influence of calcium and magnesium containing fixatives of the ultrastructure of parathyroids
TRANSCRIPT

Micron and Microscopica Acta, Vol. 18, No 4, pp. 259-271. 1987. Printed in Great Britain.
0739%6260/87 $3.00+0.00 I&J 1988 Pergamon Journals Ltd.
INFLUENCE OF CALCIUM AND MAGNESIUM CONTAINING FIXATIVES OF THE ULTRASTRUCTURE OF PARATHYROIDS
PETER WILD,* GUISEPPE BERTONI,* ELISABETH M. SCHRANER* and RUDOLF BEGLINGER?
*Institute of Veterinary Anatomy, Laboratory for Electron Microscopy, University of Ziirich, Winterthurerstrasse 260, CH-8057 Ziirich Switzerland and tCiba-Geigy Ltd., CH-4000 Basel, Switzerland
(Received 25 Jury 1987)
Abstract-The effect of Ca2+ and Mg’+ on feline parathyroid cells during perfusion fixation with glutaraldehyde and subsequent immersion in 0~0, was investigated. Both Ca” and Mg” may exert a stabilizing or destabilizing effect on cell membranes and on elements of the cytoskeleton. The effect depends (1) on the ion concentration, (2) on the buffer concentration and (3) on the fixative. Stabilization due to Ca2+ or
Mg ” during glutaraldehyde fixation is not altered during subsequent osmication but both cations may cause destabilization during osmication in tissue prefixed without cations. Ca’+ and Mg*+ also reduce cell volume in combination with low osmolar buffer but they prevent cells from excessive shrinkage due to high osmolar buffers. Ca2+ and Mg ‘+ alone or in combination reduce swelling of RER, extraction ofcellular material and loss of subcellular compartments, such as secretory granules, under optimal conditions. Ca’+, however, provokes formation ofdark (shrunken) and light (swollen) cells accompanied by loss ofsubcellularcomponents when used in low concentration during osmication. Low concentrations of Mg2+ added to glutaraldehyde exert similar effects. Stabilization of membranes is assumed to be due to the binding capacity of Ca2+ and Mgz+ to both phospholipids and proteins. The influence ofCa2 + and Mg’+ to changes in cell volume is considered likely to be the result of ionic interaction in the cytoplasmic gel, the maintenance of cell volume being a matter of equilibrium between the swelling pressure of the cytoplasmic gel and the osmotic pressure of the fixative solution.
Index key words: Cat, parathyroid, fixation, glutaraldehyde, OsO,, Ca, Mg, TEM.
INTRODUCTION
Calcium and-to a lesser extent-magnesium have been used for many years as additives to buffers during fixation in order to prevent extraction of substances occurring during dehyd- ration, to stabilize membranes and to prevent cells from shrinkage (Hayat, 1981; Bullock, 1984). We investigated the influence of Ca2+ and Mg2+ on the ultrastructure of parathyroid (PT) cells during glutaraldehyde and/or osmium fixa- tion. We used this model because PT cells react very sensitively to changes in fixation considering the fixative itself (Wild et al., 1987), buffers, additives and fixation method. Differences in fixation quality resulting from immersion and vascular perfusion, respectively, were reported elsewhere (Wild and Manser, 1985; Wild et al., 1986). In this study, we will show that both Ca2+ and Mg2+ may influence the morphology of PT cells during vascular perfusion with glutaralde-
hyde and/or subsequent immersion fixation with oso,.
MATERIALS AND METHODS
One to 3 year old female cats were anaesthe- tized with Na pentobarbital, 6&70 mg per kg body wt. They were perfused via the left cardiac ventricle with 2.5% glutaraldehyde (GA) in 0.05 M Na cacodylate without or with 6.5% sucrose, pH 7.4, 100 and 300 mOsm, respec- tively, or 0.1 M Na cacodylate, pH 7.4, 200 mOsm, containing various concentrations of CaCl, and MgCl, (Tables l-3) for 15-20 min with a pressure of about 120 cm H,O. After perfusion, PT glands were carefully removed, disected and immersed in the same fixative for 1 h at 4”C, washed in the same buffer for i to 1 h at 4”C, and post-fixed with 1% 0~0, in 0.05,0.075 or 0.1 M Na cacodylate containing various
259

260 Peter Wild, Giuseppe Bertoni, Elisabeth M. Schraner and Rudolf Beglinger
Table 1. Main alterations in parathyroid tissue provoked by Ca*+ (PM) and/or Mg2+ (PM) added to 2.5% glutaraldehyde in 0.05 M Na cacodylate and to 1% 0~0, in 0.05,0.075 or 0.1 M Na cacodylate
Figures Glutaraldehyde oso,
CaCI, MaCI, CaCI, MgCl, Main alterations
2 0
3 250 0 0 0 500 0 0 0
4 0 0 250 0
10 250 0 250
5 0 0 500 500 0 500
6 0 250 0 9 0 250 0
7 0 500 0 0 0 500 0 500
8 0 0 0 250 0 0 0 500
250 250
11 500 12 500
250 250
13 0 0 0 0
250 0 0 250 250 250 250 0 0 250 500 250
500 0 0 500 250 500
0 2.50 250 0 250 500 0 500 250 0 500 500
0 0 0
0
0 0
0 250
Uniformity, intact RER and GC, many SG
Uniformity, good contrast of membranes
Dark cells: shrinkage, vacuolization of RER; Light cells: swelling, loss of membranes
Uniformity, wide intercellular spaces
Uniformity, distinct plasma membranes, narrow intercellular spaces
Dark cells: dense cytoplasmic matrix, vacuolization of RER, loss of GC; light cells: light matrix, loss of membranes
Uniformity, distinct plasma membrane, little RER and GC, narrow intercellular spaces
Dark cells: shrinkage, disrupted mitochondrial cristae; Light cells: swell- ing, disruption and loss of membranes
Uniformity, distinct plasma membranes, narrow intercellular spaces
GC: Golgi complex; SG: secretory granules.
