influence of nonionic organic solutes on various reactions of energy conservation and utilization
TRANSCRIPT

INFLUENCE OF NONIONIC ORGANIC SOLUTES ON VARIOUS REACTIONS OF ENERGY
CONSERVATION AND UTILIZATION
Thomas E. Conover
Depart men! of Biological Chemistry Hahnemann Medical College
Philadelphia, Pennsylvania 19102
Unlike many of the papers to be included in this monograph, this paper will not elaborate on the various unique properties of dimethyl sulfoxide; rather it will dis- cuss some of the effects that it shows in common with a broader class of com- pounds, the water-soluble, organic solvent. In apology, I must confess that the main thrust of this work was not toward dimethyl sulfoxide at all, although the effects of this solvent have been studied. The major prototype chosen in this work was gly- cerol; I hope, however, to indicate the various similarities in their effects. It might be mentioned here that although most of the compounds studied can be called solvents, they have been studied in aqueous solutions, and therefore will be referred to as solutes.
The study of enzymic reactions in aqueous solutions of organic solvents goes back to the work of Nelson and Schubert,’ whose studies on the effect of alcohol on the hydrolysis of sucrose catalyzed by “invertase” established the role of water in this enzymic reaction in 1928. Numerous other workers have studied purified single- enzyme systems by this means.*J In such systems, the effect of solvent can often be attributed to its effect of decreasing the water concentration in the reactions that in- volve water as a p a r t i ~ i p a n t . ~
The problem of applying such an approach to a system as complex and ill-under- stood as that of mitochondria1 energy conservation is obviously a more precarious task, although water, since it is a primary product of ATP synthesis, must be inti- mately involved in these processes. Indeed, Mitchell’s chemiosmotic theory‘ notwithstanding, a major reason, if not the primary one, for the vital importance of the integrity of the mitochondrial membrane in the processes of energy conservation lies in the need to exclude water, both environmental water and, perhaps, that produced at the sites of energy coupling.
In spite of such reservations, this problem was examined. At its initiation, this investigation was guided by the fond hope that interference with the active water concentrations by highly soluble, biologically innocuous solutes might lead t o a sta- bilization of high-energy intermediates and to a possible increase in the P:O ratio and the rate of energy-linked reactions.
Eflect of Glycerol on Respiration and Oxidative Phosphorylation in Submitochondrial Particles
In these studies, submitochondrial particles prepared from bovine heart mitochondria by sonication were Such preparations have the advantage of good stability, and do not present the problem of osmotic effects due to high solute concentration, which might be anticipated with intact mitochondria. When such a preparation was incubated with varying concentrations of glycerol in aqueous me-
24

Conover: Influence of Nonionic Organic Solu tes 25
2 4 6 8 %, GLYCEROL, MOLAR
1 , I I I I 2 4 6 8
GLYCEROL CONCENTRATION, MOLAR
FIGURE 1. Effect of glycerol on respiration and oxidative phosphorylation in submitochon- drial particles. The reaction medium contained 20 pmol triethanolamine-HCI (pH 7.4). 8 pmol MgCI,, 4 pmol ATP, 1 pmol EDTA, 20 pmol S*P-orthophosphate (lo4 cpmlpmol, pH 7.4); 24 pmol glucose, 0.2 mg hexokinase, 2 mg bovine serum albumin, and 1.5-3 mg submitochondrial particles in a volume of 2.0 ml. The substrates were 20 pmol succinate with 2 pmol malonate in one case, and 40 pmol m-8-hydroxybutyrate with 0.5 pmol NAD+ in the other. The glycerol concentration was as indicated. Oxygen consumption was followed polarographically, and the 32P-orthophosphate esterification was determined after deproteinization, as described elsewhere.E The oxygen content of mixtures of glycerol and water was determined from values given in the International Critical Tables.' The temperature was 25'C.
