intracellular hiv tat expression in astrocytes promotes astrocyte
TRANSCRIPT

HIV-Tat expression in astrocytes induces neurotoxicity
INTRACELLULAR HUMAN IMMUNODEFICIENCY VIRUS TAT EXPRESSION IN ASTROCYTES PROMOTES
ASTROCYTE SURVIVAL BUT INDUCES POTENT NEUROTOXICITY AT DISTANT SITES VIA AXONAL
TRANSPORT
Ashok Chauhan1, Jadwiga Turchan1, Chava Pocernich2, Anna Bruce-Keller3, Susan Roth4, D.Allan Butterfield2,
Eugene O. Major5, Avindra Nath1
Departments of Neurology1, Johns Hopkins University, Departments of Chemistry2, Anatomy and
Neurobiology3 and Microbiology and Immunology4, University of Kentucky, Lexington, KY, Laboratory of
Molecular Medicine and Neuroscience5, NINDS, NIH, Bethesda, MD
Running Title: HIV-Tat expression in astrocytes induces neurotoxicity
Correspondence to:
A. Nath, Department of Neurology, Johns Hopkins University, Pathology 509, 600 N Wolfe St, Baltimore, MD,
21287. Tele: 443-287-4657; Fax: 410-614-1008. e-mail: [email protected]
1
Copyright 2003 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on January 24, 2003 as Manuscript M209381200 by guest on February 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
Summary
The HIV-Tat protein has been implicated in the neuropathogenesis of HIV infection. However, its role in
modulating astroglial-neuronal relationships is poorly understood. Astrocyte infection with HIV has been
associated with rapid progression of dementia. We thus initially transfected astrocytes with HIV proviral DNA and
confirmed Tat production in these cells. Subsequently, using stably Tat producing asytocyte cell lines, we
observed that Tat promoted astrocyte survival by causing a prominent antioxidant effect and resistance to cell
injury in these cells. Tat was released extracellularly where it could be taken up by other cells. Tat remained
functionally active following uptake and caused LTR transactivation in lymphocytic and astrocytic cell lines
Tat released from astrocytes caused mitochondrial dysfunction, trimming of neurites and cell death in neurons. Tat
neurotoxicity was attenuated by anti-Tat antibodies, kynurenate or heparan sulfate. The neurotoxic effects of Tat
were caused at concentrations lower than that needed to cause LTR transactivation. When Tat expressing cells
were injected into the rat dentate gyrus, Tat was taken up by granule cells and transported along neuronal
pathways to the CA3 region where it caused glial cell activation and neurotoxicity. The arginine rich domain of Tat
was essential for both the LTR transactivation and the neurotoxic properties of Tat. Thus HIV-Tat is a potent
neurotoxin that may act at distant sites while at the same time it assures its production by preventing cell death in
astrocytes where it is produced.
Key Words: Tat, Neurotoxicity, Astrocyte, HIV, LTR-GFP, Mitochondria, Gene Array.
2
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
Introduction
The brain is a frequent target in patients with human immunodeficiency virus (HIV) infection resulting in a
dementing illness termed HIV dementia (HIVD). The pathological features associated with HIVD include microglial
cell activation, astrocytosis, decreased synaptic and dendritic density and selective neuronal loss (1). Neuronal
damage occurs through toxic substances released from infected microglia or macrophages and possibly
astrocytes (2). The virus predominantly infects microglia and macrophages where it causes a productive infection.
Astrocytes are also frequently infected with HIV but produce limited viral replication (2). The relevance of this viral
infection remains unknown. Astrocytes maintain a barrier between blood and brain and provide neuronal support
functions. Therefore, astrocytes may serve as a reservoir for the virus or induce neuronal damage by loss of
neuronal support functions or release of cellular and viral products. Hence infection of these cells could potentially
have long term consequences on cerebral function.
A striking feature is the presence of Tat transcripts in brains of HIV infected individuals (3,4). HIV Tat is a
non structural regulatory protein of 15 kD, that transactivates viral and cellular genes. It is produced in the early
phase of infection and is actively released from infected lymphoid cells extracellularly (5). Extracellular Tat may be
internalized by uninfected cells or it may interact with the cell membrane initiating a cascade of events (6). Tat
protein has both direct and indirect neurotoxic activity mediated by interaction with glutamate receptors, by
disruption of cytokine network and by reducing the neuroprotective effects of astrocytes (7-11). However, neither is
the role of intracellularly expressed Tat on astrocyte function known nor is the relative role of Tat uptake by
neurons or its membrane interactions understood.
Tat can produce oxidative stress in microglia cells (12), monocytes (13) and T-lymphocytes. In the latter,
intracellular expression of Tat leads to down regulation of mitochondrial superoxide dismutase there by causing
impaired mitochondrial membrane potential (14,15). Oxidative damage is difficult to measure directly, however,
oxidized endogenous macromolecules such as free protein carbonyl and 4-hydroxynonenal (HNE) can serve as
indicators of oxidative damage (16). Lipid peroxidation leads to the formation of HNE, a lipophilic alkenal that
forms stable adducts on mitochondrial proteins (17). It has been suggested to be key mediator of oxidative stress
3
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
induced cell death (18). Hence in this study, we have determined the effect of Tat on mitochondrial function and
other indicators of oxidative stress in astrocytes and neurons.
Experimental Procedures.
Plasmids.
Tat 86 from HIVHXB-2 was cloned from pGEX-Tat into pcDNA3 vector at BamH1/EcoR1 driven by a CMV
promoter. Further, Tat-86 and a deletion mutant of Tat-86 from which amino acids 48-56 were deleted by PCR
(∆Tat), was cloned in frame upstream of green fluorescent protein (GFP) gene driven by CMV promoter in pEGFP-
N1 (Clonetech) (Fig 1A). The HIV long terminal repeat (LTR) driven GFP construct was made in pEGFP vector by
deleting the CMV promoter and inserting the LTR at Sal1 and Sma1. The PCR cloned sequences were verified by
double strand DNA sequencing. Tetracycline (tet) “on” system was used for generation of inducible constructs.
Tat-86 was cloned downstream of a tet responsive element in pTREX vector (Clonetech). Reverse tetracycline
transactivator (rtta) was first cloned in pEGFP vector (Clonetech) at BamH1/ EcoR1 and further subcloned in
GFAP promoter driven vector pGfaLac-1 at BamH1/ BglII (Fig 1B). LTR-gag GFP construct was obtained from Q-
BIOgene, USA.
Cell culture.
Human fetal brain specimens were obtained from fetuses of 12-14 weeks gestational age. Neuronal
cultures were prepared as described previously (19). Briefly, the cells were mechanically dissociated, suspended
in opti-MEM with 5% heat inactivated fetal bovine serum (FBS), 0.2% N2 supplement and 1% antibiotic solution
(penicillin G 104 units/ml, streptomycin 10 mg/ml and amphotericin B 25 ug/ml). The dissociated cells were cultured
for at least one month prior to use in experiments. Human fetal astrocytes, rat C6 glioma cells, SVGA (a human
astrocytic subclone of the SVG cells) (20), and Hela cells were maintained in DMEM with 2mM L-glutamine, 10%
FBS and antibiotic solution.
