international journal of food microbiology manuscript
TRANSCRIPT

International Journal of Food
Microbiology
Manuscript Draft
Manuscript Number: FOOD-D-20-00197
Title: Multivariate analysis of microbial and volatile compounds profile
of Mexican Criollo cocoa beans fermented by artisanal methods with
consecutive turning
Article Type: Full Length Article
Keywords: HS-SPME/GC-MS; key-aroma markers; MALDI-TOF; Principal
Component Analysis; PLS-DA.
Abstract: The effect of two artisanal methods of fermentation of Mexican
Criollo cocoa with a different turning start time on the microbial
dynamics and volatile compounds generated were studied and analyzed by
multivariate analysis (Principal Component Analysis and Partial Least
Square Discriminant Analysis). The turning start time at 48h stimulated a
microbial profile with yeasts domain such as Hanseniaspora opuntiae,
Pichia manshurica, and Candida carpophila, favoring the production of
several key-aroma markers associated with cocoa quality such as
phenylethyl acetate, 2-phenylacetaldehyde, 3-methylbutanal, 2-phenylethyl
alcohol, 2,3-butanedione, 3-methylbutanoic acid, and 2-methylpropanoic
acid. While an immediate turning start time favored an aerobic
environment that stimulated the rapid growth of Acetobacter pasteurianus,
Bacillus subtilis, and higher biodiversity of LAB such as Lactobacillus
plantarum and Pedioccoccus acidilactici, increasing the ethyl acetate
production. Thus offering cocoa producers a scientific-based tool for the
selection of the fermentation technique based on the volatile compounds
profile in fermented cocoa.

Highlights
The microbial dynamics and volatile compounds generated in two traditional
fermentation methods of Mexican Criollo cocoa with a different start time of
turning were evaluated.
The fermentation methods evaluated did not change the fermentation time, but the
volatile compounds profile.
The multivariate analysis revealed the volatile compounds related to the different
turning methods
The turning start time at 48h stimulated a yeast domain that favors the production of
compounds recognized as key-aroma markers that are associated with cocoa quality.
An immediate turning start favors a mostly aerobic environment that stimulated the
rapid growth of genus Bacillus spp. which favor the ethyl acetate production.
*Highlights (for review)

1
Multivariate analysis of microbial and volatile compounds profile of Mexican Criollo 1
cocoa beans fermented by artisanal methods with consecutive turning 2
3
Velasquez-Reyes Dulce1, Gschaedler Anne2, Kirchmayr Manuel2, Avendaño-Arrazate 4
Carlos3, Rodriguez-Campos Jacobo1, Calva-Estrada Sergio1, Lugo-Cervantes Eugenia1 5
6
1Food Technology Departament. Centro de Investigación y Asistencia en Tecnología y Diseño del 7
Estado de Jalisco (CIATEJ) A.C., Camino Arenero 1227, El Bajío, Zapopan, Jalisco, Mexico. C.P. 8
45019. 9
2Industrial Biotechnology Departament, Centro de Investigación y Asistencia en Tecnología y 10
Diseño del Estado de Jalisco (CIATEJ) A.C., Camino Arenero 1227, El Bajío, Zapopan, Jalisco, 11
Mexico. C.P. 45019. 12
3Genetic Departament, Instituto Nacional de Investigaciones Forestales Agricolas y Pecuarias 13
(INIFAP), C. E. Rosario Izapa, Chiapas. Km. 18. Carretera Tapachula-Cacahoatan, Tuxtla Chico, 14
Chiapas, Mexico. CP. 30780. 15
16
17
18
*Corresponding author: [email protected] 19
20
21
*Manuscript with Line NumbersClick here to view linked References

2
Multivariate analysis of microbial and volatile compounds profile of Mexican Criollo 22
cocoa beans fermented by artisanal methods with consecutive turning 23
24
Abstract 25
The effect of two artisanal methods of fermentation of Mexican Criollo cocoa with 26
a different turning start time on the microbial dynamics and volatile compounds generated 27
were studied and analyzed by multivariate analysis (Principal Component Analysis and 28
Partial Least Square Discriminant Analysis). The turning start time at 48h stimulated a 29
microbial profile with yeasts domain such as Hanseniaspora opuntiae, Pichia manshurica, 30
and Candida carpophila, favoring the production of several key-aroma markers associated 31
with cocoa quality such as phenylethyl acetate, 2-phenylacetaldehyde, 3-methylbutanal, 2-32
phenylethyl alcohol, 2,3-butanedione, 3-methylbutanoic acid, and 2-methylpropanoic acid. 33
While an immediate turning start time favored an aerobic environment that stimulated the 34
rapid growth of Acetobacter pasteurianus, Bacillus subtilis, and higher biodiversity of LAB 35
such as Lactobacillus plantarum and Pedioccoccus acidilactici, increasing the ethyl acetate 36
production. Thus offering cocoa producers a scientific-based tool for the selection of the 37
fermentation technique based on the volatile compounds profile in fermented cocoa. 38
39
Keywords: HS-SPME/GC-MS, key-aroma markers, MALDI-TOF, Multivariate analysis, 40
Principal Component Analysis, PLS-DA. 41
42

3
1. Introduction 43
México represents the 13th
worldwide producer of cocoa beans (Theobroma cacao L.) 44
with an annual production of 28,363 ton in 2018 (SIAP, 2018). The cocoa aroma potential 45
is the most important quality attribute that determines the economic value and acceptability 46
of chocolate and derived products (Magagna et al., 2017). The cocoa aroma depends on the 47
genotype, geographical origin, and post-harvest processing (fermentation, drying, and 48
roasting), imparting a high variability to the composition and quality of the cocoa flavor 49
(Kongor et al., 2016). Several chocolate brands are interested in controlling each stage in 50
the manufacture of their product, focusing on the principal and crucial process, the 51
fermentation (Crafack et al., 2014; Menezes et al., 2016). 52
53
The cocoa fermentation is a spontaneous process that involves a microbial succession 54
responsible for the production of aroma precursor compounds and the development of 55
pigments characteristic of cocoa inside the beans through biochemical reactions 56
(Papalexandratou et al., 2019). The traditional fermentation process involves the extraction 57
of the cocoa beans from the pod. Then, the beans are placed in either heaps, boxes (of 58
wooden or plastic), or trays, and are covered with plantain leaves and left to ferment for 5-7 59
days according to the cocoa origin and genotype (Guehi et al., 2010; Moreira et al., 2013). 60
Throughout the process, a wide range of yeasts and fermenting bacteria appears, increasing 61
the cocoa mass temperature above 45 ºC, and producing a liquid rich in ethanol, lactic acid, 62
and acetic acid that change the color and volume of the beans, and yield several aroma 63
precursors by proteolysis and carbohydrates hydrolysis (Camu et al., 2008a). The 64
fermentation method generally is based on experience of generation to generation, resulting 65

4
in very heterogeneous process applied in the world that implies a greater variation in 66
microbial growth and in the generated metabolites according to the cocoa genotype, origin, 67
production method, batch size, pod ripeness, pod storage, and fermentation conditions 68
(Guehi et al., 2010). One of the common differences between the fermentative cocoa 69
processes is the turning method. The application of two turning times (the first at 48 h, and 70
a second at 96 h) during the fermentation of African cocoa has been studied (Camu et al., 71
2008b; Guehi et al., 2010; Hamdouche et al., 2019). In the cocoa-pulp turning the 72
oxygenation is favored, increasing the growth of Acetic Acid Bacteria (AAB) and 73
producing higher acetic acid amount, a key metabolite in the production of aroma 74
precursors, and decreasing the fermentation time. 75
76
In Mexico, producers apply different fermentation techniques for freshly cut cocoa in 77
search to improve the aroma profile and decrease the fermentation time. Some of them 78
involve the consecutive turning of cocoa pulp with a different turning start time (at 24 h or 79
at 48 h). However, there are no scientific studies that support the fermentation techniques 80
applied. Therefore, the aim of this study was to investigate the effect of two artisanal 81
fermentation methods from Mexican Criollo cocoa with a different turning start time on the 82
microbial dynamics and volatile compounds produced through the multivariate analysis 83
(Principal Component Analysis and Partial Least Square Discriminant Analysis). 84
85
2. Materials and methods 86
2.1 Fermentation process, monitoring, and sampling 87

