increased apical sodium-dependent glucose …...research article increased apical sodium-dependent...
TRANSCRIPT

RESEARCH ARTICLE
Increased apical sodium-dependent glucose transporterabundance in the ctenidium of the giant clam Tridacna squamosaupon illuminationChristabel Y. L. Chan1, Kum C. Hiong1, Celine Y. L. Choo1, Mel V. Boo1, Wai P. Wong1, Shit F. Chew2
and Yuen K. Ip1,3,*
ABSTRACTGiant clams contain phototrophic zooxanthellae, and live in nutrient-deficient tropical waters where light is available. We obtained thecomplete cDNA coding sequence of a homolog of mammalian sodium/glucose cotransporter 1 (SGLT1) – SGLT1-like – from the ctenidium ofthe fluted giant clam, Tridacna squamosa. SGLT1-like had a host originand was expressed predominantly in the ctenidium. Molecularcharacterizations reveal that SGLT1-like of T. squamosa couldtransport urea, in addition to glucose, as other SGLT1s do. It has anapical localization in the epithelium of ctenidial filaments and waterchannels, and the apical anti-SGLT1-like immunofluorescence wasstronger in individuals exposed to light than to darkness. Furthermore,the protein abundance of SGLT1-like increased significantly in thectenidium of individuals exposed to light for 12 h, although the SGLT1-like transcript level remained unchanged. As expected, T. squamosacould perform light-enhanced glucose absorption, which was impededby exogenous urea. These results denote the close relationshipsbetween light-enhanced glucose absorption and light-enhancedSGLT1-like expression in the ctenidium of T. squamosa. Althoughglucose absorption could be trivial compared with the donation ofphotosynthates from zooxanthellae in symbiotic adults, SGLT1-likemight be essential for the survival of aposymbiotic larvae, leading toits retention in the symbiotic stage. A priori, glucose uptake throughSGLT1-like might be augmented by the surface microbiome throughnutrient cycling, and the absorbed glucose could partially fulfill themetabolic needs of the ctenidial cells. Additionally, SGLT1-like couldpartake in urea absorption, as T. squamosa is known to conduct light-enhanced urea uptake to benefit the nitrogen-deficient zooxanthellae.
KEYWORDS: Carbohydrate, Calcification, Nitrogen, Symbiodinium,Urea, Zooxanthellae
INTRODUCTIONTropical waters are poor in nutrients owing to a lack of overturn, andtherefore referred to as ‘deserts’ of the sea. To compensate for nutrientshortage in tropical waters, some marine invertebrates, includinggiant clams and scleractinian corals, acquire and maintain symbiotic
relationships with certain dinoflagellates (zooxanthellae) (Trench,1987). Giant clams are common members of coral reefs throughoutthe tropical Indo-Pacific (Neo et al., 2017). They can harbor threegenera of dinoflagellates of the family Symbiodiniaceae –Symbiodinium (formerly Symbiodinium clade A), Cladocopium(formerly Symbiodinium clade C) and Durusdinium (formerlySymbiodinium clade D) (Takabayashi et al., 2004; Hernawan,2008; LaJeunesse et al., 2018) – extracellularly in a branchedtubular system. These symbionts live inside the zooxanthellal tubuleslocated mainly in the extensible and colorful outer mantle (Nortonet al., 1992), where they conduct photosynthesis during insolation.They transfer >95% of photosynthates to the host, which cangenerally satisfy the host’s energy requirements (Fitt, 1993; Griffithsand Klumpp, 1996). With that, giant clams can perform light-enhanced shell formation (calcification) and maintain a high growthrate in nutrient-deficient tropical waters where light is available(Lucas et al., 1989). As the photosynthesizing zooxanthellae require asupply of inorganic carbon, research in the past has focused oninorganic carbon uptake and metabolism in giant clams (Rees et al.,1993b; Baillie and Yellowlees, 1998; Leggat et al., 2002, 2005;Yellowlees et al., 2008). Nonetheless, giant clams can also obtainsome nutrients through filter feeding (Fankboner and Reid, 1986),possible digestion of zooxanthellae in the digestive tract (Reid et al.,1984), and uptake of dissolved organic molecules from the externalmedium (Fitt, 1993).
