institut curie-recherche, umr2027 cnrs/ic centre … · tissulaire. les nouvelles données en...
TRANSCRIPT

Données nouvelles en radiobiologie
parDietrich AVERBECK
Institut Curie-Recherche, UMR2027 CNRS/ICCentre Universitaire d’0rsay
91405 [email protected]
A.M.T.S.N
Salons de l’Aveyron,17, rue de l’Aubrac à Paris 12ème
Journée de formation continue des médecins du travail habilités nucléaire

Introduction (1)Introduction (1)
• Pendant sa vie, l’homme est exposé le plus souvent à de faibles doses de rayonnements ionisants (RI), à l’exception de traitements radiothérapeutiques ou d’accidents (Chernobyl). L’évaluation des risques des faibles doses pour la santé de l’homme constitue donc un enjeu important pour la radioprotection.
• Les études épidémiologiques n’ont pas assez de puis sance statistique pour déterminer les risques engendrés par le s très faibles expositions. Donc, les études fondamen tales mécanistiques sont essentielles pour mieux comprendre les effets biologiques des faibles doses de RI et p our évaluer les risques.

IntroductionIntroduction (2)
• La recherche récente a fait apparaître de nouvelles données qui mettent en question plusieurs concepts radiobiologiques précédemment établis concernant les effets de faibles doses.
• Voir le rapport récent de l’Académie Nationale de Médecine et de l’Académie des Sciences du 30 Mars 2005 sur la relation dose-effet et l’estimation des effets cancérogènes des faibles doses de rayonnements ionisants.

Objectifs de l’exposé:
• Montrer les progrès réalisés dans la compréhension de certains mécanismes moléculaires et cellulaires, notamment ceux impliqués dans les effets de faibles dose de rayonnement ionisant

Exposition aux radiations ionisantes
Régions de haut niveau de radioactivité 50 à 100 mSv/an(Inde (Kérala), Brésil, Iran)
Radioactivité naturelle en France 2,4 mSv/an(Rayons cosmiques: 0,4 mSv/an, expositions telluriques: 0,5 mSv/an, internes: 1,5 mSv/an)
Montagne à 1500 m d’altitude 3,6 mSv/an
Scanner (tomographie) 6 mSv /an
Diagnostic (radiographie des poumons…) 1 mSv/an____________________________________________________Réglementation : Limite d’exposition
pour le grand public 1 mSv/anpour les travailleurs 20 mSv/an
____________________________________________________

Evaluation du risque d’une exposition aux RI
Risque relatif
Exposition
Effets avérés
0X
RR
• Relation linéairesans seuil (RLSS)
Effetshypothétiques
RR
• quadratique
• Hormésis
• Réglementation

Etudes épidémiologiques
Hiroshima-Nagasaki Leucémies seuil # 150 mSv76.000 H. 200 mSv Cancers solides NS < 100 mSv
Tumeurs solides : curvilinéairesCIRC 1995 Leucémies NS < 400 mSv96.000 T. Nucléaire Cancers solides NS
CIRC 2004600.000 T. nucléaire
Radiologues > 1960 Leucémies NS220.000 H. 10 - 50 mSv / an Cancers solides NS
Navigants Leucémies NS47.000 H. 1,5 - 6 mSv / an Cancers solides NS
Mélanomes
(selon Aurengo et al. 2005)

Consensus sur la radiocarcinogenèse
• Les études épidémiologiques (survivants de Hiroshima et Nagasakiet expositions accidentelles) montrent que le risque de cancer croît à partir de 100-200 mGy de RI.
• Les expositions environnementales ou diagnostiques sont beaucoupplus faibles.
� Discussion actuelle sur les effets éventuels des irra diations diagnostiques (tomographie..) ou en dehors des champs d’irradiation lors d’une radiothérapie conformationnelle
� Discussion sur la contamination interne avec des radionucléïdes

