inhibition of metastatic lung cancer in c57bl/6 mice by...
TRANSCRIPT

International Immunopharmacology 14 (2012) 570–579
Contents lists available at SciVerse ScienceDirect
International Immunopharmacology
j ourna l homepage: www.e lsev ie r .com/ locate / in t imp
Inhibition of metastatic lung cancer in C57BL/6 mice by liposome encapsulated alltrans retinoic acid (ATRA)
Siddikuzzaman 1, V.M. Berlin Grace ⁎Department of Biotechnology, Karunya University, Coimbatore-641 114, Tamil Nadu, India
⁎ Corresponding author. Tel.: +91 9487846617; fax:E-mail addresses: [email protected] (
[email protected] (V.M.B. Grace).1 Tel.: +91 9042166990.
1567-5769/$ – see front matter © 2012 Elsevier B.V. Allhttp://dx.doi.org/10.1016/j.intimp.2012.09.008
a b s t r a c t
a r t i c l e i n f oArticle history:Received 4 September 2012Received in revised form 12 September 2012Accepted 12 September 2012Available online 27 September 2012
Keywords:All-trans retinoic acid (ATRA)LiposomeLung cancer and HPLC
The purpose of this study was to investigate whether all trans retinoic acid (ATRA) incorporated in liposomecomposed of distearoylphosphatidylcholine (DSPC/cholesterol) could inhibit the metastatic lung cancer inmice more efficiently than free ATRA. Metastatic lung cancer model was developed by intravenous injectionof B16F10 cells and it is also referred as melanoma model. In this present study, C57BL/6 mice were dividedinto several groups as per experimental design and the free ATRA and liposome encapsulated ATRA weregiven for 21 days at a dose of 0.60 mg/kg body weight/day after cell line implantation. After 21 days, micewere sacrificed at different time interval for ATRA level analysis in serum and lung tissue by HPLC methodand the remaining mice were kept for anticancer study. The ATRA level increased significantly in serumand lung tissue in liposome encapsulated ATRA treated mice. In cancer bearing mice, tumor nodule formationdecreased and life span increased after receiving liposome encapsulated ATRA treatment than free ATRAtreated mice. This result implies that the liposome encapsulated ATRA has maintained more ATRA concentra-tion in lung tissue and showed more inhibition on the lung tumor nodule formation. The results indicate apossible use of liposome encapsulated ATRA in prevention of lung metastasis.
© 2012 Elsevier B.V. All rights reserved.
1. Introduction
Lung cancer is more common in older adults with highest mortal-ity rate. This may be due to metastasis. There is no drug presentlyavailable to specifically inhibit the metastasis of cancer cells. Thereis a cascade of events leading to the metastasis of tumors, wheretumor cells disseminate from the primary tumor, migrate throughthe basement membrane, survive in the circulatory system, invadeinto a secondary site, and start to proliferate [1,2].
All-trans retinoic acid (ATRA) is an active metabolite of vitamin Athat the body needs in small amount to function and stay healthy [3].ATRA is considered as an anti-cancer chemotherapy drug and classi-fied as a member of the “retinoid family” which includes retinal, ret-inol, retinoic acid, and retinyl ester. ATRA is a promising anticanceragent, which has been investigated in the chemoprevention andtreatment of cancer [4]. ATRA has been used as first choice drug foracute promyelocytic leukemia (APL) [5]. ATRA and its other active de-rivatives are potent modulators of cell growth, differentiation, andapoptosis in a variety of cell types [6,7]. They can also be used as che-motherapeutic and chemopreventive agents in a variety of malignan-cies, such as leukemias, uterine leiomyomas, as well as colon, gastric,solid tumor and breast cancers [8–10]. Retinoic acid (RA) has also
+91 4222615615.Siddikuzzaman),
rights reserved.
been suggested to be efficacious in treating lung cancer [11,12].Retinoic acid derivatives are already in clinical use for different dis-eases, such as acute leukemia, for which ATRA is used as a differenti-ating agent [13]. Anticancer activity of ATRA is achieved by binding toretinoic acid receptors present on the nuclear membrane of cancercells, leading to the induction of cell growth inhibition, differentiationor apoptosis [14]. Growth of tumor tissue is determined by the bal-ance between cell proliferation and apoptosis. The inability of cancercells to undergo apoptosis may provide a growth advantage, and cellswith defective apoptosis may therefore have a better chance of sur-vival. Thus, induction of apoptosis was recognized as an ideal strategyfor cancer chemotherapy. Researchers revealed that the induction ofapoptosis by ATRA incorporated in cationic liposome is composed of1,2 dioleoyl-3-trimethylammonium propane (DOTAP)/cholesterol inA549 human lung carcinoma cells, which are insensitive (resistant)to the growth inhibitory effects of ATRA in vitro [15]. In mammarycarcinoma cell lines, ATRA has been shown to induce growth inhibi-tion by triggering either cell cycle arrest or apoptosis or both[16–18]. Another study showed the inhibition of murine liver metas-tasis by ATRA incorporated in O/W emulsions, which could accumu-late in the liver [19]. These results provide evidence that thetargeting of ATRA by a drug carrier could improve the efficacy of can-cer metastasis and differentiation therapy. To date, there are few pub-lished reports on cancer differentiation therapy in the lung [20]. SinceATRA is expected to be selectively toxic to cancer cells, the passivetargeting of the lung organ by encapsulation may be enough for thetherapy of lung metastasis.

571Siddikuzzaman, V.M.B. Grace / International Immunopharmacology 14 (2012) 570–579
The poor aqueous solubility of ATRA can be a major drawback forits parenteral administration. Incorporation of drugs in lipid-basedcarriers such as liposome and lipid emulsions is known as an attrac-tive means to overcome the solubility limitations of drugs [21].ATRA contains a carboxyl group, thus ATRA would be incorporatedmore stably in cationic liposome. Gradual decrease in ATRA level inblood circulation observed after long term oral treatment with ATRAhowever, has been shown to be, probably due to the induced cyto-chrome P-450-dependent metabolism of ATRA [22]. Attempts havebeen made to develop i.v. injectable formulation of ATRA by loadingin carriers such as solid nano particles, cyclodextrins, liposome etc.in order to improve its aqueous solubility [23,24]. Several studieshave shown that the liposome can be a novel injectable formulationof ATRA [25]. In addition to the poor solubility, ATRA has shorterhalf life and is also known to be chemically unstable [26]. This studywas therefore designed to enhance the chemo preventive/therapeuticeffect of ATRA on lung metastasis by encapsulating it with liposomewhich might help the ATRA for its persistence in circulation and toreach the lung tissue. In this study, the ATRA level analysis in serumand lung tissue, toxicity and anticancer activity were evaluated aftertreatment with ATRA incorporated in DSPC/cholesterol liposome inB16F10 cell implanted C57BL/6 lung cancer bearing mice model. Re-sults were compared between the free ATRA and the encapsulatedATRA in lung cancer model.
2. Materials and methods
2.1. Animals
C57BL/6 male mice (4–6 weeks old) were purchased from the Na-tional Institute of Nutrition (Hyderabad, Andhra Pradesh, India). Theanimals were kept in well-ventilated cages in a facility maintainedat 22 (±1) °C, with a 40 (±10) % relative humidity, and a 12-hlight–dark during the experimental period. Mice were providedaccess to normal mouse chow (Sai Feeds, Bangalore, India) andfiltered water ad libitum. All the animal experiments were carriedout according to the rules and regulation of the Animal EthicsCommittee of the Government of India.
