interactions between gastrointestinal parasites and the gut...
TRANSCRIPT

GHENT UNIVERSITY
FACULTY OF VETERINARY MEDICINE
Academic year 2015 - 2016
Interactions between gastrointestinal parasites and the gut
microflora
by
Hannah JONES
Promoters: Prof. dr. Peter Geldhof Literature Review
Drs. Oonagh Paerewijck as part of the Master's Dissertation
© 2016 Hannah JONES


Disclaimer
Universiteit Gent, its employees and/or students, give no warranty that the information provided in this
thesis is accurate or exhaustive, nor that the content of this thesis will not constitute or result in any
infringement of third-party rights.
Universiteit Gent, its employees and/or students do not accept any liability or responsibility for any use
which may be made of the content or information given in the thesis, nor for any reliance which may
be placed on any advice or information provided in this thesis.

GHENT UNIVERSITY
FACULTY OF VETERINARY MEDICINE
Academic year 2015 - 2016
Interactions between gastrointestinal parasites and the gut
microflora
by
Hannah JONES
Promoters: Prof. dr. Peter Geldhof Literature Review
Drs. Oonagh Paerewijck as part of the Master's Dissertation
© 2016 Hannah JONES

PREFACE
I would like to thank my promotor for his supervision and guidance throughout my work. A
special thank you to my parents/family and Rolf Koenen for their support and help, not only
when working on this thesis, but throughout my entire study during the past few years.

ABSTRACT ........................................................................................................................... 1
SAMENVATTING .................................................................................................................. 1
LITERATURE STUDY ........................................................................................................... 3
1. INTRODUCTION ........................................................................................................ 3
2. EFFECT PER SPECIES ............................................................................................. 5
2.1. PIGS........................................................................................................................ 5
2.2. RODENTS ............................................................................................................... 8
2.2.1. Rats .................................................................................................................. 8
2.2.2. Mice .................................................................................................................. 8
2.2.2.1. Trichuris muris infections ............................................................................ 8
2.2.2.2 Heligmosomoides polygyrus bakeri infections ............................................10
2.3 HUMANS .................................................................................................................14
2.3.1 Protozoans ........................................................................................................14
2.3.2 Helminths ..........................................................................................................14
2.4 DOGS AND CATS ...................................................................................................18
2.5 RUMINANTS ...........................................................................................................19
3. DISCUSSION ............................................................................................................20
4. REFERENCES ..........................................................................................................22

1
ABSTRACT The intestinal microbiome is a unique ecosystem which is of great importance for the overall health of
an individual, forming a symbiotic relationship between host and its microbiota. It is known that the
microbiome has an influence on the host’s immune system, regulating it and forming a crosstalk
interaction. As intestinal parasites can survive in the same niche, and have been thought to alter the
immune system directly, so it was hypothesised that they possibly had an influence on the gut
microbiome too. In this relatively new research topic, several studies have shown that there is
crosstalk between parasites and the intestinal microbiota of the hosts they invade. Studies using
experimental infections with helminths in pigs, small ruminants and rodents have been executed to
see if a shift in microbiota occurred as a result of the infection. In most studies there were quite some
changes on species-level, rather than a shift in diversity of the microbiota. In humans, dogs and cats,
the studies were performed on naturally infected individuals. Shifts were also noted in different
metabolic pathways, such as reduced carbohydrate metabolism, for instance, in Trichuris suis infected
pigs. Pigs, and their intestinal helminth Trichuris suis, are often used as a good model for the human
infection with Trichuris trichiura.
SAMENVATTING Het onderzoeken of er een interactie is tussen gastro-intestinale parasieten en het microbioom van de
gastheer dat ze koloniseren, is een redelijk recent onderwerp. Men is er steeds vanuit gegaan dat
parasieten een directe invloed hadden op de gastheer zelf, met o.a. het aantasten van de mucus en
epitheliale wand in het gastro-intestinaal stelsel, waardoor er een reactie ontstaat van het
immuunsysteem van de gastheer.
Met deze artikels wordt de hypothese aangekaart dat er een eventuele wisselwerking is tussen het
immuunsysteem van de gastheer, de parasiet en de microbiota, waardoor het koloniseren van de
gastheer door een parasiet een heel stuk complexer zou verlopen dan tot nu toe werd gedacht.
De verschillende artikels bespreken de invloeden van verschillende parasieten op de microbiota bij
varkens, ratten, muizen, geiten, honden, katten en ook bij de mens. Bij infectie met verschillende
parasieten, voornamelijk helminthen, werd er aangetoond dat er wel degelijk een verandering aan de
samenstelling van de intestinale microbiota ontstaat. Bij varkens die geïnfecteerd werden met de
helminth Trichuris suis is er bijvoorbeeld een verhoging in Mucispirillum en zijn er veranderingen in
verschillende metabole wegen ontdekt, en zelfs een verhoging van Campylobacter in geïnfecteerde
dieren. Bij ratten en muizen als proefdieren werden er vooral veranderingen ter hoogte van de
Firmicutes en Lactobacillaceae gevonden, die sterk verhoogd waren in met Trichuris muris
geïnfecteerde muizen en in met Hymenolepis diminuta geïnfecteerde ratten. Bij proefdieren
geïnfecteerd met de helminth Heligmosomoides polygyrus is aangetoond dat er zowel een
verschuiving naar meer Enterobacteriaceae als naar Lactobacillaceae was in de microbiota van de
dunne darm. H. polygyrus zorgde er ook voor dat er meer korte keten vrije-vetzuren geproduceerd
werden, wat ervoor zorgde dat de muizen minder gevoelig werden voor bepaalde allergieën zoals o.a.

2
asthma. De aanwezigheid van bepaalde species in de normale microbiota bepalen de overleving van
deze helminth in de darm van de gastheer.
Bij de mens werden verschillende studies uitgevoerd. In 2 studies werd er geen verschil aangetoond
tussen geïnfecteerde mensen met o.a. Trichuris trichiuria en Necator americanus en niet-
geïnfecteerde mensen. Er was wel een duidelijk verschil te zien in de samenstelling van de microbiota
van een groep kinderen in Ecuador en westerlingen. Drie studies vonden wel een shift in het
microbioom en ook veranderingen in metabolische pathways, net zoals dit gezien werd bij de varkens
en andere proefdieren.
Carnivoren, zoals honden en katten, en herkauwers werden eveneens bestudeerd en met duidelijk
resultaat. Giardia duodenalis, Ancylostoma caninum en Cystoisospora infecties bij carnivoren gaven
een verhoogde aanwezigheid van Firmicutes en een verlaagde aanwezigheid van Bacteroidetes in
geïnfecteerde honden, en een hogere aanwezigheid van Bifidobacterium in geïnfecteerde katten. De
verhoogde aanwezigheid van Firmicutes zorgde hier ook voor verhoogde productie van korte keten
vrije-vetzuren, net zoals dit het geval was bij H. polygyrus infecties bij de muis.
Een belangrijk gevolg bij infectie met Haemonchus contortus bij geiten was de verhoging van de pH in
de lebmaag. Hierdoor kan de helminth beter overleven. De aanwezigheid van deze helminth zorgde
ook voor een shift naar minder Butyrivibrio in geïnfecteerde geiten, waardoor de butyraat biosynthese,
dat van belang is voor het dempen van ontstekingsreacties, in het gedrang kwam.
Naast de interacties die hier besproken werden tussen de parasiet en het microbioom zijn er nog
andere verbanden die in acht moeten genomen worden tussen de gastheer en het microbioom, de
parasiet en de gastheer-immuniteit.

3
LITERATURE STUDY
1. INTRODUCTION
The intestinal tract in mammals has a very unique ecosystem of microorganisms, which is referred to
as the ‘microbiome’. The most important bacterial phyla are those of the Firmicutes, Bacteroidetes,
Actinobacteria and Proteobacteria (Kinross et al., 2011). We know that the gut-microbiota is vital to the
overall health of an individual, and that the relationship between host and microbiome is a symbiotic
one, with both parties benefiting from each other (Gandolfo, 2015). The microorganisms rely on the
host for nutrition which is presented to them in a form of mucus to support the bacterial metabolism,
and in return the host relies on them for some key functions related to nutrition, education of the
immune system and prevention of infection and disease (Wu et al., 2013). In essence, the microbiome
forms a unique ‘organ’ and delivers a significant contribution to the metabolism and production of our
much needed and essential vitamins and energy (Smits et al., 2014). The composition of this
microbiome is different and unique in each individual, even between individuals of the same species.
When the microbial composition changes in time of disease, the term dysbiosis is used. The
alternation in composition of the microbiome, and thus, the forming of a dysbiosis can be influenced by
diet, environment, ethnicity, genetics and medication use, especially the use of antibiotics. When using
antibiotics, a shift of bacteria takes place, with a decrease in a few key species and a decrease in the
overall microbial diversity. In a situation like this, it is possible for other, potentially pathogenic, species
to thrive and multiply and therefore create an unhealthy microbiome (Gandolfo, 2015). If there is a
case of dysbiosis, the digestion and absorption of nutrients will be adversely affected too, which can
result in a lower feeding efficiency and lower growth rates. The gut epithelium could also become
damaged, which results in a higher chance of infection (Smits et al., 2014).
The microbiome is also necessary for the regulation and programming of the immune system. This
happens through a ‘cross-talk’ mechanism between the microbiota and the mucosal immune system of
the gut, especially in the jejunum and ileum through the Peyer’s patches. The colon and caecum
microbiota play an important role in the fermentation process of the host’s nutrients. The competence
of the microbiome determines whether and how the body will react to harmful components, but at the
same time it determines whether the body should not react to different components. The latter is called
immunotolerance. This equilibrium is partially directed and induced through microbial components,
several of the body’s own components and food particles. In this way the body constantly screens the
intestines for foreign components that it would need to react against, or not. This screening happens
through receptors, with the most important being the Toll-Like-Receptors on the surface of the gut
epithelium (Smits et al., 2014).
This thesis will mainly focus on intestinal parasites, as they seem to have an influence on the
gastrointestinal microbiome. Most common gastrointestinal parasites are the Helminths, which are
multicellular organisms and can be divided into 2 groups, the Nematodes (roundworms), which are by
far the most common infectious worms in animals and humans, and then the group of the
Platyhelminths. The latter consists of Trematodes (flukes), and Cestodes (tapeworms) (van Riet et al.,

4
2007). Unicellular parasites, such as Giardia and Cryptosporidium can also colonise the gut and cause
damage directly to the epithelial barrier.
At this point in this relatively new research area, the question arises whether parasites have an effect
on the microbiome and, conversely, if the microbiome has an effect on the intestinal parasites it holds.

