insight into the molecular basis of schistosoma ... · 3/3/2016 · original article insight into...
TRANSCRIPT
Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/297656950
InsightintothemolecularbasisofSchistosomahaematobium-inducedbladdercancerthroughurineproteomics
ARTICLEinTUMORBIOLOGY·MARCH2016
ImpactFactor:3.61·DOI:10.1007/s13277-016-4997-y
READS
19
10AUTHORS,INCLUDING:
JoséManuelCorreiadaCosta
NationalInstituteofHealthDr.RicardoJor…
78PUBLICATIONS1,241CITATIONS
SEEPROFILE
FranciscoAmado
UniversityofAveiro
168PUBLICATIONS2,126CITATIONS
SEEPROFILE
RitaFerreira
UniversityofAveiro
125PUBLICATIONS1,514CITATIONS
SEEPROFILE
LuciolaraSantos
InstitutoPortuguêsdeOncologia
118PUBLICATIONS1,116CITATIONS
SEEPROFILE
Allin-textreferencesunderlinedinbluearelinkedtopublicationsonResearchGate,
lettingyouaccessandreadthemimmediately.
Availablefrom:JoséManuelCorreiadaCosta
Retrievedon:15March2016

1 23
Tumor BiologyTumor Markers, Tumor Targeting andTranslational Cancer Research ISSN 1010-4283 Tumor Biol.DOI 10.1007/s13277-016-4997-y
Insight into the molecular basis ofSchistosoma haematobium-inducedbladder cancer through urine proteomics
Carina Bernardo, Maria Cláudia Cunha,Júlio Henrique Santos, José M. Correiada Costa, Paul J. Brindley, Carlos Lopes,et al.

1 23
Your article is protected by copyright and all
rights are held exclusively by International
Society of Oncology and BioMarkers (ISOBM).
This e-offprint is for personal use only
and shall not be self-archived in electronic
repositories. If you wish to self-archive your
article, please use the accepted manuscript
version for posting on your own website. You
may further deposit the accepted manuscript
version in any repository, provided it is only
made publicly available 12 months after
official publication or later and provided
acknowledgement is given to the original
source of publication and a link is inserted
to the published article on Springer's
website. The link must be accompanied by
the following text: "The final publication is
available at link.springer.com”.

ORIGINAL ARTICLE
Insight into the molecular basis of Schistosomahaematobium-induced bladder cancer through urine proteomics
Carina Bernardo1,2 & Maria Cláudia Cunha3 & Júlio Henrique Santos1,4 &
José M. Correia da Costa4,5 & Paul J. Brindley6 & Carlos Lopes1 & Francisco Amado3 &
Rita Ferreira3 & Rui Vitorino7,8 & Lúcio Lara Santos1,9
Received: 1 December 2015 /Accepted: 25 February 2016# International Society of Oncology and BioMarkers (ISOBM) 2016
Abstract Infection due to Schistosoma haematobium is car-cinogenic. However, the cellular and molecular mechanismsunderlying urogenital schistosomiasis (UGS)-induced carci-nogenesis have not been well defined. Conceptually, earlymolecular detection of this phenomenon, through non-invasive procedures, seems feasible and is desirable.Previous analysis of urine collected during UGS suggests thatestrogen metabolites, including depurinating adducts, may beuseful for this purpose. Here, a new direction was pursued: theidentification of molecular pathways and potential biomarkersin S. haematobium-induced bladder cancer by analyzing theproteome profiling of urine samples from UGS patients.GeLC-MS/MS followed by protein-protein interaction analy-
sis indicated oxidative stress and immune defense systemsresponsible for microbicide activity are the most representa-tive clusters in UGS patients. Proteins involved in immunity,negative regulation of endopeptidase activity, and inflamma-tion were more prevalent in UGS patients with bladder cancer,whereas proteins with roles in renal system process, sensoryperception, and gas and oxygen transport were more abundantin subjects with urothelial carcinoma not associated withUGS. These findings highlighted a Th2-type immune re-sponse induced by S. haematobium, which seems to be furthermodulated by tumorigenesis, resulting in high-grade bladdercancer characterized by an inflammatory response and com-plement activation alternative pathway. These findings
Carina Bernardo and Maria Cláudia Cunha contributed equally to thiswork.
Electronic supplementary material The online version of this article(doi:10.1007/s13277-016-4997-y) contains supplementary material,which is available to authorized users.
* Lúcio Lara [email protected]
1 Experimental Pathology and Therapeutics Group - Research Center,Portuguese Oncology Institute - Porto (IPO-Porto), Rua Dr. AntónioBernardino de Almeida, 4200-072 Porto, Portugal
2 Mass Spectrometry Group, QOPNA, Department of Chemistry,University of Aveiro, Campus Universitário de Santiago,3810-193 Aveiro, Portugal
3 School of Health Sciences, University of Minho, Campus Gualtar,4710-057 Braga, Portugal
4 Center for the Study of Animal Science, CECA/ICETA, Universityof Porto, Rua D. Manuel II, Apt 55142, 4051-401 Porto, Portugal
5 Center for Parasite Biology and Immunology (CIBP), NationalInstitute of Health Dr. Ricardo Jorge (INSA), Rua AlexandroHerculano, 4000-055 Porto, Portugal
6 Department of Microbiology, Immunology & Tropical Medicine,and Research Center for Neglected Diseases of Poverty, School ofMedicine & Health Sciences, George Washington University,Washington, DC 20037, USA
7 Department of Medical Sciences, Institute for Biomedicine –iBiMED, University of Aveiro, Campus Universitário de Santiago,Agra do Crasto, 3810-193 Aveiro, Portugal
8 Department of Physiology and Cardiothoracic Surgery, Faculty ofMedicine, University of Porto, 4200-072 Porto, Portugal
9 Department of Surgical Oncology, Portuguese Oncology Institute -Porto (IPO-Porto), Rua Dr. António Bernardino de Almeida,4200-072 Porto, Portugal
Tumor Biol.DOI 10.1007/s13277-016-4997-y
Author's personal copy

established a starting point for the development ofmultimarker strategies for the early detection of UGS-induced bladder cancer.