2.2. Chemicals
ATRA was purchased from Sigma Chem. Co. (St. Louis, MO).Distearoyl-L-phosphatidylcholine (DSPC) and cholesterol were pur-chased from Sigma-Aldrich Co. (St. Louis, MO, USA). All other chemicalsused were of analytical grade.
2.3. Tumor cell line
The B16F10 melanoma cell line was purchased from the NationalCentre for Cell Science (NCCS, Pune, India). The cells were maintainedin RPMI 1640 medium buffered with 2 g/l of HEPES and sodium bicar-bonate, and supplemented with dextrose, penicillin, streptomycinand 10% of fetal bovine serum. The cells were maintained in a humid-ified atmosphere containing 5% CO2 at 37 °C. When needed for exper-iments the cells were harvested with trypsin:EDTA (0.05:0.03 [w/v])solution, and thenwashed in phosphate-buffered saline (PBS, pH 7.4).For the animal experiments, the recovered cells were adjusted to1×106 cells/ml in PBS and then 100 μl of the suspension was injectedinto the tail-vein.
2.4. Preparation of liposome
The liposome preparation process was modified from relevant lit-erature [27]. The liposome, DSPC:cholesterol:ATRA (70:20:10 molarratios), were prepared with or without ATRA of total lipid liposome.The mixture, with or without ATRA, was first dissolved in chloroform.
After vacuum drying and desiccation, pH 7.4 phosphate-buffered sa-line was added for hydration. The preparation was then sonicatedand centrifuged at 100,000×g to remove unentrapped ATRA.
2.5. Characterization of liposome
The mean particle size of the liposome was measured using aZetasizerNano ZS instrument (Malvern Instruments Ltd.,Worcestershire,UK) [27]. The liposomal loading of ATRAwas estimated by absorbance at340 nm after dissolving ATRA incorporated in liposome in ethanol sincethe ATRA alone is soluble in ethanol.
2.6. In-vivo drug toxicity studies in C57BL/6 mice
A group of six C57BL/6 mice was given graded doses of the ATRA(i.e., 0.5–1.0 mg/kg-body weight) via intra-peritoneal (IP) and con-tinuously observed for 5 h for any changes in various autonomicand behavioral responses. Thereafter, the mice were examined dailyfor 50 days to allow for extended observations of any ATRA-inducedmortality.
2.7. Determination of the in-vitro drug toxicity activity of ATRA by trypanblue dye exclusion method
Short term toxicity studies by trypan blue exclusionmethod is a verysimple method which can be carried out within a short time of 3 h.B16F10 cells (1×106 cells) were incubatedwith various concentrations(5 μg/ml, 15 μg/ml, 25 μg/ml, and 50 μg/ml) of ATRA in a final volumeof 1.0 ml for 3 h at 37 °C. After incubation the viability of the cellswere determined by trypan blue dye exclusion method [28].
2.8. In vitro toxicity of ATRA towards B16F-10 melanoma cells by MTTassay
In vitro toxicity of the ATRA was determined using B16F10 cells.Cells were seeded in 96-well flat-bottom plates (2×106 cells/well)and allowed to adhere for 24 h at 37 °C with 5% CO2 atmosphere. Dif-ferent concentrations of free ATRA (5–50 μg/ml) and free liposomewere incubated further for 48 h. Before the 4th hour of the comple-tion of incubation, 20 μl of MTT (3-(4,5-dimethylthiazol-2-Yl)-2,5-diphenyltetrazolium bromide) (5 mg/ml) was added [29,30].Same experiment was carried out for without ATRA treatment i.e. con-trol set also. Percentage of dead cells was determined using an ELISAplate reader set to record absorbance at 570 nm.
2.9. To study the level of ATRA in serum and lung tissue
2.9.1. Experimental groups for ATRA level analysis by reverse phase HPLCC57BL/6 mice were divided into 5 groups of 15 mice each to carry
out the level of ATRA present in lung and serum in the lung tumor bear-ingmicemodel. B16F10 cell linewas injected (1×106 cells in 0.1 ml) bytail vein to groups I, IV and V. Free ATRA and encapsulated ATRA weregiven for respective group at a dose of 0.60 mg/kg/body weight/dayfor 21 days after cell line implantation. Extra 3 mice were kept for nor-mal healthy animal to assay the normal level of ATRA in serum and lungtissue.
Group I = Cell line alone injectedGroup II = Free ATRA aloneGroup III = Liposome encapsulated ATRA aloneGroup IV = Cell line injected+free ATRAGroup V = Cell line injected+encapsulated ATRA
2.9.2. Sample collection for ATRA analysisAfter 21st dose, three mice from each group were sacrificed by
cervical dislocation at different time points such as 1, 3, 6, 12 and

572 Siddikuzzaman, V.M.B. Grace / International Immunopharmacology 14 (2012) 570–579
24 h respectively and on the same day the extra 3 normal healthy micewere also sacrificed by cervical dislocation. Bloodwas collected by heartpuncture and lung tissue was dissected out for ATRA level analysis.
Serum was prepared by centrifugation of the blood for 10 min at4 °C and 1500×g and stored at −80 °C until analysis. Lung tissuewas cut into pieces with scissor (200 mg) and washed with cold sa-line solution. Lung tissues were homogenized and supernatant wassubjected for analysis of ATRA level in lung tissue.
2.9.3. Sample processing for HPLC analysisConcentration of ATRA was measured by HPLC [31]. Prior to injec-
tion into the chromatograph, serum samples were subjected to thefollowing extraction method which was modified from relevant liter-ature to deproteinate at dark room [32]: 100 μl of acetonitrile wasadded to 100 μl of serum sample and the mixture was shaken byhand (for 10 s), placed in the refrigerator at −4 °C for 10 min, andthen centrifuged (2000×g for 5 min). The supernatant was trans-ferred to a clean tube and stored at −20 °C until injection (200 μl)into the chromatograph.
Tissueswere homogenizedwith 250 μl of 10% ascorbic acid and then250 μl of ethanol was added to each tube and vortexed for 1 min. Later2 ml of n-hexane was added and vortexed. The sample was thencentrifuged at 2000 rpm for 10 min. The hexane containing upperlayer was removed, evaporated and dried under nitrogen stream. Theresidue was dissolved in 250 μl of methanol for further HPLC analysis.
2.9.4. Pharmacokinetic study of ATRA in serum and lung tissueAll samples were analyzed for ATRA concentration by reversed-
phase HPLC in a Shimadzu Prominence HPLC system with LC 10 ATpumps, SPD M20 A detector, with Rheodyne injector loop volume of20 μl. The column was a Phenomenex Luna (250 mm×4.6 mm) col-umn, 5 μmparticle size, and themobile phasewas amixture of acetoni-trile and water (45:65, v/v) delivered at a flow rate of 1 ml/min.Wavelength used for the study was 310 nm.
The pharmacokinetic parameters were assessed by analysis of theATRA level presence in the serum and lung tissue at a particular timepoint. Elimination rate constant (ERC), Cmax, Tmax, terminal half-life(t1/2), area under the concentration–time curve up to last time(AUC0–t) or infinite time (AUC0–∞) were analyzed.
2.9.5. Standard curve calibration for ATRA level analysisCalibration graphs were constructed by linear least squares re-
gression analysis of the plot of the peak height against the concentra-tion of ATRA for serum and lung tissue respectively. For serumsample, 5 standards cover the range of 200 to 1000 ng/ml and simi-larly for lung tissue sample, 5 standards cover the range of 400 to2000 ng/ml. The concentration of ATRA in serum and lung tissue sam-ples was determined by interpolation in the calibration graph of thecorresponding peak-height value obtained in the chromatograms.
