increasing frequency of bite wounds with increasing population density in eurasian badgers, meles...
TRANSCRIPT

ANIMAL BEHAVIOUR, 2004, 67, 745e751doi:10.1016/j.anbehav.2003.08.021
Increasing frequency of bite wounds with increasing population
density in Eurasian badgers, Meles meles
D. W. MACDONALD, B. J. HARMSEN, P. J. JOHNSON & C. NEWMAN
Wildlife Conservation Research Unit, Department of Zoology, University of Oxford
(Received 9 March 2003; initial acceptance 28 April 2003;
final acceptance 30 August 2003; MS. number: 7644)
Eurasian badgers sometimes live in territorial, mixed-sex groups; the adaptive significance of this is notunderstood, but members generally interact amicably. None the less, badgers occasionally fight and inflictsometimes severe wounds on one another. Based on 498 badger life histories, from first emergence as a cubuntil death, documented during a 10-year trapping study at Wytham Woods, Oxfordshire, U.K., thepatterns and rates of bite wounding and consequential scarring were examined. Male badgers receivedmore wounds and more severe wounds than did females. Wounding rates for both sexes increasedsignificantly with age, and there was evidence that heavier individuals received most wounds. No seasonalpattern in wounding rates was apparent. During the study, the badger population size increased three-foldand wounding rates, particularly in males, showed a density-dependent increase. The rate of bitewounding increased with group size, and this increase was more marked among males than amongfemales. Among males, but not females, the rate of bite wounding also increased with the number ofbadgers living in adjoining territories.
� 2004 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved.
Despite prolonged study, the social system of Eurasianbadgers has so far eluded more than superficial un-derstanding (Kruuk 1989; Woodroffe & Macdonald 1992;da Silva et al. 1994; Neal & Cheeseman 1996; Macdonaldet al. 2002a). Evidence is emerging that ectoparasitecontrol is one fundamental form of cooperation in badgersociety (Stewart & Macdonald 2003; Johnson et al., inpress) whereas evidence of alloparental behaviour remainspreliminary (Woodroffe 1993). There is, however, noevidence of a stable social hierarchy within badger groups(Macdonald et al. 2002a), but a conspicuous feature ofbadger society is that individuals fight both within andbetween groups, as revealed by video observations of ourstudy population (Stewart et al. 1997b).Fighting most often involves ritualized escalations of
aggressive behaviour, starting with jaw-to-jaw contests,which can proceed to mutual neck biting, which,although dangerous, provides neither badger with a clearadvantage. Severe fighting involves badgers trying toavoid exposing their heads to attack, so strikes are directedagainst the opponent’s body. An advantage in sucha contest is achieved if one combatant locks a bite on tothe opponent’s rump and circles swiftly to prevent themanoeuvre being reciprocated. The bitten individual will
Correspondence: D. W. Macdonald, Wildlife Conservation ResearchUnit, Department of Zoology, University of Oxford, South Parks Road,Oxford OX1 3PS, U.K. (email: [email protected]).
740003e3472/03/$30.00/0 � 2004 The Association
ultimately pull away and savage, extensive rump woundscan be inflicted in this way on the skin and subcutaneoustissue of the recipient. Later examination of fresh woundsoften reveals secondary infection, the presence of maggotsand necrotic tissue, so there are additional chronic healthimplications to severe wounding. Successful healing ofa wound produces inelastic granular tissue, often nodular,which might provide considerable protection againstfuture bites (Keeble 1999). Badgers have an unresolvedrole in the transmission of bovine tuberculosis (bTB; Krebset al. 1997). Mycobacterium bovis, the bacterium responsi-ble for bTB, has been isolated from the swabbed woundsof infected badgers and thus bite wounding may be animportant factor in the spread of bTB within badgerpopulations (Gallagher & Nelson 1979; Clifton-Hadleyet al. 1993).The contexts of these fights are not yet understood, but
their consequences are clearly visible, and quantifiable inthe form of variously severe wounds or subsequently scars.We have live-trapped, and examined closely, a substantialproportion of our study population at Wytham Woods,typically four times annually since 1987 (Macdonald &Newman 2002), and have recorded the incidence of theseinjuries. Thus, we have the opportunity to evaluate howfrequently badgers incur injuries from bites, and how thisincidence varies with their sex, age and weight. Further-more, during our study, the population size and groupsizes have increased and changed in spatial organization,
5for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved.

ANIMAL BEHAVIOUR, 67, 4746
revealing density dependence in some parameters(Macdonald et al. 2002b), enabling us to ask also whetherthe frequency of bite wounding, a measure of socialtension, varies with local population density. In this study,we addressed these questions with the aim of furtherelucidating the nature of badger society.
