interactions between propionic acid bacteria and thermophilic lactic
TRANSCRIPT

Lait (1999) 79, 79-92© InralElsevier, Paris
79
Original article
Interactions between propionic acid bacteriaand thermophilic lactic acid bacteria
Augustin Baer*, Irène Ryba
Dairy Research Institute, CH-3097 Liebefeld-Bem, Switzerland
Abstract - The growth of propionic acid bacteria was directly dependent on the concentration offree amino acids in cheese, regardless of the strains of lactobacilli contained in the raw mixed cultures,suggesting that the growth of these bacteria was mainly dependent on the proteolytic activity of lac-tic acid bacteria. The stimulating effect of S. thermophilus was poor probably because this species pro-duces only low amounts of amino acids in milk or cheese. Lactobacilli, which undergo cell Iysisshortly after cheese manufacture, produce large amounts of amino acids. Facultatively heterofer-mentative lactobacilli were shown to reduce the risk of secondary fermentation in Emmental cheese.This interaction was not due to their production of diacetyl, formate or acetate. Propionibacteriaable to grow at 14 "C have a greater tendency to induce secondary fermentation compared to thoseunable to grow at this temperature. © InralElsevier, Paris.
propionic acid bacteria / lactic acid bacteria / facultatively heterofermentative Iactobacilli /interaction / secondary fermentation
Résumé - Interaction entre les bactéries propioniques et les bactéries lactiques thermophiles.L'étude présentée a porté sur l'interaction entre les bactéries propioniques (BP) et 23 levains lactiques,30 de leurs souches isolées, 36 souches isolées de produits laitiers fabriqués hors de la Suisse, une cul-ture de S. thermophilus ainsi que la flore naturelle de deux laits de mélange. La croissance des bac-téries lactiques a été réalisée dans des expériences de simulation de fabrication fromagère dans du laitUHT, dont le pH a été maintenu au niveau de celui du fromage par l'ajout de carbonate de calcium.Les lactosérums obtenus ont été utilisés comme source d'azote pour la croissance des BP. Les résul-tats ont montré que l'intensité de la croissance des BP variait en fonction du lactosérum utilisé. Tou-tefois, les différences observées ne dépendaient pas spécifiquement de la souche des bactéries lactiques,mais uniquement de la concentration en acides aminés. Les différences entre les intensités de crois-sance ont pu être supprimées en uniformisant la concentration des acides aminés ajoutés, ce qui sug-gère que la croissance des BP dépend essentiellement de l'activité protéolytique des bactéries lactiques.L'effet stimulant de S. thermophilus sur la croissance des BP a été peu prononcé à cause de la faibleconcentration des acides aminés libérés. Par contre, les lactobacilles, dont la lyse cellulaire apparaîtrapidement après la fabrication du fromage, produisent de grandes quantités d'acides aminés quistimulent la croissance des BP. Il a été démontré que l'addition de lactobacilles hétérofermentaires
Oral communication at the 2nd Symposium on Propionibacteria, Cork, Ireland, June 25-27, 1998.* Correspondence and reprints. [email protected]

80 A. Baer, I. Ryba
facultatifs aux levains pouvaient réduire le risque de fermentation secondaire dans les fromagesd'Emmental en diminuant l'intensité de la croissance des BP. Cette interaction n'est pas due à laproduction de diacétyle, formiate ou acétate par les lactobacilles hétérofermentaires. Les BP capablesde croître à 14 "C ont tendance à induire la fermentation secondaire, ce qui n'est pas le cas dessouches de BP incapables de croître à cette basse température. © InralElsevier, Paris.
bactérie propionique / lactobacille thermophile / lactobacille hétérofermentaire facultatif /interaction / fermentation secondaire
1. INTRODUCTION
In Switzerland, Emmental cheese is man-ufactured exclusively from raw milk. Thethermophilic raw mixed-cultures (RMC)used contain approximately the same num-bers of S. thermophilus and L. delbrueckiissp. lactis. These cultures were collectedfrom naturally soured raw milks, used asstarters in chee se factories 40 years ago andchecked for their quality by manufacturingexperimental cheeses. Subcultures of thebest of the se cultures, containing S. ther-mophilus and L. delbrueckii ssp. lactis only,were kept frozen or Iyophilized. A liquidmixed-culture of propionic acid bacteria(PAB) is also added to the milk. PAB growduring ripening and confer the typical nuttytas te to this type of chee se [II, 12, 16].However, PAB are also responsible for twofaults, brown spots and secondary fermen-tation, which are mostly encountered incheese manufactured in winter.The causeof the appearance of brown spots is due toinsufficient growth of PAB [1] and this faulthas almost disappeared from Emmentalcheese, whereas it is still observed in semi-hard, Swiss cheeses such as Appenzell,probably due to contaminating PAB in themilk. Secondary fermentation, thought tobe caused by excessive growth ofPAB dur-ing ripening [24, 26], occurs sporadicallyand alters the quality of Emmental cheese.Raw mixed cultures favouring late fermen-tation contain L. helveticus, suggesting thatthe high proteolytic activity of this bacteriumproduces nitrogenous compounds whichaccelerate the growth rate of PAB at the
beginning of ripening [14, 19,20]. It hasalso been observed that the addition of fac-ultatively heterofermentative lactobacilli(FHL) to the milk reduces the risk of sec-ondary fermentation [23] and growth ofPAB [15, 17]. These observations indicatethat sorne lactic acid bacteria may have aspecifie influence on the growth of PAB.
