interactions between the human pathogen parahaemolyticus ...multiplicity of infection (moi) of...
TRANSCRIPT

Interactions between the human pathogen Vibrio parahaemolyticus and common marine microalgae
ABSTRACT Vibrio parahaemolyticus is a gastrointestinal pathogen that is abundant in coastal marine environments. Elevated numbers of V. parahaemolyticus cells have been correlated with marine microalgae blooms, particularly blooms of diatoms and dinoflagellates, but the nature of the relationship between V. parahaemolyticus and microalgae is unknown. We performed in vitro assays using 27 environmental V. parahaemolyticus strains and various phototrophs; a diatom, a dinoflagellate, unarmored and armored forms of a coccolithophore, and two species of cyanobacteria. The V. parahaemolyticus strains we employed contained different combinations of virulence-correlated genes, the hemolysin genes tdh and trh, the Type III Secretion System 2 (T3SS2) marker gene vscC2, and the Type VI Secretion System (T6SS) marker gene vipA1. We determined that all V. parahaemolyticus strains, even strains in which no virulence factor genes were detected, were able to cause decreases in diatom, dinoflagellate, and unarmored coccolithophore biomass in vitro. No correlation between content of any virulence gene and damage to microalgae was apparent. We hypothesize that marine microalgae represent a reservoir of nutrients that the copiotroph V. parahaemolyticus can utilize in salt marsh environments, which are often poor in labile carbon and energy sources. This helps to explain the recent correlations between V. parahaemolyticus and microalgae blooms in such environments.
KEYWORDS: Vibrio parahaemolyticus, tdh, trh, T3SS2, T6SS, microalgae. INTRODUCTION Vibrio parahaemolyticus, a common organism in coastal environments, is a significant and sometimes pandemic human pathogen responsible for an estimated 34,000 cases of seafood-associated gastroenteritis per year in the United States [1]. Most cases of V. parahaemolyticus-induced gastroenteritis are self-limiting and relatively mild, but infections can be deadly in immunocompromised individuals. The common mode of transmission of this bacterium to the human host is ingestion of raw or undercooked shellfish, primarily oysters. In addition, some strains of V. parahaemolyticus can infect wounds and some produce systemic infections, while others are apparently non-pathogenic. Elevated densities of V. parahaemolyticus most often occur during the warm months and at warm locations [2, 3] but recently large vibriosis outbreaks have occurred at locations not considered typical for this organism [4, 5]. V. parahaemolyticus not only persists but can increase in population size very rapidly in coastal marine environments [3, 6, 7]. It is not understood how this copiotrophic organism acquires carbon and other nutrients in coastal marine ecosystems where levels of utilizable soluble (labile) carbon and energy sources are typically quite low [8-10]. Even considering the known catabolic versatility of this species [3], rapid growth opportunities in many coastal ecosystems would seem infrequent at best. The abundance of V. parahaemolyticus as
Department of Biological Sciences, University of South Carolina, Columbia, South Carolina, 29208, USA.
Savannah L. Klein, Katherine E. Haney, Thomas M. Hornaday, India B. Gartmon and Charles R. Lovell*
*Corresponding author: [email protected]
Current Trends in Microbiology
Vol. 12, 2018

free-living cells in water is typically low (< 2,000 cells per liter) [6], but this organism can be very abundant in surficial sediment and in infaunal burrows [3, 6]. V. parahaemolyticus also occurs at higher levels in shellfish [2, 7, 11], and in association with algal blooms [12-16]. Thus, significant reservoirs exist even when no outbreak is underway [17]. The population expansion of V. parahaemolyticus that predicates an outbreak may be supported by means other than the typically low abundance, and largely refractory, dissolved organic carbon pool found in relatively low human impact coastal marine systems. Potential for a V. parahaemolyticus outbreak has often been predicted on the basis of local temperature, salinity, turbidity, and chlorophyll a concentrations [18-20]. In addition, some correlations between V. parahaemolyticus densities and certain algal taxa, specifically diatoms and dinoflagellates [12, 21, 22], have been reported and elevated levels of V. parahaemolyticus can occur during dinoflagellate and diatom blooms [13, 14]. The interaction between microalgae and V. parahaemolyticus could be commensalistic, based on soluble exudates released from algal cells lysed by viruses [23, 24], inefficient grazing by zooplankton [25, 26], or from undamaged algal cells. Or, perhaps, the microalgae themselves serve as supplemental carbon sources that V. parahaemolyticus can utilize. We examined the ability of V. parahaemolyticus to cause damage to healthy phototrophs. Chlorophyll a served as an indicator of phototroph biomass. Six phototrophs, including three species of microalgae and two species of cyanobacteria were incubated with several strains of V. parahaemolyticus. The phototrophs employed are abundant in marine environments and present a variety of cell wall surface structures and properties, providing insight into associations between susceptibility of the microalgae to V. parahaemolyticus predation and cell wall features. MATERIALS AND METHODS
V. parahaemolyticus strain isolation and characterization V. parahaemolyticus strains were isolated from the pristine North Inlet estuary near Georgetown, SC, USA (33°20’N, 79°12’W) in August and September
Savannah L. Klein et al.
2011 as described previously [27]. The North Inlet-Winyah Bay National Estuarine Research Reserve protects the third largest watershed on the east coast of the United States; and North Inlet is a bar built oligotrophic salt marsh where human impact is negligible [28, 29]. Samples were diluted and plated directly onto Thiosulfate Citrate Bile Salts Sucrose agar (TCBS) (BD, NJ). The presumptive identification of all V. parahaemolyticus strains used in this study was confirmed by recA sequence analysis [27] using the PCR primers and protocols of Thompson et al. [30].
Virulence gene PCR screening Two virulence-related hemolysin genes, tdh and trh, have been correlated with pathogenesis in V. parahaemolyticus, and these hemolysin genes are frequently used as molecular markers for strain virulence [27, 31]. Additional virulence factors, specifically secretion systems, have been discovered with recent sequencing of V. parahaemolyticus genomes [32-34]. The Type III Secretion System (T3SS2) has also been implicated in V. parahaemolyticus virulence [32] and the outer membrane protein gene, vscC2 is a useful marker for this structure [35, 36]. The Type VI Secretion System (T6SS) has also been detected in some V. parahaemolyticus isolates [37]. This secretion system has not been implicated in the pathogenicity of V. parahaemolyticus to humans, but T6SS producing V. parahaemolyticus strains have been shown to cause damage to other prokaryotes in vitro when incubated on a surface [37]. Its impacts on eukaryotic microalgae are presently unknown. Strains were screened for the T6SS marker gene vipA1 using the PCR primers and protocols of Salomon et al. [37]. V. parahaemolyticus strains were grown overnight at 37 ºC in Saline Luria-Bertani Broth (SLB; per L 27 g NaCl, 10 g Tryptone, 5 g Yeast Extract) and boiled extracts (15 min at 95-100 ºC) were prepared. All PCR reactions were completed within three days of DNA extraction and 1 μl of boiled DNA extract was used per reaction. PCR products were resolved on a 1.5% agarose gel and sequenced using an ABI Prism 3730 DNA analyzer to confirm gene identity. Sequences were analyzed using the Kimura 2 parameter model with Mega version 7 [38]. Sequence data obtained from this work were

Vibrio parahaemolyticus interactions with phototrophs
final and initial time points was determined by the formula: ((Tfinal – Tinitial) / (Tinitial)) x 100. Before and after incubation, aliquots were observed under a Nikon Eclipse TS100 microscope to determine the effect of V. parahaemolyticus clinical strains ATCC 17802T, ATCC 33846, and environmental strain 5-10-J5-4 on unarmored E. huxleyi. Microalgal cell counts were performed using a hemocytometer after the 24-h co-incubation. Each V. parahaemolyticus strain was tested in 12 wells per 96 well plate. For true replication, each 96 well plate was repeated three times. Controls included replicates of phototrophs in appropriate media and replicates of phototrophs in artificial seawater (with no V. parahaemolyticus added), against which experimental replicates were compared. Vibrio pacinii, an avirulent Vibrio [43], was used as a non-V. parahaemolyticus, heterotrophic bacterial control. Changes in chlorophyll a fluorescence were compared to controls (phototrophs suspended in artificial seawater without V. parahaemolyticus) using one way analysis of variance (ANOVA). Multiple comparisons were made versus the control group using the Holm-Sidak method (SigmaPlot, 2016). The significance level used was 0.05. All 29 V. parahaemolyticus strains were tested against the eukaryotic algae. A subset of these strains, some having the antibacterial mechanism T6SS (vipA1-positive) and some lacking it, was also tested against the cyanobacteria. We also used the T6SS-bearing POR1 strain, a derivative of the V. parahaemolyticus RIMD 2210633 reference strain, and two POR1 derivatives. The POR1 derivatives were the T6SS-knockout strain, POR1∆hcp1, and the T6SS de-repressed strain, POR1∆hns.
