inhibition of the bacteriolytic effect of β-lactam-antibiotics on staphylococcus aureus by the...
TRANSCRIPT

APMIS 98: 7 1-8 1, 1990
Inhibition of the bacteriolytic effect of P-lactam-antibiotics on Staphylococcus aureus by
the polyanionic drugs suramin and Evans Blue*
JORG WECKE, MARITA FRANZ and PETER GIESBRECHT
Robert Koch-Institut des Bundesgesundheitsamtes, Berlin, Germany
Wecke, J., Franz, M. & Giesbrecht, P. Inhibition of the bacteriolytic effect of P-lactam-antibiotics on Staphylococcus aureus by the polyanionic drugs suramin and Evans Blue. APMIS 98: 7 1-8 1, 1990.
The anionic polyelectrolytes suramin and Evans Blue inhibited different autolytic systems involved in wall growth and wall turnover of growing staphylococci and in wall autolysis of resting bacteria. Moreover, both substances lowered the P-lactam-induced pre-lytic release of cytoplasmic constituents from staphylococci, and inhibited the P-lactam-induced bacteriolysis as well as the loss of viability. The protective effects of these sulfonated drugs against bacteriolysis were also monitored by electron microscopy. Some medical implications of our results are discussed.
Key words: Staphylococcus aureus ; bacteriolysis; inhibition; penicillin; latamoxefi suramin; Evans Blue.
Jorg Wecke, Robert Koch-Institut, Nordufer 20, D-1000 Berlin 65, Federal Republic of Germany.
In previous studies it was demonstrated that anion- ic polyelectrolytes, such as certain anticoagulants, were effective in reducing the release of a specific wall label from bacteria (Ginsburg & Sela 1976). This inhibition was recently found to be caused by an interaction between the anticoagulants and the staphylococcal cell wall ( Wecke et al. 1986). It was also shown that such anionic anticoagulants were capable of suppressing the penicillin-induced bac- teriolysis ( Wecke et al. 1987).
Another substance belonging to the same group of negatively charged compounds is suramin. Apart from its therapeutic effect against try- panosomiasis and onchocerciasis, this drug repre- sents a potent competitive inhibitor of the reverse transcriptase of a number of animal retroviruses
Received December 12, 1988. Accepted August 1, 1989. *Dedicated to Prof. Dr. Georg Henneberg on the occa- sion of his 80th birthday.
(de Clercq 1979). Its virustatic activity was ascribed to its high content of sulfonic acid residues (Chan- dra et al. 1985). Recently, suramin was shown to inhibit in vitro the infectivity and the cytopathic effect of HIV (Mitsuya et al. 1984). Likewise, a virustatic in vivo effect of suramin was demon- strated in a clinical study with AIDS-patients (Broder et al. 1985). The anionic dye Evans Blue, which is structurally related to suramin, was de- scribed as an inhibitor of the cytopathic effect of HIV on ATH8 cells, too (Balzarini et al. 1986b).
So far, however, virtually nothing is known about the possible simultaneous interference of suramin or Evans Blue with the action of those antibiotics which have to be used at high doses against opportunistic bacterial infections during advanced stages of AIDS. Therefore, the aim of this study was to investigate whether suramin and Evans Blue would interact with untreated staphy- lococci and would interfere with the action of some j3-lactam antibiotics on staphylococci.
71

WECKE et al.
MATERIALS AND METHODS
Bacterial strains and cultivation Staphylococcus aureus strain SG 5 1 1 Berlin from the
culture collection of the Robert Koch-Institute was used throughout. The bacteria were transferred from agar plates to flasks containing peptone broth (2.5% peptone) and cultivated for 22 h at 37 "C. Then the suspensions were diluted with fresh medium and, after growth for another 2 h at 37 "C under shaking, the resulting log-phase cultures were adjusted to an optical density at 578 nm (OD) of 0.5 by addition of fresh medium. Penicillin G was added to some of these cultures to establish a concentration of 0.1 Fg penicillin per ml. This drug concentration corresponded to 5 times the MIC and had earlier proved optimal for rapid bacteriolysis with the strain in use. Bacteriolysis was almost complete after 4 - 5 h. Bacterial growth was monitored by an "Eppendorf" photometer and the cells were harvested by a Sorvall RC 2B centrifuge ( 1 0,000 x g for 15 rnin).
Chemicals Penicillin G (Serva, FRG) amoxycillin (Becham, UK)
and latamoxef (a gift from Dr T. Nishino, Kyoto) were the p-lactam compounds used. Suramin was purchased from Bayer AG (Leverkusen, FRG), Evans Blue (EB) from Merck (Darmstadt, FRG) and lysostaphin from Sigma (Tauflcirchen, FRG). 14C-N-acetylglucosamine and 'H-uracil were obtained from Amersham/Buchler (Braunschweig, FRG).
Assay of viable counts Viability measurements were performed using a spiral
plater model CU (Spiral System Instruments Inc., USA).
Labeling of intracellular components of staphylococci Bacteria from the logarithmic growth phase were
transferred to 25 ml fresh peptone broth, containing 150 p1 (5,6-3H)-uracil (37 M Bq/ml). After four division cycles (about 120 rnin), the bacteria were harvested by centrifugation and washed with peptone broth.
Aliquots were withdrawn from the cultures and after centrifugation (12,000 x g for 5 min) supernatants were monitored by a liquid scintillation counter (Rackbeta, LKB, Sweden).
Labeling of cell walls Cell wall labeling of staphylococci with I4C-N-acetyl-
glucosamine ( I4C-NAGA) and radioactivity measure- ments were performed as described in a previous study ( Wecke et al. 1982).
Isolation and fractionation of staphylococcal cell walls After cultivation for 1 h or 22 h, bacteria were har-
vested by centrifugation at 10,000 x g for 10 min. After washing with saline, the bacterial cells were disintegrated for 3 - 6 rnin in a "Dyno-mill" TYPE KDL (Wily A. Bachofen, Basel, Switzerland) filled with glass beads. The temperature was kept below 4 "C. After separation from the glass beads by glass filters, the cell debris was shaken
72
for 30 rnin at 60 "C in a sodium dodecylsulfate solution to remove the cytoplasmic membranes. After five wash- ings with water, the debris was incubated with trypsin (0.2 mg/ml) in 0.15 M Trisbuffer for 48 h at 37 "C.
