increased dihydroceramide/ceramide ratio mediated...
TRANSCRIPT

Nuria Barbarroja12 Sergio Rodriguez-Cuenca1 Heli Nygren3 Antonio Camargo14
Ana Pirraco15 Joana Relat6 Irene Cuadrado7 Vanessa Pellegrinelli1
Gema Medina-Gomez1 Chary Lopez-Pedrera2 Francisco J Tinahones89
J David Symons1011 Scott A Summers12 Matej Oresic313 and Antonio Vidal-Puig114
Increased DihydroceramideCeramide RatioMediated by Defective Expression of degs1Impairs Adipocyte Differentiation and FunctionDiabetes 2015641180ndash1192 | DOI 102337db14-0359
Adipose tissue dysfunction is an important determinantof obesity-associated lipid-induced metabolic compli-cations Ceramides are well-known mediators of lipid-induced insulin resistance in peripheral organs such asmuscle DEGS1 is the desaturase catalyzing the last stepin the main ceramide biosynthetic pathway Functionalsuppression of DEGS1 activity results in substantialchanges in ceramide species likely to affect fundamentalbiological functions such as oxidative stress cell sur-vival and proliferation Here we show that degs1 ex-pression is specifically decreased in the adipose tissueof obese patients and murine models of genetic and nu-tritional obesity Moreover loss-of-function experimentsusing pharmacological or genetic ablation of DEGS1 inpreadipocytes prevented adipogenesis and decreasedlipid accumulation This was associated with elevatedoxidative stress cellular death and blockage of the cellcycle These effects were coupled with increased dihy-droceramide content Finally we validated in vivo thatpharmacological inhibition of DEGS1 impairs adipocyte
differentiation These data identify DEGS1 as a newpotential target to restore adipose tissue function andprevent obesity-associated metabolic disturbances
Dysfunction of white adipose tissue (WAT) and impaireddifferentiation of new adipocytes may lead to lipid leakageand inappropriate accumulation of ectopic lipids inperipheral organs causing lipotoxicity and the metabolicsyndrome (1) The toxic effects of lipids are determinedby both their quantity and their qualitative characteristics(2) Whereas it is well documented that specific species ofsphingolipids and ceramides (Cers) mediate lipotoxicity inliver and muscle (34) the contribution of specific lipotoxicspecies to WAT dysfunction in the context of obesity is stillnot well defined It is known that in rodent WAT Cersincrease in response to a high-fat diet (HFD) (56) concom-itantly with the onset of insulin resistance (6) Lipid anal-ysis of WAT from human obese subjects has produced
1Metabolic Research Laboratories Wellcome Trust-MRC Institute of MetabolicScience Addenbrookersquos Hospital University of Cambridge Cambridge UK2Instituto Maimoacutenides de Investigacioacuten Biomeacutedica de Coacuterdoba Reina SofiaUniversity Hospital Coacuterdoba Spain3VTT Technical Research Centre of Finland Espoo Finland4Lipids and Atherosclerosis Research Unit Instituto Maimoacutenides de Investiga-cioacuten Biomeacutedica de Coacuterdoba Reina Sofia University Hospital Coacuterdoba Spain5Department of Biochemistry (U38-FCT) Faculty of Medicine University ofPorto Porto Portugal6Departament de Bioquiacutemica i Biologia Molecular Universitat de BarcelonaBarcelona Spain7Departamento de Farmacologiacutea Universidad Complutense de Madrid MadridSpain8CIBER in Physiopathology of Obesity and Nutrition (CB0603) Instituto de SaludCarlos III Madrid Spain9Instituto de Investigacioacuten Biomeacutedica de MaacutelagaHospital Virgen de la VictoriaMalaga Spain10College of Health University of Utah Salt Lake City UT11Division of Endocrinology Metabolism and Diabetes University of Utah SaltLake City UT
12Program in Cardiovascular and Metabolic Disorders Duke-National Universityof Singapore Graduate Medical School Singapore Singapore13Steno Diabetes Center Gentofte Denmark14Wellcome Trust Sanger Institute Hinxton UK
Corresponding authors Nur ia Barbarroja nur iabarbarro jaextsjuntadeandaluciaes Sergio Rodriguez-Cuenca sr441medschlcamacukand Antonio Vidal-Puig ajv22medschlcamacuk
Received 1 March 2014 and accepted 20 October 2014
This article contains Supplementary Data online at httpdiabetesdiabetesjournalsorglookupsuppldoi102337db14-0359-DC1
NB and SR-C contributed equally to this work
GM-G is currently affiliated with Departamento de Bioquiacutemica Fisiologiacutea yGeneacutetica Molecular Universidad Rey Juan Carlos Madrid Spain
copy 2015 by the American Diabetes Association Readers may use this article aslong as the work is properly cited the use is educational and not for profit andthe work is not altered
1180 Diabetes Volume 64 April 2015
METABOLISM
conflicting results showing either increased or decreasedCer levels in obese and insulin-resistant patients (78)
Cers can be synthesized from sphingomyelins but themain contributor to their biosynthesis is the de novopathway The final reaction of the de novo pathway iscatalyzed by a D4-dihydroceramide (DhCer) desaturase(DEGS1) that adds a 45-trans-double bound on the sphin-goid base of the DhCer (9) The other enzyme DEGS2catalyzes the synthesis of phytoceramides (10) whoseexpression is restricted to skin intestine and kidney
There is evidence that downregulation of degs1 in-creases the DhCerCer ratio in different cellular modelsAlthough DhCer was considered an inactive precursorof Cers (11) recent studies (1213) have suggested theirrelevance as modulators of cell cycle apoptosis autophagyor oxidative stress processes that a priori are expectedto compromise the development and function of adiposetissue (AT)
Since DEGS1 is the key enzyme regulating the DhCerCer ratio understanding its regulation is important indetermining the pathophysiological relevance of this path-way in AT Of note degs1 has recently been identifiedin genome-wide association studies as a candidate geneassociated with fat mass accumulation in mice (14) fur-ther suggesting that DEGS1 may be relevant for theadaptive accretion of AT Moreover DEGS1 could beconsidered an attractive therapeutic target for obesity-associated insulin resistance since fenretinide has beenclaimed to improve insulin sensitivity by inhibitingDEGS1 (15) although other molecular targets cannotbe ruled out
Here we show that DEGS1 expression is selectivelyperturbed in the WAT of murine models of nutritionaland genetically induced obesity and in the WAT of mor-bidly obese (MO) patients In vitro analysis revealed thatboth pharmacological inhibition and genetic ablation ofDEGS1 result in impaired adipocyte differentiation andlipid accumulation effects mediated by increased DhCercontent In vivo pharmacological inhibition of DEGS1also resulted in impaired adipocyte differentiation De-creased levels of DEGS1 were associated with increasedoxidative stress accelerated cellular death and blockageof cell cycle We also show data supporting that DEGS1expression is regulated by peroxisome proliferatorndashactivated receptor (PPAR)g
RESEARCH DESIGN AND METHODS
Mice HusbandryAnimals were housed at 22ndash24degC with 12-h lightdark cyclesFood and water were available ad libitum Lean wild-type(WT) PPARg222 and PPARg2obob double knockoutmice (16) were used for profiling purposes Mice were feda normal chow diet (D12450B) or an HFD (D12451) fromResearch Diets
For the pharmacological inhibition of DEGS1 10-week-old male C57BL6 WT mice were used Mice were fedan HFD for 5 weeks before C8-cyclopropenylceramide
(C8-CPPC) (Matreya) administration All protocols usedwere approved by the UK Home Office
Intraperitoneal Injection of CyclopropenylceramideMice were distributed in two groups (n = 8) and admin-istered daily vehicle (2-hydroxypropyl betacyclodextrine)or C8-CPPC (2 mgkgday) for 9 days via intraperitonealinjection Mice were culled at the end of the experimentand WAT was removed for gene expression and micros-copy analysis No differences in food intake lean massand fat content were observed during the treatment
Ex Vivo Experiments in Isolated Mature AdipocytesAdipocytes from gonadal WAT of 16-week-old C57BL6 micewere obtained by collagenase type II digestion at 37degC Afterdigestion adipocytes were placed in DMEM with or without1 mmolL C8-CPPC every 5 h for a total period of 20 h
Retroviral Short Hairpin RNA Constructs for DEGS1RNAi-Ready pSIREN-RetroQ vectors (BD Biosciences)were used to target degs1 in 3T3-L1 cells Sequences tar-geting degs1 were ligated into the pSIREN vector as de-scribed in the manufacturerrsquos instructions Retroviruseswere generated by transfecting BOSC cells (AmericanType Culture Collection) with the pSIREN plasmids usingFuGene6 (Roche) Supernatant with the viral content wasused to transfect 3T3-L1 preadipocytes Twenty-fourhours after retroviral infection the cells were selectedwith puromycin (4 mgmL)
Culture Differentiation and Treatment of 3T3-L1PreadipocytesCells were differentiated into adipocytes (day 9) accord-ingly to the protocol described by Roberts et al (17) withor without rosiglitazone 01 mmolL Lipid accumulationwas assessed by Oil Red O solution (18)
Pharmacological Inhibition of DEGS1 Activity In Vitro
Effects on DifferentiationAt day 0 of differentiation 3T3-L1 cells were treated withC8-CPPC 1 mmolL andor rosiglitazone 01 mmolL for 96 h
Effects on LipolysisOn day 8 of differentiation after 48 h of treatment withC8-CPPC 1 mmolL the cells were exposed to norepineph-rine (NE) (1028 and 1027 molL) for 6 h
Effects on Insulin SignalingOn day 8 of differentiation after 48 h of treatment withC8-CPPC 1 mmolL the cells were exposed to insulin (10and 100 nmolL) for 15 min
Effects on GLUT4 and AdiponectinOn day 8 of differentiation after 48 h of treatment withC8-CPPC 05ndash1 mmolL the cells were exposed to insulin
Effects on AMPKOn day 8 of differentiation after 48 h of treatment withC8-CPPC 05ndash1 mmolL the cells were exposed to insulin(10 and 100 nmolL) rosiglitazone (1 and 10 nmolL)and metformin (100 nmolL) for 24 h
diabetesdiabetesjournalsorg Barbarroja and Associates 1181
DhCer Treatment3T3-L1 preadipocytes were treated at day 0 of differentia-tion with the induction cocktail and C2DhCer at 50 mmolLfor 3 days and a second batch of cells were exposed at day 3of differentiation and treated with C2DhCer at 50 mmolLfor 3 days All batches were taken until final differentiationat day 9
Human SamplesThe cohort included 28 MO and 6 nonobese subjects withno alterations to lipid or glucose metabolism as controlsubjects (Table 1) Approval for the study was given by theethics committee and all patients gave their informedconsent Visceral AT biopsy samples were obtained fromMO patients undergoing bariatric surgery (Scopinaro pro-cedure) or laparoscopic surgery (hiatus hernia repair orcholecystectomies) for the lean subjects
Western BlottingProtein extracts were prepared using the Nuclear andCytoplasmic Extraction Reagents Kit (Pierce) according tothe manufacturerrsquos instructions Immunoblots were incu-bated with the following antibodies PPARg CEBPb andCyclinA (Santa Cruz Biotechnology) CyclinB1 D1 D3 andE1 cdk4 AKT Ser473-pAKT p4442 MAPK phospho-p4442 MAPK Ser660-pHSL (p-hormone-sensitive lipase)Ser565-pHSL total HSL ATGL caveolin-1 plin1 glut4 adi-ponectin and AMPK (Cell Signaling Technology) and adfpadrp abhd5 and anti-b-actin (Abcam)
RT-PCRRNA was extracted using TRI Reagent (Sigma) and reversetranscribed to cDNA Real-time PCR using SYBRgreenwas performed according to the manufacturerrsquos instruc-tions (ABI) Primer sequences were obtained from PrimerBlast (19) Expression of genes was corrected by the
geometrical average of 18s b2m b-act and 36b4 usingBestkeeper (20)
Apoptosis Combined Annexin VPropidium IodideStainingViability was assessed by using an Annexin Vpropidiumiodide kit (Bender MedSystems) according to the manufac-turerrsquos recommendations Binding of fluorescein-conjugatedAnnexin V and propidium iodide was measured by FACSCalibur(BD Biosciences)
Cellular ProliferationCell viability was assessed using an colorimetric assay(Roche) following the protocol supplied by the manufac-turer Cell proliferation was analyzed by quantification ofthe incorporation of BrdU (Roche)
Boron-Dipyrromethene Staining Reactive OxygenStress Production and Mitochondrial ContentCells were incubated with boron-dipyrromethene (BODIPY)at 4degC or alternatively with 20 mmolL 2979-dichlorodihy-drofluorescein diacetate or 100 nmolL Mitotracker (Invi-trogen) at 37degC for 30 min and analyzed on a FACSCaliburcell analyzer
Cellular Oxygen ConsumptionCells were exposed to oligomycin (1 mmolL) C8-CPPC(09 mmolL) and antimycinrotenone (1 mmolL each)O2 consumption was measured using the XF24 analyzer(Seahorse Bioscience) for a period of 90 min
Whole-Mount Confocal MicroscopyGonadal AT was fixed in 4 paraformaldehyde Sampleswere incubated at 4degC with mouse Pref-1 (preadipocytefactor-1) or rabbit Ki67 primary antibodies Nuclei andneutral lipids were stained with Hoechst 33342 and BODIPY493503 respectively Sample examination was performedusing a Zeiss 510 confocal laser scanning microscope (CarlZeiss) Pref-1+ Ki67+ and BODIPY+ cells and total cells (upto 913 cellsfield) were automatically counted by usingImageJ software in three independent fields in each tissue
Light Microscopy AnalysisSamples for AT hematoxylin-eosin staining were preparedas described previously (16) Adipocyte sizes were mea-sured using Cell P (Olympus Soft Imaging SolutionsGmbH) Between 1000 and 3000 adipocytes from eachmouse were used to obtain the mean cell area
Luciferase Reporter AssayHuman embryonic kidney 293 cells were transfectedusing Lipofectamine LTX (Invitrogen) following themanufacturerrsquos instructions Seventy-five nanograms ofthe reporter plasmid (3xPPRE TKLuc) and 375 ng of theeukaryotic expression vector (pSV-PPARg) were cotrans-fected to each well The plasmid pRL-CMV (5 ngwell)was included as an internal control Twenty-four hourspost-transfection cells were treated as indicated (DMSOrosiglitazone 10 mmolL GW1929 10 mmolL andC2DhCer C16DhCer and Cer 100 mmolL) Luciferaseassays were performed using the dual luciferase reporterassay system (Promega)
Table 1mdashClinical characteristics of control subjects and MOpatients
Control subjects MO patients
Sex nMale 3 14Female 3 14
Age years 5217 6 51 4157 6 296
Weight kg 7400 6 071 1555 6 732
Height cm 16260 6 160 16553 6 249
BMI kgm2 2504 6 055 5651 6 168
Serum insulin IUmL 1123 6 158 3009 6 230
HOMA-IR 351 6 048 818 6 078
Serum glucose mmolL 603 6 032 574 6 026
Serum cholesterol mmolL 468 6 040 492 6 025
HDL cholesterol mmolL 097 6 023 110 6 009
Triglycerides mmolL 108 6 020 142 6 019
Values are reported as the mean 6 SEM unless otherwise in-dicated HOMA-IR HOMA of insulin resistance
1182 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
LipidomicsCells were mixed with 09 NaCl and sonicated for 5 minat 5degC and 40 kHz Samples were spiked with internalstandard The samples were extracted with chloroformmethanol (21) The lower phase was collected and mixedwith the labeled standard mixture (three stable isotope-labeled reference compounds) Lipid extracts were ana-lyzed on a Q-ToF Premier Mass Spectrometer (Waters)combined with an Acquity Ultra Performance LiquidChromatographyMass Spectrometry system (Waters)The data were processed using MZmine software Thelipids were quantified by normalizing with correspond-ing internal standard
Statistical AnalysisStudent t test (unpaired) ANOVA and Duncan test wereused for the statistical analysis Statistical significance wasset at P 005 and P 001 The Spearman correlationwas calculated to estimate the linear correlations betweenvariables at P 001
RESULTS
DEGS1 Is Downregulated in WAT in Obese MurineModelsTissue distribution analysis showed that degs1 is presentin most tissues and it is particularly highly expressed inAT liver and muscle whereas degs2 mRNA is detectedonly in intestine (Fig 1A) Degs1 expression in vivo waspositively correlated with WAT mass in lean healthy mice(Fig 1B) However this correlation was disrupted in mu-rine models of nutritional and genetically induced obesity(HFD and obob) where degs1 mRNA expression was de-creased versus controls (Fig 1C and F) Of note the dys-regulation of degs1 in obesity was limited to WAT as itsexpression in liver or skeletal muscle was not affected ineither HFD-fed mice (Fig 1D and E) or obob mice (Fig1G and H) DEGS1 was preferentially expressed in ma-tured adipocytes versus stromal vascular fraction in leanand obob mice (Supplementary Fig 1) and downregulatedin obob adipocytes versus lean adipocytes Moreover thisdownregulation of degs1 in total WAT of HFD-fed and obobmice was also recapitulated in visceral AT of MO patients(Supplementary Fig 2A)
To determine whether the decrease in degs1 expressionwas part of a global adaptation of de novo Cer synthesispathway in obesity the expression of other genes in thispathway was measured Expression of the sptlc1 and sptlc2subunits was not changed in human visceral AT or in obobWAT but sptlc2 expression was increased in HFD WAT(Supplementary Fig 2B and C) Moreover both HFD andobob mouse WAT exhibited a reduction in a subset of Cersynthases suggesting a possible defect in the synthesis ofa specific subset of Cers
We then confirmed that modulators of obesity-associatedinflammation may contribute to the downregulation ofdegs1 as treatment of 3T3-L1 adipocytes with tumornecrosis factor-a (5 and 10 ngmL) for 48 h resulted ina dose-dependent downregulation of degs1 mRNA
expression (388 6 36 and 457 6 73 reductionrespectively)
We then investigated a link between DEGS1 and theadipogenic program It is known that obese and insulin-resistant murine models and humans have reducedexpression of pparg2 in WAT Furthermore we observedthat expression of pparg2 and degs1 was directly corre-lated in 3T3-L1 after treatment with rosiglitazone (Sup-plementary Fig 3A) To validate this in vivo we analyzedthe regulation of degs1 in AT liver and skeletal muscle ofpparg2KO and POKO mice (16) WAT of pparg2KO andPOKO mice expressed significantly lower degs1 mRNAlevels compared with WT mice (Supplementary Fig 3B)This association was restricted to WAT since the expres-sion of degs1 in other metabolic organs such as skeletalmuscle or liver was not affected when pparg2 was absent(Supplementary Fig 3C and D)
DEGS1 Controls Important Cellular Functions Such asProliferation Survival and Oxidative Stress in 3T3-L1AdipocytesA stable degs1 knockdown 3T3-L1 cell line (65) (Fig 2A)resulted in inhibition of cell growth after 24 h (Fig 2B andC) Degs1 knockdown also induced cell death (137) andapoptosis (63) (Fig 2E) These were associated with a de-crease in cyclin-A and cdk2 levels in degs1 KD proliferat-ing cells (Fig 2F) as well as elevated levels of Bax andcaspase-3 mRNA (Fig 2D) Elevated reactive oxygen speciesproduction (Fig 2I) along with an upregulation of the ex-pression of antioxidant genes were observed in degs1 KDpreadipocytes (Fig 2H) This was accompanied by impairedmitochondrial oxygen consumption (Fig 2G) withoutchanges in the number of mitochondria (Fig 2J)
DEGS1 Is Required for Adipocyte DifferentiationExpression of degs1 is increased during normal differen-tiation of 3T3-L1 adipocytes (Fig 3A) To investigatewhether ablation of DEGS1 affected adipogenesis 3T3-L1degs1 KD cells were induced to differentiate At day 9degs1 KD cells showed impaired lipid accumulation versuscontrols (Fig 3B) Moreover pref1 mRNA expressiona marker for preadipocytes was not decreased in degs1KD cells at day 9 suggesting that a relevant number ofcells remained in the preadipocyte stage (Fig 3C) In ad-dition the expressions of proadipogenic and lipogenicgenes were downregulated (Fig 3D) Interestingly treat-ment with rosiglitazone only partially improved the dif-ferentiation and lipid accumulation rates of degs1 KD cells(Supplementary Fig 4A and B)
We tested whether DEGS1 expression may affect themitotic clonal expansion (MCE) In degs1 KD cells ppargand cebpb were downregulated compared with controls attime 0 and 24 h following differentiation induction (Sup-plementary Fig 5A and B) The expression of PPARg andseveral cyclins involved in adipogenesis were measured atearlier time points We noted a strong effect of degs1depletion on PPARg2 mRNA from time 0 Additionallyreduced levels of degs1 blocked MCE as indicated by the
diabetesdiabetesjournalsorg Barbarroja and Associates 1183
inhibition of cyclin D1 and cdk2 expression between time0 and 4 h after the induction (Supplementary Fig 5A andB) Cyclin E and cyclin D3 expression was also inhibitedbut at later time points
Pharmacological Inhibition of DEGS1 RecapitulatesDEGS1 KD Antiadipogenic PhenotypeUse of C8-CPPC a selective inhibitor of DEGS1 (21) fur-ther confirmed the relevance of DEGS1 in adipogenesisSimilarly to degs1 KD cells 3T3-L1 cells were induced todifferentiate and were treated simultaneously withC8-CPPC from time 0 h every 6 h for 48 h C8-CPPC de-creased the expression of proadipogenic transcription fac-tors such as PPARg and CEBPb as well as lipogenic genes(Fig 4A and C) The antiadipogenic effect of C8-CPPC per-sisted after rosiglitazone treatment (Fig 4A and C) Asexpected analysis of neutral lipids showed that 3T3-L1 cellstreated with C8-CPPC for 96 h accumulated significantlyless lipids compared with untreated cells (Fig 4D)
Pharmacological Inhibition of DEGS1 Also ImpairsAdipocyte Differentiation In VivoWe administered C8-CPPC intraperitoneally to mice fedan HFD for 5 weeks an experimental protocol that isknown to trigger adipocyte hyperplasia Molecular analysisof the WAT from mice treated with C8-CPPC presentedhigher levels of pref-1+ cells suggesting an increase in thenumber of preadipocytes compared with controls Inter-estingly a significant number of preadipocytes showedevidence of increased proliferation (Ki67+) whereas thenumber of differentiating preadipocytes (pref1+ BODIPY+)was significantly smaller in comparison with controls (Fig5A) These data were reinforced at the mRNA level byshowing increased expression of pref-1 and gata2 (Fig5B) and the presence of fewer small adipocytes in treatedmice (Supplementary Fig 6) Altogether these data sug-gest that C8-CPPCndashmediated inhibition of DEGS1 im-paired the capacity of preadipocytes to differentiate intoadipocytes in vivo
Figure 1mdashA Tissue distribution of degs1 B Correlation of degs1 mRNA expression and fat pad size The Spearman correlation coef-ficients were calculated to estimate the linear correlations between variables The rejection level for a null hypothesis was P lt 001 Dataare from 14 lean mice CndashE Expression levels of degs1 mRNA in response to HFD FndashH Expression levels of degs1 mRNA in obob miceValues are the mean 6 SEM for six to eight animals per group P lt 005 vs WT AU arbitrary units BAT brown AT gWAT gonadal WATIngFat inguinal fat RetroFat retroperitoneal fat SKM skeletal muscle
1184 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
DEGS1 Is Required for Lipid Accumulation BasalLipolysis and Glucose Uptake in Mature AdipocytesWe next focused on the effects of DEGS1 inhibition infully differentiated adipocytes by treating isolated matureadipocytesmdashfrom the gonadal WAT of C57BL6 micemdashwith C8-CPPC for 24 h C8-CPPC caused a decrease in theexpression of lipid metabolism genes as well as an increasein antioxidant genes (Supplementary Fig 7) recapitulatingour observations in 3T3-L1 preadipocytes
We further investigated the effects of inhibiting DEGS1on lipolytic activity and insulin signaling and measuredadiponectin expression as a representative fingerprintof global adipocyte homeostasis in mature differentiated3T3-L1 adipocytes Thus we observed that basal lipolyticactivity was decreased in mature differentiated 3T3-L1adipocytes when treated with C8-CPPC in nonstimulated
conditions (Fig 6A) Systematic evaluation of the lipolyticaxis showed that under basal conditions phosphorylation ofSer565 was also substantially increased by C8-CPPC con-comitantly with a minor downregulation of Ser660 phosp-horylation (Fig 6BndashD) suggesting a decreased HSL activityMoreover total levels of HSL were also decreased inC8-CPPCndashtreated cells These data suggest that pharmaco-logical inhibition of DEGS1 in mature adipocytes disruptsthe lipolytic response under nonstimulated conditions a de-fect that was superseded in the presence of NE
Finally we characterized the effects of pharmacologicalinhibition of DEGS1 on insulin signaling in 3T3-L1 matureadipocytes treated with C8-CPPC for 48 h as describedabove and subsequently incubated in the presence ofincreasing doses of insulin No major differences wereobserved in the phosphorylation of AKT in C8-CPPCndashtreated
Figure 2mdashA mRNA expression of degs1 in 3T3-L1 treated with short hairpin RNA (shRNA) against degs1 B and C Cell proliferation (XTTand BrdU assays every 24 h until 96 and 72 h respectively) D mRNA expression of Cdk2 Bcl2 Bax and Caspase3 E Apoptosis and celldeath rate F Protein expression of Cdk2 G Oxygen consumption rate H mRNA expression of antioxidant machinery genes I Reactiveoxygen species production J Mitochondria levels All these experiments were perfomed in degs1 KD and WT 3T3-L1 cells Values are themean 6 SEM of three separate experiments performed in triplicate ETC electron transport chain MFI mean fluorescence intensity PIpropidium iodide P lt 005
diabetesdiabetesjournalsorg Barbarroja and Associates 1185
cells after acute insulin stimulation (Fig 6E) Howeverwe found that Glut4 protein levels were increased inC8-CPPCndashtreated cells suggesting that glucose uptakemay be increased (Fig 6F) Interestingly we also did notobserve any differences in either adiponectin levels in re-sponse to C8-CPPC or in AMPK phosphorylation a knowninducer of adiponectin although increased levels of totalAMPK were observed (Fig 6G and H)
Pharmacological and Genetic Inhibition of DEGS1Increases DhCerCer Ratio in 3T3-L1 CellsWe confirmed that C8-CPPC increased the DhCerCerratio in 3T3L-1 preadipocytes upon the inhibition ofDEGS1 (Fig 4E) Similarly degs1 KD preadipocytes (day 0)exhibited an increased DhCerCer ratio versus controls(Supplementary Fig 8A) Downregulation of degs1
expression in degs1 KD cells was accompanied by a down-regulation of serine palmitoyltransferase (sptlc1) and Cersynthase 6 (Cers6) (Supplementary Fig 8A) suggestive ofa homeostatic readjustment of the biosynthetic pathwayto compensate for the dysfunction of degs1 Other genesrelated to Cer homeostasis such as neutral ceramidase(ncdase) and glycosylceramide synthase (gcs) were alsodownregulated
Curiously at day 9 degs1 KD cells despite impaireddifferentiation showed a complete restoration of theDhCerCer ratio (indicated by the normalization of theCer pool) (Supplementary Fig 8B) This could be partiallyexplained by a compensation mechanism mediated bydownregulation of neutral ceramidases and the conversionof sphyngomyelins to Cers (upregulation of sphyngomye-linase 2) (Supplementary Fig 8B)
Figure 3mdashA mRNA expression of degs1 in 3T3-L1 cells during adipocyte differentiation B Lipid accumulation at day 9 of differentiationOil Red O staining C mRNA expression of PREF1 in WT and degs1 KD cells during differentiation D mRNA expression of genes involvedin adipocyte differentiation and lipid accumulation Values are the mean 6 SEM of three separate experiments performed in triplicate FASfatty acid synthase Insig insulin-induced gene LPL lipoprotein lipase P lt 005
1186 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
The Impairment of Adipocyte Differentiation DuringEarly Adipogenesis by DEGS1 Inhibition IsRecapitulated by DhCers Per SeWe finally sought to validate whether DhCer by itself couldmediate the effects of DEGS1 inhibition on adipocytedifferentiation in our models of genetic or pharmacologicalinhibition of degs1 3T3-L1cells were treated with C2DhCerduring 1) the MCE at time 0 and 2) at day 3 after differ-entiation DhCer inhibited lipid accumulation (Fig 7A) andthe expression of genes involved during the early stages ofadipogenesis (Fig 7B)
Since synthetic sphingolipid analogs are known tomodulate PPAR activity (22) we investigated whetherboth C2DhCer and C16DhCer could exert a repressiveeffect on PPARg activation in the presence of the PPARgligands (Fig 7C) Our results show that DhCer can also
block the ligand-mediated transactivation of PPARg Sim-ilar results were obtained with Cers indicating that theseeffects were not a consequence of the unsaturation of thesphingoid moiety (data not shown)
DISCUSSION
The DhCerCer ratio has an important homeostaticregulatory role in the cell contributing to cell survivalautophagy and oxidative stress (1213) Here we identifyDEGS1 as an essential metabolic enzyme which is dysreg-ulated in obese states and contributes to AT dysfunctionOur complementary in vitro and in vivo approaches revealthat reduced DEGS1 function impairs adipogenesis andlipogenesis programs and increases oxidative stress
DEGS1 is expressed in WAT and correlates in rodentswith fat mass in healthy states an association that is
Figure 4mdash3T3-L1 cells treated with C8-CPPC andor rosiglitazone A and B mRNA expression of genes involved in adipocyte differen-tiation and lipid accumulation at 24 and 48 h after induction C Protein expression levels of PPARg2 and cEBPa D Lipid content after 48and 96 h of differentiation Values are the mean 6 SEM of two separate experiments performed in triplicate One-way ANOVA was used toanalyze the statistical significance between treatments at 24 and 48 h Significant differences (Duncan test P lt 005) are indicated withdifferent letters E DhCer Cer and hexosylceramide (HexCer) levels after 48 h of differentiation Values are the mean 6 SEM of twoseparate experiments performed in triplicate cEBP CCAATenhancer binding protein d day FITC fluorescein isothiocyanate MFI meanfluorescence intensity Rosi rosiglitazone P lt 005
diabetesdiabetesjournalsorg Barbarroja and Associates 1187
disrupted in severe forms of obesity The expression ofDEGS1 in liver or muscle is not affected by obesity fur-ther indicating that the WAT is particularly susceptible tochanges in Cer metabolism in obese states The relevanceto human obesity is confirmed by the downregulation ofDEGS1 in the AT of MO patients and by recent reports(2324) showing that DhCer rather than Cer content pos-itively correlates with BMI and waist circumference incohorts of overweight obese subjects To date only a singlefunctional mutation in the degs1 gene has been reportedin humans where patients demonstrate increased serumDhCer decreased cholesterol esters and decreased waist-to-hip ratio (25) Globally considered these data suggest
the existence of an association between DEGS1 functionand fat mass
The molecular mechanism leading to the defective ATexpression of degs1 in human and rodent obesity is un-known Here we have shown that tumor necrosis factor-aa mediator of obesity-induced inflammation and insulinresistance decreases the transcriptional expression ofdegs1 in cultured adipocytes which suggests that lowchronic inflammation may contribute to DEGS1 dysfunc-tion Other obesity-associated processes such as in-creased oxidative stress and hypoxia have been shownto impair DEGS1 activity and increase DhCer levels (26)Our results also show a positive association between
Figure 5mdashC8-CPPC inhibitor increases proliferation and decreases lipid accumulation in preadipocytes in vivo Ten-week-old mice weregiven a 45 HFD for 5 weeks before being treated intraperitoneally with vehicle or C8-CPPC inhibitor (2 mgkgday) over 9 daysA Representative images of immunofluorescence analysis of gonadal AT of control (vehicle) and C8-CPPCndashtreated mice are presentedwith Pref-1 (cyan) and Ki67 (red white arrows) Nuclei and lipids are respectively stained with Hoechst stain (blue) and BODIPY (green)Scale bars 100 or 20 mm Quantification of preadipocytes (left graph) (percentage of Pref-1+ cellstotal cells) and (right graph) Ki67+ (whitebars)BODIPY+ (dark gray bars) cells among preadipocytes (Pref-1+ cells) (light gray bars) B mRNA expression of preadipocyte markersand pparg degs1 adiponectin caspase 3 and fas in gonadal AT of control and C8-CPPCndashtreated mice n = 7ndash8 mice per experimentalgroup Values are the mean 6SEM P lt 005 P lt 00001 vs vehicle
1188 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
Figure 6mdashA Glycerol release in 3T3-L1 adipocytes exposed for 48 h to C8-CPPC and stimulated with NE for 6 h BndashD Results fromWestern blot analysis for phosphorylated (p) HSLHSL ratio and total HSL in 3T3-L1 adipocytes exposed for 48 h to C8-CPPC andstimulated with NE for 6 h Graphs show the mean 6 SEM of two separated experiments P lt 005 E Effects of pharmacologicalinhibition of DEGS1 on insulin (ins) signaling Results from Western blot analysis for p-AKT and AKT in 3T3-L1 adipocytes exposed 48 or 72h with C8-CPPC 1 mmolL and increased concentrations of insulin (10ndash100 nmolL) for 15 min F and G Results from Western blot analysisfor Glut4 and adiponectin in 3T3-L1 adipocytes exposed with C8-CPPC 05ndash1 mmolL and insulin (100 nmolL) for 48 h H Results fromWestern blot analysis for AMPK in 3T3-L1 adipocytes exposed to C8-CPPC 05ndash1 mmolL and insulin (Ins) (100 nmolL) rosiglitazone (Rosi)(1 nmolL) and metformin (Metf) (100 nmolL) for 48 h aP lt 005 vs untreated cells (Ins Rosi or Metf effect) bP lt 005 vs 0 mmolLC8-CPPC (C8-CPPC effect) AU arbitrary units IOD integrated optical density
diabetesdiabetesjournalsorg Barbarroja and Associates 1189
levels of the proadipogenic pparg2 isoform and degs1in AT and given the documented downregulation ofpparg2 in AT in obese insulin-resistant states it is con-ceivable that decreased levels of pparg2 may contributeto the defective expression of degs1 associated withobesity
Studies in rodents have shown that the number of Cersis increased in the WAT of animals fed an HFD (5) con-comitantly with the onset of insulin resistance (6) Resultsfrom obob mice and obese human studies are more un-clear where both increased or decreased levels of Cershave been reported (78) This suggests that Cer synthesismay be dependent on the severity of the obese state andthe strength of the adaptive homeostatic responsesattempting to restore their normal balance
Our pharmacological and gene ablation studies haveshown that inhibition of DEGS1 is associated with oxida-tive stress cellular death impaired adipocyte differentia-tion impaired lipid accumulation and impaired basallipolysis in mature adipocytes
Our in vitro studies revealed that the defect in theadipogenesis program caused by depletion of DEGS1occurs during the early stages of adipogenesis (MCE)when PPARg controls the expression of genes that regu-late the cell cycle by directly interacting with cyclin D3Another cyclin involved in adipocyte differentiation iscyclin D1 and here we show that downregulation ofDEGS1 blocks PPARg activity and cyclins such as D1D3 E as well as decreasing cdk2 which modulates PPARgactivity during adipogenesis These data are consistent
Figure 7mdashA and B DhCers impair adipocyte differentiation during early adipogenesis 3T3-L1 preadipocytes treated with C2DhCer(50 mmolL) for a period of 48 h at day 0 or day 3 of differentiation Oil Red O staining and mRNA expression of adipogenesis and lipidmetabolism genes Values are the mean 6 SEM of two separate experiments performed in triplicate P lt 005 vs control C DhCersdecrease ligand-mediated PPARg transactivation Cells were treated with DMSO as a control group and rosiglitazone 10 mmolL GW192910 mmolL and C2 and C16DhCers and Cers 100 mmolL as indicated Graphs represent the average of three independent experimentsP lt 005 vs control cells P lt 005 vs rosiglitazone ampP lt 005 vs GW1929 CTL control
1190 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
with studies showing that downregulation of degs1 bysmall interfering RNA inhibits cell growth and arreststhe cell cycle in cancer cells (12) and where its overexpres-sion increased cell migration and metastasis (27)