More than a century ago, Putter (1909) suggested that marineorganisms might be able to obtain nutrients by directly absorbingcertainmolecules from the ambient seawater. Since then, evidence hasbeen gathered to substantiate the absorption of isotopically labelledsugars and amino acids against their concentration gradients throughepidermal tissues of annelids, echinoderms and pogonophores(Stephens and Schinske, 1961; Ferguson, 1967; Ahearn andGomme, 1975; Davis and Stephens, 1984; Manahan, 1989; Pajoret al., 1989). Péquignat (1973) provided categorical evidence tosupport the presence of a similar epidermal route of absorption forglucose and amino acids in the ctenidium (gill) and mantle of thefilibranch bivalve Mytilus edulis. Subsequently, by measuringD-glucose transport in brush-border membrane vesicles derived fromthe ctenidium of M. edulis, the functional presence of a sodium-dependent glucose transporter (SGLT) in its labial palps has beenconfirmed (Pajor et al., 1989). Furthermore, in vitro incubation of thectenidium of the oysterCrassostrea gigas in artificial seawater revealsthat it can absorb D-glucose and D-galactose, but not 3-O-methyl-D-glucose, via an active carrier-mediated system (Bamford and Gingles,1974). In the absence of exogenous Na+, glucose uptake occurs bysimple diffusion, but an Na+-dependent carrier-mediated processsensitive to phlorizin [a potent inhibitor of sodium/glucosecotransporter 1 (SGLT1); Panayotova-Heiermann et al., 1996]Received 1 November 2018; Accepted 5 March 2019
1Department of Biological Sciences, National University of Singapore, Kent Ridge,Singapore 117543, Republic of Singapore. 2Natural Sciences and ScienceEducation, National Institute of Education, Nanyang Technological University,1 Nanyang Walk, Singapore 637616, Republic of Singapore. 3The Tropical MarineScience Institute, National University of Singapore, Kent Ridge, Singapore 119227,Republic of Singapore.
*Author for correspondence ([email protected])
Y.K.I., 0000-0001-9124-7911
1
© 2019. Published by The Company of Biologists Ltd | Journal of Experimental Biology (2019) 222, jeb195644. doi:10.1242/jeb.195644
Journal
ofEx
perim
entalB
iology

inhibition is operating to absorb glucose in the ctenidium and themantle of C. gigas. A kinetic analysis of glucose uptake confirms thepresence of a saturable component at low substrate concentrations, anda diffusive component at high substrate concentrations. Subsequently,a sequence of SGLT-like has been obtained from the oyster C. gigas(Huvet et al., 2004) that has a high mRNA expression level in thectenidium and the mantle edge but low expression in other tissues/organs (Hanquet et al., 2011).SGLTs cotransport Na+ and glucose down the electrochemical
potential gradient of Na+ but up against the concentration gradient ofglucose across cell membranes. Six isoforms of the SGLT genebelonging to the SLC5 gene family have been identified in humans(Wright et al., 2011). All SGLTs have 14 transmembrane regions(TMs) in topology (Wright et al., 2011). In humans, SGLT1 isexpressed mainly in the intestine and kidney, where it functionsas a glucose/galactose transporter. In mouse intestine, SGLT1 isexpressed in the brush-border membranes, and glucose absorptionacross these membranes disappears in SGLT1-deficient mouse,indicating that intestinal glucose absorption is mediatedpredominantly by SGLT1 (Gorboulev et al., 2012).Until now, it was unknownwhether giant clams express a homolog