Dose response To determine the effect of V. parahaemolyticus dosage on the in vitro experiments, bacterial cultures were serially diluted (107 to 102 cells mL-1) in artificial seawater. These dilutions were incubated with unarmored E. huxleyi, the concentration of which was not altered. RESULTS
Eukaryotic microalgae in vitro experiments Twenty nine V. parahaemolyticus strains, 27 of which were environmental strains isolated from
submitted to the NCBI GenBank and assigned the accession numbers KX171447- KX171449.
Cultivation of microalgae and cyanobacteria Phototroph cultures were obtained from the Bigelow National Center for Marine Algae and Microbiota (Bigelow Center, East Boothbay, ME). Three species of eukaryotic microalgae were used in this project, the diatom Thallasiosira pseudonana (CCMP 1335), the dinoflagellate Prorocentrum minimum (CCMP 695), and two strains of the coccolithophore, Emiliania huxleyi (CCMP 371 and CCMP 373). T. pseudonana and P. minimum are common in North Inlet and E. huxleyi CCMP 371 is a coccolith-producing (armored) form that causes extensive blooms. E. huxleyi CCMP 373 is an unarmored mutant phenotype. Two species of cyanobacteria were also used, Prochlorococcus marinus (CCMP 1986) and Synechococcus bacillaris (CCMP 1333). All of the phototrophs employed have ubiquitous distributions worldwide and are frequently found in the same environments where V. parahaemolyticus occurs [39-41]. The phototrophs chosen had a variety of cell wall surface structures [39-42] and each required its own growth medium (S1). Microalgae and cyanobacteria were grown at 23 °C with an 11h light, 13h dark cycle.
In vitro experiments V. parahaemolyticus strains were grown in SLB at 37 °C with shaking. At 5 h, cultures were in exponential growth phase and yielded approximately 2 x 107 cells mL-1. Cultures were centrifuged (600 x g), the supernatants discarded, and cell pellets resuspended in a mixture of artificial sea salts (Instant Ocean, 33 ppt). Phototroph cultures were grown for 5 days, reaching approximately 2.0 x 105 cells mL-1, then harvested by centrifugation (1075 x g) for 10 min, and the supernatants removed. Cells were resuspended in 33 ppt Instant Ocean. Bacterial strains and phototroph cultures were combined in 96 well microplates with a multiplicity of infection (MOI) of approximately 100:1. Chlorophyll a fluorescence was used as an indicator of phototroph condition, and was measured immediately after co-inoculation (Tinitial) using a SpectraMax Gemini EM microplate reader (Molecular Devices, Sunnyvale, CA). Microplates were then incubated with light for 24 h at 25 °C. After incubation, chlorophyll a fluorescence was measured (Tfinal). The percent difference between
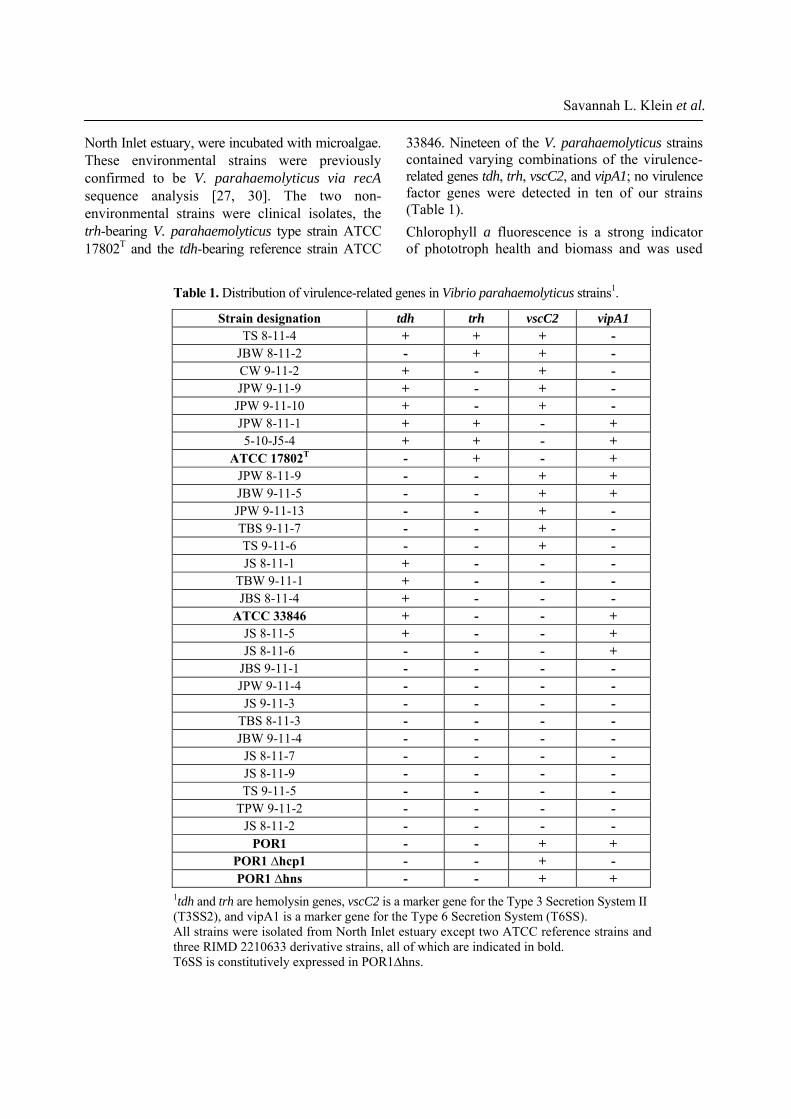
33846. Nineteen of the V. parahaemolyticus strains contained varying combinations of the virulence-related genes tdh, trh, vscC2, and vipA1; no virulence factor genes were detected in ten of our strains (Table 1). Chlorophyll a fluorescence is a strong indicator of phototroph health and biomass and was used
North Inlet estuary, were incubated with microalgae. These environmental strains were previously confirmed to be V. parahaemolyticus via recA sequence analysis [27, 30]. The two non-environmental strains were clinical isolates, the trh-bearing V. parahaemolyticus type strain ATCC 17802T and the tdh-bearing reference strain ATCC
Savannah L. Klein et al.
Table 1. Distribution of virulence-related genes in Vibrio parahaemolyticus strains1.
Strain designation tdh trh vscC2 vipA1 TS 8-11-4 + + + -
JBW 8-11-2 - + + - CW 9-11-2 + - + - JPW 9-11-9 + - + - JPW 9-11-10 + - + - JPW 8-11-1 + + - + 5-10-J5-4 + + - +
ATCC 17802T - + - + JPW 8-11-9 - - + + JBW 9-11-5 - - + + JPW 9-11-13 - - + - TBS 9-11-7 - - + - TS 9-11-6 - - + - JS 8-11-1 + - - -
TBW 9-11-1 + - - - JBS 8-11-4 + - - -
ATCC 33846 + - - + JS 8-11-5 + - - + JS 8-11-6 - - - +
JBS 9-11-1 - - - - JPW 9-11-4 - - - - JS 9-11-3 - - - -
TBS 8-11-3 - - - - JBW 9-11-4 - - - -
JS 8-11-7 - - - - JS 8-11-9 - - - - TS 9-11-5 - - - -
TPW 9-11-2 - - - - JS 8-11-2 - - - -
POR1 - - + + POR1 ∆hcp1 - - + - POR1 ∆hns - - + +
1tdh and trh are hemolysin genes, vscC2 is a marker gene for the Type 3 Secretion System II (T3SS2), and vipA1 is a marker gene for the Type 6 Secretion System (T6SS). All strains were isolated from North Inlet estuary except two ATCC reference strains and three RIMD 2210633 derivative strains, all of which are indicated in bold. T6SS is constitutively expressed in POR1∆hns.

