Wageningen
Netherlands pp 153ndash172
Placinta CM DrsquoMello JPF Macdonald AMC 1999 A review of worldwide contamination of
cereal grains and animal feed with Fusarium mycotoxins Animal Feed Science and
Technology 7821ndash37
Plank B Schuh M Binder EM 2009 Investigations on the effect of two feed additives
Biomin BBSH 797 and Mycofix Plus (R) 3E as detoxificants of DON contaminated feed
of piglets Wien Tierarztl Monatssch 9655ndash71
Poapolathep A Ohtsuka R Kiatipattanasakul W Ishigami N Nakayama H Doi K 2002
Nivalenol-induced apoptosis in thymus spleen and Peyerrsquos patches of mice Experimental
and Toxicological Pathology 53441ndash446
201
Poppenberger B Berthiller F Lucyshyn D Sieberer T Schuhmacher R Krska R Kuchler K
Gloumlssl J Luschnig C Adam G 2003 Detoxification of the Fusarium mycotoxin
deoxynivalenol by a UDP-glucosyltransferase from Arabidopsis thaliana Journal of
Biological Chemistry 27847905ndash47914
Prelusky DB Trenholm HL 1993 The efficacy of various classes of anti-emetics in
preventing deoxynivalenol-induced vomiting in swine Natural Toxins 1296ndash302
Proctor RH Hohn TM McCormick SP 1995 Reduced virulence of Gibberella zeae caused
by disruption of a trichothecene toxin biosynthetic gene Molecular Plant-Microbe
Interactions 8593-601
Puchenkova SG 1996 Enterobacteria in areas of water along the Crimean Coast
Mikrobiolohichnyĭ zhurnal 58 3-7
Qi BW Hao BL 2004 Whole-proteome prokaryote phylogeny without sequence alignment
a K-string composition approach Journal of Molecular Evolution 581ndash11
Quarta A Mita G Haidukowski M Logrieco A Mule G Visconti A 2006 Multiplex PCR
assay for the identification of nivalenol 3- and 15-acetyl-deoxynivalenol chemotypes in
Fusarium FEMS Microbiological Letters 2597ndash13
Rehr B Klemme JH 1989 Formate dependent nitrate and nitrite reduction to ammonia by
Citrobacter freundii and competition with denitrifying bacteria Antonie van Leeuwenhoek
Journal of Microbiology 56311ndash321
Rocha O Ansari K Doohan FM 2005 Effects of trichothecene mycotoxins on eukaryotic
cells A review Food Additives and Contaminants 22369ndash378
Roscoe V Lombaert GA Huzel V et al 2008 Mycotoxins in breakfast cereals from the
Canadian retail market a 3-year survey Food Additives and Contaminants 25347ndash355
202
Rotter BA Prelusky DB Pestka JJ 1996 Toxicology of deoxynivalenol (vomitoxin) Journal
of Toxicology and Environment Health 481ndash34
Ryu JC Ohtsubo K Izurniyarna N Nakamura K Tanaka T Yamarnura H Ueno Y 1988
The acute and chronic toxicities of nivalenol in mice Fundamental and Applied
Toxicology 1138ndash47
Saier MH Roseman S 1972 Inducer exclusion and repression of enzyme synthesis in
mutants of Salmonella typhimurium defective in enzyme I of the phosphoenolpyruvate
sugar phosphotransferase system The Journal of Biological Chemistry 247972ndash975
Saier MHJ Tran CV Barabote RD 2006 TCDB the transporter classification database for
membrane transport protein analyses and information Nucleic Acids Research 34D181ndash
186
Saitou N Nei M 1987 The neighbor-joining method a new method for reconstructing
phylogenetic trees Molecular Biology and Evolution 4406ndash425
Savelieva O Kotova I Roelofsen W Stams AJM Netrusov A 2004 Utilization of
aminoaromatic acids by a methanogenic enrichment culture and by a novel Citrobacter
freundii strain Archives of Microbiology 181163ndash170
Schaafsma AW Tamburic-Ilinic L Miller JD Hooker DC 2001 Agronomic considerations
for reducing deoxynivalenol in wheat grain Canadian Journal of Plant Pathology 23279ndash
285
Schaafsma AW 2002 Economic changes imposed by mycotoxins in food grains case study
of deoxynivalenol in winter wheat Advances in Experimental Medicine and Biology
504271ndash276
203
Scharf C Riethdorf S Ernst H Engelmann S Voumllker U Hecker M 1998 Theoredoxin is an
essential protein induced by heat shock and other stresses in Bacillus subtilis Journal of
Bacteriology 1801869-1877
Schatzmayr G Zehner F Taubel M Schatzmayr D Klimitsch A Loibner AP Binder EM
2006 Microbiologicals for deactivating mycotoxins Molecular Nutrition and Food
Research 50543-551
Schollenberger M Muumlller H-M Ruumlfle R Suchy S Plank S Drochner W 2006 Natural
occurrence of 16 Fusarium toxins in grains and feedstuffs of plant origin from Germany
Mycopathologia 16143ndash52
Scientific Committee for Food 2002 Opinion on Fusarium toxins Part 6 Group evaluation
of T-2 toxin HT-2 toxin nivalenol and deoxynivalenol (available at http
wwweuropaeuintcommfoodfsscscfout123_enpdf)
Schulman S Wang B Li W Rapoport TA 2010 Vitamin K epoxide reductase prefers ER
membrane-anchored thioredoxin-like redox partners Proceedings of the National
Academy of Sciences 10715027ndash15032
Scott PM 1997 Multi-year monitoring of Canadian grains and grain-based foods for
trichothecenes and zearalenone Food Additives and Contaminants 14333-339
Seitz LM Bechtel DB 1985 Chemical physical and microscopal studies of scab-infected
hard red winter wheat Journal of Agricultural and Food Chemistry 33373-7
Sergent T Parys M Garsou S Pussemier L Schneider YJ Larondelle Y 2006
Deoxynivalenol transport across human intestinal Caco-2 cells and its effects on cellular
metabolism at realistic intestinal concentrations Toxicological Letters 164167ndash176
204
Sharma J Fulekar MH 2009 Potential of Citrobacter freundii for bioaccumulation of
heavy metalndashcopper Biology and Medicine 17-14
Sherif SO Salama EE Abdel-Wahhab MA 2009 Mycotoxins and child health the need for
health risk assessment International Journal of Hygiene and Environmental Health
212347ndash368
Shifrin VI Anderson P 1999 Trichothecene mycotoxins trigger a ribotoxic stress response
that activates c-Jun N-terminal kinase and p38 mitogen-activated protein kinase and
induces apoptosis The Journal of Biological Chemistry 27413985ndash13992
Shima J Takase S Takahashi Y Iwai Y Fujimoto H Yamazaki M Ochi K 1997 Novel
detoxification of the trichothecene mycotoxin deoxynivalenol by a soil bacterium isolated
by enrichment culture Applied and Environmental Microbiology 633825ndash3830
Singh J Behal A Singla N Joshi A Birbian N et al 2009 Metagenomics Concept
methodology ecological inference and recent advances Biotechnology Journal 4480ndash
494
Smith TK McMillan EG Castillo JB 1997 Effect of feeding blends of Fusarium mycotoxin-
contaminated grains containing deoxynivalenol and fusaric acid on growth and feed
consumption of immature swine Journal of Animal Science 752184ndash2191
Smits WK Dubois JYF Bron S van Dijl JM Kuipers OP 2005 Tricksy business
transcriptome analysis reveals the involvement of thioredoxin in redox homeostasis
oxidative stress sulfur metabolism and cellular differentiation in Bacillus subtilis Journal
of Bacteriology 1873921ndash3930
205
Sprando RL Collins TFX Black TN Olejnik N Rorie JI Eppley RM Ruggles DI 2005
Characterization of the effect of deoxynivalenol on selected male reproductive endpoints
Food Chemistry and Toxicology 43623ndash635
Starkey DE Ward TJ Aoki T Gale LR Kistler HC Geiser DM Suga H Toacuteth B Varga J
ODonnell K 2007 Global molecular surveillance reveals novel Fusarium head blight
species and trichothecene toxin diversity Fungal Genetics and Biology 441191ndash1204
Stevenson BS Schmidt TM 1997 Growth rate-dependent expression of RNA from plasmid-
borne rRNA operons in Escherichia coli Journal of Bacteriology 1801970ndash1972
Stothard P Wishart DS 2005 Circular genome visualization and exploration using
CGView Bioinformatics 21537ndash539
Suenaga H Ohnuki T Miyazaki K 2007 Functional screening of a metagenomic library for
genes involved in microbial degradation of aromatic compounds Environmental
Microbiology 92289-97
Surai P Mezes M Fotina TI Denev SD 2010 Mycotoxins in Human Diet A Hidden
Danger In Modern Dietary Fat Intakes in Disease Promotion Edited by F De Meester et
al Nutrition and Health DOI 101007978-1-60327-571-2_18
Sutton JC 1982 Epidemiology of wheat head blight and maize ear rot caused by Fusarium
graminearum Canadian Journal of Plant Pathology 4195
Swaving J De Bont JAM Westphal A De Kok A 1996 A novel type pyridine nucleotide-
disulfide oxidoreductase is essential for NAD+- and NADPH-dependent degradation of
epoxyalkanes by Xanthobacter strain Py2 Journal of Bacteriology 1786644ndash6
206
Swanson SP Rood JHD Behrens JC Sanders PE 1987 Preparation and characterization of
the deepoxy trichothecenes deepoxy HT-2 deepoxy T-2 triol deepoxy T-2 tetraol
deepoxy 15-monoacetoxyscirpenol and deepoxy scirpentriol Applied and Environmental
Microbiology 532821ndash2826
Swanson SP Helaszek C Buck WB Rood JHD Haschek WM 1988 The role of intestinal
microflora in the metabolism of trichothecene mycotoxins Food Chemistry and
Toxicology 26823ndash829
Tian F Yu Y Chen B Li H Yao Y-F Guo X-K 2009 Bacterial archaeal and eukaryotic
diversity in Arctic sediment as revealed by 16S rRNA and 18S rRNA gene clone libraries
analysis Polar Biology 3293-103
Tatusov RL Galperin MY Natale DA Koonin EV et al 2000 Artemis sequence
visualization and annotation Bioinformatics 16944-945
Trail F Haixin X Loranger R Gadoury D 2002 Physiological and environmental aspects of
ascospore discharge in Gibberella zeae (anamorph Fusarium graminearum) Mycologia
94181-189
Tsuge K Ohata Y Shoda M 2001 Gene yerP involved in surfactin self-resistance in
Bacillus subtilis Antimicrobial Agents and Chemotherapy 453566ndash3573
Tucker 2001 The ldquoyellow rainrdquo controversy Lesson for arms control compliance The
Nonproliferation Review 825-42
Ueno Y 1985 The toxicology of mycotoxins Critical Reviews in Toxicology 1499-132
Ueno Y Nakayama K Ishii K Tashiro F Minoda Y Omori T Komagata K 1983
Metabolism of T-2 toxin in Curtobacterium sp strain 114-2 Applied and Environmental
Microbiology 46120ndash127
207
van Elsas JD Speksnijder AJ van Overbeek LS 2008 A procedure for the metagenomics
exploration of disease suppressive soils Journal of Microbiological Methods 75515ndash522
van Ginkel CG de Bont JAM 1986 Isolation and characterization of alkene-utilizing
Xanthobacter spp Archives of Microbiology 145403ndash407
van Lanen SG Shen B 2006 Microbial genomics for the improvement of natural product
discovery Current Opinion in Microbiology 9252ndash260
Varga J Toacuteth B 2005 Novel strategies to control mycotoxins in feeds a review Acta
Veterinaria Hungarica 53189-203
Voigt CA Schafer W Salomon S 2005 A secreted lipase of Fusarium graminearum is a
virulence factor required for infection of cereals The Plant Journal 42364ndash375
Vogelgsang S Hecker A Musa T Dorn B Forrer HR 2011 On farm experiments over five
years in a grain maizewinter wheat rotation Effect of maize residue treatments on
Fusarium graminearum infection and deoxynivalenol contamination in wheat In Proc
7th
Canadian Workshop on Fusarium Head Blight edited by J Gilbert A Tekauz N
Sweetland and K Slusarenko Winnipeg Manitoba Canada p 41
Voumllkl A Vogler B Schollenberger M Karlovsky P 2004 Microbial detoxification of
mycotoxin deoxynivalenol Journal of Basic Microbiology 44147ndash156
Wagacha JM Muthomi JW 2007 Fusarium culmorum Infection process mechanisms of
mycotoxin production and their role in pathogenesis in wheat Crop Protection 26877-
885
Wakulinski W 1989 Phytotoxicity of the secondary metabolites of fungi causing wheat head
fusariosis (head blight) Acta Physiologiae Plantarum 11301ndash306
208
Wang JT Chang SC Chen YC Luh KT 2000 Comparison of antimicrobial susceptibility of
Citrobacter freundii isolates in two different time periods The Journal of Microbiology
Immunology and Infection 33258-62
Wannemacher RJ Stanley L Wiener MD 2000 Trichothecenes mycotoxins In Medical
aspects of chemical and biological warfare edited by Zajtchuk R Washington DC
Department of the Army pp 656-676
Westphal AH Swaving J Jacobs L de Kok A 1998 Purification and characterization of a
flavoprotein involved in the degradation of epoxyalkanes by Xanthobacter Py2 European
Journal of Biochemistry 257160ndash168
Whalen JG Mully TW Enlgish JC 2007 Spontaneous Citrobacter freundii infection in an
immunocompetent patient Archives of Dermatology 143124-5
White 2007 The physiology and biochemistry of prokaryotes Third edition Oxford
University Press New York Oxford p 628
Wilcke WF Morey RV 1995 Natural-air corn drying in the upper Midwest University of
Minnesota Minnesota Extension Service Publication BU-6577-E pp 20
Windels CE 2000 Economic and social impacts of Fusarium headblight changing farms and
rural communities in the Northern Great Plains Phytopathology 9017ndash21
Winzeler EA Shoemaker DD Astromoff A Liang H Anderson K Andre B Bangham R
Benito R Boeke JD Bussey H et al 1999 Functional characterization of the S
cerevisiae genome by gene deletion and parallel analysis Science 285901ndash906
Yang GH Jarvis BB Chung YJ Pestka JJ 2000 Apoptosis induction by the satratoxins and
other trichothecene mycotoxins relationship to ERK p38 MAPK and SAPKJNK
activation Toxicology and Applied Pharmacology 164149ndash160
209
Yang Q Yin GM Guo YL Zhang DF Chen SJ Xu ML 2010 A major QTL for resistance
to Gibberella stalk rot in maize Theoretical and Applied Genetics 121673ndash687
Yazar S Omurtag GZ 2008 Fumonisins trichothecenes and zearalenone in cereals
International Journal of Molecular Science 92062ndash2090
Yoshizawa T Morooka N 1973 Deoxynivalenol and its monoacetate new mycotoxins from
Fusaruium roseum and moldy barley Agricultural and Biological Chemistry 372933ndash
2934
Yoshizawa T Takeda H Ohi T 1983 Structure of a novel metabolite from deoxynivalenol a
trichothecene mycotoxin in animals Agricultural and Biological Chemistry 472133ndash
2135
Yoshizawa T 1993 Red-mold diseases and natural occurrence in Japan In Trichothecenes-
Chemistry Biology and Toxicological Aspects edited by Y Ueno Elsevier Amsterdam
Netherlands p 195-209
Yoshizawa T Sakamoto T Okamoto K 1984 In vitro formation of 3rsquo-hydroxy T-2 and 3rsquo-
hydroxy HT-2 toxins from T-2 by liver homogenates from mice and monkeys Applied
and Environmental Microbiology 47130-134
Young JC Subryan LM Potts D McLaren ME Gobran FH 1986 Reduction in levels of
deoxynivalenol in contaminated wheat by chemical and physical treatment Journal of
Agricultural and Food Chemistry 34461ndash465
Young JC Trenholm HL Friend DW Prelusky DB 1987 Detoxification of deoxynivalenol
with sodium bisulfite and evaluation of the effects when pure mycotoxin or contaminated
corn was treated and given to pigs Journal of Agricultural and Food Chemistry 35259ndash
261
210
Young JC Zhu H Zhou T 2006 Degradation of trichothecene mycotoxins by aqueous
ozone Food Chemistry and Toxicology 44417ndash424
Young JC Zhou T Yu H Zhu H Gong J 2007 Degradation of trichothecene mycotoxins by
chicken intestinal microbes Food Chemistry and Toxicology 45136-143
Yu Z Morrison M 2004 Comparisons of different hypervariable regions of rrs genes for use
in fingerprinting of microbial communities by PCR-denaturing gradient gel
electrophoresis Applied and Environmental Microbiology 704800-4806
Yu H Zhou T Gong J Young C Su X Li X Zhu H Tsao R Yang R 2010 Isolation of
deoxynivalenol-transforming bacteria from the chicken intestines using the approach of
PCR-DGGE guided microbial selection BMC Microbiology 10182
Zeibdawi A Steele M Savoie A 2011 An overview of CFIAs mycotoxin monitoring
program in livestock feeds In Proc 7th
Canadian Workshop on Fusarium Head Blight
edited by J Gilbert A Tekauz N Sweetland and K Slusarenko Winnipeg Manitoba
Canada p 25
Zhang J-B Li H-P Dang F-J Qu B Xu Y-B Zhao C-S Lia Y-C 2007 Determination of the
trichothecene mycotoxins chemotypes and associated geographical distribution and
phylogenetic species of the Fusarium graminearum clade from China Mycological
Research 111967ndash975
Zhao JS Manno D Thiboutot S Ampleman G Hawari J 2007 Shewanella canadensis sp
nov and Shewanella atlantica sp nov manganese dioxide- and hexahydro-135-trinitro-
135-triazine-reducing psychrophilic marine bacteria International Journal of Systemic
Evolution and Microbiology 572155ndash2162
211
Zhao X-Q 2011 Genome-based studies of marine microorganisms to maximize the diversity
of natural products discovery for medical treatments Evidence-Based Complementary
and Alternative Medicine Article ID 384572 doi1011552011384572
ZhaoY Park RD Muzzarelli RA 2010 Chitin deacetylases properties and applications
Marine Drugs 824ndash46
Zhou HR Lau AS Pestka JJ 2003a Role of double-stranded RNA-activated protein kinase R
(PKR) in deoxynivalenol-induced ribotoxic stress response Toxicological Science
74335ndash344
Zhou HR Islam Z Pestka JJ 2003b Rapid sequential activation of mitogen-activated protein
kinases and transcription factors precedes proinflammatory cytokine mRNA expression in
spleen of mice exposed to the trichothecene vomitoxin Toxicological Sciences 72130ndash
142
Zhou T He J Gong J 2008 Microbial transformation of trichothecene mycotoxins World
Mycotoxin Journal 123-30
Zhou G Zhang H He Y He L 2010 Identification of a chitin deacetylase producing bacteria
isolated from soil and its fermentation optimization African Journal of Microbiology
Research 42597-2603

v
Agricultural Research and Development Fund (AAFCBARD) project for providing me
financial support
Mike Bryan a very supportive person helped provide solutions to my software issues
that improved my exposure and caliber at conference awards competitions Several student
competition awards honored and inspired me to work hard and lead to the successful
completion of my thesis research I would like to convey my appreciation to the judges of
different student award committees who recognized my work They are Society for Industrial
Microbiology and Biotechnology conference 2010 (USA) Food Safety Research Forum-
Ontario Ministry of Agriculture and Rural Affairs annual meeting 2010 (Canada)
International Mycological Conference 2009 (UK) and Southwestern Ontario Regional
Association of Canadian Phytopathological Society annual meeting 2009-2011 (Canada)
I would like to express my very special thanks to Prof Golam Hafeez Kennedy my
brother-in-law who is like my elder brother In many aspects I am indebt to him for his
cooperations which aided me to achieve my current status
Lastly and most importantly I wish to give my special thanks to my wife Mimi and
lovely daughter Racy whose endless love sacrifice support and encouragement enabled me
to cross the tough long road and made possible to achieve my goal and aspirations Racy
once you told me that one of your desires for me is to finish my PhD which is done now I
know you want to be a scientist which is a great ambition I wish you all the best in your
future aims Racy
vi
TABLE OF CONTENTS
DEDICATIONShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipiii
ACKNOWLEDGEMENTShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipiv
TABLE OF CONTENTShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipvi
LIST OF TABLEShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipix
LIST OF FIGUREShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipx
LIST OF ABBREVIATIONShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipxiii
10 GENERAL INTRODUCTION AND LITERATURE REVIEW helliphelliphelliphelliphelliphelliphelliphellip1
11 Backgroundhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip1
12 Plant pathogenesis and trichothecene productionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip3
13 Biosynthesis of food contaminating TChelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip6
14 Occurrence of TC mycotoxins in food and feed helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip11
15 Significance of deoxynivalenol and trichotheceneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip16
16 Toxic effects of deoxynivalenol and trichotheceneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip19
17 Cellular and molecular mechanisms of DON toxicity in animals and plantshelliphelliphelliphellip22
18 Strategies to control Fusarium head blight and trichotheceneshelliphelliphelliphelliphelliphelliphelliphelliphellip26
19 Isolation of DONTC de-epoxidating micro-organisms and geneshelliphelliphelliphelliphelliphelliphelliphellip28
110 The bacterial species Citrobacter freundiihelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip30
111 Hypotheses and objectiveshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip31
vii
20 AEROBIC AND ANAEROBIC DE-EPOXIDATION OF MYCOTOXIN
DEOXYNIVALENOL BY BACTERIA ORIGINATING FROM AGRICULTURAL
SOIL33
21 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip33
22 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
23 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip37
24 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip45
25 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip62
26 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip64
30 ISOALTION AND CHARACTERIZATION OF A NOVEL SOIL BACTERIUM
CAPABLE OF DETOXIFYING ELEVEN TRICHOTHECENE MYCOTOXINS65
31 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip65
32 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip66
33 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip68
34 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip75
35 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip96
36 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip100
40 CONSTRUCTION AND FUNCTIONAL SCREENING OF GENOMIC DNA
LIBRARY FOR ISOLATION OF DON DE-EPOXYDIZING GENE(S) FROM
BACTERIAL STRAIN ADS47helliphelliphelliphelliphelliphelliphelliphellip102
41 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip102
42 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip103
43 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip104
viii
44 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip112
45 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip118
46 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip121
50 GENOME SEQUENCING CHARACTERIZATION AND COMPARATIVE
ANALYSIS OF A TRICHOTHECENE MYCOTOXIN DETOXIFYING
CITROBACTER FREUNDII STRAIN ADS47helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip122
51 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip122
52 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip123
53 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip125
54 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip129
55 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip162
56 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip168
60 CONCLUSIONhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip171
61 Summary helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip171
62 Research contributions helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip174
63 Future workhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip175
70 REFERENCEShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip176
ix
LIST OF TABLES
Table 1-1 Fusaria produced type-A and type-B trichothecene mycotoxinshelliphelliphelliphelliphelliphelliphellip9
Table 1-2 Worldwide occurrence of deoxynivalenol (DON) in animal feedshelliphelliphelliphelliphellip12
Table 1-3 Occurrence of DON in Canadian grains grains foods and beershelliphelliphelliphelliphelliphelliphellip13
Table 1-4 Legislative maximum tolerated levels of trichothecene mycotoxins in Canadahellip14
Table 1-5 In vitro and in vivo synergistic effects of different food contaminating
mycotoxins helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip18
Table 3-1 Microbial transformation of type-A and type-B tricthothecene mycotoxins
after 72 h of incubation under aerobic conditions and at room temperaturehelliphelliphelliphelliphelliphellip95
Table 4-1 Analysis of Fosmid DNA library for deoxynivalenol (DON) de-epoxidation
capability helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip117
Table 5-1 General features of C freundii strain ADS47 genomehelliphelliphelliphelliphelliphelliphelliphelliphellip132
Table 5-2 Genome comparisons between strain ADS47 and five Citrobacter species141
Table 5-3 Functional categorized gene abundances among the six Citrobacter genomeshellip145
Table 5-4 Unique genes of C freundii strain ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip156
Supplemental Table S5-1 List of unique genes of strain ADS47 gene IDs and their
functional roleshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip169
x
LIST OF FIGURES
Figure 1-1 Structures of trichothecene mycotoxins produced by Fusariahelliphelliphelliphelliphelliphelliphelliphellip7
Figure 1-2 Cellular and molecular mechanisms of deoxynivalenol toxicity in
eukaryotic organismshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip23
Figure 2-1 Mechanism of microbial DON de-epoxidationhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip36
Figure 2-2 LC-UV-MS chromatographic analysis of DON and its de-epoxidated product
dE-DON obtained by culturing with selected soil microbeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip46
Figure 2-3 Time course of relative biotransformation of DON to dE-DON by the initial
soil suspension culture in MSB mediumhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip49
Figure 2-4 Microbial de-epoxidation of DON after five days of incubationhelliphelliphelliphelliphelliphellip51
Figure 2-5 Analysis of microbial population reduction by terminal restriction fragment
length polymorphism (T-RFLP)helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip53
Figure 2-6 Microbial genera identified in the enriched DON de-epoxidizing microbial
culturehelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip56
Figure 2-7 Effects of culture conditions on DON to dE-DON transformation by the
enriched microbial culturehelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip58
Figure 2-8 Microbial DON to dE (de-epoxy)-DON transformation activity under aerobic
and anaerobic conditionshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip61
Figure 3-1 Colony color shape and size of the mycotoxin degrading bacterial strain
ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip76
Figure 3-2 Dendrogram based on 16S-rDNA sequences for strain ADS47 and 11
reference Citrobacter specieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip77
xi
Figure 3-3 Effects of DON concentration on bacterial growth and de-epoxidation
activitieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip79
Figure 3-4 Bacterial growth and DON de-epoxidation capabilities under aerobic and
anaerobic conditions helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip82
Figure 3-5 Effects of media on bacterial growth and DON de-epoxidation activityhellip85
Figure3-6 Effects of temperature on microbial growth and DON de-epoxydation helliphelliphellip87
Figure 3-7 Influence of pH on microbial growth and DON de-epoxydation capabilityhellip89
Figure 3-8 Effects of sodium azide (SA) on bacterial DON de-epoxidation activityhelliphelliphellip91
Figure 3-9 Determination of DON to deepoxy-DON transformation by bacterial culture
filtrate and cell extracthelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip92
Figure 3-10 Determination of microbial de-epoxidation and de-acetylation of a type-A
trichothecene mycotoxin (HT-2 toxin) by LC-UV-MShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip94
Supplemental Figure S3-1 Flow chart depicted the procedures of enrichment and isolation
of DON de-epoxidation bacterial strain ADS47helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip101
Figure 4-1 Schematic overview of the construction and functional screening of Fosmid
DNA libraryhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip105
Figure 4-2 Genetic map of copy controlled Fosmid vector (pCC1FOS)helliphelliphelliphelliphelliphelliphellip108
Figure 4-3 Preparation of 40 kb size insert DNA from a DON de-epoxidizing bacterium113
Figure 4-4 Production of genomic DNA library in E coli EPI300-T1 host strainhelliphelliphelliphellip114
Figure 4-5 Confirmation of the Fosmid DNA library construction in E coli helliphelliphelliphelliphellip 116
Figure 5-1 Co-linearity between C freundii strain ADS47 genome and C koseri helliphelliphellip129
Figure 5-2 Circular genome map of C freundii strain ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphellip133
xii
Figure 5-3 Size distribution of the predicted genes identified in C freundii strain
ADS47helliphelliphellip 135
Figure 5-4 Functional classification of the predicted coding sequences (CDS) of
C freundii strain ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip137
Figure 5-5 Phylogeny of the trichothecene detoxifying C freundii strain ADS47 139
Figure 5-6 Comparisons of coding sequences between C freundii strain ADS47 and
five Citrobacter specieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip143
Figure 5-7 Determination of the locus enterocyte effacement (LEE) operons in
Citrobacter species by comparing C rodentium helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip151
Figure 5-8 Locations of unique genes of C freundii strain ADS467helliphelliphelliphelliphelliphelliphelliphellip157
Figure 5-9 Unique genomic regions and associated genes in strain ADS47 compared
to five Citrobacter specieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip159
xiii
LIST OF ABBREVIATIONS
APCI= atmospheric pressure chemical ionization
AE= attaching and effacing
A= aerobic conditions
ABC= ATP-binding cassette
ACT= artemis comparative tools
AN= anaerobic conditions
3-ADON= 3-acetyl-deoxynivalenol
15-ADON= 15-acetyl-deoxynivalenol
ATA= alimentary toxic aleukia
bp= base pair
BGI= Beijing genomics institute
BLAST= basic local alignment search tool
BRIG= blast ring image generator
Bw= body weight
C= carbon
CDS= coding sequence
CFIA= Canadian food inspection agency
CHEF= clamped homogeneous electric fields
CMB= corn meal broth
CVTree= composition vector tree
DAS= diacetoxyscirpenol
dH2O= distilled water
DOGT1= UDP-glucosyltransferase
DON= deoxynivalenol
dsRNA= double stranded RNA
dE= de-epoxy
dE-DON= de-epoxy DON
E coli= Escherichia coli
ER=endoplasmic reticulum
FAO= Food amp Agriculture Organization
FM= full medium
FHB= Fusarium head blight
FUS= fusarenonX
gDNA= genomic DNA
GC= guanine cytosine
GER= Gebberella ear rot
GPI= glycosylphosphatidylinositol
h= hour
Hck= hematopoietic cell kinase
Kb= kilobase
xiv
Kg= kilogram
LC-UV-MS= liquid chromatography-ultraviolet-
mass spectrometry
LEE= locus of enterocyte effacement
LD50= lethal dose of 50 animal
LB= Luria Bertani
LMP= low melting point
MAPK= mitogen-activated protein kinase
microg= microgram
mg= milligram
mL= milliliter
microL= microliter
microM= micromole
min= minute
Mb= megabases
MCS=multiple cloning site
MSA= MSB+ agar
MSB= mineral salts broth
MM= minimal medium
MSC= mineral salts + 03 Bacto Peptone
MS-DON= mineral salts + 200 microgmL DON
mRNA= messenger RNA
MSY= mineral salts + yeast extract
mz= mass ion ratio
N= nitrogen
Na-Cl= sodiumndashclorine
NADH= (reduced) nicotinamide adenine
dinucleotide
NAD(P)= nicotinamide adenine
dinucleotide phosphate
NCBI= national center for biotechnology
information
NEO= neosolaniol
ng= nanogram
NIV= nivalenol
OD600nm= optical density at 600
nanometer wavelength
OMAFRA= Ontario Ministry of
Agriculture and Rural Affairs
Paa= phenylacetic acid
PCR= polymerase chain reaction
PDB= potato dextrose broth
ppm= parts per million
PFGE= pulsed field gel electrophoresis
PKR= protein kinase
xv
PNDO= pyridine nucleotide-disulfide
oxidoreductase
QTL= quantitative trait locus
RAST= rapid annotation subsystem technology
RDP= ribosomal database project
Rpl3= 60S ribosomal protein L3
rRNA= ribosomal RNA
s= second
SA= sodium azide
SDR= short chain dehydrogenasereductase
SE= soil extract
SIM= selected ion monitoring
SOAP= short oligonucleotide analysis package
sp= species
sRNA= small RNA
TC= trichothecene
TCA= tricarboxylic acid cycle
TSB= tryptic soy broth
TF= transcription factor
tRNA= transfer RNA
TIC= total ion count
TRAP= tripartite ATP-independent
periplasmic
T-RFLP=terminal restriction fragment
length polymorphism
TRI101= trichothecene 3-O-
acetyltransferase
YEB= yeast extract broth
VER= verrucarol
VKOR= vitamin K epoxide reductase
wv= weight by volume
1
CHAPTER 1
10 GENERAL INTRODUCTION AND LITERATURE REVIEW
11 Background
According to the Food and Agriculture Organization (FAO) of the United Nations
more than 25 of the worldrsquos food crops are contaminated with mycotoxins every year
causing serious adverse effects on livestock industry crop production and international trade
(Boutrif and Canet 1998 Desjardins 2006 Binder et al 2007 Surai et al 2010) The negative
effects of mycotoxins on the consumers are diverse including organ toxicity mutagenicity
carcinogenicity neurotoxicity teratogenicity modulation of the immune system reproductive
system effects and growth retardation Mycotoxins are toxic secondary metabolites produced
by various fungal species during crop colonization in field andor post harvest conditions
(FAO 1991 Barrett 2000) Aspergillus Penicillum and Fusarium are the predominant fungal
genera that accumulate economically important mycotoxins such as aflatoxins zearalenone
trichothecenes fumonisins ochratoxins and ergot alkaloids in agricultural commodities
Humans and animals are principally exposed to mycotoxins through consumption of
contaminated foodfeed and animal products such as milk meat and eggs (Fink-Gremmels
1989 Hollinger and Ekperigin 1999)
Trichothecenes (TCs) produced by a variety of toxigenic Fusarium species are
probably the most food and feed contaminating mycotoxins in the temperate regions of
America Europe and Asia (Creppy 2002 Yazar and Omurtag 2008) The fungal pathogen
accumulates various type-A (eg T-2 toxin HT-2 toxin) and type-B trichothecenes (eg
2
deoxynivalenol nivalenol and their derivatives) in cereals Among them type-B
trichothecenes particularly deoxynivalenol (DON) andor its derivatives appear to be the
most commonly found mycotoxins in grain and grain products worldwide impairing human
health and productivity of farm animals (Placinta et al 1999 Sherif et al 2009) A number of
human and animal disease outbreaks associated with TC contaminated grain food and feed
consumption have been reported in Asia (Japan India and central Asia) Europe New
Zealand and United States of America (USA) (Pestka and Smolinski 2005 Desjardins 2006)
A severe outbreak of Alimentary Toxic Aleukia (ATA) mycotoxicosis killed 100000 people
in the Orenburg region of Russia (Eriksen 2003) In 1996 losses of over $100 million were
estimated in Ontario because of costs related to managing DON (Schaafsma 2002)
Historically T-2 toxin DON and nivalenol (NIV) have likely been used as a biological
warfare in Laos Kampuchea and Afghanistan and there is concern because of the potential
use of these compounds for bioterrorism (Wannemacher et al 2000 Tucker 2001)
Despite the extensive efforts that have been made to prevent TC contamination with
the current varieties and production practices its presence in agricultural commodities is
likely unavoidable There are no crop cultivars currently available that are completely
resistant to Fusarium head blightGibberella ear rot (FHBGER) diseases Chemical or
physical methods cannot safely remove TC mycotoxins from grain completely or they are not
economically viable (Varga and Toacuteth 2005) Therefore an alternative strategy to address the
issue is of great importance (Awad et al 2010 Karlovsky 2011)
3
12 Plant pathogenesis and trichothecene production
Cereals infested by the ascomyceteous fungi may cause Gibberella ear rot (GER) in
maize (Zea mays L) and Fusarium head blight (FHB also known as lsquoscabrsquo) of small grain
cereals [wheat (Triticum aestivum L) barley (Hordeum vulgare L)] and other small grain
cereals (Goswami and Kistler 2004) As many as 17 fungal species have been reported to be
associated with FHBGER which include F graminearum Schwabe [telemorph Gibberella
zeae (Schwein) Petch] F culmorum (WG Smith) Sacc F sporotrichioides Scherb F
poae (Peck) Wollenw Microdochium nivale (Fr) Samuels amp IC Hallett and F avenaceum
(Fr) Sacc (Osborne and Stein 2007 Audenaert et al 2009) The two species F
graminearum and F culmorum are the principal species causing crop damage Among them
F graminearum is the widely adapted and most damaging species across the world including
North America (Dyer et al 2006) F culmorum tend to be the main species in the cooler parts
of North Central and Western Europe (Wagacha and Muthomi 2007)
The fungi overwinter mainly on crop residues as saprophytes where they produce
asexual (micro- and macro-conidia) andor sexual (ascospores) fruiting structures which act
as the initial source of inoculums in the subsequent crop season (Sutton 1982 Wagacha and
Muthomi 2007) Ascospores are primarily dispersed by air (Trail et al 2002) and conidia
move by rain-splash (Paul et al 2004) After they land on maize silks and florets of small
grain cereals infection occurs through natural openings (eg stomata) or occasionally by
direct penetration The pathogens can invade maize kernels through wounds caused in the
husks by insects or birds Infection may occur throughout the crop growing season but
disease incidence and severity is facilitated by warm temperatures (gt20degC) high humidity
(gt80) and frequent precipitation that coincides with anthesis (Osborne and Stein 2007
4
Wagacha and Muthomi 2007) The Fusarium pathogen initially infects as biotroph which
becomes a necrotroph as the disease progresses After penetration Fusaria produce different
extracellular cell wall degrading enzymes that trigger infection and hyphal progression
towards the developing earkernel where further colonization occurs and development of
FHBGER diseases (Voigt et al 2005 Miller et al 2007 Kikot et al 2009) FHB disease is
characterized by a bleached appearance around the middle of the head Severely infected
grains have a pinkish discoloration and can be shriveled In maize the disease produces a
reddishndashbrown discoloration of the kernels and progresses from the tip to the bottom of
affected ears
The fungi associated with FHB and GER contaminate cereals by accumulating various
host-nonspecific TC mycotoxins (Brennen et al 2007) TCs have typically been associated
with cereals grown in temperate climatic conditions including Canada (Doohan et al 2003)
DON production by F graminearum and F culmorum is maximal at warm temperatures (22-
28˚C) high grain moisture contents (gt17) and humid conditions (gt70 RH) (Martins and
Martins 2002 Hope et al 2005 Llorens et al 2006) Grain can be contaminated with TCs in
the field as well as in storage if conditions for fungal growth are optimal and grain contains
high moisture (gt17) (Larsen et al 2004 Birzele et al 2002)
Populations of FHBGER causing pathogens are dynamic Genetic variability between
and within Fusarium species appears to have occurred to allow for their adaptation to new
agro-climates (Desjardins 2006 Fernando et al 2011) Three strain specific Fusarium
chemotypes NIV 3-ADON (3-acetyl-deoxynivalenol) and 15-ADON (15- acetyl-
deoxynivalenol) have been identified which correlate with the geographical distributions
(Starkey et al 2007 Zhang et al 2007 Audenaert et al 2009) The NIV chemotype mainly
5
produced by F culmorum accumulates NIV and 4-NIV (Fusarinon X) is predominant in
Asia Africa and Europe (Lee et al 2001) The 3-ADON chemotype which synthesizes DON
and 3-ADON is the dominant strain in EuropeFar East (Larsen et al 2004) In North
America 15-ADON is the main chemotype which produces DON and 15-ADON (Goswami
and Kistler 2004) Significant Fusarium population shifting was observed in the past and in
recent years (Kant et al 2011) A dynamic change from F culmorum to F gramninearum
and DON to NIV producing chemotyes occurred in North West European countries The 3-
ADON chemotype which is uncommon in North America was found in North Dakota and
Minnesota (Goswami and Kistler 2004) F pseudograminearum which historically occurs in
Australia Africa and North West Pacific America has been found in Western Canada (Larsen
et al 2004 Kant et al 2011) In Manitoba a replacement of 15-ADON chemotype by the
more phytoxic 3-ADON type was observed in recent years (Fernando et al 2011) All
together the FHBGER associated pathogen complex is highly dynamic which not only
reduces crop yield but also decreases grain quality by accumulating various mycotoxins
6
13 Biosynthesis of food contaminating TC
TCs are fungal secondary metabolites that were initially identified in Trichotheceium
roseum Thus the name of this class of compounds follows the name of the genus
Trichotheceium (Freeman and Morrison 1949) There have been 180 TCs identified in various
fungal species belonging to the genera Fusarium Myrothecium Stachybotrys Trichoderma
and Thrichothecium (Eriksen 2003 Eriksen et al 2004) The FHBGER associated TCs are
mainly synthesized by different Fusaria and are characterized as tricyclic sesquiterpenoids
with a double bond at position C9-10 and a C12-13 epoxide ring
7
Figure 1-1 Structures of trichothecene mycotoxins produced by Fusaria (a) General
structure of trichothecenes R1 = H OH OiValeryl keto R2 = H OH R3= OH OAc R4 =
H OH OAc R5= OH OAc (b) Type-A trichothecene mycotoxin (c) Type-B trichothecene
mycotoxin which contain a carbonyl functional group at C8 position (Young et al 2007)
(a) (b)
(c)
8
The fungal metabolites are small in size (molecular weight 250-550) heat stable and
non-volatile molecules (Pestka 2007 Yazar and Omurtag 2008) Food contaminating TCs are
divided into two groups type-A and type-B (Figure 1-1b and 1c) Type-A is characterized by
the lack of a ketone function at C8 position and include T-2 toxin HT-2 toxin
diacetoxyscirpenol (DAS) and neosolaniol (NEO) which are mainly produced by F
sporotrichioides and F poae (Creppy 2002 Quarta et al 2006) The most frequently found
TC belongs to type-B which contains carbonyl functions at C8 positions DON NIV and
their acetylated derivatives The two types are further differentiated by the presence or
absence of functional hydroxyl (-OH) and acetyl (CH3-CO-) groups at C-3 C-4 C-7 C-8 and
C-15 positions (Table 1-1)
The syntheses of various TCs are speciesstrain specific and are genetically controlled
but influenced by environmental conditions (Desjardins 2006 Paterson and Lima 2010) Until
now 15 Fusaria catalytic genes and regulators involved in TC productions have been
identified (Merhej et al 2011) These biosynthetic genes are located at three loci on different
chromosomes The core trichothecene (TRI) gene cluster is comprised of 12 genes localized
in a 25-kb genomic region (Kimura et al 2001) The remaining three genes are encoded by
the TRI1-TRI16 and TRI101 loci (Gale et al 2005 Alexander et al 2009) Fusaria produce
TC following a number of oxygenation isomerization acetylation hydroxylation and
esterification reaction steps (Desjardins 2006 Desjardins and Proctor 2007 Kimura et al
2007 Merhej et al 2011) In brief farnesyl pyrophosphate is initially cyclized into
trichodiene by TRI5 which codes for trichodiene synthase In the next four steps
isotrichotriol is produced by TRI4 (a multifunctional cytochrome P450) via C-2 hydroxylation
and 1213 epoxidation followed by two hydroxylation reactions
9
Table 1-1 Fusaria produced type-A and type-B trichothecene mycotoxins Molecular weight
(MW) and acetyl (-R) hydroxyl (-OH) and ketone (=O) functions at different carbon (C)
position of the tricyclic rings A-types T-2 toxin HT-2 toxin DAS (diacetoxyscirpenol)
NEO (neosolaniol) VER (verrucarol) B-type DON (deoxynivalenol) 3-ADON (3-acetyl-
deoxynivalenol) 15-ADON (15-acetyl-deoxynivalenol) NIV (nivalenol) and FusX
(fusarenon X) (Young et al 2007)
Trichothecenes MW R1C3 R2C4 R4C7 R3C15 R5C8
Type-A
T-2 toxin 466 OH OCOCH3 H OCOCH3 OCOCH2CH(CH3)2
HT-2 toxin 424 OH OH H OCOCH3 OCOCH2CH(CH3)2
DAS 366 OH OCOCH3 H OCOCH3 H
NEO 382 OH OCOCH3 OH OCOCH3 H
VER 266 H OH H OH H
Type-B
DON 296 OH H OH OH =O
3-ADON 338 OCOCH3 H OH OH =O
15-ADON 338 OH H OH OCOCH3 =O
NIV 312 OH OH OH OH =O
FusX 354 OH OCOCH3 OH OH =O
10
The next two non-enzymatic reactions results in isotrichodemol Eventually the
trichothecene skeleton is produced via the formation of a C-O bond between the C-2 oxygen
and C-11 Subsequent acetylation at C-3 by TRI101 hydroxylation at C-15 by TRI1 and
catalytic activity of TRI3 leads to calonectrin production The pathway described above is
common for both type-A (eg T-2 toxin HT-2 toxin) and type-B (eg DON NIV and their
acetylated derivatives) TCs
The following steps are speciesstrain specific In DON producing F graminearum
strains calonectrin is converted to either 3-ADON or 15-ADON followed by DON production
by the catabolic activities of TRI1 and TRI8 The biosynthesis of 3-ADON or 15-ADON is
controlled by variation in the esterase coding sequences of TRI8 F graminearum produces
NIV from calonectrin via synthesis of 4-NIV followed by NIV by TRI1 and TRI8 gene
actions NIV can be produced either from DON by hydroxylation at C-4 or via
transformation of calonectrin to 315-diacetoxyscirpenol (315-DAS) followed by addition of
a ketone at C-8 position by the activity of the TRI7 and TRI13 genes 315-DAS is the
principal substrate for synthesis of type-A TCs by F sporotrichioides the main type-A
producer The biosynthetic pathway of T-2 toxin involves C-4 acetylation of 315-DAS by the
TRI7 gene product and TRI8 triggered by TRI1 315-DAS is also the substrate for HT-2
toxin synthesis In the absence of TRI7 expression HT-2 toxin is produced through
hydroxylation of 315-DAS and isovalerate addition to C-8 position (Foroud and Eudes 2009)
11
14 Occurrence of TC mycotoxins in food and feed
TC producing Fusaria are adapted to variable climatic conditions and agricultural
ecosystems As a consequence this class of mycotoxins particularly DON is frequently
detected in foods and feeds including breakfast cereals bakery products snack foods beer
and animal feed derived from cereals in most part of the world (Pittet 2001 Desjardins 2006
Lancova et al 2008) A two-year survey during 2003-2005 conducted by Romer Lab Inc
revealed unacceptable levels of DON in animal feedsfeed raw materials (such as maize
wheat wheat bran) in Asia and Oceania (1062 microgg) central Europe (802 microgg) Mid-West
USA (55 microgg) northern Europe (551 microgg) and southern Europe (303 microgg) (Binder et al
2007) (Table 1-2)
Quite high levels of DON contamination (19 microgg) were detected in north Asian and
Chinese feedfeed materials Recently five-year on-farm studies in Switzerland showed that
74 of the wheat grain and straw samples were contaminated with high levels of DON
(average 5 microgg) The amount is higher than the European maximum limit of 125 microgg
(Vogelgsang et al 2011)
The occurrences of TCs in Canadian agricultural commodities are indicated in Table
1-3 Seventeen surveys conducted during the 1980s and 1990s showed up to 100 DON
incidence in Ontario wheat and maize samples with an 18 average incidence (Scott 1997) A
three-year survey recently (2006-08) conducted by the Ontario Ministry of Agriculture and
Rural Affairs (OMAFRA) detected high levels of DON in spring (maximum 26 microgg) and
winter wheat (maximum 29 microgg) samples (Ana Matu personal communication) The DON
levels were much higher than Health Canadarsquos maximum limit of 2 microgg for uncleaned soft
wheat for staple foods (Table 1-4)
12
Table 1-2 Worldwide occurrence of DON in animal feed Feed and feed raw material
samples were collected from animal farmsfeed production sites during 2003-2005 The
survey was conducted by Romer Labs Inc (httpwwwromerlabscom)
Country
region
Sample
type
Incidence
()
Maximum
DON content
(microgg)
Mean DON
content
(microgg)
Survey
period
Source
North
AsiaChina
Animal
feed
071 1899 092
Binder et
al 2007
South East
Asia
014 152 016
South Asia 070 1062 049
Oceania 2003-05
Australia 041 186 030
Northern
Europe
070 551 059
Central
Europe
066 802 057
Southern
Europe
052 303 030
13
Table 1-3 Occurrence of DON in Canadian grains grain foods and beers Infant cereal foods
oat barley soy and multi-grain based foods
Province Sample
type
Incidence
()
Maximum
DON
contents
(microgg)
Multiple
mycotoxins
samples ()
Survey
period
Sources
Ontario Spring
Wheat
70 260 Yes 2006-08 OMAFRA1
Ontario Winter
wheat
49 290 Yes 2006-08 OMAFRA
Ontario Maize 78 140 Yes 2009-10 Limay-Rios
and
Schaafsma
2010
Ontario Maize - 1000 Yes 2011 AampL
Biologicals
Canada Breakfast
cereals
46 094 430 1999-01 Roscoe et al
2008
Canada Animal
feeds
73 425 Yes 1990-10 Zeibdawi et
al 2011
Canada Beers 58 0052 50 1990s Scott 1997
Canada Infant
Cereals
63 098 Yes 1997-99 Lombaert et
al 2003
1
Ontario Ministry of Agriculture and Rural Affairs
2 microgmL
14
Table 1-4 Legislative maximum tolerated levels of trichothecene mycotoxins in Canada
according to Canadian Food Inspection Fact Sheet April 23 2009 DON- deoxynivalenol
DAS- diacetoxyscirpenol
Products DON
(microgg)
T-2 toxin
(microgg)
HT-2 toxin
(microgg)
DAS
(microgg)
Uncleaned soft wheat for human
consumption
2 - - -
Cattlepoultry diets 5 lt1 01 lt1
Swinedairy cattle feeds 1 lt1 0025 lt2
15
High DON incidence (78) was observed in maize samples collected from
southwestern Ontario fields with moderate levels (lt1 microgg) of contamination during 2009 and
2010 (Limay-Rios and Schaafsma 2010) Some samples had high DON levels (14 microgg) In
the 2011 crop season extremely high levels of DON (gt100 microgg) were found in some
southern Ontario maize samples (AampL Biologicals London ON Personal communication)
In a Canadian Food Inspection Agency (CFIA) 20 year survey (1990-2010) 73 of
animal feed samples collected from feed mills farms and retail stores across Canada were
contaminated with DON and for more than 25 of the samples the mycotoxin levels were
above the maximum limit (ge 1 microgg) (Zeibdawi et al 2011) The highest level of DON (15
microgg) detected in the 2006-2007 samples were much lower than the levels found in the 1999-
2000 samples which contained a maximum of 425 microgg DON Importantly this study
reported a significant number of samples contaminated with multiple mycotoxins DON NIV
3-ADON 15-ADON T-2 toxin HT-2 toxin DAS and NEO
Earlier studies in the 1990s detected high levels of DON content in Eastern Canadian
cereals maize (maximum 175 microgg) wheat (maximum 916 microgg) barley (maximum 911
microgg) and oats (maximum 12 microgg) (Campbell et al 2002) The study identified a significant
number of samples (15-367) that contained multiple mycotoxins and up to 33 of the
wheat samples exceeded 1 microgg DON Low amounts but high incidences of TCs were found
in cereal-based foods collected from Canadian retail stores Sixty three percent of infant
cereal foods 46 of breakfast cereals and 58 of beers were contaminated with one or
multiple mycotoxins (Scott 1997 Lombaert et al 2003 Roscoe et al 2008) The maximum
amount of DON was found in infant foods (098 microgg) followed by breakfast cereals (094
microgg) and beers (005 microgmL)
16
15 Significance of deoxynivalenol and trichothecenes
For most cereals including wheat maize rice barley oats and rye the risk of
contamination with TCs resulted in grain quality reduction and FHBGER disease progression
(Bai et al 2002 Jansen et al 2005) The impacts of TC contamination in agricultural
commodities are diverse including reduced livestock production induced human illnesses
and enhanced FHB disease severity which may result in the reduction of crop yield Exposure
of farm animals (particularly pigs) to DON is a continuing threat to the livestock industries in
Canada USA and Europe (DrsquoMello et al 1999) Ingestion of TC-contaminated feeds
generally causes reduction of feed intake feed refusal poor feed conversion retardation of
weight gain increased disease susceptibility and reduced reproductive capacities (Binder et al
2007) High doses of DON result in acute toxic effects in pigs (eg emesis) while low doses
(ge1 microgmL) cause chronic effects (eg growth retardation in pigs) (Rotter et al 1996
Accensi et al 2006) Reduction of growth and reproduction ability was observed in
experimental rats exposured to much higher doses of TC (Sprando et al 2005 Collins et al
2006) Several animal disease outbreaks that occurred in different continents have been
attributed to consumption of feed contaminated with DON NIV and DAS (Desjardins 2006)
The consequences of human exposure to TCs include liver damage gastricintestinal
lesions and central nervous system toxicity resulting in anorexia nausea hypotensionshock
vomiting diarrhea and death (Pestka 2007) Human mycotoxicosis (illnesses and outbreaks)
by consumption of type-A and type-B TC mycotoxins in Europe New Zealand Russia
Japan South America India and Vietnam have been reported (Bhat et al 1989 Yoshizawa
1993 Luo 1994 Desjardins 2006) Severe outbreaks of Alimentary Toxic Aleukia (ATA)
associated with TC killed thousands of people in the Orenburg region of Russia during the
17
1930rsquos and 1940rsquos (Eriksen 2003) TCs (T-2 DON and NIV) were likely used for biological
warfare in Laos Kampuchea and Afghanistan and its potential application for bioterrorism is
an additional concern associated with TC (Tucker 2001 Bennett and Klich 2003)
TCs are phytotoxic and may contribute to FHB disease severity in cereals (Proctor et
al 1995 Maier et al 2006 Masuda et al 2007) Studies demonstrated that DON caused
wilting chlorosis inhibition of seedling growth and necrosis in cereals (Cossette and Miller
1995 Mitterbauer et al 2004 Rocha et al 2005) DON-mediated host cell damage and
subsequent necrotrophic fungal spread has been shown to lead to FHBGER disease
development and higher kernel damage (Procter et al 1995 Mitterbauer et al 2004 Maier et
al 2005) This suggests that DON contributes to yield losses as well as to reductions in the
market value of cereals (Osborne and Stein 2007) Severely infected grain may contain more
than 60 microgg DON which is unacceptable to the milling and brewing industries (Okubara et
al 2002) Significant crop losses due to FHB disease epidemics and mycotoxin contamination
have been reported in Ontario (Schaafsma 2002) FHB disease epidemics during 1998 to 2000
caused huge crop losses resulting in farm failures in the northern Great Plains in the USA
(Windels 2000) In that period FHB associated losses were estimated at US $ 27 billion in
the region (Nganje et al 2002)
Importantly Fusaria infested grains are often co-contaminated with multiple
mycotoxins which further increase health risks as interactions between the mycotoxins may
cause greater toxic effects than expected (Schollenberger et al 2006) This implies that an
apparently low level of contamination of a single mycotoxin can have severe effects on
consumers because of synergistic effects among multiple co-occuring mycotoxins (Surai et al
2010)
18
A number of studies demonstrated synergistic toxic effects among Fusarium
mycotoxins on pigs turkeys lamb chicks as well as in in vitro Caco-2 cell and yeast
bioassays (Table 1-5)
Table 1-5 In vitro and in vivo synergistic effects of different food contaminating mycotoxins
DON-deoxynivalenol DAS-diacetoxyscirpenol FB1-fumonisins 1 NIV-nivalenol ZEN-
zearelenone FA-fusaric acid Afl-aflatoxin
Mycotoxins source Interacting
mycotoxins
Experimental
animalcells
Effects Reference
Naturally contaminated
cereals
DON FA Pigs Inhibit growth Smith et al
1997
DON contaminated
wheat and FB1
DON FB1 Pigs Weight loss Harvey et al
1996
Culture materials
contained mycotoxins
T-2 FB1 Turkey poults Weight loss oral
lesion
Kubena et al
1997
Inoculated rice and
pure mycotoxin
DAS Afl Lamb Weight reduction Harvey et al
1995
Naturally DON
contaminated wheat
and pure T-2 toxin
DON T-2 Chicks Reduced body
weight significantly
Kubena et al
1989
Pure mycotoxins DAS Afl Chicks Reduced body
weight
Kubena et al
1993
Pure mycotoxins T-2 Afl Chicks Reduced weight
oral lesion
Kubena et al
1990
Pure mycotoxins DON
ZENFB1
Human intestinal
Caco-2 cells
Increased DNA
fragmentations
Kouadio et al
2007
Pure mycotoxins DON NIV Yeast bioassay Synergistic effects Madhyastha et
al 1994
Pure mycotoxins T-2 HT-2 Yeast bioassay Synergistic effects Madhyastha et
al 1994
19
16 Toxic effects of deoxynivalnol and trichothecenes
As small molecules TCs simply enter into cells and interact with different cell
components resulting in disruption of cellular functions and causing various toxic effects that
include inhibition of protein synthesis and ribosomal fragmentation (Eudes et al 2000
Minervini et al 2004 Pestka 2007) Simple absorption of DON into human intestinal Caco-2
cells and crossing human placenta barriers through passive diffusion has been reported
(Sergent et al 2006 Nielsen et al 2011)
TC may cause various toxicities in animals and plants including apoptosis
immunotoxicity chromatin condensation disruption of cellularmitochondrion membranes
disruption of gut tissues inhibition of protein and ribosome synthesis breakage of ribosome
and cell damage The toxicity of TC is influenced by structural variability (ie position
number and type of functional groups attached to the tricyclic nucleus and C-C double bond at
C9-10 position) (Desjardins 2006) Nevertheless epoxide function at the C12-13 position is
the key for TC cytotoxicity (Swanson et al 1988 Eriksen et al 2004)
Different in vitro and in vivo studies revealed that the method and period of exposure
amount of TC and types of celltissue and host species have significant effects on TCrsquos
cytotoxicity (Scientific Committee for Food 2002 Rocha et al 2005 Nielsen et al 2009)
Administration of 10 mgkg body weight (bw) NIV caused apoptosis in mice thymus Peyerrsquos
patch and spleen tissues (Poapolathep et al 2002) DON and NIV at 887 mgkg bw for 3 days
a week for four weeks induced immunotoxicity in experimental mice determined by
estimating increased immunoglobulins (IgA) and decreased IgM levels (Gouze et al 2007)
Exposure of mice to 1 mgkg bw of DON induced skeletal malformations and reduced mating
tendency while significant resorption of embryos observed at the dose of gt10 mgkg bw
20
(Khera et al 1982) At high acute doses (4972 mgkg bw) DONNIV and type-A
trichothecenes caused mice death because of intestinal haemorrhage necrosis of bone
marrowlymphoid tissues and kidneyheart lesions (Yoshizawa and Morooka 1973
Yoshizawa et al 1994 Forsell et al 1987 Ryu et al 1988) In a separate study 15-ADON
showed more toxicity than DON when the mycotoxins were orally administered to mice
(Forsell et al 1987)
Studies suggested that type-A TCs are more toxic in mammals than type-B which
means that functional variability at the C-8 position of TC has a significant role in toxicity
The toxicity of various TC mycotoxins in animals can be ranked in the order of T-2
toxingtHT-2 toxingtNIVgtDONge3-ADONgt15-ADON according to the results from human cell
lines and mice model systems bioassays (Ueno et al 1983 Pestka et al 1987 Nielsen et al
2009) Moreover animal feeding assays demonstrated that monogastric animals are most
sensitive to DON The order of sensitivity of animals to DON is
pigsgtrodentsgtdogsgtcatsgtpoultrygtruminants (Pestka 2005) The reasons for the variations in
DON toxicity species might be due to differences in metabolism absorption distribution and
elimination of the mycotoxin among animal species (Pestka 2007)
To my knowledge the mechanism of TC uptake by plant cells has not been studied
Nonetheless cereals are exposed to TC by toxigenic Fusaria infection which may cause
various toxicities such as cell damage wilting chlorosis necrosis program cell death and
inhibition of seedling growth in sensitive plants (eg wheat maize and barley) (Cossette and
Miller 1995 Rocha et al 2005 Boddu et al 2007) As animals TC inhibits protein synthesis
in non-host A thaliana (Nishiuchi et al 2006) Immunogold assays showed that the cellular
targets of DON in wheat and maize were cell membrane ribosome endoplasmic reticulum
21
vacuoles and mitochondria Modulation of cell membrane integrity and electrolyte losses
were observed by direct DON interference in maize and wheat cell assays (Cossette and
Miller 1995 Mitterbauer et al 2004) DON-mediated mitochondrion membrane alterations
disruption of host cells enzymatic activity apoptosis and necrosis have been observed in
cereals (Mitterbauer et al 2004 Rocha et al 2005) In contrast to animal type-B is more toxic
to plants than type-A DON showed 8- and 13-fold phytotoxicity to wheat coleoptiles than T-
2 and HT-2 toxins respectively (Eudes et al 2000) By compilation of different studies TC
phytoxicity can be ordered as DONge3-ADONgtT-2 toxingtDAS (Wakulinski 1989 McLean
1996 Eudes et al 2000)
22
17 Cellular and molecular mechanisms of DON toxicity in animals and plants
The underlying cellular and molecular mechanisms of DONTC toxicity are evolving
Exposure to DON proceeds with the up and down regulation of numerous critical genes of
eukaryotic organisms which causes various negative effects on animal and plant organisms
(Figure 1-2)
One of the early events of DON toxicity occurs when the mycotoxin binds to the
active ribosome resulting in blocking protein translation through different routes by
damaging the ribosome (Shifrin and Anderson 1999) degrading or altering conformational
changes of 28S-rRNA (Alisi et al 2008 Li and Pestka 2008) and inducing expression of
miRNAs This results in down regulation of the ribosome associated genes (He et al 2010a
Pestka 2010) Both animal and plant cell assays demonstrated that DON bound to the peptidyl
transferase (Rpl3) domain of the 60S subunit of ribosome and blocked protein chain
elongation and termination at high DON concentrations while peptide chain termination and
ribosome breakage occurred at a low level of DON exposure (Zhou et al 2003a)
23
Figure 1-2 Cellular and molecular mechanisms of deoxynivalenol toxicity in eukaryotic
organisms DON is simply absorbed by the cell and interacts with the cytoplasmic membrane
(CM) mitochondrion (MT) membrane and ribosome Disruption of the CM and alteration of
the MT membrane causes a loss of electrolytes and affects the function of the MT
respectively DON binds to the active ribosomes which leads to ribosome damage and
induces micro RNAs (miRNA) Expression of miRNA impairs the synthesis of ribosome
associated protein The subsequent transduction and phosphorylation of the RNA-activated
protein kinase (PKR) and hematopoietic cell kinase (Hck) activate the mitogen-activated
protein kinase (MAPK) cascade Phosphorylation of the MAPKs induces targeted
transcription factors (TF) which results in ribotoxis stress response upregulation of
proinflammatory cytokines and dysregulation of neuroendocrine signaling (derived from
Pestka 2007 and 2010)
24
The consequence of DON interaction with ribosomes leads to cellular and molecular
changes which are complex and not fully understood However it is well-demonstrated that
the activation of the mitogen-activated protein kinase (MAPK) and subsequent molecular
actions play key roles in the cytotoxicity of TC Upon binding to the ribosome DON rapidly
induced interactions between the protein kinases and the ribosomal 60S subunit as observed in
mononuclear phagocytes (Bae and Pestka 2008) Ribosomal binding to DON activated the
ribosome associated kinases for instances protein kinase R (PKR) hematopoietic cell kinase
(Hck) and P38
mitogen-activated protein kinase The protein kinases are induced by double-
stranded RNA (dsRNA) or possibly by the damaged ribosomal peptides (Iordanov et al 1997
Shifrin and Anderson 1999 Zhou et al 2003b Pestka 2007) Subsequent phosphorylation of
kinases (eg PKRHckP38
) activates the MAPK cascade resulting in ribotoxis stress
response upregulation of proinflammatory cytokines and dysregulation of neuroendocrine
signaling (Pestka and Smolinki 2005 Pestka 2010) Studies suggested that high doses of
DON induces ribotoxis stress response (apoptosis) preceded by immunosuppression and
disruption of intestinal permeabilitytissue functions (Yang et al 2000 Pestka 2008) Further
study observed stabilization of the mRNA and expression of transcription factors for critical
genes when cells were exposed to a low concentration of DON (Chung et al 2003)
The MAPK pathway proceeds with upregulation of a broad range of proinflammatory
cytokine genes and dysregulation of neuroendocrine signaling within the central and enteric
nervous systems Modulation of neuroendocrine signaling by DON increased the level of
serotonin resulted in vomiting reduced food intake and growth retardation in experimental
animalscell lines (Prelusky and Trenholm 1993 Li 2007) Upregulation of proinflammatory
cytokines and chemokines genes was determined in mice liver spleen kidney and lung
25
following in 1 h of exposure to DON A subsequent shock-like effect was observed in mice
because of a ldquocytokine stormrdquo (Pestka 2008) Induction of proinflammatory cytokines also led
to various cellular effects on the central nervous system interference of the innate immune
system and growth hormone signaling These contribute to anorexia growth retardation
aberrant immune response and tissue dysfunction as demonstrated in experimental animal
systems (Pestka 2010)
DON-mediated growth retardation in mice via suppression of growth hormones
(insulin-like growth factor 1 and insulin-like growth factor acid-labile subunit) has been
reported by Amuzie et al (2009) Disruption of gut tissues by DONNIV and subsequent
impairment of intestinal transporter activity was demonstrated in a human epithelial cell
model (Marseca et al 2002) The study proposed that alteration of gut tissue permeability
may cause gastroenteritis induction and growth retardation in livestock animals Growth
retardation of livestock may also occur via dysregulation of neuroendocrine signaling-
mediated induction of emesis and anorexia (Pestka et al 2010)
The molecular mechanism of TC toxicity in plant systems has not been well
documented A recent study showed the upregulation of numerous defense related genes in
wheat upon exposure to DON (Desmond et al 2008) The study further suggested that DON-
induced program cell death in wheat occurred through a hydrogen peroxide signaling
pathway This mechanism had not been observed in mammals
26
18 Strategies to control FHB and trichothecenes
During the past decades many pre- and post-harvest strategies have been applied to
protect cereal crops from Fusarium infestation and to eliminate TC from contaminated
agricultural commodities (FAO 2001 Munkvold 2003 Varga and Toth 2005 Kabak and
Dobson 2009) Some of the above crop protection techniques have been patented (He and
Zhou 2010) In spite of those efforts TC contamination of cereals appears to be unavoidable
using the currently available methods
Pre-harvest control strategies involving cultural practices (eg crop rotation tillage
early planting) and fungicide applications partially reduced Fusarium infection and DON
contamination in cereals (Dill-Macky and Jones 2000 Schaafsma et al 2001) However
repeated application of fungicides may lead to the evolution of Fusarium strains resistant to
fungicides with greater mycotoxin production ability (DrsquoMello et al 1998) At post-harvest
reduction of grain moisture by artificial drying storage at low temperatures (1-4degC) and
proper ventilation of the storage room have shown to be effective in inhibiting fungal growth
and mycotoxin production in infected grain (Midwest Plan Service 1987 Wilcke and Morey
1995) This technology has become less applicable particularly in developing countries
because of the frequent observation required to maintain proper storage conditions (Hell et al
2000)
Removal of Fusarium infected grains by physical methods (eg mechanical
separation density segregation colour sorting etc) was unable to eliminate TC completely
(Abbas et al 1985 He et al 2010b) This is partly because FHB symptomless kernels may
contain a considerable amount of DON which are not eliminated by physical separation
procedures (Seitz and Bechtel 1985) Several studies showed that milling and food processing
27
steps were unable to completely remove DON from grain-based foods (Jackson and
Bullerman 1999 Placinta et al 1999 Kushiro 2008) Detoxification of TC by chemical
treatments (eg ozone ammonia chlorine hydrogen peroxide and sodium metabisulfite) has
been demonstrated (Young et al 1986 Young et al 1987 Young et al 2006) The problem
with these techniques is that the application of such chemicals is not economically viable and
it also has a negative impact on grain quality (Karlovsky 2011) Although the technique has
been popular and showed effectiveness to certain mycotoxins utilization of absorbents to
inhibit mycotoxin absorption by the livestock gastrointestinal tract appeared to be ineffective
for TC (Huwig et al 2001)
The introduction of Fusarium resistant cultivars would be the best strategy to address
the FHB and associated mycotoxin problems in cereals However breeding for cultivars
resistant to FHBGER is difficult and time consuming as natural resistance against Fusarium
is quantitative in nature FHB resistant quantitative trait loci (QTLs) have been identified in
several cereal genotypes (eg Sumai 3 Ning 7840 Bussard) (Bernando et al 2007 Golkari