METHODS
We conducted this study at Wytham Woods, 5 km north-west of Oxford, U.K. (OS ref: SP 462080). Wytham’sbadgers have been studied continually since the 1970s(Kruuk 1978), and we have attempted to mark the entirepopulation annually since 1987 (Macdonald & Newman2002). Under U.K. Home Office and English NatureGovernment licences, we set steel mesh cage traps(85!37!38 cm), baited with peanuts, without priorprebaiting, to catch badgers at each sett and at associatedoutlying holes in the study area on at least four occasionsannually: early January (winter), MayeJune (spring),JulyeAugust (summer) and OctobereNovember (au-tumn). Trapping was suspended between February andApril (inclusive), to avoid disturbance during the finaltrimester of gestation and the period during whichaltricial young are highly dependent. To ensure saturationtrapping, more traps than the estimated number ofresident badgers were set at each sett. Traps were set for3 successive days. Once captured, badgers were transferredto a holding cage and removed to a processing area wherethey were anaesthetized with ketamine hydrochloride0.2 ml/kg body weight (Hunt 1976; Mackintosh et al.1976; Thornton et al., in press). As a prerequisite to alldemographic analyses, badgers were permanently markedwith a tattoo in the inguinal region (Cheeseman & Harris1982) when first caught; thus the identity of eachindividual could be logged against the site and date ofcapture. After 3 h to allow them to recover fromanaesthesia, all badgers were assessed and released at thesite of capture.Each time a badger was captured, we noted the details of
scars and wounds (which we collectively refer to here as‘injuries’), recording details of severity, freshness andlocation. Since 1987, the detail recorded has been pro-gressively refined, culminating for the years 1993e2002 inrecords that noted the sites (head/neck, rump, back,underside) of fresh injuries (which we refer to here as‘wounds’) and older scars. So, analysis of body-site-specificinjuries could use only the subset of data from1993e2002; analyses of nonsite-specific injuries coulddraw upon the full sequence of data. Although weoccasionally found injuries from other sources (e.g.entanglement in wire, glass cuts), most injuries wereclearly identifiable as bite wounds. Wounds were definedas fresh bites (open/haemorrhaging, infected/suppuratingor lightly scabbed), occasionally with entire areas ofepidermis pulled away and severe subcutaneous traumato soft tissues; these could impair mobility and sometimesevidenced severe secondary infection. Scars were definedas areas of healed epithelialized granular tissue indicativeof previous wounds. Severe wounds are a subset of
wounds in general, which were also rated as minor andmoderate; the same applies to scars. Wounds alwaysbecome scars; however, only the largest wounds producescars visible through the fur.
Each time an individual was trapped, we recordeda variety of other biometric parameters, of which thefollowing are relevant to this analysis.
Maximum weight. We defined this as the heaviestweight recorded for an individual captured at least threetimes as an adult over the 10-year period, including atleast one measurement during autumn or winter. We alsoused weight recorded at each capture as a predictor ofwhether wounds were recorded at the same capture event.
Age. As the study progressed, the age of an increasingpercentage of the population was known. We grouped allbadgers of a known age throughout the 10-year periodand calculated the percentage of badgers within each ageclass for which wounds of any degree were recorded. Wealso made a vertical analysis of age-specific wounding for1999 to verify any trends seen, because the percentage ofmature known-age badgers increased during the study.
Number of animals per social group. Setts were assignedto social groups on the basis of bait-marking data (Kruuk1978; Feore & Montgomery 1999; Delahay et al. 2000).The number of males, females and total number ofindividuals per social group were calculated for each yearon the basis of animals caught at these setts. Macdonald &Newman (2002) reported a robust relation betweenannual trapping returns and retrospective MinimumNumber Alive (MNA) estimates, with a mean G SE of86.11 G 1.99% of the MNA-estimated population beingreturned by the trapping effort between 1988 and 1996.We therefore used numbers trapped as an estimate ofpopulation size.
Number of individuals in neighbouring social groups.Bait marking revealed social group territories to becontiguous over the study area, forming a tessellatingmosaic; thus, each social group bordered several neigh-bouring ranges, which we defined as neighbouring socialgroups. The number of neighbouring males and femalesfor each social group per year was established from thetrapping record.
Percentage of badgers recorded with wounds at leastonce per year. The percentage of wounded badgers withina social group per year (hereafter referred to as thewounding frequency) was calculated as:
Predictive Variables Analysed
We analysed the influence of sex, age, weight, seasonand group size on the wounding pattern. For the sexvariable, we carried out analyses at both the individuallevel, where the response was therefore binary (woundedor not wounded), and the social group level where
% badgerswounded
¼
total number of wounded badgers
within social group A at year t
total number of badgers
within social group A at year t
!100
MACDONALD ET AL.: BITE WOUNDS IN BADGERS 747
responses were proportions of badgers. For the age vari-able, we treated badgers as adult at 1 year. We definedseasons as winter (December, January, February), spring(March, April, May), summer (June, July, August) andautumn (September, October, November).Concerning group size, we knew that the density of
badgers at Wytham Woods was increasing throughout thestudy (Macdonald & Newman 2002). We investigatedwhether, at a smaller spatial scale, the size of individualsocial groups had any additional effect, and whether thenumber of badgers in adjacent social groups influencedwounding rates within social groups.We constructed a GLM model predicting wounding
frequencies, which included social group identity asa blocking factor and year as a continuous predictor.Additional terms for numbers of badgers in a social groupand in neighbouring groups were then included in themodel. Because these potential predictors are correlatedwith ‘year’, only sequential (type 1) models are informa-tive. We also examined the effect of discriminatingbetween the sexes for group members and neighbours aspredictors. This allowed us to investigate whether, forexample, wounding frequencies of males were affected byvariation in the number of neighbouring males but notfemales. Initial analyses showed that for some of ourresponses of interest, there was evidence that the maineffects of sex and the number of neighbouring badgersshowed significant interaction. Models were thereforeconstructed separately for males and females and also forthe two wound severities. The outputs from these modelsare not independent. Responses used as proportions werearcsine square-root transformed.
