insights into the origin and evolution of the plant hormone … · plant hormones modulate plant...
TRANSCRIPT

Insights into the Origin and Evolution of the PlantHormone Signaling Machinery1
Chunyang Wang2, Yang Liu, Si-Shen Li*, and Guan-Zhu Han2*
Jiangsu Key Laboratory for Microbes and Functional Genomics, Jiangsu Engineering and Technology ResearchCenter for Microbiology, College of Life Sciences, Nanjing Normal University, Nanjing, Jiangsu 210023, China(C.W., G.-Z.H.); State Key Laboratory of Crop Biology, Shandong Agricultural University, Tai’an, Shandong271018, China (C.W., Y.L., S.-S.L.); and Department of Ecology and Evolutionary Biology, University ofArizona, Tucson, Arizona 85721 (G.-Z.H.)
Plant hormones modulate plant growth, development, and defense. However, many aspects of the origin and evolution of planthormone signaling pathways remain obscure. Here, we use a comparative genomic and phylogenetic approach to investigate theorigin and evolution of nine major plant hormone (abscisic acid, auxin, brassinosteroid, cytokinin, ethylene, gibberellin,jasmonate, salicylic acid, and strigolactone) signaling pathways. Our multispecies genome-wide analysis reveals that: (1) auxin,cytokinin, and strigolactone signaling pathways originated in charophyte lineages; (2) abscisic acid, jasmonate, and salicylic acidsignaling pathways arose in the last common ancestor of land plants; (3) gibberellin signaling evolved after the divergence ofbryophytes from land plants; (4) the canonical brassinosteroid signaling originated before the emergence of angiosperms butlikely after the split of gymnosperms and angiosperms; and (5) the origin of the canonical ethylene signaling pathway postdatesshortly the emergence of angiosperms. Our findings might have important implications in understanding the molecularmechanisms underlying the emergence of land plants.
Hormones play a central role in regulating plantgrowth, development, and defense. To date, a total of 10structurally unrelated small molecules (abscisic acid[ABA], auxin [AUX], brassinosteroids [BRs], cytokinins[CKs], ethylene [ETH], GAs, jasmonates [JAs], nitricoxide, salicylic acid [SA], and strigolactones [SLs]) havebeen recognized as plant hormones (Santner andEstelle, 2009). Many aspects of plant hormone signal-ing biology, such as transport and signal transduction,have been well characterized (Santner and Estelle, 2009,and refs. therein; for a schematic overview, see Fig. 1).Remarkably, as the intermediates between hormonesand target genes, the receptors of nine plant hormones(except nitric oxide) have been identified (Chang et al.,1993; Inoue et al., 2001; Wang et al., 2001; Dharmasiri
et al., 2005; Murase et al., 2008; Fujii et al., 2009; Sheardet al., 2010; Fu et al., 2012; Zhou et al., 2013).
ABA plays an important role in the responses ofplants to environmental stresses, especially drought(Fujii et al., 2009). The PYRABACTIN RESISTANCE(PYR)/PYRABACTIN RESISTANCE-LIKE (PYL)/REGULATORY COMPONENT OF ABSCISIC ACIDRECEPTOR (RCAR) family of START proteins (PYRsfor short) has been identified as ABA receptors (Fujiiet al., 2009). In the absence of ABA, the positive regu-lator SUBCLASS III SUCROSE NONFERMENTING1-RELATED PROTEIN KINASE2 (SnRK2) is inactivatedby groupA PROTEIN PHOSPHATASE2C (PP2C) throughdephosphorylation. The binding of ABA to receptorsbreaks the interaction of groupA PP2C and SnRK2, whichactivates the function of SnRK2 by autophosphorylation.The activated SnRK2 phosphorylates the ABA-responsivetranscription factors, such as ABSCISIC ACID-RESPONSIVEELEMENT-BINDING FACTORs (ABFs) and ABSCISICACID-INSENSITIVE5 (ABI5). Moreover, ABSCISIC ACID-INSENSITIVE5-BINDING PROTEINs (AFPs) facilitate thedegradation of ABI5 (Kelley and Estelle, 2012). There arethree different types of ABA transporters, ABSCISICACID-IMPORTING TRANSPORTER (AIT), ATP-BINDINGCASSETTE G25 (ABCG25), and PLEIOTROPIC DRUGRESISTENCE (PDR; Boursiac et al., 2013).
AUX plays a central role in plant growth and de-velopment by affecting cell expansion and division(Dharmasiri et al., 2005). The AUX receptors TRANSPORTINHIBITOR RESPONSE1 (TIR1) and AUXIN-SIGNALINGF-BOX PROTEIN (AFB) perceive AUX in cells (Dharmasiriet al., 2005). When the AUX level is low, an endoplasmic
1 This work was supported by the National Key TechnologiesR&D Program of China (grant no. 2011BAD35B03) and the PriorityAcademic Program Development of Jiangsu Higher Education Insti-tutions.
2 These authors contributed equally to the article.* Address correspondence to [email protected] and guanzhu@
email.arizona.edu.C.W. and G.-Z.H. performed most of the experiments; Y.L. pro-
vided technical assistance; G.-Z.H. designed the experiments and an-alyzed the data; G.-Z.H. and S.-S.L. conceived the project; G.-Z.H.wrote the article with input from C.W.; S.-S.L. supervised the projectand complemented the writing.
The author responsible for distribution of materials integral to thefindings presented in this article in accordance with the policy de-scribed in the Instructions for Authors (www.plantphysiol.org) is:Guan-Zhu Han ([email protected]).
www.plantphysiol.org/cgi/doi/10.1104/pp.114.247403
872 Plant Physiology�, March 2015, Vol. 167, pp. 872–886, www.plantphysiol.org � 2014 American Society of Plant Biologists. All Rights Reserved.

reticulum-localized AUXIN-BINDING PROTEIN1 (ABP1)increases the stability of AUXIN/INDOLE-3-ACETICACIDs (AUX/IAAs), possibly by inhibiting AUX/IAAs
recruitment to the S-PHASE KINASE-ASSOCIATEDPROTEIN1-CULLIN-F-BOX (SCFTIR1/AFB) E3 ligase complex(Tromas et al., 2013); however, the detailed mechanism
Figure 1. Schematic diagrams of nine major plant hormone signaling pathways: AUX signaling (A), JA signaling (B), GA sig-naling (C), SL signaling (D), CK signaling (E), BR signaling (F), ETH signaling (G), ABA signaling (H), and SA signaling (I). Fordetailed molecular mechanisms, see the introduction. ER, Endoplasmic reticulum; NM, nuclear membrane.
Plant Physiol. Vol. 167, 2015 873
Origin of Plant Hormone Signaling Pathways

remains unclear. Then, AUX/IAAs and TOPLESS (TPL)repress the activity of POSITIVE AUXIN RESPONSEFACTORs (ARFs+) and NEGATIVE AUXIN RESPONSEFACTORs (ARFs2). When the AUX level is high, AUXincreases the affinity of TIR1/AFBs with AUX/IAAs,which leads to the degradation of AUX/IAAs by theubiquitin-proteasome pathway via the SCFTIR1/AFB E3 li-gase complex and the release of ARFs+/ARFs2. Therelease of ARFs+/ARFs2 activates or represses the ex-pression of AUX target genes and the hormone response(Guilfoyle and Hagen, 2007). The PIN-FORMED FAMILYproteins (PINs) function as AUX transporters and play akey role in generating the AUX gradients (Mravec et al.,2009). The AUXIN1/LIKE AUXIN1 genes encode a high-affinity AUX influx carrier (Péret et al., 2012). GRETCHENHAGEN3 (GH3) can conjugate AUX to amino acids,maintaining the level of AUX (Mashiguchi et al., 2011).
BRs are the plant-specific steroid hormones that reg-ulate a wide range of developmental and physiologicalprocesses of plants, such as cell elongation, photomor-phogenesis, seed germination, flowering, male fertility,and senescence (Wang et al., 2012). BRASSINOSTEROIDINSENSITIVE1 (BRI1) and BRASSINOSTEROIDINSENSITIVE1-ASSOCIATED RECEPTOR KINASE1(BAK1) act as BR receptors (Wang et al., 2001). When the BRlevel is low, the kinase domain of BRI1 interacts withBRASSINOSTEROID INSENSITIVE1 KINASE INHIBITOR(BKI1), BRASSINOSTEROID-SIGNALING KINASE(BSK), and CONSTITUTIVE DIFFERENTIAL GROWTH1(CDG1). The transcription factors BRASSINAZOLE-RESISTANT1 (BZR1) and BZR2 are phosphorylatedby BRASSINOSTEROID-INSENSITIVE2 (BIN2) andthen retained by the 14-3-3 (GENERAL REGULATORYFACTOR [GRF]) proteins. When the BR level is high,BRs bind BRI1, triggering the disassociation with BKI1and the association with coreceptor BAK1 and thenactivating BRI1. The activated BRI1 phosphorylatesBSKs and CDG1, which sequentially phosphorylatesBRASSINOSTEROID INSENSITIVE1 SUPPRESSOR1(BSU1). BSU1 dephosphorylates and inactivates BIN2.PP2A dephosphorylates BZRs, which results in the re-lease of BZRs from the GRF proteins. BZRs then bindand regulate the BR-responsive genes.