4
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
Cell transfection
C6 (rat glioma cell line), SVGA (human fetal astrocytes cell line), Hela or primary human astrocytes were
transfected with endotoxin free plasmids (Qiagen) using lipofectamine plus (Invitrogen) as per supplier’s protocol.
Cells were monitored for GFP fluorescence or stained for Tat using a monoclonal antibody (APR 352, NIBSC,
U.K.). For establishing stably transfected cell lines, 72 hrs post transfection, the cells were cultured in DMEM
containing G418 (Life Technology) at 1.5 mg/ml. After one week, GFP florescent clones were picked under a
florescent microscope using trypsin soaked filter discs and sub-cultured for few weeks in G418 containing
selection media. The cells were used within 30 passages and maintained in 500 ug/ml G418. We established
SVGA-Tat, doxycycline inducible SVGA-Tat (SVGA13), SVGA-LTR-CAT, SVGA-LTR-GFP, SVGA-pcDNA (SVGA-
neo), C6-Tat, C6-Tat-GFP, C6-∆Tat-GFP, C6-pcDNA (C6-neo) and Hela-Tat cell lines for the present study. Tat
levels in culture supernatants were determined by a sandwich ELISA, where a Tat monoclonal antibody was
coated at the bottom of the plate and Tat polyclonal antisera (raised in rabbit against HIV-1 Tat72) was used as
the secondary antibody (4). The sensitivity of detection was 50 pg/ml. Highly purified recombinant Tat protein
produced in our laboratory was used as a standard (6).
RNA isolation and RT-PCR.
In each case, total RNA was extracted from 3-4x 106 cells by Trizol reagent (Life technology) and treated
with RNase free DNase. 1ug RNA was reverse transcribed as described previously (21) into cDNA using random
hexamer primers and 200 units of superscript reverse transcriptase (Gibco/BRL) at 45oC for 1 hr and 70oC for 10
min in 20 ul total volume. 50 ng of cDNA was used in PCR amplification with 25 pM of each primer, 0.1mM dNTP
mix, 1.25 unit platinum taq polymerase and 1.2mM Mg++. The following primers were used;
Tat; Tat-AC1 5’-ATGGAGCCAGTAGATCCTAG-3’, Tat-AC2, 5’-TCATTGCTTTGATAGAGAAACTTG-3’; rTta;
rTta1, 5’-AATCGAAGGTTTAACCCG-3’, rTta2, 5’-TTGATCTTCCAATACGCAACC-3’; iNOS, iNOS1 5’-
TGTGCCACCTCCAGTCCAGTGACA-3’, iNOS2 5’-GCTCATCTCCCGTCAGTTGGTAGG-3’; FASL, FASL1 5’-
TGGGGATGTTTCAGCTCTTC-3’, FASL2 5’-TCATCATCTTCCCCTCCATC-3’; GAPDH, GAPDH1 5’-
5
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
ACCACAGTCCATGCCATCAC-3’, GAPDH2 5’-TCCACCACCCTGTTGCTGTA-3’. The amplified products were
analyzed on 1% agarose gels with ethidium bromide and visualized by a UV transilluminator.
Immunofluorescence.
Cells grown in 60 mm dishes 24-48 hrs post transfection were fixed in 4% paraformaldehyde (PFA) for 15
min. The cells were washed with phosphate buffered saline (PBS), treated with 0.2% triton X-100 for 10 min at
room temperature (RT) and then blocked with 5% bovine serum albumin (BSA) in PBS for one hr. The cells were
incubated at 4°C for 18 hrs with either of the following antisera diluted in blocking buffer; monoclonal anti-Tat
(1:1000); anti-microtubule associated protein (MAP-2 ,a neuronal marker, 1:400; Chemicon) and anti-GFAP
(astrocyte marker, 1:400; Sigma). The cells were washed 4 times in PBS followed by incubation with anti-mouse
antisera labeled with Alexa 488 or 568 (Molecular probes) for 30 min at RT. After 4 washes in PBS, specimens
were visualized by fluorescent confocal microscopy.
Transactivation assay.
HIV-1 LTR-CAT based transactivation assays were performed using either Hela-LTR-CAT or SVGA-LTR-
CAT cells. A stable SVGA-LTR-CAT cell line was established using the LTR-CAT plasmid and selection with G418
for three weeks. The presence of LTR-CAT was confirmed following a transient transfection with pcDNA-Tat86.
Alternatively, C6-Tat-GFP, C6 -∆Tat-GFP, SVGA-Tat and SVGA13 were transfected with LTR-CAT plasmid. CAT
activity was monitored by quantitative ELISA. The supernatants from these cells were collected at 24 hrs, 48 hrs
and 96 hrs which was then added to LTR-CAT or LTR-GFP cells by a complete media exchange. The
supernatants were used fresh without undergoing any freeze thaw cycles. For detecting transcellular Tat activity,
Tat expressing cells were co-cultured with SVGA-LTR-GFP cells or a lymphocytic cell line D3R5 LTR-GFP (22) in
equal ratios for 48-96 hrs in 1.5% FBS containing medium with 50 µM chloroquine (to minimize the degradation of
endocytosed Tat). In select experiments polyclonal Tat antisera at 1:200 final concentration was included for
neutralization of Tat activity and observed by fluorescent microscopy.
6
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
mRNA profiles of apoptotic and chemokine related genes
Total RNA was isolated at different time intervals using Trizol reagent (Life technology) from the Tat or
pcDNA transfected primary astrocytes. 6-7 ug RNA was labeled using reverse transcription step and labeled cDNA
was hybridized at 68°C for 18 hrs with superarray nylon membranes for chemokine and apoptotic gene profile
(Super Array). The signals were quantified by a phospho-imager. The signals were normalized to GAPDH and
ratios of test and control were expressed as fold increase or decrease.
Protein carbonyl and 4-hydroxynonenol assays.
SVGA-Tat and SVGA-neo cells were treated with10 µM 3-nitroproprionic acid (3-NP) for varying amounts
of time at 37°C. Cells were collected in Locke’s buffer and sonicated for 5 sec. The level of protein oxidation was
determined by an oxidized protein detection kit (OxyblotTM, ONCOR) (23). Samples were incubated for 20 min
with 12% SDS and 2,4-dinitrophenylhydrazine (DNPH) in 10% trifluoroacetic acid, vortexing every 5 min, and then
neutralized with OxyblotTM Neutralization solution. 600 ng protein was blotted onto nitrocellulose paper by the slot
blotting technique. Membranes were incubated with blocking buffer (PBS with 3% BSA, 0.01% sodium azide and
0.2% Tween-20) for 30 min at RT, exposed to rabbit anti-DNPH protein antibody (1:150) for 90 min followed by
anti-rabbit IgG coupled to alkaline phosphatase (1:15,000), for 2 hrs at RT. Membranes were washed after each
step in PBS with 0.2% Tween-20 and developed with SigmaFastTM chromogen. Blots were analyzed by computer
assisted imaging software, Scion Imaging. Samples for HNE detection were similarly analyzed by slot blotting
technique except that rabbit-anti-HNE antibody (Calbiochem; 1:4,000) was used as a primary antibody.