Vibrio parahaemolyticus interactions with phototrophs
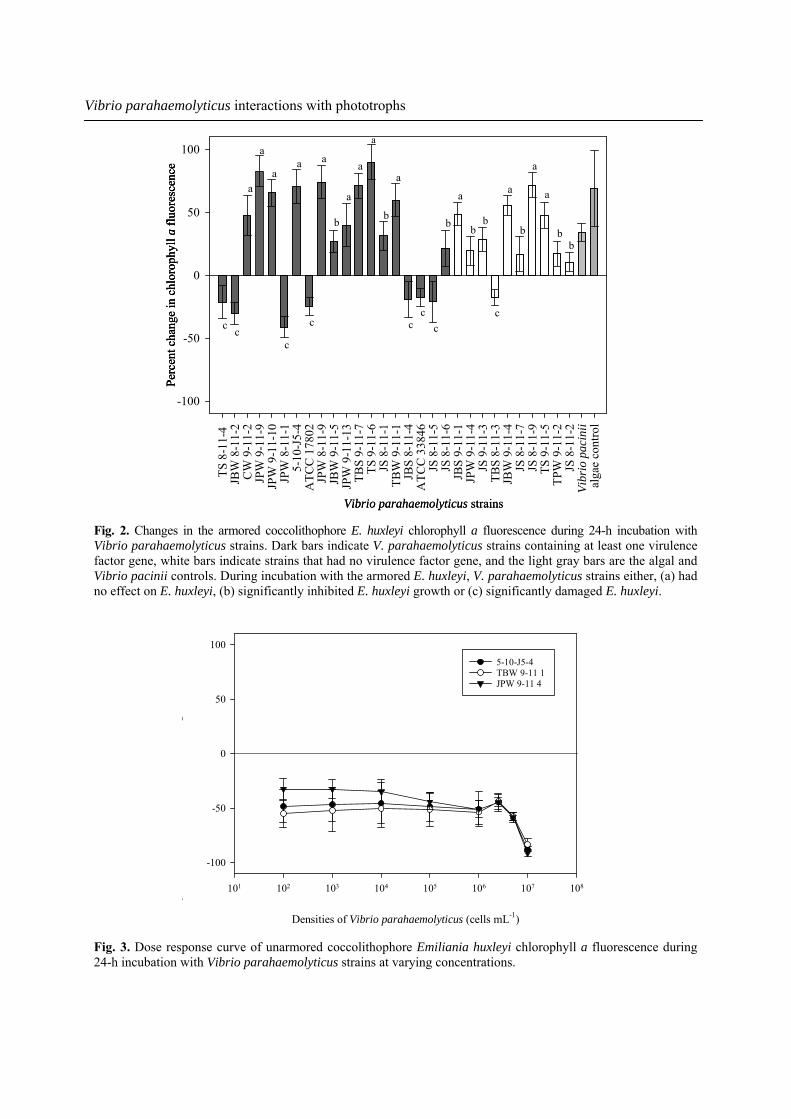
Dose response We conducted dose response experiments (Fig. 3), challenging unarmored E. huxleyi with three of our environmental V. parahaemolyticus strains at levels of 107 to 102 cells mL-1. We found that even at low V. parahaemolyticus concentrations, this pathogen was able to reduce the biomass of unarmored E. huxleyi. As expected, the degree of chlorophyll a loss decreased at lower V. parahaemolyticus concentrations. For example, V. parahaemolyticus strain TBW 9-11-1 caused an 83.1% decrease in unarmored E. huxleyi chlorophyll a fluorescence at 107 cells mL-1, while at 102 cells mL-1 a 55.2% decrease in chlorophyll a fluorescence was observed.
vipA1 in environmental V. Parahaemolyticus strainsSix of our 27 (22%) environmental V. parahaemolyticus strains carried the T6SS marker gene vipA1. vipA1 was also detected in the clinical reference strains ATCC 17802T and ATCC 33846. The T6SS is only expressed when V. parahaemolyticus is also expressing lateral flagella on a surface [36]; however, we did not find the T6SS marker gene vipA1 exclusively in strains isolated from surfaces. Three environmental strains that contained vipA1 were isolated from sediment (JS 8-11-5, JS 8-11-6, 5-10-J5-4); the other three were isolated from water (JPW 8-11-1, JPW 8-11-9, JBW 9-11-5). Sequence data recovered from these amplicons confirmed amplification of the vipA1 gene.
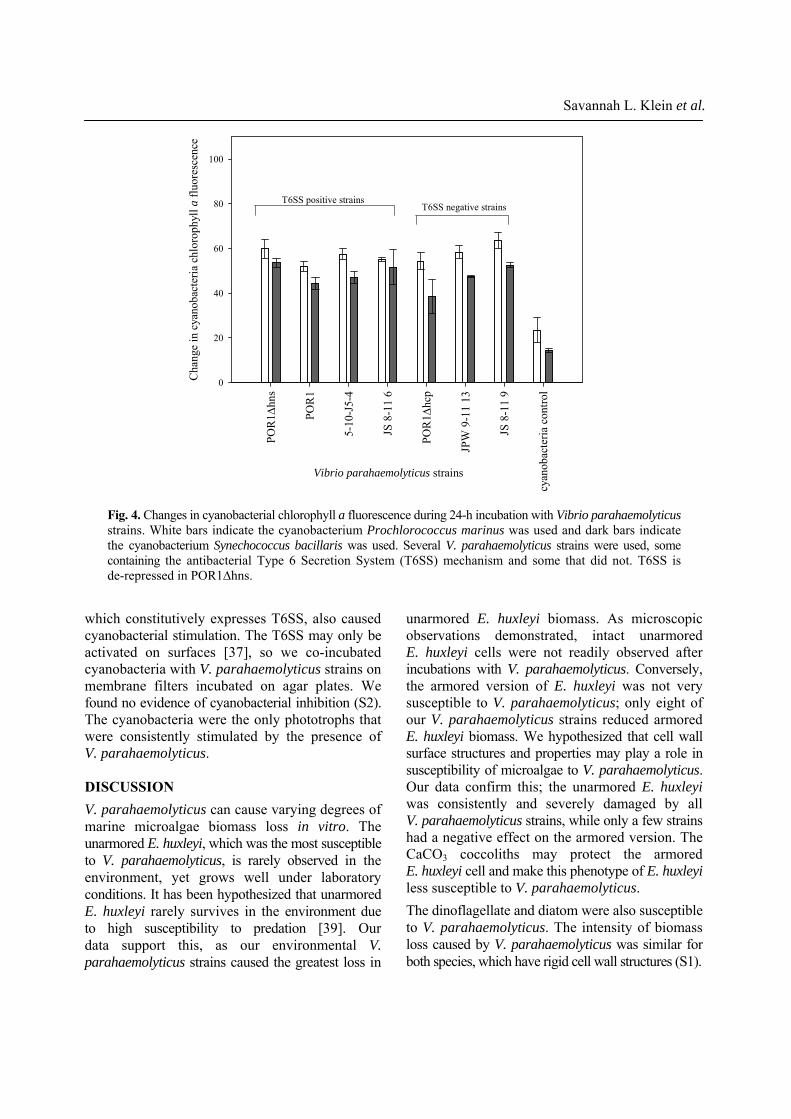
Cyanobacteria in vitro experiments Cyanobacteria were exposed to a subset of our environmental V. parahaemolyticus strains; some that contained the antibacterial mechanism T6SS and some that did not. We found that regardless of content of vipA1, all V. parahaemolyticus strains tested stimulated cyanobacterial growth (Fig. 4). No decrease of biomass was observed in either species of cyanobacteria, rather there was a significant increase in chlorophyll a fluorescence during co-incubation with V. parahaemolyticus (One Way ANOVA, all p values < 0.001). There was no significant difference in cyanobacterial stimulation between strains that contained T6SS and strains that did not (Student’s t-test, p values 0.214 and 0.252). The de-repressed strain POR1∆hns,
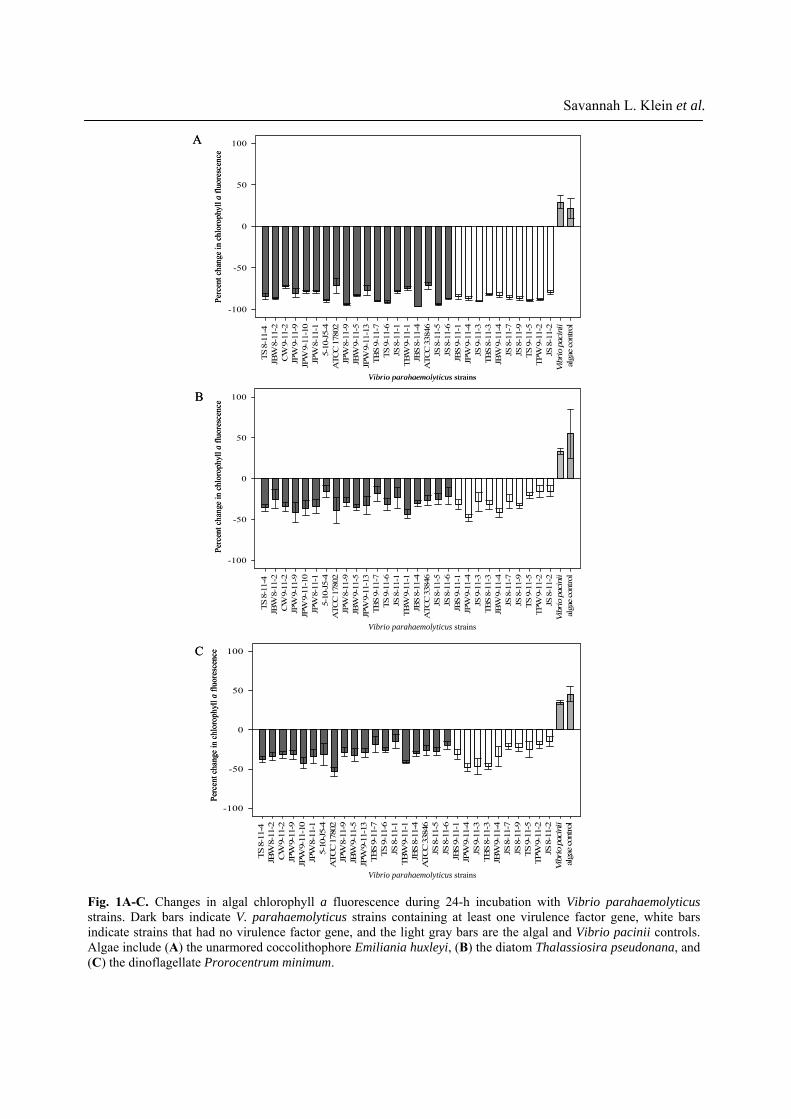
to quantitatively measure the condition of each alga when exposed to V. parahaemolyticus. The unarmored coccolithophore E. huxleyi was most susceptible to V. parahaemolyticus, with chlorophyll a decreasing by 71.5-96.3% (Fig. 1A) after incubation. Variability among replicates was extremely low. No correlations could be made between content of virulence gene(s) and unarmored E. huxleyi biomass loss. Instead, consistently strong decreases in chlorophyll a fluorescence were observed in the presence of all V. parahaemolyticus strains (One Way ANOVA, all p values < 0.001). Chlorophyll a decreases were accompanied by strong decreases in microscopically visible cells (S3). The avirulent Vibrio control, V. pacinii, did not cause any decreases in chlorophyll a in unarmored E. huxleyi or any other microalga. All V. parahaemolyticus strains tested caused a significant loss in the diatom T. pseudonana and the dinoflagellate P. minimum biomass (Figs. 1B and 1C). The decreases in chlorophyll a from these species were not as extreme as those of unarmored E. huxleyi, amounting to 15-50% (T. pseudonana: 15.3 to 48.0%; P. minimum 14.7 to 53.3%). Compared to controls, all V. parahaemolyticus strains tested caused significant chlorophyll