The pharmacological inhibition of DEGS1 in vivovalidated the results obtained in vitro and strengthenedthe concept that functional degs1 is required for the fulldifferentiation of preadipocytes into mature adipocytes
We also provide evidence that both pharmacologicaland genetic ablation of degs1 which according to ourresults in in vitro models may directly block adipocytedifferentiation and lipid deposition certainly increasedDhCer levels in in vitro models In this line our resultsshowed that the treatment of adipocytes with DhCercauses impaired adipogenesis and lipid accumulationThis effect on lipid accumulation was more severewhen the treatment was administered during the first3 days of differentiation suggesting a direct impact ofDhCer accumulation during early stages of adipogenesisWe also provide evidence showing that accumulation ofDhCers can also directly repress the transcriptional ac-tivity of PPARg and hence could contribute to impair-ment of the capacity of the preadipocytes to developa full adipogenic program
These observations strengthen the concept that DhCersare not merely inert precursors of Cers Nevertheless themolecular mechanism linking DhCers accumulation anddysregulation of the cell cycle and other cellular eventsremains elusive One possible mechanism is that changesin the DhCerCer ratio may disrupt membrane-dependentstructures by altering the levels of cholesterol andorcaveolin in lipid rafts which are known to be highlysensitive to a sphingomyelin pool Alternatively changesin the DhCerCer ratio may disturb the global phospho-lipidome potentially altering membrane-associated pro-cesses relevant to adipocyte function These are importantquestions to address in future research
In summary our results indicate that defects in DEGS1in the context of the metabolic syndrome may compro-mise AT expansion and function through the combinedinhibition of adipogenesis promotion of cell death andoxidative stress due to the direct accumulation of DhCersThus our data suggest that the selective manipulation ofDEGS1 andor its substrates in WAT may be a pathophy-siologically advantageous strategy to improve AT homeo-stasis and ameliorate the burden of obesity-associatedmetabolic complications
Acknowledgments The authors thank Dr Carobbio Mrs Peirce andMs Philips (Wellcome Trust-Medical Research Council Institute of MetabolicScience University of Cambridge UK) for their excellent technical advice sup-port and suggestionsFunding This work was funded by the Medical Research Council (MRC) theMetabolic Diseases Unit of the MRC European Commission grant FP7-ETHERPATHSand the British Heart FoundationDuality of Interest No potential conflicts of interest relevant to this articlewere reported
Author Contributions NB helped to develop the hypothesis design theexperiments perform the in vitro and ex vivo experiments collect and analyze thedata hold and characterize the human cohorts collect gene expression data fromadipose depots and analyze the data discuss the manuscript coordinate anddirect the project and write the manuscript SR-C helped to develop thehypothesis design the experiments perform the in vitro and ex vivo experimentscollect and analyze the data discuss the manuscript coordinate and direct theproject and write the manuscript HN and MO helped to develop analyticalplatforms perform and analyze lipidomic experiments and discuss the manu-script AC AP JR IC VP GM-G and CL-P helped to perform the invitro and ex vivo experiments collect and analyze the data and discuss themanuscript FJT helped to hold and characterize the human cohorts collectgene expression data from adipose depots and analyze the data and discussthe manuscript JDS and SAS helped to design the experiments and dis-cuss the manuscript AV-P helped to develop the hypothesis design theexperiments discuss the manuscript coordinate and direct the project andwrite the manuscript AV-P is the guarantor of this work and as such had fullaccess to all the data in the study and takes responsibility for the integrity of thedata and the accuracy of the data analysis
References1 Montague CT OrsquoRahilly S The perils of portliness causes and con-
sequences of visceral adiposity Diabetes 200049883ndash8882 Carobbio S Rodriguez-Cuenca S Vidal-Puig A Origins of metabolic com-
plications in obesity ectopic fat accumulation The importance of the qualitative
aspect of lipotoxicity Curr Opin Clin Nutr Metab Care 201114520ndash5263 Chavez JA Summers SA Lipid oversupply selective insulin resistance
and lipotoxicity molecular mechanisms Biochim Biophys Acta 20101801
252ndash2654 Chavez JA Summers SA A ceramide-centric view of insulin resistance Cell
Metab 201215585ndash5945 Shah C Yang G Lee I Bielawski J Hannun YA Samad F Protection from
high fat diet-induced increase in ceramide in mice lacking plasminogen activator
inhibitor 1 J Biol Chem 200828313538ndash135486 Turner N Kowalski GM Leslie SJ et al Distinct patterns of tissue-specific
lipid accumulation during the induction of insulin resistance in mice by high-fat
feeding Diabetologia 2013561638ndash16487 Blachnio-Zabielska AU Koutsari C Tchkonia T Jensen MD Sphingolipid
content of human adipose tissue relationship to adiponectin and insulin re-
sistance Obesity (Silver Spring) 2012202341ndash23478 Błachnio-Zabielska AU Pułka M Baranowski M et al Ceramide metabo-
lism is affected by obesity and diabetes in human adipose tissue J Cell Physiol
2012227550ndash5579 Ternes P Franke S Zaumlhringer U Sperling P Heinz E Identification and
characterization of a sphingolipid delta 4-desaturase family J Biol Chem 2002
27725512ndash2551810 Mizutani Y Kihara A Igarashi Y Identification of the human sphingolipid C4-
hydroxylase hDES2 and its up-regulation during keratinocyte differentiation
FEBS Lett 200456393ndash9711 Ahn EH Schroeder JJ Induction of apoptosis by sphingosine sphinganine
and C(2)-ceramide in human colon cancer cells but not by C(2)-dihydroceramide
Anticancer Res 2010302881ndash288412 Kraveka JM Li L Szulc ZM et al Involvement of dihydroceramide desaturase
in cell cycle progression in human neuroblastoma cells J Biol Chem 2007282
16718ndash1672813 Siddique MM Li Y Wang L et al Ablation of dihydroceramide desaturase 1
a therapeutic target for the treatment of metabolic diseases simultaneously stim-
ulates anabolic and catabolic signaling Mol Cell Biol 2013332353ndash236914 Parks BW Nam E Org E et al Genetic control of obesity and gut microbiota
composition in response to high-fat high-sucrose diet in mice Cell Metab 2013
17141ndash152
diabetesdiabetesjournalsorg Barbarroja and Associates 1191
15 Bikman BT Guan Y Shui G et al Fenretinide prevents lipid-induced insulinresistance by blocking ceramide biosynthesis J Biol Chem 201228717426ndash1743716 Medina-Gomez G Gray SL Yetukuri L et al PPAR gamma 2 preventslipotoxicity by controlling adipose tissue expandability and peripheral lipid me-tabolism PLoS Genet 20073e6417 Roberts LD Virtue S Vidal-Puig A Nicholls AW Griffin JL Metabolic pheno-typing of a model of adipocyte differentiation Physiol Genomics 200939109ndash11918 Ramiacuterez-Zacariacuteas JL Castro-Muntildeozledo F Kuri-Harcuch W Quantitation ofadipose conversion and triglycerides by staining intracytoplasmic lipids with Oilred O Histochemistry 199297493ndash49719 Ye J Coulouris G Zaretskaya I Cutcutache I Rozen S Madden TL Primer-BLAST a tool to design target-specific primers for polymerase chain reactionBMC Bioinformatics 20121313420 Pfaffl MW Tichopad A Prgomet C Neuvians TP Determination of stablehousekeeping genes differentially regulated target genes and sample integrityBestKeepermdashExcel-based tool using pair-wise correlations Biotechnol Lett 200426509ndash51521 Triola G Fabriagraves G Llebaria A Synthesis of a cyclopropene analogue of cer-amide a potent inhibitor of dihydroceramide desaturase Angew Chem Int Ed Engl2001401960ndash1962
22 Tsuji K Satoh S Mitsutake S et al Evaluation of synthetic sphingolipidanalogs as ligands for peroxisome proliferator-activated receptors Bioorg MedChem Lett 2009191643ndash164623 Mamtani M Meikle PJ Kulkarni H et al Plasma dihydroceramide speciesassociate with waist circumference in Mexican American families Obesity (SilverSpring) 201422950ndash95624 Weir JM Wong G Barlow CK et al Plasma lipid profiling in a largepopulation-based cohort J Lipid Res 2013542898ndash290825 Curran J Meikle P Weir J Jowett J Deep sequencing in extended pedi-grees reveals a major rare non-synonymous variant influencing the de novoceramide synthesis pathway Late-breaking abstract presented at the AmericanSociety of Human Genetics 2013 Meeting 22ndash26 October 2013 at the BostonConvention and Exhibition Center Boston Massachusetts26 Idkowiak-Baldys J Takemoto JY Grilley MM Structure-function studies ofyeast C-4 sphingolipid long chain base hydroxylase Biochim Biophys Acta 2003161817ndash2427 Zhou W Ye XL Sun ZJ Ji XD Chen HX Xie D Overexpression of de-generative spermatocyte homolog 1 up-regulates the expression of cyclin D1 andenhances metastatic efficiency in esophageal carcinoma Eca109 cells MolCarcinog 200948886ndash894
1192 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015

conflicting results showing either increased or decreasedCer levels in obese and insulin-resistant patients (78)
Cers can be synthesized from sphingomyelins but themain contributor to their biosynthesis is the de novopathway The final reaction of the de novo pathway iscatalyzed by a D4-dihydroceramide (DhCer) desaturase(DEGS1) that adds a 45-trans-double bound on the sphin-goid base of the DhCer (9) The other enzyme DEGS2catalyzes the synthesis of phytoceramides (10) whoseexpression is restricted to skin intestine and kidney
There is evidence that downregulation of degs1 in-creases the DhCerCer ratio in different cellular modelsAlthough DhCer was considered an inactive precursorof Cers (11) recent studies (1213) have suggested theirrelevance as modulators of cell cycle apoptosis autophagyor oxidative stress processes that a priori are expectedto compromise the development and function of adiposetissue (AT)
Since DEGS1 is the key enzyme regulating the DhCerCer ratio understanding its regulation is important indetermining the pathophysiological relevance of this path-way in AT Of note degs1 has recently been identifiedin genome-wide association studies as a candidate geneassociated with fat mass accumulation in mice (14) fur-ther suggesting that DEGS1 may be relevant for theadaptive accretion of AT Moreover DEGS1 could beconsidered an attractive therapeutic target for obesity-associated insulin resistance since fenretinide has beenclaimed to improve insulin sensitivity by inhibitingDEGS1 (15) although other molecular targets cannotbe ruled out
Here we show that DEGS1 expression is selectivelyperturbed in the WAT of murine models of nutritionaland genetically induced obesity and in the WAT of mor-bidly obese (MO) patients In vitro analysis revealed thatboth pharmacological inhibition and genetic ablation ofDEGS1 result in impaired adipocyte differentiation andlipid accumulation effects mediated by increased DhCercontent In vivo pharmacological inhibition of DEGS1also resulted in impaired adipocyte differentiation De-creased levels of DEGS1 were associated with increasedoxidative stress accelerated cellular death and blockageof cell cycle We also show data supporting that DEGS1expression is regulated by peroxisome proliferatorndashactivated receptor (PPAR)g
RESEARCH DESIGN AND METHODS
Mice HusbandryAnimals were housed at 22ndash24degC with 12-h lightdark cyclesFood and water were available ad libitum Lean wild-type(WT) PPARg222 and PPARg2obob double knockoutmice (16) were used for profiling purposes Mice were feda normal chow diet (D12450B) or an HFD (D12451) fromResearch Diets
For the pharmacological inhibition of DEGS1 10-week-old male C57BL6 WT mice were used Mice were fedan HFD for 5 weeks before C8-cyclopropenylceramide
(C8-CPPC) (Matreya) administration All protocols usedwere approved by the UK Home Office
Intraperitoneal Injection of CyclopropenylceramideMice were distributed in two groups (n = 8) and admin-istered daily vehicle (2-hydroxypropyl betacyclodextrine)or C8-CPPC (2 mgkgday) for 9 days via intraperitonealinjection Mice were culled at the end of the experimentand WAT was removed for gene expression and micros-copy analysis No differences in food intake lean massand fat content were observed during the treatment
Ex Vivo Experiments in Isolated Mature AdipocytesAdipocytes from gonadal WAT of 16-week-old C57BL6 micewere obtained by collagenase type II digestion at 37degC Afterdigestion adipocytes were placed in DMEM with or without1 mmolL C8-CPPC every 5 h for a total period of 20 h
Retroviral Short Hairpin RNA Constructs for DEGS1RNAi-Ready pSIREN-RetroQ vectors (BD Biosciences)were used to target degs1 in 3T3-L1 cells Sequences tar-geting degs1 were ligated into the pSIREN vector as de-scribed in the manufacturerrsquos instructions Retroviruseswere generated by transfecting BOSC cells (AmericanType Culture Collection) with the pSIREN plasmids usingFuGene6 (Roche) Supernatant with the viral content wasused to transfect 3T3-L1 preadipocytes Twenty-fourhours after retroviral infection the cells were selectedwith puromycin (4 mgmL)
Culture Differentiation and Treatment of 3T3-L1PreadipocytesCells were differentiated into adipocytes (day 9) accord-ingly to the protocol described by Roberts et al (17) withor without rosiglitazone 01 mmolL Lipid accumulationwas assessed by Oil Red O solution (18)
Pharmacological Inhibition of DEGS1 Activity In Vitro
Effects on DifferentiationAt day 0 of differentiation 3T3-L1 cells were treated withC8-CPPC 1 mmolL andor rosiglitazone 01 mmolL for 96 h
Effects on LipolysisOn day 8 of differentiation after 48 h of treatment withC8-CPPC 1 mmolL the cells were exposed to norepineph-rine (NE) (1028 and 1027 molL) for 6 h
Effects on Insulin SignalingOn day 8 of differentiation after 48 h of treatment withC8-CPPC 1 mmolL the cells were exposed to insulin (10and 100 nmolL) for 15 min
Effects on GLUT4 and AdiponectinOn day 8 of differentiation after 48 h of treatment withC8-CPPC 05ndash1 mmolL the cells were exposed to insulin
Effects on AMPKOn day 8 of differentiation after 48 h of treatment withC8-CPPC 05ndash1 mmolL the cells were exposed to insulin(10 and 100 nmolL) rosiglitazone (1 and 10 nmolL)and metformin (100 nmolL) for 24 h
diabetesdiabetesjournalsorg Barbarroja and Associates 1181
DhCer Treatment3T3-L1 preadipocytes were treated at day 0 of differentia-tion with the induction cocktail and C2DhCer at 50 mmolLfor 3 days and a second batch of cells were exposed at day 3of differentiation and treated with C2DhCer at 50 mmolLfor 3 days All batches were taken until final differentiationat day 9
Human SamplesThe cohort included 28 MO and 6 nonobese subjects withno alterations to lipid or glucose metabolism as controlsubjects (Table 1) Approval for the study was given by theethics committee and all patients gave their informedconsent Visceral AT biopsy samples were obtained fromMO patients undergoing bariatric surgery (Scopinaro pro-cedure) or laparoscopic surgery (hiatus hernia repair orcholecystectomies) for the lean subjects
Western BlottingProtein extracts were prepared using the Nuclear andCytoplasmic Extraction Reagents Kit (Pierce) according tothe manufacturerrsquos instructions Immunoblots were incu-bated with the following antibodies PPARg CEBPb andCyclinA (Santa Cruz Biotechnology) CyclinB1 D1 D3 andE1 cdk4 AKT Ser473-pAKT p4442 MAPK phospho-p4442 MAPK Ser660-pHSL (p-hormone-sensitive lipase)Ser565-pHSL total HSL ATGL caveolin-1 plin1 glut4 adi-ponectin and AMPK (Cell Signaling Technology) and adfpadrp abhd5 and anti-b-actin (Abcam)
RT-PCRRNA was extracted using TRI Reagent (Sigma) and reversetranscribed to cDNA Real-time PCR using SYBRgreenwas performed according to the manufacturerrsquos instruc-tions (ABI) Primer sequences were obtained from PrimerBlast (19) Expression of genes was corrected by the
geometrical average of 18s b2m b-act and 36b4 usingBestkeeper (20)
Apoptosis Combined Annexin VPropidium IodideStainingViability was assessed by using an Annexin Vpropidiumiodide kit (Bender MedSystems) according to the manufac-turerrsquos recommendations Binding of fluorescein-conjugatedAnnexin V and propidium iodide was measured by FACSCalibur(BD Biosciences)
Cellular ProliferationCell viability was assessed using an colorimetric assay(Roche) following the protocol supplied by the manufac-turer Cell proliferation was analyzed by quantification ofthe incorporation of BrdU (Roche)
Boron-Dipyrromethene Staining Reactive OxygenStress Production and Mitochondrial ContentCells were incubated with boron-dipyrromethene (BODIPY)at 4degC or alternatively with 20 mmolL 2979-dichlorodihy-drofluorescein diacetate or 100 nmolL Mitotracker (Invi-trogen) at 37degC for 30 min and analyzed on a FACSCaliburcell analyzer
Cellular Oxygen ConsumptionCells were exposed to oligomycin (1 mmolL) C8-CPPC(09 mmolL) and antimycinrotenone (1 mmolL each)O2 consumption was measured using the XF24 analyzer(Seahorse Bioscience) for a period of 90 min
Whole-Mount Confocal MicroscopyGonadal AT was fixed in 4 paraformaldehyde Sampleswere incubated at 4degC with mouse Pref-1 (preadipocytefactor-1) or rabbit Ki67 primary antibodies Nuclei andneutral lipids were stained with Hoechst 33342 and BODIPY493503 respectively Sample examination was performedusing a Zeiss 510 confocal laser scanning microscope (CarlZeiss) Pref-1+ Ki67+ and BODIPY+ cells and total cells (upto 913 cellsfield) were automatically counted by usingImageJ software in three independent fields in each tissue
Light Microscopy AnalysisSamples for AT hematoxylin-eosin staining were preparedas described previously (16) Adipocyte sizes were mea-sured using Cell P (Olympus Soft Imaging SolutionsGmbH) Between 1000 and 3000 adipocytes from eachmouse were used to obtain the mean cell area
Luciferase Reporter AssayHuman embryonic kidney 293 cells were transfectedusing Lipofectamine LTX (Invitrogen) following themanufacturerrsquos instructions Seventy-five nanograms ofthe reporter plasmid (3xPPRE TKLuc) and 375 ng of theeukaryotic expression vector (pSV-PPARg) were cotrans-fected to each well The plasmid pRL-CMV (5 ngwell)was included as an internal control Twenty-four hourspost-transfection cells were treated as indicated (DMSOrosiglitazone 10 mmolL GW1929 10 mmolL andC2DhCer C16DhCer and Cer 100 mmolL) Luciferaseassays were performed using the dual luciferase reporterassay system (Promega)
Table 1mdashClinical characteristics of control subjects and MOpatients
Control subjects MO patients
Sex nMale 3 14Female 3 14
Age years 5217 6 51 4157 6 296
Weight kg 7400 6 071 1555 6 732
Height cm 16260 6 160 16553 6 249
BMI kgm2 2504 6 055 5651 6 168
Serum insulin IUmL 1123 6 158 3009 6 230
HOMA-IR 351 6 048 818 6 078
Serum glucose mmolL 603 6 032 574 6 026
Serum cholesterol mmolL 468 6 040 492 6 025
HDL cholesterol mmolL 097 6 023 110 6 009
Triglycerides mmolL 108 6 020 142 6 019
Values are reported as the mean 6 SEM unless otherwise in-dicated HOMA-IR HOMA of insulin resistance
1182 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
LipidomicsCells were mixed with 09 NaCl and sonicated for 5 minat 5degC and 40 kHz Samples were spiked with internalstandard The samples were extracted with chloroformmethanol (21) The lower phase was collected and mixedwith the labeled standard mixture (three stable isotope-labeled reference compounds) Lipid extracts were ana-lyzed on a Q-ToF Premier Mass Spectrometer (Waters)combined with an Acquity Ultra Performance LiquidChromatographyMass Spectrometry system (Waters)The data were processed using MZmine software Thelipids were quantified by normalizing with correspond-ing internal standard
Statistical AnalysisStudent t test (unpaired) ANOVA and Duncan test wereused for the statistical analysis Statistical significance wasset at P 005 and P 001 The Spearman correlationwas calculated to estimate the linear correlations betweenvariables at P 001
RESULTS
DEGS1 Is Downregulated in WAT in Obese MurineModelsTissue distribution analysis showed that degs1 is presentin most tissues and it is particularly highly expressed inAT liver and muscle whereas degs2 mRNA is detectedonly in intestine (Fig 1A) Degs1 expression in vivo waspositively correlated with WAT mass in lean healthy mice(Fig 1B) However this correlation was disrupted in mu-rine models of nutritional and genetically induced obesity(HFD and obob) where degs1 mRNA expression was de-creased versus controls (Fig 1C and F) Of note the dys-regulation of degs1 in obesity was limited to WAT as itsexpression in liver or skeletal muscle was not affected ineither HFD-fed mice (Fig 1D and E) or obob mice (Fig1G and H) DEGS1 was preferentially expressed in ma-tured adipocytes versus stromal vascular fraction in leanand obob mice (Supplementary Fig 1) and downregulatedin obob adipocytes versus lean adipocytes Moreover thisdownregulation of degs1 in total WAT of HFD-fed and obobmice was also recapitulated in visceral AT of MO patients(Supplementary Fig 2A)
To determine whether the decrease in degs1 expressionwas part of a global adaptation of de novo Cer synthesispathway in obesity the expression of other genes in thispathway was measured Expression of the sptlc1 and sptlc2subunits was not changed in human visceral AT or in obobWAT but sptlc2 expression was increased in HFD WAT(Supplementary Fig 2B and C) Moreover both HFD andobob mouse WAT exhibited a reduction in a subset of Cersynthases suggesting a possible defect in the synthesis ofa specific subset of Cers
We then confirmed that modulators of obesity-associatedinflammation may contribute to the downregulation ofdegs1 as treatment of 3T3-L1 adipocytes with tumornecrosis factor-a (5 and 10 ngmL) for 48 h resulted ina dose-dependent downregulation of degs1 mRNA
expression (388 6 36 and 457 6 73 reductionrespectively)
We then investigated a link between DEGS1 and theadipogenic program It is known that obese and insulin-resistant murine models and humans have reducedexpression of pparg2 in WAT Furthermore we observedthat expression of pparg2 and degs1 was directly corre-lated in 3T3-L1 after treatment with rosiglitazone (Sup-plementary Fig 3A) To validate this in vivo we analyzedthe regulation of degs1 in AT liver and skeletal muscle ofpparg2KO and POKO mice (16) WAT of pparg2KO andPOKO mice expressed significantly lower degs1 mRNAlevels compared with WT mice (Supplementary Fig 3B)This association was restricted to WAT since the expres-sion of degs1 in other metabolic organs such as skeletalmuscle or liver was not affected when pparg2 was absent(Supplementary Fig 3C and D)
DEGS1 Controls Important Cellular Functions Such asProliferation Survival and Oxidative Stress in 3T3-L1AdipocytesA stable degs1 knockdown 3T3-L1 cell line (65) (Fig 2A)resulted in inhibition of cell growth after 24 h (Fig 2B andC) Degs1 knockdown also induced cell death (137) andapoptosis (63) (Fig 2E) These were associated with a de-crease in cyclin-A and cdk2 levels in degs1 KD proliferat-ing cells (Fig 2F) as well as elevated levels of Bax andcaspase-3 mRNA (Fig 2D) Elevated reactive oxygen speciesproduction (Fig 2I) along with an upregulation of the ex-pression of antioxidant genes were observed in degs1 KDpreadipocytes (Fig 2H) This was accompanied by impairedmitochondrial oxygen consumption (Fig 2G) withoutchanges in the number of mitochondria (Fig 2J)
DEGS1 Is Required for Adipocyte DifferentiationExpression of degs1 is increased during normal differen-tiation of 3T3-L1 adipocytes (Fig 3A) To investigatewhether ablation of DEGS1 affected adipogenesis 3T3-L1degs1 KD cells were induced to differentiate At day 9degs1 KD cells showed impaired lipid accumulation versuscontrols (Fig 3B) Moreover pref1 mRNA expressiona marker for preadipocytes was not decreased in degs1KD cells at day 9 suggesting that a relevant number ofcells remained in the preadipocyte stage (Fig 3C) In ad-dition the expressions of proadipogenic and lipogenicgenes were downregulated (Fig 3D) Interestingly treat-ment with rosiglitazone only partially improved the dif-ferentiation and lipid accumulation rates of degs1 KD cells(Supplementary Fig 4A and B)
We tested whether DEGS1 expression may affect themitotic clonal expansion (MCE) In degs1 KD cells ppargand cebpb were downregulated compared with controls attime 0 and 24 h following differentiation induction (Sup-plementary Fig 5A and B) The expression of PPARg andseveral cyclins involved in adipogenesis were measured atearlier time points We noted a strong effect of degs1depletion on PPARg2 mRNA from time 0 Additionallyreduced levels of degs1 blocked MCE as indicated by the
diabetesdiabetesjournalsorg Barbarroja and Associates 1183
inhibition of cyclin D1 and cdk2 expression between time0 and 4 h after the induction (Supplementary Fig 5A andB) Cyclin E and cyclin D3 expression was also inhibitedbut at later time points
Pharmacological Inhibition of DEGS1 RecapitulatesDEGS1 KD Antiadipogenic PhenotypeUse of C8-CPPC a selective inhibitor of DEGS1 (21) fur-ther confirmed the relevance of DEGS1 in adipogenesisSimilarly to degs1 KD cells 3T3-L1 cells were induced todifferentiate and were treated simultaneously withC8-CPPC from time 0 h every 6 h for 48 h C8-CPPC de-creased the expression of proadipogenic transcription fac-tors such as PPARg and CEBPb as well as lipogenic genes(Fig 4A and C) The antiadipogenic effect of C8-CPPC per-sisted after rosiglitazone treatment (Fig 4A and C) Asexpected analysis of neutral lipids showed that 3T3-L1 cellstreated with C8-CPPC for 96 h accumulated significantlyless lipids compared with untreated cells (Fig 4D)
Pharmacological Inhibition of DEGS1 Also ImpairsAdipocyte Differentiation In VivoWe administered C8-CPPC intraperitoneally to mice fedan HFD for 5 weeks an experimental protocol that isknown to trigger adipocyte hyperplasia Molecular analysisof the WAT from mice treated with C8-CPPC presentedhigher levels of pref-1+ cells suggesting an increase in thenumber of preadipocytes compared with controls Inter-estingly a significant number of preadipocytes showedevidence of increased proliferation (Ki67+) whereas thenumber of differentiating preadipocytes (pref1+ BODIPY+)was significantly smaller in comparison with controls (Fig5A) These data were reinforced at the mRNA level byshowing increased expression of pref-1 and gata2 (Fig5B) and the presence of fewer small adipocytes in treatedmice (Supplementary Fig 6) Altogether these data sug-gest that C8-CPPCndashmediated inhibition of DEGS1 im-paired the capacity of preadipocytes to differentiate intoadipocytes in vivo
Figure 1mdashA Tissue distribution of degs1 B Correlation of degs1 mRNA expression and fat pad size The Spearman correlation coef-ficients were calculated to estimate the linear correlations between variables The rejection level for a null hypothesis was P lt 001 Dataare from 14 lean mice CndashE Expression levels of degs1 mRNA in response to HFD FndashH Expression levels of degs1 mRNA in obob miceValues are the mean 6 SEM for six to eight animals per group P lt 005 vs WT AU arbitrary units BAT brown AT gWAT gonadal WATIngFat inguinal fat RetroFat retroperitoneal fat SKM skeletal muscle
1184 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
DEGS1 Is Required for Lipid Accumulation BasalLipolysis and Glucose Uptake in Mature AdipocytesWe next focused on the effects of DEGS1 inhibition infully differentiated adipocytes by treating isolated matureadipocytesmdashfrom the gonadal WAT of C57BL6 micemdashwith C8-CPPC for 24 h C8-CPPC caused a decrease in theexpression of lipid metabolism genes as well as an increasein antioxidant genes (Supplementary Fig 7) recapitulatingour observations in 3T3-L1 preadipocytes
We further investigated the effects of inhibiting DEGS1on lipolytic activity and insulin signaling and measuredadiponectin expression as a representative fingerprintof global adipocyte homeostasis in mature differentiated3T3-L1 adipocytes Thus we observed that basal lipolyticactivity was decreased in mature differentiated 3T3-L1adipocytes when treated with C8-CPPC in nonstimulated
conditions (Fig 6A) Systematic evaluation of the lipolyticaxis showed that under basal conditions phosphorylation ofSer565 was also substantially increased by C8-CPPC con-comitantly with a minor downregulation of Ser660 phosp-horylation (Fig 6BndashD) suggesting a decreased HSL activityMoreover total levels of HSL were also decreased inC8-CPPCndashtreated cells These data suggest that pharmaco-logical inhibition of DEGS1 in mature adipocytes disruptsthe lipolytic response under nonstimulated conditions a de-fect that was superseded in the presence of NE
Finally we characterized the effects of pharmacologicalinhibition of DEGS1 on insulin signaling in 3T3-L1 matureadipocytes treated with C8-CPPC for 48 h as describedabove and subsequently incubated in the presence ofincreasing doses of insulin No major differences wereobserved in the phosphorylation of AKT in C8-CPPCndashtreated
Figure 2mdashA mRNA expression of degs1 in 3T3-L1 treated with short hairpin RNA (shRNA) against degs1 B and C Cell proliferation (XTTand BrdU assays every 24 h until 96 and 72 h respectively) D mRNA expression of Cdk2 Bcl2 Bax and Caspase3 E Apoptosis and celldeath rate F Protein expression of Cdk2 G Oxygen consumption rate H mRNA expression of antioxidant machinery genes I Reactiveoxygen species production J Mitochondria levels All these experiments were perfomed in degs1 KD and WT 3T3-L1 cells Values are themean 6 SEM of three separate experiments performed in triplicate ETC electron transport chain MFI mean fluorescence intensity PIpropidium iodide P lt 005
diabetesdiabetesjournalsorg Barbarroja and Associates 1185
cells after acute insulin stimulation (Fig 6E) Howeverwe found that Glut4 protein levels were increased inC8-CPPCndashtreated cells suggesting that glucose uptakemay be increased (Fig 6F) Interestingly we also did notobserve any differences in either adiponectin levels in re-sponse to C8-CPPC or in AMPK phosphorylation a knowninducer of adiponectin although increased levels of totalAMPK were observed (Fig 6G and H)
Pharmacological and Genetic Inhibition of DEGS1Increases DhCerCer Ratio in 3T3-L1 CellsWe confirmed that C8-CPPC increased the DhCerCerratio in 3T3L-1 preadipocytes upon the inhibition ofDEGS1 (Fig 4E) Similarly degs1 KD preadipocytes (day 0)exhibited an increased DhCerCer ratio versus controls(Supplementary Fig 8A) Downregulation of degs1
expression in degs1 KD cells was accompanied by a down-regulation of serine palmitoyltransferase (sptlc1) and Cersynthase 6 (Cers6) (Supplementary Fig 8A) suggestive ofa homeostatic readjustment of the biosynthetic pathwayto compensate for the dysfunction of degs1 Other genesrelated to Cer homeostasis such as neutral ceramidase(ncdase) and glycosylceramide synthase (gcs) were alsodownregulated
Curiously at day 9 degs1 KD cells despite impaireddifferentiation showed a complete restoration of theDhCerCer ratio (indicated by the normalization of theCer pool) (Supplementary Fig 8B) This could be partiallyexplained by a compensation mechanism mediated bydownregulation of neutral ceramidases and the conversionof sphyngomyelins to Cers (upregulation of sphyngomye-linase 2) (Supplementary Fig 8B)
Figure 3mdashA mRNA expression of degs1 in 3T3-L1 cells during adipocyte differentiation B Lipid accumulation at day 9 of differentiationOil Red O staining C mRNA expression of PREF1 in WT and degs1 KD cells during differentiation D mRNA expression of genes involvedin adipocyte differentiation and lipid accumulation Values are the mean 6 SEM of three separate experiments performed in triplicate FASfatty acid synthase Insig insulin-induced gene LPL lipoprotein lipase P lt 005
1186 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
The Impairment of Adipocyte Differentiation DuringEarly Adipogenesis by DEGS1 Inhibition IsRecapitulated by DhCers Per SeWe finally sought to validate whether DhCer by itself couldmediate the effects of DEGS1 inhibition on adipocytedifferentiation in our models of genetic or pharmacologicalinhibition of degs1 3T3-L1cells were treated with C2DhCerduring 1) the MCE at time 0 and 2) at day 3 after differ-entiation DhCer inhibited lipid accumulation (Fig 7A) andthe expression of genes involved during the early stages ofadipogenesis (Fig 7B)
Since synthetic sphingolipid analogs are known tomodulate PPAR activity (22) we investigated whetherboth C2DhCer and C16DhCer could exert a repressiveeffect on PPARg activation in the presence of the PPARgligands (Fig 7C) Our results show that DhCer can also
block the ligand-mediated transactivation of PPARg Sim-ilar results were obtained with Cers indicating that theseeffects were not a consequence of the unsaturation of thesphingoid moiety (data not shown)
DISCUSSION
The DhCerCer ratio has an important homeostaticregulatory role in the cell contributing to cell survivalautophagy and oxidative stress (1213) Here we identifyDEGS1 as an essential metabolic enzyme which is dysreg-ulated in obese states and contributes to AT dysfunctionOur complementary in vitro and in vivo approaches revealthat reduced DEGS1 function impairs adipogenesis andlipogenesis programs and increases oxidative stress
DEGS1 is expressed in WAT and correlates in rodentswith fat mass in healthy states an association that is
Figure 4mdash3T3-L1 cells treated with C8-CPPC andor rosiglitazone A and B mRNA expression of genes involved in adipocyte differen-tiation and lipid accumulation at 24 and 48 h after induction C Protein expression levels of PPARg2 and cEBPa D Lipid content after 48and 96 h of differentiation Values are the mean 6 SEM of two separate experiments performed in triplicate One-way ANOVA was used toanalyze the statistical significance between treatments at 24 and 48 h Significant differences (Duncan test P lt 005) are indicated withdifferent letters E DhCer Cer and hexosylceramide (HexCer) levels after 48 h of differentiation Values are the mean 6 SEM of twoseparate experiments performed in triplicate cEBP CCAATenhancer binding protein d day FITC fluorescein isothiocyanate MFI meanfluorescence intensity Rosi rosiglitazone P lt 005
diabetesdiabetesjournalsorg Barbarroja and Associates 1187
disrupted in severe forms of obesity The expression ofDEGS1 in liver or muscle is not affected by obesity fur-ther indicating that the WAT is particularly susceptible tochanges in Cer metabolism in obese states The relevanceto human obesity is confirmed by the downregulation ofDEGS1 in the AT of MO patients and by recent reports(2324) showing that DhCer rather than Cer content pos-itively correlates with BMI and waist circumference incohorts of overweight obese subjects To date only a singlefunctional mutation in the degs1 gene has been reportedin humans where patients demonstrate increased serumDhCer decreased cholesterol esters and decreased waist-to-hip ratio (25) Globally considered these data suggest
the existence of an association between DEGS1 functionand fat mass
The molecular mechanism leading to the defective ATexpression of degs1 in human and rodent obesity is un-known Here we have shown that tumor necrosis factor-aa mediator of obesity-induced inflammation and insulinresistance decreases the transcriptional expression ofdegs1 in cultured adipocytes which suggests that lowchronic inflammation may contribute to DEGS1 dysfunc-tion Other obesity-associated processes such as in-creased oxidative stress and hypoxia have been shownto impair DEGS1 activity and increase DhCer levels (26)Our results also show a positive association between