CKs influence many aspects of plant growth anddevelopment, such as cell division, chloroplast de-velopment, leaf senescence, and root and shoot growthand branching (Schaller et al., 2011). Similar to the bac-terial two-component regulatory system (TCS), the CKsignaling pathway contains multistep phosphorylation(Schaller et al., 2011). The CK receptors ARABIDOPSISHISTIDINE KINASE2 (AHK2), AHK3, and AHK4autophosphorylate themselves when they bind CKsand then phosphorylate ARABIDOPSIS HISTIDINEPHOSPHOTRANSFER PROTEINs (AHPs). AHPs carryphosphate groups into the nucleus, where they phos-phorylate the transcription factors B-type RESPONSEREGULATORs that positively regulate the CK responses.A-type RESPONSE REGULATORs are the negative reg-ulators of the CK responses and contend for phosphategroups with B-type RESPONSE REGULATORs (Gruhn
and Heyl, 2013). CYTOKININ RESPONSES FACTORspositively regulate the transcription response to CKs. CKsare transported by ABCG14 in the long distance fromroot to shoot (Zhang et al., 2014).
ETH regulates both development and defense pro-cesses of plants, such as fruit ripening, senescence, ab-scission, and the responses to biotic and abiotic stresses(Qiao et al., 2012). ETH is perceived by its receptorETHYLENE RESPONSE1 (Chang et al., 1993). In theabsence of ETH, the receptors promote the kinase activityof CONSTITUTIVE TRIPLE1 (CTR1). CTR1 phosphory-lates the positive regulator ETHYLENE INSENSITIVE2(EIN2), repressing the ETH responses. EIN3 is de-graded via the SCF E3 ligase complex with ETHYLENEINSENSITIVE3-BINDING F-BOX PROTEIN1/2 (EBFs),whereas EIN2 is degraded via the SCF complex withETHYLENE INSENSITIVE2-TARGETING PROTEIN1/2 (ETP1/2). ETH promotes the accumulation of EIN2 bydown-regulating ETP1/2. In the presence of ETH, thereceptors and CTR1 are inactivated, triggering the de-phosphorylation and cleavage of EIN2. The C-terminaldomain of EIN2 is translocated to the nucleus, where itstabilizes EIN3. EIN3 triggers the ETH responses by bind-ing to ETHYLENE RESPONSE FACTOR1/2 (Kendrickand Chang, 2008). MITOGEN-ACTIVATED PROTEINKINASE KINASE KINASE (MAP3K) is a negative regu-lator in the ethylene response (Kumar and Sharma, 2014).
GAs regulate the germination, elongation growth,and sex determination of plants (Murase et al., 2008).When GAs are absent, their receptor GIBBERELLIN-INSENSITIVE DWARF1 (GID1) is in the passive stateand the repressor DELLA-CONTAINED PROTEIN(DELLA) inhibits the activity of the transcription factorPHYTOCHROME-INTERACTING FACTOR (PIF) andthus represses the GA responses. The bioactive GAscan change the conformation of their receptor GID1,which increases the affinity between GID1 and DELLAand forms the GA-GID1-DELLA complex (Muraseet al., 2008; Shimada et al., 2008). The complex in-creases the interaction between DELLA and SLEEPY1/2 (SLY1/2). Consequently, DELLA is degraded by theubiquitin-proteasome pathway via the SCFSLY1/2 com-plex. The degradation of DELLA releases PIFs, per-mitting the GA responses (Santner and Estelle, 2009).SPINDLY negatively regulates the GA responses,potentially by stabilizing the DELLA protein (Sun,2008).
JAs control plant defense against wounding, herbivores,and certain pathogens and are also crucial for plant fertilityand reproduction (Sheard et al., 2010). JASMONATERESISTANT1 (JAR1; also known as GH3-11) conjugatesJAs into biologically active status. The active JAs are per-ceived by their receptor CORONATINE INSENSITIVE1(COI1; Sheard et al., 2010). In the absence of active JAs,JASMONATE ZIM-DOMAIN PROTEINs (JAZs), NOVELINTERACTOR OF JASMONATE ZIM-DOMAINPROTEIN (NINJA), and TPL work together to bind thetranscription factorMYC-RELATED TRANSCRIPTIONALACTIVATOR (MYCs) and inhibit their function (Pauwelset al., 2010). Binding the active JAs, COI1 interact with
874 Plant Physiol. Vol. 167, 2015
Wang et al.

JAZs, which leads to the degradation of JAZs by theSCFCOI1 complex and the release of MYCs (Pauwelset al., 2010). Consequently, the JA-responsive genes aretranscribed by MYCs.SA regulates the immunity of plants and induces
systemic acquired resistance when plants are facingpathogen challenge (Fu et al., 2012). The SA responsemechanism was characterized in the model plantArabidopsis (Arabidopsis thaliana) recently (Fu et al., 2012).NONEXPRESSER OF PATHOGENESIS-RELATEDGENE1 (NPR1) is a master regulator of systemic ac-quired resistance. NPR3 and NPR4 have been identifiedas the SA receptors (Fu et al., 2012). They perceive SAwith different affinities: NPR4 has a higher affinity toSA than NPR3. When the SA concentration is low,NPR4 binds SA instead of NPR3, which in turn inhibitsthe degradation of NPR1 and allows the basal resis-tance. The pathogen challenge increases the SA level,forming an SA concentration gradient (Fu et al., 2012).In the infection sites with the highest SA level, SA bindsto NPR3, which promotes NPR1 degradation to allowprogrammed cell death. In the cells with lower SA level,NPR1 accumulates and activates the transcription factors,such as WRKYs and TGACG SEQUENCE-SPECIFICBINDING PROTEINs (TGAs), to turn on the defense-related genes (Ülker and Somssich, 2004; Fu et al.,2012).SLs, a newly identified class of plant hormones, in-
hibit plant shoot branching and modulate seed ger-mination and seedling photomorphogenesis (Zhouet al., 2013). The protein DWARF14 (D14) perceivesSLs, which can interact with D3 in rice (Oryza sativa;MOREAXILLARY BRANCHES2 [MAX2] in Arabidopsis).When the SL level is low, D53 in rice (SUPPRESSOR OFMAX2-1-LIKE PROTEINS [SMXLs] in Arabidopsis) mayinteract with TPL and repress the TCP proteins. TCPtranscription factor (FINE CULM1 in rice, TB1 in maize[Zea mays], and BRANCHED1 [BRC1] in Arabidopsis),thus inhibiting the SL-responsive genes (de Saint Germainet al., 2013; Jiang et al., 2013). When the SL level increases,SLs promote the interaction of D3-D14 and lead to thedegradation of D53 by the ubiquitin-proteasome system.The TCP transcription factor is then released, triggering thedownstream response (Zhou et al., 2013).The previous evolutionary and comparative ge-
nomic analyses suggest that AUX and SL signalingpathways were, at least in part, already present beforethe emergence of land plants; that ABA and CK sig-naling pathways emerged while plants were coloniz-ing land; and that GA, JA, ETH, and BR signalingpathways evolved after the evolutionary split of mossand vascular plants (Rensing et al., 2008; Pils and Heyl,2009; Banks et al., 2011; Hauser et al., 2011; Boot et al.,2012; Delaux et al., 2012; Thaler et al., 2012; Viaeneet al., 2013). However, many details of the origin andevolutionary mode of the plant hormone signalingmachinery remain obscure and controversial.The accumulating plant genome-scale data and the
understanding of plant hormone signaling biologyoffer a great opportunity to investigate the origin and
evolution of the plant hormone signaling machinery.We focus on nine major plant hormone signalingpathways (i.e. ABA, AUX, BR, CK, ETH, GA, JA, SA,and SL signaling pathways) because these signalingpathways have been well characterized (Santner andEstelle, 2009; Fig. 1). We performed multispecies com-parative genomic and phylogenetic analyses of the im-portant components of each plant hormone signalingpathway using genome-scale data of plant species thatrepresent all the major plant lineages (SupplementalTable S1). Our study provides a comprehensive view ofthe origin and evolutionary mechanisms of these majorplant hormone signaling pathways.
RESULTS
Plant Species and Ortholog Identification
Our data set (Supplemental Table S1) includes thecomplete genome sequences of nine species that rep-resent all the major plant lineages (Chlamydomonasreinhardtii, Volvox carteri, Ostreococcus tauri, Cyanidioschyzonmerolae, Klebsormidium flaccidum, Physcomitrella patens,Selaginella moellendorffii, Arabidopsis, and rice). We alsoused the transcriptome data of one liverwort (Marchantiapolymorpha; Sharma et al., 2013) and four charophytes(Nitella hyalina, Nitella mirabilis, Penium margaritaceum,and Spiragyra pratensis; Timme et al., 2012). Charophytesare a group of freshwater green algae from which landplants were descended (Qiu et al., 2006; Finet et al.,2010). The previous studies of plant hormone signalingevolution (Rensing et al., 2008; Pils and Heyl, 2009; DeSmet et al., 2011) used P. patens as the most basal lineageof land plants. Here, we used the liverwortM. polymorphaas the most basal plant lineage, as liverworts are the sistergroup of all the other land plants (including P. patens) andrepresent the earliest branching land plant lineage (Qiuet al., 2006).
To determine the origin of plant hormone signalingmachinery, we identified the orthologs of plant hor-mone signaling components in the 14 plant species(Supplemental Table S1), because orthologs are morelikely to retain similar functions in the course of evo-lution (Tatusov et al., 1997; Chen and Zhang, 2012).Homologs are genes/proteins that share a certain de-gree of similarity with each other, whereas orthologsare genes/proteins of different species that originatedfrom a single gene of the last common ancestor (Chenand Zhang, 2012). Homologs do not necessarily meanthey are orthologs, because homologs might share se-quence similarity only in certain regions (e.g. conserveddomains). Thus, high similarity between the proteinsof different species does not guarantee that they areorthologs and/or share similar functions (Tatusov et al.,1997; Chervitz et al., 1998). Here, we employed acombination of similarity search, phylogenetic recon-struction, and conserved domain analysis approaches(see “Materials and Methods”) to systemically identifythe orthologs of plant hormone signaling components.