Mitochondrial trans-membrane potential (δ Ψ) activity in neurons
Mitochondrial function was monitored by a fluorescent dye JC-1 to measure mitochondrial δ Ψ as
previously described (24). When loaded in the cells, JC-1 produces a green florescence at low membrane
potential and red at high membrane potential. At the end of the experimental treatment, the cells were incubated
7
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
with JC-1 (10 µM) at 37oC for 30 min and then washed in Locke’s solution. Fluorescence measurements were
made with excitation at 485 nm and emission at 527 nm and 590 nm. The levels of fluorescence at both emission
wavelengths were expressed as ratios of fluorescence measurements from which the percentage of the mean
ratios compared to the untreated samples was calculated and analyzed using ANOVA.
Neurotoxicity assay
Stable cell lines expressing Tat as SVGA-Tat, C6-Tat, C6-Tat-GFP, C6-∆Tat-GFP, SVGA13 and control
cell lines, SVGA-neo and C6-neo were maintained in DMEM+10% FBS. These cells were placed in the upper
chambers of transwells and neurons in the bottom of the 6 or 24 well plate in medium containing 0.5 to 1.0% FBS.
Tet-inducible cells (SVGA13) were induced 1 day prior to and upto 6 days after co-culture with 1 µg/ml
doxycycline. In select experiments either heparan sulfate (1.3µM), dextran sulfate (0.5µM) or kynurenate (10µM)
were added to the bottom chamber of the transwell. Cell death was monitored by trypan blue exclusion assay as
described previously (19). Five random fields were photographed and neuronal counts were determined by a
blinded investigator. At least 200 cells were counted in each field. Each experiment was done in triplicate and 3
independent experiments were conducted. The results are expressed as mean percentage of dead cells.
Implantation of Tat expressing cells in rat brain.
C6Tat-GFP cells were sorted by a flow sorter. 7.5 × 104 C6-Tat-GFP or C6-neo cells were unilaterally
injected into the molecular layer of the dentate gyrus of 12 week old Sprague-Dawley rats using the following
stereotactic coordinates: 3.8 mm anterior and 1.5 mm lateral to Bregma, and 3.2 mm dorsal to the dural surface.
Animals were monitored for 10 days post-surgery and then euthanized under deep anesthesia by perfusion with
normal saline followed by 4% PFA. The brains were removed, post-fixed for 16 hrs in 4% PFA, and then
cryopreserved in 30% sucrose for 72 hrs. 30 um sections were prepared from the hippocampus. Sections at the
needle tract and 300 um posterior to the injection site were processed for Nissl stain (25). Sections at the
injection site were also immunostained for Tat using a monoclonal Tat antibody.
8
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
Results.
Intracellular expression of Tat in astrocytes.
To determine if astrocytes can support the expression of Tat protein and to explore the subcellular
localization of Tat, we transfected primary human astrocytes or SVGA cells with full length HIV-1 infectious
molecular DNA clone pNL4-3. Tat expression was demonstrated by immunostaining (Fig. 2Aa and b) and was
further confirmed by LTR trans-activation in pNL4-3 transfected SVGA-LTR-GFP cells (Fig 2Ad). No
immunostaining for Tat or LTR transactivation was noted in pcDNA transfected cells (Fig 2Ac and e). Pattern of
Tat expression was also monitored in primary human fetal astrocytes transfected with Tat-GFP plasmid (Fig 2B).
The expression of Tat in astrocytes was confirmed by immunostaining for GFAP and Tat-GFP fluorescence (Fig.
2B b). Tat expression occurred within 24 hrs of transfection. Tat was expressed in the cytoplasm and nucleus with
a strong signal in the nucleolus as revealed by green fluorescence (Fig. 2B a, d, e). The nuclei were counter
stained red with propidium iodide (c). Primary astrocytes transfected with ∆Tat-GFP revealed ∆Tat expression
diffusely throughout the cell with no demarcation of nuclear boundary or no nucleolar pattern (Fig 2B f, g).
Stable transfection of C6 and SVGA cells with Tat.
C6 Tat-GFP and C6 ∆Tat-GFP clones were established by G418 selection and multiple subcloning by
FACS. The pattern of Tat and ∆Tat expression in these stable cell lines was similar to that of transiently
transfected astrocytes with the respective plasmids and did not change with prolonged culture. The expression of
Tat in stable cell lines was further confirmed by detection of Tat mRNA by RT-PCR (Fig. 3A) or LTR-CAT
transactivation assay following transfection of these cells with the LTR-CAT plasmid (Fig.3B). ∆Tat expressing
cells showed basal level of transactivation activity (Fig 3B). Clones of C6-Tat, SVGA-Tat and SVGA13 cells that
showed high CAT activity were used for further studies.
Functional expression of Tat and cell to cell transmission.
9
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
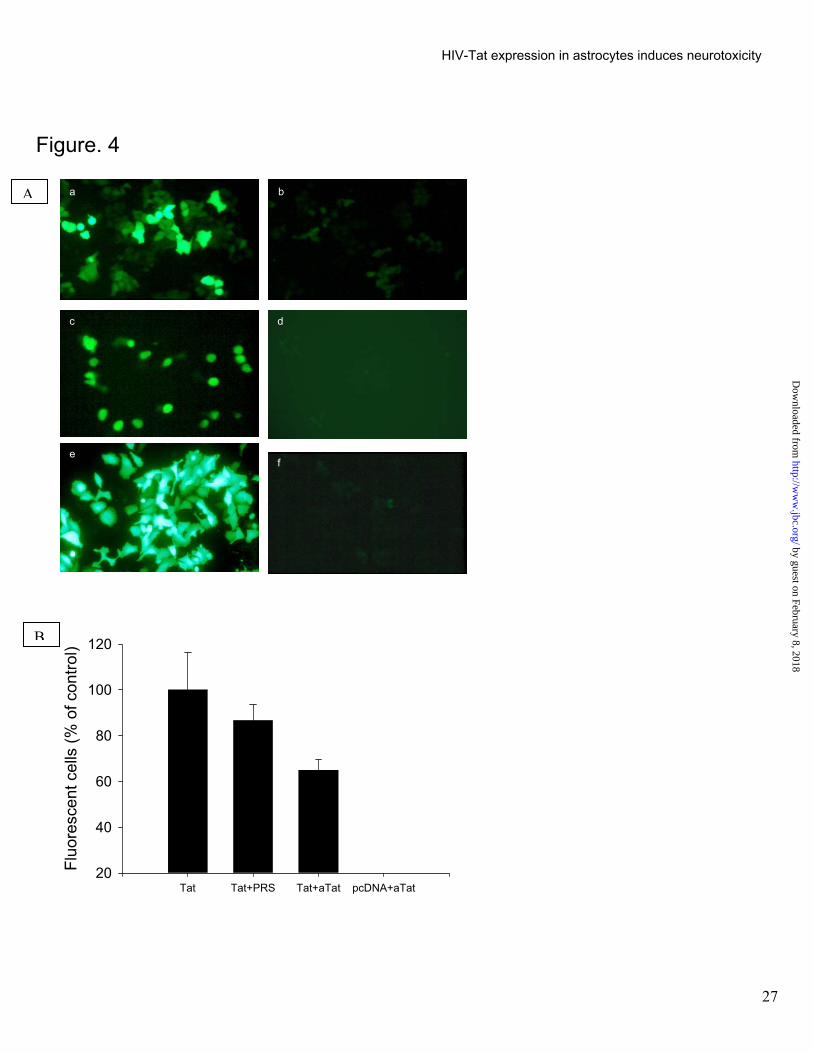
To determine if biologically active Tat can be released from Tat expressing astrocytes and taken up by non
expressing cells, SVGA-Tat cells were overlaid with SVGA-LTR-GFP cells. GFP expression occurred in 48 to 96
hrs of co-culture (Fig. 4A a). No GFP expression was noted when these cells were co-cultures with SVGA-neo
cells.(Fig 4A b) Co-culture of C6-Tat or SVGA-Tat with D3R5-LTR-GFP cells also revealed GFP fluorescence (Fig.