of SGLT1 (SGLT1-like) in their ctenidia, and whether they canabsorb glucose from the ambient seawater. Nonetheless, giant clamsare known to display light-dependent physiological properties,including light-enhanced shell formation (Sano et al., 2012; Ipet al., 2017a) and light-enhanced nitrogen uptake (Wilkerson andTrench, 1986; Chan et al., 2018). In addition, the gene and/or proteinexpression levels of some of their enzymes and transporters are alsolight dependent (Hiong et al., 2017a,b; Boo et al., 2017, 2018; Ipet al., 2015, 2017a,b, 2018; Koh et al., 2018; Chan et al., 2018; Chewet al., 2019). As such, it is highly probable that giant clams canincrease the expression of SGLT1-like/SGLT1-like in the ctenidiumand the rate of glucose uptake during light exposure.Therefore, the present study was undertaken to clone and sequence
SGLT1-like from the ctenidium of the fluted giant clam, Tridacnasquamosa. Sequence similarity analysis was conducted to verify thatthe SGLT1-like of T. squamosawas derived from the host. Molecularcharacterization of the deduced SGLT1-like amino acid sequencewasperformed to elucidate its roles in the co-transport of Na+ andglucose. Furthermore, the gene expression of SGLT1-like in variousorgans and tissues of T. squamosa was examined. The hypothesistested was that SGLT1-like was expressed predominantly in thectenidium, where glucose uptake could occur. Based on the deducedamino acid sequences, a custom-made anti-SGLT1-like polyclonalantibody was produced commercially, and immunofluorescencemicroscopy was performed to test the proposition that SGLT1-likewas localized in the apical membrane of the epithelial cells of thectenidial filament and water channels, where it could engage in activeglucose absorption. In addition, quantitative real-time PCR (qPCR)and western blotting were performed to examine whether the geneand/or protein expression of SGLT1-like/SGLT1-like in the ctenidiumof T. squamosa could be upregulated by light. To verify thatT. squamosa could indeed perform light-enhanced glucoseabsorption, the rates of glucose absorption in T. squamosa exposedto darkness (control) or to light were determined. Efforts were alsomade to elucidate the possible relationship between glucose and ureaabsorption in T. squamosa, as SGLT1 is known to have the capacityto transport urea (Panayotova-Heiermann and Wright, 2001; Bankirand Yang, 2012). As a crucial part of the coral reef, giant clams aresensitive to adverse changes in their environment, particularly inconnection with global warming and ocean acidification. Resultsobtained from this study may shed light on the mechanisms of
glucose (and urea) uptake in giant clams, which may offer insightsinto ways to enhance their growth and survivorship in this rapidlychanging climate.
MATERIALS AND METHODSAnimalsAdult Tridacna squamosa Lamarck 1819 (mass=500±180 g,N=42)were procured from Xanh Tuoi Tropical Fish (Ho Chi Minh City,Vietnam). The giant clams were maintained in tanks as described byIp et al. (2015) but with slight modifications. The water temperaturewas maintained at 26±1°C, the salinity was 30–32 and thepH ranged between 8.1 and 8.3. The carbonate hardness was143–179 ppm and the calcium concentration was 280–400 ppm.Each tank was illuminated from the top by two sets of Aquazonic T5lighting systems (Yi Hu Fish Farm Trading, Singapore), and eachsystem consisted of two sun and two actinic blue fluorescence tubes(39 Weach). Using a Skye SKP 200 display meter connected with aSKP 215 PAR Quantum sensor (Skye Instruments, UK), the lightintensity at the level of the giant clams was determined as∼100 μmol m−2 s−1. Institutional approval was not necessary forresearch on giant clams (National University of SingaporeInstitutional Animal Care and Use Committee).