Keywords Urogenital schistosomiasis . Immune response .
Squamous cell carcinoma . GeLC-MS/MS . Urine proteomics
Introduction
More than 20% of cancers in the developing world are causedby infections [1]. The blood fluke Schistosoma haematobiumis of particular concern; infection with this worm, urogenitalschistosomiasis (UGS), is carcinogenic and classified as group1 carcinogen [2]. In addition to directly damaging the devel-opment, health, and prosperity of infected populations, infec-tion with these parasites leads to squamous cell carcinoma(SCC) of the urinary bladder [2–4]. Squamous cell carcinomais the common form of bladder cancer in rural Africa whereS. haematobium is prevalent [5, 6]. In contrast, the majority ofbladder cancer in developing countries and regions not en-demic for UGS is transitional (or urothelial) cell carcinoma(TCC) [7]. The epidemiologic association between SCC of thebladder with UGS is based both on case-control studies and onthe correlation of bladder cancer incidence with prevalence ofinfection with S. haematobium in different geographic loca-tions. UGS is usually a chronic disease; the adult, egg-producing schistosomes live for many years, re-infections oc-cur frequently, and UGS-SCC of the bladder appears relative-ly early, often by middle age (see [4]).
The cellular and molecular mechanisms linkingS. haematobium infection with cancer formation are not de-fined; however, recent findings implicate S. haematobium an-tigens as the direct cause of alterations in the urothelium inmice [8]. In addition, an estrogen-DNA adduct-mediated path-way may be involved in the carcinogenesis of the bladderassociated with S. haematobium infection (reviewed in [4,8]). These novel estrogen-like metabolites occur in the schis-tosome and in sera and urine during UGS [9]. LC-MS/MS anal-yses provide deeper insights into the carcinogenesis of UGS-induced bladder cancer and highlight potential biomarkers fordiagnosis and/or prognosis of this neglected tropical disease-linked malignancy [10]. Previous findings from animal studieswith chemically induced urothelial carcinoma highlight thepotential of mass spectrometry-based urine proteomics forthe identification of the biological processes underlying dis-ease pathogenesis [11]. This analysis supported the involve-ment of response to selenium and to drugs; neutral lipid me-tabolism to disease pathogenesis at earlier stages; and inflam-mation, immune response, and wound healing at advancedstages. In this sense, a novel exploratory approach was per-formed: a comprehensive analysis of urine proteome profile inthree groups of patients: with UGS, with UGS and bladder
cancer, and patients with bladder cancer without schistosomi-asis. Here, we present findings from a new approach, focusedon the urine proteome, targeting a better understanding ofcarcinogenesis due to infection with S. haematobium.
Materials and methods
Characterization of study population
Six patients from the archive of the ICETA-CECA R&D unitwere enrolled as participants in the present study. These studyparticipants were followed between December 2011 andSeptember 2012 at the Clínica da Sagrada Esperança and atthe Américo Boavida University Hospital, Luanda, Angolawhere they were diagnosed with UGS after microscopic iden-tification of eggs in the urine and cystoscopy examination, asdescribed elsewhere [12]. Three other participants of the pres-ent investigation, who had been diagnosed with urothelialcancer at IPO-Porto, Portugal, were also enrolled in the study.These latter cases had no history of S. haematobium infectionand were considered as a control group. Urine samples wereobtained according to a collaborative program between theseinstitutions and IPO-Porto after obtaining informed consentfrom all participants [10]. This study was approved by the localethics committee and followed the Declaration of Helsinki.
Participants were assigned to one of three groups: group S(patients with UGS and without bladder cancer), group S + T(patients with UGS and bladder cancer), and group T (patientswith urothelial cancer from geographical location non-endemic for UGS). Table 1 summarizes the demographicand clinical characteristics of the enrolled participants. Mostparticipants were female and those with UGS had ready andfrequent access to the river near their residence, the probablesource of transmission. Group S was characterized by youngerpatients (average age, 26 years) and presented chronic inflam-mation of the bladder although they did not have histologicalsigns of carcinoma of the urinary bladder. The average age ofgroup S + T was 52 years, and participants in this group pre-sented squamous cell carcinoma (SCC) of the bladder in ad-dition to chronic inflammation. Participants in both groups Sand S + T passed eggs of S. haematobium in the urine, andcystoscopy exhibited schistosome egg deposition and granu-lomatous lesions in the bladder as shown in Fig. 1. The meanage of participants in group T was 70 years; these personspresented high-grade urothelial carcinoma of the bladder andinflammatory changes. All the enrolled participants had a his-tory of hematuria.