Analysis
Binary logistic regressions, predicting whether badgerswere wounded or scarred, which included badger ID asa random factor, were performed using SAS (GENMODProcedure, SPSS Inc., Chicago, U.S.A.) to predict the effectof weight, season and sex on whether a wound wasrecorded at a trapping event. The effect of social group sizewas investigated with the SAS GLM procedure (SPSS Inc.).For other analyses, we used SPSS for Windows v. 10 (SASInstitute 1985).
RESULTS
Demographic Differences in Bite Wounds
SexA greater proportion of males than females received bite
wounds (Table 1). This is based on 498 individual capturesequences in 1990e1999 varying in length between twocaptures separated by several months and 50 capturesspaced over more than 12 years. The statistical signifi-cance of this sex bias varied with the severity category ofbite wounds (Table 1). Males incurred more rump woundsand scars, and more wounds overall than did females.
AgeThe percentage of wounded badgers increased with age
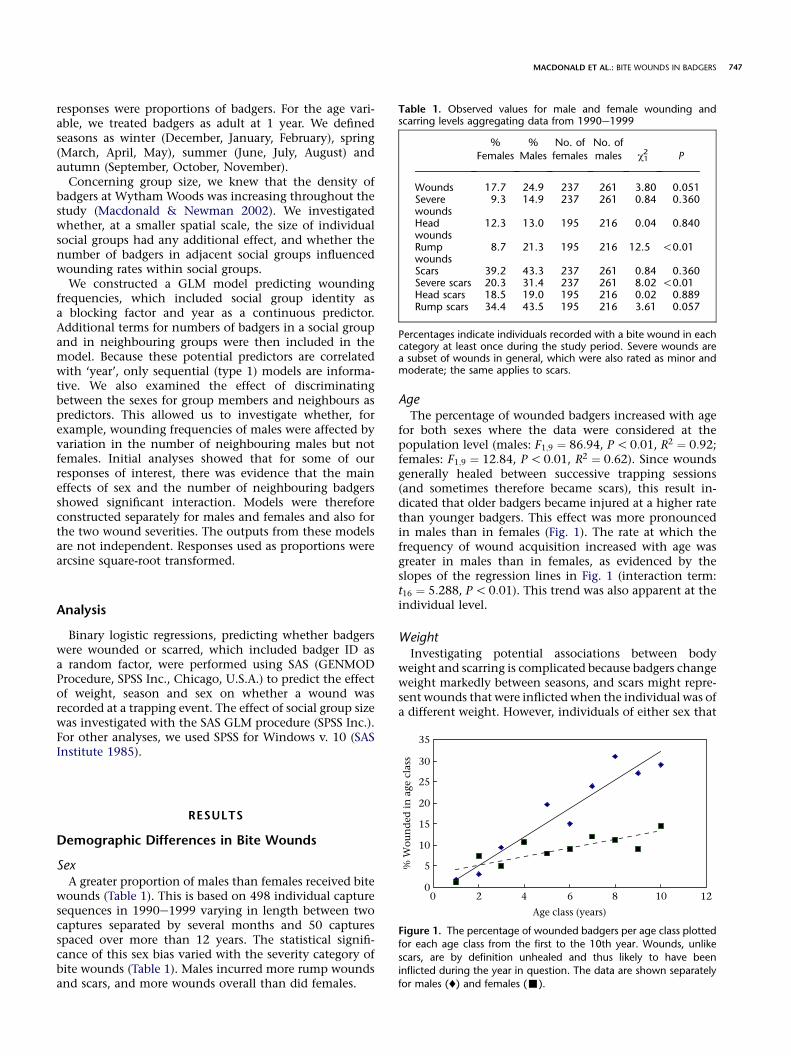
for both sexes where the data were considered at thepopulation level (males: F1;9 ¼ 86:94, P!0:01, R2 ¼ 0:92;females: F1;9 ¼ 12:84, P!0:01, R2 ¼ 0:62). Since woundsgenerally healed between successive trapping sessions(and sometimes therefore became scars), this result in-dicated that older badgers became injured at a higher ratethan younger badgers. This effect was more pronouncedin males than in females (Fig. 1). The rate at which thefrequency of wound acquisition increased with age wasgreater in males than in females, as evidenced by theslopes of the regression lines in Fig. 1 (interaction term:t16 ¼ 5:288, P!0:01). This trend was also apparent at theindividual level.