a decreases in both algae (One Way ANOVA, all p values <0.001), regardless of virulence gene content. The armored E. huxleyi showed highest variability when incubated with V. parahaemolyticus (Fig. 2), with three different results observed: (a) Some V. parahaemolyticus strains had no effect (13 strains, One Way ANOVA, p values 0.09-0.88, no growth stimulation). (b) Some produced significant inhibition of armored E. huxleyi growth (8 strains, One Way ANOVA, p values < 0.001 to 0.009). When exposed to these strains, the armored E. huxleyi grew (i.e.: the chlorophyll a fluorescence (biomass) increased) but this growth was significantly less than the control with no V. parahaemolyticus. (c) Significant losses in E. huxleyi biomass, as seen by decreases in chlorophyll a fluorescence (8 strains, One Way ANOVA, all p values < 0.001). Once again, we found no correlation between content of virulence factor gene(s) and decreases in chlorophyll a fluorescence. For example, strain TBS 8-11-3 caused significant armored E. huxleyi biomass loss, yet no virulence factor genes were detected in this strain.
TS 8
-11-
4 JB
W 8
-11-
2C
W 9
-11-
2JP
W 9
-11-
9JP
W 9
-11-
10JP
W 8
-11-
15-
10-J
5-4
ATC
C 1
7802
JPW
8-1
1-9
JBW
9-1
1-5
JPW
9-1
1-13
TBS
9-11
-7TS
9-1
1-6
JS 8
-11-
1TB
W 9
-11-
1JB
S 8-
11-4
ATC
C 3
3846
JS 8
-11-
5JS
8-1
1-6
JBS
9-11
-1JP
W 9
-11-
4JS
9-1
1-3
TBS
8-11
-3JB
W 9
-11-
4JS
8-1
1-7
JS 8
-11-
9TS
9-1
1-5
TPW
9-1
1-2
JS 8
-11-
2Vi
brio
pac
inii
alga
e co
ntro
l
-100
-50
0
50
100
TS 8
-11-
4 JB
W 8
-11-
2C
W 9
-11-
2JP
W 9
-11-
9JP
W 9
-11-
10JP
W 8
-11-
15-
10-J
5-4
ATC
C 1
7802
JPW
8-1
1-9
JBW
9-1
1-5
JPW
9-1
1-13
TBS
9-11
-7TS
9-1
1-6
JS 8
-11-
1TB
W 9
-11-
1JB
S 8-
11-4
ATC
C 3
3846
JS 8
-11-
5JS
8-1
1-6
JBS
9-11
-1JP
W 9
-11-
4JS
9-1
1-3
TBS
8-11
-3JB
W 9
-11-
4JS
8-1
1-7
JS 8
-11-
9TS
9-1
1-5
TPW
9-1
1-2
JS 8
-11-
2Vi
brio
pac
inii
alga
e co
ntro
l-100
-50
0
50
100
TS 8
-11-
4 JB
W 8
-11-
2C
W 9
-11-
2JP
W 9
-11-
9JP
W 9
-11-
10JP
W 8
-11-
15-
10-J
5-4
ATC
C 1
7802
JPW
8-1
1-9
JBW
9-1
1-5
JPW
9-1
1-13
TBS
9-11
-7TS
9-1
1-6
JS 8
-11-
1TB
W 9
-11-
1JB
S 8-
11-4
ATC
C 3
3846
JS 8
-11-
5JS
8-1
1-6
JBS
9-11
-1JP
W 9
-11-
4JS
9-1
1-3
TBS
8-11
-3JB
W 9
-11-
4JS
8-1
1-7
JS 8
-11-
9TS
9-1
1-5
TPW
9-1
1-2
JS 8
-11-
2Vi
brio
pac
inii
alga
e co
ntro
l
-100
-50
0
50
100
Perc
ent c
hang
e in
chl
orop
hyll
aflu
ores
cenc
ePe
rcen
t cha
nge
in c
hlor
ophy
ll a
fluor
esce
nce
Perc
ent c
hang
e in
chl
orop
hyll
aflu
ores
cenc
e
Vibrio parahaemolyticus strains
Vibrio parahaemolyticus strains
Vibrio parahaemolyticus strains
A
B
C
TS 8
-11-
4 JB
W 8
-11-
2C
W 9
-11-
2JP
W 9
-11-
9JP
W 9
-11-
10JP
W 8
-11-
15-
10-J
5-4
ATC
C 1
7802
JPW
8-1
1-9
JBW
9-1
1-5
JPW
9-1
1-13
TBS
9-11
-7TS
9-1
1-6
JS 8
-11-
1TB
W 9
-11-
1JB
S 8-
11-4
ATC
C 3
3846
JS 8
-11-
5JS
8-1
1-6
JBS
9-11
-1JP
W 9
-11-
4JS
9-1
1-3
TBS
8-11
-3JB
W 9
-11-
4JS
8-1
1-7
JS 8
-11-
9TS
9-1
1-5
TPW
9-1
1-2
JS 8
-11-
2Vi
brio
pac
inii
alga
e co
ntro
l
-100
-50
0
50
100
TS 8
-11-
4 JB
W 8
-11-
2C
W 9
-11-
2JP
W 9
-11-
9JP
W 9
-11-
10JP
W 8
-11-
15-
10-J
5-4
ATC
C 1
7802
JPW
8-1
1-9
JBW
9-1
1-5
JPW
9-1
1-13
TBS
9-11
-7TS
9-1
1-6
JS 8
-11-
1TB
W 9
-11-
1JB
S 8-
11-4
ATC
C 3
3846
JS 8
-11-
5JS
8-1
1-6
JBS
9-11
-1JP
W 9
-11-
4JS
9-1
1-3
TBS
8-11
-3JB
W 9
-11-
4JS
8-1
1-7
JS 8
-11-
9TS
9-1
1-5
TPW
9-1
1-2
JS 8
-11-
2Vi
brio
pac
inii
alga
e co
ntro
l-100
-50
0
50
100
TS 8
-11-
4 JB
W 8
-11-
2C
W 9
-11-
2JP
W 9
-11-
9JP
W 9
-11-
10JP
W 8
-11-
15-
10-J
5-4
ATC
C 1
7802
JPW
8-1
1-9
JBW
9-1
1-5
JPW
9-1
1-13
TBS
9-11
-7TS
9-1
1-6
JS 8
-11-
1TB
W 9
-11-