dium, and the oxygen uptake and the inorganic phosphate esterification in the presence of a hexokinase-glucose coupling system were followed, the results shown in FIGURE I were obtained. With a very active substrate such as succinate, a linear decrease in the oxygen consumption was observed as the concentration of glycerol was increased. With the NAD+-linked substrate @-hydroxybutyrate, which is oxidized at a considerably lower rate than succinate, little effect was seen until the concentration of glycerol exceeded 3.0 M. This concentration of glycerol decreased succinate oxidation to a rate similar t o that of the @-hydroxybutyrate oxidation. At concentrations of glycerol above 3.0 M, the rate of respiration with @- hydroxybutyrate showed a linear decline of activity with the increasing glycerol con- centration, parallel to that of succinate. It is presumed that the rate-limiting step in @-hydroxybutyrate oxidation is the dehydrogenase step; and inhibition of glycerol did not become apparent until the respiratory chain was sufficiently inhibited to be- come limiting. The P:O ratio was affected in the opposite manner. The ratio with @- hydroxybutyrate was rapidly decreased at low concentrations of glycerol, pre- sumably because phosphate esterification was being inhibited while oxygen consumption was not. With succinate, the P:O ratio declined less sharply, since suc- cinate oxidation was also being inhibited at these concentrations.
It appears, therefore, that electron transport is inhibited in a simple linear relation with increasing concentrations of glycerol. No simple relation in the case of

26 Annals New York Academy of Sciences
the P:O ratio was apparent, however, nor was there any apparent stabilization of energy coupling which it had been hoped would be observed with the increased gly- cerol concentrations. I t may be pointed out, however, that a t least a portion of the phosphate esterification reaction seemed to be quite resistant to glycerol inhibition. Whether this might be due to a higher stability of one specific site of phosphate esterification or to a nonspecific resistance of the process of phosphate esterification to high concentrations of glycerol is not clear.
Effect of Organic Solutes on Electron Transport in Submitochondrial Particles
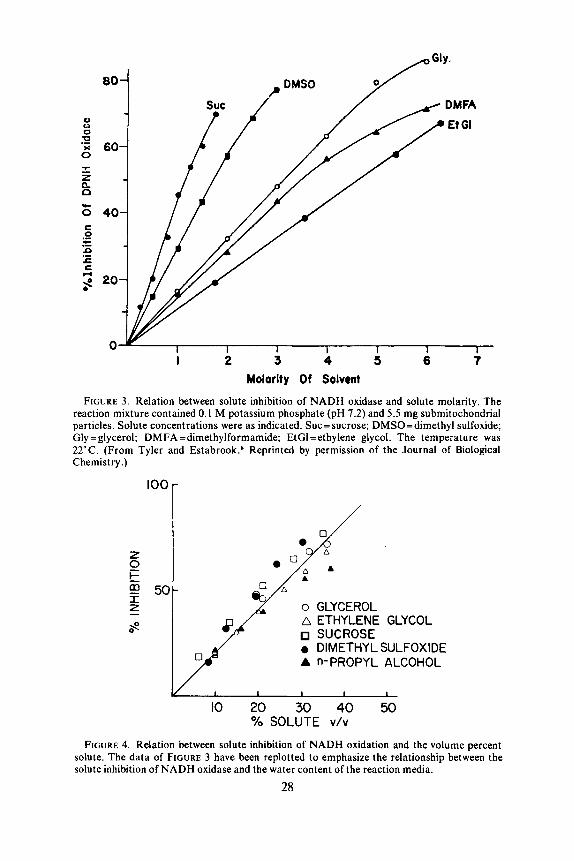
Tyler and Estabrooks have studied the electron transport chain in similar systems by means of spectrophotometric measurement of the components of the chain, for which they used a dual-wavelength recording spectrophotometer. Since the rate of reduction of each individual component may be followed in this manner, it is possible to determine both the steady-state level of reduction of the component and the rate of substrate oxidation. In FIGURE 2 the values they obtained for both cytochrome c + c , components (551-540 nm) and cytochrome a + a3 components (605-625 nm) are shown. It can be seen that the inhibition of NADH oxidation was linear with glycerol concentration, giving 50% inhibition at about 3.0 M. This is similar to the data shown in FIGURE 1 for succinate oxidation. It is most interesting, however, that a t all concentrations of glycerol the steady-state reduction level of the components did not vary, but acted as if they were frozen in place. This is very different from the situation that appears with the usual site-specific inhibitors, such as antimycin A or cyanide, which produce marked changes in the steady-state level of reduction, depending on the site of inhibition.* This constant level of reduction at steady state must result from a nonspecific inhibition of all components, which ap- parently affects all components in a quantitatively similar manner. This, then, might be termed a nonspecific inhibition, and must affect some element common to all components and their reactions.s