5
Samples of Criollo cocoa pods were harvested in October 2018 in Chiapas, Mexico. 88
Cocoa beans were spontaneously fermented by the wooden box method. The freshly cut 89
pods were opened using a machete in the same place where they were fermented. The beans 90
were extracted manually and divided into two batches. Each batch was distributed into 91
individual wooden boxes (0.79 m x 0.80 m x 0.75 m each one). Two experimental 92
treatments were evaluated: B1) a spontaneous fermentation with first turning time at 24 93
hours after placing the beans in the fermentation box with consecutive turnings each 24 94
hours; and B2) a spontaneous fermentation with the first turning time at 48 hours after 95
placing the beans in the fermentation box with consecutive turnings each 24 hours. The 96
method selection was made following the artisanal practices applied. The turning of the 97
cocoa beans was manual, moving the total mass from one box to another empty box to 98
ensure uniform aeration during the process. The fermentation time was 7 days for both 99
batches. The end of the fermentation was based on temperature, pH changes, and the death 100
of the bean embryo. A sample of the beans mass was collected for each fermentation time 101
(0, 24, 48, 72, 96, 120, 144 h). The microbial and chemical analyses were carried out 102
immediately. The environmental temperature, box temperature, pH value and soluble solids 103
(ºBx) were measured throughout the fermentation. The pH was measured using a pHmeter 104
(pH Hanna HI98108). Soluble solids were measured using a refractometer. And the 105
temperature was measured in the middle of the fermenting mass with a digital thermometer 106
(Fluke t3000 FC). 107
108
2.2. Microbial ecology analysis 109

6
For the microbial analysis, 5 g of beans mass sample was vortexed with 10 mL of 110
physiological solution for 5 min. The supernatant was placed into a 1.5 mL sterile 111
Eppendorf tube and then different dilutions were prepared, depending on the expected 112
microbial counts. Then were plated on Wallerstein (WL) agar (Sigma-Aldrich) for yeast 113
enumeration, MRS agar (Sigma-Aldrich), and GYC agar (25 g/L D-glucose, 5 g/L yeast 114
extract, 10 g/L calcium carbonate, 7.5 g/L agar). Incubation was started in situ at 115
environmental temperature (24-34 ºC) for 24 h. Then, the plates with 30-300 colonies were 116
used for counting by the morphology of the colony. The Colony-Forming Units (CFU) was 117
recorded and isolated. Microbial morphology served as isolation criteria, for yeast, the 118
isolation was performed on Yeast-extract Peptone Dextrose agar (YPD), while the bacteria 119
were isolated on MRS and GYC agar. 120
121
2.2.1. MALDI-TOF sample preparation, measurement and data analysis 122
Isolates were grown on plates using specific culture medium and were incubated at 28 123
ºC for 18 h. Using the direct transfer method, the biological material (single colony) as a 124
thin film directly onto a spot on a MALDI target plate and covered with 1 µL of a saturated 125
solution of α-cyano-4-hydroxycinnamic acid (CHCA). Each MALDI-TOF sample was 126
spotted in duplicated to evaluate reproducibility. Samples were then analyzed in a Matrix-127
Assisted Laser Desorption/Ionization–Time-Of-Flight Mass Spectrometry (MALDI-TOF 128
MS) Microflex spectrometer (Bruker Daltonics), using the MALDI Biotyper 3.1 automatic 129
system. 130
131
2.3. Volatile compounds analysis 132

7
The analysis of volatile compounds was performed using the methodology described by 133
Rodriguez-Campos et al. (2011). The volatile compounds were extracted by Head-Space 134
Solid Phase Micro-Extraction (HS-SPME) using a 50/30 μm 135
divinylbenzene/carboxene/polydimethylsiloxane (DVB/CAR/PDMS) fiber (Supelco). The 136
sample (2 g) was placed in a head-space vial, and sealed with a PolyTetraFluoroEthylene 137
(PTFE) cap. The sample was equilibrated at 60 ºC for 15 minutes and then, the fiber was 138
exposed for 30 min at the same temperature. The volatile compounds were analyzed by Gas 139
Chromatography-Mass Spectrometry (GC-MS), equipped with an Innowax capillary 140
column (60 m x 0.25 mm id x 0.25 μm film thickness). And the identification of volatile 141
compounds was made comparing the mass spectra of the compounds in the samples with 142
the database of the National Institute of Standards and Technology (NIST/EPA/NIH v2.0 d 143
2004 Library, Gaithersburg, MD, USA) with a match of at least 80%. The average relative 144
abundance (n=3) of each compound was reported as a percentage of normalized area of the 145
corresponding peak. Aroma descriptors for the compounds identified were obtained by 146
bibliographic review. 147
148
2.4. Data processing and multivariate analysis (PCA and PLS-DA) 149
All analyses were performed in triplicate (n = 3). The experimental data are 150
presented as the mean and Standard Deviation (SD). An analysis of variance was performed 151
(ANOVA) and Tukey multiple range tests were conducted to determine the significant 152
differences among the means. The mean differences were considered significant at p<0.05. 153
Data analysis was performed using the XLSTAT 2019. 2. 3 (Addinsoft, Boston, USA). 154
155

8
The variables used in the Principal component analysis (PCA) represent the 156
normalized area of the chromatographic peaks obtained by GC-MS, and the CFU 157
corresponding to the main microorganisms identified during fermentation. Each variable 158
corresponds to the average of experimental points (analysis) done in triplicate. PCA bi-plot 159
was constructed to identify differences in the dynamics of microbial growth and volatile 160
compounds generated in the two fermentation treatments studied. The data set consisted of 161
12 observations (different fermentation times of both treatments) and 60 variables (52 162
volatile compounds, and 8 microorganisms). Previously, the complete matrix of volatile 163
compounds (70 volatile compounds) was filtered selecting only those compounds that were 164
identified at the beginning and end of the fermentation. Subsequently, all the set of 165
variables was transformed by mean-centering for the analysis. 166
167
Partial Least Square Discriminant Analysis (PLS-DA) was applied to classify the 168
volatile compounds generated according to the fermentation treatment used. PLS-DA 169
reduces the number of variables used in the model combining the variables in order to 170
calculate the factors that most correlated with a class (treatment in this case). For the 171
analysis, the complete matrix of the relative abundance data of each compound identified 172
by GC-MS as qualitative dependent variables (Y) the type of treatment (B1 or B2), as 173
quantitative explicative variables (X) the volatile compounds, and as qualitative explicative 174
variables the different fermentation times for both treatments. The PLS-DA model was 175
evaluated by exhaustive cross-validation. The data were transformed by mean-centering 176
with a reduction of the aligned data based with a 95% confidence interval. 177
178

9
3. Results and Discussion 179
3.1. Physicochemical changes 180
The soluble solids (ºBrix), pH, environmental temperature, and temperature inside the 181
fermentation boxes evaluated throughout the process are shown in Figure 1. The mentioned 182
parameters are critical in the microbial enzymatic activity generated during the cocoa beans 183
fermentation (Camu et al., 2008a; Koné et al., 2016). After cacao pods opening, the initial 184
ºBrix and pH values were 13.30–13.33, and 3.53–3.59 respectively, in both fermentations 185
batches. The soluble solids of cocoa-pulp are related to the sugar content (between 10–186
15%), citric acid (1–3%), and pectin (1–1.5%) (Santander-Muñoz et al., 2019). And the 187
acidity of the initial pH of cocoa-pulp is attributed to the citric acid content mainly 188
(Lagunes-Gálvez et al., 2007). The behavior of pH, ºBx, and box temperature during 189
fermentation, were statistically different between the two treatments evaluated (p<0.05) 190
(Figure 1). The pH values and box temperature were increasing throughout the 191
fermentation, while the soluble solids were decreasing. In the treatment B1 (with initial 192
turning at 24 h), the ºBx showed a higher decreasing rate compared to the treatment B2 193
(with initial turning at 48 h). The microbial activity and metabolites generated during the 194
cocoa fermentation lead to an increase in temperature and pH (Moreira et al., 2017). The 195
rapid decrease in soluble solids in the cocoa bean mass may be associated with the highest 196
number of turning applied in B1 treatment compared to B2. Turning favors the deposition 197
of mucilaginous liquid of cocoa beans, rich in sugars, at the bottom of the fermentation box, 198
leading to the decrease of the substrate available for ethanol-producing yeasts at the 199
beginning of the fermentation (Schwan & Wheals, 2004). The decrease of soluble solids 200
from treatment B1 was correlated with a higher rate of temperature rise compared to 201

10
treatment B2. In B1, the maximum box temperature was 46.5 ± 2.33 ºC at 48 h, while in B2 202
it was observed at 72 h. The environmental temperature was the same for both treatments. 203
The results could be associated with the oxygenation incorporated with the turning at 24 h 204
in the B1 treatment, which could stimulate the growth of Acetic Acid Bacteria (AAB) 205
characterized by a highly exothermic metabolism generating an increase in temperature 206
above 45 °C (Papalexandratou et al., 2019; Ramos et al., 2020; Schwan & Wheals, 2004). 207
The pH increase is associated with the activity of Lactic Acid Bacteria (LAB) that 208
consumes the fresh pulp citric acid as an alternative energy source for yeasts and LAB, 209
causing that the pulp pH increase from the onset with values around 3.5–4.0 (Illeghems et 210
al., 2015; Nielsen et al., 2013; Papalexandratou et al., 2019). The pH of the cocoa-bean 211
mass was increased during the fermentation time. In B2, the higher pH value (5.59 ± 0.03) 212
was observed at 120 h, while B1 showed a higher pH (5.60 ± 0.51) at 144 h (Figure 1). 213
Although during the fermentation acetic acid is also being produced, this occurs within the 214
bean cotyledon, having a minimal impact on the pH of the pulp (Camu et al., 2008a). 215
216
3.2. Microbial community dynamics during fermentation processing. 217
The evolution of colony-forming units (CFU) of yeasts, LAB, AAB, and spore-218
forming bacteria throughout fermentation for all experiments is shown in Figure 2. At the 219
beginning of the fermentation, both treatments showed a high content of yeasts. The initial 220
average yeast counts were between 6.71−7.28 log CFU/g, constituted by Hanseniaspora 221
opuntiae, and Pichia manshurica. The initial yeast count was higher than reported by 222
Papalexandratou et al. (2019) at the beginning of the fermentation of Nicaraguan Criollo 223
cocoa beans (6.00−7.50 log CFU/g). Among the most frequently isolated yeasts at the 224