5
2. EFFECT PER SPECIES
2.1. PIGS
Trichuris suis is a gastrointestinal helminth in pigs, which is often used in experimental infections in
pigs as a model for Trichuris trichiura infections in humans.
In Wu et al. (2012), pigs were experimentally infected with T. suis for 21 days, and among these
infected pigs a first group had adult worms and the second group were worm-free, and each group
had a control group. The overall majority of the bacterial phyla in control pigs were Bacteroidetes,
Firmicutes, Spirochaetes,and Proteobacteria, in descending order of abundance and counting for
>99,4% of the assigned sequences in the pig’s proximal colon microbiome, which can be assigned to
as the ‘core microbiota’. Of these phyla, Prevotella was most common, then the genus of
Oscillobacter, followed by Treponema, Succinivibrio, Anaerovibrio, and Roseburia (Wu et al., 2012, Li
et al., 2012).
In a study by Li et al. (2012), the changes in porcine proximal colon microflora with an experimental
infection with Trichuris suis was examined by using metagenomics tools, such as gas
chromatography-mass spectrometry (GC-MS). Seven female piglets were taken from four different
litters. They were all placed in the same conditions and were fed the same feed, and no antibiotics
were used during the study. Four piglets were inoculated with a single dose (2*104
eggs/piglet) of
infective T. suis eggs at the age of 3 months old (Hill et al., 1993). The other 3 pigs received a dose of
the supporting medium of phosphate-buffered saline (PBS) as a placebo, and therefore served as
uninfected controls. They were all killed 21 days post inoculation and the luminal contents of the
colons were collected and examined. Different changes and shifts in metabolic potentials of the host’s
proximal colon microbiota were observed as a result of infection with T. suis. These included
categories such as general function repair, replication, recombination and repair, carbohydrate
transport and metabolism and cell wall/membrane/envelope biogenesis (Li et al., 2012).
Both studies showed significant changes in the microbiome composition. First of all, the study by Li et
al. (2012) noted that the infected pigs had a significant change in composition on both phylum and
genus levels compared to the control pigs (table 1). Especially two phyla were significantly altered by
infection, these were the Proteobacteria and Deferribacteres. Of the 25 classes and 64 families,
abundances of two classes and seven families were significantly impacted by infection (p<0,05). Also
17 genera, which is about 13% of all genera present, were also significantly affected (p<0,05). For
example, the relative abundance of Oscillibacter, which was the second-most-abundant genus in the
proximal colon of the control pigs, decreased from 7,8% to 2,8% in infected pigs. A second example
was the abundance of Succinivibrio, which decreased from 3,6% in control pigs to 0,4% in infected
pigs. There was an 86-fold increase in the relative abundance of Mucispirillium to 0,09% in infected
pigs. In conclusion, the infection with T. suis did result in a significant change in the proximal colon
microbiota composition, although it did not seem to change the biodiversity (Li et al., 2012). Infection
resulted in a significant change in approximately 13% of the microbial genera in the proximal colon,

6
with or without adult worms, so rather as a cause of the initial infection (Wu et al., 2012), which causes
local tissue disruption occurs in the intestinal mucosal layer, with eosinophil and mast cell infiltration,
crypt hyperplasia, goblet cell hyperplasia and mucosal hypertrophy (Mansfield et al., 1996, Kringel et
al., 2006). The infected pigs seemed to have a greater possible colonisation and appearance of
bacteria and viruses as a result of the infection with T. suis (Li et al., 2012).
In both studies, the clearest was the repression on the metabolism of starch and sucrose, fructose and
mannose, pyruvate and galactose, and the decrease in lysine biosynthesis (Wu et al, 2012, Li et al.,
2012). It is not yet clear if the worm itself altered the compounds needed for the metabolic reactions,
or that they affected the upstream metabolism of compounds formed in the small intestine (Wu et al.,
2012). One interesting finding was that the colon of the infected pigs contained oleic acid, which was
not detected in the control pigs. This oleic acid has an antibacterial activity, which then may have a
contribution to the shift in the microbiome composition and its lipolytic properties, which work mucosal
pro-inflammatory. Especially genera that consist of carbohydrate- and/or fibre-utilising bacteria, such
as Fibrobacter species and Ruminococcus, were significantly affected by infection with T. suis. The
presence of carbohydrates influence the intestinal morphology and mucine biosynthesis in the porcine
large intestine, and the metabolism of these carbohydrates is controlled by the intestinal microbiota
Thomsen et al., 2006). If this carbohydrate metabolism is reduced by infection, there is a decrease in
short-chain fatty acids that are absorbed through the epithelial barrier, which then means there is a
reduced circulation of energy for the cells and therefore a reduced local epithelial cell function. This
can then in turn affect the local microbiota composition. This problem could be solved by feeding the
pigs foods rich in carbohydrates. Helminth infections demand a high demand of essential amino acids,
which are deprived through this infection as the helminths compromise the biosynthesis capacity for
these essential amino acids. Also Ruminococcus, which is one of the most important cellulolytic
bacteria established in the gastrointestinal tract of animals, was significantly reduced. The luminal pH
was not altered during infection (Wu et al., 2012, Li et al., 2012).
Another effect which was discovered was the influence the helminth had on Campylobacter, which is
also part of the normal microbiome in pigs and normally does not cause infections, unless the pigs are
concomitant infected by T.suis. The abundance of Campylobacter in the colon of infected pigs with
worms was 3 times higher than in non-infected pigs. The helminths up-regulate IL-4, which enhances
the internalisation of intestinal pig epithelial cells by Campylobacter. However, infected but worm-free
pigs had a significant decrease in Campylobacter (table 1). This could be explained as such, that the
helminths cause for an increased uptake and antigen processing of Campylobacter, with therefore
have a higher induction of local anti-Campylobacter antibody responses (Wu et al.,2012, Mansfield et
al., 1996, Mansfield et al., 2004).
In a study by Zaiss et al. (2015), pigs that were experimentally infected with Ascaris suum eggs
showed an increase in short chain fatty acids (SCFAs) at 8 weeks post infection. This was mostly due
to an increase in propionate, butyrate and acetate concentrations in infected pigs (table 1).
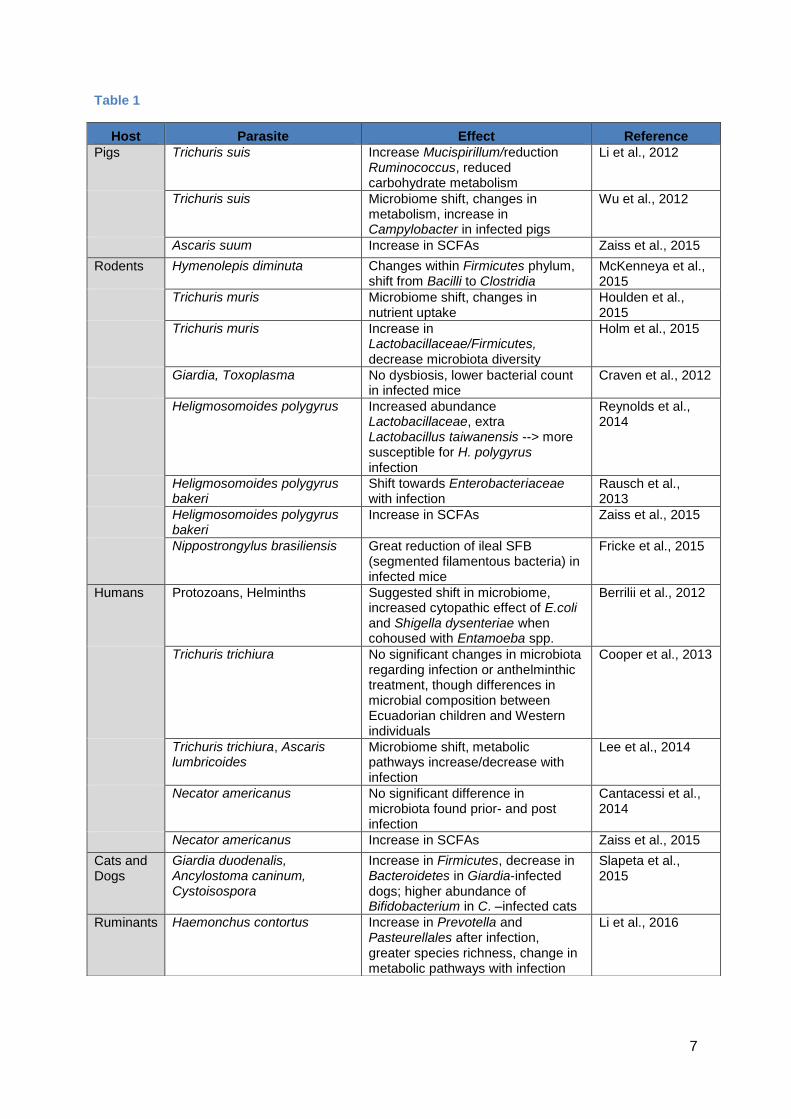
7
Table 1
Host Parasite Effect Reference
Pigs Trichuris suis Increase Mucispirillum/reduction Ruminococcus, reduced carbohydrate metabolism
Li et al., 2012
Trichuris suis Microbiome shift, changes in metabolism, increase in Campylobacter in infected pigs
Wu et al., 2012
Ascaris suum Increase in SCFAs Zaiss et al., 2015
Rodents Hymenolepis diminuta Changes within Firmicutes phylum, shift from Bacilli to Clostridia
McKenneya et al., 2015
Trichuris muris Microbiome shift, changes in nutrient uptake
Houlden et al., 2015
Trichuris muris Increase in Lactobacillaceae/Firmicutes,
decrease microbiota diversity
Holm et al., 2015
Giardia, Toxoplasma No dysbiosis, lower bacterial count
in infected mice Craven et al., 2012
Heligmosomoides polygyrus Increased abundance Lactobacillaceae, extra Lactobacillus taiwanensis --> more susceptible for H. polygyrus infection
Reynolds et al., 2014
Heligmosomoides polygyrus bakeri
Shift towards Enterobacteriaceae with infection
Rausch et al., 2013
Heligmosomoides polygyrus bakeri
Increase in SCFAs Zaiss et al., 2015
Nippostrongylus brasiliensis Great reduction of ileal SFB (segmented filamentous bacteria) in infected mice
Fricke et al., 2015
Humans Protozoans, Helminths Suggested shift in microbiome, increased cytopathic effect of E.coli and Shigella dysenteriae when cohoused with Entamoeba spp.
Berrilii et al., 2012
Trichuris trichiura No significant changes in microbiota regarding infection or anthelminthic treatment, though differences in microbial composition between Ecuadorian children and Western individuals
Cooper et al., 2013
Trichuris trichiura, Ascaris lumbricoides
Microbiome shift, metabolic pathways increase/decrease with infection
Lee et al., 2014
Necator americanus No significant difference in microbiota found prior- and post infection
Cantacessi et al., 2014
Necator americanus Increase in SCFAs Zaiss et al., 2015
Cats and Dogs
Giardia duodenalis, Ancylostoma caninum, Cystoisospora
Increase in Firmicutes, decrease in Bacteroidetes in Giardia-infected dogs; higher abundance of Bifidobacterium in C. –infected cats
Slapeta et al., 2015
Ruminants Haemonchus contortus Increase in Prevotella and Pasteurellales after infection, greater species richness, change in metabolic pathways with infection
Li et al., 2016

