investigation into the insect biodiversity of grasslands surrounding the reedbed restoration at...
TRANSCRIPT

1
Grassland insects after gravel extraction
Project report submitted for the Quarry Life Award 2014
Alvin J. Helden
Animal and Environmental Research Group, Department of Life Sciences, Anglia Ruskin University, East Road, Cambridge, CB1 1PT, UK
Ribautodelphax imitans (female)

2
Contents
ABSTRACT 3
INTRODUCTION 4
OBJECTIVES 4
METHODS 5
Location and site details 5
Invertebrate sampling and identification 5
Statistical methods 6
Invertebrate density and habitat area 6
RESULTS 6
Invertebrates recorded 6
Comparison of grazed and ungrazed grassland 7
Comparison of older and more recently restored grassland 8
DISCUSSION 8
CONCLUSIONS 10
ACKNOWLEDGEMENTS 11
REFERENCES 11
APPENDIX 1. SPECIES LIST 14
APPENDIX 2. MAPS & PHOTOGRAPHS 17
APPENDIX 3. GRAPHS 20

3
Abstract 1. Habitat restoration following gravel extraction has been long-associated with
freshwater lake creation. However at Needingworth, to the north-west of Cambridge, Hanson/Heidelberg Cement are working with the RSPB to create a major new wetland nature reserve to be called Ouse Fen.
2. As well as open water and reedbed, the Needingworth site has areas of grassland, which in themselves have the potential to provide valuable habitat for invertebrates.
3. We used suction sampling to collect grassland invertebrates to: a) find out more about the Hemiptera community, and in particular the Auchenorrhyncha (leafhoppers an allies) present; b) to test whether the number and diversity of invertebrates was higher in ungrazed areas of grassland compared to those grazed by cattle; and c) to investigate whether the abundance and diversity in an older area of restoration exceeded than in a newer area.
4. All invertebrates collected were sorted and counted at order level and the Hemiptera were identified to species, genus or morphospecies.
5. In total 26092 were sampled, consisting of 3296 Araneae, 2462 Coleoptera, 7085 Diptera, 8765 Hemiptera, 2782 Hymenoptera and 1702 of other orders. The number of Hemiptera identified further was 5987, representing 81 species.
6. Five species of Auchenorrhyncha were found which were local or had more specific conservation interest (Table X): Euscelidius variegatus (Kbm.), Psammotettix striatus (L.), Eurybregma nigrolineata Scott., Xanthodelphax straminea (Ståland) Ribautodelphax imitans (Rib.).
7. Abundance of total invertebrates and of all orders, except for the Diptera, which showed the reverse pattern, was greater in ungrazed than grazed grassland. Calculations based on the numbers collected and area sampled indicated that the presence of ungrazed grassland resulted in 14.9% greater invertebrate abundance.
8. Diptera were more abundant in the more recently restored grasslands, while most other orders showed no difference with restoration age. This indicates that invertebrate populations can quite rapidly establish following restoration of ex-gravel extraction sites.
9. The dominant Hemiptera at the site were typically pioneer and eurytopic species, typical of many restored grassland but the presence of several rare and poorly known suggests Needingworth has specific value for the conservation of some rare Hemiptera. Management of the site should take into account the requirements of invertebrate communities.
10. The higher populations and species richness of invertebrates in ungrazed areas than grazed grassland, shows that setting aside ungrazed areas has been effective in enhancing invertebrate populations at Needingworth. This should be continued as part of wider strategy to provide a range of grassland management to enhance invertebrate biodiversity, as new areas of site are restored after gravel extraction.

4
Introduction Although by its very nature, quarrying is destructive of ecosystems, the extraction of gravel has a long association with the subsequent re-creation of habitats and the establishment of biodiversity they support. In particular this has occurred through the natural flooding of gravel pits, creating networks of freshwater lakes and some associated terrestrial habitats such as scrub and woodland. There are good examples of this in the Colne Valley and Lee Valley to the north-west and north of Greater London, respectively.
Needingworth Quarry to the north-west of Cambridge, UK is the location for a different approach following gravel extraction. Hanson/Heidelberg Cement has been collaborating with the Royal Society for the Protection of Birds (RSPB) to carry out a phased restoration to create a very large reedbed-focussed wetland nature reserve, called Ouse Fen (Aggregates Business Europe, 2010; RSPB, 2011) (Fig. 1 – for all figures see appendixes).
As gravel extraction is completed in sections of Needingworth, these areas are being restored into wetland habitats, including large areas of reedbed. The open water and reedbeds provide much of the focus of the RSPB's efforts, given the importance they have for birds, some of which are rare and threatened in the UK, as well as other wildlife. However reedbed restoration sites, typified by Needingworth also include areas of wet or dry rough grassland, with varying degrees of management through grazing, mowing and fencing. These grasslands in themselves potentially provide a valuable habitat for biodiversity, especially given the context of the considerable intensification of agricultural and other grasslands that has taken place over recent decades. This project aimed to investigate the grassland invertebrates present at Needingworth, with a particular focus on the Hemiptera. In particular we were interested in finding out how grassland management and age since restoration may affect the invertebrates communities.
Objectives In this project we aimed to investigate the insect biodiversity of grasslands surrounding the reedbed restoration units at Needingworth/Ouse Fen. The principal objective was to increase our knowledge of insect communities in these habitats at Needingworth/Ouse Fen and to inform management practice decisions in relation to these grasslands, which it is hoped will help to further enhance the biodiversity value of the restoration project. In particular we were interested in how leafhopper communities are affected by the level of grazing by cattle and the age of grassland since restoration. We had a specific focus on the Hemiptera and in especially the leafhoppers, planthoppers and froghoppers (Auchenorrhyncha), which are common in grassland and have been found to be useful indicators of management intensity effects (Nickel & Hildebrandt, 2003).
Our first hypothesis was that overall invertebrate populations, as well as the species richness and abundance of Hemiptera would be greater in fenced, ungrazed plots that had been created for the development of scrub, than in more open, cattle-grazed grassland within the older area of restoration.
Our second hypothesis was that Hemiptera species richness and overall invertebrate populations would be more abundant in the older area of restoration than the newer area.
Both these hypotheses were tested and the Hemiptera community characterised in more detail, using suction sampling to collect invertebrates at Needingworth in June and July 2014.