Sample collection and preparation
During patient interviews, the participants provided one sam-ple of urine of ~50 mL, collected mid-stream during
Tumor Biol.
Author's personal copy

micturition. The urine was subjected centrifugation for 2 min,at 4000×g, 4 °C; after which, the supernatant was passedthrough 10-kDa filters (Vivaspin, Sartorius) in order to con-centrate samples and remove salts. The retentate was resus-pended in 100μL of 6M urea, 2M thiourea, 1%CHAPS, and0.5 % SDS. Protein content was estimated using the RC-DCassay kit (Bio-Rad).
Urinary proteome characterizationthrough GeLC-MS/MS
Proteins in the urines were analyzed as described [13]. Briefly,20 μg protein was separated by SDS-PAGE, using 12.5 %polyacrylamide gels, stained with BlueSafe® solution; afterwhich, gel lanes were cut into 16 fragments for trypsin diges-tion. The resulting peptide mixture was extracted from the gelfractions, lyophilized in a speed vacuum centrifuge, resus-pended in 20 μL 0.1 % trifluoroacetic acid/5 % acetonitrile,and loaded onto a Pepmap100 capillary analytical C18 col-umn with 3 mm particle size (Dionex/LC Packings) at a flowrate of 300 nL/min (Ultimate 3000 (Dionex)). Peptides elutedfrom the column were mixed with a matrix solution (2 mg/mLα-cyano-4-hydroxycinnamic acid in 70 % acetonitrile/0.3 %
trifluoroacetic acid in a fraction micro-collector (Probot;Dionex/LC Packings) and directly deposited onto the liquidchromatography-MALDI plates. Subsequently, MALDIplates were analyzed using a 4800 MALDI-time of flight(TOF)/TOF Analyzer (ABSCIEX). A signal/noise thresholdof 50 was used to select peaks for MS/MS analysis. The spec-tra were processed using the TS2Mascot (v1.0; MatrixScience Ltd); after which, peptide/protein identification wasdetermined using Mascot, v.2.1.0.4 (Matrix Science Ltd).Searches were performed in Mascot using Homo sapiens re-lease 13102014. A MS tolerance of 30 ppm was found forprecursor ions and 0.3 Da for fragment ions, as well as twomissed cleavages and methionine oxidation as a variable mod-ification. Confidence levels of ≥95 % were accepted as a pos-itive identification for the protein. A minimal Mascot peptidescore of 30 was determined by a reverse database search,which revealed a false-positive rate <5 % for identifiedproteins.
The abundance of identified proteins was estimated by cal-culating the emPAI [14]. The theoretically observable peptideswere determined by the in silico digestion of mature proteinsusing the output of the program Protein Digestion Simulator,http://panomics.pnnl.gov/software/. The observed peptides
Table 1 Demographic and clinical characterization of the participants enrolled in the study
Age Sex Benign/pre-malignant lesions Tumor type
Group S (N= 3) 19 M Chronic inflammation –
40 F Chronic inflammation –
18 F Chronic inflammation –
Group S + T (N= 3) 48 F Chronic inflammation, hyperplasia High-grade SCC (pT2/3NxMx)
46 F Chronic inflammation, epidermoid metaplasia High-grade SCC (pT2/3NxMx)
63 F Chronic inflammation, epidermoidmetaplasia
High-grade SCC (pT2/3NxMx)
Group T (N= 3) 55 M Inflammation and regenerative changes High-grade urothelial carcinoma(pT2bN0M0), CIS
65 M Inflammation High-grade urothelial carcinoma (pT2bN1M0)
82 F Granulomatous inflammation Urothelial carcinoma (pT2bN0M0)
Group S patients infected with Schistosoma haematobium,Group S + T patients infected with S. haematobium and with bladder cancer,Group T patientswith bladder cancer without schistosome infection
Fig. 1 Representative image ofschistosome lesions in the bladderobserved through cystoscopy (a)and bladder ultrasound (b)
Tumor Biol.
Author's personal copy

were unique parent ions including those with two missedcleavages. Mean protein emPAI values were log2transformed for protein ratio calculation.
Results
In order to characterize the profile of urinary proteins, GeLC-MS/MSwas performed and a total of 535 distinct proteins wereretrieved with a p≤0.05 (Supplementary Table S1). Accordingto the PANTHER bioinformatic tool, most of these proteinswere associated with binding, catalytic, structural molecule,and receptor activities and were involved in biological process-es including metabolic, cellular, developmental, response tostimulus, localization, and immune response processes.
Among all proteins detected in the urines, only 74 werecommon to the three groups of patients (Fig. 2); these includedalbumin, protein AMBP, and S100A9. Curiously, uromodulinand osteopontin, typically found in the urine [13], were notidentified in group S + T. Normalized abundance analysis ofcommon proteins between group S/group S + T evidencedintestinal maltase-glucoamylase (MGA), plasma serine prote-ase inhibitor (IPSP), leukocyte elastase inhibitor (ILEU), andmyeloperoxidase (MPO) as the top four abundant in group S.Cystatin-C (CYTC), apolipoprotein A-I (APOA1), haptoglo-bin (HPT), and alpha-1-acid glycoprotein 1 (A1AG1) werethe most abundant proteins in group S + T (Fig. 3a). The
analysis of common proteins between group S + T/group Trevealed that cytochrome c (CYC), protein S100-A9, andalpha-1-antitrypsin (A1AT) were the most abundant proteinsin urine of the group S + T. Hemoglobin subunit beta (HBB),zinc-alpha-2-glycoprotein (ZA2G), and kininogen-1 (KNG1)were the most abundant proteins detected in group T (Fig. 3b).