Figure 5mdashC8-CPPC inhibitor increases proliferation and decreases lipid accumulation in preadipocytes in vivo Ten-week-old mice weregiven a 45 HFD for 5 weeks before being treated intraperitoneally with vehicle or C8-CPPC inhibitor (2 mgkgday) over 9 daysA Representative images of immunofluorescence analysis of gonadal AT of control (vehicle) and C8-CPPCndashtreated mice are presentedwith Pref-1 (cyan) and Ki67 (red white arrows) Nuclei and lipids are respectively stained with Hoechst stain (blue) and BODIPY (green)Scale bars 100 or 20 mm Quantification of preadipocytes (left graph) (percentage of Pref-1+ cellstotal cells) and (right graph) Ki67+ (whitebars)BODIPY+ (dark gray bars) cells among preadipocytes (Pref-1+ cells) (light gray bars) B mRNA expression of preadipocyte markersand pparg degs1 adiponectin caspase 3 and fas in gonadal AT of control and C8-CPPCndashtreated mice n = 7ndash8 mice per experimentalgroup Values are the mean 6SEM P lt 005 P lt 00001 vs vehicle
1188 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
Figure 6mdashA Glycerol release in 3T3-L1 adipocytes exposed for 48 h to C8-CPPC and stimulated with NE for 6 h BndashD Results fromWestern blot analysis for phosphorylated (p) HSLHSL ratio and total HSL in 3T3-L1 adipocytes exposed for 48 h to C8-CPPC andstimulated with NE for 6 h Graphs show the mean 6 SEM of two separated experiments P lt 005 E Effects of pharmacologicalinhibition of DEGS1 on insulin (ins) signaling Results from Western blot analysis for p-AKT and AKT in 3T3-L1 adipocytes exposed 48 or 72h with C8-CPPC 1 mmolL and increased concentrations of insulin (10ndash100 nmolL) for 15 min F and G Results from Western blot analysisfor Glut4 and adiponectin in 3T3-L1 adipocytes exposed with C8-CPPC 05ndash1 mmolL and insulin (100 nmolL) for 48 h H Results fromWestern blot analysis for AMPK in 3T3-L1 adipocytes exposed to C8-CPPC 05ndash1 mmolL and insulin (Ins) (100 nmolL) rosiglitazone (Rosi)(1 nmolL) and metformin (Metf) (100 nmolL) for 48 h aP lt 005 vs untreated cells (Ins Rosi or Metf effect) bP lt 005 vs 0 mmolLC8-CPPC (C8-CPPC effect) AU arbitrary units IOD integrated optical density
diabetesdiabetesjournalsorg Barbarroja and Associates 1189
levels of the proadipogenic pparg2 isoform and degs1in AT and given the documented downregulation ofpparg2 in AT in obese insulin-resistant states it is con-ceivable that decreased levels of pparg2 may contributeto the defective expression of degs1 associated withobesity
Studies in rodents have shown that the number of Cersis increased in the WAT of animals fed an HFD (5) con-comitantly with the onset of insulin resistance (6) Resultsfrom obob mice and obese human studies are more un-clear where both increased or decreased levels of Cershave been reported (78) This suggests that Cer synthesismay be dependent on the severity of the obese state andthe strength of the adaptive homeostatic responsesattempting to restore their normal balance
Our pharmacological and gene ablation studies haveshown that inhibition of DEGS1 is associated with oxida-tive stress cellular death impaired adipocyte differentia-tion impaired lipid accumulation and impaired basallipolysis in mature adipocytes
Our in vitro studies revealed that the defect in theadipogenesis program caused by depletion of DEGS1occurs during the early stages of adipogenesis (MCE)when PPARg controls the expression of genes that regu-late the cell cycle by directly interacting with cyclin D3Another cyclin involved in adipocyte differentiation iscyclin D1 and here we show that downregulation ofDEGS1 blocks PPARg activity and cyclins such as D1D3 E as well as decreasing cdk2 which modulates PPARgactivity during adipogenesis These data are consistent
Figure 7mdashA and B DhCers impair adipocyte differentiation during early adipogenesis 3T3-L1 preadipocytes treated with C2DhCer(50 mmolL) for a period of 48 h at day 0 or day 3 of differentiation Oil Red O staining and mRNA expression of adipogenesis and lipidmetabolism genes Values are the mean 6 SEM of two separate experiments performed in triplicate P lt 005 vs control C DhCersdecrease ligand-mediated PPARg transactivation Cells were treated with DMSO as a control group and rosiglitazone 10 mmolL GW192910 mmolL and C2 and C16DhCers and Cers 100 mmolL as indicated Graphs represent the average of three independent experimentsP lt 005 vs control cells P lt 005 vs rosiglitazone ampP lt 005 vs GW1929 CTL control
1190 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
with studies showing that downregulation of degs1 bysmall interfering RNA inhibits cell growth and arreststhe cell cycle in cancer cells (12) and where its overexpres-sion increased cell migration and metastasis (27)
The pharmacological inhibition of DEGS1 in vivovalidated the results obtained in vitro and strengthenedthe concept that functional degs1 is required for the fulldifferentiation of preadipocytes into mature adipocytes
We also provide evidence that both pharmacologicaland genetic ablation of degs1 which according to ourresults in in vitro models may directly block adipocytedifferentiation and lipid deposition certainly increasedDhCer levels in in vitro models In this line our resultsshowed that the treatment of adipocytes with DhCercauses impaired adipogenesis and lipid accumulationThis effect on lipid accumulation was more severewhen the treatment was administered during the first3 days of differentiation suggesting a direct impact ofDhCer accumulation during early stages of adipogenesisWe also provide evidence showing that accumulation ofDhCers can also directly repress the transcriptional ac-tivity of PPARg and hence could contribute to impair-ment of the capacity of the preadipocytes to developa full adipogenic program
These observations strengthen the concept that DhCersare not merely inert precursors of Cers Nevertheless themolecular mechanism linking DhCers accumulation anddysregulation of the cell cycle and other cellular eventsremains elusive One possible mechanism is that changesin the DhCerCer ratio may disrupt membrane-dependentstructures by altering the levels of cholesterol andorcaveolin in lipid rafts which are known to be highlysensitive to a sphingomyelin pool Alternatively changesin the DhCerCer ratio may disturb the global phospho-lipidome potentially altering membrane-associated pro-cesses relevant to adipocyte function These are importantquestions to address in future research
In summary our results indicate that defects in DEGS1in the context of the metabolic syndrome may compro-mise AT expansion and function through the combinedinhibition of adipogenesis promotion of cell death andoxidative stress due to the direct accumulation of DhCersThus our data suggest that the selective manipulation ofDEGS1 andor its substrates in WAT may be a pathophy-siologically advantageous strategy to improve AT homeo-stasis and ameliorate the burden of obesity-associatedmetabolic complications
Acknowledgments The authors thank Dr Carobbio Mrs Peirce andMs Philips (Wellcome Trust-Medical Research Council Institute of MetabolicScience University of Cambridge UK) for their excellent technical advice sup-port and suggestionsFunding This work was funded by the Medical Research Council (MRC) theMetabolic Diseases Unit of the MRC European Commission grant FP7-ETHERPATHSand the British Heart FoundationDuality of Interest No potential conflicts of interest relevant to this articlewere reported
Author Contributions NB helped to develop the hypothesis design theexperiments perform the in vitro and ex vivo experiments collect and analyze thedata hold and characterize the human cohorts collect gene expression data fromadipose depots and analyze the data discuss the manuscript coordinate anddirect the project and write the manuscript SR-C helped to develop thehypothesis design the experiments perform the in vitro and ex vivo experimentscollect and analyze the data discuss the manuscript coordinate and direct theproject and write the manuscript HN and MO helped to develop analyticalplatforms perform and analyze lipidomic experiments and discuss the manu-script AC AP JR IC VP GM-G and CL-P helped to perform the invitro and ex vivo experiments collect and analyze the data and discuss themanuscript FJT helped to hold and characterize the human cohorts collectgene expression data from adipose depots and analyze the data and discussthe manuscript JDS and SAS helped to design the experiments and dis-cuss the manuscript AV-P helped to develop the hypothesis design theexperiments discuss the manuscript coordinate and direct the project andwrite the manuscript AV-P is the guarantor of this work and as such had fullaccess to all the data in the study and takes responsibility for the integrity of thedata and the accuracy of the data analysis
References1 Montague CT OrsquoRahilly S The perils of portliness causes and con-
sequences of visceral adiposity Diabetes 200049883ndash8882 Carobbio S Rodriguez-Cuenca S Vidal-Puig A Origins of metabolic com-
plications in obesity ectopic fat accumulation The importance of the qualitative
aspect of lipotoxicity Curr Opin Clin Nutr Metab Care 201114520ndash5263 Chavez JA Summers SA Lipid oversupply selective insulin resistance
and lipotoxicity molecular mechanisms Biochim Biophys Acta 20101801
252ndash2654 Chavez JA Summers SA A ceramide-centric view of insulin resistance Cell
Metab 201215585ndash5945 Shah C Yang G Lee I Bielawski J Hannun YA Samad F Protection from
high fat diet-induced increase in ceramide in mice lacking plasminogen activator
inhibitor 1 J Biol Chem 200828313538ndash135486 Turner N Kowalski GM Leslie SJ et al Distinct patterns of tissue-specific
lipid accumulation during the induction of insulin resistance in mice by high-fat
feeding Diabetologia 2013561638ndash16487 Blachnio-Zabielska AU Koutsari C Tchkonia T Jensen MD Sphingolipid
content of human adipose tissue relationship to adiponectin and insulin re-
sistance Obesity (Silver Spring) 2012202341ndash23478 Błachnio-Zabielska AU Pułka M Baranowski M et al Ceramide metabo-
lism is affected by obesity and diabetes in human adipose tissue J Cell Physiol
2012227550ndash5579 Ternes P Franke S Zaumlhringer U Sperling P Heinz E Identification and
characterization of a sphingolipid delta 4-desaturase family J Biol Chem 2002
27725512ndash2551810 Mizutani Y Kihara A Igarashi Y Identification of the human sphingolipid C4-
hydroxylase hDES2 and its up-regulation during keratinocyte differentiation
FEBS Lett 200456393ndash9711 Ahn EH Schroeder JJ Induction of apoptosis by sphingosine sphinganine
and C(2)-ceramide in human colon cancer cells but not by C(2)-dihydroceramide
Anticancer Res 2010302881ndash288412 Kraveka JM Li L Szulc ZM et al Involvement of dihydroceramide desaturase
in cell cycle progression in human neuroblastoma cells J Biol Chem 2007282
16718ndash1672813 Siddique MM Li Y Wang L et al Ablation of dihydroceramide desaturase 1
a therapeutic target for the treatment of metabolic diseases simultaneously stim-
ulates anabolic and catabolic signaling Mol Cell Biol 2013332353ndash236914 Parks BW Nam E Org E et al Genetic control of obesity and gut microbiota
composition in response to high-fat high-sucrose diet in mice Cell Metab 2013
17141ndash152
diabetesdiabetesjournalsorg Barbarroja and Associates 1191
15 Bikman BT Guan Y Shui G et al Fenretinide prevents lipid-induced insulinresistance by blocking ceramide biosynthesis J Biol Chem 201228717426ndash1743716 Medina-Gomez G Gray SL Yetukuri L et al PPAR gamma 2 preventslipotoxicity by controlling adipose tissue expandability and peripheral lipid me-tabolism PLoS Genet 20073e6417 Roberts LD Virtue S Vidal-Puig A Nicholls AW Griffin JL Metabolic pheno-typing of a model of adipocyte differentiation Physiol Genomics 200939109ndash11918 Ramiacuterez-Zacariacuteas JL Castro-Muntildeozledo F Kuri-Harcuch W Quantitation ofadipose conversion and triglycerides by staining intracytoplasmic lipids with Oilred O Histochemistry 199297493ndash49719 Ye J Coulouris G Zaretskaya I Cutcutache I Rozen S Madden TL Primer-BLAST a tool to design target-specific primers for polymerase chain reactionBMC Bioinformatics 20121313420 Pfaffl MW Tichopad A Prgomet C Neuvians TP Determination of stablehousekeeping genes differentially regulated target genes and sample integrityBestKeepermdashExcel-based tool using pair-wise correlations Biotechnol Lett 200426509ndash51521 Triola G Fabriagraves G Llebaria A Synthesis of a cyclopropene analogue of cer-amide a potent inhibitor of dihydroceramide desaturase Angew Chem Int Ed Engl2001401960ndash1962
22 Tsuji K Satoh S Mitsutake S et al Evaluation of synthetic sphingolipidanalogs as ligands for peroxisome proliferator-activated receptors Bioorg MedChem Lett 2009191643ndash164623 Mamtani M Meikle PJ Kulkarni H et al Plasma dihydroceramide speciesassociate with waist circumference in Mexican American families Obesity (SilverSpring) 201422950ndash95624 Weir JM Wong G Barlow CK et al Plasma lipid profiling in a largepopulation-based cohort J Lipid Res 2013542898ndash290825 Curran J Meikle P Weir J Jowett J Deep sequencing in extended pedi-grees reveals a major rare non-synonymous variant influencing the de novoceramide synthesis pathway Late-breaking abstract presented at the AmericanSociety of Human Genetics 2013 Meeting 22ndash26 October 2013 at the BostonConvention and Exhibition Center Boston Massachusetts26 Idkowiak-Baldys J Takemoto JY Grilley MM Structure-function studies ofyeast C-4 sphingolipid long chain base hydroxylase Biochim Biophys Acta 2003161817ndash2427 Zhou W Ye XL Sun ZJ Ji XD Chen HX Xie D Overexpression of de-generative spermatocyte homolog 1 up-regulates the expression of cyclin D1 andenhances metastatic efficiency in esophageal carcinoma Eca109 cells MolCarcinog 200948886ndash894
1192 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015

DhCer Treatment3T3-L1 preadipocytes were treated at day 0 of differentia-tion with the induction cocktail and C2DhCer at 50 mmolLfor 3 days and a second batch of cells were exposed at day 3of differentiation and treated with C2DhCer at 50 mmolLfor 3 days All batches were taken until final differentiationat day 9
Human SamplesThe cohort included 28 MO and 6 nonobese subjects withno alterations to lipid or glucose metabolism as controlsubjects (Table 1) Approval for the study was given by theethics committee and all patients gave their informedconsent Visceral AT biopsy samples were obtained fromMO patients undergoing bariatric surgery (Scopinaro pro-cedure) or laparoscopic surgery (hiatus hernia repair orcholecystectomies) for the lean subjects
Western BlottingProtein extracts were prepared using the Nuclear andCytoplasmic Extraction Reagents Kit (Pierce) according tothe manufacturerrsquos instructions Immunoblots were incu-bated with the following antibodies PPARg CEBPb andCyclinA (Santa Cruz Biotechnology) CyclinB1 D1 D3 andE1 cdk4 AKT Ser473-pAKT p4442 MAPK phospho-p4442 MAPK Ser660-pHSL (p-hormone-sensitive lipase)Ser565-pHSL total HSL ATGL caveolin-1 plin1 glut4 adi-ponectin and AMPK (Cell Signaling Technology) and adfpadrp abhd5 and anti-b-actin (Abcam)
RT-PCRRNA was extracted using TRI Reagent (Sigma) and reversetranscribed to cDNA Real-time PCR using SYBRgreenwas performed according to the manufacturerrsquos instruc-tions (ABI) Primer sequences were obtained from PrimerBlast (19) Expression of genes was corrected by the
geometrical average of 18s b2m b-act and 36b4 usingBestkeeper (20)
Apoptosis Combined Annexin VPropidium IodideStainingViability was assessed by using an Annexin Vpropidiumiodide kit (Bender MedSystems) according to the manufac-turerrsquos recommendations Binding of fluorescein-conjugatedAnnexin V and propidium iodide was measured by FACSCalibur(BD Biosciences)
Cellular ProliferationCell viability was assessed using an colorimetric assay(Roche) following the protocol supplied by the manufac-turer Cell proliferation was analyzed by quantification ofthe incorporation of BrdU (Roche)
Boron-Dipyrromethene Staining Reactive OxygenStress Production and Mitochondrial ContentCells were incubated with boron-dipyrromethene (BODIPY)at 4degC or alternatively with 20 mmolL 2979-dichlorodihy-drofluorescein diacetate or 100 nmolL Mitotracker (Invi-trogen) at 37degC for 30 min and analyzed on a FACSCaliburcell analyzer
Cellular Oxygen ConsumptionCells were exposed to oligomycin (1 mmolL) C8-CPPC(09 mmolL) and antimycinrotenone (1 mmolL each)O2 consumption was measured using the XF24 analyzer(Seahorse Bioscience) for a period of 90 min
Whole-Mount Confocal MicroscopyGonadal AT was fixed in 4 paraformaldehyde Sampleswere incubated at 4degC with mouse Pref-1 (preadipocytefactor-1) or rabbit Ki67 primary antibodies Nuclei andneutral lipids were stained with Hoechst 33342 and BODIPY493503 respectively Sample examination was performedusing a Zeiss 510 confocal laser scanning microscope (CarlZeiss) Pref-1+ Ki67+ and BODIPY+ cells and total cells (upto 913 cellsfield) were automatically counted by usingImageJ software in three independent fields in each tissue
Light Microscopy AnalysisSamples for AT hematoxylin-eosin staining were preparedas described previously (16) Adipocyte sizes were mea-sured using Cell P (Olympus Soft Imaging SolutionsGmbH) Between 1000 and 3000 adipocytes from eachmouse were used to obtain the mean cell area
Luciferase Reporter AssayHuman embryonic kidney 293 cells were transfectedusing Lipofectamine LTX (Invitrogen) following themanufacturerrsquos instructions Seventy-five nanograms ofthe reporter plasmid (3xPPRE TKLuc) and 375 ng of theeukaryotic expression vector (pSV-PPARg) were cotrans-fected to each well The plasmid pRL-CMV (5 ngwell)was included as an internal control Twenty-four hourspost-transfection cells were treated as indicated (DMSOrosiglitazone 10 mmolL GW1929 10 mmolL andC2DhCer C16DhCer and Cer 100 mmolL) Luciferaseassays were performed using the dual luciferase reporterassay system (Promega)
Table 1mdashClinical characteristics of control subjects and MOpatients
Control subjects MO patients
Sex nMale 3 14Female 3 14
Age years 5217 6 51 4157 6 296
Weight kg 7400 6 071 1555 6 732
Height cm 16260 6 160 16553 6 249
BMI kgm2 2504 6 055 5651 6 168
Serum insulin IUmL 1123 6 158 3009 6 230
HOMA-IR 351 6 048 818 6 078
Serum glucose mmolL 603 6 032 574 6 026
Serum cholesterol mmolL 468 6 040 492 6 025
HDL cholesterol mmolL 097 6 023 110 6 009
Triglycerides mmolL 108 6 020 142 6 019
Values are reported as the mean 6 SEM unless otherwise in-dicated HOMA-IR HOMA of insulin resistance
1182 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
LipidomicsCells were mixed with 09 NaCl and sonicated for 5 minat 5degC and 40 kHz Samples were spiked with internalstandard The samples were extracted with chloroformmethanol (21) The lower phase was collected and mixedwith the labeled standard mixture (three stable isotope-labeled reference compounds) Lipid extracts were ana-lyzed on a Q-ToF Premier Mass Spectrometer (Waters)combined with an Acquity Ultra Performance LiquidChromatographyMass Spectrometry system (Waters)The data were processed using MZmine software Thelipids were quantified by normalizing with correspond-ing internal standard
Statistical AnalysisStudent t test (unpaired) ANOVA and Duncan test wereused for the statistical analysis Statistical significance wasset at P 005 and P 001 The Spearman correlationwas calculated to estimate the linear correlations betweenvariables at P 001
RESULTS
DEGS1 Is Downregulated in WAT in Obese MurineModelsTissue distribution analysis showed that degs1 is presentin most tissues and it is particularly highly expressed inAT liver and muscle whereas degs2 mRNA is detectedonly in intestine (Fig 1A) Degs1 expression in vivo waspositively correlated with WAT mass in lean healthy mice(Fig 1B) However this correlation was disrupted in mu-rine models of nutritional and genetically induced obesity(HFD and obob) where degs1 mRNA expression was de-creased versus controls (Fig 1C and F) Of note the dys-regulation of degs1 in obesity was limited to WAT as itsexpression in liver or skeletal muscle was not affected ineither HFD-fed mice (Fig 1D and E) or obob mice (Fig1G and H) DEGS1 was preferentially expressed in ma-tured adipocytes versus stromal vascular fraction in leanand obob mice (Supplementary Fig 1) and downregulatedin obob adipocytes versus lean adipocytes Moreover thisdownregulation of degs1 in total WAT of HFD-fed and obobmice was also recapitulated in visceral AT of MO patients(Supplementary Fig 2A)
To determine whether the decrease in degs1 expressionwas part of a global adaptation of de novo Cer synthesispathway in obesity the expression of other genes in thispathway was measured Expression of the sptlc1 and sptlc2subunits was not changed in human visceral AT or in obobWAT but sptlc2 expression was increased in HFD WAT(Supplementary Fig 2B and C) Moreover both HFD andobob mouse WAT exhibited a reduction in a subset of Cersynthases suggesting a possible defect in the synthesis ofa specific subset of Cers
We then confirmed that modulators of obesity-associatedinflammation may contribute to the downregulation ofdegs1 as treatment of 3T3-L1 adipocytes with tumornecrosis factor-a (5 and 10 ngmL) for 48 h resulted ina dose-dependent downregulation of degs1 mRNA
expression (388 6 36 and 457 6 73 reductionrespectively)
We then investigated a link between DEGS1 and theadipogenic program It is known that obese and insulin-resistant murine models and humans have reducedexpression of pparg2 in WAT Furthermore we observedthat expression of pparg2 and degs1 was directly corre-lated in 3T3-L1 after treatment with rosiglitazone (Sup-plementary Fig 3A) To validate this in vivo we analyzedthe regulation of degs1 in AT liver and skeletal muscle ofpparg2KO and POKO mice (16) WAT of pparg2KO andPOKO mice expressed significantly lower degs1 mRNAlevels compared with WT mice (Supplementary Fig 3B)This association was restricted to WAT since the expres-sion of degs1 in other metabolic organs such as skeletalmuscle or liver was not affected when pparg2 was absent(Supplementary Fig 3C and D)
DEGS1 Controls Important Cellular Functions Such asProliferation Survival and Oxidative Stress in 3T3-L1AdipocytesA stable degs1 knockdown 3T3-L1 cell line (65) (Fig 2A)resulted in inhibition of cell growth after 24 h (Fig 2B andC) Degs1 knockdown also induced cell death (137) andapoptosis (63) (Fig 2E) These were associated with a de-crease in cyclin-A and cdk2 levels in degs1 KD proliferat-ing cells (Fig 2F) as well as elevated levels of Bax andcaspase-3 mRNA (Fig 2D) Elevated reactive oxygen speciesproduction (Fig 2I) along with an upregulation of the ex-pression of antioxidant genes were observed in degs1 KDpreadipocytes (Fig 2H) This was accompanied by impairedmitochondrial oxygen consumption (Fig 2G) withoutchanges in the number of mitochondria (Fig 2J)
DEGS1 Is Required for Adipocyte DifferentiationExpression of degs1 is increased during normal differen-tiation of 3T3-L1 adipocytes (Fig 3A) To investigatewhether ablation of DEGS1 affected adipogenesis 3T3-L1degs1 KD cells were induced to differentiate At day 9degs1 KD cells showed impaired lipid accumulation versuscontrols (Fig 3B) Moreover pref1 mRNA expressiona marker for preadipocytes was not decreased in degs1KD cells at day 9 suggesting that a relevant number ofcells remained in the preadipocyte stage (Fig 3C) In ad-dition the expressions of proadipogenic and lipogenicgenes were downregulated (Fig 3D) Interestingly treat-ment with rosiglitazone only partially improved the dif-ferentiation and lipid accumulation rates of degs1 KD cells(Supplementary Fig 4A and B)
We tested whether DEGS1 expression may affect themitotic clonal expansion (MCE) In degs1 KD cells ppargand cebpb were downregulated compared with controls attime 0 and 24 h following differentiation induction (Sup-plementary Fig 5A and B) The expression of PPARg andseveral cyclins involved in adipogenesis were measured atearlier time points We noted a strong effect of degs1depletion on PPARg2 mRNA from time 0 Additionallyreduced levels of degs1 blocked MCE as indicated by the
diabetesdiabetesjournalsorg Barbarroja and Associates 1183
inhibition of cyclin D1 and cdk2 expression between time0 and 4 h after the induction (Supplementary Fig 5A andB) Cyclin E and cyclin D3 expression was also inhibitedbut at later time points
Pharmacological Inhibition of DEGS1 RecapitulatesDEGS1 KD Antiadipogenic PhenotypeUse of C8-CPPC a selective inhibitor of DEGS1 (21) fur-ther confirmed the relevance of DEGS1 in adipogenesisSimilarly to degs1 KD cells 3T3-L1 cells were induced todifferentiate and were treated simultaneously withC8-CPPC from time 0 h every 6 h for 48 h C8-CPPC de-creased the expression of proadipogenic transcription fac-tors such as PPARg and CEBPb as well as lipogenic genes(Fig 4A and C) The antiadipogenic effect of C8-CPPC per-sisted after rosiglitazone treatment (Fig 4A and C) Asexpected analysis of neutral lipids showed that 3T3-L1 cellstreated with C8-CPPC for 96 h accumulated significantlyless lipids compared with untreated cells (Fig 4D)
Pharmacological Inhibition of DEGS1 Also ImpairsAdipocyte Differentiation In VivoWe administered C8-CPPC intraperitoneally to mice fedan HFD for 5 weeks an experimental protocol that isknown to trigger adipocyte hyperplasia Molecular analysisof the WAT from mice treated with C8-CPPC presentedhigher levels of pref-1+ cells suggesting an increase in thenumber of preadipocytes compared with controls Inter-estingly a significant number of preadipocytes showedevidence of increased proliferation (Ki67+) whereas thenumber of differentiating preadipocytes (pref1+ BODIPY+)was significantly smaller in comparison with controls (Fig5A) These data were reinforced at the mRNA level byshowing increased expression of pref-1 and gata2 (Fig5B) and the presence of fewer small adipocytes in treatedmice (Supplementary Fig 6) Altogether these data sug-gest that C8-CPPCndashmediated inhibition of DEGS1 im-paired the capacity of preadipocytes to differentiate intoadipocytes in vivo
Figure 1mdashA Tissue distribution of degs1 B Correlation of degs1 mRNA expression and fat pad size The Spearman correlation coef-ficients were calculated to estimate the linear correlations between variables The rejection level for a null hypothesis was P lt 001 Dataare from 14 lean mice CndashE Expression levels of degs1 mRNA in response to HFD FndashH Expression levels of degs1 mRNA in obob miceValues are the mean 6 SEM for six to eight animals per group P lt 005 vs WT AU arbitrary units BAT brown AT gWAT gonadal WATIngFat inguinal fat RetroFat retroperitoneal fat SKM skeletal muscle
1184 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
DEGS1 Is Required for Lipid Accumulation BasalLipolysis and Glucose Uptake in Mature AdipocytesWe next focused on the effects of DEGS1 inhibition infully differentiated adipocytes by treating isolated matureadipocytesmdashfrom the gonadal WAT of C57BL6 micemdashwith C8-CPPC for 24 h C8-CPPC caused a decrease in theexpression of lipid metabolism genes as well as an increasein antioxidant genes (Supplementary Fig 7) recapitulatingour observations in 3T3-L1 preadipocytes
We further investigated the effects of inhibiting DEGS1on lipolytic activity and insulin signaling and measuredadiponectin expression as a representative fingerprintof global adipocyte homeostasis in mature differentiated3T3-L1 adipocytes Thus we observed that basal lipolyticactivity was decreased in mature differentiated 3T3-L1adipocytes when treated with C8-CPPC in nonstimulated
conditions (Fig 6A) Systematic evaluation of the lipolyticaxis showed that under basal conditions phosphorylation ofSer565 was also substantially increased by C8-CPPC con-comitantly with a minor downregulation of Ser660 phosp-horylation (Fig 6BndashD) suggesting a decreased HSL activityMoreover total levels of HSL were also decreased inC8-CPPCndashtreated cells These data suggest that pharmaco-logical inhibition of DEGS1 in mature adipocytes disruptsthe lipolytic response under nonstimulated conditions a de-fect that was superseded in the presence of NE
Finally we characterized the effects of pharmacologicalinhibition of DEGS1 on insulin signaling in 3T3-L1 matureadipocytes treated with C8-CPPC for 48 h as describedabove and subsequently incubated in the presence ofincreasing doses of insulin No major differences wereobserved in the phosphorylation of AKT in C8-CPPCndashtreated
Figure 2mdashA mRNA expression of degs1 in 3T3-L1 treated with short hairpin RNA (shRNA) against degs1 B and C Cell proliferation (XTTand BrdU assays every 24 h until 96 and 72 h respectively) D mRNA expression of Cdk2 Bcl2 Bax and Caspase3 E Apoptosis and celldeath rate F Protein expression of Cdk2 G Oxygen consumption rate H mRNA expression of antioxidant machinery genes I Reactiveoxygen species production J Mitochondria levels All these experiments were perfomed in degs1 KD and WT 3T3-L1 cells Values are themean 6 SEM of three separate experiments performed in triplicate ETC electron transport chain MFI mean fluorescence intensity PIpropidium iodide P lt 005
diabetesdiabetesjournalsorg Barbarroja and Associates 1185
cells after acute insulin stimulation (Fig 6E) Howeverwe found that Glut4 protein levels were increased inC8-CPPCndashtreated cells suggesting that glucose uptakemay be increased (Fig 6F) Interestingly we also did notobserve any differences in either adiponectin levels in re-sponse to C8-CPPC or in AMPK phosphorylation a knowninducer of adiponectin although increased levels of totalAMPK were observed (Fig 6G and H)
Pharmacological and Genetic Inhibition of DEGS1Increases DhCerCer Ratio in 3T3-L1 CellsWe confirmed that C8-CPPC increased the DhCerCerratio in 3T3L-1 preadipocytes upon the inhibition ofDEGS1 (Fig 4E) Similarly degs1 KD preadipocytes (day 0)exhibited an increased DhCerCer ratio versus controls(Supplementary Fig 8A) Downregulation of degs1
expression in degs1 KD cells was accompanied by a down-regulation of serine palmitoyltransferase (sptlc1) and Cersynthase 6 (Cers6) (Supplementary Fig 8A) suggestive ofa homeostatic readjustment of the biosynthetic pathwayto compensate for the dysfunction of degs1 Other genesrelated to Cer homeostasis such as neutral ceramidase(ncdase) and glycosylceramide synthase (gcs) were alsodownregulated
Curiously at day 9 degs1 KD cells despite impaireddifferentiation showed a complete restoration of theDhCerCer ratio (indicated by the normalization of theCer pool) (Supplementary Fig 8B) This could be partiallyexplained by a compensation mechanism mediated bydownregulation of neutral ceramidases and the conversionof sphyngomyelins to Cers (upregulation of sphyngomye-linase 2) (Supplementary Fig 8B)
Figure 3mdashA mRNA expression of degs1 in 3T3-L1 cells during adipocyte differentiation B Lipid accumulation at day 9 of differentiationOil Red O staining C mRNA expression of PREF1 in WT and degs1 KD cells during differentiation D mRNA expression of genes involvedin adipocyte differentiation and lipid accumulation Values are the mean 6 SEM of three separate experiments performed in triplicate FASfatty acid synthase Insig insulin-induced gene LPL lipoprotein lipase P lt 005
1186 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
The Impairment of Adipocyte Differentiation DuringEarly Adipogenesis by DEGS1 Inhibition IsRecapitulated by DhCers Per SeWe finally sought to validate whether DhCer by itself couldmediate the effects of DEGS1 inhibition on adipocytedifferentiation in our models of genetic or pharmacologicalinhibition of degs1 3T3-L1cells were treated with C2DhCerduring 1) the MCE at time 0 and 2) at day 3 after differ-entiation DhCer inhibited lipid accumulation (Fig 7A) andthe expression of genes involved during the early stages ofadipogenesis (Fig 7B)
Since synthetic sphingolipid analogs are known tomodulate PPAR activity (22) we investigated whetherboth C2DhCer and C16DhCer could exert a repressiveeffect on PPARg activation in the presence of the PPARgligands (Fig 7C) Our results show that DhCer can also
block the ligand-mediated transactivation of PPARg Sim-ilar results were obtained with Cers indicating that theseeffects were not a consequence of the unsaturation of thesphingoid moiety (data not shown)
DISCUSSION
The DhCerCer ratio has an important homeostaticregulatory role in the cell contributing to cell survivalautophagy and oxidative stress (1213) Here we identifyDEGS1 as an essential metabolic enzyme which is dysreg-ulated in obese states and contributes to AT dysfunctionOur complementary in vitro and in vivo approaches revealthat reduced DEGS1 function impairs adipogenesis andlipogenesis programs and increases oxidative stress
DEGS1 is expressed in WAT and correlates in rodentswith fat mass in healthy states an association that is
Figure 4mdash3T3-L1 cells treated with C8-CPPC andor rosiglitazone A and B mRNA expression of genes involved in adipocyte differen-tiation and lipid accumulation at 24 and 48 h after induction C Protein expression levels of PPARg2 and cEBPa D Lipid content after 48and 96 h of differentiation Values are the mean 6 SEM of two separate experiments performed in triplicate One-way ANOVA was used toanalyze the statistical significance between treatments at 24 and 48 h Significant differences (Duncan test P lt 005) are indicated withdifferent letters E DhCer Cer and hexosylceramide (HexCer) levels after 48 h of differentiation Values are the mean 6 SEM of twoseparate experiments performed in triplicate cEBP CCAATenhancer binding protein d day FITC fluorescein isothiocyanate MFI meanfluorescence intensity Rosi rosiglitazone P lt 005
diabetesdiabetesjournalsorg Barbarroja and Associates 1187
disrupted in severe forms of obesity The expression ofDEGS1 in liver or muscle is not affected by obesity fur-ther indicating that the WAT is particularly susceptible tochanges in Cer metabolism in obese states The relevanceto human obesity is confirmed by the downregulation ofDEGS1 in the AT of MO patients and by recent reports(2324) showing that DhCer rather than Cer content pos-itively correlates with BMI and waist circumference incohorts of overweight obese subjects To date only a singlefunctional mutation in the degs1 gene has been reportedin humans where patients demonstrate increased serumDhCer decreased cholesterol esters and decreased waist-to-hip ratio (25) Globally considered these data suggest
the existence of an association between DEGS1 functionand fat mass
The molecular mechanism leading to the defective ATexpression of degs1 in human and rodent obesity is un-known Here we have shown that tumor necrosis factor-aa mediator of obesity-induced inflammation and insulinresistance decreases the transcriptional expression ofdegs1 in cultured adipocytes which suggests that lowchronic inflammation may contribute to DEGS1 dysfunc-tion Other obesity-associated processes such as in-creased oxidative stress and hypoxia have been shownto impair DEGS1 activity and increase DhCer levels (26)Our results also show a positive association between
Figure 5mdashC8-CPPC inhibitor increases proliferation and decreases lipid accumulation in preadipocytes in vivo Ten-week-old mice weregiven a 45 HFD for 5 weeks before being treated intraperitoneally with vehicle or C8-CPPC inhibitor (2 mgkgday) over 9 daysA Representative images of immunofluorescence analysis of gonadal AT of control (vehicle) and C8-CPPCndashtreated mice are presentedwith Pref-1 (cyan) and Ki67 (red white arrows) Nuclei and lipids are respectively stained with Hoechst stain (blue) and BODIPY (green)Scale bars 100 or 20 mm Quantification of preadipocytes (left graph) (percentage of Pref-1+ cellstotal cells) and (right graph) Ki67+ (whitebars)BODIPY+ (dark gray bars) cells among preadipocytes (Pref-1+ cells) (light gray bars) B mRNA expression of preadipocyte markersand pparg degs1 adiponectin caspase 3 and fas in gonadal AT of control and C8-CPPCndashtreated mice n = 7ndash8 mice per experimentalgroup Values are the mean 6SEM P lt 005 P lt 00001 vs vehicle
