induction of interferon regulatory factor 1 expression in human dermal endothelial cells by...
TRANSCRIPT

ORIGINAL ARTICLESee related Commentary on page xiii
Induction of Interferon Regulatory Factor 1 Expression in HumanDermal Endothelial Cells by Interferon-c and Tumor NecrosisFactor-a Is Transcriptionally Regulated and Requires Iron
Amy K. Gira,� Katherine A. Casper,� Kristen B. Otto,� Shubhada M. Naik,� S.Wright Caughman,�w andRobert A. Swerlick�w�Department of Dermatology, Emory University School of Medicine, Atlanta, Georgia, USA; wDepartment of Veteran A¡airs Medical Center, Atlanta,Georgia, USA
Interferon regulatory factor-1 is a transcription factorthat is linked to the expression of genes important inthe initiation of the in£ammatory response and the con-trol of cell cycle. In this study, we determined that thegeneration of interferon regulatory factor-1 expressionin human dermal microvascular endothelial cells wastranscriptionally mediated by tumor necrosis factor-aor interferon-c via iron-dependent pathways. The induc-tion of interferon regulatory factor-1 protein and the up-regulation of interferon regulatory factor-1 mRNAlevels was inhibited when cells were pretreated with theiron chelators 2-2-dipyridyl or deferoxamine.This inhi-bition of interferon regulatory factor-1 expression wasassociated with loss of interferon regulatory factor-1binding to the interferon-stimulated response elementas assessed by electrophoretic mobility shift assay. Addi-tion of exogenous iron with the iron chelator resulted in
reconstitution of cytokine responsiveness, thus demon-strating iron as the target for the chelator e¡ect. Bothtumor necrosis factor-a and interferon-c-induced inter-feron regulatory factor-1 gene transcription, as assessedby the measurement of unspliced, nascent, heteroge-neous nuclear RNA, and treatment with iron chelatorsblocked tumor necrosis factor-a or interferon-c media-ted interferon regulatory factor-1 gene transcription.Iron was not essential, however, for the association ofinterferon regulatory factor-1 mRNA with polyribo-somes, suggesting iron was not essential for interferonregulatory factor-1 protein translation. Through suchinhibitory regulation on pro-in£ammatory transcrip-tion factors, iron chelators may serve as anti-in£amma-tory agents. Key words: transcription factor/cytokine. J InvestDermatol 121:1191 ^1196, 2003
Interferon regulatory factor (IRF)-1, which was initiallyidenti¢ed as a regulator of interferon (IFN)-b gene expres-sion, belongs to a nine-member family of transcription fac-tors characterized by a highly conserved amino-terminalDNA binding domain (Sato et al, 2001; Taniguchi et al,
2001). IRF-1 acts as a transcriptional activator and has been linkedto the expression of a number of pro-in£ammatory cytokinesand cell cycle regulatory genes (Miyamoto et al, 1988; Sanceauet al, 1995; Lechleitner et al, 1998; Sato et al, 2001). Speci¢cally,IRF-1 induces gene expression by binding to interferon-stimu-lated response elements (ISRE) and regulates the transcriptionof pro-in£ammatory genes, including vascular cell adhesion mo-lecule (VCAM)-1, inducible nitric oxide synthase (iNOS), inter-leukin-12, interleukin-15, cyclooxygenase II, angiotensin II type 2receptor, and interleukin 1b converting enzyme (Neish et al, 1995;Sanceau et al, 1995). IRF-1 is involved in the di¡erentiation of
CD8þ T cells, T helper 1 subsets of CD4þ T cells, and naturalkiller cells (Mamane et al, 1999). It has also been implicated as atumor suppressor gene due to its role in promoting apoptosis,and loss of IRF-1, which has been mapped to 5q31.1, has beenlinked to the development of human hematopoietic malignancies(Miyamoto et al, 1988; Harada et al, 1989; Tanaka and Taniguchi,2000).In human umbilical vein endothelial cells, low levels of IRF-1
protein may be present in unstimulated cells, but markedly in-creased expression results within 4 h after treatment with eithertumor necrosis factor (TNF)-a or IFN-g (Lechleitner et al, 1998;Ochi et al, 2002). IRF-1 has also been shown to be induced byviral infection, leukocyte inhibitory factor, and lipopolysacchar-ide (Sims et al, 1993; Harada et al, 1994). Evidence exists for thepresence of both TNF-a-induced nuclear factor (NF)-kB activa-tion and IFN-g-induced Jak/STAT signaling pathways in the reg-ulation of IRF-1 expression. Limited studies have been done inendothelial cells, particularly microvascular endothelium andcell-speci¢c di¡erences in cytokine-induced IRF-1 expressionmay exist (Lechleitner et al, 1998; Liu et al, 2001).Iron, which is present intracellularly in its ferrous form (Fe2þ ),
is a necessary element for cell growth and proliferation (Conrad,1998); however, excess ferrous iron is capable of acting as a catalystin the Fenton reaction to generate reactive oxygen species andto increase lipid peroxidation, both of which can induce tissuedamage and promote tissue in£ammation (McCord, 1998).In£ammatory skin diseases in which excess iron has been found
Address correspondence and reprint requests to: Robert A. Swerlick,MD, Department of Dermatology,WMB 5311, Emory University Schoolof Medicine, 1639 Pierce Drive, Atlanta, Georgia 30322, USA. Email:[email protected]: DFO, deferoxamine; DP, 2;2-dipyridyl; HDMEC, hu-
man dermal microvascular endothelial cells; iNOS, inducible nitric oxidesynthase; IRF-1, interferon regulatory factor 1; hnRNA, heterogeneousnuclear RNA.