2.10. Anti cancer study of encapsulated ATRA in B16F10 cell implantedC57BL/6 mice
2.10.1. Drug administrationFree ATRA was suspended in olive oil and liposome encapsulated
ATRA was dissolved in phosphate buffer saline to the desired concen-tration (0.60 mg/kg body weight/day). For the studies, the free ATRAand liposome ATRA were administered intraperitoneally (IP) startingon the day the B16F10 tumor cells were injected (1×106 cells in0.1 ml) by tail vein and thereafter the free ATRA and liposomalATRA were given for 21 days by IP.
2.10.2. Experimental groups for anti cancer studyGroup I = NormalGroup II = Cell line alone injected
Group III = Cell line injected+olive oilGroup IV = Cell line injected+free liposomeGroup V = Cell line injected+free ATRAGroup VI = Cell line injected+encapsulated ATRA
2.10.3. Determination of the effect of ATRA on the hematological parameters,body weight and relative lung weight
Blood was collected from the tail vein and various parameterssuch as total WBC count [33], differential count (Leishman's stain)and hemoglobulin (Hb) content [34] were recorded on the 22ndday. Body weight was taken before and at the end of experimentand lung tissue weight was also noted down after sacrification.
2.10.4. Lung nodule formation and tumor indexThe effect of ATRA having the activity to suppress the lung metas-
tasis was studied using C57BL/6 mice which were injected withB16F10 mouse melanoma cell line having high metastasis activityfrom small inoculums. ATRA treatment was carried out as mentionedearlier. On the 22nd day after the tumor inoculation six mice fromeach group were sacrificed and their lung was excised out, weighed,and placed in phosphate-buffered saline (PBS, pH 7.4). The numberof the lung tumor nodules on the surface of the lung was countedand the percentage inhibition of lung tumor nodule formation wascalculated.
Tumor index was calculated for the control and treated mice usingthe following formula: Tumor index=lung weights×average gradefor the group. The lung tumor was graded based on a scale of 1–5:b10 tumor foci — Grade 1; 10–100 foci — Grade 2; one lobe of lungis full of tumors — Grade 3; both lobes are full of tumors — Grade 4;and, if lungs are full of tumors and tumors were growing into chestwall — Grade 5 [35,36].
2.10.5. Histopathology analysisSix animals from each groupwere sacrificed on Day 22 and lung tis-
sue was collected and kept in 10% formaldehyde for histopathologyanalysis and a portion was blocked in paraffin after processing. Sections(4–5 μmthickness)were taken and subjected to histopathological anal-ysis using hematoxylin and eosin (H&E) stain as per the standard proto-col [37]. Histologic changes were observed under light microscope; allresults were verified and certified by histopathologist.
2.10.6. TUNEL assay for DNA fragmentation in lung tissueTo detect apoptosis, in situ DNA fragmentation in lung tissue was
performed by using Dead End Colorimetric TUNEL System (Promega,Madison, WI) according to the recommendations of the manufactur-er. Briefly, paraffin embedded lung sections from both normal healthymice, cancer control and treated mice were deparaffinized and thenwashed with PBS followed by fixation with 4% paraformaldehydeand permeabilized with proteinase K solution. Tissue sections werenext incubated with equilibration buffer, bio‐tinylated nucleotidemix and TdT at 37 °C for 60 min. The activity of TdT was terminatedby addition of 2× saline sodium citrate (0.9% NaCl and sodium cit-rate). The endogenous peroxidase was blocked by 0.3% H2O2. Tissuesections were then incubated with streptavidin HRP solution for30 min and followed by incubation with substrate. After mountingin 80% glycerol the slides were observed under a light microscope.For the quantification of TUNEL expression (i.e. TUNEL score), positivecells (intense brown) were counted in six random fields [38]. Cellsthat had granular brown pigment in the nucleoplasm were scoredas apoptosis positive.
2.10.7. Study on survival rate and increase in life span of lung cancerbearing C57BL/6 mice
Six mice of each group were kept for their survival rate study, andthe percentage increase in life span (% ILS) was calculated using the

Fig. 2. In vitro toxicity activity of ATRA towards B16F10 cells byMTT Assay. Percentageof cell death caused by ATRA is shown. Values shown are expressed as mean±SD(n=3 determinations/dose).
573Siddikuzzaman, V.M.B. Grace / International Immunopharmacology 14 (2012) 570–579
following formula: % ILS=T−C/T×100, where T represents thenumber of survival days of the treated animals, and C represents thenumber of survival days of the control animals.
2.11. Statistical analysis
All data were expressed as mean (±SD). The statistical analysiswas done using one-way analysis of variance (ANOVA) followed bya Dunnett's test (using Graphpad InStat version 3.00 for WindowsXP; GraphPad Software, Inc., La Jolla, CA). A p-valueb0.05 was consid-ered significant.
3. Results
3.1. Size determination of ATRA encapsulated liposome, free liposomeand drug entrapment efficiency
The encapsulation of ATRA was done successfully. The mean sizeof the liposome alone was found to be 141.8±7.2 nm and that ofwith ATRA was 182.6±4.8 nm. The percent entrapment of the drugin liposome was found to be 80–82%.
3.2. In-vivo drug toxicity studies in C57BL/6 mice
The drug did not show any predictable changes in the autonomicand behavioral responses in C57BL/6 mice. All the mice lived healthyeven after 50 days at a dose of 1.0 mg/kg body weight of C57BL/6mice.
3.3. Determination of the in-vitro drug toxicity activity by trypan blueand MTT assay
The effect of ATRA towards B16F10 melanoma cell line was deter-mined by trypan blue exclusion method and result is given in Fig. 1.The ATRA has shown toxicity effect toward B16F10 tumor cells.50 μg/ml ATRA had shown a maximum 19% cell death.
The toxicity of ATRA towards B16F-10 melanoma cell line by MTTassay is shown in Fig. 2. Free ATRA was found to have a dose depen-dent toxicity to B16F-10 melanoma cell line. Free liposome showedno toxicity towards the B16F10 cells (data not shown).
3.4. HPLC analysis study
The ATRA levels in serum and lung tissues and the pharmacokinet-ic parameters that were quite consistent are given in Figs. 3 and 4respectively.
In non cancer mice model, encapsulated ATRA treatment showedthe serum ATRA level as 0.601±.04 μg/ml at 1st hour and 0.273±.012 μg/ml at 12th hour respectively, while ATRA level in free ATRA
Fig. 1. Determination of the in vitro drug toxicity activity of ATRA. The percentage ofdead cells was then determined by the trypan blue exclusion method. Values areexpressed in mean±SD (n=3).
group was found to be 0.423±.04 μg/ml at 1st hour and 0.111±.008 μg/ml at 12th hour respectively (Fig. 3). The t1/2 was found tobe 9.03 and 5.42 h for encapsulated ATRA and free ATRA respectivelyin non-cancer bearing mice model and that of 9.03 and 7.2 h respec-tively in lung cancer bearing mice model. The AUC0–t′was found to be4.86 and 2.67 μg h/ml for encapsulated ATRA and free ATRA innon-cancer bearing mice model and that of 4.86 and 2.15 μg h/ml re-spectively in lung cancer bearing mice model (Table 1). ATRA levelwas measured in serum sample up to 12th hour because of instru-ment limitation which could not measure the ATRA level on 24thhour.