Protein-protein interaction analysis of common proteins togroup S and group S + T indicated an upregulation ofmyeloperoxidase (MPO), cathelicidin antimicrobial peptide(CAMP), lactotransferrin (LTF), and azurocidin (AZU1)(green nodes) and a downregulation of the proteins involvedin negative regulation of endopeptidase activity, acute inflam-matory response, and platelet degranulation (red nodes) in theurine of participants of group S (Fig. 4a).
Protein-protein interaction analysis of common proteins togroup S + Tand group Tmainly demonstrated an upregulationof proteins involved in negative regulation of glycoproteinmetabolic process, acute inflammatory response, positive reg-ulation of protein secretion (S100 calcium binding protein A8(S100A8) and S100 calcium binding protein A9 (S100A9)),regulation of immune response in group S + T (green nodes),and a downregulation of renal system processes and gas trans-port (mutant hemoglobin alpha 2 globin chain (HBA2) andkininogen-1 (KNG1)) (red nodes) (Fig. 4b).
Seventeen proteins were only identified in the urine of theschistosome-infected individuals: these proteins are mainlyinvolved in B and T cell activation, apoptosis signaling, andinflammation mediated by chemokine and cytokine signalingpathway (Supplementary Figure S2—blue nodes). In the urineof individuals with bladder cancer and UGS, there were 42exclusive proteins mainly involved in metabolic, cellular, andimmune system processes such as complement activation(adipsin-CFD; complement factor B—CFB; complement fac-tor H—CFH; complement component 9—C9), leukocyte-mediated cytotoxicity, acute inflammatory response, regula-tion of inflammatory response, regulation of macrophage celldifferentiation, and regulation of endopeptidase activity(Supplementary Figure S2—yellow nodes). Twenty-five pro-teins were only identified in the urine of the patients withbladder cancer without UGS. Structural molecules and cata-lytic activity are their most significant molecular functions.Concerning biological processes, they were mainly associatedwith metabolic, cell adhesion (cadherin family), tumor cellmetastasis (lysosomal associated membrane protein 2—LAMP2), regulation of tumor growth (peroxiredoxin 2—PRDX2; vasorin—VASN) and immune response activities(Supplementary Figure S2—pink nodes).
Fig. 2 Venn diagram representing the distribution of proteins identifiedin the urine samples per group:Group S schistosome infection,Group S +T schistosome infection and bladder cancer, Group T bladder cancerwithout schistosome infection) (http://bioinfogp.cnb.csic.es/tools/venny/)
�Fig. 3 Normalized abundance of urinary proteins in group S and group S+ T (a) and in group S + T and T (b). Group S schistosome infection,Group S + T schistosome infection and bladder cancer, Group T bladdercancer without schistosome infection. Protein accession number hascorrespondence to protein name in Supplementary Table S1
Tumor Biol.
Author's personal copy

Tumor Biol.
Author's personal copy
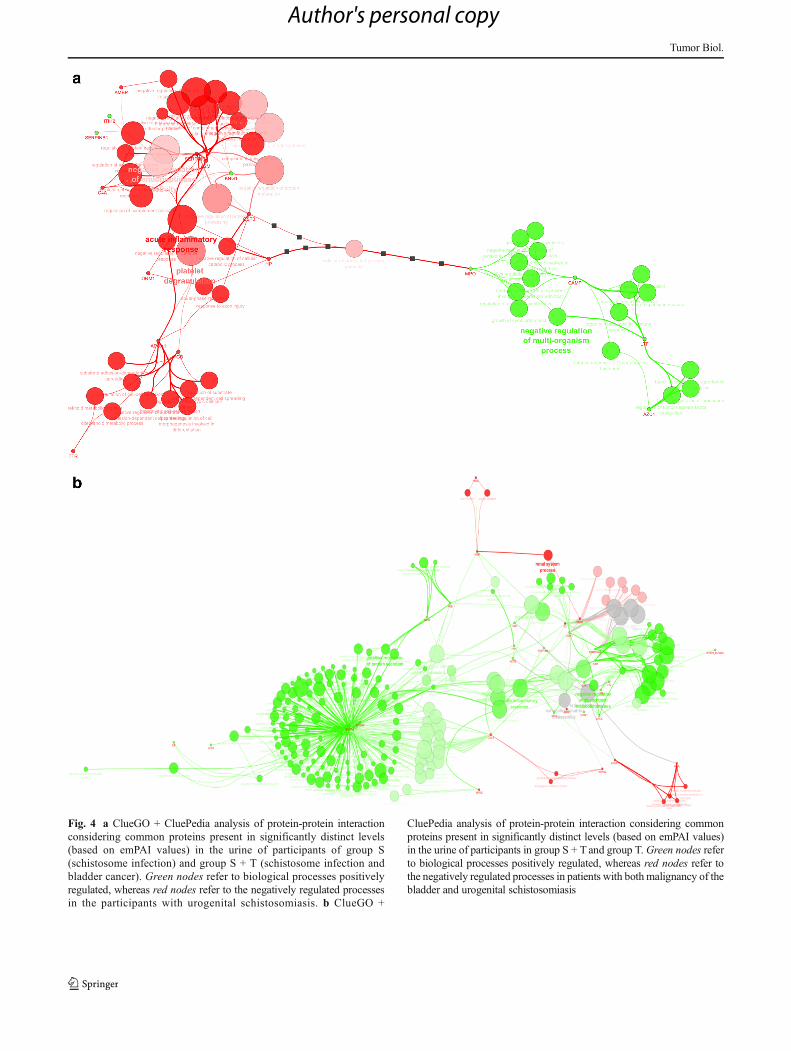
Fig. 4 a ClueGO + CluePedia analysis of protein-protein interactionconsidering common proteins present in significantly distinct levels(based on emPAI values) in the urine of participants of group S(schistosome infection) and group S + T (schistosome infection andbladder cancer). Green nodes refer to biological processes positivelyregulated, whereas red nodes refer to the negatively regulated processesin the participants with urogenital schistosomiasis. b ClueGO +
CluePedia analysis of protein-protein interaction considering commonproteins present in significantly distinct levels (based on emPAI values)in the urine of participants in group S + T and group T. Green nodes referto biological processes positively regulated, whereas red nodes refer tothe negatively regulated processes in patients with bothmalignancy of thebladder and urogenital schistosomiasis
Tumor Biol.