The purpose of this work was to studythe influence of the proteolytic activity ofthe natural milk f1ora, and single and mixedstrains of thermophilic lactic acid bacteria(LAB) on the growth of PAB, during simu-lated cheese manufacture. These conditionsare important since rennet participates inchee se proteolysis at the very beginning ofcheese manufacture; the presence of resid-ual chymosin activity has been shown inEmmental cheese, in spite of the high cook-ing tempe rature used [5]. Rennet can alsoinfluence the growth of raw mixed culturebacteria [3], contrary to our earlier resultsobtained with a raw mixed culture [2]. Theseexperimental conditions were not taken intoconsideration in similar studies on the influ-ence of lactic acid bacteria on PAB growth[7,20,22].
2. MATERIALS AND METHODS
2.1. Chemicals
Ali chemicals were of analytical grade. Tryp-ticase peptone was from Difco (Chemie Brun-schwig, Basel, Switzerland). Other chemicalswere from Merck (Merck-Schuchardt, Zürich,Switzerland). Sterile filters of 0.20 um and0.45 um were from Sartorius (Sartorius AG, GÔ!-

Interactions Propionibacteria and Lactobacilli
tingen, Gerrnany). Resazurin was purchased fromSigma (Division of Fluka Chemie, Buchs,Switzerland).
2.2. Bacteria
The PAB used are mixed cultures which haverecently replaced the old PAB culture used sincethe end of the thirties. The PAB, as weil as23 RMCs containing approximately the samenumber of S. thermophilus and L. delbrueckiissp lactis and the pure cultures of S. thennophilus,L. rhamnosus, and L. casei were obtained fromour Department of Microbiology. The PAB andRMCs used are produced exclusively by the Fed-erai Dairy Research Institute. Thirty pure cul-tures of L. delbrueckii ssp lactis and 3 strains ofS. thermophilus were isolated from 10 RMCs.Thirty six strains of therrnophilic lactic acid bac-teria were isolated from various dairy productsobtained from Italy, Portugal, Minorca, Turkey,Tadjikistan, Slovenia and Siberia.
2.3. Growth ofPAB
PAB were grown in 10 mL of the definedmedium,described by Crow [9], modified byommission of cysteine and (NH4)2S04 so as toavoid growth of PAB in the absence of a nitrogensource. The filter-sterilized wheys obtained fromthe simulation assay (1.0 mL·1 0 mL -1 medium)were used as a nitrogen source and the growthof the bacteria was followed photometrically at650 nm [2].
2.4. Simulation assay
Simulation assays using RMCs were per-formed in screw capped 50-mL vials, asdescribed elsewhere [2, 3]. Simulation assaysusing pure cultures of lactic acid bacteria wasrnodified as follows: 50-mL glass vials stopperedwith but yI rubber stoppers to which 1.2 g ofCaCO] and a magnetic bar were added and ster-ilized. The assay was performed in 40 mL UHTmilk containing 100 ug filter-sterilized resazurinas a redox indicator. After addition of 8 ul, liq-uid rennet, to coagulate the milk in approximately26 min, and breaking the curd by vigorouslyshaking the vial, the head-space of the vial wastlushed with sterile filtered CO2 for 20 min. Thetemperature profile used in Emmental manufac-ture over the first 24 h, (heating to 53 "C in
81
35 min, maintaining this temperature for 45 min,and cooling to 30 "C in 22 h) was used. The vialswere then incubated at 30 "C over 12 d. Exceptfor the renneting phase, the simulations werestirred constantly during incubation.
2.5. Determinationof the proteolytic activity
Free amino acids (FAA) were deterrnined inthe whey of the simulation assays using cad-mium ninhydrin [4]. X-Prolyl dipeptidyl pepti-dase activity was deterrnined according to Caseyand Meyer [8], using Gly-Pro-AMC (BachemAG, 4416 Bubendorf, Switzerland) as substrate.
3. RESULTS AND DISCUSSION
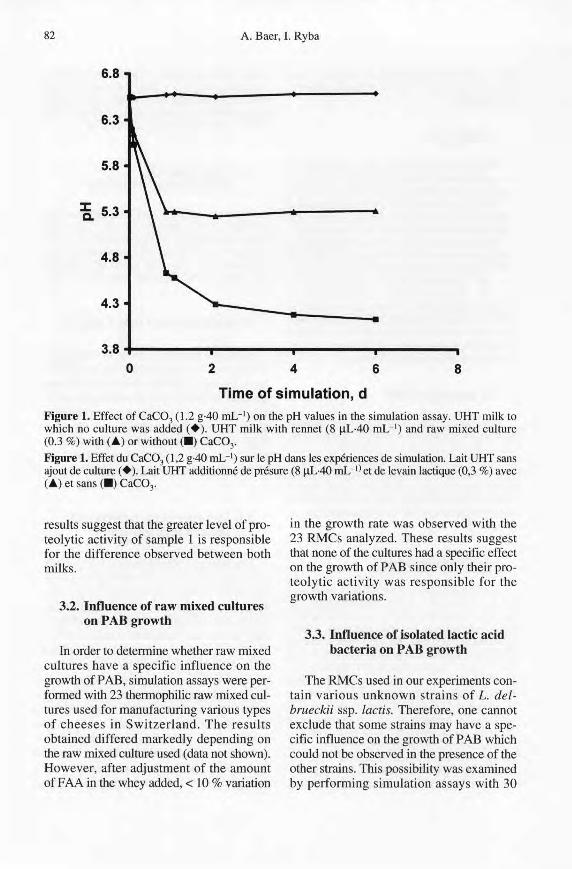
Since the growth of LAB in simulationassays can vary depending on the RMCsused, leading to the production of differentamounts of lactic acid, it was important toascertain that the pH value was similar tothat of chee se at the beginning of ripening.This condition was obtained, even with thestrongest lactic acid producing RMC, byadding CaC03 and mixing (figure 1).
