integrating bionenergetics, spatial scales and population...
TRANSCRIPT

Integrating Bionenergetics, Spatial Scales and Population Dynamics for Environmental Flow Assessments
Roger M. Nisbet1, Kurt E. Anderson2, Laure Pecquerie1, Lee Harrison1
1. University of California, Santa Barbara2. University of California, Riverside an

Disconnect between management objectives and management actions1
1. Locke, A., C. Stalnaker, et al. (2008). Integrated Approaches to Riverine Resource Stewardship: Case Studies, Science, Law. Pedople, and Policy. Cheyanne, WY, Instream Flow Council.

Ecological Dynamics and Instream Flow Assessments
Starting point1: “tools lacking recognition of the many dynamic feedbacks among physical and biological components of the river environment are unlikely to provide sufficient descriptions of how population or community viability will respond to changes in the flow regime.”
Recommendations included: • improving bioenergetic-based models of population dynamics to allow them
to address flow variability in streams and rivers and
• testing methods to understand the effects of spatial variability on population and community responses to changes in flow regime.
Current research:Our contribution relates to these recommendations:• Full life cycle Dynamic Energy Budget (DEB) model of Pacific salmon• Food distribution models for riverine life stages
1. Anderson et al. 2006. Ecological dynamics and the management of instream flow needs in rivers and streams. Frontiers in Ecology and Environment 4: 309-318.
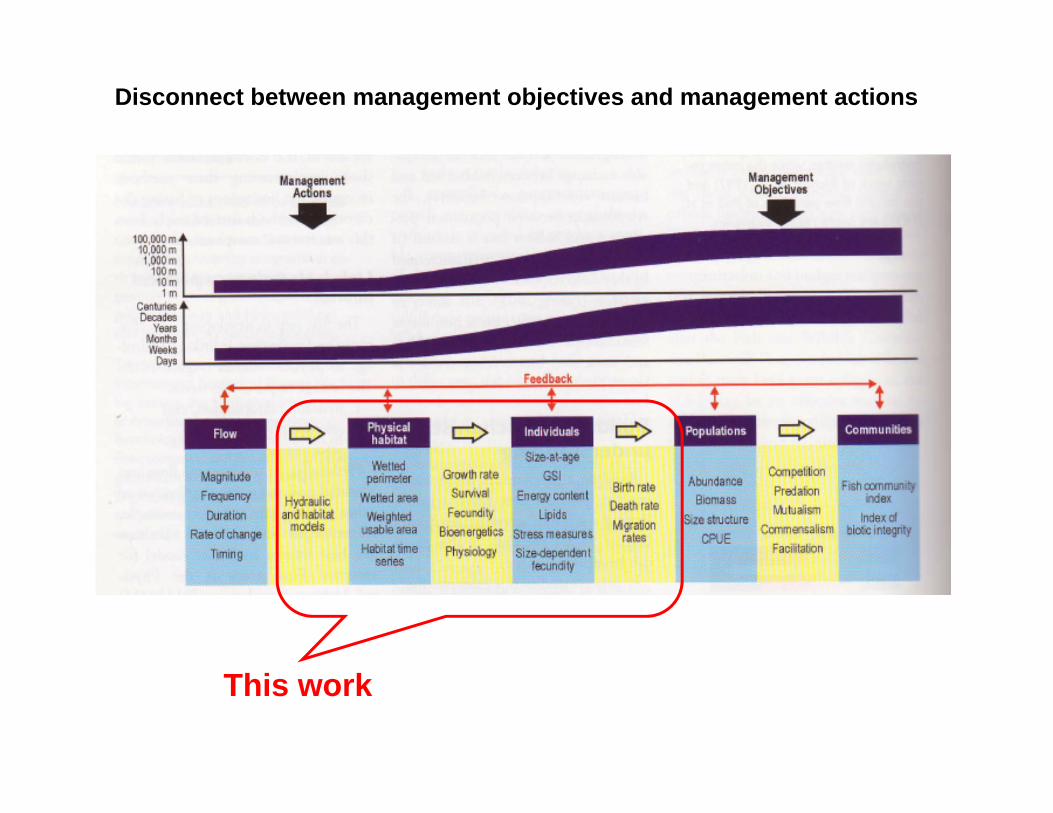
Disconnect between management objectives and management actions
This work

Full life cycle model for Pacific salmon based on Dynamic Energy Budget (DEB) theory.
1. Kooijman, S.A.L.M. Dynamic Energy Budget Theory for Metabolic Organization. Cambridge University Press, 2010
DEB theory1: conceptual framework that integrates info from all life stages (embryo, juvenile, adult)
Multiple stressors (limited food, high temperature, disease, parasitism, contaminants) can be modeled
Use of data from other salmon species when data on focal species lackingor to help parameter estimation (e.g. to provide priors for Bayesian methods)
This project: Synthesis of data from five salmon species to test the assumptions and
predictions of the DEB model – essential prerequisite to applications
Use of information from the data synthesis to parameterize the model for Chinook salmon (Oncorhynchus tshawytscha) – for work in Merced River
Calculations of sensitivity of salmon population growth rate to changes in food delivery rate that in turn are influenced by changes in flow regime

Spatial variability in food for young salmon
Recent ecological theory1 provides methodology relating habitat variability to population distributions • Applicable to benthic invertebrates - food for young salmon • Untested in real rivers with complex geometry and flow • Opens possibility of modeling effects of habitat variability over larger
stretches of river This project: Uses a 2-D hydraulic model of a re-engineered section of the Merced
River to describe the transport and settlement of macroinvertebrates –essential prerequisite to applications
Evaluates the validity of 1-D approximations to Merced River hydrology –new efficient methodology for habitat descriptions
Explores the influence of macroinvertebrate transport in a variable flow environment on characteristic length scale calculations for the Merced River – defines appropriate spatial scale for habitat descriptions
1. Anderson, K.E., Nisbet, R. M. and Diehl, S. 2006. Spatial scaling of consumer-resource interactions in advection dominated systems. American Naturalist, 168: 358-372.