Exposure of animals to experimental conditions for tissuecollectionAt the end of the 12 h dark period of the 12 h:12 h light:dark regime,one batch of giant clam (N=5; control) was killed and sampled. Theother 15 individuals were separated into three batches (N=5 forevery time point) and exposed to 3, 6 or 12 h of light beforebeing killed for tissue sampling. They were anesthetized in 0.2%phenoxyethanol, and forced open to cut the adductor muscles. Then,samples of outer mantle, inner mantle, ctenidium, adductor muscle,foot muscle, hepatopancreas and kidney were excised. The tissuesamples were dabbed dry and then freeze-clamped with aluminumtongs in liquid nitrogen. All samples were stored at −80°C prior toprocessing. Tissue samples for immunofluorescence microscopywere harvested separately from another batch of individuals that hadbeen exposed to either darkness or light for 12 h (N=4 for eachcondition) followed with anesthetization in 0.2% phenoxyethanol.
Extraction of mRNA and DNA synthesisExtraction of the total RNA from the ctenidium of T. squamosawas achieved using TRI Reagent® (Sigma-Aldrich, St Louis,MO, USA). Further purification of extracted total RNA wasaccomplished using the RNeasy Plus Mini Kit (Qiagen, Hilden,Germany), and the concentration of total RNAwas verified using aShimadzu BioSpec-nano spectrophotometer (Shimadzu, Tokyo,Japan). Electrophoresis was used to verify total RNA integrity. TheRevertAid™ first-strand cDNA synthesis kit (Thermo FisherScientific, Waltham, MA, USA) was used to reverse transcribethe purified total RNA into cDNA.
Sequence analysis of the isolated geneThe partial SGLT1-like sequence was isolated using primers (given 5′to 3′; forward: GGWTGGGTGTTTGTMCCTGT; reverse: GGWT-GCACSAAYATYGCMTATC; where W, M, S and Y representdegenerate bases) designed from the homologous regions ofCrassostrea gigas sodium glucose cotransporter (AY551098.1),Octopus bimaculoides sodium/glucose cotransporter 4-like (XM_014916561.1), Homo sapiens solute carrier family 5 member 1(NM_000343.3; SGLT1) and Acropora digitifera sodium/glucosecotransporter 1-like (XM_015901071.1). The PCR reaction was
2
RESEARCH ARTICLE Journal of Experimental Biology (2019) 222, jeb195644. doi:10.1242/jeb.195644
Journal
ofEx
perim
entalB
iology

conducted using DreamTaq™ polymerase (Thermo Fisher Scientific)in a 9902 Veriti 96-well thermal cycler (Applied Biosystems,Carlsbad, CA, USA). Following the methods previously described(Hiong et al., 2017a,b) with minor modifications, PCR and cloningexperiments were used to isolate and analyze the gene. For the PCRexperiments, initial denaturation was at 95°C held for 3 min, followedby 40 cycles of denaturation, annealing and extension at 95°C for 30 s,55°C for 30 s and 72°C for 1.5 min. A final extension for 10 min washeld at 72°C. An absence of isoforms was observed from the analysisof multiple clones of SGLT1-like fragments. 5′ and 3′ RACE(SMARTer™ RACE cDNA amplification kit, ClontechLaboratories, Mountain View, CA, USA) PCR was performed withspecific primers (forward: GCCAGGTACAGGTTCAAGTGTA-CAGACT; reverse: ATCCAATGCCAAAGCGGGTACAATTCTG)to amplify and isolate the full coding sequence of SGLT1-like. TheBigDye Terminator v3.1 Cycle Sequencing Kit (Thermo FisherScientific) with ethanol/sodium acetate precipitation was used toprepare the samples for gene sequencing. Sequencing was performedusing a 3130XL Genetic Analyzer (Thermo Fisher Scientific). Thesequence was subsequently analyzed and assembled using BioEditversion 7.2.5. The SGLT1-like sequence has been deposited intoGenBank with accession number MF073182.The SGLT1-like amino acid sequence was translated from the
SGLT1-like nucleotide sequence using the ExPASy Proteomicserver (http://web.expasy.org/translate/). To confirm the identity ofSGLT1-like from T. squamosa, the deduced amino acid sequencewas aligned with selected SGLT and SGLT1-like sequences fromvarious species of mollusk and other animals to generate a sequencesimilarity table (see Table 1 for details). The TMs of the deducedamino acid sequence were predicted using the TMpred server ofthe ExPASy portal (https://embnet.vital-it.ch/software/TMPRED_form.html).