Etapes de la cancérogenèse
Initiation Promotion Progression Cancer
Tumeur Métastases
Rayonnementsionisants
dommagesde l’ADN mutations
réparation de l ’ADN
instabilitégénétique
sélection clonale
cellules malignes
induction de gènes
transduction des signaux
Influence du microenvironnement
tissulaire

Les nouvelles données en radiobiologie
1 - Conception classique (<1985) : Relation linéaire sans seuilCertaines mutations provoquent un cancerProbabilité de mutation augmente proportionnellement à la dose
2 - 1985-2000: Mécanismes de sauvegarde- Réparation de l’ADN : fidèleou fautive- Élimination des cellules lésées
par la mort mitotique (lésion empêche la division)par apoptose (mort programmée)
3 - Conception Contemporaine (2000-2008)- Nocivité de RI n’est pasconstante avec la dose- Mécanismes de défense sont activés à de très faibles doses- Probabilité de réparation varie avec la dose de 4 à 100 mSv,
l’apoptose intervient

Plusieurs phénomènes biologiques modifient la réponse aux faibles doses de RI
• Les défenses antioxydantes• La signalisation des dommages radioinduits• La réparation des dommages radioinduits• La radiosensibilité individuelle• L’effet bystander• La réponse adaptative• L’instabilité génétique• Les systèmes immunologiques de surveillance

Stress quotidien et défense cellulaire
• Stress interneIl est essentiellement dû à la vie à 37°C et au mét abolisme oxydatif qui génère des espèces réactives d’oxygène (ERO)---> anion superoxyde (0 2
•-), peroxyde d’hydrogène (H 2O2) et radical hydroxyl (OH •)
Ces espèces attaquent tous les constituants cellulaires (ADN, protéines) et peuvent provoquer des dysfonctions de la cellule, des effets létaux et mutagènes. ---> maladies neurodégénératives, cardio-vasculaires etc.

Mise en place de la défense cellulaire
Les cellules sont naturellement dotées d’une grande réactivité permettant leur adaptation à l’environnement et leur défense contre les stress interne et externe.
• Découverte récente majeure :La détection et la signalisation des dommages sont cruciales pour la mise en place de la défense cellulaire.

Mécanismes de défense
• Élimination des agents métaboliques toxiques par des antioxydants et des enzymes de détoxification (SOD, catalase etc.)
• Élimination des dommages par 5 systèmes de réparation enzymatiques:réparation de mesappariements de bases, excision de bases, excision de nucléotides, recombinaison homologue, religation non homologue (NHEJ)
• Élimination des cellules endommagées- par apoptose- par nécrose- par des cellules immunocompétentes

Interaction distincte des rayonnements ionisants
L’endommagement de l’ADN des cellules par l’irradiat ion ionisante estconsidéré comme l’événement le plus délétère car ceci peut initierla mutagenèse, l’instabilité génomique et la cancéro genèse.
Les dommages de l’ADN sont induits soitdirectement par un dépôt d’énergie soitindirectement par radiolyse de l’eau et les radicaux libres d’oxygène générés.
1 Gy de rayons ionisants induit:1000 cassures simple brin d’ADN
150 pontages protéine/ADN2500 dommages de bases
40 cassures double brin d’ADN(très délétères)
+ lésions multiples localisées (LMDS)

Dommages Endogènes Radio-induits/cellule/jour/Gy
_______________________________________________________________Cassures simple brin 10 000 à 55 000 1000
Pertes de bases 12 600 ?
Dommages de bases 3 200 2000
Cassures double brin 8 40
Pontages ADN/ADN 8 30
Pontages ADN/protéine quelques-unes 150
Lésions multiples localisées quelques-unes ? 130 ?_______________________________________________________________
Dommages induits dans l’ADN