The inhibition of lysostaphine-induced lysis of staphylo- coccal cell walls
The radiolabelled walls of staphylococci suspended in buffer (ammonium acetate pH 6.9) were pretreated with suramin (250 pg/ml) or EB (100 pg/ml) for 1 h at 4 "C. Afterwards cell walls were washed twice with ammoni- um acetate buffer. Following this treatment, the EB- treated cell walls appeared as a blue colored pellet after centrifugation (10,000 x g , 15 rnin), while the superna- tant was undyed. Then lysostaphin ( 10 pg/ml) was added at room temperature. The wall lysis was measured by determination of released wall label in the supernatant after centrifugation (10,000 x g , 15 rnin). The radioac- tivity measurements were performed as described in a previous study (Wecke et al. 1982).
Thin sectioning technique Staphylococci were prepared for thin-sectioning by
fixation with 2.5% glutaraldehyde (Serva, FRG) in 0.1 M cacodylate buffer pH 7.2 for 1 h at room temperature. After washing with cacodylate buffer the cells were postfixed with 1.5% osmiumtetroxide (Serva, FRG) + 1.65% potassium dichromate in 0.1 M cacodylate buffer for 1 h at room temperature. The fixed samples were washed again with the same buffer and poststained en bloc with 0.5% uranyl acetate in distilled water for 1 hat room temperature. After dehydration using a series of increasing ethanol concentrations, the samples were infiltrated by the LR White plastic monomer (Science Services, FRG) and then polymerized for 24 h at 60 "C.
Thin sections were cut with diamond knives (Dupont, USA) on a Reichert OM U 3 ultramicrotome, and stained with lead citrate for 3 - 5 rnin (Reynolds 1963) prior to examination in a Philips 400 electron micro- scope.
RESULTS
Action of suramin or Evans Blue on staphylococci (a) Growing staphylococci. After the addition of
suramin (100 - 1000 pg/ml) to growing staphylo- cocci, no disturbance of growth, as determined by optical density measurements, could be detected. However, the release of wall label within 2 h was remarkably reduced after addition of suramin or Evans Blue to growing staphylococci (Fig. 1). This result demonstrated a suppression of the activity of wall autolytic enzymes participating in wall turnover of staphylococci. The ultrathin sections of suramin-treated bacteria revealed thickened cross walls, while the peripheral wall seemed to be unchanged (Fig. 2). Sometimes, a layering of the
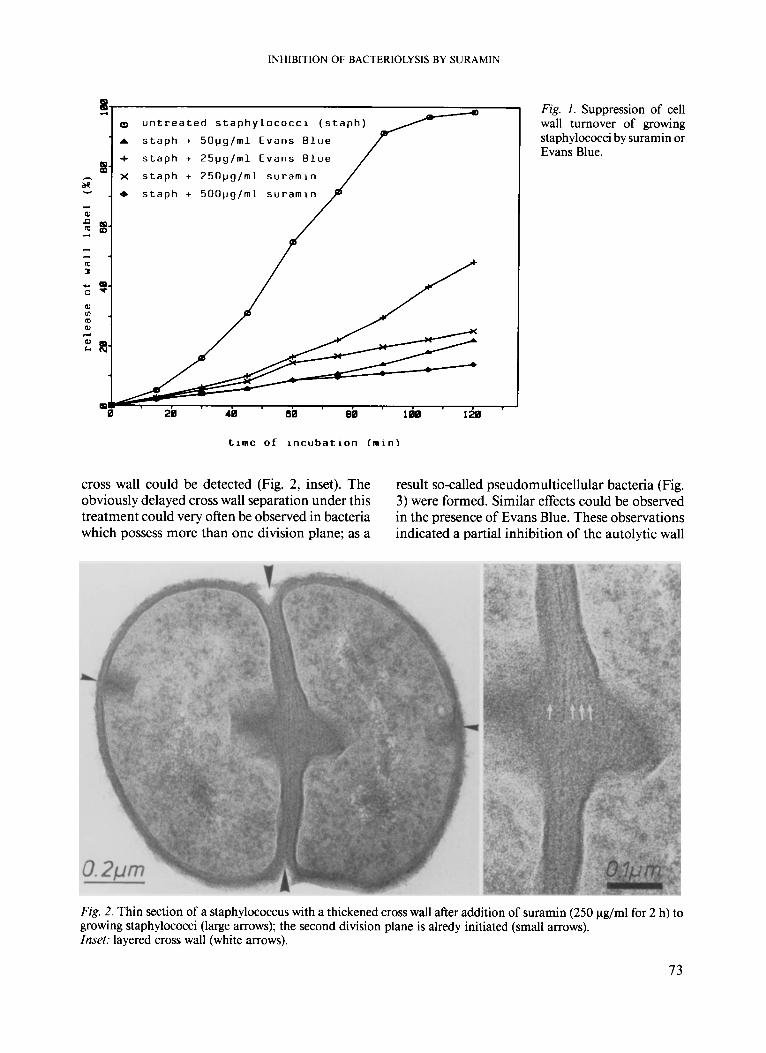
INHIBITION OF BACTERIOLYSIS BY SURAMIN
Q u n t r e a t e d s t a p h y l o c o c c i ( s t a p h )
A s t a p h + 5 0 p g / m l Evans B l u e
+ s t a p h + 2 5 p g / m l Evans B l u e
e s t a p h + 5 0 0 p g / m l s u r a m i n
Fig. I . Suppression of cell wall turnover of growing staphylococci by suramin or Evans Blue.
t i m e of i n c u b a t i o n I m i n )
cross wall could be detected (Fig. 2, inset). The obviously delayed cross wall separation under this treatment could very often be observed in bacteria which possess more than one division plane; as a
result so-called pseudomulticellular bacteria (Fig. 3) were formed. Similar effects could be observed in the presence of Evans Blue. These observations indicated a partial inhibition of the autolytic wall
Fig. 2. Thin section of a staphylococcus with a thickened cross wall after addition of suramin (250 pg/ml for 2 h) to growing staphylococci (large arrows); the second division plane is alredy initiated (small arrows). Inset: layered cross wall (white arrows).
