inverse methods in systems biology · inverse methods in systems biology ... inverse bifurcation:...
TRANSCRIPT

24th TBI Winterseminar, Bled 2009
Inverse Methods in Systems Biology
James Lu
Johann Radon Institute for Computational and Applied Mathematics (RICAM)
Austrian Academy of Sciences
James Lu Inverse Methods in Systems Biology

Systems Biology
◮ Cells are complex systems
James Lu Inverse Methods in Systems Biology

Systems Biology
◮ Cells are complex systems
◮ In the face of continuously changing environments and itsstate, cells need to respond appropriately:
James Lu Inverse Methods in Systems Biology

Systems Biology
◮ Cells are complex systems
◮ In the face of continuously changing environments and itsstate, cells need to respond appropriately:
◮ inputs: nutrients, repellants, heat shock, DNA damage, ...
◮ responses: movement, growth, protein production, death
James Lu Inverse Methods in Systems Biology

Systems Biology
◮ Complex networks of genes and pathways are involved inmediating the various inputs to the appropriate responses
Signal transduction network: Hanahan and Weinberg, Cell (2000)
James Lu Inverse Methods in Systems Biology

Forward Problems
◮ Consider ODE models: y(t) = f (y(t),q)
James Lu Inverse Methods in Systems Biology

Forward Problems
◮ Consider ODE models: y(t) = f (y(t),q)
◮ The Forward analysis may include:◮ numerical integration for a set of given parameters◮ sensitivity analysis◮ bifurcation analysis
reactions→ dydt = f (y,q)→ {y(t), bifurcations}
5000 10000 15000 20000
0.2
0.4
0.6
0.8
1
1.2
Signal S = 0.095
A
5000 10000 15000 20000
0.2
0.4
0.6
0.8
1
1.2
Signal S = 1
A
5000 10000 15000 20000
0.2
0.4
0.6
0.8
1
1.2
Signal S = 0.01
A
5000 10000 15000 20000
0.2
0.4
0.6
0.8
1
1.2
Signal S = 0.087
A
0.05 0.06 0.07 0.08 0.09 0.1 0.110
0.2
0.4
0.6
0.8
1
1.2
H
LP
LP
Continuation parameter tG‘S
Sta
te c
ompo
nent
tG‘A
[t]
LPC
LPC
James Lu Inverse Methods in Systems Biology

Inverse Problems
◮ In Inverse Problems, one looks for the causes of observedand desired effects
reactions← dydt = f (y,q)← {y(t), bifurcations}
James Lu Inverse Methods in Systems Biology

Inverse Problems
◮ In Inverse Problems, one looks for the causes of observedand desired effects
reactions← dydt = f (y,q)← {y(t), bifurcations}
◮ Parameter identification from time-course data
James Lu Inverse Methods in Systems Biology

Inverse Problems
◮ In Inverse Problems, one looks for the causes of observedand desired effects
reactions← dydt = f (y,q)← {y(t), bifurcations}
◮ Parameter identification from time-course data◮ Inverse bifurcation: infer model mechanisms that can
achieve the desired bifurcation behaviors
◮ maximize the parameter domain for oscillations◮ place bifurcation points at the desired locations
James Lu Inverse Methods in Systems Biology

Inverse Problems
◮ In Inverse Problems, one looks for the causes of observedand desired effects
reactions← dydt = f (y,q)← {y(t), bifurcations}
◮ Parameter identification from time-course data◮ Inverse bifurcation: infer model mechanisms that can
achieve the desired bifurcation behaviors
◮ maximize the parameter domain for oscillations◮ place bifurcation points at the desired locations
◮ Inverse problems are typically ill-posed :
◮ solutions may not exist or unique◮ the solution may not depend on the data in a continuous
manner
James Lu Inverse Methods in Systems Biology

Inverse Problems
◮ In Inverse Problems, one looks for the causes of observedand desired effects
reactions← dydt = f (y,q)← {y(t), bifurcations}
◮ Parameter identification from time-course data◮ Inverse bifurcation: infer model mechanisms that can
achieve the desired bifurcation behaviors
◮ maximize the parameter domain for oscillations◮ place bifurcation points at the desired locations
◮ Inverse problems are typically ill-posed :
◮ solutions may not exist or unique◮ the solution may not depend on the data in a continuous
manner
◮ To numerically tackle inverse problems, regularizationstrategies are needed
James Lu Inverse Methods in Systems Biology