FIGURE 3 shows the effect of various other organic solutes, including dimethyl sulfoxide, on the oxidation of NADH by submitochondrial particles. One of these solutes, sucrose, has been previously reported to inhibit electron transport and oxi- dative phosphorylation a t higher concentrations. It may be seen that although the concentration required to produce a 50% inhibition of respiration varied widely, sucrose being the most effective and ethylene glycol the least, the inhibitions all showed a simple linear relationship with concentration up to a t least 60 or 70% in- hibition. Furthermore, these inhibitions were all reversible, and all appeared t o be of the same nonspecific type.s
It has been suggested that one effect of glycerol might be to increase the viscosity of the medium, thereby perhaps hindering the complex interaction of components.12 The data of Tyler and Estabrook* would clearly eliminate such a consideration, since there would appear to be little relation between viscosity and the degree of in- hibition. For instance, dimethyl sulfoxide proved to be an effective nonspecific in- hibitor, but solutions of dimethyl sulfoxide do not show much of an increase in vis- cosity, a t least compared to solutions of sucrose or glycerol.8
Since our concern here is with the modification of the water content of the me- dium by the solutes, the inhibitions may better be expressed in relation to the volume of water or solute as a percentage of the total solvent volume. In FIGURE 4, the data of FIGURE 3 have been replotted in terms of the percentage of solute, on a volume/

100
80
60
40
20.
0 .
Conover: Influence of Nonionic Organic Solutes
*/* W Inhibition / Molarity
Reduction in Steady State
I I i I I I .o 2x) 3.0 4.0 5.0
~
Glycerol Concentration Molar
27
FIGURE 2. Effect of glycerol on the NADH oxidase and the steady-state reduction of endogenous cytochrome components. Reaction mixtures contained 0.1 M potassium phos- phate buffer (pH 7.2), 0.18 mM NADH, and 6 mg submitochondrial particles. The glycerol concentration was as indicated. The temperature was 22" C. The kinetics of cytochrome reduc- tion were recorded in a dual-wavelength spectrophotometer, set at 55 1-540 mp (cytochrome c + c,) or 605-625 mp (cytochrome (I + a3). Two plots of percentage inhibition versus molarity are shown, which were based on the degree of inhibition calculated from the spec- trophotometric traces obtained at the two pairs of wavelength settings used. The percentage steady-state reduction of cytochrome was calculated as the absorbance change at steady state relative to the overall change between the fully oxidized and the fully reduced condition. For more details, see the original paper. (From Tyler and Estabrook.8 Reproduced by permission of the Journal of Biological Chemistry.)
volume basis, which would be one way of expressing this relationship. Tyler and Estabrook used a g/g basis, which did not always appear to give as good a fit for suc- rose in our results.8 It may be seen that with all of these compounds, a congruent relation now appears to exist between the inhibition of electron transport and the water content. One might note that dimethyl sulfoxide is perhaps the most effective inhibitor among these compounds, which suggests that it may either have other effects or may be in some way more effective in reducing the active water content. Nevertheless, the conclusion is implied that the nonspecific inhibitory effect of the various solutes was related to a reduction in the water content of the reaction mix- tures, and was independent of the chemical nature of the substance used.
0- I I I I I 1 I I 2 3 4 5 6 7
Molarity Of Solvent
FIGURE 3. Relation between solute inhibition of NADH oxidase and solute molarity. The reaction mixture contained 0.1 M potassium phosphate (pH 7.2) and 5.5 mg submitochondrial particles. Solute concentrations were as indicated. Suc=sucrose; DMSO=dimethyl sulfoxide; Gly =glycerol; DMFA=dimethylformamide: EtGl=ethylene glycol. The temperature was 22°C. (From Tyler and Estabrook.* Reprinted by permission of the Journal of Biological Chemistry.)
2 0 t E? 50 I z - I
o GLYCEROL a ETHYLENE GLYCOL
SUCROSE 0 DIMETHYL SULFOXIDE A n-PROPYL ALCOHOL
1 I 1
10 20 30 40 50 O/o SOLUTE V/V
FIGURE 4. Relation between solute inhibition of NADH oxidation and the volume percent solute. The data of FIGURE 3 have been replotted to emphasize the relationship between the solute inhibition of NADH oxidase and the water content of the reaction media.
