increased plasma cholinesterase activity succinyl- choline
TRANSCRIPT

Journal of Clinical InvestigationVol. 45, No. 3, 1966
Increased Plasma Cholinesterase Activity and Succinyl-choline Resistance: A Genetic Variant *
HARVEYW. NEITLICH t(From the Clinical Investigation Branch, Clinical Research Division, Edgewood Arsenal, Md.)
Quantitative variations of plasma cholinesteraseactivity (also called serum cholinesterase, pseudo-cholinesterase, nonspecific cholinesterase, or cho-linesterase II) are associated with several diseases.Slightly elevated levels have been observed in thy-rotoxicosis (1), schizophrenia (2), hypertension(3), acute emotional disorders, and after concus-sion (4). Markedly elevated levels, two to threetimes normal, occur with the nephrotic syndrome(5). Plasma cholinesterase activity is decreasedin various disorders of the liver (6), in carcinoma(7), and after anticholinesterase drugs (8).
Low plasma cholinesterase activity may signifythe presence of a qualitatively different enzyme.Some surgical patients develop prolonged apneaafter succinylcholine, a muscle relaxant normallyhydrolyzed rapidly by cholinesterase; their plasmahydrolyzes cholinesterase substrates more slowlythan usual (9). Several cholinesterase inhibitorssuch as dibucaine, physostigmine, and hexametho-nium are weak inhibitors of the atypical cholines-terase from these patients (10). Under standard-ized conditions, dibucaine inhibits the usual en-zyme more than 70%o, but the atypical enzyme isinhibited only 20%o (11). The per cent inhibitionby dibucaine has become known as the dibucainenumber. People with dibucaine numbers of about20 are very sensitive to succinylcholine.
Kalow and Staron determined the dibucainenumbers of 1,556 healthy and mentally ill subjects(12) and demonstrated that two allelic, codomi-nant, autosomal genes control the presence of theusual and atypical cholinesterase. Ninety-sevenper cent of the population have dibucaine num-bers higher than 70 and are homozygous for theusual cholinesterase; 0.03%o have dibucaine num-
* Submitted for publication September 3, 1965; ac-cepted December 2, 1965.
tAssistant resident in medicine, University Hospital,Boston, Mass. Address requests for reprints to Dr.Harvey W. Neitlich, 3 Pama Gardens, Apt. 3, Brighton,Mass. 02135.
bers around 20 and are homozygous for the atypi-cal enzyme; 3%o are intermediate and have mix-tures of the two enzymes.
Kalow's studies of the genetics of plasma cholin-esterase inheritance have been expanded, and twofurther alleles have been reported. One is a "si-lent" gene that results in complete absence of cho-linesterase activity in the homozygote (13); an-other cholinesterase genotype is characterized bya normal or slightly low dibucaine number but aresistance to sodium fluoride at standard condi-tions (14).
Harris, Hopkinson, Robson, and Whittaker de-tected a further phenotype that can only be demon-strated electrophoretically (15). Family studiessuggested that the extra band they reported is dueto a gene that is nonallelic to the previously de-scribed genes (16).
Recently, a normal man was found to have aplasma cholinesterase activity three times higherthan usual. The inheritance, characterization, andsignificance of this cholinesterase form the sub-ject of this report.
MethodsSubjects. The propositus is a 26-year-old male with
normal physical and psychiatric examinations. He hashad the usual childhood diseases, but has no history ofsevere illnesses and no operations. He is allergic topenicillin. Laboratory studies, including complete bloodcount, urinalysis, blood urea nitrogen, sulfobromophtha-lein retention, alkaline phosphatase, serum glutamic ox-alacetic and pyruvic transaminases, cholesterol, protein-bound iodine, serum protein electrophoresis, electrocardio-gram, electroencephalogram, and chest X-ray, werenormal.
His relatives were examined, and blood was obtainedfor evaluation. They had no history of mental illness orphysical abnormalities.
Plasma cholinesterase activity was determined on1,029 military personnel between the ages of 17 and 35(average, 22). The subjects selected as controls for thetests described in this paper had normal plasma cholin-esterase activities and normal dibucaine and fluoridenumbers.