5
Methods
Location and site details Needingworth/Ouse Fen is located approximately 1 km north of the village of Over, Cambridgeshire. The site is centred on national grid reference TL377726 and at present the area restored after gravel extraction covers a total area of approximately 148 ha. The main habitats within the site can be divided into wetlands, composed of open water, reedbed and marshland vegetation, and a mixture of grassland and developing scrub (Fig. 2). Restoration, by Hanson and the RSPB, is being carried out in stages. The first area to be restored, from 2003 onwards (2003 restoration), was the area of approximately 69.9 ha to the south of the conveyor belt (Fig. 2) (RSPB, 2011). This was followed by restoration, from 2011 (2011 restoration), of the 78.2 ha area to the north of the conveyor belt.
In the 2003 restoration (Fig. 2) most of the grassland (23.7 ha, 33%) is being managed by grazing using a herd of 20 cattle, giving an approximate density of 0.85 cattle.ha-1 during the spring and summer. Of the remaining area, 38.0 ha (54%) are open water, reedbed or other wet vegetation. There are 8.3 ha (11%) of ungrazed grassland, which is fenced with the aim of eventual succession into scrub (Fig. 3). In most of these ungrazed areas there has been some low-density planting of woody vegetation, such as hawthorn Crataegus monogyna Jacq.
Management in the 2011 restoration, at least in 2014, has not involved grazing. Much of the area (52.4 ha, 67%) is covered with wetland habitats (Fig. 2) with most of the rest (18.7 ha, 23% of the 2011 restoration), being grassland managed with occasional mowing to control thistles and other similar dominant plants. One section of 7.1 ha (9%) in the central eastern part of the 2011 restoration (Fig. 2) is left unmanaged.
Habitat areas were calculated using satellite photographs from the Ordnance Survey, Get a Map (Ordnance Survey, 2014) and the Google Maps Area Calculator Tool (Daft Logic, 2014).
Invertebrate sampling and identification Invertebrates were sampled from 26 grassland locations (Fig. 2) on three dates, 6 June, 25 June and 21 July 2014, using a Vortis suction sampler (Arnold, 1994) (Fig. 4). In the 2003 restoration a sample was taken on each date from each of eight fenced ungrazed areas. These were paired with a sample from an adjacent area of grazed grassland. On the same dates, 10 samples were taken from the 2011 restoration. Eight were taken from randomly positioned points in the managed grassland areas and two from the unmanaged area (Fig. 2).
Each suction sample consisted of ten 16-second sucks (Brook et al., 2008) covering a total area of 0.2m2 (10 x 0.02m2). Invertebrates collected were preserved in 70% ethanol solution.
Invertebrates were removed from the samples an initially identified as: Araneae, Coleoptera, Diptera, Hemiptera, Hymenoptera or as other orders, although Collembola and Acari were excluded. The numbers of each order were counted.
Subsequently the Hemiptera from 6 June and 21 July were identified further. Adults Hemiptera were identified either to species level, or when this was not possible to genus or morphospecies, using: (Biedermann & Niedringhaus, 2009; della Giustina, 1989; Hawkins, 2003; Hodkinson & White, 1979; Le Quesne, 1960, 1965, 1969; Le Quesne & Payne, 1981; Ribaut, 1952; Stroyan, 1977; Taylor et al., 1984). Nymphs were identified to species level whenever possible but some could only be

6
assigned to morphospecies, genus, or in the case of some very early instars, to family. Auchenorrhyncha nymph identification was done with the aid of (Stöckman et al., 2013).
Statistical methods Statistical modelling of invertebrate data was carried out using R version 2.15.1 (R Core Team, 2012). Prior to modelling data were combined from the two sample dates to give a single replicate for each sampling location.
Grazed-ungrazed comparisons were made using generalised linear mixed models, with the lmer function from the lme4 package (Bates et al., 2012). In each model the response variable was the number of invertebrates, either in total or by order, with habitat type (grazed or ungrazed grassland) being the single explanatory variable. Sample pair was used as a random effect, and Poisson error structure was defined in all models. The number of Hemiptera species was modelled in the same way.
Models of invertebrates and age since restoration were similar. The response variables were the number of individuals or species. The single explanatory variable was the age since restoration (2003 and 2011 restoration). The comparison was made between the managed grassland of the 2011 restoration and the ungrazed habitat in the 2003 restoration. The models were generalised linear models, with Poisson error structure.
A species accumulation curve was calculated using the accumresult and accumplot functions from the BiodiversityR package (Bates et al., 2012), using Hemiptera species data. This gave some measure of the completeness of sampling. The specpool function from the vegan package (Oksanen et al., 2012) was used to calculate species richness estimators, which extrapolate to give a measure of the full number of species present.
Invertebrate density and habitat area Density estimates derived from the generalised linear models were combined with habitat area data to calculate the number of invertebrates that would be expected to occur in the 2003 restoration. This was done for the present 23.7ha grazed and 8.3ha ungrazed but also for other possible combinations of the two grassland habitat types, creating a model of the area of invertebrate abundance in relation to ungrazed grassland area. The relationship between number of individuals and area of ungrazed grassland was modelled with a linear regression.
Results
Invertebrates recorded The total number of invertebrates sampled at Needingworth was 26092, consisting of 3296 Araneae, 2462 Coleoptera, 7085 Diptera, 8765 Hemiptera, 2782 Hymenoptera and 1702 of other orders. The number of Hemiptera identified further was 5987, representing 81 identified species (Appendix 1). Five species of which were local or more specific conservation interest: Euscelidius variegatus (Kbm.), nationally notable (B); Psammotettix striatus (L.), Red Data Book K (insufficiently known); Eurybregma nigrolineata Scott. and Xanthodelphax stramineus (Stål), both local; and Ribautodelphax imitans (Rib.), Red Data Book K (Stewart, 2012).
Of the Hemiptera species found, the Heteroptera were represented by two species of Tingidae, nine Miridae, three Nabidae, one Berytinidae, four Lygaeidae and two Pentatomidae. Amongst the Auchenorrhyncha there were six Delphacidae, two