3.1. Influence of the natural milkflora on PAB growth
The influence of the natural flora of rawmilk was studied by incubating two bulkraw milks, to which rennet and CaC03 wereadded, at 30 "C for 4 d. After incubation,the microflora and the concentration of FAAand lactate were measured (table 1). Wheyfrom sample 1 stimulated the growth ofPAB-A much more than that from sample 2(figure 2A). When the volume of whey fromsample 1 was adjusted in order to obtain thesame final concentration of ami no acids ineach test tube, the two growth curves werevery similar (figure 2B), and independentof the bacteria responsible for the productionof amino acids. Sample 1 contained about5 times more FAA than sample 2. These
82
6.8
6.3
5.8
i 5.3
4.8
4.3
3.80
A. Baer, I.Ryba
2 4 6 8
lime of simulation, dFigure 1. Effect of CaCa3 (1.2 g·40 mL-1) on the pH values in the simulation assay. UHT milk towhich no culture was added (+). UHT milk with rennet (8 IlL·40 ml..") and raw mixed culture(0.3 %) with (.~) or without (.) CaCa3•
Figure 1. Effet du CaCa3 (1,2 g-40 mL-I) sur le pH dans les expériences de simulation. Lait UHT sansajout de culture (+). Lait UHT additionné de présure (8 IlL·40 mL-1) et de levain lactique (0,3 %) avec(~) et sans (.) CaCa3.
results suggest that the greater level of pro-teolytic activity of sample 1 is responsiblefor the difference observed between bothmilks.
3.2. Influence of raw rnixed cultureson PAB growth
In order to determine whether raw mixedcultures have a specifie influence on thegrowth of PAB, simulation assays were per-formed with 23 thermophilic raw mixed cul-tures used for manufacturing various typesof cheeses in Switzerland. The resultsobtained differed markedly depending onthe raw mixed culture used (data not shown).However, after adjustment of the amountof FAA in the whey added, < lO % variation
in the growth rate was observed with the23 RMCs analyzed. These results suggestthat none of the cultures had a specifie effecton the growth of PAB since only their pro-teolytic activity was responsible for thegrowth variations.
3.3. Influence of isolated lactic acidbacteria on PAB growth
The RMCs used in our experiments con-tain various unknown strains of L. del-brueckii ssp. lactis. Therefore, one cannotexclude that sorne strains may have a spe-cifie influence on the growth of PAB whichcould not be observed in the presence of theother strains. This possibility was examinedby performing simulation assays with 30

Interactions Propionibacteria and Lactobacilli 83
0- vi § ~ i ~ pure cultures of L. delbrueckii ssp. lactis"uv '"cv. » E;'g.S! ~ isolated from 10 RMCs. However, the"0 -
E.::: If) 0\ <1) '"C ..... ..0 01)
N t: o ~ 0 '- growth of the lactobacilli was not satisfac-E g 00 »'" tr. a'~ ;;;<"') -a ~ ~ ?f ê ~ tory and gave nonreproducible results. Since• v t: ~<r;- '" '<1) .- , ~ ::s
< CIl;" ~~ e s satisfactory growth of lactobacilli wasu, t: ~.- t: l-<:::::I .. u obtained with RMCs containing strepto-] ~ o U'~ 0
..... ~ ('f'),~
v- ~ ~ c ~ ....... cocci, 3 pure cultures of S. thermophilus,~ , 2~ ~ .è?? eS....J "'Tt-- lSg - u 00 .... also isolated from the RMCs, were addedu"':' .9 >'("1') ~'" 0....J E ~ ,,~ .::::§ ..~ 0 to the pure cultures of lactobacilli to simu-JE 0:.;' ~ c'~:O
("')~ ~~..ê e late the RMCs. The results obtained were~ t: ::a § '~'~ similar to those obtained with RMCs. Itwas'" '""0 ..'~'û::r: ~::l
v- -e- "0 '2 s ~.-; therefore assumed that the streptococci pro-~ , .. '"~:: 0'<1) ll.) U ::s ("1') duced substances which favoured the growthr- 00 "-<.n ~ 0 en ...0 "0 ::l t:~....lU
'" 0 <"') <1) U "'~Œo of the lactobacilli. The lactobacilli were then....J E ~ t: • t: 0'" .-Cl E .n '<1) "0<1) ("')grown in whey obtained after simulation::l ~ ("1"') >.. -: , es
U '<1) .. - <1)t: ~ ~~rn--< assays performed with streptococci alone.- t:"0 0 u.~.S! U
'c t:~o "8-eC: but the results again were not satisfactorySS "'E-<g'j "o::r: ~ ~'~ ~ as they were similar to those obtained in theen uOJ.) x x .g~~ -d' § .~'v o ' "g.'§ absence of streptococci. Assuming that theâ 8~ If) <"') ro('f')o..c0000 ~- 001-011,)0
t: -'<: <1) 7 tE ....;,~ effect of streptococci on the growth of lac-:; ~ :;:"'O~=,. ';:: ::r: ("') '"
-civ E-<....lS ~~c.5~ tobacilli may be due to an improvement inOJ.)
v t: ::r:="3 cOo :;: the anaerobie growth conditions produced0("') ..t:
'" '" '" ~OU ....l,";'g g.::l ~ ~ .,;-...:3 by the NADH-oxidase of streptococci [21],'" E :.2 2.J oh ~ ::r:.. '"~ LI.. ro ,::'j '<1.)