Plant Physiol. Vol. 167, 2015 875
Origin of Plant Hormone Signaling Pathways
First, we used the BLAST algorithmswith the Arabidopsis(occasionally P. patens) proteins as queries to identify theirhomologs in the genomes of the other plant species. Next,we performed phylogenetic analysis to identify potential
orthologs. Third, because many protein domains have adegree of evolutionary independence and can combinewith each other in different forms (Chervitz et al., 1998),we compared their conserved domain architecture with
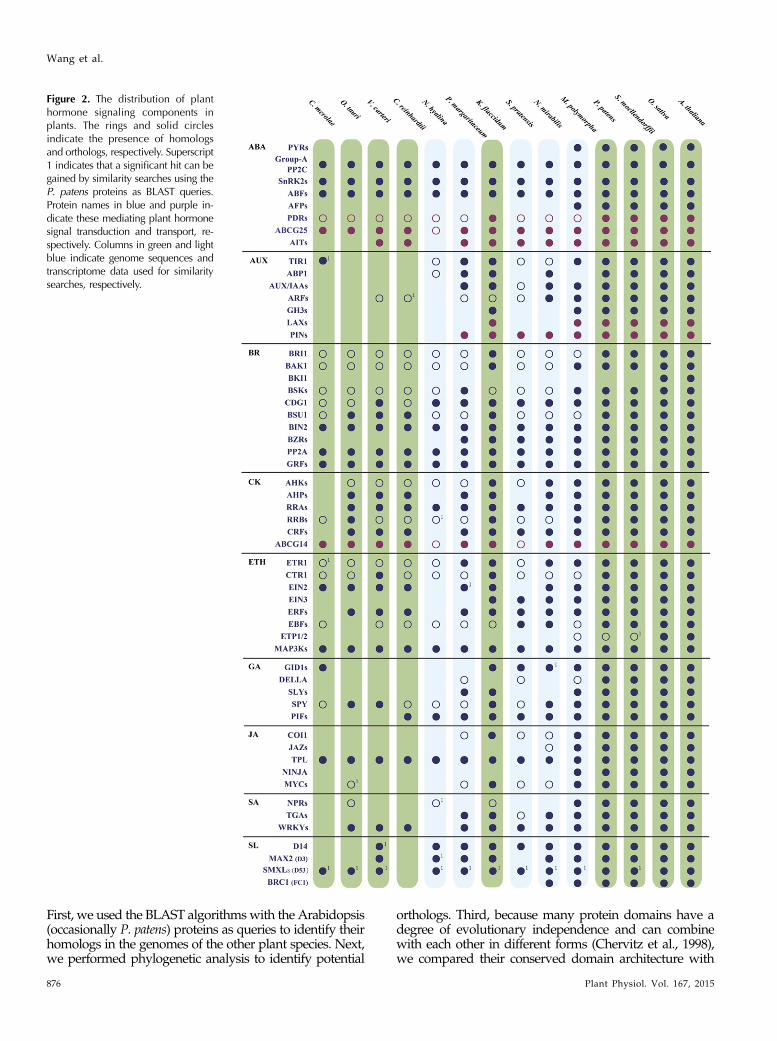
Figure 2. The distribution of planthormone signaling components inplants. The rings and solid circlesindicate the presence of homologsand orthologs, respectively. Superscript1 indicates that a significant hit can begained by similarity searches using theP. patens proteins as BLAST queries.Protein names in blue and purple in-dicate these mediating plant hormonesignal transduction and transport, re-spectively. Columns in green and lightblue indicate genome sequences andtranscriptome data used for similaritysearches, respectively.
876 Plant Physiol. Vol. 167, 2015
Wang et al.

that of Arabidopsis proteins. Only the potential orthologscontaining the same conserved domain architecture as theArabidopsis proteins are defined as orthologs in thisstudy. The orthologs identified using this approach arelikely to have similar functionality, given that homologssharing the same domain architecture tend to have similarfunctions (Hegyi and Gerstein, 2001). Because gene ex-pression is usually tissue specific, developmental stagespecific, and/or stress induced, transcriptome data (es-pecially when its coverage is low) sometimes cannot re-cover all the gene information of an organism’s genome(Wang et al., 2014). Therefore, regarding the ortholog iden-tification with transcriptome data of M. polymorphaand four charophytes (N. hyalina, N. mirabilis, P.margaritaceum, and S. pratensis), the presence of ortho-logs indicates the presence of orthologs but the absenceof orthologs does not necessarily indicate the absence oforthologs (Wang et al., 2014).To better understand the molecular mechanisms,
we also predicted the protein structures of the ortho-logs of eight hormone (ABA, AUX, BR, CK, ETH,GA, JA, and SL) receptors using a homology-modelingapproach based on their available experimental three-dimensional structures (Müller-Dieckmann et al., 1999;Hayashi et al., 2008; Murase et al., 2008; Nishimuraet al., 2009; Sheard et al., 2010; Hothorn et al., 2011a,2011b; Zhao et al., 2013). The similarity betweenpredicted protein structure and the experimentalstructure is evaluated by the template modeling (TM)score. A TM score of greater than 0.17 indicates thatthe protein structures compared are similar and dis-tinguishable from random structure pairs (Xu and Zhang,2010).
ABA Signaling Originated in the Last Common Ancestorof Land Plants
Our comparative analyses identified the orthologs ofall the core components mediating the ABA response(ABFs, AFPs, group A PP2C, PYRs, and SnRK2s) andtwo ABA transporters (ABCG25 and AITs) in the landplant genomes (Fig. 2). Algal genomes encode most ofthe ABA response and transport components, includ-ing ABFs, group A PP2C, SnRK2s, PDRs, ABCG25,and AITs. Because PDRs can be identified in the ge-nome of K. flaccidum, their absence in other charophytesand M. polymorpha is possibly due to the short nature ofthe assembled transcriptome sequences. The orthologsof AFPs and the ABA receptor PYR proteins wereidentified only in the land plant genomes, suggestingthat both proteins first occurred in land plants. Inter-estingly, the predicted structures of the P. patens andM. polymorpha ABA receptor orthologs share signif-icant similarity with the experimental structure ofthe Arabidopsis protein (TM score = 0.307 and 0.333,respectively; Supplemental Fig. S1). The binding sites ofthe land plant ABA receptor orthologs are well con-served (Supplemental Fig. S2). These results suggestthat the ABA signaling machinery originated at least inthe last common ancestor of land plants (Fig. 3). Consistentwith our hypothesis, the group A PP2C-mediated ABAresponse mechanism appears to function inM. polymorpha(Tougane et al., 2010).
AUX Signaling Might Originate in Charophytes
The previous analysis of the moss (P. patens) genomesuggests that the AUX signaling machinery originatedbefore the divergence of moss and vascular plants(Rensing et al., 2008). Further analyses of the charophyteEST library identified several, but not all, components(including ARFs, AUX/IAAs, and PINs) of the AUXsignaling and transport machinery, which leads to thehypothesis that the canonical AUX signaling was, at leastin part, already present in the charophyte genomes (Lauet al., 2009; De Smet et al., 2011; Viaene et al., 2013). Ourcomparative genomic analyses show that the genomes ofK. flaccidum and land plants encode the orthologs of allthe core components (except ABP1 in M. polymorpha andARFs in K. flaccidum) mediating the AUX response andtransport (Fig. 2). The ABP1 protein ortholog was notfound in the transcriptome assembly of M. polymorpha,but it can be found in several charophytes (N. mirabilis, P.margaritaceum, and K. flaccidum). This distribution patternmight be due to either the specific loss of ABP1 in thelineage ofM. polymorpha or, more likely, the low coverageor short nature of M. polymorpha transcriptome data. Asfor the ARF protein, the ARF ortholog was not identifiedin the K. flaccidum genome but was found in the genomeof N. mirabilis (Fig. 2; Supplemental Fig. S1). Interestingly,the AUX/IAA ortholog of K. flaccidum contained all thetypical conserved domains (I, II, III, and IV) of AUX/IAAproteins identified in previous studies (Supplemental Fig.S2); typical domains I, II, III, and IV act as transcriptional
Figure 3. The origin of nine major plant hormone signaling path-ways in plants. The green bars indicate the emergence of all thecomponents of each specific hormone signaling machinery.Branches in blue and purple indicate algae and land plants, re-spectively. The phylogenetic relationship among the plant speciesused in this study is based on Qiu et al. (2006), Finet et al. (2010),and Bowman (2013).
Plant Physiol. Vol. 167, 2015 877
Origin of Plant Hormone Signaling Pathways

repressor, degradation motif, dimerization domain, anddimerization domain, respectively (Paponov et al., 2009).The AUX/IAA ortholog of N. mirabilis clusters togetherwith land plant AUX/IAA proteins, although it lacksdomains I and II, which might be due to the short natureof the assembled transcriptome sequences. Nevertheless,all the core AUX signaling components seem to be al-ready present in charophytes. Moreover, the main AUXbiosynthesis pathway (TAA/YUC) already existed in thegenome of K. flaccidum (Wang et al., 2014). The AUXhormone has been found to be present in algal lineagesand to have effects on algal development (Tarakhovskayaet al., 2007; Lau et al., 2009). These lines of evidencesuggest that AUX signaling is likely to originate incharophytes and thus to account for the mechanism ofthe AUX responses in algal species (Fig. 3).