1JB
S 8-
11-4
ATC
C 3
3846
JS 8
-11-
5JS
8-1
1-6
JBS
9-11
-1JP
W 9
-11-
4JS
9-1
1-3
TBS
8-11
-3JB
W 9
-11-
4JS
8-1
1-7
JS 8
-11-
9TS
9-1
1-5
TPW
9-1
1-2
JS 8
-11-
2Vi
brio
pac
inii
alga
e co
ntro
l
-100
-50
0
50
100
Perc
ent c
hang
e in
chl
orop
hyll
aflu
ores
cenc
ePe
rcen
t cha
nge
in c
hlor
ophy
ll a
fluor
esce
nce
Perc
ent c
hang
e in
chl
orop
hyll
aflu
ores
cenc
e
Vibrio parahaemolyticus strains
Vibrio parahaemolyticus strains
Vibrio parahaemolyticus strains
A
B
C
Savannah L. Klein et al.
Fig. 1A-C. Changes in algal chlorophyll a fluorescence during 24-h incubation with Vibrio parahaemolyticus strains. Dark bars indicate V. parahaemolyticus strains containing at least one virulence factor gene, white bars indicate strains that had no virulence factor gene, and the light gray bars are the algal and Vibrio pacinii controls. Algae include (A) the unarmored coccolithophore Emiliania huxleyi, (B) the diatom Thalassiosira pseudonana, and (C) the dinoflagellate Prorocentrum minimum.
Densities of Vibrio parahaemolyticus (cells mL-1)
101 102 103 104 105 106 107 108
hi
fl
-100
-50
0
50
1005-10-J5-4 TBW 9-11 1 JPW 9-11 4
TS 8
-11-
4 JB
W 8
-11-
2C
W 9
-11-
2JP
W 9
-11-
9JP
W 9
-11-
10JP
W 8
-11-
15-
10-J
5-4
ATC
C 1
7802
JPW
8-1
1-9
JBW
9-1
1-5
JPW
9-1
1-13
TBS
9-11
-7TS
9-1
1-6
JS 8
-11-
1TB
W 9
-11-
1JB
S 8-
11-4
ATC
C 3
3846
JS 8
-11-
5JS
8-1
1-6
JBS
9-11
-1JP
W 9
-11-
4JS
9-1
1-3
TBS
8-11
-3JB
W 9
-11-
4JS
8-1
1-7
JS 8
-11-
9TS
9-1
1-5
TPW
9-1
1-2
JS 8
-11-
2Vi
brio
pac
inii
alga
e co
ntro
l
-100
-50
0
50
100
c cc
c cc
cc
b b
bb
bb
bb
a
a
aa a
a
a
a
a
a a
a
a
Perc
ent c
hang
e in
chl
orop
hyll
aflu
ores
cenc
e
Vibrio parahaemolyticus strains
TS 8
-11-
4 JB
W 8
-11-
2C
W 9
-11-
2JP
W 9
-11-
9JP
W 9
-11-
10JP
W 8
-11-
15-
10-J
5-4
ATC
C 1
7802
JPW
8-1
1-9
JBW
9-1
1-5
JPW
9-1
1-13
TBS
9-11
-7TS
9-1
1-6
JS 8
-11-
1TB
W 9
-11-
1JB
S 8-
11-4
ATC
C 3
3846
JS 8
-11-
5JS
8-1
1-6
JBS
9-11
-1JP
W 9
-11-
4JS
9-1
1-3
TBS
8-11
-3JB
W 9
-11-
4JS
8-1
1-7
JS 8
-11-
9TS
9-1
1-5
TPW
9-1
1-2
JS 8
-11-
2Vi
brio
pac
inii
alga
e co
ntro
l
-100
-50
0
50
100
c cc
c cc
cc
b b
bb
bb
bb
a
a
aa a
a
a
a
a
a a
a
a
TS 8
-11-
4 JB
W 8
-11-
2C
W 9
-11-
2JP
W 9
-11-
9JP
W 9
-11-
10JP
W 8
-11-
15-
10-J
5-4
ATC
C 1
7802
JPW
8-1
1-9
JBW
9-1
1-5
JPW
9-1
1-13
TBS
9-11
-7TS
9-1
1-6
JS 8
-11-
1TB
W 9
-11-
1JB
S 8-
11-4
ATC
C 3
3846
JS 8
-11-
5JS
8-1
1-6
JBS
9-11
-1JP
W 9
-11-
4JS
9-1
1-3
TBS
8-11
-3JB
W 9
-11-
4JS
8-1
1-7
JS 8
-11-
9TS
9-1
1-5
TPW
9-1
1-2
JS 8
-11-
2Vi
brio
pac
inii
alga
e co
ntro
l
-100
-50
0
50
100
c cc
c cc
cc
b b
bb
bb
bb
a
a
aa a
a
a
a
a
a a
a
a
Perc
ent c
hang
e in
chl
orop
hyll
aflu
ores
cenc
e
Vibrio parahaemolyticus strains
Perc
ent c
hang
e in
chl
orop
hyll
aflu
ores
cenc
e
Vibrio parahaemolyticus strains
Vibrio parahaemolyticus interactions with phototrophs
Fig. 2. Changes in the armored coccolithophore E. huxleyi chlorophyll a fluorescence during 24-h incubation withVibrio parahaemolyticus strains. Dark bars indicate V. parahaemolyticus strains containing at least one virulence factor gene, white bars indicate strains that had no virulence factor gene, and the light gray bars are the algal and Vibrio pacinii controls. During incubation with the armored E. huxleyi, V. parahaemolyticus strains either, (a) had no effect on E. huxleyi, (b) significantly inhibited E. huxleyi growth or (c) significantly damaged E. huxleyi.
Fig. 3. Dose response curve of unarmored coccolithophore Emiliania huxleyi chlorophyll a fluorescence during 24-h incubation with Vibrio parahaemolyticus strains at varying concentrations.