28

Conover: Influence of Nonionic Organic Solutes 29
Effect of Organic Solutes on Energy-linked Reactions in Submitochondrial Particles
When we turn to the effects of these compounds on energy-conservation reac- tions, the situation becomes more complex, as is shown in FIGURE 5 . In this figure the effects of glycerol on a number of reactions related t o energy conservation are shown. These are oxidative phosphorylation, the 32Pi -ATP exchange reaction, AT- Pase activity, and the succinate-linked reduction of NAD+ driven by ATP. The ex- change reaction and the ATPase activity represent what is thought to be a partial reversal of the phosphate esterification reaction. The succinate-linked reduction of NAD+ represents the total reversal of oxidative phosphorylation, in that a reversal of electron transport driven by the hydrolysis of ATP is measured. It can be seen
o P:O RATIO 0 NADt REDUCTION 8 ATPose ACTIVITY 0 f'i32-ATP EXCHANGE
I I Y 10 20 30 40 50
O/O glycerol - v/v FIGURE 5. The effect of glycerol on related reactions of oxidative ,phosphorylation in
submitochondrial particles. The reaction medium for oxidative phosphorylation was as described in the legend to FIGURE 1, with succinate as substrate. The reaction medium for the 32Pi-ATP exchange contained 16 pmol ATP, 16 pmol MgCl,, 1 pmol EDTA, 40 pmol 32P-or- thophosphate (10' cpm/pmol, pH 7.4), and 1 mg submitochondrial particles, in a final volume of 1.0 ml. The reaction medium for the ATPase assay contained 50 pmol triethanolamine-HCl (pH 7.4). 3.0 pmol MgCI,, 6.0 pmol ATP, 0.8 pmol 2,4-dinitrophenol, 5.0 pmol phos- phoenolpyruvate, 0.05 mg pyruvate kinase, and 0.4 mg submitochondrial particles, in a volume of 1.0 ml. The reaction medium for the energy-linked reduction of NAD+ by succinate contained 50 pmol triethanolamine-HC1 (pH 7.4). 10 pmol MgCI,, 1 pmol ATP, 0.5 pmol NAD+, 10 pmol succinate, 1 pmol Na& 0.5 pmol EDTA, 1 mg bovine serum albumin, and 0.4 mg submitochondrial particles, in a fiqal volume of 2.0 ml. The glycerol concentration was as indicated in each case. The temperature was 25'C for all assays. 0----0 = P:O ratio (activity without glycerol, 0.88); .----a = a'q -ATP exchange reaction (activity without glycerol, 0.14 pmollmglmin); e----e = ATPase activity (activity without glycerol, 0.65 pmollmglmin); @-----ID = energy-linked NAD+ reduction by succinate (activity without glycerol, 0.027 pmollmglmin). (From Conover.' Reproduced by permission of the Journal of Biological Chemistry.)

30 Annals New York Academy of Sciences
that these reactions do not show the linear inhibition relative to concentration ob- served with electron transport, but rather a complex family of sigmoid curves. Low concentrations of glycerol often actually stimulated activity, as is shown here for NAD+ reduction. This phenomenon was commonly observed with the ATPase activity, and occasionally even with the exchange reaction as well. This was followed by a rather sharp drop in activity as the concentrations increased. It is possible that the sigmoidicity may result from the observed stimulation by the solute and a si- multaneously occurring linear inhibition. On the other hand, oxidative phos- phorylation as measured by the P:O ratio declined rather less rapidly, as we noted earlier. This was undoubtedly due in part t o the simultaneous inhibition of both respiration and phosphate esterification. However, when the actual rate of or- thopho~phate-~~P esterification during both oxidative phosphorylation and the 32Pi - ATP exchange under similar conditions was determined (FIGURE ti), it can be seen that the exchange was considerably more sensitive to inhibition by the organic solute. Although the rate of the exchange was more rapid in the absence of glycerol, it actually dropped below that of oxidative phosphorylation at higher concentrations of solute. This is possibly related to the fact that the 32Pi-ATP exchange reaction in contrast to oxidative phosphorylation involves the reactions of phosphate esterification in both forward and reverse directions. Indeed, as mentioned, all the reactions shown in FIGURE 5 require the reversal of these reactions, except oxidative phosphorylation. It is possible that the reverse reaction is somewhat more sensitive to this inhibition by organic solutes.