et al 2009 Haumlberle et al 2009 Yang et al 2010) The 3B 5A and 6B QTLs for type-I
(resistant to initial infection) and type-II (resistant to fungal spread in the kernel) resistances
have been identified on different wheat chromosomes (Buerstmayr et al 2009) Type-I
(Qfhsifa-5A) and type-II (Fhb1Fhb2-3B) QTLs from Sumai 3 are being used in many
breeding programs and pyramiding of the QTLs in spring wheat showed additional effects
Nonetheless the key genes involved in FHB resistance in cereals are yet to be identified This
makes it difficult to integrate the resistance genes into elite cultivars
28
Cultivar development by direct incorporation of TCDON resistance genes may be an
alternative way of protecting grains from these mycotoxins contaminations Many genes from
distantly related species have been integrated into crop cultivars using a transgenic approach
with great economic benefits (Brookes and Barfoot 2006) DON transforming trichothecene
3-O-acetyltransferase (TRI101 from Fusarium sporotrichioides) and UDP-
glucosyltransferase (from Arabidopsis thaliana) genes expressed in cereal cultivars showed
some DONDAS detoxifying ability under in vitro and controlled conditions (Kimura et al
1998 Okubara et al 2002 Poppenberger et al 2003 Ohsato et al 2007) By contrast no
reduction of mycotoxins in either of the above cases was observed under field conditions
(Manoharan et al 2006) Importantly the transformed acetylated and glycosylated
compounds could revert to the original substances or convert to even more toxic products
(Gareis et al 1990 Alexander 2008)
19 Isolation of DONTC de-epoxidating micro-organisms and genes
An enzyme with the capability of de-epoxidizing DONTC appears to be a potential
source for completely removing TC from contaminated cereals (Karlovsky 2011) However
such trichothecene detoxifying genes do not exist in cereals and other plants (Boutigny et al
2008) Because of this several research groups have attempted to obtain microorganisms that
encode DONTC de-epoxidation genes Mixed microbial culture capable of de-epoxydizing
trichothecenes have been isolated from chicken gut (He et al 1992) cow ruminal fluid (King
et al 1984 Swanson et al 1987 Fuchs et al 2002) pig gut (Kollarczik et al 1994) and
digesta of chicken (Young et al 2007) rats (Yoshizawa et al 1983) sheep (Westlake et al
1989) and fish (Guan et al 2009) All of the microbial cultures cited above showed de-
29
epoxydation capabilities only under anaerobic conditions realtively high temperatures and at
lowhigh pH except microbes from fish intestine that showed efficient de-epoxidation at low
temperatures and wide range of temperatures (Guan et al 2009) Such enzymes active under
anaerobic conditions high temperatues and low pH may have potential applications as feed
additives and the eubacterial strain BBSH 797 is an animal feed additive product with the
brand name Mycofixreg by BIOMIN (He et al 2010b) Biotechnological applications of
anaerobically-active enzymes from the above microbial sources seem to be inappropriate
under the aerobic and moderate conditions at which cereals plants are grown
Most of these efforts have failed to isolate single DONTC degrading microbes
effective under aerobic conditions because of inappropriate microbial growth conditions
under laboratory conditions Earlier attempts were made to obtain TC de-epoxidizing aerobic
bacteria but the culture did not retain DONNIV de-epoxidation activity after reculturing
(Ueno 1983 He et al 1992) In addition the co-metabolic nature of the microbial TC de-
epoxidation reaction has limited the use of mycotoxins as the sole nutrient source for selective
isolation of the target microorganisms Several DONTC de-epoxidation microbial strains
including Bacillus strain LS100 and Eubacterial strain BBSH 797 have been isolated from
chicken gut and rumen fluids through multi-step enrichment procedures (Fuchs et al 2002
Yu et al 2010) A DON to 3-keto DON transforming bactial strain was islated from soil by
prolonged incubation with high concentration DON (Shima et al 1997)
Microbial genesenzymes with TC de-epoxidation activity are not known Based on
the microbial TC de-epoxidation activity of a mixed microbial culture it has been proposed
that ceratin reductases or oxidoreductases may catalyze such a biochemical reaction (Islam et
al 2012) With the advent of molecular techniques many novel genes with
30
industrialagricultural value have been cloned by reverse genetics (genomic DNA library
preparation and functional screening of the library clones) forward genetics (transposon
mutagenesis) and comparative genomics methods (Frey et al 1998 Koonin et al 2001
Suenaga et al 2007 Chung et al 2008 Ansorge 2009) The tremendous advancement of next
generation sequencing technology and bioinformatics tools may facilitate the identification of
the microbial genes responsible for DONTC detoxification (Ansorge 2009) Isolation of TC
de-epoxidation genes may play a major role in producing safe foods and feeds derived from
grains
110 The bacterial species Citrobacter freundii
Citrobacter freundii is a bacterial species belonging to the family Enterobacteriace It
is a facultatively anaerobic and Gram-negative bacillus (Wang et al 2000) The bacterium
survives almost everywhere including animalhuman intestines blood water and soils
(Wang et al 2000 Whalen et al 2007) C freundii is known as an opportunistic human
pathogen and causes a variety of respiratory tract urinary tract and blood infections and
responsible for serious neonatal meningistis disease (Badger et al 1999 Whalen et al 2007)
Interestingly this bacterium plays a positive environmental role through nitrogen recycling
(Puchenkova 1996) Strains of C freundii also contribute to the environment by sequenstering
heavy metals and degrading xenobiotic compounds (Kumar et al 1999 Sharma and Fulekar
2009)
31
111 Hypotheses and objectives
The study was based on the following hypotheses
i Certain microorganisms in DON contaminated agricultural soils are able to transform DON
to de-epoxy DON (de-epoxidation) under aerobic conditions and moderate temperatures
ii DON de-epoxidation may be achieved by a single microbial strain
iii Microbial DON detoxifying genes and associated regulators are able to express and
function in a closely related host organism
iv DON detoxifying microbial strains carry genes for de-epoxidation activity that are absent
from related species that have not evolved under DON selection pressure
The objectives of this research were to
i Select microbial cultures from soil having DON de-epoxidation activity under aerobic
conditions and moderate temperatures by analyzing potential DON contaminated soils in
various growth conditions namely aerobicanaerobic conditions different growth media
temperature and pH
ii Select for organisms with DON de-epoxidation activity from the mixed microbial culture
by treating with antibiotics heating subculturing on agar medium and incubating for a
prolonged period in media with a high DON concentration
iii Identify microbial de-epoxidation genes by producing a microbial genomic DNA library
from the DON de-epoxidizing strain followed by screening of the clones for DON de-
epoxidation
32
iv Determine microbial de-epoxidation genes by sequencing of the genome and comparative
analysis of the genome with closely related species and identification of putative DON
detoxification genes
33
CHAPTER 2
20 AEROBIC AND ANAEROBIC DE-EPOXIDATION OF MYCOTOXIN
DEOXYNIVALENOL BY BACTERIA ORIGINATING FROM AGRICULTURAL
SOIL
Published in World Journal of Microbiology and Biotechnology 2012 287-13
21 Abstract
Soil samples collected from different cereal crop fields in southern Ontario Canada
were screened to obtain microorganisms capable of transforming deoxynivalenol (DON) to
de-epoxy DON (dE-DON) Microbial DON to dE-DON transformation (ie de-epoxidation)
was monitored by using liquid chromatography-ultraviolet-mass spectrometry (LC-UV-MS)
The effects of growth substrates temperature pH incubation time and aerobic versus
anaerobic conditions on the ability of the microbes to de-epoxidize DON were evaluated A
soil microbial culture showed 100 DON to dE-DON biotransformation in mineral salts
broth (MSB) after 144 h of incubation Treatments of the culture with selective antibiotics
followed an elevated temperature (50ordmC) for 15 h and subculturing on agar medium
considerably reduced the microbial diversity Partial 16S-rRNA gene sequence analysis of the
bacteria in the enriched culture indicated the presence of at least Serratia Citrobacter
Clostridium Enterococcus Stenotrophomonas and Streptomyces The enriched culture
completely de-epoxidized DON after 60 h of incubation Bacterial de-epoxidation of DON
occurred at pH 60 to 75 and a wide array of temperatures (12 to 40ordmC) The culture showed
34
rapid de-epoxidation activity under aerobic conditions compared to anaerobic conditions This
is the first report on microbial DON to dE-DON transformation under aerobic conditions and
moderate temperatures The culture could be used to detoxify DON contaminated feed and
might be a potential source for gene(s) for DON de-epoxidation
Keywords biotransformation de-epoxidation deoxynivalenol soil bacteria vomitoxin
22 Introduction
Deoxynivalenol (DON commonly known as lsquovomitoxinrsquo) 3α 7α 15-trihydroxy-
1213-epoxytrichothec-9-en-8-one is a trichothecene produced by toxigenic Fusarium species
causing Fusarium head blight (FHB) or Gibberella ear rot (GER) disease on small grain
cereals and maize respectively (Starkey et al 2007 Foroud and Eudes 2009) DON
contamination in cereals and maize is widespread and can lead to tremendous losses to the
growers It has also detrimental effects on human and livestock health (Pestka and Smolinski
2005 Rocha et al 2005) The toxin interferes with the eukaryotic ribosomal peptidyl
transferase (Rpl3) activity resulting in human and animal mycotoxicosis (Ueno 1985 Bae
and Pestka 2008) Common effects of DON toxicity in animals include diarrhea vomiting
feed refusal growth retardation immunosuppression reduced ovarian function and
reproductive disorders (Pestka and Smolinski 2005 Pestka 2007) In plants DON causes a
variety of effects viz inhibition of seed germination seedling growth green plant
regeneration root and coleoptile growth (cited in Rocha et al 2005) In addition DON
enhances disease severity in sensitive host plants (Maier et al 2006)
35
Avoidance of DON contamination or reduction of toxin levels in contaminated grains
remains a challenging task Physical and chemical processes are either inappropriate or
prohibitively expensive to use to decrease the DON content of agricultural commodities
(Kushiro 2008 He et al 2010b) The use of Fusarium resistant crop varieties might be the
best method to reduce DON contamination However breeding for FHB or GER resistant
cultivars appears to be very difficult and time consuming as natural sources of resistance
against Fusarium-caused diseases is quantitative in nature (Ali et al 2005 Buerstmayr et al
2009) As an alternative expressing genes for enzymatic detoxification of DON in elite
cultivars may be an effective strategy for protecting the value of feeds and foods (Karlovsky
1999) Heterologous expression of eukaryotic trichothecene 3-O-acetyltransferase and UDP-
glucosyltransferase genes in cereals and Arabidopsis have resulted in reduced toxicity via
acetylation and glycosylation of the DON (Poppenberger et al 2003 Ohsato et al 2007)
It has been proposed that enzymatic de-epoxidation of trichothecene toxins could be
used to limit DON contamination (Zhou et al 2008 Foroud and Eudes 2009) DON belongs
to type B-trichothecenes which contain an epoxide ring at C12-13 and the epoxide is highly
reactive and mainly responsible for toxicity (Figure 2-1) (Swanson et al 1988 Eriksen et al
2004)
36
Figure 2-1 Mechanism of microbial DON de-epoxidation Removal of the oxygen atom from
the epoxide ring results in formation of a C=C double bond at C-12 position which reduces
cytotoxicity (Swanson et al 1988 Eriksen et al 2004)
Microbial DON de-epoxidation by single bacterial strains and by mixed cultures has
been reported (King et al 1984 Swanson et al 1987 He et al 1992 Kollarczik et al 1994
Fuchs et al 2002 Young et al 2007 Guan et al 2009 Yu et al 2010) However all of these
microbes originated from fish and other animals and were strictly anaerobic The recovered
microbial cultures and single strains only showed de-epoxidation of trichothecenes at low
(le15ordmC) and high (gt30degC) temperatures Such enzymes might have limited applications for
toxin decontamination in cereal grains The aim of the current study was to isolate the
microbes from soil capable of transforming DON to a de-epoxy metabolite under aerobic
conditions and moderate temperatures
De-epoxydation
Deoxynivalenol (DON)
De-epoxyDeoxynivalenol (dE-DON)
37
23 Materials and methods
231 Soil samples
The upper layer (0-30 cm) of soils along with crop debris were randomly collected
from cereal (eg maize wheat barley) crop fields during 2004-2006 in Southern Ontario
Canada Detailed of the crops and locations of the fields have been described in Jianwei Hersquos
Master Degree thesis (He 2007) Briefly soil samples were collected in sterile plastic bags
and transported to the laboratory Samples were sieved to remove large roots stones and
stored at 4ordmC until used The samples were mixed with grounded mouldy corn (containing 95
microgg DON) + F graminearum and incubated at 22-24degC for six weeks Five samples were
prepared for DON de-epoxidation activity analysis by dispersing 5 g soil in 50 mL distilled
water and storing at4degC until further use
232 Culture media
Media used in the study were nutrient broth (NB) Luria Bertani (LB) minimal
medium (MM) (He 2007) six mineral salts + 05 bacto peptone (MSB) (He 2007) tryptic
soy broth (TSB) corn meal broth (CMB) (Guan et al 2009) potato dextrose broth (PDB) six
mineral salts + 05 yeast extract broth (MSY) and soil extract (SE) MM was composed of
sucrose-100 g K2HPO4-25 g KH2PO4-25 g (NH4)2HPO4-10 g MgSO47H2O-02 g
FeSO4-001 g MnSO47H2O- 0007 g water- 985 mL and MSB was composed of the same
salts as MM but was supplemented with Bactotrade Peptone (05 wv) as the sole C source
instead of sucrose To prepare soil extract 100 g of the composite soil was homogenized in 1
L water The soil particles and plant debris were removed by passing the soil suspension
through Whatmanreg grade 3 filter paper (Sigma-Aldrich Oakville Ontario Canada) The
38
extract was autoclaved at 121degC for 30 min Media and ingredients were purchased either
from Fisher Scientific Inc (Fair Lawn New Jersey USA) Sigma-Aldrich (Oakvile Ontario
Canada) or Difco (Sparks Maryland USA) Prior to autoclaving pH of the broth media was
adjusted to 70 Solid media were prepared by adding 15 agar
233 Screening of soil samples
Frozen soil suspensions were thawed and vortexed to homogenize the microbial
population An aliquot (100 microL) of the suspension was immediately transferred to MSB PDB
and MM broth media DON at 50 microgmL was added to the microbial suspension cultures The
cultures were incubated at 22-24degC with continuous shaking at 180 revolutions per minute
(rpm) The extent of DON de-epoxidation was determined at 24 h 48 h 72 h 96 h 120 h and
144 h of incubation
234 Determination of de-epoxidation product
Microbial DON de-epoxidation was measured according to Guan et al (2009)
Samples were prepared by mixing equal volume of microbial culture and methanol and
allowed to stand for 30 min at room temperature Microorganisms in the mixture were
removed by passing them through a 045 microm Millipore filter (Whatmanreg Florham Park New
Jersey USA) The filtrates were used for de-epoxidation analysis by a liquid chromatography-
ultraviolet-mass spectrometry (LC-UV-MS) Identification and quantification of DON and its
transformation product was achieved using a Finnigan LCQ DECA ion trap LC-MS
instrument (ThermoFinnigan San Jose CA USA) The equipment detects 30-2000 mz
(molecular mass ion ratio) compounds and able to determine 001 micromL DON The HPLC
39
system was equipped with a Finnigan Spectra System UV6000LP ultraviolet (UV) detector
and an Agilent Zorbax Eclipse XDB C18 column (46times150 mm 35 μm) The binary mobile
phase consisted of solvent A (methanol) and solvent B (water) The system was run with a
gradient program 25 A at 0 min 25ndash41 A over 5 min isocratically at 41 A for 2 min
and 41ndash25 A over 1 min There was a 3 min post-run at its starting condition to allow for
the equilibration of the column The flow-rate was set at 10 mLmin UV absorbance was
measured at 218 nm Atmospheric pressure chemical ionization (APCI) positive ion mode
was used for MS detection A DON standard was used to adjust the instrument to its
maximum response The system was operated as follows shear gas and auxiliary flow-rates
were set at 95 and 43 (arbitrary units) voltages on the capillary tube lens offset multipole 1
offset multipole 2 offset and entrance lens were set at 3000 minus1700 minus370 minus850 minus3500
and minus4200 V respectively capillary and vaporizer temperatures were set at 200 and 450degC
respectively and the discharge needle current was set at 7 μA Identification of mycotoxin
was achieved by comparing their retention times UV spectra and mass spectrum with DON
and de-epoxy DON (Biopure Union MO USA) as standards Quantification of DON and
product was based on peak areas related to their UV and MS calibration curves
40
235 Biological DON de-epoxidation
To help identify the types of microorganisms causing the DON de-epoxidation MSB
medium was supplemented with either 100 microgmL penicillin G or streptomycin sulfate for
suppressing gram positive or negative bacterial growth and activities respectively Three
replications of antibiotic-treated media were inoculated with 100 microL of microbial culture
capable of de-epoxidizing DON The antibiotic-treated cultures were incubated at 27degC
spiked with 50 microgmL DON Controls were prepared without adding antibiotics in the
microbial culture Inhibition of DON de-epoxidation was analyzed after five day of
incubation
236 Reduction of microbial diversity by antibiotic treatments heating and
subculturing on agar medium
In order to reduce non-DON degrading microbial populations the DON de-
epoxidation capable cultures were sequentially treated with antibiotics having diverse modes
of action including i) dichloramphenicol that kills various gram positive Gram-negative and
anaerobic bacteria ii) trimethoprim that kills Gram-positive and negative bacteria iii)
penicillin that is lethal to Gram-positive bacteria and iv) cyclohexamide to kill filamentous
fungi and yeasts Antibiotics were purchased from Sigma-Aldrich Antibiotic stock solutions
(10 mgmL) were prepared with sterile water or methanol Microbial cultures in MSB
medium were treated with 100 microgmL of each antibiotic followed by incubation of the culture
at 50ordmC for 15 hours to kill undesirded microbial species After heating cultures were treated
with 50 microgmL DON and incubated at 27ordmC for five days DON de-epoxidation activity of the
microbial cultures was determined following the procedure described above Cultures that
41
showed gt99 de-epoxidation were considered for subsequent antibiotic and heat treatments
Aliquots of the achieved culture were spread on MSB agar plates and grown at room
temperature for few days Colonies appeared on the plates were collected and incubated in
MSB medium supplemented with 50 microgmL DON De-epoxidation ability of the subcultured
microbial cultures was determined after five days of incubation at 27degC by LC-UV-MS
method
237 Analysis of microbial population diversity
Terminal Restriction Fragment Length Polymorphism (T-RFLP) analysis
Initially microbial diversity was determined by terminal restriction fragment length
polymorphism (T-RFLP) fingerprint analysis (Bruce and Hughes 2000) Genomic DNA from
microbial cultures with and without enrichment was extracted using DNeasy Plant DNA
Extraction Mini Kit (Qiagen Inc Mississauga Ontario Canada) 16S-rDNA was amplified
by PCR using the HEX-labeled forward primer Bac8f
(5-agagtttgatcctggctcag-3) and
unlabeled reverse primer Univ1492r (5-ggttaccttgttacgactt-3) (Fierer and Jackson 2006)
Fifty microlitter PCR mixture for each sample was prepared having 1x HotStarTaq Master
Mix (Qiagen) 05 microM of each primer 50 microg of BSA and 50 ng of DNA The 35 PCR cycles
followed 60 s at 94degC 30 s at 50degC and 60 s at 72degC After size verification by agarose-gel
electrophoresis 50 ng of purified PCR products was digested for 3 h in a 20 microL reaction
mixture with 5 U of HhaI restriction enzyme (New England Biolabs Ipswich MA USA)
The size and relative abundance of the labeled T-RFLP fragments were separated by capillary
electrophoresis in an automated DNA sequencer equipped with fluorescence detector
(Applied Biosystem 3730 DNA Analyzer) The DNA fragments were analyzed with the Peak
42
Scannertrade Software v10 to create an electropherogram with the genetic fingerprints of the
microbial community
Analysis of V3-V5 region of the 16S-rDNA
To define the bacterial genera present in the enrichment culture the 16S-rDNA was
amplified using two sets of bacterial universal primers Primarily 16S-rDNA was PCR
amplified using the primers 46f-5rsquogcytaacacatgcaagtcga3rsquo (Kaplan and Kitts 2004) and
1371r-5rsquogtgtgtacaagncccgggaa3rsquo (Zhao et al 2007) PCR was performed in a total volume of
25 μL which contained 06 μM of each primer 1x of HotStarTaqreg Master Mix (Qiagen Inc)
and 20-40 ng of DNA The PCR cycles included the following steps 95ordmC (15 min) 35 cycles
of 94ordmC (20 s) 52ordmC (15 s) 72ordmC (1 min 30 s) and a 72ordmC (7 min) extension step The second
pair of universal bacterial primers were 27f-5rsquoagagtttgatcmtggctcag3rsquoand 1492r-
5rsquoggttaccttgttacgactt3rsquo (Lane 1991) PCR was conducted with 25 cycles following the
procedures of Tian et al (2009) Amplification was carried out in a GeneAmpreg PCR System
9700 (Applied Biosystems) Furthermore the PCR products were used as templates to
amplify the hypervariable V3-V5 region of the 16S-rDNA using primers 357f-
5rsquocctacgggaggcagcag3rsquo and 907r7 5rsquoccgtcaattcctttgagttt3rsquo (Yu and Morrison 2004)
After purification of the PCR products by QIAquick PCR purification kit two
amplicon libraries were prepared by ligating to pCR-TOPO vector (Invitrogen) and the library
clones were transformed into TOPO10 competent cells according to Invitrogenrsquos TOPO
cloning manual The clone libraries were plated on LB plates containing 25 μgmL
kanamycin Fifty colonies from each library were randomly picked up from the overnight
plates and inserts were sequenced as described by Geng et al (2006) PCR products were
43
sequenced with an automated ABI 3730 DNA analyzer (Applied Biosystems) The sequence
similarities of the amplified V3-V5 region were compared with the ribosomal database project
(RDP) (httprdpcmemsuedu) Sequences with greater than 95 identity were assigned to
the same bacterial genus
238 Factors influencing DON de-epoxidation
DON de-epoxidation ability of the enriched microbial culture was investigated under
different conditions as described below
Maintaince of active culture
To retain microbial de-epoxidation activity the active culture has been maintained in
MSB medium containing 50 microgmL DON
Media growth temperature and pH
Overnight microbial cultures (100 μL) were added to 2 mL of NB LB MM TSB
MSB CMB MSY PDB or SE broths The pH of the media was adjusted to 70 and the
experiments were conducted at 27degC temperature The effect of temperature was studied by
incubating the culture in MSB medium (pH 7) at 12 20 25 30 and 40ordmC To determine the
effects of pH MSB medium pH was adjusted to 55 60 65 70 75 and 80 Media with
different pHs were inoculated with aliquots of the overnight active culture and then incubated
at 27ordmC The cultures were supplemented with 50 μgmL DON before incubation Three
replications for each medium temperature and pH condition were evaluated
44
De-epoxidation under aerobic and anaerobic conditions
Three replicate microbial cultures of five mL each were grown in sterile 15 mL
centrifuge tubes under aerobic conditions and in an anaerobic chamber filled with 95 CO2
and 5 H2 (COY Laboratory Products Inc Ann Arbor MI USA) Three days after initiation
half (25 mL) of the cultures from anaerobic conditions was transferred to new centrifuge
tubes and brought to aerobic conditions Similarly half of the aerobically grown cultures were
placed into an anaerobic chamber Cultures moved from anaerobic to aerobic conditions were
shaken after opening the lids of the tubes in a laminar flow every eight hours intervals to
maintain aerobic conditions in the culture Lids of the culture tubes transferred to anaerobic
conditions were kept loose to allow oxygen to escape from the culture Two days after
transferring to the new conditions the cultures in both conditions were treated with 50 microgmL
DON and incubated at 27degC Every 12 h 500 microL were removed from each tube and analyzed
for microbial DON de-epoxidation activity by LC-UV-MS
239 Data analysis
Data were processed using Microsoft Excel Program 2007 Statistical analysis was
done using General Linear Model procedures in SAS version 91 (SAS Institute Inc Cary
NC USA 2002-03) The results were the average of three replications stated as mean plusmn
standard error Differences between means were considered significant at plt 005 according
to Tukeyrsquos-HSD test
45
24 Results
241 Analyses of DON to dE-DON biotransformation
A typical LC-UV-MS analysis where there is incomplete de-epoxidation of DON by a
particular soil suspension culture is shown in Figure 2-2 Analysis by LC with UV detection
and integration of peak areas showed 662 conversion of DON to dE-DON (Figure 2-2a)
Analysis LC with MS detection (expressed as total ion counts) indicated 643 conversion
(Figure 2-2b) LC analyses with UV and TIC (total ion count) detection are subject to
interferences due to co-eluting contaminants and therefore integration of small peaks was
very difficult making quantification inaccurate MS detection by SIM (selected ion
monitoring) can eliminate the influence of co-eluting contaminants For example the SIM LC
chromatogram for DON (Figure 2-2c) was generated by using the major ions resulting from
the APCI positive ion MS of DON (Figure 2-2e)
Similarly the SIM LC chromatogram for dE-DON (Figure 2-2d) was derived from the
MS ions of dE-DON (Figure 2-2f) The signal to noise ratios (SN) of peaks observed by SIM
were at least 20 times greater than those detected by TIC because there is reduced likelihood
of co-eluting peaks having the same ions Integration of the SIM peaks revealed a 616
conversion The three methods gave similar results but the SIM LC results were considered
to be the most accurate Using that method the conversion of DON to dE-DON by the mixed
culture occurred most quickly at 48 h and was completed by 144 h (Figure 2-3)
46
47
Figure 2-2 LC-UV-MS chromatographic analysis of DON and its de-epoxidated product dE-
DON obtained by culturing with selected soil microbes (a) LC-UV chromatogram of product
mixture monitored at 218 nm (b) LC-MS total ion chromatogram (TIC) of product mixture
(c) selected ion monitoring (SIM) chromatogram of cultured DON at mz 231 249 267 279
and 297 (d) SIM chromatogram of cultured dE-DON at mz 215 233 245 251 263 and 281
(e) atmospheric pressure chemical ionization (APCI) positive ion mass spectrum of DON (f)
APCI positive ion mass spectrum of dE-DON
48
242 Screening of soil samples
Initially samples originating from soils were analyzed in three types of culture media
to support the growth of fungi yeast and bacteria However cultures growing in PDB which
is a suitable substrate for many fungi and yeasts did not permit transform DON to dE-DON
(data not shown by microorganisms) A define medium (ie MM) was also used in the
sample screening assay None of the soil samples showed de-epoxidation activity in MM
(data not shown) From five soil samples one soil sample culture showed de-epoxidation of
DON in MSB The culture in MSB converted gt99 DON to dE-DON within 144 h (Figure 2-
3) Conversion began after a lag of 48 h and was linear thereafter with a rate of 1 per hour
49
Figure 2-3 Time course of relative biotransformation of DON to dE-DON by the initial soil
suspension culture in MSB medium The culture contained 50 μgmL DON at time 0 Culture
with three replications was incubated at room temperature with continuous shaking at 180
rpm The conversion of DON to dE-DON was analysed by LC-UV-MS at 24 h intervals Blue
and red lines are the percent conversion of DON and recovery of dE-DON respectively
50
243 Evidence of biological DON de-epoxidation
Treatment of active microbial culture with 100 microgmL of streptomycin completely
inhibited DON de-epoxidation In contrast the same concentration of penicillin G did not
affect activity of the culture (Figure 2-4) Streptomycin is effective against Gram-negative
bacteria while penicillin G is lethal to Gram-positive bacteria These experiments provided
evidence that DON de-epoxidation may be caused by certain Gram-negative bacteria
51
Figure 2-4 Microbial de-epoxidation of DON after five days of incubation The active DON
de-epoxidation culture was treated with either 100 microgmL penicillin G or streptomycin
(Strepto) in MSB Cultures having three replications were added 50 microgmL DON and
incubated at room temperatures with continuous shaking at 180 rpm For controls cultures
were without antibiotics DON to dE-DON transformation was monitored after five day of
incubation by LC-UV-MS method
0
20
40
60
80
100
120
Penicillin Strepto Control
DO
N d
e-e
po
xyd
ati
on
(
)
Antibiotics
52
244 Reduction of the microbial diversity
Plating the suspension cultures onto MSB agar revealed colonies with a variety of
sizes morphologies and colours reflecting a mixed culture (data not shown) To reduce the
complexity of the culture three steps of selection pressures were applied namely treatment
with antibiotics heat shocks and subculturing on solid medium The above indicated
treatments did not significantly affect DON de-epoxidation of the microbial cultures (data not
shown) T-RFLP profiles of the mixed culture before treatments showed six distinct peaks of
about 110 150 200 220 340 and 420 bp restriction fragments with the 110 bp peak being
the predominant one (Figure 2-5a)
After treatment with antibiotics and heat only three peaks at 110 150 and 325 bp
remained (Figure 2-5b) suggesting that there was a reduction in the microbial diversity
without affecting DON de-epoxidation After enrichments the bacteria species in the culture
changed their growth patterns as the electropherogram produced different sizes of pick
lengths compared to untreated culture
53
(b)
(a)
Rela
tive flu
ore
scen
ce
32000
0
0 500
16S-rDNA fragment size (bp)
100 200 300 400
Rela
tive flu
ore
scen
ce
32000
0
500
16S-rDNA fragment size (bp)
0 300200100 400
54
Figure 2-5 Analysis of microbial population reduction by terminal restriction fragment
length polymorphism (T-RFLP) (a) Restriction fragments of the 16S-rDNA of the untreated
culture and (b) fragmentation patterns of the enriched culture Picks along the X-axis showed
the fragmentation patterns of the 16S-rDNA which is related to the bacterial species diversity
The length of picks (along the Y-axis) showed abundance of terminal restriction fragments
related to the abundance of particular bacterial speciesgenera
55
245 Microbial population structures in the enriched culture
Microbial diversity in the enriched culture was determined by analyzing the sequences
of the V3-V5 region (~586 bp) amplified from the 16S-rDNAs of the microorganisms it
contained The sequence analysis showed the presence of Serratia Citrobacter Clostridium
Enterococcus Stenotrophomonas and Streptomyces in the enriched culture (Figure 2-6)
Out of 100 V3-V5 sequences 54 sequences were found to be belonging to Serratia
the predominant species of the culture The remaining sequences matched with 12
Citrobacter 8 Clostridium 14 Enterococcus 8 Stenotrophomonas and 4 Streptomyces
species
56
Figure 2-6 Microbial genera identified in the enriched DON de-epoxidizing microbial
culture The bacterial genera were determined by comparing the hypervariable regions (V3-
V5) of the 16S-rDNA using ribosomal database project (httprdpcmemsuedu) X-axis
showed the identified bacterial genera and Y-axis demonstrated the abundance of the V3-V5
sequences belonging to different bacterial genera
54
12 8
14
8 4
0
20
40
60 S
eq
uen
ce
fre
qu
en
cy
Microbial genera
57
246 Effects of media growth temperature pH and oxygen requirements on DON de-
epoxidation
Of the nine media that were tested DON de-epoxidation by the enriched microbial
culture only occurred in NB and MSB (Figure 2-7a) It appeared that C andor N
concentrations in the media were crucial for the enzymatic activity because less or more than
05 Bacto Peptone in MSB reduced the microbial DON transformation ability
DON de-epoxidation activity occurred over a wide range of temperatures from 12 to
40ordmC (Figure 2-7b) The fastest rates of DON de-epoxidation occurred between 25 to 40ordmC
De-epoxidation was observed within a relatively narrow range of pH (60 to 75) with
the most rapid DON biotransformation activity observed at pH 70 (Figure 2-7c) At pH 70
the enriched culture showed DON de-epoxidation activity 125 times faster than at pH 60 or
75 At pH 55 and 80 there was no observable DON de-epoxidation
58
0
20
40
60
80
100
120
12degC
20degC
25degC
30degC
40degC
DO
N d
e-e
po
xyd
ati
on
(
)
Temperature
0
20
40
60
80
100
120
DO
N d
e-e
pxyd
ati
on
(
)
Media
(a)
(b)
59
Figure 2-7 Effects of culture conditions on DON to de-epoxy DON transformation by the
enriched microbial culture For the media experiment (a) all the media were adjusted to pH 7
and the incubation temperature was 27degC For the temperature experiment (b) MSB pH 7 was
used For the pH experiment (c) MSB at 27degC was used Microbial cultures were inoculated
with 50 μgmL DON De-epoxidation was determined after 72 h of incubation Media MSY
(mineral salts + 05 yeast extract) LB (Luria Bertani) TSB (tryptic soy broth) MSB
(mineral salts + 05 bacto peptone) SE (soil extract) CMB (corn meal broth) PDB (potato
dextrose broth) and MM (minimal medium)
0
20
40
60
80
100
120
55
60
65
70
75
80
DO
N d
e-e
po
xyd
ati
on
(
)
pH
(c)
60
The enriched culture showed DON de-epoxidation activity under both aerobic and
anaerobic conditions (Figure 2-8) The fastest transformation (100 de-epoxidation after 60 h
of incubation) occurred in cultures (A) that were grown under aerobic conditions throughout
the culture growth period A medium rate of de-epoxidation (100 de-epoxidation after 72 h)
was observed in cultures that were initially grown in either aerobic or anaerobic conditions
followed by transferring to anaerobic or aerobic conditions (A+AN and AN+A) respectively
The lowest rate occurred (100 de-epoxidation after 96 h) in cultures grown in continual
anaerobic conditions (AN) The means comparison of microbial DON de-epoxidation
efficiencies at 60 h of incubation showed that cultures continually grown in conditions A and
AN were statistically different Biotransformation abilities of the cultures that were switched
from one conditions to another (A+AN and AN+A) were statistically similar among each
other but different from cultures that were only grown in conditions A or AN (plt005)
61
Figure 2-8 Microbial DON to dE (de-epoxy DON)-DON transformation activity under
aerobic and anaerobic conditions A = culture incubated and examined the activity only under
aerobic conditions A+AN = culture initially was grown in aerobic conditions and then
transferred to anaerobic conditions AN = cultures grown throughout the assay under
anaerobic conditions AN+A = culture initially grown under anaerobic conditions and then
transferred to aerobic conditions Differences of lower case letters are significantly different
in de-epoxidation capability (after 60 h) among incubation conditions according to Tukeyrsquos
HSD test (p lt 005) The experiment was conducted with three replications
62
25 Discussion
The current study describes a novel bacterial culture isolated from agricultural soil that
was capable of transforming DON to de-epoxy DON under aerobic and anaerobic conditions
at moderate temperatures Because it has been speculated that DON translocates to soil from
contaminated crops and DON that enters into the soil could be metabolized by microbes
(Voumllkl et al 2004) soil samples for the current study were mainly collected from fields where
cereal crops contaminated with DON were grown
In our initial sample screening steps upwards of 144 h of incubation were required to
achieve gt99 DON de-epoxidation Such a long period may have been due to the presence of
fast growing microorganisms that had suppressive effects on the growth or activity of the
DON transforming bacteria (Davis et al 2005) The sequential use of different antibiotics
with different modes of actions followed by thermal treatments and subculturing on solid
medium resulted in a reduction in the number of bacterial species as determined by T-RFLP
analysis Analysis of V3-V5 region of the 16S-rDNA showed that five bacterial genera were
present in the enriched culture The culture showed higher DON de-epoxidation activity (only
60 h required for gt99 biotransformation) compared to initial soil suspension culture that
required 144 h to complete the de-epoxidation This microbial culture could be used to isolate
the DON de-epoxidizing gene(s) applying metagenomics approaches (Daniel 2005)
The culture showed complete de-epoxidation activity both in MSB and NB media
However we have found that the use of nutritionally rich NB medium enhanced non-target
bacterial (eg Serratia lequifaciences) growth but this did not occur in MSB which is a
comparably poor medium To avoid the possible suppressive and detrimental effects of the
fast growing bacteria on the target bacteria MSB was used for enrichment and
63
characterization of the culture An important characteristic of the DON de-epoxidation
activity observed for the microbial culture in the current study is its activity at moderate
temperature (25-30degC) In contrast previously isolated cultures showed optimal activities at
low (le15degC) or high temperatures (37degC) (Guan et al 2009) Also the culture showed DON
de-epoxidation activity at neutral pH range (pH 60-75) in contrast to previous reports which
found that microbial trichothecene de-epoxidation occurred at acidic and basic pHs (Swanson
et al 1987 He et al 1992 Kollarczik et al 1994 Young et al 2007) However a microbial
culture from fish gut showed de-epoxidation under wide range of pHs (Guan et al 2009)
Probably the pH range reflects the environment in which the bacteria naturally grow which in
this case is similar to the soil pH of a typical crop field in Southern Ontario of Canada The
advantage of the present finding is that DON de-epoxidation activity of the culture was
optimal in a temperature range that matches the temperature range of activity for most arable
plants Therefore it is reasonable to suggest that the gene(s)enzyme(s) responsible for the
activity have potential application for enzymatic detoxification of DON in cereals (Karlovsky
1999)
The current observation that bacterial DON de-epoxidation activity occurred under
both aerobic and anaerobic conditions was interesting because DON de-epoxidation is a
reductive chemical reaction that might be expected to be promoted by anaerobic conditions
However there are previous reports of aerobic de-epoxidation of eldrindieldrin by soil
bacteria (Matsumoto et al 2008) and Goodstadt and Ponting (2004) described aerobic
reductive de-epoxidation by vitamin K epoxide reductasephylloquinone reductase A
definitive determination of the oxygen sensitivity of the enzyme responsible for DON de-
epoxidation will ultimately come from experiments with the purified protein
64
In conclusion this study reports on a novel bacterial source from soil that efficiently
de-epoxidizes DON at moderate temperatures under both aerobic and anaerobic conditions
This research may lead to the isolation of microbes and gene(s) encoding enzyme(s) capable
of de-epoxidizing DON that could create transgenic plants with reduced DON contamination
thus improving food and feed safety and reducing negative effects of the toxin on public
health
26 Acknowledgements
The authors gratefully acknowledged the Natural Sciences and Engineering Research
Council of Canada (NSERC) for awarding a scholarship to Rafiqul Islam for PhD program
The research was supported by the Agriculture and Agri-Food Canada Binational
Agricultural Research and Development Fund (AAFCBARD) project We are thankful for
the contributions of Jianwei He Xiu-Zhen Li and Honghui Zhu
65
CHAPTER 3
30 ISOLATION AND CHARACTERIZATION OF A NOVEL SOIL BACTERIUM
CAPABLE OF DETOXIFYING ELEVEN TRICHOTHECENE MYCOTOXINS
31 Abstract
Deoxynivalenol (DON vomitoxin) is one of the most commonly detected food and
feed contaminating mycotoxins worldwide In order to develop a control method effective
under aerobic conditions and moderate temperatures a bacterial strain ADS47 was isolated
capable of transforming DON to the less toxic de-epoxy DON The bacterial strain is
facultatively anaerobic Gram-negative rod shaped with an average cell size of 150 X 072
microm and has peritrichous (multiple) flagella The 16S-rRNA gene sequence had gt99
nucleotide identity to Citrobacter freundii a bacterium that belongs to the gamma subclass of
proteobacteria Strain ADS47 showed near 100 de-epoxidation of DON after 42 h of
incubation in nutrient broth (NB) under aerobic conditions moderate temperature (28˚C) and
neutral pH (70) However de-epoxidation could occur at 10-45˚C and pH 60-75 Under
anaerobic conditions near 100 de-epoxidation was observed after 72 h mainly during the
stationary growth phase Microbial DON de-epoxidation ability increased with increasing
DON concentrations in the culture media Inhibition of bacterial de-epoxidation by sodium
azide suggested that an active electron transport chain was required In addition to DON
strain ADS47 was able to de-epoxidize andor de-acetylate five type-A and five type-B
trichothecene mycotoxins This is the first report of a bacterial isolate capable of de-
epoxidizing DON and trichothecene mycotoxins under aerobic conditions and moderate
66
temperatures Strain ADS47 could potentially be used in bioremediation to eliminate DON
and other trichothecene mycotoxins from contaminated food and feed
Keywords aerobic de-epoxidation Citrobacter freundii isolation soil bacterium
trichothecenes
32 Introduction
Cereals are often contaminated with a variety of type-A and type-B trichothecene
mycotoxins produced by a number of toxigenic Fusarium species (Placinta et al 1999
Binder et al 2007 Foroud and Eudes 2009) Trichothecenes are tricyclic sesquiterpenes with
a double bond at the C9 10 position and a C12 13 epoxy group (Desjardins 2006) The two
types are distinguished by the presence of a ketone at position C8 in type-B which is absent
in type-A (Yazar and Omurtag 2008) The type-B toxin deoxynivalenol (DON) is the most
frequently found mycotoxin in food and feed around the world (Larsen et al 2004 Desjardins
2006 Starkey et al 2007) Different Fusarium species associated with Fusarium head blight
(FHB) or Gibberella ear rot (GER) disease of small grain cereals and maize are mainly
responsible for contaminating grains with DON and other type-A and type-B trichothecenes
These can cause serious mycotoxicosis in humans animals and plants (Ueno 1985 DrsquoMello
et al 1999 Bae and Pestka 2008) Common effects of DONTC toxicity in animals include
diarrhea vomiting feed refusal growth retardation immunosuppression reduced ovarian
functionsreproductive disorders and even death (Pestka and Smolinski 2005 Pestka 2007)
DON enhances FHB disease severity in wheat and maize (Proctor et al 1995 Desjardins
1996 Maier et al 2006)
67
Reduction of mycotoxin levels in contaminated agricultural products remains a
challenging task Current physical and chemical processes to decrease DON content are either
ineffective or prohibitively expensive (Kushiro 2008 He et al 2010b) However the use of
detoxifying microbes to biodegrade mycotoxins is an alternative (Awad et al 2010
Karlovsky 2011) The toxicity of DON is mainly due to the epoxide and there is a significant
reduction of DON toxicity (54) when it is reduced to produce de-epoxy DON Related
mycotoxins also show reductions in toxicity when they are de-epoxidized such as NIV (55)
and T-2 toxin (400) (Swanson et al 1988 Eriksen et al 2004) Bacillus arbutinivorans
strain LS100 and Eubacterial strain BBSH 797 originating from chicken intestine and rumen
fluid respectively are capable of cleaving the epoxide ring of DON under anaerobic
conditions and high temperature (37degC) (Fuchs et al 2002 Yu et al 2010) Strain BBSH 797
which can de-epoxidize DON scirpentriol and T-2 triol trichothecene mycotoxins has been
used as an additive for poultry and swine diets (Karlovsky 2011) However there are no
reports of microbes able to de-epoxidze DON and other trichothecenes under aerobic
conditions and at moderate temperatures
To develop an effective method for eliminating DON and trichothecene mycotoxins
from contaminated grains soils from fields with FHBGER infected crops were screened for
DON de-epoxidizing microbes (refer to chapter 2 Islam et al 2012) One soil sample yielded
a mixed microbial culture was partially enriched for bacteria with DON de-epoxidizing
activity (chapter 2) The current study describes the purification and characterization of a
novel DON de-epoxidizing bacterium from this culture
68
33 Materials and methods
331 Microbial source
A mixed microbial culture originating from a screen of more than 150 separate soil
samples collected from maize and wheat fields near Guelph Ontario Canada (Islam et al
2012) was used for enrichment purification and characterization of the DON de-epoxidizing
bacterium
332 Media and chemicals
The media used were nutrient broth (NB) 110 strength NB (110 NB) minimal
medium (MM) consisting of six mineral salts (MS) + 100g per liter sucrose (Islam et al
2012) MS + 05 Bacto Peptone (MSB) MS + 03 Bacto Peptone (MSC) MS + 05
yeast extract (MSY) MS + 200 microgmL DON (MS-DON) sterile water-soluble soil extract
(SE) (Islam et al 2012) potato dextrose broth (PDB) and Luria Bertani (LB) Prepared media
and media components were obtained from Fisher Scientific (Fair Lawn New Jersey USA)
Sigma-Aldrich (Oakville ON Canada) andor Difco (Sparks Maryland USA) Media pH
was adjusted as required prior to autoclaving For solid media preparation 15 agar was
added DON and all trichothecene mycotoxins were purchased from Biomin (Romer Lab
Union MO USA)
333 Isolation of the DON de-epoxidizing bacterial strain
Aliquots (20 microL) of the microbial culture partially enriched for DON de-epoxidizing
ability (Islam et al 2012) was transferred to 05 mL MSB (pH 70) in sterile 15 mL
Eppendorf tubes After addition of 200 microgmL pure DON the culture was incubated at room
69
temperature with continuous shaking at 220 revolutions per minute (rpm) Seven to ten days
later 20 microL of the culture was transferred to fresh MSB supplemented with 200 microgmL DON
The culture was then incubated for 7 to 10 days under the above conditions This procedure
was repeated 12 times
After three months of repeated transfers in broth with 200 microgmL DON 100 microL of the
culture was serially diluted in 09 mL MSB containing 05 Tweenreg20 (Sigma Aldrich) to
reduce bacterial cell aggregation (Mabagala 1997) The dilutions were vigorously shaken for
ten minutes A 100 microL aliquot from each dilution was spread on MSB + agar (MSA) The
MSA plates were incubated at room temperature until single colonies appeared
Single colonies from plates were transferred to 1 mL MSB+ 50 microgmL DON and
incubated at room temperature with continuous shaking at 180 rpm under aerobic conditions
After three days the DON de-epoxidation ability was examined by mixing an equal volume
of the bacterial culture and methanol and incubating for 30 min at room temperature to lyse
the cells Insoluble materials in the mixture were removed by passing it through a 045 microm
Millipore filter (Whatmanreg Florham Park New Jersey USA) The level of de-epoxy DON
(dE-DON also known as DOM-1) in the filtrate was determined by liquid chromatography-
ultraviolet-mass spectrometry (LC-UV-MS) (Guan et al 2009)
Individual colonies that showed de-epoxidation activity were serially diluted in
MSB+05 Tweenreg20 and plated onto MSA Single colonies that appeared on the agar plate
were incubated in MSB as described above and retested for DON de-epoxidation ability To
ensure that pure cultures were obtained this was repeated 10 times and individual colonies
were selected on MSA each time
70
334 Identification of a DON de-epoxidizing bacterium
Strain ADS47 with DON de-epoxidation ability was obtained from a single colony
after 10 serial transfers and was grown on MSA at room temperature under aerobic
conditions for 72 h Colony color shape edge and elevation were observed The cell
morphology of the bacterium was examined with a transmission electron microscope at the
University of Guelph Guelph Ontario Canada Whole bacterial cells were placed directly on
a formvar and carbon-coated 200-mesh copper grid They were washed two times by touching
the grids to a drop of clean dH2O and the excess water was blotted off The samples were
negatively stained using 1 Uranyl Acetate (aq) for five seconds Electron microscopy was
performed on a Philips CM10 transmission electron microscope at 60kV under standard
operating conditions (Philips Optics Eindhoven Netherlands) Gram staining was performed
using the procedure described in the commercial gram stain kit from BD Canada
(Mississauga Ontario Canada)
Total bacterial DNA was extracted using a DNeasy Plant DNA Extraction Mini Kit
(Qiagen Inc) The 16S-rDNA was PCR amplified using the primers 46f-
5rsquogcytaacacatgcaagtcga3rsquo (Kaplan and Kitts 2004) and 1371r-5rsquogtgtgtacaagncccgggaa3rsquo
(Zhao et al 2007) PCR was performed in 25 microL containing 06 microM of each primer 1x of
HotStarTaqreg Master Mix (Qiagen Inc) and ~40 ng of DNA The PCR cycles were 95ordmC (15
min) 35 cycles of 94ordmC (20 s) 52ordmC (15 s) 72ordmC (1 min 30 s) and a 72ordmC (7 min) extension
step Amplification was carried out in a GeneAmpreg PCR System 9700 (Applied Biosystems)
PCR products were sequenced with an automated ABI 3730 DNA analyzer (Applied
Biosystems) The 16S-rRNA gene sequence similarity was performed by BlastN using the
Ribosomal Database Project (RDP) (httprdpcmemsueduseqmatch)
71
A phylogenetic tree was constructed by comparing the partial 16S-rDNA sequence of
strain ADS47 with the 11 known Citrobacter species (Bornstein and Schafer 2006 Brenner et
al 1993) The Citrobacter strains were C freundii [FN997616] C koseri [AF025372] C
amalonaticus [AF025370] C farmeri [AF025371] C youngae [AB273741] C braakii
[AF025368] C werkmanii [AF025373] C sedlakii [AF025364] C rodentium [AF025363]
C gillenii [AF025367] and C murliniae [AF025369] Escherichia coli O157H7 [AE005174]
was used as an out group strain The analysis was performed using distance matrix with a gap
opening of 100 and a gap extension of 01 by a server based ClustalW2 program
(httpwwwebiacukToolsmsaclustalw2) A dissimilarity tree was created and viewed
using a TreeView X software (Page 1996)
335 Effects of DON concentration aeration substrate temperature pH and antibiotic
To determine if DON concentration can affect the growth of the isolated bacteria
tubes containing 5 mL of NB or MSB with 20 200 or 500 microgmL of DON were inoculated
with strain ADS47 that had been grown for 16 h with continuous shaking at 220 rpm and then
adjusted to cell density at 025 OD600nm (107 cellsmL) The cultures were incubated at 28degC
with continuous shaking at 180 rpm and 05 mL was removed every 12 h to measure
OD600nm The culture (05 mL) was then analysed for DON de-epoxidation activity by LC-
UV-MS (Guan et al 2009) as described above Three replicate cultures were tested
To examine the effect of aeration five mL of NB or MSB containing 50 microgmL DON
were inoculated with an overnight culture of strain ADS47 prepared as described above and
incubated at 28˚C with continuous shaking at 180 rpm One mL of culture was removed every
six hours and bacterial growth was determined by measuring cell density at OD600nm At the
72
same time points culture samples were analysed for DON to dE-DON transformation ability
(Guan et al 2009) The experiment was repeated with MSB medium containing 50 microgmL
DON under anaerobic conditions as described by Islam et al (2012) with DON to dE-DON
transformation ability being determined every 6 h Three replicate cultures were prepared for
each assay
The effect of substrate on DON de-epoxidation ability was tested after growing
ADS47 for 72 h in NB 110 NB MM MS-DON MSB MSC MSY SE PDB or LB
containing 50 microgmL DON For this assay the media were maintained at pH70plusmn02 and 28degC
for 72 h The effect of temperature was examined with MSB containing 50 microgmL DON at 10
15 20 25 30 35 40 45 or 50degC The effect of pH on growth of strain ADS47 was examined
with MSB adjusted to pH 55 60 65 70 75 or 80 and the cultures were incubated at 28degC
for 72 h Three replicate cultures were used for each assay
Overnight cultures of ADS47 in MSB were treated with 001 and 0001 (wv) of
an antibiotic sodium azide (SA) (Sigma-Aldrich) Controls were prepared in MSB without
SA Three replicate cultures were spiked with 50 microgmL DON DON to dE-DON
transformation ability of the bacterial culture was analysed (Guan et al 2009) after 72 h at
28degC with continuous shaking at 180 rpm Three replicate cultures were tested
336 Localization of DON to dE-DON transformation ability in the bacterium
Bacterial cell extracts were prepared using B-PER II Bacterial Protein Extraction
Reagent following the manufacturerrsquos instructions (PI 78260 Thermo Scientific Rockford
IL USA) ADS47 was grown in 15 mL MSB for 48 h under the growth conditions described
above A culture (05 mL) was centrifuged at 5000 x g for 10 minutes and the pellet was
73
collected B-PER II reagent (2 mL) was added to the pellet and the cells were lysed by
pipetting the suspension up and down several times The cell lysate was treated with 20 microL
Halt Protease Inhibitor Cocktail (Thermo Scientific Rockford IL USA) Culture filtrate was
prepared by passing 05 mL of the culture through a 022 microm Millipore filter (Fisher
Scientific) The remaining 05 mL culture was used as control Cell lysate culture filtrate and
control were incubated with 50 microgmL DON and incubated at 28degC DON to dE-DON
transformation ability of the cell lysate culture filtrate and control were analysed after 72 h
incubation (Guan et al 2009) Three replicate cultures were tested
337 Detoxification of additional trichothecene mycotoxins
Strain ADS47 was evaluated for its ability to degrade five type-A trichothecene
mycotoxins (HT-2 toxin T-2 toxin T2-triol diacetoxyscirpenol and neosolaniol) and five
type-B trichothecene mycotoxins (nivalenol verrucarol fusarenon X 3-acetyl-
deoxynivalenol and 15-acetyl-deoxynivalenol) One and a half mL NB was added with 100
microgmL of each mycotoxin and then inoculated with 100 microL of ADS47 culture grown
overnight as previously described The culture of NB + mycotoxin + ADS47 was incubated at
28degC with continuous shaking at 180 rpm and DON to dE-DON transformation ability of the
culture was analysed by LC-UV-MS (Guan et al 2009) Five replicate cultures were tested for
each trichothecene mycotoxin
74
338 Statistical analysis
Data were processed using Microsoft Excel Program 2007 Statistical analysis was
performed using General Linear Model procedure in SAS version 91 (SAS Institute Cary
NC USA 2002-03) Replications were averaged and stated as mean plusmn standard error The
mean differences were considered significant at plt 005 according to Tukeyrsquos-HSD test
75
34 Results
341 Isolation and identification of a DON de-epoxidizing bacterium
A DON de-epoxidizing bacterium designated strain ADS47 was isolated after 12
transfers over three months from cultures growing in MSB containing 200 microgmL DON
Strain ADS47 produced small to medium size circular convex and white colonies on MSA at
room temperature under aerobic conditions (Figure 3-1a) The bacterium is rod shaped and
150 x 072 microm in size (Figure 3-1b) It has numerous flagella that cover the surface of the cell
(peritrichous flagella) and is Gram-negative
BlastN analysis of the 16S-rDNA sequence of strain ADS47 showed the most
similarity to the eleven known Citrobacter species in the gamma proteobacteria of the
Enterobacteriaceae In particular the 16S-rDNA nucleotide sequence of ADS47 was more
than 99 identical to C fruendii (Figure 3-2) The next most similar sequences were a cluster
of C werkmanii and C murliniae followed by more distantly related C braakii C gilleni
and C youngae A separate cluster was formed by C rodentium C farmeri C sedlakii C
koseri and C amalonaticus
76
Figure 3-1 (a) Colony color shape and size of the mycotoxin degrading bacterial strain
ADS47 on mineral salts agar (MSA) medium (b) Transmission electron microscopic (TEM)
image of the Citrobacter freundii strain ADS47 The bacterium showed multiple flagella
(peritrichous flagella) and numerous fimbriae around the cell surface
(a) (b)
77
Figure 3-2 Dendrogram based on 16S-rDNA sequences for strain ADS47 and 11 reference
Citrobacter species An E coli O157H7 strain was considered as an out group species The
tree was constructed using a genetic distance matrix by ClustalW 2 and viewed by
TreeViewX software The distance was converted to scale and shown by branch lengths (scale
bar 001 which indicated the 16S-rDNA nucleotide sequence variability between the bacterial
species) GenBank accession numbers for 16S-rDNA sequences of the species used for
comparison are shown in materials and methods section (334)
78
342 Effects of DON concentrations on bacterial DON de-epoxidation
Growth of ADS47 aerobically in MSB with 20 200 and 500 microgmL DON was not
significantly different from that observed in control cultures without DON (Figure 3-3a)
Similarly increasing DON concentrations did not affect bacterial growth in NB medium
(Figure 3-3b)
DON de-epoxidation activity determined as microgmL DON convertedcell density
(OD600) was measured lowest in MSB+20 microgmL DON and highest in MSB+ 500 microgmL
DON containing cultures (Figure 3-3c) The highest rate of dE-DON formation was 115 microgh
in ADS47 cultures in MSB with 500 microgmL DON between 48 to 72 h All of the DON was
converted in the cultures containing 20 microgmL DON and 200 microgmL DON between 48-60 h
and 60-72 h respectively In 200 microgmL and 500 microgmL of DON cultures the rate of de-
epoxidation increased significantly after 36 h
79
00
04
08
12
16
0 24 36 48 60 72 96
OD
600n
m
Incubation time (h)
Control
20 microgmL
200 microgmL
500 microgmL
(b)
(a)
80
Figure 3-3 Effects of DON concentration on bacterial growth and de-epoxidation activities
Strain ADS47 was grown in MSB and NB media with three different DON concentrations 20
microgmL 200 microgmL and 500 microgmL Control cultures were prepared without DON Cultures
were grown at 28degC with continuous shaking at 180 rpm (a) Microbial density (OD600 nm) in
MSB with different DON concentrations was measured at 24 36 48 60 72 and 96 h (b)
Microbial growth (OD600 nm) in NB having different DON concentration was measured at 24
48 72 and 96 h (c) Bacterial DON de-epoxidation activity in 20 200 and 500 microgmL DON
containing cultures at different time points DON de-epoxidation was measured in microgmL per
OD600 nm Three replicate cultures for each assay were analysed
(C)
81
343 Influence of growth conditions and antibiotic on microbial DON de-epoxydation
Aerobic and anaerobic conditions
ADS47 grew more quickly uner aerobic conditions under MSB+50 microgmL DON than
in MSB+50 microgmL DON under anaerobic conditions (Figure 3-4a) Aerobic growth was
faster in NB+50 microgmL DON compared to MSB+50 microgmL DON
However the DON de-epoxidation activities on a cell density basis were very similar
in NB+50 microgmL DON compared to MSB+50 microgmL DON (Figure 3-4b) Also DON de-
epoxidation activity of ADS47 under aerobic conditions in MSB+50 microgmL DON was higher
than in the same medium under anaerobic conditions (Figure 3-4b)
82
(b)
(a)
83
Figure 3-4 Bacterial growth and DON de-epoxidation capabilities under aerobic and
anaerobic conditions (a) Microbial growth under aerobic conditions in MSB and NB medium
and anaerobic conditions in MSB medium (b) Microbial DON de-epoxidation ability under
aerobic conditions in MSB and NB media and anaerobic conditions in MSB medium
Cultures media having 50 microgmL DON were inoculated with overnight culture of strain
ADS47 Initial microbial concentration in the cultures was adjusted to 025 OD600nm Cultures
were incubated at 28degC having three replications Bacterial growth at OD600nm and DON de-
epoxidation in microgmL per OD600nm were measured at six hour interval
84
Media temperature and pH
Growth of ADS47 in media containing 50 microgmL DON under aerobic conditions at 72
h was three-fold higher in NB and LB than in SE two-fold higher than in 110 NB and MSC
and did not occur in MS-DON (Figure 3-5a) Growth in MM MSB and MSY was
intermediate
Over the 72 h DON de-epoxidation per cell density was highest in MSB and MSY and
approximately 25 lower in NB and LB and lower by 75 and 87 in 110 NB and MSC
respectively (Figure 3-5b) No DON de-epoxidation was observed in MS-DON in which
DON was used as a carbon source Bacteria grown in MM broth which contains sucrose as a
sole carbon source were unable to de-epoxidize DON No DON de-epoxidation was observed
in SE and PDB grown cultures
Growth of ADS47 under aerobic conditions for 72 h in MSB+50 microgmL DON was
greatest between 25-40degC but no growth occurred at 50degC (Figure 3-6a) DON de-
epoxidation activity was greatest and equivalent between 20-40degC and lower at 10 15 and
45degC temperatures (Figure 3-6b)
ADS47 growth under aerobic conditions for 72 h in MSB+50 microgmL DON was
limited to neutral pHs (ie 60 - 75) (Figure 3-7a) The transformation of DON to dE-DON
occurred at the same pH range and DON de-epoxidation activity was not significantly
different between pH 60 to 75 (Figure 3-7b)
85
b
c
a
b
a
d c
b b
(a)
(b)
b
c
b
a a
d
86
Figure 3-5 Effects of media on bacterial growth and DON de-epoxidation activity (a)
Microbial growth (at OD600nm) on different culture media Media pHs were adjusted to
70plusmn02 (b) Bacterial DON to dE-DON transformation in microgmL by OD600nm in different
media after 72 h The cultures having three replications were incubated for 72 h at 28degC and
DON de-epoxidation was measured The Media NB = nutrient broth 1NB = 110 strength
NB MM = minimal medium MS-DON = six mineral salts + 200 microgmL DON as sole C
source MSB = six mineral salts + 05 bacto peptone MSC = six mineral salts broth + 03
bacto peptone MSY = six mineral salts with 05 yeast extract SE = soil extract PDB =
potato dextrose broth and LB = Luria Bertani Means with the same label are not significantly
different according to Tukeyrsquos HSD test (p lt 005)
87
(a)
(b)
88
Figure 3-6 Effects of temperature on microbial growth and DON de-epoxydation capability
(a) Microbial growth (at OD600nm) at different temperatures after 72 h of incubation (b)
Bacteria DON de-epoxidation in microgmL by OD600nm in different temperatures The most
efficient temperature range (ge40 microgmL DON de-epoxidation) is marked by dot lines The
experiment was conducted in MSB medium at pH 70plusmn02 with three replications
89
(a)
(b)
90
Figure 3-7 Influence of pH on microbial growth and DON de-epoxydation (a) Microbial
growth (OD600nm) in MSB at different pH (b) Bacterial DON to de-epoxy-DON
transformation in microgmL by OD600nm in different pH Dot lines indicated the efficient de-
epoxidation (ge40 microgmL DON conversion) pH range Experiment was conducted at 28degC
with three replications
91
Antibiotic
Treatment of ADS47 at 72 h in MSB+50 microgmL DON with sodium azide (SA) at
0001 and 001 reduced DON de-epoxidation activity to 5 (25 microgmL) and 0
respectively The control with no SA added was able to de-epoxidize DON completely
(100) (Figure 3-8)
Figure 3-8 Effects of sodium azide (SA) on bacterial DON de-epoxidation activity
Microbial culture in MSB+50 microgmL DON was treated with 001 (SA-1) and 0001 (SA-
2) sodium azide In control (C) no SA was added The experiments with three replications
were incubated for 72 h at 28degC and DON de-epoxidation was measured by LC-UV-MS
method
0
20
40
60
80
100
120
SA1 SA2 C
D
ON
de
-ep
oxyd
ize
d
92
344 Localization of bacterial DON de-epoxidation
A cell lysate prepared from ADS47 grown for 48 h in MSB transformed 20 (10
microgmL) of DON added at 50 microgmL to dE-DON within 72 h (Figure 3-9) In contrast a cell-
free culture filtrate did not show DON de-epoxidation activity
Figure 3-9 Determination of DON to de-epoxy DON transformation by bacterial culture
filtrate and cell extract The treatments having three replications were spiked with 50 microgmL
DON and incubated at 28degC for 72 h In control strain ADS47 was incubated in MSB broth
with 50 microgmL DON DON de-epoxidation activity in percentage was examined
0
20
40
60
80
100
120
Filtrate Extract Control
D
ON
de
-ep
oxyd
ize
d
93
345 Bacterial detoxification (de-epoxidtaion and de-acetylation) of other trichothecene
mycotoxins
Aerobic cultures of ADS47 grown for 72 h in NB+100 microgmL HT-2 converted 74 of
the toxin to de-epoxy HT-2 toxin and 26 to a de-acetylated product (Figure 3-10 and Table
3-1)
In a culture treated with T2-toxin 46 was converted to de-epoxy T-2 16 was
converted to an unknown compound and 38 was not converted In ADS47 culture with
diacetoxyscirpenol (DAS) 61 was transformed into de-epoxy DAS and 39 was de-
acetylated DAS (39) Neosolaniol (NEO) was transformed to de-epoxy NEO (55) and
de-acetylated NEO (45) T2-triol was transformed in to de-epoxy T2-triol (47) and de-
acetylated T2-triol (53)
Aerobic cultures of ADS47 were also grown for 72 h in NB+100 microgmL of one of five
type-B trichothecenes For nivalenol (NIV) 92 was de-epoxydized while the remaining 8
NIV was transformed to DON 3-acetyl-deoxynivalenol (3-ADON) was transformed into
three products de-epoxy NIV (5) de-epoxy DON (54) and DON (41) For 15-
acetyldeoxynivalenol (15-ADON) 8 was transformed to de-epoxy 15-ADON and 92 to
de-epoxy DON (dE-DON) FusarenonX (FUS) had 95 transformed into de-epoxy FUS
while 97 of verrucarol (VER) was transformed to de-epoxy VER The remaining 5 FUS
and 3 VER were not transformed
94
Figure 3-10 Determination of microbial de-epoxidation and de-acetylation of HT-2 toxin by
LC-UV-MS Bacterial culture in NB was added with 100 microgmL HT-2 toxin Bacterial HT-2
toxin transformation ability was examined after 72 h of incubation at 28degC The upper panel
illustrated mass spectrometry chromatograph of the de-acetylated (red peak) and de-
epoxydized (blue peak) products Arrows indicated the targeted acetyl (red) and epoxy (blue)
functional groups of HT-toxin The middle and lower panels showed the mass spectra of the
de-acetylated (dA) and de-epoxidized (dE) compounds Arrows on the upper panel indicated
the targeted acetyl (red) and epoxy (blue) functional groups of HT-2 toxin Red and blue
arrows on the middle and lower panels pointed at the degradation sites of HT-2 toxin
95
Table 3-1 Microbial transformation of type-A and type-B tricthothecene mycotoxins after 72
h of incubation under aerobic conditions at room temperature DAS=diacetoxyscirpenol
DON=deoxynivalenol FUS=FusarenonX 3AON=3-acetyl-deoxynivalenol and
15ADON=15-acetyl-deoxynivalenol
1 Convertion of trichothecene mycotoxins to either DON or an unknown compound
Trichothecene
mycotoxin
Untransformed
trichothecenes ()
Transformed products of
trichothecenes ()
De-
epoxy
De-
acetylate
Other1
Type-A
HT-2 toxin 0 74 26 -
T-2 toxin 38 46 - 16
(unknown)
DAS 0 61 39 -
Neosolanol 0 55 45 -
T2-triol 0 47 53 -
Type-B
DON 0 100 - -
Nivalenol 0 92 - 8 (DON)
3-ADON 0 59 41 (DON)
15-ADON 0 100 - -
FUS 5 95 - -
Varrucarol 3 97 - -
96
35 Discussion
The ADS47 strain of C freundii was isolated in the present study by a prolonged
incubation of a mixed microbial culture with a high concentration DON since the bacterium
efficiently de-epoxidized (detoxified) DON and number of other trichothecene mycotoxins
under aerobic and anaerobic conditions at moderate temperatures and neutral pH The total
procedures for enrichment and isolation of the strain ADS47 is depicted in supplemental