Table 2. Main alterations in parathyroid tissue provoked by Ca2+ (PM) and/or Mg*+ (PM) added to 2.5% glutaraldehyde and/or 1% 0~0, in 0.1 M Na cacodylate
Figures Glutaraldehyde oso,
CaCI, MgCf, CaCl, MgCl, Main alterations
0
250 14 500
250
15 0 0 250 0 500 0 500 0
0 0 500 0
16 0 250 0 250 0 500 0 500
0
250 0
500
17 0 0
0 0
250 Dark-light variation, little RER, many 500 SG
0 0
0 0
250
0 Uniformity: narrow RER, small GC, many SG Distinct GC, vacuolization of RER, disruption of mitochondrial cristae, few SG, shrinkage of some cells Uniformity: dense cytoplasmic matrix, few SG Dark-light variation, some dilation of RER, few SG
Chijbmity: dense cytoplasmic matrix, intact GC, little RER, many SG, distinct plasma membrane

Ca’+ and Mg ‘+ Dependent Changes in Parathyroids 261
Table 3. Effect of Ca2+ (FM) or Mg2+ (FM) on parathyroid tissue during post-fixation with 1% 0~0, in 0.05 M Na cacodylate containing 6.5% sucrose after perfusion fixation with 2.5% glutaraldehyde in ion-
free 0.05 M Na cacodylate containing 6.5% sucrose
Figures Glutaraldehyde oso, CaCI, MgC1, CaCl, MgCl, Main alterations
18 0 0 0 0 19 0 0 250 0
20 0 0 500 0
21 0 0 1000 0
22 0 0 0 250
23 0 0 0 500
24 0 0 0 1000
Shrinkage, slight dark-light variation Uniformity, reduced shrinkage of cytoplasm linijiirmity, reduced shrinkage, large intercellular spaces Light cells: little shrinkage; Dark cells: shrunken, indistinct Enhanced shrinkage, slight dark-light variation Unijormity, little shrinkage, rather large intercellular spaces Light cells: little shrinkage; Dark cells: shrunken, indistinct, large intercellular spaces
concentrations of CaCl, and MgCl, (Tables l-3) for 1 h at 4°C. The blocks were then dehydrated in a graded series of ethanol starting at 70% and, after two changes in propylene oxide, embedded in epon. Semithin sections were stained with toluidine blue and ultrathin sections with Mg many1 acetate and lead citrate.
In order to get an idea of possible changes in cell volume and subcellular compartments due to Ca2+ and Mg2+ we estimated the mean nuclear volume, the mean cell volume and the distribu- tion of secretory granules of PT tissue fixed with glutaraldehyde in 0.05 M Na cacodylate fol-
lowed by 0~0, in 0.075 M Na cacodylate (Fig. 1). On 10 randomly taken photographs per sample (final magnification approximately x 6700) the volume density of the nuclear and cytoplasmic compartment was estimated by point counting (Weibel, 1979) using a projection unit and a HP 981% table calculator. The numerical density of nuclei was calculated apply- ing the formula from Weibel and Gomez (1962). The mean nuclear and the mean cell volume were calculated from these data (Wild and Manser, 1985). The number of secretory granules were counted on 10 randomly taken photographs per
800
600
400
200
cz 010 2510 0109 510 510 0125s OK) 015 010 0125’) O/g 2515 2Y5 C&J, 010 010 25x) 010 510 o/o m.5 010 015 012.5 010 2.545 2.515
GA oso4
Fig. 1. Estimation of nuclear volume (unshaded) and cell volume (shaded) expressed in um3, and of the number of secretory granules (*) expressed per unit area (2000 urn’) cytoplasmic compartment of PT tissue fixed with 2.5% GA in 0.05 M Na cacodylate and 1% 0~0, in 0.075 M Na cacodylate containing different concentrations of Ca2+ and Mg’+. (a) Concentrations which led to extreme dark-light variations and poor preservation of the
ultrastructure (see Figs. 4, 6 and 9).