v 0. -o E C'! ,;; CIl t: simulation assays were performed by flush-ï@ '0 0_ ::;;:: Vi d '~ ;;;
"" o-~ c: ..... 0 dl ing the head space of the simulation vial..>:: '" v OJ.) X x "'t-~~ ,S! § ~ E::l E .:; ... <2 ~t--CIlO E ~ 8 oo.~ with COz' Under these conditions, theu NU".D tJ .~ o --:d~ § ~ ~ §
::: .D /\ ~.C; ,~ U -'" ~ e!' growth of lactobacilli was similar tç that~ 0
'" ... U c, 0... v 8'2] .- c '- .... obtained in the presence of streptococci, as0 >< <r; -g 0 ~ ~
~::l cS a enV '- r.I'J en ci measured by the concentration of D-Iactate'0 t: <1) ~ = <1)'-
V '" '" "'011>= C (Il 'lU..cproduced. The growth curves ofPAB, usingc ......- ro '2 .E 2-5 v E ro § g
~ ~ E ~"'" '0 '" ~u.n wheys of pure cultures of lactobacilli0 v '2 -'<: .0 "'0 ... 0 c::l '" =çtî § ~,~ <1.)t: 0- OJ.) 00 E::r:j adjusted to the same concentration of amino.g .. '_ "'C . ...;'50 ... -
~~~N ..0' !i:~" acids, were very similar, whereas differ-'vs 0 ~'?'l x x0 C t: ;:l ~'a ....l .. o~U0. ,- "''''" "'T If) ....l::::;Œ",
o Vi co ences were observed when wheys were usedE "8 ... u o\",f ~ Ij) 000
v U E '" ("')0 .. '2 =E:::15
~.~ ~ A~ V before adjustment of the concentration ofu u ~o::~
OJ 's ~ ~::],~ ('f') -§ 'ü d) 0 arnino acids (data not shown). These resultsE [) E1·-u ô:) CIl $-o~oo-§ confirm those obtained with RMCs and'5h ê ~'B·5
0 v'" -o
<1)'- U = ~>.g·a,[)could be due to the strains of lactobacilliô ::l 00 ~ .g.~ ~ 1::::::: - 1: ~0- ....JI ............ CI') ~ CI:l ~ t:::E 's .- - Q) Q) present in the RMCs, which may have been0 ::r:O!J x x g. ~.~ ~ 0:: :EL~ a <1).... :.2 u...<2 0\ ...... ;..::::;;; E o ~2"""'~..c= selected over the years by successive cul-o u ~ 'Q) E <C c:Q)CI:lO...q-'s 0
u /\ o $-0 Q) •• Oro~§~N"
'0 :E ....lc, = < "0 "0 ';:: CIlU turing during the development of mixed cui-t: t: ::t.g ~ < E C"l ~ S 0 tures. This may explain why the character-ro 0 ~~b~ ro $-0 c: CI:l00
OJ '';:: ] ~ 2:; ~~O~('f') istics of the strains pre senti y used are very'vs -o -o EU~§/~U '(3 - 00 ~~]~'S 0 ::3 0 .- $-0 Q)similar.0. ",' ~oo~~;.§.D'?'l c: 'Q) >.. ~
v E 92 x x e ~ ~ s Q) ('f') \-0 0 ::3..c 0 [)~~.§~ In order to study a more heterogeneousu u u u ~ '~ ';j 'Ë,2 '" /\ r. ~ \-0 \-0 ~ 1)
....J 1) c..:;'j Q :;'jQ)..cQ
co ... ~ E'=: .- Oû-O tU collection of lactobacilli, 36 strains, able to~ ~ g ~ ::: ~ c: ;[i c:... ::l g ~ ~ ~:~ grow at 45 "C and only producing D-Iac-~ == ,-:: u, u,... ...E-,".:..i-< -g '" .,;- '" E::ë ::ë ~ - - N c'j ~ tate, isolated from milk products manufac-~ ~ :§ ~~::r:~ o 0 \-0 Oû c:
E-o E-o -N E-<....l"-"- j'2~ 5~ tured outside Switzerland, were evaluated

0.6
EO.S
1:Cl."
0.4caC0ai 0.3ceD..-0oC 0.2~0..C)
0.1
00 10 20 30 40 so
Time of incubation, h
Figure 2. Influence of the natural flora of two milks on PAB growth. Before (A) and after (B) adjust-ment of the concentration of amino acids. Milk 1 (.); milk 2 (0). The natural flora was enriched byincubation of the milks at 30 "C for 4 d after addition of rennet (8 llL·40 mL -1) and CaCO)" In B, thevolume of the two wheys was adjusted so that the same level of amino acids was added to the definedmedium.Figure 2. Influence de la flore naturelle de deux laits de mélange sur la croissance des BP avant(A) et après (B) ajustement de la concentration en acides aminés. Lait A (.) ; lait B (0). La flore natu-relle a été enrichie par incubation des laits crus à 30 "C pendant 4 j, après adjonction de présure(811L·40 ml..") et de CaCO)" En B, le volume des deux lactosérums a été ajusté.pour ajouter lamême concentration d'acides aminés au milieu défini.

Interactions Propionibacteria and Lactobacilli
20 30lime of incubation, h
Figure 3. Growth of PAB in the defined medium in the presence of peptone (l g·L-I) (.~), wheyafter the growth of S. thermophilus in the simulation assay (0); further addition to the whey of3 mrnol-L -lof a mixture of pure amino acids, corresponding to the concentrations of FAA determinedin Emmental cheese (.).Figure 3. Croissance des BP dans le milieu défini en présence de peptone(l g·L-I) (.6.), lactosérumobtenu après croissance de S. thermophilus en expérience de simulation (0) et après addition sup-plémentaire au lactosérum de 3 mmol-L:' d'une solution d'acides aminés purs, correspondant auxconcentrations des acides aminés libres dans un fromage d'emmental (.).
in the simulation assay. Again the resultsobtained showed that the amounts of liber-ated amino acids varied depending on thestrains examined but no difference in thegrowth of PAB could be detected when theamino acid concentrations were adjusted tothe same level (data not shown). Theseresults confirm those obtained with RMCsand isolates from the same RMCs, suggest-ing that, under our experimental conditions,free amino acids are the sole factor influ-encing PAB growth.