Figure S3-1 The reductive de-epoxidation activity may be due to a cytoplasmic enzyme
which would be in agreement with a previous study that showed de-epoxidation of
epoxyalkanes was catalyzed by a bacterial cytoplasmic flavoprotein (Westphal et al 1998)
The ability of the currently isolated strain of C freundii to biodegrade environmental
pollutants such as aminoaromatic acids and heavy metals (copper) is also likely mediated by
cytoplasmic enzymes since these compounds need to be taken up by the cells before they are
metabolized (Savelieva et al 2004 Sharma and Fulekar 2009) In contrast oxidative
transformation of DON to 3-keto DON by microbial extracellular enzyme has been reported
(Shima et al 1997)
Isolation of a trichothecene de-epoxidizing microorganism from a mixed microbial
community is very challenging Although trichothecenes are toxic to eukaryotes prokaryotes
appear to be resistant to these mycotoxins (He et al 1992 Rocha et al 2005) The present
study showed that the bacteria grew well in media with high concentrations of DON (500
microgmL) Bacterial de-epoxidation of DON involves the removal of an oxygen atom from the
functional epoxy ring of the tricyclic DON structure (Yoshizawa et al 1983) Such co-
metabolic degradation does not provide the bacteria with any nutrients from the reacting
compounds (Horvath 1972 Fedorak and Grabic-Galic 1991) This is supported by the results
97
of the current study which showed that strain ADS47 was unable to grow in media with DON
as a sole carbon source and increasing the concentration of DON did not increase ADS47
growth in MSB Hence DON could not be used as sole nutrient source in the mixed culture
enrichment process
Many previous attempts to obtain DON de-epoxidizing microbes have failed
particularly under aerobic conditions because media or laboratory conditions were
inappropriate to support growth or initially active cultures lost de-epoxidizing function after
serial transfer (Karlovsky 2011)
Media composition appears to be a crucial factor for microbial trichothecene de-
epoxidation activity (Voumllkl et al 2004) Effective biotransformation of trichothecene
mycotoxins by the fish digesta microbial culture C133 occurred in full media (FM) but a low
level of de-epoxidation occurred in media having a single nitrogen source and no
transformation activity was observed in the common bacterial medium LB (Guan et al
2009) Zhou et al (2010) also showed that chitin de-acetylation by soil bacterial strains
occurred more rapidly in yeast extract compared to common bacterial media Therefore a
screening strategy for microbial DON de-epoxidation ability should use different media in
parallel as was done in this study
This study observed efficient de-epoxidation in common bacterial media (NB and LB)
as well as in mineral salts medium containing yeast extract (MSY) or 05 Bacto Peptone
(MSB) However reduction of Bacto-Peptone to 03 (MSC) significantly reduced the
bacterial de-epoxidation activity suggesting that the activity is sensitive to the availability of
protein and amino acids The study found that medium containing sucrose as carbon source
(MM) and a commonly used medium PDB blocked microbial DON de-epoxidation even
98
though bacteria grew in those growth media These suggest that media containing high sugar
contents had inhibitory effects on microbial de-epoxidation enzymatic activity Previously
sugars have been found to have negative effects on the production andor activity of bacterial
enzymes (Saier and Roseman 1971)
Losses of microbial functions under laboratory conditions limited researchers to obtain
economically important microbial genetic sources The soil borne Curtobacterium sp strain
114-2 efficiently de-epoxydized nivalenol but unfortunately the strain lost its activity in
latter generations (Ueno et al 1983) Instability of de-epoxidation activity of ADS47
observed in cultures grown on agar media without DON resulted in the loss of DON de-
epoxidation activity by more than 80 of the colonies after one generation However strain
ADS47 retained de-epoxidation activity after ten subcultures in MSB (data not shown)
The non-specific de-epoxidation of DON made it impossible to use DON as a sole
substrate C source for selective isolation of ADS47 Thus a number of techniques were
utilized to select ADS47 and suppress other bacteria including the use of a variety of
antibiotics and culture at elevated temperatures (Islam et al 2012) Selection of the isolate
ADS47 required repeated subculturing and prolonged exposure to media containing 200
microgmL DON After three months of selection with DON culture that was obtained had a
DON de-epoxidation activity that was up to 30-fold higher than the mixed culture As growth
of ADS47 was not affected by 200 microgmL DON the prolonged incubation may have reduced
populations of other bacteria that are more sensitive to DON Enrichment and isolation of a
DON to 3-keto DON transforming bacterial strain E3-39 by prolonged incubation of a soil
microbial culture with 200 microgmL DON was reported by Shima et al (1997)
99
This study demonstrated that microbial DON de-epoxidation occurred mainly at the
stationary growth phase A DON degrading strain of Citrobacter may have gained energy
from DON by co-metabolic cleavage of the epoxy ring under less nutrient conditions
Microbial use of energy from co-metabolic reaction has been suggested by Chiu et al (2005)
The rate of de-epoxidation activity increased when ADS47 grew at higher concentrations of
DON Taken together a prolonged exposure to high concentration DON may have resulted in
non-DON metabolizing microbial populations declining by DON toxicity while facilitated
the population of DON metabolizing bacterium by providing energy
DON is a small molecule and is simply absorbed by cells (Sergent et al 2006)
Microbial DON de-epoxidation occurred in intact and lysed bacterial cells In addition
treating the ADS47 culture with SA which blocks gram negative bacterial electron transport
and activity inhibited DON de-epoxidation activity (Lichstein and Malcolm 1943 Erecinska
and Wilson 1981) implying that an active electron transport system required for microbial de-
epoxidation reaction (He et al 1992) The data suggest that a cytoplasmic epoxy reductase is
involved in the DON de-epoxidation reaction in the ADS47 microbial strain that was isolated
in the present study
Since agricultural commodities are often contaminated with multiple fungal
mycotoxins the ability of bacteria like ADS47 to degrade mycotoxins by several
mechanisms may be an advantage De-acetylation might be another mechanism of microbial
mycotoxin detoxification since toxicity of trichothecenes also relies on the acetyl functional
groups (Desjardins 2006) Strain ADS47 showed efficient de-epoxidation and de-acetylation
of several type-A and type-B trichothecene mycotoxins that are commonly detected in cereals
in association with DON In conclusion the soil-borne gram-negative bacterium C freundii
100
strain ADS47 is capable of detoxifying trichothecene mycotoxins by both de-epoxidation and
de-acetylation under aerobic and anaerobic conditions neutral pH and a range of
temperatures ADS47 has great potential to detoxify DON contaminated animal feed
Furthermore isolation of novel bacterial de-epoxidation genes might be used to generate
transgenic plants that can detoxify DON in contaminated seed Alternatively contaminated
seed might be treated with manufacture de-epoxidation enzyme(s) These applications can
improve the quality and safety of agricultural commodities that are prone to trichothecene
mycotoxin contamination
36 Acknowledgements
Thank to the Natural Sciences and Engineering Research Council of Canada (NSERC) for a
scholarship to Rafiqul Islam The work was funded by the Agriculture and Agri-Food Canada
Binational Agricultural Research and Development Fund (AAFCBARD) project We
appreciate Honghui Zhu for technical assistance
101
Supplemental Figure S3-1 Flow chart depicts the procedures of enrichment and isolation of
DON de-epoxidation bacterial strain ADS47 Steps 1 to 4 describe the optimization of
microbial DON to dE-DON transformation activity and removal of undesired microbial
species Step 5 to 7 shows the culture enrichment by incubation with high concentration DON
and isolation of the single strain ADS47
Step 1 Screening of soil samples
Step 2 Optimization of cultural conditions
Step 3 Elimination of undesired microbial species
Step 4 Removal of untargeted microbial species
Step 5 High concentration DON enrichment
Step 6 Separation of bacterial cells and dilution plating
Step 7 Single colony analysis
Active culture was sequentially treated with different mode of actions antibiotics heating cultures at 50degC and subculturing on agar media
Colonies were tested for their DON de-epoxidation ability Purified the single colony via several times subculturing on agar medium
Optimized the culture conditions ie growth substrates temperatures pH and oxygen requirements
Obtained DON de-epoxidation capable microbial culture obtained by analyzing soil samples collected from cereal fields
Serially diluted active culture was grown on agar media Collected washed colonies having DON de-epoxidation activity
The partially enriched culture that retained DON de-epoxidation activity was incubated with 200 microgmL DON in MSB media with continuous shaking at 220 microgmL for 7 d The procedure was repeated 12 times
The enriched culture was serially diluted and treated dilutes with 05 Tween 20 After vigorously vortex for ten min aliquot cultures were spread on different agar media plates to produce single colonies
102
CHAPTER 4
40 CONSTRUCTION AND FUNCTIONAL SCREENING OF GENOMIC DNA
LIBRARY FOR ISOLATION OF DON DE-EPOXYDIZING GENE(S) FROM
CITROBACTER FREUNDII STRAIN ADS47
41 Abstract
Enzymatic activity-based screening of a genomic DNA library is a robust strategy for
isolating novel microbial genes A genomic DNA library of a DON detoxifying bacterium
(Citrobacter freundii strain ADS47) was constructed using a copy control Fosmid vector
(pCC1FOSTM
) and E coli EPI300-T1R as host strain A total of 1250 E coli clones containing
large DNA inserts (~40 kb) were generated which resulted in about 10-fold theoretical
coverage of the ADS47 genome The efficiency of E coli transformation with the
recombinant Fosmid vector was examined by restriction digestion of Fosmids extracted from
15 randomly selected library clones The fragmentation patterns showed that all of the clones
contained an 81 kb Fosmid backbone and ~40 kb of insert DNA except one that had a small
insert This suggests that a DNA library was prepared with expected DNA inserts The de-
epoxidation ability of the library was analysed by incubating clones in Luria Bertani (LB)
broth medium supplemented with 20 microgmL DON with induction (by addition of L-arabinose)
or without induction None of the 1000 clones showed DON de-epoxidation activity The
potential DON de-epoxidation gene(s) of ADS47 may not have been expressed in E coli or
might have been expressed but lacked additional factors required for DON de-epoxidation
103
Keywords DNA library fosmid vector functional library screening DON de-epoxidation
activity
42 Introduction
Microorganisms contain numerous valuable genes and biocatalysts which may have
significant roles in agricultural productivity (Jube and Borthakur 2007) Various techniques
have been used to isolate novel genes from different organisms including transposon
mutagenesis (Frey et al 1998 Tsuge et al 2001) gene knockout (Winzeler et al 1999)
whole genome sequencing (Krause et al 2006) comparative genomics (Koonin et al 2001)
and suppression subtractive hybridization (Dai et al 2010)
The genomic DNA library preparation and activity-based screening of the library is a
relatively new technique which has been efficiently used for isolation of microbial novel
enzymes and compounds such as lipases proteases oxidoreductases antibiotic resistance
genes membrane proteins and antibiotics (Henne et al 2000 Gupta et al 2002 Knietsch et
al 2003 Gabor 2004 Suenaga et al 2007 van Elsas et al 2008) The strength of the
technique is that genes from microbial sources could be obtained without prior sequence
information and not requiring culturing the organisms under laboratory conditions Using this
technique an unknown gene or a set of genes could be obtained by determining particular
phenotypes of the library clones Eventually the gene of interest from the positive clones is
isolated by using a forward (eg mutation) or reverse genetics method (eg subcloning the
positive clones into a microbial expression vector) followed by functional analysis of the
subclones
104
Although this strategy has been used effectively for isolating novel microbial genes
several factors need to be considered in order for the method to be successful These factors
include DNA quality vector type (eg plasmid cosmid fosmid bacterial artificial
chromosome etc) vector copy number and host strains (Daniel 2005 Singh et al 2009) The
Fosmid vector-based technique has been used successfully to clone bacterial genes including
xenobiotics degradation genes antifungal gene clusters and antibiotic resistance genes from
soil metagenomes (Hall 2004 Suenaga et al 2007 Chung et al 2008) This study reports the
construction and activity-based screening of a Fosmid DNA library in an E coli host with the
aim of isolating a novel DON de-epoxidation gene from Citrobacter freundii strain ADS47
(Chapter 3)
43 Materials and methods
431 Genomic DNA library construction
The genomic DNA library was prepared using a Copy Control Fosmid Library
Production Kit (Epicentre Biotechnologies Madison Wisconsin USA) following the
manufacturerrsquos instructions as depicted in Figure 4-1
105
Figure 4-1 Schematic overview of the construction and functional screening of a Fosmid
DNA library
106
Citrobacter freundii strain ADS47 was grown in NB medium at 28degC with continuous
shaking at 220 rpm The high molecular weight bacterial genomic DNA was extracted from
the overnight culture using the Gentra Puregene YeastBacteria Kit (Qiagen Inc Mississauga
Ontario Canada) The purity of the DNA was confirmed by determining OD260280 at ~190 by
a NanoDrop spectrophotometer A 5 microL sample having ~25 microg high molecular weight DNA
was sheared by passing through a 200-microL pipette tip and achieved a sufficient amount of
DNA about 40 kb in size Blunt ended 5rsquo-phosphorylated insert DNA was generated by
thoroughly mixing it with End-Repair Enzyme reagents on ice The 20 microL mixture was
incubated for 45 min at room temperature followed by incubation at 70degC for 10 min to
inactivate the End-Repair Enzyme activity
The treated DNA was size fractionated by a Low Melting Point-Pulsed Field Gel
Electrophoresis (LMP-PFGE) on the clamped homogeneous electric fields (CHEF-DR III)
system (Bio-RAD Laboratories) using 1 UltraPure Low Melting Point Agarose (Invitrogen)
The PFGE was conditioned according to manufacturerrsquos instructions as follows initial switch
time-42 s final switch time-761 s run time 16 h 50 min angle-120deg gradient- 6 Vcm
temperature-14degC and running buffer-05 TBE DNA size markers for the 42 kb Fosmid
control DNA (Epicentre Biotechnologies) and low range Pulse Field Gel (PFG) markers (New
England Biolabs Inc) were loaded adjacent to the DNA sample slots After electrophoresis a
portion of the gel that contained DNA size markers was cut off The gel was stained in SYBR
Safe DNA gel stain (Invitrogen) for 30 min visualized and marked on the 23 42 and 485 kb
marker bands A slice of PFGE gel containing 40 plusmn 10 kb size DNA was cut off with a sterile
razor blade by comparing the marker sizes DNA from the gel was recovered by digesting it
with GELase enzyme ( 1 U100 microL melted agarose) in 1X GELase Buffer followed by a
107
sodium acetate-ethanol precipitation The DNA with the desired size was stored at 4degC for
next dayrsquos ligation to a cloning-ready Fosmid vector (pCC1FOSTM
)
Ligation was performed in a 10 microL reaction mixture by thoroughly mixing 10X Fast-
Link Ligation Buffer 10mM ATP 025 microg of ~40 kb insert DNA 05 microg Fosmid Copy
Control Vector (pCC1FOS) (Figure 4-2) and 2 U Fast-Link DNA lipase The mixture was
incubated for 4 h at room temperature preceded by inactivating the ligation reaction by
heating at 70degC for 15 min The recombinant Fosmid vector was stored at -20degC for later use
Ten microL of the ligated DNA was thawed on ice and transferred to a pre-thawed
MaxPlax Lambda Packaging Extract tube containing 25 microL extract By pipetting several
times the mixture was incubated at 30degC for lamda packaging After 2 h of packaging an
additional 25 microL packaging extract was added into the mixture and incubated for a further 2 h
at 30degC The packaged phage particles were volumed to 05 mL by adding ~400 microL Phage
Dilution Buffer and 25 microL chloramphenicol
Transformation was performed via transduction by mixing the packaged particles with
the overnight cultured E coli EPI300-T1R cells (1 10 vv ratio) in LB supplemented with 10
mM MgSO4 and 02 Maltose Transformation was completed by incubation at 37degC for 1 h
The transformed E coli cells were spread on an LB agar plate containing 125 microgmL
chloramphenicol The plates were incubated overnight at 37degC Colonies that appeared on the
plates were individually transferred to 96-well microtiter plates having 200 microL LB medium
supplemented with 10 glycerol and 125 microgmL chloramphenicol The microtiter plates
were incubated at 37degC for 24 h with continuous shaking at 180 rpm The prepared library
clones were stored at -80degC for further analysis and functional screening
108
109
Figure 4-2 Genetic map of copy controlled Fosmid vector (pCC1FOSTM
) The 813 kb
Fosmid vector backbone shows a multiple cloning site (MCS) chloramphenicol resistance
selectable marker (CmR) replication initiation fragment (redF) inducible high copy number
origin (oriV) replication initiation control (repE) and F-factor-based partitioning genes (ParA
ParB ParC) The vector contains a cos site for lamda packaging (not shown) and lacZ gene
for bluewhite colony selection The vector map was provided by the Epicentre
Biotechnologies (httpwwwepibiocomitemaspid=385)
110
432 Confirmation of library construction
The quality of the DNA library prepared was determined by examining 15 E coli
clones randomly selected A 15 mL LB broth supplemented with 125 microgmL
chloramphenicol was inoculated by each of the transformed clones To obtain higher Fosmid
DNA yield 1 microL autoinduction solution (L-arabinose) to increase the copy number of the
Fosmid vector (Epicentre) was added to the cultures After an overnight incubation of the
cultures at 37degC with continuous shaking at 220 rpm Fosmid DNA was extracted using a
Qiagen Large-Construct Kit followed by restriction digestion with Not I enzyme A 20 microL
digestion mixture containing 05 microg Fosmid DNA 1X digestion buffer and 05 U Not I was
incubated at 37degC for 15 h The partially digested DNA and a 42 kb DNA size marker
(Fosmid control DNA) were ran on 08 agrose gel for 15 h at 100 V The linear Fosmid
vector backbone (813 kb) and insert DNA fragments were visualized on the gel and the
image was recorded
433 Functional screening of the clones
Eppendorf tubes (15 mL) containing 500 microL LB supplemented with 125 microgmL
chloramphenicol were inoculated with a single clone (400 clones tested) or a combination of
12 library clones (600 tested) from microtiter plates stored at -80degC One half of the cultures
in each group (single or combined clones) were induced to increase the Fosmid copy number
by adding a 1 microL (500X) autoinduction solution (contained L-arabinose) to the culture tubes
L-arabinose induces the expression of a mutant trfA gene of the E coli host strain which
leads to the initiation of Fosmid replication from a low-copy to a high-copy number origin of
replication (oriV) The cultures were spiked with 20 microgmL DON and incubated at 37degC for
111
72 h with continuous shaking at 180 rpm The DON to de-epoxy DON transformation ability
of the clone cultures was examined by a liquid chromatography-ultraviolet-mass spectrometry
(LC-UV-MS) method as described by Islam et al (2012)
112
44 Results
441 Production of the DNA library
High molecular weight intact (non-degraded) and good quality (OD600280 ~190)
genomic DNA was obtained from the DON de-epoxidizing bacterial strain ADS47 (data not
shown) Pipetting of the DNA extract resulted in an adequate amount of insert DNA (gt025
microg) of approximately 40 kb size as showed on a LMP-PFGE (Figure 4-3)
A DNA library in E coli host was constructed using LMP-PFGE purified DNA The
library was comprised of 1250 E coli transformants which were selected on LB agar plates
supplemented with chloramphenicol (Figure 4-4)
113
Figure 4-3 Preparation of a 40 kb size insert DNA from a DON de-epoxidizing bacterium
Genomic DNA was extracted from the overnight culture of a DON de-epoxidizing
Citrobacter freundii strain ADS47 After fragmentation the DNA was size fractionated by
Low Melting Point- Pulse Field Gel Electrophoresis (LMP-PFGE) with markers for the PFGE
(M1) and 42 kb Fosmid control DNA (M2) S1 S2 S3= different DNA samples extracted
from bacterial strain ADS47 The red dots area indicates the section of the gel containing
genomic DNA of the expected size (~40 kb)
114
Figure 4-4 Production of the genomic DNA library in E coli EPI300-T1 host strain
Approximately 40 kb long blunt ended genomic DNA from a DON de-epoxidizing bacterial
strain ADS47 was ligated to a Copy Control Fosmid vector pCC1FOS The construct was
packaged into lamda bacteriophage followed by a transduction-mediated transformation of E
coli host cells The clones which appeared on the LB agar plate supplemented with 125
microgmL chloramphenicol are expected to be transformed by the recombinant Fosmid vector
containing insert DNA
115
442 Conformation of the library construction
All of the clones tested acquired the Fosmid vector Out of 15 clones examined 14
received the recombinant Fosmid vector that contain the expected size insert DNA as
determined by an 81 kb vector backbone and at least one high molecular weight insert DNA
band (~40 kb) on agarose gel (Figure 4-5) In contrast one clone (lane 4) showed an 81 kb
vector backbone and a low molecular weight band which is also a fragment of the Fosmid
vector The result indicated that approximately 94 of the library clones were transformed
with the recombinant vector
443 Activity-based screening of the library
Out of 500 single copy number clones (200 individual and 300 combined clones)
none showed DON to de-epoxy DON transformation activity (Table 4-1) An additional 500
high copy number clones (200 single and 300 combined clones) were unable to de-epoxidize
DON as well
116
Figure 4-5 Confirmation of the Fosmid DNA library production in E coli Fosmid DNA
collected from 15 clones was digested with Not I and run on 08 agarose gel at 100 V for 15
h The presence of the vector backbone (81 kb) and the insert DNA fragment (~40 kb) in the
clones was determined by comparing a 42 kb molecular size marker (M) Lanes (1-15) denote
the recombinant Fosmid DNA extracted from 15 E coli clones
117
Table 4-1 Analysis of Fosmid DNA library clones for deoxynivalenol (DON) de-epoxidation
capability
Type of clones Analysis of
singlecombined
clones
Number of
clones
examined
Number of clones
showed DON de-
epoxidation activity
Without induction
(single copy number clones)
Singly 200 0
Combination of 12 300 0
With induction (multi-copy
number clones)
Singly 200 0
Combination of 12 300 0
De-epoxidation activity of the clones was examined alone or in combination of 12 clones
without or with induction by adding L-arabinose The clones grown in LB supplemented with
125 microgmL and 20 microgmL DON were analysed for DON to de-epoxy DON transformation
ability by a liquid chromatography-ultra violet- mass spectrum (LC-UV-MS)
118
45 Discussion
While a Fosmid DNA library with the expected genome coverage of a DON de-
epoxidizing bacterial genome was produced the function-derived screening of the library
clones did not identify any clones that showed DON de-epoxidation enzymatic activity This
is unlikely due to lack of coverage of the bacterial genome as 1000 clones analyzed in the
study should have provided eight fold coverage of the genome Therefore it was likely due to
a lack of expression of the enzyme(s) for DON de-epoxidizing activity under the conditions
used
Thus far there is only one report of heterologous expression of bacterial epoxide
reductase gene in another bacterium A high level of heterologous expression of a
Synechococcus sp vitamin K epoxide reductase gene was achieved in E coli with codon
optimization and placing the gene under the control of a prokaryotic T7 promoter in an
expression vector (Li et al 2010) Therefore heterologous expression of bacterial epoxide
reducatse gene using its own promoter has not yet been demonstrated The success of the
method used in this chapter relies on an intact copy of the gene or operon being cloned the
selection of an appropriate cloning vector a host organism that can recognize the cloned C
fruendii promoter and a suitable library screening method for DON degradation by E coli
(Mocali and Benedetti 2010)
The unbiased cloning efficiency of the Fosmid vector its ability to contain large insert
DNA high stability in E coli as single copy number and autoinduction to increase copy
number in host cells made it appear to be a suitable method for the present work (Kim et al
1992 Lee et al 2004) Library production with large insertions is a major advantage as gene
cluster involved in particular pathway could be trapped into single clone and their functions
119
could be analysed The pathway of microbial DON de-epoxidation activity and the genes
encoding the activity are not known Therefore it was reasonable to prepare a library with
large inserts to ensure the possibility of cloning single as well as whole pathway genes of
interest for functional screening A further advantage of the method is that Fosmid clones are
inducible which allows detecting poorly expressed genes in a heterologous host by inducing
high copy numbers (Daniel 2005)
One of the limitations of the method is that the functions of the target genes rely
mostly on the host organismrsquos transcription and translation machineries Therefore selection
of the host organism is critical for activity-based screening of library clones in heterologous
hosts A number of host strains including E coli Streptomyces lividans Rhizobium
leguminosarum and Pseudomonas aeruginosa have been used for functional screening
activities from diverse organisms (Daniel 2005) The present work attempted to isolate C
freundii strain ADS47 genes encoding DON de-epoxidation enzymes Both strain ADS47 and
E coli belong to gamma proteobacteria which suggest that E coli would be an appropriate
host organism for heterologous expression of ADS47 genes Poor expression of heterologous
genes could be another limitation of the method which may be overcome by overexpression
to increase the catalytic activity of less expressed genes However clones with DON de-
epoxidation activity were not identified in this study even the Fosmid copy number was
increased by induction
In general a large number of clones must be analysed to obtain the gene of interest
using an activity-based screening technique (Lee et al 2004 van Elsas et al 2008) In this
study the constructed DNA library comprised of 1160 clones having approximately ~40 kb
insert DNA suggesting that the prepared library achieved ~10 fold coverage of the ADS47
120
genome (48 Mb) which ensured gt99 probability of the presence of all DNA sequence in
the library In the produced library three clones (lanes 3 10 and 12) showed one intact band
greater than 42 kb in size on the agarose gel It is presumed that the Fosmid DNA of the three
samples was not digested because of insufficient amount of restriction enzyme added to the
digestion mixture About half of the samples demonstrated multiple bands suggesting that the
insert DNA might have contained multiple Not I restriction sites
However not every heterologous gene may necessarily express or function in the E
coli host (Gabor et al 2004) The functionality of the gene requires expression of the genes
and proper folding of the protein which may be influenced by environmental conditions (eg
substrate temperature and pH) and genetic factors In this study library clones were screened
in growth conditions at which strain ADS47 performed the best de-epoxidation activity The
expression of genes may be controlled by the promoter suitable ribosome binding site and
transndashacting factors (eg special transcription factors inducers chaperones cofactors proper
modifying enzymes or an appropriate secretion system) (Gabor et al 2004) Those factors
have to be supplied by the cloned DNA fragments or host cells to allow any heterologous
gene to be functional Since DON de-epoxidation genes of strain ADS47 evolved in a distinct
environmental condition the expression and activity of the gene(s) might have required
specific factorsregulators that were absent in E coli Even though the Fosmid based library
screening technique facilitated isolation of many microbial genes clones having de-
epoxidation activity were not identified using such technique in this study
121
46 Acknowledgements
We appreciate the Agriculture and Agri-Food CanadaBinational Agricultural
Research and Development Fund (AAFCBARD) project for providing the financial support
for this work The authors thank Honghui Zhu for analyzing numerous samples by liquid
chromatography ultraviolet mass spectrum method
122
CHAPTER 5
50 GENOME SEQUENCING CHARACTERIZATION AND COMPARATIVE
ANALYSIS OF CITROBACTER FREUNDII STRAIN ADS47
51 Abstract
The genome of a trichothecene mycotoxin detoxifying C freundii strain ADS47
originating from soil was sequenced and analyzed Strain ADS47 contains a circular
chromosome of 4878242 bp that encodes 4610 predicted protein coding sequences (CDSs)
with an average gene size of 893 bp Rapid Annotation Subsystems Technology (RAST)
analysis assigned functions to 2821 (62) of the CDSs while 1781 (38) CDSs were
assigned as uncharacterized or hypothetical proteins Comparative bioinformatics analyses
were done for strain ADS47 C freundii Ballerup 785139 Citrobacter sp 30_2 C youngae
ATCC 29220 C koseri ATCC BAA-895 and C rodentium ICC168 There were a significant
number of common orthologous genes (2821) were found in the six Citrobacter genomes
implying a common evolutionary relationship ADS47 is most closely related to the
opportunistic human pathogen C freundii Ballerup The human and mouse pathogenic
species C rodentium and C koseri are the most distantly related to ADS47 and non-
humananimal pathogens C youngae and Citrobacter sp 30_2 were closely related ADS47
C koseri C youngae and Citrobacter sp 30_2 lacked the clinically important locus for
enterocyte effacement (LEE)-encoded type-III secretion systems which are present in C
rodentium and C freundii Ballerup Strain ADS47 had 270 unique CDSs many of which are
located in the large unique genomic regions potentially indicating horizontal gene transfer
123
Among the unique CDSs nine were reductasesoxidoreductases and a de-acetylase that may
be involved in trichothecene mycotoxin de-epoxidation and de-acetylation reactions
Keywords Citrobacter freundii comparative genomics genome sequencing unique genes
reductases
52 Introduction
According to the Food and Agriculture Organization (FAO) at least 25 of the
worldrsquos food crops are contaminated with mycotoxins causing serious negative impacts on
livestock and international trade (Boutrif and Canet 1998 Desjardins 2006 Binder et al
2007) Trichothecenes (TCs) produced by a variety of toxigenic Fusarium species are
probably the most prevalent food and feed contaminating mycotoxins It is estimated that
infections by the fungus caused more than $1 billion in crop yield losses in USA and Canada
alone (Windels 2000 Schaafsma et al 2002 Yazar and Omurtag 2008) TC mycotoxins can
have adverse affects on human health including organ toxicity mutagenicity carcinogenicity
neurotoxicity modulation of the immune system negative reproductive system affects and
growth retardation (DrsquoMello et al 1999 Pestka et al 2007) A strain of Citrobacter freundii
ADS47 was isolated from soil in southern Ontario that is capable of detoxifying (ie de-
epoxidation and de-acetylation) several TC mycotoxins including deoxynivalenol nivalenol
and T2-toxin (Chapters 2-3)
Citrobacter freundii is a gram negative motile rod shaped facultatively anaerobic
bacterium belonging to the family of Enterobacteriaceae (Wang et al 2000 Whalen et al
2007) The opportunistic human pathogen is responsible for a number of infections including
124
nosocomial infections of respiratory tract urinay tract blood and causes neonatal meningitis
(Badger et al 1999 Wang et al 2000) C freundii exists almost ubiquitously in soil water
wastewater feces food and the human intestine Biodegradations of tannery effluents copper
aromatic amino acids by C freundii strains isolated from water mine tailings and soil
respectively have been previously reported (Kumar et al 1999 Savelieva et al 2004 Sharma
and Fulekar 2009) Because of their biochemical capabilities bacteria have played important
roles in industrial biotechnology including de-epoxidation and de-acetylation activities
(Demain and Adrio 2008 Plank et al 2009 Zhao et al 2010) However the identification of
genes for de-epoxidation activity against trichothecene mycotoxins is limited (Boutigny et al
2008)
The advent of molecular techniques such as next generation sequencing technologies
and the related bioinformatics tools has enabled the discovery of novel genes and the
enzymes they encode in pathogenic and beneficial bacteria (van Lanen and Shen 2006
Ansorge 2009 Zhao 2011) In the present study the genome of mycotoxin-detoxifying strain
C freundii ADS47 was sequenced and subjected to comparative genomic analyses with other
Citrobacter species to identify putative mycotoxin-detoxifying genes
125
53 Materials and methods
531 Sequencing quality control and assemble of ADS47 genome
Genomic DNA of the strain ADS47 was extracted using Gentra Puregene
YeastBacteria Kit (Qiagen Inc Mississauga Ontario Canada) and purity of the DNA was
confirmed by determining OD 260280 at ~190 using a Nanodrop spectrophotometer
The genomic DNA was sent to be sequenced by the Illumina HiSeq 2000 sequencing
analyzer at Beijing Genomics Institute (BGI) Hong Kong
(httpwwwbgisequencecomhomeservicessequencing-servicesde-novo-sequencing) The
sequencing workflow involved short read library preparation cluster generation sequencing
and data analysis Briefly 3-5 ug of bacterial gDNA was fragmented to produce a 500 bp
insert library by preparative electrophoresis through a 08 agarose gel The fragmented
DNAs were blunt-ended by treating with a mixture of enzymes (T4 DNA polymerase klenow
fragment) and subsequently adapters were ligated to the blunt ends Each of the DNA
fragments was amplified to a cluster which was performed in the flow cell by template
hybridization and bridge amplification Millions of clusters were then sequenced by
sequencing-by-synthesis technology leading to base calls The raw data was filtered to remove
low abundant sequences from the assembly according to 15-mer frequencies The low
complexity reads low quality (leQ20) base calls and adapter contaminations (24 bp overlap
between adapter and reads) were eliminated The quality of the sequences was analysed by
GC content and depth correlative analysis (for GC biasness) K-mer analysis was performed
using 15 bp of the short reads for heterologous sequence identification
126
The cleaned short reads (approximately 90 bp) were self-assembled using the Short
Oligonucleotide Analysis Package (SOAP) denovo (httpsoapgenomicsorgcn) (Li et al
2008) The assembly errors were corrected by filling gaps and proofreading single base pairs
using mapping information The assembly produced 441 contigs with an N50 contig size of
29222 bp (the largest contig size 114839 bp) and 80 scaffolds with an N50 scaffold size of
325726 bp (largest scaffold 662452 bp) with an total sequence length of 4878242 bp which
constitutes 33-fold genome coverage The scaffolds were ordered and oriented according to a
reference genome C koseri ATCC BAA-895 the closest completely sequenced species of
the newly sequenced bacterial genome by alignment with the MUMmer program v322
(httpmummersourceforgenet) (Delcher et al 2002) Genome coverage was determined by
mapping the reads to the assembled genome with Bowtie software (Langmead et al 2009) A
preliminary genome annotation was performed to define the origin of replication by
identifying colocalization of multiple genes ie gyrB recF dnaN and dnaA (usually found
near the origin of replication) and comparison with the C koseri reference genome Synteny
analysis between the reference bacterium and assembled scaffolds was performed with the
MUMmer program
532 Gene function annotation and classification
The genome of ADS47 was annotated by Rapid Annotation using Subsystems
Technology v40 (RAST) a server-based genome analysis system (httprastnmpdrorg)
(Aziz et al 2008) A circular genome map was constructed using CGView program
(httpstothardafnsualbertacacgview_server) (Stothard and Wishart 2005)
127
533 Molecular phylogenetic analysis
A robust phylogenetic tree was produced using Composition Vector Tree (CVTree)
web-based program v20 (httpcvtreecbipkueducn) (Qi and Hao 2004) by comparing
whole genome amino acid sequences of strain ADS47 two completely sequenced Citrobacter
genomes [C rodentium ICC168 (NC_013716) and C koseri ATCC BAA-8959 NC_009792)]
and three partially sequenced Citrobacter genomes [C freundii strain Ballerup 785139
(CACD00000000) Citrobacter sp 30-2 (NZ_ACDJ00000000) and C youngae ATCC 29220
(NZ_ABWL00000000)] E coli O157H7 strain EDL933 (NC_002655) was used as an
outgroup species The tree was constructed from the generated pairwise distance matrix data
using Neighbor-Joining Program in PHYLIP v369 package (Saitou and Nei 1987 Felsenstein
1989)
534 Comparative genomic analysis
Genome-wide comparative analyses between C freundii ADS47 C freundii strain
Ballerup 785139 Citrobacter sp 30-2 C youngae ATCC 29220 C Koseri ATCC BAA-895
and C rodentium ICC168 was carried out The nucleotide sequence idendities were
determined by pairwise alignments of the ADS47 genome with the five other Citrobacter
genomes using MUMmer software (Delcher et al 2002) and GC content total CDSs genome
coverage by CDSs and number of stable RNAs were determined by RAST program
Orthologous CDS groups among the total set of 29079 CDSs derived from the six
Citrobacter genomes were determined using OrthoMCL software v202
(httporthomclsourceforgenet) based on all-against-all reciprocal BLASTP alignment of
amino acid sequences (Li et al 2003) Genes unique to each strain were identified as those
128
CDSs not included in any orthologous group of the five other Citrobacter genomes BLASTP
comparisons were visualized with Blast Ring Image Generator (BRIG) software v095
(httpsourceforgenetprojectsbrig) (Alikhan et al 2011)
Protein sequences of the five Citrobacter genomes pooled from the GenBank data
bases were compared with ADS47 A circular map was developed by BRIG software that
showed the unique genomic regionsgenes of ADS47 The unique genes were grouped
according to their functional similarities previously identified by RAST software and
genomic regions possibly acquired by horizontal transfer were identified using RAST
software
129
54 Results
541 Assembly and organization of the ADS47 bacterial genome
Scaffolds derived from Citrobacter freundii strain ADS47 were ordered and oriented
by alignment to Citrobacter koseri ATCC BAA-895 the most closely related genome that has
been completely sequenced The genomes of ADS47 and C koseri displayed close synteny
(Figure 5-1)
Figure 5-1 Co-linearity between C freundii strain ADS47 genome and C koseri Scaffolds
derived from ADS47 genome sequences (Y- axis) were aligned and assembled by comparison
with the closest completely sequenced genome C koseri ATCC BAA-875 (X- axis)
130
542 An overview of the ADS47 genome
General features of the C freundii strain ADS47 are presented in Table 5-1 Some
notable features are that strain ADS47 contains 4881122 bp circular chromosome with an
average GC content of 5202 The genome contains 4610 CDSs which comprise 8784 of
the whole genome Annotation by RAST assigned 2821 (62) CDSs to known biological
functions and remaining 1789 (38) genes were classified to unknown or hypothetical
proteins
The GC content in the coding regions is significantly higher (5318) than in the non-
coding intergenic region (4379) which covers 1215 (592781 bp) of the genome The
intergenic region is comprised of 018 repeat sequences (total 8999 bp) 5 (009)
transposons (4806 bp) 025 stable RNA (12091 bp) and the remaining ~115 is
classified as regulatory and other functions The ADS47 genome contains 62 tRNAs 3
rRNAs and 34 copies of sRNAs The average lengths of the tRNAs rRNAs and sRNAs are
785 885 and 13441 bp respectively There are also 42 phageprophage-related genes
identified in the genome
Figure 5-2 shows the distribution of the 26 categorized CDSs rRNA tRNA GC
content and GC skew It appears that ADS47 genome contains equal quantity of genes in
leading and lagging strands and showed occasional clustering (likely operons) of genes in the
two strands The genome contains three rRNAs (two large subunit and a small subunit) and 62
tRNAs which are distributed across the genome A consistent GC content was observed in
the ADS47 genome however 7-10 low GC content (high black color peaks) genomic regions
were found at locations origin of replication 600 2000 3000 3300 3550 3850 4300 and
4800 kbp This is indicative of possible horizontally transferred genes or insertions in those
131
genomic locations GC skew denotes the excess of C over G in certain genomic locations
Positive GC skew (+) (green color) indicates the skew value in the leading strand while
negative skew (-) (purple color) in the lagging strand As in many bacterial genomes the
ADS47 genome map shows the separation of positive and negative GC skew at the origin of
replication (base pair position 1) The GC skew in the leading strand is highly skewed (high
green color peak) indicating that possible horizontal gene transfer occurred into the ADS47
genome
The CDSs ranged from ~100 bp to ~2000 bp and have an average size of 893 bp
(Figure 5-3) The highest numbers of CDS (770) were 600-800 bp long A considerable
number of genes (220) were gt2000 bp in size Some of the biological functions for the
smallest genes (100-200 bp) are sulfur carrier protein methionine sulfoxide reductase
ribonuclease P protein component while the largest genes (gt2000 bp) are ferrous iron
transport protein B translation elongation factor G pyruvate flavodoxin oxidoreductase and
tetrathionate reductase
132
Table 5-1 General features of the C freundii strain ADS47 genome
Feature Chromosome
Genome size 4881122 bp
Overall GC content 5202
Number of genes 4610
Genes with assigned function (number and ) 2821 (62 )
Genes without assigned function (number and ) 1789 (38 )
Total length of protein coding sequences 4285461 bp
Genome coverage by coding sequences () 8784
Average length of coding sequences 893 bp
GC content in the coding region 5318
GC content in the non-coding region 4379
Length and percentage of intergenic region 592781 bp and 1215
Total repeat sequences and genome coverage 8999 bp and 018
Total transposons sequences and genome coverage 4806 and 009
Number average length and genome coverage of tRNA 62 copies 785 bp and 0099
Number average length and genome coverage of rRNA 3 copies 885 bp and 0067
Number average length and genome coverage of sRNA 34 copies 13441 bp and 0093
133
134
Figure 5-2 Circular genome map of C freundii strain ADS47 Map was created with the
CGView software From outside to inside the first circle indicates the genomic position in kb
Different functional categories of predicted coding sequences (CDS) are distinguished by
colours with clockwise transcribed CDSs in the second circle and anticlockwise transcribed
CDSs in the third circle Bars on the fourth and fifth circles indicate the rRNAs and tRNAs
respectively Sixth and seventh circles show the coverage and GC content respectively The
innermost circle demonstrates the GC skew + (green) and GC skew- (purple)
135
Figure 5-3 Size distribution of the predicted genes identified in C freundii strain ADS47
The X axis identifies the gene size categories (bp) and Y axis the abundance of genes
136
543 Annotation and functional classification of ADS47 genome
RAST analysis identified predicted CDSs in the genome of Citrobacter strain ADS47
assigned functions and grouped the annotated genes into 26 functional subsystem categories
out of 27 No CDS was found belonging to the photosynthesis category A graphical
presentation of the distribution of the CDSs into each of the 26 categories is given in
Figure 5-4
A large portion (38) of the CDSs belong to hypothetical or unknown functions
Among the 62 known or predicted proteins the highest numbers (835) belong to the
carbohydrate category followed by amino acids and derivatives (468) and
cofactorsvitaminsprosthetic groups (324) A noticeable number of stress response (236) and
respiration (198) categories genes were identified in the bacterial genome The ADS47
genome encoded by 171 membrane transport genes 113 virulencedefense genes and 126
genes involved in regulationcell signaling categories Two hundred and one CDSs were
grouped into the miscellaneous category whose functions are different than the indicated
RAST categories The RAST annotations further divided the categorized CDSs into
subcategories based on similar functional roles which is discussed in section 545
137
Figure 5-4 Functional classification of the predicted coding sequences (CDS) of C freundii
strain ADS47 The annotation was performed by Rapid Annotation using Subsystems
Technology (RAST) program
138
544 Molecular phylogenetic relationships
Genome-wide phylogenetic analysis based on comparisons of whole genome amino
acid sequences of two completely sequenced Citrobacter genomes (C koseri and C
rodentium) and three partially sequenced Citrobacter genome sequences (C freundii
Citrobacter sp and C youngae) illustrates the relationships of ADS47 to five Citrobacter
species (Figure 5-5)
C youngae ATCC 29220 originating from human feces and two pathogenic species
(C koseri ATCC BAA-895 and C rodentium strain ICC168) are progressively more distantly
related to ADS47 The present analysis is consistent with the 16S-rRNA sequence based
phylogenetic analysis (Chapter 3) which showed that the closest bacterial strain to ADS47 is
C freundii (Figure 3-2) This confirmed the identification of strain ADS47 as the gamma
proteobacterial species C freundii made in Chapter 3
139
Figure 5-5 Phylogeny of the trichothecene detoxifying C freundii strain ADS47 The
dendrogram was constructed by comparing the ADS47 genome (amino acid sequences) with
five Citrobacter genomes and a E coli strain (out group species) using the CVTree software
The calculated pairwise distances matrix by Protdist program was transformed into a tree by
neighbor-joining program The dissimilarity tree was drawn to scale with branch lengths
(scale bar 002 indicated the amino acid substitutions among the genomes) The C koseri C
rodentium C freundii Citrobacter sp C youngae and E coli protein sequences were
uploaded from GenBank (Benson et al 2009)
140
545 Comparative genomics
Genome-wide comparative analyses between the genome of strain ADS47 and the five
previously sequenced Citrobacter genomes showed that 872 of the total nucleotides
between ADS47 and C freundii Ballerup were identical The values for the comparisons
between C freundii strain ADS47 were 8212 for Citrobacter sp 30-2 7637 for C
youngae 5981 for C koseri and 4213 for C rodentium (Table 5-2)
The two human and mouse pathogenic species namely C rodentium and C koseri
and the bacteria originally isolated from human feces (C youngae) have higher GC contents
(546 and 532 respectively) compared to C freundii strain ADS47 C freundii strain
Ballerup and Citrobacter sp 30_2 which have slightly lower GC contents (~52) The two
pathogenic species contain 22 copies of rRNA which is significantly higher than the 9 found
in C freundii strain Ballerup 6 in Citrobacter sp 30_2 and 3 in C freundii strain ADS47 but
less than the 25 copies in C youngae The genome of C freundii strain ADS47 C freundii
strain Ballerup C youngae and Citrobacter sp 30-2 had a similar percentage of the genome
comprising coding sequences (865-88) C koseri and C rodentium genomes contained the
highest (90) and lowest (84) genome coverage by coding sequences respectively
141
Table 5-2 Genome comparisons between strain ADS47 and five Citrobacter species The
base nucleotide sequence identities were performed using MUMmer DNAdff script program
(Delcher et al 2002) The features (GC content total CDS percentage CDS and number of
ribosomal RNA) were determined using RAST program (Aziz et al 2008)
Genome feature Bacterial species
C
freundii
ADS47
C freundii
Ballerup
Citrobacter
sp 30_2
C
youngae
C koseri C
rodentium
Genome length
(bp)
4878242
4904641
5125150
5154159
4720462
5346659
Base similarity to
ADS47 ()
NA 8715 8212 7637 5981 4213
GC () 5202 5218 516 5265 530 546
Number of coding
sequences
4610 4685 4728 5278 4978 4800
Genome coverage
with coding
sequences ()
880 865 866 872 900 840
rRNA copies 3 9 6 25 22 22
Total stable RNAs 99 81 78 105 120 142
142
The number of orthologs shared between ADS47 and other Citrobacter species is
illustrated in Figure 5-6 ADS47 shares the smallest number of orthologous CDSs with C
rodentium and C koseri 3250 and 3341 respectively The highest number of shared CDS
were found with C freundii Ballerup (3706) followed by C youngae (3693) and Citrobacter
sp 30_2 (3676) A total of 2821 CDSs were found to be common to all of the six Citrobacter
genomes analysed (data not shown) Nonetheless significant diversity exists between C
freundii ADS47 and the other species as a considerable number of unshared genes were
identified from the comparisons with C freundii Ballerup [979 (20)] Citrobacter sp 30_2
[1064 (23)] C youngae [1654 (31)] C koseri [1693 (34)] and C rodentium [1634
(34)]
143
Figure 5-6 Comparisons of coding sequences between strain C freundii ADS47 and five
Citrobacter species Coding sequences of ADS47 were compared with C freundii Ballerup
Citrobacter sp 30_2 C youngae C koseri and C rodentium genomes based on all-versus-all
reciprocal FASTA comparison of at least 30 amino acid identity and gt70 coverage of the
total length using OrthoMCL program A Venn diagram demonstrates the shared and
unshared (indicated in the brackets) coding sequences
144
Table 5-3 shows differences in the 26 RAST categorized genes between the six
Citrobacter genomes Even though a low abundance of virulencediseasedefense category
genes were found in the animalhuman pathogenic bacteria of C rodentium (85) and C koseri
(92) significantly higher mycobacterium virulence genes (13) were identified in this two
species The genomes of the two closely related strains C freundii ADS47 and C freundii
Ballerup lack this type of virulence gene Instead elevated numbers of genes for resistance to
antibiotic and toxic compounds were found in C freundii ADS47 (94) C freundii Ballerup
(88) and Citrobacter sp (110) compared to other three species Copper homeostasis and
cobalt-zinc-cadmium resistance genes were identified in strain C freundii ADS47 (13 and 21
respectively) and closely related C freundii Ballerup (12 and 22) and Citobacter sp (18 and
22) Significantly lower numbers of these genes were present in C rodentium (6 and 4) and C
koseri (7 and 8) C rodentium and C koseri lack mercuric reductase mercuric resistance and
arsenic resistance genes perhaps because being animalhuman pathogens but these genes
were found in the remaining four genomes The C koseri genome is richer in betandashlactamase
genes (5) compared to the remaining genomes which encodes 1-3 genes
145
Table 5-3 Functional categorized gene abundances among the genomes of C freundii
ADS47 C freundii Ballerup Citrobacter sp 30_2 C youngae C koseri and C rodentium
The genomes were classified into different functional categories with the RAST program
which classified the total CDSs
RAST subsystem category Citrobacter species
C
freundii
ADS47
C
freundii
Ballerup
C
sp
C
youngae
C
koseri
C
rodentium
Cofactorsvitaminsprosthetic
groupspigments
323
317
310
311
325
304
Cell wall and capsule 279 272 248 262 247 262
Virulence disease and defense 113 107 134 100 92 85
Potassium metabolism 50 53 33 32 30 28
Miscellaneous 210 228 200 185 245 185
Phageprophagetransposable
element plasmid
24 75 25 4 6 49
Membrane transport 170 156 171 195 171 172
Iron acquisition and metabolism 80 81 86 62 113 27
RNA metabolism 220 222 217 215 238 212
Nucleosides and nucleotides 119 112 110 111 120 114
Protein metabolism 249 280 269 298 287 254
Cell division and cell cycle 30 29 32 29 38 29
Motility and chemotaxis 133 124 90 147 143 133
Regulation and cell signaling 126 130 124 135 132 123
Secondary metabolism 21 22 20 21 0 0
DNA metabolism 145 167 165 155 147 152
Fatty acids lipids and isoprenoids 121 149 122 121 134 133
Nitrogen metabolism 50 52 53 49 56 58
Dormancy and sporulation 3 3 3 4 3 3
146
Table 5-3 (continued)
RAST subsystem category
Citrobacter species
C
freundii
ADS47
C
freundii
Ballerup
C
sp
C
youngae
C
koseri
C
rodentium
Respiration 198 184 180 191 166 162
Stress response 236 235 170 177 179 174
Metabolism of aromatic
compounds
20 20 28 25 17 48
Amino acids and derivatives 482 476 450 466 428 416
Sulfur metabolism 48 50 52 57 56 56
Phosphorus metabolism 50 49 43 42 43 28
Carbohydrates 837 803 708 731 733 737
147
Strains C freundii ASD47 and C freundii Ballerup have much more potassium
metabolism category genes (50 and 53 respectively) than the other bacterial species which
code for 28-33 proteins Larger numbers of genes associated with the glutathione regulated
potassium efflux system were also identified in C freundii ASD47 with 23 and C freundii
Ballerup with 26 compared to the remaining four species that encode only five genes All of
the six genomes contained phageprophage-related genes with the maximum identified in C
freundii Ballerup (75) followed by C rodentium (49) Citrobacter sp (25) and C freundii
ADS47 (24) Lower numbers of phageprophage-related genes were found in C koseri (6) and
C youngae (4)
The six Citrobacter genomes showed remarkable variability in their secretion systems
The type-III secretion operon genes are only found in C rodentium and C freundii Ballerup
Type-II secretion genes are predominant in C rodentium (12) followed by ADS47 (9)
Type-IV secretion system is uncommon in Citrobacter species since it was only
identified in Citrobacter sp where it is represented by 19 genes Variable numbers of type-VI
secretion system genes are present in the five genomes except C koseri which lacks such
genes C youngae has the highest number (43) of type-VI secretion proteins The other strains
have a range of type-VI secretion proteins in particular 27 were identified in C freundii
Ballerup 15 in C freundii ADS47 and 8 in Citrobacter sp Type-VII (chaperoneusher)
secretion system genes were found in all of the six Citrobacter species with the largest (42)
and smallest (13) numbers of these genes occurring in C koseri and C freundii Ballerup
respectively
Although a relatively similar number of ABC transporters (37-42) were identified in
the six genomes C freundii ADS47 contain the largest number of tripartite ATP-independent
148
periplasmic (TRAP) transporter genes (7) By contrast the closely related C freundii
Ballerup lacks such transporter system An identical number of iron acquisition and
metabolism category genes were found in C freundii ADS47 (80) and C freundii Ballerup
(81) while the two pathogenic species C rodentium and C koseri code for the lowest (27)
and highest (113) numbers of such genes C koseri is rich in siderophore related genes (64)
but the remaining five genomes have smaller numbers of siderophore-associated genes (20-
22)
Variable numbers of motility and chemotaxis related genes were found in the
Citrobacter genomes with the largest number occurring in C youngae (147) followed by C
koseri (143) C freundii ADS47 (133) C rodentium (133) C freundii Ballerup (124) and
Citrobacter sp (90) The two pathogenic species (C rodentium and C koseri) encode a
remarkably low number of quorum sensing genes (2) In contrast C youngae codes for the
highest number of quorum sensing genes (19) A common and distinctive feature of the two
pathogenic species is the lack of genes required for secondary metabolism In contrast the
non-pathogenic species contain a common set of genes for degrading a range of
phenylpropanoids and cinnamic acid (20-22)
The lowest number of genes related to phospholipidsfatty acids biosynthesis and
metabolism were found in C freundii ADS47 (66) compared to C freundii Balllerup (94) and
C koseri (98) The smallest numbers of nitratenitrite ammonification genes were found in
ADS47 (31) and its closest genome C freundii Ballerup (32) In contrast the two pathogens
encode the highest number of such proteins (39-40)
C rodentium and C koseri code for significantly low numbers of respiratory category
genes (162-165) while C freundii ADS47 contains the highest number of such genes (198)
149
Further analysis of the subcategory genes determined that the two gut species (C youngae and
Citrobacter sp 30_2) contain the largest numbers of genes encoding anaerobic reductases
(22) and identical numbers of formate dehydrogenases (17) The largest numbers of formate
dehydrogenase genes were identified in C freundii Ballerup (32) and C freundii ADS47 (30)
but the two pathogenic species differed from the other species by encoding the lowest number
(13) of c-type cytochrome biogenesis genes
A higher number of aromatic compound metabolism category genes were found in the
mouse pathogen C rodentium (48) compared to the human pathogen (C koseri) which
contain 17 genes A significantly higher number of genes involved in central aromatic
intermediate metabolism (26) were found in C rodentium compared to the mouse pathogen
(C koseri) and other genomes which only contain 6-8 such genes The two C freundii strains
ADS47 and Ballerup lacked the anaerobic degradation of aromatic compound
(hydroxyaromatic decarboxylase family) genes
In spite of the abundance of stress response category genes strain ADS47 has only a
small number of desiccation stress genes (5) compared to the other species which ranged
from 9 to10 However ADS47 and Ballerup code for higher number of genes related to
oxidative stress (94 and 91 respectively) and cold shock (8 and 7 respectively) than the other
species
The smallest numbers of amino acids and its derivatives category genes were found in
the two pathogens C rodentium and C koseri (416 and 428 respectively) while the largest
number of these genes were identified in strain ADS47 (482) and C freundii Ballerup (476)
Despite the similar number of sulfur metabolism category genes determined across the six
species only ADS47 lacks taurine utilization genes Strains ADS47 and Ballerup were found
150
to be rich in the phosphorous metabolism gene category (49-50) while C rodentium contains
a significantly decreased number of these category genes (28)
The bioinformatics analyses found the largest number of carbohydrate category genes
in ADS47 (837) followed by Ballerup (803) The gut bacteria (Citrobacter sp 30_2) codes for
the least number of genes assigned to the carbohydrate category (708) while C koseri C
rodentium and C youngae contained 731-737 in this category genes The analyses also
determined that there are relatively small numbers of TCA and Entner Doudoroff pathway
genes in C rodentium and C youngae compared to the other four species and relatively large
numbers of fermentation-related genes in ADS47 (81) and C freundii (85) compared to the
other four species
A comparison of the pathogenicity operons [locus of enterocyte effacement (LEE)] of
C rodentium with other five Citrobacter genomes revealed the absence of this virulence
determinant gene region in the other four Citrobacter species including ADS47 (Figure 5-7)
The analysis showed the presence of LEE operon genes in C freundii strain Ballerup as no
gap was observed in that genomic location while C freundii ADS47 C koseri C youngae
and Citrobacter sp 30_2 lacked that loci (7a) Further ananlysis showed alignments between
C rodentium and E coli LEE operon genes (Figure 7b) In contrast no alignment of such
genes was observed in ADS47 genome
151
(a)
152
Figure 5-7 (a) Blast Ring Image Generator (BRIG) output based of Citrobacter rodentium
(NC_013716) compared against ADS47 C freundii C koseri C youngae E coli and
Citrobacter sp genomes The targeted comparative genome analysis was performed based on
locus of enterocyte effacement (LEE) operons of C rodentium (Petty et al 2011) The
innermost rings show GC content (black) and GC skew (purple and green) The remaining
rings show BLAST comparisons of the other Citrobacter and E coli genomes against C
rodentium The region shown in red is LEE operon genes (b) The absence of LEE operons in
the ADS47 genome comparing to C rodentium and E coli Analysis was performed and
visualized by Artemis Comparison Tool software v132
(httpwwwsangeracukresourcessoftwareact) (Tatusov et al 2000 Craver et al 2005)
LEE operons
(b)
153
546 Analysis of the unique genes in C freundii ADS47
Two hundred and seventy genes unique to ADS47 were identified from a comparison
of its genome to the other five Citrobacter species In general the unique genes are
distributed throughout the ADS47 genome but some clusters of unique genes occurred
including eight unique genomic regions (Figure 5-8) Six large unique regions are located at
767204-775090 (~8 kb) 1025572-1042510 (17 kb) 2798289-2807217 (~9 kb)
3045220-3055912 (~11 kb) 3930366-3943075 (12 kb) and 4022195-4032444 (10 kb)
Out of 270 unique genes 147 (54 ) are hypothetical proteins The remaining 123
(46) genes are grouped into reductasesoxidoreductases (9 genes) de-acetylase (1)
hydrolases (2) transferases (4) sulfatasesarylsulfatases (3) bacteriocins (3) type VI
secretion proteins (2) kinases (4) transporters (11) membraneexported proteins (8)
regulators (12) integraserecombinases (5) phagesprophagestransposaes related genes (27)
and miscellaneous (32) that includes DNA helicases ligases fimbrial proteins and
chromosome partitioning genes (Table 5-4)
The unique regions encode various functional genes that may cause trichothecene de-
epoxidation or de-acetylation reactions The 8 kb unique regions contains six genes encoding
glycosyltransferases and two copies of oxidoreductase-like coding sequences (CMD_0711
and CMD_0712) that showed 39 identity to the Marinobacter aquaeolei F420
hydrogenasedehydrogenase beta subunit The largest (17 kb) unique genomic region codes
for 16 genes including dehydrogenase hydrolase bacteriocin (pyocin) integrase and a short
chain dehydrogenasereductase-like (SDR) (CMD_0979) gene (Figure 5-9a)
154
The SDR like gene is 49 amino acid similarity to an oxidoreductase of the
phytopathogenic Dickeya dadantii and the enzyme contains a domain with 77 homology to
the Dickeya zeae NAD(P) binding site The 9 kb unique genomic section contains 8 genes
including a NaCl symporter a dehydratase (CMD_2679) with 86 homology to
Enterobacteriaceae bacterium dihydroxy-acid dehydratase and a putative oxidoreductase
(CMD_2683) having 37 similarity with oxopropionate reductase of Bacillus sp The unique
region also encodes a short chain dehydrogenasereductase (SDR) (CMD_2684) that shows
85 identity to Enterobacteriaceae bacterium (Figure 5-9b) A unique thiol-disulfide and
thioredoxin like reductase (CMD_3721) was identified in a 10 kb unique region that contains
13 genes (Figure 5-9c) This gene is 59 identical to Oligotropha carboxidovorans thiol-
disulfide isomerase and thioredoxin In addition several reductasesoxidoreductase-like
unique genes were identified in the ADS47 genome including an oxidoreductase
(CMD_1024) with 27 identity to Canadidatus nitrospira NADH-quinone oxidoreductase
membrane L Another oxidoreductase-like gene identified in a different location of the
ADS47 genome (CMD_1393) shows 87 homology to Klebsiella pneumoniae
deoxygluconate dehydrogenase contains a SDR active site A putative nitrate reductase gene
(CMD_3184) has a low level of identity (27) with an uncultured bacterial respiratory nitrate
reductase The ~11 kb unique region encodes sulfatasesarylsulfatses
(CMD_2898CMD_2905) and related genes Furthermore ADS47 contains a unique
glycosylphosphatidylinositol (GPI) anchor biosynthesis operon comprised of four genes
including a N-acetylglucosaminyl-phosphatidylinositol de-acetylase (CMD_2431) which
showed 78 similarity to E coli de-acetylase (Figure 5-9e)
155
Other unique genespredicted genes in the ADS47 genome include
polysaccharidephenazine biosynthesis genes (CMD_0710CMD_1406) heavy metal kinase
(CMD_0989) two component heavy metal regulator (CMD_0990) nitrate reductase regulator
(CMD_3477) putative potassiumzinc transporters (CMD_0973CMD_4600) sodium
symporter (CMD_2678) (tripartite ATP-independent periplasmic transporters) TRAP
transporters (CMD_3693 CMD_3694 CMD_3695 CMD_4337) glutamineribose ABC
transporters (CMD_3176CMD_3544) toxin transcription regulator (CMD_3464) fimbrial
proteinchaperone (CMD_1695 CMD_2434 CMD_3277 CMD_3278) and ParB
chromosomeplasmid partitioning proteins (CMD_4033CMD_4034)
Majority of the unique genes and their distributions are shown in comparative circular
genome maps (Figure 5-8)
156
Table 5-4 Unique genes of C freundii strain ADS47 Genes were categorized according to
the functional similarity The unique genes were determined by comparing ADS47 with five
sequenced Citrobacter species using OrthoMCL software
Unique gene classes Number
Total unique genes
270
Reductases oxidoreductases 9
De-acetylase 1
Hydrolase 2
Transferase 4
Sulfatase 3
Bacteriocin 2
Type VI secretion 2
Kinase 4
Transporter 9
Membraneexported protein 8
Regulator 12
Integraserecombinase 5
Phageprophagetransposae 27
Miscellaneous 34
Hypothetical protein 147
157
158
Figure 5-8 Locations of the unique genes of C freundii strain ADS47 The unique genes
were determined by comparing the total CDS of strain C freundii ADS47 with C freundii
Ballerup Citrobacter sp C youngae C koseri and C rodentium CDSs by OrthoMCL
program The map was developed by Blast Ring Generator (BRIG) software First second
and third circles from inside to out side show the genome size GC and GC skew of strain
ADS47 Forward (Fwd) and reversed (Rev) transcribed CDSs of ADS47 are shown in green
(forth circle) and red (fifth circle) respectively The next five rings represent the circular
genome maps of five Citrobacter species distinguished by different ring colours (C
rodentium as sky blue C koseri as dark blue C youngae as light blue C freundii Ballerup as
yellow and Citrobacter sp as green) The degree of orthologous gene identity between
ADS47 and other five genomes are shown by color intensity (light color= 75 gene identity
ash color= 50 gene identity) Gaps across five genome rings indicate the absence of
genesregions compared to ADS47 Unique genes are shown surrounding the map rings and
the identities of some of the genes are given Arrows indicate putative
reductasesoxidoreductases and de-acetylase
159
(a)
(b)
160
(c)
(d)
161
Figure 5-9 Unique genomic regions and associated genes in C freundii ADS47 compared to
five Citrobacter species RAST server based analysis compared the unique genesregions of
ADS47 with the publicly available databases The upper panels demonstrate the large unique
regions defined by yellow boxes and genes located in the genomic regions are indicated by
arrows The lower panels are closely related genesoperons found in distantly related species
The unique regions that contain potential trichothecene de-epoxidation genes are shown in (a)
short chain dehydrogenasereductase (SDR) and dehydrogenase (b) putative oxidoreductase
dehydratase and SDR (c) thiol-disulfide isomerase and thioredoxin oxidoreductase and (d)
unique glycosylphosphatidylinositol (GPI) biosynthesis operon having N-acetylglucosaminyl-
phosphatidylinositol de-acetylase gene
162
55 Discussion
The de novo sequenced ADS47 genome was oriented and assembled using a
completely sequenced C koseri genome as reference strain The synteny analysis between
ADS47 and C koseri showed very similar overall organization of the two genomes The high