1 3
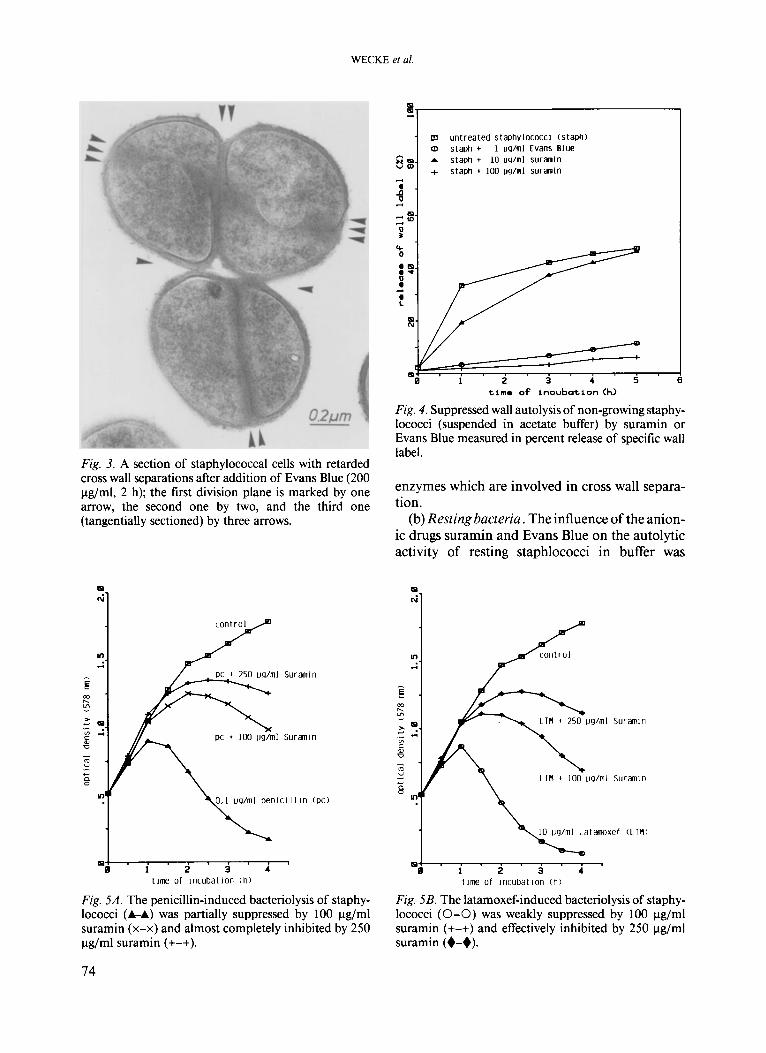
WECKE et al.
Fig. 3. A section of staphylococcal cells with retarded cross wall separations after addition of Evans Blue (200 pg/ml, 2 h); the first division plane is marked by one arrow, the second one by two, and the third one (tangentially sectioned) by three arrows.
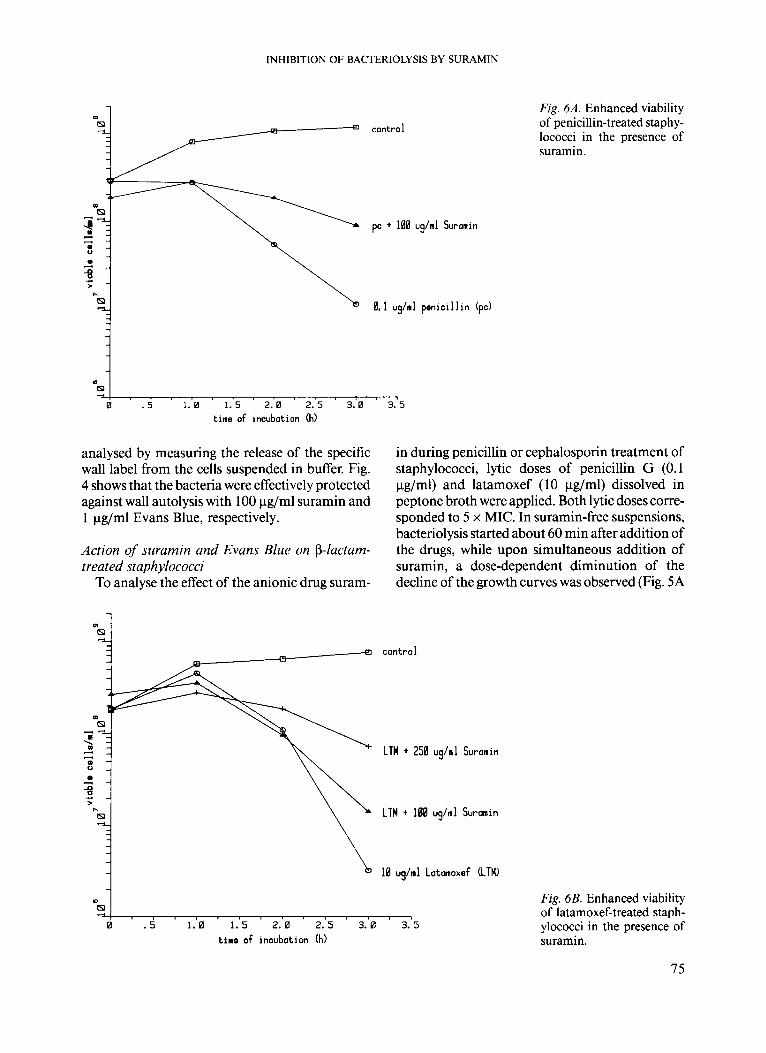
/& pc + 100 ug/ml Suranin
0 1 2 3 4 m4 . . . . . . - .
time of incubation ( h )
Fig. 5A. The penicillin-induced bacteriolysis of staphy- lococci (A-A) was partially suppressed by 100 pg/ml suramin (x-x) and almost completely inhibited by 250 pg/ml suramin (+-+).