Vibrio parahaemolyticus strains
POR
1 Δhn
s
POR
1
5-10
-J5-
4
JS 8
-11
6
POR
1 Δhc
p
JPW
9-1
1 13
JS 8
-11
9
cyan
obac
teria
con
trol
Cha
nge
in c
yano
bact
eria
chl
orop
hyll
a flu
ores
cenc
e
0
20
40
60
80
100
T6SS positive strainsT6SS negative strains
Savannah L. Klein et al.
unarmored E. huxleyi biomass. As microscopic observations demonstrated, intact unarmored E. huxleyi cells were not readily observed after incubations with V. parahaemolyticus. Conversely, the armored version of E. huxleyi was not very susceptible to V. parahaemolyticus; only eight of our V. parahaemolyticus strains reduced armored E. huxleyi biomass. We hypothesized that cell wall surface structures and properties may play a role in susceptibility of microalgae to V. parahaemolyticus. Our data confirm this; the unarmored E. huxleyi was consistently and severely damaged by all V. parahaemolyticus strains, while only a few strains had a negative effect on the armored version. The CaCO3 coccoliths may protect the armored E. huxleyi cell and make this phenotype of E. huxleyi less susceptible to V. parahaemolyticus. The dinoflagellate and diatom were also susceptible to V. parahaemolyticus. The intensity of biomass loss caused by V. parahaemolyticus was similar for both species, which have rigid cell wall structures (S1).
which constitutively expresses T6SS, also caused cyanobacterial stimulation. The T6SS may only be activated on surfaces [37], so we co-incubated cyanobacteria with V. parahaemolyticus strains on membrane filters incubated on agar plates. We found no evidence of cyanobacterial inhibition (S2). The cyanobacteria were the only phototrophs that were consistently stimulated by the presence of V. parahaemolyticus. DISCUSSION V. parahaemolyticus can cause varying degrees of marine microalgae biomass loss in vitro. The unarmored E. huxleyi, which was the most susceptible to V. parahaemolyticus, is rarely observed in the environment, yet grows well under laboratory conditions. It has been hypothesized that unarmored E. huxleyi rarely survives in the environment due to high susceptibility to predation [39]. Our data support this, as our environmental V. parahaemolyticus strains caused the greatest loss in
Fig. 4. Changes in cyanobacterial chlorophyll a fluorescence during 24-h incubation with Vibrio parahaemolyticusstrains. White bars indicate the cyanobacterium Prochlorococcus marinus was used and dark bars indicate the cyanobacterium Synechococcus bacillaris was used. Several V. parahaemolyticus strains were used, some containing the antibacterial Type 6 Secretion System (T6SS) mechanism and some that did not. T6SS is de-repressed in POR1∆hns.

Vibrio parahaemolyticus interactions with phototrophs
Cellulose thecal plates protect P. minimum and a silica frustule covers T. pseudonana. Although these microalgae are covered by cell walls composed of differing materials, they were similarly susceptible to V. parahaemolyticus. Correlations between elevated V. parahaemolyticus densities and dinoflagellate and diatom blooms have been reported [12, 21, 22]. This may be due to V. parahaemolyticus causing damage to these marine microalgae, leading to nutrient acquisition (predation) by V. parahaemolyticus. Certainly, release of dissolved organic carbon (DOC) from microalgae due to excretion may also contribute to the association between V. parahaemolyticus and algal blooms, but direct predation on algae by V. parahaemolyticus presents an interesting additional aspect to this association. We found no correlation between content of tdh, trh, or vscC2 and microalgae biomass loss. The hemolysin genes tdh and trh have long been used as molecular markers of V. parahaemolyticus virulence. However, our results are consistent with recent reports [44-46] that destruction of eukaryotic cells does not exclusively rely on these hemolysin genes. Content of the T3SS marker gene vscC2 was also not correlated with algal loss. Ten of the V. parahaemolyticus strains used contained no known virulence factors, yet were able to cause decreases in algae chlorophyll a fluorescence. Our data suggest that another mechanism(s) by which V. parahaemolyticus damages these eukaryotes must exist. Cyanobacteria were tested against a subset of our V. parahaemolyticus strains, some that contained the antibacterial mechanism T6SS, and some that did not. In addition, we used a T6SS knockout mutant and a T6SS de-repressed strain. All strains of V. parahaemolyticus caused cyanobacterial growth stimulation. We found no evidence that V. parahaemolyticus can decrease cyanobacteria biomass. Cyanobacteria grow better in non-axenic laboratory cultures (Lovell and many others, personal observations) and our finding of cyanobacterial stimulation when incubated with V. parahaemolyticus is consistent with this observation. Perhaps V. parahaemolyticus and other heterotrophic bacteria consume or neutralize some inhibitory byproducts of cyanobacterial growth. In vitro, the presence of V. parahaemolyticus results
in cyanobacterial stimulation and microalgae biomass loss. In the environment, V. parahaemolyticus may affect phototrophic population dynamics. Further experimentation is needed to determine if V. parahaemolyticus can induce changes in marine phototroph populations, perhaps by selective predation upon phototrophs having more susceptible cell wall structures. The in vitro experiments performed used high doses of V. parahaemolyticus to assure observation of damage if such occurred. We also determined what would happen when the unarmored E. huxleyi was incubated with varying concentrations of V. parahaemolyticus, including concentrations that more accurately mimic V. parahaemolyticus densities observed in the environment. As shown by our dose response curves, the unarmored E. huxleyi was susceptible to V. parahaemolyticus at low concentrations. These low concentrations (103 or 102 cells mL-1) are similar to V. parahaemolyticus concentrations found in surficial sediment and shellfish [3, 6, 7, 11]. V. parahaemolyticus at low, “environmental” doses can still damage unarmored E. huxleyi, meaning that this interaction is certainly possible in specific environments. High concentrations of marine microalgae and other phototrophs are often found in surficial sediment in salt marshes along the US east coast; in North Inlet estuary, where our V. parahaemolyticus strains were isolated, chlorophyll a in the sediment can reach as high as 101.5 mg chlorophyll a m-2 [47, 48]. V. parahaemolyticus concentrations in surficial sediment, particularly around fiddler crab burrows, can reach levels as high as 103 cells mL-1 [3]. Clearly, phototrophs and V. parahaemolyticus occur, and even bloom, in the same environments. Utilization of phototrophs as an additional nutritional resource in the sediment, as well as other areas rich in both V. parahaemolyticus and phototrophs, may be a mechanism supporting persistence of V. parahaemolyticus. Acquisition of carbon and energy sources in estuaries having low availability of labile resources is imperative for V. parahaemolyticus survival and propagation. High densities of V. parahaemolyticus in oligotrophic environments prove that this copiotrophic human pathogen has mechanisms for gathering carbon under resource-poor conditions. We consider that marine microalgae represent a reservoir of

Savannah L. Klein et al.