Part II
Modeling the life cycle of Pacific salmon using Dynamic Energy Budget (DEB) theory
1. Motivations for considering DEB theory
2. Properties of the model distinct from other approaches
3. Validation of the model (inter- and intra-species levels)
4. Questions we will address with the model
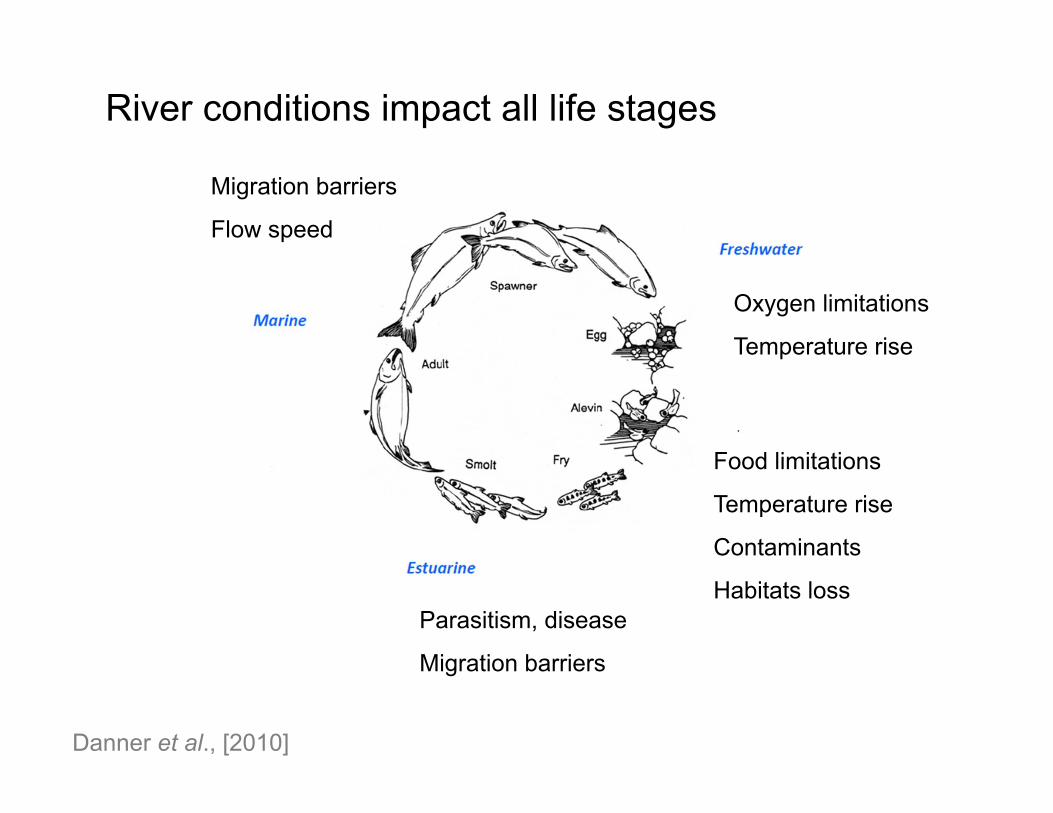
Danner et al., [2010]
River conditions impact all life stages
Oxygen limitations
Temperature rise
Food limitations
Temperature rise
Contaminants
Habitats lossParasitism, disease
Migration barriers
Migration barriers
Flow speed
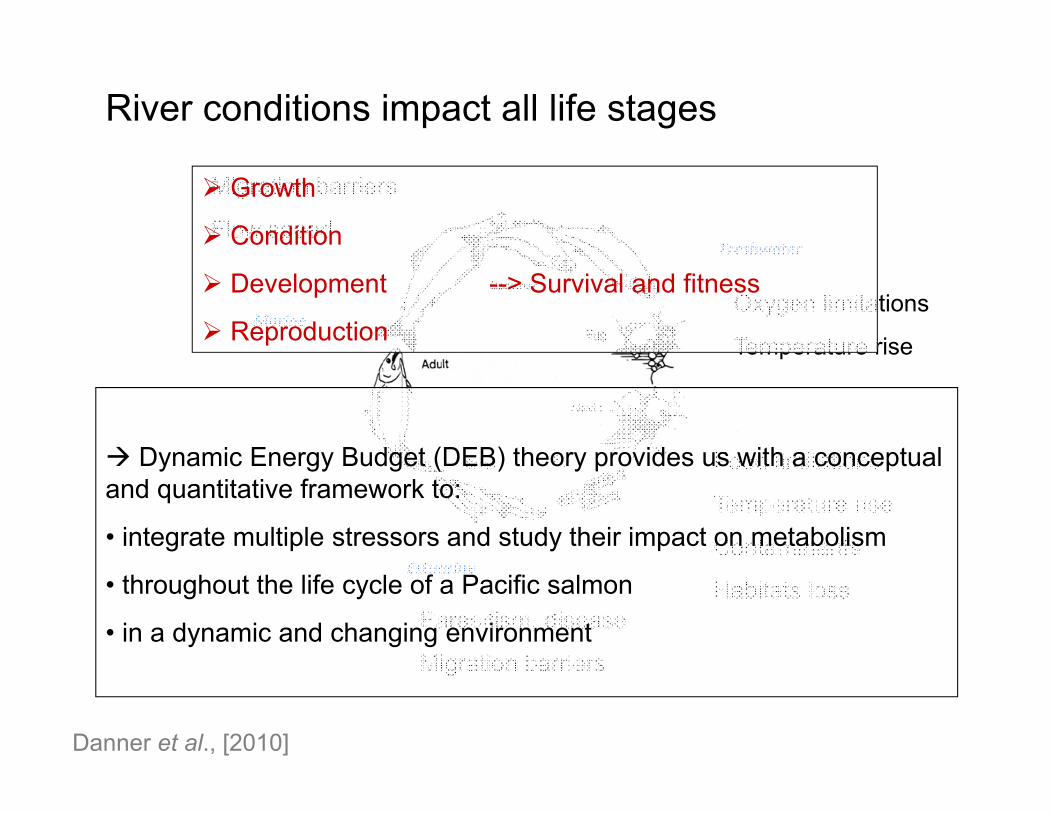
Danner et al., [2010]
River conditions impact all life stages
Oxygen limitations
Temperature rise
Food limitations
Temperature rise
Contaminants
Habitats lossParasitism, disease
Migration barriers
Migration barriers
Flow speed
Growth
Condition
Development --> Survival and fitness
Reproduction
Dynamic Energy Budget (DEB) theory provides us with a conceptual and quantitative framework to:
• integrate multiple stressors and study their impact on metabolism
• throughout the life cycle of a Pacific salmon
• in a dynamic and changing environment

maturity
1-maturity
maintenance
development
food faecesassimilation
reserve
structure
somaticmaintenance
growth
Life events in a standard DEB model
reproductionbuffer
reproduction

maturity
1-maturity
maintenance
development
food faecesassimilation
reserve
structurestructure
somaticmaintenance
growth
reproductionbuffer
reproduction
maturity
1-maturity
maintenance
development
maturitymaturity
1-maturity
maintenance
development
1-maturity
maintenance
development
food faecesassimilation
food faecesassimilation
reserve
structurestructure
somaticmaintenance
growth
reproductionbuffer
reproduction
reproductionbuffer
reproduction
Model simulations
INPUTS DEB MODEL
Food density
Temperature
Flow
Weight
Fecundity / egg size
OUTPUTS
Length

Properties of the model distinct from other approachesSame parameters for all stages – 12 parameters + 6 for migrations
• Chinook embryo model (Beer and Anderson, 1997): Weight and age at emergence as a function of temperature;12 parameters
Large number of processes: consumption, growth and maintenance + development and reproduction + mortality when limited resources (oxygen, food)• Wisconsin model (Madejian et al., 2004), Net-rate of energy intake
(NREI) (Hayes et al., 2007): consumption, growth and maintenance
Maternal effect: egg size depends on female condition
Rules for parameter variations among related species

Calibration and validation of the model
Inter-species level:• Standard DEB model• Parameter variation (genetic variability)• Length at spawning for parameter variation:
Pink < sockeye < coho < chum < Chinook• Embryo stage (egg weight, fry length and age)• Adult stage (female length, fecundity and egg weight)
Intra-species level:• Limited number of new rules and parameters (time window for
migration decision)• Variations in environmental conditions (phenotypic variability)• Age, length and weight when migrating ocean (smolts) river (returning adults)
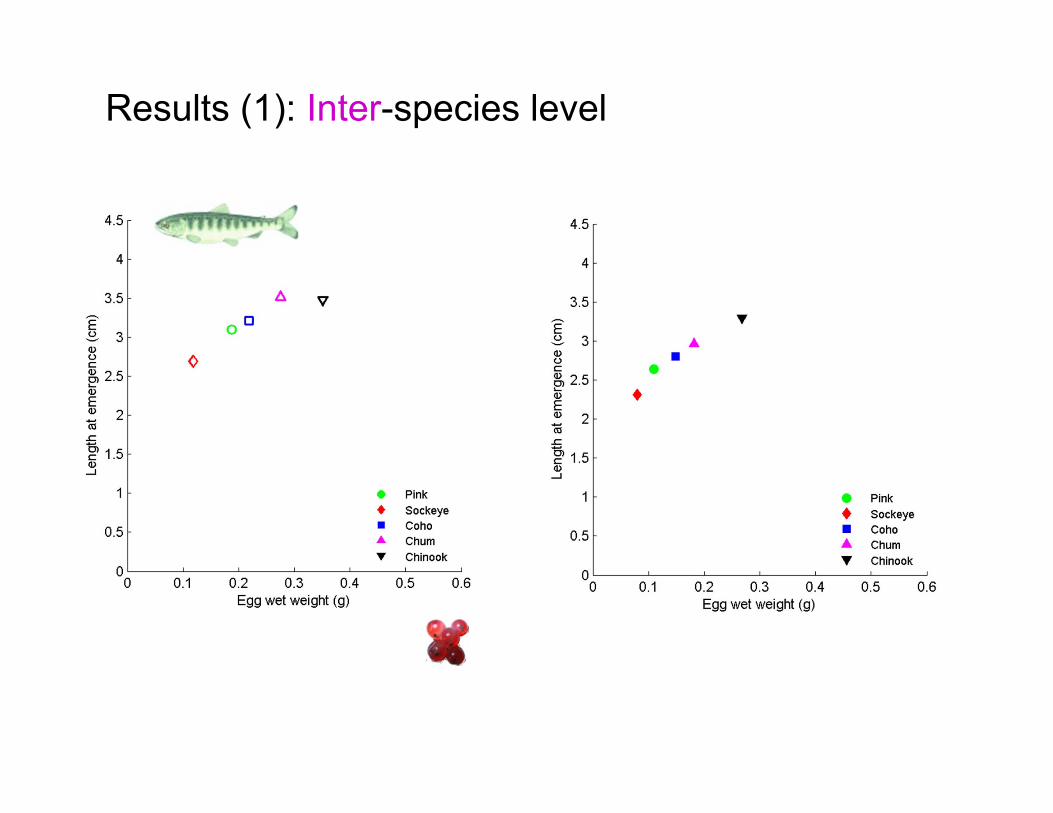
Results (1): Inter-species level

Life-history traits Observations Agreement
Female length at spawning pink < sockeye < coho < chum < chinook
Reference for our comparison
1) Reproductive material
pink < sockeye < coho < chum < chinook Yes
2) Fecundity pink < coho ≈ chum < sockeye < chinook Yes
3) Egg wet weight sockeye < pink < coho < chum < chinook Yes
4) Length at emergence sockeye < pink < coho < chum ≈ chinook Yes
5) Age at emergence
coho < chum < pink < chinook ≈ sockeye (5°C)
coho < chum ≈ chinook < pink ≈ sockeye (10°C)
Right order of magnitude but
not the rank
Results (1): Inter-species level