262 Peter Wild, Giuseppe Bertoni, Elisabeth M. Schraner and Rudolf Beglinger
sample at higher magnification (final magnifica- tion approximately x 15,550) and expressed per
because fixation with a given ion concentratic
unit area cytoplasm. was not repeated so that the data lack statistic, analysis.
RESULTS
The influence of Ca2 + and Mg2 + on the ultrastructure of PT cells during both aldehyde and osmium fixation are explicitly shown in Tables l-3 and Figs. l-24.
Eflect during glutaraldehyde jixation
Ca2+ led to cell shrinkage, dilation of RER and disruption of membranes in combination with 0.1 M Na cacodylate (Table 2) whereas MgZf provoked dark cells with shrunken cyto- plasm and light cells with little subcellular components only at low concentration in combi- nation with 0.05 M Na cacodylate (Table 1).
The shrinkage effect of hypertonic buffe (Figs. 18-24; Table 3) was partly compensated t 500 or 1000 uM CaCl, and-to a great extent-by 500 uM MgCl,. Both 1000 uLF CaCl, and MgCl, gave rise to enlarged interce lular spaces and to some shrunken indistin’ cells. Enlargement of intercellular spaces ovc large areas were observed also in PT tissue fixe with GA and 0~0, in 0.1 M buffer. Th enlargement was only diminished by simu taneous use of 250 uM CaCl, and 500 uE MgCl, during both GA and 0~0, fixation (nc shown).
Influence on membranes
Effect during 0~0, jixation
250 uM CaCl, in 0~0, prepared in 0.05 M Na cacodylate (Table 1) provoked similar cell vari- ants as 250 uM MgCl, did during GA fixation. Increasing the concentration of CaCl, to 500 uM or addition of at least 250 uM CaCl, to GA prior to osmication or simultaneous use of at least 250 uM MgCl, prevented cells from disturbance of membrane integrity. 0.1 M buffer (Table 2) also reduced the effect of Ca2 +.
Ca2 ’ provoked loss of intracellular men branes, i.e. membranes of RER, Golgi complc and secretory granules, when introduced durir oxmium fixation at low concentration in tl presence of 0.05 M Na cacodylate. At high, concentration, it enhanced contrast mainly of tl plasma membrane.
Mg 2 + had no obvious effect in 0~0, buffered with 0.05, 0.075 or 0.1 M Na cacodylate after perfusion with GA in 0.05 M Na cacodylate (Table 1). However, the undesirable effect which became manifest after introducing 250 uM MgCl, to GA in 0.05 M buffer could not be avoided by addition of MgCl, or CaCl, to the 0~0, fixatives. In contrast, Mg2+ in combina- tion with 0.1 M Na cacodylate (after perfusion with GA in the same buffer) led to dark and light variants which did not arise when Mg2+ was also used during fixation with GA in 0.1 M Na cacodylate prior to osmication (Table 2).
Mg2+ gave rise to a similar loss of membran’ including inner membranes of mitochondr when added to GA at low concentration and enhancement of membrane contrast at high’ concentrations. In order to get an idea about lo of subcellular compartments we counted tl number of secretory granules and expressed the per unit area cytoplasm (2000 urn’). Figure shows that 250 uM CaCl, induced loss secretory granules during osmium fixatic whereas 500 uM CaCl, led to some loss durir GA fixation. MgCl, exerted a similar effe during GA fixation. Combination of Ca2+ ar Mg2+ prevented loss of secretory granules du ing osmication but not during GA fixation.
DISCUSSION
Effect on cell volume
The stereologic data (Fig. 1) obtained from PT tissue perfused in the presence of 0.05 M buffers and post-fixed in the presence of 0.075 M buffers show that both the nuclear and cytoplasmic compartment were reduced by Ca2+ and/or Mg2+-with two exceptions-during GA or 0~0, fixation. These data have to be interpreted cautiously (1) because of the low number (about 200) of nuclei per sample considered and (2)
Parathyroid glands are believed to be car posed of dark and light chief cells, multinucleatc syncytial cells, atrophic cells, and oxyphil ccl (e.g. Altenahr, 1972; Roth and Capen, 197 Thiele, 1977; Capen, 1980; Meuten et al., 198~ Recently, comparative studies of PT glands fixc by perfusion or immersion revealed that da: and light chief cells, atrophic cells and multinu leated syncytial cells of various mammals ari during immersion fixation (Setoguti 1977; Lar son et al., 1984; Moreira and Goncalves, 198 Wild and Manser, 1985; Wild et al., 198

Ca’+ and Mg 2f Dependent Changes in Parathyroids 263
Figs. 2-13. PT cells fixed with 2.5% GA in 0.05 M Na cacodylate and 1% 0~0, in 0.05,0.075 or 0.1 M Na cacodylate containing different concentrations of Ca2+ and Mg2+. Since the buffer concentration does not influence obviously the alterations provoked by Cal+ and Mg’+ during osmication, it is not indicated. All
figures x 7200.