preservation of genes among five Citrobacter species and ADS47 were emphasized by the
close alignments observed by Blast Ring Image Generator (BRIG) outputs obtained from
these genomes
Genome-wide phylogenetic analyses based on a large number of common CDSs
(2821) observed within the six Citrobacter genomes confirmed the species identification
based on 16S-rDNA sequencing of the trichothecene mycotoxin-detoxifying C freundii strain
ADS47 The base sequence idendity was 8715 between these two genomes They shared
~80 CDS and had a similar GC content (52) Citrobacter are Gram-negative rod shaped
and facultatively anaerobic bacteria belonging to the family of Enterobacteriaceae (Wang et
al 2000 Whalen et al 2007) Strains ADS47 and Ballerup of C freundii are both aerobic
gram-negative bacilli that have numerous flagella and inhabit in soil (Cong et al 2011 Islam
et al 2012) C freundii is considered to have a positive environmental role by reducing nitrate
to nitrite (Rehr and Klemme 1989) which is an important step in the nitrogen cycle
C freundii C rodentium and C koseri are animalhuman pathogenic species C
rodentium is a natural mice pathogen causing hemorrhagic colitis and hemolytic uremic
syndrome (Gareau et al 2010) C koseri causes human disease (Badger et al 1999 Kariholu
et al 2009) particularly neonatal meningitis and C freundii is an opportunistic human
pathogen responsible for a various types of infections like respiratory tract urinary tract and
blood (Whalen et al 2007) C freundii is very common representing approximately 29 of
163
all opportunistic human infections (Hodges et al 1978) Outside of animals they can be found
in soil water sewages and different organs of diseased animals (Wang et al 2000) These
pathogens code for large numbers of mycobacterium virulence operonsgenes which were
absent in the three other genomes However they have fewer genes involved in environmental
stress such as copper homeostasis genes and no mercuricarsenic resistance and secondary
metabolism category genes unlike the other Citrobacter genomes Nonetheless considerable
variability exists between the two pathogens correlates with C rodentium causing disease in
mice C koseri causing meningistis in human and C freundii causing infections in different
human organsThere were attaching and effacing (AE) lesion forming (LEE) operons in C
rodentium and C freundii but not in C koseri Those genes have shown to be acquired by
lateral gene transfer playing major roles in virulence of enteropathogenic and
enterohemorrhagic E coli and C rodentium (Mundy et al 2005 Petty et al 2010 Petty et al
2011 The ~356 kb pathogenic LEE gene island is comprised of five operons (LEE1 to
LEE5) that code for ~40 virulence genes that include type-III secretion genes and associated
effectors (Berdichevsky et al 2005) In addition an elevated number of type-II and -VI
secretion proteins identified in C rodentium which may act as additional virulence factors
(Petty et al 2010) Because it lacks type-III secretion apparatus genes it is likely that C
koseri uses a different strategy to cause human neonatal meningitis (Badger et al 1999 Ong
et al 2010)
To adapt to different environmental niches or hosts bacteria like Citrobacter species
frequently gainlose genes increase repeat sequences and recombine genomic regions using
different mechanisms (Parkhill et al 2001 Petty et al 2011) The existence of large numbers
of phageprophagetransposae related genes in the Citrobacter genomes particularly in C
164
freundii Ballerup suggests that their genomes are unstable For example C freundii Ballerup
and C koseri might have acquired LEE operons by horizontal transfer via phage mediated
transpositions Interestingly although ADS47 is closely related to C freundii Ballerup it
lacks the pathogen-related LEE gene island and mycobacterium virulence operons required
for invasion and intracellular survival in macrophages (Gao et al 2006) Instead ADS47 has a
larger number and variety of stress-responsive genes such as efflux genes and bacteriocins
(pyocin hemolysin) genes that provide protection against heavy metals and antibiotics and
increase their competitiveness against other bacteria These findings suggest that the strain
ADS47 is not an opportunistic animal pathogen but has evolved to be highly competitive in
the soil micro-environment (Arnold 1998 Bengoechea and Skurnik 2000) Perhaps ADS47
evolved trichothecene degrading genes for adaptation to soils containing trichothecene
mycotoxins contaminated soils (Gadd and Griffiths 1978 Sharma and Fulekar 2009 Han et
al 2011) Conversely the absence or reduced number of stress-responsive genes in C
rodentium and C koseri may imply that those two pathogenic species are not exposed to such
stressful and xenobiotic-contaminated environments as ADS47
This interpretation is supported by the observation that the two of the three
animalhuman pathogenic species contained high of rRNA copy number but lower numbers
were found in ADS47 C freundii Ballerup and Citrobacter sp 30_2 Bacteria maintain large
number of rRNA for higher production of ribosome during high growth rates (Stevenson et al
1997 Klappenbach et al 2000) such as might occur during the establishment of an infection
However a large copy number of rRNAs may not be metabolically favourable for bacteria
such as ADS47 that evolved in a highly competitive environment as that costs higher cellular
energy (Hsu et al 1994)
165
C freundii Ballerup seems to be an exception in that it is similar to the other animal
pathogens for containing virulent factors (ie LEE-operon genes) but more like the
environmental species for encoding high number genes for stress defence heavy metal and
toxic compounds resistances
The comparison of the genes in C freundii ADS47 with the unique ability to detoxify
trichothecenes with genes in other Citrobacter species allowed approximately 270 unique
genes to be identified in ADS47 Some of those genes may be involved in trichothecene de-
epoxidation or de-acetylation Currently the pathways and microbial enzymes leading to
trichothecene de-epoxidation and de-acetylation are unknown but based on the required
reaction ie the elimination of an oxygen atom from epoxide moiety it is likely that epoxide
reductases or epoxide oxidoreductases are responsible for such biodegradation activity (He et
al 1992)
From database searches three enzymes having epoxide reductase activity were found
These are a vitamin K epoxide reductase (VKOR) a monooxygenase PaaABCE and pyridine
nucleotide-disulfide oxidoreductase (PNDO) which reduce vitamin K epoxide phenylacetyl-
CoA and epoxyalkanes respectively (Swaving et al 1996 Li et al 2004 Teufel et al 2012)
In mammals VKOR plays important roles in vitamin K cycle by reducing vitamin K epoxide
to vitamin K and its homologues that are found in plants bacteria and archaea (Goodstadt
2004 Li et al 2010) The microbial multi-component PaaABCE monooxygenase catalyzes
degradation of variety of aromatic compounds or environmental contaminants via epoxidation
of phenylacetic acid (Paa) and NADPH-dependant de-epoxidation of epoxy-Paa (Luengo et
al 2001 Teufel et al 2012) PNDO is a unique type of NADP-dependent oxidoreductase
identified in Xanthobacter strain Py2 that was obtained by propane enrichment procedure
166
(van Ginkel and de Bont 1986) However no orthologs of these genes were found in ADS47
genome
Nine ADS47-unique reductaseoxidoreductase-like genes are potential candidates for
the trichothecene de-epoxidation enzyme Genes belong to short chain
dehydrogenasereductase (SDR) family are structurally and functionally diverse and cause
different enzymatic activities including xenobiotic degradation (Kavanagh et al 2008) This is
a promising candidate for the trichothecene de-epoxidation gene Genes belong to dehydratase
family catalyze reduction reactions by cleaving carbon-oxygen bonds of organic compounds
that result in removal of oxygen in various biosynthetic pathways (Flint et al 1993) This
mechanism may be relevant to microbial trichothecene de-epoxidation reaction since the latter
involves removal of the oxygen atom forming the epoxide in trichothecene mycotoxins
(Young et al 2007)
Another ADS47-unique gene was for a nitrate reductase that may be a candidate de-
epoxidation gene Under stress conditions such an enzyme causes reductive removal of an
oxygen molecule (Morozkina and Zvyagilskaya 2007)
A fourth type of ADS47-unique gene is for a thiol-disulfide isomerasethioredoxin like
(Trx) oxidoreductase The thiol-disulfide isomerase and thioredoxin oxidoreductases contain a
thioredoxin like domain with a CXXC catalytic motif which catalyzes various biological
processes that allow the organism to adapt to environmental stress and develop competence to
sporulate (Scharf et al 1998 Petersohn et al 2001 Smits et al 2005) The membrane-
anchored Trxndashlike domain initiates the vitamin K de-epoxidation reaction by reducing the
active motif (CXXC) of VKOR (Schulman et al 2010)
167
The present study did not find any homologues of VKOR in ADS47 genome but we
identified two copies of structurally similar quinone reductase protein disulfide bond
formation (DsbB) gene (CMD_1044 and CMD_2641) Both VKOR and DsbB contain four
transmembrane segments an active site CXXC motif and cofactor quinone and use a
common electron transfer pathway for reducing substrates (Li et al 2010 Malojcic et al
2008) As VKOR bacterial DsbB requires a Trx-like partner (eg DsbA) for activity Further
experiments are required to examine if DON de-epoxidation occurs by the catalytic activity of
DsbB coupled with a unique thiol-disulfide isomerase and thioredoxin oxidoreductase present
in ADS47 genome
The analysis also identified an ADS47-unique glycosylphosphatidylinositol (GPI)
biosynthetic operon that is comprised of four genes including an N-
acetylglucosaminylphosphatidylinositol de-acetylase The GPI membrane anchor protein
synthesis is initiated by transferring N-acetylglucosamine from uridine diphosphate N-
acetylglucosamine to phosphatidylinositol by N-acetylglucosaminyltransferase an
endoplasmic reticulum (ER) membrane-bound protein complex (Hong and Kinoshita 2009)
In the second step of the pathway N-acetylglucosamine-phosphatidylinositol is de-acetylated
to produce glucosaminyl-phosphatidylinositol by the catalytic activity of a zinc
metalloenzyme N-acetylglucosaminylphosphatidylinositol de-acetylase which is localized on
the cytoplasmic side of the ER (Nakamura et al 1997 Hong and Kinoshita 2009) It is
possible that the identified unique N-acetylglucosaminylphosphatidylinositol de-acetylase
gene of ADS47 may be involved in trichothecene de-acetylation activity
168
Further analysis is needed to determine which of these candidate gene or genes may be
involved in tricthothecene mycotoxin de-epoxidation and de-acetylation activities
Identification of gene responsible for the trichothecene detoxification activities of C freundii
ADS47 opens many potential applications for its use in agri-based industries to contribute to
cropanimal production and food safety (Karlovsky 2011)
56 Acknowledgement
We gratefully acknowledge Agriculture and Agri-Food CanadaBinational
Agricultural Research and Development Fund (AAFCBARD) project for providing the
financial supports of the work
169
Supplemental Table S5-1 List of unique genes of strain ADS47 gene IDs and their
functional roles The unique genes were determined by comparing the ADS47 genome with
five sequenced Citrobacter species (C freundii Citrobacter sp C youngae C koseri and C
rodentium) using OrthoMCL (Li et al 2003)
Unique gene
classes
Number Gene ID and functions
Total unique
genes
270 123 known functions 147 hypothetical proteins
Reductases
oxidoreductases
9 CMD_0711 coenzyme F420 hydrogenasedehydrogenase beta subunit
CMD_0979 Short chain dehydrogenase reductase (SDR) Oxidoreductase
SDR dehydrogenase CMD_1024 NADH-quinone oxidoreductase
membrane L CMD_1393 oxidoreductase SDR putative deoxygluconate
dehydrogenase (NAD+ 3-oxidoreductase) CMD_2679
Dehydratasedihydroxy acid dehydratase CMD_2683 oxidoreductase
oxopropionate reductase CMD_2684 short-chain dehydrogenasereductase
SDR CMD_3184 nitrate reductase alfa subunit CMD_3721 thiol-disulfide
isomerase and thioredoxin oxidoreductase
Deacetylase 1 CMD_2431 N-acetylglucosaminylphosphatidylinositol deacetylase
superfamily
Hydrolase 2 CMD_0991 Dienelactone hydrolase CMD_2895 Putative glycosyl
hydrolase
Transferase 4 CMD_1640 D12 class N6 adenine-specific DNA methyltransferase
CMD_2432 Glycosyltransferase CMD_2433 putative acetyltransferase
CMD_3023 adenine-specific DNA modification methyltransferase
Sulfatase 3 CMD_2898 Possible sulfatase arylsulfatasearylsulfatransferase
CMD_2902 Arylsulfatasesulfatase CMD_2905 Putative
sulfatasearylsulfatase
Bacteriocin 3 CMD_0982 Thermosatble hemolysin superfamily CMD_0993 Pyocin S2
superfamily (bacteriocin that kills other bacterial strain) CMD_3705 S-type
pyocin containing domain protein
Type VI secretion 2 CMD_1802 EvpB family type VI secretion protein CMD_2364 type VI
effector Hcp1
Kinase 4 CMD_0989 Heavy metal Sensor kinase CMD_1794 Polo kinase kinase
(PKK) superfamily CMD_3124 predicted diacylglycerol kinase CMD_4266
putative signal transduction kinase
Transporter 11 CMD_0973 TrkA-N domain-containing protein (K uptake transporter)
CMD_1407 Branched-chain amino acid transport system carrier protein
CMD_2678 Sodium Symporter CMD_3176 glutamine ABC transporter
CMD_3544 Ribose ABC transport system CMD_3693 TRAP-type C4-
dicarboxylate transport system binding protein CMD_3694 TRAP-type C4-
dicarboxylate transport system binding protein small permease component
(predicted N-acetylneuraminate transporter) CMD_3695 TRAP-type C4-
dicarboxylate transport system large permease CMD_4337 23-diketo-L-
gulonate TRAP transporter small permease protein yiaM component
CMD_4338 23-diketo-L-gulonate TRAP transporter large permease protein
yiaN CMD_4600 putative transporter involved in Zinc uptake (YciA
protein)
170
Supplmental Table S5-1 (continued)
Unique gene
classes
Number Gene ID and functions
Membrane
exported protein
8 CMD_0713 putative membrane protein wzy CMD_0983 Putative exported
protein precursor CMD_0986 Putative transmembrane protein
CMD_0987 putative exported protein precursor CMD_1458 putative
exported protein CMD_2903 Putative exported protein CMD_2904
putative secreted 5-nucleotidase CMD_4291 (S) putative integral
membrane export protein
Regulator 12 CMD_0977 probable transcriptional regulator protein CMD_0980 Putative
transcription activator TenA family CMD_0990 Two component heavy
metal response transcription regulator CMD_1766 Leucine responsive
regulatory protein CMD_2677 transcriptional regulator GntR CMD_2909
putative arylsulfatase regulator CMD_3279 Uncharacterized outer
membrane usher protein yqiG precursor CMD_3464 toxin transcription
regulatory protein CMD_3465 EAL regulator protein (signaling)
CMD_3475 putative transcription regulator CMD_3477 Fumarate and
nitrate reduction regulatory protein (for anaerobic adaptation) CMD_3812
putative regulator from phage
Integrase
recombinase
5 CMD_0785 CMD_0992 CMD_0994 CMD_4032 CMD_4424
Phage prophage
transposae related
genes
27 Phages CMD_1608 CMD_1609 CMD_1610 CMD_1611 CMD_1619
CMD_1631 CMD_1913 CMD_1914 CMD_1915 CMD_1917
CMD_1920 CMD_1921 CMD_1936 CMD_3351 CMD_3804
CMD_3805 CMD_3807 CMD_3808 CMD_4581 CMD_4609
CMD_4610 Transposae CMD_0252 CMD_0398 Prophage CMD_1645
CMD_3806 CMD_4425 CMD_4597
Miscellaneous 32 CMD_0095 ATP-dependent DNA helicase Rep CMD_0656 LysA protein
CMD_0710 Polysaccharide biosynthesis family protein CMD_0978 TPR
repeat containing protein CMD_0981 AMP dependant synthatase and
ligase CMD_0985 Cupin superfamily CMD_0987 flavodoxin
CMD_1404 Putative endoribonuclease CMD_1405 Putative PAC domain
protein CMD_1406 Phenazine (PhzF) biosynthesis protein CMD_1695
Putative fimbrial protein CMD_1793 HPChromosome segregation ATPase
domain protein CMD_1922 FRG uncharacterized protein domain
CMD_2206 L-fucose isomerase like protein CMD_2434 CS12 fimbria
outer membrane usher protein CMD_2680 HpcHHpaI aldosecitrate lyase
protein family CMD_2681 Aldose -1 epimerase CMD_2682
Gluconolaconase LRE like region CMD_2900 Putative cytoplasmic
protein CMD_3009 putative restriction endonuclease CMD_3277 Putative
yqi fimbrial adhesion CMD_3278 Hypothetical fimbrial chaperone yqiH
CMD_3340 Pea-VEAacid family CMD_3518 3-hydroxy-3-methylglutaryl
CoA synthase CMD_3801 putative reverse transcriptase CMD_3840 beta-
D-galactosidase alpha subunit CMD_4033 ParB family chromosome
partitioning protein CMD_4034 ParBrepB chromosomeplasmid
partitioning protein CMD_4035 RelASpoT protein CMD_4532 putative
DNA ligase -like protein CMD_4541 Antitermination protein Q and
CMD_4585 putative ATP dependent helicase
171
CHAPTER 6
60 CONCLUSION
61 Summary
Contamination of grain with mycotoxins including deoxynivalenol (DON) and other
trichothecenes is becoming a limiting factor for cereal and animal production (Binder et al
2007) To address the issue this study obtained a novel microbial source for DON
detoxification and determined candidate genes for the function which may have potential
applications to detoxify grain and grain-derived foodfeed contaminated with DON and TC
mycotoxins
From a screen of soil samples from crop field microorganism was isolated that is able
to de-epoxidize the most frequently detected foodfeed contaminating mycotoxin DON The
effects of media compositions temperature pH and aerobicanaerobic conditions on
microbial detoxification activity were assessed From 150 analyzed soil samples a mixed soil
microbial culture that showed DON de-epoxidation activity under aerobic conditions and
moderate temperature (~24degC) was obtained This culture exhibited a slow rate of microbial
DON to de-epoxyDON transformation (de-epoxidation) possibly because of competition
between de-epoxidizing and other microorganisms in the culture
The mixed microbial culture was enriched for DON detoxifying bacteria by reducing
the non-targeted microbial species through antibiotic treatments heating and subculturing on
agar medium and selecting colonies The enriched culture had higher DON de-epoxidation
activity than the initial culture Because of low abundance of DON detoxifying
microorganisms further culture enrichments were required to increase the target microbial
172
population Reculture of the partially enriched microbial community in nutritonally poor MSB
medium with high level of DON increased the DON de-epoxidizing bacterial population
Serial dilution plating and selecting single colonies eventually led to isolation of a DON-
detoxifying bacterial strain named ADS47
Phenotypic and 16S-rDNA sequence analyses provided evidence that the bacterium is
a strain of C freundii Strain ADS47 is a gram negative rod-shaped facultative anaerobe and
has peritrichous flagella Strain ADS47 was unable to use DON as sole carbon source
however increasing DON concentrations in growth media enhanced its DON metabolism
efficiency The pure bacterial culture was shown to be able to rapidly de-epoxidize DON
under aerobic conditions moderate temperatures and neutral pH Strain ADS47 more quickly
de-epoxidizes DON in aerobic conditions than the anaerobic conditions In addition to DON
strain ADS47 showed capability to de-epoxidize andor de-acetylate 10 different type-A and
type-B TC mycotoxins which often co-contaminant grain with DON
To clone the DON detoxifying genes a DON detoxifying bacterial DNA library in E
coli host was constructed followed by screening of the library clones for DON detoxification
activity The study produced a library giving approximately 10 fold coverage of the genome
of strain ADS47 Restriction digestion analysis determined that most of the clones contained
ADS47 DNA inserts This indicates that a good DNA library was produced which had gt99
probability of containing all the DNA of the ADS47 genome However activity-based
screening of the library could not identify any clones with DON de-epoxidation ability It is
assumed that bacterial DON de-epoxidation genes evolved in a certain environmental niche
which may be required for the DON de-epoxidation to be expressed and active but was not
supplied by the host E coli cells
173
An attempt was made to identify the bacterial genes encoding DON de-epoxidation
and de-acetylation enzymes by sequencing the genome of strain ADS47 and comparing its
sequence to the genomes of related bacteria that are not known to posses DON de-epoxidation
activity A RAST server based bioinformatics analysis determined that the 48 Mb circular
genome of ADS47 codes for 4610 predicted CDS A genome wide phylogenetic analysis
determined that strain ADS47 was gt87 identical to the pathogen C freundii strain Ballerup
The relationship to two other animalhuman pathogenic species C rodentium and C koseri
was more distant This result is consistent with the 16S-rDNA based phylogenetic analysis
It is assumed that the microbial DON detoxifying genes were acquired by horizontal
gene transfer or other means during the adaptation of ADS47 in mycotoxin contaminated soil
If it is correct then this means that those newly acquired genes are not common in other
related microbial species and no other Citrobacter species is known to degrade DON Hence
the study identified 270 unique genes to ASDS47 with a subset of 10 genes for reductases
oxidoreductases and a de-acetylase which were considered to be candidate genes for the
enzymes that have DON detoxifying activity Furthermore a targeted comparative genome
analysis showed that the mycotoxin detoxifying ADS47 genome does not harbour the
clinically important LEE encoded typendashIII secretion system and mycobacterium virulence
genes This suggests that strain ADS47 is not likely to be an animal pathogenic bacterium
unlike C freundii Ballerup which encodes the pathogenic relevance LEE operons
174
62 Research contributions
The research made four major contributions which may aid in reducing the risks of
animals and humans from DONTC exposure Firstly the study identified a novel bacterial
strain ADS47 that is able to detoxify DON and 10 related mycotoxins over a wide range of
temperatures under aerobic and anaerobic conditions The discovery has led to a patent
application for strain ADS47 supporting the novelty and importance of the finding This
bacterial strain could be used to detoxify trichothecene mycotoxins in contaminated animal
feed thus reducing food safety risks
Secondly the study resulted in the development of a method for isolating DON de-
epoxidizing microbes from soil as well as optimizing the conditions for bacterial DONTC
detoxification activity to increase the effectiveness usefulness of the strain
Thirdly the thesis work resulted in availability of the genome sequence of a
trichothecene detoxifying microbe which may ultimately lead to an understanding of the
cellularmolecular mechanisms of DONTC de-epoxidation and de-acetylation
genespathways The genome analysis resulted in identification of several candidate genes for
DONTC de-epoxidation and de-acetylation activities Genes which show de-epoxidation
activity could be used to develop DONTC resistant cereal cultivars Fourthly strain ADS47
lacks major animal pathogenicity genes (in particular the LEE-operons) as well as does not
encodes mycobacterium virulence genes Thus strain ADS47 has a great potential to be
utilized to detoxify cereal grain and may be used as a feed additive for livestockfishery All
together the reported work may have significant applications to produce and supply
DONTC-free food and feed
175
63 Further research
This research provides the basis for developing biological detoxification of DON and
related mycotoxins This goal might be achieved by using DON de-epoxidizing gene to
develop resistant plants enzymatic detoxification of contaminated feed and use of the bacteria
as a feed additive Follow-up studies may reach the above indicated goals Because the
bacterium showed de-epoxidation activity under aerobicanaerobic conditions and under wide
range of temperatures it is likely to have an application for enzymatic detoxification of
contaminated animal feed Therefore additional studies on bacterial efficiency to detoxify
contaminated cereal and grain-derived feed need to be conducted Although strain ADS47
does not encode major animal pathogenicity genes the bacterium must be directly tested on
animals to see if it could negatively affect animal digestive systems If not then the potential
for ADS47 is considerable for its use as a feed additive for pig poultry and fish Animal
feeding trials will also need to be performed to determine if DON detoxification is sufficient
to be cost effective as a feed additive In order to develop DONTC resistant cereal cultivars
the gene responsible to DON degrading activity among the presently identified candidate
genes (reductases oxidoreductases and de-acetylase) will need to be expressed in other
organisms like E coli and a codon modified version adapted for expression in plants like
corn and wheat
176
70 REFERENCES
Abbas HK Mirocha CJ Pawlosky RJ Pusch DJ 1985 Effect of cleaning milling and
baking on deoxynivalenol in wheat Applied and Environmental Microbiology 50482ndash
486
Accensi F Pinton P Callu P Abella-Bourges N Guel WJF Grosjean F Oswald IP 2006
Ingestion of low doses of deoxynivalenol does not affect haematological biochemical or
immune responses of piglets Journal of Animal Science 841935ndash1942
Agrawal D Mahapatra AK 2005 Vertically acquired neonatal Citrobacter brain abscessndash
case report and review of the literature Journal of Clinical Neuroscience 12188ndash190
Alexander NJ 2008 The TRI101 story engineering wheat and barley to resist Fusarium head
blight World Mycotoxin Journal 131-37
Alexander NJ Proctor RH McCormick SP 2009 Genes gene clusters and biosynthesis of
trichothecenes and fumonisins in Fusarium Toxin Reviews 28198ndash215
Ali ML Taylor JH Jie L Sun G William M Kasha KJ Reid LM Pauls KP 2005
Molecular mapping of QTLs for resistance to Gibberella ear rot in corn caused by
Fusarium graminearum Genome 48521-533
Alikhan N-F Petty NK Zakour NLB Beatson SA 2011 BLAST Ring Image Generator
(BRIG) simple prokaryote genome comparisons BMC Genomics 12402
Alisi A Spaziani A Anticoli S Ghidinelli M Balsano C 2008 PKR is a novel functional
direct player that coordinates skeletal muscle differentiation via p38MAPKAKT
pathways Cellular Signalling 20534ndash542
177
Amuzie CJ Shinozuka J Pestka JJ 2009 Induction of suppressors of cytokine signaling by
the trichothecene deoxynivalenol in the mouse Toxicological Science 111277ndash287
Ansorge WJ 2009 Next generation DNA sequencing techniques New Biotechnology
25195-203
Arnold JW Senter SD 1998 Use of digital aroma technology and SPME GC-MS to compare
volatile compounds produced by bacteria isolated from processed poultry Journal of the
Science of Food and Agriculture 78343ndash348
Audenaert K Van Broeck R Bekaert B De Witte F Heremans B Messens K Hofte
M Haesaert G 2009 Fusarium head blight (FHB) in Flanders population
diversity inter-species associations and DON contamination in commercial winter
wheat varieties European Journal of Plant Pathology 125445-458
Awad WA Ghareeb K Bohm J Zentek J 2010 Decontamination and detoxification
strategies for the Fusarium mycotoxin deoxynivalenol in animal feed and the
effectiveness of microbial biodegradation Food Additives and Contaminants 27510-520
Aziz K Bartels D Best AA DeJongh M Disz T Edwards RA Formsma K Gerdes S Glass
EM Kubal M Meyer F Olsen GJ Olson R Osterman AL Overbeek RA McNeil LK
Paarmann D Paczian T Parrello1 B Pusch GD Reich C Stevens R Vassieva O
Vonstein V Wilke A Zagnitko O 2008 The RAST Server Rapid Annotations using
Subsystems Technology BMC Genomics 975
Badger JL Stins MF Kim KS 1999 Citrobacter freundii invades and replicates in human
brain microvascular endothelial cells Infection and Immunology 674208ndash4215
Bae HK Pestka JJ 2008 Deoxynivalenol induces p38 interaction with the ribosome in
monocytes and macrophages Toxicological Science 10559-66
178
Bai GH Desjardins AE Plattner RD 2002 Deoxynivalenol-nonproducing Fusarium
graminearum causes initial infection but does not cause disease spread in wheat spikes
Mycopathologia 15391ndash98
Barrett JR 2000 Mycotoxins of molds and maladies Environmental Health Perspectives
108A20ndashA23
Bengoechea JA Skurnik M 2000 Temperature-regulated efflux pumppotassium antiporter
system mediates resistance to cationic antimicrobial peptides in Yersinia Molecular
Microbiology 3767ndash80
Bennett JW Klich M 2003 Mycotoxins Clinical Microbiology Reviews 16497ndash516
Benson DA Karsch-Mizrachi I Lipman DJ Ostell J Sayers EW 2009 GenBank Nucleic
Acids Research 37D26-31
Berdichevsky T Friedberg D Nadler C Rokney A Oppenheim A Rosenshine I 2005 Ler is
a negative autoregulator of the LEE1 operon in neuropathogenic Escherichia coli Journal
of Bacteriology 187349ndash357
Bernardo A Bai GH Guo PG Xiao K Guenzi AC Ayoubi P 2007 Fusarium graminearum-
induced changes in gene expression between Fusarium head blight-resistant and
susceptible wheat cultivars Functional and Integrative Genomics 769ndash77
Bhat RV Beedu SR Ramakrishna Y Munshi KL 1989 Outbreak of trichothecene
mycotoxicosis associated with consumption of mould-damaged wheat production in
Kashmir Valley India The Lancet 33335ndash37
Binder EM Tan LM Chin LJ Handl J Richard J 2007 Worldwide occurrence of
mycotoxins in commodities feeds and feed ingredients Animal Feed Science and
Technology 137265-282
179
Birzele B Meier A Hindorf H Kraumlmer J Dehne H-W 2002 Epidemiology of Fusarium
infection and deoxynivalenol content in winter wheat in the Rhineland Germany
European Journal of Plant Pathology 108667ndash673
Boddu J Cho S Muehlbauer GJ 2007 Transcriptome analysis of trichothecene-induced gene
expression in barley Molecular PlantndashMicrobe Interactions 201364ndash1375
Borenshtein D Schauer DB 2006 The Genus Citrobacter Prokaryotes 690ndash98
Boutigny A-L Richard-Forget F Barreau C 2008 Natural mechanisms for cereal resistance
to the accumulation of Fusarium trichothecenes European Journal of Plant Pathology
121411ndash423
Boutrif E Canet C 1998 Mycotoxin prevention and control FAO programmes Revue de
Meacutedecine Veacuteterinaire 149681-694
Brennan JM Leonard G Fagan B Cooke BM Ritieni A Ferracane R et al 2007
Comparison of commercial European wheat cultivars to Fusarium infection of head and
seedling tissue Plant Pathology 5655ndash64
Brenner DJ Grimont PA Steigerwalt AG Fanning GR Ageron E Riddle CF 1993
Classification of Citrobacteria by DNA hybridization designation of Citrobacter farmeri
sp nov Citrobacter youngae sp nov Citrobacter braakii sp nov Citrobacter
werkmanii sp nov Citrobacter sedlakii sp nov and three unnamed Citrobacter
genomospecies International Journal of Systematic Bacteriology 43645-58
Brookes G Barfoot P 2006 GM Crops The first ten years-global socio-economic and
environmental Impacts no 36 ISAAA (International Service for the Acquisition of Agri-
Biotech Applications) Ithaca New York
180
Bruce KD Hughes MR 2000 Terminal restriction fragment length polymorphism monitoring
of genes amplified directly from bacterial communities in soils and sediments Molecular
Biotechnology 16261ndash269
Buerstmayr H Ban T Anderson JA 2009 QTL mapping and marker-assisted selection for
Fusarium head blight resistance in wheat A review Plant Breeding 1281ndash26
Campbell H Choo TM Vigier B Underhill L 2002 Comparison of mycotoxin profiles
among cereal samples from eastern Canada Canadian Journal of Botany 80526ndash532
Carver et al 2005 ACT the Artemis Comparison Tool Bioinformatics 213422-3423
Carver et al 2009 DNAPlotter circular and linear interactive genome visualization
Bioinformatics 25119-120
Chen CY et al 2003 Comparative genome analysis of Vibrio vulnificus a marine pathogen
Genome Research 132577ndash2587
Chiu TC Yen JH Hsieh YN Wang YS 2005 Reductive transformation of dieldrin under
anaerobic sediment culture Chemosphere 601182ndash1189
Chung YJ Zhou HR Pestka JJ 2003 Transcriptional and posttranscriptional roles for p38
mitogen-activated protein kinase in upregulation of TNF-alpha expression by
deoxynivalenol (vomitoxin) Toxicology and Applied Pharmacology 193188ndash201
Chung EJ Lim HK Kim J-C Choi GJ Park EJ Lee MH Chung YR Lee S-W 2008 Forest
soil metagenome gene cluster involved in antifungal activity expression in Escherichia
coli Applied and Environmental Microbiology 74723-730
Collins TFX Sprando RL Black TN Olejnik N Eppley RM Hines FA Rorie J Ruggles DI
2006 Effects of deoxynivalenol (DON vomitoxin) on in utero development in rats Food
and Chemical Toxicology 44747ndash757
181
Cong Y Wang J Chen Z Xiong K Xu Q Hu F 2011 Characterization of swarming motility
in Citrobacter freundii FEMS Microbiology Letters 371160-171
Cossette F Miller JD 1995 Phytotoxic effect of deoxynivalenol and Gibberella ear rot
resistance of corn Natural Toxins 3383ndash388
Creppy EE 2002 Update of survey regulation and toxic effects of mycotoxins in Europe
Toxicological Letters 12719-28
Dai JS Wang D Guerlebeck C Laturnus S Guenther Z Lu SC Ewers C 2010 Suppression
subtractive hybridization identifies an autotransporter adhesin gene of E coli IMT5155
specifically associated with avian pathogenic Escherichia coli (APEC) BMC
Microbiology 10236
Daniel R 2005 The metagenomics of soil Nature Review Microbiology 3470-478
Davis KER Joseph SJ Janssen PH 2005 Effects of growth medium inoculum size and
incubation time on culturability and isolation of soil bacteria Applied and Environmental
Microbiology 71826-834
Delcher AL Phillippy A Carlton J Salzberg SL 2002 MUMmer 21 NUCmer and
PROmer are described in Fast Algorithms for Large-scale Genome Alignment and
Comparisonrdquo Nucleic Acids Research 302478-2483
Demain AL Adrio JL 2008 Contributions of microorganisms to industrial biology
Molecular Biotechnology 3841ndash455
Deng W Li Y Vallance BA Finlay BB 2001 Locus of enterocyte efffacement from
Citrobacter rodentium sequence analysis and evidence for horizontal transfer among
attaching and effacing pathogens Infection and Immunity 696323ndash6335
182
Desjardins AE Proctor RH Bai G McCormick SP Shaner G Beuchley G Hohn TM 1996
Reduced virulence of trichothecene-nonproducing mutants of Gibberella zeae in wheat
field tests Molecular Plant-Microbe Interactions 9775-781
Desjardins AE 2006 Fusarium mycotoxins chemistry genetics and biology The American
Phytopathological Society St Paul Minnesota USA p 260
Desjardins AE Proctor RH 2007 Molecular biology of Fusarium mycotoxins International
Journal of Food Microbiology 11947-50
Desmond OJ Manners JM Stephens AE MacLean DJ Schenk PM Gardiner DM Munn
AL Kazan K 2008 The Fusarium mycotoxin deoxynivalenol elicits hydrogen peroxide
production programmed cell death and defence responses in wheat Molecular Plant
Pathology 9435-445
Dill-Macky R Jones RK 2000 The effect of previous crop residues and tillage on Fusarium
head blight of wheat Plant Disease 8471ndash76
DrsquoMello JPF Macdonald AMC Dijksma WTP 1998 3-Acetyl deoxynivalenol and esterase
production in a fungicide-insensitive strain of Fusarium culmorum Mycotoxin Research
149ndash18
DrsquoMello JPF Placinta CM Macdonald AMC 1999 Fusarium mycotoxins a review of
global implications of animal health welfare and productivity Animal Feed Science and
Technology 80183-205
Doohan FM Brennan J Cooke BM 2003 Influence of climatic factors on Fusarium species
pathogenic to cereals European Journal of Plant Pathology 109755ndash768
183
Dyer RB Kendra DF Brown DW 2006 Real-time PCR assay to quantify Fusarium
graminearum wild-type and recombinant mutant DNA in plant material Journal of
Microbiological Methods 67534ndash542
Erecinska M Wilson DF 1981 Inhibitors of cytochrome c oxidase In inhibitors of
mitochondrial function edited by M Erecinska and DF Wilson Pergamon Press Ltd
Oxford pp 145-160
Eriksen GS 2003 Metabolism and toxicity of trichothecene Doctoral thesis Swedish
University of Agricultural Sciences Uppsala Sweden
Eriksen GS Pettersson H Lundh T 2004 Comparative cytotoxicity of deoxynivalenol
nivalenol their acetylated derivatives and de-epoxy metabolites Food Chemistry and
Toxicology 42619-624
Eudes F Comeau A Rioux S Collin J 2000 Phytotoxicity of eight mycotoxins associated
with Fusarium in wheat head blight Canadian Journal of Plant Pathology 22286ndash292
FAO (Food and Agriculture Organization) 1991 Food Nutrition and Agriculture Food for
the Future volume 1 FAO Rome
FAO (Food and Agriculture Organization) 2001 FNP73 Manual of the application of
HACCP system in mycotoxin prevention and control FAO Food and Nutrition paper no
73 FAO Rome
Fedorak PM Grbic-Galic D 1991 Aerobic microbial cometabolism of benzothiophene and
3-methylbenzothiophene Applied and Environmental Microbiology 57932-940
Felsenstein J 1989 PHYLIP - Phylogeny Inference Package (Version 32) Cladistics 5164-
166
184
Fernando WGD Amarasinghe C Demoz B Surujdeo‐Maharaj S Gauthier V Guerrieri G
Al‐Taweel K Brucircleacute‐Babel A Tamburic‐Illincic L Gilbert J 2011 What do we know of
the Fusarium graminearum chemotypes their epidemiology genetics and infection
process in Canada In Proc 7th
Canadian Workshop on Fusarium Head Blight edited by
J Gilbert A Tekauz N Sweetland and K Slusarenko Winnipeg Manitoba Canada p 45
Fierer N Jackson RB 2006 The diversity and biogeography of soil bacterial community
Proceedings of the National Academy of Sciences 103626-631
Fink-Gremmels J 1989 The significance of mycotoxin assimilation for meat animals
Deutsche Tierarztliche Wochenschrift 96360-363
Flint DH Emptage MH Finnegan MG Fu W Johnson MK 1993 The role and properties of
the iron-sulfur cluster in Escherichia coli dihydroxy-acid dehydratase Journal of
Biological Chemistry 26814732ndash14742
Foroud NA Eudes F 2009 Trichothecenes in cereal grains International Journal of
Molecular Science 10147-173
Forsell JH Jensen R Tai JH Witt M Lin WS Pestka JJ 1987 Comparison of acute
toxicities of deoxynivalenol (vomitoxin) and 15-acetyldeoxynivalenol in the B6C3F1
mouse Food and Chemical Toxicology 25155ndash162
Frey M Stettner C Gierl A 1998 A general method for gene isolation in tagging approaches
amplification of insertion mutagenised sites (AIMS) The Plant Journal 13717-721
Freeman GG Morrison RI 1949 The isolation and chemical properties of trichothecin an
antifungal substance from Trichothecium roseum Link Biochemical Journal 441ndash5
185
Fuchs E Binder EM Heidler D Krska R 2002 Structural characterization of metabolites
after the microbial degradation of type A trichothecenes by the bacterial strain BBSH 797
Food Additives and Contaminants 19379ndash386
Gabor EM Alkema WBL Janssen DB 2004 Quantifying the accessibility of the
metagenome by random expression cloning techniques Environmental Microbiology
6879ndash886
Gadd GM Griffiths AJ 1978 Microorganisms and heavy metal toxicity Microbial Ecology
4303ndash317
Gale LR Bryant JD Calvo S Giese H 2005 Chromosome complement of the fungal plant
pathogen Fusarium graminearum based on genetic and physical mapping and cytological
observations Genetics 171985ndash1001
Gao LY Pak M Kish R Kajihara K Brown EJ 2006 A mycobacterial operon essential for
virulence in vivo and invasion and intracellular persistence in macrophages Infection and
Immunity 741757ndash1767
Gareau MG Wine E Reardon C et al 2010 Probiotics prevent death caused by Citrobacter
rodentium infection in neonatal mice Journal of Infectious Diseases 20181e91
Gareis M Bauer J Thiem J Plank G Grabley S Gedek B 1990 Cleavage of
zearalenoneglycoside a masked mycotoxin during digestion in swine Journal of
Veterinary Medicine 37236-240
Geng L Xin W Haung DW Feng G 2006 A universal cloning vector using vaccinia
topoisomerase I Molecular Biotechnology 3323-28
186
Golkari S Gilbert J Ban T Procunier JD 2009 QTLspecific microarray gene expression
analysis of wheat resistance to Fusarium head blight in Sumai-3 and two susceptible
NILs Genome 52409ndash418
Goodstadt L Ponting CP 2004 Vitamin K epoxide reductase homology active site and
catalytic mechanism Trends in Biochemical Sciences 29289-292
Goswami RS Kistler HC 2004 Heading for disaster Fusarium graminearum on cereal
crops Molecular Plant Pathology 5515-525
Gouze ME Laffitte J Printon P Dedieux G Galinier A Thouvenot JP Loiseau N Oswald
IP Galtier P 2007 Effect of subacute oral doses of nivalenol on immune and metabolic
defence systems in mice Veterinary Research 38635ndash646
Guan S He J Young JC Zhu H Li X Ji C Zhou T 2009 Transformation of trichothecene
mycotoxins by microorganisms from fish digesta Aquaculture 290290ndash295
Gupta R Beg QK Lorenz P 2002 Bacterial alkaline proteases molecular approaches and
industrial applications Applied Microbiology and Biotechnology 5915ndash32
Haberle J Schweizer G Schondelmaier J Zimmermann G Hart L 2009 Mapping of QTL
for resistance against Fusarium head blight in the winter wheat population
PelikanBussardNing8026 Plant Breeding 12827ndash35
Hall BG 2004 Predicting the evolution of antibiotic resistance genes Nature Reviews
Microbiology 2430-435
Han JI et al 2011 Complete genome sequence of the metabolically versatile plant growth-
promoting endophyte Variovorax paradoxus S110 Journal of Bacteriology 1931183ndash
1190
187
Harvey RB Edrington TS Kubena LF Elissalde MH Corrier DE Rottinghaus GE 1995
Effect of aflatoxin and diacetoxyscirpenol in ewe lambs Bulletin of Environmental
Contamination and Toxicology 54325-330
Harvey RB Edrington TS Kubena LF Elissalde MH Casper HH Rottinghaus GE Turk JR
1996 Effects of dietary fumonisin B1-containing culture material deoxynivalenol-
contaminated wheat or their combination on growing barrows American Journal of
Veterinary Research 571790-1794
He J 2007 Microbial strategies for management of Fusarium graminearum and
transformation of deoxynivalenol Master of Science Thesis University of Guelph
Guelph Ontario Canada
He K Vines L Pestka JJ 2010a Deoxynivalenol-induced modulation of microRNA
expression in RAW 2647 macrophages-A potential novel mechanism for translational
inhibition Toxicologist 114310
He J Zhou T Young JC Boland GJ Scott PM 2010b Chemical and biological
transformations for trichothecene mycotoxins in human and animal food chains A review
Trends in Food Science and Technology 2167-76
He J Zhou T 2010 Patented techniques for detoxification of mycotoxins in feeds and food
matrices Rec Pat Food Nutr Agric 296ndash104
He P Young LG Forsberg C 1992 Microbial transformation of deoxynivalenol (vomitoxin)
Applied and Environmental Microbiology 583857-3863
Hell K Cardwell KF Setamou M Poehling H-M 2000 The influence of storage practices on
aflatoxins contamination in maize in four agroecological zones in Benin West Africa
Journal of Stored Product Research 36365ndash82
188
Henne A Schmitz RA Bomeke M Gottschalk G Daniel R 2000 Screening of
environmental DNA libraries for the presence of genes conferring lipolytic activity on
Escherichia coli Applied and Environmental Microbiology 663113ndash3116
Hodges GR Degener CE Barnes WG 1978 Clinical significance of Citrobacter isolates
American Journal of Clinical Pathology 7037ndash40
Hollinger K Ekperigin HE 1999 Mycotoxicosis in food producing animals The Veterinary
Clinics of North America Food Animal Practice 15133-165
Hong Y Kinoshita T 2009 Trypanosome glycosylphosphatidylinositol biosynthesis Korean
Journal of Parasitology 47197ndash204
Hope RJ Aldred D Magan N 2005 Comparison of environmental profiles for growth and
deoxynivalenol production by Fusarium culmorum and F graminearum on wheat grain
Letters in Applied Microbiology 40295ndash300
Horvath RS 1972 Microbial co-metabolism and the degradation of organic compounds in
nature Bacteriological Reviews 36146ndash155
Hsu D Shih LM Zee YC 1994 Degradation of rRNA in Salmonella strains a novel
mechanism to regulate the concentrations of rRNA and ribosomes Journal of
Bacteriology 1764761ndash4765
Huwig A Freimund S Kappeli O Dutler H 2001 Mycotoxin detoxification of animal feed
by different adsorbents Toxicological Letters 122179ndash188
Iordanov MS Pribnow D Magun JL Dinh TH Pearson JA Magun BE 1997 Ribotoxic
stress response activation of the stress activated protein kinase JNK1 by inhibitors of the
peptidyl transferase reaction and by sequence-specific RNA damage to the sarcinricin
loop in the 28S rRNA Molecular and Cellular Biology 173373ndash3381
189
Islam R Zhou T Young JC Goodwin PH Pauls KP 2012 Aerobic and anaerobic de-
epoxidation of mycotoxin deoxynivalenol by bacteria originating from agricultural soil
World Journal of Microbiology and Biotechnology 287-13
Jackson LS Bullerman LB 1999 Effect of processing on Fusarium mycotoxins Advances in
Experimental Medicine and Biology 459243-261
Jansen C von Wettstein D Schafer W Kogel K-H Felk A Maier FJ 2005 Infection patterns
in barley and wheat spikes inoculated with wild-type and trichodiene synthase gene
disrupted Fusarium graminearum Proceedings of the National Academy of Sciences
10216892ndash16897
Jube S Borthakur D 2007 Expression of bacterial genes in transgenic tobacco methods
applications and future prospects Electronic Journal of Biotechnology 10452-467
Kabak B Dobson AD 2009 Biological strategies to counteract the effects of mycotoxins
Journal of Food Protection 722006-2016
Kant P Reinprecht Y Martin CJ Islam R Pauls KP 2011 Integration of biotechnologies
disease resistance pathology and Fusarium In Comprehensive Biotechnology edited by
Moo-Young M Second Edition 4729-743
Kaplan CW Kitts CL 2004 Bacterial succession in a petroleum land treatment unit Applied
and Environmental Microbiology 701777-1786
Kaprelyants AS Kell DB 1996 Do bacteria need to communicate with each other for
growth Trends in Microbiology 4237-42
190
Kariholu U Rawal J Namnyak S 2009 Neonatal Citrobacter koseri meningitis and brain
abscess The Internet Journal of Pediatrics and Neonatology 10
(httpwwwispubcomjournalthe-internet-journal-of-pediatrics-and-
neonatologyvolume-10-number-1neonatal-citrobacter-koseri-meningitis-and-brain-
abscesshtml) (accessed on April 19 2012)
Karlovsky P 1999 Biological detoxification of fungal toxins and its use in plant breeding
feed and food production Natural Toxins 71ndash23
Karlovsky P 2011 Biological detoxification of the mycotoxin deoxynivalenol and its use in
genetically engineered crops and feed additives Applied Microbiology and
Biotechnology 91491-504
Kavanagh KL Jornvall H Persson B Oppermann U 2008 Medium and short-chain
dehydrogenasereductase gene and protein families the SDR superfamily functional and
structural diversity within a family of metabolic and regulatory enzymes Cellular and
Molecular Life Sciences 653895ndash3906
Khera KS Whalen C Angers G Vesonder RF Kuiper-Goodman T 1982 Embryotoxicity of
4-deoxynivalenol (vomitoxin) in mice Bulletin of Environmental Contamination and
Toxicology 29487ndash491
Kikot GE Hours RA Alconada TM 2009 Contribution of cell wall degrading enzymes to
pathogenesis of Fusarium graminearum a review Journal of Basic Microbiology 49
231ndash241
Kim UJ Shizuya H Jong PJ Birren B Simon MI 1992 Stable propogation of cosmid sized
human DNA inserts in a F factor based vector Nucleic Acids Research 201083-1065
191
Kimura M Kaneko I Komiyama M Takatsuki A Koshino H Yoneyama K Yamaguchi I
1998 Trichothecene 3-O-acetyltransferase protects both the producing organism and
transformed yeast from related mycotoxins cloning and characterization of Tri101
Journal of Biological Chemistry 2731654-1661
Kimura M Anzai H Yamaguchi I 2001 Microbial toxins in plantndashpathogen interactions
biosynthesis resistance mechanisms and significance The Journal of General and
Applied Microbiology 47149ndash160
Kimura M Tokai T Takahashi-Ando N Ohsato S Fujimura M 2007 Molecular and genetics
studies of Fusarium trichothecene biosynthesis pathways genes and evolution
Bioscience Biotechnology and Biochemistry 712105-2123
King RR McQueen RE Levesque D Greenhalgh R 1984 Transformation of deoxynivalenol
(vomitoxin) by rumen microorganisms Journal of Agricultural and Food Chemistry
32181ndash1183
Klappenbach JA Dunbar JM Schmidt TM 2000 rRNA operon copy number reflects
ecological strategies of bacteria Applied and Environmental Microbiology 661328ndash
1333
Knietsch A Waschkowitz T Bowien S Henne A Daniel R 2003 Metagenomes of complex
microbial consortia derived from different soils as sources for novel genes conferring
formation of carbonyls from short-chain polyols on Escherichia coli Journal of Molecular
Microbiology and Biotechnology 546ndash56
Kollarczik B Gareis M Hanelt M 1994 In vitro transformation of the Fusarium mycotoxins
deoxynivalenol and zearalenone by the normal gut microflora of pigs Natural Toxins
2105ndash110
192
Koonin EV Wolf YI Aravind L 2001 Prediction of the archaeal exosome and its
connections with the proteasome and the translation and transcription machineries by a
comparative-genomic approach Genome Research 11240ndash252
Kouadio JH Dano SD Moukha S Mobio TA Creppy EE 2007 Effects of combinations of
Fusarium mycotoxins on the inhibition of macromolecular synthesis malondialdehyde
levels DNA methylation and fragmentation and viability in Caco-2 cells Toxicon
49306ndash317
Krause L Diaz NN Bartels D Edwards RA Puumlhler A et al 2006 Finding novel genes in
bacterial communities isolated from the environment Bioinformatics 22281ndash289
Kubena LF Huff WE Harvey RB Phillips TD Rottinghaus GE 1989 Individual and
combined toxicity of deoxynivalenol and T-2 toxin in broiler chicks Poultry Science
68622ndash626
Kubena LF Harvey RB Huff WE Corrier DE Phillips TD Rottinghaus GE 1990 Efficacy
of hydrated sodium calcium aluminosilicate to reduce the toxicity of aflatoxin and T-2
toxin Poultry Science 691078ndash1086
Kubena LF Harvey RB Huff WE Elissalde MH Yersin AG Phillips TD Rottinghaus GE
1993 Efficacy of hydrated sodium calcium aluminosilicate to reduce the toxicity of
aflatoxin and diacetoxyscirpenol Poultry Science 7251ndash59
Kubena LF Edrington TS Harvey RB Buckley SA Phillips TD Rottinghaus GE Casper
HH 1997 Individual and combined effects of fumonisin B1 present in Fusarium
moniliforme culture material and T-2 toxin or deoxynivalenol in broiler chicks Poultry
Science 761239ndash1247
193
Kumar RA Gunasekaran P Lakshmanan M 1999 Biodegradation of tannic acid by
Citrobacter freundii isolated from a tannery effluent Journal of Basic Microbiology
39161-168
Kushiro M 2008 Effects of milling and cooking processes on the deoxynivalenol content in
wheat International Journal Molecular Science 92127-2145
Lane DJ 1991 16S23S rRNA sequencing In Nucleic acid techniques in bacterial
systematic edited by Stackebrandt E and Goodfellow M Wiley J and Sons Inc New
York pp 115-175
Lancova K Hajslova J Poustka J Krplova A Zachariasova M Dostalek P Saschambula L
2008 Transfer of Fusarium mycotoxins and ldquomaskedrdquo deoxynivalenol (deoxynivalenol-3-
glucoside) from field barley through malt to beer Food Additives and Contaminants
25732ndash744
Langmead B Trapnell C Pop M Salzberg SL 2009 Ultrafast and memory-efficient
alignment of short DNA sequences to the human genome Genome Biology 10R25
Larsen JC Hunt J Perrin I Ruckenbauer P 2004 Workshop on trichothecenes with a focus
on DON summary report Toxicological Letters 1531-22
Lee T Oh DW Kim HS Lee J Kim YH Yun SH Lee YW 2001 Identification of
deoxynivalenol- and nivalenol-producing chemotypes of Gibberella zeae by using PCR
Applied and Environmental Microbiology 672966ndash2972
Lee S-W Won K Lim HK Kim J-C Choi GJ Cho KY 2004 Screening for novel lipolytic
enzymes from uncultured soil microorganisms Applied and Environmental Microbiology
65720-726
194
Li L Christian J Stoeckert J David SR 2003 OrthoMCL Identification of Ortholog Groups
for Eukaryotic Genomes Genome Research 132178-2189
Li T Chang CY Jin DY Lin PJ Khvorova A Stafford DW 2004 Identification of the gene
for vitamin K epoxide reductase Nature 427541ndash544
Li Y 2007 Sensory signal transduction in the vagal primary afferent neurons Current
Medicinal Chemistry 142554ndash2563
Li MX Pestka JJ 2008 Comparative induction of 28S ribosomal RNA cleavage by ricin and
the trichothecenes deoxynivalenol and T-2 toxin in the macrophage Toxicological
Science 10567ndash78
Li R Li Y Kristiansen K Wang J 2008 SOAP short oligonucleotide alignment program
Bioinformatics 24713-4
Li W Schulman S Dutton RJ Boyd D Beckwith J Rapoport TA 2010 Structure of a
bacterial homologue of vitamin K epoxide reductase Nature 463507-512
Lichstein HC Malcolm HS 1943 Studies of the effect of sodium azide on microbic growth
and respiration Journal of Bacteriology 47221ndash230
Limay-Rios V Schaafsma AW 2011 2010 Update on the development of an integrated
mycotoxin management system in Ontario corn 30th
Centralia Swine Research Update
Kirkton Ontario Canada
Llorens A Hinojo MJ Mateo R Gonzalez-Jaen MT Valle- Algarra FM Logrieco A Jimenez
M 2006 Characterization of Fusarium spp isolates by PCR-RFLP analysis of the
intergenic spacer region of the rRNA gene (rDNA) International Journal of Food
Microbiology 106287ndash306
195
Lombaert GA Pellaers P Roscoe V Mankotia M Neil R Scott PM 2003 Mycotoxins in
infant cereal foods from the Canadian retail market Food Additives and Contaminants
20494ndash504
Luengo JM Garciacutea JL Olivera ER 2001 The phenylacetyl-CoA catabolon a complex
catabolic unit with broad biotechnological applications Molecular Microbiology
391434ndash1442
Luo X 1994 Food poisoning caused by Fusarium toxins In Proceedings of the Second
Asian Conference on Food Safety ILSI Thailand pp 129ndash136
Mabagala RB 1997 The effect of populations of Xanthomonas campestris pv phaseoli in
bean reproductive tissues on seed infection of resistant and susceptible bean genotypes
European Journal of Plant Pathology 103175ndash181
Madhyastha MS Marquardt RR Abramson D 1994 Structurendashactivity relationships and
interactions among trichothecene mycotoxins as assessed by yeast bioassay Toxicon
321147ndash1152
Maier FJ Miedaner T Hadeler B Felk A Salomon S Lemmens M Kassner H Schaumlfer W
2006 Involvement of trichothecenes in fusarioses of wheat barley and maize evaluated
by gene disruption of the trichodiene synthase (Tri5) gene in three field isolates of
different chemotype and virulence Molecular Plant Pathology 7449ndash461
Malojcic G Owen RL Grimshaw JP Glockshuber R 2008 Preparation and structure of the
charge-transfer intermediate of the transmembrane redox catalyst DsbB FEBS Letters
5823301ndash3307
196
Manoharan M Dahleen LS Hohn TM Neate SM Yu XH Alexander NJ et al 2006
Expression of 3-OH trichothecene acetyltransferase in barley (Hordeum vulgare L) and
effects on deoxynivalenol Plant Science 171699ndash706
Maresca M Mahfoud R Garmy N Fantini J 2002 The mycotoxin deoxynivalenol affects
nutrient absorption in human intestinal epithelial cells The Journal of Nutrition
1322723ndash2731
Martins ML Martins HM 2002 Influence of water activity temperature and incubation time
on the simultaneous production of deoxynivalenol and zearalenone in corn (Zea mays) by
Fusarium graminearum Food Chemistry 79315ndash318
Masuda D Ishida M Yamaguchi K Yamaguchi I Kimura M Nishiuchi T 2007 Phytotoxic
effects of trichothecenes on the growth and morphology of Arabidopsis thaliana Journal
of Experimental Botany 581617ndash1626
Matsumoto E Kawanaka Y Yun SJ Oyaizu H 2008 Isolation of dieldrin- and endrin-
degrading bacteria using 12-epoxycyclohexane as a structural analog of both compounds
Applied Microbiology and Biotechnology 801095-1103
McLean M 1996 The phytotoxicity of Fusarium metabolites an update since 1989
Mycopathologia 133163ndash179
Merhej J Richard-Forget F Barreau C 2011 Regulation of trichothecene biosynthesis in
Fusarium recent advances and new insights Applied Microbiology and Biotechnology
91519ndash528
Midwest Plan Service 1987 Grain Drying Handling Storage Handbook Ames Iowa Iowa
State University 2nd edition
197
Miller SS Reid LM Harris LJ 2007 Colonization of maize silks by Fusarium graminearum
the causative organism of Gibberella ear rot Canadian Journal of Botany 85369-376
Minervini F Fornelli F Flynn KM 2004 Toxicity and apoptosis induced by mycotoxins
nivalenol deoxynivalenol and fumonisin B1 in a human erythroleukemia cell line
Toxicology In Vitro 1821ndash28
Mitterbauer R Poppenberger B RaditschnigA Lucyshyn D Lemmens M Glossl J Adam G
2004 Toxin-dependent utilization of engineered ribosomal protein L3 limits trichothecene
resistance in transgenic plants Plant Biotechnology Journal 2329ndash340
Mocali S Benedetti A 2010 Exploring research frontiers in microbiology the challenge of
metagenomics in soil microbiology Research Microbiology 161497-505
Morozkina EV Zvyagilskaya RA 2007 Nitrate reductases structure functions and effect of
stress factors Biochemistry 721151ndash1160
Munkvold GP 2003 Cultural and genetic approaches to managing mycotoxins in maize
Annual Review of Phytopathology 4199-116
Mundy R MacDonald TT Dougan G Frankel G Wiles S 2005 Citrobacter rodentium of
mice and man Cellular Microbiology 71697ndash1706
Nakamura N Inoue N Watanabe R Takahashi M Takeda J Stevens VL Kinoshita T 1997
Expression cloning of PIG-L a candidate N-acetylglucosaminyl-phosphatidylinositol
deacetylase The Journal of Biological Chemistry 27215834ndash15840
Nganje WE Bangsund DA Leistritz FL Wilson WW Tiapo NM 2002 Estimating the
economic impact of a crop disease the case of Fusarium head blight in US wheat and
barley National Fusarium Head Blight Forum Proceedings East Lansing Michigan State
University pp 275ndash281
198
Nielsen C Lippke H Didier A Dietrich R Martlbauer E 2009 Potential of deoxynivalenol
to induce transcription factors in humanhepatoma cells Molecular Nutrition and Food
Research 53479ndash491
Nielsen JKS Vikstroumlm AC Turner P Knudsen LE 2011 Deoxynivalenol transport across
the human placental barrier Food and Chemical Toxicology 492046-2052
Nishiuchi T Masuda D Nakashita H Ichimura K Shinozaki K Yoshida S et al 2006
Fusarium phytotoxin trichothecenes have an elicitor-like activity in Arabidopsis thaliana
but the activity differed significantly among their molecular species Molecular Plantndash
Microbe Interactions 19512ndash520
Ohsato S Ochiai-Fukuda T Nishiuchi T Takahashi-Ando N Koizumi S Hamamoto H
Kudo T Yamaguchi I Kimura M 2007 Transgenic rice plants expressing trichothecene
3-O-acetyltransferase show resistance to the Fusarium phytotoxin deoxynivalenol Plant
Cell Reports 26531-538
Okubara PA Blechl AE McCormick SP Alexander NJ Dill-Macky R Hohn TM 2002
Engineering deoxynivalenol metabolism in wheat through the expression of a fungal
trichothecene acetyltransferase gene Theoritical and Applied Genetics 10674-83
Ong CL Beatson SA Totsika M Forestier C McEwan AG Schembri MA 2010 Molecular
analysis of type 3 fimbrial genes from Escherichia coli Klebsiella and Citrobacter
species BMC Microbiology 10183
Osborne LE Stein JM 2007 Epidemiology of Fusarium head blight on small-grain cereals
International Journal of Food Microbiology 119103-108
Page RDM 1996 TREEVIEW An application to display phylogenetic trees on personal
computers Computer Applications in the Biosciences 12357-358
199
Parkhill J Dougan G James KD Thomson NR Pickard D Wain J Churcher C Mungall KL
Bentley SD Holden MT et al 2001 Complete genome sequence of a multiple drug
resistant Salmonella enterica serovar Typhi CT18 Nature 413848ndash852
Paterson RRM Lima N 2010 How will climate change affect mycotoxins in food Food
Research International 431902ndash14
Paul PA El-Allaf SM Lipps PE Madden LV 2004 Rain splash dispersal of Gibberella zeae
within wheat canopies in Ohio Phytopathology 941342-1349
Pestka JJ Lin WS Miller ER 1987 Emetic activity of the trichothecene 15-
acetyldeoxynivalenol in pigs Food and Chemical Toxicology 25855ndash858
Pestka JJ Smolinski AT 2005 Deoxynivalenol Toxicology and potential effects on humans
Journal of Toxicology Environmental Health 839ndash69
Pestka JJ 2007 Deoxynivalenol toxicity mechanisms and animal health risks Animal Feed
Science and Technology 137283-298
Pestka JJ 2008 Mechanisms of deoxynivalenol-induced gene expression and apoptosis Food
Additives and Contaminants 251128-1140
Pestka J2010 Toxicological mechanisms and potential health effects of deoxynivalenol and
nivalenol World Mycotoxin Journal 3323ndash347
Petersohn A Brigulla M Haas S Hoheisel JD Voumllker U Hecker M 2001 Global Analysis
of the General Stress Response of Bacillus subtilis Journal of Bacteriology 1835617-
5631
200
Petty NK Bulgin R Crepin VF Cerdeno-Tarraga AM Schroeder GN Quail MA Lennard N
Corton C Barron A Clark L Toribio AL Parkhill J Dougan G Frankel G Thomson NR
2010 The Citrobacter rodentium genome sequence reveals convergent evolution with
human pathogenic Escherichia coli Journal of Bacteriology 192525ndash538
Petty NK Feltwell T Pickard D Clare S Toribio AL et al 2011 Citrobacter rodentium is
an unstable pathogen showing evidence of significant genomic flux PLoS Pathology
7e1002018 doi101371journalppat1002018
Pittet A 2001 Natural occurrence of mycotoxins in foods and feeds A decade in review In
Mycotoxins and phytotoxins in perspective at the turn of the millennium edited by De
Koe WJ Samson RA Van Egmond HP Gilbert J Sabino M W J de Koe Wageningen
Netherlands pp 153ndash172
Placinta CM DrsquoMello JPF Macdonald AMC 1999 A review of worldwide contamination of
cereal grains and animal feed with Fusarium mycotoxins Animal Feed Science and
Technology 7821ndash37
Plank B Schuh M Binder EM 2009 Investigations on the effect of two feed additives
Biomin BBSH 797 and Mycofix Plus (R) 3E as detoxificants of DON contaminated feed
of piglets Wien Tierarztl Monatssch 9655ndash71
Poapolathep A Ohtsuka R Kiatipattanasakul W Ishigami N Nakayama H Doi K 2002
Nivalenol-induced apoptosis in thymus spleen and Peyerrsquos patches of mice Experimental
and Toxicological Pathology 53441ndash446
201
Poppenberger B Berthiller F Lucyshyn D Sieberer T Schuhmacher R Krska R Kuchler K
Gloumlssl J Luschnig C Adam G 2003 Detoxification of the Fusarium mycotoxin
deoxynivalenol by a UDP-glucosyltransferase from Arabidopsis thaliana Journal of
Biological Chemistry 27847905ndash47914
Prelusky DB Trenholm HL 1993 The efficacy of various classes of anti-emetics in
preventing deoxynivalenol-induced vomiting in swine Natural Toxins 1296ndash302
Proctor RH Hohn TM McCormick SP 1995 Reduced virulence of Gibberella zeae caused
by disruption of a trichothecene toxin biosynthetic gene Molecular Plant-Microbe
Interactions 8593-601
Puchenkova SG 1996 Enterobacteria in areas of water along the Crimean Coast
Mikrobiolohichnyĭ zhurnal 58 3-7
Qi BW Hao BL 2004 Whole-proteome prokaryote phylogeny without sequence alignment
a K-string composition approach Journal of Molecular Evolution 581ndash11
Quarta A Mita G Haidukowski M Logrieco A Mule G Visconti A 2006 Multiplex PCR
assay for the identification of nivalenol 3- and 15-acetyl-deoxynivalenol chemotypes in
Fusarium FEMS Microbiological Letters 2597ndash13
Rehr B Klemme JH 1989 Formate dependent nitrate and nitrite reduction to ammonia by
Citrobacter freundii and competition with denitrifying bacteria Antonie van Leeuwenhoek
Journal of Microbiology 56311ndash321
Rocha O Ansari K Doohan FM 2005 Effects of trichothecene mycotoxins on eukaryotic
cells A review Food Additives and Contaminants 22369ndash378
Roscoe V Lombaert GA Huzel V et al 2008 Mycotoxins in breakfast cereals from the
Canadian retail market a 3-year survey Food Additives and Contaminants 25347ndash355
202
Rotter BA Prelusky DB Pestka JJ 1996 Toxicology of deoxynivalenol (vomitoxin) Journal
of Toxicology and Environment Health 481ndash34
Ryu JC Ohtsubo K Izurniyarna N Nakamura K Tanaka T Yamarnura H Ueno Y 1988
The acute and chronic toxicities of nivalenol in mice Fundamental and Applied
Toxicology 1138ndash47
Saier MH Roseman S 1972 Inducer exclusion and repression of enzyme synthesis in
mutants of Salmonella typhimurium defective in enzyme I of the phosphoenolpyruvate
sugar phosphotransferase system The Journal of Biological Chemistry 247972ndash975
Saier MHJ Tran CV Barabote RD 2006 TCDB the transporter classification database for
membrane transport protein analyses and information Nucleic Acids Research 34D181ndash
186
Saitou N Nei M 1987 The neighbor-joining method a new method for reconstructing
phylogenetic trees Molecular Biology and Evolution 4406ndash425
Savelieva O Kotova I Roelofsen W Stams AJM Netrusov A 2004 Utilization of
aminoaromatic acids by a methanogenic enrichment culture and by a novel Citrobacter
freundii strain Archives of Microbiology 181163ndash170
Schaafsma AW Tamburic-Ilinic L Miller JD Hooker DC 2001 Agronomic considerations
for reducing deoxynivalenol in wheat grain Canadian Journal of Plant Pathology 23279ndash
285
Schaafsma AW 2002 Economic changes imposed by mycotoxins in food grains case study
of deoxynivalenol in winter wheat Advances in Experimental Medicine and Biology
504271ndash276
203
Scharf C Riethdorf S Ernst H Engelmann S Voumllker U Hecker M 1998 Theoredoxin is an
essential protein induced by heat shock and other stresses in Bacillus subtilis Journal of
Bacteriology 1801869-1877
Schatzmayr G Zehner F Taubel M Schatzmayr D Klimitsch A Loibner AP Binder EM
2006 Microbiologicals for deactivating mycotoxins Molecular Nutrition and Food
Research 50543-551
Schollenberger M Muumlller H-M Ruumlfle R Suchy S Plank S Drochner W 2006 Natural
occurrence of 16 Fusarium toxins in grains and feedstuffs of plant origin from Germany
Mycopathologia 16143ndash52
Scientific Committee for Food 2002 Opinion on Fusarium toxins Part 6 Group evaluation
of T-2 toxin HT-2 toxin nivalenol and deoxynivalenol (available at http
wwweuropaeuintcommfoodfsscscfout123_enpdf)
Schulman S Wang B Li W Rapoport TA 2010 Vitamin K epoxide reductase prefers ER
membrane-anchored thioredoxin-like redox partners Proceedings of the National
Academy of Sciences 10715027ndash15032
Scott PM 1997 Multi-year monitoring of Canadian grains and grain-based foods for
trichothecenes and zearalenone Food Additives and Contaminants 14333-339
Seitz LM Bechtel DB 1985 Chemical physical and microscopal studies of scab-infected
hard red winter wheat Journal of Agricultural and Food Chemistry 33373-7
Sergent T Parys M Garsou S Pussemier L Schneider YJ Larondelle Y 2006
Deoxynivalenol transport across human intestinal Caco-2 cells and its effects on cellular
metabolism at realistic intestinal concentrations Toxicological Letters 164167ndash176
204
Sharma J Fulekar MH 2009 Potential of Citrobacter freundii for bioaccumulation of
heavy metalndashcopper Biology and Medicine 17-14
Sherif SO Salama EE Abdel-Wahhab MA 2009 Mycotoxins and child health the need for
health risk assessment International Journal of Hygiene and Environmental Health
212347ndash368
Shifrin VI Anderson P 1999 Trichothecene mycotoxins trigger a ribotoxic stress response
that activates c-Jun N-terminal kinase and p38 mitogen-activated protein kinase and
induces apoptosis The Journal of Biological Chemistry 27413985ndash13992
Shima J Takase S Takahashi Y Iwai Y Fujimoto H Yamazaki M Ochi K 1997 Novel
detoxification of the trichothecene mycotoxin deoxynivalenol by a soil bacterium isolated
by enrichment culture Applied and Environmental Microbiology 633825ndash3830
Singh J Behal A Singla N Joshi A Birbian N et al 2009 Metagenomics Concept
methodology ecological inference and recent advances Biotechnology Journal 4480ndash
494
Smith TK McMillan EG Castillo JB 1997 Effect of feeding blends of Fusarium mycotoxin-
contaminated grains containing deoxynivalenol and fusaric acid on growth and feed
consumption of immature swine Journal of Animal Science 752184ndash2191
Smits WK Dubois JYF Bron S van Dijl JM Kuipers OP 2005 Tricksy business
transcriptome analysis reveals the involvement of thioredoxin in redox homeostasis
oxidative stress sulfur metabolism and cellular differentiation in Bacillus subtilis Journal
of Bacteriology 1873921ndash3930
205
Sprando RL Collins TFX Black TN Olejnik N Rorie JI Eppley RM Ruggles DI 2005
Characterization of the effect of deoxynivalenol on selected male reproductive endpoints
Food Chemistry and Toxicology 43623ndash635
Starkey DE Ward TJ Aoki T Gale LR Kistler HC Geiser DM Suga H Toacuteth B Varga J
ODonnell K 2007 Global molecular surveillance reveals novel Fusarium head blight
species and trichothecene toxin diversity Fungal Genetics and Biology 441191ndash1204
Stevenson BS Schmidt TM 1997 Growth rate-dependent expression of RNA from plasmid-
borne rRNA operons in Escherichia coli Journal of Bacteriology 1801970ndash1972
Stothard P Wishart DS 2005 Circular genome visualization and exploration using
CGView Bioinformatics 21537ndash539
Suenaga H Ohnuki T Miyazaki K 2007 Functional screening of a metagenomic library for
genes involved in microbial degradation of aromatic compounds Environmental
Microbiology 92289-97
Surai P Mezes M Fotina TI Denev SD 2010 Mycotoxins in Human Diet A Hidden
Danger In Modern Dietary Fat Intakes in Disease Promotion Edited by F De Meester et