CI untreated StaDhYlOCOCCl (Staph) 0 staph + 1 ug/ml Evans Blue A staph + 10 ug/ml suranin + staph + 100 ug/ml suranin
4
a m a * d
9
m N
0 1 2 3 4 5 m
timr o f inoubation (h)
Fig. 4. Suppressed wall autolysis of non-growing staphy- lococci (suspended in acetate buffer) by suramin or Evans Blue measured in percent release of specific wall label.
enzymes which are involved in cross wall separa- tion.
(b) Resting bacteria. The influence of the anion- ic drugs suramin and Evans Blue on the autolytic activity of resting staphlococci in buffer was
m i . . . . . . . . 4 0 1 2 3 4
time of incubation (h)
Fig. 5B. The latamoxef-induced bacteriolysis of staphy- lococci (0-0) was weakly suppressed by 100 pg/ml suramin (+-+) and effectively inhibited by 250 pg/ml suramin (+-+).
74
INHIBITION OF BACTERIOLYSIS BY SURAMIN
- \ pc + 100 ug/ml Suromin - -
m
m n - .- >
0 , l ug/ml penicillin (pc)
0 .5 1. 0 1. 5 2.0 2.5 3.0 3. 5 time o f incubation (h)
analysed by measuring the release of the specific wall label from the cells suspended in buffer. Fig. 4 shows that the bacteria were effectively protected against wall autolysis with 100 pg/ml suramin and 1 pg/ml Evans Blue, respectively.
Action of suramin and Evans Blue on p-lactam- treated staphylococci
To analyse the effect of the anionic drug suram-
1
Fig. 6A. Enhanced viability of penicillin-treated staphy- lococci in the presence of suramin.
in during penicillin or cephalosporin treatment of staphylococci, lytic doses of penicillin G (0.1 pg/ml) and latamoxef (10 pg/ml) dissolved in peptone broth were applied. Both lytic doses corre- sponded to 5 x MIC. In suramin-free suspensions, bacteriolysis started about 60 min after addition of the drugs, while upon simultaneous addition of suramin, a dose-dependent diminution of the decline of the growth curves was observed (Fig. 5A
- C \ - - m
m
3
LTH + 250 ug/nl Suranin
- .I
> LTH + 1B0 ug/nl Suranin
10 ug/nl Lotonoxef (LTH)
0 .5 1.0 1.5 2.0 2 . 5 3.0 3.5 t ine o f incubation (h)
Fig. 6B. Enhanced viability of latamoxef-treated staph- ylococci in the presence of suramin.
75
WECKE et al.
A 0,lpg/ml prnioillin (PO) + PO + 100pg/d muramin
X po + 10p9/ml Evan. Blur
PO + 100pg/ml Evan- Blur
mJ 0 1s 30 4s 60 98
time of incubation tmln)
Fig. 7. Suppression of the release of the cytoplasm- specific label (3H-uracil) from penicillin-treated staphy- lococci by suramin or Evans Blue.
and 5B). Basically the same inhibitory effects were observed when suramin was replaced by Evans Blue (data not shown).
It was of particular medical interest to note that the inhibition of the P-lactam-induced bacterioly- sis of staphylococci also yielded considerably higher viability rates (Fig. 6A and 6B).
In order to contribute to a better understanding of the inhibitory effects of suramin and Evans Blue on P-lactam-induced bacteriolysis, the specific cytoplasmic label 3H-uracil was introduced into staphylococcal cells and its release was followed after additon of penicillin and of penicillin plus the anionic substances respectively, to the growth medium. These experiments showed that also the penicillin-induced pre-lytic release of the cytoplas- mic label proved to be remarkably reduced in the presence of suramin or Evans Blue (Fig. 7). The most effective suppression totalling up to more than 50Yo after 60 min was observed in the pres- ence of 0.1 pg/ml penicillin + 100 pg/ml Evans Blue.
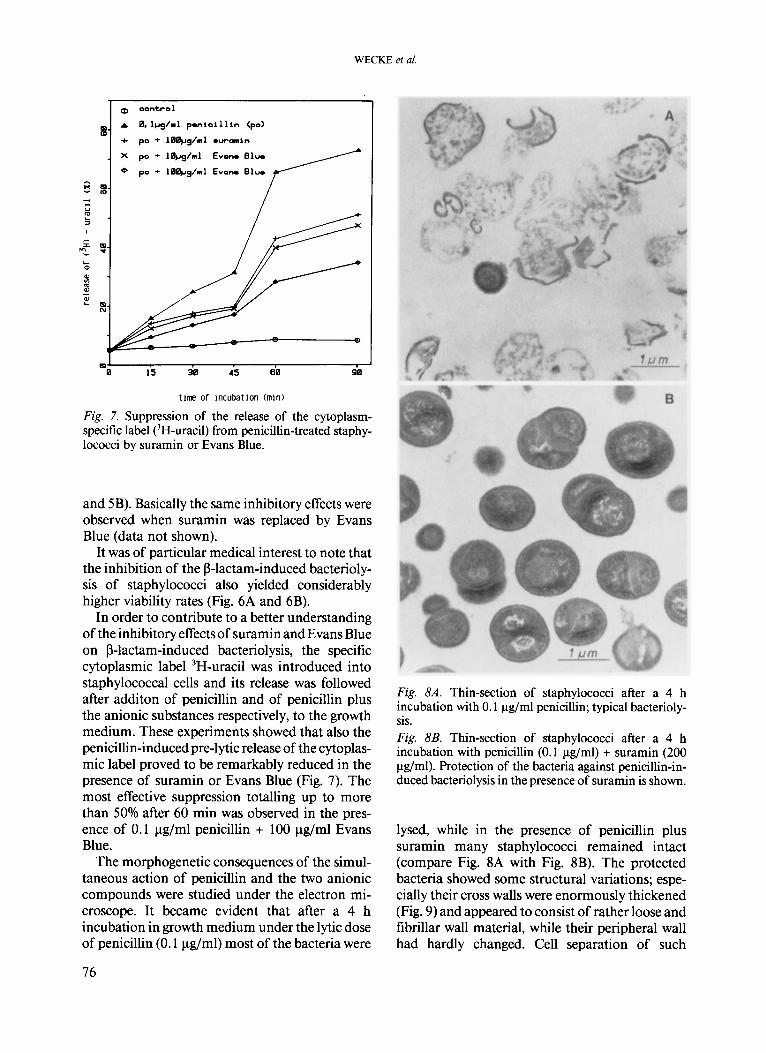
The morphogenetic consequences of the simul- taneous action of penicillin and the two anionic compounds were studied under the electron mi- croscope. It became evident that after a 4 h incubation in growth medium under the lytic dose of penicillin (0.1 pg/ml) most of the bacteria were
Fig. 8A. Thin-section of staphylococci after a 4 h incubation with 0.1 Fg/ml penicillin; typical bacterioly- sis. Fig. 8B. Thin-section of staphylococci after a 4 h incubation with penicillin (0.1 pg/ml) + suramin (200 pg/ml). Protection of the bacteria against penicillin-in- duced bacteriolysis in the presence of suramin is shown.