4A c), while no effect was noted when co-cultured with SVGA-neo or C6-neo cells (Fig. 4A d) SVGA cells when
directly co-transfected with LTR-GFP and Tat plasmids revealed higher level of LTR based GFP expression within
24 hrs of transfection (Fig. 4A e, f). Together, these observations demonstrate the LTR transactivation in both
astrocytic and lymphocytic cells was specific for the Tat expressing cells. To further confirm that the effect was due
to release of Tat from the Tat expressing cells and uptake by the LTR containing cells we repeated the above
experiments in the presence of Tat antisera. The cell to cell Tat mediated LTR transactivation activity was partially
inhibited by Tat antisera which is strongly suggestive of release of Tat extracellularly. The inability of the antisera
to block the LTR-transactivation completely, is most likely related to poor access to Tat by the antisera in areas of
cell to cell contact. The transactivation signal was specific for Tat since no effect was noted with preimmune rabbit
sera or by co-incubation of the LTR-GFP cells with SVGA-neo cells (Fig. 4B). Tat secretion in 48 hrs post
transfection period from 8x105 cells was estimated as 100 ng/ ml by ELISA. However, culture supernatants
containing Tat from the stable Tat expressing cells collected at 24, 48 and 96 hrs post media change (Tat
concentration less than 50 pg), did not reveal transactivation of LTR when incubated with the LTR-CAT or LTR-
GFP cells suggesting that small amount of Tat secreted may not be sufficient for LTR transactivation. This is
consistent with previous observations where the minimum Tat concentration required for LTR transactivation and
HIV replication was determined to be> 100 ng / ml (26).
Effect of intracellularly expressed Tat on Astrocyte mRNA profile for apoptosis and chemokine genes.
Since Tat was stably expressed in astrocytes, we determined the effect of Tat expression on apoptotic
genes as well as chemokine and cytokine gene expression profiles. Interestingly, primary astrocytes transfected
10
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
with Tat plasmid for 24 –36 showed that TNF-α and DAXX mRNA,s were elevated 2-4 fold (Table-1) while others
were unchanged. Surprisingly, we did not see any effect on MCP-1 or MIP-1α which was previously shown to
increase in astrocytes after extracellular treatment with Tat protein (27). Other mRNAs like, Caspase gene family,
iNOS, TRAIL, Bcl gene family and chemokine genes were unaffected, The results were also confirmed by RT-PCR
(not shown). Further, TNF-α and MCP-1 were undetectable in the 48 hrs conditioned medium from C6-Tat and
SVGA-Tat cells by quantitative ELISA. To rule out the under expression of Tat, Tat expression was monitored in
these cells by RT-PCR and immunostaining. Further cells expressing the Tat-GFP construct were used as a
control. Nearly 95% cells showed green fluorescence.
Protein carbonyl and HNE levels in Tat transfected cell lines.
To further explore the possible effects of Tat on vulnerability of astrocytes to oxidative stress, we analyzed
protein and lipid peroxidation products after challenging the cells with mitochondrial toxin 3-nitro-propionic acid
(3NP). Oxidized endogenous products protein carbonyl and HNE were measured as markers of oxidative stress
(16). Tat expressing astrocytic cells showed decreased oxidation products (HNE and protein carbonyl) when
compared to the neo-cells (Fig 5). These effects were specific for astrocytes as Hela cells expressing Tat did not
have a similar protective effect (Fig. 5).
Dissipation of neuronal mitochondrial δ Ψ by Astrocyte expressed Tat.
To determine the effect of Tat expressing cells on neurons, stable Tat producing or neo-C6 cells were co-
cultured with primary neurons using transwells and analyzed at various time points. The mitochondrial δ Ψ of
neurons was unaffected within the first 48hrs but after 72hrs membrane potential decreased significantly in
neurons co-cultured with C6-Tat cells as compared to C6-neo cells or C6- ∆Tat cells (Fig. 6). These observations
were confirmed by the use of doxycycline regulated expression of Tat in SVGA cells (Fig 6 ii). Doxycycline takes
approximately 72 hrs for induction of Tat. Hence in keeping with the above observation we noted a significant
decrease in mitochondrial δ Ψ at about 7 days after adding doxycycline to the media. No significant toxicity was
11
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
noted in similar cultures where doxycycline was not added or when doxycycline was added to the neuronal
cultures similarly incubated with SVGA neo cells where the Tat gene was not present (Fig. 6 ii).
These observations were further confirmed by collecting supernatants from stably Tat producing cells after
48 hrs of culture and applying it to primary neurons for different times. The C6 Tat conditioned media decreased
the neuronal mitochondrial membrane potential at 6 hrs post treatment (not shown).
Attenuation of astrocyte expressed Tat effect on neuronal mitochondrial δ Ψ by kynurenate, heparan
sulfate or dextran sulfate.
Previous studies have shown that Tat-induced neurotoxicity is mediated via excitatory amino acid receptors
and Tat binds to heparan sulfate and dextran sulfate (6,19). Hence we incubated Tat expressing astrocytes with
neurons in transwells in the presence of each of these compounds. All these compounds attenuated the
neurotoxicity measured as change in mitochondrial δ Ψ. In control cultures where staurosporine (mitochondrial
toxin) was incubated in the presence of heparan sulfate, no protective effect was noted (Fig. 6 iii), thus indicating
that extracellular Tat is responsible for the neurotoxicity.
Astrocyte expressed Tat induces neuronal cell death
To determine the effect of Tat expressing astrocytes on neuronal survival, we exposed neuronal cultures to
C6 Tat cells in transwells. Significant neuronal cell death occurred with C6 Tat cells (P<0.05). However, C6 ∆Tat
cells did not produce significant neurotoxicity (Fig. 7 i) indicating that amino acids 48-56 are essential for
neurotoxicity. To confirm the neurotoxic properties of Tat, SVGA-13 cells were cultured in transwells with neurons.