Inverse Problems
◮ Forward operator equation:
F(q) = y
James Lu Inverse Methods in Systems Biology

Inverse Problems
◮ Forward operator equation:
F(q) = y
◮ Inverse problem: determining q from y
James Lu Inverse Methods in Systems Biology

Inverse Problems
◮ Forward operator equation:
F(q) = y
◮ Inverse problem: determining q from y◮ Typically ill-posed (in the sense of Hadamard):
◮ non-uniqueness;◮ instability of inversion
James Lu Inverse Methods in Systems Biology

Inverse Problems
◮ Forward operator equation:
F(q) = y
◮ Inverse problem: determining q from y◮ Typically ill-posed (in the sense of Hadamard):
◮ non-uniqueness;◮ instability of inversion
◮ Variational regularization: add penalty term
minq‖F(q)− y‖+ µR(q)
James Lu Inverse Methods in Systems Biology

Inverse Problems
◮ Forward operator equation:
F(q) = y
◮ Inverse problem: determining q from y◮ Typically ill-posed (in the sense of Hadamard):
◮ non-uniqueness;◮ instability of inversion
◮ Variational regularization: add penalty term
minq‖F(q)− y‖+ µR(q)
◮ While stabilizing ill-posed problems, regularization bringsbias to the solution
James Lu Inverse Methods in Systems Biology

Inverse Problems
◮ Forward operator equation:
F(q) = y
◮ Inverse problem: determining q from y◮ Typically ill-posed (in the sense of Hadamard):
◮ non-uniqueness;◮ instability of inversion
◮ Variational regularization: add penalty term
minq‖F(q)− y‖+ µR(q)
◮ While stabilizing ill-posed problems, regularization bringsbias to the solution
◮ For biological problems, usually want to find solutions thatare sparse, i.e., having as few non-zero entries aspossible: Ockam’s razor
James Lu Inverse Methods in Systems Biology

Sparsity-Promoting Regularization
◮ As the regularization term, consider smoothed functionalsR
n→ R: lp,ε(q) = ∑i(q2i + ε)p/2
James Lu Inverse Methods in Systems Biology

Sparsity-Promoting Regularization
◮ As the regularization term, consider smoothed functionalsR
n→ R: lp,ε(q) = ∑i(q2i + ε)p/2
◮ Convex only within the box {q : |qi|<√
ε, 0 < i≤ n}
−0.2 −0.1 0 0.1 0.2 0.30
0.02
0.04
0.06
0.08
0.1
x
l 2(x)
−0.2 −0.1 0 0.1 0.2 0.30
0.1
0.2
x
l 0.1,
1e−
4(x)
James Lu Inverse Methods in Systems Biology

Sparsity-Promoting Regularization
◮ As the regularization term, consider smoothed functionalsR
n→ R: lp,ε(q) = ∑i(q2i + ε)p/2
◮ Convex only within the box {q : |qi|<√
ε, 0 < i≤ n}◮ Recent applications of sparse solutions using non-convex
penalty:◮ Exact reconstruction of sparse signals via nonconvex
minimization, R. Chartrand (2007)◮ Compressive sensing using l1 re-weighting, E. Candes, S.
P. Boyd, M. Wakin et al. (2007)◮ Log-det heuristic for matrix rank minimization with
applications to Hankel and Euclidean distance matrices, M.Fazel, H. Hindi and S. P Boyd (2003)
James Lu Inverse Methods in Systems Biology

Inverse Bifurcation: the G1/S module of cell cycle
M. Swat, A. Kel, H. Herzel, Bioinformatics (2004)
James Lu Inverse Methods in Systems Biology

Inverse Bifurcation: the G1/S module of cell cycle
0 0.5 1 1.5 2 2.5 3 3.5 4
0
2
4
6
8
10
12
14
SN1
SN2
TC
G0/G
1 phase
Fm
(x10−3)
E2F
1
S phase
(CyclinD/cdk4,6 activation threshold)
M. Swat, A. Kel, H. Herzel, Bioinformatics (2004)
James Lu Inverse Methods in Systems Biology