Results (2): Intra-species level - Embryo stage
Patterns Observations Agreement
1) Length at emergence as a function of egg wet weight
Larger eggs produce larger fry Yes
2) Weight-Length relationship at emergence
Allometric Yes
3) Age at emergence as a function of egg wet weight
In Chinook, age at emergence slightly increases with egg weight at 10C or stay constant at other
temperatures)
No
4) Length at emergence as a function of temperature
In Chinook, length at emergence decreases with temperature No
5) Age at emergence as a function of temperature
Age at emergence decreases with temperature Yes
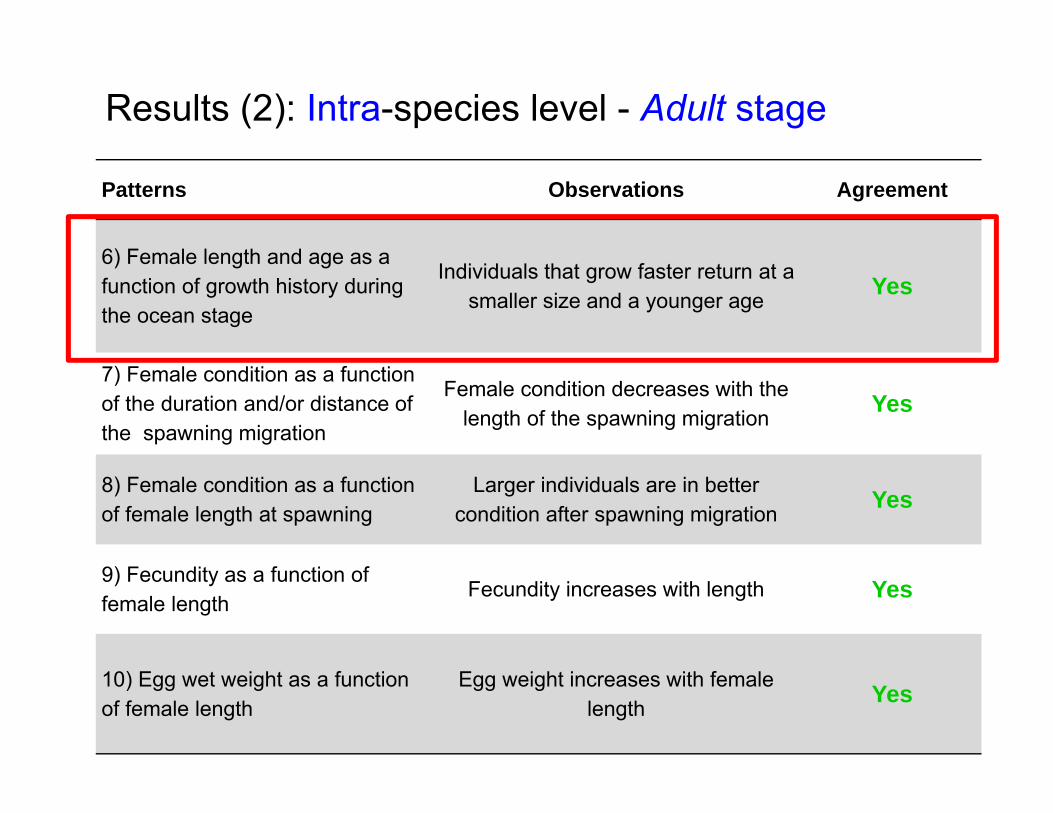
Patterns Observations Agreement
6) Female length and age as a function of growth history during the ocean stage
Individuals that grow faster return at a smaller size and a younger age Yes
7) Female condition as a function of the duration and/or distance of the spawning migration
Female condition decreases with the length of the spawning migration Yes
8) Female condition as a function of female length at spawning
Larger individuals are in better condition after spawning migration Yes
9) Fecundity as a function of female length
Fecundity increases with length Yes
10) Egg wet weight as a function of female length
Egg weight increases with female length Yes
Results (2): Intra-species level - Adult stage

Results (3) – Calibration to Chinook data
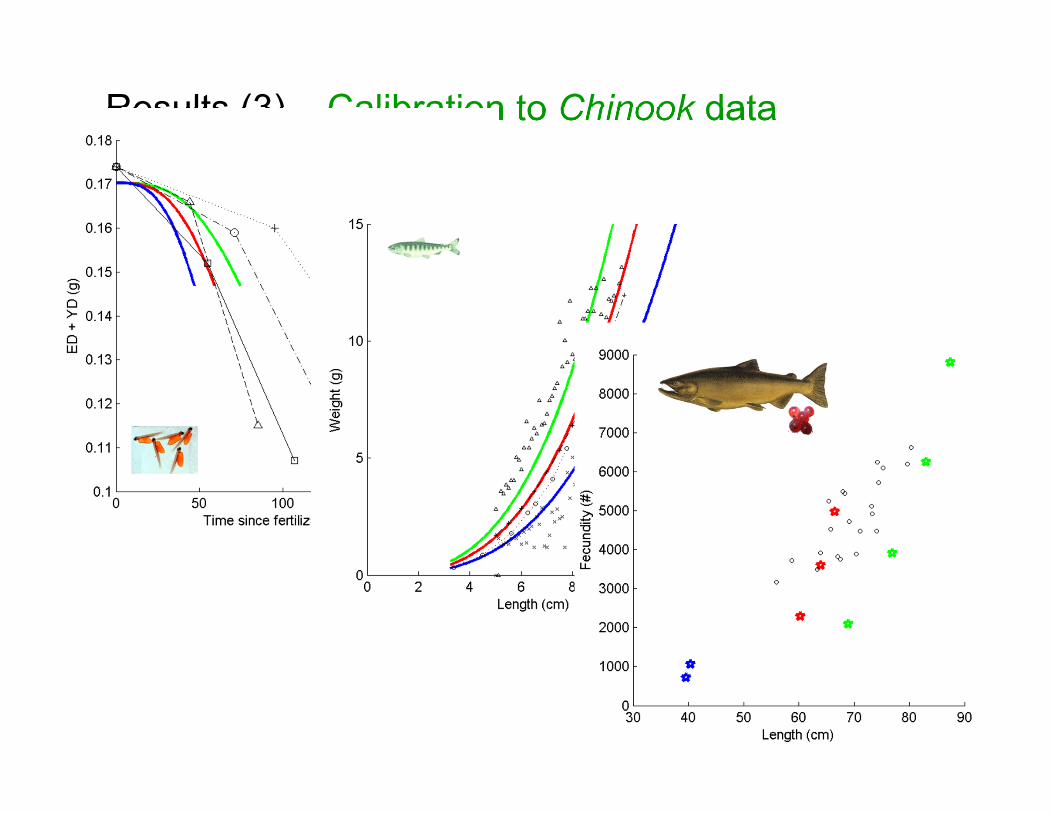
Results (3) – Calibration to Chinook data

Summary Part II
• We have a generic model for the life cycle of a Pacific salmon
• We need more details for the impact of temperature on metabolic processes (cf Coho embryo development for instance)
• Model captures most of the variation in life-history traits among the 5 species of Pacific salmon in North-America
• Model captures patterns at the intra-species level and variation in age and length at migration in particular
• Good fit of the model to Chinook data

Further developments
Short-term: • Include more data for Chinook model (Bayesian framework)• Juveniles: individual growth AND development rates in varying
flow conditions• Eggs: oxygen limitations• Analyzing otolith and scale patterns to reconstruct individual food
histories
Long-term:• Coupling with 2D model (river, coastal ocean)• Adults: survival during migration, female condition after
migration• Long-term population growth rates (individual growth, stage
transitions, fecundity, survival)• Application to Steelhead

Part III
Spatial variability in food delivery for juvenile salmonids
1. Goals
2. Modeling framework
3. Calibration/validation of 2D flow model
4. Flow-Drift Simulations

Project Goals Spatial variability in food for juvenile salmon
1.Formulate 2D numerical models of flow and transport of macroinvertebrates
– Improve food delivery functions in Bioenergetics/DEB models
– Test in simple river under range of flows
2. 2D results used as input to simpler 1D flow and drift model that can be applied over larger spatial scales (Anderson)

Field SiteRobinson Reach, Merced River
o Recently re-engineered reach of the Merced River, CA.
o Single-thread, meandering planform, with alternating deep pools and shallow riffles.
o Utilized existing topographic and hydraulic data sets that were collected with collaborators Tom Dunne (UCSB) and Carl Legleiter (U Wyoming).