Author's personal copy

Discussion
In order to expand our understanding of disease pathogenesisand to envision potential disease biomarkers for the early di-agnosis of UGS and UGS-induced bladder cancer, profiling ofthe urine proteome was performed using a GeLC-MS/MSapproach. Urine is a desirable and accessible biofluid becauseof the comparative non-invasiveness and simplicity of speci-men collection and previous data on the identification of pu-tative disease biomarkers using MS-based proteomics [13].
Urine proteome profile modulated by infectionwith S. haematobium
The proteins exclusively assigned to study participants withUGS but free of bladder cancer (group S) were predicted tobe mainly involved in leukocyte-mediated cytotoxicity, regula-tion of inflammatory response, and regulation of cytokine bio-synthetic processes. Indeed, the expression of nuclear factor ofkappa light polypeptide gene enhancer in B-cells 1 (NF-κB1),one of the singular proteins detected in this group, is known tobe required within dendritic cells for optimal Th2 responsesfollowing exposure to eggs of Schistosoma mansoni, a potentand natural Th2-inducing stimulus [15]. A similar mechanismseems to be present during UGS. Immune cells and their cyto-kines have been associated with carcinogenesis and T cell-infiltrating tumors correlated with patient outcome [16]. In ad-dition, neutrophil-expressed elastase (ELANE) was anotherprotein only detected in this group. Elastases are effectors usedby neutrophils, eosinophils, and macrophages to attack devel-opmental stages of S. mansoni. These cells attach to the surfaceof the parasites and secrete substances such as cationic proteins,hydrolytic enzymes, and oxidants that cause damage to theschistosomes [17]. Detection of ELANE in the urine duringUGS raises the possibility that the immune response during thisdiscrete schistosome infection triggers the same mechanismknown from Schistosoma mansoni infection. Furthermore, theprotein roundabout homolog 4 (Robo4) was only detected inthis group of patients. Robo4 functions as a receptor for Slitproteins that participate in vascular patterning and angiogenesis[18]. Proteins associated with oxidative stress including cata-lase and glutathione peroxidase 3 were also detected. As de-scribed for infection with S. mansoni, the main organs affectedin this disease are shifted to a pro-oxidant state during thecourse of the infection [19, 20], which could explain the detec-tion of these oxidative stress parameters in the urine of partic-ipants with UGS.
Significant higher levels of myeloperoxidase (MPO),cathelicidin antimicrobial peptide (CAMP), lactotransferrin(LTF), and azurocidin (AZU1) were also found in the urineof patients from group S (Fig. 3a). These proteins share a rolein non-specific immune defense systems responsible for mi-crobicide activity [21–23] and are associated with diverse
cancers [24–26], but, to the extent of our knowledge, directassociation with bladder cancer has not been established.CAMP might also modulate the innate immune response totissue injury and wound healing [27, 28]. Helminthic parasitessuppress the classical pro-inflammatory responses and inducea Th2-type immune response that rapidly and effectively healstissue. However, the outcome of this type of immune responseis a tolerance by the host for the parasite that remains in situfor many years. This facilitates successfully parasitism by theschistosome including continuous release of schistosome ova[29], increasing the risk of bladder cancer that is detected at anaverage age of 50 years. Azurocidin, also known as heparin-binding protein (HBP) and CAP37, is released from secretoryvesicles and azurophilic granules of activated neutrophils. Ithas antimicrobial activity, mainly against Gram-negative bac-teria [30]. Azurocidin in the urine has been reported duringurinary tract infection in children [31].