Gene expression in various tissues and organsThe mRNA expression of SGLT1-like in the outer mantle, innermantle, ctenidium, adductor muscle, foot muscle, kidney andhepatopancreas of T. squamosa was investigated qualitatively byPCR using SGLT1-like specific primers (forward: AAATCTAT-CCAATGCCAAAGCGG; reverse: TTCTATCCCACAAATCCA-CTGACC). DreamTaq™ polymerase (Thermo Fisher Scientific)was used for each PCR reaction, with each reaction having a totalvolume of 10 µl. For the thermal cycling, initial denaturation was
accomplished through holding 95°C for 3 min, followed by 35cycles of 95°C (30 s) for denaturation, 55°C (30 s) for annealing and72°C (1 min) for extension, and one final extension at 72°C(10 min). Electrophoresis in 1% agarose gel was used to separate thePCR products.
Determination of mRNA expression by quantitative real-timePCR (qPCR)Absolute quantification through qPCR was performed using aStepOnePlus™Real-Time PCR System (Thermo Fisher Scientific).cDNA (4 µg) was synthesized from RNA using random hexamerprimers and the RevertAid™ first-strand cDNA synthesis kit. ThemRNA expression level of SGLT1-like in the ctenidium wasdetermined through the use of specific qPCR primers (forward:TCTTACACAGCCGATTGACGA; reverse: ATCCGAACACTG-AGATGTCCT). The amplification efficiency for SGLT1-like was86.6%. The absolute quantification of transcripts was calculatedusing the plasmid standard curve expressed as copy number pernanogram total RNA, with methodology following previouspublications (Hiong et al., 2017a,b).
AntibodiesA custom-made rabbit polyclonal anti-SGLT1-like antibody wasdeveloped by a commercial firm (GenScript, Piscataway, NJ, USA)against residues 243–256 (PPDNSMNLIRSYDD) of SGLT1-like ofT. squamosa, and used for immunofluorescence microscopy andimmunoblotting. α-Tubulin was chosen as the reference protein forwestern blotting, and the anti-α-tubulin antibody (12G10)was procuredfrom the Developmental Studies Hybridoma Bank (Department ofBiological Sciences, University of Iowa, Iowa City, IA, USA)
Immunofluorescence microscopyThe immuno-labeling of ctenidial samples of T. squamosa with theanti-SGLT1-like antibody (1.35 µg ml−1) was performed followingthe methods of Hiong et al. (2017b) with Alexa Fluor 488, and goatanti-rabbit (Invitrogen, 2.5 µg ml−1) secondary labeling in green.Nuclei were counterstained using 4′6′-diamidino-2-phenylindole(DAPI) (Sigma-Aldrich; 50 ng ml−1). Slides were mounted inProLong Gold Antifade Mountant (Life Technologies, USA) andcured in the dark at room temperature prior to storing at 4°C forfurther image acquisition. Image acquisition was conducted with afluorescence microscope (Olympus BX60 equipped with a DP73
Table 1. Percentage similarities between the deduced amino acid sequence of sodium/glucose cotransporter 1-like (SGLT1-like) from Tridacnasquamosa and SGLT sequences from other species obtained from GenBank
Classification Species (accession number) Protein Similarity (%)
Mollusca Lottia gigantea (XP_009051606.1) Solute carrier 5,6-like 86.0Crassostrea gigas (AAT44356.1) Sodium/glucose cotransporter 84.3
Brachiopoda Lingula anatina (XP_013421875.1) Sodium/glucose cotransporter 4-like 80.0Priapulus caudatus (XP_014663590.1) Sodium/glucose cotransporter 4-like 78.3
Echinodermata Strongylocentrotus purpuratus (XP_798065.2) Sodium/glucose cotransporter 4 74.7Hemichordata Branchiostoma belcheri (XP_019620396.1) Sodium/glucose cotransporter 4-like 72.2
Branchiostoma floridae (XP_002587561.1) Solute carrier 5,6-like 72.1Chordata Homo sapiens (NP_000334.1) Sodium/glucose cotransporter 1 71.8
Mus musculus (AAF17249.1) Sodium/glucose cotransporter 1 70.8Bos taurus (NP_777031.1) Sodium/glucose cotransporter 1 70.7
Arthropoda Limulus polyphemus (XP_013789754.1) Sodium/glucose cotransporter 4-like 70.5Plantae Vigna angularis (KOM56425.1) Solute carrier 5,6-like 32.7
Vitis vinifera (CAN65174.1) Solute carrier 5,6-like 30.7Fusarium oxysporum (EGU86911.1) Solute carrier 5,6-like 31.0Talaromyces islandicus (CRG86372.1) Solute carrier 5,6-like 30.2
Fungi Paracoccidioides brasiliensis (EEH21459.2) Solute carrier 5,6-like 12.9
Sequences are arranged in order of descending similarity.