Fig. 2. No Ca2’ and Mg2+ at all led to uniform appearance of PT cells.
Fig. 3. Addition of 250 uM CaCl, (or 500 FM, not shown) to GA only increased contrast of membranes.
Fig. 4.250 uM CaCI, used only during osmication provoked cell shrinkage, dilation of RER and disruption of membranes giving rise to PT cell variants.
Fig. 5. Addition of 500 uM CaCI, only to osmium, results in uniformity, the cells showing distinct plasma membranes but small GC.
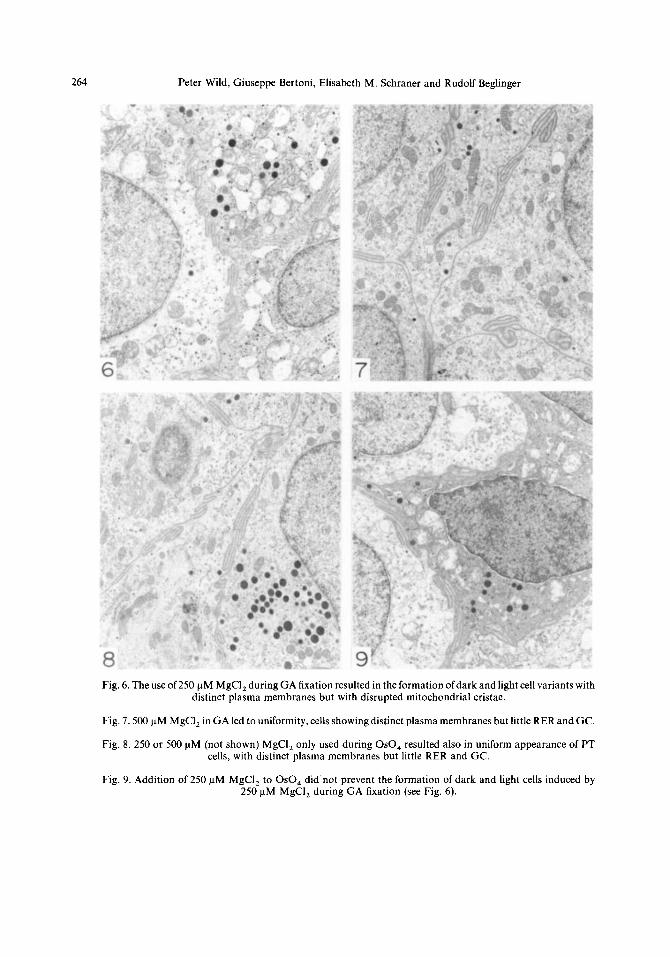
264 Peter Wild, Giuseppe Bertoni, Elisabeth M. Schraner and Rudolf Beglinger
Fig. 6. The use of 250 nM MgCI, during GA fixation resulted in the formation of dark and light cell variants with distinct plasma membranes but with disrupted mitochondrial cristae.
Fig. 7.500 uM MgCI, in GA led to uniformity, cells showing distinct plasma membranes but little RER and GC.
Fig. 8. 250 or 500 uM (not shown) MgCI, only used during 0~0, resulted also in uniform appearance of PT cells, with distinct plasma membranes but little RER and GC.
Fig. 9. Addition of 250 uM MgCI, to 0~0, did not prevent the formation of dark and light cells induced by 250 pM MgCI, during GA fixation (see Fig. 6).

Ca 2 + and Mg 2÷ Dependent Changes in Parathyroids 265
Fig. 10.250 ~M CaCI 2 in GA prevented the formation of cell variants induced by 250 l-tM CaCI 2 added to OsO 4 (see Fig. 4).
Fig. 11. 250 laM (not shown) or 500 pM CaC12 added to GA did not prevent the formation of shrunk dark cells and light cells which were provoked by 250 p,M MgCI 2 during GA fixation (see Fig. 6).
Fig, 12. Addition of 250 laM (not shown) or 500 ~tM CaC12 to both GA and OsO 4 also did not prevent the . formation of dark and light cells induced by 250 p.M MgCI 2 during GA fixation (see Fig. 6).