3.4. Influence of S. thermophiluson PAB growth
A simulation assay using 3 strains ofS. thermophilus isolated from RMCs was
1.4
1.2
E1: 10II)(1)
C0 0.8
aictno
0.6....0.c:~0 0.4...
C)
0.2
10
85
performed. The amount of FAA present inthe whey after 4-d incubation at 30 "C wasequivalent to 3.2 mmol-leucine-L:'. At thislow concentration, which is almost the sameas the concentration of FAA in milk, thegrowth of PAB was poor when comparedto the growth observed on addition of pep-tone (1.0 g·L-I). However, better growthwas observed after further addition of3 mmol of a mixture of amino acids, corre-sponding to the concentrations of FAAfound in Emmental cheese [25] (figure 3),demonstrating that amino acids are probablythe limiting factor for PAB growth in ourexperimental conditions. Important varia-tions in the stimulatory effect of S. ther-mophilus have previously been describedby Piveteau et al. [20] who have shown that
40 50

86 A. Baer, 1. Ryba
stimulation was dependent on both the strainof S. thermophilus as weil as the PAB strainused. The low production of FAA by S. ther-mophilus compared to lactobacilli may bedue to the different rate of autolysis of thetwo bacterial species. Amino peptidases areintracellular enzymes which are liberatedinto the growth medium only after celllysis[10]. Lysis of thermophilic streptococci inchee se is a slow process since it is detectedonly after 30 d of ripening in model chee se[18].
In contrast, FAA can be detected after0.7 d in the simulation assay (figure 4A).The production of FAA coincided with thelysis of lactobacilli, since significantamounts of X-prolyl dipeptidyl peptidasewere detected at the same time (figure 48).
3.5. Relationship between FAAin cheese and secondaryfermentation
Secondary fermentation in Emmentalcheese, characterized by cracks or splits inthe chee se body, is always accompanied byincreased proteolysis of casein as identifiedby high values of the trichloroacetic acidsoluble N (TCAS-N) fraction. An increasein propionate production together with adecrease in the concentration of lactic acidwas also observed in cheeses with a sec-ondary fermentation, suggesting that astronger propionic acid fermentation occurs[25, 26]. Since the intensity of propionicacid fermentation is related to PAB growth,it is assumed that the amount of FAA incheese is correlated with the intensity ofsecondary fermentation. Analyses were per-formed on 30 Emmental cheeses made onthe same day in various cheese factories,using thetwo new types of PAB cultures(20 chee ses with culture A, 10 cheeses withculture B). The raw mixed cultures usedvaried according to the choice of cheesemakers. After 24 h, the cheeses wereremoved to the same ripening cellar. Fig-ure 5 shows a plot of the degree of sec-
ondary fermentation, estimated by a pro-fessional panel on an arbitrary scale of 1,corresponding to good chee ses, to 3 for pro-nounced secondary fermentation, againstthe amount of FAA, measured after 180 d ofripening. A good correlation between thetwo parameters was found (r = 0.735),which suggests that the amount of FAAplays a key role in the development of sec-ondary fermentation. Since the TCAS-Nfraction is also increased during secondaryfermentation and since this fraction containsFAA as weil as small peptides, one cannotexclude peptides as the cause, as has beenproposed previously [2]. This assumptionwas tested by plotting the intensity of sec-ondary fermentation after 180 d of ripeningagainst the values obtained by substractingFAA values (mrnol-kg ") from TCAS-Nvalues (mmol-kg ") to obtain the amount ofpeptides. As can be seen fromfigure 6, nocorrelation exists between the two param-eters.
According to these results, PAB clearlyprefer FAA to peptides for their growth incheese, which conflicts with a previousstudy [2]. In our previous work, we showedthat commercial casarnino acids, consideredas a FAA source, resulted in competitiveinhibition on PAB growth, when added topeptone. However, it was discovered, afterpublication, that addition of the pure aminoacids did not cause inhibition, suggestingthat the inhibition was due to sorne sub-stance in the casamino solution, such as salt,which is present at high levels in casaminoacids (unpublished results).
3.6. Influence of facultativelyheterofermentative lactobacillion PAB growth
It has been shown that the addition offacultatively heterofermentative lactobacilli(FHL), such as L. casei or L. rhamnosus,together with thermophilic raw mixed cul-tures reduces, but does not completely sup-press the risk of secondary fermentation in

30
.... 25:.....cD.520u:::1Q)..Jcr 15W'0E 10E
~ 5LI.
00 1
Time of simulation, d
14
12
t: 10:êE 8Et:
>. 6'0...Q..>< 4
2
0••
0 10 20 30 40 50 60 70Free amino acids, mmol Eq. Leucine.ml,"
Interactions Propionibacteria and Lactobacilli 87
A
2 63 4 5
Figure 4. (A) Liberation of free amino acids by a pure culture of L. delbrueckii ssp. lactis in a sim-ulation assay as a function of incubation time. (B) Liberation of ami no acids as a function of celllysisby the same culture as in A. Celllysis was measured by the amount of X-prolyl-dipeptidyl aminopep-tidase liberated, determined by using G1y-Pro-AMC as substrate.Figure 4. (A) Libération des acides aminés dans une expérience de simulation avec une culture purede L. delbrueckii ssp. lactis en fonction du temps d'incubation. (B) libération des acides aminés enfonction de la lyse de la même culture que sous (A). La lyse cellulaire a été déterminée par la mesurede la libération de la X-prolyl-dipeptidyl aminopeptidase en utilisant le substrat Gly-Pro-AMC.