7
Aphrophoridae and 22 species of Cicadellidae. There were 26 species of Aphidoidea, two species of Psylloidea and one species of Coccoidea (Appendix 1).
The most abundant species was the leafhopper (Cicadellidae) Arthaldeus pascuellus (Fall.), with a total of 1279 individuals. Six other species were represented by more than 100 specimens: Javesella pellucida (F.) (Delphacidae), Anoscopus serratulae (F.) (Cicadellidae), Zyginidia scutellaris (H.-S.) (Cicadellidae), Uroleucon Mordvilko sp. (Aphidoidea), Euscelis incisus (Kbm.) (Cicadellidae) and Atheroides serratulus Haliday (Aphidoidea) (Appendix 1).
Although 81 species of Hemiptera were recorded, the species accumulation curve for samples taken throughout the Needingworth site, suggests that there are many more species that remain to be recorded (Fig. 5). Species richness estimators, which give an estimate of the actual number of species that may be present, gave the following: 119 ± 19 (SE) (Chao), 110 ± 8 (first order jack-knife), 128 (second order jack-knife), and 94 ± 4 (bootstrap). The mean of these four values is 112.8 species, suggesting that a further 32 unrecorded species occur at Needingworth.
Comparison of grazed and ungrazed grassland The total number of invertebrates recorded in the ungrazed grassland habitat was significantly higher than that in grazed (z=30.05 d.f.=14 p<0.001) (Fig. 6). The same pattern was true in five out of the six invertebrate orders (Fig. X): Araneae (z=12.34 d.f.=14 p<0.001), Coleoptera (z=4.22 d.f.=14 p<0.001), Hemiptera (z=28.42 d.f.=14 p<0.001), Hymenoptera (z=20.88 d.f.=14 p<0.001), other orders. The Diptera showed the opposite pattern, with greater abundance in the grazed habitat (z=-2.86 d.f.=14 p=0.004) (Fig. 7).
The generalised linear mixed model estimates of invertebrate density (two sampling dates combined) are shown in Table 1. Table 1. Estimates, derived from generalised linear mixed models, of the density at 0.2 and 1m2, of invertebrates in grazed and ungrazed grassland at Needingworth (Aran, Araneae; Col, Coleoptera; Dipt, Diptera; Hem, Hemiptera; Hym, Hymenoptera; Others, other orders). Habitat Invertebrate order Aran Col Dipt Hem Hym Others Overall a) Abundance/0.2m2 Grazed 70 72 237 175 76 21 668 Ungrazed 131 91 215 423 110 119 1117 b) Abundance/1m2 Grazed 350 360 1185 875 380 105 3340 Ungrazed 655 455 1075 2115 550 595 5585
As with abundance, the number of species of Hemiptera sampled was greater in ungrazed than grazed grassland (z=3.51 d.f.=14 p<0.001) (Fig. 8). Statistical model estimates of the number of species per 0.2m2 over the two sampling dates were: 13.8 and 21.1 for grazed and ungrazed, respectively.

8
The model of invertebrate abundance and ungrazed area in the 2003 restoration (Fig. 9), indicated that if all the grassland was grazed there would be 14.9% less grassland invertebrates. Conversely a doubling of the ungrazed area, from 8.3 to 16.6ha, would lead to a 14.9% increase. At 8.3ha the ungrazed area covers 25.9% of the area under grass but 16.6ha would cover 51.9%. If the ungrazed area were increased by a factor of 1.5 (12.4ha, 38.9%) there would be a 7.4% increase in invertebrates. The addition of 1ha would result in 1.8% more invertebrates.
Comparison of older and more recently restored grassland There was no significant difference in the overall invertebrate abundance of the 2003 restoration and the 2011 restoration (t=0.07 d.f.=14 p=0.946) (Fig. 10). There was also no difference in the abundance of Araneae (t=0.71 d.f.=14 p=0.491), Coleoptera (t=1.03 d.f.=14 p=0.319), Hemiptera (t=-1.0 d.f.=14 p=0.326) and Hymenoptera (t=0.26 d.f.=14 p=0.796). There were more Diptera in the 2011 restoration than the 2003 restoration (t=2.6 d.f.=14 p=0.021) but the pattern was reversed in the other orders (t=-8.2 d.f.=14 p<0.001) (Fig 11).
There was no significant difference in the number of Hemiptera between the two ages of restoration (t=-0.9 d.f.=14 p=0.389).
Discussion The Hemiptera community at Needingworth was a typical of many lowland grasslands in the UK, such as Coe Fen in Cambridge (Blake et al., 2011; Helden et al., 2011). Five of the seven most abundant species, A. pascuellus, J. pellucida, A. serratulae, Z. scutellaris and E. incisus are all associated with ruderal habitats and are often found in pastures and other fertilised grasslands (Nickel, 2003). Of the other two species, Uroleucon feeds on thistles (Compositae: Cynareae) (Blackman, 2010), which are typical of ruderal sites, and A. serratulus is described as common and widespread (Stroyan, 1977). Such pioneer and eurytopic species are typical of many restored grassland (Biedermann et al., 2005; Blake et al., 2011; Nickel & Achtziger, 2005). Most of the emphasis on grassland restoration has been following intensive agricultural land use, such as arable cropping. Evidence so far suggests that in such systems, due to the difficulties of high nutrient levels and problems in restoring the moisture conditions, the re-establishment of the original insect communities may take many years (Biedermann et al., 2005; Nickel & Achtziger, 2005).
Although the Needingworth community was dominated by species typical of disturbed grasslands, there were also a number of more unusual species, suggesting the site is potentially valuable for grassland fauna. Five Hemiptera, three Delphacidae and two Cicadellidae sampled have a UK status of local or greater conservation interest. E. variegatus is described as very local (Kirby, 1992; Stewart, 2012). Although it has mainly been found in southern Britain and in East Anglia, and has been associated with the Breckland region of Norfolk and Suffolk, there are several records of it to the south-west of Cambridge and nearby at Wicken Fen. P. striatus, a very rare and local species that is classed as insufficiently known (RDB K), has only been found in two UK ten-kilometre squares, both within Breckland (NBN Gateway, 2014). Its presence at Needingworth, extends its known UK distribution to a third such square. Ribautodelphax imitans, also RDB K, has been listed as a Biodiversity Action Plan (BAP) species of insect that had been found on only four sites in the UK, and only at two of these in the last 35 years (JNCC, 2010). However more recently it