11
beginning of cocoa fermentation were H. opuntiae and P. manschurica, possibly due to its 225
tolerance capacity of low pH by the high-citrate concentration (Meersman et al., 2013; 226
Papalexandratou et al., 2019). In the consecutive fermentation stages, the microbial 227
community was characterized per pronounced differences between treatments. In the case 228
of the yeast community, the fermentation with B2 treatment (with initial turning at 48 h) 229
favored a higher growth compared to the B1 treatment. The B2 treatment was characterized 230
by a greater predominance and yeast biodiversity constituted by H. opuntiae, P. 231
manshurica, and Candida carpophila, from 24-120 h (Figure 2B). While in B1 only P. 232
manshurica is maintained until 120 h, H. opuntiae growth only covered the period of 0-72 233
h, and C. carpophila, was not detected (Figure 2A). The yeasts play an important role at the 234
beginning of the cocoa bean fermentation for the reduction of pectin in sugars for the 235
production of ethanol, decreasing the viscosity of the mass and causing cocoa pulp drainage 236
at the bottom of the fermentation box (De Vuyst & Weckx, 2016; Garcia-Armisen et al., 237
2010). Prolonging the turning start time of the cocoa-bean mass could have favored an 238
anaerobic environment and a higher amount of substrate available for the growth and 239
biodiversification of yeasts community (Figueroa-Hernández et al., 2019; Koné et al., 2016; 240
Serra et al., 2019). Also, the rapid rise in temperature above 45 °C in treatment B1 (Figure 241
1) could cause growth inhibition of some species of yeast compared to B2 in which the 242
temperature peak was reached at a later time (Daniel et al., 2009). The growth of 243
Kodamaea ohmeri was in the last stages of B1 fermentation (Figure 2A), where the yeasts 244
H. opuntiae and P. manshurica are no longer detectable, which was not identified in B2. 245
This can be associated with the high concentration of acetic acid that acts as a yeast growth 246

12
inhibitor, to which K. ohmeri has demonstrated resistance, taking advantage of the sugars 247
available in the medium (Sharma et al., 2018). 248
249
After the yeast domain, the subsequent bacterial community appears after 48 hours 250
for both treatments, but with different dynamics and biodiversity. In treatment B1 (Figure 251
1A), the bacterial community begins with the predominance of Lactic Acid Bacteria (LAB), 252
such as Lactobacillus plantarum and Pedioccoccus acidilactici, Acetic Acid Bacteria 253
(AAB) such as Acetobacter pasteurianus, and spore-forming bacteria such as Bacillus 254
subtilis. While in treatment B2 (Figure 2B), the microbial community was dominated by 255
Lactobacillus plantarum at 48 h, followed by Acetobacter pasteurianus after 72 h, and 256
finally Bacillus subtilis at the end of fermentation. Pedioccoccus acidilactici was not 257
identified in B2. BAL and AAB constitute the microbial groups that dominate the typical 258
fermentative process of cocoa after yeasts. Lactobacillus plantarum is the main LAB 259
identified in cocoa fermentation and is involved in the assimilation of glucose, citric acid, 260
and fructose for the production of ethanol, lactic acid, and mannitol (Nielsen et al., 2013). 261
Acetobacter pasteurianus is an AAB that participates in the ethanol oxidation in acetic acid 262
(Illeghems et al., 2015; Ramos et al., 2020; Saltini et al., 2013). And Bacillus spp. is 263
involved in depectinization of the cocoa-pulp increasing the permeability of the beans that 264
connecting the reactions occurring in the outer part of the bean (by microbial activity) 265
triggering reactions inside (Camu et al., 2008b; Ouattara et al., 2017). A. pasteurianus and 266
Bacillus spp. growth is favored under aerobic conditions and high temperature (Camu, 267
González, et al., 2008b; Hamdouche et al., 2019; Illeghems et al., 2015; Ouattara et al., 268
2017; Saltini et al., 2013). Therefore, the greater oxygenation incorporated in treatment B1 269

13
may have rapidly stimulated the growth of AAB and Bacillus spp. compared to treatment 270
B2. An antagonistic interaction has been reported in the growth of co-cultures of yeast 271
strains (Pichia spp.) and Bacillus subtilis in simulated cocoa-pulp media (Ouattara et al., 272
2020), which could explain the lower growth of the Bacillus genus in the B2 treatment in 273
which yeasts dominate fermentation. A. pasteurianus is a competitive species in an acid- 274
and ethanol-rich environment that favors to poor or no growth of L. plantarum (Camu et al., 275
2008b), which explains the decrease in LAB in later stages in which the maximum peak of 276
AAB appears in B1 treatment (Figure 2A), compared to B2 in which the AAB peak appears 277
after 48h, favoring a greater LAB growth in the final fermentation stages (Figure 2B). And 278
Pediococcus acidilactici are other LAB identified on cocoa fermentation, and its 279
inoculation in the B1 treatment could have been done since the direct environment with the 280
greater turning applied (Camu et al., 2008b; Miguel et al., 2017). 281
282
3.3. Volatile compounds profile 283
Seventy volatile compounds were identified during the cocoa fermentation 284
treatments evaluated. The volatile compounds were classified into six chemical classes: 285
volatile acids, alcohols, aldehydes, esters, ketones, and pyrazines. The percentage of 286
individual relative abundance for each compound at different fermentation times and the 287
associated aroma descriptor are presented in Table 1. 288
289
3.3.1. Volatile acids 290
Acetic acid, propanoic acid, 3-methylbutanoic and 2-methylpropanoic acid were the 291
main volatile acids identified in the cocoa fermentations studied (Table 1). Volatile acids 292

14
represented between 46.72-48.16% of the final volatile profile in both treatments. Acetic 293
acid, 3-methylbutanoic and 2-methylpropanoic are recognized as cocoa key-aroma markers 294
and are responsible for imparting a sour, rancid, and intense vinegar-like perception that 295
affects cocoa aroma quality (Magagna et al., 2017). The highest production of volatile acids 296
was observed in the B2 treatment. Possibly due to the increased growth of alcohol-297
producing yeasts due AAB are able to convert alcohol by dehydrogenation to acids such as 298
propanoic, and 2-methylpropanoic, and by the degradation of citrate mediated by LAB 299
(Ramos et al., 2020). 300
301
3.3.2. Alcohols 302
Twenty-one alcohols were identified during fermentations evaluated. Phenylethyl 303
alcohol, ethanol, 2-pentanol, 2-methyl-1-propanol, 2-pentanol, and 3-methyl-1-butanol 304
were the main. The final profile and percentage of alcohol between treatments was 305
different. The final profile of the B1 treatment was characterized by a higher content of 3-306
methyl-1-butanol, 2-ethyl-1-hexanol, guaiacol, and 2-phenylethyl alcohol. And treatment 307
B2 was characterized by a higher content of 2-pentanol, 2,3-butanediol, 4,5-octanediol, and 308
benzyl alcohol. The B1 treatment showed a final volatile profile with a higher percentage of 309
alcohol compared to the B2 treatment. High alcohol contents are desirable to obtain cocoa 310
products with flowery and candy notes (Rodriguez-Campos et al., 2012), due to alcohols 311
such as 2-phenylethyl alcohol is recognized as cocoa key-aroma marker related to the 312
honey-like aroma characteristic of Criollo cocoa (Cevallos-Cevallos et al., 2018), and 2,3-313
butanediol that is associated with a sweet chocolate odor descriptor (Moreira et al., 2017). 314
Yeasts such as H. opuntiae have been associated with 2-phenylethyl alcohol production 315