Fig, 13. 250 pM MgCI 2 or 500 I, tM MgCI 2 to OsO4entirely blocked the formation of dark and light cell variants provoked by 250 I-tM CaCI 2 during osmium fixation (see Fig. 4).

266 Peter Wild, Giuseppe Bertoni, Elisabeth M. Schraner and Rudolf Beglinger
Figs. 1417. PT cells fixed with 2.5% GA in 0.1 M Na cacodylate containing various concentrations of CaCl, and MgCI, and post-fixed with 1% 0~0, in the same buffer. All figures x 7200.
Fig. 14. In contrast to 0.05 M buffer (see Fig. 3), 250 PM (not shown) or 500 uM CaCI, in GA caused cell shrinkage and dilation of RER in some cells.
Fig. 15. 250 uM CaCl, or 500 uM (not shown) in 0~0, led to minor dark-light variation.
Fig. 16. 250 uM MgCI, added to GA in 0.1 M Na cacodylate resulted in uniformity (compare the effect of 250 uM MgCl, added to GA to 0.05 M Na cacodylate, Fig. 6).
Fig. 17.250 uM CaCl, added to OsO,-and to a lesser extent 500 uM MgCI, (not showntprovoked dark and light cell variants with little disruption of membranes (compare Fig. 8).

Ca’+ and Mg’+ Dependent Changes in Parathyroids 267
proving the idea of Stoeckel and Porte (1966) that variations in PT cell morphology rather depend on fixation than on functional diversity. On the other hand, Roth and Raisz (1966) found that incubation of rat PT glands in vitro in the presence of different concentrations of Ca’+, which largely control together with Mg*+ PT secretory activity through a unique negative feedback mechanism, led to changes in cell morphology: high calcium concentration resulted in an increased number of light cells and low calcium concentration in an increased number of dark cells. These findings gave rise to the idea that dark cells represent active forms and light cells resting forms of a secretory cycle because the changes were reversible. Alteration in serum calcium concentration in intact rats with subsequent perfusion fixation, however, yielded a high uniformity in PT cell morphology, irrespective of the state of stimulation or suppres- sion of secretory activity (Wild et al., 1982, 1984, 1985). We routinely used 2.5% GA in 0.1 M Na cacodylate containing 250 uM CaCl, and 500 uM MgCl, for perfusion and 1% 0~0, in 0.1 M s-collidine containing 5% sucrose, 500 uM CaCl, and 1000 uM MgCl, for post- fixation.
The effect of divalent cations during fixation is not well understood. Possible interactions between Ca*+ and subcellular components are discussed in detail by Hayat (1981) stating that Ca*+ bind to polypeptides as well as phospho- lipids. The binding capacity of Ca* + and Mg* + to phospholipids has been shown by Hauser and Dawson (1967). CaCl, and MgCl, is utilized to precipitate isolated plasma membranes, MgCl, having the advantage of preventing hydrolysis of membrane phospholipids by calcium-activated phospholipases (Hauser et al., 1980).
Stabilization is probably achieved by binding of cations to membranes, e.g. of intestinal epithelial cells (Oschman and Wall, 1972), neural cells (Palay et al., 1962; Roozemond, 1969; Oschman et al., 1974), spermatozoa (Roomans, 1975) and of algae (Lin and Sommerfeld, 1978). Both Ca*+ and Mg2+ may have a stabilizing or destabilizing effect on filaments and micro- tubules which depends on the ion concentration (Harris, 1962; Shigenaka et al., 1974; Schliwa, 1977; Aoki and Tavassoli, 1981).
The effect of Ca*+ and Mg2+ is clearly demonstrated in PT tissue. 250 uM MgCl, added to 0.05 M Na cacodylate for buffering GA was responsible for destabilization of membranes
resulting in swelling of RER combined with cellular shrinkage or loss of membranes and cytoplasmic ground substance ending in the formation of extreme dark-light variations. This destabilizing effect was omitted either by increas- ing the Mg*+ concentration or the buffer concen- tration. 250 or 500 uM CaCl,, on the other hand, stabilized membranes during fixation with GA buffered with 0.05 M Na cacodylate whereas CaCl, in combination with 0.1 M Na cacodylate led to destabilization, which was more severe when 250 uM CaCl, was used. These results thus demonstrate (1) that the effect of Mg*+ and Ca*+ depends on the ion concentration and on the buffer concentration, the latter being more important for Ca* + . (2) The effect of Ca*+ and Mg* + on PT cells is similar considering mem- brane stability but it is inversely related to the buffer concentration. The buffer concentration used for prefixation with GA largely influenced the action of Ca*+ and Mg2+ during post- fixation with 0~0, whereas the buffer concentra- tion used during osmication had little effect. Mg*+ stabilized cellular components during osmication when 0.05 M Na cacodylate was used to buffer GA. 250 uM CaCl, destabilized and 500 uM CaCl, stabilized membranes when used under the same condition. If the buffer concen- tration for GA was increased to 0.1 M, both Ca*+ and Mg*+ caused mild alterations consi- dering disruption of membranes and loss of secretory granules and cytoplasmic matrix mimicking dark and light PT’cells or PT cells in active and resting stages. Thus one may argue that different Ca*+ concentrations used for stimulation or suppression PT secretory activity in oitro (Roth and Raisz, 1966) might have been responsible for the formation of the varying number of light and dark cells during subsequent fixation because tiny rests of Ca*+ might have been sufficient to introduce such changes. On the other hand, we were not able to prevent the formation of dark and light variants during immersion fixation unless we introduced an aldehyde mixture consisting of GA, formalde- hyde and acrolein (Wild et al., 1987). It is not clear why dark and light cells arise during fixation. The possibility that the changes reflect different functional stages can be excluded because cells originating from the same gland behaved dissimilarly in different fixative solu- tions.