al Nutrition and Health DOI 101007978-1-60327-571-2_18
Sutton JC 1982 Epidemiology of wheat head blight and maize ear rot caused by Fusarium
graminearum Canadian Journal of Plant Pathology 4195
Swaving J De Bont JAM Westphal A De Kok A 1996 A novel type pyridine nucleotide-
disulfide oxidoreductase is essential for NAD+- and NADPH-dependent degradation of
epoxyalkanes by Xanthobacter strain Py2 Journal of Bacteriology 1786644ndash6
206
Swanson SP Rood JHD Behrens JC Sanders PE 1987 Preparation and characterization of
the deepoxy trichothecenes deepoxy HT-2 deepoxy T-2 triol deepoxy T-2 tetraol
deepoxy 15-monoacetoxyscirpenol and deepoxy scirpentriol Applied and Environmental
Microbiology 532821ndash2826
Swanson SP Helaszek C Buck WB Rood JHD Haschek WM 1988 The role of intestinal
microflora in the metabolism of trichothecene mycotoxins Food Chemistry and
Toxicology 26823ndash829
Tian F Yu Y Chen B Li H Yao Y-F Guo X-K 2009 Bacterial archaeal and eukaryotic
diversity in Arctic sediment as revealed by 16S rRNA and 18S rRNA gene clone libraries
analysis Polar Biology 3293-103
Tatusov RL Galperin MY Natale DA Koonin EV et al 2000 Artemis sequence
visualization and annotation Bioinformatics 16944-945
Trail F Haixin X Loranger R Gadoury D 2002 Physiological and environmental aspects of
ascospore discharge in Gibberella zeae (anamorph Fusarium graminearum) Mycologia
94181-189
Tsuge K Ohata Y Shoda M 2001 Gene yerP involved in surfactin self-resistance in
Bacillus subtilis Antimicrobial Agents and Chemotherapy 453566ndash3573
Tucker 2001 The ldquoyellow rainrdquo controversy Lesson for arms control compliance The
Nonproliferation Review 825-42
Ueno Y 1985 The toxicology of mycotoxins Critical Reviews in Toxicology 1499-132
Ueno Y Nakayama K Ishii K Tashiro F Minoda Y Omori T Komagata K 1983
Metabolism of T-2 toxin in Curtobacterium sp strain 114-2 Applied and Environmental
Microbiology 46120ndash127
207
van Elsas JD Speksnijder AJ van Overbeek LS 2008 A procedure for the metagenomics
exploration of disease suppressive soils Journal of Microbiological Methods 75515ndash522
van Ginkel CG de Bont JAM 1986 Isolation and characterization of alkene-utilizing
Xanthobacter spp Archives of Microbiology 145403ndash407
van Lanen SG Shen B 2006 Microbial genomics for the improvement of natural product
discovery Current Opinion in Microbiology 9252ndash260
Varga J Toacuteth B 2005 Novel strategies to control mycotoxins in feeds a review Acta
Veterinaria Hungarica 53189-203
Voigt CA Schafer W Salomon S 2005 A secreted lipase of Fusarium graminearum is a
virulence factor required for infection of cereals The Plant Journal 42364ndash375
Vogelgsang S Hecker A Musa T Dorn B Forrer HR 2011 On farm experiments over five
years in a grain maizewinter wheat rotation Effect of maize residue treatments on
Fusarium graminearum infection and deoxynivalenol contamination in wheat In Proc
7th
Canadian Workshop on Fusarium Head Blight edited by J Gilbert A Tekauz N
Sweetland and K Slusarenko Winnipeg Manitoba Canada p 41
Voumllkl A Vogler B Schollenberger M Karlovsky P 2004 Microbial detoxification of
mycotoxin deoxynivalenol Journal of Basic Microbiology 44147ndash156
Wagacha JM Muthomi JW 2007 Fusarium culmorum Infection process mechanisms of
mycotoxin production and their role in pathogenesis in wheat Crop Protection 26877-
885
Wakulinski W 1989 Phytotoxicity of the secondary metabolites of fungi causing wheat head
fusariosis (head blight) Acta Physiologiae Plantarum 11301ndash306
208
Wang JT Chang SC Chen YC Luh KT 2000 Comparison of antimicrobial susceptibility of
Citrobacter freundii isolates in two different time periods The Journal of Microbiology
Immunology and Infection 33258-62
Wannemacher RJ Stanley L Wiener MD 2000 Trichothecenes mycotoxins In Medical
aspects of chemical and biological warfare edited by Zajtchuk R Washington DC
Department of the Army pp 656-676
Westphal AH Swaving J Jacobs L de Kok A 1998 Purification and characterization of a
flavoprotein involved in the degradation of epoxyalkanes by Xanthobacter Py2 European
Journal of Biochemistry 257160ndash168
Whalen JG Mully TW Enlgish JC 2007 Spontaneous Citrobacter freundii infection in an
immunocompetent patient Archives of Dermatology 143124-5
White 2007 The physiology and biochemistry of prokaryotes Third edition Oxford
University Press New York Oxford p 628
Wilcke WF Morey RV 1995 Natural-air corn drying in the upper Midwest University of
Minnesota Minnesota Extension Service Publication BU-6577-E pp 20
Windels CE 2000 Economic and social impacts of Fusarium headblight changing farms and
rural communities in the Northern Great Plains Phytopathology 9017ndash21
Winzeler EA Shoemaker DD Astromoff A Liang H Anderson K Andre B Bangham R
Benito R Boeke JD Bussey H et al 1999 Functional characterization of the S
cerevisiae genome by gene deletion and parallel analysis Science 285901ndash906
Yang GH Jarvis BB Chung YJ Pestka JJ 2000 Apoptosis induction by the satratoxins and
other trichothecene mycotoxins relationship to ERK p38 MAPK and SAPKJNK
activation Toxicology and Applied Pharmacology 164149ndash160
209
Yang Q Yin GM Guo YL Zhang DF Chen SJ Xu ML 2010 A major QTL for resistance
to Gibberella stalk rot in maize Theoretical and Applied Genetics 121673ndash687
Yazar S Omurtag GZ 2008 Fumonisins trichothecenes and zearalenone in cereals
International Journal of Molecular Science 92062ndash2090
Yoshizawa T Morooka N 1973 Deoxynivalenol and its monoacetate new mycotoxins from
Fusaruium roseum and moldy barley Agricultural and Biological Chemistry 372933ndash
2934
Yoshizawa T Takeda H Ohi T 1983 Structure of a novel metabolite from deoxynivalenol a
trichothecene mycotoxin in animals Agricultural and Biological Chemistry 472133ndash
2135
Yoshizawa T 1993 Red-mold diseases and natural occurrence in Japan In Trichothecenes-
Chemistry Biology and Toxicological Aspects edited by Y Ueno Elsevier Amsterdam
Netherlands p 195-209
Yoshizawa T Sakamoto T Okamoto K 1984 In vitro formation of 3rsquo-hydroxy T-2 and 3rsquo-
hydroxy HT-2 toxins from T-2 by liver homogenates from mice and monkeys Applied
and Environmental Microbiology 47130-134
Young JC Subryan LM Potts D McLaren ME Gobran FH 1986 Reduction in levels of
deoxynivalenol in contaminated wheat by chemical and physical treatment Journal of
Agricultural and Food Chemistry 34461ndash465
Young JC Trenholm HL Friend DW Prelusky DB 1987 Detoxification of deoxynivalenol
with sodium bisulfite and evaluation of the effects when pure mycotoxin or contaminated
corn was treated and given to pigs Journal of Agricultural and Food Chemistry 35259ndash
261
210
Young JC Zhu H Zhou T 2006 Degradation of trichothecene mycotoxins by aqueous
ozone Food Chemistry and Toxicology 44417ndash424
Young JC Zhou T Yu H Zhu H Gong J 2007 Degradation of trichothecene mycotoxins by
chicken intestinal microbes Food Chemistry and Toxicology 45136-143
Yu Z Morrison M 2004 Comparisons of different hypervariable regions of rrs genes for use
in fingerprinting of microbial communities by PCR-denaturing gradient gel
electrophoresis Applied and Environmental Microbiology 704800-4806
Yu H Zhou T Gong J Young C Su X Li X Zhu H Tsao R Yang R 2010 Isolation of
deoxynivalenol-transforming bacteria from the chicken intestines using the approach of
PCR-DGGE guided microbial selection BMC Microbiology 10182
Zeibdawi A Steele M Savoie A 2011 An overview of CFIAs mycotoxin monitoring
program in livestock feeds In Proc 7th
Canadian Workshop on Fusarium Head Blight
edited by J Gilbert A Tekauz N Sweetland and K Slusarenko Winnipeg Manitoba
Canada p 25
Zhang J-B Li H-P Dang F-J Qu B Xu Y-B Zhao C-S Lia Y-C 2007 Determination of the
trichothecene mycotoxins chemotypes and associated geographical distribution and
phylogenetic species of the Fusarium graminearum clade from China Mycological
Research 111967ndash975
Zhao JS Manno D Thiboutot S Ampleman G Hawari J 2007 Shewanella canadensis sp
nov and Shewanella atlantica sp nov manganese dioxide- and hexahydro-135-trinitro-
135-triazine-reducing psychrophilic marine bacteria International Journal of Systemic
Evolution and Microbiology 572155ndash2162
211
Zhao X-Q 2011 Genome-based studies of marine microorganisms to maximize the diversity
of natural products discovery for medical treatments Evidence-Based Complementary
and Alternative Medicine Article ID 384572 doi1011552011384572
ZhaoY Park RD Muzzarelli RA 2010 Chitin deacetylases properties and applications
Marine Drugs 824ndash46
Zhou HR Lau AS Pestka JJ 2003a Role of double-stranded RNA-activated protein kinase R
(PKR) in deoxynivalenol-induced ribotoxic stress response Toxicological Science
74335ndash344
Zhou HR Islam Z Pestka JJ 2003b Rapid sequential activation of mitogen-activated protein
kinases and transcription factors precedes proinflammatory cytokine mRNA expression in
spleen of mice exposed to the trichothecene vomitoxin Toxicological Sciences 72130ndash
142
Zhou T He J Gong J 2008 Microbial transformation of trichothecene mycotoxins World
Mycotoxin Journal 123-30
Zhou G Zhang H He Y He L 2010 Identification of a chitin deacetylase producing bacteria
isolated from soil and its fermentation optimization African Journal of Microbiology
Research 42597-2603

vi
TABLE OF CONTENTS
DEDICATIONShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipiii
ACKNOWLEDGEMENTShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipiv
TABLE OF CONTENTShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipvi
LIST OF TABLEShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipix
LIST OF FIGUREShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipx
LIST OF ABBREVIATIONShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipxiii
10 GENERAL INTRODUCTION AND LITERATURE REVIEW helliphelliphelliphelliphelliphelliphelliphellip1
11 Backgroundhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip1
12 Plant pathogenesis and trichothecene productionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip3
13 Biosynthesis of food contaminating TChelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip6
14 Occurrence of TC mycotoxins in food and feed helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip11
15 Significance of deoxynivalenol and trichotheceneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip16
16 Toxic effects of deoxynivalenol and trichotheceneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip19
17 Cellular and molecular mechanisms of DON toxicity in animals and plantshelliphelliphelliphellip22
18 Strategies to control Fusarium head blight and trichotheceneshelliphelliphelliphelliphelliphelliphelliphelliphellip26
19 Isolation of DONTC de-epoxidating micro-organisms and geneshelliphelliphelliphelliphelliphelliphelliphellip28
110 The bacterial species Citrobacter freundiihelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip30
111 Hypotheses and objectiveshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip31
vii
20 AEROBIC AND ANAEROBIC DE-EPOXIDATION OF MYCOTOXIN
DEOXYNIVALENOL BY BACTERIA ORIGINATING FROM AGRICULTURAL
SOIL33
21 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip33
22 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
23 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip37
24 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip45
25 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip62
26 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip64
30 ISOALTION AND CHARACTERIZATION OF A NOVEL SOIL BACTERIUM
CAPABLE OF DETOXIFYING ELEVEN TRICHOTHECENE MYCOTOXINS65
31 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip65
32 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip66
33 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip68
34 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip75
35 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip96
36 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip100
40 CONSTRUCTION AND FUNCTIONAL SCREENING OF GENOMIC DNA
LIBRARY FOR ISOLATION OF DON DE-EPOXYDIZING GENE(S) FROM
BACTERIAL STRAIN ADS47helliphelliphelliphelliphelliphelliphelliphellip102
41 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip102
42 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip103
43 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip104
viii
44 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip112
45 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip118
46 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip121
50 GENOME SEQUENCING CHARACTERIZATION AND COMPARATIVE
ANALYSIS OF A TRICHOTHECENE MYCOTOXIN DETOXIFYING
CITROBACTER FREUNDII STRAIN ADS47helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip122
51 Abstract helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip122
52 Introduction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip123
53 Materials and methodshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip125
54 Results helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip129
55 Discussionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip162
56 Acknowledgementshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip168
60 CONCLUSIONhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip171
61 Summary helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip171
62 Research contributions helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip174
63 Future workhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip175
70 REFERENCEShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip176
ix
LIST OF TABLES
Table 1-1 Fusaria produced type-A and type-B trichothecene mycotoxinshelliphelliphelliphelliphelliphelliphellip9
Table 1-2 Worldwide occurrence of deoxynivalenol (DON) in animal feedshelliphelliphelliphelliphellip12
Table 1-3 Occurrence of DON in Canadian grains grains foods and beershelliphelliphelliphelliphelliphelliphellip13
Table 1-4 Legislative maximum tolerated levels of trichothecene mycotoxins in Canadahellip14
Table 1-5 In vitro and in vivo synergistic effects of different food contaminating
mycotoxins helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip18
Table 3-1 Microbial transformation of type-A and type-B tricthothecene mycotoxins
after 72 h of incubation under aerobic conditions and at room temperaturehelliphelliphelliphelliphelliphellip95
Table 4-1 Analysis of Fosmid DNA library for deoxynivalenol (DON) de-epoxidation
capability helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip117
Table 5-1 General features of C freundii strain ADS47 genomehelliphelliphelliphelliphelliphelliphelliphelliphellip132
Table 5-2 Genome comparisons between strain ADS47 and five Citrobacter species141
Table 5-3 Functional categorized gene abundances among the six Citrobacter genomeshellip145
Table 5-4 Unique genes of C freundii strain ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip156
Supplemental Table S5-1 List of unique genes of strain ADS47 gene IDs and their
functional roleshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip169
x
LIST OF FIGURES
Figure 1-1 Structures of trichothecene mycotoxins produced by Fusariahelliphelliphelliphelliphelliphelliphelliphellip7
Figure 1-2 Cellular and molecular mechanisms of deoxynivalenol toxicity in
eukaryotic organismshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip23
Figure 2-1 Mechanism of microbial DON de-epoxidationhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip36
Figure 2-2 LC-UV-MS chromatographic analysis of DON and its de-epoxidated product
dE-DON obtained by culturing with selected soil microbeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip46
Figure 2-3 Time course of relative biotransformation of DON to dE-DON by the initial
soil suspension culture in MSB mediumhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip49
Figure 2-4 Microbial de-epoxidation of DON after five days of incubationhelliphelliphelliphelliphelliphellip51
Figure 2-5 Analysis of microbial population reduction by terminal restriction fragment
length polymorphism (T-RFLP)helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip53
Figure 2-6 Microbial genera identified in the enriched DON de-epoxidizing microbial
culturehelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip56
Figure 2-7 Effects of culture conditions on DON to dE-DON transformation by the
enriched microbial culturehelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip58
Figure 2-8 Microbial DON to dE (de-epoxy)-DON transformation activity under aerobic
and anaerobic conditionshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip61
Figure 3-1 Colony color shape and size of the mycotoxin degrading bacterial strain
ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip76
Figure 3-2 Dendrogram based on 16S-rDNA sequences for strain ADS47 and 11
reference Citrobacter specieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip77
xi
Figure 3-3 Effects of DON concentration on bacterial growth and de-epoxidation
activitieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip79
Figure 3-4 Bacterial growth and DON de-epoxidation capabilities under aerobic and
anaerobic conditions helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip82
Figure 3-5 Effects of media on bacterial growth and DON de-epoxidation activityhellip85
Figure3-6 Effects of temperature on microbial growth and DON de-epoxydation helliphelliphellip87
Figure 3-7 Influence of pH on microbial growth and DON de-epoxydation capabilityhellip89
Figure 3-8 Effects of sodium azide (SA) on bacterial DON de-epoxidation activityhelliphelliphellip91
Figure 3-9 Determination of DON to deepoxy-DON transformation by bacterial culture
filtrate and cell extracthelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip92
Figure 3-10 Determination of microbial de-epoxidation and de-acetylation of a type-A
trichothecene mycotoxin (HT-2 toxin) by LC-UV-MShelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip94
Supplemental Figure S3-1 Flow chart depicted the procedures of enrichment and isolation
of DON de-epoxidation bacterial strain ADS47helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip101
Figure 4-1 Schematic overview of the construction and functional screening of Fosmid
DNA libraryhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip105
Figure 4-2 Genetic map of copy controlled Fosmid vector (pCC1FOS)helliphelliphelliphelliphelliphelliphellip108
Figure 4-3 Preparation of 40 kb size insert DNA from a DON de-epoxidizing bacterium113
Figure 4-4 Production of genomic DNA library in E coli EPI300-T1 host strainhelliphelliphelliphellip114
Figure 4-5 Confirmation of the Fosmid DNA library construction in E coli helliphelliphelliphelliphellip 116
Figure 5-1 Co-linearity between C freundii strain ADS47 genome and C koseri helliphelliphellip129
Figure 5-2 Circular genome map of C freundii strain ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphellip133
xii
Figure 5-3 Size distribution of the predicted genes identified in C freundii strain
ADS47helliphelliphellip 135
Figure 5-4 Functional classification of the predicted coding sequences (CDS) of
C freundii strain ADS47 helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip137
Figure 5-5 Phylogeny of the trichothecene detoxifying C freundii strain ADS47 139
Figure 5-6 Comparisons of coding sequences between C freundii strain ADS47 and
five Citrobacter specieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip143
Figure 5-7 Determination of the locus enterocyte effacement (LEE) operons in
Citrobacter species by comparing C rodentium helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip151
Figure 5-8 Locations of unique genes of C freundii strain ADS467helliphelliphelliphelliphelliphelliphelliphellip157
Figure 5-9 Unique genomic regions and associated genes in strain ADS47 compared
to five Citrobacter specieshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip159
xiii
LIST OF ABBREVIATIONS
APCI= atmospheric pressure chemical ionization
AE= attaching and effacing
A= aerobic conditions
ABC= ATP-binding cassette
ACT= artemis comparative tools
AN= anaerobic conditions
3-ADON= 3-acetyl-deoxynivalenol
15-ADON= 15-acetyl-deoxynivalenol
ATA= alimentary toxic aleukia
bp= base pair
BGI= Beijing genomics institute
BLAST= basic local alignment search tool
BRIG= blast ring image generator
Bw= body weight
C= carbon
CDS= coding sequence
CFIA= Canadian food inspection agency
CHEF= clamped homogeneous electric fields
CMB= corn meal broth
CVTree= composition vector tree
DAS= diacetoxyscirpenol
dH2O= distilled water
DOGT1= UDP-glucosyltransferase
DON= deoxynivalenol
dsRNA= double stranded RNA
dE= de-epoxy
dE-DON= de-epoxy DON
E coli= Escherichia coli
ER=endoplasmic reticulum
FAO= Food amp Agriculture Organization
FM= full medium
FHB= Fusarium head blight
FUS= fusarenonX
gDNA= genomic DNA
GC= guanine cytosine
GER= Gebberella ear rot
GPI= glycosylphosphatidylinositol
h= hour
Hck= hematopoietic cell kinase
Kb= kilobase
xiv
Kg= kilogram
LC-UV-MS= liquid chromatography-ultraviolet-
mass spectrometry
LEE= locus of enterocyte effacement
LD50= lethal dose of 50 animal
LB= Luria Bertani
LMP= low melting point
MAPK= mitogen-activated protein kinase
microg= microgram
mg= milligram
mL= milliliter
microL= microliter
microM= micromole
min= minute
Mb= megabases
MCS=multiple cloning site
MSA= MSB+ agar
MSB= mineral salts broth
MM= minimal medium
MSC= mineral salts + 03 Bacto Peptone
MS-DON= mineral salts + 200 microgmL DON
mRNA= messenger RNA
MSY= mineral salts + yeast extract
mz= mass ion ratio
N= nitrogen
Na-Cl= sodiumndashclorine
NADH= (reduced) nicotinamide adenine
dinucleotide
NAD(P)= nicotinamide adenine
dinucleotide phosphate
NCBI= national center for biotechnology
information
NEO= neosolaniol
ng= nanogram
NIV= nivalenol
OD600nm= optical density at 600
nanometer wavelength
OMAFRA= Ontario Ministry of
Agriculture and Rural Affairs
Paa= phenylacetic acid
PCR= polymerase chain reaction
PDB= potato dextrose broth
ppm= parts per million
PFGE= pulsed field gel electrophoresis
PKR= protein kinase
xv
PNDO= pyridine nucleotide-disulfide
oxidoreductase
QTL= quantitative trait locus
RAST= rapid annotation subsystem technology
RDP= ribosomal database project
Rpl3= 60S ribosomal protein L3
rRNA= ribosomal RNA
s= second
SA= sodium azide
SDR= short chain dehydrogenasereductase
SE= soil extract
SIM= selected ion monitoring
SOAP= short oligonucleotide analysis package
sp= species
sRNA= small RNA
TC= trichothecene
TCA= tricarboxylic acid cycle
TSB= tryptic soy broth
TF= transcription factor
tRNA= transfer RNA
TIC= total ion count
TRAP= tripartite ATP-independent
periplasmic
T-RFLP=terminal restriction fragment
length polymorphism
TRI101= trichothecene 3-O-
acetyltransferase
YEB= yeast extract broth
VER= verrucarol
VKOR= vitamin K epoxide reductase
wv= weight by volume
1
CHAPTER 1
10 GENERAL INTRODUCTION AND LITERATURE REVIEW
11 Background
According to the Food and Agriculture Organization (FAO) of the United Nations
more than 25 of the worldrsquos food crops are contaminated with mycotoxins every year
causing serious adverse effects on livestock industry crop production and international trade
(Boutrif and Canet 1998 Desjardins 2006 Binder et al 2007 Surai et al 2010) The negative
effects of mycotoxins on the consumers are diverse including organ toxicity mutagenicity
carcinogenicity neurotoxicity teratogenicity modulation of the immune system reproductive
system effects and growth retardation Mycotoxins are toxic secondary metabolites produced
by various fungal species during crop colonization in field andor post harvest conditions
(FAO 1991 Barrett 2000) Aspergillus Penicillum and Fusarium are the predominant fungal
genera that accumulate economically important mycotoxins such as aflatoxins zearalenone
trichothecenes fumonisins ochratoxins and ergot alkaloids in agricultural commodities
Humans and animals are principally exposed to mycotoxins through consumption of
contaminated foodfeed and animal products such as milk meat and eggs (Fink-Gremmels
1989 Hollinger and Ekperigin 1999)
Trichothecenes (TCs) produced by a variety of toxigenic Fusarium species are
probably the most food and feed contaminating mycotoxins in the temperate regions of
America Europe and Asia (Creppy 2002 Yazar and Omurtag 2008) The fungal pathogen
accumulates various type-A (eg T-2 toxin HT-2 toxin) and type-B trichothecenes (eg
2
deoxynivalenol nivalenol and their derivatives) in cereals Among them type-B
trichothecenes particularly deoxynivalenol (DON) andor its derivatives appear to be the
most commonly found mycotoxins in grain and grain products worldwide impairing human
health and productivity of farm animals (Placinta et al 1999 Sherif et al 2009) A number of
human and animal disease outbreaks associated with TC contaminated grain food and feed
consumption have been reported in Asia (Japan India and central Asia) Europe New
Zealand and United States of America (USA) (Pestka and Smolinski 2005 Desjardins 2006)
A severe outbreak of Alimentary Toxic Aleukia (ATA) mycotoxicosis killed 100000 people
in the Orenburg region of Russia (Eriksen 2003) In 1996 losses of over $100 million were
estimated in Ontario because of costs related to managing DON (Schaafsma 2002)
Historically T-2 toxin DON and nivalenol (NIV) have likely been used as a biological
warfare in Laos Kampuchea and Afghanistan and there is concern because of the potential
use of these compounds for bioterrorism (Wannemacher et al 2000 Tucker 2001)
Despite the extensive efforts that have been made to prevent TC contamination with
the current varieties and production practices its presence in agricultural commodities is
likely unavoidable There are no crop cultivars currently available that are completely
resistant to Fusarium head blightGibberella ear rot (FHBGER) diseases Chemical or
physical methods cannot safely remove TC mycotoxins from grain completely or they are not
economically viable (Varga and Toacuteth 2005) Therefore an alternative strategy to address the
issue is of great importance (Awad et al 2010 Karlovsky 2011)
3
12 Plant pathogenesis and trichothecene production
Cereals infested by the ascomyceteous fungi may cause Gibberella ear rot (GER) in
maize (Zea mays L) and Fusarium head blight (FHB also known as lsquoscabrsquo) of small grain
cereals [wheat (Triticum aestivum L) barley (Hordeum vulgare L)] and other small grain
cereals (Goswami and Kistler 2004) As many as 17 fungal species have been reported to be
associated with FHBGER which include F graminearum Schwabe [telemorph Gibberella
zeae (Schwein) Petch] F culmorum (WG Smith) Sacc F sporotrichioides Scherb F
poae (Peck) Wollenw Microdochium nivale (Fr) Samuels amp IC Hallett and F avenaceum
(Fr) Sacc (Osborne and Stein 2007 Audenaert et al 2009) The two species F
graminearum and F culmorum are the principal species causing crop damage Among them
F graminearum is the widely adapted and most damaging species across the world including
North America (Dyer et al 2006) F culmorum tend to be the main species in the cooler parts
of North Central and Western Europe (Wagacha and Muthomi 2007)
The fungi overwinter mainly on crop residues as saprophytes where they produce
asexual (micro- and macro-conidia) andor sexual (ascospores) fruiting structures which act
as the initial source of inoculums in the subsequent crop season (Sutton 1982 Wagacha and
Muthomi 2007) Ascospores are primarily dispersed by air (Trail et al 2002) and conidia
move by rain-splash (Paul et al 2004) After they land on maize silks and florets of small
grain cereals infection occurs through natural openings (eg stomata) or occasionally by
direct penetration The pathogens can invade maize kernels through wounds caused in the
husks by insects or birds Infection may occur throughout the crop growing season but
disease incidence and severity is facilitated by warm temperatures (gt20degC) high humidity
(gt80) and frequent precipitation that coincides with anthesis (Osborne and Stein 2007
4
Wagacha and Muthomi 2007) The Fusarium pathogen initially infects as biotroph which
becomes a necrotroph as the disease progresses After penetration Fusaria produce different
extracellular cell wall degrading enzymes that trigger infection and hyphal progression
towards the developing earkernel where further colonization occurs and development of
FHBGER diseases (Voigt et al 2005 Miller et al 2007 Kikot et al 2009) FHB disease is
characterized by a bleached appearance around the middle of the head Severely infected
grains have a pinkish discoloration and can be shriveled In maize the disease produces a
reddishndashbrown discoloration of the kernels and progresses from the tip to the bottom of
affected ears
The fungi associated with FHB and GER contaminate cereals by accumulating various
host-nonspecific TC mycotoxins (Brennen et al 2007) TCs have typically been associated
with cereals grown in temperate climatic conditions including Canada (Doohan et al 2003)
DON production by F graminearum and F culmorum is maximal at warm temperatures (22-
28˚C) high grain moisture contents (gt17) and humid conditions (gt70 RH) (Martins and
Martins 2002 Hope et al 2005 Llorens et al 2006) Grain can be contaminated with TCs in
the field as well as in storage if conditions for fungal growth are optimal and grain contains
high moisture (gt17) (Larsen et al 2004 Birzele et al 2002)
Populations of FHBGER causing pathogens are dynamic Genetic variability between
and within Fusarium species appears to have occurred to allow for their adaptation to new
agro-climates (Desjardins 2006 Fernando et al 2011) Three strain specific Fusarium
chemotypes NIV 3-ADON (3-acetyl-deoxynivalenol) and 15-ADON (15- acetyl-
deoxynivalenol) have been identified which correlate with the geographical distributions
(Starkey et al 2007 Zhang et al 2007 Audenaert et al 2009) The NIV chemotype mainly
5
produced by F culmorum accumulates NIV and 4-NIV (Fusarinon X) is predominant in
Asia Africa and Europe (Lee et al 2001) The 3-ADON chemotype which synthesizes DON
and 3-ADON is the dominant strain in EuropeFar East (Larsen et al 2004) In North
America 15-ADON is the main chemotype which produces DON and 15-ADON (Goswami
and Kistler 2004) Significant Fusarium population shifting was observed in the past and in
recent years (Kant et al 2011) A dynamic change from F culmorum to F gramninearum
and DON to NIV producing chemotyes occurred in North West European countries The 3-
ADON chemotype which is uncommon in North America was found in North Dakota and
Minnesota (Goswami and Kistler 2004) F pseudograminearum which historically occurs in
Australia Africa and North West Pacific America has been found in Western Canada (Larsen
et al 2004 Kant et al 2011) In Manitoba a replacement of 15-ADON chemotype by the
more phytoxic 3-ADON type was observed in recent years (Fernando et al 2011) All
together the FHBGER associated pathogen complex is highly dynamic which not only
reduces crop yield but also decreases grain quality by accumulating various mycotoxins
6
13 Biosynthesis of food contaminating TC
TCs are fungal secondary metabolites that were initially identified in Trichotheceium
roseum Thus the name of this class of compounds follows the name of the genus
Trichotheceium (Freeman and Morrison 1949) There have been 180 TCs identified in various
fungal species belonging to the genera Fusarium Myrothecium Stachybotrys Trichoderma
and Thrichothecium (Eriksen 2003 Eriksen et al 2004) The FHBGER associated TCs are
mainly synthesized by different Fusaria and are characterized as tricyclic sesquiterpenoids
with a double bond at position C9-10 and a C12-13 epoxide ring
7
Figure 1-1 Structures of trichothecene mycotoxins produced by Fusaria (a) General
structure of trichothecenes R1 = H OH OiValeryl keto R2 = H OH R3= OH OAc R4 =
H OH OAc R5= OH OAc (b) Type-A trichothecene mycotoxin (c) Type-B trichothecene
mycotoxin which contain a carbonyl functional group at C8 position (Young et al 2007)
(a) (b)
(c)
8
The fungal metabolites are small in size (molecular weight 250-550) heat stable and
non-volatile molecules (Pestka 2007 Yazar and Omurtag 2008) Food contaminating TCs are
divided into two groups type-A and type-B (Figure 1-1b and 1c) Type-A is characterized by
the lack of a ketone function at C8 position and include T-2 toxin HT-2 toxin
diacetoxyscirpenol (DAS) and neosolaniol (NEO) which are mainly produced by F
sporotrichioides and F poae (Creppy 2002 Quarta et al 2006) The most frequently found
TC belongs to type-B which contains carbonyl functions at C8 positions DON NIV and
their acetylated derivatives The two types are further differentiated by the presence or
absence of functional hydroxyl (-OH) and acetyl (CH3-CO-) groups at C-3 C-4 C-7 C-8 and
C-15 positions (Table 1-1)
The syntheses of various TCs are speciesstrain specific and are genetically controlled
but influenced by environmental conditions (Desjardins 2006 Paterson and Lima 2010) Until
now 15 Fusaria catalytic genes and regulators involved in TC productions have been
identified (Merhej et al 2011) These biosynthetic genes are located at three loci on different
chromosomes The core trichothecene (TRI) gene cluster is comprised of 12 genes localized
in a 25-kb genomic region (Kimura et al 2001) The remaining three genes are encoded by
the TRI1-TRI16 and TRI101 loci (Gale et al 2005 Alexander et al 2009) Fusaria produce
TC following a number of oxygenation isomerization acetylation hydroxylation and
esterification reaction steps (Desjardins 2006 Desjardins and Proctor 2007 Kimura et al
2007 Merhej et al 2011) In brief farnesyl pyrophosphate is initially cyclized into
trichodiene by TRI5 which codes for trichodiene synthase In the next four steps
isotrichotriol is produced by TRI4 (a multifunctional cytochrome P450) via C-2 hydroxylation
and 1213 epoxidation followed by two hydroxylation reactions
9
Table 1-1 Fusaria produced type-A and type-B trichothecene mycotoxins Molecular weight
(MW) and acetyl (-R) hydroxyl (-OH) and ketone (=O) functions at different carbon (C)
position of the tricyclic rings A-types T-2 toxin HT-2 toxin DAS (diacetoxyscirpenol)
NEO (neosolaniol) VER (verrucarol) B-type DON (deoxynivalenol) 3-ADON (3-acetyl-
deoxynivalenol) 15-ADON (15-acetyl-deoxynivalenol) NIV (nivalenol) and FusX
(fusarenon X) (Young et al 2007)
Trichothecenes MW R1C3 R2C4 R4C7 R3C15 R5C8
Type-A
T-2 toxin 466 OH OCOCH3 H OCOCH3 OCOCH2CH(CH3)2
HT-2 toxin 424 OH OH H OCOCH3 OCOCH2CH(CH3)2
DAS 366 OH OCOCH3 H OCOCH3 H
NEO 382 OH OCOCH3 OH OCOCH3 H
VER 266 H OH H OH H
Type-B
DON 296 OH H OH OH =O
3-ADON 338 OCOCH3 H OH OH =O
15-ADON 338 OH H OH OCOCH3 =O
NIV 312 OH OH OH OH =O
FusX 354 OH OCOCH3 OH OH =O
10
The next two non-enzymatic reactions results in isotrichodemol Eventually the
trichothecene skeleton is produced via the formation of a C-O bond between the C-2 oxygen
and C-11 Subsequent acetylation at C-3 by TRI101 hydroxylation at C-15 by TRI1 and
catalytic activity of TRI3 leads to calonectrin production The pathway described above is
common for both type-A (eg T-2 toxin HT-2 toxin) and type-B (eg DON NIV and their
acetylated derivatives) TCs
The following steps are speciesstrain specific In DON producing F graminearum
strains calonectrin is converted to either 3-ADON or 15-ADON followed by DON production
by the catabolic activities of TRI1 and TRI8 The biosynthesis of 3-ADON or 15-ADON is
controlled by variation in the esterase coding sequences of TRI8 F graminearum produces
NIV from calonectrin via synthesis of 4-NIV followed by NIV by TRI1 and TRI8 gene
actions NIV can be produced either from DON by hydroxylation at C-4 or via
transformation of calonectrin to 315-diacetoxyscirpenol (315-DAS) followed by addition of
a ketone at C-8 position by the activity of the TRI7 and TRI13 genes 315-DAS is the
principal substrate for synthesis of type-A TCs by F sporotrichioides the main type-A
producer The biosynthetic pathway of T-2 toxin involves C-4 acetylation of 315-DAS by the
TRI7 gene product and TRI8 triggered by TRI1 315-DAS is also the substrate for HT-2
toxin synthesis In the absence of TRI7 expression HT-2 toxin is produced through
hydroxylation of 315-DAS and isovalerate addition to C-8 position (Foroud and Eudes 2009)
11
14 Occurrence of TC mycotoxins in food and feed
TC producing Fusaria are adapted to variable climatic conditions and agricultural
ecosystems As a consequence this class of mycotoxins particularly DON is frequently
detected in foods and feeds including breakfast cereals bakery products snack foods beer
and animal feed derived from cereals in most part of the world (Pittet 2001 Desjardins 2006
Lancova et al 2008) A two-year survey during 2003-2005 conducted by Romer Lab Inc
revealed unacceptable levels of DON in animal feedsfeed raw materials (such as maize
wheat wheat bran) in Asia and Oceania (1062 microgg) central Europe (802 microgg) Mid-West
USA (55 microgg) northern Europe (551 microgg) and southern Europe (303 microgg) (Binder et al
2007) (Table 1-2)
Quite high levels of DON contamination (19 microgg) were detected in north Asian and
Chinese feedfeed materials Recently five-year on-farm studies in Switzerland showed that
74 of the wheat grain and straw samples were contaminated with high levels of DON
(average 5 microgg) The amount is higher than the European maximum limit of 125 microgg
(Vogelgsang et al 2011)
The occurrences of TCs in Canadian agricultural commodities are indicated in Table
1-3 Seventeen surveys conducted during the 1980s and 1990s showed up to 100 DON
incidence in Ontario wheat and maize samples with an 18 average incidence (Scott 1997) A
three-year survey recently (2006-08) conducted by the Ontario Ministry of Agriculture and
Rural Affairs (OMAFRA) detected high levels of DON in spring (maximum 26 microgg) and
winter wheat (maximum 29 microgg) samples (Ana Matu personal communication) The DON
levels were much higher than Health Canadarsquos maximum limit of 2 microgg for uncleaned soft
wheat for staple foods (Table 1-4)
12
Table 1-2 Worldwide occurrence of DON in animal feed Feed and feed raw material
samples were collected from animal farmsfeed production sites during 2003-2005 The
survey was conducted by Romer Labs Inc (httpwwwromerlabscom)
Country
region
Sample
type
Incidence
()
Maximum
DON content
(microgg)
Mean DON
content
(microgg)
Survey
period
Source
North
AsiaChina
Animal
feed
071 1899 092
Binder et
al 2007
South East
Asia
014 152 016
South Asia 070 1062 049
Oceania 2003-05
Australia 041 186 030
Northern
Europe
070 551 059
Central
Europe
066 802 057
Southern
Europe
052 303 030
13
Table 1-3 Occurrence of DON in Canadian grains grain foods and beers Infant cereal foods
oat barley soy and multi-grain based foods
Province Sample
type
Incidence
()
Maximum
DON
contents
(microgg)
Multiple
mycotoxins
samples ()
Survey
period
Sources
Ontario Spring
Wheat
70 260 Yes 2006-08 OMAFRA1
Ontario Winter
wheat
49 290 Yes 2006-08 OMAFRA
Ontario Maize 78 140 Yes 2009-10 Limay-Rios
and
Schaafsma
2010
Ontario Maize - 1000 Yes 2011 AampL
Biologicals
Canada Breakfast
cereals
46 094 430 1999-01 Roscoe et al
2008
Canada Animal
feeds
73 425 Yes 1990-10 Zeibdawi et
al 2011
Canada Beers 58 0052 50 1990s Scott 1997
Canada Infant
Cereals
63 098 Yes 1997-99 Lombaert et
al 2003
1
Ontario Ministry of Agriculture and Rural Affairs
2 microgmL
14
Table 1-4 Legislative maximum tolerated levels of trichothecene mycotoxins in Canada
according to Canadian Food Inspection Fact Sheet April 23 2009 DON- deoxynivalenol
DAS- diacetoxyscirpenol
Products DON
(microgg)
T-2 toxin
(microgg)
HT-2 toxin
(microgg)
DAS
(microgg)
Uncleaned soft wheat for human
consumption
2 - - -
Cattlepoultry diets 5 lt1 01 lt1
Swinedairy cattle feeds 1 lt1 0025 lt2
15
High DON incidence (78) was observed in maize samples collected from
southwestern Ontario fields with moderate levels (lt1 microgg) of contamination during 2009 and
2010 (Limay-Rios and Schaafsma 2010) Some samples had high DON levels (14 microgg) In
the 2011 crop season extremely high levels of DON (gt100 microgg) were found in some
southern Ontario maize samples (AampL Biologicals London ON Personal communication)
In a Canadian Food Inspection Agency (CFIA) 20 year survey (1990-2010) 73 of
animal feed samples collected from feed mills farms and retail stores across Canada were
contaminated with DON and for more than 25 of the samples the mycotoxin levels were
above the maximum limit (ge 1 microgg) (Zeibdawi et al 2011) The highest level of DON (15
microgg) detected in the 2006-2007 samples were much lower than the levels found in the 1999-
2000 samples which contained a maximum of 425 microgg DON Importantly this study
reported a significant number of samples contaminated with multiple mycotoxins DON NIV
3-ADON 15-ADON T-2 toxin HT-2 toxin DAS and NEO
Earlier studies in the 1990s detected high levels of DON content in Eastern Canadian
cereals maize (maximum 175 microgg) wheat (maximum 916 microgg) barley (maximum 911
microgg) and oats (maximum 12 microgg) (Campbell et al 2002) The study identified a significant
number of samples (15-367) that contained multiple mycotoxins and up to 33 of the
wheat samples exceeded 1 microgg DON Low amounts but high incidences of TCs were found
in cereal-based foods collected from Canadian retail stores Sixty three percent of infant
cereal foods 46 of breakfast cereals and 58 of beers were contaminated with one or
multiple mycotoxins (Scott 1997 Lombaert et al 2003 Roscoe et al 2008) The maximum
amount of DON was found in infant foods (098 microgg) followed by breakfast cereals (094
microgg) and beers (005 microgmL)
16
15 Significance of deoxynivalenol and trichothecenes
For most cereals including wheat maize rice barley oats and rye the risk of
contamination with TCs resulted in grain quality reduction and FHBGER disease progression
(Bai et al 2002 Jansen et al 2005) The impacts of TC contamination in agricultural
commodities are diverse including reduced livestock production induced human illnesses
and enhanced FHB disease severity which may result in the reduction of crop yield Exposure
of farm animals (particularly pigs) to DON is a continuing threat to the livestock industries in
Canada USA and Europe (DrsquoMello et al 1999) Ingestion of TC-contaminated feeds
generally causes reduction of feed intake feed refusal poor feed conversion retardation of
weight gain increased disease susceptibility and reduced reproductive capacities (Binder et al
2007) High doses of DON result in acute toxic effects in pigs (eg emesis) while low doses
(ge1 microgmL) cause chronic effects (eg growth retardation in pigs) (Rotter et al 1996
Accensi et al 2006) Reduction of growth and reproduction ability was observed in
experimental rats exposured to much higher doses of TC (Sprando et al 2005 Collins et al
2006) Several animal disease outbreaks that occurred in different continents have been
attributed to consumption of feed contaminated with DON NIV and DAS (Desjardins 2006)
The consequences of human exposure to TCs include liver damage gastricintestinal
lesions and central nervous system toxicity resulting in anorexia nausea hypotensionshock
vomiting diarrhea and death (Pestka 2007) Human mycotoxicosis (illnesses and outbreaks)
by consumption of type-A and type-B TC mycotoxins in Europe New Zealand Russia
Japan South America India and Vietnam have been reported (Bhat et al 1989 Yoshizawa
1993 Luo 1994 Desjardins 2006) Severe outbreaks of Alimentary Toxic Aleukia (ATA)
associated with TC killed thousands of people in the Orenburg region of Russia during the
17
1930rsquos and 1940rsquos (Eriksen 2003) TCs (T-2 DON and NIV) were likely used for biological
warfare in Laos Kampuchea and Afghanistan and its potential application for bioterrorism is
an additional concern associated with TC (Tucker 2001 Bennett and Klich 2003)
TCs are phytotoxic and may contribute to FHB disease severity in cereals (Proctor et
al 1995 Maier et al 2006 Masuda et al 2007) Studies demonstrated that DON caused
wilting chlorosis inhibition of seedling growth and necrosis in cereals (Cossette and Miller
1995 Mitterbauer et al 2004 Rocha et al 2005) DON-mediated host cell damage and
subsequent necrotrophic fungal spread has been shown to lead to FHBGER disease
development and higher kernel damage (Procter et al 1995 Mitterbauer et al 2004 Maier et
al 2005) This suggests that DON contributes to yield losses as well as to reductions in the
market value of cereals (Osborne and Stein 2007) Severely infected grain may contain more
than 60 microgg DON which is unacceptable to the milling and brewing industries (Okubara et
al 2002) Significant crop losses due to FHB disease epidemics and mycotoxin contamination
have been reported in Ontario (Schaafsma 2002) FHB disease epidemics during 1998 to 2000
caused huge crop losses resulting in farm failures in the northern Great Plains in the USA
(Windels 2000) In that period FHB associated losses were estimated at US $ 27 billion in
the region (Nganje et al 2002)
Importantly Fusaria infested grains are often co-contaminated with multiple
mycotoxins which further increase health risks as interactions between the mycotoxins may
cause greater toxic effects than expected (Schollenberger et al 2006) This implies that an
apparently low level of contamination of a single mycotoxin can have severe effects on
consumers because of synergistic effects among multiple co-occuring mycotoxins (Surai et al
2010)
18
A number of studies demonstrated synergistic toxic effects among Fusarium
mycotoxins on pigs turkeys lamb chicks as well as in in vitro Caco-2 cell and yeast
bioassays (Table 1-5)
Table 1-5 In vitro and in vivo synergistic effects of different food contaminating mycotoxins
DON-deoxynivalenol DAS-diacetoxyscirpenol FB1-fumonisins 1 NIV-nivalenol ZEN-
zearelenone FA-fusaric acid Afl-aflatoxin
Mycotoxins source Interacting
mycotoxins
Experimental
animalcells
Effects Reference
Naturally contaminated
cereals
DON FA Pigs Inhibit growth Smith et al
1997
DON contaminated
wheat and FB1
DON FB1 Pigs Weight loss Harvey et al
1996
Culture materials
contained mycotoxins
T-2 FB1 Turkey poults Weight loss oral
lesion
Kubena et al
1997
Inoculated rice and
pure mycotoxin
DAS Afl Lamb Weight reduction Harvey et al
1995
Naturally DON
contaminated wheat
and pure T-2 toxin
DON T-2 Chicks Reduced body
weight significantly
Kubena et al
1989
Pure mycotoxins DAS Afl Chicks Reduced body
weight
Kubena et al
1993
Pure mycotoxins T-2 Afl Chicks Reduced weight
oral lesion
Kubena et al
1990
Pure mycotoxins DON
ZENFB1
Human intestinal
Caco-2 cells
Increased DNA
fragmentations
Kouadio et al
2007
Pure mycotoxins DON NIV Yeast bioassay Synergistic effects Madhyastha et
al 1994
Pure mycotoxins T-2 HT-2 Yeast bioassay Synergistic effects Madhyastha et
al 1994
19
16 Toxic effects of deoxynivalnol and trichothecenes
As small molecules TCs simply enter into cells and interact with different cell
components resulting in disruption of cellular functions and causing various toxic effects that
include inhibition of protein synthesis and ribosomal fragmentation (Eudes et al 2000
Minervini et al 2004 Pestka 2007) Simple absorption of DON into human intestinal Caco-2
cells and crossing human placenta barriers through passive diffusion has been reported
(Sergent et al 2006 Nielsen et al 2011)
TC may cause various toxicities in animals and plants including apoptosis
immunotoxicity chromatin condensation disruption of cellularmitochondrion membranes
disruption of gut tissues inhibition of protein and ribosome synthesis breakage of ribosome
and cell damage The toxicity of TC is influenced by structural variability (ie position
number and type of functional groups attached to the tricyclic nucleus and C-C double bond at
C9-10 position) (Desjardins 2006) Nevertheless epoxide function at the C12-13 position is
the key for TC cytotoxicity (Swanson et al 1988 Eriksen et al 2004)
Different in vitro and in vivo studies revealed that the method and period of exposure
amount of TC and types of celltissue and host species have significant effects on TCrsquos
cytotoxicity (Scientific Committee for Food 2002 Rocha et al 2005 Nielsen et al 2009)
Administration of 10 mgkg body weight (bw) NIV caused apoptosis in mice thymus Peyerrsquos
patch and spleen tissues (Poapolathep et al 2002) DON and NIV at 887 mgkg bw for 3 days
a week for four weeks induced immunotoxicity in experimental mice determined by
estimating increased immunoglobulins (IgA) and decreased IgM levels (Gouze et al 2007)
Exposure of mice to 1 mgkg bw of DON induced skeletal malformations and reduced mating
tendency while significant resorption of embryos observed at the dose of gt10 mgkg bw
20
(Khera et al 1982) At high acute doses (4972 mgkg bw) DONNIV and type-A
trichothecenes caused mice death because of intestinal haemorrhage necrosis of bone
marrowlymphoid tissues and kidneyheart lesions (Yoshizawa and Morooka 1973
Yoshizawa et al 1994 Forsell et al 1987 Ryu et al 1988) In a separate study 15-ADON
showed more toxicity than DON when the mycotoxins were orally administered to mice
(Forsell et al 1987)
Studies suggested that type-A TCs are more toxic in mammals than type-B which
means that functional variability at the C-8 position of TC has a significant role in toxicity
The toxicity of various TC mycotoxins in animals can be ranked in the order of T-2
toxingtHT-2 toxingtNIVgtDONge3-ADONgt15-ADON according to the results from human cell
lines and mice model systems bioassays (Ueno et al 1983 Pestka et al 1987 Nielsen et al
2009) Moreover animal feeding assays demonstrated that monogastric animals are most
sensitive to DON The order of sensitivity of animals to DON is
pigsgtrodentsgtdogsgtcatsgtpoultrygtruminants (Pestka 2005) The reasons for the variations in
DON toxicity species might be due to differences in metabolism absorption distribution and
elimination of the mycotoxin among animal species (Pestka 2007)
To my knowledge the mechanism of TC uptake by plant cells has not been studied
Nonetheless cereals are exposed to TC by toxigenic Fusaria infection which may cause
various toxicities such as cell damage wilting chlorosis necrosis program cell death and
inhibition of seedling growth in sensitive plants (eg wheat maize and barley) (Cossette and
Miller 1995 Rocha et al 2005 Boddu et al 2007) As animals TC inhibits protein synthesis
in non-host A thaliana (Nishiuchi et al 2006) Immunogold assays showed that the cellular
targets of DON in wheat and maize were cell membrane ribosome endoplasmic reticulum
21
vacuoles and mitochondria Modulation of cell membrane integrity and electrolyte losses
were observed by direct DON interference in maize and wheat cell assays (Cossette and
Miller 1995 Mitterbauer et al 2004) DON-mediated mitochondrion membrane alterations
disruption of host cells enzymatic activity apoptosis and necrosis have been observed in
cereals (Mitterbauer et al 2004 Rocha et al 2005) In contrast to animal type-B is more toxic
to plants than type-A DON showed 8- and 13-fold phytotoxicity to wheat coleoptiles than T-
2 and HT-2 toxins respectively (Eudes et al 2000) By compilation of different studies TC
phytoxicity can be ordered as DONge3-ADONgtT-2 toxingtDAS (Wakulinski 1989 McLean
1996 Eudes et al 2000)
22
17 Cellular and molecular mechanisms of DON toxicity in animals and plants
The underlying cellular and molecular mechanisms of DONTC toxicity are evolving
Exposure to DON proceeds with the up and down regulation of numerous critical genes of
eukaryotic organisms which causes various negative effects on animal and plant organisms
(Figure 1-2)
One of the early events of DON toxicity occurs when the mycotoxin binds to the
active ribosome resulting in blocking protein translation through different routes by
damaging the ribosome (Shifrin and Anderson 1999) degrading or altering conformational
changes of 28S-rRNA (Alisi et al 2008 Li and Pestka 2008) and inducing expression of
miRNAs This results in down regulation of the ribosome associated genes (He et al 2010a
Pestka 2010) Both animal and plant cell assays demonstrated that DON bound to the peptidyl
transferase (Rpl3) domain of the 60S subunit of ribosome and blocked protein chain
elongation and termination at high DON concentrations while peptide chain termination and
ribosome breakage occurred at a low level of DON exposure (Zhou et al 2003a)
23
Figure 1-2 Cellular and molecular mechanisms of deoxynivalenol toxicity in eukaryotic
organisms DON is simply absorbed by the cell and interacts with the cytoplasmic membrane
(CM) mitochondrion (MT) membrane and ribosome Disruption of the CM and alteration of
the MT membrane causes a loss of electrolytes and affects the function of the MT
respectively DON binds to the active ribosomes which leads to ribosome damage and
induces micro RNAs (miRNA) Expression of miRNA impairs the synthesis of ribosome
associated protein The subsequent transduction and phosphorylation of the RNA-activated
protein kinase (PKR) and hematopoietic cell kinase (Hck) activate the mitogen-activated
protein kinase (MAPK) cascade Phosphorylation of the MAPKs induces targeted
transcription factors (TF) which results in ribotoxis stress response upregulation of
proinflammatory cytokines and dysregulation of neuroendocrine signaling (derived from
Pestka 2007 and 2010)
24
The consequence of DON interaction with ribosomes leads to cellular and molecular
changes which are complex and not fully understood However it is well-demonstrated that
the activation of the mitogen-activated protein kinase (MAPK) and subsequent molecular
actions play key roles in the cytotoxicity of TC Upon binding to the ribosome DON rapidly
induced interactions between the protein kinases and the ribosomal 60S subunit as observed in
mononuclear phagocytes (Bae and Pestka 2008) Ribosomal binding to DON activated the
ribosome associated kinases for instances protein kinase R (PKR) hematopoietic cell kinase
(Hck) and P38
mitogen-activated protein kinase The protein kinases are induced by double-
stranded RNA (dsRNA) or possibly by the damaged ribosomal peptides (Iordanov et al 1997
Shifrin and Anderson 1999 Zhou et al 2003b Pestka 2007) Subsequent phosphorylation of
kinases (eg PKRHckP38
) activates the MAPK cascade resulting in ribotoxis stress
response upregulation of proinflammatory cytokines and dysregulation of neuroendocrine
signaling (Pestka and Smolinki 2005 Pestka 2010) Studies suggested that high doses of
DON induces ribotoxis stress response (apoptosis) preceded by immunosuppression and
disruption of intestinal permeabilitytissue functions (Yang et al 2000 Pestka 2008) Further
study observed stabilization of the mRNA and expression of transcription factors for critical
genes when cells were exposed to a low concentration of DON (Chung et al 2003)
The MAPK pathway proceeds with upregulation of a broad range of proinflammatory
cytokine genes and dysregulation of neuroendocrine signaling within the central and enteric
nervous systems Modulation of neuroendocrine signaling by DON increased the level of
serotonin resulted in vomiting reduced food intake and growth retardation in experimental
animalscell lines (Prelusky and Trenholm 1993 Li 2007) Upregulation of proinflammatory
cytokines and chemokines genes was determined in mice liver spleen kidney and lung
25
following in 1 h of exposure to DON A subsequent shock-like effect was observed in mice
because of a ldquocytokine stormrdquo (Pestka 2008) Induction of proinflammatory cytokines also led
to various cellular effects on the central nervous system interference of the innate immune
system and growth hormone signaling These contribute to anorexia growth retardation
aberrant immune response and tissue dysfunction as demonstrated in experimental animal
systems (Pestka 2010)
DON-mediated growth retardation in mice via suppression of growth hormones
(insulin-like growth factor 1 and insulin-like growth factor acid-labile subunit) has been
reported by Amuzie et al (2009) Disruption of gut tissues by DONNIV and subsequent
impairment of intestinal transporter activity was demonstrated in a human epithelial cell
model (Marseca et al 2002) The study proposed that alteration of gut tissue permeability
may cause gastroenteritis induction and growth retardation in livestock animals Growth
retardation of livestock may also occur via dysregulation of neuroendocrine signaling-
mediated induction of emesis and anorexia (Pestka et al 2010)
The molecular mechanism of TC toxicity in plant systems has not been well
documented A recent study showed the upregulation of numerous defense related genes in
wheat upon exposure to DON (Desmond et al 2008) The study further suggested that DON-
induced program cell death in wheat occurred through a hydrogen peroxide signaling
pathway This mechanism had not been observed in mammals
26
18 Strategies to control FHB and trichothecenes
During the past decades many pre- and post-harvest strategies have been applied to
protect cereal crops from Fusarium infestation and to eliminate TC from contaminated
agricultural commodities (FAO 2001 Munkvold 2003 Varga and Toth 2005 Kabak and
Dobson 2009) Some of the above crop protection techniques have been patented (He and
Zhou 2010) In spite of those efforts TC contamination of cereals appears to be unavoidable
using the currently available methods
Pre-harvest control strategies involving cultural practices (eg crop rotation tillage
early planting) and fungicide applications partially reduced Fusarium infection and DON
contamination in cereals (Dill-Macky and Jones 2000 Schaafsma et al 2001) However
repeated application of fungicides may lead to the evolution of Fusarium strains resistant to
fungicides with greater mycotoxin production ability (DrsquoMello et al 1998) At post-harvest
reduction of grain moisture by artificial drying storage at low temperatures (1-4degC) and
proper ventilation of the storage room have shown to be effective in inhibiting fungal growth
and mycotoxin production in infected grain (Midwest Plan Service 1987 Wilcke and Morey
1995) This technology has become less applicable particularly in developing countries
because of the frequent observation required to maintain proper storage conditions (Hell et al
2000)
Removal of Fusarium infected grains by physical methods (eg mechanical
separation density segregation colour sorting etc) was unable to eliminate TC completely
(Abbas et al 1985 He et al 2010b) This is partly because FHB symptomless kernels may
contain a considerable amount of DON which are not eliminated by physical separation
procedures (Seitz and Bechtel 1985) Several studies showed that milling and food processing
27
steps were unable to completely remove DON from grain-based foods (Jackson and
Bullerman 1999 Placinta et al 1999 Kushiro 2008) Detoxification of TC by chemical
treatments (eg ozone ammonia chlorine hydrogen peroxide and sodium metabisulfite) has
been demonstrated (Young et al 1986 Young et al 1987 Young et al 2006) The problem
with these techniques is that the application of such chemicals is not economically viable and
it also has a negative impact on grain quality (Karlovsky 2011) Although the technique has
been popular and showed effectiveness to certain mycotoxins utilization of absorbents to
inhibit mycotoxin absorption by the livestock gastrointestinal tract appeared to be ineffective
for TC (Huwig et al 2001)
The introduction of Fusarium resistant cultivars would be the best strategy to address
the FHB and associated mycotoxin problems in cereals However breeding for cultivars
resistant to FHBGER is difficult and time consuming as natural resistance against Fusarium
is quantitative in nature FHB resistant quantitative trait loci (QTLs) have been identified in
several cereal genotypes (eg Sumai 3 Ning 7840 Bussard) (Bernando et al 2007 Golkari
et al 2009 Haumlberle et al 2009 Yang et al 2010) The 3B 5A and 6B QTLs for type-I
(resistant to initial infection) and type-II (resistant to fungal spread in the kernel) resistances
have been identified on different wheat chromosomes (Buerstmayr et al 2009) Type-I
(Qfhsifa-5A) and type-II (Fhb1Fhb2-3B) QTLs from Sumai 3 are being used in many
breeding programs and pyramiding of the QTLs in spring wheat showed additional effects
Nonetheless the key genes involved in FHB resistance in cereals are yet to be identified This
makes it difficult to integrate the resistance genes into elite cultivars
28
Cultivar development by direct incorporation of TCDON resistance genes may be an
alternative way of protecting grains from these mycotoxins contaminations Many genes from
distantly related species have been integrated into crop cultivars using a transgenic approach
with great economic benefits (Brookes and Barfoot 2006) DON transforming trichothecene
3-O-acetyltransferase (TRI101 from Fusarium sporotrichioides) and UDP-
glucosyltransferase (from Arabidopsis thaliana) genes expressed in cereal cultivars showed
some DONDAS detoxifying ability under in vitro and controlled conditions (Kimura et al
1998 Okubara et al 2002 Poppenberger et al 2003 Ohsato et al 2007) By contrast no
reduction of mycotoxins in either of the above cases was observed under field conditions
(Manoharan et al 2006) Importantly the transformed acetylated and glycosylated
compounds could revert to the original substances or convert to even more toxic products
(Gareis et al 1990 Alexander 2008)
19 Isolation of DONTC de-epoxidating micro-organisms and genes
An enzyme with the capability of de-epoxidizing DONTC appears to be a potential
source for completely removing TC from contaminated cereals (Karlovsky 2011) However
such trichothecene detoxifying genes do not exist in cereals and other plants (Boutigny et al
2008) Because of this several research groups have attempted to obtain microorganisms that
encode DONTC de-epoxidation genes Mixed microbial culture capable of de-epoxydizing
trichothecenes have been isolated from chicken gut (He et al 1992) cow ruminal fluid (King
et al 1984 Swanson et al 1987 Fuchs et al 2002) pig gut (Kollarczik et al 1994) and
digesta of chicken (Young et al 2007) rats (Yoshizawa et al 1983) sheep (Westlake et al
1989) and fish (Guan et al 2009) All of the microbial cultures cited above showed de-
29
epoxydation capabilities only under anaerobic conditions realtively high temperatures and at
lowhigh pH except microbes from fish intestine that showed efficient de-epoxidation at low
temperatures and wide range of temperatures (Guan et al 2009) Such enzymes active under
anaerobic conditions high temperatues and low pH may have potential applications as feed
additives and the eubacterial strain BBSH 797 is an animal feed additive product with the
brand name Mycofixreg by BIOMIN (He et al 2010b) Biotechnological applications of
anaerobically-active enzymes from the above microbial sources seem to be inappropriate
under the aerobic and moderate conditions at which cereals plants are grown
Most of these efforts have failed to isolate single DONTC degrading microbes
effective under aerobic conditions because of inappropriate microbial growth conditions
under laboratory conditions Earlier attempts were made to obtain TC de-epoxidizing aerobic
bacteria but the culture did not retain DONNIV de-epoxidation activity after reculturing
(Ueno 1983 He et al 1992) In addition the co-metabolic nature of the microbial TC de-
epoxidation reaction has limited the use of mycotoxins as the sole nutrient source for selective
isolation of the target microorganisms Several DONTC de-epoxidation microbial strains
including Bacillus strain LS100 and Eubacterial strain BBSH 797 have been isolated from
chicken gut and rumen fluids through multi-step enrichment procedures (Fuchs et al 2002
Yu et al 2010) A DON to 3-keto DON transforming bactial strain was islated from soil by
prolonged incubation with high concentration DON (Shima et al 1997)
Microbial genesenzymes with TC de-epoxidation activity are not known Based on
the microbial TC de-epoxidation activity of a mixed microbial culture it has been proposed
that ceratin reductases or oxidoreductases may catalyze such a biochemical reaction (Islam et
al 2012) With the advent of molecular techniques many novel genes with
30
industrialagricultural value have been cloned by reverse genetics (genomic DNA library
preparation and functional screening of the library clones) forward genetics (transposon
mutagenesis) and comparative genomics methods (Frey et al 1998 Koonin et al 2001
Suenaga et al 2007 Chung et al 2008 Ansorge 2009) The tremendous advancement of next
generation sequencing technology and bioinformatics tools may facilitate the identification of
the microbial genes responsible for DONTC detoxification (Ansorge 2009) Isolation of TC
de-epoxidation genes may play a major role in producing safe foods and feeds derived from
grains
110 The bacterial species Citrobacter freundii
Citrobacter freundii is a bacterial species belonging to the family Enterobacteriace It
is a facultatively anaerobic and Gram-negative bacillus (Wang et al 2000) The bacterium
survives almost everywhere including animalhuman intestines blood water and soils
(Wang et al 2000 Whalen et al 2007) C freundii is known as an opportunistic human
pathogen and causes a variety of respiratory tract urinary tract and blood infections and
responsible for serious neonatal meningistis disease (Badger et al 1999 Whalen et al 2007)
Interestingly this bacterium plays a positive environmental role through nitrogen recycling
(Puchenkova 1996) Strains of C freundii also contribute to the environment by sequenstering
heavy metals and degrading xenobiotic compounds (Kumar et al 1999 Sharma and Fulekar
2009)
31
111 Hypotheses and objectives
The study was based on the following hypotheses
i Certain microorganisms in DON contaminated agricultural soils are able to transform DON
to de-epoxy DON (de-epoxidation) under aerobic conditions and moderate temperatures
ii DON de-epoxidation may be achieved by a single microbial strain
iii Microbial DON detoxifying genes and associated regulators are able to express and
function in a closely related host organism
iv DON detoxifying microbial strains carry genes for de-epoxidation activity that are absent
from related species that have not evolved under DON selection pressure
The objectives of this research were to
i Select microbial cultures from soil having DON de-epoxidation activity under aerobic
conditions and moderate temperatures by analyzing potential DON contaminated soils in
various growth conditions namely aerobicanaerobic conditions different growth media
temperature and pH
ii Select for organisms with DON de-epoxidation activity from the mixed microbial culture
by treating with antibiotics heating subculturing on agar medium and incubating for a
prolonged period in media with a high DON concentration
iii Identify microbial de-epoxidation genes by producing a microbial genomic DNA library
from the DON de-epoxidizing strain followed by screening of the clones for DON de-
epoxidation
32
iv Determine microbial de-epoxidation genes by sequencing of the genome and comparative
analysis of the genome with closely related species and identification of putative DON
detoxification genes
33
CHAPTER 2
20 AEROBIC AND ANAEROBIC DE-EPOXIDATION OF MYCOTOXIN
DEOXYNIVALENOL BY BACTERIA ORIGINATING FROM AGRICULTURAL
SOIL
Published in World Journal of Microbiology and Biotechnology 2012 287-13
21 Abstract
Soil samples collected from different cereal crop fields in southern Ontario Canada
were screened to obtain microorganisms capable of transforming deoxynivalenol (DON) to
de-epoxy DON (dE-DON) Microbial DON to dE-DON transformation (ie de-epoxidation)
was monitored by using liquid chromatography-ultraviolet-mass spectrometry (LC-UV-MS)
The effects of growth substrates temperature pH incubation time and aerobic versus
anaerobic conditions on the ability of the microbes to de-epoxidize DON were evaluated A
soil microbial culture showed 100 DON to dE-DON biotransformation in mineral salts
broth (MSB) after 144 h of incubation Treatments of the culture with selective antibiotics
followed an elevated temperature (50ordmC) for 15 h and subculturing on agar medium
considerably reduced the microbial diversity Partial 16S-rRNA gene sequence analysis of the
bacteria in the enriched culture indicated the presence of at least Serratia Citrobacter
Clostridium Enterococcus Stenotrophomonas and Streptomyces The enriched culture
completely de-epoxidized DON after 60 h of incubation Bacterial de-epoxidation of DON
occurred at pH 60 to 75 and a wide array of temperatures (12 to 40ordmC) The culture showed
34
rapid de-epoxidation activity under aerobic conditions compared to anaerobic conditions This
is the first report on microbial DON to dE-DON transformation under aerobic conditions and
moderate temperatures The culture could be used to detoxify DON contaminated feed and
might be a potential source for gene(s) for DON de-epoxidation
Keywords biotransformation de-epoxidation deoxynivalenol soil bacteria vomitoxin
22 Introduction