lysed, while in the presence of penicillin plus suramin many staphylococci remained intact (compare Fig. 8A with Fig. 8B). The protected bacteria showed some structural variations; espe- cially their cross walls were enormously thickened (Fig. 9) and appeared to consist of rather loose and fibrillar wall material, while their peripheral wall had hardly changed. Cell separation of such
76

INHIBITION OF BACTERIOLYSIS BY SURAMIN
Fig. 9. A thin-section of a staphylococcal cell after a 4 h incubation with 0.1 Fg/ml penicillin + 100 pg/ml Evans Blue. Only one daughter cell has retained the ability to produce a new cross wall. (The first division plane is marked by one arrow, the second by two arrows). The cross walls are enormously thickened, while the periph- eral wall hardly appears to be affected.
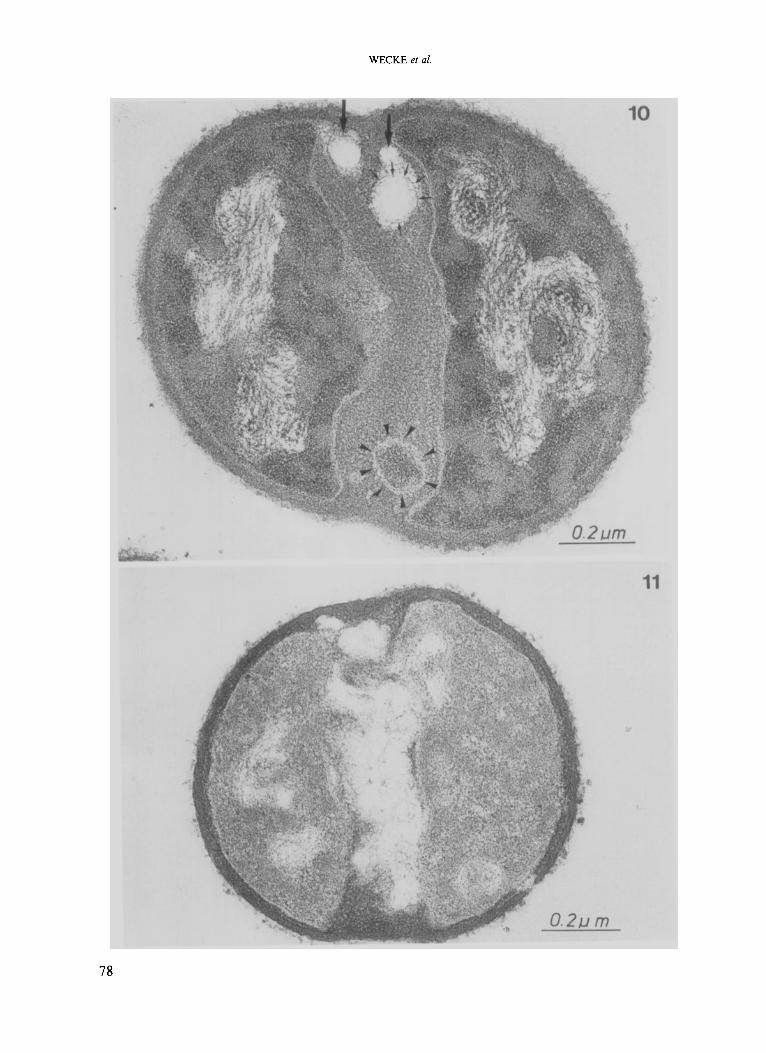
staphylococci was almost entirely blocked, which resulted in the formation of pseudomulticellular bacteria. In some cases, only one of the two daughter cells had produced a new cross wall in the second division plane, while the other did not reveal any morphogenetic activity (Fig. 9). Lytic organelles, which were deformed to a greater or lesser extent, so-called murosomes (Giesbrecht et al. 1985), located at the former initiation site of the cross wall could also be demonstrated (Fig. 10). After longer cultivation periods in the presence of penicillin plus protective substances, some wall autolysis within the cross wall was detected (Figs. 10). Unexpectedly, after an even longer period of cultivation, a more or less complete lysis of the cross wall could be observed (Fig. 1 I ) , whereas the peripheral wall remained almost entirely intact under suramin or Evans Blue protection.