Urine proteome profile modulated by urogenitalschistosomiasis-related bladder cancer
Integrated analysis, undertaken with the assistance of theClueGo + CluePedia bioinformatic tool, of unique proteinsidentified in the urine of patients with UGS as well as blad-der cancer (group S + T) revealed an upregulation of acuteinflammatory response and complement activation alterna-tive pathway. Complement factor H (CFH), a properdin in-hibitor, has the principal function of regulating the alterna-tive pathway of the complement system and has beenappointed as a bladder cancer marker [32]. Complementcomponent 9 (C9) was also detected in this group. C9 is aconstituent of the membrane attack complex (MAC) thatplays a key role in the innate and adaptive immune responseby forming pores in the plasma membrane of target cells. Onthe other hand, neither CFH nor C9 occurred in the urine ofpatients in group T (with bladder cancer but without UGS),which suggested that different immune responses might beinfluential for chemically induced bladder cancer comparedto UGS-induced bladder cancer. Also, C-reactive protein(CRP) was only detected in this group, supporting the oc-currence of inflammation in these patients. In this group, webelieve that inflammation is not a consequence of cancersince UGS-induced bladder cancer follows the long-termgranulomatous inflammatory reaction to eggs of the parasitelodged in the bladder wall. Instead, inflammation is an earlyevent in this disease pathogenesis. Furthermore, most ofthese exclusive proteins are associated with inflammationmediated by chemokine, cytokine signaling pathways, epi-dermal growth factor (EGF), and fibroblast growth factor(FGF) signaling pathways. EGF and FGF activate down-stream signaling pathways including RAS/MAPK that playkey roles in bladder cancer cell proliferation [33, 34].
Tumor Biol.
Author's personal copy

Urine proteome profile modulated by bladder cancer
Bioinformatic analysis of the unique proteins identified in theurine of bladder cancer patients without schistosome infection(group T) indicated the involvement of these proteins in met-abolic, cellular, and immune system processes. Proteins suchas lysosomal-associated membrane protein 2 (LAMP2),peroxiredoxin 2 (PRDX2), and cadherin 1 (CADH1) playpivotal roles in blood coagulation and cadherin signaling path-ways implicated in tumor cell metastasis, signaling cascadesof growth factors, and tumor necrosis factor-α and are in-volved inmechanisms regulating cell-cell adhesions, mobility,and proliferation of epithelial cells [35–37].
Two of the eight proteins common to groups S + T and Twere identified as S100A8 and S100A9. Although the role ofS100 proteins in carcinogenesis is poorly understood, theyappear to be essential for the development and progressionof many cancers through cell survival and apoptosis path-ways. In animal models of bladder cancer, several S100 pro-teins were found differentially expressed in bladder tumorsrelative to healthy urothelium [38]. The upregulation of thesetwo proteins in addition to α-2-macroglobulin (A2MG),cystatin 3 (CST3), complement C4-A (C4A), and others sug-gested an increase of the inflammatory response in the S + Tpatients when compared to the group T. Downregulation ofkininogen-1 was detected in the urine of group S + T partic-ipants. Kininogens are inhibitors of thiol proteases and HMW-kininogen plays a pivotal role in blood coagulation by helpingto position optimally prekallikrein and factor XI next to factorXII. Along with the downregulation of plasma serine proteaseinhibitor also seen in this group, it might reflect the involve-ment of coagulation in the tumor microenvironment, as hasbeen previously suggested [39].
Taken together, these new findings indicated that UGS rep-resents a pathogenic stimulus that induces oxidative stress andtriggers a Th2-type immune response. The Th2 phenotypeincludes upregulation of host antimicrobial peptides andNF-κB, as is induced by antigens such as the omega-1 factorsecreted by the egg granuloma during infection withS. mansoni [40]. These events play an important though notcompletely understood role in carcinogenesis, as they seem toincrease the risk of high-grade bladder cancer that is typicallydiagnosed in middle-aged individuals with a history of chron-ic UGS [41]. Inflammation and activation of the alternativecomplement pathway are the biological processes upregulatedin this situation. A prolonged inflammatory response mightlead to increased DNA mutation rates, due to previous alter-ations in suppressor genes such as TP53 (expressed by accu-mulation of p53 in the urothelium) and overall genetic insta-bility, characterized by high levels of 8-nitroguanine and 8-hydroxy-2′-deoxyguanosine [10, 42, 43]. Our findings alsosupport the hypothesis, as recently proposed [16], that mostcancers originate from a biological or chemical stimulus and
are followed by chronic inflammation, fibrosis, and a changein the tissue microenvironment that leads to a pre-cancerousniche. Future work involving a larger cohort of participantswill be important to disclose the putative diagnostic value ofmultimarker strategies including proteins from these upregu-lated biological processes, such as azurocidin, CAMP,NF-κB1, CFH, MPO, C9, and CRP for the early diagnosisof urogenital schistosomiasis-induced bladder cancer.
Acknowledgments This work was supported by the PortugueseFoundation for Science and Technology (FCT), European Union,QREN, FEDER, and COMPETE for funding the QOPNA; by iBiMEDresearch unit (project PEst-C/QUI/UI0062/2013, UID/BIM/04501/2013,UID/IC/00051/2013, and COST action BM1305) and PhD fellowshipSFRH/BD/80855/2011 (CB); and by the Portuguese Mass SpectrometryNetwork (RNEM). The authors also acknowledge Clínica SagradaEsperança and Serviço de Urologia do Hospital Américo Boavida fromLuanda, Angola.
Compliance with ethical standards Urine samples were obtained ac-cording to a collaborative program between Clínica da SagradaEsperança, Américo Boavida University Hospital and IPO-Porto afterobtaining informed consent from all participants. This study was ap-proved by the local ethics committee and followed the Declaration ofHelsinki.