Flow ModelingMIKE 21 Code (DHI)
o Model Inputs– Topography– Roughness– Discharge– Stage
o Outputs– Depth– Velocity

Drift ModelingMIKE 21 Code (DHI)
o LaGrangian Particle Tracking Algorithm– Particle concentration– Particle trajectory
o Vertical Profile– Assumed logarithmic form
o Transport Processes– Invertebrates released at 0.6*h
o Settlement Processes– Accounts for the time an organism
spends in the drift given its settling velocity (ωs)
– Invertebrates removed from simulation once settled out of drift
o Dispersion– Random-walk approach– Values calculated as a function of the
eddy viscosity
30 40 50 60 70 80U(cm/s)
0
0.2
0.4
0.6
0.8
1
Z/h
Zo
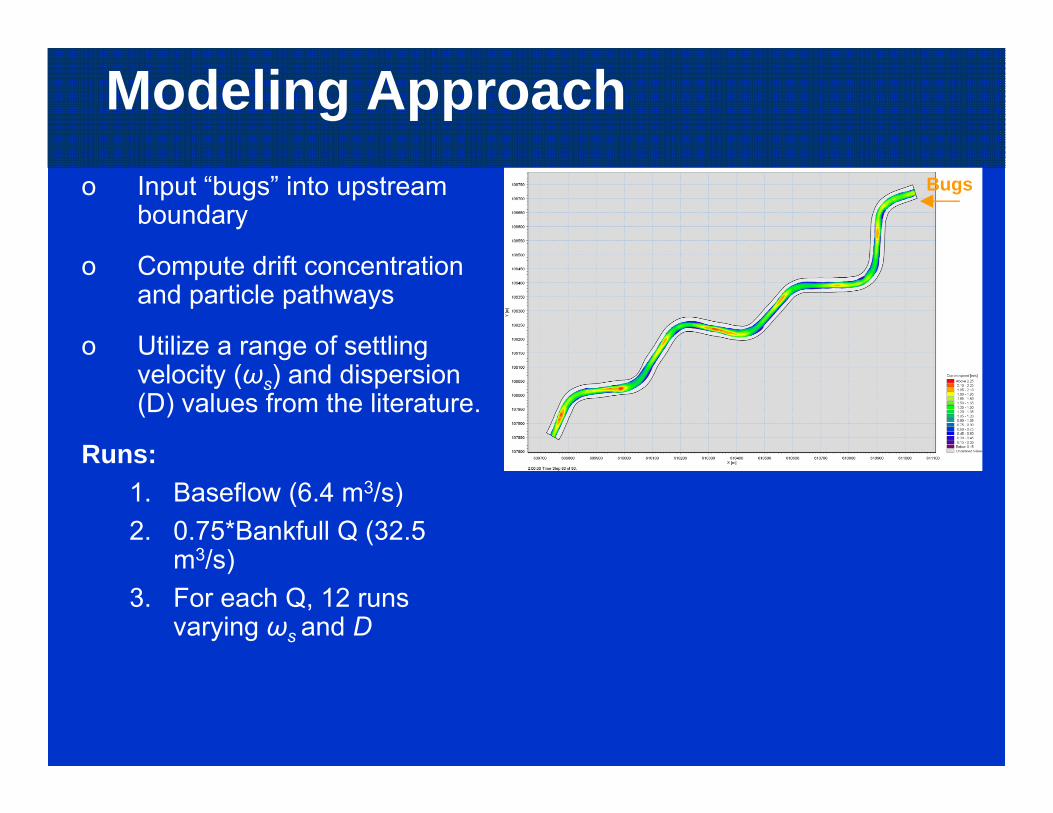
Modeling Approacho Input “bugs” into upstream
boundary
o Compute drift concentration and particle pathways
o Utilize a range of settling velocity (ωs) and dispersion (D) values from the literature.
Runs:1. Baseflow (6.4 m3/s)2. 0.75*Bankfull Q (32.5
m3/s)3. For each Q, 12 runs
varying ωs and D
Bugs

Model CalibrationWater Surface Elevation (Q = 32.5 m3/s)
o WSE measured along right and left banks (Q = 6.4 and 32.5 m3/s)
o Good overall agreement (WSE RMSE = 0.037 m)
WSE – Left Bank WSE – Right Bank
0 200 400 600 800Streamwise Distance (m)
58.5
59
59.5
60
60.5
61
Wat
er S
urfa
ce E
leva
tion
(m)
Obs (RB)Pred (RB)
58.5 59.5 60.5Observed WSE (m)
58.5
59.5
60.5
Pre
dict
ed W
SE (m
) 1:1
0 200 400 600 800Streamwise Distance (m)
58.5
59
59.5
60
60.5
61
Wat
er S
urfa
ce E
leva
tion
(m)
Obs. (LB)Pred. (LB)
58.5 59.5 60.5Observed WSE (m)
58.5
59.5
60.5
Pre
dict
ed W
SE
(m) 1:1
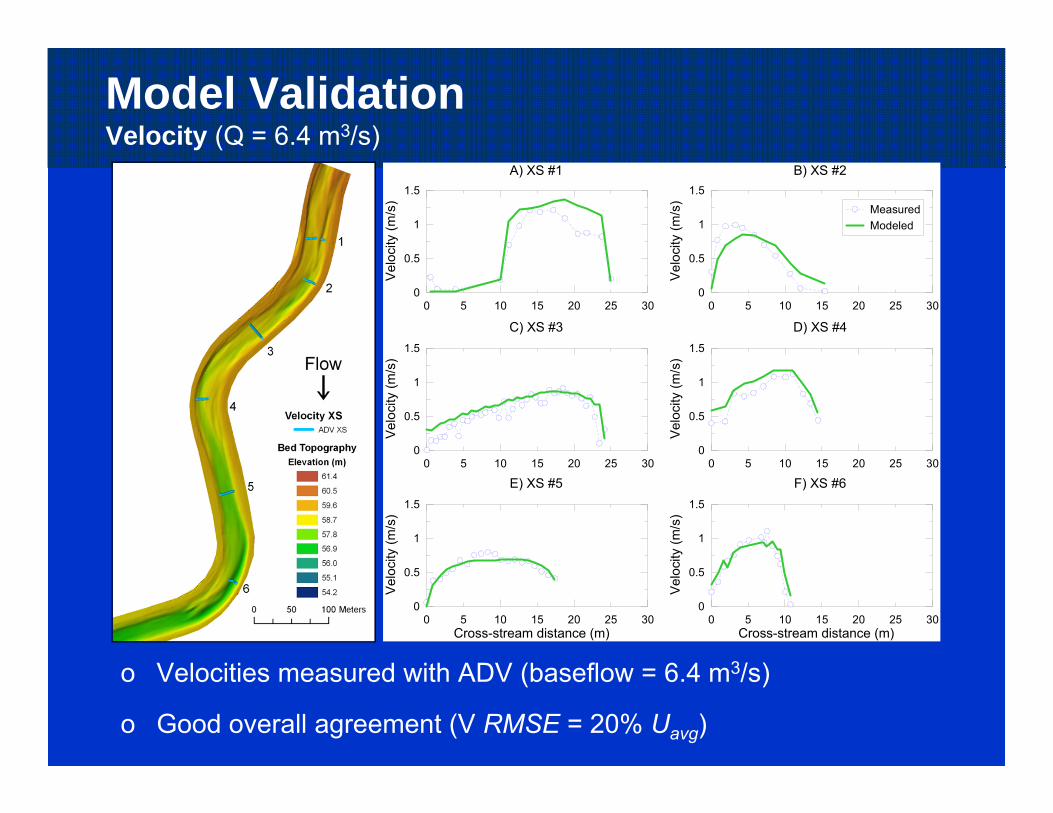
Model ValidationVelocity (Q = 6.4 m3/s)
0 10 20 305 15 250
0.5
1
1.5
Vel
ocity
(m/s
)
0 10 20 305 15 250
0.5
1
1.5
Vel
ocity
(m/s
)
0 10 20 305 15 250
0.5
1
1.5
Vel
ocity
(m/s
)
0 10 20 305 15 250
0.5
1
1.5
Vel
ocity
(m/s
)
0 10 20 305 15 25Cross-stream distance (m)
0
0.5
1
1.5
Vel
ocity
(m/s
)
0 10 20 305 15 25Cross-stream distance (m)
0
0.5
1
1.5
Vel
ocity
(m/s
)
MeasuredModeled
A) XS #1
C) XS #3
E) XS #5
B) XS #2
D) XS #4
F) XS #6
o Velocities measured with ADV (baseflow = 6.4 m3/s)
o Good overall agreement (V RMSE = 20% Uavg)