3
RESEARCH ARTICLE Journal of Experimental Biology (2019) 222, jeb195644. doi:10.1242/jeb.195644
Journal
ofEx
perim
entalB
iology

CCD digital camera) with the appropriate fluorescent filter sets(Ip et al., 2018). A DIC slider (U-DICT, Olympus) was used toproduce differential interference contrast (DIC) images of thetissue structures such as ctenidial filaments and water channels.Micrographs were collected using CellSens Imaging Software(Olympus) under optimal exposure settings of 200–400 ms. Imageswere further processed with Adobe Photoshop CC (Adobe Systems,San Jose, CA, USA) with adjusted brightness and contrast.
Western blottingThe extraction of protein from ctenidial samples was performedfollowing the methods of Hiong et al. (2017b). Twenty microgramsof ctenidial proteins was subjected to 8% SDS-PAGE and thenblotted onto polyvinylidene difluoride membrane. The membraneswere blocked with Pierce Fast blocking buffer for 30 min at 25°C,followed by incubation with anti-SGLT1-like (1.25 μg ml−1) oranti-α-tubulin (12G10, 0.1 μg ml−1) antibodies for 1 h at 25°C.Subsequently, the membranes were incubated in horseradishperoxidase-conjugated secondary antibodies (Santa CruzBiotechnology; 1:10,000) for 1 h at 25°C. Blots were washedwith TBST (0.05% Tween 20 in Tris-buffered saline: 20 mmol l−1
Tris HCl; 500 mmol l−1 NaCl, pH 7.6) three times and developedwith the ECL system (Thermo Fisher Scientific). Scanning andquantification of the protein bands were performed as described byIp et al. (2017a,b). The quantitation of the SGLT1-like proteinabundance was normalized with that of α-tubulin. Prior toimmunoblotting, immunizing peptide (Genscript) was incubatedwith anti-SGLT1-like antibody for 1 h to further determine thespecificity of the custom-made antibody.