88 A. Baer, 1. Ryba
200•0)~
-..Q)C •'u:::::l 150Q)
.....J0-
UJ
0E
100E •~IL
50 '---L-L-_--'- __ '--_-'--_----L_--'
0.5 1.0 1.5 2.0 2.5 3.0 3.5Degree of secondary fermentation
Figure 5. Correlation between the degree of secondary fermentation in chee se at 180 d of ripeningand the amount of free amino acids. Secondary fermentation was estimated by a professional panelon an arbitrary scale of 1, corresponding to good cheeses, to 3 for pronounced secondary fermentation.Figure 5. Corrélation entre l'intensité de la fermentation secondaire dans le fromage après 180 j dematuration et la concentration en acides aminés. La fermentation secondaire a été estimée par unjury professionnel selon une échelle arbitraire allant de 1 pour les bons fromages, à 3 pour ceuxatteints de fermentation secondaire.
Emmental cheeses [23]. Consequentlynumerous chee se makers, in Switzerland,use FHL. In cheeses made using FHL, sec-ondary fermentation was observed in 12 outof 20 cheeses manufactured with the newPAB culture A, whereas none was detectedin 10 cheeses made with the new PAB cul-ture B. The average values of propionate,lactate and FAA measured in the 3 groups ofcheeses are given in table Il. The propionateconcentration as well as proteolysis arehigher in cheeses with secondary fermen-tation. However, secondary fermentation,accompanied by an increase of about 20 %of propionate, was observed in spite of thepresence of FHL, but only in cheeses man-ufactured with PAB culture A. This sug-gests that this mixed culture may be lesssensitive to inhibition by FHL than those ofculture B. To study this hypothesis, simu-lation experiments were performed using
L. casei and L. rhamnosus alone or in addi-tion to the most used raw mixed culture inEmmental manufactory; the wheys obtainedwere then tested for their activity on thegrowth of the two new PAB cultures. Noinhibition could be detected on either PABculture A or B. On the contrary, anincreasein FAA concentration was detected in sorneexperiments, thus favouring the growth rateof PAB. According to Jimeno et al. [15], theproduction of diacetyl, formate and acetateby FHL could be responsible for the growthinhibition of PAB. The concentration ofthese metabolites producing 5 % inhibitionon the growth of the 2 PAB cultures, wasdetermined by adding increasing amountsof metabolites to the defined growthmedium. From table Ill, one can see thatthe minimal inhibitory concentration of for-mate, diacetyl and acetate is at least 10 timeshigher than the concentrations found in

Interactions Propionibacteria and Lactobacilli
~ 450
89
•0EE •~
400 •LL.en •:::J •C •
•Ez 350
1(J) •<t:Ü1-
300 L.-_--l...-_-L __ '--_...l-_--L_--l
0.5 1.0 1.5 2.0 2.5 3.0 3.5Degree of secondary fermentation
Figure 6. Correlation between the degree of secondary fermentation in cheese at 180 d ofripeningand the amount of smaIl peptides. Peptides were determined by subtracting the values of the free aminoacids (FAA) from the nitrogen soluble in trichloroacetic acid (TCAS-N).Figure 6. Corrélation entre l'intensité de la fermentation secondaire dans le fromage de '180 j dematuration et la concentration en petits peptides. Les peptides ont été déterminés par soustraction desacides aminés (FAA) de l'azote soluble dans l'acide trichloroacétique (TCAS-N).
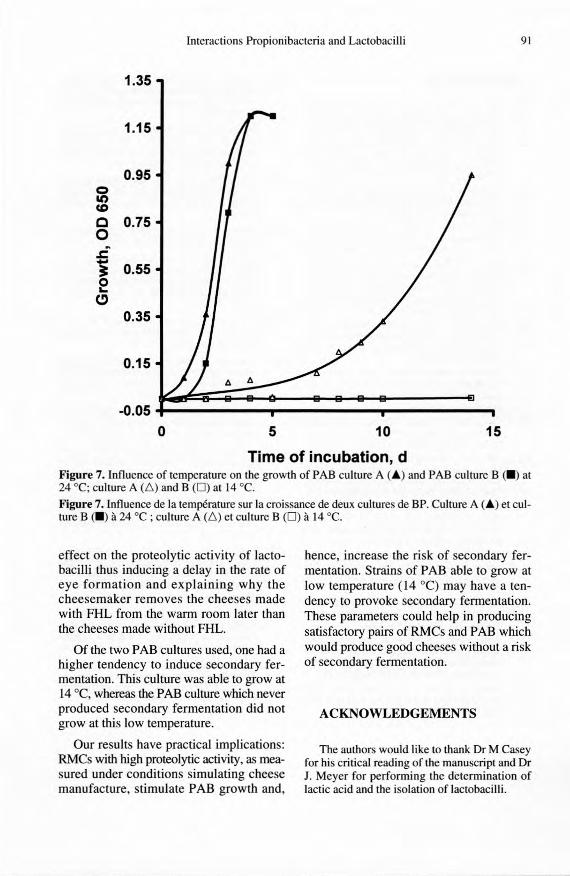
ity of cheese.They also suggest that the abil-ity of sorne PAB strains to grow at low tem-peratures should be taken into considera-tion when producing new PAB cultures.Similar results were published by Hettingaet al. [13], who demonstrated that the abil-ity to grow at low temperatures was strain-dependent.