15
(Hu et al., 2018). In addition to the production of alcohols, the yeasts also participate in 316
their esterification, o alcoholic compounds are oxidized to ester with the presence of 317
oxygen by LAB (Cevallos-Cevallos et al., 2018; Hamdouche et al., 2019; Schlüter et al., 318
2020). So that the greater growth of yeasts in the fermentation B2 could have favored a 319
greater conversion of these alcohols into their derived esters. 320
321
3.3.3. Aldehydes 322
Twelve aldehydes were identified during fermentations evaluated, constituted by 3-323
methylbutanal, 2-methylbutanal, butanedial, 2-methylpropanal, 2-methylpentanal, nonanal, 324
benzaldehyde, phenylacetaldehyde, and benzenebutanal. The percentage of aldehydes in the 325
final volatile profile was similar between the treatments (around 25.91-30.85%). The final 326
profile of treatment B1 was characterized by phenylacetaldehyde, nonanal, and pentanal. 327
While the B2 profile was characterized by benzaldehyde, 2-methylpropanal, butanedial, 2-328
methylbutanal, and 3-methylbutanal. Phenylacetaldehyde and benzaldehyde are odor-active 329
cocoa aldehydes related to honey- and almond-like aroma descriptors (Hinneh et al., 2018), 330
and 3-methylbutanal is cataloged as a cocoa key-aroma marker associated with a chocolate 331
odor (Magagna et al., 2017). Therefore, high aldehyde content is favorable for the quality 332
of cocoa (Menezes et al., 2016). The aldehydes are produced from the oxidation of primary 333
alcohols produced by yeasts, and from amino acids such as leucine, isoleucine (for 3-334
methylbutanal) and phenylalanine (for benzaldehyde and phenylacetaldehyde) mainly by 335
enzymatic activity of LAB, whose activity is favored when the temperature increased and 336
reached its maximum (Hamdouche et al., 2019). 337
338

16
3.3.4. Esters 339
Twenty-one esters compounds were identified, represented by ethyl acetate, 340
isobutyl acetate, 2-pentanol acetate, ethyl octanoate, ethyl benzeneacetate, 2-phenylethyl 341
acetate, and ethyl palmitate, among others. At the end of fermentation, B2 presented a 342
volatile profile with a higher percentage and a number of esters compared to B1 that only 343
presented 2-phenylethyl acetate. The esters are important compounds in the profile of cocoa 344
products, and characteristic of products derived from Criollo cocoa, due to their attractive 345
sweet, flowery, and fruity aroma descriptors, such as 2-phenylethyl acetate associated with 346
perception like honey, and for which it is recognized as a key-aroma marker of cocoa 347
(Koné et al., 2016; Magagna et al., 2017; Moreira et al., 2018). The esters are produced 348
from the esterification of alcohols mediated by microorganisms such as yeasts, and some 349
LAB (Cevallos-Cevallos et al., 2018; Moreira et al., 2018; Ouattara et al., 2020). Yeasts 350
such as H. opuntiae have been associated with 2-phenylethyl acetate, and ethyl hexanoate 351
production (Hu et al., 2018). Esters are mostly produced by synergy of yeasts and LAB 352
during the anaerobic stage of fermentation (da Silva Vale et al., 2019; Hamdouche et al., 353
2019). The results could indicate the greater production of esters in B2 due to the lower 354
aeration of cocoa bean-mass compared to B1. 355
356
3.3.5. Ketones 357
Nine ketones represented by 4-hydroxy-2-butanone, 2,3-butanedione, 3-methyl-2-358
butanone, 4-methyl-2-hexanone, 3-hydroxy-2-butanone, 2-nonanone, and acetophenone 359
were identified in the fermentations evaluated. A higher percentage of ketones were 360
observed in the early stages of fermentation, acetophenone principally. At the end of the 361

17
fermentation, the B2 treatment presented a higher percentage and number of ketones, 362
highlighting the presence of 2,3-butanedione, following by 3-hydroxy-2-butanone, and 363
acetophenone, compare to B1 treatment. Ketones such as 2,3-butanedione, and 3-hydroxy-364
2-butanone are recognized technological markers of cocoa processing and are important 365
contributors to the typical buttery and creamy aroma of chocolate (Magagna et al., 2017). 366
Ketones are compounds naturally present in cocoa, and their content decreases throughout 367
fermentation by degradation or microbial catabolism (Hamdouche et al., 2019). However, 368
some ketones such as acetophenone are produced by H. opuntiae (Hu et al., 2018), and 3-369
hydroxy-2-butanone can be produced by bacteria of the genus Bacillus spp. favoring its 370
increase in anaerobic conditions and high temperatures (Ouattara et al., 2020). 371
372
3.3.6. Pyrazines 373
Only the presence of 2,5-dimethylpyrazine was detected in the last stages of both 374
fermentations, observing a slightly higher production in the B1 treatment (0.51±0.04) 375
compared to B2 (0.28±0.01). Pyrazines in cocoa beans are formed only during the roasting 376
process. However, some methylpyrazines can originate during fermentation due to the 377
enzymatic activity of Bacillus spp. (Koné et al., 2016). Therefore, the higher production of 378
2,5-dimethylpyrazine could be associated with the greater growth of Bacillus during B1 379
fermentation. 380
381
3.4. Multivariate analysis of the volatile compounds produced during the cocoa 382
fermentations 383

18
The principal component analysis (PCA) was performed to evaluate the effect of the 384
first turning time on the growth dynamics of the main microorganisms (yeasts, BAL, AAB 385
and spore-forming bacteria) and volatile compounds generated during cocoa fermentation. 386
Figure 3 shows the first two Principal Components (PC), explaining together 56.15% of the 387
total variance (30.80% for PC1 and 25.35% for PC2). The PCA allowed a natural grouping 388
of the observations separating the initial time, intermediate times and final time of both 389
fermentations in different quadrants of the biplot. PC2 separated the initial fermentation 390
times in the bottom of the plot (with negative scoring) mainly related to 2-heptanol, 2-391
heptanone, 2-hydroxyacetophenone, butanal, and the microorganisms H. opuntiae and B. 392
subtilis. While the fermentation time of 72 h, and the final time (144 h) were placed in the 393
top of the biplot, with positive scoring of PC2, related to 2-methylpropanal, 2,3-394
butanedione, 3-methyl-2-butanol, 4-methyl-2,3-pentanediol, 4,5-octanediol, phenylalcohol, 395
methyl-2-methoxy-butanoate, 2-methylpropanoic acid, 3-methylbutanoic acid, and 2,5-396
dimethylpyrazine. The PC1 allowed separating the final profile of each treatment. The B1 397
treatment was located in the upper quadrant of the plot with negative scoring related to 398
ethyl acetate and Pediococcus acidilactici. And the B2 treatment was located with positive 399
scoring, mainly related to butanal, nonanal, phenylacetaldehyde, acetophenone, 2-400
heptanone, 4-methyl-2-hexanone, 2-phenylethyl alcohol, 2-pentanol, 2,3-butanediol, 2-401
heptanol, and ethyl octanoate, as well as the microorganisms H. opuntiae and B. subtilis. 402
403
Subsequently, supervised classification was carried out. Partial Least Square 404
Discriminant Analysis (PLS-DA) model was constructed to classify volatile compounds 405
generated in fermentation with the type of treatment used, in order to maximize the 406

19
separation between classes, and find the most relevant compounds for each turning 407
treatment (B1 and B2). PLS-DA scores biplot of volatile compounds is shown in Figure 4. 408
Seventeen compounds such as 3-metilpentanoic acid, 2-nonanone, 3-methyl-2-butanone, 409
acetophenone, 4-hydroxy-2-butanone, phenyleacetaldehyde, benzaldehyde, 3-410
methylbutanal, 1-phenylethyl acetate, 2-pentanol acetate, 2,3-butanediol diacetate, ethyl 411
hexanoate, 2-hydroxy-ethyl propanoate, ethyl decanoate, 2-phenylethyl alcohol, 2-412
pentadecanol, and 2-decanol were highly correlated with the B2 treatment, for being placed 413
farther from the center, particularly between the inner and outer circles in the biplot. PLS-414
DA allows to visualize that the compounds associated as key-aroma markers of cocoa, such 415
as phenylacetaldehyde, 3-methylbutanal, 2-phenylethyl alcohol, 2-phenylethyl acetate, 2,5-416
dimethylpyrazine, 3-methylbutanoic acid, 2-methylpropanoic acid, and 2,3-butanedione are 417
mostly correlated with the B2 treatment (with initial turning at 48 h). While B1 (with initial 418
turning at 24 h) showed a higher correlation with the key-aroma marker 3-hydroxy-2-419
butanone. 420
421
The results can be associated with the fact that the greater aeration incorporated into 422
the cocoa-bean mass in the B1 treatment caused an aerobic environment that caused the 423
decline of yeasts and the increase of AAB and spore-forming bacteria of the genus Bacillus 424
spp (Guehi et al., 2010; Hamdouche et al., 2019; Ouattara et al., 2020). The latter microbial 425
genus has been reported as a producer of 3-hydroxy-2-butanone in simulated cocoa pulp 426
media (Ouattara et al., 2020). In addition, Pediococcus acidilactici was identified in 427
treatment B1, which in co-inoculation with yeasts of the genus Pichia spp. has shown a 428
higher production of ethyl acetate than in isolation (da Silva-Vale et al., 2019). 429