380
INCREASEDPLASMACHOLINESTERASEACTIVITY
Experimental procedures. Diisopropyl fluorophosphate(DFP),1 an irreversible noncompetitive inhibitor mainlyof plasma cholinesterase, was administered intramuscu-larly in graded doses to the propositus and seven normalvolunteers. DFP was diluted in saline and used immedi-ately, since its half-life in solution is 16 hours. Thetotal dose was 8 to 12 sAg per kg divided into three or fourinjections. Each injection was separated by 45 to 60minutes.
A continuous recording of the plasma cholinesteraseactivity of the propositus and one control subject was ob-tained to insure that the time interval between injectionswould be sufficient for the cholinesterase activity to sta-bilize. Intermittent blood samples were also drawn fromthe propositus and seven control subjects for cholines-terase activity assay before each injection and 1 hour afterthe last injection. After the last assay, an excess ofDFP (0.15 mg, intramuscularly, per man) was given toinactivate the plasma cholinesterase activity completely.Blood was then drawn daily to measure the rate of cho-linesterase regeneration.
Succinylcholine2 (0.1 to 0.2 mg per kg) was givenintravenously to the propositus and five normal volun-teers to determine the pharmacological influence of ele-vated cholinesterase activity. The injection lasted 5 sec-
1 Merck Sharp and Dohme, Philadelphia, Pa.2Abbott Laboratories, North Chicago, Ill.
110 -
100 -
60 -
50s
40 '
30 .
20 -
10 '
LI Y vr °~- i- V-
` 9'O @& A
onds. Grip strength was measured every 20 seconds witha Collins hand dynamometer until the strength returnedto control levels. Oxygen saturation was measured con-tinuously with a Waters ear oximeter. This experi-ment was conducted three times on each subject on suc-cessive days.
Laboratory methods. Individual blood samples weredrawn from the antecubital vein and mixed with heparin[02 ml heparin (1,000 U.S.P. U per ml) per 8 ml blood].Plasma was obtained immediately by centrifugation. Thedeterminations were usually performed the same day.When this was not possible, the plasma was refrigeratedat 40 C for less than 1 week or frozen. No change inactivity occurred with these storage procedures.
The plasma cholinesterase activity of the volunteerswas measured at least once by an automated procedureusing De la Huerga, Yesenick, and Popper's modificationof Hestrin's colorimetric technique (17). Undilutedplasma and acetylcholine (0.05 M) were used as reac-tants. Results are expressed in micromoles of acetylcho-line hydrolyzed per hour by 1 ml of plasma at 370 C.
Another method for cholinesterase assay was the ultra-violet spectrophotometric determination described byKalow and Lindsay (18). Plasma, diluted 1 to 50, wasused, in contrast to the previous methods, which requiredundiluted plasma. Benzoylcholine (2 X 10' M) wasused as the substrate. The dibucaine numbers and fluo-