9
has been found in the Cambridge area, at Coe Fen near the centre of the city, where it is the focus of an ongoing research project by Anglia Ruskin University (Helden et al., 2012).
Given the presence of these very interesting species, further sampling may yet reveal the presence of other undiscovered species with similar conservation status. It is difficult to know how many species might be added to the 81 found at Needingworth so far. The species accumulation curve suggests that there are plenty more to find. The species richness estimators suggest a further 32 may be present. However the number may be higher than this, as the estimates themselves have a tendency to increase with sampling intensity (Chao et al., 2009; Magurran, 2004).
The number of Hemiptera species per sample site showed no difference between the 2003 and 2011 restored areas of Needingworth. Similarly neither the overall abundance, nor that of four of the five main orders of grassland invertebrates differed significantly. This suggests that many aspects of the invertebrate community have become established within a few years of restoration. However this apparently rapid and successful colonisation was not uniform, as the number of Diptera were significantly higher in the 2011 restoration and the reverse was true of the other orders. As neither group has been identified further, the reasons for this must largely remain conjecture at present. From initial, non-quantitative examination of the other orders, the difference may be related to detritivores, such as woodlice. Their populations may be slower to establish after restoration, due to limited dispersal and lack of a dead organic matter in younger restoration areas. It is hoped that further examination of the samples will be possible, which would help to investigate this further. Due to their high vagility Diptera may move widely within the area and their numbers in any given area may be affected by factors outside the immediate habitat area concerned. For example many Diptera are associated with wetland habitats and high numbers in the 2011 restoration may be related to the characteristics of the water bodies.
Another issue that ought to be taken into consideration regarding the comparison of data from the two ages of restoration, is that of experimental design. As all 2003 restoration sites were spatially separated from all 2011 there is a possibility of spatial autocorrelation and so samples within each area are nested; in effect pseudoreplicates. Without replication of restoration age this is impossible to avoid at a site when there is just one area within each category. Therefore in such circumstances it must always be remembered that any differences may be affected be a genuine treatment effect (age since restoration) or the spatial separation. However this problem is probably less important an issue given the lack of significant treatment effects found at Needingworth and that the whole site has been restored from a more or less zero biodiversity baseline.
Nestedness of treatments was not an issue with the grazed-ungrazed comparisons within the 2003 restoration area. Modelling indicated that Hemiptera species richness, and the abundance of all invertebrates was significantly higher in ungrazed areas, despite the grazing intensity being relatively low (0.85 cattle.ha-1) (Dumont et al., 2007; Kruess & Tscharntke, 2002). This was also true of all orders apart from the Diptera. As with the previous comparisons their vagility and the potential influence of wetland habitats may have resulted in the contrasting pattern in Diptera.
Reductions in invertebrate biodiversity due to grazing have been widely reported in the literature (Morris, 1969, 1971, 1973; Sheridan et al., 2008). Ungrazed grasslands, whether large-scale or in small patches, such as grass islets (Helden et al., 2010) are less disturbed and typically have longer vegetation, with greater niche diversity, reduced temperature variation and increased shelter compared to grazed areas (Morris, 2000). Based on this, ungrazed grasslands have been set up along field

10
margins and water courses to increase biodiversity in agricultural systems (Anderson et al., 2013; Sheridan et al., 2008; Woodcock et al., 2007). In effect the same has occurred at Needingworth, with the areas fenced off from grazing facilitating an increase in invertebrate populations. At present, in the 2003 restoration area, the ungrazed areas account for 25.9% of the grassland. If this were doubled to 51.9%, our data suggest a 14.9% increase in overall invertebrate numbers. This would be beneficial not only for the invertebrates themselves but also enhance their contribution to ecosystem processes, being food for other organisms, including birds, and in their role as pollinators and decomposers. Larger invertebrate numbers may help to increase the stability of populations of rare species and so enhance the conservation status of the grasslands. Of course these benefits may have to balanced against losses of grazing area, which may reduce feeding opportunities for birds such as Anas penelope (wigeon) and several species of geese (Vickery & Gill, 1999), and may result in a reduction in income from renting out grazing rights. Whether an expansion of ungrazed areas is desirable will depend of the relative value of these alternatives and is a matter of judgement for land managers.
No areas have been fenced off in the 2011 restoration area, as at present the grasslands there are not grazed. However it is believed that grazing will be used as a management tool in future. Given the results of this study, we recommend that areas are fenced off and allowed to remain ungrazed. Eventually without any intervention any such area will go through a successional sequence as scrub develops and the area eventually becomes woodland. It is believed this was the original plan for the ungrazed areas in the 2003 restoration. However retaining some ungrazed grassland on a permanent basis would also be beneficial to biodiversity and could be achieved by strategies such as rotational management to prevent succession (Morris, 2000). Furthermore as indicated in indicated by a number of authors, a diversity of management approaches within the grasslands of Needingworth would most likely be the optimal strategy for maintaining grassland invertebrate communities on the site (Biedermann et al., 2005; Blake et al., 2011; Morris, 2000; Nickel & Achtziger, 2005).
Conclusions • Needingworth, as with many restored grasslands has a Hemiptera fauna
dominated by species typical of ruderal and human impacted sites. However also present are several rare and poorly known species, and others may be found with further sampling effort. This suggests the site has potential for a less eurytopic/pioneer dominated community and has specific value for the conservation of some rare Hemiptera. Management of the site should take into account the requirements of invertebrate communities.
• Age since restoration (11 years or 4 years) had little effect on invertebrates generally of Hemiptera more specifically, although there were large differences in decomposers. This indicates that invertebrate communities can quite rapidly establish following restoration of ex-gravel extraction sites, although the community structure may be limited largely to eurytopic/pioneer species.
• Populations of invertebrates were higher in ungrazed areas than grazed grassland. Setting aside ungrazed areas has been effective in enhancing invertebrate populations at Needingworth. This should be continued as part of wider strategy to provide a range of grassland management to enhance invertebrate biodiversity, as new areas of site are restored after gravel extraction.