Manuscript received December 17, 2002; revised April 1, 2003; acceptedfor publication May 14, 2003
0022-202X/03/$15.00 . Copyright r 2003 by The Society for Investigative Dermatology, Inc.
1191

include psoriasis (Molin and Wester, 1973), venous ulceration(Ackerman et al, 1988), atopic dermatitis (David et al, 1990), andultraviolet-irradiated skin (Bissett et al, 1991, 1994).There is a growing list of iron-regulated genes, including
iNOS, protein kinase C-b, acid phosphatase type 5 (Acp5)/tar-trate resistant acid phosphatase, hypoxia inducible factor 1a, andp21WAF1/CIP1 (Boldt, 1999; Alcantara et al, 2001). The mechanismsthrough which iron regulates gene transcription are multiple.Wehave previously investigated the role of iron in TNF-a-inducedVCAM-1 gene expression in human dermal microvascular en-dothelial cells (HDMEC) (Koo et al, 2003), and found TNF-amediated induction of VCAM-1 to be iron dependent. Further-more, we observed that a portion of this iron dependence ismediated via iron-dependent induction of IRF-1.In this study, we examined the role of iron in the regulation of
IRF-1 expression in HDMEC.We observed that bothTNF-a andIFN-g induction of IRF-1 required iron for transcriptional acti-vation of the IRF-1 gene and that loss of IRF-1 expression wasaccompanied by loss of IRF-1 binding to the ISRE on a targetpromoter.
MATERIALS AND METHODS
Cell culture Cultures of HDMEC immortalized with SV40 large T(5A32 cells) were utilized and passaged as described previously (Ades et al,1992). Cells were grown in £asks coated with 0.1% gelatin in MCDB 131media (Mediatech, Herndon, Virginia) supplemented with 10% (v/v) fetalbovine serum (Hyclone, Logan, Utah), 1% penicillin/streptomycin(Mediatech), 2 mM L-glutamine (Mediatech), 0.25 mg cyclic adenosinemonophosphate per mL, and 1 mg hydrocortisone per mL (Sigma, StLouis, Missouri) and maintained at 371C in a humidi¢ed atmosphere of5% CO2.
Reagents and antibodies The speci¢c iron chelators 2-2-dipyridyl(DP) and deferoxamine (DFO) (Syvertsen and McKinley-McKee, 1984;Horky et al, 1998; Liu and Hider, 2002), which, respectively, bind ferrousiron (Fe2þ ) and ferric iron (Fe3þ), were obtained from Sigma. Recom-binant human TNF-a and IFN-g were purchased from R&D Systems,Inc. (Minneapolis, Minnesota). Anti-IRF-1 antibody was obtained fromSanta Cruz Biotechnology Inc. (Santa Cruz, California).
Measurement of IRF-1 mRNA and heterogeneous nuclear IRF-1RNA (hnRNA) expression 5A32 cells were either pretreated with DPor DFO for 24 h followed by treatment with IFN-g or TNF-a. mRNAexpression of IRF-1 was measured by real-time quantitative polymerasechain reaction (PCR). Brie£y, total RNA was isolated with TriReagent(Sigma) from 5A32 HDMEC according to the manufacturer’s instruc-tions. cDNA was prepared using the Superscript Preampli¢cation Systemfor 1st Strand cDNA Synthesis (Gibco, Carlsbad, California) using therandom primer protocol. Real-time PCR was done using SYBR greentechnology with Perkin-Elmer 5700 Gene-Amp Detection System withIRF-1 primers (forward sequence CTT TCG CTG TGC CAT GAA CTCand reverse sequence AGC TGC CCT TGT TCC TGC T). Data werenormalized with primers for GAPDH housekeeping gene. Relativequantities were determined by generating a standard curve from dilutionsof cDNA containing the message of interest. Data were then expressed asfold increase over control.IRF-1 hnRNA expression was measured to assess the e¡ect of iron
chelation on the transcription of the IRF-1 gene using a modi¢cation ofthe above technique (Elferink and Reiners, 1996). Prior to reversetranscription, total RNAwas treated with RNAse-free DNAse for 1 h atroom temperature to destroy any potential contaminating genomic DNA.DNAse was inactivated by heating at 751C for 30 min. IRF-1 primers wereselected from an intron segment that would amplify only nascent,unspliced hnRNA (forward sequence AAT TTT GTT GGA TAT GTTTTC TCA TTT C located at intron 2 and reverse sequence CTG GCATTG TCA CTG TAT CTC TTT G located at intron 2). All primers weresynthesized by Emory University Microchemical Facility.