The use of Ca* + or Mg* + during GA and 0~0, fixation showed that stabilization achieved

268 Peter Wild, Giuseppe Bertoni, Elisabeth M. Schraner and Rudolf Beglinger
through one of these cations during GA fixation was not destroyed during osmication indicating that the action of both Ca2+ and Mg2+ is of first order during GA fixation in the context of double fixation. Nevertheless, volume changes may be caused during osmication as shown for Ca2+ and Mg2+ added to buffer with high osmolarity. Both, Ca2+ and Mg2+ suppressed cell shrinkage whereas they slightly reduced cell volume or prevented cells from swelling when they are added during GA and/or 0~0, fixation in combination with 0.05 M buffer, with the excep- tion of Mg2 + which seem to cause swelling during GA fixation. Again, the effect depends on the ion and buffer concentration which may explain the discrepancy to the results of Tooze (1964) who reported shrinkage of erythrocytes due to Ca2+ added in form of 10 mM CaCl, whereas 45 mM CaCl, prevented erythrocytes from swelling (Millonig and Marinozzi, 1968). The effect on cell volume in combination with 0.1 M buffer was not investigated. It is note- worthy, however, that the enlargement of inter- cellular spaces was not influenced by Ca2+ or Mg2+ but by the combination of the two (Wild et al., 1986).
It has to be borne in mind that the effect of cations added to fixatives on cells may be enhanced during subsequent dehydration (Boyde and Maconnachie, 1980). The question arises how and where Ca2 + and Mg2 + act in affecting cell volume during fixation and sub- sequent dehydration. GA makes cell membrane permeable (Harper, 1986) so that many compo- nents including sugars, proteins, lipids and ions
may be extracted during GA fixation (Coetzee and van der Merwe, 1984) and osmication (Luft and Wood, 1963; Pentilla et al., 1974). Since Harper (1986) showed that La3+ enter cells one may assume that other ions such as Ca2+ and Mg* + may do the same. Ca2+, therefore, enter- ing the cell may activate elements of the cyto- skeleton (review by Bennett and Weeds, 1986). Whether Ca2+ exerts such an effect under fixation conditions is unknown. However, the cytoplasm is considered to be a living gel (Porter, 1984). The gelled state of the cytoplasm seems likely to play a major role in the maintenance of cell volume in living cells (Lechene, 1985). Hence, it is reasonable to assume that the swelling equilibrium of the cytoplasmic gel contributes much to the maintenance of cell volume under fixation conditions. The volume of living cells thus will be maintained in the course of fixation and subsequent treatment if the osmotic pressure of the fixative equals the swelling pressure of the cytoplasmic gel at its physiological swelling equilibrium. Of course, the swelling pressure is influenced by the fixative itself, such as GA because of its cross-linking ability, and by ions which may be extracted or which possibly enter the cytoplasmic gel when added to the fixative. If the swelling equilibrium of the cytoplasmic gel is responsible for maintaining cell volume during GA fixation, it also may influence volume changes during osmication and subsequent dehydration. Consequently, volume changes occuring during osmication and dehydration can be influenced by addition of ions, such as Ca2+ and Mg2 + to 0~0, and to the dehydrating agent
Legend for page 269
Figs. 18-24. PT cells of the same animal fixed with 2.5% GA in 0.05 M Na cacodylate containing 6.5% sucrose and post-fixed with 1% 0~0, in the same buffer containing various concentrations of CaCl, and MgCI,. All
figures x 5040.
Fig. 18. Marked shrinkage after osmication containing no ions.
Fig. 19. 250 uM CaCl, slightly reduced cytoplasmic shrinkage.
Fig. 20. Reduced shrinkage and enlarged intracellular spaces by 500 uM CaCI,.
Fig. 21, 1000 uM MgCl, led to the formation of indistinct dark cells.