The target structure of the anionic substances In order to gain information about the mecha-
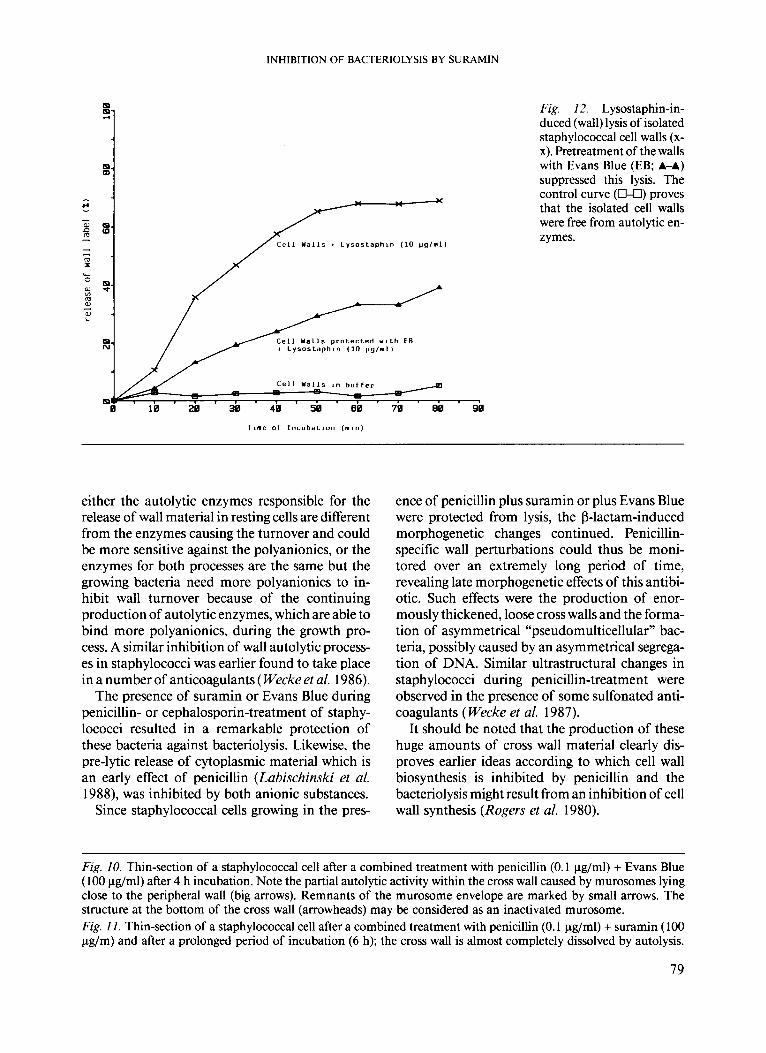
nism of autolysis inhibition, isolated and au- tolysin-free staphylococcal cell walls pretreated with Evans Blue were used as target. Then lysostaphin as a mixture of wall lytic enzymes was added. As indicated in Fig. 12 the lysostaphin-in- duced release of wall label was remarkably reduced
by Evans Blue. These results demonstrated that autolysin-free cell walls have been protected by Evans Blue, even against the lysostaphin-induced wall lysis.
DISCUSSION
Treatment of growing staphylococci with suramin or Evans Blue resulted in certain ultrastructural changes, although bacterial growth appeared to be unaffected. The most conspicuous alteration proved to be a considerable thickening of the cross wall, while the peripheral wall retained its normal appearance. This might be explained by a contin- ued apposition of wall material at the cross wall, while cross wall separation was delayed. The delay might be caused by an inhibition of wall autolytic enzymes which are known to be responsible not only for cross wall separation but also for cell wall growth. A similar inhibition of cross wall separa- tion has been reported to occur also in Clostridium sporogenes and in Bacillus megaterium after addi- tion of suramin (Shaikh & Lominski 1975; Shaikh et al. 1978), although the observed chain forma- tion of these bacteria was caused by higher doses of suramin.
The inhibition of several other enzymes by suramin was described earlier. Preferably cationic enzymes such as cathepsin D were affected by suramin (Buys et al. 1978: Ackerman et al. 1983). N-acetyl-glucosaminidase was inhibited by suram- in, as well ( Warr & Jakab 1983); this enzyme, although of human source, could be related, at least in terms of its function, to certain wall autolytic enzymes of bacteria, the so-called glu- cosidases. Autolytic enzymes responsible for wall turnover of growing bacteria have obviously been suppressed too. Such wall autolysins participating in bacterial cell wall turnover should preferably be located in the peripheral wall (Bliimel et al. 1979). The third process that could be inhibited by suramin and Evans Blue was the release of wall components from bacteria resting in buffer as mediated by cell wall autolytic enzymes. Our results indicated that wall autolysis of such non- growing cells was more effectively inhibited by Evans Blue than by the other sulfonated drug, the suramin. The doses of polyanionics which were needed to inhibit the wall autolysis were approxi- mately ten times lower than the doses to inhibit the cell wall turnover of growing cells. The reasons for this difference have to be discussed in two ways:
77
WECKE el al.
11
78
INHIBITION OF BACTERIOLYSIS BY SURAMIN
T i m e of Incubation ( m i ” )
Fig. 12. Lysostaphin-in- duced (wall) lysis of isolated staphylococcal cell walls (x- x). Pretreatment of the walls with Evans Blue (EB; A-A) suppressed this lysis. The control curve (0-0) proves that the isolated cell walls were free from autolytic en- zymes.
C e l l Halls p r o t e c t e d w i t h El3 + L y s o s t a p h i n (10 p g l n l )
I 10 20 30 40 SB BB 70 am 90
either the autolytic enzymes responsible for the release of wall material in resting cells are different from the enzymes causing the turnover and could be more sensitive against the polyanionics, or the enzymes for both processes are the same but the growing bacteria need more polyanionics to in- hibit wall turnover because of the continuing production of autolytic enzymes, which are able to bind more polyanionics, during the growth pro- cess. A similar inhibition of wall autolytic process- es in staphylococci was earlier found to take place in a number of anticoagulants ( Wecke et al. 1986).