0 SUCCINATE OXIDATION @ p -HYDROXYBUTYRATE
OXIDATION pi32-ATP EXCHANGE
10 20 30 40 50 */,, glycerol - v/v
FIGURE 6. A comparison of the rates of orthophosphate -*lP esterification during oxidative phosphorylation and the -ATP exchange reaction in the presence of glycerol. The condi- tions for the determination of these rates were those described in the legends to FIGURES 1 and 5. o---o = Oxidative phosphorylation with succinate as substrate; @----a = oxidative phosphorylation with 6-hydroxybutyrate as substrate; o---• = -ATP exchange reaction. (From Conover.e Reproduced by permission of the Journal of Biological Chemistry.)

Conover: Influence of Nonionic Organic Solutes 3 1
i n n a z
10 20 30 40 50 '10 glycerol - v/v
FIGURE 7. The effect of glycerol on the energy-linked transhydrogenase activity of submitochondrial particles. The initial reaction contained 50 pmol triethanolamine-HCI (pH 7.4). 0.5 pmol NADP+, 0.1 pmol NAD+, 2 pmol Na,S, 0.5 pmol EDTA, 85 pmol ethanol, 0.03 mg alcohol dehydrogenase, 1 mg bovine serum albumin, and 0.4 rng submitochondrial parti- cles, in a volume of 2.0 ml. After the initial rate of NADP+ reduction had been recorded, 20 pmol MgCl, were added, and the rate was recorded again. Finally, I pmol ATP was added and the rate was recorded again. The temperature was 25' C. 0----0 = initial rate; a----o = rate after addition of MgCI,; o---• = rate after addition of ATP. (From Conover.a Reproduced by permission of the Journal of Biological Chemistry.)
FIGURE 7 shows another possible example of this: the effects of glycerol on the NADH-NADP+ energy-driven transhydrogenase activity of submitochondrial par- ticles. This rather unusual reaction of mitochondria shows an inhibition by organic solutes that is logarithmic with solute concentration, for reasons that are not under- stood a t this time. Among the reactions of energy conservation in mitochondria, this one frequently shows rather unique ~ r 0 p e r t i e s . l ~ I t can be seen that the effect of glycerol on the initial rate, in the absence of ATP, was less marked than its effect on the rate after the addition of Mg++ and ATP to drive the reaction.
That we are still talking about a nonspecific inhibition similar to that observed with electron transport can be seen in FIGURES 8 and 9, which demonstrate the simi- larity of the inhibitions by a variety of the organic solutes. FIGURE 8 shows the effects on the 3zPi -ATP exchange. The polyhydroxyl compounds glycerol, ethylene glycol, and sucrose all showed excellent agreement when the results were plotted by percent volume against concentration. Dimethylformamide, however, was some- what more inhibitory, for unexplained reasons. Dimethyl sulfoxide again appeared to give an inhibition similar to the others, although as with electron transport, it seemed to be somewhat more effective than the polyols. By contrast, n-propyl alcohol was shown to be a much more inhibitory compound, producing a marked, irreversible inhibition of these reactions. This is presumably related to the more asymmetric polarity of primary alcohols and their possible effects on membrane

100
z o GLYCEROL 0 n ETHYLENE GLYCOL k u SUCROSE I 50 0 DIMETHYLSULFOXIDE z - A DIMETHYLFORMAMIDE
x n-PROPYL ALCOHOL 8
10 20 30 40 50 O/' SOLUTE V / V
FIGURE 8. The effect of various organic solutes on the "Q-ATP exchange reaction of submitochondrial particles. The conditions for the assay of the 32Pi -ATP exchange reaction were those described in the legend to FIGURE 5. o--o=Glycerol; A----A=ethykne glycol; O----O = sucrose; o---• =dimethyl sulfoxide; A--A= dimethylformamide; x ---x = n-propyl alcohol.
100
> > t 0
t .- .- 50
,\"
o GLYCEROL A ETHYLENE GLYCOL
SUCROSE 0 DIMETHYL SULFOXIDE A DIMETHYL FORMAMIDE X n-PROPYL ALCOHOL
A
10 2 0 30 40 50 O/,, solute - v/v
FIGURE 9 . The effect of various organic solutes on the ATPase activity of submitochondrial particles. The reaction mixture in each case contained 80 pmol triethanolamine-HCI (pH 7.4), 8 pmol MgCI,, 8pmol ATP, 2 pmol 2,4-dinitrophenol, and 0.2 mg submitochondrial particles, in a total volume of 4.0 ml. The concentrations of solute were as indicated. The tempera- ture was 25°C. o--o=glycerol; &---&=ethylene glycol; n---o=sucrose; *---• =dimethyl sulfoxide; r-----=dimethylformamide; x ---x= n-propyl alcohol.