Dommages multiples localisés radio-induits
• Ces dommages existent mais il est difficile de les identifier au niveau cellulaire et de déterminer leur nocivité (mutagénicité et carcinogénicité).
• Certaines études sur des lésions complexes artificielles suggèrent une certaine mutagénicité, d’autres travaux au niveau cellulaire suggèrent plutôt que la cytotoxicité est prédominante.

dommage de base
cassure simple brincassure double brin
Dommages radio-induits, signalisation et effets
�Signalisation et activation de H2AX
�Induction de gènes
�Arrêt du cycle cellulaire
réparationmort cellulaire
correcteincorrecte
survie
instabilité génétique
carcinogenèse
transformation cellulaire
mutation
Activation de H2AX

CDB radioinduites détectées par l’activation (phosphorylation) de l’histone H2AX
� L’induction de CDB est corrélée avec la formation de foyers de γγγγ-H2AX(Sedelnikova OA et al. Radiat. Res. 2002;158:486-492)
�L’induction de CDB dans des fibroblastes humains prim aires est linéaire à partir de 1,2 mGy
(Rothkamm and Löbrich ,PNAS 2003;100:5057-5062)
Les foyers de γγγγH2AX sur les sites de CDB sont quantifiables par des anticorps spécifiques(marquage par immuno-fluorescence)

Induction et réparation de CDB radioinduites
A: fibroblastes MRC-5 B: fibroblastes 180 BR : déficients dans la réparation (ligase IV)
(Rothkamm et Löbrich, PNAS 2003;100:5057-5062)

Induction et réparation de CDBaprès de faibles doses de rayons X
Absence de réparation à très faible dose (1,2 mGy) !
(Rothkamm et Löbrich, PNAS 2003;100:5057-5062)

Induction de CDB en tomographie chez les patients
(M.Löbrich et al. 2005PNAS; 102:8984-9)

Réparation de CDB Réparation de CDB ((γγγγγγγγ--H2AX)H2AX)dans les lymphocytes humains après CT scandans les lymphocytes humains après CT scan
(M.Löbrich et al. 2005 PNAS; 102:8984-9)

Mise en évidence d’un défaut de réparationchez un patient après une faible dose (tomographie)
HOM-85
(M.Löbrich et al. 2005PNAS; 102:8984-9)

1. Réversion in situ du dommage.
2. Excision de dommagesa) Réparation de mésappariementsb) Excision de bases modifiées• Action spécifique de DNA-N-glycosylasessur un type de lésion (base altérée)
• Réparation de CSB (intervention de PARP1)
3. Excision de nucléotides modifiés
4. Réparation post-réplicative par recombinaison (homologue)
réplication brèche incorporation réparation
ouCSB
PARP1
5. Réparation par recombinaison homologue(échange entre deux brins des deux ADN)
6. Réparation par religation non-homologue(après façonnage, ligation avec perte de matériel. Ligation avec délétion)
CDB
CDB
Stratégies de réparation

Systèmes de réparation de l’ADN (1)
Systèmes de réparation fidèles:Excision de dommages suivant le schéma: reconnaissan ce du
dommage, excision, remplissage de la brèche par une A DN polymérase et ligation
1. Excision des bases incorporées par erreur au cours de la réplication (‘mismatch repair’ ou ‘MMR’)
2. Excision enzymatique spécifique de bases modifié es (oxydées) et réparation des cassures simple brin (brèches) de l’ ADN.Ceci implique un façonnage des cassures simple bri n (CSB) complexes par des endonucléases, la protéine nucléotide kinase PN K et le complexe XRCC1/ ligase III (‘Base excision repair’ ou ‘BER’) . Cette réparation concerne les dommages de bases et les CSB complexes
3. Excision de nucléotides modifiés

Systèmes de réparation de l’ADN (2)concernant les cassures double brin
Système de réparation fidèle :4. Réparation par recombinaison homologue selon le schéma:
détection du dommage (CDB), façonnage des terminais ons des fragments, échange avec un ADN double brin homolog ue, synthèse et religation par une ligase.
Système de réparation non ou peu fidèle :5. Religation non homologue des CDB ( ‘non homo logous
endjoining’ ou ‘NHEJ’) selon le schéma: détection d u dommage (CDB), façonnage, religation par la ligase IV
---> Concerne les CDB et les lésions multiples loca lisées (‘locally multiply damaged sites’ ou ‘LMDS’).