Doxycycline treated SVGA-13 cells induced significant neuronal cell death first noted at 4 days of induction
(P<0.001) with maximal neurotoxicity at 7 days post induction (P<0.001) compared to controls where doxycycline
was omitted or with SVGA-neo cells similarly treated with doxycycline (Fig. 7 ii). Further, neuronal cell death could
be neutralized using polyclonal Tat antisera but not preimmune antisera (7 iii) indicating that extracellular release
of Tat is required for neurotoxicity.
12
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
Neurite trimming in neurons by Tat.
Neurite fragmentation is a common phenomenon before the neurons collapse in response to toxic
substances and loss of dendrites is a prominent histopathological feature of patients with HIV encephalitis (28).
Hence we monitored morphological changes in neurons following culture with Tat expressing astrocytes. Direct co-
culture of C6-Tat GFP cells with primary human fetal neurons for 72 hours induced prominent neurite trimming or
loss as shown in figure 8, following Map-2 staining (red florescence). Tat production was confirmed by direct
visualization of GFP florescence.
Hippocampal neurotoxicity induced by Tat-producing C6 cells in vivo.
To determine if Tat-producing C6 cells could elicit neurotoxicity in vivo, C6-Tat or -neo cells were
unilaterally injected into the rat dorsal hippocampus. Nissl-stained sections from the injection site revealed that C6-
Tat or C6-neo cells produced local damage of the dentate granule cells, but only animals with C6-Tat cells had
extensive damage in the CA4/3 pyramidal cell layers (Fig. 9a). 300 um posterior to the injection site, only minimal
signs of neuronal injury were observed in animals with C6-neo cells, while extensive damage to the dentate gyrus
and CA4/3 pyramidal cells was observed in animals with C6-Tat cells (Fig 9b, c). Tat immunoreactivity at the
injection site was localized to the C6 cells (Fig 9d), indicating that these cells are able to survive and produce Tat
in vivo. Additionally, Tat immunoreactivity was present in the CA4/3 region in cells that morphologically resembled
both microglial cells and neurons (Fig 9d), raising the possibility that constitutively produced Tat from the C6 cells
could be secreted and taken up by adjacent cells in the brain.
Discussion.
To determine the effect of HIV-1 Tat on astroglia-neuronal relationships, we used constitutively and
inducible expressed Tat in astrocytes in contrast to previous studies that used recombinant Tat protein (29). We
used two different astrocytic cell lines and two approaches for intracellular Tat expression, i.e., direct expression
through CMV promoter and tet-inducible expression using a human GFAP promoter. Similar results were obtained
13
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
with both cell lines and both approaches for Tat expression. Our experimental model took advantage of a unique
observation that intracellularly expressed Tat was not toxic to astrocytes. In fact it had increased resistance to
oxidizing agents such as 3NP. We also noted overexpression of DAXX mRNA. Activation of physiological levels of
DAXX has been associated with induction of apoptosis however, overexpression has an anti-apoptotic effect (30).
This is in keeping with observations that HIV infection of astrocytes is non-cytopathic (31). We were thus able to
produce stably Tat expressing astrocytic cell lines. We expressed Tat in astrocytes because Tat can be detected
in HIV infected astrocytes in vivo (4) and in vitro as shown in this study. Production of Tat was monitored by mRNA
analysis, immunostaining, HIV-LTR transactivation assays and ELISA. Further, Tat released from these cells was
biologically active since it transactivated HIV-LTR in lymphocytic and astrocytic cell lines. These observations are
consistent with a previous study where Tat was taken up by neurons and was able to transactivate the HIV-LTR
(10). A unique pattern of Tat expression was noted intracellularly in astrocytes. Tat was highly concentrated in the
nucleolus, followed by the nucleus and cytoplasm. Deletion of the amino acids 48-56 resulted in diffuse staining
throughout the cytoplasm and nucleus confirming that this region of Tat is required for its nuclear localization (32-
34). Previous studies have shown that the intact arginine rich domain of Tat (aa 49-57) is required for nuclear or
nucleolar expression or localization (35-37). Despite the diffusion of ∆Tat into the nucleus, it failed to transactivate
HIV-LTR. In astrocytic cells, LTR transactivation occurs by two mechanisms; interaction of Tat with TAR RNA of
LTR or via interaction of Tat with NF- kappa B-binding sites in the LTR(38). The arginine rich domain is thus
essential for both modes of LTR transactivation and as discussed below also essential for Tat-induced
neurotoxicity. Hence this region may be an important target for development of therapeutic approaches. The
significance of Tat localization to the nucleolus remains undetermined. Interestingly, we were unable to detect an
effect of intracellularly expressed Tat on iNOS, TNF-∝, MCP-1, cytokines and chemokines production in astrocytes
using super array which was further confirmed by RT-PCR and ELISA for TNF-∝ and MCP-1 (data not shown).
Recently, it was shown that transiently transfected Tat plasmids induced iNOS and produced NO in an
astrocytoma cell line U373MG (39). However, our gene array and RT-PCR data on primary astrocytes as well as
the astrocyte cell line SVGA do not support these observations. Similarly, a macrophage cell line stably expressing
14
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
Tat inhibited induction of iNOS by gamma interferon (40). In previous studies, others and we have shown that
extracellular Tat has profound effects on cytokine and chemokine production by astrocytes (41). Thus intra and
extracellular Tat may have opposing effects.
We present several lines of evidence to conclusively show that extracellularly released Tat is neurotoxic.
Supernatants from the Tat expressing cell lines produced neurotoxicity as manifested by destablization of
mitochondria, neurite trimming and neuronal cell death. The neurotoxicity could be blocked by kynurenate,
heparan sulfate, dextran sulfate and antisera to Tat, clearly demonstrating that the neurotoxicity is mediated
directly via Tat. Kynurenate blocks the effect of Tat on glutaminergic receptors on neurons (19), while heparan
sulfate and dextran sulfate bind directly to Tat (5). This confirms previous studies showing that recombinant Tat
protein was toxic to neurons. However, the concentration of Tat necessary to cause the neurotoxicity was much
lower in the current study suggesting that Tat produced under physiological conditions by mammalian cells is a
potent neurotoxin. Another study showed that antisera to Tat could block neurotoxicity produced by supernatants
from HIV infected macrophages confirming the relevance of these observations(24).
We noted cell to cell movement of Tat among astrocytes and from astrocytes to lymphocytes using LTR-
GFP reporter expression indicator cells that were exposed to Tat producing astrocytes. This function was also
dependent upon amino acids 48-56 since no effect was seen with ∆Tat. Interestingly, this required co-culture of
Tat expressing cells with the indicator cell lines while culture supernatants from the stably Tat expressing cells
failed to induce LTR transactivation. This was most likely due to lower concentrations of Tat in the supernatants.