Inverse Bifurcation: the G1/S module of cell cycle
0 0.5 1 1.5 2 2.5 3 3.5 4
0
2
4
6
8
10
12
14
SN1
SN2
TC
G0/G
1 phase
Fm
(x10−3)
E2F
1
S phase
(CyclinD/cdk4,6 activation threshold)
M. Swat, A. Kel, H. Herzel, Bioinformatics (2004)
◮ Which of the interactions play important roles in controllingthe geometry of the bifurcation diagram?
James Lu Inverse Methods in Systems Biology

Inverse Bifurcation: the G1/S module of cell cycle
◮ Map various bifurcation phenotypes to parameter sets
James Lu Inverse Methods in Systems Biology

Inverse Bifurcation: the G1/S module of cell cycle
◮ Map various bifurcation phenotypes to parameter sets◮ Consider the following 3 modes of transformations of the
nominal bifurcation diagram:
James Lu Inverse Methods in Systems Biology

Inverse Bifurcation: the G1/S module of cell cycle
◮ Map various bifurcation phenotypes to parameter sets◮ Consider the following 3 modes of transformations of the
nominal bifurcation diagram:
James Lu Inverse Methods in Systems Biology

Inverse Bifurcation: the G1/S module of cell cycle
◮ Map various bifurcation phenotypes to parameter sets◮ Consider the following 3 modes of transformations of the
nominal bifurcation diagram:
James Lu Inverse Methods in Systems Biology
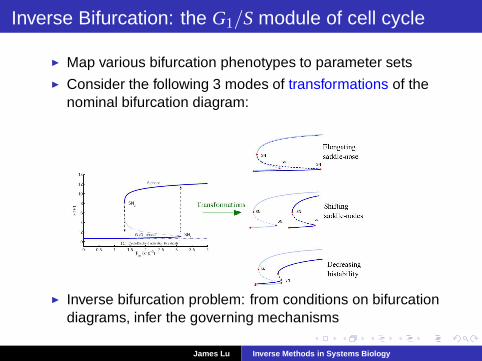
Inverse Bifurcation: the G1/S module of cell cycle
◮ Map various bifurcation phenotypes to parameter sets◮ Consider the following 3 modes of transformations of the
nominal bifurcation diagram:
◮ Inverse bifurcation problem: from conditions on bifurcationdiagrams, infer the governing mechanisms
James Lu Inverse Methods in Systems Biology

Inverse Bifurcation: effect of sparsity-promiting penalty
0 2 4 6 80
2
4
6
8
SN
SN
SN
Fm
E2F
1
Mammalian G1/S transition
−15 −10 −5 0
k43k67k76k23k28k89k98
aJ15J18J68J13J63
Km9phipRB
phiCycDiphiAp1
phipRBpphipRBppphiCycEi
phiCycEaJ65J62J61
Km1k61k16k25
Km4J11
k3k1
k34J12
Km2phiCycDa
phiE2F1k2kp
% change
Par
amet
er li
st
(a) l0.1,10−4 regularization
James Lu Inverse Methods in Systems Biology

Inverse Bifurcation: effect of sparsity-promiting penalty
0 2 4 6 80
2
4
6
8
SN
SN
SN
Fm
E2F
1
Mammalian G1/S transition
−15 −10 −5 0
k43k67k76k23k28k89k98
aJ15J18J68J13J63
Km9phipRB
phiCycDiphiAp1
phipRBpphipRBppphiCycEi
phiCycEaJ65J62J61
Km1k61k16k25
Km4J11k3k1
k34J12
Km2phiCycDa
phiE2F1k2kp
% change
Par
amet
er li
st
(b) l0.1,10−4 regularization
0 2 4 6 80
2
4
6
8
SN
SN
SN
Fm
E2F
1
Mammalian G1/S transition
−4 −3 −2 −1 0 1 2 3
k43k67k76k28k89k98
aJ15J18J68J13
Km9phipRB
phipRBppphiCycEi
phiCycEaJ63J61k23J65
Km1J62
phipRBpk61k16k25
Km4phiCycDi
k1J11k2
k34phiCycDa
k3J12
phiAp1phiE2F1
Km2kp
% change
Par
amet
er li
st
(c) l2 regularization
James Lu Inverse Methods in Systems Biology