WeightInvestigating potential associations between body
weight and scarring is complicated because badgers changeweight markedly between seasons, and scars might repre-sent wounds that were inflicted when the individual was ofa different weight. However, individuals of either sex that
Table 1. Observed values for male and female wounding andscarring levels aggregating data from 1990e1999
%
Females
%
Males
No. of
females
No. of
males c12 P
Wounds 17.7 24.9 237 261 3.80 0.051Severewounds
9.3 14.9 237 261 0.84 0.360
Headwounds
12.3 13.0 195 216 0.04 0.840
Rumpwounds
8.7 21.3 195 216 12.5 !0.01
Scars 39.2 43.3 237 261 0.84 0.360Severe scars 20.3 31.4 237 261 8.02 !0.01Head scars 18.5 19.0 195 216 0.02 0.889Rump scars 34.4 43.5 195 216 3.61 0.057
Percentages indicate individuals recorded with a bite wound in eachcategory at least once during the study period. Severe wounds area subset of wounds in general, which were also rated as minor andmoderate; the same applies to scars.
Figure 1. The percentage of wounded badgers per age class plottedfor each age class from the first to the 10th year. Wounds, unlike
scars, are by definition unhealed and thus likely to have been
inflicted during the year in question. The data are shown separately
for males (¤) and females (-).
ANIMAL BEHAVIOUR, 67, 4748
had been scarred during their lifetime had a significantlyhigher maximum weight than those that had never beenrecorded as being scarred (considering only adult badgers),which suggested at least a generalized association betweenmaximum weight and bite-wounding rates (males:Nscarred ¼ 99, mean 11.74 kg, Nnot scarred ¼ 74, mean11.14 kg, t test: t172 ¼ 1:97, P ¼ 0:04; females: Nscarred ¼11:12 kg, Nnot scarred ¼ 68, mean 10.56 kg, t141 ¼ 1:96, P ¼0:05).Whenwe explored this further using a logisticmodel(GENMOD) predicting wounding status (modelling theprobability that woundedZ ‘yes’), this provided inconclu-sive evidence for a maximum weight effect (c2
1 ¼ 3:09,P ¼ 0:07). The effect was, however, in the expected di-rection (parameter estimate G SEZ 0.10 G 0.06). Therewas no evidence for seasonal variation in wounding rates(c2
3 ¼ 6:12, P ¼ 0:10). The interaction terms including sexand both season and weight were nonsignificant (sex*sea-son: c2
3 ¼ 2:27, P ¼ 0:52; weight*sex: c21 ¼ 1:67, P ¼ 0:20).
Themaineffect of sexwas,however, significant, confirminga difference in male/female wounding frequencies at thelevel of individual badgers (c2
1 ¼ 5:85, P ¼ 0:016). Assuggested at the population level (Fig. 1), the sex effectdiffered with age (c2
1 ¼ 6:41, P ¼ 0:011).
SeasonThe logistic model showed no significant seasonal
pattern to the incidence of wounding.
Group sizePopulation trends. Thepercentageof individualswounded
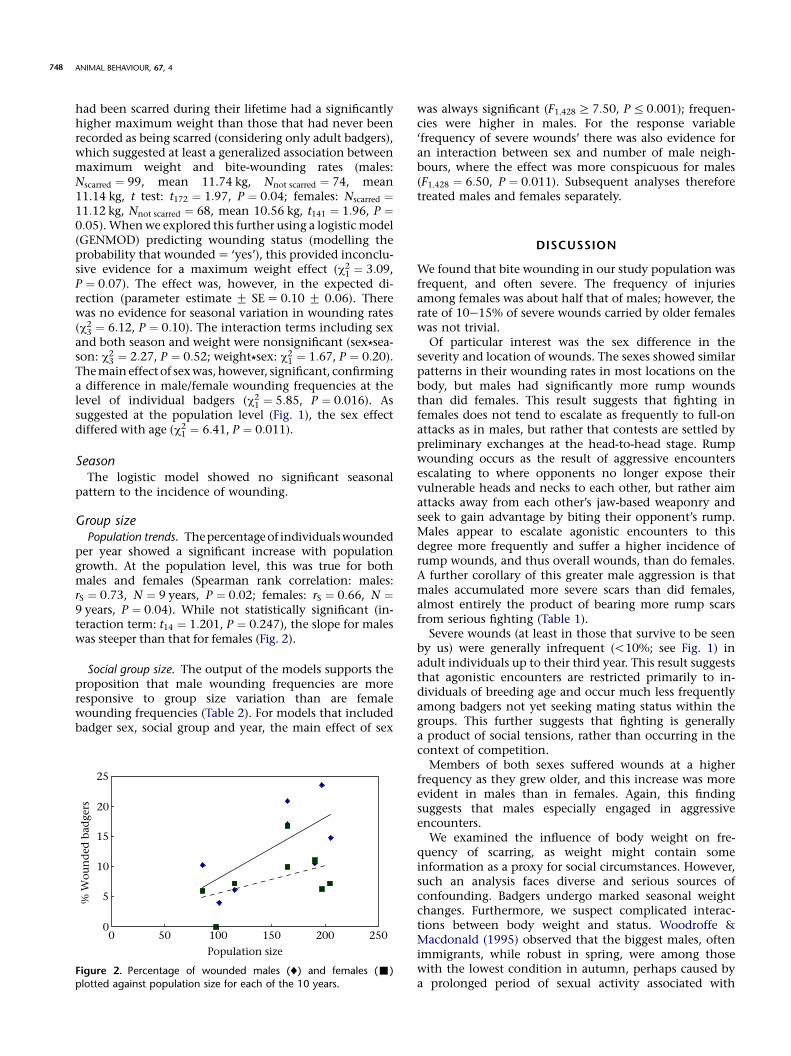
per year showed a significant increase with populationgrowth. At the population level, this was true for bothmales and females (Spearman rank correlation: males:rS ¼ 0:73, N ¼ 9 years, P ¼ 0:02; females: rS ¼ 0:66, N ¼9 years, P ¼ 0:04). While not statistically significant (in-teraction term: t14 ¼ 1:201, P ¼ 0:247), the slope for maleswas steeper than that for females (Fig. 2).