Conflicts of interest None
References
1. De Martel C, Ferlay J, Franceschi S, et al. Global burden of cancersattributable to infections in 2008: a review and synthetic analysis.Lancet Oncol. 2012;13:607–15. doi:10.1016/S1470-2045(12)70137-7.
2. IARC A review of human carcinogens. Part B: biological agents.In: IARC Work. Gr. Eval. Carcinog. Risks to humans. Lyon,France; 2012. p. 371–384.
3. Pagano JS, Blaser M, Buendia M-A, et al. Infectious agents andcancer: criteria for a causal relation. Semin Cancer Biol. 2004;14:453–71. doi:10.1016/j.semcancer.2004.06.009.
4. Brindley PJ, da Costa JMC, Sripa B.Why does infection with somehelminths cause cancer? Trends Cancer. 2015. doi:10.1016/j.trecan.2015.08.011.
5. Mostafa MH, Sheweita SA, O’Connor PJ. Relationship betweenschistosomiasis and bladder cancer. Clin Microbiol Rev. 1999;12:97–111.
6. Zhong X, Isharwal S, Naples JMM, et al. Hypermethylation ofgenes detected in urine from Ghanaian adults with bladder pathol-ogy associated with Schistosoma haematobium infection. PLoSOne. 2013;8:e59089. doi:10.1371/journal.pone.0059089.
7. Kaufman DS, Shipley WU, Feldman AS. Bladder cancer. Lancet.2009;374:239–49. doi:10.1016/S0140-6736(09)60491-8.
8. Botelho MC, Machado JC, Brindley PJ, Correia da Costa JM.Targeting molecular signaling pathways of Schistosomahaematobium infection in bladder cancer. Virulence. 2011;2:267–79.
9. Correia da Costa JM, Vale N, Gouveia MJ, et al. Schistosome andliver fluke derived catechol-estrogens and helminth associated can-cers. Front Genet. 2014;5:444. doi:10.3389/fgene.2014.00444.
Tumor Biol.
Author's personal copy

10. Gouveia MJ, Santos J, Brindley PJ, et al. Estrogen-like metabolitesand DNA-adducts in urogenital schistosomiasis-associated bladdercancer. Cancer Lett. 2015;359:226–32. doi:10.1016/j.canlet.2015.01.018.
11. Ferreira R, Oliveira P, Martins T, et al. Comparative proteomicanalyses of urine from rat urothelial carcinoma chemically inducedby exposure to N-butyl-N-(4-hydroxybutyl)-nitrosamine. MolBiosyst. 2015;11:1594–602. doi:10.1039/c4mb00606b.
12. Santos J, Chaves J, Araújo H, et al. Comparison of findings usingultrasonography and cystoscopy in urogenital schistosomiasis in apublic health centre in rural Angola. S Afr Med J. 2015;105:312–5.doi:10.7196/SAMJ.8564.
13. Caseiro A, Barros A, Ferreira R, et al. Pursuing type 1 diabetesmellitus and related complications through urinary proteomics.Transl Res. 2014;163:188–99. doi:10.1016/j.trsl.2013.09.005.
14. Ishihama Y. Exponentially modified protein abundance index(emPAI) for estimation of absolute protein amount in proteomicsby the number of sequenced peptides per protein. Mol CellProteomics. 2005;4:1265–72. doi:10.1074/mcp.M500061-MCP200.
15. Artis D, Kane CM, Fiore J, et al. Dendritic cell-intrinsic expressionof NF-B1 is required to promote optimal Th2 cell differentiation. JImmunol. 2005;174:7154–9. doi:10.4049/jimmunol.174.11.7154.
16. Brücher BL, Jamall IS. Epistemology of the origin of cancer: a newparadigm. BMC Cancer. 2014;14:331. doi:10.1186/1471-2407-14-331.
17. Freudenstein-Dan A, Gold D, Fishelson Z. Killing of schistosomesby elastase and hydrogen peroxide: implications for leukocyte-mediated schistosome killing. J Parasitol. 2003;89:1129–35. doi:10.1645/GE-96R.
18. Park KW, Morrison CM, Sorensen LK, et al. Robo4 is a vascular-specific receptor that inhibits endothelial migration. Dev Biol.2003;261:251–67. doi:10.1016/S0012-1606(03)00258-6.
19. de Oliveira RB, Senger MR, Vasques LM, et al. Schistosomamansoni infection causes oxidative stress and alters receptor foradvanced glycation endproduct (RAGE) and tau levels in multipleorgans in mice. Int J Parasitol. 2013;43:371–9. doi:10.1016/j.ijpara.2012.12.006.
20. Pascal M, Abdallahi OM, Elwali NE, et al. Hyaluronate levels andmarkers of oxidative stress in the serum of Sudanese subjects at riskof infection with Schistosoma mansoni. Trans R Soc Trop MedHyg. 2000;94:66–70.
21. Kajer TB, Fairfull-Smith KE, Yamasaki T, et al. Inhibition ofmyeloperoxidase- and neutrophil-mediated oxidant production bytetraethyl and tetramethyl nitroxides. Free Radic Biol Med.2014;70:96–105. doi:10.1016/j.freeradbiomed.2014.02.011.
22. Khan AA, Rahmani AH, Aldebasi YH, Aly SM. Biochemical andpathological studies on peroxidases—an updated review. Glob JHealth Sci. 2014;6:35689. doi:10.5539/gjhs.v6n5p87.