Mutations de gènes affectant la réparation de l’ADN et prédisposant au cancer
BRCA1/2 Carcinome du sein/utérus Facteur de transcription?
MYH Polypose adénomat. APC- Elimination de Ade
en face de OxoGua
FANCC Lymphome ?
NBS Leucémie Endonucléase
BLM Lymphome RecQ hélicase
ATM Leucémie PI-3 kinase, signalisation
XPA-G Cancers de la peau Excision de nucléotides
MSH2, MLH1,PMS Cancer du colon Mésappariement de bases
Gène Cancer FonctionP53 Plusieurs types de cancer Facteur de transcription

Sensibilité individuelle et polymorphismes ou mutat ions de gènes de réparation
La sensibilité individuelle est rare et n’est pas détectable dans les études de populations (études épidémiologiques)
•Des études récentes sur l’induction de micronoyaux par de faibles doses de rayons ionisants (Angelini S et al. Mutat Res.2005) suggèrent que des polymorphismes de gènes de réparation (tels que XRCC3, XRCC1 et XPD) contribuent aux dommages génétiques chez des individus chroniquement exposés.•De plus, des différences dans la capacité de réparation ont été mises en évidence chez des patients, en radiothérapie et radiodiagnostic (tomographie).

Induction de familles de gènes différentes à de faibl eset à de hautes doses
(selon Franco N et al. Radiat. Res. 163, 2005)
A 1 cGy sont induits les gènes impliqués dans l’homéostasie, le stress,la signalisation cellulaire, le cytosquelette, la synthèse d’ARN,les fonctions membranaires, mais pas encore les gènes de réparation de l’ADN

Induction de phosphoprotéines différentes dans les fibroblastes humains après faible et haute doses
de rayons X ( Yang F et al. J Proteome Res. 2006;5:1252-1260)
A haute dose (4 Gy) ð phosphorylation de protéines impliquées dans les voies de la signalisation et de l’apoptose.A faible dose (2 mGy) ð phosphorylation de protéines impliquées dans le métabolisme plus général

Effets nouveaux
Notion ancienne : Focalisation sur les dommages radio-induits au niveau de
l’ADN (patrimoine génétique) précurseurs de l’induction de cancers
Notion récente : La cible n’est pas seulement l’ADN: Les dommages radio-
induits dans l’ADN et également dans le cytoplasme et les membranes sont reconnus et donnent lieu aussi à une signalisation intra-cellulaire qui détermine les conséquences biologiques
De plus, il existe une signalisation inter-cellulairedéclenchée par l’irradiation et affectant les cellules voisines
--->effet de voisinage ou effet « bystander ».

Paradigme conventionnel: Induction ciblée de dommages dans les cellules irradiées et transmission
des dommages aux cellules filles
Celluleinitiée
MutationsDommages
chromosomiques
Instabilité génomique

Existence d’effets non ciblés et retardés aux faibles doses de
rayonnements ionisants
• Effet “bystander” ou effet de voisinage:observé dans les cellules non directement touchées ounon traversées par le rayonnement et impliquantl’émission de signaux par les cellules irradiées
• Instabilité génomique radioinduite :observée dans la progénie des cellules irradiées qui ontété ou n’ont pas été directement touchées par un dépôtd’énergie du rayonnement
Volume endommagé > volume irradié

Effet de voisinage (“bystander”) radio-induit:
irradiation des cellules par microfaisceaux
Quelques cellules sont directement irradiées, mais les cellules voisines sont également affectées
Cellule irradiée Particule αFaible dose