The presence of suramin or Evans Blue during penicillin- or cephalosporin-treatment of staphy- lococci resulted in a remarkable protection of these bacteria against bacteriolysis. Likewise, the pre-lytic release of cytoplasmic material which is an early effect of penicillin (Labischinski et al. 1988), was inhibited by both anionic substances.
Since staphylococcal cells growing in the pres-
ence of penicillin plus suramin or plus Evans Blue were protected from lysis, the P-lactam-induced morphogenetic changes continued. Penicillin- specific wall perturbations could thus be moni- tored over an extremely long period of time, revealing late morphogenetic effects of this antibi- otic. Such effects were the production of enor- mously thickened, loose cross walls and the forma- tion of asymmetrical “pseudomulticellular” bac- teria, possibly caused by an asymmetrical segrega- tion of DNA. Similar ultrastructural changes in staphylococci during penicillin-treatment were observed in the presence of some sulfonated anti- coagulants ( Wecke et al. 1987).
It should be noted that the production of these huge amounts of cross wall material clearly dis- proves earlier ideas according to which cell wall biosynthesis is inhibited by penicillin and the bacteriolysis might result from an inhibition of cell wall synthesis (Rogers et al. 1980).
Fig. 10. Thin-section of a staphylococcal cell after a combined treatment with penicillin (0.1 pg/ml) + Evans Blue (100 pg/ml) after 4 h incubation. Note the partial autolytic activity within the cross wall caused by murosomes lying close to the peripheral wall (big arrows). Remnants of the murosome envelope are marked by small arrows. The structure at the bottom of the cross wall (arrowheads) may be considered as an inactivated murosome. Fig. 11. Thin-section of a staphylococcal cell after a combined treatment with penicillin (0.1 pg/ml) + suramin (100 pg/m) and after a prolonged period of incubation (6 h); the cross wall is almost completely dissolved by autolysis.
79

WECKE et al.
As for the fate of the lytic organelle -like vesicles of the cell wall, the murosomes, known for punch- ingpores into the peripheral wall for cell separation of untreated staphylococci (and initiating the peni- cillin-induced bacteriolysis, Giesbrecht et al. 1985), our present data indicate that the muro- somes apparently became inactive when penicillin and the anionic substances were applied simulta- neously. Only after a prolonged period of treat- ment with penicillin plus suramin, could a certain restricted activity of murosomes still be observed within the cross wall, either close to the peripheral wall (Le., at the former initiation site of the cross wall) or in the middle (at the initiation site of the next division plane). The observation that even the most extended cross wall lysis was not accompa- nied by any subsequent lysis of the peripheral wall was astonishing; it suggested that both anionic substances were unable to penetrate into the region of the cross wall proper.
Basically, two possibilities should be discussed when explaining the mechanism of autolysis sup- pression caused by anionic substances: i) direct inhibition of autolytic enzymes, ii) alteration of the bacterial cell wall as target structure of wall autolytic enzymes. Since the P-lactam-induced release of cytoplasmic constituents was inhibited already long before bacteriolysis (triggered by au- tolytic enzymes) started, the autolytic wall en- zymes cannot be considered to function as a primary target of the anionic substances. More probably, an interaction of these substances with the cell wall must be assumed; this is consistent with our findings that isolated staphylococcal cell walls freed from cytoplasmic membrane and au- tolytic enzymes by SDS- and trypsin-treatment apparently represented the target for anionic sub- stances because of the suppressed lysostaphin-in- duced wall lysis.
A direct interaction of the anionic substances with the lytic agent lysostaphin is improbable because the cell walls were pretreated with suramin or Evans Blue and afterwards carefully washed before addition of lysostaphin. Altogether, the data suggested an interaction of the anionic substance with the target structure of wall autolysins: the bacterial cell wall. Therefore, the inhibition of the different cell wall degradation processes including the penicillin-induced bacteriolysis may be caused by the changed target structure, so that autolytic or other wall lytic enzymes are no longer able to bind so effectively because of the steric hindrance.
80
Nevertheless an additional, restricted direct inter- action of the anionic substances used with wall autolytic enzymes can at present not be totally excluded.
Our findings regarding the protection of p-lac- tam-treated staphylococci by suramin or Evans Blue are also considered to be of therapeutic importance. This type of anionic drugs has the ability to suppress the replication of HIV, either alone (Balzarini et al. 1986 a; Balzarini et al. 1986 b) or in combination with other drugs (Resnick et al. 1986).
Since therapeutic plasma levels of suramin were reported to exceed 100 pg/ml for several weeks (Collins et al. 1986), any penicillin-therapy for opportunistic infections with staphylococci would be hampered due to an inhibited destruction of the bacteria and an increased level of the survival rates of these bacteria. Another possible implication of our results should be noted; the lysis resistance of the staphylococcal cell wall including those consid- erable amounts of material from the thickened cross walls, might result in an enhanced overall amount of bacterial wall material preserved within the human body. This consideration should be of some interest, since chronic inflammatory pro- cesses may be triggered by large amounts of lysis-resistant wall material (Ginsburg et al. 1985, Giesbrecht & Wecke 1987).
This study was supported by a grant from the Deutsche Forschungsgemeinschaft. The skillful technical assis- tance of Mrs Albrecht, Mr Kniffke, Mr Nurnberg and Mrs Schotes is much appreciated.
REFERENCES
1. Ackermann, N. R., Jubb, S., Trimble, B., Marlowe, S., Miram, L. & Maloney, P.: Divergent effects of suramin on in vitro and in vivo assays of cartilage degradation. J. Pharmacol. Exp. Ther. 225: 243-250, 1983.
2. Balzarini, J., Mitsuya, H., De Clercq, E. & Broder, S.: Comparative inhibitory effects of suramin and other selectedcompounds on the infectivity and replication of human T-cell lymphotropic virus (HTLV-III)/ lymphadenopathy-associated virus (LAV). Int. J. Cancer 37: 451-457, 1986a.