20
430
On the other hand, the B2 treatment, by prolonging the anaerobic conditions 431
stimulated the growth of yeasts and LAB that favored its total dominance during the whole 432
fermentation. A higher production of acetate esters has been associated with greater 433
synergy between co-inoculation of yeasts and LAB (da Silva-Vale et al., 2019). Some yeast 434
isolated from cocoa fermentation, such as H. opuntiae and Pichia spp., have been 435
recognized as producers of ethanol, 2-phenylethyl alcohol, esters (such as 2-phenylethyl 436
acetate, ethyl hexanoate, and 2-pentanol acetate), and aldehydes (such as 3-methyl butanal, 437
and phenylacetaldehyde), and acetophenone (Cevallos-Cevallos et al., 2018; Hu et al., 438
2018; Ouattara et al., 2020). Subsequently, by incorporating aeration into the cocoa-pulp, 439
aerobic conditions favored the growth of AAB, which from the alcohols generated by 440
yeasts, produced a higher content of acids such as propanoic acid, 3-methylpropanoic acid, 441
and 3-methyl butanoic acid (Ramos et al., 2020). All the aforementioned compounds were 442
co-correlated with the B2 treatment. 443
444
Conclusion 445
The effect of two traditional methods of fermentation of Mexican Criollo cocoa 446
with different start time of turning the cocoa-pulp mass on the microbial dynamics and 447
volatile compounds generated using multivariate analysis was studied. The multivariate 448
analysis indicate that 3-metilpentanoic acid, 2-nonanone, 3-methyl-2-butanone, 449
acetophenone, 4-hydroxy-2-butanone, phenyleacetaldehyde, benzaldehyde, 3-450
methylbutanal, 1-phenylethyl acetate, 2-pentanol acetate, 2,3-butanediol diacetate, ethyl 451
hexanoate, 2-hydroxy-ethyl propanoate, ethyl decanoate, 2-phenylethyl alcohol, 2-452

21
pentadecanol, and 2-decanol are volatile compounds are related to the start time of the 453
cocoa-bean mass turning. The results indicate that the production of volatile compounds 454
during cocoa fermentation, rather than being produced by specific microbial taxa, is closely 455
related to a synergism and/or antagonistic mechanism between yeasts, LAB, AAB and 456
spore-forming bacteria. The turning start time at 48h stimulated a microbial profile with 457
yeast domain that favors the production of compounds such as acetate esters, aldehydes 458
(such as 2-phenylacetaldehyde and 3-methylbutanal), alcohols (2-phenylethyl alcohol), 459
acetophenone, and increased production of volatile acids recognized as key-aroma markers 460
that are associated with cocoa quality. While an immediate turning start favors a mostly 461
aerobic environment that generates a decline of yeasts stimulating a rapid growth of LAB, 462
AAB and spore-forming bacteria of the genus Bacillus spp. which favor a greater 463
production of ethyl acetate mainly. 464
465
The fermentation methods evaluated did not change the fermentation time, but the 466
volatile compounds profile. These results offer Mexican Criollo cocoa producers an 467
overview of the different volatile compound profiles as a tool for the selection of the 468
fermentative technique. This suggests deepen the changes in the aroma profile on the 469
improvement of Mexican Criollo cocoa quality using regional artisanal fermentation 470
methods, and generate a greater understanding of the interaction of the generated microbial 471
profile associated with the genotype and origin of the cocoa bean. 472
473
Acknowledgments 474

22
The authors acknowledge the support of the SAGARPA Conacyt grant (project number: 475
2017-02-291417). 476
References 477
Camu, N., De Winter, T., Addo, S. K., Takrama, J. S., Bernaert, H., & De Vuyst, L. (2008). 478
Fermentation of cocoa beans: influence of microbial activities and polyphenol 479
concentrations on the flavour of chocolate. Journal of the Science of Food and 480
Agriculture, 88(13), 2288–2297. 481
Camu, N., González, Á., De Winter, T., Van Schoor, A., De Bruyne, K., Vandamme, P., 482
Takrama, J. S., Addo, S. K., & De Vuyst, L. (2008). Influence of turning and 483
environmental contamination on the dynamics of populations of lactic acid and acetic 484
acid bacteria involved in spontaneous cocoa bean heap fermentation in Ghana. Applied 485
and Environmental Microbiology, 74(1), 86–98. 486
Cevallos-Cevallos, J. M., Gysel, L., Maridueña-Zavala, M. G., & Molina-Miranda, M. J. 487
(2018). Time-related changes in volatile compounds during fermentation of bulk and 488
fine-flavor cocoa (Theobroma cacao) Beans. Journal of Food Quality, 2018. 489
Crafack, M., Keul, H., Eskildsen, C. E., Petersen, M. A., Saerens, S., Blennow, A., 490
Skovmand-Larsen, M., Swiegers, J. H., Petersen, G. B., Heimdal, H., & Nielsen, D. S. 491
(2014). Impact of starter cultures and fermentation techniques on the volatile aroma 492
and sensory profile of chocolate. Food Research International, 63, 306–316. 493
da Silva Vale, A., de Melo Pereira, G. V., de Carvalho Neto, D. P., Rodrigues, C., 494
Pagnoncelli, M. G. B., & Soccol, C. R. (2019). Effect of Co-Inoculation with Pichia 495
fermentans and Pediococcus acidilactici on metabolite produced during fermentation 496
and volatile composition of coffee beans. Fermentation, 5(3), 1–17. 497

23
Daniel, H. M., Vrancken, G., Takrama, J. F., Camu, N., De Vos, P., & De Vuyst, L. (2009). 498
Yeast diversity of Ghanaian cocoa bean heap fermentations. FEMS Yeast Research, 499
9(5), 774–783. 500
De Vuyst, L., & Weckx, S. (2016). The cocoa bean fermentation process: from ecosystem 501
analysis to starter culture development. Journal of Applied Microbiology, 121(1), 5–502
17. 503
Figueroa-Hernández, C., Mota-Gutierrez, J., Ferrocino, I., Hernández-Estrada, Z. J., 504
González-Ríos, O., Cocolin, L., & Suárez-Quiroz, M. L. (2019). The challenges and 505
perspectives of the selection of starter cultures for fermented cocoa beans. 506
International Journal of Food Microbiology, 301(January), 41–50. 507
Garcia-Armisen, T., Papalexandratou, Z., Hendryckx, H., Camu, N., Vrancken, G., De 508
Vuyst, L., & Cornelis, P. (2010). Diversity of the total bacterial community associated 509
with Ghanaian and Brazilian cocoa bean fermentation samples as revealed by a 16 S 510
rRNA gene clone library. Applied Microbiology and Biotechnology, 87(6), 2281–511
2292. 512
Guehi, S. T., Dabonne, S., Ban-Koffi, L., Kedjebo, D. K., & Zahouli, G. I. B. (2010). Effect 513
of turning beans and fermentation method on the acidity and physical quality of raw 514
cocoa beans. Advance Journal of Food Science and Technology, 2(3), 163–171. 515
Hamdouche, Y., Meile, J. C., Lebrun, M., Guehi, T., Boulanger, R., Teyssier, C., & Montet, 516
D. (2019). Impact of turning, pod storage and fermentation time on microbial ecology 517
and volatile composition of cocoa beans. Food Research International, 119(January), 518
477–491. 519
Hinneh, M., Semanhyia, E., Van de Walle, D., De Winne, A., Tzompa-Sosa, D. A., 520

24
Scalone, G. L. L., de Meulenaer, B., Messens, K., Van Durme, J., Afoakwa, E. O., De 521
Cooman, L., & Dewettinck, K. (2018). Assessing the influence of pod storage on 522
sugar and free amino acid profiles and the implications on some Maillard reaction 523
related flavor volatiles in Forastero cocoa beans. Food Research International, 524
111(May), 607–620. 525
Hu, L., Wang, J., Ji, X., Liu, R., Chen, F., & Zhang, X. (2018). Selection of non-526
Saccharomyces yeasts for orange wine fermentation based on their enological traits 527
and volatile compounds formation. Journal of Food Science and Technology, 55(10), 528
4001–4012. 529
Illeghems, K., Weckx, S., & De Vuyst, L. (2015). Applying meta-pathway analyses 530
through metagenomics to identify the functional properties of the major bacterial 531
communities of a single spontaneous cocoa bean fermentation process sample. Food 532
Microbiology, 50, 54–63. 533
Koné, M. K., Guéhi, S. T., Durand, N., Ban-Koffi, L., Berthiot, L., Tachon, A. F., Brou, K., 534
Boulanger, R, & Montet, D. (2016). Contribution of predominant yeasts to the 535
occurrence of aroma compounds during cocoa bean fermentation. Food Research 536
International, 89, 910–917. 537
Kongor, J. E., Hinneh, M., de Walle, D. Van, Afoakwa, E. O., Boeckx, P., & Dewettinck, 538
K. (2016). Factors influencing quality variation in cocoa (Theobroma cacao) bean 539
flavour profile - A review. Food Research International, 82, 44–52. 540
Lagunes Gálvez, S., Loiseau, G., Paredes, J. L., Barel, M., & Guiraud, J. P. (2007). Study 541
on the microflora and biochemistry of cocoa fermentation in the Dominican Republic. 542
International Journal of Food Microbiology, 114(1), 124–130. 543