Uptake of glucose by T. squamosaExperiments were designed to demonstrate glucose absorption inT. squamosa by monitoring the reduction in glucose concentrationin the external medium, which represented the sum of glucose influxand efflux. It was not feasible to determine increases in glucoseconcentration in the hemolymph related to glucose absorption, asthe hemolymph has high and fluctuating concentrations of glucose(500–600 mmol l−1) owing to the donation of photosynthates,including glucose, from zooxanthellae (Deane and O’Brien, 1980;Rees et al., 1993a). Uptake of radiolabeled glucose was not adoptedin the present study, because it would reflect at best theunidirectional influx of exogenous glucose without informationon the possible glucose efflux.Sixteen individuals of T. squamosa were randomly selected at the
end of a 12 h:12 h light:dark regime, and transferred separately to clearplastic tanks (21.5×11.5×12.5 cm, length×width×height) in completedarkness. Each plastic tank contained eight volumes (8×mass ofclam) of aerated artificial seawater, and the individual clam wasallowed to acclimatize therein for 2 h in darkness. Then, concentratedglucose solution was added to the artificial seawater to make a finalconcentration of 50 µmol l−1. Although natural seawater has relativelylow glucose concentrations (∼0.23 μmol l−1; Mopper et al., 1980),preliminary results indicated that 50 µmol l−1 of exogenous glucosewas needed for the giant clam to sustain a relatively linear uptake ofglucose for 6 h. Approximately 2 min was required for the glucoseconcentration to become homogeneous; hence, 2 min after theaddition of glucose was taken as time 0.To examine the effects of light on glucose uptake by T. squamosa,
five giant clams were exposed to light at 100–105 µmol m−2 s−1, andanother five giant clams were kept as controls in darkness. In order toexamine whether urea would interfere with glucose absorption, threeindividuals were exposed to light for 6 h in seawater containing
50 µmol l−1 glucose, while another three individuals were exposed tolight for 6 h in seawater containing 50 µmol l−1 glucose+50 µmol l−1
urea. The experiment lasted only 6 h because preliminary resultsobtained indicated that the glucose concentration would decrease by∼50% over a 6-h period. Waters containing glucose or glucose+urea,but without giant clams, were regarded as blanks. Water sampleswere collected at 0, 2, 4 or 6 h, and kept at−20°C. The glucose assay,following the methods of Bergmeyer et al. (1974), was performedwithin 7 days of the uptake experiment. Briefly, the water sampleswere pre-incubated in 1.5 ml of reaction mixture containing250 mmol l−1 triethanolamine (pH 7.5), 2.5 mmol l−1 MgSO4,0.8 mmol l−1 NADP, 10 mmol l−1 ATP and 2.1 i.u. glucose-6-phosphate dehydrogenase. After incubation at 25°C for 5 min, theabsorbance was determined at 340 nm using a Shimadzu UV160UV-VIS spectrophotometer and the reaction was initiated by theaddition of 2.8 i.u. hexokinase. The absorbance was recorded after10 min and the change in absorbance was used for calculation.Freshly prepared glucose solution was used as a standard forcomparison. Percentage changes were used to express the decreasesin the glucose concentration in the external medium, because ofminor variations in the initial glucose concentrations at hour 0. Therate of glucose absorption was expressed as µmol g–1 h–1.
Statistical analysisValues are given as means±s.e.m. Student’s t-test for independentsamples was applied to compare the differences between two means.As for multiple means, the homogeneity of variance among themeans was analyzed using Levene’s test. Subsequently, one-wayANOVA was used to evaluate the difference between the means ofdata sets, followed by Dunnett’s T3 post hoc test or Tukey’s post hoctest, where appropriate. Statistical analysis of data was performedusing SPSS Statistics version 19 (IBM Corporation, Armonk, NY,USA) with the significance level set at P<0.05.
RESULTSSGLT1-like from T. squamosa: analysis of nucleotidesequence and deduced amino acid sequenceThe full cDNA coding sequence of SGLT1-like consisting of 1946 bpwas obtained from the ctenidium ofT. squamosa (GenBank accessionno.: MF073182). The deduced SGLT1-like of T. squamosacomprised 649 amino acids with an estimated molecular mass of72.4 kDa. It had the highest sequence similarity with the SGLTs ofmolluscs (84.3–86.0%; Table 1), followed by those of brachiopods,echinoderms, hemichordates and chordates (70.5–80.0%), but it hadlow sequence similarity with the SGLTs of plants (30.7–34.6%). Acomparison with the SGLT sequence of the green algaAuxenochlorella protothecoides indicated only 34.6% similarity(Table 1). Hence, it can be confirmed that the SGLT1-like obtainedfrom T. squamosa was derived from the host clam and not thezooxanthellae.