Results: Flow FieldQ = 6.4 m3/s
o Velocity is uniform through straight riffles
o Peak velocity located in curved pools
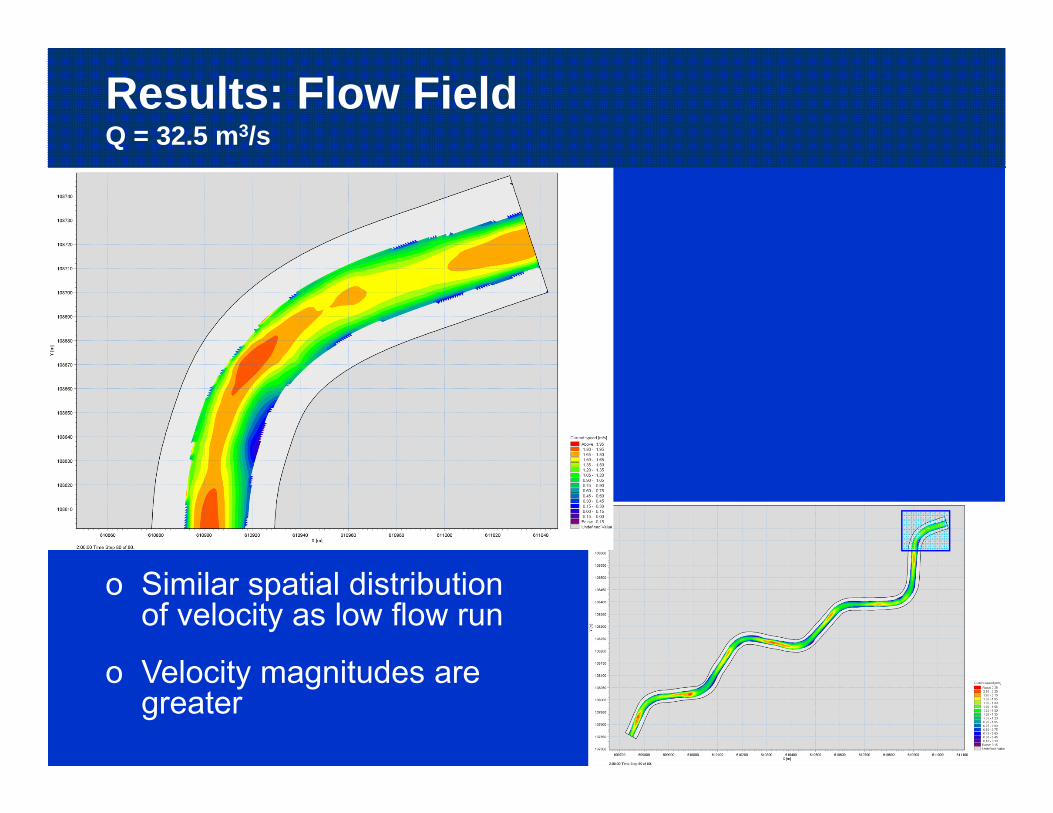
Results: Flow FieldQ = 32.5 m3/s
o Similar spatial distribution of velocity as low flow run
o Velocity magnitudes are greater

Q (m3/s)
s
(m/s)LEV (m2/s)
D h and D v
(m2/s)6.4 0.0005 0.003 06.4 0.0005 0.003 0.0036.4 0.0005 0.003 0.0156.4 0.0005 0.003 0.036.4 0.005 0.003 06.4 0.005 0.003 0.0036.4 0.005 0.003 0.0156.4 0.005 0.003 0.036.4 0.05 0.003 06.4 0.05 0.003 0.0036.4 0.05 0.003 0.0156.4 0.05 0.003 0.0332.5 0.0005 0.01 032.5 0.0005 0.01 0.0132.5 0.0005 0.01 0.0532.5 0.0005 0.01 0.132.5 0.005 0.01 032.5 0.005 0.01 0.0132.5 0.005 0.01 0.0532.5 0.005 0.01 0.132.5 0.05 0.01 032.5 0.05 0.01 0.0132.5 0.05 0.01 0.0532.5 0.05 0.01 0.1
Model Simulations24 Runs: With and Without Dispersion

0 400 800 1200 1600 2000Streamwise Distance (m)
0
5
10
15
20
Num
ber I
ndiv
idua
ls
Low flow (w/Disp)Low Flow (no/Disp)High Flow (w/Disp)High Flow (no/Disp)
0 400 800 1200 1600 2000Streamwise Distance (m)
0
5
10
15
20
Num
ber I
ndiv
idua
ls
0 400 800 1200 1600 2000Streamwise Distance (m)
0
5
10
15
20
Num
ber I
ndiv
idua
ls
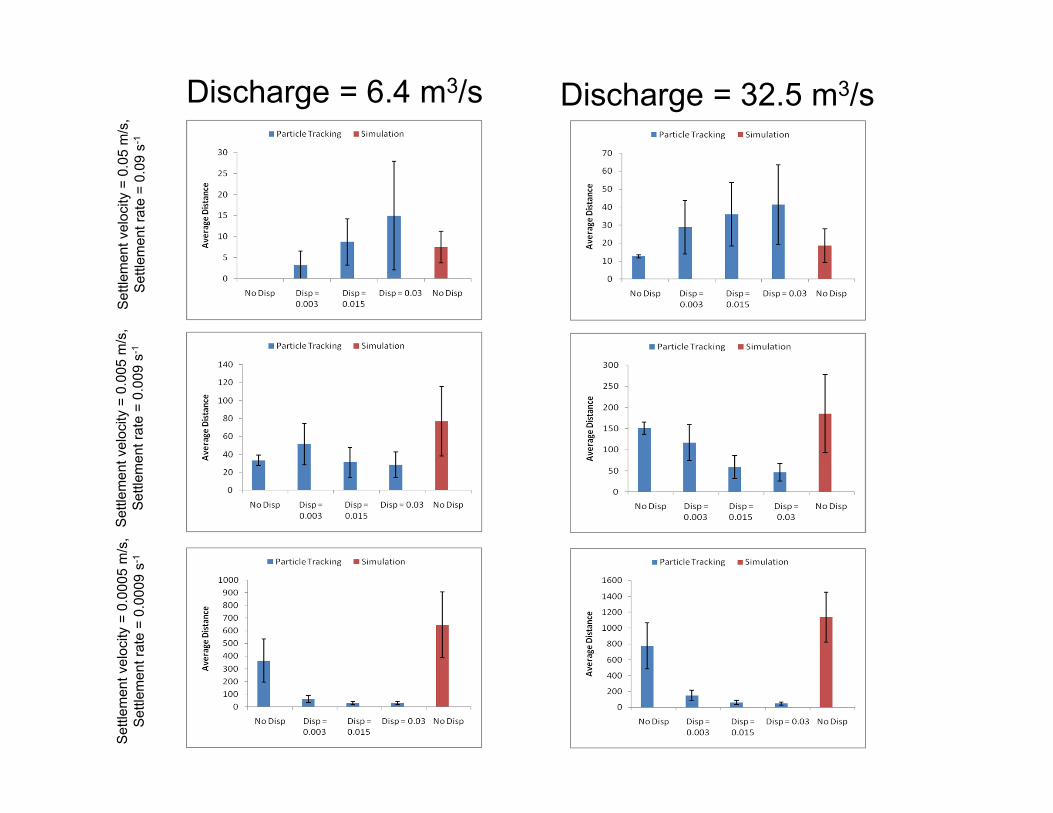
Dispersal DistancesVariable Q, D and ωs
o Travel distances decrease with higher settling velocity
o Without dispersion– Particles settle out over
a small range of distances.
o At low flow– Dispersion increases the
average dispersal distance and the variance.
o At high flow– Dispersion decreases
the mean dispersal distance.
A) ωs = 0.05 (m/s)
B) ωs = 0.005 (m/s)
C) ωs = 0.0005 (m/s)
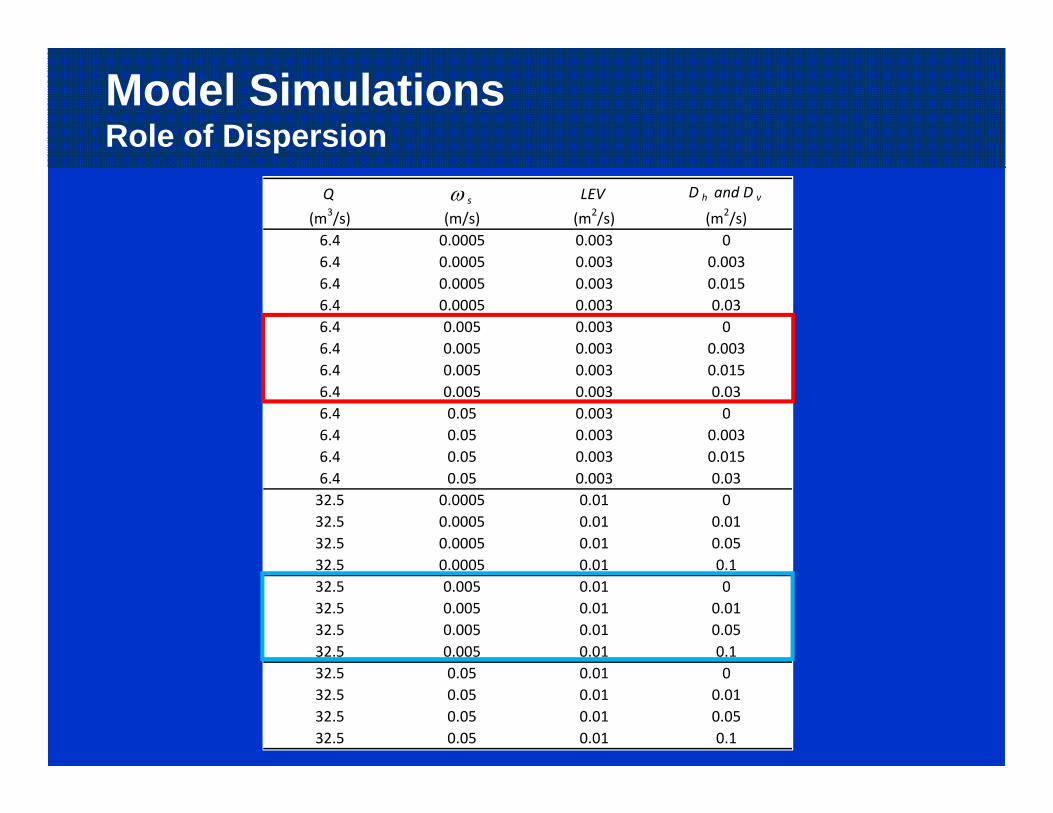
Model SimulationsRole of Dispersion
Q (m3/s)
s
(m/s)LEV (m2/s)
D h and D v
(m2/s)6.4 0.0005 0.003 06.4 0.0005 0.003 0.0036.4 0.0005 0.003 0.0156.4 0.0005 0.003 0.036.4 0.005 0.003 06.4 0.005 0.003 0.0036.4 0.005 0.003 0.0156.4 0.005 0.003 0.036.4 0.05 0.003 06.4 0.05 0.003 0.0036.4 0.05 0.003 0.0156.4 0.05 0.003 0.0332.5 0.0005 0.01 032.5 0.0005 0.01 0.0132.5 0.0005 0.01 0.0532.5 0.0005 0.01 0.132.5 0.005 0.01 032.5 0.005 0.01 0.0132.5 0.005 0.01 0.0532.5 0.005 0.01 0.132.5 0.05 0.01 032.5 0.05 0.01 0.0132.5 0.05 0.01 0.0532.5 0.05 0.01 0.1
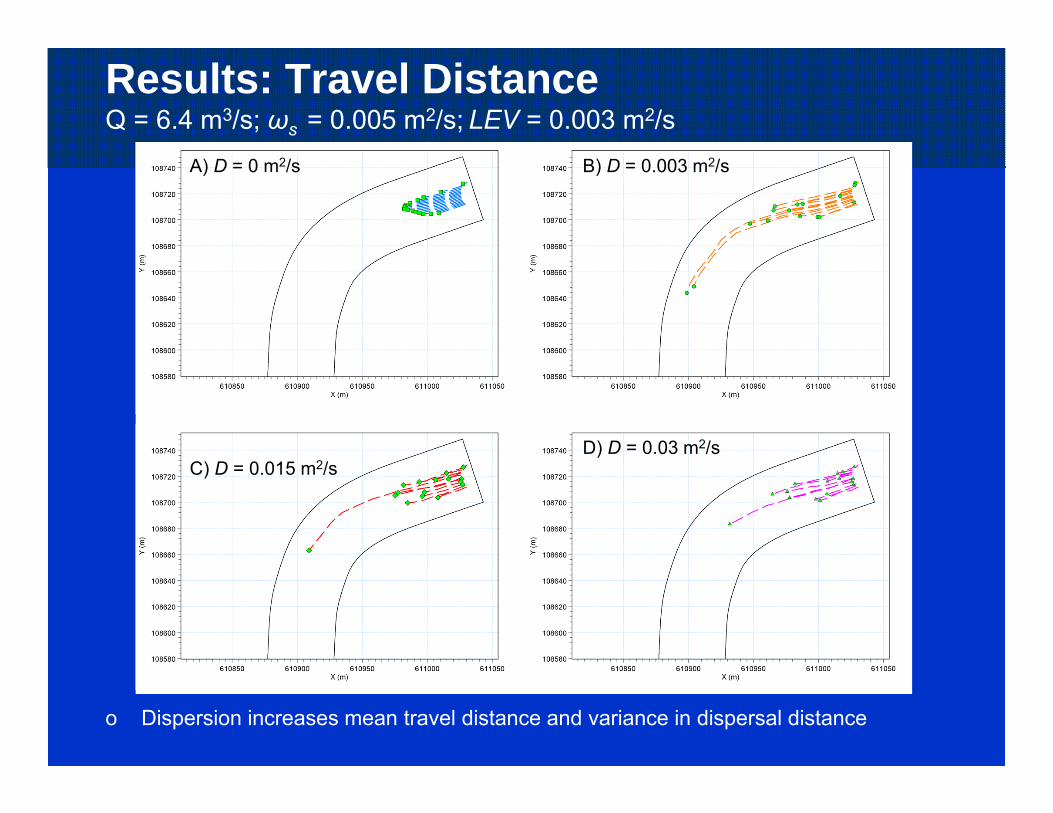
Results: Travel DistanceQ = 6.4 m3/s; ωs = 0.005 m2/s; LEV = 0.003 m2/s
o Dispersion increases mean travel distance and variance in dispersal distance
A) D = 0 m2/s B) D = 0.003 m2/s
C) D = 0.015 m2/sD) D = 0.03 m2/s