Deoxynivalenol (DON commonly known as lsquovomitoxinrsquo) 3α 7α 15-trihydroxy-
1213-epoxytrichothec-9-en-8-one is a trichothecene produced by toxigenic Fusarium species
causing Fusarium head blight (FHB) or Gibberella ear rot (GER) disease on small grain
cereals and maize respectively (Starkey et al 2007 Foroud and Eudes 2009) DON
contamination in cereals and maize is widespread and can lead to tremendous losses to the
growers It has also detrimental effects on human and livestock health (Pestka and Smolinski
2005 Rocha et al 2005) The toxin interferes with the eukaryotic ribosomal peptidyl
transferase (Rpl3) activity resulting in human and animal mycotoxicosis (Ueno 1985 Bae
and Pestka 2008) Common effects of DON toxicity in animals include diarrhea vomiting
feed refusal growth retardation immunosuppression reduced ovarian function and
reproductive disorders (Pestka and Smolinski 2005 Pestka 2007) In plants DON causes a
variety of effects viz inhibition of seed germination seedling growth green plant
regeneration root and coleoptile growth (cited in Rocha et al 2005) In addition DON
enhances disease severity in sensitive host plants (Maier et al 2006)
35
Avoidance of DON contamination or reduction of toxin levels in contaminated grains
remains a challenging task Physical and chemical processes are either inappropriate or
prohibitively expensive to use to decrease the DON content of agricultural commodities
(Kushiro 2008 He et al 2010b) The use of Fusarium resistant crop varieties might be the
best method to reduce DON contamination However breeding for FHB or GER resistant
cultivars appears to be very difficult and time consuming as natural sources of resistance
against Fusarium-caused diseases is quantitative in nature (Ali et al 2005 Buerstmayr et al
2009) As an alternative expressing genes for enzymatic detoxification of DON in elite
cultivars may be an effective strategy for protecting the value of feeds and foods (Karlovsky
1999) Heterologous expression of eukaryotic trichothecene 3-O-acetyltransferase and UDP-
glucosyltransferase genes in cereals and Arabidopsis have resulted in reduced toxicity via
acetylation and glycosylation of the DON (Poppenberger et al 2003 Ohsato et al 2007)
It has been proposed that enzymatic de-epoxidation of trichothecene toxins could be
used to limit DON contamination (Zhou et al 2008 Foroud and Eudes 2009) DON belongs
to type B-trichothecenes which contain an epoxide ring at C12-13 and the epoxide is highly
reactive and mainly responsible for toxicity (Figure 2-1) (Swanson et al 1988 Eriksen et al
2004)
36
Figure 2-1 Mechanism of microbial DON de-epoxidation Removal of the oxygen atom from
the epoxide ring results in formation of a C=C double bond at C-12 position which reduces
cytotoxicity (Swanson et al 1988 Eriksen et al 2004)
Microbial DON de-epoxidation by single bacterial strains and by mixed cultures has
been reported (King et al 1984 Swanson et al 1987 He et al 1992 Kollarczik et al 1994
Fuchs et al 2002 Young et al 2007 Guan et al 2009 Yu et al 2010) However all of these
microbes originated from fish and other animals and were strictly anaerobic The recovered
microbial cultures and single strains only showed de-epoxidation of trichothecenes at low
(le15ordmC) and high (gt30degC) temperatures Such enzymes might have limited applications for
toxin decontamination in cereal grains The aim of the current study was to isolate the
microbes from soil capable of transforming DON to a de-epoxy metabolite under aerobic
conditions and moderate temperatures
De-epoxydation
Deoxynivalenol (DON)
De-epoxyDeoxynivalenol (dE-DON)
37
23 Materials and methods
231 Soil samples
The upper layer (0-30 cm) of soils along with crop debris were randomly collected
from cereal (eg maize wheat barley) crop fields during 2004-2006 in Southern Ontario
Canada Detailed of the crops and locations of the fields have been described in Jianwei Hersquos
Master Degree thesis (He 2007) Briefly soil samples were collected in sterile plastic bags
and transported to the laboratory Samples were sieved to remove large roots stones and
stored at 4ordmC until used The samples were mixed with grounded mouldy corn (containing 95
microgg DON) + F graminearum and incubated at 22-24degC for six weeks Five samples were
prepared for DON de-epoxidation activity analysis by dispersing 5 g soil in 50 mL distilled
water and storing at4degC until further use
232 Culture media
Media used in the study were nutrient broth (NB) Luria Bertani (LB) minimal
medium (MM) (He 2007) six mineral salts + 05 bacto peptone (MSB) (He 2007) tryptic
soy broth (TSB) corn meal broth (CMB) (Guan et al 2009) potato dextrose broth (PDB) six
mineral salts + 05 yeast extract broth (MSY) and soil extract (SE) MM was composed of
sucrose-100 g K2HPO4-25 g KH2PO4-25 g (NH4)2HPO4-10 g MgSO47H2O-02 g
FeSO4-001 g MnSO47H2O- 0007 g water- 985 mL and MSB was composed of the same
salts as MM but was supplemented with Bactotrade Peptone (05 wv) as the sole C source
instead of sucrose To prepare soil extract 100 g of the composite soil was homogenized in 1
L water The soil particles and plant debris were removed by passing the soil suspension
through Whatmanreg grade 3 filter paper (Sigma-Aldrich Oakville Ontario Canada) The
38
extract was autoclaved at 121degC for 30 min Media and ingredients were purchased either
from Fisher Scientific Inc (Fair Lawn New Jersey USA) Sigma-Aldrich (Oakvile Ontario
Canada) or Difco (Sparks Maryland USA) Prior to autoclaving pH of the broth media was
adjusted to 70 Solid media were prepared by adding 15 agar
233 Screening of soil samples
Frozen soil suspensions were thawed and vortexed to homogenize the microbial
population An aliquot (100 microL) of the suspension was immediately transferred to MSB PDB
and MM broth media DON at 50 microgmL was added to the microbial suspension cultures The
cultures were incubated at 22-24degC with continuous shaking at 180 revolutions per minute
(rpm) The extent of DON de-epoxidation was determined at 24 h 48 h 72 h 96 h 120 h and
144 h of incubation
234 Determination of de-epoxidation product
Microbial DON de-epoxidation was measured according to Guan et al (2009)
Samples were prepared by mixing equal volume of microbial culture and methanol and
allowed to stand for 30 min at room temperature Microorganisms in the mixture were
removed by passing them through a 045 microm Millipore filter (Whatmanreg Florham Park New
Jersey USA) The filtrates were used for de-epoxidation analysis by a liquid chromatography-
ultraviolet-mass spectrometry (LC-UV-MS) Identification and quantification of DON and its
transformation product was achieved using a Finnigan LCQ DECA ion trap LC-MS
instrument (ThermoFinnigan San Jose CA USA) The equipment detects 30-2000 mz
(molecular mass ion ratio) compounds and able to determine 001 micromL DON The HPLC
39
system was equipped with a Finnigan Spectra System UV6000LP ultraviolet (UV) detector
and an Agilent Zorbax Eclipse XDB C18 column (46times150 mm 35 μm) The binary mobile
phase consisted of solvent A (methanol) and solvent B (water) The system was run with a
gradient program 25 A at 0 min 25ndash41 A over 5 min isocratically at 41 A for 2 min
and 41ndash25 A over 1 min There was a 3 min post-run at its starting condition to allow for
the equilibration of the column The flow-rate was set at 10 mLmin UV absorbance was
measured at 218 nm Atmospheric pressure chemical ionization (APCI) positive ion mode
was used for MS detection A DON standard was used to adjust the instrument to its
maximum response The system was operated as follows shear gas and auxiliary flow-rates
were set at 95 and 43 (arbitrary units) voltages on the capillary tube lens offset multipole 1
offset multipole 2 offset and entrance lens were set at 3000 minus1700 minus370 minus850 minus3500
and minus4200 V respectively capillary and vaporizer temperatures were set at 200 and 450degC
respectively and the discharge needle current was set at 7 μA Identification of mycotoxin
was achieved by comparing their retention times UV spectra and mass spectrum with DON
and de-epoxy DON (Biopure Union MO USA) as standards Quantification of DON and
product was based on peak areas related to their UV and MS calibration curves
40
235 Biological DON de-epoxidation
To help identify the types of microorganisms causing the DON de-epoxidation MSB
medium was supplemented with either 100 microgmL penicillin G or streptomycin sulfate for
suppressing gram positive or negative bacterial growth and activities respectively Three
replications of antibiotic-treated media were inoculated with 100 microL of microbial culture
capable of de-epoxidizing DON The antibiotic-treated cultures were incubated at 27degC
spiked with 50 microgmL DON Controls were prepared without adding antibiotics in the
microbial culture Inhibition of DON de-epoxidation was analyzed after five day of
incubation
236 Reduction of microbial diversity by antibiotic treatments heating and
subculturing on agar medium
In order to reduce non-DON degrading microbial populations the DON de-
epoxidation capable cultures were sequentially treated with antibiotics having diverse modes
of action including i) dichloramphenicol that kills various gram positive Gram-negative and
anaerobic bacteria ii) trimethoprim that kills Gram-positive and negative bacteria iii)
penicillin that is lethal to Gram-positive bacteria and iv) cyclohexamide to kill filamentous
fungi and yeasts Antibiotics were purchased from Sigma-Aldrich Antibiotic stock solutions
(10 mgmL) were prepared with sterile water or methanol Microbial cultures in MSB
medium were treated with 100 microgmL of each antibiotic followed by incubation of the culture
at 50ordmC for 15 hours to kill undesirded microbial species After heating cultures were treated
with 50 microgmL DON and incubated at 27ordmC for five days DON de-epoxidation activity of the
microbial cultures was determined following the procedure described above Cultures that
41
showed gt99 de-epoxidation were considered for subsequent antibiotic and heat treatments
Aliquots of the achieved culture were spread on MSB agar plates and grown at room
temperature for few days Colonies appeared on the plates were collected and incubated in
MSB medium supplemented with 50 microgmL DON De-epoxidation ability of the subcultured
microbial cultures was determined after five days of incubation at 27degC by LC-UV-MS
method
237 Analysis of microbial population diversity
Terminal Restriction Fragment Length Polymorphism (T-RFLP) analysis
Initially microbial diversity was determined by terminal restriction fragment length
polymorphism (T-RFLP) fingerprint analysis (Bruce and Hughes 2000) Genomic DNA from
microbial cultures with and without enrichment was extracted using DNeasy Plant DNA
Extraction Mini Kit (Qiagen Inc Mississauga Ontario Canada) 16S-rDNA was amplified
by PCR using the HEX-labeled forward primer Bac8f
(5-agagtttgatcctggctcag-3) and
unlabeled reverse primer Univ1492r (5-ggttaccttgttacgactt-3) (Fierer and Jackson 2006)
Fifty microlitter PCR mixture for each sample was prepared having 1x HotStarTaq Master
Mix (Qiagen) 05 microM of each primer 50 microg of BSA and 50 ng of DNA The 35 PCR cycles
followed 60 s at 94degC 30 s at 50degC and 60 s at 72degC After size verification by agarose-gel
electrophoresis 50 ng of purified PCR products was digested for 3 h in a 20 microL reaction
mixture with 5 U of HhaI restriction enzyme (New England Biolabs Ipswich MA USA)
The size and relative abundance of the labeled T-RFLP fragments were separated by capillary
electrophoresis in an automated DNA sequencer equipped with fluorescence detector
(Applied Biosystem 3730 DNA Analyzer) The DNA fragments were analyzed with the Peak
42
Scannertrade Software v10 to create an electropherogram with the genetic fingerprints of the
microbial community
Analysis of V3-V5 region of the 16S-rDNA
To define the bacterial genera present in the enrichment culture the 16S-rDNA was
amplified using two sets of bacterial universal primers Primarily 16S-rDNA was PCR
amplified using the primers 46f-5rsquogcytaacacatgcaagtcga3rsquo (Kaplan and Kitts 2004) and
1371r-5rsquogtgtgtacaagncccgggaa3rsquo (Zhao et al 2007) PCR was performed in a total volume of
25 μL which contained 06 μM of each primer 1x of HotStarTaqreg Master Mix (Qiagen Inc)
and 20-40 ng of DNA The PCR cycles included the following steps 95ordmC (15 min) 35 cycles
of 94ordmC (20 s) 52ordmC (15 s) 72ordmC (1 min 30 s) and a 72ordmC (7 min) extension step The second
pair of universal bacterial primers were 27f-5rsquoagagtttgatcmtggctcag3rsquoand 1492r-
5rsquoggttaccttgttacgactt3rsquo (Lane 1991) PCR was conducted with 25 cycles following the
procedures of Tian et al (2009) Amplification was carried out in a GeneAmpreg PCR System
9700 (Applied Biosystems) Furthermore the PCR products were used as templates to
amplify the hypervariable V3-V5 region of the 16S-rDNA using primers 357f-
5rsquocctacgggaggcagcag3rsquo and 907r7 5rsquoccgtcaattcctttgagttt3rsquo (Yu and Morrison 2004)
After purification of the PCR products by QIAquick PCR purification kit two
amplicon libraries were prepared by ligating to pCR-TOPO vector (Invitrogen) and the library
clones were transformed into TOPO10 competent cells according to Invitrogenrsquos TOPO
cloning manual The clone libraries were plated on LB plates containing 25 μgmL
kanamycin Fifty colonies from each library were randomly picked up from the overnight
plates and inserts were sequenced as described by Geng et al (2006) PCR products were
43
sequenced with an automated ABI 3730 DNA analyzer (Applied Biosystems) The sequence
similarities of the amplified V3-V5 region were compared with the ribosomal database project
(RDP) (httprdpcmemsuedu) Sequences with greater than 95 identity were assigned to
the same bacterial genus
238 Factors influencing DON de-epoxidation
DON de-epoxidation ability of the enriched microbial culture was investigated under
different conditions as described below
Maintaince of active culture
To retain microbial de-epoxidation activity the active culture has been maintained in
MSB medium containing 50 microgmL DON
Media growth temperature and pH
Overnight microbial cultures (100 μL) were added to 2 mL of NB LB MM TSB
MSB CMB MSY PDB or SE broths The pH of the media was adjusted to 70 and the
experiments were conducted at 27degC temperature The effect of temperature was studied by
incubating the culture in MSB medium (pH 7) at 12 20 25 30 and 40ordmC To determine the
effects of pH MSB medium pH was adjusted to 55 60 65 70 75 and 80 Media with
different pHs were inoculated with aliquots of the overnight active culture and then incubated
at 27ordmC The cultures were supplemented with 50 μgmL DON before incubation Three
replications for each medium temperature and pH condition were evaluated
44
De-epoxidation under aerobic and anaerobic conditions
Three replicate microbial cultures of five mL each were grown in sterile 15 mL
centrifuge tubes under aerobic conditions and in an anaerobic chamber filled with 95 CO2
and 5 H2 (COY Laboratory Products Inc Ann Arbor MI USA) Three days after initiation
half (25 mL) of the cultures from anaerobic conditions was transferred to new centrifuge
tubes and brought to aerobic conditions Similarly half of the aerobically grown cultures were
placed into an anaerobic chamber Cultures moved from anaerobic to aerobic conditions were
shaken after opening the lids of the tubes in a laminar flow every eight hours intervals to
maintain aerobic conditions in the culture Lids of the culture tubes transferred to anaerobic
conditions were kept loose to allow oxygen to escape from the culture Two days after
transferring to the new conditions the cultures in both conditions were treated with 50 microgmL
DON and incubated at 27degC Every 12 h 500 microL were removed from each tube and analyzed
for microbial DON de-epoxidation activity by LC-UV-MS
239 Data analysis
Data were processed using Microsoft Excel Program 2007 Statistical analysis was
done using General Linear Model procedures in SAS version 91 (SAS Institute Inc Cary
NC USA 2002-03) The results were the average of three replications stated as mean plusmn
standard error Differences between means were considered significant at plt 005 according
to Tukeyrsquos-HSD test
45
24 Results
241 Analyses of DON to dE-DON biotransformation
A typical LC-UV-MS analysis where there is incomplete de-epoxidation of DON by a
particular soil suspension culture is shown in Figure 2-2 Analysis by LC with UV detection
and integration of peak areas showed 662 conversion of DON to dE-DON (Figure 2-2a)
Analysis LC with MS detection (expressed as total ion counts) indicated 643 conversion
(Figure 2-2b) LC analyses with UV and TIC (total ion count) detection are subject to
interferences due to co-eluting contaminants and therefore integration of small peaks was
very difficult making quantification inaccurate MS detection by SIM (selected ion
monitoring) can eliminate the influence of co-eluting contaminants For example the SIM LC
chromatogram for DON (Figure 2-2c) was generated by using the major ions resulting from
the APCI positive ion MS of DON (Figure 2-2e)
Similarly the SIM LC chromatogram for dE-DON (Figure 2-2d) was derived from the
MS ions of dE-DON (Figure 2-2f) The signal to noise ratios (SN) of peaks observed by SIM
were at least 20 times greater than those detected by TIC because there is reduced likelihood
of co-eluting peaks having the same ions Integration of the SIM peaks revealed a 616
conversion The three methods gave similar results but the SIM LC results were considered
to be the most accurate Using that method the conversion of DON to dE-DON by the mixed
culture occurred most quickly at 48 h and was completed by 144 h (Figure 2-3)
46
47
Figure 2-2 LC-UV-MS chromatographic analysis of DON and its de-epoxidated product dE-
DON obtained by culturing with selected soil microbes (a) LC-UV chromatogram of product
mixture monitored at 218 nm (b) LC-MS total ion chromatogram (TIC) of product mixture
(c) selected ion monitoring (SIM) chromatogram of cultured DON at mz 231 249 267 279
and 297 (d) SIM chromatogram of cultured dE-DON at mz 215 233 245 251 263 and 281
(e) atmospheric pressure chemical ionization (APCI) positive ion mass spectrum of DON (f)
APCI positive ion mass spectrum of dE-DON
48
242 Screening of soil samples
Initially samples originating from soils were analyzed in three types of culture media
to support the growth of fungi yeast and bacteria However cultures growing in PDB which
is a suitable substrate for many fungi and yeasts did not permit transform DON to dE-DON
(data not shown by microorganisms) A define medium (ie MM) was also used in the
sample screening assay None of the soil samples showed de-epoxidation activity in MM
(data not shown) From five soil samples one soil sample culture showed de-epoxidation of
DON in MSB The culture in MSB converted gt99 DON to dE-DON within 144 h (Figure 2-
3) Conversion began after a lag of 48 h and was linear thereafter with a rate of 1 per hour
49
Figure 2-3 Time course of relative biotransformation of DON to dE-DON by the initial soil
suspension culture in MSB medium The culture contained 50 μgmL DON at time 0 Culture
with three replications was incubated at room temperature with continuous shaking at 180
rpm The conversion of DON to dE-DON was analysed by LC-UV-MS at 24 h intervals Blue
and red lines are the percent conversion of DON and recovery of dE-DON respectively
50
243 Evidence of biological DON de-epoxidation
Treatment of active microbial culture with 100 microgmL of streptomycin completely
inhibited DON de-epoxidation In contrast the same concentration of penicillin G did not
affect activity of the culture (Figure 2-4) Streptomycin is effective against Gram-negative
bacteria while penicillin G is lethal to Gram-positive bacteria These experiments provided
evidence that DON de-epoxidation may be caused by certain Gram-negative bacteria
51
Figure 2-4 Microbial de-epoxidation of DON after five days of incubation The active DON
de-epoxidation culture was treated with either 100 microgmL penicillin G or streptomycin
(Strepto) in MSB Cultures having three replications were added 50 microgmL DON and
incubated at room temperatures with continuous shaking at 180 rpm For controls cultures
were without antibiotics DON to dE-DON transformation was monitored after five day of
incubation by LC-UV-MS method
0
20
40
60
80
100
120
Penicillin Strepto Control
DO
N d
e-e
po
xyd
ati
on
(
)
Antibiotics
52
244 Reduction of the microbial diversity
Plating the suspension cultures onto MSB agar revealed colonies with a variety of
sizes morphologies and colours reflecting a mixed culture (data not shown) To reduce the
complexity of the culture three steps of selection pressures were applied namely treatment
with antibiotics heat shocks and subculturing on solid medium The above indicated
treatments did not significantly affect DON de-epoxidation of the microbial cultures (data not
shown) T-RFLP profiles of the mixed culture before treatments showed six distinct peaks of
about 110 150 200 220 340 and 420 bp restriction fragments with the 110 bp peak being
the predominant one (Figure 2-5a)
After treatment with antibiotics and heat only three peaks at 110 150 and 325 bp
remained (Figure 2-5b) suggesting that there was a reduction in the microbial diversity
without affecting DON de-epoxidation After enrichments the bacteria species in the culture
changed their growth patterns as the electropherogram produced different sizes of pick
lengths compared to untreated culture
53
(b)
(a)
Rela
tive flu
ore
scen
ce
32000
0
0 500
16S-rDNA fragment size (bp)
100 200 300 400
Rela
tive flu
ore
scen
ce
32000
0
500
16S-rDNA fragment size (bp)
0 300200100 400
54
Figure 2-5 Analysis of microbial population reduction by terminal restriction fragment
length polymorphism (T-RFLP) (a) Restriction fragments of the 16S-rDNA of the untreated
culture and (b) fragmentation patterns of the enriched culture Picks along the X-axis showed
the fragmentation patterns of the 16S-rDNA which is related to the bacterial species diversity
The length of picks (along the Y-axis) showed abundance of terminal restriction fragments
related to the abundance of particular bacterial speciesgenera
55
245 Microbial population structures in the enriched culture
Microbial diversity in the enriched culture was determined by analyzing the sequences
of the V3-V5 region (~586 bp) amplified from the 16S-rDNAs of the microorganisms it
contained The sequence analysis showed the presence of Serratia Citrobacter Clostridium
Enterococcus Stenotrophomonas and Streptomyces in the enriched culture (Figure 2-6)
Out of 100 V3-V5 sequences 54 sequences were found to be belonging to Serratia
the predominant species of the culture The remaining sequences matched with 12
Citrobacter 8 Clostridium 14 Enterococcus 8 Stenotrophomonas and 4 Streptomyces
species
56
Figure 2-6 Microbial genera identified in the enriched DON de-epoxidizing microbial
culture The bacterial genera were determined by comparing the hypervariable regions (V3-
V5) of the 16S-rDNA using ribosomal database project (httprdpcmemsuedu) X-axis
showed the identified bacterial genera and Y-axis demonstrated the abundance of the V3-V5
sequences belonging to different bacterial genera
54
12 8
14
8 4
0
20
40
60 S
eq
uen
ce
fre
qu
en
cy
Microbial genera
57
246 Effects of media growth temperature pH and oxygen requirements on DON de-
epoxidation
Of the nine media that were tested DON de-epoxidation by the enriched microbial
culture only occurred in NB and MSB (Figure 2-7a) It appeared that C andor N
concentrations in the media were crucial for the enzymatic activity because less or more than
05 Bacto Peptone in MSB reduced the microbial DON transformation ability
DON de-epoxidation activity occurred over a wide range of temperatures from 12 to
40ordmC (Figure 2-7b) The fastest rates of DON de-epoxidation occurred between 25 to 40ordmC
De-epoxidation was observed within a relatively narrow range of pH (60 to 75) with
the most rapid DON biotransformation activity observed at pH 70 (Figure 2-7c) At pH 70
the enriched culture showed DON de-epoxidation activity 125 times faster than at pH 60 or
75 At pH 55 and 80 there was no observable DON de-epoxidation
58
0
20
40
60
80
100
120
12degC
20degC
25degC
30degC
40degC
DO
N d
e-e
po
xyd
ati
on
(
)
Temperature
0
20
40
60
80
100
120
DO
N d
e-e
pxyd
ati
on
(
)
Media
(a)
(b)
59
Figure 2-7 Effects of culture conditions on DON to de-epoxy DON transformation by the
enriched microbial culture For the media experiment (a) all the media were adjusted to pH 7
and the incubation temperature was 27degC For the temperature experiment (b) MSB pH 7 was
used For the pH experiment (c) MSB at 27degC was used Microbial cultures were inoculated
with 50 μgmL DON De-epoxidation was determined after 72 h of incubation Media MSY
(mineral salts + 05 yeast extract) LB (Luria Bertani) TSB (tryptic soy broth) MSB
(mineral salts + 05 bacto peptone) SE (soil extract) CMB (corn meal broth) PDB (potato
dextrose broth) and MM (minimal medium)
0
20
40
60
80
100
120
55
60
65
70
75
80
DO
N d
e-e
po
xyd
ati
on
(
)
pH
(c)
60
The enriched culture showed DON de-epoxidation activity under both aerobic and
anaerobic conditions (Figure 2-8) The fastest transformation (100 de-epoxidation after 60 h
of incubation) occurred in cultures (A) that were grown under aerobic conditions throughout
the culture growth period A medium rate of de-epoxidation (100 de-epoxidation after 72 h)
was observed in cultures that were initially grown in either aerobic or anaerobic conditions
followed by transferring to anaerobic or aerobic conditions (A+AN and AN+A) respectively
The lowest rate occurred (100 de-epoxidation after 96 h) in cultures grown in continual
anaerobic conditions (AN) The means comparison of microbial DON de-epoxidation
efficiencies at 60 h of incubation showed that cultures continually grown in conditions A and
AN were statistically different Biotransformation abilities of the cultures that were switched
from one conditions to another (A+AN and AN+A) were statistically similar among each
other but different from cultures that were only grown in conditions A or AN (plt005)
61
Figure 2-8 Microbial DON to dE (de-epoxy DON)-DON transformation activity under
aerobic and anaerobic conditions A = culture incubated and examined the activity only under
aerobic conditions A+AN = culture initially was grown in aerobic conditions and then
transferred to anaerobic conditions AN = cultures grown throughout the assay under
anaerobic conditions AN+A = culture initially grown under anaerobic conditions and then
transferred to aerobic conditions Differences of lower case letters are significantly different
in de-epoxidation capability (after 60 h) among incubation conditions according to Tukeyrsquos
HSD test (p lt 005) The experiment was conducted with three replications
62
25 Discussion
The current study describes a novel bacterial culture isolated from agricultural soil that
was capable of transforming DON to de-epoxy DON under aerobic and anaerobic conditions
at moderate temperatures Because it has been speculated that DON translocates to soil from
contaminated crops and DON that enters into the soil could be metabolized by microbes
(Voumllkl et al 2004) soil samples for the current study were mainly collected from fields where
cereal crops contaminated with DON were grown
In our initial sample screening steps upwards of 144 h of incubation were required to
achieve gt99 DON de-epoxidation Such a long period may have been due to the presence of
fast growing microorganisms that had suppressive effects on the growth or activity of the
DON transforming bacteria (Davis et al 2005) The sequential use of different antibiotics
with different modes of actions followed by thermal treatments and subculturing on solid
medium resulted in a reduction in the number of bacterial species as determined by T-RFLP
analysis Analysis of V3-V5 region of the 16S-rDNA showed that five bacterial genera were
present in the enriched culture The culture showed higher DON de-epoxidation activity (only
60 h required for gt99 biotransformation) compared to initial soil suspension culture that
required 144 h to complete the de-epoxidation This microbial culture could be used to isolate
the DON de-epoxidizing gene(s) applying metagenomics approaches (Daniel 2005)
The culture showed complete de-epoxidation activity both in MSB and NB media
However we have found that the use of nutritionally rich NB medium enhanced non-target
bacterial (eg Serratia lequifaciences) growth but this did not occur in MSB which is a
comparably poor medium To avoid the possible suppressive and detrimental effects of the
fast growing bacteria on the target bacteria MSB was used for enrichment and
63
characterization of the culture An important characteristic of the DON de-epoxidation
activity observed for the microbial culture in the current study is its activity at moderate
temperature (25-30degC) In contrast previously isolated cultures showed optimal activities at
low (le15degC) or high temperatures (37degC) (Guan et al 2009) Also the culture showed DON
de-epoxidation activity at neutral pH range (pH 60-75) in contrast to previous reports which
found that microbial trichothecene de-epoxidation occurred at acidic and basic pHs (Swanson
et al 1987 He et al 1992 Kollarczik et al 1994 Young et al 2007) However a microbial
culture from fish gut showed de-epoxidation under wide range of pHs (Guan et al 2009)
Probably the pH range reflects the environment in which the bacteria naturally grow which in
this case is similar to the soil pH of a typical crop field in Southern Ontario of Canada The
advantage of the present finding is that DON de-epoxidation activity of the culture was
optimal in a temperature range that matches the temperature range of activity for most arable
plants Therefore it is reasonable to suggest that the gene(s)enzyme(s) responsible for the
activity have potential application for enzymatic detoxification of DON in cereals (Karlovsky
1999)
The current observation that bacterial DON de-epoxidation activity occurred under
both aerobic and anaerobic conditions was interesting because DON de-epoxidation is a
reductive chemical reaction that might be expected to be promoted by anaerobic conditions
However there are previous reports of aerobic de-epoxidation of eldrindieldrin by soil
bacteria (Matsumoto et al 2008) and Goodstadt and Ponting (2004) described aerobic
reductive de-epoxidation by vitamin K epoxide reductasephylloquinone reductase A
definitive determination of the oxygen sensitivity of the enzyme responsible for DON de-
epoxidation will ultimately come from experiments with the purified protein
64
In conclusion this study reports on a novel bacterial source from soil that efficiently
de-epoxidizes DON at moderate temperatures under both aerobic and anaerobic conditions
This research may lead to the isolation of microbes and gene(s) encoding enzyme(s) capable
of de-epoxidizing DON that could create transgenic plants with reduced DON contamination
thus improving food and feed safety and reducing negative effects of the toxin on public
health
26 Acknowledgements
The authors gratefully acknowledged the Natural Sciences and Engineering Research
Council of Canada (NSERC) for awarding a scholarship to Rafiqul Islam for PhD program
The research was supported by the Agriculture and Agri-Food Canada Binational
Agricultural Research and Development Fund (AAFCBARD) project We are thankful for
the contributions of Jianwei He Xiu-Zhen Li and Honghui Zhu
65
CHAPTER 3
30 ISOLATION AND CHARACTERIZATION OF A NOVEL SOIL BACTERIUM
CAPABLE OF DETOXIFYING ELEVEN TRICHOTHECENE MYCOTOXINS
31 Abstract
Deoxynivalenol (DON vomitoxin) is one of the most commonly detected food and
feed contaminating mycotoxins worldwide In order to develop a control method effective
under aerobic conditions and moderate temperatures a bacterial strain ADS47 was isolated
capable of transforming DON to the less toxic de-epoxy DON The bacterial strain is
facultatively anaerobic Gram-negative rod shaped with an average cell size of 150 X 072
microm and has peritrichous (multiple) flagella The 16S-rRNA gene sequence had gt99
nucleotide identity to Citrobacter freundii a bacterium that belongs to the gamma subclass of
proteobacteria Strain ADS47 showed near 100 de-epoxidation of DON after 42 h of
incubation in nutrient broth (NB) under aerobic conditions moderate temperature (28˚C) and
neutral pH (70) However de-epoxidation could occur at 10-45˚C and pH 60-75 Under
anaerobic conditions near 100 de-epoxidation was observed after 72 h mainly during the
stationary growth phase Microbial DON de-epoxidation ability increased with increasing
DON concentrations in the culture media Inhibition of bacterial de-epoxidation by sodium
azide suggested that an active electron transport chain was required In addition to DON
strain ADS47 was able to de-epoxidize andor de-acetylate five type-A and five type-B
trichothecene mycotoxins This is the first report of a bacterial isolate capable of de-
epoxidizing DON and trichothecene mycotoxins under aerobic conditions and moderate
66
temperatures Strain ADS47 could potentially be used in bioremediation to eliminate DON
and other trichothecene mycotoxins from contaminated food and feed
Keywords aerobic de-epoxidation Citrobacter freundii isolation soil bacterium
trichothecenes
32 Introduction
Cereals are often contaminated with a variety of type-A and type-B trichothecene
mycotoxins produced by a number of toxigenic Fusarium species (Placinta et al 1999
Binder et al 2007 Foroud and Eudes 2009) Trichothecenes are tricyclic sesquiterpenes with
a double bond at the C9 10 position and a C12 13 epoxy group (Desjardins 2006) The two
types are distinguished by the presence of a ketone at position C8 in type-B which is absent
in type-A (Yazar and Omurtag 2008) The type-B toxin deoxynivalenol (DON) is the most
frequently found mycotoxin in food and feed around the world (Larsen et al 2004 Desjardins
2006 Starkey et al 2007) Different Fusarium species associated with Fusarium head blight
(FHB) or Gibberella ear rot (GER) disease of small grain cereals and maize are mainly
responsible for contaminating grains with DON and other type-A and type-B trichothecenes
These can cause serious mycotoxicosis in humans animals and plants (Ueno 1985 DrsquoMello
et al 1999 Bae and Pestka 2008) Common effects of DONTC toxicity in animals include
diarrhea vomiting feed refusal growth retardation immunosuppression reduced ovarian
functionsreproductive disorders and even death (Pestka and Smolinski 2005 Pestka 2007)
DON enhances FHB disease severity in wheat and maize (Proctor et al 1995 Desjardins
1996 Maier et al 2006)
67
Reduction of mycotoxin levels in contaminated agricultural products remains a
challenging task Current physical and chemical processes to decrease DON content are either
ineffective or prohibitively expensive (Kushiro 2008 He et al 2010b) However the use of
detoxifying microbes to biodegrade mycotoxins is an alternative (Awad et al 2010
Karlovsky 2011) The toxicity of DON is mainly due to the epoxide and there is a significant
reduction of DON toxicity (54) when it is reduced to produce de-epoxy DON Related
mycotoxins also show reductions in toxicity when they are de-epoxidized such as NIV (55)
and T-2 toxin (400) (Swanson et al 1988 Eriksen et al 2004) Bacillus arbutinivorans
strain LS100 and Eubacterial strain BBSH 797 originating from chicken intestine and rumen
fluid respectively are capable of cleaving the epoxide ring of DON under anaerobic
conditions and high temperature (37degC) (Fuchs et al 2002 Yu et al 2010) Strain BBSH 797
which can de-epoxidize DON scirpentriol and T-2 triol trichothecene mycotoxins has been
used as an additive for poultry and swine diets (Karlovsky 2011) However there are no
reports of microbes able to de-epoxidze DON and other trichothecenes under aerobic
conditions and at moderate temperatures
To develop an effective method for eliminating DON and trichothecene mycotoxins
from contaminated grains soils from fields with FHBGER infected crops were screened for
DON de-epoxidizing microbes (refer to chapter 2 Islam et al 2012) One soil sample yielded
a mixed microbial culture was partially enriched for bacteria with DON de-epoxidizing
activity (chapter 2) The current study describes the purification and characterization of a
novel DON de-epoxidizing bacterium from this culture
68
33 Materials and methods
331 Microbial source
A mixed microbial culture originating from a screen of more than 150 separate soil
samples collected from maize and wheat fields near Guelph Ontario Canada (Islam et al
2012) was used for enrichment purification and characterization of the DON de-epoxidizing
bacterium
332 Media and chemicals
The media used were nutrient broth (NB) 110 strength NB (110 NB) minimal
medium (MM) consisting of six mineral salts (MS) + 100g per liter sucrose (Islam et al
2012) MS + 05 Bacto Peptone (MSB) MS + 03 Bacto Peptone (MSC) MS + 05
yeast extract (MSY) MS + 200 microgmL DON (MS-DON) sterile water-soluble soil extract
(SE) (Islam et al 2012) potato dextrose broth (PDB) and Luria Bertani (LB) Prepared media
and media components were obtained from Fisher Scientific (Fair Lawn New Jersey USA)
Sigma-Aldrich (Oakville ON Canada) andor Difco (Sparks Maryland USA) Media pH
was adjusted as required prior to autoclaving For solid media preparation 15 agar was
added DON and all trichothecene mycotoxins were purchased from Biomin (Romer Lab
Union MO USA)
333 Isolation of the DON de-epoxidizing bacterial strain
Aliquots (20 microL) of the microbial culture partially enriched for DON de-epoxidizing
ability (Islam et al 2012) was transferred to 05 mL MSB (pH 70) in sterile 15 mL
Eppendorf tubes After addition of 200 microgmL pure DON the culture was incubated at room
69
temperature with continuous shaking at 220 revolutions per minute (rpm) Seven to ten days
later 20 microL of the culture was transferred to fresh MSB supplemented with 200 microgmL DON
The culture was then incubated for 7 to 10 days under the above conditions This procedure
was repeated 12 times
After three months of repeated transfers in broth with 200 microgmL DON 100 microL of the
culture was serially diluted in 09 mL MSB containing 05 Tweenreg20 (Sigma Aldrich) to
reduce bacterial cell aggregation (Mabagala 1997) The dilutions were vigorously shaken for
ten minutes A 100 microL aliquot from each dilution was spread on MSB + agar (MSA) The
MSA plates were incubated at room temperature until single colonies appeared
Single colonies from plates were transferred to 1 mL MSB+ 50 microgmL DON and
incubated at room temperature with continuous shaking at 180 rpm under aerobic conditions
After three days the DON de-epoxidation ability was examined by mixing an equal volume
of the bacterial culture and methanol and incubating for 30 min at room temperature to lyse
the cells Insoluble materials in the mixture were removed by passing it through a 045 microm
Millipore filter (Whatmanreg Florham Park New Jersey USA) The level of de-epoxy DON
(dE-DON also known as DOM-1) in the filtrate was determined by liquid chromatography-
ultraviolet-mass spectrometry (LC-UV-MS) (Guan et al 2009)
Individual colonies that showed de-epoxidation activity were serially diluted in
MSB+05 Tweenreg20 and plated onto MSA Single colonies that appeared on the agar plate
were incubated in MSB as described above and retested for DON de-epoxidation ability To
ensure that pure cultures were obtained this was repeated 10 times and individual colonies
were selected on MSA each time
70
334 Identification of a DON de-epoxidizing bacterium
Strain ADS47 with DON de-epoxidation ability was obtained from a single colony
after 10 serial transfers and was grown on MSA at room temperature under aerobic
conditions for 72 h Colony color shape edge and elevation were observed The cell
morphology of the bacterium was examined with a transmission electron microscope at the
University of Guelph Guelph Ontario Canada Whole bacterial cells were placed directly on
a formvar and carbon-coated 200-mesh copper grid They were washed two times by touching
the grids to a drop of clean dH2O and the excess water was blotted off The samples were
negatively stained using 1 Uranyl Acetate (aq) for five seconds Electron microscopy was
performed on a Philips CM10 transmission electron microscope at 60kV under standard
operating conditions (Philips Optics Eindhoven Netherlands) Gram staining was performed
using the procedure described in the commercial gram stain kit from BD Canada
(Mississauga Ontario Canada)
Total bacterial DNA was extracted using a DNeasy Plant DNA Extraction Mini Kit
(Qiagen Inc) The 16S-rDNA was PCR amplified using the primers 46f-
5rsquogcytaacacatgcaagtcga3rsquo (Kaplan and Kitts 2004) and 1371r-5rsquogtgtgtacaagncccgggaa3rsquo
(Zhao et al 2007) PCR was performed in 25 microL containing 06 microM of each primer 1x of
HotStarTaqreg Master Mix (Qiagen Inc) and ~40 ng of DNA The PCR cycles were 95ordmC (15
min) 35 cycles of 94ordmC (20 s) 52ordmC (15 s) 72ordmC (1 min 30 s) and a 72ordmC (7 min) extension
step Amplification was carried out in a GeneAmpreg PCR System 9700 (Applied Biosystems)
PCR products were sequenced with an automated ABI 3730 DNA analyzer (Applied
Biosystems) The 16S-rRNA gene sequence similarity was performed by BlastN using the
Ribosomal Database Project (RDP) (httprdpcmemsueduseqmatch)
71
A phylogenetic tree was constructed by comparing the partial 16S-rDNA sequence of
strain ADS47 with the 11 known Citrobacter species (Bornstein and Schafer 2006 Brenner et
al 1993) The Citrobacter strains were C freundii [FN997616] C koseri [AF025372] C
amalonaticus [AF025370] C farmeri [AF025371] C youngae [AB273741] C braakii
[AF025368] C werkmanii [AF025373] C sedlakii [AF025364] C rodentium [AF025363]
C gillenii [AF025367] and C murliniae [AF025369] Escherichia coli O157H7 [AE005174]
was used as an out group strain The analysis was performed using distance matrix with a gap
opening of 100 and a gap extension of 01 by a server based ClustalW2 program
(httpwwwebiacukToolsmsaclustalw2) A dissimilarity tree was created and viewed
using a TreeView X software (Page 1996)
335 Effects of DON concentration aeration substrate temperature pH and antibiotic
To determine if DON concentration can affect the growth of the isolated bacteria
tubes containing 5 mL of NB or MSB with 20 200 or 500 microgmL of DON were inoculated
with strain ADS47 that had been grown for 16 h with continuous shaking at 220 rpm and then
adjusted to cell density at 025 OD600nm (107 cellsmL) The cultures were incubated at 28degC
with continuous shaking at 180 rpm and 05 mL was removed every 12 h to measure
OD600nm The culture (05 mL) was then analysed for DON de-epoxidation activity by LC-
UV-MS (Guan et al 2009) as described above Three replicate cultures were tested
To examine the effect of aeration five mL of NB or MSB containing 50 microgmL DON
were inoculated with an overnight culture of strain ADS47 prepared as described above and
incubated at 28˚C with continuous shaking at 180 rpm One mL of culture was removed every
six hours and bacterial growth was determined by measuring cell density at OD600nm At the
72
same time points culture samples were analysed for DON to dE-DON transformation ability
(Guan et al 2009) The experiment was repeated with MSB medium containing 50 microgmL
DON under anaerobic conditions as described by Islam et al (2012) with DON to dE-DON
transformation ability being determined every 6 h Three replicate cultures were prepared for
each assay
The effect of substrate on DON de-epoxidation ability was tested after growing
ADS47 for 72 h in NB 110 NB MM MS-DON MSB MSC MSY SE PDB or LB
containing 50 microgmL DON For this assay the media were maintained at pH70plusmn02 and 28degC
for 72 h The effect of temperature was examined with MSB containing 50 microgmL DON at 10
15 20 25 30 35 40 45 or 50degC The effect of pH on growth of strain ADS47 was examined
with MSB adjusted to pH 55 60 65 70 75 or 80 and the cultures were incubated at 28degC
for 72 h Three replicate cultures were used for each assay
Overnight cultures of ADS47 in MSB were treated with 001 and 0001 (wv) of
an antibiotic sodium azide (SA) (Sigma-Aldrich) Controls were prepared in MSB without
SA Three replicate cultures were spiked with 50 microgmL DON DON to dE-DON
transformation ability of the bacterial culture was analysed (Guan et al 2009) after 72 h at
28degC with continuous shaking at 180 rpm Three replicate cultures were tested
336 Localization of DON to dE-DON transformation ability in the bacterium
Bacterial cell extracts were prepared using B-PER II Bacterial Protein Extraction
Reagent following the manufacturerrsquos instructions (PI 78260 Thermo Scientific Rockford
IL USA) ADS47 was grown in 15 mL MSB for 48 h under the growth conditions described
above A culture (05 mL) was centrifuged at 5000 x g for 10 minutes and the pellet was
73
collected B-PER II reagent (2 mL) was added to the pellet and the cells were lysed by
pipetting the suspension up and down several times The cell lysate was treated with 20 microL
Halt Protease Inhibitor Cocktail (Thermo Scientific Rockford IL USA) Culture filtrate was
prepared by passing 05 mL of the culture through a 022 microm Millipore filter (Fisher
Scientific) The remaining 05 mL culture was used as control Cell lysate culture filtrate and
control were incubated with 50 microgmL DON and incubated at 28degC DON to dE-DON
transformation ability of the cell lysate culture filtrate and control were analysed after 72 h
incubation (Guan et al 2009) Three replicate cultures were tested
337 Detoxification of additional trichothecene mycotoxins
Strain ADS47 was evaluated for its ability to degrade five type-A trichothecene
mycotoxins (HT-2 toxin T-2 toxin T2-triol diacetoxyscirpenol and neosolaniol) and five
type-B trichothecene mycotoxins (nivalenol verrucarol fusarenon X 3-acetyl-
deoxynivalenol and 15-acetyl-deoxynivalenol) One and a half mL NB was added with 100
microgmL of each mycotoxin and then inoculated with 100 microL of ADS47 culture grown
overnight as previously described The culture of NB + mycotoxin + ADS47 was incubated at
28degC with continuous shaking at 180 rpm and DON to dE-DON transformation ability of the
culture was analysed by LC-UV-MS (Guan et al 2009) Five replicate cultures were tested for
each trichothecene mycotoxin
74
338 Statistical analysis
Data were processed using Microsoft Excel Program 2007 Statistical analysis was
performed using General Linear Model procedure in SAS version 91 (SAS Institute Cary
NC USA 2002-03) Replications were averaged and stated as mean plusmn standard error The
mean differences were considered significant at plt 005 according to Tukeyrsquos-HSD test
75
34 Results
341 Isolation and identification of a DON de-epoxidizing bacterium
A DON de-epoxidizing bacterium designated strain ADS47 was isolated after 12
transfers over three months from cultures growing in MSB containing 200 microgmL DON
Strain ADS47 produced small to medium size circular convex and white colonies on MSA at
room temperature under aerobic conditions (Figure 3-1a) The bacterium is rod shaped and
150 x 072 microm in size (Figure 3-1b) It has numerous flagella that cover the surface of the cell
(peritrichous flagella) and is Gram-negative
BlastN analysis of the 16S-rDNA sequence of strain ADS47 showed the most
similarity to the eleven known Citrobacter species in the gamma proteobacteria of the
Enterobacteriaceae In particular the 16S-rDNA nucleotide sequence of ADS47 was more
than 99 identical to C fruendii (Figure 3-2) The next most similar sequences were a cluster
of C werkmanii and C murliniae followed by more distantly related C braakii C gilleni
and C youngae A separate cluster was formed by C rodentium C farmeri C sedlakii C
koseri and C amalonaticus
76
Figure 3-1 (a) Colony color shape and size of the mycotoxin degrading bacterial strain
ADS47 on mineral salts agar (MSA) medium (b) Transmission electron microscopic (TEM)
image of the Citrobacter freundii strain ADS47 The bacterium showed multiple flagella
(peritrichous flagella) and numerous fimbriae around the cell surface
(a) (b)
77
Figure 3-2 Dendrogram based on 16S-rDNA sequences for strain ADS47 and 11 reference
Citrobacter species An E coli O157H7 strain was considered as an out group species The
tree was constructed using a genetic distance matrix by ClustalW 2 and viewed by
TreeViewX software The distance was converted to scale and shown by branch lengths (scale
bar 001 which indicated the 16S-rDNA nucleotide sequence variability between the bacterial
species) GenBank accession numbers for 16S-rDNA sequences of the species used for
comparison are shown in materials and methods section (334)
78
342 Effects of DON concentrations on bacterial DON de-epoxidation
Growth of ADS47 aerobically in MSB with 20 200 and 500 microgmL DON was not
significantly different from that observed in control cultures without DON (Figure 3-3a)
Similarly increasing DON concentrations did not affect bacterial growth in NB medium
(Figure 3-3b)
DON de-epoxidation activity determined as microgmL DON convertedcell density
(OD600) was measured lowest in MSB+20 microgmL DON and highest in MSB+ 500 microgmL
DON containing cultures (Figure 3-3c) The highest rate of dE-DON formation was 115 microgh
in ADS47 cultures in MSB with 500 microgmL DON between 48 to 72 h All of the DON was
converted in the cultures containing 20 microgmL DON and 200 microgmL DON between 48-60 h
and 60-72 h respectively In 200 microgmL and 500 microgmL of DON cultures the rate of de-
epoxidation increased significantly after 36 h
79
00
04
08
12
16
0 24 36 48 60 72 96
OD
600n
m
Incubation time (h)
Control
20 microgmL
200 microgmL
500 microgmL
(b)
(a)
80
Figure 3-3 Effects of DON concentration on bacterial growth and de-epoxidation activities
Strain ADS47 was grown in MSB and NB media with three different DON concentrations 20
microgmL 200 microgmL and 500 microgmL Control cultures were prepared without DON Cultures
were grown at 28degC with continuous shaking at 180 rpm (a) Microbial density (OD600 nm) in
MSB with different DON concentrations was measured at 24 36 48 60 72 and 96 h (b)
Microbial growth (OD600 nm) in NB having different DON concentration was measured at 24
48 72 and 96 h (c) Bacterial DON de-epoxidation activity in 20 200 and 500 microgmL DON
containing cultures at different time points DON de-epoxidation was measured in microgmL per
OD600 nm Three replicate cultures for each assay were analysed
(C)
81
343 Influence of growth conditions and antibiotic on microbial DON de-epoxydation
Aerobic and anaerobic conditions
ADS47 grew more quickly uner aerobic conditions under MSB+50 microgmL DON than
in MSB+50 microgmL DON under anaerobic conditions (Figure 3-4a) Aerobic growth was
faster in NB+50 microgmL DON compared to MSB+50 microgmL DON
However the DON de-epoxidation activities on a cell density basis were very similar
in NB+50 microgmL DON compared to MSB+50 microgmL DON (Figure 3-4b) Also DON de-
epoxidation activity of ADS47 under aerobic conditions in MSB+50 microgmL DON was higher
than in the same medium under anaerobic conditions (Figure 3-4b)
82
(b)
(a)
83
Figure 3-4 Bacterial growth and DON de-epoxidation capabilities under aerobic and
anaerobic conditions (a) Microbial growth under aerobic conditions in MSB and NB medium
and anaerobic conditions in MSB medium (b) Microbial DON de-epoxidation ability under
aerobic conditions in MSB and NB media and anaerobic conditions in MSB medium
Cultures media having 50 microgmL DON were inoculated with overnight culture of strain
ADS47 Initial microbial concentration in the cultures was adjusted to 025 OD600nm Cultures
were incubated at 28degC having three replications Bacterial growth at OD600nm and DON de-
epoxidation in microgmL per OD600nm were measured at six hour interval
84
Media temperature and pH
Growth of ADS47 in media containing 50 microgmL DON under aerobic conditions at 72
h was three-fold higher in NB and LB than in SE two-fold higher than in 110 NB and MSC
and did not occur in MS-DON (Figure 3-5a) Growth in MM MSB and MSY was
intermediate
Over the 72 h DON de-epoxidation per cell density was highest in MSB and MSY and
approximately 25 lower in NB and LB and lower by 75 and 87 in 110 NB and MSC
respectively (Figure 3-5b) No DON de-epoxidation was observed in MS-DON in which
DON was used as a carbon source Bacteria grown in MM broth which contains sucrose as a
sole carbon source were unable to de-epoxidize DON No DON de-epoxidation was observed
in SE and PDB grown cultures
Growth of ADS47 under aerobic conditions for 72 h in MSB+50 microgmL DON was
greatest between 25-40degC but no growth occurred at 50degC (Figure 3-6a) DON de-
epoxidation activity was greatest and equivalent between 20-40degC and lower at 10 15 and
45degC temperatures (Figure 3-6b)
ADS47 growth under aerobic conditions for 72 h in MSB+50 microgmL DON was
limited to neutral pHs (ie 60 - 75) (Figure 3-7a) The transformation of DON to dE-DON
occurred at the same pH range and DON de-epoxidation activity was not significantly
different between pH 60 to 75 (Figure 3-7b)
85
b
c
a
b
a
d c
b b
(a)
(b)
b
c
b
a a
d
86
Figure 3-5 Effects of media on bacterial growth and DON de-epoxidation activity (a)
Microbial growth (at OD600nm) on different culture media Media pHs were adjusted to
70plusmn02 (b) Bacterial DON to dE-DON transformation in microgmL by OD600nm in different
media after 72 h The cultures having three replications were incubated for 72 h at 28degC and
DON de-epoxidation was measured The Media NB = nutrient broth 1NB = 110 strength
NB MM = minimal medium MS-DON = six mineral salts + 200 microgmL DON as sole C
source MSB = six mineral salts + 05 bacto peptone MSC = six mineral salts broth + 03
bacto peptone MSY = six mineral salts with 05 yeast extract SE = soil extract PDB =
potato dextrose broth and LB = Luria Bertani Means with the same label are not significantly
different according to Tukeyrsquos HSD test (p lt 005)
87
(a)
(b)
88
Figure 3-6 Effects of temperature on microbial growth and DON de-epoxydation capability
(a) Microbial growth (at OD600nm) at different temperatures after 72 h of incubation (b)
Bacteria DON de-epoxidation in microgmL by OD600nm in different temperatures The most
efficient temperature range (ge40 microgmL DON de-epoxidation) is marked by dot lines The
experiment was conducted in MSB medium at pH 70plusmn02 with three replications
89
(a)
(b)
90
Figure 3-7 Influence of pH on microbial growth and DON de-epoxydation (a) Microbial
growth (OD600nm) in MSB at different pH (b) Bacterial DON to de-epoxy-DON
transformation in microgmL by OD600nm in different pH Dot lines indicated the efficient de-
epoxidation (ge40 microgmL DON conversion) pH range Experiment was conducted at 28degC
with three replications
91
Antibiotic
Treatment of ADS47 at 72 h in MSB+50 microgmL DON with sodium azide (SA) at
0001 and 001 reduced DON de-epoxidation activity to 5 (25 microgmL) and 0
respectively The control with no SA added was able to de-epoxidize DON completely
(100) (Figure 3-8)
Figure 3-8 Effects of sodium azide (SA) on bacterial DON de-epoxidation activity
Microbial culture in MSB+50 microgmL DON was treated with 001 (SA-1) and 0001 (SA-
2) sodium azide In control (C) no SA was added The experiments with three replications
were incubated for 72 h at 28degC and DON de-epoxidation was measured by LC-UV-MS
method
0
20
40
60
80
100
120
SA1 SA2 C
D
ON
de
-ep
oxyd
ize
d
92
344 Localization of bacterial DON de-epoxidation
A cell lysate prepared from ADS47 grown for 48 h in MSB transformed 20 (10
microgmL) of DON added at 50 microgmL to dE-DON within 72 h (Figure 3-9) In contrast a cell-
free culture filtrate did not show DON de-epoxidation activity
Figure 3-9 Determination of DON to de-epoxy DON transformation by bacterial culture
filtrate and cell extract The treatments having three replications were spiked with 50 microgmL
DON and incubated at 28degC for 72 h In control strain ADS47 was incubated in MSB broth
with 50 microgmL DON DON de-epoxidation activity in percentage was examined
0
20
40
60
80
100
120
Filtrate Extract Control
D
ON
de
-ep
oxyd
ize
d
93
345 Bacterial detoxification (de-epoxidtaion and de-acetylation) of other trichothecene
mycotoxins
Aerobic cultures of ADS47 grown for 72 h in NB+100 microgmL HT-2 converted 74 of
the toxin to de-epoxy HT-2 toxin and 26 to a de-acetylated product (Figure 3-10 and Table
3-1)
In a culture treated with T2-toxin 46 was converted to de-epoxy T-2 16 was
converted to an unknown compound and 38 was not converted In ADS47 culture with
diacetoxyscirpenol (DAS) 61 was transformed into de-epoxy DAS and 39 was de-
acetylated DAS (39) Neosolaniol (NEO) was transformed to de-epoxy NEO (55) and
de-acetylated NEO (45) T2-triol was transformed in to de-epoxy T2-triol (47) and de-
acetylated T2-triol (53)
Aerobic cultures of ADS47 were also grown for 72 h in NB+100 microgmL of one of five
type-B trichothecenes For nivalenol (NIV) 92 was de-epoxydized while the remaining 8
NIV was transformed to DON 3-acetyl-deoxynivalenol (3-ADON) was transformed into
three products de-epoxy NIV (5) de-epoxy DON (54) and DON (41) For 15-
acetyldeoxynivalenol (15-ADON) 8 was transformed to de-epoxy 15-ADON and 92 to
de-epoxy DON (dE-DON) FusarenonX (FUS) had 95 transformed into de-epoxy FUS
while 97 of verrucarol (VER) was transformed to de-epoxy VER The remaining 5 FUS
and 3 VER were not transformed
94
Figure 3-10 Determination of microbial de-epoxidation and de-acetylation of HT-2 toxin by
LC-UV-MS Bacterial culture in NB was added with 100 microgmL HT-2 toxin Bacterial HT-2
toxin transformation ability was examined after 72 h of incubation at 28degC The upper panel
illustrated mass spectrometry chromatograph of the de-acetylated (red peak) and de-
epoxydized (blue peak) products Arrows indicated the targeted acetyl (red) and epoxy (blue)
functional groups of HT-toxin The middle and lower panels showed the mass spectra of the
de-acetylated (dA) and de-epoxidized (dE) compounds Arrows on the upper panel indicated
the targeted acetyl (red) and epoxy (blue) functional groups of HT-2 toxin Red and blue
arrows on the middle and lower panels pointed at the degradation sites of HT-2 toxin
95
Table 3-1 Microbial transformation of type-A and type-B tricthothecene mycotoxins after 72
h of incubation under aerobic conditions at room temperature DAS=diacetoxyscirpenol
DON=deoxynivalenol FUS=FusarenonX 3AON=3-acetyl-deoxynivalenol and
15ADON=15-acetyl-deoxynivalenol
1 Convertion of trichothecene mycotoxins to either DON or an unknown compound
Trichothecene
mycotoxin
Untransformed
trichothecenes ()
Transformed products of
trichothecenes ()
De-
epoxy
De-
acetylate
Other1
Type-A
HT-2 toxin 0 74 26 -
T-2 toxin 38 46 - 16
(unknown)
DAS 0 61 39 -
Neosolanol 0 55 45 -
T2-triol 0 47 53 -
Type-B
DON 0 100 - -
Nivalenol 0 92 - 8 (DON)
3-ADON 0 59 41 (DON)
15-ADON 0 100 - -
FUS 5 95 - -
Varrucarol 3 97 - -
96
35 Discussion
The ADS47 strain of C freundii was isolated in the present study by a prolonged
incubation of a mixed microbial culture with a high concentration DON since the bacterium
efficiently de-epoxidized (detoxified) DON and number of other trichothecene mycotoxins
under aerobic and anaerobic conditions at moderate temperatures and neutral pH The total
procedures for enrichment and isolation of the strain ADS47 is depicted in supplemental
Figure S3-1 The reductive de-epoxidation activity may be due to a cytoplasmic enzyme
which would be in agreement with a previous study that showed de-epoxidation of
epoxyalkanes was catalyzed by a bacterial cytoplasmic flavoprotein (Westphal et al 1998)
The ability of the currently isolated strain of C freundii to biodegrade environmental
pollutants such as aminoaromatic acids and heavy metals (copper) is also likely mediated by
cytoplasmic enzymes since these compounds need to be taken up by the cells before they are
metabolized (Savelieva et al 2004 Sharma and Fulekar 2009) In contrast oxidative
transformation of DON to 3-keto DON by microbial extracellular enzyme has been reported
(Shima et al 1997)
Isolation of a trichothecene de-epoxidizing microorganism from a mixed microbial
community is very challenging Although trichothecenes are toxic to eukaryotes prokaryotes
appear to be resistant to these mycotoxins (He et al 1992 Rocha et al 2005) The present
study showed that the bacteria grew well in media with high concentrations of DON (500
microgmL) Bacterial de-epoxidation of DON involves the removal of an oxygen atom from the
functional epoxy ring of the tricyclic DON structure (Yoshizawa et al 1983) Such co-
metabolic degradation does not provide the bacteria with any nutrients from the reacting
compounds (Horvath 1972 Fedorak and Grabic-Galic 1991) This is supported by the results
97
of the current study which showed that strain ADS47 was unable to grow in media with DON
as a sole carbon source and increasing the concentration of DON did not increase ADS47
growth in MSB Hence DON could not be used as sole nutrient source in the mixed culture
enrichment process
Many previous attempts to obtain DON de-epoxidizing microbes have failed
particularly under aerobic conditions because media or laboratory conditions were
inappropriate to support growth or initially active cultures lost de-epoxidizing function after
serial transfer (Karlovsky 2011)
Media composition appears to be a crucial factor for microbial trichothecene de-
epoxidation activity (Voumllkl et al 2004) Effective biotransformation of trichothecene
mycotoxins by the fish digesta microbial culture C133 occurred in full media (FM) but a low
level of de-epoxidation occurred in media having a single nitrogen source and no
transformation activity was observed in the common bacterial medium LB (Guan et al
2009) Zhou et al (2010) also showed that chitin de-acetylation by soil bacterial strains
occurred more rapidly in yeast extract compared to common bacterial media Therefore a
screening strategy for microbial DON de-epoxidation ability should use different media in
parallel as was done in this study
This study observed efficient de-epoxidation in common bacterial media (NB and LB)
as well as in mineral salts medium containing yeast extract (MSY) or 05 Bacto Peptone
(MSB) However reduction of Bacto-Peptone to 03 (MSC) significantly reduced the
bacterial de-epoxidation activity suggesting that the activity is sensitive to the availability of
protein and amino acids The study found that medium containing sucrose as carbon source
(MM) and a commonly used medium PDB blocked microbial DON de-epoxidation even
98
though bacteria grew in those growth media These suggest that media containing high sugar
contents had inhibitory effects on microbial de-epoxidation enzymatic activity Previously
sugars have been found to have negative effects on the production andor activity of bacterial
enzymes (Saier and Roseman 1971)
Losses of microbial functions under laboratory conditions limited researchers to obtain
economically important microbial genetic sources The soil borne Curtobacterium sp strain
114-2 efficiently de-epoxydized nivalenol but unfortunately the strain lost its activity in
latter generations (Ueno et al 1983) Instability of de-epoxidation activity of ADS47
observed in cultures grown on agar media without DON resulted in the loss of DON de-
epoxidation activity by more than 80 of the colonies after one generation However strain
ADS47 retained de-epoxidation activity after ten subcultures in MSB (data not shown)
The non-specific de-epoxidation of DON made it impossible to use DON as a sole
substrate C source for selective isolation of ADS47 Thus a number of techniques were
utilized to select ADS47 and suppress other bacteria including the use of a variety of
antibiotics and culture at elevated temperatures (Islam et al 2012) Selection of the isolate
ADS47 required repeated subculturing and prolonged exposure to media containing 200
microgmL DON After three months of selection with DON culture that was obtained had a
DON de-epoxidation activity that was up to 30-fold higher than the mixed culture As growth
of ADS47 was not affected by 200 microgmL DON the prolonged incubation may have reduced
populations of other bacteria that are more sensitive to DON Enrichment and isolation of a
DON to 3-keto DON transforming bacterial strain E3-39 by prolonged incubation of a soil
microbial culture with 200 microgmL DON was reported by Shima et al (1997)
99
This study demonstrated that microbial DON de-epoxidation occurred mainly at the
stationary growth phase A DON degrading strain of Citrobacter may have gained energy
from DON by co-metabolic cleavage of the epoxy ring under less nutrient conditions
Microbial use of energy from co-metabolic reaction has been suggested by Chiu et al (2005)
The rate of de-epoxidation activity increased when ADS47 grew at higher concentrations of
DON Taken together a prolonged exposure to high concentration DON may have resulted in
non-DON metabolizing microbial populations declining by DON toxicity while facilitated
the population of DON metabolizing bacterium by providing energy
DON is a small molecule and is simply absorbed by cells (Sergent et al 2006)
Microbial DON de-epoxidation occurred in intact and lysed bacterial cells In addition
treating the ADS47 culture with SA which blocks gram negative bacterial electron transport
and activity inhibited DON de-epoxidation activity (Lichstein and Malcolm 1943 Erecinska
and Wilson 1981) implying that an active electron transport system required for microbial de-
epoxidation reaction (He et al 1992) The data suggest that a cytoplasmic epoxy reductase is
involved in the DON de-epoxidation reaction in the ADS47 microbial strain that was isolated
in the present study
Since agricultural commodities are often contaminated with multiple fungal
mycotoxins the ability of bacteria like ADS47 to degrade mycotoxins by several
mechanisms may be an advantage De-acetylation might be another mechanism of microbial
mycotoxin detoxification since toxicity of trichothecenes also relies on the acetyl functional
groups (Desjardins 2006) Strain ADS47 showed efficient de-epoxidation and de-acetylation
of several type-A and type-B trichothecene mycotoxins that are commonly detected in cereals
in association with DON In conclusion the soil-borne gram-negative bacterium C freundii
100
strain ADS47 is capable of detoxifying trichothecene mycotoxins by both de-epoxidation and
de-acetylation under aerobic and anaerobic conditions neutral pH and a range of
temperatures ADS47 has great potential to detoxify DON contaminated animal feed
Furthermore isolation of novel bacterial de-epoxidation genes might be used to generate
transgenic plants that can detoxify DON in contaminated seed Alternatively contaminated
seed might be treated with manufacture de-epoxidation enzyme(s) These applications can
improve the quality and safety of agricultural commodities that are prone to trichothecene
mycotoxin contamination
36 Acknowledgements
Thank to the Natural Sciences and Engineering Research Council of Canada (NSERC) for a
scholarship to Rafiqul Islam The work was funded by the Agriculture and Agri-Food Canada
Binational Agricultural Research and Development Fund (AAFCBARD) project We
appreciate Honghui Zhu for technical assistance
101
Supplemental Figure S3-1 Flow chart depicts the procedures of enrichment and isolation of
DON de-epoxidation bacterial strain ADS47 Steps 1 to 4 describe the optimization of
microbial DON to dE-DON transformation activity and removal of undesired microbial
species Step 5 to 7 shows the culture enrichment by incubation with high concentration DON
and isolation of the single strain ADS47
Step 1 Screening of soil samples
Step 2 Optimization of cultural conditions
Step 3 Elimination of undesired microbial species
Step 4 Removal of untargeted microbial species
Step 5 High concentration DON enrichment
Step 6 Separation of bacterial cells and dilution plating
Step 7 Single colony analysis
Active culture was sequentially treated with different mode of actions antibiotics heating cultures at 50degC and subculturing on agar media
Colonies were tested for their DON de-epoxidation ability Purified the single colony via several times subculturing on agar medium
Optimized the culture conditions ie growth substrates temperatures pH and oxygen requirements
Obtained DON de-epoxidation capable microbial culture obtained by analyzing soil samples collected from cereal fields
Serially diluted active culture was grown on agar media Collected washed colonies having DON de-epoxidation activity