11
Acknowledgements This project was carried out as part of the Quarry Life Award. We thank Hanson/Heidelberg Cement and the RSPB for the opportunity and permission to sample invertebrates at the site. Fieldwork, invertebrate sorting and initial identification was carried out with the help of James Chipps, Luiza Becker Pereira and the following members of the Anglia Ruskin University Wildlife Society: Steve Allain, Claire Cable, Mark Goodman, Toni Mott, David Pereira and Liam Smith. The work was supported by an Undergraduate Research Taster Placement from the Faculty of Science & Technology, Anglia Ruskin University, awarded to James Chipps.
References
Aggregates Business Europe (2010) Restoration at sand and gravel quarry. http://www.aggbusiness.com/sections/health-safety-environment/features/restoration-at-sand-and-gravel-quarry/ Accessed 19.9.14.
Anderson, A., Carnus, T., Helden, A.J., Sheridan, H., & Purvis, G. (2013) The influence of conservation field margins in intensively managed grazing land on communities of five arthropod trophic groups. Insect Conservation and Diversity, 6, 201-211.
Arnold, A.J. (1994) Insect suction sampling without nets, bags or filters. Crop Protection, 13, 73-76.
Bates, D., Maechler, M., & Bolker, B. (2012) lme4: Linear mixed-effects models using S4 classes. R package version 0.999999-0. http://CRAN.R-project.org/package=lme4.
Biedermann, R., Achtziger, R., Nickel, H., & Stewart, A.J.A. (2005) Conservation of grassland leafhoppers: a brief review. Journal of Insect Conservation, 9, 229-243.
Biedermann, R. & Niedringhaus, R. (2009) The Plant- and Leafhoppers of Germany. Identification key to all species. WABV - Fründ, Scheeßel.
Blackman, R.L. (2010) Aphids - Aphidinae (Macrosiphini). Handbooks for the Identification of British Insects 2 (7). Royal Entomological Society of London.
Blake, R.J.B.R.J., Woodcock, B.A., Ramsay, A.J., Pilgrim, E.S., Brown, V.K., Tallowin, J.R., & Potts, S.G. (2011) Novel margin management to enhance Auchenorrhyncha biodiversity in intensive grasslands. Agriculture Ecosystems & Environment, 140, 506-513.
Brook, A.J., Woodcock, B.A., Sinka, M., & Vanbergen, A.J. (2008) Experimental verification of suction sampler capture efficiency in grasslands of differing vegetation height and structure. Journal of Applied Ecology, 45, 1357-1363.
Chao, A., Colwell, R.K., Lin, C.W., & Gotelli, N.J. (2009) Sufficient sampling for asymptotic minimum species richness estimators. Ecology, 90, 1125-1133.
Daft Logic (2014) Google Maps Area Calculator Tool. http://www.daftlogic.com/projects-google-maps-area-calculator-tool.htm Accessed 19.9.14.
della Giustina, W. (1989) Faune de France. France et Iles Anglo-Normandes 73. Homoptères Cicadellidae Volume 3. Compléments aux ouvrages d'Henri Ribaut. Fédération Française des Sociétés de Sciences Naturelles., Paris.
Dumont, B., Rook, A.J., Coran, C., & Rover, K.U. (2007) Effects of livestock breed and grazing intensity on biodiversity and production in grazing systems. 2. Diet selection. Grass and Forage Science, 62, 159-171.
Hawkins, R.D. (2003) Sheildbugs of Surrey. Surrey Wildlife Trust, Woking.