Social group size. The output of the models supports theproposition that male wounding frequencies are moreresponsive to group size variation than are femalewounding frequencies (Table 2). For models that includedbadger sex, social group and year, the main effect of sex
Figure 2. Percentage of wounded males (¤) and females (-)plotted against population size for each of the 10 years.
was always significant (F1;428 R7:50, P%0:001); frequen-cies were higher in males. For the response variable‘frequency of severe wounds’ there was also evidence foran interaction between sex and number of male neigh-bours, where the effect was more conspicuous for males(F1;428 ¼ 6:50, P ¼ 0:011). Subsequent analyses thereforetreated males and females separately.
DISCUSSION
We found that bite wounding in our study population wasfrequent, and often severe. The frequency of injuriesamong females was about half that of males; however, therate of 10e15% of severe wounds carried by older femaleswas not trivial.
Of particular interest was the sex difference in theseverity and location of wounds. The sexes showed similarpatterns in their wounding rates in most locations on thebody, but males had significantly more rump woundsthan did females. This result suggests that fighting infemales does not tend to escalate as frequently to full-onattacks as in males, but rather that contests are settled bypreliminary exchanges at the head-to-head stage. Rumpwounding occurs as the result of aggressive encountersescalating to where opponents no longer expose theirvulnerable heads and necks to each other, but rather aimattacks away from each other’s jaw-based weaponry andseek to gain advantage by biting their opponent’s rump.Males appear to escalate agonistic encounters to thisdegree more frequently and suffer a higher incidence ofrump wounds, and thus overall wounds, than do females.A further corollary of this greater male aggression is thatmales accumulated more severe scars than did females,almost entirely the product of bearing more rump scarsfrom serious fighting (Table 1).
Severe wounds (at least in those that survive to be seenby us) were generally infrequent (!10%; see Fig. 1) inadult individuals up to their third year. This result suggeststhat agonistic encounters are restricted primarily to in-dividuals of breeding age and occur much less frequentlyamong badgers not yet seeking mating status within thegroups. This further suggests that fighting is generallya product of social tensions, rather than occurring in thecontext of competition.
Members of both sexes suffered wounds at a higherfrequency as they grew older, and this increase was moreevident in males than in females. Again, this findingsuggests that males especially engaged in aggressiveencounters.
We examined the influence of body weight on fre-quency of scarring, as weight might contain someinformation as a proxy for social circumstances. However,such an analysis faces diverse and serious sources ofconfounding. Badgers undergo marked seasonal weightchanges. Furthermore, we suspect complicated interac-tions between body weight and status. Woodroffe &Macdonald (1995) observed that the biggest males, oftenimmigrants, while robust in spring, were among thosewith the lowest condition in autumn, perhaps caused bya prolonged period of sexual activity associated with
MACDONALD ET AL.: BITE WOUNDS IN BADGERS 749
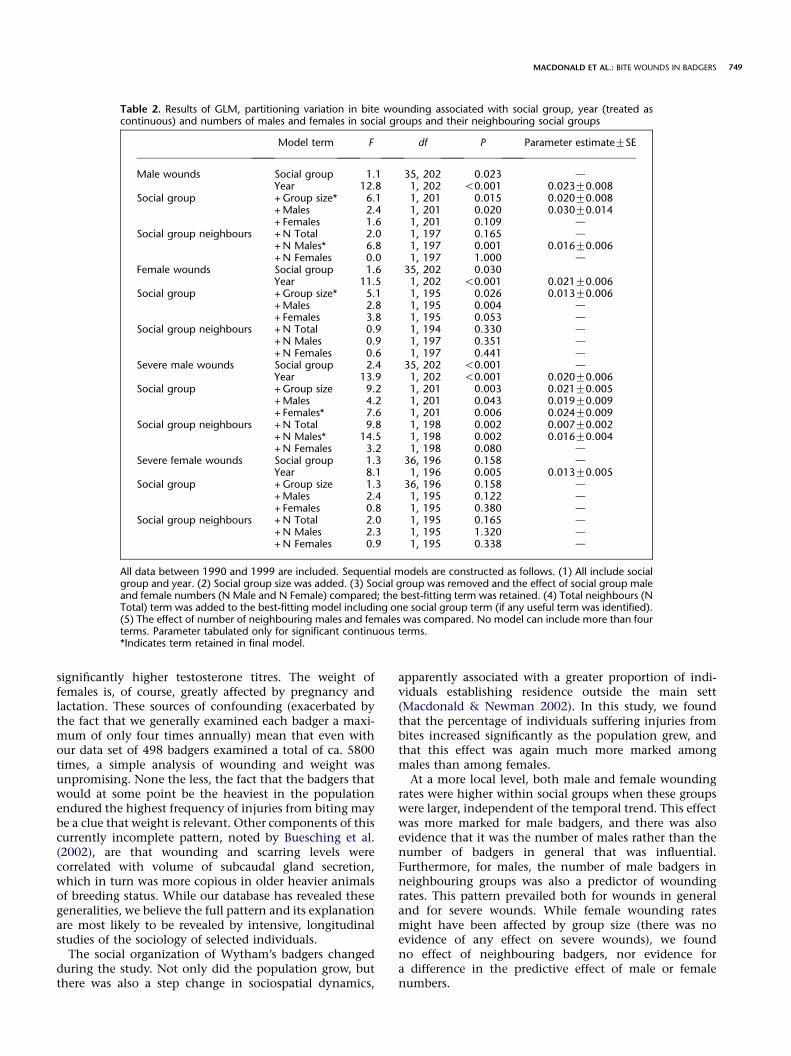
Table 2. Results of GLM, partitioning variation in bite wounding associated with social group, year (treated ascontinuous) and numbers of males and females in social groups and their neighbouring social groups
Model term F df P Parameter estimateGSE
Male wounds Social group 1.1 35, 202 0.023 dYear 12.8 1, 202 !0.001 0.023G0.008
Social group + Group size* 6.1 1, 201 0.015 0.020G0.008+Males 2.4 1, 201 0.020 0.030G0.014+ Females 1.6 1, 201 0.109 d
Social group neighbours + N Total 2.0 1, 197 0.165 d+ N Males* 6.8 1, 197 0.001 0.016G0.006+ N Females 0.0 1, 197 1.000 d