The partially enriched culture that retained DON de-epoxidation activity was incubated with 200 microgmL DON in MSB media with continuous shaking at 220 microgmL for 7 d The procedure was repeated 12 times
The enriched culture was serially diluted and treated dilutes with 05 Tween 20 After vigorously vortex for ten min aliquot cultures were spread on different agar media plates to produce single colonies
102
CHAPTER 4
40 CONSTRUCTION AND FUNCTIONAL SCREENING OF GENOMIC DNA
LIBRARY FOR ISOLATION OF DON DE-EPOXYDIZING GENE(S) FROM
CITROBACTER FREUNDII STRAIN ADS47
41 Abstract
Enzymatic activity-based screening of a genomic DNA library is a robust strategy for
isolating novel microbial genes A genomic DNA library of a DON detoxifying bacterium
(Citrobacter freundii strain ADS47) was constructed using a copy control Fosmid vector
(pCC1FOSTM
) and E coli EPI300-T1R as host strain A total of 1250 E coli clones containing
large DNA inserts (~40 kb) were generated which resulted in about 10-fold theoretical
coverage of the ADS47 genome The efficiency of E coli transformation with the
recombinant Fosmid vector was examined by restriction digestion of Fosmids extracted from
15 randomly selected library clones The fragmentation patterns showed that all of the clones
contained an 81 kb Fosmid backbone and ~40 kb of insert DNA except one that had a small
insert This suggests that a DNA library was prepared with expected DNA inserts The de-
epoxidation ability of the library was analysed by incubating clones in Luria Bertani (LB)
broth medium supplemented with 20 microgmL DON with induction (by addition of L-arabinose)
or without induction None of the 1000 clones showed DON de-epoxidation activity The
potential DON de-epoxidation gene(s) of ADS47 may not have been expressed in E coli or
might have been expressed but lacked additional factors required for DON de-epoxidation
103
Keywords DNA library fosmid vector functional library screening DON de-epoxidation
activity
42 Introduction
Microorganisms contain numerous valuable genes and biocatalysts which may have
significant roles in agricultural productivity (Jube and Borthakur 2007) Various techniques
have been used to isolate novel genes from different organisms including transposon
mutagenesis (Frey et al 1998 Tsuge et al 2001) gene knockout (Winzeler et al 1999)
whole genome sequencing (Krause et al 2006) comparative genomics (Koonin et al 2001)
and suppression subtractive hybridization (Dai et al 2010)
The genomic DNA library preparation and activity-based screening of the library is a
relatively new technique which has been efficiently used for isolation of microbial novel
enzymes and compounds such as lipases proteases oxidoreductases antibiotic resistance
genes membrane proteins and antibiotics (Henne et al 2000 Gupta et al 2002 Knietsch et
al 2003 Gabor 2004 Suenaga et al 2007 van Elsas et al 2008) The strength of the
technique is that genes from microbial sources could be obtained without prior sequence
information and not requiring culturing the organisms under laboratory conditions Using this
technique an unknown gene or a set of genes could be obtained by determining particular
phenotypes of the library clones Eventually the gene of interest from the positive clones is
isolated by using a forward (eg mutation) or reverse genetics method (eg subcloning the
positive clones into a microbial expression vector) followed by functional analysis of the
subclones
104
Although this strategy has been used effectively for isolating novel microbial genes
several factors need to be considered in order for the method to be successful These factors
include DNA quality vector type (eg plasmid cosmid fosmid bacterial artificial
chromosome etc) vector copy number and host strains (Daniel 2005 Singh et al 2009) The
Fosmid vector-based technique has been used successfully to clone bacterial genes including
xenobiotics degradation genes antifungal gene clusters and antibiotic resistance genes from
soil metagenomes (Hall 2004 Suenaga et al 2007 Chung et al 2008) This study reports the
construction and activity-based screening of a Fosmid DNA library in an E coli host with the
aim of isolating a novel DON de-epoxidation gene from Citrobacter freundii strain ADS47
(Chapter 3)
43 Materials and methods
431 Genomic DNA library construction
The genomic DNA library was prepared using a Copy Control Fosmid Library
Production Kit (Epicentre Biotechnologies Madison Wisconsin USA) following the
manufacturerrsquos instructions as depicted in Figure 4-1
105
Figure 4-1 Schematic overview of the construction and functional screening of a Fosmid
DNA library
106
Citrobacter freundii strain ADS47 was grown in NB medium at 28degC with continuous
shaking at 220 rpm The high molecular weight bacterial genomic DNA was extracted from
the overnight culture using the Gentra Puregene YeastBacteria Kit (Qiagen Inc Mississauga
Ontario Canada) The purity of the DNA was confirmed by determining OD260280 at ~190 by
a NanoDrop spectrophotometer A 5 microL sample having ~25 microg high molecular weight DNA
was sheared by passing through a 200-microL pipette tip and achieved a sufficient amount of
DNA about 40 kb in size Blunt ended 5rsquo-phosphorylated insert DNA was generated by
thoroughly mixing it with End-Repair Enzyme reagents on ice The 20 microL mixture was
incubated for 45 min at room temperature followed by incubation at 70degC for 10 min to
inactivate the End-Repair Enzyme activity
The treated DNA was size fractionated by a Low Melting Point-Pulsed Field Gel
Electrophoresis (LMP-PFGE) on the clamped homogeneous electric fields (CHEF-DR III)
system (Bio-RAD Laboratories) using 1 UltraPure Low Melting Point Agarose (Invitrogen)
The PFGE was conditioned according to manufacturerrsquos instructions as follows initial switch
time-42 s final switch time-761 s run time 16 h 50 min angle-120deg gradient- 6 Vcm
temperature-14degC and running buffer-05 TBE DNA size markers for the 42 kb Fosmid
control DNA (Epicentre Biotechnologies) and low range Pulse Field Gel (PFG) markers (New
England Biolabs Inc) were loaded adjacent to the DNA sample slots After electrophoresis a
portion of the gel that contained DNA size markers was cut off The gel was stained in SYBR
Safe DNA gel stain (Invitrogen) for 30 min visualized and marked on the 23 42 and 485 kb
marker bands A slice of PFGE gel containing 40 plusmn 10 kb size DNA was cut off with a sterile
razor blade by comparing the marker sizes DNA from the gel was recovered by digesting it
with GELase enzyme ( 1 U100 microL melted agarose) in 1X GELase Buffer followed by a
107
sodium acetate-ethanol precipitation The DNA with the desired size was stored at 4degC for
next dayrsquos ligation to a cloning-ready Fosmid vector (pCC1FOSTM
)
Ligation was performed in a 10 microL reaction mixture by thoroughly mixing 10X Fast-
Link Ligation Buffer 10mM ATP 025 microg of ~40 kb insert DNA 05 microg Fosmid Copy
Control Vector (pCC1FOS) (Figure 4-2) and 2 U Fast-Link DNA lipase The mixture was
incubated for 4 h at room temperature preceded by inactivating the ligation reaction by
heating at 70degC for 15 min The recombinant Fosmid vector was stored at -20degC for later use
Ten microL of the ligated DNA was thawed on ice and transferred to a pre-thawed
MaxPlax Lambda Packaging Extract tube containing 25 microL extract By pipetting several
times the mixture was incubated at 30degC for lamda packaging After 2 h of packaging an
additional 25 microL packaging extract was added into the mixture and incubated for a further 2 h
at 30degC The packaged phage particles were volumed to 05 mL by adding ~400 microL Phage
Dilution Buffer and 25 microL chloramphenicol
Transformation was performed via transduction by mixing the packaged particles with
the overnight cultured E coli EPI300-T1R cells (1 10 vv ratio) in LB supplemented with 10
mM MgSO4 and 02 Maltose Transformation was completed by incubation at 37degC for 1 h
The transformed E coli cells were spread on an LB agar plate containing 125 microgmL
chloramphenicol The plates were incubated overnight at 37degC Colonies that appeared on the
plates were individually transferred to 96-well microtiter plates having 200 microL LB medium
supplemented with 10 glycerol and 125 microgmL chloramphenicol The microtiter plates
were incubated at 37degC for 24 h with continuous shaking at 180 rpm The prepared library
clones were stored at -80degC for further analysis and functional screening
108
109
Figure 4-2 Genetic map of copy controlled Fosmid vector (pCC1FOSTM
) The 813 kb
Fosmid vector backbone shows a multiple cloning site (MCS) chloramphenicol resistance
selectable marker (CmR) replication initiation fragment (redF) inducible high copy number
origin (oriV) replication initiation control (repE) and F-factor-based partitioning genes (ParA
ParB ParC) The vector contains a cos site for lamda packaging (not shown) and lacZ gene
for bluewhite colony selection The vector map was provided by the Epicentre
Biotechnologies (httpwwwepibiocomitemaspid=385)
110
432 Confirmation of library construction
The quality of the DNA library prepared was determined by examining 15 E coli
clones randomly selected A 15 mL LB broth supplemented with 125 microgmL
chloramphenicol was inoculated by each of the transformed clones To obtain higher Fosmid
DNA yield 1 microL autoinduction solution (L-arabinose) to increase the copy number of the
Fosmid vector (Epicentre) was added to the cultures After an overnight incubation of the
cultures at 37degC with continuous shaking at 220 rpm Fosmid DNA was extracted using a
Qiagen Large-Construct Kit followed by restriction digestion with Not I enzyme A 20 microL
digestion mixture containing 05 microg Fosmid DNA 1X digestion buffer and 05 U Not I was
incubated at 37degC for 15 h The partially digested DNA and a 42 kb DNA size marker
(Fosmid control DNA) were ran on 08 agrose gel for 15 h at 100 V The linear Fosmid
vector backbone (813 kb) and insert DNA fragments were visualized on the gel and the
image was recorded
433 Functional screening of the clones
Eppendorf tubes (15 mL) containing 500 microL LB supplemented with 125 microgmL
chloramphenicol were inoculated with a single clone (400 clones tested) or a combination of
12 library clones (600 tested) from microtiter plates stored at -80degC One half of the cultures
in each group (single or combined clones) were induced to increase the Fosmid copy number
by adding a 1 microL (500X) autoinduction solution (contained L-arabinose) to the culture tubes
L-arabinose induces the expression of a mutant trfA gene of the E coli host strain which
leads to the initiation of Fosmid replication from a low-copy to a high-copy number origin of
replication (oriV) The cultures were spiked with 20 microgmL DON and incubated at 37degC for
111
72 h with continuous shaking at 180 rpm The DON to de-epoxy DON transformation ability
of the clone cultures was examined by a liquid chromatography-ultraviolet-mass spectrometry
(LC-UV-MS) method as described by Islam et al (2012)
112
44 Results
441 Production of the DNA library
High molecular weight intact (non-degraded) and good quality (OD600280 ~190)
genomic DNA was obtained from the DON de-epoxidizing bacterial strain ADS47 (data not
shown) Pipetting of the DNA extract resulted in an adequate amount of insert DNA (gt025
microg) of approximately 40 kb size as showed on a LMP-PFGE (Figure 4-3)
A DNA library in E coli host was constructed using LMP-PFGE purified DNA The
library was comprised of 1250 E coli transformants which were selected on LB agar plates
supplemented with chloramphenicol (Figure 4-4)
113
Figure 4-3 Preparation of a 40 kb size insert DNA from a DON de-epoxidizing bacterium
Genomic DNA was extracted from the overnight culture of a DON de-epoxidizing
Citrobacter freundii strain ADS47 After fragmentation the DNA was size fractionated by
Low Melting Point- Pulse Field Gel Electrophoresis (LMP-PFGE) with markers for the PFGE
(M1) and 42 kb Fosmid control DNA (M2) S1 S2 S3= different DNA samples extracted
from bacterial strain ADS47 The red dots area indicates the section of the gel containing
genomic DNA of the expected size (~40 kb)
114
Figure 4-4 Production of the genomic DNA library in E coli EPI300-T1 host strain
Approximately 40 kb long blunt ended genomic DNA from a DON de-epoxidizing bacterial
strain ADS47 was ligated to a Copy Control Fosmid vector pCC1FOS The construct was
packaged into lamda bacteriophage followed by a transduction-mediated transformation of E
coli host cells The clones which appeared on the LB agar plate supplemented with 125
microgmL chloramphenicol are expected to be transformed by the recombinant Fosmid vector
containing insert DNA
115
442 Conformation of the library construction
All of the clones tested acquired the Fosmid vector Out of 15 clones examined 14
received the recombinant Fosmid vector that contain the expected size insert DNA as
determined by an 81 kb vector backbone and at least one high molecular weight insert DNA
band (~40 kb) on agarose gel (Figure 4-5) In contrast one clone (lane 4) showed an 81 kb
vector backbone and a low molecular weight band which is also a fragment of the Fosmid
vector The result indicated that approximately 94 of the library clones were transformed
with the recombinant vector
443 Activity-based screening of the library
Out of 500 single copy number clones (200 individual and 300 combined clones)
none showed DON to de-epoxy DON transformation activity (Table 4-1) An additional 500
high copy number clones (200 single and 300 combined clones) were unable to de-epoxidize
DON as well
116
Figure 4-5 Confirmation of the Fosmid DNA library production in E coli Fosmid DNA
collected from 15 clones was digested with Not I and run on 08 agarose gel at 100 V for 15
h The presence of the vector backbone (81 kb) and the insert DNA fragment (~40 kb) in the
clones was determined by comparing a 42 kb molecular size marker (M) Lanes (1-15) denote
the recombinant Fosmid DNA extracted from 15 E coli clones
117
Table 4-1 Analysis of Fosmid DNA library clones for deoxynivalenol (DON) de-epoxidation
capability
Type of clones Analysis of
singlecombined
clones
Number of
clones
examined
Number of clones
showed DON de-
epoxidation activity
Without induction
(single copy number clones)
Singly 200 0
Combination of 12 300 0
With induction (multi-copy
number clones)
Singly 200 0
Combination of 12 300 0
De-epoxidation activity of the clones was examined alone or in combination of 12 clones
without or with induction by adding L-arabinose The clones grown in LB supplemented with
125 microgmL and 20 microgmL DON were analysed for DON to de-epoxy DON transformation
ability by a liquid chromatography-ultra violet- mass spectrum (LC-UV-MS)
118
45 Discussion
While a Fosmid DNA library with the expected genome coverage of a DON de-
epoxidizing bacterial genome was produced the function-derived screening of the library
clones did not identify any clones that showed DON de-epoxidation enzymatic activity This
is unlikely due to lack of coverage of the bacterial genome as 1000 clones analyzed in the
study should have provided eight fold coverage of the genome Therefore it was likely due to
a lack of expression of the enzyme(s) for DON de-epoxidizing activity under the conditions
used
Thus far there is only one report of heterologous expression of bacterial epoxide
reductase gene in another bacterium A high level of heterologous expression of a
Synechococcus sp vitamin K epoxide reductase gene was achieved in E coli with codon
optimization and placing the gene under the control of a prokaryotic T7 promoter in an
expression vector (Li et al 2010) Therefore heterologous expression of bacterial epoxide
reducatse gene using its own promoter has not yet been demonstrated The success of the
method used in this chapter relies on an intact copy of the gene or operon being cloned the
selection of an appropriate cloning vector a host organism that can recognize the cloned C
fruendii promoter and a suitable library screening method for DON degradation by E coli
(Mocali and Benedetti 2010)
The unbiased cloning efficiency of the Fosmid vector its ability to contain large insert
DNA high stability in E coli as single copy number and autoinduction to increase copy
number in host cells made it appear to be a suitable method for the present work (Kim et al
1992 Lee et al 2004) Library production with large insertions is a major advantage as gene
cluster involved in particular pathway could be trapped into single clone and their functions
119
could be analysed The pathway of microbial DON de-epoxidation activity and the genes
encoding the activity are not known Therefore it was reasonable to prepare a library with
large inserts to ensure the possibility of cloning single as well as whole pathway genes of
interest for functional screening A further advantage of the method is that Fosmid clones are
inducible which allows detecting poorly expressed genes in a heterologous host by inducing
high copy numbers (Daniel 2005)
One of the limitations of the method is that the functions of the target genes rely
mostly on the host organismrsquos transcription and translation machineries Therefore selection
of the host organism is critical for activity-based screening of library clones in heterologous
hosts A number of host strains including E coli Streptomyces lividans Rhizobium
leguminosarum and Pseudomonas aeruginosa have been used for functional screening
activities from diverse organisms (Daniel 2005) The present work attempted to isolate C
freundii strain ADS47 genes encoding DON de-epoxidation enzymes Both strain ADS47 and
E coli belong to gamma proteobacteria which suggest that E coli would be an appropriate
host organism for heterologous expression of ADS47 genes Poor expression of heterologous
genes could be another limitation of the method which may be overcome by overexpression
to increase the catalytic activity of less expressed genes However clones with DON de-
epoxidation activity were not identified in this study even the Fosmid copy number was
increased by induction
In general a large number of clones must be analysed to obtain the gene of interest
using an activity-based screening technique (Lee et al 2004 van Elsas et al 2008) In this
study the constructed DNA library comprised of 1160 clones having approximately ~40 kb
insert DNA suggesting that the prepared library achieved ~10 fold coverage of the ADS47
120
genome (48 Mb) which ensured gt99 probability of the presence of all DNA sequence in
the library In the produced library three clones (lanes 3 10 and 12) showed one intact band
greater than 42 kb in size on the agarose gel It is presumed that the Fosmid DNA of the three
samples was not digested because of insufficient amount of restriction enzyme added to the
digestion mixture About half of the samples demonstrated multiple bands suggesting that the
insert DNA might have contained multiple Not I restriction sites
However not every heterologous gene may necessarily express or function in the E
coli host (Gabor et al 2004) The functionality of the gene requires expression of the genes
and proper folding of the protein which may be influenced by environmental conditions (eg
substrate temperature and pH) and genetic factors In this study library clones were screened
in growth conditions at which strain ADS47 performed the best de-epoxidation activity The
expression of genes may be controlled by the promoter suitable ribosome binding site and
transndashacting factors (eg special transcription factors inducers chaperones cofactors proper
modifying enzymes or an appropriate secretion system) (Gabor et al 2004) Those factors
have to be supplied by the cloned DNA fragments or host cells to allow any heterologous
gene to be functional Since DON de-epoxidation genes of strain ADS47 evolved in a distinct
environmental condition the expression and activity of the gene(s) might have required
specific factorsregulators that were absent in E coli Even though the Fosmid based library
screening technique facilitated isolation of many microbial genes clones having de-
epoxidation activity were not identified using such technique in this study
121
46 Acknowledgements
We appreciate the Agriculture and Agri-Food CanadaBinational Agricultural
Research and Development Fund (AAFCBARD) project for providing the financial support
for this work The authors thank Honghui Zhu for analyzing numerous samples by liquid
chromatography ultraviolet mass spectrum method
122
CHAPTER 5
50 GENOME SEQUENCING CHARACTERIZATION AND COMPARATIVE
ANALYSIS OF CITROBACTER FREUNDII STRAIN ADS47
51 Abstract
The genome of a trichothecene mycotoxin detoxifying C freundii strain ADS47
originating from soil was sequenced and analyzed Strain ADS47 contains a circular
chromosome of 4878242 bp that encodes 4610 predicted protein coding sequences (CDSs)
with an average gene size of 893 bp Rapid Annotation Subsystems Technology (RAST)
analysis assigned functions to 2821 (62) of the CDSs while 1781 (38) CDSs were
assigned as uncharacterized or hypothetical proteins Comparative bioinformatics analyses
were done for strain ADS47 C freundii Ballerup 785139 Citrobacter sp 30_2 C youngae
ATCC 29220 C koseri ATCC BAA-895 and C rodentium ICC168 There were a significant
number of common orthologous genes (2821) were found in the six Citrobacter genomes
implying a common evolutionary relationship ADS47 is most closely related to the
opportunistic human pathogen C freundii Ballerup The human and mouse pathogenic
species C rodentium and C koseri are the most distantly related to ADS47 and non-
humananimal pathogens C youngae and Citrobacter sp 30_2 were closely related ADS47
C koseri C youngae and Citrobacter sp 30_2 lacked the clinically important locus for
enterocyte effacement (LEE)-encoded type-III secretion systems which are present in C
rodentium and C freundii Ballerup Strain ADS47 had 270 unique CDSs many of which are
located in the large unique genomic regions potentially indicating horizontal gene transfer
123
Among the unique CDSs nine were reductasesoxidoreductases and a de-acetylase that may
be involved in trichothecene mycotoxin de-epoxidation and de-acetylation reactions
Keywords Citrobacter freundii comparative genomics genome sequencing unique genes
reductases
52 Introduction
According to the Food and Agriculture Organization (FAO) at least 25 of the
worldrsquos food crops are contaminated with mycotoxins causing serious negative impacts on
livestock and international trade (Boutrif and Canet 1998 Desjardins 2006 Binder et al
2007) Trichothecenes (TCs) produced by a variety of toxigenic Fusarium species are
probably the most prevalent food and feed contaminating mycotoxins It is estimated that
infections by the fungus caused more than $1 billion in crop yield losses in USA and Canada
alone (Windels 2000 Schaafsma et al 2002 Yazar and Omurtag 2008) TC mycotoxins can
have adverse affects on human health including organ toxicity mutagenicity carcinogenicity
neurotoxicity modulation of the immune system negative reproductive system affects and
growth retardation (DrsquoMello et al 1999 Pestka et al 2007) A strain of Citrobacter freundii
ADS47 was isolated from soil in southern Ontario that is capable of detoxifying (ie de-
epoxidation and de-acetylation) several TC mycotoxins including deoxynivalenol nivalenol
and T2-toxin (Chapters 2-3)
Citrobacter freundii is a gram negative motile rod shaped facultatively anaerobic
bacterium belonging to the family of Enterobacteriaceae (Wang et al 2000 Whalen et al
2007) The opportunistic human pathogen is responsible for a number of infections including
124
nosocomial infections of respiratory tract urinay tract blood and causes neonatal meningitis
(Badger et al 1999 Wang et al 2000) C freundii exists almost ubiquitously in soil water
wastewater feces food and the human intestine Biodegradations of tannery effluents copper
aromatic amino acids by C freundii strains isolated from water mine tailings and soil
respectively have been previously reported (Kumar et al 1999 Savelieva et al 2004 Sharma
and Fulekar 2009) Because of their biochemical capabilities bacteria have played important
roles in industrial biotechnology including de-epoxidation and de-acetylation activities
(Demain and Adrio 2008 Plank et al 2009 Zhao et al 2010) However the identification of
genes for de-epoxidation activity against trichothecene mycotoxins is limited (Boutigny et al
2008)
The advent of molecular techniques such as next generation sequencing technologies
and the related bioinformatics tools has enabled the discovery of novel genes and the
enzymes they encode in pathogenic and beneficial bacteria (van Lanen and Shen 2006
Ansorge 2009 Zhao 2011) In the present study the genome of mycotoxin-detoxifying strain
C freundii ADS47 was sequenced and subjected to comparative genomic analyses with other
Citrobacter species to identify putative mycotoxin-detoxifying genes
125
53 Materials and methods
531 Sequencing quality control and assemble of ADS47 genome
Genomic DNA of the strain ADS47 was extracted using Gentra Puregene
YeastBacteria Kit (Qiagen Inc Mississauga Ontario Canada) and purity of the DNA was
confirmed by determining OD 260280 at ~190 using a Nanodrop spectrophotometer
The genomic DNA was sent to be sequenced by the Illumina HiSeq 2000 sequencing
analyzer at Beijing Genomics Institute (BGI) Hong Kong
(httpwwwbgisequencecomhomeservicessequencing-servicesde-novo-sequencing) The
sequencing workflow involved short read library preparation cluster generation sequencing
and data analysis Briefly 3-5 ug of bacterial gDNA was fragmented to produce a 500 bp
insert library by preparative electrophoresis through a 08 agarose gel The fragmented
DNAs were blunt-ended by treating with a mixture of enzymes (T4 DNA polymerase klenow
fragment) and subsequently adapters were ligated to the blunt ends Each of the DNA
fragments was amplified to a cluster which was performed in the flow cell by template
hybridization and bridge amplification Millions of clusters were then sequenced by
sequencing-by-synthesis technology leading to base calls The raw data was filtered to remove
low abundant sequences from the assembly according to 15-mer frequencies The low
complexity reads low quality (leQ20) base calls and adapter contaminations (24 bp overlap
between adapter and reads) were eliminated The quality of the sequences was analysed by
GC content and depth correlative analysis (for GC biasness) K-mer analysis was performed
using 15 bp of the short reads for heterologous sequence identification
126
The cleaned short reads (approximately 90 bp) were self-assembled using the Short
Oligonucleotide Analysis Package (SOAP) denovo (httpsoapgenomicsorgcn) (Li et al
2008) The assembly errors were corrected by filling gaps and proofreading single base pairs
using mapping information The assembly produced 441 contigs with an N50 contig size of
29222 bp (the largest contig size 114839 bp) and 80 scaffolds with an N50 scaffold size of
325726 bp (largest scaffold 662452 bp) with an total sequence length of 4878242 bp which
constitutes 33-fold genome coverage The scaffolds were ordered and oriented according to a
reference genome C koseri ATCC BAA-895 the closest completely sequenced species of
the newly sequenced bacterial genome by alignment with the MUMmer program v322
(httpmummersourceforgenet) (Delcher et al 2002) Genome coverage was determined by
mapping the reads to the assembled genome with Bowtie software (Langmead et al 2009) A
preliminary genome annotation was performed to define the origin of replication by
identifying colocalization of multiple genes ie gyrB recF dnaN and dnaA (usually found
near the origin of replication) and comparison with the C koseri reference genome Synteny
analysis between the reference bacterium and assembled scaffolds was performed with the
MUMmer program
532 Gene function annotation and classification
The genome of ADS47 was annotated by Rapid Annotation using Subsystems
Technology v40 (RAST) a server-based genome analysis system (httprastnmpdrorg)
(Aziz et al 2008) A circular genome map was constructed using CGView program
(httpstothardafnsualbertacacgview_server) (Stothard and Wishart 2005)
127
533 Molecular phylogenetic analysis
A robust phylogenetic tree was produced using Composition Vector Tree (CVTree)
web-based program v20 (httpcvtreecbipkueducn) (Qi and Hao 2004) by comparing
whole genome amino acid sequences of strain ADS47 two completely sequenced Citrobacter
genomes [C rodentium ICC168 (NC_013716) and C koseri ATCC BAA-8959 NC_009792)]
and three partially sequenced Citrobacter genomes [C freundii strain Ballerup 785139
(CACD00000000) Citrobacter sp 30-2 (NZ_ACDJ00000000) and C youngae ATCC 29220
(NZ_ABWL00000000)] E coli O157H7 strain EDL933 (NC_002655) was used as an
outgroup species The tree was constructed from the generated pairwise distance matrix data
using Neighbor-Joining Program in PHYLIP v369 package (Saitou and Nei 1987 Felsenstein
1989)
534 Comparative genomic analysis
Genome-wide comparative analyses between C freundii ADS47 C freundii strain
Ballerup 785139 Citrobacter sp 30-2 C youngae ATCC 29220 C Koseri ATCC BAA-895
and C rodentium ICC168 was carried out The nucleotide sequence idendities were
determined by pairwise alignments of the ADS47 genome with the five other Citrobacter
genomes using MUMmer software (Delcher et al 2002) and GC content total CDSs genome
coverage by CDSs and number of stable RNAs were determined by RAST program
Orthologous CDS groups among the total set of 29079 CDSs derived from the six
Citrobacter genomes were determined using OrthoMCL software v202
(httporthomclsourceforgenet) based on all-against-all reciprocal BLASTP alignment of
amino acid sequences (Li et al 2003) Genes unique to each strain were identified as those
128
CDSs not included in any orthologous group of the five other Citrobacter genomes BLASTP
comparisons were visualized with Blast Ring Image Generator (BRIG) software v095
(httpsourceforgenetprojectsbrig) (Alikhan et al 2011)
Protein sequences of the five Citrobacter genomes pooled from the GenBank data
bases were compared with ADS47 A circular map was developed by BRIG software that
showed the unique genomic regionsgenes of ADS47 The unique genes were grouped
according to their functional similarities previously identified by RAST software and
genomic regions possibly acquired by horizontal transfer were identified using RAST
software
129
54 Results
541 Assembly and organization of the ADS47 bacterial genome
Scaffolds derived from Citrobacter freundii strain ADS47 were ordered and oriented
by alignment to Citrobacter koseri ATCC BAA-895 the most closely related genome that has
been completely sequenced The genomes of ADS47 and C koseri displayed close synteny
(Figure 5-1)
Figure 5-1 Co-linearity between C freundii strain ADS47 genome and C koseri Scaffolds
derived from ADS47 genome sequences (Y- axis) were aligned and assembled by comparison
with the closest completely sequenced genome C koseri ATCC BAA-875 (X- axis)
130
542 An overview of the ADS47 genome
General features of the C freundii strain ADS47 are presented in Table 5-1 Some
notable features are that strain ADS47 contains 4881122 bp circular chromosome with an
average GC content of 5202 The genome contains 4610 CDSs which comprise 8784 of
the whole genome Annotation by RAST assigned 2821 (62) CDSs to known biological
functions and remaining 1789 (38) genes were classified to unknown or hypothetical
proteins
The GC content in the coding regions is significantly higher (5318) than in the non-
coding intergenic region (4379) which covers 1215 (592781 bp) of the genome The
intergenic region is comprised of 018 repeat sequences (total 8999 bp) 5 (009)
transposons (4806 bp) 025 stable RNA (12091 bp) and the remaining ~115 is
classified as regulatory and other functions The ADS47 genome contains 62 tRNAs 3
rRNAs and 34 copies of sRNAs The average lengths of the tRNAs rRNAs and sRNAs are
785 885 and 13441 bp respectively There are also 42 phageprophage-related genes
identified in the genome
Figure 5-2 shows the distribution of the 26 categorized CDSs rRNA tRNA GC
content and GC skew It appears that ADS47 genome contains equal quantity of genes in
leading and lagging strands and showed occasional clustering (likely operons) of genes in the
two strands The genome contains three rRNAs (two large subunit and a small subunit) and 62
tRNAs which are distributed across the genome A consistent GC content was observed in
the ADS47 genome however 7-10 low GC content (high black color peaks) genomic regions
were found at locations origin of replication 600 2000 3000 3300 3550 3850 4300 and
4800 kbp This is indicative of possible horizontally transferred genes or insertions in those
131
genomic locations GC skew denotes the excess of C over G in certain genomic locations
Positive GC skew (+) (green color) indicates the skew value in the leading strand while
negative skew (-) (purple color) in the lagging strand As in many bacterial genomes the
ADS47 genome map shows the separation of positive and negative GC skew at the origin of
replication (base pair position 1) The GC skew in the leading strand is highly skewed (high
green color peak) indicating that possible horizontal gene transfer occurred into the ADS47
genome
The CDSs ranged from ~100 bp to ~2000 bp and have an average size of 893 bp
(Figure 5-3) The highest numbers of CDS (770) were 600-800 bp long A considerable
number of genes (220) were gt2000 bp in size Some of the biological functions for the
smallest genes (100-200 bp) are sulfur carrier protein methionine sulfoxide reductase
ribonuclease P protein component while the largest genes (gt2000 bp) are ferrous iron
transport protein B translation elongation factor G pyruvate flavodoxin oxidoreductase and
tetrathionate reductase
132
Table 5-1 General features of the C freundii strain ADS47 genome
Feature Chromosome
Genome size 4881122 bp
Overall GC content 5202
Number of genes 4610
Genes with assigned function (number and ) 2821 (62 )
Genes without assigned function (number and ) 1789 (38 )
Total length of protein coding sequences 4285461 bp
Genome coverage by coding sequences () 8784
Average length of coding sequences 893 bp
GC content in the coding region 5318
GC content in the non-coding region 4379
Length and percentage of intergenic region 592781 bp and 1215
Total repeat sequences and genome coverage 8999 bp and 018
Total transposons sequences and genome coverage 4806 and 009
Number average length and genome coverage of tRNA 62 copies 785 bp and 0099
Number average length and genome coverage of rRNA 3 copies 885 bp and 0067
Number average length and genome coverage of sRNA 34 copies 13441 bp and 0093
133
134
Figure 5-2 Circular genome map of C freundii strain ADS47 Map was created with the
CGView software From outside to inside the first circle indicates the genomic position in kb
Different functional categories of predicted coding sequences (CDS) are distinguished by
colours with clockwise transcribed CDSs in the second circle and anticlockwise transcribed
CDSs in the third circle Bars on the fourth and fifth circles indicate the rRNAs and tRNAs
respectively Sixth and seventh circles show the coverage and GC content respectively The
innermost circle demonstrates the GC skew + (green) and GC skew- (purple)
135
Figure 5-3 Size distribution of the predicted genes identified in C freundii strain ADS47
The X axis identifies the gene size categories (bp) and Y axis the abundance of genes
136
543 Annotation and functional classification of ADS47 genome
RAST analysis identified predicted CDSs in the genome of Citrobacter strain ADS47
assigned functions and grouped the annotated genes into 26 functional subsystem categories
out of 27 No CDS was found belonging to the photosynthesis category A graphical
presentation of the distribution of the CDSs into each of the 26 categories is given in
Figure 5-4
A large portion (38) of the CDSs belong to hypothetical or unknown functions
Among the 62 known or predicted proteins the highest numbers (835) belong to the
carbohydrate category followed by amino acids and derivatives (468) and
cofactorsvitaminsprosthetic groups (324) A noticeable number of stress response (236) and
respiration (198) categories genes were identified in the bacterial genome The ADS47
genome encoded by 171 membrane transport genes 113 virulencedefense genes and 126
genes involved in regulationcell signaling categories Two hundred and one CDSs were
grouped into the miscellaneous category whose functions are different than the indicated
RAST categories The RAST annotations further divided the categorized CDSs into
subcategories based on similar functional roles which is discussed in section 545
137
Figure 5-4 Functional classification of the predicted coding sequences (CDS) of C freundii
strain ADS47 The annotation was performed by Rapid Annotation using Subsystems
Technology (RAST) program
138
544 Molecular phylogenetic relationships
Genome-wide phylogenetic analysis based on comparisons of whole genome amino
acid sequences of two completely sequenced Citrobacter genomes (C koseri and C
rodentium) and three partially sequenced Citrobacter genome sequences (C freundii
Citrobacter sp and C youngae) illustrates the relationships of ADS47 to five Citrobacter
species (Figure 5-5)
C youngae ATCC 29220 originating from human feces and two pathogenic species
(C koseri ATCC BAA-895 and C rodentium strain ICC168) are progressively more distantly
related to ADS47 The present analysis is consistent with the 16S-rRNA sequence based
phylogenetic analysis (Chapter 3) which showed that the closest bacterial strain to ADS47 is
C freundii (Figure 3-2) This confirmed the identification of strain ADS47 as the gamma
proteobacterial species C freundii made in Chapter 3
139
Figure 5-5 Phylogeny of the trichothecene detoxifying C freundii strain ADS47 The
dendrogram was constructed by comparing the ADS47 genome (amino acid sequences) with
five Citrobacter genomes and a E coli strain (out group species) using the CVTree software
The calculated pairwise distances matrix by Protdist program was transformed into a tree by
neighbor-joining program The dissimilarity tree was drawn to scale with branch lengths
(scale bar 002 indicated the amino acid substitutions among the genomes) The C koseri C
rodentium C freundii Citrobacter sp C youngae and E coli protein sequences were
uploaded from GenBank (Benson et al 2009)
140
545 Comparative genomics
Genome-wide comparative analyses between the genome of strain ADS47 and the five
previously sequenced Citrobacter genomes showed that 872 of the total nucleotides
between ADS47 and C freundii Ballerup were identical The values for the comparisons
between C freundii strain ADS47 were 8212 for Citrobacter sp 30-2 7637 for C
youngae 5981 for C koseri and 4213 for C rodentium (Table 5-2)
The two human and mouse pathogenic species namely C rodentium and C koseri
and the bacteria originally isolated from human feces (C youngae) have higher GC contents
(546 and 532 respectively) compared to C freundii strain ADS47 C freundii strain
Ballerup and Citrobacter sp 30_2 which have slightly lower GC contents (~52) The two
pathogenic species contain 22 copies of rRNA which is significantly higher than the 9 found
in C freundii strain Ballerup 6 in Citrobacter sp 30_2 and 3 in C freundii strain ADS47 but
less than the 25 copies in C youngae The genome of C freundii strain ADS47 C freundii
strain Ballerup C youngae and Citrobacter sp 30-2 had a similar percentage of the genome
comprising coding sequences (865-88) C koseri and C rodentium genomes contained the
highest (90) and lowest (84) genome coverage by coding sequences respectively
141
Table 5-2 Genome comparisons between strain ADS47 and five Citrobacter species The
base nucleotide sequence identities were performed using MUMmer DNAdff script program
(Delcher et al 2002) The features (GC content total CDS percentage CDS and number of
ribosomal RNA) were determined using RAST program (Aziz et al 2008)
Genome feature Bacterial species
C
freundii
ADS47
C freundii
Ballerup
Citrobacter
sp 30_2
C
youngae
C koseri C
rodentium
Genome length
(bp)
4878242
4904641
5125150
5154159
4720462
5346659
Base similarity to
ADS47 ()
NA 8715 8212 7637 5981 4213
GC () 5202 5218 516 5265 530 546
Number of coding
sequences
4610 4685 4728 5278 4978 4800
Genome coverage
with coding
sequences ()
880 865 866 872 900 840
rRNA copies 3 9 6 25 22 22
Total stable RNAs 99 81 78 105 120 142
142
The number of orthologs shared between ADS47 and other Citrobacter species is
illustrated in Figure 5-6 ADS47 shares the smallest number of orthologous CDSs with C
rodentium and C koseri 3250 and 3341 respectively The highest number of shared CDS
were found with C freundii Ballerup (3706) followed by C youngae (3693) and Citrobacter
sp 30_2 (3676) A total of 2821 CDSs were found to be common to all of the six Citrobacter
genomes analysed (data not shown) Nonetheless significant diversity exists between C
freundii ADS47 and the other species as a considerable number of unshared genes were
identified from the comparisons with C freundii Ballerup [979 (20)] Citrobacter sp 30_2
[1064 (23)] C youngae [1654 (31)] C koseri [1693 (34)] and C rodentium [1634
(34)]
143
Figure 5-6 Comparisons of coding sequences between strain C freundii ADS47 and five
Citrobacter species Coding sequences of ADS47 were compared with C freundii Ballerup
Citrobacter sp 30_2 C youngae C koseri and C rodentium genomes based on all-versus-all
reciprocal FASTA comparison of at least 30 amino acid identity and gt70 coverage of the
total length using OrthoMCL program A Venn diagram demonstrates the shared and
unshared (indicated in the brackets) coding sequences
144
Table 5-3 shows differences in the 26 RAST categorized genes between the six
Citrobacter genomes Even though a low abundance of virulencediseasedefense category
genes were found in the animalhuman pathogenic bacteria of C rodentium (85) and C koseri
(92) significantly higher mycobacterium virulence genes (13) were identified in this two
species The genomes of the two closely related strains C freundii ADS47 and C freundii
Ballerup lack this type of virulence gene Instead elevated numbers of genes for resistance to
antibiotic and toxic compounds were found in C freundii ADS47 (94) C freundii Ballerup
(88) and Citrobacter sp (110) compared to other three species Copper homeostasis and
cobalt-zinc-cadmium resistance genes were identified in strain C freundii ADS47 (13 and 21
respectively) and closely related C freundii Ballerup (12 and 22) and Citobacter sp (18 and
22) Significantly lower numbers of these genes were present in C rodentium (6 and 4) and C
koseri (7 and 8) C rodentium and C koseri lack mercuric reductase mercuric resistance and
arsenic resistance genes perhaps because being animalhuman pathogens but these genes
were found in the remaining four genomes The C koseri genome is richer in betandashlactamase
genes (5) compared to the remaining genomes which encodes 1-3 genes
145
Table 5-3 Functional categorized gene abundances among the genomes of C freundii
ADS47 C freundii Ballerup Citrobacter sp 30_2 C youngae C koseri and C rodentium
The genomes were classified into different functional categories with the RAST program
which classified the total CDSs
RAST subsystem category Citrobacter species
C
freundii
ADS47
C
freundii
Ballerup
C
sp
C
youngae
C
koseri
C
rodentium
Cofactorsvitaminsprosthetic
groupspigments
323
317
310
311
325
304
Cell wall and capsule 279 272 248 262 247 262
Virulence disease and defense 113 107 134 100 92 85
Potassium metabolism 50 53 33 32 30 28
Miscellaneous 210 228 200 185 245 185
Phageprophagetransposable
element plasmid
24 75 25 4 6 49
Membrane transport 170 156 171 195 171 172
Iron acquisition and metabolism 80 81 86 62 113 27
RNA metabolism 220 222 217 215 238 212
Nucleosides and nucleotides 119 112 110 111 120 114
Protein metabolism 249 280 269 298 287 254
Cell division and cell cycle 30 29 32 29 38 29
Motility and chemotaxis 133 124 90 147 143 133
Regulation and cell signaling 126 130 124 135 132 123
Secondary metabolism 21 22 20 21 0 0
DNA metabolism 145 167 165 155 147 152
Fatty acids lipids and isoprenoids 121 149 122 121 134 133
Nitrogen metabolism 50 52 53 49 56 58
Dormancy and sporulation 3 3 3 4 3 3
146
Table 5-3 (continued)
RAST subsystem category
Citrobacter species
C
freundii
ADS47
C
freundii
Ballerup
C
sp
C
youngae
C
koseri
C
rodentium
Respiration 198 184 180 191 166 162
Stress response 236 235 170 177 179 174
Metabolism of aromatic
compounds
20 20 28 25 17 48
Amino acids and derivatives 482 476 450 466 428 416
Sulfur metabolism 48 50 52 57 56 56
Phosphorus metabolism 50 49 43 42 43 28
Carbohydrates 837 803 708 731 733 737
147
Strains C freundii ASD47 and C freundii Ballerup have much more potassium
metabolism category genes (50 and 53 respectively) than the other bacterial species which
code for 28-33 proteins Larger numbers of genes associated with the glutathione regulated
potassium efflux system were also identified in C freundii ASD47 with 23 and C freundii
Ballerup with 26 compared to the remaining four species that encode only five genes All of
the six genomes contained phageprophage-related genes with the maximum identified in C
freundii Ballerup (75) followed by C rodentium (49) Citrobacter sp (25) and C freundii
ADS47 (24) Lower numbers of phageprophage-related genes were found in C koseri (6) and
C youngae (4)
The six Citrobacter genomes showed remarkable variability in their secretion systems
The type-III secretion operon genes are only found in C rodentium and C freundii Ballerup
Type-II secretion genes are predominant in C rodentium (12) followed by ADS47 (9)
Type-IV secretion system is uncommon in Citrobacter species since it was only
identified in Citrobacter sp where it is represented by 19 genes Variable numbers of type-VI
secretion system genes are present in the five genomes except C koseri which lacks such
genes C youngae has the highest number (43) of type-VI secretion proteins The other strains
have a range of type-VI secretion proteins in particular 27 were identified in C freundii
Ballerup 15 in C freundii ADS47 and 8 in Citrobacter sp Type-VII (chaperoneusher)
secretion system genes were found in all of the six Citrobacter species with the largest (42)
and smallest (13) numbers of these genes occurring in C koseri and C freundii Ballerup
respectively
Although a relatively similar number of ABC transporters (37-42) were identified in
the six genomes C freundii ADS47 contain the largest number of tripartite ATP-independent
148
periplasmic (TRAP) transporter genes (7) By contrast the closely related C freundii
Ballerup lacks such transporter system An identical number of iron acquisition and
metabolism category genes were found in C freundii ADS47 (80) and C freundii Ballerup
(81) while the two pathogenic species C rodentium and C koseri code for the lowest (27)
and highest (113) numbers of such genes C koseri is rich in siderophore related genes (64)
but the remaining five genomes have smaller numbers of siderophore-associated genes (20-
22)
Variable numbers of motility and chemotaxis related genes were found in the
Citrobacter genomes with the largest number occurring in C youngae (147) followed by C
koseri (143) C freundii ADS47 (133) C rodentium (133) C freundii Ballerup (124) and
Citrobacter sp (90) The two pathogenic species (C rodentium and C koseri) encode a
remarkably low number of quorum sensing genes (2) In contrast C youngae codes for the
highest number of quorum sensing genes (19) A common and distinctive feature of the two
pathogenic species is the lack of genes required for secondary metabolism In contrast the
non-pathogenic species contain a common set of genes for degrading a range of
phenylpropanoids and cinnamic acid (20-22)
The lowest number of genes related to phospholipidsfatty acids biosynthesis and
metabolism were found in C freundii ADS47 (66) compared to C freundii Balllerup (94) and
C koseri (98) The smallest numbers of nitratenitrite ammonification genes were found in
ADS47 (31) and its closest genome C freundii Ballerup (32) In contrast the two pathogens
encode the highest number of such proteins (39-40)
C rodentium and C koseri code for significantly low numbers of respiratory category
genes (162-165) while C freundii ADS47 contains the highest number of such genes (198)
149
Further analysis of the subcategory genes determined that the two gut species (C youngae and
Citrobacter sp 30_2) contain the largest numbers of genes encoding anaerobic reductases
(22) and identical numbers of formate dehydrogenases (17) The largest numbers of formate
dehydrogenase genes were identified in C freundii Ballerup (32) and C freundii ADS47 (30)
but the two pathogenic species differed from the other species by encoding the lowest number
(13) of c-type cytochrome biogenesis genes
A higher number of aromatic compound metabolism category genes were found in the
mouse pathogen C rodentium (48) compared to the human pathogen (C koseri) which
contain 17 genes A significantly higher number of genes involved in central aromatic
intermediate metabolism (26) were found in C rodentium compared to the mouse pathogen
(C koseri) and other genomes which only contain 6-8 such genes The two C freundii strains
ADS47 and Ballerup lacked the anaerobic degradation of aromatic compound
(hydroxyaromatic decarboxylase family) genes
In spite of the abundance of stress response category genes strain ADS47 has only a
small number of desiccation stress genes (5) compared to the other species which ranged
from 9 to10 However ADS47 and Ballerup code for higher number of genes related to
oxidative stress (94 and 91 respectively) and cold shock (8 and 7 respectively) than the other
species
The smallest numbers of amino acids and its derivatives category genes were found in
the two pathogens C rodentium and C koseri (416 and 428 respectively) while the largest
number of these genes were identified in strain ADS47 (482) and C freundii Ballerup (476)
Despite the similar number of sulfur metabolism category genes determined across the six
species only ADS47 lacks taurine utilization genes Strains ADS47 and Ballerup were found
150
to be rich in the phosphorous metabolism gene category (49-50) while C rodentium contains
a significantly decreased number of these category genes (28)
The bioinformatics analyses found the largest number of carbohydrate category genes
in ADS47 (837) followed by Ballerup (803) The gut bacteria (Citrobacter sp 30_2) codes for
the least number of genes assigned to the carbohydrate category (708) while C koseri C
rodentium and C youngae contained 731-737 in this category genes The analyses also
determined that there are relatively small numbers of TCA and Entner Doudoroff pathway
genes in C rodentium and C youngae compared to the other four species and relatively large
numbers of fermentation-related genes in ADS47 (81) and C freundii (85) compared to the
other four species
A comparison of the pathogenicity operons [locus of enterocyte effacement (LEE)] of
C rodentium with other five Citrobacter genomes revealed the absence of this virulence
determinant gene region in the other four Citrobacter species including ADS47 (Figure 5-7)
The analysis showed the presence of LEE operon genes in C freundii strain Ballerup as no
gap was observed in that genomic location while C freundii ADS47 C koseri C youngae
and Citrobacter sp 30_2 lacked that loci (7a) Further ananlysis showed alignments between
C rodentium and E coli LEE operon genes (Figure 7b) In contrast no alignment of such
genes was observed in ADS47 genome
151
(a)
152
Figure 5-7 (a) Blast Ring Image Generator (BRIG) output based of Citrobacter rodentium
(NC_013716) compared against ADS47 C freundii C koseri C youngae E coli and
Citrobacter sp genomes The targeted comparative genome analysis was performed based on
locus of enterocyte effacement (LEE) operons of C rodentium (Petty et al 2011) The
innermost rings show GC content (black) and GC skew (purple and green) The remaining
rings show BLAST comparisons of the other Citrobacter and E coli genomes against C
rodentium The region shown in red is LEE operon genes (b) The absence of LEE operons in
the ADS47 genome comparing to C rodentium and E coli Analysis was performed and
visualized by Artemis Comparison Tool software v132
(httpwwwsangeracukresourcessoftwareact) (Tatusov et al 2000 Craver et al 2005)
LEE operons
(b)
153
546 Analysis of the unique genes in C freundii ADS47
Two hundred and seventy genes unique to ADS47 were identified from a comparison
of its genome to the other five Citrobacter species In general the unique genes are
distributed throughout the ADS47 genome but some clusters of unique genes occurred
including eight unique genomic regions (Figure 5-8) Six large unique regions are located at
767204-775090 (~8 kb) 1025572-1042510 (17 kb) 2798289-2807217 (~9 kb)
3045220-3055912 (~11 kb) 3930366-3943075 (12 kb) and 4022195-4032444 (10 kb)
Out of 270 unique genes 147 (54 ) are hypothetical proteins The remaining 123
(46) genes are grouped into reductasesoxidoreductases (9 genes) de-acetylase (1)
hydrolases (2) transferases (4) sulfatasesarylsulfatases (3) bacteriocins (3) type VI
secretion proteins (2) kinases (4) transporters (11) membraneexported proteins (8)
regulators (12) integraserecombinases (5) phagesprophagestransposaes related genes (27)
and miscellaneous (32) that includes DNA helicases ligases fimbrial proteins and
chromosome partitioning genes (Table 5-4)
The unique regions encode various functional genes that may cause trichothecene de-
epoxidation or de-acetylation reactions The 8 kb unique regions contains six genes encoding
glycosyltransferases and two copies of oxidoreductase-like coding sequences (CMD_0711
and CMD_0712) that showed 39 identity to the Marinobacter aquaeolei F420
hydrogenasedehydrogenase beta subunit The largest (17 kb) unique genomic region codes
for 16 genes including dehydrogenase hydrolase bacteriocin (pyocin) integrase and a short
chain dehydrogenasereductase-like (SDR) (CMD_0979) gene (Figure 5-9a)
154
The SDR like gene is 49 amino acid similarity to an oxidoreductase of the
phytopathogenic Dickeya dadantii and the enzyme contains a domain with 77 homology to
the Dickeya zeae NAD(P) binding site The 9 kb unique genomic section contains 8 genes
including a NaCl symporter a dehydratase (CMD_2679) with 86 homology to
Enterobacteriaceae bacterium dihydroxy-acid dehydratase and a putative oxidoreductase
(CMD_2683) having 37 similarity with oxopropionate reductase of Bacillus sp The unique
region also encodes a short chain dehydrogenasereductase (SDR) (CMD_2684) that shows
85 identity to Enterobacteriaceae bacterium (Figure 5-9b) A unique thiol-disulfide and
thioredoxin like reductase (CMD_3721) was identified in a 10 kb unique region that contains
13 genes (Figure 5-9c) This gene is 59 identical to Oligotropha carboxidovorans thiol-
disulfide isomerase and thioredoxin In addition several reductasesoxidoreductase-like
unique genes were identified in the ADS47 genome including an oxidoreductase
(CMD_1024) with 27 identity to Canadidatus nitrospira NADH-quinone oxidoreductase
membrane L Another oxidoreductase-like gene identified in a different location of the
ADS47 genome (CMD_1393) shows 87 homology to Klebsiella pneumoniae
deoxygluconate dehydrogenase contains a SDR active site A putative nitrate reductase gene
(CMD_3184) has a low level of identity (27) with an uncultured bacterial respiratory nitrate
reductase The ~11 kb unique region encodes sulfatasesarylsulfatses
(CMD_2898CMD_2905) and related genes Furthermore ADS47 contains a unique
glycosylphosphatidylinositol (GPI) anchor biosynthesis operon comprised of four genes
including a N-acetylglucosaminyl-phosphatidylinositol de-acetylase (CMD_2431) which
showed 78 similarity to E coli de-acetylase (Figure 5-9e)
155
Other unique genespredicted genes in the ADS47 genome include
polysaccharidephenazine biosynthesis genes (CMD_0710CMD_1406) heavy metal kinase
(CMD_0989) two component heavy metal regulator (CMD_0990) nitrate reductase regulator
(CMD_3477) putative potassiumzinc transporters (CMD_0973CMD_4600) sodium
symporter (CMD_2678) (tripartite ATP-independent periplasmic transporters) TRAP
transporters (CMD_3693 CMD_3694 CMD_3695 CMD_4337) glutamineribose ABC
transporters (CMD_3176CMD_3544) toxin transcription regulator (CMD_3464) fimbrial
proteinchaperone (CMD_1695 CMD_2434 CMD_3277 CMD_3278) and ParB
chromosomeplasmid partitioning proteins (CMD_4033CMD_4034)
Majority of the unique genes and their distributions are shown in comparative circular
genome maps (Figure 5-8)
156
Table 5-4 Unique genes of C freundii strain ADS47 Genes were categorized according to
the functional similarity The unique genes were determined by comparing ADS47 with five
sequenced Citrobacter species using OrthoMCL software
Unique gene classes Number
Total unique genes
270
Reductases oxidoreductases 9
De-acetylase 1
Hydrolase 2
Transferase 4
Sulfatase 3
Bacteriocin 2
Type VI secretion 2
Kinase 4
Transporter 9
Membraneexported protein 8
Regulator 12
Integraserecombinase 5
Phageprophagetransposae 27
Miscellaneous 34
Hypothetical protein 147
157
158
Figure 5-8 Locations of the unique genes of C freundii strain ADS47 The unique genes
were determined by comparing the total CDS of strain C freundii ADS47 with C freundii
Ballerup Citrobacter sp C youngae C koseri and C rodentium CDSs by OrthoMCL
program The map was developed by Blast Ring Generator (BRIG) software First second
and third circles from inside to out side show the genome size GC and GC skew of strain
ADS47 Forward (Fwd) and reversed (Rev) transcribed CDSs of ADS47 are shown in green
(forth circle) and red (fifth circle) respectively The next five rings represent the circular
genome maps of five Citrobacter species distinguished by different ring colours (C
rodentium as sky blue C koseri as dark blue C youngae as light blue C freundii Ballerup as
yellow and Citrobacter sp as green) The degree of orthologous gene identity between
ADS47 and other five genomes are shown by color intensity (light color= 75 gene identity
ash color= 50 gene identity) Gaps across five genome rings indicate the absence of
genesregions compared to ADS47 Unique genes are shown surrounding the map rings and
the identities of some of the genes are given Arrows indicate putative
reductasesoxidoreductases and de-acetylase
159
(a)
(b)
160
(c)
(d)
161
Figure 5-9 Unique genomic regions and associated genes in C freundii ADS47 compared to
five Citrobacter species RAST server based analysis compared the unique genesregions of
ADS47 with the publicly available databases The upper panels demonstrate the large unique
regions defined by yellow boxes and genes located in the genomic regions are indicated by
arrows The lower panels are closely related genesoperons found in distantly related species
The unique regions that contain potential trichothecene de-epoxidation genes are shown in (a)
short chain dehydrogenasereductase (SDR) and dehydrogenase (b) putative oxidoreductase
dehydratase and SDR (c) thiol-disulfide isomerase and thioredoxin oxidoreductase and (d)
unique glycosylphosphatidylinositol (GPI) biosynthesis operon having N-acetylglucosaminyl-
phosphatidylinositol de-acetylase gene
162
55 Discussion
The de novo sequenced ADS47 genome was oriented and assembled using a
completely sequenced C koseri genome as reference strain The synteny analysis between
ADS47 and C koseri showed very similar overall organization of the two genomes The high
preservation of genes among five Citrobacter species and ADS47 were emphasized by the
close alignments observed by Blast Ring Image Generator (BRIG) outputs obtained from
these genomes
Genome-wide phylogenetic analyses based on a large number of common CDSs
(2821) observed within the six Citrobacter genomes confirmed the species identification
based on 16S-rDNA sequencing of the trichothecene mycotoxin-detoxifying C freundii strain
ADS47 The base sequence idendity was 8715 between these two genomes They shared
~80 CDS and had a similar GC content (52) Citrobacter are Gram-negative rod shaped
and facultatively anaerobic bacteria belonging to the family of Enterobacteriaceae (Wang et
al 2000 Whalen et al 2007) Strains ADS47 and Ballerup of C freundii are both aerobic
gram-negative bacilli that have numerous flagella and inhabit in soil (Cong et al 2011 Islam
et al 2012) C freundii is considered to have a positive environmental role by reducing nitrate
to nitrite (Rehr and Klemme 1989) which is an important step in the nitrogen cycle
C freundii C rodentium and C koseri are animalhuman pathogenic species C
rodentium is a natural mice pathogen causing hemorrhagic colitis and hemolytic uremic
syndrome (Gareau et al 2010) C koseri causes human disease (Badger et al 1999 Kariholu
et al 2009) particularly neonatal meningitis and C freundii is an opportunistic human
pathogen responsible for a various types of infections like respiratory tract urinary tract and
blood (Whalen et al 2007) C freundii is very common representing approximately 29 of
163
all opportunistic human infections (Hodges et al 1978) Outside of animals they can be found
in soil water sewages and different organs of diseased animals (Wang et al 2000) These
pathogens code for large numbers of mycobacterium virulence operonsgenes which were
absent in the three other genomes However they have fewer genes involved in environmental
stress such as copper homeostasis genes and no mercuricarsenic resistance and secondary
metabolism category genes unlike the other Citrobacter genomes Nonetheless considerable
variability exists between the two pathogens correlates with C rodentium causing disease in
mice C koseri causing meningistis in human and C freundii causing infections in different
human organsThere were attaching and effacing (AE) lesion forming (LEE) operons in C
rodentium and C freundii but not in C koseri Those genes have shown to be acquired by
lateral gene transfer playing major roles in virulence of enteropathogenic and
enterohemorrhagic E coli and C rodentium (Mundy et al 2005 Petty et al 2010 Petty et al
2011 The ~356 kb pathogenic LEE gene island is comprised of five operons (LEE1 to
LEE5) that code for ~40 virulence genes that include type-III secretion genes and associated
effectors (Berdichevsky et al 2005) In addition an elevated number of type-II and -VI
secretion proteins identified in C rodentium which may act as additional virulence factors
(Petty et al 2010) Because it lacks type-III secretion apparatus genes it is likely that C
koseri uses a different strategy to cause human neonatal meningitis (Badger et al 1999 Ong
et al 2010)
To adapt to different environmental niches or hosts bacteria like Citrobacter species
frequently gainlose genes increase repeat sequences and recombine genomic regions using
different mechanisms (Parkhill et al 2001 Petty et al 2011) The existence of large numbers
of phageprophagetransposae related genes in the Citrobacter genomes particularly in C
164
freundii Ballerup suggests that their genomes are unstable For example C freundii Ballerup
and C koseri might have acquired LEE operons by horizontal transfer via phage mediated
transpositions Interestingly although ADS47 is closely related to C freundii Ballerup it
lacks the pathogen-related LEE gene island and mycobacterium virulence operons required
for invasion and intracellular survival in macrophages (Gao et al 2006) Instead ADS47 has a
larger number and variety of stress-responsive genes such as efflux genes and bacteriocins
(pyocin hemolysin) genes that provide protection against heavy metals and antibiotics and
increase their competitiveness against other bacteria These findings suggest that the strain
ADS47 is not an opportunistic animal pathogen but has evolved to be highly competitive in
the soil micro-environment (Arnold 1998 Bengoechea and Skurnik 2000) Perhaps ADS47
evolved trichothecene degrading genes for adaptation to soils containing trichothecene
mycotoxins contaminated soils (Gadd and Griffiths 1978 Sharma and Fulekar 2009 Han et
al 2011) Conversely the absence or reduced number of stress-responsive genes in C
rodentium and C koseri may imply that those two pathogenic species are not exposed to such
stressful and xenobiotic-contaminated environments as ADS47
This interpretation is supported by the observation that the two of the three
animalhuman pathogenic species contained high of rRNA copy number but lower numbers
were found in ADS47 C freundii Ballerup and Citrobacter sp 30_2 Bacteria maintain large
number of rRNA for higher production of ribosome during high growth rates (Stevenson et al
1997 Klappenbach et al 2000) such as might occur during the establishment of an infection
However a large copy number of rRNAs may not be metabolically favourable for bacteria
such as ADS47 that evolved in a highly competitive environment as that costs higher cellular
energy (Hsu et al 1994)
165
C freundii Ballerup seems to be an exception in that it is similar to the other animal
pathogens for containing virulent factors (ie LEE-operon genes) but more like the
environmental species for encoding high number genes for stress defence heavy metal and
toxic compounds resistances
The comparison of the genes in C freundii ADS47 with the unique ability to detoxify
trichothecenes with genes in other Citrobacter species allowed approximately 270 unique
genes to be identified in ADS47 Some of those genes may be involved in trichothecene de-
epoxidation or de-acetylation Currently the pathways and microbial enzymes leading to
trichothecene de-epoxidation and de-acetylation are unknown but based on the required
reaction ie the elimination of an oxygen atom from epoxide moiety it is likely that epoxide
reductases or epoxide oxidoreductases are responsible for such biodegradation activity (He et
al 1992)
From database searches three enzymes having epoxide reductase activity were found
These are a vitamin K epoxide reductase (VKOR) a monooxygenase PaaABCE and pyridine
nucleotide-disulfide oxidoreductase (PNDO) which reduce vitamin K epoxide phenylacetyl-
CoA and epoxyalkanes respectively (Swaving et al 1996 Li et al 2004 Teufel et al 2012)
In mammals VKOR plays important roles in vitamin K cycle by reducing vitamin K epoxide
to vitamin K and its homologues that are found in plants bacteria and archaea (Goodstadt
2004 Li et al 2010) The microbial multi-component PaaABCE monooxygenase catalyzes
degradation of variety of aromatic compounds or environmental contaminants via epoxidation
of phenylacetic acid (Paa) and NADPH-dependant de-epoxidation of epoxy-Paa (Luengo et
al 2001 Teufel et al 2012) PNDO is a unique type of NADP-dependent oxidoreductase
identified in Xanthobacter strain Py2 that was obtained by propane enrichment procedure
166
(van Ginkel and de Bont 1986) However no orthologs of these genes were found in ADS47
genome
Nine ADS47-unique reductaseoxidoreductase-like genes are potential candidates for
the trichothecene de-epoxidation enzyme Genes belong to short chain
dehydrogenasereductase (SDR) family are structurally and functionally diverse and cause
different enzymatic activities including xenobiotic degradation (Kavanagh et al 2008) This is
a promising candidate for the trichothecene de-epoxidation gene Genes belong to dehydratase
family catalyze reduction reactions by cleaving carbon-oxygen bonds of organic compounds
that result in removal of oxygen in various biosynthetic pathways (Flint et al 1993) This
mechanism may be relevant to microbial trichothecene de-epoxidation reaction since the latter
involves removal of the oxygen atom forming the epoxide in trichothecene mycotoxins
(Young et al 2007)
Another ADS47-unique gene was for a nitrate reductase that may be a candidate de-
epoxidation gene Under stress conditions such an enzyme causes reductive removal of an
oxygen molecule (Morozkina and Zvyagilskaya 2007)
A fourth type of ADS47-unique gene is for a thiol-disulfide isomerasethioredoxin like
(Trx) oxidoreductase The thiol-disulfide isomerase and thioredoxin oxidoreductases contain a
thioredoxin like domain with a CXXC catalytic motif which catalyzes various biological
processes that allow the organism to adapt to environmental stress and develop competence to
sporulate (Scharf et al 1998 Petersohn et al 2001 Smits et al 2005) The membrane-
anchored Trxndashlike domain initiates the vitamin K de-epoxidation reaction by reducing the
active motif (CXXC) of VKOR (Schulman et al 2010)
167
The present study did not find any homologues of VKOR in ADS47 genome but we
identified two copies of structurally similar quinone reductase protein disulfide bond
formation (DsbB) gene (CMD_1044 and CMD_2641) Both VKOR and DsbB contain four