was supported by South Carolina Sea Grant award NA140R4170082 Subaward N256 to Charles R. Lovell. Savannah Klein would like to thank the Slocum-Lunz foundation for funding. Katherine Haney would like to thank the University of South Carolina Honors College for funding. We would like to thank Dr. Tammi Richardson for access to algal cultures and media, and the Orth Lab at the University of Texas Southwestern Medical Center for providing the RIMD derivative strains. SUPPLEMENTARY MATERIAL S1. Surface structure of microalgae and cyanobacteria Two species of cyanobacteria were used in this study, Prochlorococcus marinus (CCMP 1986) and Synechococcus bacillaris (CCMP 1333). As is typical of gram negative prokaryotes, P. marinus and S. bacillaris have cell walls composed of peptidoglycan surrounded by an outer membrane and a glycocalyx of polysaccharides and polypeptides. Three species of eukaryotic microalgae were used, the diatom Thallasiosira pseudonana (CCMP 1335), the dinoflagellate Prorocentrum minimum (CCMP 695), and two strains of the coccolithophore, Emiliania huxleyi (CCMP 371 and CCMP 373).E. huxleyi CCMP 371 is a coccolith-producing (armored) form that often causes extensive blooms. E. huxleyi CCMP 373 is an unarmored mutant phenotype. The bloom producing armored form can have as many as thirty CaCO3 coccoliths forming multiple layers to protect the cell interior. The unarmored E. huxleyi lacks coccoliths and these strains are rarely isolated from the environment. Unarmored cells are thought to arise from armored cells via mutation; reversion back to the coccolith-forming morphology has not been reported [39]. The centric diatom T. pseudonana is a model organism and was the first diatom chosen forgenome sequencing [40]. As is the case for all diatoms, the cell wall, or frustule of T. pseudonana, iscomposed of amorphous hydrated silica in a species-specific three-dimensional structure [42]. The “petri dish” shape of centric diatoms is due to two unequalsilicate halves (valves) that are connected by a series
nutrients that V. parahaemolyticus can utilize. Primary producer biomass in salt marshes is dominated by cordgrasses, such as Spartina alterniflora; however, we do not think that S. alterniflora, and other macroflora, are the key to V. parahaemolyticus nutrient-acquisition in salt marshes. Up to 80% of vascular plant biomass is comprised of recalcitrant lignocellulose and its breakdown products [49]. Up to 40% of dissolved organic carbon in salt marshes also consists of recalcitrant lignocellulose [50]. Benthic marine phototrophs are responsible for almost half of the primary production in salt marsh ecosystems like the North Inlet estuary [47] and may be a preferred source of easily degraded carbon for heterotrophic bacteria. Salt marsh microphototroph populations primarily consist of cyanobacteria and diatoms and these organisms occur at highest biomass in the surficial sediment, where elevated V. parahaemolyticus densities are also found. We propose that it is no coincidence V. parahaemolyticus populations are correlated with marine microalgae as marine microalgae provide a nutrient-rich resource that V. parahaemolyticus can utilize in an otherwise nutrient-restricted system. CONCLUSION We performed in vitro assays co-incubating V. parahaemolyticus with various phototrophs and discovered that all V. parahaemolyticus strains, with or without known virulence-correlated genes, can cause significant decreases in marine microalgalbiomass. There was no correlation between known V. parahaemolyticus virulence genes and microalgal destruction, suggesting other mechanisms of virulence. We hypothesize that marine microalgae can act as a source of nutrients for V. parahaemolyticus in the environment, providing an explanation for recent correlations found between increased V. parahaemolyticus abundance and microalgal blooms. Our study provides insight into how this copiotrophic organism is able to persist in an environment that may be poor in readily utilized organic carbon and energy sources. ACKNOWLEDGEMENTS We thank the Belle W. Baruch Institute for Coastal and Marine Science for access to the North Inlet estuary in Georgetown, SC. This work

Na2MoO4·2H2O, 3.93 x 10-8 M CuSO4·5H2O) and with F/2 vitamins (2.96 x 10-7 M thiamine·HCl, 2.05 x 10-9 M biotin, 3.69 x 10-10 M cyanocobalamin). Armored E. huxleyi was grown in F/50, which is a 1/25 dilution of F/2-Si. T. pseudonana was grown in F/2+Si (add 1.06 x 10-4 M Na2SiO3·9H2O to F/2-Si recipe). The cyanobacterium P. marinus was grown in Pro99 medium (5 x 10-5 M NaH2PO4·H2O, 8 x 10-4 M NH4Cl) supplemented with pro99 trace metals (1.17 x 10-6 M Na2EDTA·2H2O, 1.17 x 10-6
M FeCl3·6H2O, 8 x 10-9 M ZnSO4·7H2O, 5 x 10-9
M CoCl2·6H2O, 9 x 10-8 M MnCl2·4H2O, 3 x 10-9
M Na2MoO4·2H2O, 1 x 10-8 M Na2SeO3, 1 x 10-8
M NiSO4·6H2O). All media recipes can be found on the Bigelow Center website (https://ncma.bigelow.org/algal-recipes). Microalgae and cyanobacteria were grown at 23 °C with an 11 h light, 13 h dark cycle.
of girdles. Additionally, to prevent silica dissolution, the frustule is covered by an organic casing made up of glycoproteins [42]. The dinoflagellate P. minimum is associated with harmful algal blooms (HABs) [41]. P. minimum cells are protected by overlapping thecal plates composed of cellulose. Phototroph cultures were obtained from the Bigelow National Center for Marine Algae and Microbiota (Bigelow Center, East Boothbay, Maine). Each algal species required its own growth medium. Unarmored E. huxleyi, the dinoflagellate P. minimum and the cyanobacteria S. bacillaris were grown in F/2-Si (8.82 x 10-4 M NaNO3, 3.62 x 10-5 M NaH2PO4·H2O) supplemented with F/2 trace metals (1.17 x 10-5 M Na2EDTA·2H2O, 1.17 x 10-5 M FeCl3·6H2O, 7.65 x 10-8 M ZnSO4·7H2O, 4.2 x 10-8 M CoCl2·6H2O, 9.1 x 10-7 M MnCl2·4H2O, 2.6 x 10-8 M
Vibrio parahaemolyticus interactions with phototrophs
por1∆hcp1
por1∆hns
S. bacillaris control
S2. Agar plate experiment Materials and Methods. Cyanobacterial (S. bacillaris) cultures were grown for 5 days in F/2-Si medium (Bigelow) at 23 °C in an environmental chamber, with an 11h light, 13h dark cycle. V. parahaemolyticus strains were grown in saline Luria-Bertani Broth (SLB per L, 10 g tryptone, 5 g yeast extract, 27g NaCl) for 5 h at 37 °C with shaking. V. parahaemolyticus cultures were spun down and resuspended in a mixture of artificial sea salts (33 ppt). V. parahaemolyticus strains and cyanobacteria were combined (MOI of 100:1) and filtered onto a sterile 0.45 μm polycarbonate filter. Filters were then aseptically placed on solid F/2-Si (Bigelow recipe, add 1.5% agarose) and were left to incubate at 23 °C, with an 11h light, 13h dark cycle for 5 days. Pictures were taken after the 5-day incubation.

Savannah L. Klein et al.
6. West, C. K. 2012, Ph.D. Thesis, University of South Carolina, Columbia, SC.
7. DePaola, A., Nordstrom, J., Bowers, C., Wells, J. and Cook, D. 2003, Appl. Environ. Microbiol., 69, 1521-1526.
8. Moran, M. A. and Hodson, R. E. 1990, Mar. Ecol. Prog. Ser., 62, 161-168.
9. Amon, R. N. W. and Benner, R. 1996, Limnol Oceanogr., 41, 41-51.
10. Kaiser, K. and Benner, R. 2009, Mar Chem., 113, 63-77.
11. Klein, S. L. and Lovell, C. R. 2016, FEMS Microbiol Ecol., 93, 1-8.
12. Turner, J. W., Good, B., Cole, D. and Lipp, E. K. 2009, ISME J., 9, 1082-1092.
13. Rehnstam-Holm, A. S., Godhe, A., Härnström, K., Saravanan, V., Collin, B. and Karunasagar, I. 2010, Aquatic Microb. Ecol., 58, 127-139.
14. Seong, K. A. and Jeong, H. J. 2011, Ocean Sci. J., 46, 105-115.
15. Turner, J. W., Malayil, L., Guadagnoli, D., Cole, D. and Lipp, E. K. 2014, Environ. Microbiol., 16, 1019-1028.
CONFLICT OF INTEREST STATEMENT None to declare. REFERENCES 1. Scallan, E., Hoekstra, R. M., Angulo, F. J.,
Tauxe, R. V., Widdowson, M-A, Roy, S. L., Jones, J. L. and Griffin, P. M. 2011, Emerg. Infect. Dis., 17, 7-15.
2. DePaola, A., Hopkins, L. H., Peeler, J. T., Wentz, B. and McPhearson, R. M. 1990, Appl. Environ. Microbiol., 56, 2299-2302.
3. Gamble, M. D. and Lovell, C. R. 2011, Appl. Environ. Microbiol., 77, 3703-3714.
4. McLaughlin, J. B., DePaola, A., Bopp, C. A., Martinek, K. A., Napolilli, N. P., Allison, C. G., Murray, S. L., Thompson, E. C., Bird, M. M. and Middaugh, J. P. 2005, New England J. Med., 353, 1463-1470.