In lung tissue, in non cancer mice model, encapsulated ATRAgroup showed the ATRA level as 0.70±.05 μg/ml at 1st hour and0.21±.03 μg/ml at 24th hour respectively. While Free ATRA group
Fig. 3. ATRA level analysis in serum sample; encapsulated ATRA showed highest concen-tration ATRA in serum in non-cancer mice compared with free ATRA. In cancer micemodel it was found more for encapsulated ATRA compared with free ATRA. Values areexpressed inmean±SD (n=3). aNormal vs. cancer alone (ns= nonsignificant); bnormalvs. free ATRA alone (pb0.01);
cnormal vs. encapsulated-ATRA alone (pb0.01); dnormal vs.
free ATRA treated in cancer group (pb0.01); enormal vs. encapsulated‐ATRA treated incancer group (pb0.01); fcancer alone vs. free ATRA alone (pb0.01); gcancer alone vs.encapsulated-ATRA alone (pb0.01); h,icancer alone vs. free ATRA treated in cancergroup (h=pb0.01 and i=ns); jcancer alone vs. encapsulated-ATRA treated in cancergroup (pb0.01).

Fig. 4. ATRA level analysis in lung tissue sample; encapsulated ATRA showed highest con-centration ATRA in serum in non-cancer mice compared with free ATRA. In cancer micemodel it was found more for encapsulated ATRA compared with free ATRA. Values areexpressed in mean±SD (n=3). aNormal vs. cancer alone (ns); b,knormal vs. free ATRAalone (b=pb0.01 and k=ns); cnormal vs. encapsulated-ATRA alone (pb0.01); d,mnormalvs. free ATRA treated in cancer group (d=pb0.01 andm=ns);
enormal vs. encapsulated-
ATRA treated in cancer group (pb0.01); f,lcancer alone vs. free ATRA alone (f=pb0.01 andl=ns); gcancer alone vs. encapsulated-ATRA alone (pb0.01); h,icancer alone vs. free ATRAtreated in cancer group (h=pb0.01 and i=ns); jcancer alone vs. encapsulated-ATRAtreated in cancer group (pb0.01).
Table 2Pharmacokinetics of ATRA in mice lung tissue.
Groups Eliminationrate constant(ERC) (h)
AUC0–t(μg h/ml)
AUC0–∞(μg h/ml)
Half life inhour (t1/2)
Cmax
(μg/ml)Tmax (hour)
I – – – – – –
II 0.140288 3.79 5.001792 2.813621 7.04 1III 0.04684 7.685 18.14609 14.79812 10 3IV 0.083078 1.634 4.940883 8.343377 2.6 1V 0.06671 7.685 13.68115 10.39053 8.6 1
574 Siddikuzzaman, V.M.B. Grace / International Immunopharmacology 14 (2012) 570–579
showed 0.71±.08 μg/ml and 0.08±.011 μg/ml ATRA levels in lungtissue sample at 1st hour and on 24th hour respectively (Fig. 4). Thet1/2 was found to be 14.79 and 4.94 h for encapsulated ATRA andfree ATRA respectively in non-cancer bearing mice model and thatof 10.39 and 8.34 h respectively in lung cancer bearing mice model.The AUC0-t′ was found to be 7.68 and 3.79 μg h/ml for encapsulatedATRA and free ATRA in non-cancer bearing mice model and that of7.68 and 1.63 μg h/ml respectively in lung cancer bearing micemodel respectively (Table 2).
3.5. Effect of ATRA on hematological parameters
The gram percentage hemoglobulin (gm% Hb) was brought tonear normal by the administration of encapsulated ATRA (11.80±0.30) and free ATRA (11.57±0.60) in comparison with cell lineimplanted alone mice (8.08±0.12) (Fig. 5). The total leukocytelevel also was found to be 9.60±0.50×103 cells/mm3 and 14.35±1.1250×103 cells/mm3 respectively in encapsulated ATRA treatedand non treated cancer bearing mice on the same day (Fig. 5). Thepercentage differential count was significantly decreased to normalrange in treated group (Fig. 5).
Table 1Pharmacokinetics of ATRA in mice serum.
Groups Eliminationrate constant(ERC) (h)
AUC0–t(μg h/ml)
AUC0–∞(μg h/ml)
Half life inhour (t1/2)
Cmax
(μg/ml)Tmax (hour)
I – – – – – –
II 0.127812 2.6775 3.545966 5.423197 0.423 1III 0.07673 4.8665 8.42442 9.033562 0.601 1IV 0.096259 2.15 3.074588 7.20085 0.212 3V 0.098322 4.8665 6.788754 9.033562 0.523 1
3.6. Effect of ATRA on body weight and relative lung weight
Effect of ATRA on body weight and lung weight is given in Fig. 6.The body weight was reduced (22.9±0.40 g) in cancer bearingmice while in encapsulated ATRA treated mice, it was increased(25.9±0.50 g). The relative lung weight was found to be reversedi.e. 1.70±0.17 g in untreated cancer bearing mice and 1.23±0.15 gfor encapsulated ATRA treated group.
3.7. Effect of encapsulated ATRA on lung tumor nodule formation andtumor index
The effect of encapsulated ATRA on inhibition of lung cancer wasassessed by counting the number of lung tumor nodule formed in therespective groups as given in Table 3. The lungs of encapsulatedATRA-treated mice showed less number of tumor nodules (12.20±3.37) compared to those mice that received the tumor cell line only(56.40±6.23), which is comparatively lesser than the values of freeATRA treated group (20.55±4.75). Thus encapsulated ATRA treatmentresulted in 78.37±1.25% inhibition in lung tumor nodule formation.
The effect of ATRA on tumor index is provided in Fig. 7. The tumorindex of tumor cell-only group was 0.82±0.02 and the average indexamong the encapsulated ATRA treated mice was significantly reducedto 0.64±0.02, where as the tumor index of free ATRA treated groupwas found to be 0.68±0.01.
3.8. Histopathologic analysis
The histological changes were completely obvious in the B16 F10cell implanted mice. Representative H&E-stained sections of lung tis-sues recovered from the mice are shown in Fig. 8. The cell lineimplanted alone mice (Gr-II) showed massive tumor growth whenobserved in the lungs of mice and it was found as same massivetumor growth for vehicle group mice (Gr-III and Gr-IV) also. This pa-thology was accompanied by a fibrosis that reduced the alveolarspace, resulting in reduction in the hosts' lung capacity. Large fibrousarea in the interalveolar septa tissue, peribronchial and perivascularregions were seen with apparent infiltration of lymphocytes. Noneof these changes were observed in normal healthy mice (Fig. 8). Inaddition, some areas were characterized by necrosis around the alve-olar passages and bronchioles. Treatment with encapsulated ATRAand free ATRA leads to mitigation in tumor mass around the alveoliand pleura and ATRA could inhibit the fibrosis formation pathwayand resulted in lesser tissue damage (Gr-V and VI). EncapsulatedATRA showed better restoration of tissue architecture comparedwith free ATRA.
3.9. TUNEL assay for DNA fragmentation in lung tissue and TUNEL score
Fig. 9 shows immunohistochemical findings of tumor specimensstained with TUNEL, which detects DNA strand breaks in apoptoticcells. No detectable apoptotic DNA fragmentation was observed inthe lung of normal mice (Gr-I). The TUNEL score was calculated andit was found that the average TUNEL positive signals of apoptoticDNA fragmentation were significantly higher in lung cancer treated

Fig. 5. Effect of encapsulated ATRA on hematological parameters: Values are expressed in mean±SD (n=6). aNormal vs. Cancer alone (pb0.01), bnormal vs. free ATRA treated in cancergroup (pb0.01), e,d,gnormal vs. encapsulated-ATRA treated in cancer group (e=pb0.05, d=ns and g=pb0.01); c,dcancer alone vs. free ATRA treated in cancer group (c=pb0.01 andd=ns), f,dcancer alone vs. encapsulated-ATRA treated in cancer group (pb0.01 and d=ns).