Propositus
ss-'D To -o Ho 0 0 ° 0 0 0 0 o "o .o 'o, Y, vo o, 'o 'o 'o
9 0 .0 .0.09 099 .0 .0 .0 .0.>9 0 0 0 .0 .0H0 0 .0 .0' '9
PLASMACHOLINESTERASEACTIVITY
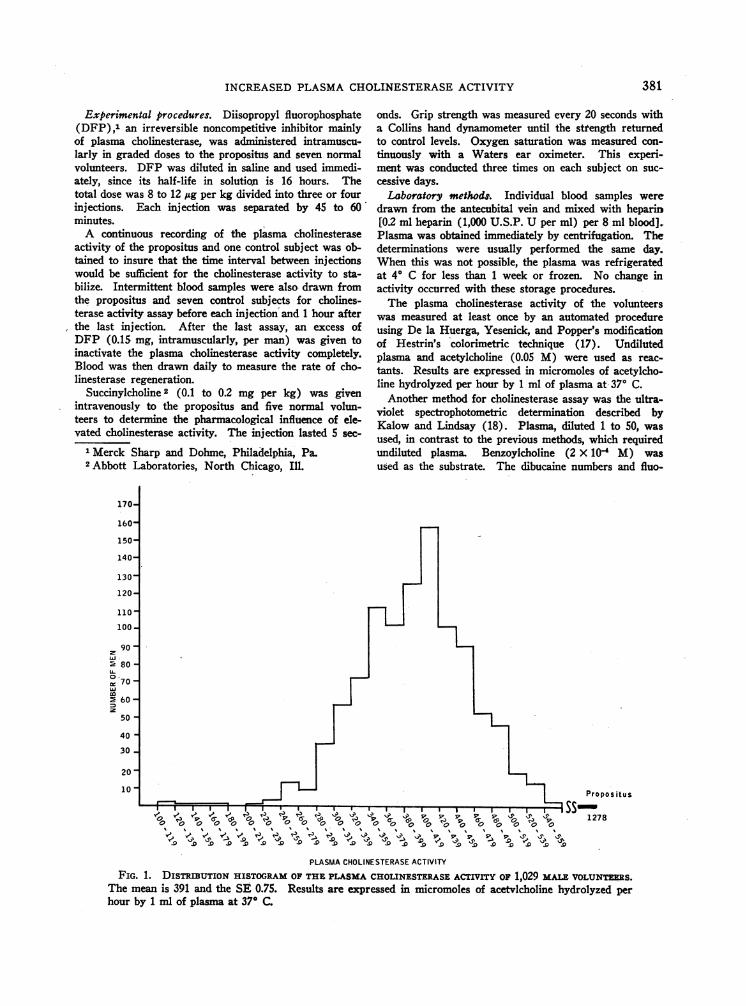
FIG. 1. DISTRIBUTION HISTOGRAMOF THEPLASMACHOLINESTERASEACTIVITY OF 1,029 MALEvoLUNTERS.The mean is 391 and the SE 0.75. Results are expressed in micromoles of acetvlcholine hydrolyzed perhour by 1 ml of plasma at 37° C.
381

382 HARVEYW. NEITLICH
KEYMALE FEMALE
Lo INCF
@ADDI
El . .:BAN[*. NOT
L)I(5 6) (409)
REASEDPSEUDOCHOLIN-rERASE1ITIONAL ELECTROPHORETICID
'TESTED
(432) ~~~~~(271) (1278) (322)(4-28 32) (1518). % 7) 2
(419) (428)(1237) (446)
FIG. 2. FAMILY DISTRIBUTION OF INCREASED PLASMA CHOLINESTERASEACTIVITY AND ADDITIONAL CHO-LINESTERASE ELECTROPHORETICBAND. The figures in parentheses indicate the cholinesterase activity; thearrow indicates the propositus: Note that each family member who has an increased cholinesterase ac-tivity also demonstrates an additional cholinesterase band with disc electrophoresis.
ride numbers were determined for the propositus, hisfamily, and the control subjects (11, 14).
For the continuous monitoring of cholinesterase activityduring the studies with DFP, 1 ml of blood per minutewas drawn through an iv catheter leading from the ante-cubital vein to a multichannel analytic system that pref-,erentially measured plasma cholinesterase since butyryl-choline was used as substrate (19).
Unidimensional disc electrophoresis was performed ac-
cording to the method of Ornstein (20). Acrylamide gel(Canalco) was prepared with Tris-glycine buffer (pH8.6). The material was stained for cholinesterase witha-naphthylacetate as substrate and fast blue B diazocoupler (21).
Results
A distribution histogram of the plasma cholin-esterase activity in 1,029 male volunteers isshown in Figure 1. The mean was 391 and theSE 0.75. The mean was higher than Kalow's se-
ries because of the younger age and male sex ofthis group (22); the distribution was similar.
The propositus's activity, 1,278 U, was more
than three times higher than the mean for the vol-unteers. Since no disease could be found to ex-
plain this elevation and since it remained constantfor 6 months, his family was studied to determinewhether the elevated cholinesterase activity mightbe familial. The family's cholinesterase activities
are shown in Figure 2. The propositus's sisterand daughter had very high enzyme activities,1,518 and 1,237 U, respectively. His mother'scholinesterase activity, 566 U, was not so strikinglyelevated, but was, nevertheless, higher than theother volunteers. Dibucaine numbers and fluoridenumbers were normal in all members of thefamily.