BR Signaling Originated before the Emergenceof Angiosperms
Our comparative genomic analysis shows that theorthologs of eight of the 10 BR signaling transductioncomponents (i.e. BAK1, BIN2, BRI1, BSU1, BZRs,CDG1, GRFs, and PP2A proteins) are present in thealgal genomes (Fig. 2), indicating that these proteinsarose before the emergence of land plants. The orthologsof BKI1 are found only in the genomes of angiosperms(Arabidopsis and rice). Furthermore, we identified aBKI1 protein ortholog in the genome of Amborellatrichopoda, which is the sister species to all other ex-tant angiosperms (Soltis et al., 2008; Finet et al., 2010;Amborella Genome Project, 2013). However, no BKI1homolog is found in the available EST data of Pinaceaespecies, although the absence of homologs in the PinaceaeEST data does not necessarily indicate the absence ofhomologs in the Pinaceae genomes. Nevertheless, theBKI1 protein seems to de novo originate at least in the lastcommon ancestor of extant angiosperms but after thedivergence of lycophytes from land plants (likely the splitbetween gymnosperms and angiosperms). The orthologsof BR receptors, BRI1 and BAK1, were identified in thegenomes of K. flaccidum and land plants. The predictedstructures of BRI1 orthologs of K. flaccidum and landplants share significant similarity with the Arabidopsisprotein structure (TM score = 0.194). The BR-binding siteis highly conserved among the land plant BRI1 orthologs(Supplemental Fig. S2). These results demonstrate thatthe canonical BR signaling machinery arose before theemergence of angiosperms and possibly after the splitbetween gymnosperms and angiosperms (Fig. 3).
CK Signaling Originated in Charophytes
The orthologs of all the core CK signaling compo-nents were identified in the genomes of land plantsand the charophyte K. flaccidum. The entire core CKsignaling and transport components cannot be identi-fied in the transcriptomes of the other four charophytes
(Fig. 2). Again, this might be due to the low coverage oftranscriptome data and/or the short nature of the as-sembled transcriptome sequences.
A previous study suggests that the CHASE domainof AHKs appeared only after plants conquered theland (Pils and Heyl, 2009). Further analyses of the S.pratensis EST data provide hints at the origin of CKreceptors in charophytes (Gruhn et al., 2014). Ouranalysis here shows that the AHK orthologs are pre-sent in the genomes of the charophytes N. mirabilis andK. flaccidum (Fig. 2), although the binding sites seemnot to be well conserved among AHK orthologs (theyare not well conserved among Arabidopsis AHKs;Supplemental Fig. S2). The orthologs of the CHASEdomain-containing AHKs were not identified in othernoncharophyte algal species (Supplemental Fig. S1).Therefore, the CHASE domain seems to be acquired bythe progenitor of AHKs in charophytes. To elucidatethe origin of the CHASE domain in plants, we recon-structed a phylogenetic tree of all currently availableCHASE domain sequences. Our phylogenetic treeshows that the CHASE domain sequences of charophyteand land plant AHKs cluster together (Shimodaira-Hasegawa test [SH-like] value = 0.808) and nest withinthe bacterial diversity (Fig. 4). The phylogenetic analysisand the absence of the CHASE domain in red algal andother green algal genomes suggest that plant speciesacquired the CHASE domain not through the ancientprimary endosymbiosis that gave birth to plastids (Reyes-Prieto et al., 2007) but through a relatively recent hori-zontal gene transfer event from bacteria to charophytespecies. An alternative hypothesis explaining the ob-served phylogenetic pattern would involve numerousindependent losses of the CHASE domain in red algae,green algae, and animals, which is highly unlikely. Takentogether, our results suggest that CK signaling originatedin charophyte species (Fig. 3).
ETH Signaling Originated Shortly after the Emergenceof Angiosperms
The orthologs of the core ETH signaling componentswere identified within the land plant genomes (exceptCTR1, EBFs, and ETP1/2; Fig. 2). As for EBFs, theirorthologs were found in the genomes of several charophytespecies and all the land plants except M. polymorpha.The EIN2 protein is present in the genomes of landplants and several charophytes. The C terminus of EIN2contained a putative nuclear localization signal (NLS)for nuclear translocation (Wen et al., 2012). However,the alignment of the C terminus of EIN2 shows that theNLS exists only in land plants (except M. polymorpha;Supplemental Fig. S2). The absence of EBFs and theNLS of EIN2 in M. polymorpha might be readily ex-plained by the low coverage/short nature of the tran-scriptome data. In the case of ETP1/2, a conserveddomain analysis shows that the Arabidopsis and riceETP1/2-like proteins have two domains, F-box andFBA_1 (Supplemental Table S2; Supplemental Fig. S1).
878 Plant Physiol. Vol. 167, 2015
Wang et al.

The FBA_1 domain interacts directly with EIN2 and isimportant for the ETP1/2 proteins to recognize theirsubstrates (Qiao et al., 2009). The ETP1/2 homologs in theS. moellendorffii and P. patens genomes do not have theFBA_1 domain. Moreover, we also found that the ho-mologs of ETP1/2 in A. trichopoda (the most basal angio-sperm lineage) do not contain the FBA_1 domain. Theseresults suggest that the FBA_1 domain was gained by theETP1/2 progenitor shortly after the emergence of angio-sperms but before the evolutionary split of monocots andeudicots. Therefore, the origin of canonical ETH signalingpostdates shortly the emergence of angiosperms but pre-dates the divergence of monocots and eudicots (Fig. 3).It appears that the receiver domain of ETH receptors
is well conserved among algal species and land plants
(Supplemental Fig. S2). The ETH-binding activity ofETH receptor-like proteins has been detected in all landplants, including M. polymorpha and P. patens (Wanget al., 2006). The ETH-mediated submergence responsesalso have been found in mosses (Yasumura et al., 2012).These facts indicate that there might be an alternativeETH response pathway that does not depend on EIN2-ETP1/2 in moss or even some charophytes.
The higher plants were thought to acquire the ETHreceptors through endosymbiosis, because similar pro-teins are found in the cyanobacterium from which theplant chloroplast arose (Mount and Chang, 2002; Schalleret al., 2011). However, our phylogenetic analyses of theirprotein domains show that the cyanobacterial proteinsare not the closest relatives of ETH receptors of plants
Figure 4. The phylogenetic relationship of the CHASE domains of prokaryotic and eukaryotic origin. The phylogeny is anapproximate maximum-likelihood (ML) tree reconstructed using FastTree 2 based on the CHASE domain sequences of bothprokaryotes and eukaryotes. SH-like values are shown near selected nodes.
Plant Physiol. Vol. 167, 2015 879
Origin of Plant Hormone Signaling Pathways

(Supplemental Fig. S3) and thus do not support theplastid origin of ETH receptors.
GA Signaling Originated after the Split of Bryophytesfrom Land Plants
Our comparative analysis shows that the land plant(except M. polymorpha) genomes encode the orthologsof all the core components of GA signaling. The ho-mologs of DELLA in the M. polymorpha genome do notshare the same domain architecture as the Arabidopsisproteins (Supplemental Fig. S1). But it is still unclearwhether the DELLA domain is truly missing in thehomologs or if the absence of the DELLA domain isdue to the short nature of the assembled transcriptomesequences. Interestingly, the orthologs of DELLA andGID1 proteins of P. patens contain no protein-proteininteraction regions (DELLA, LExLE, and VHYNPmotifs in the DELLA protein and residues Ile-24, Phe-27, and Tyr-31 in the GID1A protein; SupplementalFig. S2; Murase et al., 2008), which is consistent withprevious experimental studies showing that GID1 andDELLA do not interact with each other in P. patens(Hirano et al., 2007; Yasumura et al., 2007; Hayashiet al., 2010). It follows that GA signaling originatedafter the evolutionary split of bryophytes and landplants (Yasumura et al., 2007).
JA Signaling Originated in the Last Common Ancestor ofLand Plants
It appears that the algal genomes encode theorthologs of three JA signaling components (COI1,TPL, and MYCs), while the land plant genomes encodethe orthologs of all the core JA signaling components.The TM scores of the predicted structures of P. patensand M. polymorpha COI1 orthologs and the experi-mental structure of the Arabidopsis protein are 0.167and 0.215, respectively. Moreover, all the predictedstructures contain a loop region that forms an abruptkink in the middle, which plays a key role in binding JA(Sheard et al., 2010). Sequence alignment shows that thebinding sites are well conserved between P. patens andM. polymorpha COI1 orthologs and Arabidopsis COI1proteins (Supplemental Fig. S2). Therefore, we hy-pothesize that JA signaling originated in the last com-mon ancestor of land plants (Fig. 3).
SA Signaling Originated in the Last Common Ancestor ofLand Plants
Our analysis shows that the algal genomes encodethe orthologs of at least two SA signaling components(i.e. TGAs and WRKY proteins) but not the orthologsof NPR proteins. Initial conserved domain analysisshows that the BTB domain is absent in theM. polymorphaNPR-like protein. However, a close analysis of the BTBdomain and M. polymorpha NPR sequences suggests
that the M. polymorpha NPR protein encodes partial BTB(Supplemental Fig. S2), which might be due to the shortnature of the assembled transcriptome sequences. Thus,the NPR protein ortholog seems to be present in theM. polymorpha genome, and the land plant genomes encodethe orthologs of all the core components of SA signaling.These results suggest the origin of SA signaling transduc-tion in the last common ancestor of land plants (Fig. 3).