575Siddikuzzaman, V.M.B. Grace / International Immunopharmacology 14 (2012) 570–579
with free ATRA and encapsulated ATRA (Gr-V and VI) respectively(pb0.01) than in the lung cancer bearing alone group (Gr-II) (Fig. 10).
3.10. Effect of encapsulated ATRA on survival rate and percentage increasein life span
Survival rates and percentage increase in life span of mice in eachtreatment groups are provided in Table 4. The lifespan of the encapsu-lated ATRA treated tumor-bearing mice was significantly increased to72.8±6.8 (pb0.01) days in comparison to that of themice that received
Fig. 6. Effect of ATRA on bodyweight and relative lungweight: Values are expressed inmean±treated in cancer group (b=ns and i=pb0.01); cnormal vs. encapsulated-ATRA treated in canalone vs. encapsulated-ATRA treated in cancer group (e=pb0.01 and j=pb0.01).
the tumor cell line only (i.e., 39.3±4.2 days). This represents an in-crease in lifespan of 85.24±2.2% in encapsulated ATRA treated groupwhen compared to free ATRA treated group (71.50±2.5%) (Table 4).As expected, normal mice (who received vehicle only) survived for>100 days.
4. Discussion
In this present study, the effectiveness of liposome encapsulatedATRA as an inhibitor of metastatic lung cancer in C57BL/6 mice was
SD (n=6). a,fNormal vs. cancer alone (a=pb0.05 and f=pb0.01); b,inormal vs. free ATRAcer group (ns); dcancer alone vs. free ATRA treated in cancer group (d=pb0.01); e,jcancer

Table 3Effect of encapsulated ATRA in the inhibition of lung tumor nodule formation.
Groups Number of lung tumornodule/mice
Percent inhibition of lungtumor nodule formation
Group I Nil NilGroup II 56.40±6.23 –
Group III 56.10±5.35 –
Group IV 56.05±5.25 –
Group V 20.55±4.75a 63.56±1.10Group VI 12.20±3.37b 78.37±1.25c
Values are expressed in mean±SD (n=6).a Tumor control vs the free ATRA treated in cancer mice group (pb0.01).b Tumor control vs the encapsulated ATRA treated in cancer mice group (pb0.01).c Free ATRA treatment vs the encapsulated ATRA treated in cancer mice group
(pb0.01).
576 Siddikuzzaman, V.M.B. Grace / International Immunopharmacology 14 (2012) 570–579
studied. The B16F10 mouse melanoma cell line was chosen on thebasis of its high metastatic potential. Specifically, the B16F10 linehas been shown to metastasize to the lungs when injected into thelateral tail vein and the majority of cells find themselves in the pul-monary tissue, but some are also localized in other organs [39] andis accepted as useful model for the study of lung metastasis [40,41].Several other research studies stated the establishment of the exper-imental lung metastasis model or pulmonary metastatic model by in-travenously injected mouse melanoma B16F10 in C57BL/6 mice[35,42,43]. Once B16F10 cells form tumor nodules in the lungs, theyin turn promote lung fibrosis and collagen deposition. However,once melanoma metastasizes, no reliable treatment is currently avail-able which affects the course of disease. After implantation of B16F10cell by tail vein it can metastasize to the lung, liver, brain, bone andlymph nodes [44,45].
It is clear that there is some association between ATRA and the de-velopment of cancers. In an earlier study, we have reported that therewas a significant reduction in serum ATRA level in uterine cervicalcancer patients [46]. While the precise role of ATRA in the cancer pro-cess still not clear, our study results have shown that the free ATRAtreatment could cause a highly significant reduction in lung tumornodule formation. Our study also could show that the reduction inlung tumor nodule formation was furthermore enhanced by treatingwith encapsulated ATRA. Whether this effect is due to enhanced kill-ing of the tumor cells or inhibitory effect on the cancer cell growth/metastasis remains to be discerned. Regardless of the exact mecha-nism, we clearly showed here that the ATRA-induced inhibition oftumor nodule formation correlated well with increased lifespan inlung tumor-bearing mice. The activity of ATRA is mediated by regula-tion of a variety of forms of gene expression through ATRA-dependent
Fig. 7. Effect of encapsulated ATRA on tumor index: Values are expressed in mean±SD (n=vs. encapsulated ATRA treated in cancer mice group (pb0.01).
activation of retinoic acid receptors (RAR) on the nuclear membraneof cancer cells, leading to the growth inhibition, differentiation, and ap-optosis of cancer cells [14]. ATRA has high affinity for ligands to theretinoic acid receptors (RARs). It induces transcriptional regulatory pro-tein and binds to DNA sequences probably within the promoter regionof genes known as retinoic acid response element (RARE) to controlthe gene function. The abnormal regulation of the RARs, loss of normallevel of RARs and loss of RAR-β expression in the presence of physiolog-ical levels of retinoic acid are reported to be associated with variousforms of cancer [47]. Several transcriptional factors like COUP-TF andtwo orphan receptors have been reported to regulate RAR-β expressionin malignant cells out of that orphan receptor TR3 that is deeply in-volved in the regulatory process of ATRA. After TR3/RXR-α heterodimerformation in the nucleus, its subsequent translocation in the cytoplasmdownregulates antiapoptotic protein like Bcl-2 and Bcl-xl [48,49] andinduces apoptotic protein Bax that results in ATRA induced apoptosisin the cancerous cell [50,51]. Desai et al. has demonstrated that thetreatment of ATRA in B16F10 mouse melanoma cells, results in inhibi-tion of cell proliferation by an induction of retinoic acid receptor-β(RAR-β) expression, an increase in protein kinase C alpha (PKCalpha)expression, and enhanced activator protein-1 (AP-1) transcriptional ac-tivity [52]. Thus ATRA can activate different growth regulator and differ-entiation pathways preferentially in B16 melanoma cells to inhibit theB16F10 cell growth. By the encapsulation of ATRA in liposome, themore concentration of ATRA was reached in lung tissue and in theserum level so that the more amount of ATRA at lung tissue mighthave increased the number of ATRA receptor RAR-β and caused the en-hanced inhibition of metastasis. Correspondingly we have observed anincreased life span for encapsulated ATRA treatedmicewhen comparedwith free ATRA treated mice. Further study is required to analyze thelevel of RAR-β expression in encapsulated ATRA treated mice.
The current study showed that the mice received B16F10 cellsonly implanted, the melanoma cells quite well and the lungs ofthese hosts were visibly riddled with metastatic tumor nodules with-in 21 days. In general, the metastatic process comprises a series ofcomplicated events that can be divided into steps involving cell motil-ity, cell adhesion, degradation of extra-cellular matrices, and cell in-vasion. Thus, in the present study, we investigated the ATRA level inserum as well as lung tissue by reverse phase HPLC method. HPLCstudy revealed that the encapsulated ATRA concentration was morein serum and lung tissue sample compared with free ATRA group.