32
Conover: Influence of Nonionic Organic Solutes 33
structure. FIGURE 9 shows a similar study of the ATPase activity, which produced very nearly the same picture. Clearly, there again appears to be a common relation- ship between the extent of inhibition by these organic solutes and the decrease in the water content of the reaction mixture.
Reversibility of Solute Inhibition of Energy-linked Reactions
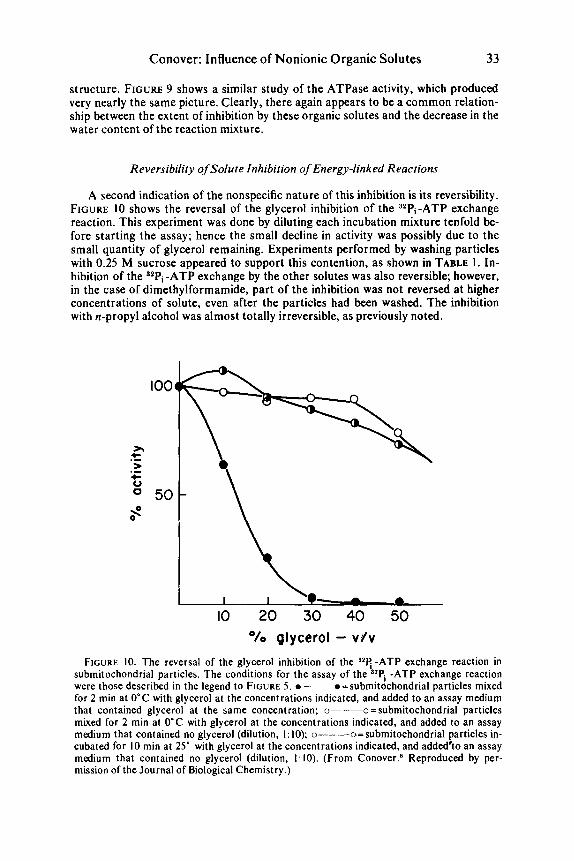
A second indication of the nonspecific nature of this inhibition is its reversibility. FIGURE 10 shows the reversal of the glycerol inhibition of the 32P,-ATP exchange reaction. This experiment was done by diluting each incubation mixture tenfold be- fore starting the assay; hence the small decline in activity was possibly due to the small quantity of glycerol remaining. Experiments performed by washing particles with 0.25 M sucrose appeared to support this contention, as shown in TABLE 1. In- hibition of the 3zPi -ATP exchange by the other solutes was also reversible; however, in the case of dimethylformamide, part of the inhibition was not reversed at higher concentrations of solute, even after the particles had been washed. The inhibition with n-propyl alcohol was almost totally irreversible, as previously noted.
)r t > t
.-
.- ' 50- ,\"
I I , \, 10 20 30 40 50
'10 glycerol - v f v FIGURE 10. The reversal of the glycerol inhibition of the 3z4 -ATP exchange reaction in
submitochondrial particles. The conditions for the assay of the ,,Pi -ATP exchange reaction were those described in the legend to FIGURE 5 . o---o =submitochondrial particles mixed for 2 rnin at 0°C with glycerol at the concentrations indicated, and added to an assay medium that contained glycerol at the same concentration; 0--0 = subrnitochondrial particles mixed for 2 min at 0°C with glycerol at the concentrations indicated, and added to an assay medium that contained no glycerol (dilution, 1: 10); o--os=subrnitochondrial particles in- cubated for 10 min at 25" with glycerol at the concentrations indicated, and added'to an assay medium that contained no glycerol (dilution, 1: 10). (From Conover.s Reproduced by per- mission of the Journal of Biological Chemistry.)