25
Magagna, F., Guglielmetti, A., Liberto, E., Reichenbach, S. E., Allegrucci, E., Gobino, G., 544
Bicche, C., & Cordero, C. (2017). Comprehensive Chemical Fingerprinting of High-545
Quality Cocoa at Early Stages of Processing: Effectiveness of Combined Untargeted 546
and Targeted Approaches for Classification and Discrimination. Journal of 547
Agricultural and Food Chemistry, 65(30), 6329–6341. 548
Meersman, E., Steensels, J., Mathawan, M., Wittocx, P. J., Saels, V., Struyf, N., Bernaert, 549
H., Vrancken, G., & Verstrepen, K. J. (2013). Detailed analysis of the microbial 550
population in Malaysian spontaneous cocoa pulp fermentations reveals a core and 551
variable microbiota. PLoS ONE, 8(12). 552
Menezes, A. G. T., Batista, N. N., Ramos, C. L., de Andrade e Silva, A. R., Efraim, P., 553
Pinheiro, A. C. M., & Schwan, R. F. (2016). Investigation of chocolate produced from 554
four different Brazilian varieties of cocoa (Theobroma cacao L.) inoculated with 555
Saccharomyces cerevisiae. Food Research International, 81, 83–90. 556
Miguel, M. G. da C. P., Reis, L. V. de C., Efraim, P., Santos, C., Lima, N., & Schwan, R. F. 557
(2017). Cocoa fermentation: Microbial identification by MALDI-TOF MS, and 558
sensory evaluation of produced chocolate. LWT - Food Science and Technology, 77, 559
362–369. 560
Moreira, I., de Figueiredo Vilela, L., da Cruz Pedroso Miguel, M. G., Santos, C., Lima, N., 561
& Freitas Schwan, R. (2017). Impact of a Microbial Cocktail Used as a Starter Culture 562
on Cocoa Fermentation and Chocolate Flavor. Molecules (Basel, Switzerland), 22(5). 563
Moreira, I. M. da V., Miguel, M. G. da C. P., Duarte, W. F., Dias, D. R., & Schwan, R. F. 564
(2013). Microbial succession and the dynamics of metabolites and sugars during the 565
fermentation of three different cocoa (Theobroma cacao L.) hybrids. Food Research 566

26
International, 54(1), 9–17. 567
Moreira, I. M. da V., Vilela, L. de F., Santos, C., Lima, N., & Schwan, R. F. (2018). 568
Volatile compounds and protein profiles analyses of fermented cocoa beans and 569
chocolates from different hybrids cultivated in Brazil. Food Research International, 570
109(September 2017), 196–203. 571
Nielsen, D. S., Crafack, M., Jespersen, L., & Jakobsen, M. (2013). The microbiology of 572
cocoa fermentation. Chocolate in Health and Nutrition. 573
Ouattara, H. G., Elias, R. J., & Dudley, E. G. (2020). Microbial synergy between Pichia 574
kudriazevii YS201 and Bacillus subtilis BS38 improves pulp degradation and aroma 575
production in cocoa pulp simulation medium. Heliyon, 6(1). 576
Ouattara, H. G., Reverchon, S., Niamke, S. L., & Nasser, W. (2017). Regulation of the 577
synthesis of pulp degrading enzymes in Bacillus isolated from cocoa fermentation. 578
Food Microbiology, 63, 255–262. 579
Papalexandratou, Z., Kaasik, K., Kauffmann, L. V., Skorstengaard, A., Bouillon, G., 580
Espensen, J. L., Hansen, L. H., Jakobsen, R. R., Blennow, A., Krych, L., Castro-Mejía, 581
J., & Nielsen, D. S. (2019). Linking cocoa varietals and microbial diversity of 582
Nicaraguan fine cocoa bean fermentations and their impact on final cocoa quality 583
appreciation. International Journal of Food Microbiology, 304(December 2018), 106–584
118. 585
Ramos, S., Salazar, M., Nascimento, L., Carazzolle, M., Pereira, G., Delforno, T., 586
Nascimento, M., de Aleluia, T., Celeghini, R., & Efraim, P. (2020). Influence of pulp 587
on the microbial diversity during cupuassu fermentation. International Journal of 588
Food Microbiology, 318(November 2019), 108465. 589

27
Rodriguez-Campos, J., Escalona-Buendía, H. B., Contreras-Ramos, S. M., Orozco-Avila, 590
I., Jaramillo-Flores, E., & Lugo-Cervantes, E. (2012). Effect of fermentation time and 591
drying temperature on volatile compounds in cocoa. Food Chemistry, 132(1), 277–592
288. 593
Rodriguez-Campos, J., Escalona-Buendía, H. B., Orozco-Avila, I., Lugo-Cervantes, E., & 594
Jaramillo-Flores, M. E. (2011). Dynamics of volatile and non-volatile compounds in 595
cocoa (Theobroma cacao L.) during fermentation and drying processes using principal 596
components analysis. Food Research International, 44(1), 250–258. 597
Saltini, R., Akkerman, R., & Frosch, S. (2013). Optimizing chocolate production through 598
traceability: A review of the influence of farming practices on cocoa bean quality. 599
Food Control, 29(1), 167–187. 600
Santander Muñoz, M., Rodríguez Cortina, J., Vaillant, F. E., & Escobar Parra, S. (2019). 601
An overview of the physical and biochemical transformation of cocoa seeds to beans 602
and to chocolate: Flavor formation. Critical Reviews in Food Science and Nutrition, 603
8398, 1–21. 604
Schlüter, A., Hühn, T., Kneubühl, M., Chatelain, K., Rohn, S., & Chetschik, I. (2020). 605
Novel time- and location-independent postharvest treatment of cocoa beans: 606
investigations on the aroma formation during “moist Incubation” of unfermented and 607
dried cocoa nibs and comparison to traditional fermentation. Journal of Agricultural 608
and Food Chemistry. 609
Schwan, R. F., & Wheals, A. E. (2004). The microbiology of cocoa fermentation and its 610
role in chocolate quality. Critical Reviews in Food Science and Nutrition, 44(4), 205–611
221. 612

28
Serra, J. L., Moura, F. G., Pereira, G. V. d. M., Soccol, C. R., Rogez, H., & Darnet, S. 613
(2019). Determination of the microbial community in Amazonian cocoa bean 614
fermentation by Illumina-based metagenomic sequencing. Lwt, 106(July 2018), 229–615
239. 616
Sharma, S., Arora, A., Sharma, P., Singh, S., Nain, L., & Paul, D. (2018). Notable mixed 617
substrate fermentation by native Kodamaea ohmeri strains isolated from Lagenaria 618
siceraria flowers and ethanol production on paddy straw hydrolysates. Chemistry 619
Central Journal, 12(1), 1–11. 620
SIAP. (201 ). Sistema de Informaci n Agr cola y Pesquera. Secretar a de Agricultura 621
anader a esarrollo ural Pesca y Alimentaci n. 622
623
624
Figure Captions 625
Figure 1. The pH, soluble solids (ºBx), environmental temperature, and box temperature 626
values at different fermentation times (0, 24, 48, 72, 96, 120, and 144 hours) of both 627
fermentation methods evaluated: B1) Fermentation of fresh cocoa beans with first turning 628
at 24 h; and B2) with first turning at 48 h. The pH and ºBx are displayed in bars with values 629
corresponding to the scale on the left side. And the box and environmental temperature 630
values are shown in lines with scale values from the right side. Different letters between pH 631
and ºBx values indicate significant differences (p<0.05). 632
633
Figure 2. Relative abundance expressed on a logaritmic scale of yeast community (left 634
side) and bacterial community (right side) at different fermentation times (0, 24, 48, 72, 96, 635

29
120, and 144 h) of both fermentation methods studied: A) Fermentation of fresh cocoa 636
beans and with first turning at 24 h; and B) with first turning at 48 h. 637
638
Figure 3. Principal Component Analysis (PCA) of the overall volatile profile and CFU of 639
the main microorganism at different fermentation times (0, 48, 72, 96, 120, and 144 h) on 640
both process evaluated (B1=green, and B2=blue). Values in parenthesis represent the 641
percentage of variance explained by each component. 642
643
Figure 4. Partial least square discriminant analysis (PLS-DA) score biplot of the volatile 644
compounds distribution according the fermentative treatments (B1=green, and B2=blue) at 645
different fermentation times (0, 48, 72, 96, 120, and 144 h). 646
647
648