12
Helden, A.J., Anderson, A., Sheridan, H., & Purvis, G. (2010) The role of grassland sward islets in the distribution of arthropods in cattle pastures. Insect Conservation and Diversity, 3, 291-301.
Helden, A.J., Dittrich, A., & Mckenzie, R. (2011). The status and distribution of Ribautodelphax imitans (Ribaut) at Coe Fen, Cambridge, with detailed of the wider Auchenorrhyncha community. (Unpublished report).
Helden, A.J., Dittrich, A., & Mckenzie, R. (2012) Ribautodelphax imitans (Ribaut) at Coe Fen, Cambridge. LEDRA. Newsletter of the Auchenorrhyncha Recording Scheme for Britain & Ireland, 5.
Hodkinson, I.D. & White, I.M. (1979) Homoptera Psylloidea. Handbooks for the Identification of British Insects 2 (5a). Royal Entomological Society of London.
JNCC (2010) Ribautodelphax imitans (Ribaut, 1953). Tall fescue planthopper. In UK priority species pages - Version 2. Joint Nature Conservation Committee.
Kirby, P. (1992) A review of the scarce and threatened Hemiptera of Great Britain. The Joint Nature Conservation Committee, Peterborough, UK.
Kruess, A. & Tscharntke, T. (2002) Contrasting responses of plant and insect diversity to variation in grazing intensity. Biological Conservation, 106, 293-302.
Le Quesne, W.J. (1960) Hemiptera Fulgoromorpha. Handbooks for the Identification of British Insects 2 (3). Royal Entomological Society of London.
Le Quesne, W.J. (1965) Hemiptera Cicadomorpha (excluding Deltocephalinae and Typhlocybinae). Handbooks for the Identification of British Insects 2 (2a). Royal Entomological Society of London.
Le Quesne, W.J. (1969) Hemiptera Cicadomorpha Deltocephalinae. Handbooks for the Identification of British Insects 2 (2b). Royal Entomological Society of London.
Le Quesne, W.J. & Payne, K.R. (1981) Cicadellidae (Typhlocybinae) with a check list of the British Auchenorhyncha (Hemiptera, Homoptera). Handbooks for the Identification of British Insects 2 (2c). Royal Entomological Society of London.
Magurran, A.E. (2004) Measuring Biological Diversity. Blackwell Publishing, Oxford, UK.
Morris, M.G. (1969) Differences between the invertebrate faunas of grazed and ungrazed chalk grassland. III. The Heteropterous fauna. Journal of Applied Ecology, 6, 475-487.
Morris, M.G. (1971) Differences between the invertebrate faunas of grazed and ungrazed chalk grassland. IV. Abundance and diversity of Homoptera-Auchenorhyncha. Journal of Applied Ecology, 8, 37-52.
Morris, M.G. (1973) The effects of seasonal grazing on the Heteroptera and Auchenorhyncha (Hemiptera) of chalk grassland. Journal of Applied Ecology, 10, 761-780.
Morris, M.G. (2000) The effects of structure and its dynamics on the ecology and conservation of arthropods in British grasslands. Biological Conservation, 95, 129-142.
NBN Gateway (2014). https://data.nbn.org.uk Accessed 19.9.14. Nickel, H. (2003) The leafhoppers and planthoppers of Germany (Hemiptera,
Auchenorrhyncha): Patterns and strategies in a highly diverse group of phytophagous insects Pensoft, Sofia-Moscow.
Nickel, H. & Achtziger, R. (2005) Do they ever come back? Responses of leafhopper communities to extensification of land use. Journal of Insect Conservation, 9, 319-333.
Nickel, H. & Hildebrandt, J. (2003) Auchenorrhyncha communities as indicators of disturbance in grasslands (Insecta, Hemiptera) - a case study from the Elbe flood plains (northern Germany). Agriculture, Ecosystems and Environment, 98, 183-199.