Inverse Bifurcation: identified module
Table: Result of hierarchical algorithm with p = 0.1,ε = 10−4
Modification Case Level j = 1 Level j = 2 Level j = 3
Elongating SN1 nose kp ↓ 14.3% k34 ↑ 31.7% φAP-1 ↓ 20.9%Km2 ↑ 6.4% φE2F1 ↑ 7.3%
Moving SN1,2 to right Km4 ↑ 269.3% J11 ↑ 191.7% k2 ↓ 39.9%kp ↑ 17.3% φE2F1 ↓ 11.7%
Km2 ↓ 10.3%Decreasing bistabiliy J11 ↑ 128.5% k1 ↑ 169.1% k2 ↓ 43.7%
kp ↑ 33.8% Km2 ↓ 21.7% φE2F1 ↓ 28.3%J12 ↓ 20.1%
James Lu Inverse Methods in Systems Biology
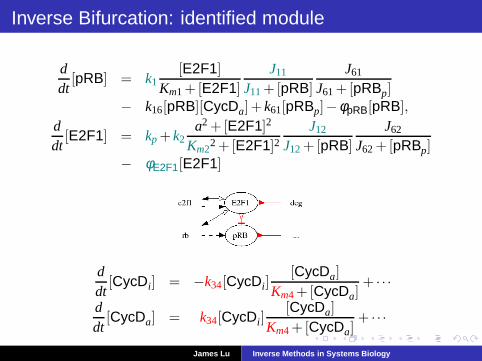
Inverse Bifurcation: identified module
ddt
[pRB] = k1[E2F1]
Km1+[E2F1]
J11
J11+[pRB]
J61
J61+[pRBp]− k16[pRB][CycDa]+ k61[pRBp]−φpRB[pRB],
ddt
[E2F1] = kp + k2a2 +[E2F1]2
Km22 +[E2F1]2
J12
J12+[pRB]
J62
J62+[pRBp]− φE2F1[E2F1]
ddt
[CycDi] = −k34[CycDi][CycDa]
Km4+[CycDa]+ · · ·
ddt
[CycDa] = k34[CycDi][CycDa]
Km4+[CycDa]+ · · ·
James Lu Inverse Methods in Systems Biology

Application: Circadian Rhythm Model
◮ Circadian rhythm underlies the 24 hr activity cycle of manyorganisms
James Lu Inverse Methods in Systems Biology

Application: Circadian Rhythm Model
◮ Circadian rhythm underlies the 24 hr activity cycle of manyorganisms
◮ Endogenous oscillator entrainable by 24 hr light-dark cycles
James Lu Inverse Methods in Systems Biology

Application: Circadian Rhythm Model
◮ Circadian rhythm underlies the 24 hr activity cycle of manyorganisms
◮ Endogenous oscillator entrainable by 24 hr light-dark cycles
◮ Circadian model for Arabidopsis thaliana proposed by Locke etal, Molecular Systems Biology (2005)
James Lu Inverse Methods in Systems Biology

Application: Circadian Rhythm Model
ODE system
James Lu Inverse Methods in Systems Biology

Application: Circadian Rhythm Model
◮ Simulation of model with nominal parameters under 12 hrlight-dark cycle
cLm
cYm
one23
20 40 60 80 100
0.2
0.4
0.6
0.8
1.0
1.2
James Lu Inverse Methods in Systems Biology

Application: Circadian Rhythm Model
◮ Evidence suggests GIGANTEA could be gene Y
Phase misfit with experimental data
J. CW Locke et al., Molecular Systems Biology (2005)
James Lu Inverse Methods in Systems Biology

Application: Circadian Rhythm Model
◮ Evidence suggests GIGANTEA could be gene Y
Phase misfit with experimental data
J. CW Locke et al., Molecular Systems Biology (2005)
James Lu Inverse Methods in Systems Biology

Application: Circadian Rhythm Model
◮ Evidence suggests GIGANTEA could be gene Y
Phase misfit with experimental data
J. CW Locke et al., Molecular Systems Biology (2005)
◮ Inverse problem: vary parameters so that Y mRNA peaksat ZT7 rather than ZT11
James Lu Inverse Methods in Systems Biology

Application: Circadian Rhythm Model
◮ The peak time t for the level of Y mRNA, satisfies
y(t) = f (y,q)fYmRNA(y(t),q) = 0
James Lu Inverse Methods in Systems Biology