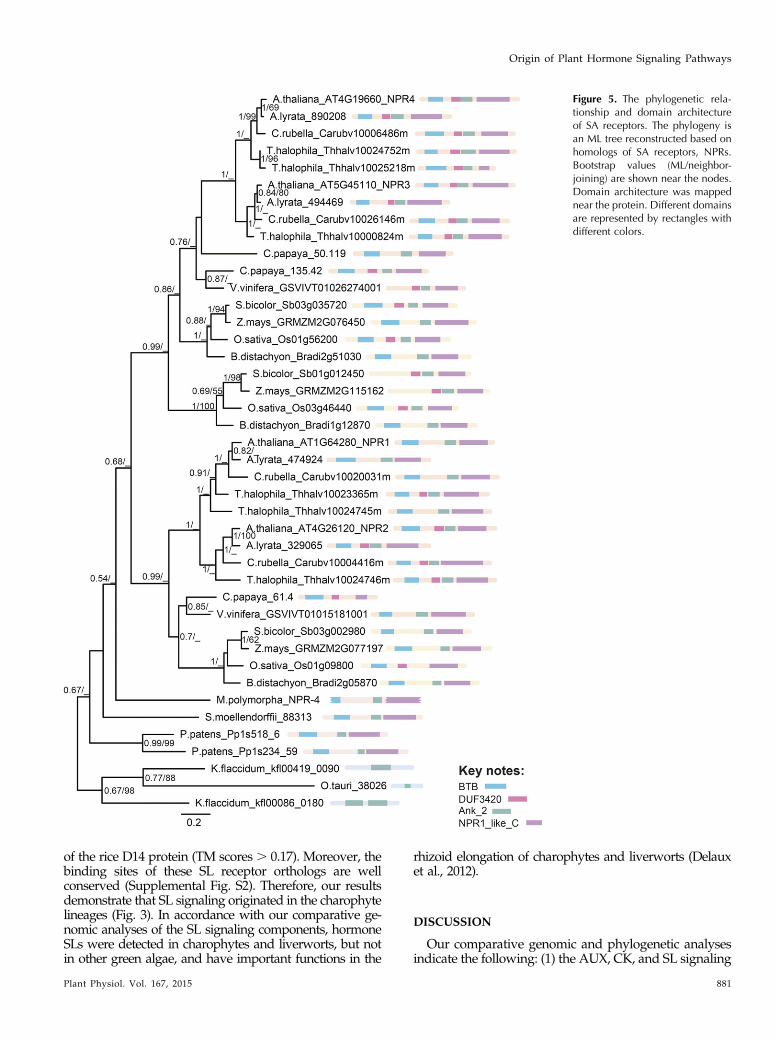
Interestingly, our phylogenetic analyses show thatboth the paralog NPR1/2 proteins and the paralogNPR3/4 proteins were generated by a gene duplicationevent in the last common ancestor of the Brassicaceae(Fig. 5). The NPR1/2 progenitor diverged from theNPR3/4 progenitor after the split between lycophytesand angiosperms (Fig. 5). Of note, the monocot ge-nomes have two copies of NPR3/4-related proteins,which are generated by a gene duplication event dif-ferent from the one generating Brassicaceae NPR3/4proteins (Fig. 5). Within the model plant Arabidopsis,the SA signaling mechanism is as follows: NPR1 is amaster regulator of systemic acquired resistance, whileNPR3 and NPR4 act as the SA receptors (Fu et al., 2012).When the SA concentration is low, NPR4, but notNPR3, binds SA, inhibiting the degradation of NPR1.During pathogen infection, in the infection site with thehighest SA level, SA binds to NPR3, promoting NPR1degradation to allow programmed cell death; in cellswith lower SA levels, NPR1 accumulates and activatesthe transcription factors to turn on the defense-relatedgenes (Ülker and Somssich, 2004; Fu et al., 2012).However, we found that the NPR copy number differsin land plant genomes. For example, there are twocopies of the NPR4-related protein and two copies ofthe NPR1-related protein in the Thellungiella halophilagenome, only one copy of the NPR3/4-related proteinin the grape (Vitis vinifera) genome, one copy of theNPR1/2-related protein in the monocot genomes, and oneand two copies of the NPR protein in the S. moellendorffiiand P. patens genomes, respectively (Fig. 5). Especially,with only one NPR3/4-related protein in the genome ofgrape, the SA signaling logic characterized in Arabidopsiscannot work, given that the NPR3/4 proteins play dif-ferent roles under different SA concentrations. It seemsthat species with different NPR copy numbers might havedifferent SA response mechanisms from Arabidopsis.
SL Signaling Originated in Charophytes
Our systematic analysis shows that the genomes ofgreen algae and land plants encode the orthologs ofMAX2 (D3), D14, and SMXLs (D53); that the TCPtranscription factor BRC1 protein was identified in thecharophyte and land plant genomes; and that the ortho-logs of all core components of SL signaling transduc-tion were identified in the genomes of the charophyteN. mirabilis and land plants. The predicted structuresof the SL receptor orthologs of P. patens,M. polymorpha,and three charophytes (N. mirabilis, K. flaccidum, andS. pratensis) are highly similar to the experimental structure
880 Plant Physiol. Vol. 167, 2015
Wang et al.
of the rice D14 protein (TM scores. 0.17). Moreover, thebinding sites of these SL receptor orthologs are wellconserved (Supplemental Fig. S2). Therefore, our resultsdemonstrate that SL signaling originated in the charophytelineages (Fig. 3). In accordance with our comparative ge-nomic analyses of the SL signaling components, hormoneSLs were detected in charophytes and liverworts, but notin other green algae, and have important functions in the
rhizoid elongation of charophytes and liverworts (Delauxet al., 2012).
DISCUSSION
Our comparative genomic and phylogenetic analysesindicate the following: (1) the AUX, CK, and SL signaling
Figure 5. The phylogenetic rela-tionship and domain architectureof SA receptors. The phylogeny isan ML tree reconstructed based onhomologs of SA receptors, NPRs.Bootstrap values (ML/neighbor-joining) are shown near the nodes.Domain architecture was mappednear the protein. Different domainsare represented by rectangles withdifferent colors.
Plant Physiol. Vol. 167, 2015 881
Origin of Plant Hormone Signaling Pathways

pathways originated in charophytes; (2) the ABA, JA,and SA signaling pathways evolved in the most recentcommon ancestor of land plants; (3) GA signaling orig-inated after the divergence of bryophytes from landplants; (4) the canonical BR signaling originated in thelast common ancestor of angiosperms, likely after thesplit between gymnosperms and angiosperms; and (5)the canonical ETH signaling originated shortly after theemergence of angiosperms but before the split ofmonocots and eudicots (Fig. 3). Although different planthormone signaling mechanisms appear to originate atdifferent time points on the evolutionary history ofplants, many signaling components are already encodedby the algal genomes.
AUX, JA, ETH, GA, and SL Signaling Originated from theSCF-Mediated Ubiquitination
AUX, JA, ETH, SL, and GA signaling pathways are alldependent on the SCF-mediated ubiquitin-proteasomesystem (Santner and Estelle, 2009; Jiang et al., 2013). In-terestingly, AUX and JA signaling pathways share acommon mechanism of hormone sensing and response.(1) Both JA and AUX receptors are F-box proteins. TheCOI1 and TIR1 proteins share highly similar crystalstructures (TM score = 0.481). Our similarity searchshows that the JA receptor (COI1) shares the highestsequence identity with the AUX receptors (TIR1/AFBs)within the Arabidopsis genome. Phylogenetic analysis ofTIR1/AFBs and COI1-related proteins shows that JAand AUX receptors are the closest relatives and descendfrom gene duplication events deep in the last commonancestor of land plants (Fig. 6). (2) JA and AUX share asimilar signal transduction logic (Santner and Estelle,2009; Pauwels et al., 2010). Both hormones function asmolecular glue between the receptors and the repressors.Both signaling pathways require the SCF-dependentdegradation of the repressors (Santner and Estelle,2009; Pauwels et al., 2010). The repressors, the AUX/IAAproteins of AUX signaling and the NINJA proteins of JAsignaling, have an EAR motif that can bind the TPL andTPL-related proteins (Pauwels et al., 2010). (3) Conjugatejasmonoyl-Ile is the bioactive form of the hormone JA(Ludwig-Müller, 2011). The JAR1 protein is the onlyGH3 protein in Arabidopsis that can conjugate JA to Ile,while some other GH3 proteins conjugate AUX but notJA (Ludwig-Müller, 2011; Supplemental Fig. S1). TheP. patens genome has two copies of the GH3 gene, PpGH3-1and PpGH3-2, both of which can catalyze the conju-gation of the hormones JA and AUX to amino acids andare thought to arise from a gene duplication of a pro-genitor gene synthesizing conjugates with JA and AUX(Ludwig-Müller, 2011). These facts and results suggestthat JA and AUX signaling pathways are likely to orig-inate from a single ancient hormone signaling and thatthe ancient hormone signaling evolved into AUX and JAsignaling pathways when the ancient receptors (TIR1/AFBs and COI1 progenitor) duplicated and divergedto bind different hormones (Supplemental Fig. S2) andregulate different target genes. Further experiments are
needed to determine whether this ancient hormonesignaling is in response to AUX, JA, or both.
Distinct from JA and AUX signaling pathways, theF-box proteins, SLY1/2 and D3, do not act as the re-ceptors of GA and SL signaling pathways and are notclosely related to AUX and JA receptors (Gagne et al.,2002). Surprisingly, the crystal structures of the GAreceptor (GID1) of Arabidopsis and the SL receptor(D14) of rice are similar (TM score = 0.187), albeit nodescribable sequence similarity could be detected. It ispossible that there is deep homology between GID1and D14 proteins. Nevertheless, these two signalingpathways appear to be independently derived fromthe SCF-mediated ubiquitination pathway.