13
Oksanen, J., Guillaume Blanchet, F., Kindt, R., Legendre, P., Minchi, P.R., O'Hara, R., Simpson, G.L., Solymos, P., Stevens, M.H.H., & Wagner, H. (2012) vegan: Community Ecology Package. R package version 2.0-4.http://CRAN.R-project.org/package=vegan.
Ordnance Survey (2014) Get a Map. http://www.getamap.ordnancesurveyleisure.co.uk/ Accessed 19.9.14.
R Core Team (2012) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL http://www.R-project.org/.
Ribaut, H. (1952) Faune de France 57. Homoptères Auchénothynques. II (Jassidae) Fédération Française des Sociétés de Sciences Naturelles., Paris.
RSPB (2011) Natural England Fund transforms Quarry into Wildlife Haven. http://www.rspb.org.uk/media/releases/details.aspx?id=279548 Accessed 19.9.14.
Sheridan, H., Finn, J.A., Culleton, N., & O'Donovan, G. (2008) Plant and invertebrate diversity in grassland field margins. Agriculture Ecosystems & Environment, 123, 225-232.
Stewart, A.J.A. (2012) Checklist of Auchenorrhyncha. http://www.ledra.co.uk/species.html Accessed 19.9.14.
Stöckman, M., Biedermann, R., Nickel, H., & Niedringhaus, R. (2013) The Nymphs of the Planthoppers and Leafhoppers of Germany. WABV - Fründ, Scheeßel.
Stroyan, H.L.G. (1977) Homoptera Aphidoidea (Part). Chaitophoridae & Callaphididae. Handbooks for the Identification of British Insects 2 (4a). Royal Entomological Society of London.
Taylor, L.R., Palmer, J.M.P., Dupuch, M.J., Cole, J., & Taylor, M.S. (1984) A handbook for the rapid identification of alate aphids of Great Britain and Europe. Rothamsted Experimental Station, Harpenden, England.
Vickery, J.A. & Gill, J.A. (1999) Managing grassland for wild geese in Britain: a review. Biological Conservation, 89, 93-106.
Woodcock, B.A., Potts, S.G., Pilgrim, E., Ramsay, A.J., Tscheulin, T., Parkinson, A., Smith, R.E.N., Gundrey, A.L., Brown, V.K., & Tallowin, J.R. (2007) The potential of grass field margin management for enhancing beetle diversity in intensive livestock farms. Journal of Applied Ecology, 44, 60-69.
14
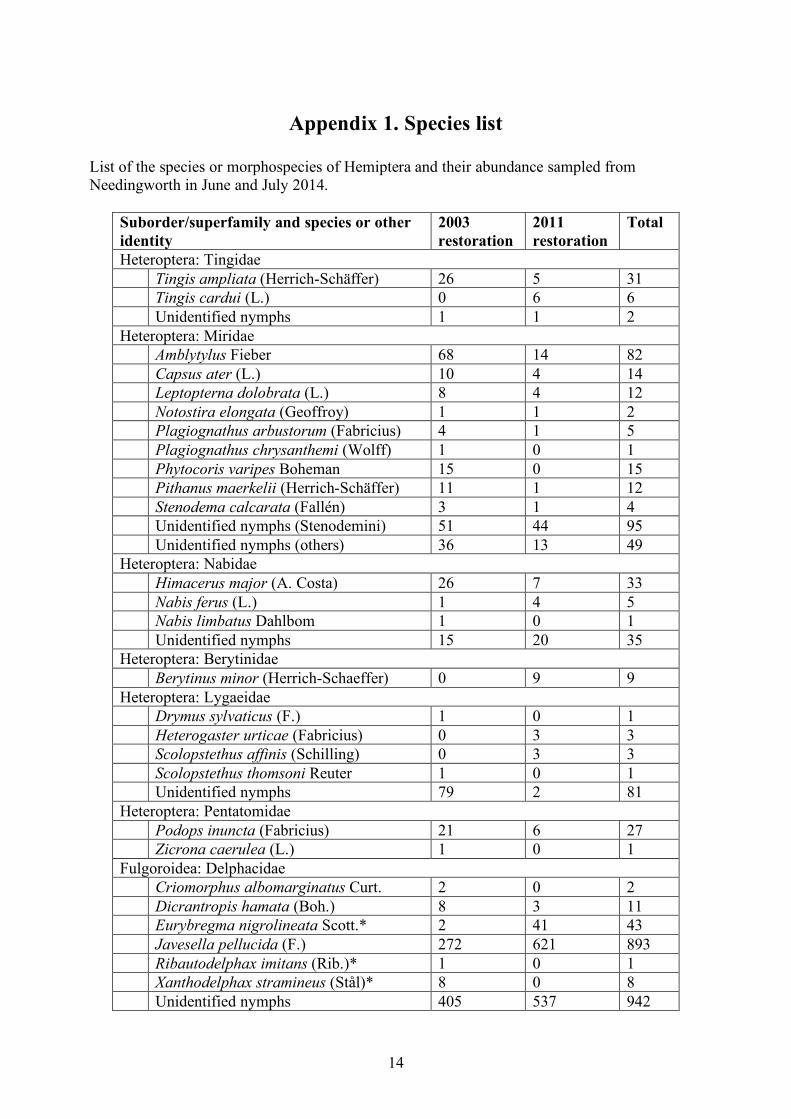
Appendix 1. Species list List of the species or morphospecies of Hemiptera and their abundance sampled from Needingworth in June and July 2014.
Suborder/superfamily and species or other identity
2003 restoration
2011 restoration
Total
Heteroptera: Tingidae Tingis ampliata (Herrich-Schäffer) 26 5 31 Tingis cardui (L.) 0 6 6 Unidentified nymphs 1 1 2 Heteroptera: Miridae Amblytylus Fieber 68 14 82 Capsus ater (L.) 10 4 14 Leptopterna dolobrata (L.) 8 4 12 Notostira elongata (Geoffroy) 1 1 2 Plagiognathus arbustorum (Fabricius) 4 1 5 Plagiognathus chrysanthemi (Wolff) 1 0 1 Phytocoris varipes Boheman 15 0 15 Pithanus maerkelii (Herrich-Schäffer) 11 1 12 Stenodema calcarata (Fallén) 3 1 4 Unidentified nymphs (Stenodemini) 51 44 95 Unidentified nymphs (others) 36 13 49 Heteroptera: Nabidae Himacerus major (A. Costa) 26 7 33 Nabis ferus (L.) 1 4 5 Nabis limbatus Dahlbom 1 0 1 Unidentified nymphs 15 20 35 Heteroptera: Berytinidae Berytinus minor (Herrich-Schaeffer) 0 9 9 Heteroptera: Lygaeidae Drymus sylvaticus (F.) 1 0 1 Heterogaster urticae (Fabricius) 0 3 3 Scolopstethus affinis (Schilling) 0 3 3 Scolopstethus thomsoni Reuter 1 0 1 Unidentified nymphs 79 2 81 Heteroptera: Pentatomidae Podops inuncta (Fabricius) 21 6 27 Zicrona caerulea (L.) 1 0 1 Fulgoroidea: Delphacidae Criomorphus albomarginatus Curt. 2 0 2 Dicrantropis hamata (Boh.) 8 3 11 Eurybregma nigrolineata Scott.* 2 41 43 Javesella pellucida (F.) 272 621 893 Ribautodelphax imitans (Rib.)* 1 0 1 Xanthodelphax stramineus (Stål)* 8 0 8 Unidentified nymphs 405 537 942
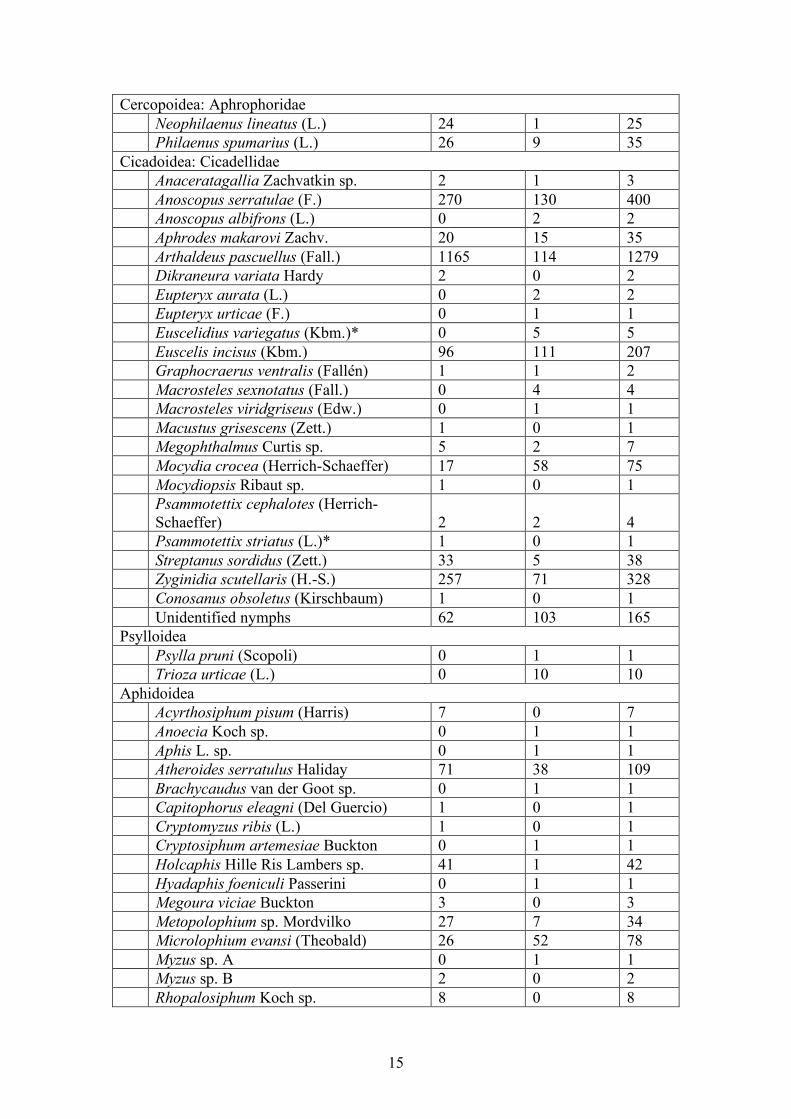
15
Cercopoidea: Aphrophoridae Neophilaenus lineatus (L.) 24 1 25 Philaenus spumarius (L.) 26 9 35 Cicadoidea: Cicadellidae Anaceratagallia Zachvatkin sp. 2 1 3 Anoscopus serratulae (F.) 270 130 400 Anoscopus albifrons (L.) 0 2 2 Aphrodes makarovi Zachv. 20 15 35 Arthaldeus pascuellus (Fall.) 1165 114 1279 Dikraneura variata Hardy 2 0 2 Eupteryx aurata (L.) 0 2 2 Eupteryx urticae (F.) 0 1 1 Euscelidius variegatus (Kbm.)* 0 5 5 Euscelis incisus (Kbm.) 96 111 207 Graphocraerus ventralis (Fallén) 1 1 2 Macrosteles sexnotatus (Fall.) 0 4 4 Macrosteles viridgriseus (Edw.) 0 1 1 Macustus grisescens (Zett.) 1 0 1 Megophthalmus Curtis sp. 5 2 7 Mocydia crocea (Herrich-Schaeffer) 17 58 75 Mocydiopsis Ribaut sp. 1 0 1 Psammotettix cephalotes (Herrich-
Schaeffer) 2 2 4 Psammotettix striatus (L.)* 1 0 1 Streptanus sordidus (Zett.) 33 5 38 Zyginidia scutellaris (H.-S.) 257 71 328 Conosanus obsoletus (Kirschbaum) 1 0 1 Unidentified nymphs 62 103 165 Psylloidea Psylla pruni (Scopoli) 0 1 1 Trioza urticae (L.) 0 10 10 Aphidoidea Acyrthosiphum pisum (Harris) 7 0 7 Anoecia Koch sp. 0 1 1 Aphis L. sp. 0 1 1 Atheroides serratulus Haliday 71 38 109 Brachycaudus van der Goot sp. 0 1 1 Capitophorus eleagni (Del Guercio) 1 0 1 Cryptomyzus ribis (L.) 1 0 1 Cryptosiphum artemesiae Buckton 0 1 1 Holcaphis Hille Ris Lambers sp. 41 1 42 Hyadaphis foeniculi Passerini 0 1 1 Megoura viciae Buckton 3 0 3 Metopolophium sp. Mordvilko 27 7 34 Microlophium evansi (Theobald) 26 52 78 Myzus sp. A 0 1 1 Myzus sp. B 2 0 2 Rhopalosiphum Koch sp. 8 0 8