Results: Travel DistanceQ = 32.5 m3/s; ωs = 0.005 m2/s; LEV = 0.01 m2/s
A) D = 0 m2/s B) D = 0.01 m2/s
C) D = 0.05 m2/s D) D = 0.1 m2/s
o Dispersion decreases mean travel distance but increases variance

2D Flow-Drift Summary
o We have a validated 2D flow model of the Merced River
o Model is capable of computing drift transport and settlement at low and high flows
o Preliminary Results:1. Invert pathways dictated by high velocity core.2. Invert travel distances:
– ↑ with flow velocity – ↓ decrease with higher ωs
3. Dispersion increases the variance in dispersal distances.
Kurt will discuss 1D flow-drift transport

Part IV: Flow regime and spatial scales of population response
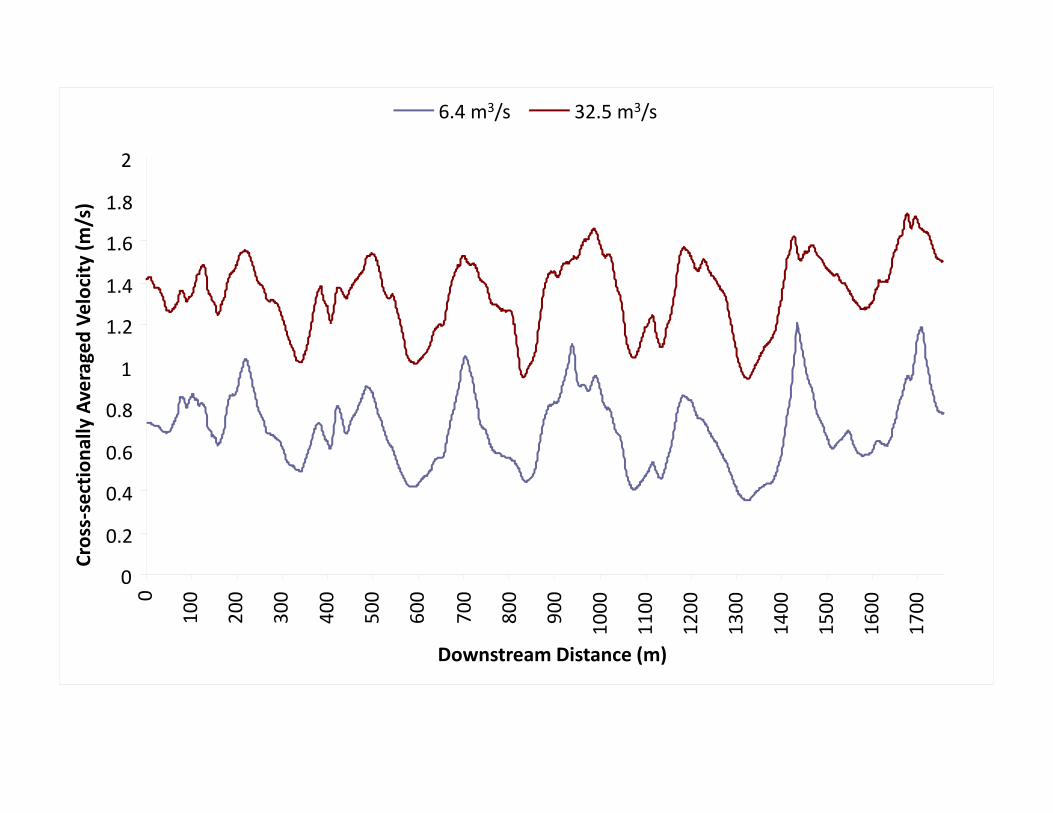
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
20
100
200
300
400
500
600
700
800
900
1000
1100
1200
1300
1400
1500
1600
1700
Downstream Distance (m)
Cross‐sectiona
lly Averaged Ve
locity (m
/s)
6.4 m3/s 32.5 m3/s
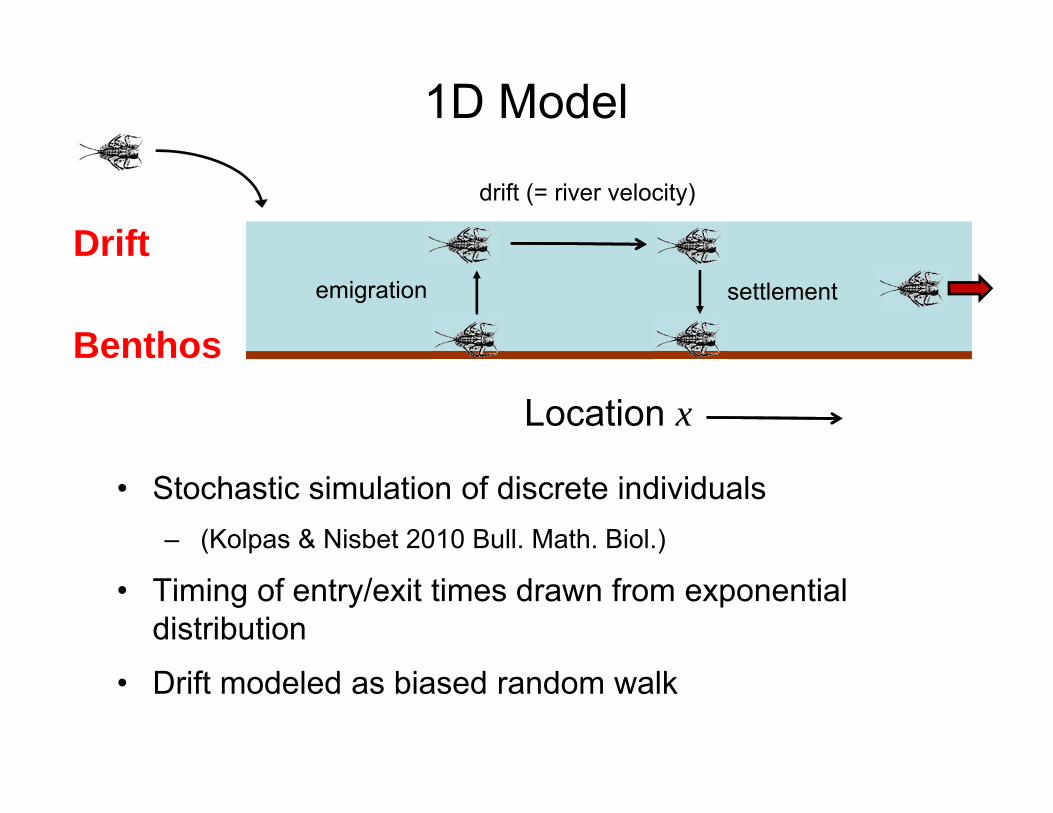
Location x
• Stochastic simulation of discrete individuals– (Kolpas & Nisbet 2010 Bull. Math. Biol.)
• Timing of entry/exit times drawn from exponential distribution
• Drift modeled as biased random walk
1D Model
Drift
Benthosemigration settlement
drift (= river velocity)