23. Yang D, Chertov O, Oppenheim JJ. Participation of mammaliandefensins and cathelicidins in anti-microbial immunity: receptorsand activities of human defensins and cathelicidin (LL-37). JLeukoc Biol. 2001;69:691–7.
24. Castillo-Tong DC, Pils D, Heinze G, et al. Association ofmyeloperoxidase with ovarian cancer. Tumor Biol. 2014;35:141–8. doi:10.1007/s13277-013-1017-3.
25. Shaheduzzaman S, Vishwanath A, Furusato B, et al. Silencing oflactotransferrin expression by methylation in prostate cancer pro-gression. Cancer Biol Ther. 2007;6:1088–95. doi:10.4161/cbt.6.7.4327.
26. Gill K, Mohanti BK, Singh AK, et al. The over expression ofcathelicidin peptide LL37 in head and neck squamous cell carcino-ma: the peptide marker for the prognosis of cancer. CancerBiomark. 2011;10:125–34. doi:10.3233/CBM-2012-0238.
27. Cotton S, Donnelly S, Robinson MW, et al. Defense peptides secret-ed by helminth pathogens: antimicrobial and/or immunomodulatormolecules? Front Immunol. 2012;3:269. doi:10.3389/fimmu.2012.00269.
28. da Costa JP, Cova M, Ferreira R, Vitorino R. Antimicrobial pep-tides: an alternative for innovative medicines? Appl MicrobiolBiotechnol. 2015;99:2023–40. doi:10.1007/s00253-015-6375-x.
29. Thivierge K, Cotton S, Schaefer DA, et al. Cathelicidin-like hel-minth defence molecules (HDMs): absence of cytotoxic, anti-microbial and anti-protozoan activities imply a specific adaptationto immunemodulation. PLoSNegl TropDis. 2013;7:e2307. doi:10.1371/journal.pntd.0002307.
30. Gabay JE, Scottt RW, Campanelli D, et al. Antibiotic proteins ofhuman polymorphonuclear leukocytes. Proc Natl Acad Sci U S A.1989;86:5610–4.
31. Kjölvmark C, Åkesson P, Linder A. Elevated urine levels ofheparin-binding protein in children with urinary tract infection.Pediatr Nephrol. 2012;27:1301–8. doi:10.1007/s00467-012-2132-x.
32. Cheng Z-Z. Complement factor H as a marker for detection ofbladder cancer. Clin Chem. 2005;51:856–63. doi:10.1373/clinchem.2004.042192.
33. di Martino E, Tomlinson DC, Knowles MA. A decade of FGFreceptor research in bladder cancer: past, present, and future chal-lenges. Adv Urol. 2012;2012:1–10. doi:10.1155/2012/429213.
34. Izumi K, Zheng Y, Li Y, et al. Epidermal growth factor inducesbladder cancer cell proliferation through activation of the androgenreceptor. Int J Oncol. 2012;41:1587–92. doi:10.3892/ijo.2012.1593.
35. Kanao H. Overexpression of LAMP3/TSC403/DC-LAMP pro-motes metastasis in uterine cervical cancer. Cancer Res. 2005;65:8640–5. doi:10.1158/0008-5472.CAN-04-4112.
36. Li G, Liu Y, Yin H, et al. E-cadherin gene promoter hypermethyla-tion may contribute to the risk of bladder cancer among Asianpopulations. Gene. 2014;534:48–53. doi:10.1016/j.gene.2013.10.027.
37. Wang T, Diaz AJ, Yun Y. The role of peroxiredoxin II inchemoresistance of breast cancer cells. Breast Cancer TargetsTher. 2014;6:73. doi:10.2147/BCTT.S61281.
38. Gebhardt C, Németh J, Angel P, Hess J. S100A8 and S100A9 ininflammation and cancer. Biochem Pharmacol. 2006;72:1622–31.doi:10.1016/j.bcp.2006.05.017.
39. Chen Y-T, Chen C-L, Chen H-W, et al. Discovery of novel bladdercancer biomarkers by comparative urine proteomics using iTRAQtechnology. J Proteome Res. 2010;9:5803–15. doi:10.1021/pr100576x.
40. Everts B, Perona-Wright G, Smits HH, et al. Omega-1, a gly-coprotein secreted by Schistosoma mansoni eggs, drives Th2responses. J Exp Med. 2009;206:1673–80. doi:10.1084/jem.20082460.
41. Salem HK, Mahfouz S. Changing patterns (age, incidence, andpathologic types) of Schistosoma-associated bladder cancer inEgypt in the past decade. Urology. 2012;79:379–83. doi:10.1016/j.urology.2011.08.072.
42. Warren W, Biggs PJ, El-Baz M, et al. Mutations in the p53 gene inschistosomal bladder cancer: a study of 92 tumours from Egyptianpatients and a comparison between mutational spectra from schis-tosomal and non-schistosomal urothelial tumours. Carcinogenesis.1995;16:1181–9.
43. Santos J, Fernandes E, Ferreira J, et al. P53 and cancer-associatedsialylated glycans are surrogate markers of cancerization of thebladder associated with Schistosoma haematobium infection.PLoS One. 2014. doi:10.1371/journal.pntd.0003329.
Tumor Biol.
Author's personal copy