8
2.2. RODENTS
Different studies have been done involving rats and mice, to see whether an infection with an intestinal
helminth significantly alters the gut microbiota (see table 1).
2.2.1. Rats
In Mckenneya et al. (2015), the cecal microbiome in rats colonised with the small intestinal helminth
Hymenolepis diminuta (a cestode helminth) was studied, and to be sure that the effects of this
infection were present from the time of birth via transmittal of immune components through the milk,
they were followed for 2 generations. The second generation of rats were experimentally infected with
the helminth at the time of weaning. All the rats, with or without helminths, were divided into two
groups, one group was exposed to lipopolysaccharide (LPS, which is a bacterial membrane
component that causes and immune response) 4 days before the microbiota analysis. This exposure
was done to show the effect and interaction that the helminth had had on and with the microbiome
after a mild bacterial infection..
A significant change was found in the rats with the helminth infection, where the abundance of
Turicibacter decreased, and that of Peptostreptococcaceae increased 2,7 fold. Originally, in animals
without the helminth, Turicibacter accounted for more than 22% of the bacterial microbiota in rat’s
small intestine, this amount dropped to about 4% in rats infected by the cestode. These changes were
mostly within the Firmicutes phylum, which holds about 20% of the total bacteria, despite the fact that
the helminth lives in the small intestine and the microbiome was assessed in the cecum. The shift was
clearest from Bacilli to Clostridia when infected with the tapeworm. The infection with LPS significantly
impacted the β-diversity (beta being the diversity between locations), but did not affect the α-diversity
(alpha being the diversity of the bacterial population in a specific location i.e. the gut) in uninfected
rats. In rats holding the helminths, this was just the other way round (McKenneya et al., 2015).
2.2.2. Mice
2.2.2.1. Trichuris muris infections In Houlden et al. (2015), mice were infected with Trichuris muris eggs (murine intestinal whipworm)
and were followed over a period of 91 days.
To define if the T. muris infection had a permanent or a temporary effect on the microbiota, a group of
5 mice was treated with an antihelmintic called mebendazole from day 41 to day 43. A control group of
uninfected mice were also given mebendazole to assess the impact of the treatment on the
microbiome. The microbiome of both naïve groups turned out to be significantly different in
comparison to both infected groups. Between the two naïve groups there was not a significant change,
neither was there between infected mice and infected mebendazole-treated mice. By day 91, the
microbiota of naïve mice, naïve mebendazole-treated mice an infected mebendazole-treated mice
were most similar and were significantly different from that of infected mice. This experiment shows
that infection with T. muris in mice causes a long-term change in the microbial communities compared
to uninfected mice. Mebendazole treatment caused no clear shift in uninfected mice, but the clearance
of worms in infected mice caused the microbiome to shift back and recover nearly to its original state.

9
The most abundant species in uninfected mice in Houlden et al. (2015), were Bacteroidetes,
Firmicutes and Proteobacteria, which made up more than 99% of all sequences. As seen in the
previous study by McKenneya et al. (2015), there were significant changes in α- and β-diversity, with a
reduction in α-diversity and a decrease in β-diversity. The clearest impact of infection was the
reduction of Bacteroidetes, and specifically Prevotella and Parabacteroides in infected mice compared
to naïve mice. The Firmicutes and Proteobacteria phyla stayed roughly similar to that of naïve mice,
which demonstrates a high stability (Houlden et al., 2015).
GC-MS and LC-MS analysis were used to see the changes in the microbiota caused by changes in
metabolic profile in the large intestine as a result of the Trichuris infection. These tests were done on
faecal samples from the mice taken on day 41. There was a significant increase in essential amino
acids and a decrease in the breakdown of carbohydrates, due to loss of Bacteroidetes. Analyses also
showed a reduction of Vitamin D2/D3 derivatives, large range of fatty acids and related metabolites,
and glycerophospholipids. Interestingly, the mice were all weighed throughout the experiment to see if
the changes in microbiota and differences in nutrient metabolisms could be a cause for less weight
gain. The tests did indeed identify that the naïve mice gained more weight than infected and infected
mebendazole-treated mice. This suggests that infection causes a small but significant impact on the
microbiota, which then impacts the nutrient uptake of the mice, and that causes the mice to reduce
weight gain, as the energy uptakes aren’t sufficient. This article notes that it is yet unclear if the origins
of the hosts metabolomic changes are caused by the hosts’ microbiota, or are a result of the helminth
which directly affects the composition of the microbiota. As the infection progressed, the microbiota
gradually changed back towards that of an uninfected mouse, but the immune cell population did not,
which suggests that such an infection with dysbiosis has, as a result, also clear impact on the immune
system (Houlden et al., 2015).
Another species whose abundance increased around day 28 of infection in Houlden et al. (2015), and
only at this particular moment, was that of Mucispirillum, which colonises the mucin layer of the gut
(Robertson et al., 2005). As was seen in in the Trichuris suis infections in pigs (Li et al., 2015, Wu et
al.,2015), around this point in time of the infection, changes occur in the epithelial cell turnover, with
goblet hyperplasia and greater mucin production. As Mucispirillum thrives in this kind of habitat, that
could explain the greater abundance of Mucispirillum. However, as mucin degradation in the gut
occurs probably as a result of the Trichuris secretions (Hasnain et al., 2012), two major components of
mucin i.e. serine and threonine (Perez-Vilar et al., 1999) levels also rise (Houlden et al., 2015). These
changes in microbial composition are in contrast to a study by Craven et al. (2012), where no
dysbiosis and bacterial invasion occurred in mice with a minimal or mild ileitis caused by Giardia and
Toxoplasma infections. It was even so that the bacterial counts found were lower than that of
uninfected control mice (Craven et al., 2012).
A second experiment with Trichuris muris was done by Holm et al. (2015), where mice were co-
housed for two weeks to ensure a stable microbiota, and later placed in separate cages to avoid
cross-contamination during the infection. Then, 10 mice were infected with T. muris, and 20 mice were