cheese. It seems, therefore, that the se com-pounds are not responsible for the inhibi-tion of PAB in chee se. However, the con-troversial results shown in table II suggestthat the two PAB cultures may be differen-tially sensitive to factors other than FHL,e.g. temperature because secondary fer-mentation appears only after about 120 ripen-ing d of storage at 12-14 "C, suggesting thatPAB are still growing and producing CO2 atthis low temperature. The growth of the twocultures at 24 "C and 14 "C, which corre-spond to the two temperatures encounteredduring cheese ripening, was studied. Theresults (figure 7) show that a slight differ-ence in the rate of growth between the twocultures exists at 24 "C, whereas only cultureA, which induces secondary fermentation, isable to grow at 14 oc. These results demon-strate that differences in PAB strains maybe of importance in determining the qual-
4. CONCLUSION
The growth of the two new PAB culturesused in Switzerland is directly dependenton the concentration of FAA in cheese,regardless of the lactobacilli strains con-tained in the RMCs. This does not excludethe possibility that PAB preferentially useonly sorne of the FAA. Neither acetate, for-mate nor diacetyl inhibited PAB growthsuggesting that the inhibitory effect of FHL

90 A. Baer, 1. Ryba
Table II. Concentrations of lactate, propionate and free amino acids in 30 cheeses manufacturedwith the two PAB cultures recently introduced, in the presence offacultatively heterofermentative lac-tobacilli.Tableau II. Concentration en lactate, propionate et acides aminés de 30 fromages fabriqués avec lesdeux nouvelles cultures de BP récemment introduites, en présence de lactobacilles hétérofermentairesfacultatifs.
PAB culture A PAB culture A PAB culture BSecondary fermentation No secondary fermentation No secondary fermentation
FAA, mmol Leucine, mfiq-kg!3 mo ripening 94.8 75.6 85.76 mo ripening 129.6 97.2 102.3
Propionate, mmol-kg!3 mo ripening 64.3 53.8 55.46 mo ripening 80.3 67.1 60.1
D + L Lactate, mmol-kg!3 mo ripening 47.0 61.5 62.66 mo ripening 24.1 45.3 57.4
20 chee ses were manufactured with culture A, 10 with culture B. Chee ses were ripened in the sa me ripeningcellar. Twelve cheeses manufactured with culture A showed secondary fermentation; no secondary fermentationwas detected in cheeses made with culture B.20 fromages ont été fabriqués avec la culture A et 10 avec la culture B. Les fromages ont été mûris dans la mêmecave. Douze fromages fabriqués avec la culture A ont présenté une fermentation secondaire; aucune fermenta-tion secondaire n'a été détectée dans les fromages fabriqués avec la culture B.
Table III. Concentrations of sorne inhibitory products in chee se and the minimum concentrationnecessary to inhibit of PAB growth by 5 to 10 %.Tableau III. Concentrations de substances inhibitrices dans le fromage et concentration minimale inhi-bitrice de 5-10 % de la croissance des BP.
Formate Acetate NaCI Diacetyl
Concentrationin cheese
5 mmol-kg!" 43 mmol·kg-1a 150 mrnol-kg!" 7 J.lmol·kg-1C
17 J.lmol·kg-1d
200 J.lmol·kg-1e
> 50 mrnol-L"! > 330 mmol-L:' > 350 mmol-L:' > 250 umolL:'Minimum inhibitoryconcentration
aSamples were steam distilled, esterified and analyzed by head space gas chromatography according to a new unpub-lished FAM method. Les échantillons ont été distillés à la vapeur, esthérifiés et analysés par chromatographie enphase gazeuse à espace de tête, selon une nouvelle méthode non publiée de la FAM.h Determined according to lOF Norm 88A: 1988 / détermination selon la norme FIL 88 A :1998.c [26]. d [27]. e [15]. Using the method described in [6] / en utilisant la méthode décrite en [6].
is not due to a biochemical process. How-ever, analyses performed in our institutehave shown that the proteolytic activity incheeses manufactured with the addition ofFHL is less intensive than in cheeses made
without FHL since the TCA-S-N fraction,as weIl as the amounts of FAA, is lower.This explains why these cheeses are lesstasty than those made without FHL. Thissuggests that FHL may have an inhibitory
Interactions Propionibacteria and Lactobacilli 91
1.35
1.15
0.950II)CDC 0.750J:- 0.55~0L-
e>0.35
0.15
-0.050 5 10 15
Time of incubation, dFigure 7. Influence of temperature on the growth of PAB culture A (Â) and PAB culture B (.) at24 -c, culture A (l'.) and B (0) at 14 oc.Figure 7. Influence de la température sur la croissance de deux cultures de BP. Culture A (Â) et cul-ture B (.) à 24 "C ; culture A (l'.) et culture B (0) à 14 oc.
effect on the proteolytic activity of lacto-bacilli thus inducing a delay in the rate ofeye formation and explaining why thecheesemaker removes the cheeses madewith FHL from the warm room later thanthe cheeses made without FHL.
Of the two PAB cultures used, one had ahigher tendency to induce secondary fer-mentation. This culture was able to grow at14 "C, whereas the PAB culture which neverproduced secondary fermentation did notgrow at this low temperature.
Our results have practical implications:RMCs with high proteolytic activity, as mea-sured under conditions simulating cheesemanufacture, stimulate PAB growth and,
hence, increase the risk of secondary fer-mentation. Strains of PAB able to grow atlow temperature (14 OC) may have a ten-dency to provoke secondary fermentation.These parameters could help in producingsatisfactory pairs of RMCs and PAB whichwould produce good chee ses without a riskof secondary fermentation.
ACKNOWLEDGEMENTS
The authors would like to thank Dr M Caseyfor his critical reading of the manuscript and DrJ. Meyer for performing the determination oflactic acid and the isolation of lactobacilli.