CK and ETH Signaling Independently Originatedfrom TCS
Both CK and ETH signaling pathways undergomultistep phosphorylation at the onset of hormoneresponses. All the CK and ETH receptors are His ki-nases and share three common domains, HATPase,HisKA, and receiver domains, resembling the compo-nents of TCS (Chang et al., 1993; Pils and Heyl, 2009;Schaller et al., 2011). Our phylogenetic analyses ofthese three domains reveal that CK and ETH receptorsoccupy different phylogenetic positions and do notform a monophyletic group (Supplemental Fig. S3).Besides the three TCS-related protein domains, CK andETH receptors have their specific domains, CHASEand GAF, respectively. It follows that CK and ETHsignaling pathways evolved from independent elabo-ration of the TCS. ETH signaling is also mediated bySCF-dependent ubiquitination (Mount and Chang, 2002).Therefore, ETH signaling seems to originate by recom-bining an ancient TCS and an SCF-mediated ubiquitin-proteasome pathway.
Some Signaling Components Underwent Gene FamilyExpansion in Vascular Plants
Some plant hormone signaling components seem toundergo significant gene family expansion in vascularplants. The following are some representative exam-ples. (1) The number of PYR proteins (ABA receptors)increases in higher plants: the genomes of P. patens andS. moellendorffii encode two and five members, whereasthe genomes of rice and Arabidopsis encode 10 and 14members, respectively (Supplemental Fig. S1). (2) TheETP1/2-related proteins of the ETH signaling pathwayarose shortly after the emergence of angiosperms. Therice genome encodes 11 ETP1/2-related proteins, whilethe Arabidopsis genome encodes 83 ETP1/2-relatedproteins (Supplemental Fig. S1). (3) The AUX/IAAs ofAUX signaling arose in charophytes. While the genomesof P. patens and S. moellendorffii encode two and fourAUX/IAAs, the genomes of Arabidopsis and rice encode25 and 27 AUX/IAAs, respectively (Supplemental Fig.S1). These plant hormone signaling components seem to
882 Plant Physiol. Vol. 167, 2015
Wang et al.

undergo a rapid gene family expansion during the evo-lutionary history of higher plants. Although the exactadvantages of these gene family expansions are unclear,
it is conceivable that they might make substantial con-tributions to the evolution and features specific to higherplants.
Figure 6. The phylogenetic relationship and domain architecture of JA and AUX receptors. The phylogeny is an ML treereconstructed based on homologs of JA and AUX receptors. Bootstrap values (ML/neighbor-joining) are shown near the nodes.Domain architecture was mapped near the protein. Different domains are represented by rectangles with different colors.Protein structures of JA and AUX receptor orthologs are shown near the corresponding proteins.
Plant Physiol. Vol. 167, 2015 883
Origin of Plant Hormone Signaling Pathways

Molecular Tinkering Might Be an Important Mechanismfor Plant Hormone Evolution
Domain rearrangement has been suggested to play arole in the evolution of the AUX hormone signalingproteins (Kalluri et al., 2007; Finet et al., 2013). For theproteins mediating plant hormone signal transductionor transport, a total of 58 different protein domainswere identified. Among these protein domains, 18appear to be plant specific, while the others seem tohave a more ancient origin (Supplemental Table S2).Moreover, the same domain might be recruited indifferent hormone signaling pathways and even indifferent components of the same signaling pathway.For example, the Pkinase domain is embedded withinMAP3K of ETH signaling, SnRK2s of ABA signaling,and BRI1, BAK1, CDG1, and BIN2 of BR signaling. Thesame domain might exist in different domain archi-tectures: BSKs of BR signaling and the CTR1 of ETHsignaling both have a Pkinase_Tyr domain, but thisdomain is combined with the TPR_11 domain in BSKsand combined with the EDR1 domain in CTR1; bothABFs of ABA signaling and TGAs of SA signalinghave a bZIP_1 domain, but TGAs have an additionaldomain, DOG1. These examples indicate that do-main shuffling did occur frequently during the originand evolution of plant hormone signaling. We be-lieve that evolutionary tinkering with the preexistingprotein domains might represent an important evo-lutionary mechanism for plant hormone signalingevolution.
Implications in the Emergence of Land Plants
The emergence of land plants represents one of themost important innovations in the history of life. Whentransiting from an aquatic to a terrestrial environment,plants evolved from organisms with a simple body of afew cells to ones with complex organs and tissues andencountered new environmental stresses, such asdrought, UV light, and pathogens (Kenrick and Crane,1997). Little is known about the biological mechanismsunderlying the morphological changes and the adap-tation to land of plants. We found that three majorhormone (ABA, JA, and SA) signaling pathways mighthave emerged while plants were colonizing the land.All of these hormone signaling pathways regulate de-fense against biotic and abiotic stresses: SA modulatesplant immunity to pathogens; JAs control plant defenseagainst wounding, herbivores, and certain pathogens;and ABA plays an important role in plant responsesto environmental stresses, especially drought. There-fore, it is highly possible that these signaling path-ways likely emerged and evolved into their currentforms as a consequence of selection pressure exertedby the biotic or abiotic stresses encountered in terres-trial environments (Hauser et al., 2011). The innova-tion of these plant hormone signaling pathways mightprovide a potential mechanism for the emergence ofland plants.
Caveats
Our analyses come with several caveats. First, thepresence of hormone signaling component orthologs doesnot necessarily indicate that these orthologs would worktogether as genuine signaling pathways. For example,orthologs of all the core components of GA are present inthe P. patens genome, but DELLA and GID1 do not in-teract with each other in the presence of GA (Hirano et al.,2007; Yasumura et al., 2007; Hayashi et al., 2010). Furtherexperiments are needed to explore the interaction of thesesignaling component orthologs. Second, hormone biologyhas been studied mainly based on the genetic analysis ofangiosperms, especially the model plant Arabidopsis.Hormone signaling might have a different/alternativeform in plants outside the model plant Arabidopsis(such as the SA and ETH signaling pathways discussedabove). Obviously, more work is needed to characterizeplant hormone signaling pathways in lower plants and/ornonmodel plants, which will provide extremely importantinsights into their biology, origin, and evolution. Third, weused a combination of similarity search, phylogenetic re-construction, and conserved domain analysis to identifyorthologs (discussed above). Only homologs with thesame domain architecture as the Arabidopsis proteins areconsidered as orthologs. This approach is a conservativeone, since some domains might not play an important rolein hormone signaling. However, these domains are func-tionally important; otherwise, they would not be con-served and identified as domains. Last, we used thetranscriptome data of several charophytes and the liver-wort M. polymorpha in this study. As discussed above,because gene expression is usually tissue specific, devel-opmental stage specific, and/or stress induced, tran-scriptome data sometimes cannot cover all the geneticinformation of the genomes. Because the coverage oftranscriptome data is sometimes low and the assembledsequences are relatively short, we cannot conclusively inferthe presence/absence of the orthologs of hormone signal-ing components in these species. Further sequencing of thegenomes of additional charophytes and M. polymorphacould help us understand the origin and evolution of planthormone signaling pathways.
MATERIALS AND METHODS
Species Selected for This Study
Genome sequences of nine plant species, Arabidopsis (Arabidopsis thaliana),rice (Oryza sativa), Selaginella moellendorffii, Physcomitrella patens, Klebsormidiumflaccidum, Volvox carteri, Chlamydomonas reinhardtii, Ostreococcus tauri, and Cya-nidioschyzon merolae, are available in public database and chosen for comparativegenomic analysis in this study (Supplemental Table S1). In addition, we alsoused publicly available assembled or raw transcriptome data from the liverwortMarchantia polymorpha and four charophytes (Nitella hyalina, Nitella mirabilis,Penium margaritaceum, and Spiragyra pratensis; Supplemental Table S1; Timmeet al., 2012). We used SeqMan NGen from DNASTAR to de novo assemble thetranscriptome data of M. polymorpha with default parameters.
Sequence Similarity Search
To identify the homologs of the core proteins implicated in the transport andsignal transduction of AUX, ABA, BR, CK, ETH, GA, JA, SA, and SL
884 Plant Physiol. Vol. 167, 2015
Wang et al.

hormones, we performed similarity searches using the BLAST (BLASTP andTBLASTN) algorithms with the Arabidopsis proteins as queries and an E-value threshold of 1025. To identify distant homologs, we performed furtherrounds of BLAST (BLASTP and TBLASTN) searches with the homologsidentified in P. patens.
Phylogenetic Analysis
Protein domain sequence alignments were also retrieved from the Pfamdatabase (Finn et al., 2014). Because most of the protein domain alignments areextremely large, we infer a phylogenetic relationship for each protein domainusing FastTree 2 with the CAT model (Price et al., 2010). The phylogeneticsupport for each split was assessed using SH-like values (Price et al., 2010). Toexplore the detailed evolutionary history of each hormone signaling compo-nent, homologous sequences were retrieved from Phytozome version 9.1(http://phytozome.jgi.doe.gov/pz/portal.html). The translated protein se-quences were aligned using MUSCLE (Edgar, 2004) and then manually edited.Neighbor-joining phylogenetic trees were reconstructed using MEGA5(Tamura et al., 2011). The phylogenetic support for each split was evaluatedwith 1,000 bootstrap replicates. Moreover, we also employed MEGA5(Tamura et al., 2011) to reconstruct ML phylogenetic trees. The phylogeneticsupport for each split was evaluated with 100 bootstrap replicates.
Ortholog Identification
We used phylogenetic analysis to identify potential orthologs. Proteinconserved domain analysis of each potential ortholog was determined usingPfam (Finn et al., 2014). Only those with the same conserved domain archi-tecture as the Arabidopsis proteins were defined as orthologs.