However, Tat released from the same cell lines was able to cause neurotoxicity suggesting that the concentrations
of Tat required for causing neurotoxicity are lower than that needed for transcellular transactivation of LTR. This
may in part be explained by the fact that Tat-induced neurotoxicity requires interaction of Tat with the neuronal cell
membrane (42) where as Tat-induced transactivation of LTR requires Tat uptake by the cell (43). Another unique
observation in vivo was that Tat released from the astrocyte cell lines was taken up by granule cells in the dentate
gyrus and transported anterogradely via mossy fibers to the CA4/3 region where it caused glial cell activation and
neurotoxicity. Thus Tat can cause pathological changes at sites distant to the site of production.
15
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
Our observations suggest the following sequence of events. During HIV replication in astrocytes, Tat is
produced as an early product. It makes the astrocytes resistant to oxidative stress thus making it an ideal reservoir
for long term survival of the viral genome. Tat localizes to the nucleolus, but small amounts are released
extracellularly. Tat may be taken up by resident cells in the brain or monocytic cells trafficing through the brain. If
these cells are latently infected with the virus, it will cause transactivation of the viral genome and viral replication.
If taken up be neurons it may be transported along neuronal pathways and cause neurotoxicity and glial cell
activation at distant sites. Small amounts of Tat are sufficient to cause excitotoxic neuronal damage that includes
mitochondrial dysfunction, neurite timming and neuronal cell death in select populations. Since mitochondria are
concentrated in neurites, the mitochondrial stress may be directly responsible for neurite trimming. Thus, HIV
infected astrocytes likely play a critical role in the neuropathogenesis of HIV infection. The Tat protein and in
particular the arginine rich region of Tat deserves close attention as a therapeutic target for HIV infection.
Acknowledgements. We thank Dr. G. Smith, for pGfalac1, Dr. M. Giacca, for pGEX Tat, Dr. Dorosky for D3R5
cells, Dr W. Atwood for SVGA cells , Dr Carlos Pardo for confocal microscopy and the NIH AIDS Research and
Reference Reagent Program and for plasmids pNL4-3 and LTR-CAT and Hela LTR-CAT cells and NIBSC, UK, for
monoclonal antibody to Tat. We thank C. Anderson, R. Reid and P. Ray for technical assistance. Supported by
grants from NINDS, NIDA, NIA and NCRR.
16
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
References
1. Masliah, E., Ge, N., Morey, M., DeTeresa, R., Terry, R. D., and Wiley, C. A. (1992) Lab Inves 66, 285-291 2. Nath, A. (1999) Sem Neurol 19, 113-128 3. Wesselingh, S. L., Power, C., Glass, J. D., Tyor, W. R., McArthur, J. C., Farber, J. M., Griffin, J. W., and
Griffin, D. E. (1993) Ann Neurol 33, 576-582 4. Hudson, L., Liu, J., Nath A, Narayan, O., Male, D., Jones, M., and Everall, I. (2000) J Neurovirol 6, 145-155 5. Chang, H. C., Samaniego, F., Nair, B. C., Buonaguro, L., and Ensoli, B. (1997) AIDS 11, 1421-1431 6. Ma, M., and Nath, A. (1997) J Virol 71, 2495-2499 7. Rappaport, J., Joseph, J., Croul, S., Alexander, G., Del Valle, L., Amini, S., and Khalili, K. (1999) J Leukoc
Biol 65, 458-465 8. Koller, H., Schaal, H., Freund, M., Garrido, S. R., von Giesen, H. J., Ott, M., Rosenbaum, C., and Arendt,
G. (2001) Eur J Neurosci 14, 1793-1799 9. New, D. R., Maggirwar, S. B., Epstein, L. G., Dewhurst, S., and Gelbard, H. A. (1998) J Biol Chem 273,
17852-17858 10. Kolson, D. L., Collman, R., Hrin, R., Balliet, J. W., Laughlin, M., McGann, K. A., Debouck, C., and
Gonzalez-Scarano, F. (1994) J Gen Virol 75, 1927-1934 11. Johnston, J. B., Zhang, K., Silva, C., Shalinsky, D. R., Conant, K., Ni, W., Corbett, D., Yong, V. W., and
Power, C. (2001) Ann Neurol 49, 230-241. 12. Nicolini, A., Ajmone-Cat, M. A., Bernardo, A., Levi, G., and Minghetti, L. (2001) J Neurochem 79, 713-716 13. Foga, I. O., Nath, A., Hasinoff, B. B., and Geiger, J. D. (1997) J Acquir Immune Defic Syndr Hum Retrovirol
16, 223-229 14. Westendorp, M. O., Shatrov, V. A., Schulze-Osthoff, K., Frank, R., Kraft, M., Los, M., Krammer, P. H.,
Droge, W., and Lehmann, V. (1995) Embo J 14, 546-554 15. Macho, A., Calzado, M. A., Jimenez-Reina, L., Ceballos, E., Leon, J., and Munoz, E. (1999) Oncogene 18,
7543-7551 16. Wilt, S. G., Dugger, N. V., Hitt, N. D., and Hoffman, P. M. (2000) J Neurosci Res 62, 440-450 17. Picklo, M. J., Amarnath, V., McIntyre, J. O., Graham, D. G., and Montine, T. J. (1999) J Neurochem 72,
1617-1624 18. Liu, W., Kato, M., Akhand, A. A., Hayakawa, A., Suzuki, H., Miyata, T., Kurokawa, K., Hotta, Y., Ishikawa,
N., and Nakashima, I. (2000) J Cell Sci 113 ( Pt 4), 635-641 19. Magnuson, D. S., Knudsen, B. E., Geiger, J. D., Brownstone, R. M., and Nath, A. (1995) Ann Neurol 37,
373-380 20. Major, E. O., Miller, A. E., Mourrain, P., Traub, R. G., de Widt, E., and Sever, J. (1985) Proc Natl Acad Sci
U S A 82, 1257-1261 21. Mori, I., Diehl, A. D., Chauhan, A., Ljunggren, H. G., and Kristensson, K. (1999) J Neurovirol 5, 355-362 22. Dorsky, D. I., and Harrington, R. D. (1999) J Acquir Immune Defic Syndr 22, 213-220 23. Butterfield, D. A., and Lauderback, C. M. (2002) Free Radic Biol Med 32, 1050-1060 24. Turchan, J., Anderson, C., Hauser, K. F., Sun, Q., Zhang, J., Liu, Y., Wise, P. M., Kruman, I., Maragos, W.,
Mattson, M. P., Booze, R., and ath, A. (2001) BMC Neurosci 2, 3 25. Bruce-Keller, A. J., Umberger, ., McFall, R., and Mattson, M. P. (1999) Ann Neurol 45, 8-15 26. Ensoli, B., Buonaguro, L., Baril
(1993) Journal of Virology 67, 2
27. Conant, K., Garzino-Demo, A.,E. O. (1998) Proceedings of th