5. Paranjype, R., Hamel, O. S., Stojanovski, A. and Liermann, M. 2012, Appl. Environ. Microbiol., 78, 8631-8638.
0
50,000
100,000
150,000
200,000
250,000
300,000
negative V. pacinii 17802T JS-8-11-7 JS-8-11-9 TS-8-11-4
Mic
roal
gal
cells
/mL
Vibrio strains
0
50,000
100,000
150,000
200,000
250,000
300,000
negative V. pacinii 17802T JS-8-11-7 JS-8-11-9 TS-8-11-4
Mic
roal
gal
cells
/mL
Vibrio strains
0
20,000
40,000
60,000
80,000
100,000
120,000
140,000
160,000
negative V. pacinii 17802 TS-8-11-4 JS-8-11-9 JS-8-11-7
Mic
roal
gal
cells
/mL
Vibrio strains
0
20,000
40,000
60,000
80,000
100,000
120,000
140,000
160,000
negative V. pacinii 17802 TS-8-11-4 JS-8-11-9 JS-8-11-7
Mic
roal
gal
cells
/mL
Vibrio strains
A B
C D
S3. Microalgal cell densities after a 24-h incubation with Vibrio parahaemolyticus strains (A-D). Algae include (A) the unarmored coccolithophore Emiliania huxleyi, (B) the armored coccolithophore Emiliania huxleyi, (C) the diatom Thalassiosira pseudonana, and (D) the dinoflagellate Prorocentrum minimum.

16. Main, C. R., Salvitti, L. R., Whereat, E. B. and Coyne, K. J. 2015, Appl. Environ. Microbiol., 81, 5703-5713.
17. Lovell, C. R. 2017, Appl. Microbiol. Biotechnol., 101, 1781-1794.
18. Carburlotto, G., Haley, B. J., Lleò, M. M., Hug, A. and Colwell, R. R. 2010, Environ. Microbiol. Rep., 2, 151-157.
19. Johnson, C., Flowers, A., Noriea, N., Zimmerman, A., Bowers, J., DePaola, A. and Grimes, D. 2010, Appl. Environ. Microbiol., 76, 7076-7084.
20. Johnson, C. N., Bowers, J. C., Griffitt, K. J., Molina, V., Clostio, R. W., Pei, S., Laws, E., Paranjpye, R. N., Strom, M. S., Chen, A., Hasan, N. A., Huq, A., Noriea, N. F., Grimes, D. J. and Colwell, R. R. 2012, Appl. Environ. Microbiol., 78, 7249-7257.
21. Eiler, A., Johansson, M. and Bertilsson, S. 2006, Appl. Environ. Microbiol., 72, 6004-6011.
22. Asplund, M. E., Rehnstam-Holm, A. S., Atnur, V., Raghunath, P., Saravanan, V., Härnström, K., Collin, B., Karunasagar, I. and Godhe, A. 2011, Environ. Microbiol., 13, 2738-2751.
23. Ortmann, A. C., Metzger, R. C., Liefer, J. D. and Novoveska, L. 2011, Aquat Microb Ecol., 65, 143-157.
24. Evans, C. and Brussaard, C. P. D. 2012, Limnol Oceanogr., 57, 1826-1837.
25. Hygum, B. H., Petersen, J. W. and Søndergaard, M. 1997, J. Plankton Res., 19, 97-111.
26. Møller, E. F. 2005, J. Plank. Res., 27, 27-35. 27. Gutierrez West, C., Klein, S. L. and Lovell,
C. R. 2013, Appl. Environ. Microbiol., 79, 2247-2252.
28. Dame, R. F., Alber, M., Allen, D., Mallin, M., Montague, C., Lewitus, A., Chalmers, A., Gardner, R., Gilman, C., Kjerfve, B., Pinckney, J. and Smith, N. 2000, Estuaries., 23, 793-819.
29. Buzzelli, C., Akman, O., Buck, T., Koepfler, E., Morris, J. and Lewitus, A. 2004, J. Coast. Res., 45, 59-74.
30. Thompson, F. L., Gevers, D., Thompson, C. C., Dawnydt, P., Naser, S., Hoste, B., Munn, C. B. and Swings, J. 2005, Appl. Environ. Microbiol., 71, 5107-5115.
31. DePaola, A. and Kaysner, C. A. 2004, U.S. Food and Drug Administration, Washington, DC. URL: http://www.fda.gov/Food/FoodScienceResearch/LaboratoryMethods/ucm070830.htm
32. Makino, K., Oshima, K., Kurokawa, K., Yokoyama, K., Uda, T., Tagomori, K., Iijima, Y., Najima, M., Nakano, M., Yamashita, A., Kubota, Y., Kimura, S., Yasunaga, T., Honda, T., Shinagawa, H., Hattori, M. and Iida, T. 2003, Lancet, 361, 743-749.
33. Park, K. S., Ono, T., Rokuda, M., Jang, M. H., Okada, K., Iida, T. and Honda, T. 2004, Infect. Immun., 72, 6659-6665.
34. Okada, N., Iida, T., Park, K. S., Goto, N., Yasunaga, T., Hiyoshi, H., Matsuda, S., Kodama, T. and Honda, T. 2009, Infect. Immun., 77, 904-913.
35. Noriea, N. F., Johnson, C. N., Griffitt, K. J. and Grimes, D. J. 2010, J. Appl. Microbiol., 109, 953-962.
36. Klein, S. L., West, C. K., Mejia, D. M. and Lovell, C. R. 2014, Appl. Environ. Microbiol., 80, 595-602.
37. Salomon, D., Gonzalez, H., Updegraff, B. L. and Orth, K. 2013, PLoS ONE, 8, e61086.
38. Tamura, K., Dudley, J., Nei, M. and Kumar, S. 2015, Mol. Biol. Evol., 24, 1596-1599.
39. Paasche, E. 2001, Phycologia, 40, 503-529. 40. Armbrust, E. V., Berges, J. A., Bowler, C.,
Green, B. R., Martinez, D., Putnam, N. H., Zhou, S., Allen, A. E., Apt, K. E., Bechner, M., Brzezinski, M. A., Chaal, B. K., Chiovitti, A., Davis, A. K., Demarest, M. S., Detter, J. C., Glavina, T., Goodstein, D., Hadi, M. Z., Hellsten, U., Hildebrand, M., Jenkins, B. D., Jurka, J., Kapitonov, V. V., Kröger, N., Lau, W. W., Lane, T. W., Larimer, F. W., Lippmeier, J. C., Lucas, S., Medina, M., Montsant, A., Obornik, M., Parker, M. S., Palenik, B., Pazour, G. J., Richardson, P. M., Rynearson, T. A., Saito, M. A., Schwartz, D. C., Thamatrokoln, K., Valentin, K., Vardi, A., Wilkerson, F. P. and Rokhsar, D. S. 2004, Science., 306, 79-86.
41. Heil, C. A., Glibert, P. M. and Fan, C. 2005, Harmful Algae., 4, 449-470.
42. Javaheri, N., Dries, R., Burson, A., Stal, L. J., Sloot, P. M. A. and Kaandorp, J. A. 2015, Scientific Reports, 5, 11652.
Vibrio parahaemolyticus interactions with phototrophs

43. Gomez-Gil, B., Thompson, F. L., Thompson, C. C. and Swings, J. 2003, Int. J. Sys. Evol. Microbiol., 53, 1569-1573.
44. Park, K. S., Ono, T., Rokuda, M., Jang, M. H., Iida, T. and Honda, T. 2004, Microbiol Immunol., 48, 313-318.
45. Lynch, T., Livingstone, S., Buenaventura, E., Lutter, E., Fedwick, J. and Buret, A. G. 2005, Infect. Immun., 73, 1275-1283.
46. Jones, J. L., Lüdeke, C. H. M., Bowers, J. C., Garrett, N., Fischer, M., Parsons, M. B.,
Bopp, C. A. and DePaloa, A. 2012, J. Clinical. Microbiol., 50, 2343-2352.
47. Pinckney, J. and Zingmark, R. G. 1993, Estuaries., 16, 887-897.
48. Pinckney, J., Piceno, Y. and Lovell, C. R. 1994, Diatom Res., 9, 143-153.
49. Hodson, R. E., Christian, R. R. and Maccubbin, A. E. 1984, Marine Biology, 81, 1-7.
50. Moran, M. A. and Hodson, R. E. 1989, Limnol. Oceanogr., 34, 1034-1047.
Savannah L. Klein et al.