Protein Structure Prediction
The structures of hormone receptor orthologs of P. patens, M. polymorpha,and charophytes were predicted using a homology-modeling approachavailable through Phyre2 (Kelley and Sternberg, 2009) and the I-TASSERonline service (Roy et al., 2010) based on the experimental crystal structuresof the hormone receptors of Arabidopsis and rice (Müller-Dieckmann et al.,1999; Hayashi et al., 2008; Murase et al., 2008; Nishimura et al., 2009; Sheardet al., 2010; Hothorn et al., 2011a, 2011b; Zhao et al., 2013). The predictedstructures were visualized using PyMOL version 1.5.0.4 (Schrödinger, LLC,2011). We also compared the predicted structures with the available experi-mental crystal structures, and the similarity between them was evaluatedusing the TM score (Xu and Zhang, 2010).
Supplemental Data
The following supplemental materials are available.
Supplemental Figure S1. Phylogenetic analysis and conserved domainarchitecture of plant hormone signaling components.
Supplemental Figure S2. The alignments of the crucial regions of planthormone receptors and other important proteins.
Supplemental Figure S3. The phylogenetic relationship of domains of ETHand CK receptors.
Supplemental Table S1. The information of plant genome-scale data usedin this study.
Supplemental Table S2. The results of protein structural domain analysisof plant hormone signaling components.
Received July 25, 2014; accepted December 30, 2014; published January 5,2015.
LITERATURE CITED
Amborella Genome Project (2013) The Amborella genome and the evolu-tion of flowering plants. Science 342: 1241089
Banks JA, Nishiyama T, Hasebe M, Bowman JL, Gribskov M, dePamphilis C,Albert VA, Aono N, Aoyama T, Ambrose BA, et al (2011) The Selaginella
genome identifies genetic changes associated with the evolution of vascularplants. Science 332: 960–963
Boot KJ, Libbenga KR, Hille SC, Offringa R, van Duijn B (2012) Polarauxin transport: an early invention. J Exp Bot 63: 4213–4218
Boursiac Y, Léran S, Corratgé-Faillie C, Gojon A, Krouk G, Lacombe B(2013) ABA transport and transporters. Trends Plant Sci 18: 325–333
Bowman JL (2013) Walkabout on the long branches of plant evolution. CurrOpin Plant Biol 16: 70–77
Chang C, Kwok SF, Bleecker AB, Meyerowitz EM (1993) Arabidopsisethylene-response gene ETR1: similarity of product to two-componentregulators. Science 262: 539–544
Chen X, Zhang J (2012) The ortholog conjecture is untestable by the currentgene ontology but is supported by RNA sequencing data. PLoS ComputBiol 8: e1002784
Chervitz SA, Aravind L, Sherlock G, Ball CA, Koonin EV, Dwight SS,Harris MA, Dolinski K, Mohr S, Smith T, et al (1998) Comparison ofthe complete protein sets of worm and yeast: orthology and divergence.Science 282: 2022–2028
Delaux PM, Xie X, Timme RE, Puech-Pages V, Dunand C, Lecompte E,Delwiche CF, Yoneyama K, Bécard G, Séjalon-Delmas N (2012) Originof strigolactones in the green lineage. New Phytol 195: 857–871
de Saint Germain A, Bonhomme S, Boyer FD, Rameau C (2013) Novelinsights into strigolactone distribution and signalling. Curr Opin PlantBiol 16: 583–589
De Smet I, Voss U, Lau S, Wilson M, Shao N, Timme RE, Swarup R, KerrI, Hodgman C, Bock R, et al (2011) Unraveling the evolution of auxinsignaling. Plant Physiol 155: 209–221
Dharmasiri N, Dharmasiri S, Estelle M (2005) The F-box protein TIR1 is anauxin receptor. Nature 435: 441–445
Edgar RC (2004) MUSCLE: multiple sequence alignment with high accuracy andhigh throughput. Nucleic Acids Res 32: 1792–1797
Finet C, Berne-Dedieu A, Scutt CP, Marlétaz F (2013) Evolution of the ARFgene family in land plants: old domains, new tricks. Mol Biol Evol 30: 45–56
Finet C, Timme RE, Delwiche CF, Marlétaz F (2010) Multigene phylogenyof the green lineage reveals the origin and diversification of land plants.Curr Biol 20: 2217–2222
Finn RD, Bateman A, Clements J, Coggill P, Eberhardt RY, Eddy SR,Heger A, Hetherington K, Holm L, Mistry J, et al (2014) Pfam: theprotein families database. Nucleic Acids Res 42: D222–D230
Fu ZQ, Yan S, Saleh A, Wang W, Ruble J, Oka N, Mohan R, Spoel SH,Tada Y, Zheng N, et al (2012) NPR3 and NPR4 are receptors for theimmune signal salicylic acid in plants. Nature 486: 228–232
Fujii H, Chinnusamy V, Rodrigues A, Rubio S, Antoni R, Park SY, CutlerSR, Sheen J, Rodriguez PL, Zhu JK (2009) In vitro reconstitution of anabscisic acid signalling pathway. Nature 462: 660–664
Gagne JM, Downes BP, Shiu SH, Durski AM, Vierstra RD (2002) The F-box subunit of the SCF E3 complex is encoded by a diverse superfamilyof genes in Arabidopsis. Proc Natl Acad Sci USA 99: 11519–11524
Gruhn N, Halawa M, Snel B, Seidl MF, Heyl A (2014) A subfamily of putativecytokinin receptors is revealed by an analysis of the evolution of the two-component signaling system of plants. Plant Physiol 165: 227–237
Gruhn N, Heyl A (2013) Updates on the model and the evolution ofcytokinin signaling. Curr Opin Plant Biol 16: 569–574
Guilfoyle TJ, Hagen G (2007) Auxin response factors. Curr Opin Plant Biol10: 453–460
Hauser F, Waadt R, Schroeder JI (2011) Evolution of abscisic acid synthesisand signaling mechanisms. Curr Biol 21: R346–R355
Hayashi K,HorieK,Hiwatashi Y, KawaideH, Yamaguchi S,HanadaA,NakashimaT, Nakajima M, Mander LN, Yamane H, et al (2010) Endogenous diterpenesderived from ent-kaurene, a common gibberellin precursor, regulate protonemadifferentiation of the moss Physcomitrella patens. Plant Physiol 153: 1085–1097
Hayashi K, Tan X, Zheng N, Hatate T, Kimura Y, Kepinski S, Nozaki H (2008)Small-molecule agonists and antagonists of F-box protein-substrate interactionsin auxin perception and signaling. Proc Natl Acad Sci USA 105: 5632–5637
Hegyi H, Gerstein M (2001) Annotation transfer for genomics: measuringfunctional divergence in multi-domain proteins. Genome Res 11: 1632–1640
Hirano K, Nakajima M, Asano K, Nishiyama T, Sakakibara H, Kojima M, KatohE, Xiang H, Tanahashi T, HasebeM, et al (2007) The GID1-mediated gibberellinperception mechanism is conserved in the lycophyte Selaginella moellendorffii butnot in the bryophyte Physcomitrella patens. Plant Cell 19: 3058–3079
Hothorn M, Belkhadir Y, Dreux M, Dabi T, Noel JP, Wilson IA, Chory J(2011a) Structural basis of steroid hormone perception by the receptorkinase BRI1. Nature 474: 467–471
Plant Physiol. Vol. 167, 2015 885
Origin of Plant Hormone Signaling Pathways

Hothorn M, Dabi T, Chory J (2011b) Structural basis for cytokinin recognitionby Arabidopsis thaliana histidine kinase 4. Nat Chem Biol 7: 766–768
Inoue T, Higuchi M, Hashimoto Y, Seki M, Kobayashi M, Kato T, TabataS, Shinozaki K, Kakimoto T (2001) Identification of CRE1 as a cytokininreceptor from Arabidopsis. Nature 409: 1060–1063
Jiang L, Liu X, Xiong G, Liu H, Chen F, Wang L, Meng X, Liu G, Yu H,Yuan Y, et al (2013) DWARF 53 acts as a repressor of strigolactonesignalling in rice. Nature 504: 401–405
Kalluri UC, Difazio SP, Brunner AM, Tuskan GA (2007) Genome-wideanalysis of Aux/IAA and ARF gene families in Populus trichocarpa.BMC Plant Biol 7: 59
Kelley DR, Estelle M (2012) Ubiquitin-mediated control of plant hormonesignaling. Plant Physiol 160: 47–55
Kelley LA, Sternberg MJ (2009) Protein structure prediction on the Web: acase study using the Phyre server. Nat Protoc 4: 363–371
Kendrick MD, Chang C (2008) Ethylene signaling: new levels of com-plexity and regulation. Curr Opin Plant Biol 11: 479–485
Kenrick P, Crane PR (1997) The origin and early evolution of plants onland. Nature 389: 33–39
Kumar R, Sharma AK (2014) Ethylene perception and signalling in ripening fruit.In P Nath, M Bouzayen, AK Matoo, JC Pech, eds, Fruit Ripening: Physiology,Signalling and Genomics. CABI, Wallingford, UK, pp 193–201
Lau S, Shao N, Bock R, Jürgens G, De Smet I (2009) Auxin signaling inalgal lineages: fact or myth? Trends Plant Sci 14: 182–188