Western blot Cells were washed with phosphate-bu¡ered saline twiceand whole cell extracts were prepared by lysis with 50 mM, pH 8.0 Tris,150 mM NaCl, 0.02% Na azide, 1 mg aprotinin per mL, 1% Triton X,1 mM phenylmethylsulfonyl £uoride. The suspension was centrifuged at13,000 g for 15 min at 41C, and the protein in the supernatant was
quanti¢ed using the Bio-Rad DC Protein Assay (Bio-Rad Laboratories,Hercules, CA).Whole cell protein extract was resolved on a 10% sodiumdodecyl sulfate^polyacrylamide gel electrophoresis using reducingconcentrations. After transfer to hybond enhanced chemiluminescencenitrocellulose (Amersham Pharmacia Biotech, Piscataway, NJ), the gel wasstained with Ponceau S (Sigma) to verify uniform loading and transfer.Membranes were blocked with 5% bovine serum albumin in TBS-T (50mmol per liter Tris^HCl, 150 mmol per liter NaCl, 0.1% Tween 20, pH7.4) overnight and subsequently incubated with primary antibodies (rabbitpolyclonal IRF-1 antibody 1:500) for 1 h at room temperature. Themembrane was washed with TBS-T three times and incubated for 1 hat room temperature with the appropriate goat antirabbit horseradishperoxidase-conjugated secondary antibody. Subsequently, the membranewas washed three times with TBS-T and analyzed by enhanced chemi-luminescence (ECL Reagent, Amersham Pharmacia Biotech).
Electrophoretic mobility shift assay Nuclear extracts were pre-pared as described previously (Naik et al, 1997). All oligonucleotideswere synthesized at the Emory University Microchemical Facility.Oligonucleotides used included the following: ISRE-like site of theiNOS promoter (forward sequence 50 -TGA AAA AAA GGG AAA CGAAGA C-30 and reverse sequence 50 -G TCT TCG TTT CCC TTT TTTTCA-30) and the NF-kB-like site of the VCAM-1 promoter (forwardsequence 50 -CTG CCC TGG GTT TCC CCT TGA AGG GAT TTCCCT CCG CCT-30 and reverse sequence 50 -AGG CGG AGG GAA ATCCCT TCA AGG GGA AAC CCA GGG CAG-30). All double-strandedoligonucleotides formed by annealing reaction were completed by ¢ll-in reaction using Klenow. The [a-32P]deoxycytidine triphosphate-labeled probe was 50 end labeled using the forward reaction with T4polynucleotide kinase. The DNA binding reaction was performed underpreviously described conditions (Naik et al, 1997). The speci¢city of theIRF-1/iNOS and NF-kB/VCAM-1 binding complexes were de¢ned byutilizing cold competition and by supershift analysis with anti-IRF-1 andanti-p65 antibodies, respectively. Cold competitor and antibodies wereadded 30 min before adding the [a-32P]deoxycytidine triphosphate-labeled probe to the binding reaction. Samples were separated on 4%native polyacrylamide gel for 4 h at 130 V. Gels were dried andautoradiography was performed.
Polyribosomal analysis Co-localization of IRF-1 mRNA withpolyribosomes was analyzed via a sucrose gradient. 5A32 cells were eithertreated with DP and/or IFN-g. Prior to mRNA extraction, the abovetreatment media was removed from plated cells and replaced with mediacontaining 150 mg cycloheximide per mL for 15 min. Total mRNA wasextracted from each condition. Brie£y, whole cell lysates were preparedby scraping cells in 500 mL of ice-cold lysis bu¡er comprised of 10 mMTris^Cl (pH 7.2), 150 mM KCl, 10 mM MgCl2, 150 mg cycloheximide permL, 0.5 mg heparin per mL, 0.5% Triton X-100, 20 mM dithiothreitol (allfrom Sigma), and 100 U RNAsin per mL (from PharMingen San Diego,CA) using diethypyrocarbonate (Sigma) treated and autoclaved water.Cells were left on ice for 10 min and then centrifuged for 10 min at15,000� g. Supernatants were gently overlayed on to a linear 10 to 50%(by weight) sucrose gradient, which was poured in 12 mL Beckmanpolyallomer tubes (14� 89 mm) containing sucrose (from Sigma) in20 mMHEPES, 250 mMKCl,10 mMMgCl2, 20 mMdithiothreitol,150 mgcycloheximide, 0.5 mg heparin per ml, 10 U RNAsin per mL.The gradientswere centrifuged at 192,072 g. for 1.25 h. Twelve 1.05 mL fractions wereextracted from each gradient. Real-time reverse transcription^PCR aspreviously described was performed to quantify mRNA amounts foundin each fraction. Reverse transcription reactions were done using a setamount of volume (10 mL) from each fraction. Percentages of mRNAfound in each fraction were calculated as ((quantity mRNA in fraction)/(total quantity mRNA in all 12 fractions))� 100.