34 Annals New York Academy of Sciences
TABLE 1 REVERSAL OF SOLUTE INHIBITION O N THE 32Pi-ATP EXCHANGE REACTION
32Pi Esterified With Solute After Washing* Solute
40%, V/V nmol/mg/min None 142 I28 Glycerol 8 126 Ethylene glycol 9 132 Sucrose I 123 Dimethyl sulfoxide 5 115 Dimethylformamide 3 98 n-Propyl alcohol 0 18
*Submitochondria1 particles, incubated 5 min at 25°C with solute, were diluted with 0.25 M sucrose, which contained 0.01 M MgC12 and 0.001 M ATP, and were centrifuged at 100,000 x g for 40 min. The assay was made as described in the legend to FIGURE 5, on particles resuspended in the sucrose medium.
Relation ofsolute Inhibition to Substrate Concentration
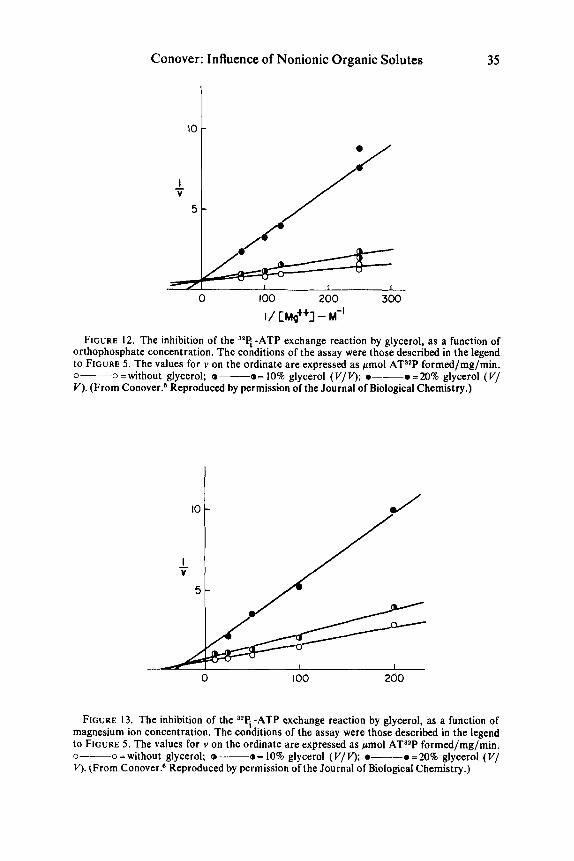
Although the submitochondrial particle is very crude in the sense that it is a complex multienzyme system, an attempt was made to study the effect of glycerol on the binding of ATP, Mg++, and orthophosphate in the 32Pi-ATP exchange reaction. Reciprocal plots of some of the data are shown in FIGURES 11, 12, and 13. Caution should perhaps be exercised in the interpretation of such plots; however, it
0 200 400 600
I /-
FIGURE 11. The inhibition of the 32Pi-ATP exchange reaction by glycerol, as a function of ATP concentration. The conditions of assay were those described in the legend to FIGURE 5 . The values for w on the ordinate are expressed as pmol AT3*P formed/mg/min. 0--0 =Without glycerol; @-PO= 10% glycerol ( V / V ) ; 0---0=20% glycerol ( V / V). (From Conover! Reproduced by permission of the Journal of Biological Chemistry.)
Conover: Influence of Nonionic Organic Solutes 35
0 I00 200 300 I / CM~++J - M-I
FIGURE 12. The inhibition of the 32q-ATP exchange reaction by glycerol, as a function of orthophosphate concentration. The conditions of the assay were those described in the legend to FIGURE 5. The values for v on the ordinate are expressed as pmol AT32P formed/mg/min. o--o=without glycerol; @-----a= 10% glycerol (V/V); .----. =20% glycerol ( V / V). (From Conover.' Reproduced by permission of the Journal of Biological Chemistry.)
J
0 I00 200
FIGURE 13. The inhibition of the '*€? -ATP exchange reaction by glycerol, as a function of magnesium ion concentration. The coiditions of the assay were those described in the legend to FIGURE 5. The values for v on the ordinate are expressed as pmol AT32P formed/mg/min. 0--0 =without glycerol; o-----o= 10% glycerol (V/V); .----.=20% glycerol (V/ V). (From Conover? Reproduced by permission of the Journal of Biological Chemistry.)