1188 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
Figure 6mdashA Glycerol release in 3T3-L1 adipocytes exposed for 48 h to C8-CPPC and stimulated with NE for 6 h BndashD Results fromWestern blot analysis for phosphorylated (p) HSLHSL ratio and total HSL in 3T3-L1 adipocytes exposed for 48 h to C8-CPPC andstimulated with NE for 6 h Graphs show the mean 6 SEM of two separated experiments P lt 005 E Effects of pharmacologicalinhibition of DEGS1 on insulin (ins) signaling Results from Western blot analysis for p-AKT and AKT in 3T3-L1 adipocytes exposed 48 or 72h with C8-CPPC 1 mmolL and increased concentrations of insulin (10ndash100 nmolL) for 15 min F and G Results from Western blot analysisfor Glut4 and adiponectin in 3T3-L1 adipocytes exposed with C8-CPPC 05ndash1 mmolL and insulin (100 nmolL) for 48 h H Results fromWestern blot analysis for AMPK in 3T3-L1 adipocytes exposed to C8-CPPC 05ndash1 mmolL and insulin (Ins) (100 nmolL) rosiglitazone (Rosi)(1 nmolL) and metformin (Metf) (100 nmolL) for 48 h aP lt 005 vs untreated cells (Ins Rosi or Metf effect) bP lt 005 vs 0 mmolLC8-CPPC (C8-CPPC effect) AU arbitrary units IOD integrated optical density
diabetesdiabetesjournalsorg Barbarroja and Associates 1189
levels of the proadipogenic pparg2 isoform and degs1in AT and given the documented downregulation ofpparg2 in AT in obese insulin-resistant states it is con-ceivable that decreased levels of pparg2 may contributeto the defective expression of degs1 associated withobesity
Studies in rodents have shown that the number of Cersis increased in the WAT of animals fed an HFD (5) con-comitantly with the onset of insulin resistance (6) Resultsfrom obob mice and obese human studies are more un-clear where both increased or decreased levels of Cershave been reported (78) This suggests that Cer synthesismay be dependent on the severity of the obese state andthe strength of the adaptive homeostatic responsesattempting to restore their normal balance
Our pharmacological and gene ablation studies haveshown that inhibition of DEGS1 is associated with oxida-tive stress cellular death impaired adipocyte differentia-tion impaired lipid accumulation and impaired basallipolysis in mature adipocytes
Our in vitro studies revealed that the defect in theadipogenesis program caused by depletion of DEGS1occurs during the early stages of adipogenesis (MCE)when PPARg controls the expression of genes that regu-late the cell cycle by directly interacting with cyclin D3Another cyclin involved in adipocyte differentiation iscyclin D1 and here we show that downregulation ofDEGS1 blocks PPARg activity and cyclins such as D1D3 E as well as decreasing cdk2 which modulates PPARgactivity during adipogenesis These data are consistent
Figure 7mdashA and B DhCers impair adipocyte differentiation during early adipogenesis 3T3-L1 preadipocytes treated with C2DhCer(50 mmolL) for a period of 48 h at day 0 or day 3 of differentiation Oil Red O staining and mRNA expression of adipogenesis and lipidmetabolism genes Values are the mean 6 SEM of two separate experiments performed in triplicate P lt 005 vs control C DhCersdecrease ligand-mediated PPARg transactivation Cells were treated with DMSO as a control group and rosiglitazone 10 mmolL GW192910 mmolL and C2 and C16DhCers and Cers 100 mmolL as indicated Graphs represent the average of three independent experimentsP lt 005 vs control cells P lt 005 vs rosiglitazone ampP lt 005 vs GW1929 CTL control
1190 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
with studies showing that downregulation of degs1 bysmall interfering RNA inhibits cell growth and arreststhe cell cycle in cancer cells (12) and where its overexpres-sion increased cell migration and metastasis (27)
The pharmacological inhibition of DEGS1 in vivovalidated the results obtained in vitro and strengthenedthe concept that functional degs1 is required for the fulldifferentiation of preadipocytes into mature adipocytes
We also provide evidence that both pharmacologicaland genetic ablation of degs1 which according to ourresults in in vitro models may directly block adipocytedifferentiation and lipid deposition certainly increasedDhCer levels in in vitro models In this line our resultsshowed that the treatment of adipocytes with DhCercauses impaired adipogenesis and lipid accumulationThis effect on lipid accumulation was more severewhen the treatment was administered during the first3 days of differentiation suggesting a direct impact ofDhCer accumulation during early stages of adipogenesisWe also provide evidence showing that accumulation ofDhCers can also directly repress the transcriptional ac-tivity of PPARg and hence could contribute to impair-ment of the capacity of the preadipocytes to developa full adipogenic program
These observations strengthen the concept that DhCersare not merely inert precursors of Cers Nevertheless themolecular mechanism linking DhCers accumulation anddysregulation of the cell cycle and other cellular eventsremains elusive One possible mechanism is that changesin the DhCerCer ratio may disrupt membrane-dependentstructures by altering the levels of cholesterol andorcaveolin in lipid rafts which are known to be highlysensitive to a sphingomyelin pool Alternatively changesin the DhCerCer ratio may disturb the global phospho-lipidome potentially altering membrane-associated pro-cesses relevant to adipocyte function These are importantquestions to address in future research
In summary our results indicate that defects in DEGS1in the context of the metabolic syndrome may compro-mise AT expansion and function through the combinedinhibition of adipogenesis promotion of cell death andoxidative stress due to the direct accumulation of DhCersThus our data suggest that the selective manipulation ofDEGS1 andor its substrates in WAT may be a pathophy-siologically advantageous strategy to improve AT homeo-stasis and ameliorate the burden of obesity-associatedmetabolic complications
Acknowledgments The authors thank Dr Carobbio Mrs Peirce andMs Philips (Wellcome Trust-Medical Research Council Institute of MetabolicScience University of Cambridge UK) for their excellent technical advice sup-port and suggestionsFunding This work was funded by the Medical Research Council (MRC) theMetabolic Diseases Unit of the MRC European Commission grant FP7-ETHERPATHSand the British Heart FoundationDuality of Interest No potential conflicts of interest relevant to this articlewere reported
Author Contributions NB helped to develop the hypothesis design theexperiments perform the in vitro and ex vivo experiments collect and analyze thedata hold and characterize the human cohorts collect gene expression data fromadipose depots and analyze the data discuss the manuscript coordinate anddirect the project and write the manuscript SR-C helped to develop thehypothesis design the experiments perform the in vitro and ex vivo experimentscollect and analyze the data discuss the manuscript coordinate and direct theproject and write the manuscript HN and MO helped to develop analyticalplatforms perform and analyze lipidomic experiments and discuss the manu-script AC AP JR IC VP GM-G and CL-P helped to perform the invitro and ex vivo experiments collect and analyze the data and discuss themanuscript FJT helped to hold and characterize the human cohorts collectgene expression data from adipose depots and analyze the data and discussthe manuscript JDS and SAS helped to design the experiments and dis-cuss the manuscript AV-P helped to develop the hypothesis design theexperiments discuss the manuscript coordinate and direct the project andwrite the manuscript AV-P is the guarantor of this work and as such had fullaccess to all the data in the study and takes responsibility for the integrity of thedata and the accuracy of the data analysis
References1 Montague CT OrsquoRahilly S The perils of portliness causes and con-
sequences of visceral adiposity Diabetes 200049883ndash8882 Carobbio S Rodriguez-Cuenca S Vidal-Puig A Origins of metabolic com-
plications in obesity ectopic fat accumulation The importance of the qualitative
aspect of lipotoxicity Curr Opin Clin Nutr Metab Care 201114520ndash5263 Chavez JA Summers SA Lipid oversupply selective insulin resistance
and lipotoxicity molecular mechanisms Biochim Biophys Acta 20101801
252ndash2654 Chavez JA Summers SA A ceramide-centric view of insulin resistance Cell
Metab 201215585ndash5945 Shah C Yang G Lee I Bielawski J Hannun YA Samad F Protection from
high fat diet-induced increase in ceramide in mice lacking plasminogen activator
inhibitor 1 J Biol Chem 200828313538ndash135486 Turner N Kowalski GM Leslie SJ et al Distinct patterns of tissue-specific
lipid accumulation during the induction of insulin resistance in mice by high-fat
feeding Diabetologia 2013561638ndash16487 Blachnio-Zabielska AU Koutsari C Tchkonia T Jensen MD Sphingolipid
content of human adipose tissue relationship to adiponectin and insulin re-
sistance Obesity (Silver Spring) 2012202341ndash23478 Błachnio-Zabielska AU Pułka M Baranowski M et al Ceramide metabo-
lism is affected by obesity and diabetes in human adipose tissue J Cell Physiol
2012227550ndash5579 Ternes P Franke S Zaumlhringer U Sperling P Heinz E Identification and
characterization of a sphingolipid delta 4-desaturase family J Biol Chem 2002
27725512ndash2551810 Mizutani Y Kihara A Igarashi Y Identification of the human sphingolipid C4-
hydroxylase hDES2 and its up-regulation during keratinocyte differentiation
FEBS Lett 200456393ndash9711 Ahn EH Schroeder JJ Induction of apoptosis by sphingosine sphinganine
and C(2)-ceramide in human colon cancer cells but not by C(2)-dihydroceramide
Anticancer Res 2010302881ndash288412 Kraveka JM Li L Szulc ZM et al Involvement of dihydroceramide desaturase
in cell cycle progression in human neuroblastoma cells J Biol Chem 2007282
16718ndash1672813 Siddique MM Li Y Wang L et al Ablation of dihydroceramide desaturase 1
a therapeutic target for the treatment of metabolic diseases simultaneously stim-
ulates anabolic and catabolic signaling Mol Cell Biol 2013332353ndash236914 Parks BW Nam E Org E et al Genetic control of obesity and gut microbiota
composition in response to high-fat high-sucrose diet in mice Cell Metab 2013
17141ndash152
diabetesdiabetesjournalsorg Barbarroja and Associates 1191
15 Bikman BT Guan Y Shui G et al Fenretinide prevents lipid-induced insulinresistance by blocking ceramide biosynthesis J Biol Chem 201228717426ndash1743716 Medina-Gomez G Gray SL Yetukuri L et al PPAR gamma 2 preventslipotoxicity by controlling adipose tissue expandability and peripheral lipid me-tabolism PLoS Genet 20073e6417 Roberts LD Virtue S Vidal-Puig A Nicholls AW Griffin JL Metabolic pheno-typing of a model of adipocyte differentiation Physiol Genomics 200939109ndash11918 Ramiacuterez-Zacariacuteas JL Castro-Muntildeozledo F Kuri-Harcuch W Quantitation ofadipose conversion and triglycerides by staining intracytoplasmic lipids with Oilred O Histochemistry 199297493ndash49719 Ye J Coulouris G Zaretskaya I Cutcutache I Rozen S Madden TL Primer-BLAST a tool to design target-specific primers for polymerase chain reactionBMC Bioinformatics 20121313420 Pfaffl MW Tichopad A Prgomet C Neuvians TP Determination of stablehousekeeping genes differentially regulated target genes and sample integrityBestKeepermdashExcel-based tool using pair-wise correlations Biotechnol Lett 200426509ndash51521 Triola G Fabriagraves G Llebaria A Synthesis of a cyclopropene analogue of cer-amide a potent inhibitor of dihydroceramide desaturase Angew Chem Int Ed Engl2001401960ndash1962
22 Tsuji K Satoh S Mitsutake S et al Evaluation of synthetic sphingolipidanalogs as ligands for peroxisome proliferator-activated receptors Bioorg MedChem Lett 2009191643ndash164623 Mamtani M Meikle PJ Kulkarni H et al Plasma dihydroceramide speciesassociate with waist circumference in Mexican American families Obesity (SilverSpring) 201422950ndash95624 Weir JM Wong G Barlow CK et al Plasma lipid profiling in a largepopulation-based cohort J Lipid Res 2013542898ndash290825 Curran J Meikle P Weir J Jowett J Deep sequencing in extended pedi-grees reveals a major rare non-synonymous variant influencing the de novoceramide synthesis pathway Late-breaking abstract presented at the AmericanSociety of Human Genetics 2013 Meeting 22ndash26 October 2013 at the BostonConvention and Exhibition Center Boston Massachusetts26 Idkowiak-Baldys J Takemoto JY Grilley MM Structure-function studies ofyeast C-4 sphingolipid long chain base hydroxylase Biochim Biophys Acta 2003161817ndash2427 Zhou W Ye XL Sun ZJ Ji XD Chen HX Xie D Overexpression of de-generative spermatocyte homolog 1 up-regulates the expression of cyclin D1 andenhances metastatic efficiency in esophageal carcinoma Eca109 cells MolCarcinog 200948886ndash894
1192 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015

LipidomicsCells were mixed with 09 NaCl and sonicated for 5 minat 5degC and 40 kHz Samples were spiked with internalstandard The samples were extracted with chloroformmethanol (21) The lower phase was collected and mixedwith the labeled standard mixture (three stable isotope-labeled reference compounds) Lipid extracts were ana-lyzed on a Q-ToF Premier Mass Spectrometer (Waters)combined with an Acquity Ultra Performance LiquidChromatographyMass Spectrometry system (Waters)The data were processed using MZmine software Thelipids were quantified by normalizing with correspond-ing internal standard
Statistical AnalysisStudent t test (unpaired) ANOVA and Duncan test wereused for the statistical analysis Statistical significance wasset at P 005 and P 001 The Spearman correlationwas calculated to estimate the linear correlations betweenvariables at P 001
RESULTS
DEGS1 Is Downregulated in WAT in Obese MurineModelsTissue distribution analysis showed that degs1 is presentin most tissues and it is particularly highly expressed inAT liver and muscle whereas degs2 mRNA is detectedonly in intestine (Fig 1A) Degs1 expression in vivo waspositively correlated with WAT mass in lean healthy mice(Fig 1B) However this correlation was disrupted in mu-rine models of nutritional and genetically induced obesity(HFD and obob) where degs1 mRNA expression was de-creased versus controls (Fig 1C and F) Of note the dys-regulation of degs1 in obesity was limited to WAT as itsexpression in liver or skeletal muscle was not affected ineither HFD-fed mice (Fig 1D and E) or obob mice (Fig1G and H) DEGS1 was preferentially expressed in ma-tured adipocytes versus stromal vascular fraction in leanand obob mice (Supplementary Fig 1) and downregulatedin obob adipocytes versus lean adipocytes Moreover thisdownregulation of degs1 in total WAT of HFD-fed and obobmice was also recapitulated in visceral AT of MO patients(Supplementary Fig 2A)
To determine whether the decrease in degs1 expressionwas part of a global adaptation of de novo Cer synthesispathway in obesity the expression of other genes in thispathway was measured Expression of the sptlc1 and sptlc2subunits was not changed in human visceral AT or in obobWAT but sptlc2 expression was increased in HFD WAT(Supplementary Fig 2B and C) Moreover both HFD andobob mouse WAT exhibited a reduction in a subset of Cersynthases suggesting a possible defect in the synthesis ofa specific subset of Cers
We then confirmed that modulators of obesity-associatedinflammation may contribute to the downregulation ofdegs1 as treatment of 3T3-L1 adipocytes with tumornecrosis factor-a (5 and 10 ngmL) for 48 h resulted ina dose-dependent downregulation of degs1 mRNA
expression (388 6 36 and 457 6 73 reductionrespectively)
We then investigated a link between DEGS1 and theadipogenic program It is known that obese and insulin-resistant murine models and humans have reducedexpression of pparg2 in WAT Furthermore we observedthat expression of pparg2 and degs1 was directly corre-lated in 3T3-L1 after treatment with rosiglitazone (Sup-plementary Fig 3A) To validate this in vivo we analyzedthe regulation of degs1 in AT liver and skeletal muscle ofpparg2KO and POKO mice (16) WAT of pparg2KO andPOKO mice expressed significantly lower degs1 mRNAlevels compared with WT mice (Supplementary Fig 3B)This association was restricted to WAT since the expres-sion of degs1 in other metabolic organs such as skeletalmuscle or liver was not affected when pparg2 was absent(Supplementary Fig 3C and D)
DEGS1 Controls Important Cellular Functions Such asProliferation Survival and Oxidative Stress in 3T3-L1AdipocytesA stable degs1 knockdown 3T3-L1 cell line (65) (Fig 2A)resulted in inhibition of cell growth after 24 h (Fig 2B andC) Degs1 knockdown also induced cell death (137) andapoptosis (63) (Fig 2E) These were associated with a de-crease in cyclin-A and cdk2 levels in degs1 KD proliferat-ing cells (Fig 2F) as well as elevated levels of Bax andcaspase-3 mRNA (Fig 2D) Elevated reactive oxygen speciesproduction (Fig 2I) along with an upregulation of the ex-pression of antioxidant genes were observed in degs1 KDpreadipocytes (Fig 2H) This was accompanied by impairedmitochondrial oxygen consumption (Fig 2G) withoutchanges in the number of mitochondria (Fig 2J)
DEGS1 Is Required for Adipocyte DifferentiationExpression of degs1 is increased during normal differen-tiation of 3T3-L1 adipocytes (Fig 3A) To investigatewhether ablation of DEGS1 affected adipogenesis 3T3-L1degs1 KD cells were induced to differentiate At day 9degs1 KD cells showed impaired lipid accumulation versuscontrols (Fig 3B) Moreover pref1 mRNA expressiona marker for preadipocytes was not decreased in degs1KD cells at day 9 suggesting that a relevant number ofcells remained in the preadipocyte stage (Fig 3C) In ad-dition the expressions of proadipogenic and lipogenicgenes were downregulated (Fig 3D) Interestingly treat-ment with rosiglitazone only partially improved the dif-ferentiation and lipid accumulation rates of degs1 KD cells(Supplementary Fig 4A and B)
We tested whether DEGS1 expression may affect themitotic clonal expansion (MCE) In degs1 KD cells ppargand cebpb were downregulated compared with controls attime 0 and 24 h following differentiation induction (Sup-plementary Fig 5A and B) The expression of PPARg andseveral cyclins involved in adipogenesis were measured atearlier time points We noted a strong effect of degs1depletion on PPARg2 mRNA from time 0 Additionallyreduced levels of degs1 blocked MCE as indicated by the
diabetesdiabetesjournalsorg Barbarroja and Associates 1183
inhibition of cyclin D1 and cdk2 expression between time0 and 4 h after the induction (Supplementary Fig 5A andB) Cyclin E and cyclin D3 expression was also inhibitedbut at later time points
Pharmacological Inhibition of DEGS1 RecapitulatesDEGS1 KD Antiadipogenic PhenotypeUse of C8-CPPC a selective inhibitor of DEGS1 (21) fur-ther confirmed the relevance of DEGS1 in adipogenesisSimilarly to degs1 KD cells 3T3-L1 cells were induced todifferentiate and were treated simultaneously withC8-CPPC from time 0 h every 6 h for 48 h C8-CPPC de-creased the expression of proadipogenic transcription fac-tors such as PPARg and CEBPb as well as lipogenic genes(Fig 4A and C) The antiadipogenic effect of C8-CPPC per-sisted after rosiglitazone treatment (Fig 4A and C) Asexpected analysis of neutral lipids showed that 3T3-L1 cellstreated with C8-CPPC for 96 h accumulated significantlyless lipids compared with untreated cells (Fig 4D)
Pharmacological Inhibition of DEGS1 Also ImpairsAdipocyte Differentiation In VivoWe administered C8-CPPC intraperitoneally to mice fedan HFD for 5 weeks an experimental protocol that isknown to trigger adipocyte hyperplasia Molecular analysisof the WAT from mice treated with C8-CPPC presentedhigher levels of pref-1+ cells suggesting an increase in thenumber of preadipocytes compared with controls Inter-estingly a significant number of preadipocytes showedevidence of increased proliferation (Ki67+) whereas thenumber of differentiating preadipocytes (pref1+ BODIPY+)was significantly smaller in comparison with controls (Fig5A) These data were reinforced at the mRNA level byshowing increased expression of pref-1 and gata2 (Fig5B) and the presence of fewer small adipocytes in treatedmice (Supplementary Fig 6) Altogether these data sug-gest that C8-CPPCndashmediated inhibition of DEGS1 im-paired the capacity of preadipocytes to differentiate intoadipocytes in vivo
Figure 1mdashA Tissue distribution of degs1 B Correlation of degs1 mRNA expression and fat pad size The Spearman correlation coef-ficients were calculated to estimate the linear correlations between variables The rejection level for a null hypothesis was P lt 001 Dataare from 14 lean mice CndashE Expression levels of degs1 mRNA in response to HFD FndashH Expression levels of degs1 mRNA in obob miceValues are the mean 6 SEM for six to eight animals per group P lt 005 vs WT AU arbitrary units BAT brown AT gWAT gonadal WATIngFat inguinal fat RetroFat retroperitoneal fat SKM skeletal muscle
1184 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
DEGS1 Is Required for Lipid Accumulation BasalLipolysis and Glucose Uptake in Mature AdipocytesWe next focused on the effects of DEGS1 inhibition infully differentiated adipocytes by treating isolated matureadipocytesmdashfrom the gonadal WAT of C57BL6 micemdashwith C8-CPPC for 24 h C8-CPPC caused a decrease in theexpression of lipid metabolism genes as well as an increasein antioxidant genes (Supplementary Fig 7) recapitulatingour observations in 3T3-L1 preadipocytes
We further investigated the effects of inhibiting DEGS1on lipolytic activity and insulin signaling and measuredadiponectin expression as a representative fingerprintof global adipocyte homeostasis in mature differentiated3T3-L1 adipocytes Thus we observed that basal lipolyticactivity was decreased in mature differentiated 3T3-L1adipocytes when treated with C8-CPPC in nonstimulated
conditions (Fig 6A) Systematic evaluation of the lipolyticaxis showed that under basal conditions phosphorylation ofSer565 was also substantially increased by C8-CPPC con-comitantly with a minor downregulation of Ser660 phosp-horylation (Fig 6BndashD) suggesting a decreased HSL activityMoreover total levels of HSL were also decreased inC8-CPPCndashtreated cells These data suggest that pharmaco-logical inhibition of DEGS1 in mature adipocytes disruptsthe lipolytic response under nonstimulated conditions a de-fect that was superseded in the presence of NE
Finally we characterized the effects of pharmacologicalinhibition of DEGS1 on insulin signaling in 3T3-L1 matureadipocytes treated with C8-CPPC for 48 h as describedabove and subsequently incubated in the presence ofincreasing doses of insulin No major differences wereobserved in the phosphorylation of AKT in C8-CPPCndashtreated
Figure 2mdashA mRNA expression of degs1 in 3T3-L1 treated with short hairpin RNA (shRNA) against degs1 B and C Cell proliferation (XTTand BrdU assays every 24 h until 96 and 72 h respectively) D mRNA expression of Cdk2 Bcl2 Bax and Caspase3 E Apoptosis and celldeath rate F Protein expression of Cdk2 G Oxygen consumption rate H mRNA expression of antioxidant machinery genes I Reactiveoxygen species production J Mitochondria levels All these experiments were perfomed in degs1 KD and WT 3T3-L1 cells Values are themean 6 SEM of three separate experiments performed in triplicate ETC electron transport chain MFI mean fluorescence intensity PIpropidium iodide P lt 005
diabetesdiabetesjournalsorg Barbarroja and Associates 1185
cells after acute insulin stimulation (Fig 6E) Howeverwe found that Glut4 protein levels were increased inC8-CPPCndashtreated cells suggesting that glucose uptakemay be increased (Fig 6F) Interestingly we also did notobserve any differences in either adiponectin levels in re-sponse to C8-CPPC or in AMPK phosphorylation a knowninducer of adiponectin although increased levels of totalAMPK were observed (Fig 6G and H)
Pharmacological and Genetic Inhibition of DEGS1Increases DhCerCer Ratio in 3T3-L1 CellsWe confirmed that C8-CPPC increased the DhCerCerratio in 3T3L-1 preadipocytes upon the inhibition ofDEGS1 (Fig 4E) Similarly degs1 KD preadipocytes (day 0)exhibited an increased DhCerCer ratio versus controls(Supplementary Fig 8A) Downregulation of degs1
expression in degs1 KD cells was accompanied by a down-regulation of serine palmitoyltransferase (sptlc1) and Cersynthase 6 (Cers6) (Supplementary Fig 8A) suggestive ofa homeostatic readjustment of the biosynthetic pathwayto compensate for the dysfunction of degs1 Other genesrelated to Cer homeostasis such as neutral ceramidase(ncdase) and glycosylceramide synthase (gcs) were alsodownregulated
Curiously at day 9 degs1 KD cells despite impaireddifferentiation showed a complete restoration of theDhCerCer ratio (indicated by the normalization of theCer pool) (Supplementary Fig 8B) This could be partiallyexplained by a compensation mechanism mediated bydownregulation of neutral ceramidases and the conversionof sphyngomyelins to Cers (upregulation of sphyngomye-linase 2) (Supplementary Fig 8B)
Figure 3mdashA mRNA expression of degs1 in 3T3-L1 cells during adipocyte differentiation B Lipid accumulation at day 9 of differentiationOil Red O staining C mRNA expression of PREF1 in WT and degs1 KD cells during differentiation D mRNA expression of genes involvedin adipocyte differentiation and lipid accumulation Values are the mean 6 SEM of three separate experiments performed in triplicate FASfatty acid synthase Insig insulin-induced gene LPL lipoprotein lipase P lt 005
1186 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
The Impairment of Adipocyte Differentiation DuringEarly Adipogenesis by DEGS1 Inhibition IsRecapitulated by DhCers Per SeWe finally sought to validate whether DhCer by itself couldmediate the effects of DEGS1 inhibition on adipocytedifferentiation in our models of genetic or pharmacologicalinhibition of degs1 3T3-L1cells were treated with C2DhCerduring 1) the MCE at time 0 and 2) at day 3 after differ-entiation DhCer inhibited lipid accumulation (Fig 7A) andthe expression of genes involved during the early stages ofadipogenesis (Fig 7B)
Since synthetic sphingolipid analogs are known tomodulate PPAR activity (22) we investigated whetherboth C2DhCer and C16DhCer could exert a repressiveeffect on PPARg activation in the presence of the PPARgligands (Fig 7C) Our results show that DhCer can also
block the ligand-mediated transactivation of PPARg Sim-ilar results were obtained with Cers indicating that theseeffects were not a consequence of the unsaturation of thesphingoid moiety (data not shown)
DISCUSSION
The DhCerCer ratio has an important homeostaticregulatory role in the cell contributing to cell survivalautophagy and oxidative stress (1213) Here we identifyDEGS1 as an essential metabolic enzyme which is dysreg-ulated in obese states and contributes to AT dysfunctionOur complementary in vitro and in vivo approaches revealthat reduced DEGS1 function impairs adipogenesis andlipogenesis programs and increases oxidative stress
DEGS1 is expressed in WAT and correlates in rodentswith fat mass in healthy states an association that is
Figure 4mdash3T3-L1 cells treated with C8-CPPC andor rosiglitazone A and B mRNA expression of genes involved in adipocyte differen-tiation and lipid accumulation at 24 and 48 h after induction C Protein expression levels of PPARg2 and cEBPa D Lipid content after 48and 96 h of differentiation Values are the mean 6 SEM of two separate experiments performed in triplicate One-way ANOVA was used toanalyze the statistical significance between treatments at 24 and 48 h Significant differences (Duncan test P lt 005) are indicated withdifferent letters E DhCer Cer and hexosylceramide (HexCer) levels after 48 h of differentiation Values are the mean 6 SEM of twoseparate experiments performed in triplicate cEBP CCAATenhancer binding protein d day FITC fluorescein isothiocyanate MFI meanfluorescence intensity Rosi rosiglitazone P lt 005
diabetesdiabetesjournalsorg Barbarroja and Associates 1187
disrupted in severe forms of obesity The expression ofDEGS1 in liver or muscle is not affected by obesity fur-ther indicating that the WAT is particularly susceptible tochanges in Cer metabolism in obese states The relevanceto human obesity is confirmed by the downregulation ofDEGS1 in the AT of MO patients and by recent reports(2324) showing that DhCer rather than Cer content pos-itively correlates with BMI and waist circumference incohorts of overweight obese subjects To date only a singlefunctional mutation in the degs1 gene has been reportedin humans where patients demonstrate increased serumDhCer decreased cholesterol esters and decreased waist-to-hip ratio (25) Globally considered these data suggest
the existence of an association between DEGS1 functionand fat mass
The molecular mechanism leading to the defective ATexpression of degs1 in human and rodent obesity is un-known Here we have shown that tumor necrosis factor-aa mediator of obesity-induced inflammation and insulinresistance decreases the transcriptional expression ofdegs1 in cultured adipocytes which suggests that lowchronic inflammation may contribute to DEGS1 dysfunc-tion Other obesity-associated processes such as in-creased oxidative stress and hypoxia have been shownto impair DEGS1 activity and increase DhCer levels (26)Our results also show a positive association between
Figure 5mdashC8-CPPC inhibitor increases proliferation and decreases lipid accumulation in preadipocytes in vivo Ten-week-old mice weregiven a 45 HFD for 5 weeks before being treated intraperitoneally with vehicle or C8-CPPC inhibitor (2 mgkgday) over 9 daysA Representative images of immunofluorescence analysis of gonadal AT of control (vehicle) and C8-CPPCndashtreated mice are presentedwith Pref-1 (cyan) and Ki67 (red white arrows) Nuclei and lipids are respectively stained with Hoechst stain (blue) and BODIPY (green)Scale bars 100 or 20 mm Quantification of preadipocytes (left graph) (percentage of Pref-1+ cellstotal cells) and (right graph) Ki67+ (whitebars)BODIPY+ (dark gray bars) cells among preadipocytes (Pref-1+ cells) (light gray bars) B mRNA expression of preadipocyte markersand pparg degs1 adiponectin caspase 3 and fas in gonadal AT of control and C8-CPPCndashtreated mice n = 7ndash8 mice per experimentalgroup Values are the mean 6SEM P lt 005 P lt 00001 vs vehicle
1188 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
Figure 6mdashA Glycerol release in 3T3-L1 adipocytes exposed for 48 h to C8-CPPC and stimulated with NE for 6 h BndashD Results fromWestern blot analysis for phosphorylated (p) HSLHSL ratio and total HSL in 3T3-L1 adipocytes exposed for 48 h to C8-CPPC andstimulated with NE for 6 h Graphs show the mean 6 SEM of two separated experiments P lt 005 E Effects of pharmacologicalinhibition of DEGS1 on insulin (ins) signaling Results from Western blot analysis for p-AKT and AKT in 3T3-L1 adipocytes exposed 48 or 72h with C8-CPPC 1 mmolL and increased concentrations of insulin (10ndash100 nmolL) for 15 min F and G Results from Western blot analysisfor Glut4 and adiponectin in 3T3-L1 adipocytes exposed with C8-CPPC 05ndash1 mmolL and insulin (100 nmolL) for 48 h H Results fromWestern blot analysis for AMPK in 3T3-L1 adipocytes exposed to C8-CPPC 05ndash1 mmolL and insulin (Ins) (100 nmolL) rosiglitazone (Rosi)(1 nmolL) and metformin (Metf) (100 nmolL) for 48 h aP lt 005 vs untreated cells (Ins Rosi or Metf effect) bP lt 005 vs 0 mmolLC8-CPPC (C8-CPPC effect) AU arbitrary units IOD integrated optical density
diabetesdiabetesjournalsorg Barbarroja and Associates 1189
levels of the proadipogenic pparg2 isoform and degs1in AT and given the documented downregulation ofpparg2 in AT in obese insulin-resistant states it is con-ceivable that decreased levels of pparg2 may contributeto the defective expression of degs1 associated withobesity
Studies in rodents have shown that the number of Cersis increased in the WAT of animals fed an HFD (5) con-comitantly with the onset of insulin resistance (6) Resultsfrom obob mice and obese human studies are more un-clear where both increased or decreased levels of Cershave been reported (78) This suggests that Cer synthesismay be dependent on the severity of the obese state andthe strength of the adaptive homeostatic responsesattempting to restore their normal balance
Our pharmacological and gene ablation studies haveshown that inhibition of DEGS1 is associated with oxida-tive stress cellular death impaired adipocyte differentia-tion impaired lipid accumulation and impaired basallipolysis in mature adipocytes
Our in vitro studies revealed that the defect in theadipogenesis program caused by depletion of DEGS1occurs during the early stages of adipogenesis (MCE)when PPARg controls the expression of genes that regu-late the cell cycle by directly interacting with cyclin D3Another cyclin involved in adipocyte differentiation iscyclin D1 and here we show that downregulation ofDEGS1 blocks PPARg activity and cyclins such as D1D3 E as well as decreasing cdk2 which modulates PPARgactivity during adipogenesis These data are consistent
Figure 7mdashA and B DhCers impair adipocyte differentiation during early adipogenesis 3T3-L1 preadipocytes treated with C2DhCer(50 mmolL) for a period of 48 h at day 0 or day 3 of differentiation Oil Red O staining and mRNA expression of adipogenesis and lipidmetabolism genes Values are the mean 6 SEM of two separate experiments performed in triplicate P lt 005 vs control C DhCersdecrease ligand-mediated PPARg transactivation Cells were treated with DMSO as a control group and rosiglitazone 10 mmolL GW192910 mmolL and C2 and C16DhCers and Cers 100 mmolL as indicated Graphs represent the average of three independent experimentsP lt 005 vs control cells P lt 005 vs rosiglitazone ampP lt 005 vs GW1929 CTL control
1190 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
with studies showing that downregulation of degs1 bysmall interfering RNA inhibits cell growth and arreststhe cell cycle in cancer cells (12) and where its overexpres-sion increased cell migration and metastasis (27)
The pharmacological inhibition of DEGS1 in vivovalidated the results obtained in vitro and strengthenedthe concept that functional degs1 is required for the fulldifferentiation of preadipocytes into mature adipocytes
We also provide evidence that both pharmacologicaland genetic ablation of degs1 which according to ourresults in in vitro models may directly block adipocytedifferentiation and lipid deposition certainly increasedDhCer levels in in vitro models In this line our resultsshowed that the treatment of adipocytes with DhCercauses impaired adipogenesis and lipid accumulationThis effect on lipid accumulation was more severewhen the treatment was administered during the first3 days of differentiation suggesting a direct impact ofDhCer accumulation during early stages of adipogenesisWe also provide evidence showing that accumulation ofDhCers can also directly repress the transcriptional ac-tivity of PPARg and hence could contribute to impair-ment of the capacity of the preadipocytes to developa full adipogenic program
These observations strengthen the concept that DhCersare not merely inert precursors of Cers Nevertheless themolecular mechanism linking DhCers accumulation anddysregulation of the cell cycle and other cellular eventsremains elusive One possible mechanism is that changesin the DhCerCer ratio may disrupt membrane-dependentstructures by altering the levels of cholesterol andorcaveolin in lipid rafts which are known to be highlysensitive to a sphingomyelin pool Alternatively changesin the DhCerCer ratio may disturb the global phospho-lipidome potentially altering membrane-associated pro-cesses relevant to adipocyte function These are importantquestions to address in future research