Female wounds Social group 1.6 35, 202 0.030Year 11.5 1, 202 !0.001 0.021G0.006
Social group + Group size* 5.1 1, 195 0.026 0.013G0.006+Males 2.8 1, 195 0.004 d+ Females 3.8 1, 195 0.053 d
Social group neighbours + N Total 0.9 1, 194 0.330 d+ N Males 0.9 1, 197 0.351 d+ N Females 0.6 1, 197 0.441 d
Severe male wounds Social group 2.4 35, 202 !0.001 dYear 13.9 1, 202 !0.001 0.020G0.006
Social group + Group size 9.2 1, 201 0.003 0.021G0.005+Males 4.2 1, 201 0.043 0.019G0.009+ Females* 7.6 1, 201 0.006 0.024G0.009
Social group neighbours + N Total 9.8 1, 198 0.002 0.007G0.002+ N Males* 14.5 1, 198 0.002 0.016G0.004+ N Females 3.2 1, 198 0.080 d
Severe female wounds Social group 1.3 36, 196 0.158 dYear 8.1 1, 196 0.005 0.013G0.005
Social group + Group size 1.3 36, 196 0.158 d+Males 2.4 1, 195 0.122 d+ Females 0.8 1, 195 0.380 d
Social group neighbours + N Total 2.0 1, 195 0.165 d+ N Males 2.3 1, 195 1.320 d+ N Females 0.9 1, 195 0.338 d
All data between 1990 and 1999 are included. Sequential models are constructed as follows. (1) All include socialgroup and year. (2) Social group size was added. (3) Social group was removed and the effect of social group maleand female numbers (N Male and N Female) compared; the best-fitting term was retained. (4) Total neighbours (NTotal) term was added to the best-fitting model including one social group term (if any useful term was identified).(5) The effect of number of neighbouring males and females was compared. No model can include more than fourterms. Parameter tabulated only for significant continuous terms.*Indicates term retained in final model.
significantly higher testosterone titres. The weight offemales is, of course, greatly affected by pregnancy andlactation. These sources of confounding (exacerbated bythe fact that we generally examined each badger a maxi-mum of only four times annually) mean that even withour data set of 498 badgers examined a total of ca. 5800times, a simple analysis of wounding and weight wasunpromising. None the less, the fact that the badgers thatwould at some point be the heaviest in the populationendured the highest frequency of injuries from biting maybe a clue that weight is relevant. Other components of thiscurrently incomplete pattern, noted by Buesching et al.(2002), are that wounding and scarring levels werecorrelated with volume of subcaudal gland secretion,which in turn was more copious in older heavier animalsof breeding status. While our database has revealed thesegeneralities, we believe the full pattern and its explanationare most likely to be revealed by intensive, longitudinalstudies of the sociology of selected individuals.The social organization of Wytham’s badgers changed
during the study. Not only did the population grow, butthere was also a step change in sociospatial dynamics,
apparently associated with a greater proportion of indi-viduals establishing residence outside the main sett(Macdonald & Newman 2002). In this study, we foundthat the percentage of individuals suffering injuries frombites increased significantly as the population grew, andthat this effect was again much more marked amongmales than among females.At a more local level, both male and female wounding
rates were higher within social groups when these groupswere larger, independent of the temporal trend. This effectwas more marked for male badgers, and there was alsoevidence that it was the number of males rather than thenumber of badgers in general that was influential.Furthermore, for males, the number of male badgers inneighbouring groups was also a predictor of woundingrates. This pattern prevailed both for wounds in generaland for severe wounds. While female wounding ratesmight have been affected by group size (there was noevidence of any effect on severe wounds), we foundno effect of neighbouring badgers, nor evidence fora difference in the predictive effect of male or femalenumbers.