16
Sipha Passerini sp. 0 1 1 Sitobion fragariae (Walker) 34 14 48 Tetraneura ulmi (Linnaeus) 0 2 2 Uroleucon Mordvilko sp. 183 67 250 Uromelan (Mordvilko) sp. 19 0 19 Aphid sp. A 2 30 32 Aphid sp. B 9 15 24 Aphid sp. C 0 1 1 Aphid sp. D 1 4 5 Aphid sp. E 0 1 1 Unidentified nymphs 138 81 219 Coccoidea Pseudococcidae 1 2 3 Total 3649 2338 5987

17
Appendix 2. Maps & photographs
Fig. 1. View of Needingworth, showing open water and reedbed surrounded by grassland.

18
Fig. 2. Map of Needingworth, showing habitat types and the locations of invertebrate sampling.

19
Fig. 3. Grassland at Needingworth. The fence separates ungrazed (left) and cattle-grazed areas (right).
Fig. 4. Sampling invertebrates at Needingworth, using a Vortis suction sampler.

20
Appendix 3. Graphs
Fig. 5. Species accumulation curve for Hemiptera at Needingworth, in June and July 2014. The shaded area represents 95% confidence intervals of the curve.
Fig. 6. Boxplots showing the overall abundance of grassland invertebrates in grazed and ungrazed grassland at Needingworth. Dark horizontal lines show the median, with the upper and lower boxes the 25th and 75th percentiles, respectively. The dashed lines indicate either 1.5 times the interquartile range or the maximum and minimum values if there are no outliers (small circles).

21
Fig. 7. Boxplots showing the abundance of grassland invertebrates of different taxonomic order, in grazed and ungrazed grassland at Needingworth.
Fig. 8. Boxplots of the number of species of Hemiptera recorded in grazed and ungrazed grassland at Needingworth.

22
Fig. 9. Model of the possible total number of grassland invertebrates, of all orders, calculated to be present in the 2003 restoration area of Needingworth, if different areas of ungrazed grassland were present. The model was based on density estimates and habitat areas. The regression line shows the model: y(individuals) = 2.25x107x(area) + 1.07x109.
Fig. 10. Boxplots of the number of grassland invertebrates of all orders, sampled in the older (R2003 – 2003 restoration) and newer (R2011 – 2011 restoration) areas of Needingworth.

23
Fig. 11. Boxplots of the number of grassland invertebrates of different taxonomic order, sampled in the older (R2003 – 2003 restoration) and newer (R2011 – 2011 restoration) areas of Needingworth.