The in vivo sensitivity of the propositus's cho-linesterase activity was tested with DFP. Thisanticholinesterase was chosen because it reactsirreversibly and stoichiometrically with plasmacholinesterase (23). Therefore, a standardizeddose should inhibit a constant amount of plasmacholinesterase activity. This relationship shouldbe independent of the original concentration ofthe enzyme. The results are shown in Figure 3.The propositus's plasma cholinesterase activitywas three to four times more sensitive to DFPthan the controls'. DFP, 1 ug per kg intramus-cularly, inhibited the propositus's plasma cholin-esterase activity 93 U. The same dose inhibitedthe controls' cholinesterase activity by a mean of25 U. After a total dose of 12 pg per kg, thepropositus's cholinesterase activity was 124 U, andthe mean of the controls was 66 U. The equationsin Figure 3 show that 13 to 15, pg per kg would
382

INCREASEDPLASMACHOLINESTERASEACTIVITY 383
have completely inhibited the cholinesterase ac- 1278Ltivity of the propositus and the controls. The per 1200icent of original cholinesterase activity inhibited 1150Io
by DFP was similar. Each microgram per kilo- 100 POPOSTUS1050- R=~~-sopoi 99
X=Actual Vakiesgram inhibited the cholinesterase activity approxi- I, ,ooo 2i142+-(93.2xmately 7% in the propositus and in the control 2 950 *- R=-.97
subjects. -~~~~~~~~~~~~~~~~~~~~~~~~~00=803.17'+(-100)xsubj ects. ~ \ - CONTROLS(7)
This experiment was repeated 15 days later, af- 1f 800 'eiimoeachsde offiter the propositus had regenerated 70%7o of his 750 . 364+ -24.8x
original cholinesterase activity. Again, his enzyme - 700- ~ 650activity was as sensitive to DFP as previously ¢ 600
(Figure 3). E 550
There is strong evidence that the regeneration of 5cholinesterase activity after inactivation by DFPrepresents resynthesis of new enzymes by the liver X 350-i(8). The comparative rates of cholinesterase ac- 300tivity regeneration of three controls and the pro- 250positus are shown in Figure 4. The per cent oforiginal activity regenerated daily was similar for 100
the controls and propositus. After 4 days, the so
propositus had regenerated 27% of his cholines- 23 4 56 7 8 9 10 11 12terase activity; the controls, 40%o. After 15 days, DOSE(1ICROGRAMS/KG)they had regenerated 70%o and 80%o, respectively. FIG. 3. THE INHIBITION OF PLASMA CHOLINESTERASE
ACTIVITY AFTER ADMINISTRATION OF DIISOPROPYL FLUORO-There was no statistical difference between the tVuro PHOSPHATE. R is the correlation coefficient; y is theslopes. equation for the line.
100- - CONTROLSy=26.58 + 3.33 xXj
1 0 0
R = + 0.90Z PROPOSITUSy=11.26 + 3.92 x_ R= + 0.96
o 80<St
(2
2 4 6 0 2 14 1
660
40.
0:
W 40 4
202 I
TIME ( DAYS
FIG. 4. RATE OF PLASMACHOLINESTERASEREGENERATION (PER CENT OF ORIGINAL ACTIVITY).'The shaded area represents 1 SE on each side of the line. There is no statistical difference be-tween the two slopes.

HARVEYW. NEITLICH
= STO-RAE BANDSGADDITIONAL SAND
04
C3
*: ~~~~~~~~~~~~~~..
- ~~~~~ ~ ~--------:2:
ALBUMINESTERASE
FIG. 5. SERUMDISC ELECTROPHORESESSTAINED FOR CHOLINESTERASE. The propositus's additional cholin-esterase band is thicker and more intense than his mother's.
The results of the disc electrophoreses stainedfor cholinesterase are shown in Figure 5. Thetwo pale bands closest to the origin represent stor-age bands. Harris and associates believe that theymay be derived from the more intense C4 bandafter a few days' storage (15). They were notpresent when fresh specimens were used. Eachsample demonstrated three light bands between therapidly migrating albumin esterase and the slowerC4 band. These three bands, called C1, C2, and C3,rapidly faded after the staining procedure. It hasbeen suggested that the C4 band contributes thegreatest proportion of the cholinesterase activity inindividuals with the usual cholinesterase genotype.
The propositus and each member of his familywith increased cholinesterase activity had a denselystaining additional band. This band was adjacentto the C4 band, but was slightly closer to the origin.It was present in fresh and stored samples fromthe propositus. This band was not observed inthe electrophoretic patterns of the members of thefamily with normal cholinesterase activity, nor wasit present in the plasma obtained from numerousvolunteers.