Communicationintercellulaire
Facteurs solublesMilieu
Particule alpha
Cellule avec noyau
Espèces réactives de
l’oxygène
TNF alpha, IL1 .?
Cellule fortement léséeCellule lésée Cellule lésée
ADN
Mort cellulaire ou survieavec mutations par délétion
Survie cellulaireavec mutations ponctuelles
Survie cellulaireavec mutations ponctuelles
Effet bystander
Effets « bystander » dans les cellules---> Expériences avec microfaisceaux
de particules alpha
ADN

Effet « bystander » sur des cellules non irradiées:Induction d’effets génotoxiques par l’intermédiaire de
cellules exposées à de faibles doses d’irradiation ionisante.
Augmentation des taux deSCE, micronoyaux, mutations ponctuelles,effets clastogèneseffets létaux certains facteurs comme TGFbêta.
L’induction a été inhibée par la SOD et la catalaseainsi que par d’autres agents tels que le lindane etc..
(Azzam EI et al. Oncogène 22,7050-7057 (2003))

Dose dépendance : Dose de RI augmente ---> pas d’augmentation de l’effet
Nature du signal dans l’effet “bystander”---> Espèces réactives d’oxygène/d’azote; signalisation par cytokines, réponses inflammatoires---> pas toutes les cellules produisent ces facteurs
Effets sur les cellules voisines:---> Induction de cassures double brin dans l’ADN(γH2AX), d’aberrations chromosomiques, de micronoyaux et d’apoptose----> l’effet n’est pas observé dans tous les types cellulaires
Effet « bystander »: Résultats in vitro

Effet « bystander »: Résultats in vivo (1)
Effets “bystander”dans la peauhumaine(modèle 3D)
Belyakov et al. PNAS 102, 14203-7 (2005)
Un effet « bystander » peut être également observé in vivo(voir aussi effets clastogènes et abscopaux).

Effet « bystander »: Résultats in vivo (2)
Effets “bystander” dans un modèle 3D de peau humaine(Belyakov et al. PNAS 102, 14203-7 (2005)

L’irradiation de cellules normales par de faibles d oses conduit à l’élimination par apoptose de cellules
précancéreuses grâce à l’effet « bystander »(D.I. Portess et al. Cancer Res. 2007; 67(3):1246--1253)
---> L’irradiation gamma de cellules non transformée s c208F conduit à l’apoptose des cellules transformées 208F src3 cocultivées.

• La signalisation (TGFbêta) est induite par de faibles doses de RI ce quiélimine les cellules précancéreuses par apoptose.
• Ceci peut être considéré comme un effet d’hormesis résultant enune réduction de la transformation cellulaire à de faibles dosesd’irradiation ionisante (Redpath et al. Radiat. Res 2003,2006; Azzam EI et al. Radiat. Res. 1996).
• Il s’agit peut-être d’un mécanisme naturel anticancéreux déclenchépar de faibles doses d’irradiation.
--->Ces effets ne sont pas compatibles avec une lin éarité sans seuil de la réponse aux radiations.
Elimination de cellules transformées par des cellules normales irradiées par de faibles doses de RI
(D.I. Portess et al. Cancer Res. 2007; 67(3):1246--1253)

Développement de tumeurs du cerveau (Médulloblastome)après irradiation X totale (WB) ou partielle (SH) dans les souris néonatales Ptch1 +/- et
souris témoins (CN)
0 10 20 30 40
0
20
40
60
80
100
WB 3 Gy (37) SH 3 Gy (46)
CN (51)
80 100
WB 0.036 Gy (34)
Me
du
llob
last
om
a f
ree
mic
e (
%)
--�Effet bystander après 3 Gy de rayons X: induction detumeurs du cerveau dans les souris prédisposées au cancer.
(Manusco M. et al. PNAS 2008;105:12445-12450)