In summary our results indicate that defects in DEGS1in the context of the metabolic syndrome may compro-mise AT expansion and function through the combinedinhibition of adipogenesis promotion of cell death andoxidative stress due to the direct accumulation of DhCersThus our data suggest that the selective manipulation ofDEGS1 andor its substrates in WAT may be a pathophy-siologically advantageous strategy to improve AT homeo-stasis and ameliorate the burden of obesity-associatedmetabolic complications
Acknowledgments The authors thank Dr Carobbio Mrs Peirce andMs Philips (Wellcome Trust-Medical Research Council Institute of MetabolicScience University of Cambridge UK) for their excellent technical advice sup-port and suggestionsFunding This work was funded by the Medical Research Council (MRC) theMetabolic Diseases Unit of the MRC European Commission grant FP7-ETHERPATHSand the British Heart FoundationDuality of Interest No potential conflicts of interest relevant to this articlewere reported
Author Contributions NB helped to develop the hypothesis design theexperiments perform the in vitro and ex vivo experiments collect and analyze thedata hold and characterize the human cohorts collect gene expression data fromadipose depots and analyze the data discuss the manuscript coordinate anddirect the project and write the manuscript SR-C helped to develop thehypothesis design the experiments perform the in vitro and ex vivo experimentscollect and analyze the data discuss the manuscript coordinate and direct theproject and write the manuscript HN and MO helped to develop analyticalplatforms perform and analyze lipidomic experiments and discuss the manu-script AC AP JR IC VP GM-G and CL-P helped to perform the invitro and ex vivo experiments collect and analyze the data and discuss themanuscript FJT helped to hold and characterize the human cohorts collectgene expression data from adipose depots and analyze the data and discussthe manuscript JDS and SAS helped to design the experiments and dis-cuss the manuscript AV-P helped to develop the hypothesis design theexperiments discuss the manuscript coordinate and direct the project andwrite the manuscript AV-P is the guarantor of this work and as such had fullaccess to all the data in the study and takes responsibility for the integrity of thedata and the accuracy of the data analysis
References1 Montague CT OrsquoRahilly S The perils of portliness causes and con-
sequences of visceral adiposity Diabetes 200049883ndash8882 Carobbio S Rodriguez-Cuenca S Vidal-Puig A Origins of metabolic com-
plications in obesity ectopic fat accumulation The importance of the qualitative
aspect of lipotoxicity Curr Opin Clin Nutr Metab Care 201114520ndash5263 Chavez JA Summers SA Lipid oversupply selective insulin resistance
and lipotoxicity molecular mechanisms Biochim Biophys Acta 20101801
252ndash2654 Chavez JA Summers SA A ceramide-centric view of insulin resistance Cell
Metab 201215585ndash5945 Shah C Yang G Lee I Bielawski J Hannun YA Samad F Protection from
high fat diet-induced increase in ceramide in mice lacking plasminogen activator
inhibitor 1 J Biol Chem 200828313538ndash135486 Turner N Kowalski GM Leslie SJ et al Distinct patterns of tissue-specific
lipid accumulation during the induction of insulin resistance in mice by high-fat
feeding Diabetologia 2013561638ndash16487 Blachnio-Zabielska AU Koutsari C Tchkonia T Jensen MD Sphingolipid
content of human adipose tissue relationship to adiponectin and insulin re-
sistance Obesity (Silver Spring) 2012202341ndash23478 Błachnio-Zabielska AU Pułka M Baranowski M et al Ceramide metabo-
lism is affected by obesity and diabetes in human adipose tissue J Cell Physiol
2012227550ndash5579 Ternes P Franke S Zaumlhringer U Sperling P Heinz E Identification and
characterization of a sphingolipid delta 4-desaturase family J Biol Chem 2002
27725512ndash2551810 Mizutani Y Kihara A Igarashi Y Identification of the human sphingolipid C4-
hydroxylase hDES2 and its up-regulation during keratinocyte differentiation
FEBS Lett 200456393ndash9711 Ahn EH Schroeder JJ Induction of apoptosis by sphingosine sphinganine
and C(2)-ceramide in human colon cancer cells but not by C(2)-dihydroceramide
Anticancer Res 2010302881ndash288412 Kraveka JM Li L Szulc ZM et al Involvement of dihydroceramide desaturase
in cell cycle progression in human neuroblastoma cells J Biol Chem 2007282
16718ndash1672813 Siddique MM Li Y Wang L et al Ablation of dihydroceramide desaturase 1
a therapeutic target for the treatment of metabolic diseases simultaneously stim-
ulates anabolic and catabolic signaling Mol Cell Biol 2013332353ndash236914 Parks BW Nam E Org E et al Genetic control of obesity and gut microbiota
composition in response to high-fat high-sucrose diet in mice Cell Metab 2013
17141ndash152
diabetesdiabetesjournalsorg Barbarroja and Associates 1191
15 Bikman BT Guan Y Shui G et al Fenretinide prevents lipid-induced insulinresistance by blocking ceramide biosynthesis J Biol Chem 201228717426ndash1743716 Medina-Gomez G Gray SL Yetukuri L et al PPAR gamma 2 preventslipotoxicity by controlling adipose tissue expandability and peripheral lipid me-tabolism PLoS Genet 20073e6417 Roberts LD Virtue S Vidal-Puig A Nicholls AW Griffin JL Metabolic pheno-typing of a model of adipocyte differentiation Physiol Genomics 200939109ndash11918 Ramiacuterez-Zacariacuteas JL Castro-Muntildeozledo F Kuri-Harcuch W Quantitation ofadipose conversion and triglycerides by staining intracytoplasmic lipids with Oilred O Histochemistry 199297493ndash49719 Ye J Coulouris G Zaretskaya I Cutcutache I Rozen S Madden TL Primer-BLAST a tool to design target-specific primers for polymerase chain reactionBMC Bioinformatics 20121313420 Pfaffl MW Tichopad A Prgomet C Neuvians TP Determination of stablehousekeeping genes differentially regulated target genes and sample integrityBestKeepermdashExcel-based tool using pair-wise correlations Biotechnol Lett 200426509ndash51521 Triola G Fabriagraves G Llebaria A Synthesis of a cyclopropene analogue of cer-amide a potent inhibitor of dihydroceramide desaturase Angew Chem Int Ed Engl2001401960ndash1962
22 Tsuji K Satoh S Mitsutake S et al Evaluation of synthetic sphingolipidanalogs as ligands for peroxisome proliferator-activated receptors Bioorg MedChem Lett 2009191643ndash164623 Mamtani M Meikle PJ Kulkarni H et al Plasma dihydroceramide speciesassociate with waist circumference in Mexican American families Obesity (SilverSpring) 201422950ndash95624 Weir JM Wong G Barlow CK et al Plasma lipid profiling in a largepopulation-based cohort J Lipid Res 2013542898ndash290825 Curran J Meikle P Weir J Jowett J Deep sequencing in extended pedi-grees reveals a major rare non-synonymous variant influencing the de novoceramide synthesis pathway Late-breaking abstract presented at the AmericanSociety of Human Genetics 2013 Meeting 22ndash26 October 2013 at the BostonConvention and Exhibition Center Boston Massachusetts26 Idkowiak-Baldys J Takemoto JY Grilley MM Structure-function studies ofyeast C-4 sphingolipid long chain base hydroxylase Biochim Biophys Acta 2003161817ndash2427 Zhou W Ye XL Sun ZJ Ji XD Chen HX Xie D Overexpression of de-generative spermatocyte homolog 1 up-regulates the expression of cyclin D1 andenhances metastatic efficiency in esophageal carcinoma Eca109 cells MolCarcinog 200948886ndash894
1192 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015

inhibition of cyclin D1 and cdk2 expression between time0 and 4 h after the induction (Supplementary Fig 5A andB) Cyclin E and cyclin D3 expression was also inhibitedbut at later time points
Pharmacological Inhibition of DEGS1 RecapitulatesDEGS1 KD Antiadipogenic PhenotypeUse of C8-CPPC a selective inhibitor of DEGS1 (21) fur-ther confirmed the relevance of DEGS1 in adipogenesisSimilarly to degs1 KD cells 3T3-L1 cells were induced todifferentiate and were treated simultaneously withC8-CPPC from time 0 h every 6 h for 48 h C8-CPPC de-creased the expression of proadipogenic transcription fac-tors such as PPARg and CEBPb as well as lipogenic genes(Fig 4A and C) The antiadipogenic effect of C8-CPPC per-sisted after rosiglitazone treatment (Fig 4A and C) Asexpected analysis of neutral lipids showed that 3T3-L1 cellstreated with C8-CPPC for 96 h accumulated significantlyless lipids compared with untreated cells (Fig 4D)
Pharmacological Inhibition of DEGS1 Also ImpairsAdipocyte Differentiation In VivoWe administered C8-CPPC intraperitoneally to mice fedan HFD for 5 weeks an experimental protocol that isknown to trigger adipocyte hyperplasia Molecular analysisof the WAT from mice treated with C8-CPPC presentedhigher levels of pref-1+ cells suggesting an increase in thenumber of preadipocytes compared with controls Inter-estingly a significant number of preadipocytes showedevidence of increased proliferation (Ki67+) whereas thenumber of differentiating preadipocytes (pref1+ BODIPY+)was significantly smaller in comparison with controls (Fig5A) These data were reinforced at the mRNA level byshowing increased expression of pref-1 and gata2 (Fig5B) and the presence of fewer small adipocytes in treatedmice (Supplementary Fig 6) Altogether these data sug-gest that C8-CPPCndashmediated inhibition of DEGS1 im-paired the capacity of preadipocytes to differentiate intoadipocytes in vivo
Figure 1mdashA Tissue distribution of degs1 B Correlation of degs1 mRNA expression and fat pad size The Spearman correlation coef-ficients were calculated to estimate the linear correlations between variables The rejection level for a null hypothesis was P lt 001 Dataare from 14 lean mice CndashE Expression levels of degs1 mRNA in response to HFD FndashH Expression levels of degs1 mRNA in obob miceValues are the mean 6 SEM for six to eight animals per group P lt 005 vs WT AU arbitrary units BAT brown AT gWAT gonadal WATIngFat inguinal fat RetroFat retroperitoneal fat SKM skeletal muscle
1184 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
DEGS1 Is Required for Lipid Accumulation BasalLipolysis and Glucose Uptake in Mature AdipocytesWe next focused on the effects of DEGS1 inhibition infully differentiated adipocytes by treating isolated matureadipocytesmdashfrom the gonadal WAT of C57BL6 micemdashwith C8-CPPC for 24 h C8-CPPC caused a decrease in theexpression of lipid metabolism genes as well as an increasein antioxidant genes (Supplementary Fig 7) recapitulatingour observations in 3T3-L1 preadipocytes
We further investigated the effects of inhibiting DEGS1on lipolytic activity and insulin signaling and measuredadiponectin expression as a representative fingerprintof global adipocyte homeostasis in mature differentiated3T3-L1 adipocytes Thus we observed that basal lipolyticactivity was decreased in mature differentiated 3T3-L1adipocytes when treated with C8-CPPC in nonstimulated
conditions (Fig 6A) Systematic evaluation of the lipolyticaxis showed that under basal conditions phosphorylation ofSer565 was also substantially increased by C8-CPPC con-comitantly with a minor downregulation of Ser660 phosp-horylation (Fig 6BndashD) suggesting a decreased HSL activityMoreover total levels of HSL were also decreased inC8-CPPCndashtreated cells These data suggest that pharmaco-logical inhibition of DEGS1 in mature adipocytes disruptsthe lipolytic response under nonstimulated conditions a de-fect that was superseded in the presence of NE
Finally we characterized the effects of pharmacologicalinhibition of DEGS1 on insulin signaling in 3T3-L1 matureadipocytes treated with C8-CPPC for 48 h as describedabove and subsequently incubated in the presence ofincreasing doses of insulin No major differences wereobserved in the phosphorylation of AKT in C8-CPPCndashtreated
Figure 2mdashA mRNA expression of degs1 in 3T3-L1 treated with short hairpin RNA (shRNA) against degs1 B and C Cell proliferation (XTTand BrdU assays every 24 h until 96 and 72 h respectively) D mRNA expression of Cdk2 Bcl2 Bax and Caspase3 E Apoptosis and celldeath rate F Protein expression of Cdk2 G Oxygen consumption rate H mRNA expression of antioxidant machinery genes I Reactiveoxygen species production J Mitochondria levels All these experiments were perfomed in degs1 KD and WT 3T3-L1 cells Values are themean 6 SEM of three separate experiments performed in triplicate ETC electron transport chain MFI mean fluorescence intensity PIpropidium iodide P lt 005
diabetesdiabetesjournalsorg Barbarroja and Associates 1185
cells after acute insulin stimulation (Fig 6E) Howeverwe found that Glut4 protein levels were increased inC8-CPPCndashtreated cells suggesting that glucose uptakemay be increased (Fig 6F) Interestingly we also did notobserve any differences in either adiponectin levels in re-sponse to C8-CPPC or in AMPK phosphorylation a knowninducer of adiponectin although increased levels of totalAMPK were observed (Fig 6G and H)
Pharmacological and Genetic Inhibition of DEGS1Increases DhCerCer Ratio in 3T3-L1 CellsWe confirmed that C8-CPPC increased the DhCerCerratio in 3T3L-1 preadipocytes upon the inhibition ofDEGS1 (Fig 4E) Similarly degs1 KD preadipocytes (day 0)exhibited an increased DhCerCer ratio versus controls(Supplementary Fig 8A) Downregulation of degs1
expression in degs1 KD cells was accompanied by a down-regulation of serine palmitoyltransferase (sptlc1) and Cersynthase 6 (Cers6) (Supplementary Fig 8A) suggestive ofa homeostatic readjustment of the biosynthetic pathwayto compensate for the dysfunction of degs1 Other genesrelated to Cer homeostasis such as neutral ceramidase(ncdase) and glycosylceramide synthase (gcs) were alsodownregulated
Curiously at day 9 degs1 KD cells despite impaireddifferentiation showed a complete restoration of theDhCerCer ratio (indicated by the normalization of theCer pool) (Supplementary Fig 8B) This could be partiallyexplained by a compensation mechanism mediated bydownregulation of neutral ceramidases and the conversionof sphyngomyelins to Cers (upregulation of sphyngomye-linase 2) (Supplementary Fig 8B)
Figure 3mdashA mRNA expression of degs1 in 3T3-L1 cells during adipocyte differentiation B Lipid accumulation at day 9 of differentiationOil Red O staining C mRNA expression of PREF1 in WT and degs1 KD cells during differentiation D mRNA expression of genes involvedin adipocyte differentiation and lipid accumulation Values are the mean 6 SEM of three separate experiments performed in triplicate FASfatty acid synthase Insig insulin-induced gene LPL lipoprotein lipase P lt 005
1186 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
The Impairment of Adipocyte Differentiation DuringEarly Adipogenesis by DEGS1 Inhibition IsRecapitulated by DhCers Per SeWe finally sought to validate whether DhCer by itself couldmediate the effects of DEGS1 inhibition on adipocytedifferentiation in our models of genetic or pharmacologicalinhibition of degs1 3T3-L1cells were treated with C2DhCerduring 1) the MCE at time 0 and 2) at day 3 after differ-entiation DhCer inhibited lipid accumulation (Fig 7A) andthe expression of genes involved during the early stages ofadipogenesis (Fig 7B)
Since synthetic sphingolipid analogs are known tomodulate PPAR activity (22) we investigated whetherboth C2DhCer and C16DhCer could exert a repressiveeffect on PPARg activation in the presence of the PPARgligands (Fig 7C) Our results show that DhCer can also
block the ligand-mediated transactivation of PPARg Sim-ilar results were obtained with Cers indicating that theseeffects were not a consequence of the unsaturation of thesphingoid moiety (data not shown)
DISCUSSION
The DhCerCer ratio has an important homeostaticregulatory role in the cell contributing to cell survivalautophagy and oxidative stress (1213) Here we identifyDEGS1 as an essential metabolic enzyme which is dysreg-ulated in obese states and contributes to AT dysfunctionOur complementary in vitro and in vivo approaches revealthat reduced DEGS1 function impairs adipogenesis andlipogenesis programs and increases oxidative stress
DEGS1 is expressed in WAT and correlates in rodentswith fat mass in healthy states an association that is
Figure 4mdash3T3-L1 cells treated with C8-CPPC andor rosiglitazone A and B mRNA expression of genes involved in adipocyte differen-tiation and lipid accumulation at 24 and 48 h after induction C Protein expression levels of PPARg2 and cEBPa D Lipid content after 48and 96 h of differentiation Values are the mean 6 SEM of two separate experiments performed in triplicate One-way ANOVA was used toanalyze the statistical significance between treatments at 24 and 48 h Significant differences (Duncan test P lt 005) are indicated withdifferent letters E DhCer Cer and hexosylceramide (HexCer) levels after 48 h of differentiation Values are the mean 6 SEM of twoseparate experiments performed in triplicate cEBP CCAATenhancer binding protein d day FITC fluorescein isothiocyanate MFI meanfluorescence intensity Rosi rosiglitazone P lt 005
diabetesdiabetesjournalsorg Barbarroja and Associates 1187
disrupted in severe forms of obesity The expression ofDEGS1 in liver or muscle is not affected by obesity fur-ther indicating that the WAT is particularly susceptible tochanges in Cer metabolism in obese states The relevanceto human obesity is confirmed by the downregulation ofDEGS1 in the AT of MO patients and by recent reports(2324) showing that DhCer rather than Cer content pos-itively correlates with BMI and waist circumference incohorts of overweight obese subjects To date only a singlefunctional mutation in the degs1 gene has been reportedin humans where patients demonstrate increased serumDhCer decreased cholesterol esters and decreased waist-to-hip ratio (25) Globally considered these data suggest
the existence of an association between DEGS1 functionand fat mass
The molecular mechanism leading to the defective ATexpression of degs1 in human and rodent obesity is un-known Here we have shown that tumor necrosis factor-aa mediator of obesity-induced inflammation and insulinresistance decreases the transcriptional expression ofdegs1 in cultured adipocytes which suggests that lowchronic inflammation may contribute to DEGS1 dysfunc-tion Other obesity-associated processes such as in-creased oxidative stress and hypoxia have been shownto impair DEGS1 activity and increase DhCer levels (26)Our results also show a positive association between
Figure 5mdashC8-CPPC inhibitor increases proliferation and decreases lipid accumulation in preadipocytes in vivo Ten-week-old mice weregiven a 45 HFD for 5 weeks before being treated intraperitoneally with vehicle or C8-CPPC inhibitor (2 mgkgday) over 9 daysA Representative images of immunofluorescence analysis of gonadal AT of control (vehicle) and C8-CPPCndashtreated mice are presentedwith Pref-1 (cyan) and Ki67 (red white arrows) Nuclei and lipids are respectively stained with Hoechst stain (blue) and BODIPY (green)Scale bars 100 or 20 mm Quantification of preadipocytes (left graph) (percentage of Pref-1+ cellstotal cells) and (right graph) Ki67+ (whitebars)BODIPY+ (dark gray bars) cells among preadipocytes (Pref-1+ cells) (light gray bars) B mRNA expression of preadipocyte markersand pparg degs1 adiponectin caspase 3 and fas in gonadal AT of control and C8-CPPCndashtreated mice n = 7ndash8 mice per experimentalgroup Values are the mean 6SEM P lt 005 P lt 00001 vs vehicle
1188 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
Figure 6mdashA Glycerol release in 3T3-L1 adipocytes exposed for 48 h to C8-CPPC and stimulated with NE for 6 h BndashD Results fromWestern blot analysis for phosphorylated (p) HSLHSL ratio and total HSL in 3T3-L1 adipocytes exposed for 48 h to C8-CPPC andstimulated with NE for 6 h Graphs show the mean 6 SEM of two separated experiments P lt 005 E Effects of pharmacologicalinhibition of DEGS1 on insulin (ins) signaling Results from Western blot analysis for p-AKT and AKT in 3T3-L1 adipocytes exposed 48 or 72h with C8-CPPC 1 mmolL and increased concentrations of insulin (10ndash100 nmolL) for 15 min F and G Results from Western blot analysisfor Glut4 and adiponectin in 3T3-L1 adipocytes exposed with C8-CPPC 05ndash1 mmolL and insulin (100 nmolL) for 48 h H Results fromWestern blot analysis for AMPK in 3T3-L1 adipocytes exposed to C8-CPPC 05ndash1 mmolL and insulin (Ins) (100 nmolL) rosiglitazone (Rosi)(1 nmolL) and metformin (Metf) (100 nmolL) for 48 h aP lt 005 vs untreated cells (Ins Rosi or Metf effect) bP lt 005 vs 0 mmolLC8-CPPC (C8-CPPC effect) AU arbitrary units IOD integrated optical density
diabetesdiabetesjournalsorg Barbarroja and Associates 1189
levels of the proadipogenic pparg2 isoform and degs1in AT and given the documented downregulation ofpparg2 in AT in obese insulin-resistant states it is con-ceivable that decreased levels of pparg2 may contributeto the defective expression of degs1 associated withobesity
Studies in rodents have shown that the number of Cersis increased in the WAT of animals fed an HFD (5) con-comitantly with the onset of insulin resistance (6) Resultsfrom obob mice and obese human studies are more un-clear where both increased or decreased levels of Cershave been reported (78) This suggests that Cer synthesismay be dependent on the severity of the obese state andthe strength of the adaptive homeostatic responsesattempting to restore their normal balance
Our pharmacological and gene ablation studies haveshown that inhibition of DEGS1 is associated with oxida-tive stress cellular death impaired adipocyte differentia-tion impaired lipid accumulation and impaired basallipolysis in mature adipocytes
Our in vitro studies revealed that the defect in theadipogenesis program caused by depletion of DEGS1occurs during the early stages of adipogenesis (MCE)when PPARg controls the expression of genes that regu-late the cell cycle by directly interacting with cyclin D3Another cyclin involved in adipocyte differentiation iscyclin D1 and here we show that downregulation ofDEGS1 blocks PPARg activity and cyclins such as D1D3 E as well as decreasing cdk2 which modulates PPARgactivity during adipogenesis These data are consistent
Figure 7mdashA and B DhCers impair adipocyte differentiation during early adipogenesis 3T3-L1 preadipocytes treated with C2DhCer(50 mmolL) for a period of 48 h at day 0 or day 3 of differentiation Oil Red O staining and mRNA expression of adipogenesis and lipidmetabolism genes Values are the mean 6 SEM of two separate experiments performed in triplicate P lt 005 vs control C DhCersdecrease ligand-mediated PPARg transactivation Cells were treated with DMSO as a control group and rosiglitazone 10 mmolL GW192910 mmolL and C2 and C16DhCers and Cers 100 mmolL as indicated Graphs represent the average of three independent experimentsP lt 005 vs control cells P lt 005 vs rosiglitazone ampP lt 005 vs GW1929 CTL control
1190 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
with studies showing that downregulation of degs1 bysmall interfering RNA inhibits cell growth and arreststhe cell cycle in cancer cells (12) and where its overexpres-sion increased cell migration and metastasis (27)
The pharmacological inhibition of DEGS1 in vivovalidated the results obtained in vitro and strengthenedthe concept that functional degs1 is required for the fulldifferentiation of preadipocytes into mature adipocytes
We also provide evidence that both pharmacologicaland genetic ablation of degs1 which according to ourresults in in vitro models may directly block adipocytedifferentiation and lipid deposition certainly increasedDhCer levels in in vitro models In this line our resultsshowed that the treatment of adipocytes with DhCercauses impaired adipogenesis and lipid accumulationThis effect on lipid accumulation was more severewhen the treatment was administered during the first3 days of differentiation suggesting a direct impact ofDhCer accumulation during early stages of adipogenesisWe also provide evidence showing that accumulation ofDhCers can also directly repress the transcriptional ac-tivity of PPARg and hence could contribute to impair-ment of the capacity of the preadipocytes to developa full adipogenic program
These observations strengthen the concept that DhCersare not merely inert precursors of Cers Nevertheless themolecular mechanism linking DhCers accumulation anddysregulation of the cell cycle and other cellular eventsremains elusive One possible mechanism is that changesin the DhCerCer ratio may disrupt membrane-dependentstructures by altering the levels of cholesterol andorcaveolin in lipid rafts which are known to be highlysensitive to a sphingomyelin pool Alternatively changesin the DhCerCer ratio may disturb the global phospho-lipidome potentially altering membrane-associated pro-cesses relevant to adipocyte function These are importantquestions to address in future research
In summary our results indicate that defects in DEGS1in the context of the metabolic syndrome may compro-mise AT expansion and function through the combinedinhibition of adipogenesis promotion of cell death andoxidative stress due to the direct accumulation of DhCersThus our data suggest that the selective manipulation ofDEGS1 andor its substrates in WAT may be a pathophy-siologically advantageous strategy to improve AT homeo-stasis and ameliorate the burden of obesity-associatedmetabolic complications
Acknowledgments The authors thank Dr Carobbio Mrs Peirce andMs Philips (Wellcome Trust-Medical Research Council Institute of MetabolicScience University of Cambridge UK) for their excellent technical advice sup-port and suggestionsFunding This work was funded by the Medical Research Council (MRC) theMetabolic Diseases Unit of the MRC European Commission grant FP7-ETHERPATHSand the British Heart FoundationDuality of Interest No potential conflicts of interest relevant to this articlewere reported
Author Contributions NB helped to develop the hypothesis design theexperiments perform the in vitro and ex vivo experiments collect and analyze thedata hold and characterize the human cohorts collect gene expression data fromadipose depots and analyze the data discuss the manuscript coordinate anddirect the project and write the manuscript SR-C helped to develop thehypothesis design the experiments perform the in vitro and ex vivo experimentscollect and analyze the data discuss the manuscript coordinate and direct theproject and write the manuscript HN and MO helped to develop analyticalplatforms perform and analyze lipidomic experiments and discuss the manu-script AC AP JR IC VP GM-G and CL-P helped to perform the invitro and ex vivo experiments collect and analyze the data and discuss themanuscript FJT helped to hold and characterize the human cohorts collectgene expression data from adipose depots and analyze the data and discussthe manuscript JDS and SAS helped to design the experiments and dis-cuss the manuscript AV-P helped to develop the hypothesis design theexperiments discuss the manuscript coordinate and direct the project andwrite the manuscript AV-P is the guarantor of this work and as such had fullaccess to all the data in the study and takes responsibility for the integrity of thedata and the accuracy of the data analysis
References1 Montague CT OrsquoRahilly S The perils of portliness causes and con-
sequences of visceral adiposity Diabetes 200049883ndash8882 Carobbio S Rodriguez-Cuenca S Vidal-Puig A Origins of metabolic com-
plications in obesity ectopic fat accumulation The importance of the qualitative
aspect of lipotoxicity Curr Opin Clin Nutr Metab Care 201114520ndash5263 Chavez JA Summers SA Lipid oversupply selective insulin resistance
and lipotoxicity molecular mechanisms Biochim Biophys Acta 20101801
252ndash2654 Chavez JA Summers SA A ceramide-centric view of insulin resistance Cell
Metab 201215585ndash5945 Shah C Yang G Lee I Bielawski J Hannun YA Samad F Protection from
high fat diet-induced increase in ceramide in mice lacking plasminogen activator
inhibitor 1 J Biol Chem 200828313538ndash135486 Turner N Kowalski GM Leslie SJ et al Distinct patterns of tissue-specific
lipid accumulation during the induction of insulin resistance in mice by high-fat
feeding Diabetologia 2013561638ndash16487 Blachnio-Zabielska AU Koutsari C Tchkonia T Jensen MD Sphingolipid
content of human adipose tissue relationship to adiponectin and insulin re-
sistance Obesity (Silver Spring) 2012202341ndash23478 Błachnio-Zabielska AU Pułka M Baranowski M et al Ceramide metabo-
lism is affected by obesity and diabetes in human adipose tissue J Cell Physiol
2012227550ndash5579 Ternes P Franke S Zaumlhringer U Sperling P Heinz E Identification and
characterization of a sphingolipid delta 4-desaturase family J Biol Chem 2002
27725512ndash2551810 Mizutani Y Kihara A Igarashi Y Identification of the human sphingolipid C4-
hydroxylase hDES2 and its up-regulation during keratinocyte differentiation
FEBS Lett 200456393ndash9711 Ahn EH Schroeder JJ Induction of apoptosis by sphingosine sphinganine
and C(2)-ceramide in human colon cancer cells but not by C(2)-dihydroceramide
Anticancer Res 2010302881ndash288412 Kraveka JM Li L Szulc ZM et al Involvement of dihydroceramide desaturase
in cell cycle progression in human neuroblastoma cells J Biol Chem 2007282
16718ndash1672813 Siddique MM Li Y Wang L et al Ablation of dihydroceramide desaturase 1
a therapeutic target for the treatment of metabolic diseases simultaneously stim-
ulates anabolic and catabolic signaling Mol Cell Biol 2013332353ndash236914 Parks BW Nam E Org E et al Genetic control of obesity and gut microbiota
composition in response to high-fat high-sucrose diet in mice Cell Metab 2013
17141ndash152
diabetesdiabetesjournalsorg Barbarroja and Associates 1191
15 Bikman BT Guan Y Shui G et al Fenretinide prevents lipid-induced insulinresistance by blocking ceramide biosynthesis J Biol Chem 201228717426ndash1743716 Medina-Gomez G Gray SL Yetukuri L et al PPAR gamma 2 preventslipotoxicity by controlling adipose tissue expandability and peripheral lipid me-tabolism PLoS Genet 20073e6417 Roberts LD Virtue S Vidal-Puig A Nicholls AW Griffin JL Metabolic pheno-typing of a model of adipocyte differentiation Physiol Genomics 200939109ndash11918 Ramiacuterez-Zacariacuteas JL Castro-Muntildeozledo F Kuri-Harcuch W Quantitation ofadipose conversion and triglycerides by staining intracytoplasmic lipids with Oilred O Histochemistry 199297493ndash49719 Ye J Coulouris G Zaretskaya I Cutcutache I Rozen S Madden TL Primer-BLAST a tool to design target-specific primers for polymerase chain reactionBMC Bioinformatics 20121313420 Pfaffl MW Tichopad A Prgomet C Neuvians TP Determination of stablehousekeeping genes differentially regulated target genes and sample integrityBestKeepermdashExcel-based tool using pair-wise correlations Biotechnol Lett 200426509ndash51521 Triola G Fabriagraves G Llebaria A Synthesis of a cyclopropene analogue of cer-amide a potent inhibitor of dihydroceramide desaturase Angew Chem Int Ed Engl2001401960ndash1962
22 Tsuji K Satoh S Mitsutake S et al Evaluation of synthetic sphingolipidanalogs as ligands for peroxisome proliferator-activated receptors Bioorg MedChem Lett 2009191643ndash164623 Mamtani M Meikle PJ Kulkarni H et al Plasma dihydroceramide speciesassociate with waist circumference in Mexican American families Obesity (SilverSpring) 201422950ndash95624 Weir JM Wong G Barlow CK et al Plasma lipid profiling in a largepopulation-based cohort J Lipid Res 2013542898ndash290825 Curran J Meikle P Weir J Jowett J Deep sequencing in extended pedi-grees reveals a major rare non-synonymous variant influencing the de novoceramide synthesis pathway Late-breaking abstract presented at the AmericanSociety of Human Genetics 2013 Meeting 22ndash26 October 2013 at the BostonConvention and Exhibition Center Boston Massachusetts26 Idkowiak-Baldys J Takemoto JY Grilley MM Structure-function studies ofyeast C-4 sphingolipid long chain base hydroxylase Biochim Biophys Acta 2003161817ndash2427 Zhou W Ye XL Sun ZJ Ji XD Chen HX Xie D Overexpression of de-generative spermatocyte homolog 1 up-regulates the expression of cyclin D1 andenhances metastatic efficiency in esophageal carcinoma Eca109 cells MolCarcinog 200948886ndash894
1192 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015

DEGS1 Is Required for Lipid Accumulation BasalLipolysis and Glucose Uptake in Mature AdipocytesWe next focused on the effects of DEGS1 inhibition infully differentiated adipocytes by treating isolated matureadipocytesmdashfrom the gonadal WAT of C57BL6 micemdashwith C8-CPPC for 24 h C8-CPPC caused a decrease in theexpression of lipid metabolism genes as well as an increasein antioxidant genes (Supplementary Fig 7) recapitulatingour observations in 3T3-L1 preadipocytes
We further investigated the effects of inhibiting DEGS1on lipolytic activity and insulin signaling and measuredadiponectin expression as a representative fingerprintof global adipocyte homeostasis in mature differentiated3T3-L1 adipocytes Thus we observed that basal lipolyticactivity was decreased in mature differentiated 3T3-L1adipocytes when treated with C8-CPPC in nonstimulated
conditions (Fig 6A) Systematic evaluation of the lipolyticaxis showed that under basal conditions phosphorylation ofSer565 was also substantially increased by C8-CPPC con-comitantly with a minor downregulation of Ser660 phosp-horylation (Fig 6BndashD) suggesting a decreased HSL activityMoreover total levels of HSL were also decreased inC8-CPPCndashtreated cells These data suggest that pharmaco-logical inhibition of DEGS1 in mature adipocytes disruptsthe lipolytic response under nonstimulated conditions a de-fect that was superseded in the presence of NE
Finally we characterized the effects of pharmacologicalinhibition of DEGS1 on insulin signaling in 3T3-L1 matureadipocytes treated with C8-CPPC for 48 h as describedabove and subsequently incubated in the presence ofincreasing doses of insulin No major differences wereobserved in the phosphorylation of AKT in C8-CPPCndashtreated
Figure 2mdashA mRNA expression of degs1 in 3T3-L1 treated with short hairpin RNA (shRNA) against degs1 B and C Cell proliferation (XTTand BrdU assays every 24 h until 96 and 72 h respectively) D mRNA expression of Cdk2 Bcl2 Bax and Caspase3 E Apoptosis and celldeath rate F Protein expression of Cdk2 G Oxygen consumption rate H mRNA expression of antioxidant machinery genes I Reactiveoxygen species production J Mitochondria levels All these experiments were perfomed in degs1 KD and WT 3T3-L1 cells Values are themean 6 SEM of three separate experiments performed in triplicate ETC electron transport chain MFI mean fluorescence intensity PIpropidium iodide P lt 005
diabetesdiabetesjournalsorg Barbarroja and Associates 1185
cells after acute insulin stimulation (Fig 6E) Howeverwe found that Glut4 protein levels were increased inC8-CPPCndashtreated cells suggesting that glucose uptakemay be increased (Fig 6F) Interestingly we also did notobserve any differences in either adiponectin levels in re-sponse to C8-CPPC or in AMPK phosphorylation a knowninducer of adiponectin although increased levels of totalAMPK were observed (Fig 6G and H)
Pharmacological and Genetic Inhibition of DEGS1Increases DhCerCer Ratio in 3T3-L1 CellsWe confirmed that C8-CPPC increased the DhCerCerratio in 3T3L-1 preadipocytes upon the inhibition ofDEGS1 (Fig 4E) Similarly degs1 KD preadipocytes (day 0)exhibited an increased DhCerCer ratio versus controls(Supplementary Fig 8A) Downregulation of degs1
expression in degs1 KD cells was accompanied by a down-regulation of serine palmitoyltransferase (sptlc1) and Cersynthase 6 (Cers6) (Supplementary Fig 8A) suggestive ofa homeostatic readjustment of the biosynthetic pathwayto compensate for the dysfunction of degs1 Other genesrelated to Cer homeostasis such as neutral ceramidase(ncdase) and glycosylceramide synthase (gcs) were alsodownregulated