Application: Circadian Rhythm Model
◮ The peak time t for the level of Y mRNA, satisfies
y(t) = f (y,q)fYmRNA(y(t),q) = 0
◮ Goal: obtain a solution that peaks at time t∗ by varying asfew parameters from the published values, q∗, as possible
James Lu Inverse Methods in Systems Biology

Application: Circadian Rhythm Model
◮ The peak time t for the level of Y mRNA, satisfies
y(t) = f (y,q)fYmRNA(y(t),q) = 0
◮ Goal: obtain a solution that peaks at time t∗ by varying asfew parameters from the published values, q∗, as possible
◮ Minimization of the objective:
‖t(q)− t∗‖2 + µ lp,ε (q−q∗)→minq
James Lu Inverse Methods in Systems Biology

Application: Circadian Rhythm Model
◮ Y mRNA solution profile for the original parameter set
0 10 20 30 40 50 60 70
0.0
0.1
0.2
0.3
0.4
Time HhrL
Y mRNA Level
James Lu Inverse Methods in Systems Biology

Application: Circadian Rhythm Model
◮ Y mRNA solution profile for the identified parameter set
0 10 20 30 40 50 60 70
0.0
0.1
0.2
0.3
0.4
Time HhrL
Y mRNA Level
James Lu Inverse Methods in Systems Biology

Application: Circadian Rhythm Model
◮ Y mRNA solution profile for the identified parameter set◮ 1 out of 54 parameters is identified
0 10 20 30 40 50 60 70
0.0
0.1
0.2
0.3
0.4
Time HhrL
Y mRNA Level
James Lu Inverse Methods in Systems Biology

Application: Circadian Rhythm Model
◮ Identified parameter: g6, Hill-constant in repression ofgene Y by LHY
James Lu Inverse Methods in Systems Biology

Inverse Eigenvalue Problems
◮ We have seen inverse bifurcation problems inferring theunderlying mechanisms controlling the qualitative aspectsof dynamics
James Lu Inverse Methods in Systems Biology

Inverse Eigenvalue Problems
◮ We have seen inverse bifurcation problems inferring theunderlying mechanisms controlling the qualitative aspectsof dynamics
◮ One may also wish to probe the possibility of differentclasses of behaviors; e.g., transition of a bistable switch toan oscillator
James Lu Inverse Methods in Systems Biology

Inverse Eigenvalue Problems
◮ We have seen inverse bifurcation problems inferring theunderlying mechanisms controlling the qualitative aspectsof dynamics
◮ One may also wish to probe the possibility of differentclasses of behaviors; e.g., transition of a bistable switch toan oscillator
◮ Infer mechanisms governing the spectrum of thedynamical system: inverse eigenvalue problems
James Lu Inverse Methods in Systems Biology

Inverse Eigenvalue Problems
◮ For the dynamical system y(t) = f (y,q), many bifurcationsof equilibrium correspond to various conditions oneigenvalues of df (y,q)
dy
James Lu Inverse Methods in Systems Biology

Inverse Eigenvalue Problems
◮ For the dynamical system y(t) = f (y,q), many bifurcationsof equilibrium correspond to various conditions oneigenvalues of df (y,q)
dy
◮ Inverse eigenvalue problems: identify the possible modelmechanisms bringing about the desired change in thespectrum
(d) Hopf (e) BT (f) Fold-Hopf (g) Hopf-Hopf
James Lu Inverse Methods in Systems Biology

Inverse Eigenvalue Problems
◮ Hybrid solution algorithm:◮ Lift-and-Project (LP)
James Lu Inverse Methods in Systems Biology

Inverse Eigenvalue Problems
◮ Hybrid solution algorithm:◮ Lift-and-Project (LP)◮ Quasi-Newton (QN)
James Lu Inverse Methods in Systems Biology

Inverse Eigenvalue Problems
◮ Hybrid solution algorithm:◮ Lift-and-Project (LP)◮ Quasi-Newton (QN)
◮ Least square formulations with sparsity regularization:
LP : J(q) = ‖A(q)−Aproj‖2F + µ lp,ε(q−q∗)QN : J(q) = ∑
i
|λi(q)−λ di |2 + µ lp,ε(q−q∗)
James Lu Inverse Methods in Systems Biology