3. Balzarini, J., Mitsuya, H., De Clercq, E. & Broder, S.: Aurintricarboxylic acid and Evans Blue represent two different classes of anionic compounds which selectively inhibit the cytopathogenicity of human T-cell lymphotropic virus type III/lymphadenopa-

INHIBITION OF BACTERIOLYSIS BY SURAMIN
thy-associated virus. Biochem. Biphys. Res. Com- mun. 136: 64-7 1, 1986b.
4. Blumel, P., Uecker, W. & Giesbrecht, P.: Zero order kinetics of cell wall turnover in Staphylococcus aureus. Arch. Microbiol. 121: 103-1 10, 1979.
5. Broder, S., Yarchoan, R., Collins, J. M., Lane, H . C., Markham, P. D., Klecker, R. W., Redfield, R. R., Mitsuya, H., Hoth, D. F., Gelmann, E., Groopman, J. E., Resnick, L., Gallo, R. C., Myers, C. E. & Fauci, A. S.: Effects of suramin on HTLV-III/LAV infec- tion presenting as Kaposi’s sarcoma or AIDS-related complex: Clinical pharmacology and suppression of virus replication in vivo. Lancet ii: 627-630, 1985.
6. Buys, C. H. C. M., Bouma, J. M. W., Gruber, M. & Wisse, E.: Induction of lysosomal storage by suram- in. Arch. Pharmacol. 304: 183-190, 1978.
7. Chandra, P., Vogel, A. & Gerber, T.: Inhibitors of retroviral DNA polymerase: Their implication in the treatment of AIDS. Cancer Res. 45: 4677-4684, 1985.
8. Collins, J. M., Klecker, Jr., R. W., Yarchoan, R., Lane, H . C., Fauci, A. S., Redfield, R. R., Broder, S. & Myers, C. E.: Clinical pharmacokinetics of suramin in patients with HTLV-III/LAV infection. J. Clin. Pharmacol. 26: 22-26, 1986.
9. De Clercq, E.: Suramin, a potent inhibitor of the reverse transcriptase of RNA tumor viruses. Cancer Letters 8: 9-22, 1979.
10. Giesbrecht, P., Labischinski, H. & Wecke, J.: A special morphogenetic wall defect and the subse- quent activity of “murosomes” as the very reason for penicillin-induced bacteriolysis in staphylococci. Arch. Microbiol. 141: 315-324, 1985.
1 1. Giesbrecht, P. & Wecke, J.: Zur Abbaubarkeit bak- terieller Zellwande und ihre Bedeutung fur die Induktion chronisch-entziindlicher Prozesse. In: Holzmann et al. (Eds.): Dermatologie und Rheuma. Springer Verlag Berlin, Heidelberg 1987, pp. 77-88.
12. Ginsburg, I., Lahav, M., Goultschin, J., Sadovnik, M., Kwa, E., Wecke, J. & Giesbrecht, P.: The interaction of Staphylococcus aureus with leuko- cytes in joint lesions: An ultrastructural study. In: Jeljaszewicz, J. (Ed.): The Staphylococci, Zbl. Bakt. Suppl. 14, Gustav Fischer Verlag, Stuttgart, New York, 1985, pp. 691-698.
13. Ginsburg, I. & Sela, M. N.: The role of leukocytes and their hydrolases in the persistence, degradation, and transport of bacterial constituents in tissues: Relation to chronic inflammatory processes in
staphylococcal, streptococcal, and mycobacterial infections and in chronic periodontal diseases. Crit. Rev. Microbniol. 44: 249-332, 1976.
14. Labischinski, H., MaidhoJ H., Franz. M., Kruger, D., Sidow, T. & Giesbrecht, P.: Biochemical and biophysical investigations into the cause of the penicillin-induced lytic death of staphylococci: Checking predictions of the murosomal model. In: Actor et al. (Eds.): Antibiotic inhibition of bacterial cell surface assembly and function. Society for Microbiology Chapter 23, Washington, 1988, pp.
15. Mitsuya, H., Popovic, M., Yarchoan, R., Matsushi- ta, S., Gallo, R. C. & Broder, S.: Suramin protection of T cells in vitro against infectivity and cytopathic effect of HTLV-111. Science 226: 172- 174, 1984.
16. Resnick, L., Markham, P. D., Veren, K., Salahuddin, S. Z. & Gallo, R. C.: In vitro suppression of HTLV-III/LAV infectivity by a combination of acyclovir and suramin. J. Infect. Dis. 154: 1027- 1030, 1986.
17. Reynolds, E. S.: The use of lead citrate at high pH as an electron-opaque stain in electron microscopy J. Cell Biol. 17: 208-212, 1963.
18. Rogers, H. J., Perkins, H. R. & Ward, J. B.: Micro- bial Cell Walls and Membranes. Chapman and Hall London, New York, 1980.
19. Shaikh, D., Shaikh, M. R. & Hoosen, A. A,: Induced chain formation in Bacillus megaterium by suramin (Bayer 205). Zentralbl. Bakteriol. 242: 137-140, 1978.
20. Shaikh, M. R. & Lominski, I.: Induction of chain formation in Clostridium sporogenes by suramin. Zentralbl. Bakteriol. 230: 237-240, 1975.
21. Warr, G. A. & Jakab, G. L.: Lung macrophage defense responses during suramin-induced lysoso- ma1 dysfunction. Exper. Mol. Pathol. 38: 193-207, 1983.
22. Wecke, J., Kwa, E., Lahav, M., Ginsburg, I. & Giesbrecht, P.: Suppression of penicillin-induced bacteriolysis of staphylococci by some anticoagu- lants. J. Antimicrob. Chemother. 20: 47-55, 1987.
23. Wecke, J., Lahav, M., Ginsburg, I. & Giesbrecht, P.: Cell wall degradation of Staphylococcus aureus by lysozyme. Arch. Microbiol. 131: 116-123, 1982.
24. Wecke, J., Lahav, M., Ginsburg, I . , Kwa, E. & Giesbrecht, P.: Inhibition of wall autolysis of staphy- lococci by sodium polyanethole sulfonate “liquoid”. Arch. Microbiol. 144: 110-1 15, 1986.
242-257.
81