Curiously at day 9 degs1 KD cells despite impaireddifferentiation showed a complete restoration of theDhCerCer ratio (indicated by the normalization of theCer pool) (Supplementary Fig 8B) This could be partiallyexplained by a compensation mechanism mediated bydownregulation of neutral ceramidases and the conversionof sphyngomyelins to Cers (upregulation of sphyngomye-linase 2) (Supplementary Fig 8B)
Figure 3mdashA mRNA expression of degs1 in 3T3-L1 cells during adipocyte differentiation B Lipid accumulation at day 9 of differentiationOil Red O staining C mRNA expression of PREF1 in WT and degs1 KD cells during differentiation D mRNA expression of genes involvedin adipocyte differentiation and lipid accumulation Values are the mean 6 SEM of three separate experiments performed in triplicate FASfatty acid synthase Insig insulin-induced gene LPL lipoprotein lipase P lt 005
1186 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
The Impairment of Adipocyte Differentiation DuringEarly Adipogenesis by DEGS1 Inhibition IsRecapitulated by DhCers Per SeWe finally sought to validate whether DhCer by itself couldmediate the effects of DEGS1 inhibition on adipocytedifferentiation in our models of genetic or pharmacologicalinhibition of degs1 3T3-L1cells were treated with C2DhCerduring 1) the MCE at time 0 and 2) at day 3 after differ-entiation DhCer inhibited lipid accumulation (Fig 7A) andthe expression of genes involved during the early stages ofadipogenesis (Fig 7B)
Since synthetic sphingolipid analogs are known tomodulate PPAR activity (22) we investigated whetherboth C2DhCer and C16DhCer could exert a repressiveeffect on PPARg activation in the presence of the PPARgligands (Fig 7C) Our results show that DhCer can also
block the ligand-mediated transactivation of PPARg Sim-ilar results were obtained with Cers indicating that theseeffects were not a consequence of the unsaturation of thesphingoid moiety (data not shown)
DISCUSSION
The DhCerCer ratio has an important homeostaticregulatory role in the cell contributing to cell survivalautophagy and oxidative stress (1213) Here we identifyDEGS1 as an essential metabolic enzyme which is dysreg-ulated in obese states and contributes to AT dysfunctionOur complementary in vitro and in vivo approaches revealthat reduced DEGS1 function impairs adipogenesis andlipogenesis programs and increases oxidative stress
DEGS1 is expressed in WAT and correlates in rodentswith fat mass in healthy states an association that is
Figure 4mdash3T3-L1 cells treated with C8-CPPC andor rosiglitazone A and B mRNA expression of genes involved in adipocyte differen-tiation and lipid accumulation at 24 and 48 h after induction C Protein expression levels of PPARg2 and cEBPa D Lipid content after 48and 96 h of differentiation Values are the mean 6 SEM of two separate experiments performed in triplicate One-way ANOVA was used toanalyze the statistical significance between treatments at 24 and 48 h Significant differences (Duncan test P lt 005) are indicated withdifferent letters E DhCer Cer and hexosylceramide (HexCer) levels after 48 h of differentiation Values are the mean 6 SEM of twoseparate experiments performed in triplicate cEBP CCAATenhancer binding protein d day FITC fluorescein isothiocyanate MFI meanfluorescence intensity Rosi rosiglitazone P lt 005
diabetesdiabetesjournalsorg Barbarroja and Associates 1187
disrupted in severe forms of obesity The expression ofDEGS1 in liver or muscle is not affected by obesity fur-ther indicating that the WAT is particularly susceptible tochanges in Cer metabolism in obese states The relevanceto human obesity is confirmed by the downregulation ofDEGS1 in the AT of MO patients and by recent reports(2324) showing that DhCer rather than Cer content pos-itively correlates with BMI and waist circumference incohorts of overweight obese subjects To date only a singlefunctional mutation in the degs1 gene has been reportedin humans where patients demonstrate increased serumDhCer decreased cholesterol esters and decreased waist-to-hip ratio (25) Globally considered these data suggest
the existence of an association between DEGS1 functionand fat mass
The molecular mechanism leading to the defective ATexpression of degs1 in human and rodent obesity is un-known Here we have shown that tumor necrosis factor-aa mediator of obesity-induced inflammation and insulinresistance decreases the transcriptional expression ofdegs1 in cultured adipocytes which suggests that lowchronic inflammation may contribute to DEGS1 dysfunc-tion Other obesity-associated processes such as in-creased oxidative stress and hypoxia have been shownto impair DEGS1 activity and increase DhCer levels (26)Our results also show a positive association between
Figure 5mdashC8-CPPC inhibitor increases proliferation and decreases lipid accumulation in preadipocytes in vivo Ten-week-old mice weregiven a 45 HFD for 5 weeks before being treated intraperitoneally with vehicle or C8-CPPC inhibitor (2 mgkgday) over 9 daysA Representative images of immunofluorescence analysis of gonadal AT of control (vehicle) and C8-CPPCndashtreated mice are presentedwith Pref-1 (cyan) and Ki67 (red white arrows) Nuclei and lipids are respectively stained with Hoechst stain (blue) and BODIPY (green)Scale bars 100 or 20 mm Quantification of preadipocytes (left graph) (percentage of Pref-1+ cellstotal cells) and (right graph) Ki67+ (whitebars)BODIPY+ (dark gray bars) cells among preadipocytes (Pref-1+ cells) (light gray bars) B mRNA expression of preadipocyte markersand pparg degs1 adiponectin caspase 3 and fas in gonadal AT of control and C8-CPPCndashtreated mice n = 7ndash8 mice per experimentalgroup Values are the mean 6SEM P lt 005 P lt 00001 vs vehicle
1188 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
Figure 6mdashA Glycerol release in 3T3-L1 adipocytes exposed for 48 h to C8-CPPC and stimulated with NE for 6 h BndashD Results fromWestern blot analysis for phosphorylated (p) HSLHSL ratio and total HSL in 3T3-L1 adipocytes exposed for 48 h to C8-CPPC andstimulated with NE for 6 h Graphs show the mean 6 SEM of two separated experiments P lt 005 E Effects of pharmacologicalinhibition of DEGS1 on insulin (ins) signaling Results from Western blot analysis for p-AKT and AKT in 3T3-L1 adipocytes exposed 48 or 72h with C8-CPPC 1 mmolL and increased concentrations of insulin (10ndash100 nmolL) for 15 min F and G Results from Western blot analysisfor Glut4 and adiponectin in 3T3-L1 adipocytes exposed with C8-CPPC 05ndash1 mmolL and insulin (100 nmolL) for 48 h H Results fromWestern blot analysis for AMPK in 3T3-L1 adipocytes exposed to C8-CPPC 05ndash1 mmolL and insulin (Ins) (100 nmolL) rosiglitazone (Rosi)(1 nmolL) and metformin (Metf) (100 nmolL) for 48 h aP lt 005 vs untreated cells (Ins Rosi or Metf effect) bP lt 005 vs 0 mmolLC8-CPPC (C8-CPPC effect) AU arbitrary units IOD integrated optical density
diabetesdiabetesjournalsorg Barbarroja and Associates 1189
levels of the proadipogenic pparg2 isoform and degs1in AT and given the documented downregulation ofpparg2 in AT in obese insulin-resistant states it is con-ceivable that decreased levels of pparg2 may contributeto the defective expression of degs1 associated withobesity
Studies in rodents have shown that the number of Cersis increased in the WAT of animals fed an HFD (5) con-comitantly with the onset of insulin resistance (6) Resultsfrom obob mice and obese human studies are more un-clear where both increased or decreased levels of Cershave been reported (78) This suggests that Cer synthesismay be dependent on the severity of the obese state andthe strength of the adaptive homeostatic responsesattempting to restore their normal balance
Our pharmacological and gene ablation studies haveshown that inhibition of DEGS1 is associated with oxida-tive stress cellular death impaired adipocyte differentia-tion impaired lipid accumulation and impaired basallipolysis in mature adipocytes
Our in vitro studies revealed that the defect in theadipogenesis program caused by depletion of DEGS1occurs during the early stages of adipogenesis (MCE)when PPARg controls the expression of genes that regu-late the cell cycle by directly interacting with cyclin D3Another cyclin involved in adipocyte differentiation iscyclin D1 and here we show that downregulation ofDEGS1 blocks PPARg activity and cyclins such as D1D3 E as well as decreasing cdk2 which modulates PPARgactivity during adipogenesis These data are consistent
Figure 7mdashA and B DhCers impair adipocyte differentiation during early adipogenesis 3T3-L1 preadipocytes treated with C2DhCer(50 mmolL) for a period of 48 h at day 0 or day 3 of differentiation Oil Red O staining and mRNA expression of adipogenesis and lipidmetabolism genes Values are the mean 6 SEM of two separate experiments performed in triplicate P lt 005 vs control C DhCersdecrease ligand-mediated PPARg transactivation Cells were treated with DMSO as a control group and rosiglitazone 10 mmolL GW192910 mmolL and C2 and C16DhCers and Cers 100 mmolL as indicated Graphs represent the average of three independent experimentsP lt 005 vs control cells P lt 005 vs rosiglitazone ampP lt 005 vs GW1929 CTL control
1190 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
with studies showing that downregulation of degs1 bysmall interfering RNA inhibits cell growth and arreststhe cell cycle in cancer cells (12) and where its overexpres-sion increased cell migration and metastasis (27)
The pharmacological inhibition of DEGS1 in vivovalidated the results obtained in vitro and strengthenedthe concept that functional degs1 is required for the fulldifferentiation of preadipocytes into mature adipocytes
We also provide evidence that both pharmacologicaland genetic ablation of degs1 which according to ourresults in in vitro models may directly block adipocytedifferentiation and lipid deposition certainly increasedDhCer levels in in vitro models In this line our resultsshowed that the treatment of adipocytes with DhCercauses impaired adipogenesis and lipid accumulationThis effect on lipid accumulation was more severewhen the treatment was administered during the first3 days of differentiation suggesting a direct impact ofDhCer accumulation during early stages of adipogenesisWe also provide evidence showing that accumulation ofDhCers can also directly repress the transcriptional ac-tivity of PPARg and hence could contribute to impair-ment of the capacity of the preadipocytes to developa full adipogenic program
These observations strengthen the concept that DhCersare not merely inert precursors of Cers Nevertheless themolecular mechanism linking DhCers accumulation anddysregulation of the cell cycle and other cellular eventsremains elusive One possible mechanism is that changesin the DhCerCer ratio may disrupt membrane-dependentstructures by altering the levels of cholesterol andorcaveolin in lipid rafts which are known to be highlysensitive to a sphingomyelin pool Alternatively changesin the DhCerCer ratio may disturb the global phospho-lipidome potentially altering membrane-associated pro-cesses relevant to adipocyte function These are importantquestions to address in future research
In summary our results indicate that defects in DEGS1in the context of the metabolic syndrome may compro-mise AT expansion and function through the combinedinhibition of adipogenesis promotion of cell death andoxidative stress due to the direct accumulation of DhCersThus our data suggest that the selective manipulation ofDEGS1 andor its substrates in WAT may be a pathophy-siologically advantageous strategy to improve AT homeo-stasis and ameliorate the burden of obesity-associatedmetabolic complications
Acknowledgments The authors thank Dr Carobbio Mrs Peirce andMs Philips (Wellcome Trust-Medical Research Council Institute of MetabolicScience University of Cambridge UK) for their excellent technical advice sup-port and suggestionsFunding This work was funded by the Medical Research Council (MRC) theMetabolic Diseases Unit of the MRC European Commission grant FP7-ETHERPATHSand the British Heart FoundationDuality of Interest No potential conflicts of interest relevant to this articlewere reported
Author Contributions NB helped to develop the hypothesis design theexperiments perform the in vitro and ex vivo experiments collect and analyze thedata hold and characterize the human cohorts collect gene expression data fromadipose depots and analyze the data discuss the manuscript coordinate anddirect the project and write the manuscript SR-C helped to develop thehypothesis design the experiments perform the in vitro and ex vivo experimentscollect and analyze the data discuss the manuscript coordinate and direct theproject and write the manuscript HN and MO helped to develop analyticalplatforms perform and analyze lipidomic experiments and discuss the manu-script AC AP JR IC VP GM-G and CL-P helped to perform the invitro and ex vivo experiments collect and analyze the data and discuss themanuscript FJT helped to hold and characterize the human cohorts collectgene expression data from adipose depots and analyze the data and discussthe manuscript JDS and SAS helped to design the experiments and dis-cuss the manuscript AV-P helped to develop the hypothesis design theexperiments discuss the manuscript coordinate and direct the project andwrite the manuscript AV-P is the guarantor of this work and as such had fullaccess to all the data in the study and takes responsibility for the integrity of thedata and the accuracy of the data analysis
References1 Montague CT OrsquoRahilly S The perils of portliness causes and con-
sequences of visceral adiposity Diabetes 200049883ndash8882 Carobbio S Rodriguez-Cuenca S Vidal-Puig A Origins of metabolic com-
plications in obesity ectopic fat accumulation The importance of the qualitative
aspect of lipotoxicity Curr Opin Clin Nutr Metab Care 201114520ndash5263 Chavez JA Summers SA Lipid oversupply selective insulin resistance
and lipotoxicity molecular mechanisms Biochim Biophys Acta 20101801
252ndash2654 Chavez JA Summers SA A ceramide-centric view of insulin resistance Cell
Metab 201215585ndash5945 Shah C Yang G Lee I Bielawski J Hannun YA Samad F Protection from
high fat diet-induced increase in ceramide in mice lacking plasminogen activator
inhibitor 1 J Biol Chem 200828313538ndash135486 Turner N Kowalski GM Leslie SJ et al Distinct patterns of tissue-specific
lipid accumulation during the induction of insulin resistance in mice by high-fat
feeding Diabetologia 2013561638ndash16487 Blachnio-Zabielska AU Koutsari C Tchkonia T Jensen MD Sphingolipid
content of human adipose tissue relationship to adiponectin and insulin re-
sistance Obesity (Silver Spring) 2012202341ndash23478 Błachnio-Zabielska AU Pułka M Baranowski M et al Ceramide metabo-
lism is affected by obesity and diabetes in human adipose tissue J Cell Physiol
2012227550ndash5579 Ternes P Franke S Zaumlhringer U Sperling P Heinz E Identification and
characterization of a sphingolipid delta 4-desaturase family J Biol Chem 2002
27725512ndash2551810 Mizutani Y Kihara A Igarashi Y Identification of the human sphingolipid C4-
hydroxylase hDES2 and its up-regulation during keratinocyte differentiation
FEBS Lett 200456393ndash9711 Ahn EH Schroeder JJ Induction of apoptosis by sphingosine sphinganine
and C(2)-ceramide in human colon cancer cells but not by C(2)-dihydroceramide
Anticancer Res 2010302881ndash288412 Kraveka JM Li L Szulc ZM et al Involvement of dihydroceramide desaturase
in cell cycle progression in human neuroblastoma cells J Biol Chem 2007282
16718ndash1672813 Siddique MM Li Y Wang L et al Ablation of dihydroceramide desaturase 1
a therapeutic target for the treatment of metabolic diseases simultaneously stim-
ulates anabolic and catabolic signaling Mol Cell Biol 2013332353ndash236914 Parks BW Nam E Org E et al Genetic control of obesity and gut microbiota
composition in response to high-fat high-sucrose diet in mice Cell Metab 2013
17141ndash152
diabetesdiabetesjournalsorg Barbarroja and Associates 1191
15 Bikman BT Guan Y Shui G et al Fenretinide prevents lipid-induced insulinresistance by blocking ceramide biosynthesis J Biol Chem 201228717426ndash1743716 Medina-Gomez G Gray SL Yetukuri L et al PPAR gamma 2 preventslipotoxicity by controlling adipose tissue expandability and peripheral lipid me-tabolism PLoS Genet 20073e6417 Roberts LD Virtue S Vidal-Puig A Nicholls AW Griffin JL Metabolic pheno-typing of a model of adipocyte differentiation Physiol Genomics 200939109ndash11918 Ramiacuterez-Zacariacuteas JL Castro-Muntildeozledo F Kuri-Harcuch W Quantitation ofadipose conversion and triglycerides by staining intracytoplasmic lipids with Oilred O Histochemistry 199297493ndash49719 Ye J Coulouris G Zaretskaya I Cutcutache I Rozen S Madden TL Primer-BLAST a tool to design target-specific primers for polymerase chain reactionBMC Bioinformatics 20121313420 Pfaffl MW Tichopad A Prgomet C Neuvians TP Determination of stablehousekeeping genes differentially regulated target genes and sample integrityBestKeepermdashExcel-based tool using pair-wise correlations Biotechnol Lett 200426509ndash51521 Triola G Fabriagraves G Llebaria A Synthesis of a cyclopropene analogue of cer-amide a potent inhibitor of dihydroceramide desaturase Angew Chem Int Ed Engl2001401960ndash1962
22 Tsuji K Satoh S Mitsutake S et al Evaluation of synthetic sphingolipidanalogs as ligands for peroxisome proliferator-activated receptors Bioorg MedChem Lett 2009191643ndash164623 Mamtani M Meikle PJ Kulkarni H et al Plasma dihydroceramide speciesassociate with waist circumference in Mexican American families Obesity (SilverSpring) 201422950ndash95624 Weir JM Wong G Barlow CK et al Plasma lipid profiling in a largepopulation-based cohort J Lipid Res 2013542898ndash290825 Curran J Meikle P Weir J Jowett J Deep sequencing in extended pedi-grees reveals a major rare non-synonymous variant influencing the de novoceramide synthesis pathway Late-breaking abstract presented at the AmericanSociety of Human Genetics 2013 Meeting 22ndash26 October 2013 at the BostonConvention and Exhibition Center Boston Massachusetts26 Idkowiak-Baldys J Takemoto JY Grilley MM Structure-function studies ofyeast C-4 sphingolipid long chain base hydroxylase Biochim Biophys Acta 2003161817ndash2427 Zhou W Ye XL Sun ZJ Ji XD Chen HX Xie D Overexpression of de-generative spermatocyte homolog 1 up-regulates the expression of cyclin D1 andenhances metastatic efficiency in esophageal carcinoma Eca109 cells MolCarcinog 200948886ndash894
1192 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015

cells after acute insulin stimulation (Fig 6E) Howeverwe found that Glut4 protein levels were increased inC8-CPPCndashtreated cells suggesting that glucose uptakemay be increased (Fig 6F) Interestingly we also did notobserve any differences in either adiponectin levels in re-sponse to C8-CPPC or in AMPK phosphorylation a knowninducer of adiponectin although increased levels of totalAMPK were observed (Fig 6G and H)
Pharmacological and Genetic Inhibition of DEGS1Increases DhCerCer Ratio in 3T3-L1 CellsWe confirmed that C8-CPPC increased the DhCerCerratio in 3T3L-1 preadipocytes upon the inhibition ofDEGS1 (Fig 4E) Similarly degs1 KD preadipocytes (day 0)exhibited an increased DhCerCer ratio versus controls(Supplementary Fig 8A) Downregulation of degs1
expression in degs1 KD cells was accompanied by a down-regulation of serine palmitoyltransferase (sptlc1) and Cersynthase 6 (Cers6) (Supplementary Fig 8A) suggestive ofa homeostatic readjustment of the biosynthetic pathwayto compensate for the dysfunction of degs1 Other genesrelated to Cer homeostasis such as neutral ceramidase(ncdase) and glycosylceramide synthase (gcs) were alsodownregulated
Curiously at day 9 degs1 KD cells despite impaireddifferentiation showed a complete restoration of theDhCerCer ratio (indicated by the normalization of theCer pool) (Supplementary Fig 8B) This could be partiallyexplained by a compensation mechanism mediated bydownregulation of neutral ceramidases and the conversionof sphyngomyelins to Cers (upregulation of sphyngomye-linase 2) (Supplementary Fig 8B)
Figure 3mdashA mRNA expression of degs1 in 3T3-L1 cells during adipocyte differentiation B Lipid accumulation at day 9 of differentiationOil Red O staining C mRNA expression of PREF1 in WT and degs1 KD cells during differentiation D mRNA expression of genes involvedin adipocyte differentiation and lipid accumulation Values are the mean 6 SEM of three separate experiments performed in triplicate FASfatty acid synthase Insig insulin-induced gene LPL lipoprotein lipase P lt 005
1186 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
The Impairment of Adipocyte Differentiation DuringEarly Adipogenesis by DEGS1 Inhibition IsRecapitulated by DhCers Per SeWe finally sought to validate whether DhCer by itself couldmediate the effects of DEGS1 inhibition on adipocytedifferentiation in our models of genetic or pharmacologicalinhibition of degs1 3T3-L1cells were treated with C2DhCerduring 1) the MCE at time 0 and 2) at day 3 after differ-entiation DhCer inhibited lipid accumulation (Fig 7A) andthe expression of genes involved during the early stages ofadipogenesis (Fig 7B)
Since synthetic sphingolipid analogs are known tomodulate PPAR activity (22) we investigated whetherboth C2DhCer and C16DhCer could exert a repressiveeffect on PPARg activation in the presence of the PPARgligands (Fig 7C) Our results show that DhCer can also
block the ligand-mediated transactivation of PPARg Sim-ilar results were obtained with Cers indicating that theseeffects were not a consequence of the unsaturation of thesphingoid moiety (data not shown)
DISCUSSION
The DhCerCer ratio has an important homeostaticregulatory role in the cell contributing to cell survivalautophagy and oxidative stress (1213) Here we identifyDEGS1 as an essential metabolic enzyme which is dysreg-ulated in obese states and contributes to AT dysfunctionOur complementary in vitro and in vivo approaches revealthat reduced DEGS1 function impairs adipogenesis andlipogenesis programs and increases oxidative stress
DEGS1 is expressed in WAT and correlates in rodentswith fat mass in healthy states an association that is
Figure 4mdash3T3-L1 cells treated with C8-CPPC andor rosiglitazone A and B mRNA expression of genes involved in adipocyte differen-tiation and lipid accumulation at 24 and 48 h after induction C Protein expression levels of PPARg2 and cEBPa D Lipid content after 48and 96 h of differentiation Values are the mean 6 SEM of two separate experiments performed in triplicate One-way ANOVA was used toanalyze the statistical significance between treatments at 24 and 48 h Significant differences (Duncan test P lt 005) are indicated withdifferent letters E DhCer Cer and hexosylceramide (HexCer) levels after 48 h of differentiation Values are the mean 6 SEM of twoseparate experiments performed in triplicate cEBP CCAATenhancer binding protein d day FITC fluorescein isothiocyanate MFI meanfluorescence intensity Rosi rosiglitazone P lt 005
diabetesdiabetesjournalsorg Barbarroja and Associates 1187
disrupted in severe forms of obesity The expression ofDEGS1 in liver or muscle is not affected by obesity fur-ther indicating that the WAT is particularly susceptible tochanges in Cer metabolism in obese states The relevanceto human obesity is confirmed by the downregulation ofDEGS1 in the AT of MO patients and by recent reports(2324) showing that DhCer rather than Cer content pos-itively correlates with BMI and waist circumference incohorts of overweight obese subjects To date only a singlefunctional mutation in the degs1 gene has been reportedin humans where patients demonstrate increased serumDhCer decreased cholesterol esters and decreased waist-to-hip ratio (25) Globally considered these data suggest
the existence of an association between DEGS1 functionand fat mass
The molecular mechanism leading to the defective ATexpression of degs1 in human and rodent obesity is un-known Here we have shown that tumor necrosis factor-aa mediator of obesity-induced inflammation and insulinresistance decreases the transcriptional expression ofdegs1 in cultured adipocytes which suggests that lowchronic inflammation may contribute to DEGS1 dysfunc-tion Other obesity-associated processes such as in-creased oxidative stress and hypoxia have been shownto impair DEGS1 activity and increase DhCer levels (26)Our results also show a positive association between
Figure 5mdashC8-CPPC inhibitor increases proliferation and decreases lipid accumulation in preadipocytes in vivo Ten-week-old mice weregiven a 45 HFD for 5 weeks before being treated intraperitoneally with vehicle or C8-CPPC inhibitor (2 mgkgday) over 9 daysA Representative images of immunofluorescence analysis of gonadal AT of control (vehicle) and C8-CPPCndashtreated mice are presentedwith Pref-1 (cyan) and Ki67 (red white arrows) Nuclei and lipids are respectively stained with Hoechst stain (blue) and BODIPY (green)Scale bars 100 or 20 mm Quantification of preadipocytes (left graph) (percentage of Pref-1+ cellstotal cells) and (right graph) Ki67+ (whitebars)BODIPY+ (dark gray bars) cells among preadipocytes (Pref-1+ cells) (light gray bars) B mRNA expression of preadipocyte markersand pparg degs1 adiponectin caspase 3 and fas in gonadal AT of control and C8-CPPCndashtreated mice n = 7ndash8 mice per experimentalgroup Values are the mean 6SEM P lt 005 P lt 00001 vs vehicle
1188 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
Figure 6mdashA Glycerol release in 3T3-L1 adipocytes exposed for 48 h to C8-CPPC and stimulated with NE for 6 h BndashD Results fromWestern blot analysis for phosphorylated (p) HSLHSL ratio and total HSL in 3T3-L1 adipocytes exposed for 48 h to C8-CPPC andstimulated with NE for 6 h Graphs show the mean 6 SEM of two separated experiments P lt 005 E Effects of pharmacologicalinhibition of DEGS1 on insulin (ins) signaling Results from Western blot analysis for p-AKT and AKT in 3T3-L1 adipocytes exposed 48 or 72h with C8-CPPC 1 mmolL and increased concentrations of insulin (10ndash100 nmolL) for 15 min F and G Results from Western blot analysisfor Glut4 and adiponectin in 3T3-L1 adipocytes exposed with C8-CPPC 05ndash1 mmolL and insulin (100 nmolL) for 48 h H Results fromWestern blot analysis for AMPK in 3T3-L1 adipocytes exposed to C8-CPPC 05ndash1 mmolL and insulin (Ins) (100 nmolL) rosiglitazone (Rosi)(1 nmolL) and metformin (Metf) (100 nmolL) for 48 h aP lt 005 vs untreated cells (Ins Rosi or Metf effect) bP lt 005 vs 0 mmolLC8-CPPC (C8-CPPC effect) AU arbitrary units IOD integrated optical density
diabetesdiabetesjournalsorg Barbarroja and Associates 1189
levels of the proadipogenic pparg2 isoform and degs1in AT and given the documented downregulation ofpparg2 in AT in obese insulin-resistant states it is con-ceivable that decreased levels of pparg2 may contributeto the defective expression of degs1 associated withobesity
Studies in rodents have shown that the number of Cersis increased in the WAT of animals fed an HFD (5) con-comitantly with the onset of insulin resistance (6) Resultsfrom obob mice and obese human studies are more un-clear where both increased or decreased levels of Cershave been reported (78) This suggests that Cer synthesismay be dependent on the severity of the obese state andthe strength of the adaptive homeostatic responsesattempting to restore their normal balance
Our pharmacological and gene ablation studies haveshown that inhibition of DEGS1 is associated with oxida-tive stress cellular death impaired adipocyte differentia-tion impaired lipid accumulation and impaired basallipolysis in mature adipocytes
Our in vitro studies revealed that the defect in theadipogenesis program caused by depletion of DEGS1occurs during the early stages of adipogenesis (MCE)when PPARg controls the expression of genes that regu-late the cell cycle by directly interacting with cyclin D3Another cyclin involved in adipocyte differentiation iscyclin D1 and here we show that downregulation ofDEGS1 blocks PPARg activity and cyclins such as D1D3 E as well as decreasing cdk2 which modulates PPARgactivity during adipogenesis These data are consistent
Figure 7mdashA and B DhCers impair adipocyte differentiation during early adipogenesis 3T3-L1 preadipocytes treated with C2DhCer(50 mmolL) for a period of 48 h at day 0 or day 3 of differentiation Oil Red O staining and mRNA expression of adipogenesis and lipidmetabolism genes Values are the mean 6 SEM of two separate experiments performed in triplicate P lt 005 vs control C DhCersdecrease ligand-mediated PPARg transactivation Cells were treated with DMSO as a control group and rosiglitazone 10 mmolL GW192910 mmolL and C2 and C16DhCers and Cers 100 mmolL as indicated Graphs represent the average of three independent experimentsP lt 005 vs control cells P lt 005 vs rosiglitazone ampP lt 005 vs GW1929 CTL control
1190 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
with studies showing that downregulation of degs1 bysmall interfering RNA inhibits cell growth and arreststhe cell cycle in cancer cells (12) and where its overexpres-sion increased cell migration and metastasis (27)
The pharmacological inhibition of DEGS1 in vivovalidated the results obtained in vitro and strengthenedthe concept that functional degs1 is required for the fulldifferentiation of preadipocytes into mature adipocytes
We also provide evidence that both pharmacologicaland genetic ablation of degs1 which according to ourresults in in vitro models may directly block adipocytedifferentiation and lipid deposition certainly increasedDhCer levels in in vitro models In this line our resultsshowed that the treatment of adipocytes with DhCercauses impaired adipogenesis and lipid accumulationThis effect on lipid accumulation was more severewhen the treatment was administered during the first3 days of differentiation suggesting a direct impact ofDhCer accumulation during early stages of adipogenesisWe also provide evidence showing that accumulation ofDhCers can also directly repress the transcriptional ac-tivity of PPARg and hence could contribute to impair-ment of the capacity of the preadipocytes to developa full adipogenic program
These observations strengthen the concept that DhCersare not merely inert precursors of Cers Nevertheless themolecular mechanism linking DhCers accumulation anddysregulation of the cell cycle and other cellular eventsremains elusive One possible mechanism is that changesin the DhCerCer ratio may disrupt membrane-dependentstructures by altering the levels of cholesterol andorcaveolin in lipid rafts which are known to be highlysensitive to a sphingomyelin pool Alternatively changesin the DhCerCer ratio may disturb the global phospho-lipidome potentially altering membrane-associated pro-cesses relevant to adipocyte function These are importantquestions to address in future research
In summary our results indicate that defects in DEGS1in the context of the metabolic syndrome may compro-mise AT expansion and function through the combinedinhibition of adipogenesis promotion of cell death andoxidative stress due to the direct accumulation of DhCersThus our data suggest that the selective manipulation ofDEGS1 andor its substrates in WAT may be a pathophy-siologically advantageous strategy to improve AT homeo-stasis and ameliorate the burden of obesity-associatedmetabolic complications
Acknowledgments The authors thank Dr Carobbio Mrs Peirce andMs Philips (Wellcome Trust-Medical Research Council Institute of MetabolicScience University of Cambridge UK) for their excellent technical advice sup-port and suggestionsFunding This work was funded by the Medical Research Council (MRC) theMetabolic Diseases Unit of the MRC European Commission grant FP7-ETHERPATHSand the British Heart FoundationDuality of Interest No potential conflicts of interest relevant to this articlewere reported
Author Contributions NB helped to develop the hypothesis design theexperiments perform the in vitro and ex vivo experiments collect and analyze thedata hold and characterize the human cohorts collect gene expression data fromadipose depots and analyze the data discuss the manuscript coordinate anddirect the project and write the manuscript SR-C helped to develop thehypothesis design the experiments perform the in vitro and ex vivo experimentscollect and analyze the data discuss the manuscript coordinate and direct theproject and write the manuscript HN and MO helped to develop analyticalplatforms perform and analyze lipidomic experiments and discuss the manu-script AC AP JR IC VP GM-G and CL-P helped to perform the invitro and ex vivo experiments collect and analyze the data and discuss themanuscript FJT helped to hold and characterize the human cohorts collectgene expression data from adipose depots and analyze the data and discussthe manuscript JDS and SAS helped to design the experiments and dis-cuss the manuscript AV-P helped to develop the hypothesis design theexperiments discuss the manuscript coordinate and direct the project andwrite the manuscript AV-P is the guarantor of this work and as such had fullaccess to all the data in the study and takes responsibility for the integrity of thedata and the accuracy of the data analysis
References1 Montague CT OrsquoRahilly S The perils of portliness causes and con-
sequences of visceral adiposity Diabetes 200049883ndash8882 Carobbio S Rodriguez-Cuenca S Vidal-Puig A Origins of metabolic com-
plications in obesity ectopic fat accumulation The importance of the qualitative
aspect of lipotoxicity Curr Opin Clin Nutr Metab Care 201114520ndash5263 Chavez JA Summers SA Lipid oversupply selective insulin resistance
and lipotoxicity molecular mechanisms Biochim Biophys Acta 20101801
252ndash2654 Chavez JA Summers SA A ceramide-centric view of insulin resistance Cell
Metab 201215585ndash5945 Shah C Yang G Lee I Bielawski J Hannun YA Samad F Protection from
high fat diet-induced increase in ceramide in mice lacking plasminogen activator
inhibitor 1 J Biol Chem 200828313538ndash135486 Turner N Kowalski GM Leslie SJ et al Distinct patterns of tissue-specific
lipid accumulation during the induction of insulin resistance in mice by high-fat
feeding Diabetologia 2013561638ndash16487 Blachnio-Zabielska AU Koutsari C Tchkonia T Jensen MD Sphingolipid
content of human adipose tissue relationship to adiponectin and insulin re-
sistance Obesity (Silver Spring) 2012202341ndash23478 Błachnio-Zabielska AU Pułka M Baranowski M et al Ceramide metabo-
lism is affected by obesity and diabetes in human adipose tissue J Cell Physiol
2012227550ndash5579 Ternes P Franke S Zaumlhringer U Sperling P Heinz E Identification and
characterization of a sphingolipid delta 4-desaturase family J Biol Chem 2002
27725512ndash2551810 Mizutani Y Kihara A Igarashi Y Identification of the human sphingolipid C4-
hydroxylase hDES2 and its up-regulation during keratinocyte differentiation
FEBS Lett 200456393ndash9711 Ahn EH Schroeder JJ Induction of apoptosis by sphingosine sphinganine
and C(2)-ceramide in human colon cancer cells but not by C(2)-dihydroceramide
Anticancer Res 2010302881ndash288412 Kraveka JM Li L Szulc ZM et al Involvement of dihydroceramide desaturase
in cell cycle progression in human neuroblastoma cells J Biol Chem 2007282
16718ndash1672813 Siddique MM Li Y Wang L et al Ablation of dihydroceramide desaturase 1
a therapeutic target for the treatment of metabolic diseases simultaneously stim-
ulates anabolic and catabolic signaling Mol Cell Biol 2013332353ndash236914 Parks BW Nam E Org E et al Genetic control of obesity and gut microbiota
composition in response to high-fat high-sucrose diet in mice Cell Metab 2013
17141ndash152
diabetesdiabetesjournalsorg Barbarroja and Associates 1191