By controlling the concentration of ATRA during the preparation ofliposome, we could get liposome encapsulated ATRA in which ATRAwas loaded at more than 82% yield. The size measurement is an im-portant parameter to predict the stability of liposome carriers. The
6). aCancer alone vs. free ATRA treated in cancer mice group (pb0.01) and bcancer alone
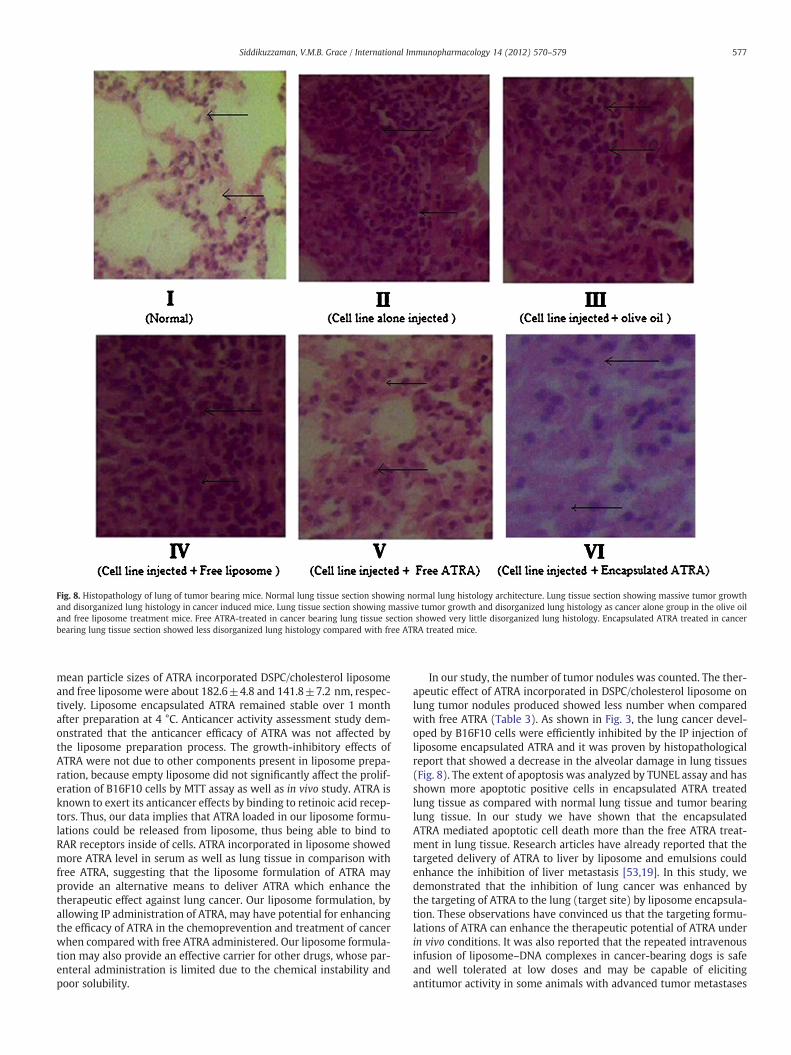
Fig. 8. Histopathology of lung of tumor bearing mice. Normal lung tissue section showing normal lung histology architecture. Lung tissue section showing massive tumor growthand disorganized lung histology in cancer induced mice. Lung tissue section showing massive tumor growth and disorganized lung histology as cancer alone group in the olive oiland free liposome treatment mice. Free ATRA-treated in cancer bearing lung tissue section showed very little disorganized lung histology. Encapsulated ATRA treated in cancerbearing lung tissue section showed less disorganized lung histology compared with free ATRA treated mice.
577Siddikuzzaman, V.M.B. Grace / International Immunopharmacology 14 (2012) 570–579
mean particle sizes of ATRA incorporated DSPC/cholesterol liposomeand free liposomewere about 182.6±4.8 and 141.8±7.2 nm, respec-tively. Liposome encapsulated ATRA remained stable over 1 monthafter preparation at 4 °C. Anticancer activity assessment study dem-onstrated that the anticancer efficacy of ATRA was not affected bythe liposome preparation process. The growth-inhibitory effects ofATRA were not due to other components present in liposome prepa-ration, because empty liposome did not significantly affect the prolif-eration of B16F10 cells by MTT assay as well as in vivo study. ATRA isknown to exert its anticancer effects by binding to retinoic acid recep-tors. Thus, our data implies that ATRA loaded in our liposome formu-lations could be released from liposome, thus being able to bind toRAR receptors inside of cells. ATRA incorporated in liposome showedmore ATRA level in serum as well as lung tissue in comparison withfree ATRA, suggesting that the liposome formulation of ATRA mayprovide an alternative means to deliver ATRA which enhance thetherapeutic effect against lung cancer. Our liposome formulation, byallowing IP administration of ATRA, may have potential for enhancingthe efficacy of ATRA in the chemoprevention and treatment of cancerwhen compared with free ATRA administered. Our liposome formula-tion may also provide an effective carrier for other drugs, whose par-enteral administration is limited due to the chemical instability andpoor solubility.
In our study, the number of tumor nodules was counted. The ther-apeutic effect of ATRA incorporated in DSPC/cholesterol liposome onlung tumor nodules produced showed less number when comparedwith free ATRA (Table 3). As shown in Fig. 3, the lung cancer devel-oped by B16F10 cells were efficiently inhibited by the IP injection ofliposome encapsulated ATRA and it was proven by histopathologicalreport that showed a decrease in the alveolar damage in lung tissues(Fig. 8). The extent of apoptosis was analyzed by TUNEL assay and hasshown more apoptotic positive cells in encapsulated ATRA treatedlung tissue as compared with normal lung tissue and tumor bearinglung tissue. In our study we have shown that the encapsulatedATRA mediated apoptotic cell death more than the free ATRA treat-ment in lung tissue. Research articles have already reported that thetargeted delivery of ATRA to liver by liposome and emulsions couldenhance the inhibition of liver metastasis [53,19]. In this study, wedemonstrated that the inhibition of lung cancer was enhanced bythe targeting of ATRA to the lung (target site) by liposome encapsula-tion. These observations have convinced us that the targeting formu-lations of ATRA can enhance the therapeutic potential of ATRA underin vivo conditions. It was also reported that the repeated intravenousinfusion of liposome–DNA complexes in cancer-bearing dogs is safeand well tolerated at low doses and may be capable of elicitingantitumor activity in some animals with advanced tumor metastases

Fig. 9. Effect of encapsulated ATRA on in situ DNA fragmentation in lung tissue of experimental mice, as detected by TUNEL assay. The apoptotic positive cells are indicated by blackarrow. There is no typical staining for apoptotic DNA fragmentation in normal healthy lung tissue. Profuse intense brown staining as indicated by black arrow, suggesting DNA frag-mentation in cancer group as well as cancer mice which received olive oil (vehicle) and free liposome alone (vehicle) treatment respectively. Lung tissue section of cancer bearingmice after free ATRA treatment showing more number of TUNEL positive cells and lung tissue section of cancer bearing mice received encapsulated ATRA treatment showing evenmore number of TUNEL positive cells in comparison with lung cancer alone group and free ATRA treated lung tissue.
578 Siddikuzzaman, V.M.B. Grace / International Immunopharmacology 14 (2012) 570–579
[54]. However, selective toxicity against cancer cells is required formore efficient systems. Therefore, in the near future, the combinationof cytokine therapy and such type of differentiation therapy could beused to improve the treatment of lung cancer.