Ludwig-Müller J (2011) Auxin conjugates: their role for plant developmentand in the evolution of land plants. J Exp Bot 62: 1757–1773
Mashiguchi K, Tanaka K, Sakai T, Sugawara S, Kawaide H, Natsume M,Hanada A, Yaeno T, Shirasu K, Yao H, et al (2011) The main auxin bio-synthesis pathway in Arabidopsis. Proc Natl Acad Sci USA 108: 18512–18517
Mount SM, Chang C (2002) Evidence for a plastid origin of plant ethylenereceptor genes. Plant Physiol 130: 10–14
Mravec J, Sk�upa P, Bailly A, Hoyerová K, K�re�cek P, Bielach A, Petrásek J,Zhang J, Gaykova V, Stierhof YD, et al (2009) Subcellular homeostasisof phytohormone auxin is mediated by the ER-localized PIN5 trans-porter. Nature 459: 1136–1140
Müller-Dieckmann HJ, Grantz AA, Kim SH (1999) The structure of thesignal receiver domain of the Arabidopsis thaliana ethylene receptorETR1. Structure 7: 1547–1556
Murase K, Hirano Y, Sun TP, Hakoshima T (2008) Gibberellin-inducedDELLA recognition by the gibberellin receptor GID1. Nature 456: 459–463
Nishimura N, Hitomi K, Arvai AS, Rambo RP, Hitomi C, Cutler SR,Schroeder JI, Getzoff ED (2009) Structural mechanism of abscisic acidbinding and signaling by dimeric PYR1. Science 326: 1373–1379
Paponov IA, Teale W, Lang D, Paponov M, Reski R, Rensing SA, Palme K(2009) The evolution of nuclear auxin signalling. BMC Evol Biol 9: 126
Pauwels L, Barbero GF, Geerinck J, Tilleman S, Grunewald W, Pérez AC,Chico JM, Bossche RV, Sewell J, Gil E, et al (2010) NINJA connects theco-repressor TOPLESS to jasmonate signalling. Nature 464: 788–791
Péret B, Swarup K, Ferguson A, Seth M, Yang Y, Dhondt S, James N,Casimiro I, Perry P, Syed A, et al (2012) AUX/LAX genes encode afamily of auxin influx transporters that perform distinct functions dur-ing Arabidopsis development. Plant Cell 24: 2874–2885
Pils B, Heyl A (2009) Unraveling the evolution of cytokinin signaling. PlantPhysiol 151: 782–791
Price MN, Dehal PS, Arkin AP (2010) FastTree 2: approximatelymaximum-likelihood trees for large alignments. PLoS ONE 5: e9490
Qiao H, Chang KN, Yazaki J, Ecker JR (2009) Interplay between ethylene,ETP1/ETP2 F-box proteins, and degradation of EIN2 triggers ethyleneresponses in Arabidopsis. Genes Dev 23: 512–521
Qiao H, Shen Z, Huang SS, Schmitz RJ, Urich MA, Briggs SP, Ecker JR(2012) Processing and subcellular trafficking of ER-tethered EIN2 con-trol response to ethylene gas. Science 338: 390–393
Qiu YL, Li L, Wang B, Chen Z, Knoop V, Groth-Malonek M, Dombrovska O,Lee J, Kent L, Rest J, et al (2006) The deepest divergences in land plants in-ferred from phylogenomic evidence. Proc Natl Acad Sci USA 103: 15511–15516
Rensing SA, Lang D, Zimmer AD, Terry A, Salamov A, Shapiro H,Nishiyama T, Perroud PF, Lindquist EA, Kamisugi Y, et al (2008) ThePhyscomitrella genome reveals evolutionary insights into the conquestof land by plants. Science 319: 64–69
Reyes-Prieto A, Weber AP, Bhattacharya D (2007) The origin and estab-lishment of the plastid in algae and plants. Annu Rev Genet 41: 147–168
Roy A, Kucukural A, Zhang Y (2010) I-TASSER: a unified platform forautomated protein structure and function prediction. Nat Protoc 5: 725–738
Santner A, Estelle M (2009) Recent advances and emerging trends in planthormone signalling. Nature 459: 1071–1078
Schaller GE, Shiu SH, Armitage JP (2011) Two-component systems andtheir co-option for eukaryotic signal transduction. Curr Biol 21: R320–R330
Schrödinger, LLC (2011) The PyMOL molecular graphics system, Version1.5.0.4. Schrödinger, LLC, Portland, OR
Sharma N, Bhalla PL, Singh MB (2013) Transcriptome-wide profiling andexpression analysis of transcription factor families in a liverwort,Marchantia polymorpha. BMC Genomics 14: 915
Sheard LB, Tan X, Mao H, Withers J, Ben-Nissan G, Hinds TR, KobayashiY, Hsu FF, Sharon M, Browse J, et al (2010) Jasmonate perception byinositol-phosphate-potentiated COI1-JAZ co-receptor. Nature 468: 400–405
Shimada A, Ueguchi-Tanaka M, Nakatsu T, Nakajima M, Naoe Y, Oh-miya H, Kato H, Matsuoka M (2008) Structural basis for gibberellinrecognition by its receptor GID1. Nature 456: 520–523
Soltis DE, Bell CD, Kim S, Soltis PS (2008) Origin and early evolution ofangiosperms. Ann N Y Acad Sci 1133: 3–25
Sun TP (2008) Gibberellin metabolism, perception and signaling pathwaysin Arabidopsis. The Arabidopsis Book 6: e0103, doi/10.1199/tab.0103
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011) MEGA5:molecular evolutionary genetics analysis using maximum likelihood, evolution-ary distance, and maximum parsimony methods. Mol Biol Evol 28: 2731–2739
Tarakhovskaya ER, Maslov YI, Shishova MF (2007) Phytohormones inalgae. Russ J Plant Physiol 54: 163–170
Tatusov RL, Koonin EV, Lipman DJ (1997) A genomic perspective onprotein families. Science 278: 631–637
Thaler JS, Humphrey PT, Whiteman NK (2012) Evolution of jasmonateand salicylate signal crosstalk. Trends Plant Sci 17: 260–270
Timme RE, Bachvaroff TR, Delwiche CF (2012) Broad phylogenomicsampling and the sister lineage of land plants. PLoS ONE 7: e29696
Tromas A, Paque S, Stierlé V, Quettier AL, Muller P, Lechner E, Genschik P,Perrot-Rechenmann C (2013) Auxin-binding protein 1 is a negative regulatorof the SCF(TIR1/AFB) pathway. Nat Commun 4: 2496
Tougane K, Komatsu K, Bhyan SB, Sakata Y, Ishizaki K, Yamato KT,Kohchi T, Takezawa D (2010) Evolutionarily conserved regulatorymechanisms of abscisic acid signaling in land plants: characterization ofABSCISIC ACID INSENSITIVE1-like type 2C protein phosphatase in theliverwort Marchantia polymorpha. Plant Physiol 152: 1529–1543
Ülker B, Somssich IE (2004) WRKY transcription factors: from DNAbinding towards biological function. Curr Opin Plant Biol 7: 491–498
Viaene T, Delwiche CF, Rensing SA, Friml J (2013) Origin and evolutionof PIN auxin transporters in the green lineage. Trends Plant Sci 18: 5–10
Wang C, Liu Y, Li SS, Han GZ (2014) Origin of plant auxin biosynthesis incharophyte algae. Trends Plant Sci 19: 741–743
Wang W, Esch JJ, Shiu SH, Agula H, Binder BM, Chang C, Patterson SE,Bleecker AB (2006) Identification of important regions for ethylenebinding and signaling in the transmembrane domain of the ETR1 ethylenereceptor of Arabidopsis. Plant Cell 18: 3429–3442
Wang ZY, Bai MY, Oh E, Zhu JY (2012) Brassinosteroid signaling networkand regulation of photomorphogenesis. Annu Rev Genet 46: 701–724
Wang ZY, Seto H, Fujioka S, Yoshida S, Chory J (2001) BRI1 is a critical com-ponent of a plasma-membrane receptor for plant steroids. Nature 410: 380–383
Wen X, Zhang C, Ji Y, Zhao Q, He W, An F, Jiang L, Guo H (2012) Acti-vation of ethylene signaling is mediated by nuclear translocation of thecleaved EIN2 carboxyl terminus. Cell Res 22: 1613–1616
Xu J, Zhang Y (2010) How significant is a protein structure similarity withTM-score = 0.5? Bioinformatics 26: 889–895
Yasumura Y, Crumpton-Taylor M, Fuentes S, Harberd NP (2007) Step-by-step acquisition of the gibberellin-DELLA growth-regulatory mecha-nism during land-plant evolution. Curr Biol 17: 1225–1230
Yasumura Y, Pierik R, Fricker MD, Voesenek LACJ, Harberd NP (2012)Studies of Physcomitrella patens reveal that ethylene-mediated submergenceresponses arose relatively early in land-plant evolution. Plant J 72: 947–959
Zhang K, Novak O, Wei Z, Gou M, Zhang X, Yu Y, Yang H, Cai Y, StrnadM, Liu CJ (2014) Arabidopsis ABCG14 protein controls the acropetaltranslocation of root-synthesized cytokinins. Nat Commun 5: 3274
Zhao LH, Zhou XE, Wu ZS, Yi W, Xu Y, Li S, Xu TH, Liu Y, Chen RZ,Kovach A, et al (2013) Crystal structures of two phytohormone signal-transducing a/b hydrolases: karrikin-signaling KAI2 and strigolactone-signaling DWARF14. Cell Res 23: 436–439
Zhou F, Lin Q, Zhu L, Ren Y, Zhou K, Shabek N, Wu F, Mao H, Dong W,Gan L, et al (2013) D14-SCF(D3)-dependent degradation of D53 regu-lates strigolactone signalling. Nature 504: 406–410
886 Plant Physiol. Vol. 167, 2015
Wang et al.