RESULTS
Previous studies examining VCAM-1 expression in HDMECidenti¢ed that TNF-a-mediated induction of IRF-1 protein wasiron dependent (Koo et al, 2003). In order to characterize the me-chanisms mediating this iron requirement and to determinewhether iron-dependent synthesis was speci¢c for TNF-a, wedetermined if TNF-a and IFN-g induction of IRF-1 proteinand mRNA could be blocked by the pretreatment with ironchelators. Both cytokines induced IRF-1 protein and mRNArapidly, with maximal levels observed after 2 h of cytokine
1192 GIRA ETAL THE JOURNAL OF INVESTIGATIVE DERMATOLOGY

stimulation (data not shown). Cultures of 5A32 endothelial cellswere pretreated with DP or DFO (250^1000 mmol) and then trea-ted withTNF-a (500 U per mL) or IFN-g (1000 U per mL) for 2h. DP or DFO pretreatment prior to IFN-g treatment resulted ina concentration-dependent inhibition of IRF-1 mRNA induc-tion (data not shown). Complete inhibition of the induction ofboth IRF-1 protein and mRNA by either TNF-a or IFN-g re-quired 24 h of DP pretreatment, and treatment with DP orDFO for 4 or 8 h resulted in only partial inhibition of TNF-aor IFN-g-induced IRF-1 expression (Fig 1A^D). Viability of5A32 cells, as assessed by trypan blue or propidium iodide exclu-sion, was not a¡ected by treatment of cells with either DP orDFO at concentrations as high as 1 mM for 24 h (data notshown).To demonstrate that the chelator e¡ect was actually mediated
via the binding of iron, we examined whether the inhibitioncould be reversed with the addition of exogenous iron. Exogen-ous iron in the form of ferric citrate was combined with DFOprior to preincubation with 5A32 cells. Addition of ferric citrateto DFO in stoichiometric ratios from 0.5:1 to 2:1 blocked DFO-mediated inhibition of IFN-g- or TNF-a-mediated induction ofIRF-1 protein and mRNA. Furthermore, at least a 1:1 ratio wasneeded to reverse the inhibitory e¡ect of DFO (Fig 1E, F). Thesedata demonstrate that iron chelator e¡ects on IRF-1 expressionare speci¢cally mediated via e¡ects on iron concentration andnot via a nonspeci¢c iron-independent mechanism.IRF-1 serves as a transcriptional activator of both pro-in£am-
matory and cell cycle regulatory genes, including iNOS (Boldt,1999; Alcantara et al, 2001). Upon appropriate cytokine stimula-tion, IRF-1 binds to ISRE located in the promoters of targetgenes (Neish et al, 1995; Sanceau et al, 1995). In order to assesswhether decreased IRF-1 expression was accompanied by loss ofbinding to ISRE in IRF-1 responsive genes, we utilized electro-
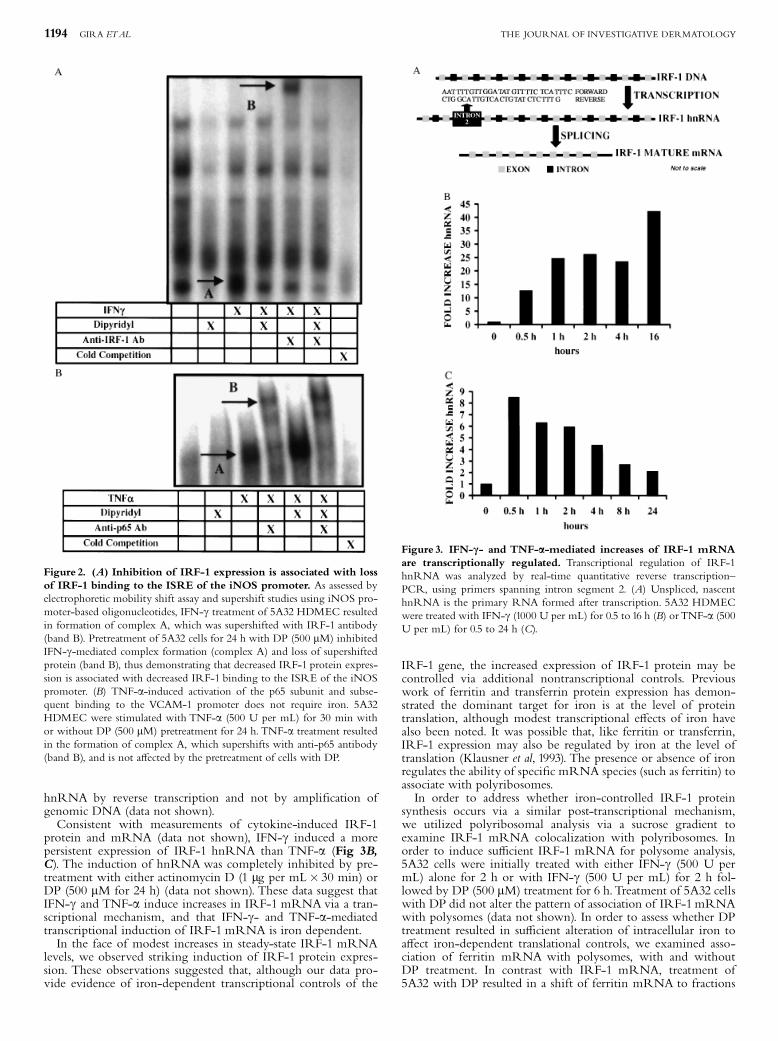
phoretic mobility shift assay. IFN-g treatment of 5A32 endothe-lial cells resulted in IRF-1 binding as assessed by electrophoreticmobility shift assay and supershift studies using iNOS promoterbased oligonucleotides. Consistent with whole cell mRNA andprotein data, pretreatment of 5A32 cells for 24 h with DP (500mM) inhibited this IFN-g-mediated complex formation, demon-strating that decreased IRF-1 protein expression is associated withcommensurate decreases in IRF-1 binding to the ISRE of theiNOS promoter (Fig 2A).This iron requirement of IFN-g- or TNF-a-induced IRF-1
expression was not due to a global e¡ect on multiple signalingpathways. 5A32 endothelial cells were pretreated with DP (500mM) for 24 h followed by treatment with TNF-a. In contrast toe¡ects on IFN-g-mediated IRF-1 binding to the iNOS promoter,pretreatment with DP did not inhibit p65 subunit binding to theVCAM-1 promoter (Fig 2B), consistent with observations madepreviously (Koo et al, 2003).Changes in steady-state IRF-1 mRNA levels may be due to
iron-mediated changes in mRNA stability or due to changes inoverall IRF-1 gene transcription. In order to address whetherIFN-g and TNF-a induction of IRF-1 was mediated via a tran-scriptional mechanism, we measured the expression of IRF-1hnRNA using primers that recognized intronic sequences speci-¢c for transiently expressed, unspliced hnRNA (Fig 3A). In-creased expression of hnRNAwas detected in 5A32 endothelialcells treated with IFN-g or TNF-a (500 U per mL) by real-timereverse transcription^PCR. To ensure that detection of nascentmessage was not the consequence of ampli¢cation of contaminat-ing genomic DNA, samples were DNAsed prior to reverse tran-scription reactions. PCR products were not detected in samplestreated with DNAase without subsequent reverse transcriptionreaction, thus demonstrating that any product detected in theexperimental samples was due to authentic ampli¢cation of