Effet « bystander »: Résultats in vivo (3)
Moelle40XY T6T6
Moelle40XY (neutrons)
Instabilité chromosomiquedans la progénie de cellules soucheshématopoïétiques(Watson et al., Cancer Res.60, 5608 - 5611 (2000))
LS174T
LS174T(125I)
Inhibition de la croissancede tumeur(Xue et al., PNAS 99, 13765-70 (2002))
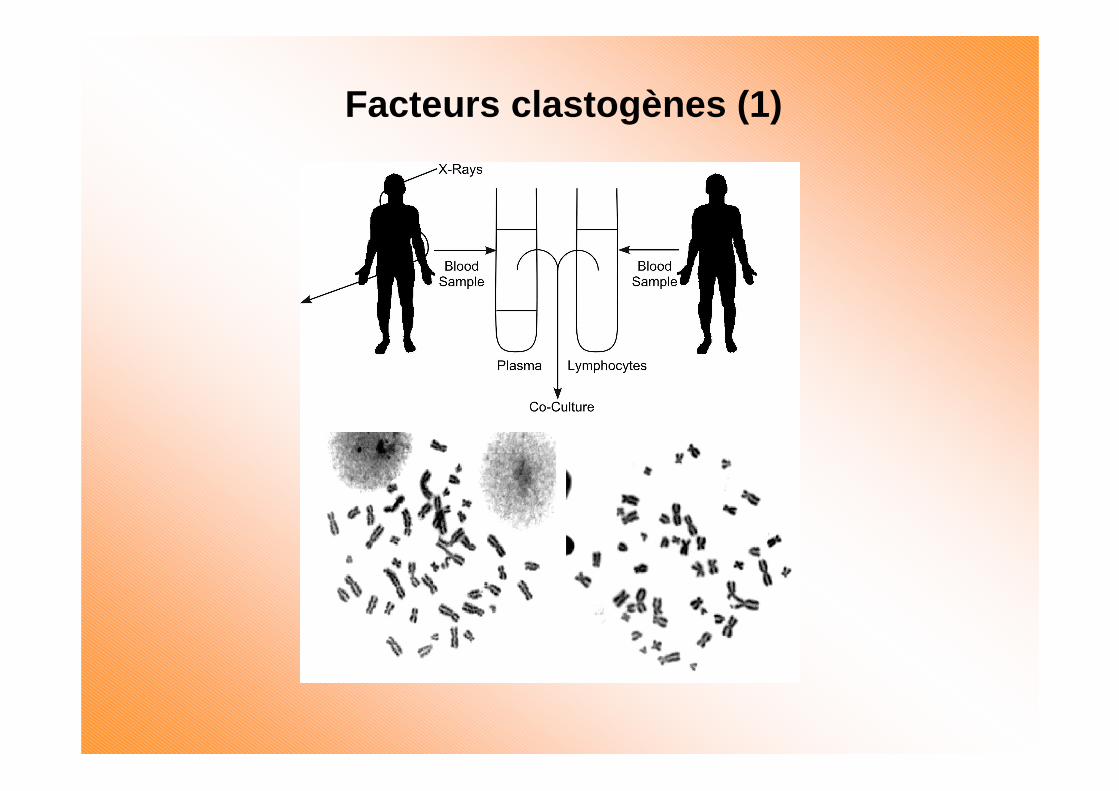
Facteurs clastogènes (1)

Instabilité génomique radio-induite
Le taux d’altérations génomiques est augmenté dans la progénie des cellules irradiées
Rayonnement
Se manifeste par:•Réarrangements chromosomiques•Formation de micronoyaux•Aneuploïdie•Apparition de mutations tardives
(avec spectre différent)•Amplification de gènes•Mort cellulaire

Lignée de hamster portant un chromosome humain util iséepour la détection de l’instabilité génomique

Détection de l’instabilité génomique dans une lignée de hamster avec chr. IV humain
Translocationsdu chr. IV

Effet abscopal chez l’homme (?)Ohba et al. Gut 43, 575-577, (1998)
Chez un homme de 76 ans souffrant de douleurs thoracique et abdominaleun CT scan montre des métastases dans l’os d’une vertèbre et des hépatocarcinomes. Une dose de 36 Gy sur l’os provoque la régression des lésions hépatiques.
Analyses rétrospectives:les concentrations des facteurs IL-1 beta, IL-2, IL-4, IL-6, HGF, etTNF-alpha dans le serum sont augmentées.