Table 1. Volatile compounds identified by HS-SPME/GC-MS in the treatments B1 and B2 at the different fermentation times (0, 24,
48, 72, 96, 120, and 144 h).
Rt Compound name Odor descriptor*
Percentage (%)a per fermentation time
B1
B2
0 24 48 72 96 120 144 0 48 72 96 120 144
Acids
18.672 Acetic acid Sour, vinegary 1.90±0.09 6.43±0.04 51.48±8.72 58.12±4.58 69.70±8.91 64.28±0.52 35.85±6.40 1.41±0.04 45.27±2.13 17.46±1.33 65.98±1.36 86.07±14.0
1
35.86±3.28
19.928 Propanoic acid Fruity, pungent 0.00 0.00 0.00 0.15±0.01 0.20±0.02 0.00 0.66±0.02 0.00 0.00 0.00 0.00 0.00 0.42±0.02
20.294 2-Methylpropanoic acid Rancid 0.00 0.00 0.00 0.29±0.01 2.84±0.15 1.94±0.16 3.14±0.46 0.00 0.00 0.29±0.01 0.77±0.01 0.83±0.01 3.15±0.07
20.959 Propanedioic acid - 0.00 0.00 0.00 0.00 0.00 0.08±0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00
21.571 3-methylbutanoic acid Rancid 0.00 0.00 0.00 0.00 0.00 0.00 7.07±0.56 0.00 0.00 0.00 0.00 0.00 8.74±0.21
21.582 3-methylpentanoic acid - 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.57±0.04 0.00 0.00 0.00
total percentage 1.90±0.09 6.43±0.04 51.48±8.72 58.55±4.52 72.73±9.07 66.30±0.66 46.72±6.82 1.41±0.04 45.27±3.74 18.42±1.34 66.75±1.37 86.90±14.0
2
48.16±3.37
Alcohols
9.195 Ethanol Ethanol-like 16.88±0.74 0.00 6.24±0.01 12.95±0.69 0.00 0.00 0.00 32.89±1.99 9.94±0.72 0.00 0.00 0.21±0.03 0.00
12.59 2-Methyl-1-propanol Wine 0.96±0.04 0.00 0.00 0.19±0.00 0.00 0.08±0.01 0.10±0.00 0.00 0.00 0.23±0.04 0.00 0.00 0.08±0.00
13.134 2-Pentanol Light, seedy, sharp 2.85±0.20 0.00 0.00 0.00 0.67±0.06 0.00 0.20±0.02 6.88±0.51 0.87±0.00 1.20±0.21 0.00 0.25±0.04 0.49±0.01
14.785 3-Methyl-1-butanol Malty, chocolate 3.91±0.27 11.49±1.25 0.00 0.57±0.11 0.00 0.00 0.62±0.02 0.00 0.00 1.94±0.08 0.00 0.00 0.16±0.00
14.826 1-Pentanol Sweet, pungent 1.48±0.11 0.00 1.18±0.13 0.00 0.96±0.02 0.67±0.06 0.00 2.88±0.35 1.31±0.24 0.00 0.19±0.02 0.32±0.02 0.34±0.01
14.965 2-Methyl-5-hexen-3-ol - 0.00 0.00 0.00 0.48±0.02 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00
16.611 2,3-Butanediol - 0.62±0.03 0.00 0.27±0.01 0.00 0.58±0.00 0.33±0.00 0.00 1.20±0.08 0.44±0.07 0.00 0.14±0.01 0.15±0.00 0.17±0.00
16.621 2-Heptanol Citrusy, fresh, lemon grass-
like
0.64±0.04 0.00 0.00 0.00 0.00 0.00 0.00 0.98±0.02 0.00 0.00 0.29±0.00 0.00 0.00
16.631 3-Methyl-2-hexanol - 0.00 1.76±0.17 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00
17.164 3-Methyl-2-butanol - 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.08±0.00
17.667 2-Methyl-5-hexen-3-ol - 0.00 0.00 0.00 0.00 0.00 0.00 0.15±0.00 0.00 0.00 0.00 0.00 0.00 0.00
19.134 2-Ethyl-1-hexanol - 1.00±0.05 1.42±0.01 0.00 0.00 0.00 0.00 1.05±0.10 0.00 0.00 0.71±0.07 0.00 0.00 0.00
19.462 2-Decanol - 0.00 1.80±0.11 0.00 0.06±0.00 0.00 0.00 0.34±0.02 0.00 0.00 0.22±0.01 0.00 0.00 0.00
19.472 2-Pentadecanol - 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 1.30±0.11 0.00 0.00 0.00
21.113 4,5-Octanediol - 0.00 0.00 0.00 0.00 0.00 0.02±0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.15±0.00
23.616 1-Phenyl-1-decanol - 0.23±0.01 0.00 0.00 0.00 0.00 0.00 0.00 0.44±0.03 0.00 0.00 0.00 0.00 0.00
23.626 α-Methyl-benzene
methanol
- 0.00 0.26±0.00 0.00 0.22±0.00 0.00 0.00 0.00 0.00 0.00 0.27±0.00 0.00 0.00 0.00
24.488 Guaiacol Spicy 0.00 0.00 0.00 0.00 0.20±0.00 0.16±0.00 2.75±0.26 0.00 0.08±0.00 0.00 1.09±0.05 0.05±0.00 0.78±0.03
24.652 Benzyl alcohol Sweet, fruity 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.23±0.02
25.288 2-Phenylethyl alcohol Honey-like 10.64±0.86 8.82±0.07 1.37±0.09 3.27±0.04 1.72±0.24 1.53±0.06 10.70±0.16 14.58±2.95 1.51±0.00 6.26±0.82 2.75±0.11 1.19±0.06 4.93±0.23
total percentage 39.13±0.83 25.50±1.20 9.06±0.24 17.76±0.77 4.13±0.32 2.78±0.14 15.78±0.58 61.65±3.48 14.15±1.31 12.10±1.12 4.45±0.19 2.17±0.18 7.39±0.29
Table