10
left as the uninfected group. From those 20 uninfected mice, 10 were put down on day 0. The
remaining mice (10 with, and 10 without infection), were sampled regularly for a period of 35 days.
Their faeces were immediately frozen on dry ice after collection, which occurred at day 13, day 20 and
day 27, which each represents a larval moulting stage resp. L2, L3, L4. After day 35, luminal contents
of the caecum and colon were collected. And finally, a control infected group of 5 mice were sacrificed
and assessed at day 35 for their worm burden. The changes in the microbiota became more
noticeable as the infection progressed, and by day 35 the infected mice had a distinctly different
microbiome to the uninfected mice.
The diversity also decreased as a result of the infection with T. muris in Holm et al. (2015), with the
decrease in α-diversity suggesting that the microbiome became less diverse in each individual,
whereas the concomitant increase in β-diversity suggests that each individual responded differently to
infection and therefore there was a larger diversity between the mice. It seemed clear that in the
experiment by Holm et al. (2015), like in the previous ones (McKenneya et al., 2015; Houlden et al.,
2015), a change was found in the diversity of the hosts microbiota in caecum and colon, with again an
increased abundance of Firmicutes, and Proteobacteria, and a decrease in Bacteroidetes. As a result,
there was a clear shift towards a gram positive microbiome as the two latter are lipopolysaccharide-
containing gram negative bacteria, whereas Firmicutes are mostly gram positive bacteria. In
comparison to the other previous articles, this one noticed a gradual increase in the relative
abundance of Lactobacillaceae after day 20 of the infection from <5% between day 0 and day 20, to
11% after day 27 and even 15% after 35 days. This increase was seen in most mice, whilst the other
changes in diversity for example were more variable among individuals. Lactobacillus, Allobaculum
and Barnesiella showed a significant increase in abundance, alongside a more than 10-fold increase
in the abundance of Mucispirillum (Holm et al., 2015).
In Holm et al. (2015), they further discovered that providing Lactobacillus casei orally to mice caused
more susceptibility to Trichuris muris infections (Dea-Ayuela et al., 2008). The microbiota are of
importance for the T. muris larvae to hatch during the infection (Hayes et al., 2010). This theory
coincides with the findings that the significant changes in microbiota were observed at the moments in
which the larvae progressed from one phase to another. So it is possible that the growth of the larvae
and the physical presence of them in the gut lumen may be of great influence on the alteration of the
microbiome. It also seems that the parasite secretes effector molecules which may influence the gut
microbiota directly or indirectly through host immune cell stimulation first (Holm et al., 2015). These
secretions, among others, consist largely of serine proteases which degrade intestinal mucin (Hasnain
et al., 2012) and therefore the helminth partially prevents the intestinal bacteria from attaching
themselves to the mucus layer from which they thrive, in order to survive in this type of niche
themselves (Van den Abbeele et al., 2011).
2.2.2.2 Heligmosomoides polygyrus bakeri infections
Reynolds et al. (2014) showed that it is the composition of the intestinal microbiota that is important,
and not the amount of bacteria present in the gut, which impacts the survival of Heligmosomoides

11
polygyrus. Especially the Lactobacillaceae, which increase in the duodenum when infected with the
helminth and therefore have a positive correlation with it, can ensure the hosts more susceptible
reaction to the helminth infection. This was shown by using germ-free mice (GF), which did not have
commensal bacteria in their gut, and they were far more vulnerable to infections with H. polygyrus than
normal mice. They examined specific pathogen-free (SPF) BALB/c mice, to find out if the composition
of the microbiota had a direct influence on the survival of the helminth. First, they treated the mice with
vancomycin, a low level of antibiotic prior to infection, but this did not result in a significant reduce in
fecal bacteria numbers, but did result in an increased H. polygyrus persistence. There was an increase
is Lactobacillaceae and Enterobacteriaceae, and a reduction of Eubacterium/Clostridium species.
Lactobacillus taiwanensis in the murine duodenum at 28 days of infection with H. polygyrus was
positively correlated with the total Lactobacillus/Lactococcus abundance and with the number of
worms inside the host. To test if this had a causal basis, the mice were given untreated drinking water
or water containing 2 X 108
colony forming units (cfu)/ml of L. Taiwanensis for one week before
infecting them with H. polygyrus. The egg-output of H. polygyrus was measured each week. On day
14, both bacteria-fed and control mice had similar amounts of eggs, but on day 21, the egg-output in
mice fed L. taiwanensis was significantly higher than that of control mice (Reynolds et al., 2014).
In Zaiss et al. (2015), an assessment was made of the anti-inflammatory activity induced in the gut
when infected with intestinal helminths, and whether this was directly regulated by the helminths
themselves, or if it involved crosstalk with the local microbiota. We know that a chronic infection with
the small intestinal helminth H. polygyrus bakeri in mice alters the composition of the gut microbiome
(Reynolds et al., 2014). This alteration allows an increase in short chain fatty acid (SCFA) production,
which is a product from bacterial fermentation of complex oligosaccharides present in the diet. The
influence of these increased SCFA on allergic asthma was shown by using infected mice which lacked
the SCFA cognate receptor GPR41, and these were far more susceptible to allergic airway disease.
This shows that the microbiota modulates the immune system directly, but also increases bacterial-
derived immune-modulatory metabolites in case of infection.
To test if this immune modulation was directly linked to the microbiota, mice were treated with
antibiotics, which eliminated 105
-106 aerobic bacteria, and 10
6 -10
7 anaerobic bacteria, and no fungal
growth was detected. These mice were then challenged with house dust mite (HDM) and the
inflammatory response was monitored. The antibiotic-treated mice did not attenuate the inflammation
at all, whilst in specific-pathogen free (SPF) mice there was a reduction of eosinophil infiltration, type 2
cell-associated cytokine concentration in the airways and HDM-specific IgG1 when infected with the
helminth. Then, the mice treated with antibiotic received the microbiota from H. p. bakeri infected mice
by co-housing them with naïve SPF mice, or H. p. bakeri infected mice for 3 weeks. This way transfer
of microbiota was possible, but no transfer of worms occurred. After the co-housing, the mice were
then again exposed to HDM. Recipient mice co-housed with helminth infected donor mice, showed a
reduced amount and proportion of eosinophils, compared to recipient mice co-housed with naïve
donors. This altered microbiome alone was sufficient enough to help protect against allergic asthma.

12
The link has been proven by epidemiological studies, showing that there is a negative correlation
between helminth infections and allergen skin test reactivity (Cooper et al., 2003).
The next experiment involved gnotobiotic mice colonised with a limited bacterial flora, called the
altered Schaedler flora (ASF), which consisted mostly of Bacteroidales and Lactobacillales orders
(Cahenzli et al., 2013), whereas helminth infected mice had an outgrowth of Clostridiales bacteria,
resulting from the parasite infection itself. Also SCFAs were significantly increased in the cecum of H.
p. bakeri infected mice compared to naïve ASF mice. To see if the increased SCFA production is
directly correlated with the alteration in microbiota during infection with helminths, they analysed the
microbiota composition and SCFA concentrations in the ceca of the mice that had been co-housed
with the naïve or helminth infected donors. The mice co-housed with the helminth infected donors had
an increase in SCFAs (Zaiss et al., 2015).
It is also possible that the parasite itself can alter the microbiota, alongside the host’s immune system.
This was tested by Rausch et al. (2013), where infection with the hookworm Heligmosomoides
polygyrus was responded to with a Th2 reaction by the mice, which altered the microbiome
composition. A second experiment with IL-4Rα-/-
mice, who are deficient in their capacity to generate a
Th2 response, showed the same intestinal microbiome changes as the first experiment, suggesting
that the hookworm can alter the composition independently from the host’s immune response (Rausch
et al., 2013).
Normally, infections with H. p. bakeri do not lead to signs of morbidity or weight loss. In this study by
Rausch et al. (2013), the amount of inflammation and changes in gut microbiota were surveyed in
different places i.e. the proximal small intestine (at the site of the helminth infection), and in the large
intestine, which is worm-free. These samples were taken on day 6 (larval development), day 14 (acute
adult phase), and 28 days post infection (chronic adult phase). At 6 and 14 days infected mice had a
mild, but significant elevated inflammatory score of the duodenum, but not of the large intestine. After
this, they assessed the luminal contents on day 14, and observed a shift towards Enterobacteriaceae
in infected mice. Especially in the small intestine, there was a significant increase in abundance of the
gram positive Lactobacilli and Clostridium species in infected mice, and in the cecal contents it was
the gram negative γ-Proteobacteria/Enterobacteriaceae and Bacteroides goup that increased in
abundance in infected mice.
It is not yet clear if it is the inflammation itself after infection with H. p. bakeri that facilitates the growth
of the gram negative Enterobacteriaceae, or that the tolerance for the new infectious circumstances by
other commensals decreases, thus making it possible for Enterobacteriaceae species to outgrow
(Rausch et al., 2013).
Both articles by Reynolds et al. (2014) and Rausch et al. (2013) can be summarised into two findings:
first of all, infection with H. polygyrus changes the composition of the small intestinal microbiota, and
second, the abundance of species in the normal microbiota can determine and alter the survival of H.
polygyrus in its host.