Figure1. DP blocks IFN-c and TNF-a in-duction of IRF-1 in HDMEC: Time course.5A32 HDMEC were pretreated with DP (1000mM) for 4, 8, or 24 h and then treated with IFN-g (1000 U per mL) or TNF-a (500 U per mL)for 2 h. IRF-1 protein expression as determinedby western blotting (A,C) and mRNA as deter-mined by real-time quantitative reverse trans-cription^PCR (B,D) were inhibited by DPpretreatment with maximal inhibition requiring24 h of pretreatment. DFO inhibition of IRF-1induction is reversed by addition of exogenousiron. 5A32 HDMEC were pretreated with DFO(500 mM) combined with varying doses of Fe3þ
for 24 h and then treated with (E) TNF-a (500 Uper mL) or (F) IFN-g (500 U per mL) for 2 h. Asdetermined by western blotting, increasing Fe3þ
amounts reversed the DFO inhibition of TNF-a-and IFN-g-induced IRF-1 protein.
IRF-1 IN ENDOTHELIAL CELLS 1193VOL. 121, NO. 5 NOVEMBER 2003
hnRNA by reverse transcription and not by ampli¢cation ofgenomic DNA (data not shown).Consistent with measurements of cytokine-induced IRF-1
protein and mRNA (data not shown), IFN-g induced a morepersistent expression of IRF-1 hnRNA than TNF-a (Fig 3B,C). The induction of hnRNAwas completely inhibited by pre-treatment with either actinomycin D (1 mg per mL� 30 min) orDP (500 mM for 24 h) (data not shown). These data suggest thatIFN-g and TNF-a induce increases in IRF-1 mRNA via a tran-scriptional mechanism, and that IFN-g- and TNF-a-mediatedtranscriptional induction of IRF-1 mRNA is iron dependent.In the face of modest increases in steady-state IRF-1 mRNA
levels, we observed striking induction of IRF-1 protein expres-sion. These observations suggested that, although our data pro-vide evidence of iron-dependent transcriptional controls of the
IRF-1 gene, the increased expression of IRF-1 protein may becontrolled via additional nontranscriptional controls. Previouswork of ferritin and transferrin protein expression has demon-strated the dominant target for iron is at the level of proteintranslation, although modest transcriptional e¡ects of iron havealso been noted. It was possible that, like ferritin or transferrin,IRF-1 expression may also be regulated by iron at the level oftranslation (Klausner et al, 1993). The presence or absence of ironregulates the ability of speci¢c mRNA species (such as ferritin) toassociate with polyribosomes.In order to address whether iron-controlled IRF-1 protein
synthesis occurs via a similar post-transcriptional mechanism,we utilized polyribosomal analysis via a sucrose gradient toexamine IRF-1 mRNA colocalization with polyribosomes. Inorder to induce su⁄cient IRF-1 mRNA for polysome analysis,5A32 cells were initially treated with either IFN-g (500 U permL) alone for 2 h or with IFN-g (500 U per mL) for 2 h fol-lowed by DP (500 mM) treatment for 6 h.Treatment of 5A32 cellswith DP did not alter the pattern of association of IRF-1mRNAwith polysomes (data not shown). In order to assess whether DPtreatment resulted in su⁄cient alteration of intracellular iron toa¡ect iron-dependent translational controls, we examined asso-ciation of ferritin mRNA with polysomes, with and withoutDP treatment. In contrast with IRF-1 mRNA, treatment of5A32 with DP resulted in a shift of ferritin mRNA to fractions
Figure 2. (A) Inhibition of IRF-1 expression is associated with lossof IRF-1 binding to the ISRE of the iNOS promoter. As assessed byelectrophoretic mobility shift assay and supershift studies using iNOS pro-moter-based oligonucleotides, IFN-g treatment of 5A32 HDMEC resultedin formation of complex A, which was supershifted with IRF-1 antibody(band B). Pretreatment of 5A32 cells for 24 h with DP (500 mM) inhibitedIFN-g-mediated complex formation (complex A) and loss of supershiftedprotein (band B), thus demonstrating that decreased IRF-1 protein expres-sion is associated with decreased IRF-1 binding to the ISRE of the iNOSpromoter. (B) TNF-a-induced activation of the p65 subunit and subse-quent binding to the VCAM-1 promoter does not require iron. 5A32HDMEC were stimulated with TNF-a (500 U per mL) for 30 min withor without DP (500 mM) pretreatment for 24 h. TNF-a treatment resultedin the formation of complex A, which supershifts with anti-p65 antibody(band B), and is not a¡ected by the pretreatment of cells with DP.