28. Masliah, E., Heaton, R. K., MarJ. A., Nelson, J. A., Atkinson, J
29. Nath, A., and Geiger, J. D. (19930. Wu, S., Loke, H. N., and Rehem31. Nath, A., Hartloper, V., Furer, M
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from
NG
lari, G., Fiorelli, V., Gendelman, R., Morgan, R., Wingfield, P., and Gallo, R. 77-287Nath, A., McArthur, J. C., Halliday, W., Power, C., Gallo, R. C., and Major, e National Academy of Sciences 95, 3117-3121 cotte, T. D., Ellis, R. J., Wiley, C. A., Mallory, M., Achim, C. L., McCutchan, . H., and Grant, I. (1997) Ann Neurol 42, 963-972 8) Prog Neurobiol 54, 19-33 tulla, A. (2002) Neoplasia 4, 486-492 ., and Fowke, K. R. (1995) J Neuropathol Exp Neurol 54, 320-330
17

HIV-Tat expression in astrocytes induces neurotoxicity
32. Ruben, S., Perkins, A., Purcell, R., Joung, K., Sia, R., Burghoff, R., Haseltine, W. A., and Rosen, C. A. (1989) J Virol 63, 1-8
33. Frankel, A. D., and Pabo, C. O. (1988) Cell 55, 1189-1193 34. Stauber, R. H., and Pavlakis, G. N. (1998) Virology 252, 126-136 35. Mann, D. A., and Frankel, A. D. (1991) EMBO Journal 10, 1733-1739 36. Nagahara, H., Vocero-Akbani, A. M., Snyder, E. L., Ho, A., Latham, D. G., Lissy, N. A., Becker-Hapak, M.,
Ezhevsky, S. A., and Dowdy, S. F. (1998) Nat Med 4, 1449-1452 37. Vives, E., Brodin, P., and Lebleu, B. (1997) J Biol Chem 272, 16010-16017 38. Yang, L., Morris, G. F., Lockyer, J. M., Lu, M., Wang, Z., and Morris, C. B. (1997) Virology 235, 48-64 39. Liu, X., Jana, M., Dasgupta, S., Koka, S., He, J., Wood, C., and Pahan, K. (2002) J Biol Chem 40. Barton, C. H., Biggs, T. E., Mee, T. R., and Mann, D. A. (1996) J Gen Virol 77 ( Pt 8), 1643-1647 41. Nath, A., Conant, K., Chen, P., Scott, C., and Major, E. O. (1999) J. Biol. Chem 274, 17098-17102 42. Cheng, J., Nath, A., Knudsen, B., Hochman, S., Geiger, J. D., M, M., and Magnuson, D. S. K. (1998)
Neuroscience 82, 97-106 43. Liu, Y., Jones, M., Hingtgen, C. M., Bu, G., Laribee, N., Tanzi, R. E., Moir, R. D., Nath, A., and He, J. J.
(2000) Nat Med 6, 1380-1387.
18
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
FIGURE LEGENDS.
Figure 1 Tat expression systems:
(A) Tat constructs. (a). Wild type Tat with demarcation of nuclear localization signal (NLS); (b) Tat-GFP and (c),
Tat GFP with deletion of 48-56 amino acids (∆Tat-GFP).
(B). Tetracycline (Tet) inducible expression. (a) GFAP promoter driven reverse Tet transactivator (rTtA)
expression; (b) rTtA induces Tet responsive element (TRE) driven Tat expression in the presence of doxycycline.
Figure 2 (A) Tat production by HIV-1 (pNL4-3) proviral DNA transfection in astrocytes.
Immunostaining for Tat after 48 hrs of transfection in (a) primary astrocytes, (b) SVGA cells transfected with
infectious pNL4-3 DNA and in (c) pcDNA transfected control SVGA cells. LTR based GFP expression in SVGA
cells (d) co-transfected with pNL4-3 and HIV-1 LTR-GFP or (e) pcDNA and HIV LTR-GFP control.
(B) Expression of wild type and mutant Tat in astrocytes.
Cells were transfected with wild type Tat-GFP or ∆Tat-GFP and observed for GFP fluorescence and immuno-
staining for GFAP after 24 hrs of transfection. (a) Primary astrocytes reveal Tat-GFP expression (green)
predominantly in nucleus but also in cytoplasm; (b) same cells immuno-stained for GFAP (red). (c) Propidium
iodide staining for nuclei (red) in SVGA Tat-GFP cells. (d) The same cell as in c, showing nucleolar Tat-GFP
(green) and (e) same cell with dual red and green fliter. (f) primary astrocytes express ∆Tat-GFP diffusely
throughout the cell with no nuclear demarcation; (g) same cells as in e stained for GFAP.
Figure 3
(A) Tat mRNA expression in astrocytes.
Each cell line was analyzed for Tat mRNA expression by RT-PCR and PCR products were shown by ethedium
bromide stained agarose gel. Lane 1, C6 Tat; lane 2, SVGA-Tat; Lane 3, SVGA-Tat13 in the absence of
19
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
doxycycline; lane 4, SVGA-Tat (tet) in the presence of doxycycline; lane 5, Hela Tat; lane 6, Hela; lane 7, positive
control (tat plasmid); lane 8, molecular weight markers.
(B) Transactivation of HIV-1 LTR-CAT in Tat expressing astrocytes.
Tat expressing astrocytes (SVGA (S) or C6 cells) were transfected with HIV LTR-CAT plasmid and quantitated for
CAT activity by ELISA. Values represent mean optical density readings of two experiments. Bars represent CAT
activity in the following cell types. 1, S-Tat ; 2, S-Tat 13 with doxycycline (Adamson #1) for 72 hrs; 3, S-Tat13; 4,
S-Neo with D; 5, C6 Tat; 6, C6 Tat-GFP; 7, C6 dTat-GFP (mutant Tat).
Figure 4. Cell to cell migration of functionally active Tat.
A. LTR transactivation in astrocytic and lymphocytic cells following co-culture with Tat expressing cells: C6-
Tat or SVGA-Tat cells were co-cultured with D3R5-LTR-GFP or SVGA-LTR-GFP cells. Green fluorescence
indicates LTR transactivation. (a), SVGA LTR-GFP cells overlaid on a monolayer of SVGA-Tat cells; (b)
control where SVGA LTR-GFP cells were overlaid on SVGA-neo cells; (c),Lymphocytic D3R5-LTR-GFP
cells were incubated with C6-Tat cells; (Adamson #1), control where D3R5-LTR-GFP cells were incubated
with C6-neo cells; (e), SVGA-LTR-GFP cells transfected with Tat plasmid; (f) SVGA LTR-GFP cells
transfected with pcDNA-3 plasmid.
B. Effect of Tat antisera on cell to cell LTR-GFP activation assay. SVGA cells transfected with 6.0 µg tat or
neo plasmid were overlaid with SVGA LTR-GFP cells followed by addition of Tat antisera (aTat) or
preimmune rabbit serum (PRS) Florescent cells were counted 24 hours later and calculated as a
percentage of the mean number of fluorescent cells in the wells without aTat or PRS. aTat partially blocked
Tat transactivation activity while PRS did not show blocking potential.
Figure 5.Tat production protects against oxidative stress in astrocytes.