13
2.2.2.3. Nippostrongylus brasiliensis infections
An article by Fricke et al. (2015) looked at the possibility of a crosstalk influence between the
nematode Nippostrongylus brasiliensis and the gut microbiota of mice. This nematode lives in the
proximal small intestine after migration through the skin and lung. It concentrates on the segmented
filamentous bacteria (SFB), which is a specific Th17-provoking commensal bacterium.
First of all, on day 11 post-inoculation, ileal samples were taken from the mice, but no significant
change in α-diversity was detected. However, changes were found in the abundance of the 3
dominant bacterial phyla, with a significant increase in Bacteroidetes and Actinobacteria and a
significant reduction in Firmicutes in infected mice compared to uninfected mice. Within the Firmicutes
phylum, a significant increase was noted in abundance of Lactobacillaceae, concurrent with a
decrease in abundance of Peptostreptococcaceae, Clostridiaceae, and Turicibacteraceae. Within the
Actinobacteria it was the Coriobacteriaceae that increased in infected mice.
Then the abundance of the SFB was examined, finding a >2000 fold reduction in the ileum of infected
mice, whilst at the same time there was a reduction of SFB found in the jejunum, cecum and proximal
colon. As SFB are prominently found in the ileal wall (Koopman et al., 1987), their reduction during
helminth infection could be due to the increased physical shredding of the mucus, or result of an
increased anti-microbial peptide (AMP) expression (Vaishnava et al., 2011).
The study showed that the host’s type 2 response to infection with N. brasiliensis inhibited the
intestinal SFB and decreased the expression of IL-17 genes. This effect is possibly the result of
modulation of host-derived AMP and mucin expression of the local microbiota during infection with an
intestinal helminth (Fricke et al., 2015).

14
2.3 HUMANS
2.3.1 Protozoans
Humans can be infected with different types of intestinal protozoans, such as the intracellular
Cryptosporidium spp., the host-specialised Entamoeba hystolytica, and the non-host-specific Giardia
duodenalis. Most of them do not cause any real harm, but may occasionally cause diarrhoea as a
result of bowel-wall damage (Berrilli et al., 2012).
Different hypotheses have been brought forward to explain the interactions between these protozoans
and the gut microbiota. One case saw a change and possible decrease in adhesive abilities in
protozoans, caused by a change in the surface saccharide ligands of superficial membranes which
altered the presence of the intracellular microflora. This change in microflora resulted in a decrease in
invasive abilities (Phillips, 1973; Dwyer and Chang, 17976). In a more recent study from Galván-
Moroyoqui et al. (2008), it was shown that in vitro phagocytosis of enteropathogenic species such as
Escherichia coli and Shigella dysenteriae, which are part of the normal human biome, and which were
co-cultured with Entamoeba hystolytica and Entamoeba dispar, increased the cytopathic effect of E.
hystolytica by increasing the expression of Gal/GalNAc lectin on the amoebic surface and the cysteine
activity. E. dispar remained avirulent though (Berrilli et al., 2012).
2.3.2 Helminths
In humans, the intestinal habitat is ideal for several types of worms. These include cestodes like
Diphyllobothrium, Taenia and Hymenolepis; digeneans such as Fasciolopsis, Heterophyes, and
Schistosoma; and finally nematodes such as Ascaris, Trichuris, Ancylostomatidae, Strongyloides and
Enterobius vermicularis (Berrilli et al., 2012). Following the previous studies done in rodents and pigs
(table 1), it has clearly been suggested that infections with helminths can cause a significant change in
the abundance, as well as in the composition of the gut microbiota.
In humans, most developed countries are struggling with lots of different autoimmune diseases (such
as Crohn’s disease, inflammatory bowel disease), whilst developing countries with a much higher
prevalence in intestinal helminth infections do not seem to have those illnesses (Sewell et al., 2002).
This problem is based on a phenomenon called the “Hygiene hypothesis” (Wills-Karp et al., 2001;
Weinstock and Elliot, 2009). A study using mainly Trichuris sp. infections, suggested that there was a
probable interaction between the microbiota and the host immune system, but that the intestinal
helminths also had an immune-regulating effect on the host. The helminths also produced molecules
that altered the microbial habitat (Bancroft et al., 2012). Balog et al. (2011), demonstrated on the base
of urinary response of human and rodent hosts to Schistosoma mansoni infection, that there was a
disturbance in metabolites associated with the intestinal microbiota and microbial co-metabolism.
Two other helminths i.e. Ascaris lumbricoides, which lives in the small intestine, and Trichuris trichiura,
where the adults thrive in the caecum, are the most common soil-transmitted helminths (STH) in
humans (Bethony et al., 2006). They have been proven to dampen inflammatory host response, and
therefore keep their own infection going (Cooper et al., 2013).

15
An article by Cooper et al. (2013) looked at infections with Trichuris trichiura in humans, and
hypothesised that the affect it had on the microbiota could be ‘undone’ by using an anthelminthic
treatment. They also evaluated the effects of such a treatment on the composition of the gut
microbiota. They then evaluated the effects of mixed infections of Trichuris trichiura and Ascaris
lumbricoides on the gut microbiota. For this experiment they evaluated 914 children from 3 different
villages in Ecuador, where a previous study had shown that there was a high prevalence of STH
infections in these areas (Moncayo et al., 2008). Two to three stool samples were taken from all
children over the period of a month, and these samples were tested for the presence of STH eggs and
larvae. The children were then divided into 3 groups. The first were the uninfected controls (n=30),
with no STH parasites detected in any of the stool samples; the second were infected with T. trichiura
only (n=17), so these showed the specific effects of T. trichiura on the microbiota; and the third had
mixed infections of A. lumbricoides and T. trichiura in all stool samples (n=50), so the effects could be
evaluated of mixed STH infections on the microbiome. For further evaluation, 121 of these children
belonging to one of the groups and aged 8 to 14 years were selected. None of these children had
taken antibiotics in the previous month, nor had they received anthelmintics in the previous 3 months;
they were also asymptomatic and had no fever at the time of sampling. All 17 children from the T.
trichiura group, and 21/30 children from the uninfected group were then treated with albendazole 400
mg twice a day for 3 days and received a single dose of 200 µg/kg ivermectine, to get rid of all STH
infections, which was also proved in the results. This was done to evaluate the effects of anthelmintic
treatment on the microbiota. New stool samples were taken at day 7 and 21 to determine the short-
term effects. There was no significant difference in microbiota between the group of children only
infected with T. trichiura, those with mixed STH infection, and those free from infection. Neither was
there a significant difference found after treatment with anthelmintica on the microbial composition. It
also showed that children with mixed STH infections showed a greater abundance of Clostridium
sensu stricto genus compared to other groups and a significantly lower overall variety in microbial
composition, and therefore linking this finding to Ascaris lumbricoides (Cooper et al., 2013).
This article shows that there were no significant changes found in the faecal microbiota composition of
all the different groups, with or without infection, and with or without treatment, in contrast to previous
studies shown in table 1. An important thing to realise when interpreting these results is that the faecal
microbiota which was collected in this study is not necessarily the same composition as the colon
microbiota (Zoetendal et al., 2002; Eckburg et al., 2005), where the infection of Trichuris spp. takes
place. As seen before in Wu et al. (2012) in table 1, that even with anthelminthic treatment and post-
infection, that the microbial composition remained largely unchanged, which could explain the
similarity in microbial composition between the uninfected and infected groups of children and that the
microbiome could have been permanently altered as such (Cooper et al., 2013).
A study by Cantacessi et al. (2014) had similar findings to Cooper et al. (2013), where faecal samples
of 8 healthy individuals were collected, after infecting them percutaneously with 20 infective L3 larvae
of the Necator americanus hookworm (Daveson et al., 2011). These were taken prior to infection (T0)
and 8 weeks post-infection (T8). The most abundant phyla found in all individuals were again the

16
Bacteroidetes, Firmicutes, and the Proteobacteria at both T0 and T8, indicating that the hookworm
infection did not alter the bacterial community composition (Cantacessi et al., 2014). The hookworm N.
americanus did, however, seem to increase the total SCFAs in infected individuals in Zaiss et al.
(2015), by increasing the local acetate, butyrate and propionate production in the gut.
In Western/industrialised countries, the most abundant bacteria come from within the Firmicutes
phylum i.e. Lachnospiraceae and Ruminococcaceae. In these Ecuadorian children in the study by
Cooper et al. (2013), it turned out that the Firmicutes species Faecalibacterium prausnitzii was the
most abundant from all microbiota, which in contrast however, has also been identified as the most
abundant in Western/industrialised individuals in a study by Walker et al. (2011). This species has also
recently been taken under the loop as a potentially anti-inflammatory species (Sokol et al., 2008). In
contrast to these similarities, some differences were also noted in the composition of microbiota from
these Ecuadorian children compared to that of Western/industrialised individuals, such as the fourth
most abundant bacteria was a member of the Succinivibrio genus of the γ-Proteobacteria, which has
not been reported in faecal samples of Western individuals, but is a similar organism to the ones
typically found in rumens (Bryant and Small, 1956). Another finding was a bacteria related to Sarcina
ventriculi, which has been found to be common in developing countries by Crowther (1971) and was
detected in 59,8% of the stool samples of the Ecuadorian children, but is rarely recovered in Western
individuals. A high abundance of Treponema spp. was also found in the Ecuadorian cohort study. In
the Bacteroidetes phylum, the highest abundance was that of the Prevotella, and only a very small
percentage Bacteroides (Cooper et al., 2013), with the latter being typically one of the most abundant
genera in Western individuals (Costello et al., 2009; Human Microbiome Project Consortium, 2012). It
has been suggested by Wu et al. (2011), that the increase of Prevotella spp. is linked to diets high in
carbohydrates and simple sugars, whilst decrease in Bacteroides spp. is linked to diets high in
saturated fats and animal proteins (Cooper et al., 2013).
A third study by Lee et al. (2014) focusses on the microbiota composition within Malaysian indigenous
populations (n=51, ages 0.4-48 years old), where the prevalence of soil transmitted helminths (STH) is
still quite high, and compared them with non-colonised individuals. These stool samples were
collected in a single field trip, and the majority (70%) of these samples contained helminths. Trichuris
spp. was the most common (54.9%), then Ascaris spp. (41.2%) and the hookworm (9.8%). One third
of the individuals were colonised by a combination of these helminths (Lee et al., 2014). Finally, the
Malaysian samples in Lee et al. (2014) were also then compared to faecal samples taken from 19
healthy adult men from New York, to see the differences in microbiota between developing and
developed countries, just as they did in Cooper et al. (2013). This analyses showed that there was a
rather great and significant difference between the microbiota of developing and developed countries
(agreeing with Cooper et al., 2013), which was far greater difference than between infected and non-
infected individuals in Malaysia. The α-diversity in Malaysian individuals was far richer than that in the
New York volunteers. Hence, the Firmicutes phylum was more abundant in New York individuals, but
the Cyanobacteria, Actinobacteria, Tenericutes, and Proteobacteria were more abundant in the
Malaysian individuals. The same differences in metabolic pathways were observed and associated