Inverse Eigenvalue Problems
◮ Hybrid solution algorithm:◮ Lift-and-Project (LP)◮ Quasi-Newton (QN)
◮ Least square formulations with sparsity regularization:
LP : J(q) = ‖A(q)−Aproj‖2F + µ lp,ε(q−q∗)QN : J(q) = ∑
i
|λi(q)−λ di |2 + µ lp,ε(q−q∗)
James Lu Inverse Methods in Systems Biology

Transition of a Bistable Switch to an Oscillator
◮ Consider a model for GATA transcription factors
James Lu Inverse Methods in Systems Biology

Transition of a Bistable Switch to an Oscillator
◮ Consider a model for GATA transcription factors◮ Scenario: duplication of a gene A→ A1, A2
James Lu Inverse Methods in Systems Biology
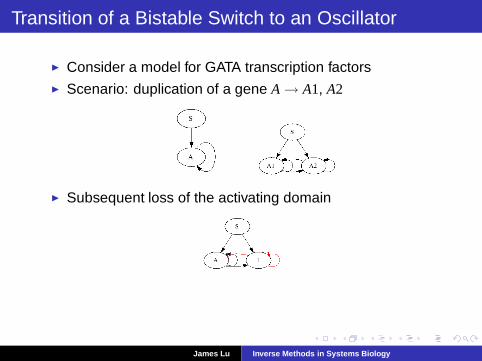
Transition of a Bistable Switch to an Oscillator
◮ Consider a model for GATA transcription factors◮ Scenario: duplication of a gene A→ A1, A2
◮ Subsequent loss of the activating domain
James Lu Inverse Methods in Systems Biology

Transition of a Bistable Switch to an Oscillator
◮ Consider a model for GATA transcription factors◮ Scenario: duplication of a gene A→ A1, A2
◮ Subsequent loss of the activating domain
◮ Can oscillations emerge via a few additional mutations?
James Lu Inverse Methods in Systems Biology

Transition of a Bistable Switch to an Oscillator
◮ Evolutionary scenario
James Lu Inverse Methods in Systems Biology

Transition of a Bistable Switch to an Oscillator
◮ Evolutionary scenario
2000 4000 6000 8000 10000
0.25
0.5
0.75
1
1.25
1.5
1.75
Solution for initial parameters
rMrA2rA1mMmA2mA1MA2cA2A1cA1
2000 4000 6000 8000 10000
0.5
1
1.5
2
2.5
3
3.5
Solution for identified parameters
rMrA2rA1mMmA2mA1MA2cA2A1cA1
James Lu Inverse Methods in Systems Biology

Transition of a Bistable Switch to an Oscillator
◮ Evolutionary scenario
2000 4000 6000 8000 10000
0.25
0.5
0.75
1
1.25
1.5
1.75
Solution for initial parameters
rMrA2rA1mMmA2mA1MA2cA2A1cA1
2000 4000 6000 8000 10000
0.5
1
1.5
2
2.5
3
3.5
Solution for identified parameters
rMrA2rA1mMmA2mA1MA2cA2A1cA1
◮ Identified reactions:
James Lu Inverse Methods in Systems Biology

Conclusions
◮ Many inverse problems arise from the modelling andanalysis of biological systems
James Lu Inverse Methods in Systems Biology

Conclusions
◮ Many inverse problems arise from the modelling andanalysis of biological systems
◮ inverse bifurcation problems: reverse engineering,matching model to data
James Lu Inverse Methods in Systems Biology

Conclusions
◮ Many inverse problems arise from the modelling andanalysis of biological systems
◮ inverse bifurcation problems: reverse engineering,matching model to data
◮ inverse eigenvalue problems: model-building, exploration ofpossible behaviors
James Lu Inverse Methods in Systems Biology

Conclusions
◮ Many inverse problems arise from the modelling andanalysis of biological systems
◮ inverse bifurcation problems: reverse engineering,matching model to data
◮ inverse eigenvalue problems: model-building, exploration ofpossible behaviors
◮ Sparsity-promoting regularization can be effective indrawing useful insights from biological models
James Lu Inverse Methods in Systems Biology

Acknowledgements
CollaboratorsRICAM: Heinz Engl, Philipp Kuegler, Stefan Mueller, Clemens ZarzerUniversity of Vienna: Peter Schuster, Christoph Flamm, Rainer MachneKeio University : Douglas Murray
Funding provided by Vienna Science and Technology Fund (WWTF) isgratefully acknowledged
Project: MA05, MA07
James Lu Inverse Methods in Systems Biology