15 Bikman BT Guan Y Shui G et al Fenretinide prevents lipid-induced insulinresistance by blocking ceramide biosynthesis J Biol Chem 201228717426ndash1743716 Medina-Gomez G Gray SL Yetukuri L et al PPAR gamma 2 preventslipotoxicity by controlling adipose tissue expandability and peripheral lipid me-tabolism PLoS Genet 20073e6417 Roberts LD Virtue S Vidal-Puig A Nicholls AW Griffin JL Metabolic pheno-typing of a model of adipocyte differentiation Physiol Genomics 200939109ndash11918 Ramiacuterez-Zacariacuteas JL Castro-Muntildeozledo F Kuri-Harcuch W Quantitation ofadipose conversion and triglycerides by staining intracytoplasmic lipids with Oilred O Histochemistry 199297493ndash49719 Ye J Coulouris G Zaretskaya I Cutcutache I Rozen S Madden TL Primer-BLAST a tool to design target-specific primers for polymerase chain reactionBMC Bioinformatics 20121313420 Pfaffl MW Tichopad A Prgomet C Neuvians TP Determination of stablehousekeeping genes differentially regulated target genes and sample integrityBestKeepermdashExcel-based tool using pair-wise correlations Biotechnol Lett 200426509ndash51521 Triola G Fabriagraves G Llebaria A Synthesis of a cyclopropene analogue of cer-amide a potent inhibitor of dihydroceramide desaturase Angew Chem Int Ed Engl2001401960ndash1962
22 Tsuji K Satoh S Mitsutake S et al Evaluation of synthetic sphingolipidanalogs as ligands for peroxisome proliferator-activated receptors Bioorg MedChem Lett 2009191643ndash164623 Mamtani M Meikle PJ Kulkarni H et al Plasma dihydroceramide speciesassociate with waist circumference in Mexican American families Obesity (SilverSpring) 201422950ndash95624 Weir JM Wong G Barlow CK et al Plasma lipid profiling in a largepopulation-based cohort J Lipid Res 2013542898ndash290825 Curran J Meikle P Weir J Jowett J Deep sequencing in extended pedi-grees reveals a major rare non-synonymous variant influencing the de novoceramide synthesis pathway Late-breaking abstract presented at the AmericanSociety of Human Genetics 2013 Meeting 22ndash26 October 2013 at the BostonConvention and Exhibition Center Boston Massachusetts26 Idkowiak-Baldys J Takemoto JY Grilley MM Structure-function studies ofyeast C-4 sphingolipid long chain base hydroxylase Biochim Biophys Acta 2003161817ndash2427 Zhou W Ye XL Sun ZJ Ji XD Chen HX Xie D Overexpression of de-generative spermatocyte homolog 1 up-regulates the expression of cyclin D1 andenhances metastatic efficiency in esophageal carcinoma Eca109 cells MolCarcinog 200948886ndash894
1192 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015

The Impairment of Adipocyte Differentiation DuringEarly Adipogenesis by DEGS1 Inhibition IsRecapitulated by DhCers Per SeWe finally sought to validate whether DhCer by itself couldmediate the effects of DEGS1 inhibition on adipocytedifferentiation in our models of genetic or pharmacologicalinhibition of degs1 3T3-L1cells were treated with C2DhCerduring 1) the MCE at time 0 and 2) at day 3 after differ-entiation DhCer inhibited lipid accumulation (Fig 7A) andthe expression of genes involved during the early stages ofadipogenesis (Fig 7B)
Since synthetic sphingolipid analogs are known tomodulate PPAR activity (22) we investigated whetherboth C2DhCer and C16DhCer could exert a repressiveeffect on PPARg activation in the presence of the PPARgligands (Fig 7C) Our results show that DhCer can also
block the ligand-mediated transactivation of PPARg Sim-ilar results were obtained with Cers indicating that theseeffects were not a consequence of the unsaturation of thesphingoid moiety (data not shown)
DISCUSSION
The DhCerCer ratio has an important homeostaticregulatory role in the cell contributing to cell survivalautophagy and oxidative stress (1213) Here we identifyDEGS1 as an essential metabolic enzyme which is dysreg-ulated in obese states and contributes to AT dysfunctionOur complementary in vitro and in vivo approaches revealthat reduced DEGS1 function impairs adipogenesis andlipogenesis programs and increases oxidative stress
DEGS1 is expressed in WAT and correlates in rodentswith fat mass in healthy states an association that is
Figure 4mdash3T3-L1 cells treated with C8-CPPC andor rosiglitazone A and B mRNA expression of genes involved in adipocyte differen-tiation and lipid accumulation at 24 and 48 h after induction C Protein expression levels of PPARg2 and cEBPa D Lipid content after 48and 96 h of differentiation Values are the mean 6 SEM of two separate experiments performed in triplicate One-way ANOVA was used toanalyze the statistical significance between treatments at 24 and 48 h Significant differences (Duncan test P lt 005) are indicated withdifferent letters E DhCer Cer and hexosylceramide (HexCer) levels after 48 h of differentiation Values are the mean 6 SEM of twoseparate experiments performed in triplicate cEBP CCAATenhancer binding protein d day FITC fluorescein isothiocyanate MFI meanfluorescence intensity Rosi rosiglitazone P lt 005
diabetesdiabetesjournalsorg Barbarroja and Associates 1187
disrupted in severe forms of obesity The expression ofDEGS1 in liver or muscle is not affected by obesity fur-ther indicating that the WAT is particularly susceptible tochanges in Cer metabolism in obese states The relevanceto human obesity is confirmed by the downregulation ofDEGS1 in the AT of MO patients and by recent reports(2324) showing that DhCer rather than Cer content pos-itively correlates with BMI and waist circumference incohorts of overweight obese subjects To date only a singlefunctional mutation in the degs1 gene has been reportedin humans where patients demonstrate increased serumDhCer decreased cholesterol esters and decreased waist-to-hip ratio (25) Globally considered these data suggest
the existence of an association between DEGS1 functionand fat mass
The molecular mechanism leading to the defective ATexpression of degs1 in human and rodent obesity is un-known Here we have shown that tumor necrosis factor-aa mediator of obesity-induced inflammation and insulinresistance decreases the transcriptional expression ofdegs1 in cultured adipocytes which suggests that lowchronic inflammation may contribute to DEGS1 dysfunc-tion Other obesity-associated processes such as in-creased oxidative stress and hypoxia have been shownto impair DEGS1 activity and increase DhCer levels (26)Our results also show a positive association between
Figure 5mdashC8-CPPC inhibitor increases proliferation and decreases lipid accumulation in preadipocytes in vivo Ten-week-old mice weregiven a 45 HFD for 5 weeks before being treated intraperitoneally with vehicle or C8-CPPC inhibitor (2 mgkgday) over 9 daysA Representative images of immunofluorescence analysis of gonadal AT of control (vehicle) and C8-CPPCndashtreated mice are presentedwith Pref-1 (cyan) and Ki67 (red white arrows) Nuclei and lipids are respectively stained with Hoechst stain (blue) and BODIPY (green)Scale bars 100 or 20 mm Quantification of preadipocytes (left graph) (percentage of Pref-1+ cellstotal cells) and (right graph) Ki67+ (whitebars)BODIPY+ (dark gray bars) cells among preadipocytes (Pref-1+ cells) (light gray bars) B mRNA expression of preadipocyte markersand pparg degs1 adiponectin caspase 3 and fas in gonadal AT of control and C8-CPPCndashtreated mice n = 7ndash8 mice per experimentalgroup Values are the mean 6SEM P lt 005 P lt 00001 vs vehicle
1188 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
Figure 6mdashA Glycerol release in 3T3-L1 adipocytes exposed for 48 h to C8-CPPC and stimulated with NE for 6 h BndashD Results fromWestern blot analysis for phosphorylated (p) HSLHSL ratio and total HSL in 3T3-L1 adipocytes exposed for 48 h to C8-CPPC andstimulated with NE for 6 h Graphs show the mean 6 SEM of two separated experiments P lt 005 E Effects of pharmacologicalinhibition of DEGS1 on insulin (ins) signaling Results from Western blot analysis for p-AKT and AKT in 3T3-L1 adipocytes exposed 48 or 72h with C8-CPPC 1 mmolL and increased concentrations of insulin (10ndash100 nmolL) for 15 min F and G Results from Western blot analysisfor Glut4 and adiponectin in 3T3-L1 adipocytes exposed with C8-CPPC 05ndash1 mmolL and insulin (100 nmolL) for 48 h H Results fromWestern blot analysis for AMPK in 3T3-L1 adipocytes exposed to C8-CPPC 05ndash1 mmolL and insulin (Ins) (100 nmolL) rosiglitazone (Rosi)(1 nmolL) and metformin (Metf) (100 nmolL) for 48 h aP lt 005 vs untreated cells (Ins Rosi or Metf effect) bP lt 005 vs 0 mmolLC8-CPPC (C8-CPPC effect) AU arbitrary units IOD integrated optical density
diabetesdiabetesjournalsorg Barbarroja and Associates 1189
levels of the proadipogenic pparg2 isoform and degs1in AT and given the documented downregulation ofpparg2 in AT in obese insulin-resistant states it is con-ceivable that decreased levels of pparg2 may contributeto the defective expression of degs1 associated withobesity
Studies in rodents have shown that the number of Cersis increased in the WAT of animals fed an HFD (5) con-comitantly with the onset of insulin resistance (6) Resultsfrom obob mice and obese human studies are more un-clear where both increased or decreased levels of Cershave been reported (78) This suggests that Cer synthesismay be dependent on the severity of the obese state andthe strength of the adaptive homeostatic responsesattempting to restore their normal balance
Our pharmacological and gene ablation studies haveshown that inhibition of DEGS1 is associated with oxida-tive stress cellular death impaired adipocyte differentia-tion impaired lipid accumulation and impaired basallipolysis in mature adipocytes
Our in vitro studies revealed that the defect in theadipogenesis program caused by depletion of DEGS1occurs during the early stages of adipogenesis (MCE)when PPARg controls the expression of genes that regu-late the cell cycle by directly interacting with cyclin D3Another cyclin involved in adipocyte differentiation iscyclin D1 and here we show that downregulation ofDEGS1 blocks PPARg activity and cyclins such as D1D3 E as well as decreasing cdk2 which modulates PPARgactivity during adipogenesis These data are consistent
Figure 7mdashA and B DhCers impair adipocyte differentiation during early adipogenesis 3T3-L1 preadipocytes treated with C2DhCer(50 mmolL) for a period of 48 h at day 0 or day 3 of differentiation Oil Red O staining and mRNA expression of adipogenesis and lipidmetabolism genes Values are the mean 6 SEM of two separate experiments performed in triplicate P lt 005 vs control C DhCersdecrease ligand-mediated PPARg transactivation Cells were treated with DMSO as a control group and rosiglitazone 10 mmolL GW192910 mmolL and C2 and C16DhCers and Cers 100 mmolL as indicated Graphs represent the average of three independent experimentsP lt 005 vs control cells P lt 005 vs rosiglitazone ampP lt 005 vs GW1929 CTL control
1190 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
with studies showing that downregulation of degs1 bysmall interfering RNA inhibits cell growth and arreststhe cell cycle in cancer cells (12) and where its overexpres-sion increased cell migration and metastasis (27)
The pharmacological inhibition of DEGS1 in vivovalidated the results obtained in vitro and strengthenedthe concept that functional degs1 is required for the fulldifferentiation of preadipocytes into mature adipocytes
We also provide evidence that both pharmacologicaland genetic ablation of degs1 which according to ourresults in in vitro models may directly block adipocytedifferentiation and lipid deposition certainly increasedDhCer levels in in vitro models In this line our resultsshowed that the treatment of adipocytes with DhCercauses impaired adipogenesis and lipid accumulationThis effect on lipid accumulation was more severewhen the treatment was administered during the first3 days of differentiation suggesting a direct impact ofDhCer accumulation during early stages of adipogenesisWe also provide evidence showing that accumulation ofDhCers can also directly repress the transcriptional ac-tivity of PPARg and hence could contribute to impair-ment of the capacity of the preadipocytes to developa full adipogenic program
These observations strengthen the concept that DhCersare not merely inert precursors of Cers Nevertheless themolecular mechanism linking DhCers accumulation anddysregulation of the cell cycle and other cellular eventsremains elusive One possible mechanism is that changesin the DhCerCer ratio may disrupt membrane-dependentstructures by altering the levels of cholesterol andorcaveolin in lipid rafts which are known to be highlysensitive to a sphingomyelin pool Alternatively changesin the DhCerCer ratio may disturb the global phospho-lipidome potentially altering membrane-associated pro-cesses relevant to adipocyte function These are importantquestions to address in future research
In summary our results indicate that defects in DEGS1in the context of the metabolic syndrome may compro-mise AT expansion and function through the combinedinhibition of adipogenesis promotion of cell death andoxidative stress due to the direct accumulation of DhCersThus our data suggest that the selective manipulation ofDEGS1 andor its substrates in WAT may be a pathophy-siologically advantageous strategy to improve AT homeo-stasis and ameliorate the burden of obesity-associatedmetabolic complications
Acknowledgments The authors thank Dr Carobbio Mrs Peirce andMs Philips (Wellcome Trust-Medical Research Council Institute of MetabolicScience University of Cambridge UK) for their excellent technical advice sup-port and suggestionsFunding This work was funded by the Medical Research Council (MRC) theMetabolic Diseases Unit of the MRC European Commission grant FP7-ETHERPATHSand the British Heart FoundationDuality of Interest No potential conflicts of interest relevant to this articlewere reported
Author Contributions NB helped to develop the hypothesis design theexperiments perform the in vitro and ex vivo experiments collect and analyze thedata hold and characterize the human cohorts collect gene expression data fromadipose depots and analyze the data discuss the manuscript coordinate anddirect the project and write the manuscript SR-C helped to develop thehypothesis design the experiments perform the in vitro and ex vivo experimentscollect and analyze the data discuss the manuscript coordinate and direct theproject and write the manuscript HN and MO helped to develop analyticalplatforms perform and analyze lipidomic experiments and discuss the manu-script AC AP JR IC VP GM-G and CL-P helped to perform the invitro and ex vivo experiments collect and analyze the data and discuss themanuscript FJT helped to hold and characterize the human cohorts collectgene expression data from adipose depots and analyze the data and discussthe manuscript JDS and SAS helped to design the experiments and dis-cuss the manuscript AV-P helped to develop the hypothesis design theexperiments discuss the manuscript coordinate and direct the project andwrite the manuscript AV-P is the guarantor of this work and as such had fullaccess to all the data in the study and takes responsibility for the integrity of thedata and the accuracy of the data analysis
References1 Montague CT OrsquoRahilly S The perils of portliness causes and con-
sequences of visceral adiposity Diabetes 200049883ndash8882 Carobbio S Rodriguez-Cuenca S Vidal-Puig A Origins of metabolic com-
plications in obesity ectopic fat accumulation The importance of the qualitative
aspect of lipotoxicity Curr Opin Clin Nutr Metab Care 201114520ndash5263 Chavez JA Summers SA Lipid oversupply selective insulin resistance
and lipotoxicity molecular mechanisms Biochim Biophys Acta 20101801
252ndash2654 Chavez JA Summers SA A ceramide-centric view of insulin resistance Cell
Metab 201215585ndash5945 Shah C Yang G Lee I Bielawski J Hannun YA Samad F Protection from
high fat diet-induced increase in ceramide in mice lacking plasminogen activator
inhibitor 1 J Biol Chem 200828313538ndash135486 Turner N Kowalski GM Leslie SJ et al Distinct patterns of tissue-specific
lipid accumulation during the induction of insulin resistance in mice by high-fat
feeding Diabetologia 2013561638ndash16487 Blachnio-Zabielska AU Koutsari C Tchkonia T Jensen MD Sphingolipid
content of human adipose tissue relationship to adiponectin and insulin re-
sistance Obesity (Silver Spring) 2012202341ndash23478 Błachnio-Zabielska AU Pułka M Baranowski M et al Ceramide metabo-
lism is affected by obesity and diabetes in human adipose tissue J Cell Physiol
2012227550ndash5579 Ternes P Franke S Zaumlhringer U Sperling P Heinz E Identification and
characterization of a sphingolipid delta 4-desaturase family J Biol Chem 2002
27725512ndash2551810 Mizutani Y Kihara A Igarashi Y Identification of the human sphingolipid C4-
hydroxylase hDES2 and its up-regulation during keratinocyte differentiation
FEBS Lett 200456393ndash9711 Ahn EH Schroeder JJ Induction of apoptosis by sphingosine sphinganine
and C(2)-ceramide in human colon cancer cells but not by C(2)-dihydroceramide
Anticancer Res 2010302881ndash288412 Kraveka JM Li L Szulc ZM et al Involvement of dihydroceramide desaturase
in cell cycle progression in human neuroblastoma cells J Biol Chem 2007282
16718ndash1672813 Siddique MM Li Y Wang L et al Ablation of dihydroceramide desaturase 1
a therapeutic target for the treatment of metabolic diseases simultaneously stim-
ulates anabolic and catabolic signaling Mol Cell Biol 2013332353ndash236914 Parks BW Nam E Org E et al Genetic control of obesity and gut microbiota
composition in response to high-fat high-sucrose diet in mice Cell Metab 2013
17141ndash152
diabetesdiabetesjournalsorg Barbarroja and Associates 1191
15 Bikman BT Guan Y Shui G et al Fenretinide prevents lipid-induced insulinresistance by blocking ceramide biosynthesis J Biol Chem 201228717426ndash1743716 Medina-Gomez G Gray SL Yetukuri L et al PPAR gamma 2 preventslipotoxicity by controlling adipose tissue expandability and peripheral lipid me-tabolism PLoS Genet 20073e6417 Roberts LD Virtue S Vidal-Puig A Nicholls AW Griffin JL Metabolic pheno-typing of a model of adipocyte differentiation Physiol Genomics 200939109ndash11918 Ramiacuterez-Zacariacuteas JL Castro-Muntildeozledo F Kuri-Harcuch W Quantitation ofadipose conversion and triglycerides by staining intracytoplasmic lipids with Oilred O Histochemistry 199297493ndash49719 Ye J Coulouris G Zaretskaya I Cutcutache I Rozen S Madden TL Primer-BLAST a tool to design target-specific primers for polymerase chain reactionBMC Bioinformatics 20121313420 Pfaffl MW Tichopad A Prgomet C Neuvians TP Determination of stablehousekeeping genes differentially regulated target genes and sample integrityBestKeepermdashExcel-based tool using pair-wise correlations Biotechnol Lett 200426509ndash51521 Triola G Fabriagraves G Llebaria A Synthesis of a cyclopropene analogue of cer-amide a potent inhibitor of dihydroceramide desaturase Angew Chem Int Ed Engl2001401960ndash1962
22 Tsuji K Satoh S Mitsutake S et al Evaluation of synthetic sphingolipidanalogs as ligands for peroxisome proliferator-activated receptors Bioorg MedChem Lett 2009191643ndash164623 Mamtani M Meikle PJ Kulkarni H et al Plasma dihydroceramide speciesassociate with waist circumference in Mexican American families Obesity (SilverSpring) 201422950ndash95624 Weir JM Wong G Barlow CK et al Plasma lipid profiling in a largepopulation-based cohort J Lipid Res 2013542898ndash290825 Curran J Meikle P Weir J Jowett J Deep sequencing in extended pedi-grees reveals a major rare non-synonymous variant influencing the de novoceramide synthesis pathway Late-breaking abstract presented at the AmericanSociety of Human Genetics 2013 Meeting 22ndash26 October 2013 at the BostonConvention and Exhibition Center Boston Massachusetts26 Idkowiak-Baldys J Takemoto JY Grilley MM Structure-function studies ofyeast C-4 sphingolipid long chain base hydroxylase Biochim Biophys Acta 2003161817ndash2427 Zhou W Ye XL Sun ZJ Ji XD Chen HX Xie D Overexpression of de-generative spermatocyte homolog 1 up-regulates the expression of cyclin D1 andenhances metastatic efficiency in esophageal carcinoma Eca109 cells MolCarcinog 200948886ndash894
1192 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015

disrupted in severe forms of obesity The expression ofDEGS1 in liver or muscle is not affected by obesity fur-ther indicating that the WAT is particularly susceptible tochanges in Cer metabolism in obese states The relevanceto human obesity is confirmed by the downregulation ofDEGS1 in the AT of MO patients and by recent reports(2324) showing that DhCer rather than Cer content pos-itively correlates with BMI and waist circumference incohorts of overweight obese subjects To date only a singlefunctional mutation in the degs1 gene has been reportedin humans where patients demonstrate increased serumDhCer decreased cholesterol esters and decreased waist-to-hip ratio (25) Globally considered these data suggest
the existence of an association between DEGS1 functionand fat mass
The molecular mechanism leading to the defective ATexpression of degs1 in human and rodent obesity is un-known Here we have shown that tumor necrosis factor-aa mediator of obesity-induced inflammation and insulinresistance decreases the transcriptional expression ofdegs1 in cultured adipocytes which suggests that lowchronic inflammation may contribute to DEGS1 dysfunc-tion Other obesity-associated processes such as in-creased oxidative stress and hypoxia have been shownto impair DEGS1 activity and increase DhCer levels (26)Our results also show a positive association between
Figure 5mdashC8-CPPC inhibitor increases proliferation and decreases lipid accumulation in preadipocytes in vivo Ten-week-old mice weregiven a 45 HFD for 5 weeks before being treated intraperitoneally with vehicle or C8-CPPC inhibitor (2 mgkgday) over 9 daysA Representative images of immunofluorescence analysis of gonadal AT of control (vehicle) and C8-CPPCndashtreated mice are presentedwith Pref-1 (cyan) and Ki67 (red white arrows) Nuclei and lipids are respectively stained with Hoechst stain (blue) and BODIPY (green)Scale bars 100 or 20 mm Quantification of preadipocytes (left graph) (percentage of Pref-1+ cellstotal cells) and (right graph) Ki67+ (whitebars)BODIPY+ (dark gray bars) cells among preadipocytes (Pref-1+ cells) (light gray bars) B mRNA expression of preadipocyte markersand pparg degs1 adiponectin caspase 3 and fas in gonadal AT of control and C8-CPPCndashtreated mice n = 7ndash8 mice per experimentalgroup Values are the mean 6SEM P lt 005 P lt 00001 vs vehicle
1188 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
Figure 6mdashA Glycerol release in 3T3-L1 adipocytes exposed for 48 h to C8-CPPC and stimulated with NE for 6 h BndashD Results fromWestern blot analysis for phosphorylated (p) HSLHSL ratio and total HSL in 3T3-L1 adipocytes exposed for 48 h to C8-CPPC andstimulated with NE for 6 h Graphs show the mean 6 SEM of two separated experiments P lt 005 E Effects of pharmacologicalinhibition of DEGS1 on insulin (ins) signaling Results from Western blot analysis for p-AKT and AKT in 3T3-L1 adipocytes exposed 48 or 72h with C8-CPPC 1 mmolL and increased concentrations of insulin (10ndash100 nmolL) for 15 min F and G Results from Western blot analysisfor Glut4 and adiponectin in 3T3-L1 adipocytes exposed with C8-CPPC 05ndash1 mmolL and insulin (100 nmolL) for 48 h H Results fromWestern blot analysis for AMPK in 3T3-L1 adipocytes exposed to C8-CPPC 05ndash1 mmolL and insulin (Ins) (100 nmolL) rosiglitazone (Rosi)(1 nmolL) and metformin (Metf) (100 nmolL) for 48 h aP lt 005 vs untreated cells (Ins Rosi or Metf effect) bP lt 005 vs 0 mmolLC8-CPPC (C8-CPPC effect) AU arbitrary units IOD integrated optical density
diabetesdiabetesjournalsorg Barbarroja and Associates 1189
levels of the proadipogenic pparg2 isoform and degs1in AT and given the documented downregulation ofpparg2 in AT in obese insulin-resistant states it is con-ceivable that decreased levels of pparg2 may contributeto the defective expression of degs1 associated withobesity
Studies in rodents have shown that the number of Cersis increased in the WAT of animals fed an HFD (5) con-comitantly with the onset of insulin resistance (6) Resultsfrom obob mice and obese human studies are more un-clear where both increased or decreased levels of Cershave been reported (78) This suggests that Cer synthesismay be dependent on the severity of the obese state andthe strength of the adaptive homeostatic responsesattempting to restore their normal balance
Our pharmacological and gene ablation studies haveshown that inhibition of DEGS1 is associated with oxida-tive stress cellular death impaired adipocyte differentia-tion impaired lipid accumulation and impaired basallipolysis in mature adipocytes
Our in vitro studies revealed that the defect in theadipogenesis program caused by depletion of DEGS1occurs during the early stages of adipogenesis (MCE)when PPARg controls the expression of genes that regu-late the cell cycle by directly interacting with cyclin D3Another cyclin involved in adipocyte differentiation iscyclin D1 and here we show that downregulation ofDEGS1 blocks PPARg activity and cyclins such as D1D3 E as well as decreasing cdk2 which modulates PPARgactivity during adipogenesis These data are consistent
Figure 7mdashA and B DhCers impair adipocyte differentiation during early adipogenesis 3T3-L1 preadipocytes treated with C2DhCer(50 mmolL) for a period of 48 h at day 0 or day 3 of differentiation Oil Red O staining and mRNA expression of adipogenesis and lipidmetabolism genes Values are the mean 6 SEM of two separate experiments performed in triplicate P lt 005 vs control C DhCersdecrease ligand-mediated PPARg transactivation Cells were treated with DMSO as a control group and rosiglitazone 10 mmolL GW192910 mmolL and C2 and C16DhCers and Cers 100 mmolL as indicated Graphs represent the average of three independent experimentsP lt 005 vs control cells P lt 005 vs rosiglitazone ampP lt 005 vs GW1929 CTL control
1190 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
with studies showing that downregulation of degs1 bysmall interfering RNA inhibits cell growth and arreststhe cell cycle in cancer cells (12) and where its overexpres-sion increased cell migration and metastasis (27)
The pharmacological inhibition of DEGS1 in vivovalidated the results obtained in vitro and strengthenedthe concept that functional degs1 is required for the fulldifferentiation of preadipocytes into mature adipocytes
We also provide evidence that both pharmacologicaland genetic ablation of degs1 which according to ourresults in in vitro models may directly block adipocytedifferentiation and lipid deposition certainly increasedDhCer levels in in vitro models In this line our resultsshowed that the treatment of adipocytes with DhCercauses impaired adipogenesis and lipid accumulationThis effect on lipid accumulation was more severewhen the treatment was administered during the first3 days of differentiation suggesting a direct impact ofDhCer accumulation during early stages of adipogenesisWe also provide evidence showing that accumulation ofDhCers can also directly repress the transcriptional ac-tivity of PPARg and hence could contribute to impair-ment of the capacity of the preadipocytes to developa full adipogenic program
These observations strengthen the concept that DhCersare not merely inert precursors of Cers Nevertheless themolecular mechanism linking DhCers accumulation anddysregulation of the cell cycle and other cellular eventsremains elusive One possible mechanism is that changesin the DhCerCer ratio may disrupt membrane-dependentstructures by altering the levels of cholesterol andorcaveolin in lipid rafts which are known to be highlysensitive to a sphingomyelin pool Alternatively changesin the DhCerCer ratio may disturb the global phospho-lipidome potentially altering membrane-associated pro-cesses relevant to adipocyte function These are importantquestions to address in future research
In summary our results indicate that defects in DEGS1in the context of the metabolic syndrome may compro-mise AT expansion and function through the combinedinhibition of adipogenesis promotion of cell death andoxidative stress due to the direct accumulation of DhCersThus our data suggest that the selective manipulation ofDEGS1 andor its substrates in WAT may be a pathophy-siologically advantageous strategy to improve AT homeo-stasis and ameliorate the burden of obesity-associatedmetabolic complications
Acknowledgments The authors thank Dr Carobbio Mrs Peirce andMs Philips (Wellcome Trust-Medical Research Council Institute of MetabolicScience University of Cambridge UK) for their excellent technical advice sup-port and suggestionsFunding This work was funded by the Medical Research Council (MRC) theMetabolic Diseases Unit of the MRC European Commission grant FP7-ETHERPATHSand the British Heart FoundationDuality of Interest No potential conflicts of interest relevant to this articlewere reported
Author Contributions NB helped to develop the hypothesis design theexperiments perform the in vitro and ex vivo experiments collect and analyze thedata hold and characterize the human cohorts collect gene expression data fromadipose depots and analyze the data discuss the manuscript coordinate anddirect the project and write the manuscript SR-C helped to develop thehypothesis design the experiments perform the in vitro and ex vivo experimentscollect and analyze the data discuss the manuscript coordinate and direct theproject and write the manuscript HN and MO helped to develop analyticalplatforms perform and analyze lipidomic experiments and discuss the manu-script AC AP JR IC VP GM-G and CL-P helped to perform the invitro and ex vivo experiments collect and analyze the data and discuss themanuscript FJT helped to hold and characterize the human cohorts collectgene expression data from adipose depots and analyze the data and discussthe manuscript JDS and SAS helped to design the experiments and dis-cuss the manuscript AV-P helped to develop the hypothesis design theexperiments discuss the manuscript coordinate and direct the project andwrite the manuscript AV-P is the guarantor of this work and as such had fullaccess to all the data in the study and takes responsibility for the integrity of thedata and the accuracy of the data analysis
References1 Montague CT OrsquoRahilly S The perils of portliness causes and con-
sequences of visceral adiposity Diabetes 200049883ndash8882 Carobbio S Rodriguez-Cuenca S Vidal-Puig A Origins of metabolic com-
plications in obesity ectopic fat accumulation The importance of the qualitative
aspect of lipotoxicity Curr Opin Clin Nutr Metab Care 201114520ndash5263 Chavez JA Summers SA Lipid oversupply selective insulin resistance
and lipotoxicity molecular mechanisms Biochim Biophys Acta 20101801
252ndash2654 Chavez JA Summers SA A ceramide-centric view of insulin resistance Cell
Metab 201215585ndash5945 Shah C Yang G Lee I Bielawski J Hannun YA Samad F Protection from
high fat diet-induced increase in ceramide in mice lacking plasminogen activator
inhibitor 1 J Biol Chem 200828313538ndash135486 Turner N Kowalski GM Leslie SJ et al Distinct patterns of tissue-specific
lipid accumulation during the induction of insulin resistance in mice by high-fat
feeding Diabetologia 2013561638ndash16487 Blachnio-Zabielska AU Koutsari C Tchkonia T Jensen MD Sphingolipid
content of human adipose tissue relationship to adiponectin and insulin re-
sistance Obesity (Silver Spring) 2012202341ndash23478 Błachnio-Zabielska AU Pułka M Baranowski M et al Ceramide metabo-
lism is affected by obesity and diabetes in human adipose tissue J Cell Physiol
2012227550ndash5579 Ternes P Franke S Zaumlhringer U Sperling P Heinz E Identification and
characterization of a sphingolipid delta 4-desaturase family J Biol Chem 2002
27725512ndash2551810 Mizutani Y Kihara A Igarashi Y Identification of the human sphingolipid C4-
hydroxylase hDES2 and its up-regulation during keratinocyte differentiation
FEBS Lett 200456393ndash9711 Ahn EH Schroeder JJ Induction of apoptosis by sphingosine sphinganine
and C(2)-ceramide in human colon cancer cells but not by C(2)-dihydroceramide
Anticancer Res 2010302881ndash288412 Kraveka JM Li L Szulc ZM et al Involvement of dihydroceramide desaturase
in cell cycle progression in human neuroblastoma cells J Biol Chem 2007282
16718ndash1672813 Siddique MM Li Y Wang L et al Ablation of dihydroceramide desaturase 1
a therapeutic target for the treatment of metabolic diseases simultaneously stim-
ulates anabolic and catabolic signaling Mol Cell Biol 2013332353ndash236914 Parks BW Nam E Org E et al Genetic control of obesity and gut microbiota
composition in response to high-fat high-sucrose diet in mice Cell Metab 2013
17141ndash152
diabetesdiabetesjournalsorg Barbarroja and Associates 1191
15 Bikman BT Guan Y Shui G et al Fenretinide prevents lipid-induced insulinresistance by blocking ceramide biosynthesis J Biol Chem 201228717426ndash1743716 Medina-Gomez G Gray SL Yetukuri L et al PPAR gamma 2 preventslipotoxicity by controlling adipose tissue expandability and peripheral lipid me-tabolism PLoS Genet 20073e6417 Roberts LD Virtue S Vidal-Puig A Nicholls AW Griffin JL Metabolic pheno-typing of a model of adipocyte differentiation Physiol Genomics 200939109ndash11918 Ramiacuterez-Zacariacuteas JL Castro-Muntildeozledo F Kuri-Harcuch W Quantitation ofadipose conversion and triglycerides by staining intracytoplasmic lipids with Oilred O Histochemistry 199297493ndash49719 Ye J Coulouris G Zaretskaya I Cutcutache I Rozen S Madden TL Primer-BLAST a tool to design target-specific primers for polymerase chain reactionBMC Bioinformatics 20121313420 Pfaffl MW Tichopad A Prgomet C Neuvians TP Determination of stablehousekeeping genes differentially regulated target genes and sample integrityBestKeepermdashExcel-based tool using pair-wise correlations Biotechnol Lett 200426509ndash51521 Triola G Fabriagraves G Llebaria A Synthesis of a cyclopropene analogue of cer-amide a potent inhibitor of dihydroceramide desaturase Angew Chem Int Ed Engl2001401960ndash1962
22 Tsuji K Satoh S Mitsutake S et al Evaluation of synthetic sphingolipidanalogs as ligands for peroxisome proliferator-activated receptors Bioorg MedChem Lett 2009191643ndash164623 Mamtani M Meikle PJ Kulkarni H et al Plasma dihydroceramide speciesassociate with waist circumference in Mexican American families Obesity (SilverSpring) 201422950ndash95624 Weir JM Wong G Barlow CK et al Plasma lipid profiling in a largepopulation-based cohort J Lipid Res 2013542898ndash290825 Curran J Meikle P Weir J Jowett J Deep sequencing in extended pedi-grees reveals a major rare non-synonymous variant influencing the de novoceramide synthesis pathway Late-breaking abstract presented at the AmericanSociety of Human Genetics 2013 Meeting 22ndash26 October 2013 at the BostonConvention and Exhibition Center Boston Massachusetts26 Idkowiak-Baldys J Takemoto JY Grilley MM Structure-function studies ofyeast C-4 sphingolipid long chain base hydroxylase Biochim Biophys Acta 2003161817ndash2427 Zhou W Ye XL Sun ZJ Ji XD Chen HX Xie D Overexpression of de-generative spermatocyte homolog 1 up-regulates the expression of cyclin D1 andenhances metastatic efficiency in esophageal carcinoma Eca109 cells MolCarcinog 200948886ndash894
1192 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
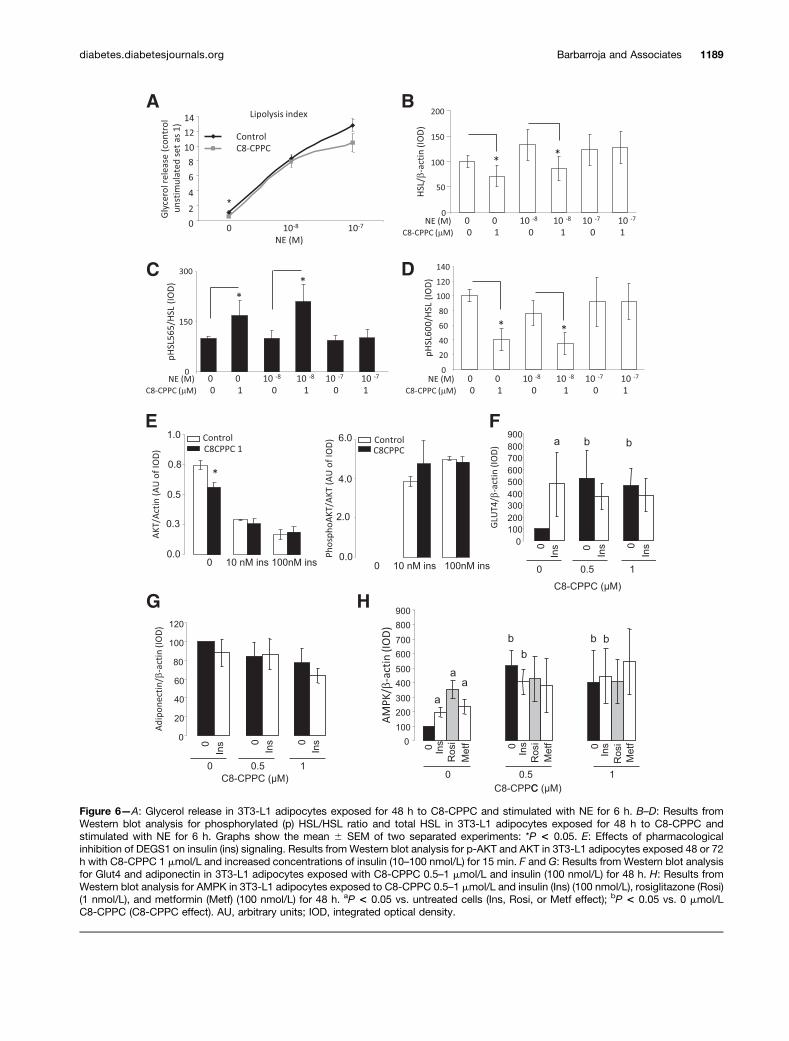
Figure 6mdashA Glycerol release in 3T3-L1 adipocytes exposed for 48 h to C8-CPPC and stimulated with NE for 6 h BndashD Results fromWestern blot analysis for phosphorylated (p) HSLHSL ratio and total HSL in 3T3-L1 adipocytes exposed for 48 h to C8-CPPC andstimulated with NE for 6 h Graphs show the mean 6 SEM of two separated experiments P lt 005 E Effects of pharmacologicalinhibition of DEGS1 on insulin (ins) signaling Results from Western blot analysis for p-AKT and AKT in 3T3-L1 adipocytes exposed 48 or 72h with C8-CPPC 1 mmolL and increased concentrations of insulin (10ndash100 nmolL) for 15 min F and G Results from Western blot analysisfor Glut4 and adiponectin in 3T3-L1 adipocytes exposed with C8-CPPC 05ndash1 mmolL and insulin (100 nmolL) for 48 h H Results fromWestern blot analysis for AMPK in 3T3-L1 adipocytes exposed to C8-CPPC 05ndash1 mmolL and insulin (Ins) (100 nmolL) rosiglitazone (Rosi)(1 nmolL) and metformin (Metf) (100 nmolL) for 48 h aP lt 005 vs untreated cells (Ins Rosi or Metf effect) bP lt 005 vs 0 mmolLC8-CPPC (C8-CPPC effect) AU arbitrary units IOD integrated optical density