The staining characteristics of the additionalband suggested that it contributed more to the totalcholinesterase activity than the other bands. Inthree of the four affected family members, the ex-tra band stained more intensely and diffusely thanthe C4 band and was much darker than C1, C2, orC3. The mother's additional band was slightlyless intense than the C4 band. This might be re-lated to her lower cholinesterase activity relativeto the other three.
The propositus's cholinesterase activity sug-gested that he might have an altered response tosuccinylcholine. His change in grip strength afteriv administration of succinylcholine was measuredand compared to that of five controls. Results areshown in Figure 6. One-tenth mg per kg affectedthe controls in 20 seconds. Grip strength, asmeasured by a hand dynamometer, decreased rap-idly and then gradually recovered in the next 2 to3 minutes. Oxygen saturation did not change, andthere were no residual effects.
The propositus's strength was not affected bythe same dose. He was then given twice the dosethat the controls received with only a slight and
384

INCREASEDPLASMACHOLINESTERASEACTIVITY
transient effect. Compared to the control subjects,the propositus was resistant to succinylcholine.
Discussion
There are two types of cholinesterase. Thesecan be differentiated by substrate specificity, ana-tomic distribution, and biological function; theircharacteristics have been reviewed recently (24).Plasma cholinesterase hydrolyzes propionylcholineand butyrylcholine more rapidly than acetylcholine.Acetylcholinesterase (specific or true cholines-terase, cholinesterase I) hydrolyzes acetylcholinemost rapidly and negligibly hydrolyzes butyryl-choline. Plasma cholinesterase is produced by theliver, and its main source is serum or plasma. Al-though the biological function of plasma cholines-terase is unknown, it is important in limiting theresponses to hydrolyzable drugs such as succinyl-choline. Acetylcholinesterase on the other hand,is found in the nervous system and red blood cellsand is important in nerve impulse transmission.There are no biological or genetic similarities be-tween the two types. The propositus's acetylcho-linesterase activity was normal, but his plasmacholinesterase activity was very high.
Increased plasma cholinesterase activity is veryrare. Kalow and his colleagues noted one indi-vidual with 2.5 times the average enzyme activityin a study of 1,556 subjects (11, 12). The sub-ject had no evidence of disease and had a normaldibucaine number. No mention of family studieswas made. If Kalow's group and the Army vol-unteers are combined, the prevalence of elevatedcholinesterase activity more than 2.5 times themean in normal individuals is approximately 2per 2,600. The presence of this unusual charac-teristic/in three members of the propositus's fam-ily supports the contention that the level is fa-milial and not acquired. The family distributionof the increased cholinesterase and the additionalelectrophoretic band suggest that inheritance is au-tosomal dominant and that each member with in-creased activity is heterozygous for the phenotype,but X-linked dominant inheritance cannot be ruledout.
The elevated cholinesterase activities found inthis family may be explained by increased enzymeproduction, slower catabolism of the enzyme, or amolecular alteration that resulted in a more activeenzyme. The response of the propositus's cho-linesterase to DFP suggests that the last hypothe-
X---X PROPOSITUSO.1Mgm/Kg
0a . PROPOSITUSO.2 Mgm/ Kg
CONTROLS0.1 Mgm/ KgMEANVALUE OF 5 CONTROLS.
100
90
80
c 70w
v 60.s
*; 50.r
0 40
- 30
Z 20
10
20 40 60 S0 100 120 140 160 180 200 220TIME ( Seconds )
FIG. 6. MUSCLESTRENGTHAFTER IV SUCCINYLCHOLINE. The shaded area represents 1 SEon each side of the line. The propositus is relatively resistant to succinylcholine.
385

HARVEYW. NEITLICH
sisvis.correct. DFP is a potent anticholinesterase.Ithashbeen'-wreported to inhibit the usual cholines-terase and the atypical enzyme described by Kalowand, Davies. in a 1: 1 ratio (10). Therefore, thepropositus's cholinesterase would be unlikely to bemore sensitive to, DFP than the usual cholines-terase. The more probable explanation for the re-
sults depicted in Figure 3 is that the propositushas a normal number of cholinesterase molecules,but some enzyme molecules are three to four timesmore active than usual. This would account forthe similar dose of DFP required to inhibit com-
pletely the cholinesterase activity of the propositusand the controls. Also, this theory would explainthe results in which the per cent of cholinesteraseactivity inhibited and the per cent regenerated af-ter DFPwere similar in the controls and the pro-positus despite very different enzyme activities.