Fig. 22. Enhanced shrinkage by 250 pM MgCl,.
Fig. 23. Little shrinkage after addition of 500 uM MgCI,.
Fig. 24. Large intercellular spaces and indistinct dark cells caused by 1000 urn MgCI,.

Ca2+ and Mg2+ Dependent Changes in Parathyroids 269

270 Peter Wild, Giuseppe Bertoni, Elisabeth M. Schraner and Rudolf Beglinger
because ions influence directly the swelling pressure ofa gel (Katchalsky and Michael, 1955). As we know, addition of l-3 mM CaCl, to OsO,, e.g., is recommended to prevent cells from swelling (Hayat, 1981).
In conclusion, stabilization of cell membranes in the course of perfusion fixation with GA may be enhanced by Ca2 + and Mg2 + . Both cations may bind to membranes and to elements of the cytoskeleton. Stabilization depends on the ion concentration and on the buffer concentration. Simultaneous use of Ca2 + and Mg2 + is recom- mended, adding Mg2 + in a concentration twice as high as Ca 2+ However, loss of membranes . (and other components) occuring during fixation or subsequent treatment is not entirely pre- vented. The mechanism of destabilization by low concentrations of Ca2+ and Mg2+ is unknown. Both Ca2+ and Mg2+ contribute also to mem- brane stabilization during post-fixation with 0~0,. Ca2+ and Mg2+ are suggested to control maintenance of cell volume by affecting the gelled state of the cytoplasm resulting in changes of the swelling pressure which, in turn, is in equilibrium with the osmotic pressure of the fixative solution. This equilibrium will maintain the original cell volume under optimal conditions.
Acknowledgements-The authors wish to thank Prof. M. A. Hayat for encouragment to do this study, Prof. J. Frewein for careful reading of the manuscript, Prof. M. C. Schaub for helpful discussions and Mrs. M. Balushev for assistance with the manuscript.
REFERENCES Altenlhr, E., 1972. Ultrastructural pathology of parathyroid
glands. Curr. Top. Pathol., 56: l-54. Aoki, M. and Tavassoli, M., 1981. OTO method for
preservation of actin filaments in electron microscopy. J. Histochem. Cytochem., 29: 682683.
Bennett, J. and Weeds, A., 1986. Calcium and the cytoskele- ton. Med. Bull., 42: 3855390.
Boyde, A. and Maconnachie, E., 1980. Treatment with lithium salts reduces ethanol dehydration shrinkage of glutaraldehyde fixed tissue. Histochemistry, 66: 181-187.
Bullock, G. R., 1984. The current status of fixation for electron microscopy: a review. J. Microsc., 133: l-15.
Capen, C. C., 1980. Parathyroid hormone, calcitonin, and cholecalciferol: the calcium regulating hormones. In: Veterinary Edocrinology and Reproduction, McDonald, L. E. (ed.), Lea and Febiger, Philadelphia, 3rd Ed., pp. 6@130.
Coetzee, J. and van der Merve, C. F., 1984. Extraction of substances during glutaraldehyde fixation of plant cells. J. Microsc., 135: 1477158.
Moreira, J. E. and Goncalves, R. P., 1985. Ultrastructural changings of the rat parathyroid gland under various fixation methods. Anat. Anz., 158: 413423.
Oschman, J. L. and Wall, B. J., 1972. Calcium binding tc intestinal membranes. J. Cell Biol., 55: 58873.
Oschman, J. L., Hall, T. A., Peters, P. D. and Wall, B. J. 1974. Association of calcium with membranes of squic giant axon. J. Cell Biol., 61: 156165.
Palay, S. L., McGee-Russell, S. M., Gordon S. Jr. and Grill0 M. A., 1962. Fixation of neural tissues for electror microscopy by perfusion with solutions of osmium tetrox. ide. J. Cell Biol., 12: 385410.
Penttila, A., Kalimo, H. and Trump, B. F., 1974. Influence o glutaraldehyde and/or osmium tetroxide on cell volume ion content, mechanical stability, and membrane per meabilitv of Ehrlich ascites tumor cells. J. Cell Biol.. 63 197-214.
Porter, K. R., 1984. The cytomatrix: a short history of it: studv. J. Cell Biol.. 99: 3~~12s.
Room&, G. M., 1975. Calcium binding to the acrosoma membrane of human spermatozoa. Expl Cell Res., 96 23330.
Roozemond, R. C., 1969. The effect of calcium chloride ant formaldehyde on the release and composition of phospho, lipids from cryostat sections of rat hypothalamus. J Histochem. Cytochem., 17: 273-279.
Roth, S. 1. and Capen, C. C., 1974. Ultrastructural ant functional correlations of the parathyroid gland. Int. Rev exp. Pathol., 13: 16lL221.