Dispersal distribution
0
0.2
0.4
0.6
0.8
1
0 5 10 15 20 25
Prop
ortio
n of
Pop
ulat
ion
in th
e D
rift
Distance Downstream
xvelocitysettlementexp
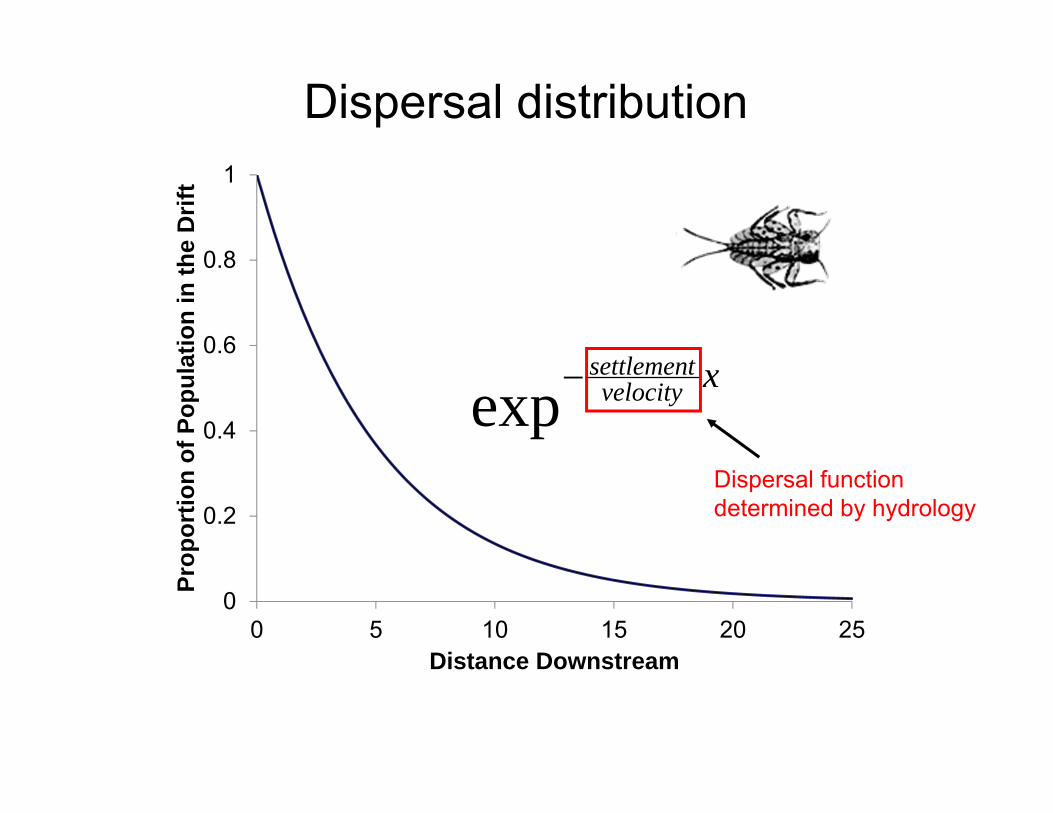
Dispersal distribution
0
0.2
0.4
0.6
0.8
1
0 5 10 15 20 25
Prop
ortio
n of
Pop
ulat
ion
in th
e D
rift
Distance Downstream
Dispersal function determined by hydrology
xvelocitysettlementexp

Dispersal distribution
y = 0.0918x1.0922
R2 = 0.435
0
5
10
15
20
25
30
35
0 10 20 30 40 50 60 70 80 90 100
Reported average stream velocity (cm/s)
Aver
age
drift
dis
tanc
e (m
)
McLay 1970Elliott 1971Ciborowski 1983Larkin and McKone 1985Allan and FeifarekLancaster et al. 1996Oldmeadow et al. 2010
y = 0.0918x1.0922
R2 = 0.435
0
5
10
15
20
25
30
35
0 10 20 30 40 50 60 70 80 90 100
Reported average stream velocity (cm/s)
Aver
age
drift
dis
tanc
e (m
)
McLay 1970Elliott 1971Ciborowski 1983Larkin and McKone 1985Allan and FeifarekLancaster et al. 1996Oldmeadow et al. 2010
Avg. distance = avg. velocity/settlement

0
50
100
150
200
250
300
350
400
450
500
0 500 1000 1500 2000 2500 3000 3500 4000 4500 5000
Time (10 sec.)
Tota
l Ind
ivid
uals
0
50
100
150
200
250
300
350
400
450
500
0 500 1000 1500 2000 2500 3000 3500 4000 4500 50002
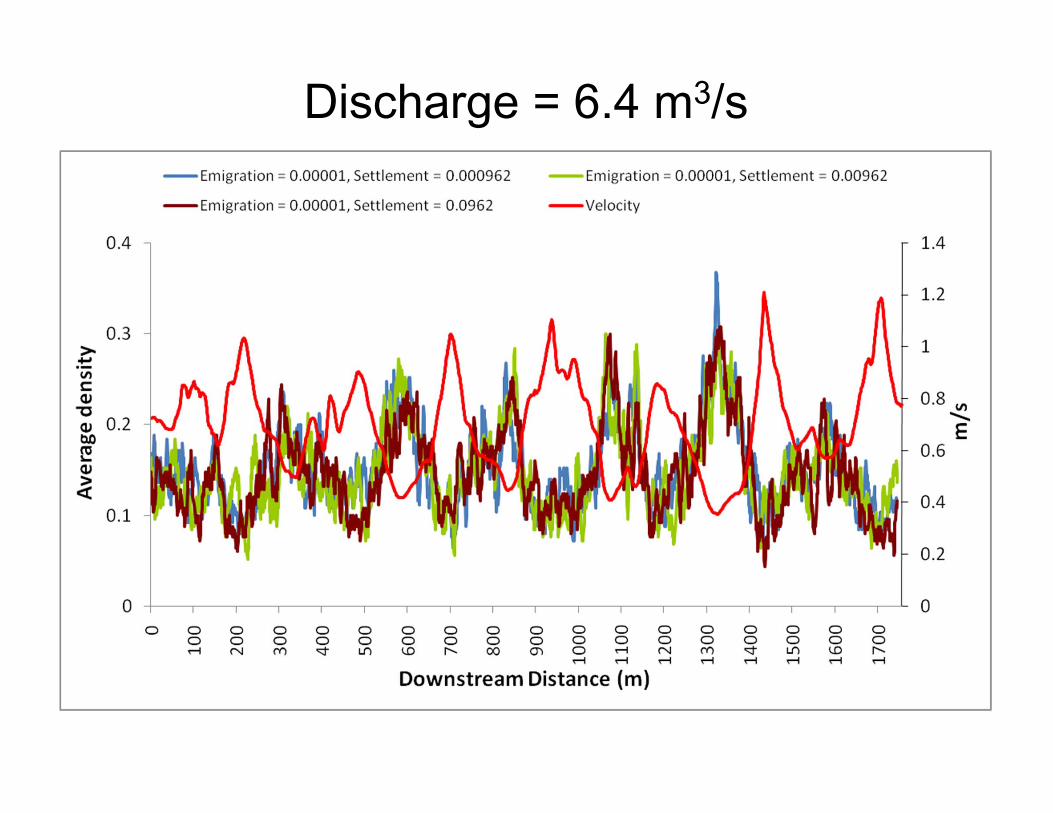
Discharge = 6.4 m3/s