In conclusion, the present study in a mice model showed the useof encapsulated ATRA that not only prevent lung cancer but also
Fig. 10. Effect of encapsulated ATRA on apoptosis in lung tissue: Values are expressedin mean±SD (n=6). The significant difference was determined between: acanceralone vs. free ATRA treated in cancer mice group (pb0.01) and acancer alone vs.encapsulated-ATRA treated in cancer group (pb0.01).
may able to inhibit lung metastasis compared with free ATRA. In thelung cancer developed by implanted with B16F10 cells in C57BL/6mice, encapsulated ATRA in liposome was particularly effective insuppressing lung cancer compared with free ATRA. The reversephase HPLC method revealed that the level of ATRA was more inserum and lung tissue in encapsulated ATRA treated group as com-pared with free ATRA group. So the liposome formulation helped toachieve more ATRA concentration at targeted site. Although many ad-ditional studies of relevant tumor models are required to improve theefficacy of ATRA incorporated into liposome. The results in this studywarrant further development of DSPC/cholesterol liposome to inves-tigate their use in clinical differentiation therapy. Even in the absenceof the mechanism in detail, the results of these studies allow us tosuggest to other investigators/clinicians about the potential utility ofencapsulated ATRA as an effective therapeutic agent against lungcancers.
Table 4Effect of encapsulated ATRA on survival rate and increase in life span (ILS).
Groups Number of days survived Increase in life spana (%)% ILS=T−C/C×100
Group I 100b –
Group II 39.3±4.20 –
Group III 39.5±6.50 –
Group IV 40.6±3.30 –
Group V 67.4±7.20c 71.50±2.50Group VI 72.8±6.80d 85.24±2.20e
Values are expressed in mean±SD (n=6).a T = no. of days ATRA treated mice survived and C = no. of days control mice
survived.b After 100 days, observations of Group I mice were halted as it is normal group and
survived longer than either other treated group.c Tumor control vs the Free ATRA treated in cancer mice group (pb0.01).d Tumor control vs the encapsulated ATRA treated in cancer mice group (pb0.01).e Free ATRA treatment vs the encapsulated ATRA treated in cancer mice group
(pb0.01).

579Siddikuzzaman, V.M.B. Grace / International Immunopharmacology 14 (2012) 570–579
Acknowledgment
The authors would like to acknowledge the University GrantsCommission (UGC) for supporting financially this study throughMaulana Azad Fellowship for Minority Students and also would liketo acknowledge the valuable technical help of Dr. Nahid Ali, SeniorPrincipal Scientist, Infectious Diseases & Immunology Division, IndianInstitute of Chemical Biology, Kolkata.
References
[1] Steeg PS. Metastasis suppressors alter the signal transduction of cancer cells. NatRev Cancer 2003;3:1-9.
[2] Stafford LJ, VaidyaKS,Welch DR.Metastasis suppressors genes in cancer. Int J BiochemCell Biol 2008;40:874-91.
[3] Siddikuzzaman, Guruvayoorappan C, Berlin Grace VM. All trans-retinoic acid andcancer. Immunopharmacol Immunotoxicol 2011;33:241-9.
[4] Orlandi M, Mantovani B, Ammar K, Avitabile E, Dal Monte P, Bartolini G. Retinoidsand cancer: antitumoral effects of ATRA, 9-cis RA and the new retinoid IIF on theHL-60 leukemic cell line. Med Princ Pract 2003;12:164-9.
[5] Peter W, Jessica LP, Kristie MO, Matthew JW, Margaret KMa, Anthony W, et al.Adaptive immunity cooperates with liposomal all-trans-retinoic acid (ATRA) tofacilitate long-term molecular remissions in mice with acute promyelocytic leu-kemia. PNAS 2002;99:9468-73.
[6] Crowe DL, Kim R, Chandraratna RA. Retinoic acid differentially regulates cancercell proliferation via dose-dependent modulation of the mitogen-activated pro-tein kinase pathway. Mol Cancer Res 2003;1:532-40.
[7] Gumireddy K, Sutton LN, Phillips PC, Reddy CD. All-trans-retinoic acid-inducedapoptosis in human medulloblastoma: activation of caspase-3/poly (ADPribose)polymerase 1 pathway. Clin Cancer Res 2003;9:405-9.
[8] Czeczuga-Semeniuk E, Anchim T, Dzieciol J, Dabrowska M, Wolczynski S. Cantransforming growth factor-β1 and retinoids modify the activity of estradioland anti-estrogens in MCF-7 breast cancer cells? Acta Biochim Pol 2004;51:733-45.
[9] LiuWJ, Zhang YW, Zhang ZX, Ding J. Alternative splicing of human telomerase reversetranscriptase may not be involved in telomerase regulation during all-trans-retinoicacid-induced HL-60 cell differentiation. J Pharmacol Sci 2004;96:106-14.
[10] Siddikuzzaman Berlin Grace VM. Evaluation of immunomodulatory and anti-tumor ac-tivity of all trans retinoic acid (ATRA) in solid tumor bearing mice. ImmunopharmacolImmunotoxicol 2012;34, http://dx.doi.org/10.3109/08923973.2012.712137.
[11] Dahl AR, Grossi IM, Houchens DP, Scovell IJ, Placke MF, Imondi AR. Inhaled isotret-inoin (13-cis-retinoic acid) is an effective lung cancer chemopreventive agent inA/J mice at low doses: a pilot study. Clin Cancer Res 2000;6:3015-24.
[12] Chang YS, Chung JH, Shin DH, Chung KY, Kim YS, Chang J. Retinoic acid receptor-βexpression in stage I non-small cell lung cancer and adjacent normal appearingbronchial epithelium. Yonsei Med J 2004;45:435-42.
[13] Marion F, Rainer WH, Bernhard F, Karl H, Konrad LM, Joachim H, et al. Alltrans-retinoic acid selectively down-regulates matrix metalloproteinase-9 (MMP-9)and up-regulates tissue inhibitor of metalloproteinase-1 (TIMP-1) in human bron-choalveolar lavage cells. Mol Med 2001;7(4):263-70.
[14] Fang J, Chen SJ, Tong JH, Wang ZG, Chen GQ, Chen Z. Treatment of acutepromyelocytic leukemia with ATRA and As2O3: a model of molecular target-basedcancer therapy. Cancer Biol Ther 2002;6:614-20.
[15] Kawakami S, Suzuki S, Yamashita F, Hashida M. Induction of apoptosis in A549human lung cancer cells by all-trans retinoic acid incorporated in DOTAP/cholesterolliposomes. J Control Release 2006;110(3):514-21.
[16] Elstner E, Muller C, Koshizuka K, Williamson EA, Park D, Asou H, et al. Ligands forperoxisome proliferator-activated receptor γ and retinoic acid receptor inhibitgrowth and induce apoptosis of human breast cancer cells in vitro and in BNXmice. Proc Natl Acad Sci 1998;95(15):8806-11.
[17] Toma S, Isnardi L, Riccardi L, Bollag W. Induction of apoptosis in MCF-7 breast car-cinoma cell line by RAR and RXR selective retinoids. Anticancer Res 1998;18(2A):935-42.
[18] Mangiarotti R, Danova M, Alberici R, Pellicciari C. All-trans retinoic acid(ATRA)-induced apoptosis is preceded by G1 arrest in human MCF-7 breast can-cer cells. Br J Cancer 1998;77(2):186-91.
[19] Chansri N, Kawakami S, Yamashita F, Hashida M. Inhibition of liver metastasis byall-trans retinoic acid incorporated into O/W emulsions in mice. Int J Pharm2006;321:42-9.
[20] Suzuki S, Kawakami S, Chansri N, Yamashita F, Hashida M. Inhibition of pulmo-nary metastasis in mice by all-trans retinoic acid incorporated in cationic lipo-somes. J Control Release 2006;116:58-63.
[21] Chen DB, Yang TZ, Lu WL, Zhang Q. In vitro and in vivo study of two types oflong-circulating solid lipid nanoparticles containing paclitaxel. Chem Pharm Bull2001;49:1444-7.