Figure 3. IFN-c- and TNF-a-mediated increases of IRF-1 mRNAare transcriptionally regulated. Transcriptional regulation of IRF-1hnRNA was analyzed by real-time quantitative reverse transcription^PCR, using primers spanning intron segment 2. (A) Unspliced, nascenthnRNA is the primary RNA formed after transcription. 5A32 HDMECwere treated with IFN-g (1000 U per mL) for 0.5 to 16 h (B) orTNF-a (500U per mL) for 0.5 to 24 h (C).
1194 GIRA ETAL THE JOURNAL OF INVESTIGATIVE DERMATOLOGY

less associated with polysomes (Fig 4). These data demonstratethat in our cell model, treatment of HDMEC with iron chelatorssequesters the intracellular iron pool as evidenced by its e¡ects onferritin mRNA association with polysomes. Furthermore, thesedata in concert with transcriptional studies provide evidence thatthe major regulatory mechanism controlling IRF-1 expression byIFN-g and TNF-a is transcriptional and not via a translationalmechanism.
DISCUSSION
IRF-1was originally identi¢ed through transcriptional studies ofthe IFN-b gene (Harada et al, 1994; Mamane et al, 1999). Subse-quent studies demonstrated IRF-1 expression in a number of celltypes after exposure to double-stranded RNA, a response that isimportant as a response to viral infection (Sims et al, 1993; Har-court et al, 2000). Promoters of genes responding to IRF-1 con-tain ISRE, to which IRF-1 binds and controls gene expression(Taniguchi et al, 2001).Because of its dual roles as a regulator of pro-in£ammatory
mediators and cell cycle, IRF-1 is an important transcription fac-tor in endothelial cells. IRF-1 controls the expression of majorhistocompatibility complex class I and II proteins and the expres-sion of pro-in£ammatory genes, such as VCAM-1 and interleu-kin-6 (Neish et al, 1995; Sanceau et al, 1995; Hobart et al, 1997). Ithas also been linked to the expression of cyclooxygenase II andthe angiotensin II type 2 receptor, implicating a role in vascularin£ammation and injury. IRF-1 also plays an important part incell proliferation and survival and has been designated as a tumorsuppressor gene. Its importance in that role is underscored by theobservation that loss of IRF-1 is associated with the developmentof human cancers, including pre-leukemic myelodysplastic syn-drome, esophageal cancers, and gastric cancers (Lin and Hiscott,1999; Tanaka and Taniguchi, 2000; Taniguchi et al, 2001).Limited studies have been done examining IRF-1 expression
in endothelial cells, particularly microvascular endothelial cells(Lechleitner et al, 1998; Harcourt et al, 2000; McMullen et al,2000; Liu et al, 2001; Ochi et al, 2002; Wagner et al, 2002; Wuet al, 2002). Increases in IRF-1 protein were accompanied bycommensurate increases in IRF-1 mRNA and hnRNA, with theinduction of hnRNA providing evidence that the upregulationof IRF-1 in 5A32 cells by either TNF-a or IFN-g was mediatedvia a transcriptional mechanism. IFN-g stimulation resulted in amore prolonged response than TNF-a stimulated, with near
maximal levels of IRF-1 protein and active transcription persist-ing after 16 h of stimulation.Previous studies demonstrated that pretreatment of HDMEC
with iron chelators prior toTNF-a stimulation prevented expres-sion of IRF-1 protein, thus providing evidence that IRF-1 induc-tion byTNF-a in endothelial cells was iron dependent (Koo et al,2003).We have extended these observations in this study to de-monstrate that induction of IRF-1 protein by either TNF-a andIFN-g requires the presence of iron. Furthermore, we havefurther demonstrated that iron is essential for transcriptional acti-vation of the IRF-1 gene induction by either TNF-a or IFN-g.Multiple pieces of evidence support the iron requirement. First,agents such as DP or DFO that bind iron inhibited induction ofIRF-1 protein expression and mRNA upregulation by eitherTNF-a or IFN-g. That the inhibition was mediated speci¢callyvia iron chelation was further supported by reversal of the chela-tor e¡ect and restoration of IRF-1 induction when iron wasadded back. Furthermore, treatment of HDMEC with iron che-lators clearly altered intracellular levels of iron as evidenced bychanges in ferritin association with polysomes (Klausner et al,1993). That the e¡ect of iron chelators was mediated primarilyby inhibition of IRF-1 gene transcription and not by iron-regu-lated translational controls was supported by our data showingiron chelators inhibited cytokine-mediated increases in nascentIRF-1 hnRNA without alteration of IRF-1 mRNA associationwith polysomes.Iron plays a crucial part in regulating the expression of genes