17
with helminth colonisation, as described above in the Malaysian people compared to the New York
subjects. These findings follow the hypothesis that socioeconomic development is associated with the
disappearance of ancestral microbiota (Wittebolle et al., 2009; Arnold et al., 2011).
The most abundant phyla, found in the study by Lee et al. (2014), were also the Firmicutes (55.9%),
Bacteroidetes (23.5%), and Proteobacteria (10.1%). This distribution was found in most individuals,
except for 2 toddlers of 3 months and 5 years old, where there was an exceptionally high abundance
of Actinobacteria (resp. 35.2% and 55.5%), and especially from the Bifidobacterium genus, which has
been observed to be an enrichment in the infant gut (De Filippo et al., 2010; Yatsunenko et al., 2012).
It was also demonstrated that the helminth-positive individuals possessed a greater richness in
species, compared to non-infected individuals. Analysis revealed that Paraprevotellaceae (strongly
associated with Trichuris-only infection), Mollicutes, Bacteroidetes, and Alphaproteobacteria were of
increased abundance in infected individuals, compared to an increased abundance in Bifidobacterium
only in helminth-negative individuals (Lee et al., 2014).
In helminth-infected individuals in Lee et al. (2014), there were metabolic functional pathways which
were more abundant that were used for genetic information processing, especially the pathways for
translation, replication and repair. Nucleotide metabolism pathways, such as those for cell growth and
death, were also significantly greater in the microbiota. These effects were seen in individuals with
STH-mixed infections and in individuals with Trichuris-only infections, indicating that the effect is
largely due to the presence of Trichuris worms. Other pathways such as those for carbohydrate and
xenobiotic metabolism were increased in the microbiome of helminth-negative individuals, whereas
positive individuals had an increased metabolism for i.e. nucleotides, amino-acids, terpenoids and
polyketides, novobiocin biosynthesis and for vitamins. It seems to be the case that it is the presence of
the Ascaris worms that cause the decrease in carbohydrate metabolism (Lee et al., 2014).

18
2.4 DOGS AND CATS
A study by Slapeta et al. (2015) compared the composition and diversity of intestinal microbiota in 40
dogs and 22 cats from Australia. These animals were naturally infected with Giardia duodenalis,
Ancylostoma caninum, and Cystoisospora, and were non-diarrhoeic. Seventeen dogs and 8 cats were
infected with G. duodenalis (a protozoan parasite), 13 dogs with the dog hookworm or A. caninum,
and another 8 cats were positive for Cystoisospora (a protozoan coccidia). Some of these animals
had been de-wormed as they came from a kennel, others were stray dogs and had had no previous
medical care or de-worming treatment.
Seventy four faecal samples were collected in total from the cats and dogs. A significant difference
was observed in microbial composition of Giardia-positive and -negative dogs, whilst no significant
difference was found between Ancylostoma-positive and -negative dogs. The influence of the
hookworm did not show any changes in faecal microbiota compared to uninfected animals, as
previously described in a study by Cantacessi et al. (2014).
The Giardia-positive group had an increased abundance in Catenibacterium, Howardella (both from
the Firmicutes phylum), and Pseudomonas (Proteobacteria). The presence of more Catenibacterium
could be stimulated by Giardia infections, which then results in a higher production of short-chain fatty
acids (SCFA), which is associated with protection against pathogens (Kageyama and Benno, 2000).
This was in contrast to the Giardia-negative group, which had a higher abundance of Bacteroides
(Bacteroidetes) and Pseudobutyrivibrio (Lachnospiraceae).
In the cat samples, the Firmicutes were increased in Giardia-negative cats, and a significant difference
was also found between Cystoisospora-positive and -negative cats. The Cystisospora infected cats
had a higher abundance of Bifidobacterium, belonging to the Actinobacteria, but a significant increase
in Firmicutes and Proteobacteria was also seen. Bifidobacterium infantis is a known probiotic in rats
and humans, which helps to decrease necrotising enterocolitis (Bergmann et al., 2013). This could
explain the higher abundance in cat microbiota after such an infection with coccidiosis.
A problem is, just like in Cooper et al. (2013) (table 1), that the faecal microbiota is not necessarily
representative for the intestinal microbiota where the parasitic infection takes place, thus sampling the
intestinal content may have given another, more detailed, outcome.

19
2.5 RUMINANTS
Haemonchus contortus is one of the most important abomasal helminth infections in small ruminants,
causing severe decreases in production in sheep and goats, even leading to mortality. It is known that
infections with parasites such as H. contortus, Teladorsagia circumcincta and Ostertagia ostertagi
quickly lead to a change in ruminants’ abomasal secretory activity, and therefore increasing the pH
(Simpson et al., 1997; Simcock et al., 1999; Li et al., 2011).
In a study by Li et al., 14 parasite-free goats were infected with 5.000 H. contortus infective larvae and
followed for 50 days, six goats were used as controls.
After infection, the goats ate a lot less, and therefore lost body weight during this experiment. The
abomasal pH increased from a normal 2.93 in the control goats to 4.50 in infected goats. This pH is
the ideal environment for the parasite egg production, and thus survival and proliferation of the
helminths is realised.
The normal abundance, in decreasing order, of bacterial phyla in the caprine abomasum were found
to be Protebacteria (37.3%), Bacteroidetes (35.5%), Firmicutes (24.5%), and Spirochaetes (0.8%).
The most common families were the Succinivibrionaceae (36.9%), Prevotellaceae (22.8%),
Lachnospiraceae (10.3%) and Ruminococcaceae (4.8%). Infection increased the abundance of
Prevotella as well as its family Prevotellaceae. This change affects the energy metabolism in the
caprine gut microbiome. The Gram-negative Pasteurellales were absent in control goats, but
significantly increased in H. contortus-infected goats, and thus compensating for the hosts’ declined
protein metabolism (table 1).
The infection did not seem to significantly impact the microbiota diversity, but it did seem to give a
greater species richness to the infected goats compared to uninfected controls, altering the
abundance of about 19% of all species. It is possible that an infection with H. contortus may have an
effect on several biological functions, especially on ABC transporters, carbohydrate metabolism, and
amino acid metabolism. Infected animals had higher potential of modifying their xenobiotics
metabolism, as the biodegradation of naphthalene, a polycyclic aromatic hydrocarbon, was higher in
infected goats. The study hypothesised that the dampening of inflammation during infection is partially
a result of modulation in the gut butyrate biosynthesis, butyrate being anti-inflammatory (Li and Li,
2008), which is caused by a change in abundance of the butyrate-producing bacteria. This change
was observed as the Butyrivibrio, a butyrate-producing bacteria, was higher in control goats than in
infected goats.