[22] Muindi J, Frankel SR, Miller Jr WH, Jakubowski A, Scheinberg DA, Young CW, et al.Continuous treatment with all-trans retinoic acid causes a progressive reductionin plasma drug concentrations: implications for relapse and retinoid “resistance”in patients with acute promyelocytic leukemia. Blood 1992;79:299-303.
[23] Ozpolat B, Lopez-Berestein G, Adamson P, Fu CJ, Williams AH. Pharmacokinetics ofintravenously administered liposomal all-trans-retinoic acid (ATRA) and orallyadministered ATRA in healthy volunteers. J Pharm Pharm Sci 2003;6:292-301.
[24] Choi Y, Kim SY, Kim SH, Yang J, Park K, Byun Y. Inhibition of tumor growth by bio-degradable microspheres containing all-trans-retinoic acid in a human head-and-neck cancer xenograft. Int J Cancer 2003;107:145-8.
[25] Lim SJ, Kim CK. Formulation parameters determining the physicochemical charac-teristics of solid lipid nanoparticles loaded with all-trans retinoic acid. Int J Pharm2002;243:135-46.
[26] Brisaert M, Gabriels M, Plaizier-Vercammen J. Investigation of the chemical stabilityof an erythromycin–tretionin lotion by theuse of an optimization system. Int J Pharm2000;197:153-60.
[27] Bhowmick S, Ali N. Identification of novel Leishmania donovani antigens that helpdefine correlates of vaccine-mediated protection in visceral leishmaniasis. PLoSOne 2009;4:1–10 (e5820).
[28] Talwar GP. Handbook of practical immunology. New Delhi: National Book Trust;1974. p. 336-9.
[29] Cole SP. Rapid chemosensitivity testing of human lung tumor cells using MTTassay. Cancer Chemother Pharmacol 1986;17:259-63.
[30] Campling BG, Pym J, Baker HM, Cole SP, Lam YM. Chemosensitivity testing ofsmall cell lung cancer using the MTT assay. Br J Cancer 1991;63:75-83.
[31] Wyss R. Chromatography of retinoids. J Chromatogr 1990;531:481-508.[32] Pikkarainen SA, ParviainenMT. Determination of retinyl palmitate and total vitamin
A content in liver and liver-based ready-to-eat foods. J Chromatogr 1992;577:163-6.[33] Benjamin NR. A rapid method for estimation of the total leukocyte count. Blood
1985;13:677-83.[34] Drabkin DL, Austin JM. Spectrometric studies; spectrometric constants for com-
mon hemoglobin derivatives in human, dog and rabbit blood. J Biol Chem1932;98:719-33.
[35] Gautam A, Waldrep JC, Densmore CL. Inhibition of experimental lung metastasisby aerosol delivery of PEI–p53 complexes. Mol Ther 2000;2:318-23.
[36] Gautam A, Waldrep JC, Densmore CL, Koshkina N, Melton S, Roberts L, et al.Growth inhibition of established B16-F10 lung metastases by sequential aerosoldelivery of p53 gene and 9-nitrocamptothecin. Gene Ther 2002;9:353-7.
[37] Booran GA, Eustis SL, Elwell MR, Montgory CA, MaKenzie WF. Pathology of thefisher rat, 1. San Diego: Academic Press; 1990. p. 405-18.
[38] Guruswamy Suresh, Lightfoot Stan, Michael AG, Raffit H, Darrell Berlin K, Todd IR,et al. Effects of retinoids on cancerous phenotype and apoptosis in organotypiccultures of ovarian carcinoma. J Natl Cancer Inst 2001;93(7):516-25.
[39] Fidler IJ. Selection of successive tumor lines for metastasis. Nature (London)1973;242:148-9.
[40] Brunda MJ, Luistro L, Warrier RR, Wright RB, Hubbard BR, Murphy M, et al.Anti-tumor and anti-metastatic activity of IL-12 against murine tumors. J ExpMed 1993;178:1223-30.
[41] Teng MW, von Scheidt B, Duret H, Towne JE, Smyth MJ. Anti-IL-23 monoclonal an-tibody synergizes in combination with targeted therapies or IL-2 to suppresstumor growth and metastases. Cancer Res 2011;71:2077-86.
[42] Yao Z, Xu-chun C, Rong L, Min-na Z, Zhi-feng Z, Jin-ping L, et al. Inhibition bytyroserleutide (YSL) on the invasion and adhesion of the mouse melanoma cell.Mol Med 2007;13(1–2):14-21.
[43] Pongpun S, Jantana Y, Suratsawadee P, Kwanjai K, Hiroyuki K, Takayuki I, et al. In-hibitory effect of liposomal rhinacanthin-N isolated from Rhinacanthus nasutus onpulmonary metastasis in mice. Biol Pharm Bull 2012;35(7):1197-200.
[44] Garbe C, McLeod GR, Buettner PG. Time trends of cutaneous melanoma inQueensland, Australia and Central Europe. Cancer 2000;89:1269-78.
[45] Morton DL, Essner RE, Kirkwood JM, Wollman RC. Malignant melanoma. In: KufeDW, Pollock RE, Weichselbaum RR, Bast Jr RC, Gansler TS, Holland JF, Frei III E, ed-itors. Holland Frei Cancer Medicine, 6. Hamilton, Canada: BC Decker Inc. 2003Chapter 10.
[46] Berlin-Grace VM, Niranjali DS, Radhakrishnan PM, Devaraj H. HPV-induced carci-nogenesis of the uterine cervix is associated with reduced serum ATRA level.Gynecol Oncol 2006;103:113-9.
[47] Toulouse A, Morin J, Dion PA, Houle B, Bradley WE. RARbeta 2 specificity in medi-ating retinoic acid inhibition of growth of lung cancer derived cells. Lung Cancer2000;28(2):127-37.
[48] Fujimura S, Suzumiya J, Yamada Y, Kuroki M, Ono J. Downregulation of Bcl-xl andactivation of caspases during retinoic acid induced apoptosis in an adult T-cellleukemia cell line. Hematol J 2003;4(5):328-35.
[49] Pettersson F, Dalgleish AG, Bissonnette RP, Colston KW. Retinoids cause apoptosisin pancreatic cancer cells via activation of RAR-gamma and altered expression ofBcl-2/Bax. Br J Cancer 2002;87(5):555-61.
[50] Palozza P, Serini S, Maggiano N, Angelini M, Boninsegna A, Di Nicuolo F, et al. In-duction of cell cycle arrest and apoptosis in human colon adenocarcinoma celllines by beta carotene through down regulation of cyclin A and Bcl-2 family pro-teins. Carcinogenesis 2002;23(1):11-8.
[51] Biswajit M, Miltu KG, Chowdhury MH. Anticancer potential of vitamin A andbeta-carotene: mechanistic approach. NSHM J Pharm Healthc Manag 2011;02:1–12.
[52] Desai SH, Boskovic G, Eastham L, Dawson M, Niles RM. Effect of receptor-selectiveretinoids on growth and differentiation pathways in mouse melanoma cells.Biochem Pharmacol 2000;59(10):1265-75.
[53] Shimizu K, Tamagawa K, TakahashiN, TakayamaK,Maitani Y. Stability and antitumoreffects of all-trans retinoic acid-loaded liposomes contained sterylglucoside mixture.Int J Pharm 2003;258:45-53.
[54] DowS, Elmslie R, Kurzman I,MacEwenG, Pericle F, Liggitt D. Phase I study of liposome–DNA complexes encoding the interleukin-2 gene in dogs with osteosarcoma lung me-tastases. Hum Gene Ther 2005;16(8):937-46.