Table 1. Continue.
Rt Compound name Odor descriptor*
Percentage (%)a per fermentation time
B1 B2
0 24 48 72 96 120 144 0 48 72 96 120 144
Aldehydes
6.867 Butanal - 0.76±0.04 0.59±0.02 0.00 0.06±0.00 0.00 0.00 0.00 01.18±0.04 0.00 0.15±0.01 0.00 0.00 0.00
7.052 2-Methylpropanal Green, pungent 0.00 0.00 0.00 0.00 0.00 0.06±0.01 0.33±0.03 0.00 0.00 0.00 0.00 0.02±0.00 1.16±0.09
8.795 2-Methylbutanal Malty, cocoa, chocolate,
almond-like
0.83±0.14 0.00 0.00 0.00 0.00 0.00 0.40±0.02 5.80±0.82 0.00 0.00 0.00 0.00 0.72±0.02
8.826 2,3-Dimethylpentanal - 0.00 1.39±0.06 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00
8.898 Butanedial - 2.74±0.53 0.00 0.00 0.00 0.00 0.07±0.00 0.00 0.00 5.14±0.07 0.00 0.00 0.00 0.11±0.00
8.918 3-Methylbutanal Malty, cocoa, chocolate 4.23±0.34 2.77±0.50 0.00 0.20±0.01 0.00 1.70±0.18 0.00 5.51±0.17 0.00 35.93±1.59 0.80±0.03 0.00 1.19±0.08
8.997 Pentanal Almond-like, malt 0.00 0.00 0.00 0.00 0.00 0.00 5.36±0.52 0.00 0.00 0.00 0.00 0.00 0.00
17.903 2-Methylpentanal - 0.00 1.35±0.01 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.34±0.01 0.00
17.985 Nonanal Fatty, waxy, pungent 1.27±0.12 1.13±0.13 0.00 0.00 0.00 0.00 0.39±0.02 1.31±0.34 0.00 0.44±0.06 0.00 0.00 0.25±0.01
20.098 Benzaldehyde Almond, burnt sugar, bitter 0.00 0.00 0.00 0.00 0.00 3.48±0.39 1.23±0.09 9.59±1.32 0.00 3.65±0.11 1.80±0.13 0.53±0.00 3.03±0.07
21.595 Phenylacetaldehyde Honey-like 37.61±5.66 38.58±0.65 16.32±2.29 3.16±0.23 14.97±1.37 17.59±1.81 23.14±4.50 0.50±0.02 29.37±0.93 17.20±1.87 22.27±1.77 7.50±0.13 19.46±0.38
23.79 Benzenebutanal - 0.00 0.00 0.00 0.00 0.57±0.01 0.63±0.02 0.00 0.00 0.00 0.00 0.13±0.02 0.61±0.00 0.00
total percentage 47.36±1.80 45.81±0.70 16.32±2.29 3.42±0.24 15.54±1.31 23.54±1.79 30.85±4.66 23.89±1.66 34.51±0.07 57.37±3.54 25.02±1.95 8.99±0.14 25.91±0.65
Esters
7.257 Methyl acetate - 0.00 0.00 0.00 0.12±0.02 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.06±0.00 0.20±0.02
8.241 Ethyl acetate Fruity, pineapple-like 3.39±0.28 0.00 19.79±1.40 10.25±0.46 0.00 0.00 0.00 0.00 2.55±0.26 0.00 0.00 0.09±0.00 0.00
11.102 Isobutyl acetate Fruity 0.00 0.00 0.00 0.24±0.01 0.00 0.00 0.00 0.00 0.00 0.17±0.03 0.00 0.00 0.00
12.262 2-Pentanol acetate Green, fruity 0.00 0.00 2.18±0.10 0.08±0.00 0.00 0.09±0.01 0.00 0.00 1.03±0.04 0.16±0.01 0.00 0.09±0.00 0.16±0.01
15.370 Ethyl hexanoate - 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.14±0.01 0.00 0.00 0.00
17.164 2-Hydroxy-ethyl propanoate - 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.73±0.03 0.00 0.00 0.00
17.855 2,3-Butanediol diacetate - 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.21±0.01 0.00 0.00 0.00
18.41 Ethyl octanoate Fruity, flowery 1.10±0.02 0.00 0.00 0.10±0.00 0.00 0.00 0.00 1.60±0.24 0.00 0.00 0.00 0.00 0.39±0.04
19.462 2-Methoxymethyl-2-
butanoato
- 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.28±0.00 0.00 0.00 0.00 0.61±0.05
21.103 Ethyl decanoate - 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.36±0.02 0.00 0.00 0.00 0.00
21.482 Isopropyl propanoate - 0.00 0.00 0.00 0.00 6.28±0.74 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00
21.493 Isopropyl methanoate - 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.49±0.02 0.00 0.00
22.58 Benzyl-3-hydroxy
propanoate
- 0.11±0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.06±0.00
23.318 1-Phenylethyl acetate Fruity, sweet 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.46±0.01 0.00 0.00 0.25±0.02
23.906 2-Phenylethyl acetate Flowery, honey-like 0.00 0.00 0.00 2.19±0.14 0.00 0.00 1.53±0.07 0.00 0.00 1.73±0.02 0.00 0.00 1.09±0.07
27.141 Isopropyl myristate - 0.00 0.00 0.00 0.51±0.07 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00
32.052 Methyl hexanoate - 0.00 0.00 0.00 0.29±0.03 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00
33.011 Ethyl palmitate Waxy, green 0.46±0.05 0.00 0.00 0.37±0.01 0.00 0.00 0.00 3.51±0.84 0.00 0.37±0.01 0.00 0.00 0.00
48.313 Ethyl linoleate - 0.14±0.02 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00
total percentage 5.20±0.35 0.00 21.97±1.50 14.15±0.71 6.28±0.74 0.09±0.01 1.53±0.07 5.11±1.08 3.86±0.30 4.33±0.08 0.49±0.02 0.25±0.01 2.76±0.21

Table 1. Continue.
Rt Compound name Odor descriptor*
Percentage (%)a per fermentation time
B1 B2
0 24 48 72 96 120 144 0 48 72 96 120 144
Ketones
7.411 4-Hydroxy-2-butanone - 0.00 0.68±0.14 0.00 0.00 0.00 0.00 0.00 0.00 0.00 4.27±0.16 0.00 0.00 0.00
10.344 2,3-Butanedione Buttery, creamy 0.00 0.00 0.00 0.00 0.00 7.29±0.10 0.00 0.00 0.00 0.00 1.87±0.10 0.99±0.02 8.69±0.20
11.102 3-Methyl-2-butanone - 0.00 0.00 0.00 0.00 0.00 0.00 0.34±0.02 0.00 0.00 0.16±0.00 0.00 0.00 0.11±0.00
13.523 3-Penten-2-one - 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.17±0.01
14.529 2-Heptanone Sweet, fruity 0.56±0.05 0.00 0.00 0.00 0.00 0.00 0.00 0.82±0.11 0.00 0.00 0.00 0.00 0.00
14.508 4-Methyl-2-hexanone - 0.35±0.00 5.71±0.08 0.00 0.00 0.00 0.00 0.00 0.69±0.07 0.00 0.00 0.08±0.00 0.00 0.27±0.03
16.457 3-Hydroxy-2-butanone Buttery, creamy 0.00 8.99±0.58 0.00 5.48±0.69 0.00 0.00 2.73±0.12 0.00 0.00 4.37±0.67 0.00 0.00 3.68±0.05
17.903 2-Nonanone Milk, green, fruity 0.00 0.00 0.00 0.00 0.00 0.00 0.53±0.01 0.00 0.00 0.50±0.06 0.00 0.00 0.00
21.739 Acetophenone Must-like, flowery, sweet, almond 5.50±0.05 6.09±0.94 1.16±0.16 0.64±0.04 1.32±0.12 0.00 1.01±0.19 6.43±1.78 2.20±0.30 2.85±0.01 1.35±0.04 0.71±0.01 2.57±0.14
total percentage 6.41±0.05 22.26±1.74 1.16±0.16 6.12±0.73 1.32±0.12 7.29±0.10 4.61±0.34 7.94±1.78 2.20±0.30 7.78±0.74 3.29±0.14 1.70±0.03 15.50±0.42
Pyrazines
17.09 2,5-Dimethylpyrazine Earthy, chocolate, nutty 0.00 0.00 0.00 0.00 0.00 0.00 0.51±0.04 0.00 0.00 0.00 0.00 0.00 0.28±0.01
total percentage 0.00 0.00 0.00 0.00 0.00 0.00 0.51±0.04 0.00 0.00 0.00 0.00 0.00 0.28±0.01
Rt= Retention time in minutes. a Percentage of compound based on the area normalization. - Indicates compound not detected. *Odor descriptor shows according Magagna et al., 2017, Rodriguez-Campos et al., 2012, Hinneh et al., 2018; and Ascrizzi et al., 2017.

Figure 1
Figure

Figure 2

Figure 3
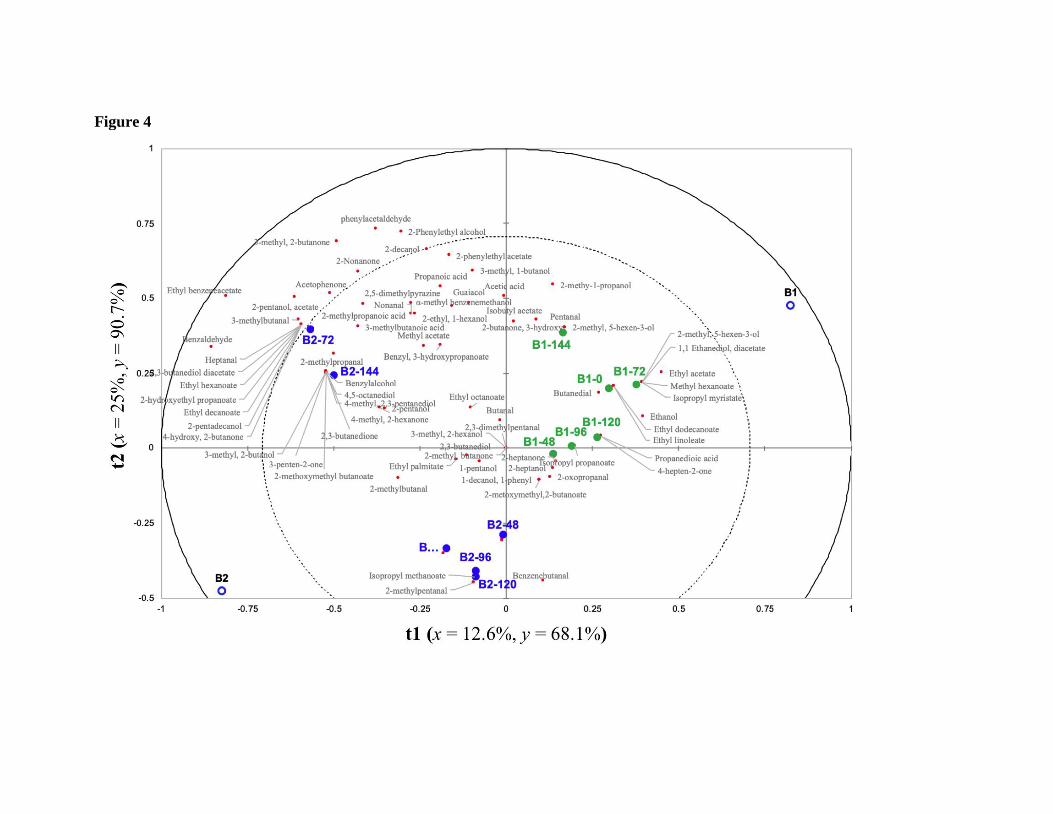
Figure 4