36 Annals N e w York Academy of Sciences
would appear that glycerol inhibited uncompetitively with regard to both ATP and orthophosphate. Unexpectedly, the reciprocal plots of activity against Mg++ con- centration appear to indicate that glycerol has a competitive type of relationship with Mg++ (or Mg-ATP, since the metal ion probably exists primarily in this form under these conditions). A similar effect was found with ethylene glycol. Un- fortunately for the purposes of this conference, dimethyl sulfoxide was not tried, but the implication is that it should act similarly.
An attempt to extend this work to submitochondrial ATPase was also made, but interpretation becomes even more hazardous, because of the apparent presence of an inhibitor in the preparations. This was suggested by a nonlinear relation between the activity and protein concentration at higher levels of protein. Pullman and Monroy reported the isolation and purification of a protein inhibitor of the coupling factor- 1 ATPase a ~ t i v i t y . ' ~ Nevertheless, with the ATPase activity, too, reciprocal plots of activity against concentration appeared to indicate that glycerol was not competitive with the substrate ATP, but was competitive with Mg++.
As one possible explanation, it is apparent that the solute may bind free Mg++ and thus diminish its active concentration. Lohmann and colleague^^^ have shown that a number of glycerol-metal complexes exist, including one that contains man- ganese ion. However, no effect on the binding of Mg++ by EDTA or ATP as de- termined by spectral measurement or enzymatic reactivity could be detected in glycerol-water systems.
I t is possible that this relationship of the inhibition and Mg++ concentration may explain the lower sensitivity of oxidative phosphorylation to solute inhibition. If the inhibition in some way primarily involved the Mg-ATP complex as a substrate, those reactions that require the reversal of ATP synthesis and therefore this substrate may be more sensitive than oxidative phosphorylation, which requires the reaction with ADP or Mg-ADP. At the present state of our knowledge, however such considerations must remain conjectural.
In summary, these data would appear to indicate that dimethyl sulfoxide acts on the systems of generation and conservation of mitochondria1 energy in a manner similar, if not identical, to that of a number of other organic solutes studied. These solutes seem to manifest their effects on a common basis, which involves a decrease in water content or, if you wish, they act as a water diluent. The effect may, of course, affect the various reactions in different ways. I t would not appear likely that water is being excluded as a substrate in these systems. More likely perhaps are effects such as decreasing proton mobility, which modifies ionization and binding constants, possible effects on hydrophobic binding between proteins, or as was dis- cussed, an effect on the Mg++ hydration or binding. In any case, these results clearly underline one aspect of the effects that dimethyl sulfoxide may show in biological systems.
References
1 . NELSON, J . M. & M. P. SCHUBERT. 1928. J. Amer. Chem. SOC. 50: 2188. 2. LUMRY, R. & H. EY RING. 1954. J. Phys. Chem. 58: 110. 3. WESTHEAD, E. W. & B. G. MALMSTROM. 1957. J . Biol. Chem. 228: 655. 4 . MITCHELL, P. 1966. Biol. Rev. Cambridge Phil. SOC. 41:445. 5. HANSEN, M. & A. L. SMITH. 1964. Biochim. Biophys. Acta81: 214. 6. CONOVER, T. E. 1969. J . Biol. Chem. 244: 254. 7. INTERNATIONAL CRITICAL TABLES. Vol. 111. 1928. McGraw-Hill Book Company. New
York, N.Y.

8. 9.
10. 11. 12.
13.
14. 15.
Conover: Influence of Nonionic Organic Solutes 31
TYLER, D. D. & R. W. ESTABROOK. 1966. J . Biol.Chern. 241: 1672. CHANCE, 9. & G. R. WILLIAMS. 1956. Advan. Enzyrnol. 17: 65. SLATER, E. C. & K. W. CLELAND. 1953. Biochem. J. 53: 557. LEHNINGER, A. L. 1961. J. Biochern. (Tokyo) 49: 553. CHANCE, B., M. NISHIMURA, S. 9. ROY & H. SCHLEYER. 1963 In Bacterial Photosyn-
thesis. H. Gest, A. San Pietro & L. P. Vernon, Eds.: 357. Antioch Press. Yellow Springs, Ohio.
MARGOLIS, S. A., H. BAUM & G. LENAZ. 1966. Biochern. Biophys. Res. Commun. 25: 133.
PULLMAN, M. E. & G. C. MONROY. 1963. J. Biol. Chem. 238: 3762. LOHMANN, W., C. F. HOWLER, A. J . MOSS, Jr. & W. H. PERKINS. 1965. Experientia 21:
31.