transmembrane segments an active site CXXC motif and cofactor quinone and use a
common electron transfer pathway for reducing substrates (Li et al 2010 Malojcic et al
2008) As VKOR bacterial DsbB requires a Trx-like partner (eg DsbA) for activity Further
experiments are required to examine if DON de-epoxidation occurs by the catalytic activity of
DsbB coupled with a unique thiol-disulfide isomerase and thioredoxin oxidoreductase present
in ADS47 genome
The analysis also identified an ADS47-unique glycosylphosphatidylinositol (GPI)
biosynthetic operon that is comprised of four genes including an N-
acetylglucosaminylphosphatidylinositol de-acetylase The GPI membrane anchor protein
synthesis is initiated by transferring N-acetylglucosamine from uridine diphosphate N-
acetylglucosamine to phosphatidylinositol by N-acetylglucosaminyltransferase an
endoplasmic reticulum (ER) membrane-bound protein complex (Hong and Kinoshita 2009)
In the second step of the pathway N-acetylglucosamine-phosphatidylinositol is de-acetylated
to produce glucosaminyl-phosphatidylinositol by the catalytic activity of a zinc
metalloenzyme N-acetylglucosaminylphosphatidylinositol de-acetylase which is localized on
the cytoplasmic side of the ER (Nakamura et al 1997 Hong and Kinoshita 2009) It is
possible that the identified unique N-acetylglucosaminylphosphatidylinositol de-acetylase
gene of ADS47 may be involved in trichothecene de-acetylation activity
168
Further analysis is needed to determine which of these candidate gene or genes may be
involved in tricthothecene mycotoxin de-epoxidation and de-acetylation activities
Identification of gene responsible for the trichothecene detoxification activities of C freundii
ADS47 opens many potential applications for its use in agri-based industries to contribute to
cropanimal production and food safety (Karlovsky 2011)
56 Acknowledgement
We gratefully acknowledge Agriculture and Agri-Food CanadaBinational
Agricultural Research and Development Fund (AAFCBARD) project for providing the
financial supports of the work
169
Supplemental Table S5-1 List of unique genes of strain ADS47 gene IDs and their
functional roles The unique genes were determined by comparing the ADS47 genome with
five sequenced Citrobacter species (C freundii Citrobacter sp C youngae C koseri and C
rodentium) using OrthoMCL (Li et al 2003)
Unique gene
classes
Number Gene ID and functions
Total unique
genes
270 123 known functions 147 hypothetical proteins
Reductases
oxidoreductases
9 CMD_0711 coenzyme F420 hydrogenasedehydrogenase beta subunit
CMD_0979 Short chain dehydrogenase reductase (SDR) Oxidoreductase
SDR dehydrogenase CMD_1024 NADH-quinone oxidoreductase
membrane L CMD_1393 oxidoreductase SDR putative deoxygluconate
dehydrogenase (NAD+ 3-oxidoreductase) CMD_2679
Dehydratasedihydroxy acid dehydratase CMD_2683 oxidoreductase
oxopropionate reductase CMD_2684 short-chain dehydrogenasereductase
SDR CMD_3184 nitrate reductase alfa subunit CMD_3721 thiol-disulfide
isomerase and thioredoxin oxidoreductase
Deacetylase 1 CMD_2431 N-acetylglucosaminylphosphatidylinositol deacetylase
superfamily
Hydrolase 2 CMD_0991 Dienelactone hydrolase CMD_2895 Putative glycosyl
hydrolase
Transferase 4 CMD_1640 D12 class N6 adenine-specific DNA methyltransferase
CMD_2432 Glycosyltransferase CMD_2433 putative acetyltransferase
CMD_3023 adenine-specific DNA modification methyltransferase
Sulfatase 3 CMD_2898 Possible sulfatase arylsulfatasearylsulfatransferase
CMD_2902 Arylsulfatasesulfatase CMD_2905 Putative
sulfatasearylsulfatase
Bacteriocin 3 CMD_0982 Thermosatble hemolysin superfamily CMD_0993 Pyocin S2
superfamily (bacteriocin that kills other bacterial strain) CMD_3705 S-type
pyocin containing domain protein
Type VI secretion 2 CMD_1802 EvpB family type VI secretion protein CMD_2364 type VI
effector Hcp1
Kinase 4 CMD_0989 Heavy metal Sensor kinase CMD_1794 Polo kinase kinase
(PKK) superfamily CMD_3124 predicted diacylglycerol kinase CMD_4266
putative signal transduction kinase
Transporter 11 CMD_0973 TrkA-N domain-containing protein (K uptake transporter)
CMD_1407 Branched-chain amino acid transport system carrier protein
CMD_2678 Sodium Symporter CMD_3176 glutamine ABC transporter
CMD_3544 Ribose ABC transport system CMD_3693 TRAP-type C4-
dicarboxylate transport system binding protein CMD_3694 TRAP-type C4-
dicarboxylate transport system binding protein small permease component
(predicted N-acetylneuraminate transporter) CMD_3695 TRAP-type C4-
dicarboxylate transport system large permease CMD_4337 23-diketo-L-
gulonate TRAP transporter small permease protein yiaM component
CMD_4338 23-diketo-L-gulonate TRAP transporter large permease protein
yiaN CMD_4600 putative transporter involved in Zinc uptake (YciA
protein)
170
Supplmental Table S5-1 (continued)
Unique gene
classes
Number Gene ID and functions
Membrane
exported protein
8 CMD_0713 putative membrane protein wzy CMD_0983 Putative exported
protein precursor CMD_0986 Putative transmembrane protein
CMD_0987 putative exported protein precursor CMD_1458 putative
exported protein CMD_2903 Putative exported protein CMD_2904
putative secreted 5-nucleotidase CMD_4291 (S) putative integral
membrane export protein
Regulator 12 CMD_0977 probable transcriptional regulator protein CMD_0980 Putative
transcription activator TenA family CMD_0990 Two component heavy
metal response transcription regulator CMD_1766 Leucine responsive
regulatory protein CMD_2677 transcriptional regulator GntR CMD_2909
putative arylsulfatase regulator CMD_3279 Uncharacterized outer
membrane usher protein yqiG precursor CMD_3464 toxin transcription
regulatory protein CMD_3465 EAL regulator protein (signaling)
CMD_3475 putative transcription regulator CMD_3477 Fumarate and
nitrate reduction regulatory protein (for anaerobic adaptation) CMD_3812
putative regulator from phage
Integrase
recombinase
5 CMD_0785 CMD_0992 CMD_0994 CMD_4032 CMD_4424
Phage prophage
transposae related
genes
27 Phages CMD_1608 CMD_1609 CMD_1610 CMD_1611 CMD_1619
CMD_1631 CMD_1913 CMD_1914 CMD_1915 CMD_1917
CMD_1920 CMD_1921 CMD_1936 CMD_3351 CMD_3804
CMD_3805 CMD_3807 CMD_3808 CMD_4581 CMD_4609
CMD_4610 Transposae CMD_0252 CMD_0398 Prophage CMD_1645
CMD_3806 CMD_4425 CMD_4597
Miscellaneous 32 CMD_0095 ATP-dependent DNA helicase Rep CMD_0656 LysA protein
CMD_0710 Polysaccharide biosynthesis family protein CMD_0978 TPR
repeat containing protein CMD_0981 AMP dependant synthatase and
ligase CMD_0985 Cupin superfamily CMD_0987 flavodoxin
CMD_1404 Putative endoribonuclease CMD_1405 Putative PAC domain
protein CMD_1406 Phenazine (PhzF) biosynthesis protein CMD_1695
Putative fimbrial protein CMD_1793 HPChromosome segregation ATPase
domain protein CMD_1922 FRG uncharacterized protein domain
CMD_2206 L-fucose isomerase like protein CMD_2434 CS12 fimbria
outer membrane usher protein CMD_2680 HpcHHpaI aldosecitrate lyase
protein family CMD_2681 Aldose -1 epimerase CMD_2682
Gluconolaconase LRE like region CMD_2900 Putative cytoplasmic
protein CMD_3009 putative restriction endonuclease CMD_3277 Putative
yqi fimbrial adhesion CMD_3278 Hypothetical fimbrial chaperone yqiH
CMD_3340 Pea-VEAacid family CMD_3518 3-hydroxy-3-methylglutaryl
CoA synthase CMD_3801 putative reverse transcriptase CMD_3840 beta-
D-galactosidase alpha subunit CMD_4033 ParB family chromosome
partitioning protein CMD_4034 ParBrepB chromosomeplasmid
partitioning protein CMD_4035 RelASpoT protein CMD_4532 putative
DNA ligase -like protein CMD_4541 Antitermination protein Q and
CMD_4585 putative ATP dependent helicase
171
CHAPTER 6
60 CONCLUSION
61 Summary
Contamination of grain with mycotoxins including deoxynivalenol (DON) and other
trichothecenes is becoming a limiting factor for cereal and animal production (Binder et al
2007) To address the issue this study obtained a novel microbial source for DON
detoxification and determined candidate genes for the function which may have potential
applications to detoxify grain and grain-derived foodfeed contaminated with DON and TC
mycotoxins
From a screen of soil samples from crop field microorganism was isolated that is able
to de-epoxidize the most frequently detected foodfeed contaminating mycotoxin DON The
effects of media compositions temperature pH and aerobicanaerobic conditions on
microbial detoxification activity were assessed From 150 analyzed soil samples a mixed soil
microbial culture that showed DON de-epoxidation activity under aerobic conditions and
moderate temperature (~24degC) was obtained This culture exhibited a slow rate of microbial
DON to de-epoxyDON transformation (de-epoxidation) possibly because of competition
between de-epoxidizing and other microorganisms in the culture
The mixed microbial culture was enriched for DON detoxifying bacteria by reducing
the non-targeted microbial species through antibiotic treatments heating and subculturing on
agar medium and selecting colonies The enriched culture had higher DON de-epoxidation
activity than the initial culture Because of low abundance of DON detoxifying
microorganisms further culture enrichments were required to increase the target microbial
172
population Reculture of the partially enriched microbial community in nutritonally poor MSB
medium with high level of DON increased the DON de-epoxidizing bacterial population
Serial dilution plating and selecting single colonies eventually led to isolation of a DON-
detoxifying bacterial strain named ADS47
Phenotypic and 16S-rDNA sequence analyses provided evidence that the bacterium is
a strain of C freundii Strain ADS47 is a gram negative rod-shaped facultative anaerobe and
has peritrichous flagella Strain ADS47 was unable to use DON as sole carbon source
however increasing DON concentrations in growth media enhanced its DON metabolism
efficiency The pure bacterial culture was shown to be able to rapidly de-epoxidize DON
under aerobic conditions moderate temperatures and neutral pH Strain ADS47 more quickly
de-epoxidizes DON in aerobic conditions than the anaerobic conditions In addition to DON
strain ADS47 showed capability to de-epoxidize andor de-acetylate 10 different type-A and
type-B TC mycotoxins which often co-contaminant grain with DON
To clone the DON detoxifying genes a DON detoxifying bacterial DNA library in E
coli host was constructed followed by screening of the library clones for DON detoxification
activity The study produced a library giving approximately 10 fold coverage of the genome
of strain ADS47 Restriction digestion analysis determined that most of the clones contained
ADS47 DNA inserts This indicates that a good DNA library was produced which had gt99
probability of containing all the DNA of the ADS47 genome However activity-based
screening of the library could not identify any clones with DON de-epoxidation ability It is
assumed that bacterial DON de-epoxidation genes evolved in a certain environmental niche
which may be required for the DON de-epoxidation to be expressed and active but was not
supplied by the host E coli cells
173
An attempt was made to identify the bacterial genes encoding DON de-epoxidation
and de-acetylation enzymes by sequencing the genome of strain ADS47 and comparing its
sequence to the genomes of related bacteria that are not known to posses DON de-epoxidation
activity A RAST server based bioinformatics analysis determined that the 48 Mb circular
genome of ADS47 codes for 4610 predicted CDS A genome wide phylogenetic analysis
determined that strain ADS47 was gt87 identical to the pathogen C freundii strain Ballerup
The relationship to two other animalhuman pathogenic species C rodentium and C koseri
was more distant This result is consistent with the 16S-rDNA based phylogenetic analysis
It is assumed that the microbial DON detoxifying genes were acquired by horizontal
gene transfer or other means during the adaptation of ADS47 in mycotoxin contaminated soil
If it is correct then this means that those newly acquired genes are not common in other
related microbial species and no other Citrobacter species is known to degrade DON Hence
the study identified 270 unique genes to ASDS47 with a subset of 10 genes for reductases
oxidoreductases and a de-acetylase which were considered to be candidate genes for the
enzymes that have DON detoxifying activity Furthermore a targeted comparative genome
analysis showed that the mycotoxin detoxifying ADS47 genome does not harbour the
clinically important LEE encoded typendashIII secretion system and mycobacterium virulence
genes This suggests that strain ADS47 is not likely to be an animal pathogenic bacterium
unlike C freundii Ballerup which encodes the pathogenic relevance LEE operons
174
62 Research contributions
The research made four major contributions which may aid in reducing the risks of
animals and humans from DONTC exposure Firstly the study identified a novel bacterial
strain ADS47 that is able to detoxify DON and 10 related mycotoxins over a wide range of
temperatures under aerobic and anaerobic conditions The discovery has led to a patent
application for strain ADS47 supporting the novelty and importance of the finding This
bacterial strain could be used to detoxify trichothecene mycotoxins in contaminated animal
feed thus reducing food safety risks
Secondly the study resulted in the development of a method for isolating DON de-
epoxidizing microbes from soil as well as optimizing the conditions for bacterial DONTC
detoxification activity to increase the effectiveness usefulness of the strain
Thirdly the thesis work resulted in availability of the genome sequence of a
trichothecene detoxifying microbe which may ultimately lead to an understanding of the
cellularmolecular mechanisms of DONTC de-epoxidation and de-acetylation
genespathways The genome analysis resulted in identification of several candidate genes for
DONTC de-epoxidation and de-acetylation activities Genes which show de-epoxidation
activity could be used to develop DONTC resistant cereal cultivars Fourthly strain ADS47
lacks major animal pathogenicity genes (in particular the LEE-operons) as well as does not
encodes mycobacterium virulence genes Thus strain ADS47 has a great potential to be
utilized to detoxify cereal grain and may be used as a feed additive for livestockfishery All
together the reported work may have significant applications to produce and supply
DONTC-free food and feed
175
63 Further research
This research provides the basis for developing biological detoxification of DON and
related mycotoxins This goal might be achieved by using DON de-epoxidizing gene to
develop resistant plants enzymatic detoxification of contaminated feed and use of the bacteria
as a feed additive Follow-up studies may reach the above indicated goals Because the
bacterium showed de-epoxidation activity under aerobicanaerobic conditions and under wide
range of temperatures it is likely to have an application for enzymatic detoxification of
contaminated animal feed Therefore additional studies on bacterial efficiency to detoxify
contaminated cereal and grain-derived feed need to be conducted Although strain ADS47
does not encode major animal pathogenicity genes the bacterium must be directly tested on
animals to see if it could negatively affect animal digestive systems If not then the potential
for ADS47 is considerable for its use as a feed additive for pig poultry and fish Animal
feeding trials will also need to be performed to determine if DON detoxification is sufficient
to be cost effective as a feed additive In order to develop DONTC resistant cereal cultivars
the gene responsible to DON degrading activity among the presently identified candidate
genes (reductases oxidoreductases and de-acetylase) will need to be expressed in other
organisms like E coli and a codon modified version adapted for expression in plants like
corn and wheat
176
70 REFERENCES
Abbas HK Mirocha CJ Pawlosky RJ Pusch DJ 1985 Effect of cleaning milling and
baking on deoxynivalenol in wheat Applied and Environmental Microbiology 50482ndash
486
Accensi F Pinton P Callu P Abella-Bourges N Guel WJF Grosjean F Oswald IP 2006
Ingestion of low doses of deoxynivalenol does not affect haematological biochemical or
immune responses of piglets Journal of Animal Science 841935ndash1942
Agrawal D Mahapatra AK 2005 Vertically acquired neonatal Citrobacter brain abscessndash
case report and review of the literature Journal of Clinical Neuroscience 12188ndash190
Alexander NJ 2008 The TRI101 story engineering wheat and barley to resist Fusarium head
blight World Mycotoxin Journal 131-37
Alexander NJ Proctor RH McCormick SP 2009 Genes gene clusters and biosynthesis of
trichothecenes and fumonisins in Fusarium Toxin Reviews 28198ndash215
Ali ML Taylor JH Jie L Sun G William M Kasha KJ Reid LM Pauls KP 2005
Molecular mapping of QTLs for resistance to Gibberella ear rot in corn caused by
Fusarium graminearum Genome 48521-533
Alikhan N-F Petty NK Zakour NLB Beatson SA 2011 BLAST Ring Image Generator
(BRIG) simple prokaryote genome comparisons BMC Genomics 12402
Alisi A Spaziani A Anticoli S Ghidinelli M Balsano C 2008 PKR is a novel functional
direct player that coordinates skeletal muscle differentiation via p38MAPKAKT
pathways Cellular Signalling 20534ndash542
177
Amuzie CJ Shinozuka J Pestka JJ 2009 Induction of suppressors of cytokine signaling by
the trichothecene deoxynivalenol in the mouse Toxicological Science 111277ndash287
Ansorge WJ 2009 Next generation DNA sequencing techniques New Biotechnology
25195-203
Arnold JW Senter SD 1998 Use of digital aroma technology and SPME GC-MS to compare
volatile compounds produced by bacteria isolated from processed poultry Journal of the
Science of Food and Agriculture 78343ndash348
Audenaert K Van Broeck R Bekaert B De Witte F Heremans B Messens K Hofte
M Haesaert G 2009 Fusarium head blight (FHB) in Flanders population
diversity inter-species associations and DON contamination in commercial winter
wheat varieties European Journal of Plant Pathology 125445-458
Awad WA Ghareeb K Bohm J Zentek J 2010 Decontamination and detoxification
strategies for the Fusarium mycotoxin deoxynivalenol in animal feed and the
effectiveness of microbial biodegradation Food Additives and Contaminants 27510-520
Aziz K Bartels D Best AA DeJongh M Disz T Edwards RA Formsma K Gerdes S Glass
EM Kubal M Meyer F Olsen GJ Olson R Osterman AL Overbeek RA McNeil LK
Paarmann D Paczian T Parrello1 B Pusch GD Reich C Stevens R Vassieva O
Vonstein V Wilke A Zagnitko O 2008 The RAST Server Rapid Annotations using
Subsystems Technology BMC Genomics 975
Badger JL Stins MF Kim KS 1999 Citrobacter freundii invades and replicates in human
brain microvascular endothelial cells Infection and Immunology 674208ndash4215
Bae HK Pestka JJ 2008 Deoxynivalenol induces p38 interaction with the ribosome in
monocytes and macrophages Toxicological Science 10559-66
178
Bai GH Desjardins AE Plattner RD 2002 Deoxynivalenol-nonproducing Fusarium
graminearum causes initial infection but does not cause disease spread in wheat spikes
Mycopathologia 15391ndash98
Barrett JR 2000 Mycotoxins of molds and maladies Environmental Health Perspectives
108A20ndashA23
Bengoechea JA Skurnik M 2000 Temperature-regulated efflux pumppotassium antiporter
system mediates resistance to cationic antimicrobial peptides in Yersinia Molecular
Microbiology 3767ndash80
Bennett JW Klich M 2003 Mycotoxins Clinical Microbiology Reviews 16497ndash516
Benson DA Karsch-Mizrachi I Lipman DJ Ostell J Sayers EW 2009 GenBank Nucleic
Acids Research 37D26-31
Berdichevsky T Friedberg D Nadler C Rokney A Oppenheim A Rosenshine I 2005 Ler is
a negative autoregulator of the LEE1 operon in neuropathogenic Escherichia coli Journal
of Bacteriology 187349ndash357
Bernardo A Bai GH Guo PG Xiao K Guenzi AC Ayoubi P 2007 Fusarium graminearum-
induced changes in gene expression between Fusarium head blight-resistant and
susceptible wheat cultivars Functional and Integrative Genomics 769ndash77
Bhat RV Beedu SR Ramakrishna Y Munshi KL 1989 Outbreak of trichothecene
mycotoxicosis associated with consumption of mould-damaged wheat production in
Kashmir Valley India The Lancet 33335ndash37
Binder EM Tan LM Chin LJ Handl J Richard J 2007 Worldwide occurrence of
mycotoxins in commodities feeds and feed ingredients Animal Feed Science and
Technology 137265-282
179
Birzele B Meier A Hindorf H Kraumlmer J Dehne H-W 2002 Epidemiology of Fusarium
infection and deoxynivalenol content in winter wheat in the Rhineland Germany
European Journal of Plant Pathology 108667ndash673
Boddu J Cho S Muehlbauer GJ 2007 Transcriptome analysis of trichothecene-induced gene
expression in barley Molecular PlantndashMicrobe Interactions 201364ndash1375
Borenshtein D Schauer DB 2006 The Genus Citrobacter Prokaryotes 690ndash98
Boutigny A-L Richard-Forget F Barreau C 2008 Natural mechanisms for cereal resistance
to the accumulation of Fusarium trichothecenes European Journal of Plant Pathology
121411ndash423
Boutrif E Canet C 1998 Mycotoxin prevention and control FAO programmes Revue de
Meacutedecine Veacuteterinaire 149681-694
Brennan JM Leonard G Fagan B Cooke BM Ritieni A Ferracane R et al 2007
Comparison of commercial European wheat cultivars to Fusarium infection of head and
seedling tissue Plant Pathology 5655ndash64
Brenner DJ Grimont PA Steigerwalt AG Fanning GR Ageron E Riddle CF 1993
Classification of Citrobacteria by DNA hybridization designation of Citrobacter farmeri
sp nov Citrobacter youngae sp nov Citrobacter braakii sp nov Citrobacter
werkmanii sp nov Citrobacter sedlakii sp nov and three unnamed Citrobacter
genomospecies International Journal of Systematic Bacteriology 43645-58
Brookes G Barfoot P 2006 GM Crops The first ten years-global socio-economic and
environmental Impacts no 36 ISAAA (International Service for the Acquisition of Agri-
Biotech Applications) Ithaca New York
180
Bruce KD Hughes MR 2000 Terminal restriction fragment length polymorphism monitoring
of genes amplified directly from bacterial communities in soils and sediments Molecular
Biotechnology 16261ndash269
Buerstmayr H Ban T Anderson JA 2009 QTL mapping and marker-assisted selection for
Fusarium head blight resistance in wheat A review Plant Breeding 1281ndash26
Campbell H Choo TM Vigier B Underhill L 2002 Comparison of mycotoxin profiles
among cereal samples from eastern Canada Canadian Journal of Botany 80526ndash532
Carver et al 2005 ACT the Artemis Comparison Tool Bioinformatics 213422-3423
Carver et al 2009 DNAPlotter circular and linear interactive genome visualization
Bioinformatics 25119-120
Chen CY et al 2003 Comparative genome analysis of Vibrio vulnificus a marine pathogen
Genome Research 132577ndash2587
Chiu TC Yen JH Hsieh YN Wang YS 2005 Reductive transformation of dieldrin under
anaerobic sediment culture Chemosphere 601182ndash1189
Chung YJ Zhou HR Pestka JJ 2003 Transcriptional and posttranscriptional roles for p38
mitogen-activated protein kinase in upregulation of TNF-alpha expression by
deoxynivalenol (vomitoxin) Toxicology and Applied Pharmacology 193188ndash201
Chung EJ Lim HK Kim J-C Choi GJ Park EJ Lee MH Chung YR Lee S-W 2008 Forest
soil metagenome gene cluster involved in antifungal activity expression in Escherichia
coli Applied and Environmental Microbiology 74723-730
Collins TFX Sprando RL Black TN Olejnik N Eppley RM Hines FA Rorie J Ruggles DI
2006 Effects of deoxynivalenol (DON vomitoxin) on in utero development in rats Food
and Chemical Toxicology 44747ndash757
181
Cong Y Wang J Chen Z Xiong K Xu Q Hu F 2011 Characterization of swarming motility
in Citrobacter freundii FEMS Microbiology Letters 371160-171
Cossette F Miller JD 1995 Phytotoxic effect of deoxynivalenol and Gibberella ear rot
resistance of corn Natural Toxins 3383ndash388
Creppy EE 2002 Update of survey regulation and toxic effects of mycotoxins in Europe
Toxicological Letters 12719-28
Dai JS Wang D Guerlebeck C Laturnus S Guenther Z Lu SC Ewers C 2010 Suppression
subtractive hybridization identifies an autotransporter adhesin gene of E coli IMT5155
specifically associated with avian pathogenic Escherichia coli (APEC) BMC
Microbiology 10236
Daniel R 2005 The metagenomics of soil Nature Review Microbiology 3470-478
Davis KER Joseph SJ Janssen PH 2005 Effects of growth medium inoculum size and
incubation time on culturability and isolation of soil bacteria Applied and Environmental
Microbiology 71826-834
Delcher AL Phillippy A Carlton J Salzberg SL 2002 MUMmer 21 NUCmer and
PROmer are described in Fast Algorithms for Large-scale Genome Alignment and
Comparisonrdquo Nucleic Acids Research 302478-2483
Demain AL Adrio JL 2008 Contributions of microorganisms to industrial biology
Molecular Biotechnology 3841ndash455
Deng W Li Y Vallance BA Finlay BB 2001 Locus of enterocyte efffacement from
Citrobacter rodentium sequence analysis and evidence for horizontal transfer among
attaching and effacing pathogens Infection and Immunity 696323ndash6335
182
Desjardins AE Proctor RH Bai G McCormick SP Shaner G Beuchley G Hohn TM 1996
Reduced virulence of trichothecene-nonproducing mutants of Gibberella zeae in wheat
field tests Molecular Plant-Microbe Interactions 9775-781
Desjardins AE 2006 Fusarium mycotoxins chemistry genetics and biology The American
Phytopathological Society St Paul Minnesota USA p 260
Desjardins AE Proctor RH 2007 Molecular biology of Fusarium mycotoxins International
Journal of Food Microbiology 11947-50
Desmond OJ Manners JM Stephens AE MacLean DJ Schenk PM Gardiner DM Munn
AL Kazan K 2008 The Fusarium mycotoxin deoxynivalenol elicits hydrogen peroxide
production programmed cell death and defence responses in wheat Molecular Plant
Pathology 9435-445
Dill-Macky R Jones RK 2000 The effect of previous crop residues and tillage on Fusarium
head blight of wheat Plant Disease 8471ndash76
DrsquoMello JPF Macdonald AMC Dijksma WTP 1998 3-Acetyl deoxynivalenol and esterase
production in a fungicide-insensitive strain of Fusarium culmorum Mycotoxin Research
149ndash18
DrsquoMello JPF Placinta CM Macdonald AMC 1999 Fusarium mycotoxins a review of
global implications of animal health welfare and productivity Animal Feed Science and
Technology 80183-205
Doohan FM Brennan J Cooke BM 2003 Influence of climatic factors on Fusarium species
pathogenic to cereals European Journal of Plant Pathology 109755ndash768
183
Dyer RB Kendra DF Brown DW 2006 Real-time PCR assay to quantify Fusarium
graminearum wild-type and recombinant mutant DNA in plant material Journal of
Microbiological Methods 67534ndash542
Erecinska M Wilson DF 1981 Inhibitors of cytochrome c oxidase In inhibitors of
mitochondrial function edited by M Erecinska and DF Wilson Pergamon Press Ltd
Oxford pp 145-160
Eriksen GS 2003 Metabolism and toxicity of trichothecene Doctoral thesis Swedish
University of Agricultural Sciences Uppsala Sweden
Eriksen GS Pettersson H Lundh T 2004 Comparative cytotoxicity of deoxynivalenol
nivalenol their acetylated derivatives and de-epoxy metabolites Food Chemistry and
Toxicology 42619-624
Eudes F Comeau A Rioux S Collin J 2000 Phytotoxicity of eight mycotoxins associated
with Fusarium in wheat head blight Canadian Journal of Plant Pathology 22286ndash292
FAO (Food and Agriculture Organization) 1991 Food Nutrition and Agriculture Food for
the Future volume 1 FAO Rome
FAO (Food and Agriculture Organization) 2001 FNP73 Manual of the application of
HACCP system in mycotoxin prevention and control FAO Food and Nutrition paper no
73 FAO Rome
Fedorak PM Grbic-Galic D 1991 Aerobic microbial cometabolism of benzothiophene and
3-methylbenzothiophene Applied and Environmental Microbiology 57932-940
Felsenstein J 1989 PHYLIP - Phylogeny Inference Package (Version 32) Cladistics 5164-
166
184
Fernando WGD Amarasinghe C Demoz B Surujdeo‐Maharaj S Gauthier V Guerrieri G
Al‐Taweel K Brucircleacute‐Babel A Tamburic‐Illincic L Gilbert J 2011 What do we know of
the Fusarium graminearum chemotypes their epidemiology genetics and infection
process in Canada In Proc 7th
Canadian Workshop on Fusarium Head Blight edited by
J Gilbert A Tekauz N Sweetland and K Slusarenko Winnipeg Manitoba Canada p 45
Fierer N Jackson RB 2006 The diversity and biogeography of soil bacterial community
Proceedings of the National Academy of Sciences 103626-631
Fink-Gremmels J 1989 The significance of mycotoxin assimilation for meat animals
Deutsche Tierarztliche Wochenschrift 96360-363
Flint DH Emptage MH Finnegan MG Fu W Johnson MK 1993 The role and properties of
the iron-sulfur cluster in Escherichia coli dihydroxy-acid dehydratase Journal of
Biological Chemistry 26814732ndash14742
Foroud NA Eudes F 2009 Trichothecenes in cereal grains International Journal of
Molecular Science 10147-173
Forsell JH Jensen R Tai JH Witt M Lin WS Pestka JJ 1987 Comparison of acute
toxicities of deoxynivalenol (vomitoxin) and 15-acetyldeoxynivalenol in the B6C3F1
mouse Food and Chemical Toxicology 25155ndash162
Frey M Stettner C Gierl A 1998 A general method for gene isolation in tagging approaches
amplification of insertion mutagenised sites (AIMS) The Plant Journal 13717-721
Freeman GG Morrison RI 1949 The isolation and chemical properties of trichothecin an
antifungal substance from Trichothecium roseum Link Biochemical Journal 441ndash5
185
Fuchs E Binder EM Heidler D Krska R 2002 Structural characterization of metabolites
after the microbial degradation of type A trichothecenes by the bacterial strain BBSH 797
Food Additives and Contaminants 19379ndash386
Gabor EM Alkema WBL Janssen DB 2004 Quantifying the accessibility of the
metagenome by random expression cloning techniques Environmental Microbiology
6879ndash886
Gadd GM Griffiths AJ 1978 Microorganisms and heavy metal toxicity Microbial Ecology
4303ndash317
Gale LR Bryant JD Calvo S Giese H 2005 Chromosome complement of the fungal plant
pathogen Fusarium graminearum based on genetic and physical mapping and cytological
observations Genetics 171985ndash1001
Gao LY Pak M Kish R Kajihara K Brown EJ 2006 A mycobacterial operon essential for
virulence in vivo and invasion and intracellular persistence in macrophages Infection and
Immunity 741757ndash1767
Gareau MG Wine E Reardon C et al 2010 Probiotics prevent death caused by Citrobacter
rodentium infection in neonatal mice Journal of Infectious Diseases 20181e91
Gareis M Bauer J Thiem J Plank G Grabley S Gedek B 1990 Cleavage of
zearalenoneglycoside a masked mycotoxin during digestion in swine Journal of
Veterinary Medicine 37236-240
Geng L Xin W Haung DW Feng G 2006 A universal cloning vector using vaccinia
topoisomerase I Molecular Biotechnology 3323-28
186
Golkari S Gilbert J Ban T Procunier JD 2009 QTLspecific microarray gene expression
analysis of wheat resistance to Fusarium head blight in Sumai-3 and two susceptible
NILs Genome 52409ndash418
Goodstadt L Ponting CP 2004 Vitamin K epoxide reductase homology active site and
catalytic mechanism Trends in Biochemical Sciences 29289-292
Goswami RS Kistler HC 2004 Heading for disaster Fusarium graminearum on cereal
crops Molecular Plant Pathology 5515-525
Gouze ME Laffitte J Printon P Dedieux G Galinier A Thouvenot JP Loiseau N Oswald
IP Galtier P 2007 Effect of subacute oral doses of nivalenol on immune and metabolic
defence systems in mice Veterinary Research 38635ndash646
Guan S He J Young JC Zhu H Li X Ji C Zhou T 2009 Transformation of trichothecene
mycotoxins by microorganisms from fish digesta Aquaculture 290290ndash295
Gupta R Beg QK Lorenz P 2002 Bacterial alkaline proteases molecular approaches and
industrial applications Applied Microbiology and Biotechnology 5915ndash32
Haberle J Schweizer G Schondelmaier J Zimmermann G Hart L 2009 Mapping of QTL
for resistance against Fusarium head blight in the winter wheat population
PelikanBussardNing8026 Plant Breeding 12827ndash35
Hall BG 2004 Predicting the evolution of antibiotic resistance genes Nature Reviews
Microbiology 2430-435
Han JI et al 2011 Complete genome sequence of the metabolically versatile plant growth-
promoting endophyte Variovorax paradoxus S110 Journal of Bacteriology 1931183ndash
1190
187
Harvey RB Edrington TS Kubena LF Elissalde MH Corrier DE Rottinghaus GE 1995
Effect of aflatoxin and diacetoxyscirpenol in ewe lambs Bulletin of Environmental
Contamination and Toxicology 54325-330
Harvey RB Edrington TS Kubena LF Elissalde MH Casper HH Rottinghaus GE Turk JR
1996 Effects of dietary fumonisin B1-containing culture material deoxynivalenol-
contaminated wheat or their combination on growing barrows American Journal of
Veterinary Research 571790-1794
He J 2007 Microbial strategies for management of Fusarium graminearum and
transformation of deoxynivalenol Master of Science Thesis University of Guelph
Guelph Ontario Canada
He K Vines L Pestka JJ 2010a Deoxynivalenol-induced modulation of microRNA
expression in RAW 2647 macrophages-A potential novel mechanism for translational
inhibition Toxicologist 114310
He J Zhou T Young JC Boland GJ Scott PM 2010b Chemical and biological
transformations for trichothecene mycotoxins in human and animal food chains A review
Trends in Food Science and Technology 2167-76
He J Zhou T 2010 Patented techniques for detoxification of mycotoxins in feeds and food
matrices Rec Pat Food Nutr Agric 296ndash104
He P Young LG Forsberg C 1992 Microbial transformation of deoxynivalenol (vomitoxin)
Applied and Environmental Microbiology 583857-3863
Hell K Cardwell KF Setamou M Poehling H-M 2000 The influence of storage practices on
aflatoxins contamination in maize in four agroecological zones in Benin West Africa
Journal of Stored Product Research 36365ndash82
188
Henne A Schmitz RA Bomeke M Gottschalk G Daniel R 2000 Screening of
environmental DNA libraries for the presence of genes conferring lipolytic activity on
Escherichia coli Applied and Environmental Microbiology 663113ndash3116
Hodges GR Degener CE Barnes WG 1978 Clinical significance of Citrobacter isolates
American Journal of Clinical Pathology 7037ndash40
Hollinger K Ekperigin HE 1999 Mycotoxicosis in food producing animals The Veterinary
Clinics of North America Food Animal Practice 15133-165
Hong Y Kinoshita T 2009 Trypanosome glycosylphosphatidylinositol biosynthesis Korean
Journal of Parasitology 47197ndash204
Hope RJ Aldred D Magan N 2005 Comparison of environmental profiles for growth and
deoxynivalenol production by Fusarium culmorum and F graminearum on wheat grain
Letters in Applied Microbiology 40295ndash300
Horvath RS 1972 Microbial co-metabolism and the degradation of organic compounds in
nature Bacteriological Reviews 36146ndash155
Hsu D Shih LM Zee YC 1994 Degradation of rRNA in Salmonella strains a novel
mechanism to regulate the concentrations of rRNA and ribosomes Journal of
Bacteriology 1764761ndash4765
Huwig A Freimund S Kappeli O Dutler H 2001 Mycotoxin detoxification of animal feed
by different adsorbents Toxicological Letters 122179ndash188
Iordanov MS Pribnow D Magun JL Dinh TH Pearson JA Magun BE 1997 Ribotoxic
stress response activation of the stress activated protein kinase JNK1 by inhibitors of the
peptidyl transferase reaction and by sequence-specific RNA damage to the sarcinricin
loop in the 28S rRNA Molecular and Cellular Biology 173373ndash3381
189
Islam R Zhou T Young JC Goodwin PH Pauls KP 2012 Aerobic and anaerobic de-
epoxidation of mycotoxin deoxynivalenol by bacteria originating from agricultural soil
World Journal of Microbiology and Biotechnology 287-13
Jackson LS Bullerman LB 1999 Effect of processing on Fusarium mycotoxins Advances in
Experimental Medicine and Biology 459243-261
Jansen C von Wettstein D Schafer W Kogel K-H Felk A Maier FJ 2005 Infection patterns
in barley and wheat spikes inoculated with wild-type and trichodiene synthase gene
disrupted Fusarium graminearum Proceedings of the National Academy of Sciences
10216892ndash16897
Jube S Borthakur D 2007 Expression of bacterial genes in transgenic tobacco methods
applications and future prospects Electronic Journal of Biotechnology 10452-467
Kabak B Dobson AD 2009 Biological strategies to counteract the effects of mycotoxins
Journal of Food Protection 722006-2016
Kant P Reinprecht Y Martin CJ Islam R Pauls KP 2011 Integration of biotechnologies
disease resistance pathology and Fusarium In Comprehensive Biotechnology edited by
Moo-Young M Second Edition 4729-743
Kaplan CW Kitts CL 2004 Bacterial succession in a petroleum land treatment unit Applied
and Environmental Microbiology 701777-1786
Kaprelyants AS Kell DB 1996 Do bacteria need to communicate with each other for
growth Trends in Microbiology 4237-42
190
Kariholu U Rawal J Namnyak S 2009 Neonatal Citrobacter koseri meningitis and brain
abscess The Internet Journal of Pediatrics and Neonatology 10
(httpwwwispubcomjournalthe-internet-journal-of-pediatrics-and-
neonatologyvolume-10-number-1neonatal-citrobacter-koseri-meningitis-and-brain-
abscesshtml) (accessed on April 19 2012)
Karlovsky P 1999 Biological detoxification of fungal toxins and its use in plant breeding
feed and food production Natural Toxins 71ndash23
Karlovsky P 2011 Biological detoxification of the mycotoxin deoxynivalenol and its use in
genetically engineered crops and feed additives Applied Microbiology and
Biotechnology 91491-504
Kavanagh KL Jornvall H Persson B Oppermann U 2008 Medium and short-chain
dehydrogenasereductase gene and protein families the SDR superfamily functional and
structural diversity within a family of metabolic and regulatory enzymes Cellular and
Molecular Life Sciences 653895ndash3906
Khera KS Whalen C Angers G Vesonder RF Kuiper-Goodman T 1982 Embryotoxicity of
4-deoxynivalenol (vomitoxin) in mice Bulletin of Environmental Contamination and
Toxicology 29487ndash491
Kikot GE Hours RA Alconada TM 2009 Contribution of cell wall degrading enzymes to
pathogenesis of Fusarium graminearum a review Journal of Basic Microbiology 49
231ndash241
Kim UJ Shizuya H Jong PJ Birren B Simon MI 1992 Stable propogation of cosmid sized
human DNA inserts in a F factor based vector Nucleic Acids Research 201083-1065
191
Kimura M Kaneko I Komiyama M Takatsuki A Koshino H Yoneyama K Yamaguchi I
1998 Trichothecene 3-O-acetyltransferase protects both the producing organism and
transformed yeast from related mycotoxins cloning and characterization of Tri101
Journal of Biological Chemistry 2731654-1661
Kimura M Anzai H Yamaguchi I 2001 Microbial toxins in plantndashpathogen interactions
biosynthesis resistance mechanisms and significance The Journal of General and
Applied Microbiology 47149ndash160
Kimura M Tokai T Takahashi-Ando N Ohsato S Fujimura M 2007 Molecular and genetics
studies of Fusarium trichothecene biosynthesis pathways genes and evolution
Bioscience Biotechnology and Biochemistry 712105-2123
King RR McQueen RE Levesque D Greenhalgh R 1984 Transformation of deoxynivalenol
(vomitoxin) by rumen microorganisms Journal of Agricultural and Food Chemistry
32181ndash1183
Klappenbach JA Dunbar JM Schmidt TM 2000 rRNA operon copy number reflects
ecological strategies of bacteria Applied and Environmental Microbiology 661328ndash
1333
Knietsch A Waschkowitz T Bowien S Henne A Daniel R 2003 Metagenomes of complex
microbial consortia derived from different soils as sources for novel genes conferring
formation of carbonyls from short-chain polyols on Escherichia coli Journal of Molecular
Microbiology and Biotechnology 546ndash56
Kollarczik B Gareis M Hanelt M 1994 In vitro transformation of the Fusarium mycotoxins
deoxynivalenol and zearalenone by the normal gut microflora of pigs Natural Toxins
2105ndash110
192
Koonin EV Wolf YI Aravind L 2001 Prediction of the archaeal exosome and its
connections with the proteasome and the translation and transcription machineries by a
comparative-genomic approach Genome Research 11240ndash252
Kouadio JH Dano SD Moukha S Mobio TA Creppy EE 2007 Effects of combinations of
Fusarium mycotoxins on the inhibition of macromolecular synthesis malondialdehyde
levels DNA methylation and fragmentation and viability in Caco-2 cells Toxicon
49306ndash317
Krause L Diaz NN Bartels D Edwards RA Puumlhler A et al 2006 Finding novel genes in
bacterial communities isolated from the environment Bioinformatics 22281ndash289
Kubena LF Huff WE Harvey RB Phillips TD Rottinghaus GE 1989 Individual and
combined toxicity of deoxynivalenol and T-2 toxin in broiler chicks Poultry Science
68622ndash626
Kubena LF Harvey RB Huff WE Corrier DE Phillips TD Rottinghaus GE 1990 Efficacy
of hydrated sodium calcium aluminosilicate to reduce the toxicity of aflatoxin and T-2
toxin Poultry Science 691078ndash1086
Kubena LF Harvey RB Huff WE Elissalde MH Yersin AG Phillips TD Rottinghaus GE
1993 Efficacy of hydrated sodium calcium aluminosilicate to reduce the toxicity of
aflatoxin and diacetoxyscirpenol Poultry Science 7251ndash59
Kubena LF Edrington TS Harvey RB Buckley SA Phillips TD Rottinghaus GE Casper
HH 1997 Individual and combined effects of fumonisin B1 present in Fusarium
moniliforme culture material and T-2 toxin or deoxynivalenol in broiler chicks Poultry
Science 761239ndash1247
193
Kumar RA Gunasekaran P Lakshmanan M 1999 Biodegradation of tannic acid by
Citrobacter freundii isolated from a tannery effluent Journal of Basic Microbiology
39161-168
Kushiro M 2008 Effects of milling and cooking processes on the deoxynivalenol content in
wheat International Journal Molecular Science 92127-2145
Lane DJ 1991 16S23S rRNA sequencing In Nucleic acid techniques in bacterial
systematic edited by Stackebrandt E and Goodfellow M Wiley J and Sons Inc New
York pp 115-175
Lancova K Hajslova J Poustka J Krplova A Zachariasova M Dostalek P Saschambula L
2008 Transfer of Fusarium mycotoxins and ldquomaskedrdquo deoxynivalenol (deoxynivalenol-3-
glucoside) from field barley through malt to beer Food Additives and Contaminants
25732ndash744
Langmead B Trapnell C Pop M Salzberg SL 2009 Ultrafast and memory-efficient
alignment of short DNA sequences to the human genome Genome Biology 10R25
Larsen JC Hunt J Perrin I Ruckenbauer P 2004 Workshop on trichothecenes with a focus
on DON summary report Toxicological Letters 1531-22
Lee T Oh DW Kim HS Lee J Kim YH Yun SH Lee YW 2001 Identification of
deoxynivalenol- and nivalenol-producing chemotypes of Gibberella zeae by using PCR
Applied and Environmental Microbiology 672966ndash2972
Lee S-W Won K Lim HK Kim J-C Choi GJ Cho KY 2004 Screening for novel lipolytic
enzymes from uncultured soil microorganisms Applied and Environmental Microbiology
65720-726
194
Li L Christian J Stoeckert J David SR 2003 OrthoMCL Identification of Ortholog Groups
for Eukaryotic Genomes Genome Research 132178-2189
Li T Chang CY Jin DY Lin PJ Khvorova A Stafford DW 2004 Identification of the gene
for vitamin K epoxide reductase Nature 427541ndash544
Li Y 2007 Sensory signal transduction in the vagal primary afferent neurons Current
Medicinal Chemistry 142554ndash2563
Li MX Pestka JJ 2008 Comparative induction of 28S ribosomal RNA cleavage by ricin and
the trichothecenes deoxynivalenol and T-2 toxin in the macrophage Toxicological
Science 10567ndash78
Li R Li Y Kristiansen K Wang J 2008 SOAP short oligonucleotide alignment program
Bioinformatics 24713-4
Li W Schulman S Dutton RJ Boyd D Beckwith J Rapoport TA 2010 Structure of a
bacterial homologue of vitamin K epoxide reductase Nature 463507-512
Lichstein HC Malcolm HS 1943 Studies of the effect of sodium azide on microbic growth
and respiration Journal of Bacteriology 47221ndash230
Limay-Rios V Schaafsma AW 2011 2010 Update on the development of an integrated
mycotoxin management system in Ontario corn 30th
Centralia Swine Research Update
Kirkton Ontario Canada
Llorens A Hinojo MJ Mateo R Gonzalez-Jaen MT Valle- Algarra FM Logrieco A Jimenez
M 2006 Characterization of Fusarium spp isolates by PCR-RFLP analysis of the
intergenic spacer region of the rRNA gene (rDNA) International Journal of Food
Microbiology 106287ndash306
195
Lombaert GA Pellaers P Roscoe V Mankotia M Neil R Scott PM 2003 Mycotoxins in
infant cereal foods from the Canadian retail market Food Additives and Contaminants
20494ndash504
Luengo JM Garciacutea JL Olivera ER 2001 The phenylacetyl-CoA catabolon a complex
catabolic unit with broad biotechnological applications Molecular Microbiology
391434ndash1442
Luo X 1994 Food poisoning caused by Fusarium toxins In Proceedings of the Second
Asian Conference on Food Safety ILSI Thailand pp 129ndash136
Mabagala RB 1997 The effect of populations of Xanthomonas campestris pv phaseoli in
bean reproductive tissues on seed infection of resistant and susceptible bean genotypes
European Journal of Plant Pathology 103175ndash181
Madhyastha MS Marquardt RR Abramson D 1994 Structurendashactivity relationships and
interactions among trichothecene mycotoxins as assessed by yeast bioassay Toxicon
321147ndash1152
Maier FJ Miedaner T Hadeler B Felk A Salomon S Lemmens M Kassner H Schaumlfer W
2006 Involvement of trichothecenes in fusarioses of wheat barley and maize evaluated
by gene disruption of the trichodiene synthase (Tri5) gene in three field isolates of
different chemotype and virulence Molecular Plant Pathology 7449ndash461
Malojcic G Owen RL Grimshaw JP Glockshuber R 2008 Preparation and structure of the
charge-transfer intermediate of the transmembrane redox catalyst DsbB FEBS Letters
5823301ndash3307
196
Manoharan M Dahleen LS Hohn TM Neate SM Yu XH Alexander NJ et al 2006
Expression of 3-OH trichothecene acetyltransferase in barley (Hordeum vulgare L) and
effects on deoxynivalenol Plant Science 171699ndash706
Maresca M Mahfoud R Garmy N Fantini J 2002 The mycotoxin deoxynivalenol affects
nutrient absorption in human intestinal epithelial cells The Journal of Nutrition
1322723ndash2731
Martins ML Martins HM 2002 Influence of water activity temperature and incubation time
on the simultaneous production of deoxynivalenol and zearalenone in corn (Zea mays) by
Fusarium graminearum Food Chemistry 79315ndash318
Masuda D Ishida M Yamaguchi K Yamaguchi I Kimura M Nishiuchi T 2007 Phytotoxic
effects of trichothecenes on the growth and morphology of Arabidopsis thaliana Journal
of Experimental Botany 581617ndash1626
Matsumoto E Kawanaka Y Yun SJ Oyaizu H 2008 Isolation of dieldrin- and endrin-
degrading bacteria using 12-epoxycyclohexane as a structural analog of both compounds
Applied Microbiology and Biotechnology 801095-1103
McLean M 1996 The phytotoxicity of Fusarium metabolites an update since 1989
Mycopathologia 133163ndash179
Merhej J Richard-Forget F Barreau C 2011 Regulation of trichothecene biosynthesis in
Fusarium recent advances and new insights Applied Microbiology and Biotechnology
91519ndash528
Midwest Plan Service 1987 Grain Drying Handling Storage Handbook Ames Iowa Iowa
State University 2nd edition
197
Miller SS Reid LM Harris LJ 2007 Colonization of maize silks by Fusarium graminearum
the causative organism of Gibberella ear rot Canadian Journal of Botany 85369-376
Minervini F Fornelli F Flynn KM 2004 Toxicity and apoptosis induced by mycotoxins
nivalenol deoxynivalenol and fumonisin B1 in a human erythroleukemia cell line
Toxicology In Vitro 1821ndash28
Mitterbauer R Poppenberger B RaditschnigA Lucyshyn D Lemmens M Glossl J Adam G
2004 Toxin-dependent utilization of engineered ribosomal protein L3 limits trichothecene
resistance in transgenic plants Plant Biotechnology Journal 2329ndash340
Mocali S Benedetti A 2010 Exploring research frontiers in microbiology the challenge of
metagenomics in soil microbiology Research Microbiology 161497-505
Morozkina EV Zvyagilskaya RA 2007 Nitrate reductases structure functions and effect of
stress factors Biochemistry 721151ndash1160
Munkvold GP 2003 Cultural and genetic approaches to managing mycotoxins in maize
Annual Review of Phytopathology 4199-116
Mundy R MacDonald TT Dougan G Frankel G Wiles S 2005 Citrobacter rodentium of
mice and man Cellular Microbiology 71697ndash1706
Nakamura N Inoue N Watanabe R Takahashi M Takeda J Stevens VL Kinoshita T 1997
Expression cloning of PIG-L a candidate N-acetylglucosaminyl-phosphatidylinositol
deacetylase The Journal of Biological Chemistry 27215834ndash15840
Nganje WE Bangsund DA Leistritz FL Wilson WW Tiapo NM 2002 Estimating the
economic impact of a crop disease the case of Fusarium head blight in US wheat and
barley National Fusarium Head Blight Forum Proceedings East Lansing Michigan State
University pp 275ndash281
198
Nielsen C Lippke H Didier A Dietrich R Martlbauer E 2009 Potential of deoxynivalenol
to induce transcription factors in humanhepatoma cells Molecular Nutrition and Food
Research 53479ndash491
Nielsen JKS Vikstroumlm AC Turner P Knudsen LE 2011 Deoxynivalenol transport across
the human placental barrier Food and Chemical Toxicology 492046-2052
Nishiuchi T Masuda D Nakashita H Ichimura K Shinozaki K Yoshida S et al 2006
Fusarium phytotoxin trichothecenes have an elicitor-like activity in Arabidopsis thaliana
but the activity differed significantly among their molecular species Molecular Plantndash
Microbe Interactions 19512ndash520
Ohsato S Ochiai-Fukuda T Nishiuchi T Takahashi-Ando N Koizumi S Hamamoto H
Kudo T Yamaguchi I Kimura M 2007 Transgenic rice plants expressing trichothecene
3-O-acetyltransferase show resistance to the Fusarium phytotoxin deoxynivalenol Plant
Cell Reports 26531-538
Okubara PA Blechl AE McCormick SP Alexander NJ Dill-Macky R Hohn TM 2002
Engineering deoxynivalenol metabolism in wheat through the expression of a fungal
trichothecene acetyltransferase gene Theoritical and Applied Genetics 10674-83
Ong CL Beatson SA Totsika M Forestier C McEwan AG Schembri MA 2010 Molecular
analysis of type 3 fimbrial genes from Escherichia coli Klebsiella and Citrobacter
species BMC Microbiology 10183
Osborne LE Stein JM 2007 Epidemiology of Fusarium head blight on small-grain cereals
International Journal of Food Microbiology 119103-108
Page RDM 1996 TREEVIEW An application to display phylogenetic trees on personal
computers Computer Applications in the Biosciences 12357-358
199
Parkhill J Dougan G James KD Thomson NR Pickard D Wain J Churcher C Mungall KL
Bentley SD Holden MT et al 2001 Complete genome sequence of a multiple drug
resistant Salmonella enterica serovar Typhi CT18 Nature 413848ndash852
Paterson RRM Lima N 2010 How will climate change affect mycotoxins in food Food
Research International 431902ndash14
Paul PA El-Allaf SM Lipps PE Madden LV 2004 Rain splash dispersal of Gibberella zeae
within wheat canopies in Ohio Phytopathology 941342-1349
Pestka JJ Lin WS Miller ER 1987 Emetic activity of the trichothecene 15-
acetyldeoxynivalenol in pigs Food and Chemical Toxicology 25855ndash858
Pestka JJ Smolinski AT 2005 Deoxynivalenol Toxicology and potential effects on humans
Journal of Toxicology Environmental Health 839ndash69
Pestka JJ 2007 Deoxynivalenol toxicity mechanisms and animal health risks Animal Feed
Science and Technology 137283-298
Pestka JJ 2008 Mechanisms of deoxynivalenol-induced gene expression and apoptosis Food
Additives and Contaminants 251128-1140
Pestka J2010 Toxicological mechanisms and potential health effects of deoxynivalenol and
nivalenol World Mycotoxin Journal 3323ndash347
Petersohn A Brigulla M Haas S Hoheisel JD Voumllker U Hecker M 2001 Global Analysis
of the General Stress Response of Bacillus subtilis Journal of Bacteriology 1835617-
5631
200
Petty NK Bulgin R Crepin VF Cerdeno-Tarraga AM Schroeder GN Quail MA Lennard N
Corton C Barron A Clark L Toribio AL Parkhill J Dougan G Frankel G Thomson NR
2010 The Citrobacter rodentium genome sequence reveals convergent evolution with
human pathogenic Escherichia coli Journal of Bacteriology 192525ndash538
Petty NK Feltwell T Pickard D Clare S Toribio AL et al 2011 Citrobacter rodentium is
an unstable pathogen showing evidence of significant genomic flux PLoS Pathology
7e1002018 doi101371journalppat1002018
Pittet A 2001 Natural occurrence of mycotoxins in foods and feeds A decade in review In
Mycotoxins and phytotoxins in perspective at the turn of the millennium edited by De
Koe WJ Samson RA Van Egmond HP Gilbert J Sabino M W J de Koe Wageningen
Netherlands pp 153ndash172
Placinta CM DrsquoMello JPF Macdonald AMC 1999 A review of worldwide contamination of
cereal grains and animal feed with Fusarium mycotoxins Animal Feed Science and
Technology 7821ndash37
Plank B Schuh M Binder EM 2009 Investigations on the effect of two feed additives
Biomin BBSH 797 and Mycofix Plus (R) 3E as detoxificants of DON contaminated feed
of piglets Wien Tierarztl Monatssch 9655ndash71
Poapolathep A Ohtsuka R Kiatipattanasakul W Ishigami N Nakayama H Doi K 2002
Nivalenol-induced apoptosis in thymus spleen and Peyerrsquos patches of mice Experimental
and Toxicological Pathology 53441ndash446
201
Poppenberger B Berthiller F Lucyshyn D Sieberer T Schuhmacher R Krska R Kuchler K
Gloumlssl J Luschnig C Adam G 2003 Detoxification of the Fusarium mycotoxin
deoxynivalenol by a UDP-glucosyltransferase from Arabidopsis thaliana Journal of
Biological Chemistry 27847905ndash47914
Prelusky DB Trenholm HL 1993 The efficacy of various classes of anti-emetics in
preventing deoxynivalenol-induced vomiting in swine Natural Toxins 1296ndash302
Proctor RH Hohn TM McCormick SP 1995 Reduced virulence of Gibberella zeae caused
by disruption of a trichothecene toxin biosynthetic gene Molecular Plant-Microbe
Interactions 8593-601
Puchenkova SG 1996 Enterobacteria in areas of water along the Crimean Coast
Mikrobiolohichnyĭ zhurnal 58 3-7
Qi BW Hao BL 2004 Whole-proteome prokaryote phylogeny without sequence alignment
a K-string composition approach Journal of Molecular Evolution 581ndash11
Quarta A Mita G Haidukowski M Logrieco A Mule G Visconti A 2006 Multiplex PCR
assay for the identification of nivalenol 3- and 15-acetyl-deoxynivalenol chemotypes in
Fusarium FEMS Microbiological Letters 2597ndash13
Rehr B Klemme JH 1989 Formate dependent nitrate and nitrite reduction to ammonia by
Citrobacter freundii and competition with denitrifying bacteria Antonie van Leeuwenhoek
Journal of Microbiology 56311ndash321
Rocha O Ansari K Doohan FM 2005 Effects of trichothecene mycotoxins on eukaryotic
cells A review Food Additives and Contaminants 22369ndash378
Roscoe V Lombaert GA Huzel V et al 2008 Mycotoxins in breakfast cereals from the
Canadian retail market a 3-year survey Food Additives and Contaminants 25347ndash355
202
Rotter BA Prelusky DB Pestka JJ 1996 Toxicology of deoxynivalenol (vomitoxin) Journal
of Toxicology and Environment Health 481ndash34
Ryu JC Ohtsubo K Izurniyarna N Nakamura K Tanaka T Yamarnura H Ueno Y 1988
The acute and chronic toxicities of nivalenol in mice Fundamental and Applied
Toxicology 1138ndash47
Saier MH Roseman S 1972 Inducer exclusion and repression of enzyme synthesis in
mutants of Salmonella typhimurium defective in enzyme I of the phosphoenolpyruvate
sugar phosphotransferase system The Journal of Biological Chemistry 247972ndash975
Saier MHJ Tran CV Barabote RD 2006 TCDB the transporter classification database for
membrane transport protein analyses and information Nucleic Acids Research 34D181ndash
186
Saitou N Nei M 1987 The neighbor-joining method a new method for reconstructing
phylogenetic trees Molecular Biology and Evolution 4406ndash425
Savelieva O Kotova I Roelofsen W Stams AJM Netrusov A 2004 Utilization of
aminoaromatic acids by a methanogenic enrichment culture and by a novel Citrobacter
freundii strain Archives of Microbiology 181163ndash170
Schaafsma AW Tamburic-Ilinic L Miller JD Hooker DC 2001 Agronomic considerations
for reducing deoxynivalenol in wheat grain Canadian Journal of Plant Pathology 23279ndash
285
Schaafsma AW 2002 Economic changes imposed by mycotoxins in food grains case study
of deoxynivalenol in winter wheat Advances in Experimental Medicine and Biology
504271ndash276
203
Scharf C Riethdorf S Ernst H Engelmann S Voumllker U Hecker M 1998 Theoredoxin is an
essential protein induced by heat shock and other stresses in Bacillus subtilis Journal of
Bacteriology 1801869-1877
Schatzmayr G Zehner F Taubel M Schatzmayr D Klimitsch A Loibner AP Binder EM
2006 Microbiologicals for deactivating mycotoxins Molecular Nutrition and Food
Research 50543-551
Schollenberger M Muumlller H-M Ruumlfle R Suchy S Plank S Drochner W 2006 Natural
occurrence of 16 Fusarium toxins in grains and feedstuffs of plant origin from Germany
Mycopathologia 16143ndash52
Scientific Committee for Food 2002 Opinion on Fusarium toxins Part 6 Group evaluation
of T-2 toxin HT-2 toxin nivalenol and deoxynivalenol (available at http
wwweuropaeuintcommfoodfsscscfout123_enpdf)
Schulman S Wang B Li W Rapoport TA 2010 Vitamin K epoxide reductase prefers ER
membrane-anchored thioredoxin-like redox partners Proceedings of the National
Academy of Sciences 10715027ndash15032
Scott PM 1997 Multi-year monitoring of Canadian grains and grain-based foods for
trichothecenes and zearalenone Food Additives and Contaminants 14333-339
Seitz LM Bechtel DB 1985 Chemical physical and microscopal studies of scab-infected
hard red winter wheat Journal of Agricultural and Food Chemistry 33373-7
Sergent T Parys M Garsou S Pussemier L Schneider YJ Larondelle Y 2006
Deoxynivalenol transport across human intestinal Caco-2 cells and its effects on cellular
metabolism at realistic intestinal concentrations Toxicological Letters 164167ndash176
204
Sharma J Fulekar MH 2009 Potential of Citrobacter freundii for bioaccumulation of
heavy metalndashcopper Biology and Medicine 17-14
Sherif SO Salama EE Abdel-Wahhab MA 2009 Mycotoxins and child health the need for
health risk assessment International Journal of Hygiene and Environmental Health
212347ndash368
Shifrin VI Anderson P 1999 Trichothecene mycotoxins trigger a ribotoxic stress response
that activates c-Jun N-terminal kinase and p38 mitogen-activated protein kinase and
induces apoptosis The Journal of Biological Chemistry 27413985ndash13992
Shima J Takase S Takahashi Y Iwai Y Fujimoto H Yamazaki M Ochi K 1997 Novel
detoxification of the trichothecene mycotoxin deoxynivalenol by a soil bacterium isolated
by enrichment culture Applied and Environmental Microbiology 633825ndash3830
Singh J Behal A Singla N Joshi A Birbian N et al 2009 Metagenomics Concept
methodology ecological inference and recent advances Biotechnology Journal 4480ndash
494
Smith TK McMillan EG Castillo JB 1997 Effect of feeding blends of Fusarium mycotoxin-
contaminated grains containing deoxynivalenol and fusaric acid on growth and feed
consumption of immature swine Journal of Animal Science 752184ndash2191
Smits WK Dubois JYF Bron S van Dijl JM Kuipers OP 2005 Tricksy business
transcriptome analysis reveals the involvement of thioredoxin in redox homeostasis
oxidative stress sulfur metabolism and cellular differentiation in Bacillus subtilis Journal
of Bacteriology 1873921ndash3930
205
Sprando RL Collins TFX Black TN Olejnik N Rorie JI Eppley RM Ruggles DI 2005
Characterization of the effect of deoxynivalenol on selected male reproductive endpoints
Food Chemistry and Toxicology 43623ndash635
Starkey DE Ward TJ Aoki T Gale LR Kistler HC Geiser DM Suga H Toacuteth B Varga J
ODonnell K 2007 Global molecular surveillance reveals novel Fusarium head blight
species and trichothecene toxin diversity Fungal Genetics and Biology 441191ndash1204
Stevenson BS Schmidt TM 1997 Growth rate-dependent expression of RNA from plasmid-
borne rRNA operons in Escherichia coli Journal of Bacteriology 1801970ndash1972
Stothard P Wishart DS 2005 Circular genome visualization and exploration using
CGView Bioinformatics 21537ndash539
Suenaga H Ohnuki T Miyazaki K 2007 Functional screening of a metagenomic library for
genes involved in microbial degradation of aromatic compounds Environmental
Microbiology 92289-97
Surai P Mezes M Fotina TI Denev SD 2010 Mycotoxins in Human Diet A Hidden
Danger In Modern Dietary Fat Intakes in Disease Promotion Edited by F De Meester et
al Nutrition and Health DOI 101007978-1-60327-571-2_18
Sutton JC 1982 Epidemiology of wheat head blight and maize ear rot caused by Fusarium
graminearum Canadian Journal of Plant Pathology 4195
Swaving J De Bont JAM Westphal A De Kok A 1996 A novel type pyridine nucleotide-
disulfide oxidoreductase is essential for NAD+- and NADPH-dependent degradation of
epoxyalkanes by Xanthobacter strain Py2 Journal of Bacteriology 1786644ndash6
206
Swanson SP Rood JHD Behrens JC Sanders PE 1987 Preparation and characterization of
the deepoxy trichothecenes deepoxy HT-2 deepoxy T-2 triol deepoxy T-2 tetraol
deepoxy 15-monoacetoxyscirpenol and deepoxy scirpentriol Applied and Environmental
Microbiology 532821ndash2826
Swanson SP Helaszek C Buck WB Rood JHD Haschek WM 1988 The role of intestinal
microflora in the metabolism of trichothecene mycotoxins Food Chemistry and
Toxicology 26823ndash829
Tian F Yu Y Chen B Li H Yao Y-F Guo X-K 2009 Bacterial archaeal and eukaryotic
diversity in Arctic sediment as revealed by 16S rRNA and 18S rRNA gene clone libraries
analysis Polar Biology 3293-103
Tatusov RL Galperin MY Natale DA Koonin EV et al 2000 Artemis sequence
visualization and annotation Bioinformatics 16944-945
Trail F Haixin X Loranger R Gadoury D 2002 Physiological and environmental aspects of
ascospore discharge in Gibberella zeae (anamorph Fusarium graminearum) Mycologia
94181-189
Tsuge K Ohata Y Shoda M 2001 Gene yerP involved in surfactin self-resistance in
Bacillus subtilis Antimicrobial Agents and Chemotherapy 453566ndash3573
Tucker 2001 The ldquoyellow rainrdquo controversy Lesson for arms control compliance The
Nonproliferation Review 825-42
Ueno Y 1985 The toxicology of mycotoxins Critical Reviews in Toxicology 1499-132
Ueno Y Nakayama K Ishii K Tashiro F Minoda Y Omori T Komagata K 1983
Metabolism of T-2 toxin in Curtobacterium sp strain 114-2 Applied and Environmental
Microbiology 46120ndash127
207
van Elsas JD Speksnijder AJ van Overbeek LS 2008 A procedure for the metagenomics
exploration of disease suppressive soils Journal of Microbiological Methods 75515ndash522
van Ginkel CG de Bont JAM 1986 Isolation and characterization of alkene-utilizing
Xanthobacter spp Archives of Microbiology 145403ndash407
van Lanen SG Shen B 2006 Microbial genomics for the improvement of natural product
discovery Current Opinion in Microbiology 9252ndash260
Varga J Toacuteth B 2005 Novel strategies to control mycotoxins in feeds a review Acta
Veterinaria Hungarica 53189-203
Voigt CA Schafer W Salomon S 2005 A secreted lipase of Fusarium graminearum is a
virulence factor required for infection of cereals The Plant Journal 42364ndash375
Vogelgsang S Hecker A Musa T Dorn B Forrer HR 2011 On farm experiments over five
years in a grain maizewinter wheat rotation Effect of maize residue treatments on
Fusarium graminearum infection and deoxynivalenol contamination in wheat In Proc
7th
Canadian Workshop on Fusarium Head Blight edited by J Gilbert A Tekauz N
Sweetland and K Slusarenko Winnipeg Manitoba Canada p 41
Voumllkl A Vogler B Schollenberger M Karlovsky P 2004 Microbial detoxification of
mycotoxin deoxynivalenol Journal of Basic Microbiology 44147ndash156
Wagacha JM Muthomi JW 2007 Fusarium culmorum Infection process mechanisms of
mycotoxin production and their role in pathogenesis in wheat Crop Protection 26877-
885
Wakulinski W 1989 Phytotoxicity of the secondary metabolites of fungi causing wheat head
fusariosis (head blight) Acta Physiologiae Plantarum 11301ndash306
208
Wang JT Chang SC Chen YC Luh KT 2000 Comparison of antimicrobial susceptibility of
Citrobacter freundii isolates in two different time periods The Journal of Microbiology
Immunology and Infection 33258-62
Wannemacher RJ Stanley L Wiener MD 2000 Trichothecenes mycotoxins In Medical
aspects of chemical and biological warfare edited by Zajtchuk R Washington DC
Department of the Army pp 656-676
Westphal AH Swaving J Jacobs L de Kok A 1998 Purification and characterization of a
flavoprotein involved in the degradation of epoxyalkanes by Xanthobacter Py2 European
Journal of Biochemistry 257160ndash168
Whalen JG Mully TW Enlgish JC 2007 Spontaneous Citrobacter freundii infection in an
immunocompetent patient Archives of Dermatology 143124-5
White 2007 The physiology and biochemistry of prokaryotes Third edition Oxford
University Press New York Oxford p 628
Wilcke WF Morey RV 1995 Natural-air corn drying in the upper Midwest University of
Minnesota Minnesota Extension Service Publication BU-6577-E pp 20
Windels CE 2000 Economic and social impacts of Fusarium headblight changing farms and
rural communities in the Northern Great Plains Phytopathology 9017ndash21
Winzeler EA Shoemaker DD Astromoff A Liang H Anderson K Andre B Bangham R
Benito R Boeke JD Bussey H et al 1999 Functional characterization of the S
cerevisiae genome by gene deletion and parallel analysis Science 285901ndash906
Yang GH Jarvis BB Chung YJ Pestka JJ 2000 Apoptosis induction by the satratoxins and
other trichothecene mycotoxins relationship to ERK p38 MAPK and SAPKJNK
activation Toxicology and Applied Pharmacology 164149ndash160
209
Yang Q Yin GM Guo YL Zhang DF Chen SJ Xu ML 2010 A major QTL for resistance
to Gibberella stalk rot in maize Theoretical and Applied Genetics 121673ndash687
Yazar S Omurtag GZ 2008 Fumonisins trichothecenes and zearalenone in cereals
International Journal of Molecular Science 92062ndash2090
Yoshizawa T Morooka N 1973 Deoxynivalenol and its monoacetate new mycotoxins from
Fusaruium roseum and moldy barley Agricultural and Biological Chemistry 372933ndash
2934
Yoshizawa T Takeda H Ohi T 1983 Structure of a novel metabolite from deoxynivalenol a
trichothecene mycotoxin in animals Agricultural and Biological Chemistry 472133ndash
2135
Yoshizawa T 1993 Red-mold diseases and natural occurrence in Japan In Trichothecenes-
Chemistry Biology and Toxicological Aspects edited by Y Ueno Elsevier Amsterdam
Netherlands p 195-209
Yoshizawa T Sakamoto T Okamoto K 1984 In vitro formation of 3rsquo-hydroxy T-2 and 3rsquo-
hydroxy HT-2 toxins from T-2 by liver homogenates from mice and monkeys Applied
and Environmental Microbiology 47130-134
Young JC Subryan LM Potts D McLaren ME Gobran FH 1986 Reduction in levels of
deoxynivalenol in contaminated wheat by chemical and physical treatment Journal of
Agricultural and Food Chemistry 34461ndash465
Young JC Trenholm HL Friend DW Prelusky DB 1987 Detoxification of deoxynivalenol
with sodium bisulfite and evaluation of the effects when pure mycotoxin or contaminated
corn was treated and given to pigs Journal of Agricultural and Food Chemistry 35259ndash
261
210
Young JC Zhu H Zhou T 2006 Degradation of trichothecene mycotoxins by aqueous
ozone Food Chemistry and Toxicology 44417ndash424
Young JC Zhou T Yu H Zhu H Gong J 2007 Degradation of trichothecene mycotoxins by
chicken intestinal microbes Food Chemistry and Toxicology 45136-143
Yu Z Morrison M 2004 Comparisons of different hypervariable regions of rrs genes for use
in fingerprinting of microbial communities by PCR-denaturing gradient gel
electrophoresis Applied and Environmental Microbiology 704800-4806
Yu H Zhou T Gong J Young C Su X Li X Zhu H Tsao R Yang R 2010 Isolation of
deoxynivalenol-transforming bacteria from the chicken intestines using the approach of
PCR-DGGE guided microbial selection BMC Microbiology 10182
Zeibdawi A Steele M Savoie A 2011 An overview of CFIAs mycotoxin monitoring
program in livestock feeds In Proc 7th
Canadian Workshop on Fusarium Head Blight
edited by J Gilbert A Tekauz N Sweetland and K Slusarenko Winnipeg Manitoba
Canada p 25
Zhang J-B Li H-P Dang F-J Qu B Xu Y-B Zhao C-S Lia Y-C 2007 Determination of the
trichothecene mycotoxins chemotypes and associated geographical distribution and
phylogenetic species of the Fusarium graminearum clade from China Mycological
Research 111967ndash975
Zhao JS Manno D Thiboutot S Ampleman G Hawari J 2007 Shewanella canadensis sp
nov and Shewanella atlantica sp nov manganese dioxide- and hexahydro-135-trinitro-
135-triazine-reducing psychrophilic marine bacteria International Journal of Systemic
Evolution and Microbiology 572155ndash2162
211
Zhao X-Q 2011 Genome-based studies of marine microorganisms to maximize the diversity
of natural products discovery for medical treatments Evidence-Based Complementary
and Alternative Medicine Article ID 384572 doi1011552011384572
ZhaoY Park RD Muzzarelli RA 2010 Chitin deacetylases properties and applications
Marine Drugs 824ndash46
Zhou HR Lau AS Pestka JJ 2003a Role of double-stranded RNA-activated protein kinase R
(PKR) in deoxynivalenol-induced ribotoxic stress response Toxicological Science
74335ndash344
Zhou HR Islam Z Pestka JJ 2003b Rapid sequential activation of mitogen-activated protein
kinases and transcription factors precedes proinflammatory cytokine mRNA expression in
spleen of mice exposed to the trichothecene vomitoxin Toxicological Sciences 72130ndash
142
Zhou T He J Gong J 2008 Microbial transformation of trichothecene mycotoxins World
Mycotoxin Journal 123-30
Zhou G Zhang H He Y He L 2010 Identification of a chitin deacetylase producing bacteria
isolated from soil and its fermentation optimization African Journal of Microbiology
Research 42597-2603