Harper, I. S., 1986. Glutaraldehyde-induced permeabiliza- Roth, S. I. and Raisz, L. G., 1966. The course and reversibilit! tion of cell membranes. J. Microsc., 141: RP33RP4. of the calcium effect on the ultrastructure of the ra
Harris, P., 1962. Some structural and functional aspects 01 the mitotic apparatus in sea urchin embryos. J. Cell Biol., 14: 475-487.
Hauser, H. and Dawson, R. M. C., 1967. The binding of calcium at lipid-water interfaces. Eur. J. Biochem., 1: 61-69.
Hauser, H., Howell, K., Dawson, R. M. C. and Bowyer, D. E., 1980. Rabbit small intestinal brush border membrane preparation and lipid composition. Biochim. biophys. Acta, 602: 567-577.
Hayat, M. A., 1981. Fixation for Electron Microscopy. Academic Press, New York.
Katchalsky, A. and Michael, L., 1955. Polyelectrolyte gels in salt solutions. J. Polymer Sci., 15: 69986.
Larsson, H. O., Lorentzon, R. and Boquist, L., 1984. Structure of the parathyroid glands, as revealed by different methods of fixation. A quantitative light- and electron-microscopic study in untreated Mongolian ger- bils. Cell Tissue Res., 235: 51-58.
Lechene, C., 1985. Cellular volume and cytoplasmic gel. Biol. Cell, 55: 1777180.
Lin, H.-P. and Sommerfeld, M. R., 1978. Stabilization 01 cytomembranes in red algae during ultrastructural fixation with calcium chloride. Trans. Am. Microsc. Sot., 97: 49-100.
Luft, J. H. and Wood, R. L., 1963. The extraction of tissue protein during and after fixation with osmium tetroxide in various buffer systems. J. Cell Biol., 19: 46A.
Meuten, D. J., Capen, C. C., Thompson, K. G. and Segre, G. V., 1984. Syncytial cells in canine parathyroid glands. Vet. Pathol., 21: 463468.
Millonig, G. and Marinozzi, V., 1968. Fixation and embed- ding in electron microscopy. Adv. Opt. Electron. Microsc.. 2: 251-341.

Ca*+ and Mg 2+ Dependent Changes in Parathyroids 271
parathyroid gland in organ culture. Lab. Invest., 15: 1187-1211.
Schhwa, M., 1977. Influence of calcium on intermicrotubule bridges within heliozoan axonemes. J. submicrosc. Cytol., 9: 221-227.
Setoguti, T., 1977. Electron microscopic studies of the parathyroid gland of senile dogs. Am. J. hat., 148: 65584.
Shigenaka, Y., Watanabe, K. and Kaneda, M., 1974. Degrading and stabilizing effects of Mg’+ ions on microtobule-containing axopodia. Expl Cell Res., 85: 391-398.
Stoeckel, M. E. and Porte, A., 1966. Observations ultrastruc- turales sur la parathyroid des souris. I. Etude chez la souris normale. Z. Zellforsch. Mikrosk. Anat., 73: 488-502.
Thiele, J., 1977. Human parathyroid gland: a freeze fracture and thin section study. Curr. Top. Pathol., 65: 31-80.
Tooze, J., 1964. Measurements of some cellular changes during the fixation of amphibian erythrocytes with osmium tetroxide solutions. J. Cell Biol., 22: 551-563.
Weibel, E. R., 1979. Stereological Methods. Vol. 1, Practical Methods for Biological Morphometry. Academic Press, New York.
Weibel, E. R. and Gomez, D. M., 1962. A principle for
counting tissue structures on random sections. J. appl. Physiol., 17: 343-348.
Wild, P. and Manser, E. M., 1985. Ultrastructural morpho- metry of parathyroid cells in rats of different ages. Cell Tissue Res., 240: 585-591.
Wild, P., Bitterli, D. and Becker, M., 1982. Quantitative changes of membranes in rat parathyroid cells related to variations of serum calcium. Lab. Invest., 47: 37@374.
Wild, P., Schraner, E. M. and Eggenberger, E., 1984. Quantitative aspects of membrane shifts in rat parathyroid cells initiated by decrease in serum calcium. Bio[. Cell, 50: 263-272.
Wild, P., Gloor, S. and Vetsch, E., 1985. Quantitative aspects of membrane behaviour in rat parathyroid cells after depression or elevation of serum calcium. Lab. Invest., 52: 49&496.
Wild, P., Schraner, E. M., Augsburger, H., Beglinger, R. and Pfister, R., 1986. Ultrastructural alterations in mammalian parathyroid glands induced by fixation. Acta anat., 126: 87-96.
Wild, P., Kellner, S. J. and Schraner, E. M. 1987. Parathyroid cell variants may be provoked during immersion fixation. Histochemistry, 87: 263-271.