diabetesdiabetesjournalsorg Barbarroja and Associates 1189
levels of the proadipogenic pparg2 isoform and degs1in AT and given the documented downregulation ofpparg2 in AT in obese insulin-resistant states it is con-ceivable that decreased levels of pparg2 may contributeto the defective expression of degs1 associated withobesity
Studies in rodents have shown that the number of Cersis increased in the WAT of animals fed an HFD (5) con-comitantly with the onset of insulin resistance (6) Resultsfrom obob mice and obese human studies are more un-clear where both increased or decreased levels of Cershave been reported (78) This suggests that Cer synthesismay be dependent on the severity of the obese state andthe strength of the adaptive homeostatic responsesattempting to restore their normal balance
Our pharmacological and gene ablation studies haveshown that inhibition of DEGS1 is associated with oxida-tive stress cellular death impaired adipocyte differentia-tion impaired lipid accumulation and impaired basallipolysis in mature adipocytes
Our in vitro studies revealed that the defect in theadipogenesis program caused by depletion of DEGS1occurs during the early stages of adipogenesis (MCE)when PPARg controls the expression of genes that regu-late the cell cycle by directly interacting with cyclin D3Another cyclin involved in adipocyte differentiation iscyclin D1 and here we show that downregulation ofDEGS1 blocks PPARg activity and cyclins such as D1D3 E as well as decreasing cdk2 which modulates PPARgactivity during adipogenesis These data are consistent
Figure 7mdashA and B DhCers impair adipocyte differentiation during early adipogenesis 3T3-L1 preadipocytes treated with C2DhCer(50 mmolL) for a period of 48 h at day 0 or day 3 of differentiation Oil Red O staining and mRNA expression of adipogenesis and lipidmetabolism genes Values are the mean 6 SEM of two separate experiments performed in triplicate P lt 005 vs control C DhCersdecrease ligand-mediated PPARg transactivation Cells were treated with DMSO as a control group and rosiglitazone 10 mmolL GW192910 mmolL and C2 and C16DhCers and Cers 100 mmolL as indicated Graphs represent the average of three independent experimentsP lt 005 vs control cells P lt 005 vs rosiglitazone ampP lt 005 vs GW1929 CTL control
1190 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
with studies showing that downregulation of degs1 bysmall interfering RNA inhibits cell growth and arreststhe cell cycle in cancer cells (12) and where its overexpres-sion increased cell migration and metastasis (27)
The pharmacological inhibition of DEGS1 in vivovalidated the results obtained in vitro and strengthenedthe concept that functional degs1 is required for the fulldifferentiation of preadipocytes into mature adipocytes
We also provide evidence that both pharmacologicaland genetic ablation of degs1 which according to ourresults in in vitro models may directly block adipocytedifferentiation and lipid deposition certainly increasedDhCer levels in in vitro models In this line our resultsshowed that the treatment of adipocytes with DhCercauses impaired adipogenesis and lipid accumulationThis effect on lipid accumulation was more severewhen the treatment was administered during the first3 days of differentiation suggesting a direct impact ofDhCer accumulation during early stages of adipogenesisWe also provide evidence showing that accumulation ofDhCers can also directly repress the transcriptional ac-tivity of PPARg and hence could contribute to impair-ment of the capacity of the preadipocytes to developa full adipogenic program
These observations strengthen the concept that DhCersare not merely inert precursors of Cers Nevertheless themolecular mechanism linking DhCers accumulation anddysregulation of the cell cycle and other cellular eventsremains elusive One possible mechanism is that changesin the DhCerCer ratio may disrupt membrane-dependentstructures by altering the levels of cholesterol andorcaveolin in lipid rafts which are known to be highlysensitive to a sphingomyelin pool Alternatively changesin the DhCerCer ratio may disturb the global phospho-lipidome potentially altering membrane-associated pro-cesses relevant to adipocyte function These are importantquestions to address in future research
In summary our results indicate that defects in DEGS1in the context of the metabolic syndrome may compro-mise AT expansion and function through the combinedinhibition of adipogenesis promotion of cell death andoxidative stress due to the direct accumulation of DhCersThus our data suggest that the selective manipulation ofDEGS1 andor its substrates in WAT may be a pathophy-siologically advantageous strategy to improve AT homeo-stasis and ameliorate the burden of obesity-associatedmetabolic complications
Acknowledgments The authors thank Dr Carobbio Mrs Peirce andMs Philips (Wellcome Trust-Medical Research Council Institute of MetabolicScience University of Cambridge UK) for their excellent technical advice sup-port and suggestionsFunding This work was funded by the Medical Research Council (MRC) theMetabolic Diseases Unit of the MRC European Commission grant FP7-ETHERPATHSand the British Heart FoundationDuality of Interest No potential conflicts of interest relevant to this articlewere reported
Author Contributions NB helped to develop the hypothesis design theexperiments perform the in vitro and ex vivo experiments collect and analyze thedata hold and characterize the human cohorts collect gene expression data fromadipose depots and analyze the data discuss the manuscript coordinate anddirect the project and write the manuscript SR-C helped to develop thehypothesis design the experiments perform the in vitro and ex vivo experimentscollect and analyze the data discuss the manuscript coordinate and direct theproject and write the manuscript HN and MO helped to develop analyticalplatforms perform and analyze lipidomic experiments and discuss the manu-script AC AP JR IC VP GM-G and CL-P helped to perform the invitro and ex vivo experiments collect and analyze the data and discuss themanuscript FJT helped to hold and characterize the human cohorts collectgene expression data from adipose depots and analyze the data and discussthe manuscript JDS and SAS helped to design the experiments and dis-cuss the manuscript AV-P helped to develop the hypothesis design theexperiments discuss the manuscript coordinate and direct the project andwrite the manuscript AV-P is the guarantor of this work and as such had fullaccess to all the data in the study and takes responsibility for the integrity of thedata and the accuracy of the data analysis
References1 Montague CT OrsquoRahilly S The perils of portliness causes and con-
sequences of visceral adiposity Diabetes 200049883ndash8882 Carobbio S Rodriguez-Cuenca S Vidal-Puig A Origins of metabolic com-
plications in obesity ectopic fat accumulation The importance of the qualitative
aspect of lipotoxicity Curr Opin Clin Nutr Metab Care 201114520ndash5263 Chavez JA Summers SA Lipid oversupply selective insulin resistance
and lipotoxicity molecular mechanisms Biochim Biophys Acta 20101801
252ndash2654 Chavez JA Summers SA A ceramide-centric view of insulin resistance Cell
Metab 201215585ndash5945 Shah C Yang G Lee I Bielawski J Hannun YA Samad F Protection from
high fat diet-induced increase in ceramide in mice lacking plasminogen activator
inhibitor 1 J Biol Chem 200828313538ndash135486 Turner N Kowalski GM Leslie SJ et al Distinct patterns of tissue-specific
lipid accumulation during the induction of insulin resistance in mice by high-fat
feeding Diabetologia 2013561638ndash16487 Blachnio-Zabielska AU Koutsari C Tchkonia T Jensen MD Sphingolipid
content of human adipose tissue relationship to adiponectin and insulin re-
sistance Obesity (Silver Spring) 2012202341ndash23478 Błachnio-Zabielska AU Pułka M Baranowski M et al Ceramide metabo-
lism is affected by obesity and diabetes in human adipose tissue J Cell Physiol
2012227550ndash5579 Ternes P Franke S Zaumlhringer U Sperling P Heinz E Identification and
characterization of a sphingolipid delta 4-desaturase family J Biol Chem 2002
27725512ndash2551810 Mizutani Y Kihara A Igarashi Y Identification of the human sphingolipid C4-
hydroxylase hDES2 and its up-regulation during keratinocyte differentiation
FEBS Lett 200456393ndash9711 Ahn EH Schroeder JJ Induction of apoptosis by sphingosine sphinganine
and C(2)-ceramide in human colon cancer cells but not by C(2)-dihydroceramide
Anticancer Res 2010302881ndash288412 Kraveka JM Li L Szulc ZM et al Involvement of dihydroceramide desaturase
in cell cycle progression in human neuroblastoma cells J Biol Chem 2007282
16718ndash1672813 Siddique MM Li Y Wang L et al Ablation of dihydroceramide desaturase 1
a therapeutic target for the treatment of metabolic diseases simultaneously stim-
ulates anabolic and catabolic signaling Mol Cell Biol 2013332353ndash236914 Parks BW Nam E Org E et al Genetic control of obesity and gut microbiota
composition in response to high-fat high-sucrose diet in mice Cell Metab 2013
17141ndash152
diabetesdiabetesjournalsorg Barbarroja and Associates 1191
15 Bikman BT Guan Y Shui G et al Fenretinide prevents lipid-induced insulinresistance by blocking ceramide biosynthesis J Biol Chem 201228717426ndash1743716 Medina-Gomez G Gray SL Yetukuri L et al PPAR gamma 2 preventslipotoxicity by controlling adipose tissue expandability and peripheral lipid me-tabolism PLoS Genet 20073e6417 Roberts LD Virtue S Vidal-Puig A Nicholls AW Griffin JL Metabolic pheno-typing of a model of adipocyte differentiation Physiol Genomics 200939109ndash11918 Ramiacuterez-Zacariacuteas JL Castro-Muntildeozledo F Kuri-Harcuch W Quantitation ofadipose conversion and triglycerides by staining intracytoplasmic lipids with Oilred O Histochemistry 199297493ndash49719 Ye J Coulouris G Zaretskaya I Cutcutache I Rozen S Madden TL Primer-BLAST a tool to design target-specific primers for polymerase chain reactionBMC Bioinformatics 20121313420 Pfaffl MW Tichopad A Prgomet C Neuvians TP Determination of stablehousekeeping genes differentially regulated target genes and sample integrityBestKeepermdashExcel-based tool using pair-wise correlations Biotechnol Lett 200426509ndash51521 Triola G Fabriagraves G Llebaria A Synthesis of a cyclopropene analogue of cer-amide a potent inhibitor of dihydroceramide desaturase Angew Chem Int Ed Engl2001401960ndash1962
22 Tsuji K Satoh S Mitsutake S et al Evaluation of synthetic sphingolipidanalogs as ligands for peroxisome proliferator-activated receptors Bioorg MedChem Lett 2009191643ndash164623 Mamtani M Meikle PJ Kulkarni H et al Plasma dihydroceramide speciesassociate with waist circumference in Mexican American families Obesity (SilverSpring) 201422950ndash95624 Weir JM Wong G Barlow CK et al Plasma lipid profiling in a largepopulation-based cohort J Lipid Res 2013542898ndash290825 Curran J Meikle P Weir J Jowett J Deep sequencing in extended pedi-grees reveals a major rare non-synonymous variant influencing the de novoceramide synthesis pathway Late-breaking abstract presented at the AmericanSociety of Human Genetics 2013 Meeting 22ndash26 October 2013 at the BostonConvention and Exhibition Center Boston Massachusetts26 Idkowiak-Baldys J Takemoto JY Grilley MM Structure-function studies ofyeast C-4 sphingolipid long chain base hydroxylase Biochim Biophys Acta 2003161817ndash2427 Zhou W Ye XL Sun ZJ Ji XD Chen HX Xie D Overexpression of de-generative spermatocyte homolog 1 up-regulates the expression of cyclin D1 andenhances metastatic efficiency in esophageal carcinoma Eca109 cells MolCarcinog 200948886ndash894
1192 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
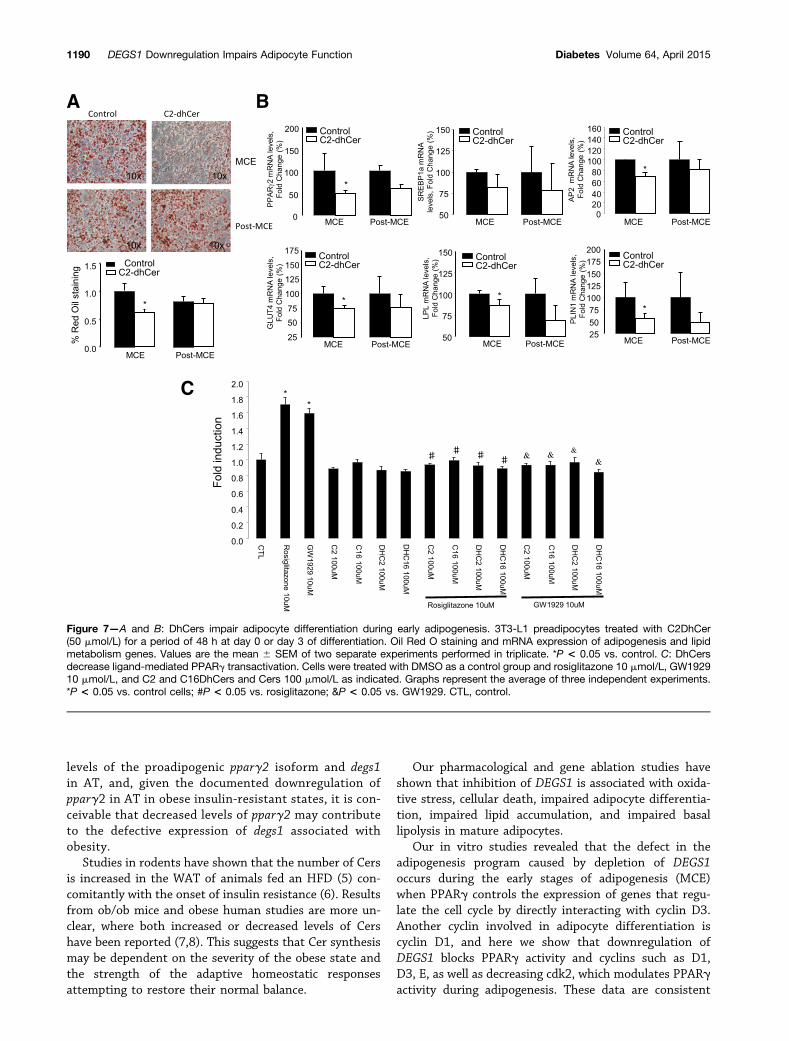
levels of the proadipogenic pparg2 isoform and degs1in AT and given the documented downregulation ofpparg2 in AT in obese insulin-resistant states it is con-ceivable that decreased levels of pparg2 may contributeto the defective expression of degs1 associated withobesity
Studies in rodents have shown that the number of Cersis increased in the WAT of animals fed an HFD (5) con-comitantly with the onset of insulin resistance (6) Resultsfrom obob mice and obese human studies are more un-clear where both increased or decreased levels of Cershave been reported (78) This suggests that Cer synthesismay be dependent on the severity of the obese state andthe strength of the adaptive homeostatic responsesattempting to restore their normal balance
Our pharmacological and gene ablation studies haveshown that inhibition of DEGS1 is associated with oxida-tive stress cellular death impaired adipocyte differentia-tion impaired lipid accumulation and impaired basallipolysis in mature adipocytes
Our in vitro studies revealed that the defect in theadipogenesis program caused by depletion of DEGS1occurs during the early stages of adipogenesis (MCE)when PPARg controls the expression of genes that regu-late the cell cycle by directly interacting with cyclin D3Another cyclin involved in adipocyte differentiation iscyclin D1 and here we show that downregulation ofDEGS1 blocks PPARg activity and cyclins such as D1D3 E as well as decreasing cdk2 which modulates PPARgactivity during adipogenesis These data are consistent
Figure 7mdashA and B DhCers impair adipocyte differentiation during early adipogenesis 3T3-L1 preadipocytes treated with C2DhCer(50 mmolL) for a period of 48 h at day 0 or day 3 of differentiation Oil Red O staining and mRNA expression of adipogenesis and lipidmetabolism genes Values are the mean 6 SEM of two separate experiments performed in triplicate P lt 005 vs control C DhCersdecrease ligand-mediated PPARg transactivation Cells were treated with DMSO as a control group and rosiglitazone 10 mmolL GW192910 mmolL and C2 and C16DhCers and Cers 100 mmolL as indicated Graphs represent the average of three independent experimentsP lt 005 vs control cells P lt 005 vs rosiglitazone ampP lt 005 vs GW1929 CTL control
1190 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015
with studies showing that downregulation of degs1 bysmall interfering RNA inhibits cell growth and arreststhe cell cycle in cancer cells (12) and where its overexpres-sion increased cell migration and metastasis (27)
The pharmacological inhibition of DEGS1 in vivovalidated the results obtained in vitro and strengthenedthe concept that functional degs1 is required for the fulldifferentiation of preadipocytes into mature adipocytes
We also provide evidence that both pharmacologicaland genetic ablation of degs1 which according to ourresults in in vitro models may directly block adipocytedifferentiation and lipid deposition certainly increasedDhCer levels in in vitro models In this line our resultsshowed that the treatment of adipocytes with DhCercauses impaired adipogenesis and lipid accumulationThis effect on lipid accumulation was more severewhen the treatment was administered during the first3 days of differentiation suggesting a direct impact ofDhCer accumulation during early stages of adipogenesisWe also provide evidence showing that accumulation ofDhCers can also directly repress the transcriptional ac-tivity of PPARg and hence could contribute to impair-ment of the capacity of the preadipocytes to developa full adipogenic program
These observations strengthen the concept that DhCersare not merely inert precursors of Cers Nevertheless themolecular mechanism linking DhCers accumulation anddysregulation of the cell cycle and other cellular eventsremains elusive One possible mechanism is that changesin the DhCerCer ratio may disrupt membrane-dependentstructures by altering the levels of cholesterol andorcaveolin in lipid rafts which are known to be highlysensitive to a sphingomyelin pool Alternatively changesin the DhCerCer ratio may disturb the global phospho-lipidome potentially altering membrane-associated pro-cesses relevant to adipocyte function These are importantquestions to address in future research
In summary our results indicate that defects in DEGS1in the context of the metabolic syndrome may compro-mise AT expansion and function through the combinedinhibition of adipogenesis promotion of cell death andoxidative stress due to the direct accumulation of DhCersThus our data suggest that the selective manipulation ofDEGS1 andor its substrates in WAT may be a pathophy-siologically advantageous strategy to improve AT homeo-stasis and ameliorate the burden of obesity-associatedmetabolic complications
Acknowledgments The authors thank Dr Carobbio Mrs Peirce andMs Philips (Wellcome Trust-Medical Research Council Institute of MetabolicScience University of Cambridge UK) for their excellent technical advice sup-port and suggestionsFunding This work was funded by the Medical Research Council (MRC) theMetabolic Diseases Unit of the MRC European Commission grant FP7-ETHERPATHSand the British Heart FoundationDuality of Interest No potential conflicts of interest relevant to this articlewere reported
Author Contributions NB helped to develop the hypothesis design theexperiments perform the in vitro and ex vivo experiments collect and analyze thedata hold and characterize the human cohorts collect gene expression data fromadipose depots and analyze the data discuss the manuscript coordinate anddirect the project and write the manuscript SR-C helped to develop thehypothesis design the experiments perform the in vitro and ex vivo experimentscollect and analyze the data discuss the manuscript coordinate and direct theproject and write the manuscript HN and MO helped to develop analyticalplatforms perform and analyze lipidomic experiments and discuss the manu-script AC AP JR IC VP GM-G and CL-P helped to perform the invitro and ex vivo experiments collect and analyze the data and discuss themanuscript FJT helped to hold and characterize the human cohorts collectgene expression data from adipose depots and analyze the data and discussthe manuscript JDS and SAS helped to design the experiments and dis-cuss the manuscript AV-P helped to develop the hypothesis design theexperiments discuss the manuscript coordinate and direct the project andwrite the manuscript AV-P is the guarantor of this work and as such had fullaccess to all the data in the study and takes responsibility for the integrity of thedata and the accuracy of the data analysis
References1 Montague CT OrsquoRahilly S The perils of portliness causes and con-
sequences of visceral adiposity Diabetes 200049883ndash8882 Carobbio S Rodriguez-Cuenca S Vidal-Puig A Origins of metabolic com-
plications in obesity ectopic fat accumulation The importance of the qualitative
aspect of lipotoxicity Curr Opin Clin Nutr Metab Care 201114520ndash5263 Chavez JA Summers SA Lipid oversupply selective insulin resistance
and lipotoxicity molecular mechanisms Biochim Biophys Acta 20101801
252ndash2654 Chavez JA Summers SA A ceramide-centric view of insulin resistance Cell
Metab 201215585ndash5945 Shah C Yang G Lee I Bielawski J Hannun YA Samad F Protection from
high fat diet-induced increase in ceramide in mice lacking plasminogen activator
inhibitor 1 J Biol Chem 200828313538ndash135486 Turner N Kowalski GM Leslie SJ et al Distinct patterns of tissue-specific
lipid accumulation during the induction of insulin resistance in mice by high-fat
feeding Diabetologia 2013561638ndash16487 Blachnio-Zabielska AU Koutsari C Tchkonia T Jensen MD Sphingolipid
content of human adipose tissue relationship to adiponectin and insulin re-
sistance Obesity (Silver Spring) 2012202341ndash23478 Błachnio-Zabielska AU Pułka M Baranowski M et al Ceramide metabo-
lism is affected by obesity and diabetes in human adipose tissue J Cell Physiol
2012227550ndash5579 Ternes P Franke S Zaumlhringer U Sperling P Heinz E Identification and
characterization of a sphingolipid delta 4-desaturase family J Biol Chem 2002
27725512ndash2551810 Mizutani Y Kihara A Igarashi Y Identification of the human sphingolipid C4-
hydroxylase hDES2 and its up-regulation during keratinocyte differentiation
FEBS Lett 200456393ndash9711 Ahn EH Schroeder JJ Induction of apoptosis by sphingosine sphinganine
and C(2)-ceramide in human colon cancer cells but not by C(2)-dihydroceramide
Anticancer Res 2010302881ndash288412 Kraveka JM Li L Szulc ZM et al Involvement of dihydroceramide desaturase
in cell cycle progression in human neuroblastoma cells J Biol Chem 2007282
16718ndash1672813 Siddique MM Li Y Wang L et al Ablation of dihydroceramide desaturase 1
a therapeutic target for the treatment of metabolic diseases simultaneously stim-
ulates anabolic and catabolic signaling Mol Cell Biol 2013332353ndash236914 Parks BW Nam E Org E et al Genetic control of obesity and gut microbiota
composition in response to high-fat high-sucrose diet in mice Cell Metab 2013
17141ndash152
diabetesdiabetesjournalsorg Barbarroja and Associates 1191
15 Bikman BT Guan Y Shui G et al Fenretinide prevents lipid-induced insulinresistance by blocking ceramide biosynthesis J Biol Chem 201228717426ndash1743716 Medina-Gomez G Gray SL Yetukuri L et al PPAR gamma 2 preventslipotoxicity by controlling adipose tissue expandability and peripheral lipid me-tabolism PLoS Genet 20073e6417 Roberts LD Virtue S Vidal-Puig A Nicholls AW Griffin JL Metabolic pheno-typing of a model of adipocyte differentiation Physiol Genomics 200939109ndash11918 Ramiacuterez-Zacariacuteas JL Castro-Muntildeozledo F Kuri-Harcuch W Quantitation ofadipose conversion and triglycerides by staining intracytoplasmic lipids with Oilred O Histochemistry 199297493ndash49719 Ye J Coulouris G Zaretskaya I Cutcutache I Rozen S Madden TL Primer-BLAST a tool to design target-specific primers for polymerase chain reactionBMC Bioinformatics 20121313420 Pfaffl MW Tichopad A Prgomet C Neuvians TP Determination of stablehousekeeping genes differentially regulated target genes and sample integrityBestKeepermdashExcel-based tool using pair-wise correlations Biotechnol Lett 200426509ndash51521 Triola G Fabriagraves G Llebaria A Synthesis of a cyclopropene analogue of cer-amide a potent inhibitor of dihydroceramide desaturase Angew Chem Int Ed Engl2001401960ndash1962
22 Tsuji K Satoh S Mitsutake S et al Evaluation of synthetic sphingolipidanalogs as ligands for peroxisome proliferator-activated receptors Bioorg MedChem Lett 2009191643ndash164623 Mamtani M Meikle PJ Kulkarni H et al Plasma dihydroceramide speciesassociate with waist circumference in Mexican American families Obesity (SilverSpring) 201422950ndash95624 Weir JM Wong G Barlow CK et al Plasma lipid profiling in a largepopulation-based cohort J Lipid Res 2013542898ndash290825 Curran J Meikle P Weir J Jowett J Deep sequencing in extended pedi-grees reveals a major rare non-synonymous variant influencing the de novoceramide synthesis pathway Late-breaking abstract presented at the AmericanSociety of Human Genetics 2013 Meeting 22ndash26 October 2013 at the BostonConvention and Exhibition Center Boston Massachusetts26 Idkowiak-Baldys J Takemoto JY Grilley MM Structure-function studies ofyeast C-4 sphingolipid long chain base hydroxylase Biochim Biophys Acta 2003161817ndash2427 Zhou W Ye XL Sun ZJ Ji XD Chen HX Xie D Overexpression of de-generative spermatocyte homolog 1 up-regulates the expression of cyclin D1 andenhances metastatic efficiency in esophageal carcinoma Eca109 cells MolCarcinog 200948886ndash894
1192 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015

with studies showing that downregulation of degs1 bysmall interfering RNA inhibits cell growth and arreststhe cell cycle in cancer cells (12) and where its overexpres-sion increased cell migration and metastasis (27)
The pharmacological inhibition of DEGS1 in vivovalidated the results obtained in vitro and strengthenedthe concept that functional degs1 is required for the fulldifferentiation of preadipocytes into mature adipocytes
We also provide evidence that both pharmacologicaland genetic ablation of degs1 which according to ourresults in in vitro models may directly block adipocytedifferentiation and lipid deposition certainly increasedDhCer levels in in vitro models In this line our resultsshowed that the treatment of adipocytes with DhCercauses impaired adipogenesis and lipid accumulationThis effect on lipid accumulation was more severewhen the treatment was administered during the first3 days of differentiation suggesting a direct impact ofDhCer accumulation during early stages of adipogenesisWe also provide evidence showing that accumulation ofDhCers can also directly repress the transcriptional ac-tivity of PPARg and hence could contribute to impair-ment of the capacity of the preadipocytes to developa full adipogenic program
These observations strengthen the concept that DhCersare not merely inert precursors of Cers Nevertheless themolecular mechanism linking DhCers accumulation anddysregulation of the cell cycle and other cellular eventsremains elusive One possible mechanism is that changesin the DhCerCer ratio may disrupt membrane-dependentstructures by altering the levels of cholesterol andorcaveolin in lipid rafts which are known to be highlysensitive to a sphingomyelin pool Alternatively changesin the DhCerCer ratio may disturb the global phospho-lipidome potentially altering membrane-associated pro-cesses relevant to adipocyte function These are importantquestions to address in future research
In summary our results indicate that defects in DEGS1in the context of the metabolic syndrome may compro-mise AT expansion and function through the combinedinhibition of adipogenesis promotion of cell death andoxidative stress due to the direct accumulation of DhCersThus our data suggest that the selective manipulation ofDEGS1 andor its substrates in WAT may be a pathophy-siologically advantageous strategy to improve AT homeo-stasis and ameliorate the burden of obesity-associatedmetabolic complications
Acknowledgments The authors thank Dr Carobbio Mrs Peirce andMs Philips (Wellcome Trust-Medical Research Council Institute of MetabolicScience University of Cambridge UK) for their excellent technical advice sup-port and suggestionsFunding This work was funded by the Medical Research Council (MRC) theMetabolic Diseases Unit of the MRC European Commission grant FP7-ETHERPATHSand the British Heart FoundationDuality of Interest No potential conflicts of interest relevant to this articlewere reported
Author Contributions NB helped to develop the hypothesis design theexperiments perform the in vitro and ex vivo experiments collect and analyze thedata hold and characterize the human cohorts collect gene expression data fromadipose depots and analyze the data discuss the manuscript coordinate anddirect the project and write the manuscript SR-C helped to develop thehypothesis design the experiments perform the in vitro and ex vivo experimentscollect and analyze the data discuss the manuscript coordinate and direct theproject and write the manuscript HN and MO helped to develop analyticalplatforms perform and analyze lipidomic experiments and discuss the manu-script AC AP JR IC VP GM-G and CL-P helped to perform the invitro and ex vivo experiments collect and analyze the data and discuss themanuscript FJT helped to hold and characterize the human cohorts collectgene expression data from adipose depots and analyze the data and discussthe manuscript JDS and SAS helped to design the experiments and dis-cuss the manuscript AV-P helped to develop the hypothesis design theexperiments discuss the manuscript coordinate and direct the project andwrite the manuscript AV-P is the guarantor of this work and as such had fullaccess to all the data in the study and takes responsibility for the integrity of thedata and the accuracy of the data analysis
References1 Montague CT OrsquoRahilly S The perils of portliness causes and con-
sequences of visceral adiposity Diabetes 200049883ndash8882 Carobbio S Rodriguez-Cuenca S Vidal-Puig A Origins of metabolic com-
plications in obesity ectopic fat accumulation The importance of the qualitative
aspect of lipotoxicity Curr Opin Clin Nutr Metab Care 201114520ndash5263 Chavez JA Summers SA Lipid oversupply selective insulin resistance
and lipotoxicity molecular mechanisms Biochim Biophys Acta 20101801
252ndash2654 Chavez JA Summers SA A ceramide-centric view of insulin resistance Cell
Metab 201215585ndash5945 Shah C Yang G Lee I Bielawski J Hannun YA Samad F Protection from
high fat diet-induced increase in ceramide in mice lacking plasminogen activator
inhibitor 1 J Biol Chem 200828313538ndash135486 Turner N Kowalski GM Leslie SJ et al Distinct patterns of tissue-specific
lipid accumulation during the induction of insulin resistance in mice by high-fat
feeding Diabetologia 2013561638ndash16487 Blachnio-Zabielska AU Koutsari C Tchkonia T Jensen MD Sphingolipid
content of human adipose tissue relationship to adiponectin and insulin re-
sistance Obesity (Silver Spring) 2012202341ndash23478 Błachnio-Zabielska AU Pułka M Baranowski M et al Ceramide metabo-
lism is affected by obesity and diabetes in human adipose tissue J Cell Physiol
2012227550ndash5579 Ternes P Franke S Zaumlhringer U Sperling P Heinz E Identification and
characterization of a sphingolipid delta 4-desaturase family J Biol Chem 2002
27725512ndash2551810 Mizutani Y Kihara A Igarashi Y Identification of the human sphingolipid C4-
hydroxylase hDES2 and its up-regulation during keratinocyte differentiation
FEBS Lett 200456393ndash9711 Ahn EH Schroeder JJ Induction of apoptosis by sphingosine sphinganine
and C(2)-ceramide in human colon cancer cells but not by C(2)-dihydroceramide
Anticancer Res 2010302881ndash288412 Kraveka JM Li L Szulc ZM et al Involvement of dihydroceramide desaturase
in cell cycle progression in human neuroblastoma cells J Biol Chem 2007282
16718ndash1672813 Siddique MM Li Y Wang L et al Ablation of dihydroceramide desaturase 1
a therapeutic target for the treatment of metabolic diseases simultaneously stim-
ulates anabolic and catabolic signaling Mol Cell Biol 2013332353ndash236914 Parks BW Nam E Org E et al Genetic control of obesity and gut microbiota
composition in response to high-fat high-sucrose diet in mice Cell Metab 2013
17141ndash152
diabetesdiabetesjournalsorg Barbarroja and Associates 1191
15 Bikman BT Guan Y Shui G et al Fenretinide prevents lipid-induced insulinresistance by blocking ceramide biosynthesis J Biol Chem 201228717426ndash1743716 Medina-Gomez G Gray SL Yetukuri L et al PPAR gamma 2 preventslipotoxicity by controlling adipose tissue expandability and peripheral lipid me-tabolism PLoS Genet 20073e6417 Roberts LD Virtue S Vidal-Puig A Nicholls AW Griffin JL Metabolic pheno-typing of a model of adipocyte differentiation Physiol Genomics 200939109ndash11918 Ramiacuterez-Zacariacuteas JL Castro-Muntildeozledo F Kuri-Harcuch W Quantitation ofadipose conversion and triglycerides by staining intracytoplasmic lipids with Oilred O Histochemistry 199297493ndash49719 Ye J Coulouris G Zaretskaya I Cutcutache I Rozen S Madden TL Primer-BLAST a tool to design target-specific primers for polymerase chain reactionBMC Bioinformatics 20121313420 Pfaffl MW Tichopad A Prgomet C Neuvians TP Determination of stablehousekeeping genes differentially regulated target genes and sample integrityBestKeepermdashExcel-based tool using pair-wise correlations Biotechnol Lett 200426509ndash51521 Triola G Fabriagraves G Llebaria A Synthesis of a cyclopropene analogue of cer-amide a potent inhibitor of dihydroceramide desaturase Angew Chem Int Ed Engl2001401960ndash1962
22 Tsuji K Satoh S Mitsutake S et al Evaluation of synthetic sphingolipidanalogs as ligands for peroxisome proliferator-activated receptors Bioorg MedChem Lett 2009191643ndash164623 Mamtani M Meikle PJ Kulkarni H et al Plasma dihydroceramide speciesassociate with waist circumference in Mexican American families Obesity (SilverSpring) 201422950ndash95624 Weir JM Wong G Barlow CK et al Plasma lipid profiling in a largepopulation-based cohort J Lipid Res 2013542898ndash290825 Curran J Meikle P Weir J Jowett J Deep sequencing in extended pedi-grees reveals a major rare non-synonymous variant influencing the de novoceramide synthesis pathway Late-breaking abstract presented at the AmericanSociety of Human Genetics 2013 Meeting 22ndash26 October 2013 at the BostonConvention and Exhibition Center Boston Massachusetts26 Idkowiak-Baldys J Takemoto JY Grilley MM Structure-function studies ofyeast C-4 sphingolipid long chain base hydroxylase Biochim Biophys Acta 2003161817ndash2427 Zhou W Ye XL Sun ZJ Ji XD Chen HX Xie D Overexpression of de-generative spermatocyte homolog 1 up-regulates the expression of cyclin D1 andenhances metastatic efficiency in esophageal carcinoma Eca109 cells MolCarcinog 200948886ndash894
1192 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015

15 Bikman BT Guan Y Shui G et al Fenretinide prevents lipid-induced insulinresistance by blocking ceramide biosynthesis J Biol Chem 201228717426ndash1743716 Medina-Gomez G Gray SL Yetukuri L et al PPAR gamma 2 preventslipotoxicity by controlling adipose tissue expandability and peripheral lipid me-tabolism PLoS Genet 20073e6417 Roberts LD Virtue S Vidal-Puig A Nicholls AW Griffin JL Metabolic pheno-typing of a model of adipocyte differentiation Physiol Genomics 200939109ndash11918 Ramiacuterez-Zacariacuteas JL Castro-Muntildeozledo F Kuri-Harcuch W Quantitation ofadipose conversion and triglycerides by staining intracytoplasmic lipids with Oilred O Histochemistry 199297493ndash49719 Ye J Coulouris G Zaretskaya I Cutcutache I Rozen S Madden TL Primer-BLAST a tool to design target-specific primers for polymerase chain reactionBMC Bioinformatics 20121313420 Pfaffl MW Tichopad A Prgomet C Neuvians TP Determination of stablehousekeeping genes differentially regulated target genes and sample integrityBestKeepermdashExcel-based tool using pair-wise correlations Biotechnol Lett 200426509ndash51521 Triola G Fabriagraves G Llebaria A Synthesis of a cyclopropene analogue of cer-amide a potent inhibitor of dihydroceramide desaturase Angew Chem Int Ed Engl2001401960ndash1962
22 Tsuji K Satoh S Mitsutake S et al Evaluation of synthetic sphingolipidanalogs as ligands for peroxisome proliferator-activated receptors Bioorg MedChem Lett 2009191643ndash164623 Mamtani M Meikle PJ Kulkarni H et al Plasma dihydroceramide speciesassociate with waist circumference in Mexican American families Obesity (SilverSpring) 201422950ndash95624 Weir JM Wong G Barlow CK et al Plasma lipid profiling in a largepopulation-based cohort J Lipid Res 2013542898ndash290825 Curran J Meikle P Weir J Jowett J Deep sequencing in extended pedi-grees reveals a major rare non-synonymous variant influencing the de novoceramide synthesis pathway Late-breaking abstract presented at the AmericanSociety of Human Genetics 2013 Meeting 22ndash26 October 2013 at the BostonConvention and Exhibition Center Boston Massachusetts26 Idkowiak-Baldys J Takemoto JY Grilley MM Structure-function studies ofyeast C-4 sphingolipid long chain base hydroxylase Biochim Biophys Acta 2003161817ndash2427 Zhou W Ye XL Sun ZJ Ji XD Chen HX Xie D Overexpression of de-generative spermatocyte homolog 1 up-regulates the expression of cyclin D1 andenhances metastatic efficiency in esophageal carcinoma Eca109 cells MolCarcinog 200948886ndash894
1192 DEGS1 Downregulation Impairs Adipocyte Function Diabetes Volume 64 April 2015