20
3. DISCUSSION
It has long been suggested that parasite infections have a direct influence on the host’s immune
system through modulating the immune response in the gut. The studies summarised in this thesis
(table 1) suggest that there could be a three way interaction between the parasites, the microbiome
and the hosts’ immune system (figure 1).
Figure 1
It is possible that the parasites influence the microflora in such a way that shifts in their phyla due to
the presence of the parasites cause the immune system to react differently to normal. Parasites may
have positive influences on the host’s immune system, educating it and providing signals that serve to
regulate inflammation and strengthen the regulatory immune response. They also dampen
inflammation responses. A branch of this subject is being studied in humans, as many developed
countries struggle with problems such as allergies, Inflammatory Bowel Disease (IBD) and Crohn’s
disease (CD). This problem is not present in developing countries, where infections with intestinal
helminths are endemic (Bancroft et al., 2012). Patients with such a disease have a different
composition of gut microflora compared to non-diseased people (Sokol et al., 2008). Infecting patients
with IBD or CD for instance with a helminth such as Trichuris trichiura, may have a positive therapeutic
effect. This possibility with its effects still needs to be examined in further depth, as it is not something
that the average patient would accept as being ‘normal’.
In the study by Li et al. (2012) for example, 3 infected pigs were selected 53 days post inoculation that
had >300 adult worms, five infected pigs with no worms, and then a further three more uninfected pigs
to compare the local changes in gene expression in the epithelial layer of the proximal colon and the
associated changes in the microbiome. High numbers of adult worms induced an increase in the
expression of several genes compared to the worm-free control pigs. Other gene-expressions were
also found to be higher in infected pigs with worms than in worm-free infected pigs (Li et al., 2012).
This suggests that the parasite has an influence not only on the metabolic pathways controlled by the
microbiota, but also seems to alter the hosts’ gene expression, which coincides with the hypothesis. It
might be of interest to use immunodeficient animal models to distinguish between direct alteration of
metabolic niche space by the helminth and alteration of niche space via alteration of the host immune
system (McKennya et al., 2015)
The interaction between host and microbiota is of great importance too. The host’s immune system
has to recognise the difference between its commensal microbiota and potentially pathogenic bacteria,
especially when these bacteria can be closely related and compete over the same niche or nutrients
(Kamada et al., 2013). Pathogens can decrease the tolerance to commensals (Hand et al., 2012), or

21
they are able to dysregulate the steady-state microbiome which can result in pathogenic infections
(Littman and Pamer, 2011; Raetz et al., 2013).
More experiments have to be done to fully understand the interactions and create a new insight on
treating inflammatory and auto-immune diseases in humans and other animals.
This field of study is still relatively new, and therefore more studies have yet to be performed, with
more subjects, to be able to definitively significantly conclude that there is, or is not, a three-way
interaction and its importance.

22
4. REFERENCES Arnold I.C., Dehzad N., Reuter S., Martin H., Becher B., et al. (2011) Helicobacter pylori infection
prevents allergic asthma in mouse models through the induction of regulatory T cells. Journal of
Clinical Investigation 121: 3088–3093.
Balog C. I., Meissner A., Goraler S., Bladergroen M.R., Vennervald B. J., Mayboroda O. A., et al.
(2011). Metabonomic investigation of human Schistosoma mansoni infection. Molecular BioSystems 7:
1473–1480.
Bancroft, A. J., Hayes, K. S., and Grencis, R. K. (2012). Life on the edge: the balance between
macrofauna, microflora and host immunity. Trends in Parasitology 28: 93–98.
Bergmann K.R., Liu S.X.L., Tian R.L., Kushnir A., Turner J.R., Li H.L., Chou P.M., Weber C.R., De
Plaen I.G. (2013) Bifidobacteria stabilize claudins at tight junctions and prevent intestinal barrier
dysfunction in mouse necrotizing enterocolitis. American Journal of Pathology 182: 1595–1606.
Berrilli F., Di Cave D., Cavallero S., D'Amelio S. (2012) Interactions between parasites and microbial
communities in the human gut. Frontiers in Cellular and Infection Microbiology 2(141): 68-73.
Bethony J., Brooker S., Albonico M., Geiger S.M., Loukas A., et al. (2006) Soil-transmitted helminth
infections: ascariasis, trichuriasis, and hookworm. Lancet 367: 1521–1532.
Bryant M.P., Small N. (1956) Characteristics of two new genera of anaerobic curved rods isolated from
the rumen of cattle. Journal of Bacteriology 72: 22–26.
Cahenzli J., Köller Y., Wyss M., Geuking M.B., and McCoy K.D. (2013). Intestinal microbial diversity
during early-life colonization shapes long-term IgE levels. Cell Host Microbe 14: 559–570.
Cantacessi C., Giacomin P., Croese J., Zakrzewski M., Sotillo J., McCann L., Nolan M.J., Mitreva M.,
Krause L., Loukas A. (2014) Impact of experimental hookworm infection on the human gut microbiota.
Journal of Infectious Diseases 210: 1431–1434.
Cooper P., Walker A.W., Reyes J., Chico M., Salter S.J., Vaca M. et al. (2013) Patent human
infections with the whipworm, Trichuris trichiura, are not associated with alterations in the faecal
microbiota. PloS one 8(10): e76573.
Cooper P.J., Chico M.E., Rodrigues L.C., Ordonez M., Strachan D., Griffin G.E., and Nutman T.B.
(2003) Reduced risk of atopy among school-age children infected with geohelminth parasites in a rural
area of the tropics. Journal of Allergy and Clinical Immunology 111: 995–1000.
Costello E.K., Lauber C.L., Hamady M., Fierer N., Gordon J.I. et al. (2009) Bacterial community
variation in human body habitats across space and time. Science 326: 1694–1697.
Craven M., Egan C.E., Dowd S.E., McDonough S.P., Dogan B., Denkers E.Y., Bowman D., Scherl
E.J., Simpson K.W., (2012) Inflammation Drives Dysbiosis and Bacterial Invasion in Murine Models of
Ileal Crohn’s Disease. PLoS ONE 7(7): e41594.
Crowther J.S. (1971) Sarcina ventriculi in human faeces. Journal of Medical Microbiology 4: 343–350.
Daveson A.J., Jones D.M., Gaze S. et al. (2011) Effect of hookworm infection on wheat challenge in
celiac disease – a randomised double-blinded placebo controlled trial. PLoS One 6:e17366.
De Filippo C., Cavalieri D., Di Paola M., Ramazzotti M., Poullet J.B., et al. (2010) Impact of diet in
shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa.
Proceedings of the National Academy of Sciences U.S.A. 107: 14691–14696.

23
Dea-Ayuela M.A., Rama-Iniguez S., Bolas-Fernandez F. (2008) Enhanced susceptibility to Trichuris
muris infection of B10Br mice treated with the probiotic Lactobacillus casei. International
Immunopharmacology 8(1):28–35.
Dwyer, D. M., and Chang, K. P. (1976). Surface membrane carbohydrate alterations of a flagellated
protozoan mediated by bacterial endosymbiotes. Proceedings of the National Academy of Sciences
U.S.A. 73: 852–856.
Eckburg P.B., Bik E.M., Bernstein C.N., Purdom E., Dethlefsen L., et al. (2005) Diversity of the human
intestinal microbial flora. Science 308: 1635–1638.
Fricke F.W., Song Y., Wang A-J., Smith A. , Grinchuk V., Pei C. , Ma B., Lu N., Urban J.F.Jr., Shea-
Donohue T., Zhao A. (2015) Type 2 immunity-dependent reduction of segmented filamentous bacteria
in mice infected with the helminthic parasite Nippostrongylus brasiliensis. Microbiome 3:40.
Galván-Moroyoqui, J. M., Del Carmen Domínguez-Robles, M., Franco, E., and Meza, I. (2008). The
interplay between Entamoeba and enteropathogenic bacteria modulates epithelial cell damage. PLoS
Neglected Tropical Diseases 2: e266.
Gandolfo F. (2015) Intro to the gut microbiome and fecal microbiota transplantation. Internet reference:
www.retroflexions.com/core-concepts/Intro-to-the-gut-microbiome-and-fecal-microbiota-
transplantation/ (consulted on 9-03-206)
Hand T.W., Dos Santos L.M., Bouladoux N., Molloy M.J., Pagán A.J., Pepper M., Maynard C.L., Elson
C.O., Belkaid Y. (2012) Acute gastrointestinal infection induces long-lived microbiota-specific T cell
responses. Science 337:1553-6
Hasnain S.Z., McGuckin M.A., Grencis R.K., Thornton D.J. (2012) Serine protease(s) secreted by the
nematode Trichuris muris degrade the mucus barrier. PLoS Neglected Tropical Diseases 6(10):e1856.
Hayes K.S., Bancroft A.J., Goldrick M., Portsmouth C., Roberts I.S., Grencis R.K. (2010) Exploitation
of the intestinal microflora by the parasitic nematode Trichuris muris. Science 328(5984):1391–4.
Hill D.E., Gamble H.R, Rhoads M.L, Fetterer R.H., Urban J.F. Jr. (1993) Trichuris suis: a zinc
metalloprotease from culture fluids of adult parasites. Experimental Parasitology 77:170-178.
Holm J.B., Sorobetea D., Kiilerich P., Ramayo-Caldas Y., Estellé J., Ma T., Madsen L., Kristiansen K.,
Svensson-Frej M. (2015) Chronic Trichuris muris Infection Decreases Diversity of the Intestinal
Microbiota and Concomitantly Increases the Abundance of Lactobacilli. PLoS One 10(5): e0125495.
Houlden A., Hayes K.S., Bancroft A.J., Worthington J.J., Wang P., Grencis R.K., Roberts I.S. (2015)
Chronic Trichuris muris Infection in C57BL/6 Mice Causes Significant Changes in Host Microbiota and
Metabolome: Effects Reversed by Pathogen Clearance. PloS One 10(5): e0125945.
Human Microbiome Project Consortium (2012) Structure, function and diversity of the healthy human
microbiome. Nature 486: 207–214.
Kageyama A., Benno Y. (2000) Catenibacterium mitsuokai gen. nov., sp, nov., a Gram-positive
anaerobic bacterium isolated from human faeces. International Journal of Systematic and Evolutionary
Microbiology 50: 1595–1599.
Kamada N., Chen G.Y., Inohara N., Núñez G. (2013) Control of pathogens and pathobionts by the gut
microbiota. Natural Immunology 14:685-90
Kinross J.M., Darzi A.W., Nicholson J.K. (2011). Gut microbiome-host interactions in health
and disease. Genome Medicine 3(3):14.