The electrophoretic patterns of plasma cholines-terase further substantiate the hypothesis that thepropositus has an additional enzyme that is dif-ferent from the usual cholinesterase. The extraband that the propositus and affected family mem-
bers possess migrated more slowly than the C4band. Using different electrophoretic techniques,Harris and associates described a band that alsomigrated more slowly -than C4, which he named C,.This band was reported to be present in 10% ofa random sample of the British population (15).However, the cholinesterase activity found in in-dividuals with this phenotype, although slightlyhigher than usual, was much lower than for thesubjects of the present report. Also, these indi-viduals were as sensitive to succinylcholine as
those with the usual cholinesterase genotype.There are numerous examples to illustrate the
relationship between genetic factors and the actionof drugs (25). Drugs may exacerbate several in-herited diseases. For example, an attack of acuteintermittent porphyria may follow the ingestionof barbiturates. Drugs may detect genetic poly-morphisms. Primaquine sensitivity is due toinherited glucose-6-phosphate dehydrogenase de-ficiency. The ability to taste phenylthiourea isgenetically controlled, and this may be related tovarious thyroid disorders.
The plasma cholinesterase polymorphism was
discovered after several surgical patients were
found to be unusually sensitive to the muscle re-
laxant, succinylcholine. Enzyme kinetics and fam-
ily studies suggested that their phenotype dif-fered from the usual one for cholinesterase. Thepropositus of the present report probably is aheterozygote, with one allele leading to the produc-tion of a more active enzyme. The latter enzymeseems to provide increased resistance to succinyl-choline.
Summary
Plasma cholinesterase activity has been deter-mined in 1,029 men. Values ranged from 92 to1,278 U with a mean of 391 U. One subject wasfound to have a consistently elevated plasma cho-linesterase activity. Examination of his familyrevealed that this cholinesterase was probably in-herited as an autosomal dominant. In vitro andin vivo studies suggest that the propositus isheterozygous for an enzyme molecule that is morethan three times as active as the usual cholines-terase.
Acknowledgments
The author is grateful to Dr. L. A. Mounter for hisexpert advice and assistance, to Miss Mary Grainger forinvaluable technical assistance in performing the electro-phoretic experiments, to Mr. W. A. Groff for the cho-linesterase determinations, and to Col. N. Bottiglieri,MC, Chief, Clinical Research Division, Edgewood Ar-senal, Md. Capt. David A. Drew, MC, collaborated inthe succinylcholine experiments, which will be publishedin greater detail.
References1. Antopol, W., L. Tuchman, and A. Schifrin. Choline-
esterase activity of human sera, with special refer-ence to hyperthyroidism. Proc. Soc. exp. Biol.(N. Y.) 1937, 36, 46.
2. Gal, E. M. Cholinesterase activity of whole bloodfrom healthy and schizophrenic individuals. Na-ture (Lond.) 1963, 198, 1118.
3. Reinfrank, R. F., and W. J. Wetstone. Serum cho-linesterase activity in hypertension, report of apilot study. Hartford Hosp. Bull. 1958, 13, 32.
4. Richter, D., and M. Lee. Serum choline-esterase anddepression. J. ment. Sci. 1942, 88, 435.
5. Vorhaus, L. J., and R. M. Kark. Serum cholin-esterase in health and disease. Amer. J. Med. 1953,14, 707.
6. Vorhaus, L. J., II, H. H. Scudamore, and R. M.Kark. Measurement of serum cholinesterase ac-tivity in the study of diseases of the liver andbiliary system. Gastroenterology 1950, 15, 304.
7. Wetstone, H. J., R. V. LaMotta, A. Bellucci, R.Tennant, and B. V. White. Studies of cholin-esterase activity. V. Serum cholinesterase in pa-
386

INCREASEDPLASMACHOLINESTERASEACTIVITY
tients with carcinoma. Ann. intern. Med. 1960, 52,102.