such as iNOS, protein kinase C-b, acid phosphatase type 5(Acp5)/tartrate resistant acid phosphatase, hypoxia inducible fac-tor 1a, and p21WAF1/CIP1 (Boldt, 1999; Alcantara et al, 2001). Inthe case of hypoxia inducible factor 1a, iron is required for genesuppression. This suppression is mediated via an iron dependentproline hydroxylase, which controls hypoxia inducible factor 1ainteraction with VHL protein (Ivan et al, 2001). In contrast, themechanisms by which iron regulates the expression of protein ki-nase C-b, Acp5 (type 5 isoenzyme of acid phosphatase/tartrateresistant acid phosphatase), and the cyclin-dependent kinaseinhibitor, p21WAF1/CIP1 (Boldt, 1999) are not well characterized.The mechanisms mediating the iron dependence of IFN-g and
TNF-a signaling appear to be distinct. Chelator treatment had noe¡ect onTNF-a-mediated NF-kB activation or nuclear transloca-tion. Previous studies have suggested that iron may be essentialfor chromatin remodeling events required for TNF-a-mediatedactivation of the VCAM-1 gene as well as other NF-kB-depen-dent genes (Koo et al, 2003). IRF-1 induction by TNF-a mayrequire similar iron sensitive chromatin remodeling events.Further de¢nition of the role of iron in these signaling pathwaysmay provide essential insights into the in£ammatory mediators inthe skin, the contributions of iron, and the potential value of ironchelators for the treatment of in£ammatory skin disorders.
This work was supported by NIAMS R01 AR39632, the Emory Skin DiseaseResearch Center (NIH P30 AR42687), and NRSA Institutional Training Grant(T32) in Dermatology.
REFERENCES
Ackerman Z, Seidenbaum M, Loewenthal E, Rubinow A: Overload of iron in theskin of patients with varicose ulcers. Possible contributing role of iron accu-mulation in progression of the disease. Arch Dermatol 124:1376^1378, 1988
Ades EW, Candal FJ, Swerlick RA, George VG, Summers S, Bosse DC, LawleyTJ:HMEC-1: Establishment of an immortalized human microvascular endothe-lial cell line. J Invest Dermatol 99:683^690, 1992
Alcantara O, Kalidas M, Baltathakis I, Boldt DH: Expression of multiple genes reg-ulating cell cycle and apoptosis in di¡erentiating hematopoietic cells is depen-dent on iron. Exp Hematol 29:1060^1069, 2001
Bissett DL, Chatterjee R, Hannon DP: Chronic ultraviolet radiation-inducedincrease in skin iron and the photoprotective e¡ect of topically applied ironchelators. Photochem Photobiol 54:215^223, 1991
Figure 4. Chelator treatment results in decreased intracellular freeiron concentrations. 5A32 cells were either treated with IFN-g (500 Uper mL) alone for 2 h or treated with IFN-g (500 U per mL) for 2 h andthen treated with DP (500 mM) for 6 h. Total mRNAwas examined usingthe described polyribosomal analysis and quanti¢ed by real-time reversetranscription^PCR. Percentages of mRNA in each fraction were calcu-lated. DP prevented ferritin mRNA colocalization with polyribosomes,thus demonstrating that chelator treatment decreases intracellular free ironconcentrations.
IRF-1 IN ENDOTHELIAL CELLS 1195VOL. 121, NO. 5 NOVEMBER 2003

Bissett DL, Oelrich DM, Hannon DP: Evaluation of a topical iron chelator in ani-mals and in human beings: Short-term photoprotection by 2-furildioxime.J Am Acad Dermatol 31:572^578, 1994
Boldt DH: New perspectives on iron: An introduction. AmJMed Sci 318:207^212, 1999Conrad ME: Introduction: Iron overloading disorders and iron regulation. Semin
Hematol 35:1^4, 1998David TJ,Wells FE, Sharpe TC, Gibbs AC, Devlin J: Serum levels of trace metals in
children with atopic eczema. Br J Dermatol 122:485^489, 1990Elferink CJ, Reiners JJ Jr: Quantitative RT-PCR on CYP1A1 heterogeneous nuclear
RNA: A surrogate for the in vitro transcription run-on assay. Biotechniques20:470^477, 1996
Harada H, Fujita T, Miyamoto M, et al: Structurally similar but functionally distinctfactors, IRF-1 and IRF-2, bind to the same regulatory elements of IFN- andIFN-inducible genes. Cell 58:729^739, 1989
Harada H,Takahashi E, Itoh S, Harada K, Hori TA,Taniguchi T: Structure and reg-ulation of the human interferon regulatory factor 1 (IRF-1) and IRF-2 genes:implications for a gene network in the interferon system. Mol Cell Biol 14:1500^1509, 1994
Harcourt JL, Hagan MK, O¡ermann MK: Modulation of double-stranded RNA-mediated gene induction by interferon in human umbilical vein endothelialcells. J Interferon Cytokine Res 20:1007^1013, 2000