20
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
Protein carbonyl and HNE products were measured in each cell line with or with out treatment with 3NP. (a)
Protein carbonyl levels were increased in Hela cell lines following 3NP treatment. No significant difference was
present between the Hela-neo (Hela) and Hela-Tat cells. (b) 3NP treatment failed to induce any significant change
in HNE products in the Hela cell lines (c) Tat expressing SVGA cells showed significantly lower production of
protein carbonyl following 3NP treatment (Adamson #1) Tat expressing SVGA cells showed significantly lower
production of HNE products following 3NP treatment. Data represents mean ± SE from 3 experiments. *P<0.05
Figure 6. Effect of Tat expressing astrocytes on neurons: Neuronal cultures were exposed to Tat expressing
astrocytes in transwells.
A (i) Mitochondrial δψ in neurons after co-culture with C6-Tat cells in transwells. Significant decrease in
mitochondrial δψ was noted following incubation with C6-Tat cells. No significant change was noted with C6-neo
or C6∆Tat cells. 3NP was used as a control. (ii) Mitochondrial δψ in neurons after co-culture with tet inducible
SVGA Tat cells in transwells. Significant decrease in mitochondrial δψ was in neurons following induction of Tat in
SVGA13 Tat cells by doxycycline. No significant change in neuronal (N) mitochondrial δψ was noted following
incubation with SVGA-neo cells similarly treated with doxycycline or SVGA13 cells without treatment with
doxycycline. 3NP was used as positive control.
(iii) Effect of kynurenate, dextran sulfate and heparan sulfate on C6-Tat induced neuronal mitochondrial effects.
Kynurenate, dextran sulfate and heparan sulfate blocked decreases in mitochondrial δψ in neurons induced by C6-
Tat cells in transwells. Heparan sulfate did not block staurosporine induced decrease in neuronal mitochondrial δψ.
In each figure data represents mean ± SE of 4 experiments. *P<0.05
Figure 7. Tat cells induced significant cell death in neurons. (i) C6-Tat cells induced significant cell death in
neurons (P< 0.05). No significant neuronal cell death was noted with C6-neo or C6-∆Tat cells. (ii) Doxycycline
induced Tat expressing SVGA13 cells caused significant neuronal cell death. No significant cell death was noted
with SVGA-neo cells similarly treated with doxycycline or with SVGA13 cells untreated with doxycycline. (iii) Tat
21
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
antisera but not preimmune rabbit sera (prs) significantly blocked neurotoxicity induced by C6-Tat cells in
transwells.
Figure 8. Neurite trimming by Tat producing astrocytes.
C6-Tat or C6-neo cells were co-cultured with primary fetal neurons and
immuno-stained for MAP-2. (a) neurons co-cultured with C6-Tat cells showing neurite shortening or trimming; (b)
neurons co-cultured with C6-neo cells showing long and extended neurites .
Figure 9. Effect of Tat-producing astrocytes on hippocampus in vivo.
Adult male rats were stereotactically injected with C6-neo cells (Left panel) or C6Tat-GFP cells (Right panel) into
the dorsal hippocampus. At 10 days sections were processed for Nissl stain and Tat immune reactivity. Nissl-
stained images (panels a-c) depict hippocampal neurons at the injection site or 300 µm posterior to the injection
site (- 300 µm) (magnification 4x or 20x). Arrows indicate damaged, pyknotic CA3 pyramidal neurons observed
only in animals injected with C6Tat-GFP cells. Immunoreactivity for Tat protein (Tat IHC; panel d) was seen at the
injection site and in the CA3 region. Images are representative of results obtained from 4 separate experiments.
22
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
Table 1. mRNA profile of Tat transfected primary human fetal astrocytes. Gene Fold Induction (21 hrs P.T) -------------------------------------------------------------------------- DAXX ↑ 4.7 TNF-α ↑ 2.65 TNF-α convertase ↑ 2.5 TRAF6 ↑ 2.9 FASL ↑ 2.6 FAS ↑ 2.7 Caspase gene family, iNOS, TRAIL, Bcl gene family, Chemokine genes were unaffected.
23
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
Figure 1
1 48 56 86 Tat-86
A
GFP 86 1Tat-86 GFP
48 56 86 GFP Tat-
86 GFP
1
NLS Deleted rTta
(a) GFAP Promoter B
rTta
(b)
Tat TRE tat Transcription Tet
24
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

Figure 2 HIV-Tat expression in astrocytes induces neurotoxicity
25
ca b
e d
b a
ed c
g f
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
Figure.3 2 3 4 5 6 7 8 A
Tat
0
0
1
1
2
2
LTR
-CA
T ac
tivity
(O.D
.)
B
1
242 bp
.0
.5
.0
.5
.0
.5
Dow
nl
S-TatS-Tat13+D
S-Tat13 S-NeoC6-Tat
C6-Tat-GFPC6-dTat-GFP
26
by guest on February 8, 2018http://w
ww
.jbc.org/oaded from
HIV-Tat expression in astrocytes induces neurotoxicity
Figure. 4
A
c
a
d
b
f e
Fluo
resc
ent c
ells
(% o
f con
trol)
B
by guest on February 8, http://w
ww
.jbc.org/D
ownloaded from
Tat Tat+PRS Tat+aTat pcDNA+aTat20
40
60
80
100
120
27
2018

HIV-Tat expression in astrocytes induces neurotoxicity
Figure. 5
Hela Hela-Tat Hela+3NP Hela-Tat+3NP0
100
200
300
400
500
600
SVGA-neo SVGA-TAT SVGA-neo SVGA-Tat0
100
200
300
400
500
600
3NP
*
Hela Hela-Tat Hela+3NP Hela-Tat+3NP
O.D
.
0
100
200
300
400
500
600
SVGA-neo SVGA-TAT SVGA-neo SVGA-Tat0
100
200
300
400
500
600
3NP
*
28
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
Figure. 6
Figure. 7
Neurons +C6-neo +C6-Tat +C6-dTat +3NP
Mito
chon
dria
l Pot
entia
l(%
cha
nge)
20
40
60
80
100
* *
Neurons +Stau +SVGA13 +SVGA +SVGA1320
40
60
80
100
**
dox
Neurons+C6Tat
+C6Tat+Kyn+C6Tat+Dex
+C6Tat+Hep+Staur+Hep
20
40
60
80
100
*
*
Neurons +C6 neo +C6Tat +C6dTat
% n
euro
nal c
ell d
eath
(mea
n+SE
)
02468
1012141618
*
Neurons +Stau +SVGA13 +SVGA +SVGA13 02468
1012141618
dox
*
*
pcDNA+Ab +prs
2468
1012141618
C6-Tat86 cells
*
29
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

HIV-Tat expression in astrocytes induces neurotoxicity
30
Figure 8
a b
Figure 9
a
b
c
d
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from

Allan Butterfield, Eugene Major and Avindra NathAshok Chauhan, Jadwiga Turchan, Chava Pocernich, Anna Bruce-Keller, Susan Roth, D.
transportastrocyte survival but induces potent neurotoxicity at distant sites via axonal Intracellular human immunodeficency virus tat expression in astrocytes promotes
published online January 24, 2003J. Biol. Chem.
10.1074/jbc.M209381200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on February 8, 2018http://w
ww
.jbc.org/D
ownloaded from