Effets transgénérationnels transmis par la lignée germinale des parents irradiés à la progéniture:
Analyses mutationnelles de deux loci de mini-satellites chez la souris( Y. Dubrova, Université de Leicester)
A: Loci Msh6-hm: P1:père; P2:progénie normale; P3: progénie avec mutation maternelle; P 4 + 5: progénie avec 2 mutations paternelles; P6: mèreB) Loci HM-2: P 1: père; P2, 4,6 t 7: progénie normale; P 3,5: progénie avec des mutations paternelles; P8: mère
---->Signification pas claire car absence de phénotype

La réponse radio-adaptative(Rigaud and Moustacchi, Mutat Res.1996; 435(2):127-34)
• Une pré-exposition à une faible dose d’irradiation ionisante (20 mGy) (à faible débit de dose) donne lieu à une résistance des cellules et des tissus à une forte dose d’exposition (quelques heures après la première dose).
• Des réponses adaptatives ont été observées pour l’induction de dommages de l’ADN, de mutations, d’aberrations chromosomiques, de micronoyaux et de transformation cellulaire in vitro et in vivo .

0
10
20
30
40
50
60
70
80
90
0 5 10 1500 E O E O
Réponse adaptative aux RIA
berr
atio
ns c
hrom
atid
ique
s
Dose (mGy)55 1010
Adapt (5 ou 10mGy) Forte dose (1,5 Gy)Analyse
chromosomique6 hr6 h

Réponse adaptative: une faible dose réduit le niveau de transformation cellulaire spontanée.
Transformation in vitropar irradiation X ( 60kVp)
Comparaison entretransformation in vitro par irradiation X (60 kVp) ouirradiation gamma (Cs-137)
Redpath et al., I.J.R.B. 79, 235-40 (2003)

Conclusions (1)• Les effets de faible dose et faible débit de dose de RI ont perdu leur
mystère: Il est désormais possible d’explorer directement les effets biologiques de doses de l’ordre du mGy et de débits de dose de l’ordre du mGy/min et de déceler les mécanismes impliqués à différents niveaux d’exposition.
• Les nouvelles données sont convergentes: Les cellules et les tissus ne réagissent pas de la même façon à haute et faible doses de RI.
• Ces observations ne confortent donc pas une extrapolat ion directe et facile des hautes vers les très faibles do ses. Toutefois, elles méritent d’être encore consolidées et modélisées.
• A de faibles doses d’irradiation ionisante plusieurs effets non ciblés ont été mis en évidence . Les phénomènes de radioadaptation et de voisinage (bystander) semblent jouer un rôle protecteur mais l’instabilité génétique pourrait au ssi jouerun rôle important dans la cancérogenèse radioinduite.

Conclusion (2)
• Les facteurs clastogènes sont souvent un signe de stress cellulaire déjà établi, par exemple dans certains syndromes humains génétiquement définis et caractérisés par des chromosomes cassés, (syndrome de Bloom, Fanconi etc..). Toutefois, ces facteurs nécessitent d’être mieux caractérisés avant qu’on puisse démontrer leur implication dans les processus de la radiocancérogenèse.
• Les effets transgénérationnels sont des effets transmissibles de parents irradiés à la progéniture plusieurs générations après exposition. Des effets intéressants ont été rapportés comme l’apparition de mutations dans des séquences dites mini-satellites transmises par la lignée germinale mais leur signification n’est pas encore très claire car aucun phénotype ne s’y trouve associé.

Merci pour votre attention !