Discharge = 32.5 m3/s
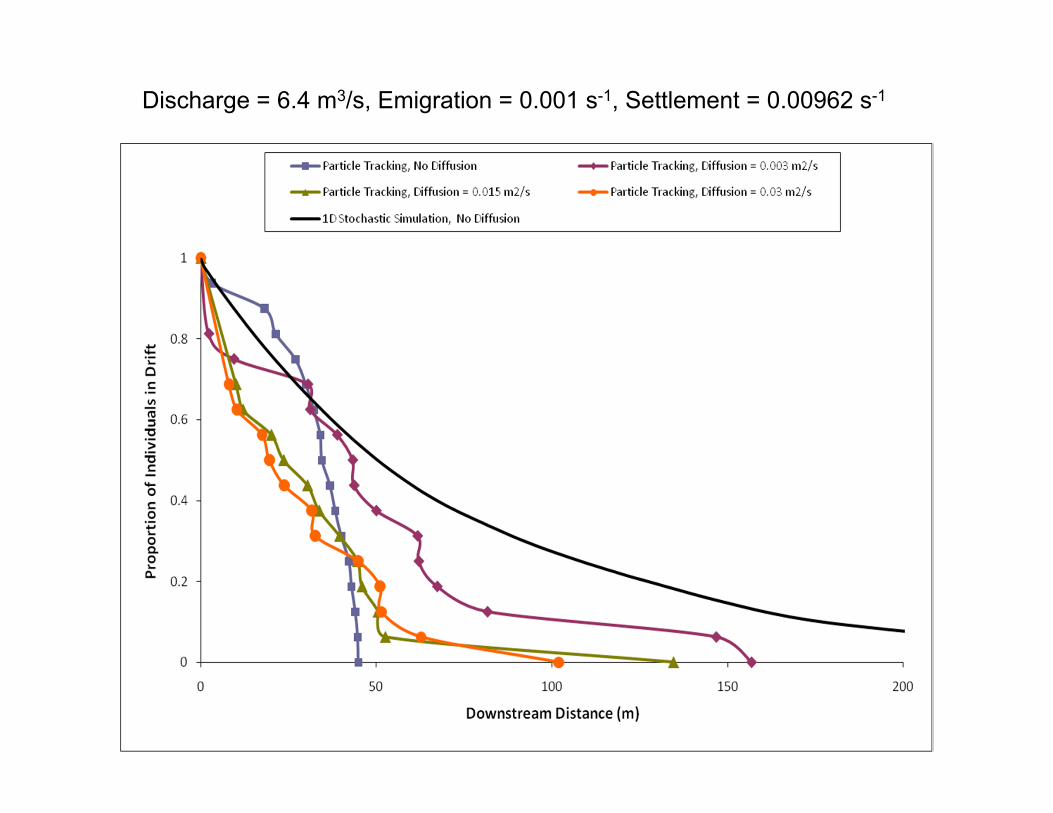
Discharge = 6.4 m3/s, Emigration = 0.001 s-1, Settlement = 0.00962 s-1
Discharge = 6.4 m3/s Discharge = 32.5 m3/sS
ettle
men
t vel
ocity
= 0
.05
m/s
,S
ettle
men
t rat
e =
0.09
s-1
Set
tlem
ent v
eloc
ity =
0.0
05 m
/s,
Set
tlem
ent r
ate
= 0.
009
s-1S
ettle
men
t vel
ocity
= 0
.000
5 m
/s,
Set
tlem
ent r
ate
= 0.
0009
s-1

Initial Conclusions• Inverts appear to follow similar trajectories at low and
high flows
• Invert pathways dictated by high velocity core
• Travel distance varies with assumed interaction with flow conditions– Qualitatively similar between 1D and 2D
• With spatially uniform rates of entry and exit from benthos, more end up in riffles –consistent with observations on Baetis
www.dfg.ca.gov

Food delivery• Tests of 1D model in more complex hydrology
• Complex structure (e.g. woody debris, boulders, gravel augmentation)
• Representation of “behavior” in inverts (entry/exit)
• Characteristic length scales to guide appropriate resolution of habitat descriptions
http://www.usbr.gov
http://www.fs.fed.us
http://www.flyfishingtraditions.com

Disconnect between management objectives and management actions1
This talk
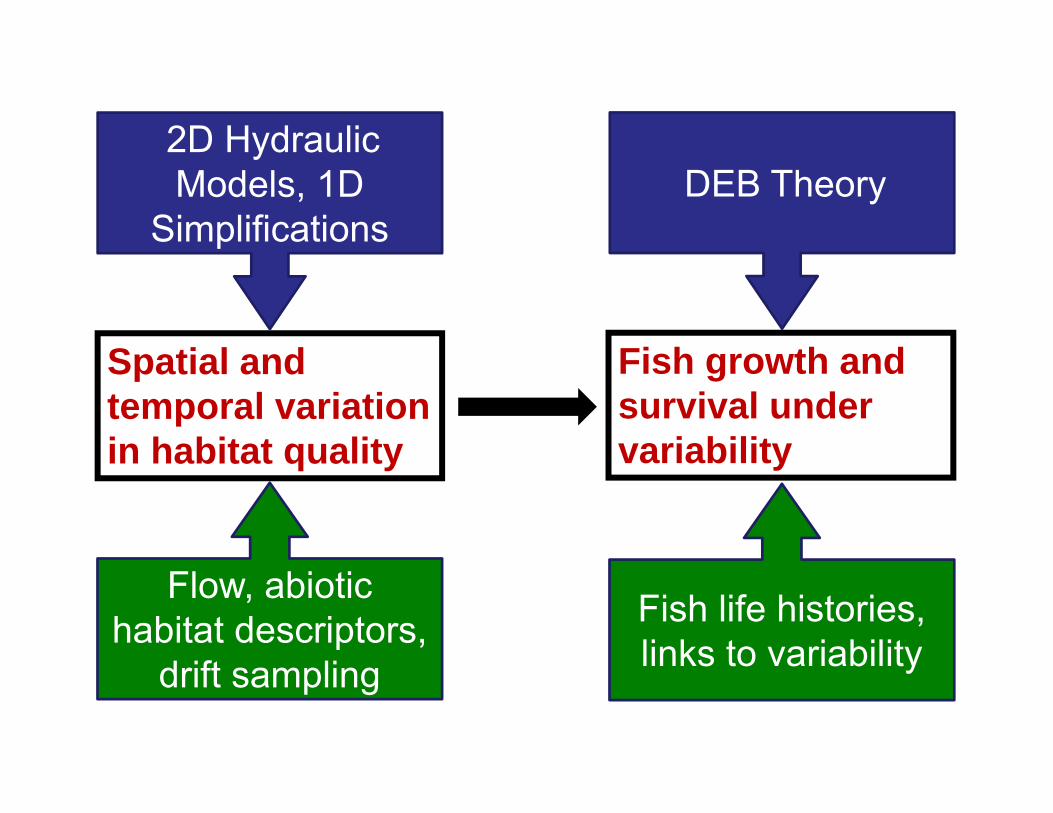
Spatial and temporal variation in habitat quality
Fish growth and survival under variability
2D Hydraulic Models, 1D
SimplificationsDEB Theory
Flow, abiotichabitat descriptors,
drift sampling
Fish life histories, links to variability

% change in flow
% change in habitat
% change in drift delivery
% change in fish growth and survival

AcknowledgementsThanks to:
Alison KolpasJonathan SarhadMargaret SimonJohn WilliamsBrad CardinaleLindsey AlbertsonTom DunneLeah JohnsonBas KooijmanCarl LegleiterAleksandra WydzgaDale KerperJulio ZysermanAlison Amenta
Funding:
California Energy Commission Public Interest Energy Research Program
National Science Foundation
Calfed Bay-Delta Authority Science Program
UC-Riverside