92 A. Baer, I. Ryba
REFERENCES[1] Baer A., Ryba L, Grand M., Ursachen der
Entstehung von braunen Tupfen im Kâse ,Schweiz. Milchw. Forschung 22 (1993) 3-7.
[2] Baer A., Influence of casein proteolysis by starterbacteria, rennet and plasmin on the growth ofpropionibacteria in Swiss-type cheese, Lait 95(1995) 391-400.
[3] Baer A., Ryba L, Fürst M., Studie über die pro-teolytische Aktivitât von Milchsâurebakterien,Int. Report FAM 18 (1996) 1-16.
[4] Baer A., Ryba L, Meyer J., Bütikofer U.,Microplate assay of free amino acids in Swisscheeses, Food Sci. Technol. 29 (1996) 58-62.
[5] Baer A., Ryba L, Casey M., Influence desenzymes coagulants et de la plasmine sur lacroissance des levains en début de fabricationde l'emmental, étude préliminaire sur milieuxmodèles, Lait 77 (1997) 189-199.
[6] Bednarski W., Jedrychowski L., Hammond E.G.,Nikolov ZL, A method for the determination ofa-dicarbonyl compounds, J. Dairy Sci. 72(1989) 2474-2477.
[7] Brunaud D., Maillard M.B., Thierry A., Inter-actions entre Lactobacillus helveticus et Propi-onibacterium. 6e col. club bact. Lac. (1994) Lyonuniversité, poster E20.
[8] Casey M.G., Meyer J., Presence of X-pro-Iyl-dipeptidyl-peptidase in lactic acid bacteria,J. Dairy Sci. 68 (1985) 3212-3215.
[9] Crow V.L., Utilization of lactate isomers byPropionibacterium freudenreichii subsp sher-manii: regulatory role for intracellular pyruvate,App!. Environ. Microbio!. 52 (1986) 352-358.
[10] Crow V.L., Coolbear T., Gopal P.K., Mar-tley F.G., McKay L.L., Riepe H., The role ofautolysis oflactic acid bacteria in the ripening ofcheese, Int. Dairy J. 5 (1995) 855-875.
[II] Gautier M., Lortal S., Boyaval P., Girard F.,Lemée R., de Carvalho A.F., Dupuis C., Lesbactéries propioniques laitières, Lait 73 (1993)257-263.
[12] Hettinga D.H., Reinbold G.W., The propionic-acid bacteria. A review: 1. Growth, J. Milk. FoodTechno. 35 (1972) (5) 295-301.
[13J Hettinga D.H., Reinbold G.W., Vedamuthu E.R.,Split defect of Swiss cheese: 1. Effect of strain ofpropionibacterium and wrapping material,J. Milk Food Technol. 37 (1974) 322-328.
[14J Hunter J.E., Frazier W.c., Gas production byassociated Swiss cheese bacteria, J. Dairy Sei. 44(1961) 2176-2186.
[15] Jimeno J., Lazaro M.J., Sollberger H., Antago-nistic interactions between propionic acid bac-teria and non-starter lactic acid bacteria, Lait 75(1995) 401-413.
[16] Langsrud T., Reinbold G.W., Flavour develop-ment and microbiology of Swiss cheese:III. Ripening and tlavour production, J. MilkFood Techno!. 36 (1973) 593-609.
[17] Martley EG., Crow, V.L., Interactions betweennon-starter microorganisms during cheese man-ufacture and ripening, Int. Dairy J. 3 (1993)461-483.
[18] Meyer J., Lyse bacterienne dans des fromages delaboratoire, Int. Report FAM, 2 (1993) 1-Q.
[19] Perez Chai a A., Pesee de Ruiz Hoigado A.P.,Oliver G., Interaction between Lactobacillushelveticus and Propionibacterium freudenre-ichii subsp. Shermanii, Microbio!. Alim. Nutr. 5(1982) 325-331.
[20] Piveteau P.G., Condon S., Cogan T.M., Inter-actions between lactic and propionic acid bac-teria, Lait 75 (1995) 331-343.
[21] Sakamoto M., Komagata K., Aerobic growthand activities of NADH oxydase and NADHperoxydase in lactic acid bacteria, J. Ferment.Bioeng. 82 (1996) 210-216.
[22] Salvat-Brunaud D., Influence des bactéries lac-tiques thermophiles sur les bactéries propi-oniques, thesis, Inra Rennes, France, 1997.
[23] Sollberger H., Der Eintluss fakultativ hetero-fermentativer Milchsiiurebakterien auf die Pro-pionsâuregârung und die Kâsequal itât imEmmentaler, Int. Report FAM 31 (1990) 1-17.
[24] Steffen C, Puhan Z., Môgliche Zusammenhângezwischen Eiweissabbau und Nachgârung imEmmentaler Kâse, Schweiz. Milchw. Forschung,5 (1976) 1-4.
[25] Steffen c., Glattli H., Nick B., VergleichendeUntersuchungen in Emmentalerkâse mit undohne Nachgârung: III. Bakteriologische undenzymatische Untersuchungen, Schweiz.Milchw. Forschung 8 (1979) 19-26.
[26] Steiger G., Flückiger E., Vergleichende Unter-suchungen in Emmentalerkâse mit und oh neNachgârung: V. Chemische und physikalischeUntersuchungen, Schweiz. Milchw. Forschung8 (1979) 39-43.
[27] Zerfiridis G.K., Vafopoulou-Mastrogiannaki A.,Litopoulu-Tzanetaki E., Changes during ripen-ing of commercial Gruyère cheese, J. Dairy Sei,67 (1984) 1397-1405.