24
Koopman J.P., Stadhouders A.M., Kennis H.M., De Boer H. (1987)The attachment of filamentous
segmented micro-organisms to the distal ileum wall of the mouse: a scanning and transmission
electron microscopy study. Lab Animal Magazine 21(1):48–52.
Kringel H., Iburg T., Dawson H., Aasted B., Roepstorff A. (2006) A time course study of immunological
responses in Trichuris suis infected pigs demonstrates induction of a local type 2 response associated
with worm burden. International Journal of Parasitology 36: 915-924.
Lee S.C., Tang M.S., Lim Y.A., Choy S.H., Kurtz Z.D., Cox L.M. et al. (2014) Helminth colonization is
associated with increased diversity of the gut microbiota. PLoS Neglected Tropical Diseases 8(5):
e2880.
Li C.J. and Li R.W. (2008) Butyrate induced cell cycle arrest in bovine cells through targeting gene
expression relevant to DNA replication apparatus. Gene Regulations and Systems Biology 2: 113–23.
Li R.W., Li W., Sun J., Yu P., Baldwin R.L., Urban J.F. (2016) The effect of helminth infection on the
microbial composition and structure of the caprine abomasal microbiome. Science Reports 6: 20606.
Li R.W., Wu S., Li W., Huang Y., Gasbarre L.C. (2011) Metagenome plasticity of the bovine abomasal
microbiota in immune animals in response to Ostertagia ostertagi infection. PLoS One 6: e24417.
Li R.W., Wu S., Li W., Navarro K., Couch R.D., Hill D., Urban J.F. Jr. (2012) Alterations in the porcine
colon microbiota induced by the gastrointestinal nematode Trichuris suis. Infection and Immunity. 80:
2150–2157.
Littman D.R., Pamer E.G. (2011) Role of the commensal microbiota in normal and pathogenic host
immune responses. Cell Host Microbe; 10:311-23
Mansfield L.S., Gauthier D.T. (2004) Lymphoglandular complexes are important colonic site for
immunoglobulin A induction against Campylobacter jejuni in a swine disease model. Comparative
medicine 54: 514-523.
Mansfield L.S., Urban J.F. Jr. (1996) The pathogenesis of necrotic proliferative colitis in swine is linked
to whipworm induced suppression of mucosal immunity to resident bacteria. Veterinary immunology
and immunopathology 50: 1-17.
McKenneya E.A., Williamson L., Yodera A.D., Rawls J.F., Bilbob & William Parker S.D. (2015)
Alteration of the rat cecal microbiome during colonization with the helminth Hymenolepis diminuta. Gut
Microbes 6(3): 182-193.
Moncayo A.L., Vaca M., Erazo S., Oviedo G., Rodriguez A., et al. (2008) Impact of long-term
treatment with ivermectin on geohelminth infections in Esmeraldas Province, Ecuador. PLoS
Neglected Tropical Diseases 2: e293.
Perez-Vilat J., Hill R.L. (1999) The Structure and Assembly of Secreted Mucins. Journal of Biology and
Chemistry 274: 31751-31754.
Phillips B. P. (1973). Entamoeba histolytica: concurrent irreversible loss of infectivity pathogenicity and
encystment potential after prolonged maintenance in axenic culture in vitro. Experimental Parasitology
34: 163–167.
Raetz M., Hwang S.H., Wilhelm C.L., Kirkland D., Benson A., Sturge C.R., Mirpuri J., Vaishnava S.,
Hou B., Defranco A.L., et al. (2013) Parasite-induced TH1 cells and intestinal dysbiosis cooperate in
IFN-γ- dependent elimination of Paneth cells. Nat Immunol; 14:136-42

25
Rausch S., Held J., Fischer A., Heimesaat M.M., Kuhl A.A., Bereswill S., et al. (2013) Small intestinal
nematode infection of mice is associated with increased enterobacterial loads alongside the intestinal
tract. PloS One. 8(9): e74026.
Reynolds L.A., Smith K.A., Filbey K.J., Harcus Y., Hewitson J.P., Redpath S.A., Valdez Y., Yebra M.J.,
Finlay B.B., Maizels R.M. (2014) Commensal-pathogen interactions in the intestinal tract: lactobacilli
promote infection with, and are promoted by, helminth parasites. Gut Microbes. 5(4):522-532.
Robertson B.R., O’Rourke J.L., Neilan B.A., Vandamme P., On S.L.W., Fox J.G., Lee A. (2005)
Mucispirillum schaedleri gen. nov., sp. nov., a spiral-shaped bacterium colonizing the mucus layer of
the gastrointestinal tract of laboratory rodents. International Journal of Systematic and Evolutionary
Microbiology, 55: 1199-1204.
Sewell D. L., Reinke E. K., Hogan L. H., Sandor M., and Fabry Z. (2002). Immunoregulation of CNS
autoimmunity by helminth and mycobacterial infections. Immunology Letters 82: 101–110.
Simcock D.C., Joblin K.N., Scott I., Burgess D.M., Rogers C.W., Pomroy W.E., Simpson H.V. (1999)
Hypergastrinaemia, abomasal bacterial population densities and pH in sheep infected with Ostertagia
circumcincta. International Journal of Parasitology 29: 1053–63.
Simpson H.V., Lawton D.E., Simcock D.C., Reynolds G.W., Pomroy W.E. (1997) Effects of adult and
larval Haemonchus contortus on abomasal secretion. International Journal for Parasitology 27: 825–
31.
Šlapeta J., Dowd S.E., Alanazi A.D., Westman M.E., Brown G.K. (2015) Differences in the faecal
microbiome of non-diarrhoeic clinically healthy dogs and cats associated with Giardia duodenalis
infection: impact of hookworms and coccidian. International Journal for Parasitology 45(9–10): 585–
594.
Smits M.M., Jansman A.J., Savelkoul H.F.J., Rebel A.J.M. (2014). De rol van microbiota voor een
evenwichtig afweersysteem, Tijdschrift voor Diergeneeskunde 6: 22-26.
Sokol H., Pigneur B., Watterlot L., Lakhdari O., Bermúdez-Humarán L.G. et al. (2008)
Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota
analysis of Crohn disease patients. Proceedings of the National Academy of Sciences U.S.A. 105:
16731–16736.
Thomsen L.E., Knudsen K.E., Hedemann M.S., Roepstorff A. (2006) The effect of dietary
carbohydrates and Trichuris suis infection on pig large intestine tissue structure, epithelial cell
proliferation and mucin characteristics. Veterinary Parasitology 142: 112-122.
Vaishnava S., Yamamoto M., Severson K.M., Ruhn K.A., Yu X., Koren O., et al. (2011) The
antibacterial lectin RegIIIgamma promotes the spatial segregation of microbiota and host in the
intestine. Science 334(6053):255–8.
Van den Abbeele P., Van de Wiele T., Verstraete W., Possemiers S. (2011) The host selects mucosal
and luminal associations of coevolved gut microorganisms: a novel concept. FEMS microbiology
reviews. 35(4):681–704.
van Riet E., Hartgers F.C., Yazdanbakhsh M. (2007). Chronic helminth infections induce
immunomodulation: consequences and mechanisms. Immunobiology 212: 475-490.
Walker A.W., Ince J., Duncan S.H., Webster L.M., Holtrop G. et al. (2011) Dominant and diet-
responsive groups of bacteria within the human colonic microbiota. The ISME Journal 5: 220–230.

26
Weinstock J. V., and Elliott D. E. (2009). Helminths and the IBD hygiene hypothesis. Inflammatory
Bowel Diseases 15: 128–133.
Wills-Karp M., Santeliz J., and Karp C. L. (2001) The germless theory of allergic disease: revisiting the
hygiene hypothesis. Nature Reviews Immunology 1: 69–75.
Wittebolle L., Marzorati M., Clement L., Balloi A., Daffonchio D. et al. (2009) Initial community
evenness favours functionality under selective stress. Nature 458: 623–626.
Wu G.D., Chen J., Hoffmann C., Bittinger K., Chen Y.Y. et al. (2011) Linking long-term dietary patterns
with gut microbial enterotypes. Science 334: 105–108.
Wu G.D., Lewis J.D. (2013). Analysis of the Human Gut Microbiome and Association With Disease.
Clinical Gastroenterological Hepatology 11(7):774-7.
Wu S., Li R.W., Li W., Beshah D., Harry et al. (2012) Worm Burden-Dependent Disruption of the
Porcine Colon Microbiota by Trichuris suis Infection. PLoS ONE 7:e35470.
Yatsunenko T., Rey F.E., Manary M.J., Trehan I., Dominguez-Bello M.G. et al. (2012) Human gut
microbiome viewed across age and geography. Nature 486: 222–227.
Zaiss M.M., Rapin A., Lebon L., Dubey L. K., Mosconi I., Sarter K., Piersigilli A., Menin L., Walker
A.W., Rougemont J., Paerewijck O., Geldhof P., McCoy K.D., Macpherson A.J., Croese J., Giacomin
P.R., Loukas A., Junt T., Marsland B.J., Harris N.L. (2015) The intestinal microbiota contributes to the
ability of helminths to modulate allergic inflammation. Immunity 43: 998–1010.
Zoetendal E.G., von Wright A., Vilpponen-Salmela T., Ben-Amor K., Akkermans A.D. et al. (2002)
Mucosa-associated bacteria in the human gastrointestinal tract are uniformly distributed along the
colon and differ from the community recovered from feces. Applied Environmental Microbiology 68:
3401–3407.