Hobart M, Ramassar V, Goes N, Urmson J, Halloran PF: IFN-regulatory factor-1plays a central role in the regulation of the expression of class I and II MHCgenes in vivo. J Immunol 158:4260^4269, 1997
Horky LL, Pluta RM, Boock RJ, Old¢eld EH: Role of ferrous iron chelator 2,20 -dipyridyl in preventing delayed vasospasm in a primate model of subarachnoidhemorrhage. J Neurosurg 88:298^303, 1998
Ivan M, Kondo K, Yang H, et al: HIFa targeted for VHL-mediated destructionby proline hydroxylation: Implications for O2 sensing. Science 292:464^468,2001
Klausner RD, Rouault TA, Harford JB: Regulating the fate of mRNA. The controlof cellular iron metabolism. Cell 72:19^28, 1993
Koo S, Casper K, Otto K, Gira A, Swerlick R: Iron chelators inhibit VCAM-1 ex-pression in human dermal microvascular endothelial cells. J Invest Dermatol120:871^879, 2003
Lechleitner S, Gille J, Johnson DR, Petzelbauer P: Interferon enhances tumornecrosis factor-induced vascular cell adhesion molecule 1 (CD106) expressionin human endothelial cells by an interferon-related factor 1-dependent path-way. J Exp Med 187:2023^2030, 1998
Lin R, Hiscott J: A role for casein kinase II phosphorylation in the regulation ofIRF-1 transcriptional activity. Mol Cell Biochem 191:169^180, 1999
Liu L, Paul A, Mackenzie CJ, Bryant C, Graham A, Plevin R: Nuclear factor kappaB is involved in lipopolysaccharide-stimulated induction of interferon regula-tory factor-1 and GAS/GAF DNA-binding in human umbilical vein endothe-lial cells. Br J Pharmacol 134:1629^1638, 2001
Liu ZD, Hider RC: Design of clinically useful iron (III)-selective chelators. Med ResRev 22:26^64, 2002
Mamane Y, Heyl Broeck C, Genin P, et al: Interferon regulatory factors: The nextgeneration. Gene 237:1^14, 1999
McCord JM: Iron, free radicals, and oxidative injury. Semin Hematol 35:5^12, 1998McMullen CB, Fleming E, Clarke G, Armstrong MA: The role of reactive oxygen
intermediates in the regulation of cytokine-induced ICAM-1 surface expres-sion on endothelial cells. Mol Cell Biol Res Commun 3:231^237, 2000
Miyamoto M, Fujita T, KumuraY, et al: Regulated expression of a gene encoding anuclear factor, IRF-1, that speci¢cally binds to IFN-beta gene regulatory ele-ments. Cell 54:903^913, 1988
Molin L, Wester PO: Iron content in normal and psoriatic epidermis. Acta DermVenereol 53:473^476, 1973
Naik SM, Shibagaki N, Li LJ, Quinlan KL, Paxton LL, Caughman SW: Interferongamma-dependent induction of human intercellular adhesion molecule-1 geneexpression involves activation of a distinct STAT protein complex. J Biol Chem272 :1283^1290, 1997
Neish AS, Read MA, Thanos D, Pine R, Maniatis T, Collins T: Endothelial inter-feron regulatory factor 1 cooperates with NF-kappa B as a transcriptional acti-vator of vascular cell adhesion molecule 1. Mol Cell Biol 15:2558^2569, 1995
Ochi H, Masuda J, Gimbrone MA: Hyperosmotic stimuli inhibit VCAM-1 expres-sion in cultured endothelial cells via e¡ects on interferon regulatory factor-1expression and activity. Eur J Immunol 32:1821^1831, 2002
Sanceau J, Kaisho T, Hirano T,Wietzerbin J: Triggering of the human interleukin-6gene by interferon-g and tumor necrosis factor-a in monocytic cells involvescooperation between interferon regulatory factor-1, NF kappa B, and Sp1 tran-scription factors. J Biol Chem 270:27920^27931, 1995
Sato M,Taniguchi T,Tanaka N:The interferon system and interferon regulatory fac-tor transcription factors�studies from gene knockout mice. Cytokine GrowthFactor Rev 12:133^142, 2001
Sims SH, ChaY, Romine MF, Gao PQ, Gottlieb K, Deisseroth AB: A novel inter-feron-inducible domain: Structural and functional analysis of the human inter-feron regulatory factor 1 gene promoter. Mol Cell Biol 13:690^702, 1993
Syvertsen C, McKinley-McKee JS: 2,2-dipyridyl binding to metal substituted horseliver alcohol dehydrogenase. J Inorg Biochem 22:43^53, 1984
Tanaka N, Taniguchi T: The interferon regulatory factors and oncogenesis. SeminCancer Biol 10:73^81, 2000
Taniguchi T, Ogasawara K,Takaoka A,Tanaka N: IRF family of transcription factorsas regulators of host defense. Annu Rev Immunol 19:623^655, 2001
Wagner AH, Gebauer M, Pollok-Kopp B, Hecker M: Cytokine-inducible CD40expression in human endothelial cells is mediated by interferon regulatoryfactor-1. Blood 99:520^525, 2002
Wu F, Tyml K,Wilson JX: Ascorbate inhibits iNOS expression in endotoxin- andIFN-gamma-stimulated rat skeletal muscle endothelial cells. FEBS Lett520:122^126, 2002
1196 GIRA ETAL THE JOURNAL OF INVESTIGATIVE DERMATOLOGY