ANIMAL BEHAVIOUR, 67, 4750
The finding that wounding rates, especially amongmales, were affected by the number of males in theneighbouring territory provides evidence on the contextsof fighting behaviour. Video surveillance of focal badgersetts in Wytham rarely records the sort of ferociousfighting that would result in the severe rump wounds weobserved through our trapping programme. Rather, weobserved, often as extensions of radiotracking surveys,that vicious fighting occurs away from the sett, in thevicinity of group territory boundaries. As male woundingrates were driven partly by the number of neighbouringmales, it would be plausible to conclude that fightingoccurs primarily between intergroup rivals, rather thanwithin groups. In accord with this hypothesis, Stewartet al. (2002) noted that females primarily use hinterlandlatrines, while males do most boundary marking withfaeces, perhaps further evidence of a greater involvementof males in warding off intruders.However, relations between neighbouring badgers re-
main puzzling. While social hierarchies within badgersocial groups have proven elusive (Macdonald et al.2002a), the existence of neatly tessellating range bordersmarkedwith latrines clearly indicates a degree of exclusivity(Stewart et al. 2002). On the other hand, there are manyobservations of badgers either meeting their neighbours atborder latrines or even during incursions without apparentaggression (e.g. Stewart et al. 2002), which seems moreconsistent with the mechanism of spatial partitioningproposed by the Passive Range Exclusion hypothesis(Stewart et al. 1997a) than with classical territorial defence.Resolving these contradictions probably lies in evaluatingdifferences based on sex, age, status and season; a furtherimportant consideration is thatnot all territorybordersmayhave the same significance; Evans et al. (1989) suggestedthat groupsmay be clustered as socially linked supergroups.Observations of bite wounding in badgers have focused
particularly on the seasonality of their occurrence.Cresswell et al. (1992), using comprehensive data collect-ed from road kills throughout the year, reported peaks inwounding levels in male, but not female, badgers duringthe spring, and suggested that they were associated withreproductive activity. In our logistic model, there was nosignificant seasonal pattern to the incidence of wounding.Cresswell et al. (1992) pointed out that wounding was notassociated with seasons when food was thought to be lessavailable.The patterns of bite wounding observed here contribute
to a picture of badger sociality that is relevant both to thetransmission of bovine tuberculosis (Tuyttens et al. 2001)and to models of badger social behaviour (Johnson et al.2002). These patterns suggest sex-specific functionswithin that society, with different roles in group dynamicsand territorial defence for males and females.
Acknowledgments
Our long-term studies of badgers at Wytham have beenlargely supported by funding to D.W.M. from the Peoples’Trust for Endangered Species, with additional supportfrom the Royal Society and the NERC, to all of which we
are grateful. We are especially grateful to Philip Riordan forhis comments and statistical advice on the manuscript.Many members of the WildCRU have participated in thisstudy, we thank them all warmly and note especially theinput of J. da Silva, R. Woodroffe, P. Stewart, D. Johnson,C. Buesching, X. Domingo-Roura and S. Baker during theperiod in which these data were gathered.
References
Buesching, C. D., Newman, C. & Macdonald, D. W. 2002.
Variations in colour and volume of the subcaudal gland secretion
of badgers (Meles meles) in relation to sex, season and individual
specific parameters. Mammalian Biology, 67, 147e156.
Cheeseman, C. L. & Harris, S. 1982. Methods of marking badgers.
Journal of Zoology, 197, 289e292.
Clifton-Hadley, R. S., Wilesmith, J. W. & Stuart, F. A. 1993.
Mycobacterium bovis in the European badger (Meles meles):epidemiological findings in tuberculous badgers from a naturally
infected population. Epidemiology and Infection, 111, 9e19.
Cresswell, W. J., Harris, S., Cheeseman, C. L. & Mallinson, P. J.1992. To breed or not to breed: an analysis of the social and
density-dependent constraints on the fecundity of female badgers
(Meles meles). Philosophical Transactions of the Royal Society ofLondon, 338, 393e407.
Delahay, R. J., Brown, J. A., Mallinson, P. J., Spyvee, P. D.,Handoll, D., Rogers, L. M. & Cheeseman, C. L. 2000. The use of
marked bait in studies of the territorial organisation of the
European badger (Meles meles). Mammal Review, 30, 73e87.
Evans, P. G. H., Macdonald, D. W. & Cheeseman, C. L. 1989.