8. Grob, D., J. L. Lilienthal, Jr., A. M. Harvey, andB. F. Jones. The administration of di-isopropylfluorophosphate (DFP) to man. I. Effect ofplasma and erythrocyte cholinesterase; generalsystemic effects; use in study of hepatic functionand erythropoiesis; and some properties of plasmacholinesterase. Bull. Johns Hopk. Hosp. 1947,81, 217.
9. Davies, R. O., A. V. Marton, and W. Kalow. Theaction of normal and atypical cholinesterase ofhuman serum upon a series of eaters of choline.Canad. J. Biochem. 1960, 38, 545.
10. Kalow, W., and R. 0. Davies. The activity of vari-ous esterase inhibitors towards atypical humanserum cholinesterase. Biochem. Pharmacol. 1959,1, 183.
11. Kalow, W., and K. Genest. A method for the de-tection of atypical forms of human serum cholin-esterase. Determination of dibucaine numbers.Canad. J. Biochem. 1957, 35, 339.
12. Kalow, W., and N. Staron. On distribution andinheritance of atypical forms of human serum cho-linesterase, as indicated by dibucaine numbers.Canad. J. Biochem. 1957, 35, 1305.
13. Liddell, J., H. Lehmann, and E. Silk. A "silent"pseudocholinesterase gene. Nature (Lond.) 1962,193, 561.
14. Harris, H., and M. Whittaker. Differential inhibi-tion of human serum cholinesterase with fluoride:recognition of two new phenotypes. Nature(Lond.) 1961, 191, 496.
15. Harris, H., D. A. Hopkinson, E. B. Robson, and M.Whittaker. Genetical studies on a new variant ofserum cholinesterase detected by electrophoresis.Ann. hum. Genet. 1963, 26, 359.
16. Harris, H., E. B. Robson, A. M. Glen-Bott, and J. A.Thornton. Evidence for non-allelism betweengenes affecting serum cholinesterase. Nature(Lond.) 1963, 200, 1185.
17. De la Huerga, J., C. Yesenick, and H. Popper. Color-imetric method for the determination of serumcholinesterase. Amer. J. clin. Path. 1952, 22, 1126.
18. Kalow, W., and H. A. Lindsay. A comparison ofoptical and manometric methods for the assay ofhuman serum cholinesterase. Canad. J. Biochem.1955, 33, 568.
19. Mounter, L. A., W. A. Groff, and V. M. Sim. Amulti-channel analytical system for continuousmonitoring of blood cholinesterase. To be pub-lished.
20. Ornstein, L. Disc electrophoresis. I. Backgroundand theory. Ann. N. Y. Acad. Sci. 1964, 121, 321.
21. Harris, H., D. A. Hopkinson, and E. B. Robson.Two-dimensional electrophoresis of pseudocholin-esterase components in normal human serum. Na-ture (Lond.) 1962, 196, 1296.
22. Shanor, S. P., G. R. Van Hees, N. Baart, E. G.Erdos, and F. F. Foldes. The influence of ageand sex on human plasma and red cell cholin-esterase. Amer. J. med. Sci. 1961, 242, 357.
23. Jansen, E. F., R. Jang, and A. K. Balls. The in-hibition of purified, human plasma cholinesterasewith diisopropyl fluorophosphate. J. biol. Chem.1952, 196, 247.
24. Augustinsson, K. Classification and comparativeenzymology of the cholinesterases and methods fortheir determination in Handbuch der experimen-tellen -Pharmakologie; Cholinesterase and Anti-cholinesterase Agents, G. B. Koelle, Subed. Ber-lin, Springer-Verlag, 1963, chap. 4.
25. Porter, I. H. Genetic basis of drug metabolism inman. Toxicol. appl. Pharmacol. 1964, 6, 499.
ERRATUM
In the paper entitled, "Gastric Secretion in Relation to Mucosal BloodFlow Studied by a Clearance Technic," by Eugene D. Jacobson, Ray H.Linford, and Morton I. Grossman, published in the January issue, line 10in the second column of page 11 is in error. Beginning with line 6, thetext should read as follows: "If the extraction ratio for aminopyrine variedgreatly with different experimental conditions, then the clearance of amino-pyrine would not be a valid method for measuring gastric mucosal bloodflow. In acute experiments we found that the gastric extraction of amino-pyrine was 58%o."
387