Social structure of the European badger (Meles meles): genetic
evidence. Journal of Zoology, 218, 587e595.
Feore, S. & Montgomery, W. I. 1999. Habitat effects on the spatial
ecology of the European badger (Meles meles). Journal of Zoology,247, 537e549.
Gallagher, J. & Nelson, J. 1979. Cause of ill health and naturaldeath in badgers in Gloucestershire. Veterinary Record, 105,546e551.
Hunt, P. S. 1976. Anaesthesis of the European badger using
ketamine hydrochloride. Veterinary Record, 98, 94.
Johnson, D. D. P., Kays, R., Blackwell, P. G. & Macdonald, D. W.2002. Does the resource dispersion hypothesis explain group
living? Trends in Ecology and Evolution, 17, 563e570.
Johnson, D. P., Stopka, P. & Macdonald, D. W. In press. Ideal flea
constraints on group living: unwanted public goods and the
emergence of cooperation. Behavioral Ecology.
Keeble, E. 1999. Surgical treatment of a territorial fight wound in
a badger (Meles meles): a novel technique. In: Proceedings of theBVZS Autumn Meeting, Nov 20e21, 1999, pp. 53e54. London:
British Veterinary Zoological Society.
Krebs, J. R., Anderson, R., Clutton-Brock, T., Morrison, I.,Young, D., Donnelly, C., Frost, S. & Woodroffe, R. 1997. BovineTuberculosis in Cattle and Badgers. London: The Ministry of
Agriculture, Fisheries and Food Publications.
Kruuk, H. 1978. Spatial organization and territorial behaviour of the
European badger Meles meles. Journal of Zoology, 184, 1e19.
Kruuk, H. 1989. The Social Badger: Ecology and Behaviour of a Group
Living Carnivore (Meles meles). Oxford: Oxford University Press.
Macdonald, D. W. & Newman, C. 2002. Population dynamics of
badgers (Meles meles) in Oxfordshire, UK: numbers, density and
cohort life histories, and a possible role of climate change inpopulation growth. Journal of Zoology, 256, 121e138.

MACDONALD ET AL.: BITE WOUNDS IN BADGERS 751
Macdonald, D. W., Stewart, P. D., Johnson, P. J., Porkert, J. &Buesching, C. 2002a. No evidence of social hierarchy
amongst feeding badgers, Meles meles. Ethology, 108,613e628.
Macdonald, D. W., Newman, C., Stewart, P. D., Domingo-Roura, X. & Johnson, P. J. 2002b. Density-dependent regulationof body mass and condition in badgers (Meles meles) from
Wytham woods. Ecology, 83, 2056e2061.
Mackintosh, C. G., MacArthur, J. A., Little, T. W. & Stuart, P.1976. The immobilization of the badger (Meles meles). British
Veterinary Journal, 132, 609e620.
Neal, E. & Cheeseman, C. L. 1996. Badgers. London: T. & A. D.
Poyser.
SAS Institute. 1985. SAS Language Guide, Personal Computer, Version
6 Edition. Carey, North Carolina: SAS Institute Inc.
da Silva, J., Macdonald, D. W. & Evans, P. G. H. 1994. Net costs of
group living in a solitary forager, the Eurasian badger. BehavioralEcology, 5, 151e158.
Stewart, P. D. & Macdonald, D. W. 2003. Badgers andbadger fleas: strategies and counter strategies. Ethology, 109,751e764.
Stewart, P. D., Anderson, C. & Macdonald, D. W. 1997a. A
mechanism for passive range exclusion: evidence from the
European badger (Meles meles). Journal of Theoretical Biology,
184, 279e289.
Stewart, P. D., Ellwood, S. A. & Macdonald, D. W. 1997b. Remote
video-surveillance of wildlife: an introduction from experience
with the European badger Meles meles. Mammal Review, 27,185e204.
Stewart, P. D., Macdonald, D. W., Newman, C. & Tattersall, F. H.2002. Behavioural mechanisms of information transmission and
reception by badgers, Meles meles, at latrines. Animal Behaviour,
63, 999e1007.
Thornton, P. D., Newman, C., Johnson, P. J., Buesching, C. D.,Baker, S. E., Slater, D., Johnson, D. D. P. & Macdonald, D. W.In press. Preliminary comparison of four anaesthetic regimensin badgers (Meles meles). Veterinary Anaesthesia and Analgesia.
Tuyttens, F. A. M., Long, B., Fawcett, T., Skinner, A., Brown, J. A.,Cheeseman, C. L., Roddam, A. W. & Macdonald, D. W. 2001.Estimating group size and population density of Eurasian badgers
Meles meles by quantifying latrine use. Journal of Applied Ecology,
38, 1114e1121.
Woodroffe, R. 1993. Alloparental behaviour in the European
badger. Animal Behaviour, 46, 413e415.
Woodroffe, R. & Macdonald, D. W. 1992. Badger clans: de-
mographic groups in an antisocial species. Journal of Zoology, 227,696e698.
Woodroffe, R. & Macdonald, D. W. 1995. Female/femalecompetition in European badgers Meles meles: effects on breeding
success. Journal of Animal Ecology, 64, 12e20.