An alignment with SGLT1 sequences from other organisms(Crassostrea gigas, Homo sapiens and Vibrio parahaemolyticus)obtained from GenBank revealed that SGLT1-like of T. squamosacontained five glucose-binding residues (D12, E85, D187, K306 andQ440) (Fig. 1). Moreover, it also contained five residues (N61, Y275,W276, S377 and S378) known to be involved in the binding of Na+
and sugar in human SGLT1-like. Particularly, the conserved residuesF436 and Q440 are known to contribute to urea permeability (Fig. 1).The backbone carbonyls of the conserved residues A59 and I62 alsocontributed to the formation of the sugar-binding site, whereasresidues W274, Y275 and W276 (indicated in the open black box inFig. 1) formed an aromatic triad contributing to the transport of sugar
4
RESEARCH ARTICLE Journal of Experimental Biology (2019) 222, jeb195644. doi:10.1242/jeb.195644
Journal
ofEx
perim
entalB
iology

molecules. There were 14 predicted TMs in the SGLT1-like ofT. squamosa. A domain analysis using PROSITE (Sigrist et al., 2002)denoted that SGLT1-like of T. squamosa displayed the profile ofSodium/Solute Symporter 3 (SSS3) family transporters.
Gene expression in various organs and tissuesThe SGLT1-like gene was strongly expressed in the ctenidium butweakly expressed in the outer mantle, foot muscle and kidney ofT. squamosa kept in darkness for 12 h (Fig. 2).
T. squamosa SGLT1-likeC. gigas SGLTH. sapiens hSGLT1V. parahaemolyticus vSGLT
T. squamosa SGLT1-likeC. gigas SGLTH. sapiens hSGLT1V. parahaemolyticus vSGLT
T. squamosa SGLT1-likeC. gigas SGLTH. sapiens hSGLT1V. parahaemolyticus vSGLT
T. squamosa SGLT1-likeC. gigas SGLTH. sapiens hSGLT1V. parahaemolyticus vSGLT
T. squamosa SGLT1-likeC. gigas SGLTH. sapiens hSGLT1V. parahaemolyticus vSGLT
T. squamosa SGLT1-likeC. gigas SGLTH. sapiens hSGLT1V. parahaemolyticus vSGLT
T. squamosa SGLT1-likeC. gigas SGLTH. sapiens hSGLT1V. parahaemolyticus vSGLT
T. squamosa SGLT1-likeC. gigas SGLTH. sapiens hSGLT1V. parahaemolyticus vSGLT
T. squamosa SGLT1-likeC. gigas SGLTH. sapiens hSGLT1V. parahaemolyticus vSGLT
T. squamosa SGLT1-likeC. gigas SGLTH. sapiens hSGLT1V. parahaemolyticus vSGLT
T. squamosa SGLT1-likeC. gigas SGLTH. sapiens hSGLT1V. parahaemolyticus vSGLT
T. squamosa SGLT1-likeC. gigas SGLTH. sapiens hSGLT1V. parahaemolyticus vSGLT
Fig. 1. A multiple sequence alignment of sodium/glucose cotransporter 1-like (SGLT1-like) of Tridacna squamosa with SGLT1 of Crassostrea gigas(AAT44356.1), hSGLT1 of Homo sapiens (NP_000334.1) and vSGLT of Vibrio parahaemolyticus (AAF80602.1). Identical or similar residues areindicated by shading. An asterisk indicates residues involved in binding of sugar molecules. An open circle indicates residues involved in binding of both sugar andNa+. An open triangle indicates residues involved in the structural backbone of the sugar binding site. A shaded circle indicates residues involved in urea binding.The transmembrane regions (TM1 to TM14) are indicated in the underlined sections, and are predicted from TMpred server of the ExPASy portal. The openblack box indicates the aromatic triad involved in the transport of sugar molecules.
5
RESEARCH ARTICLE Journal of Experimental Biology (2019) 222, jeb195644. doi:10.1242/jeb.195644
Journal
ofEx
perim
entalB
iology
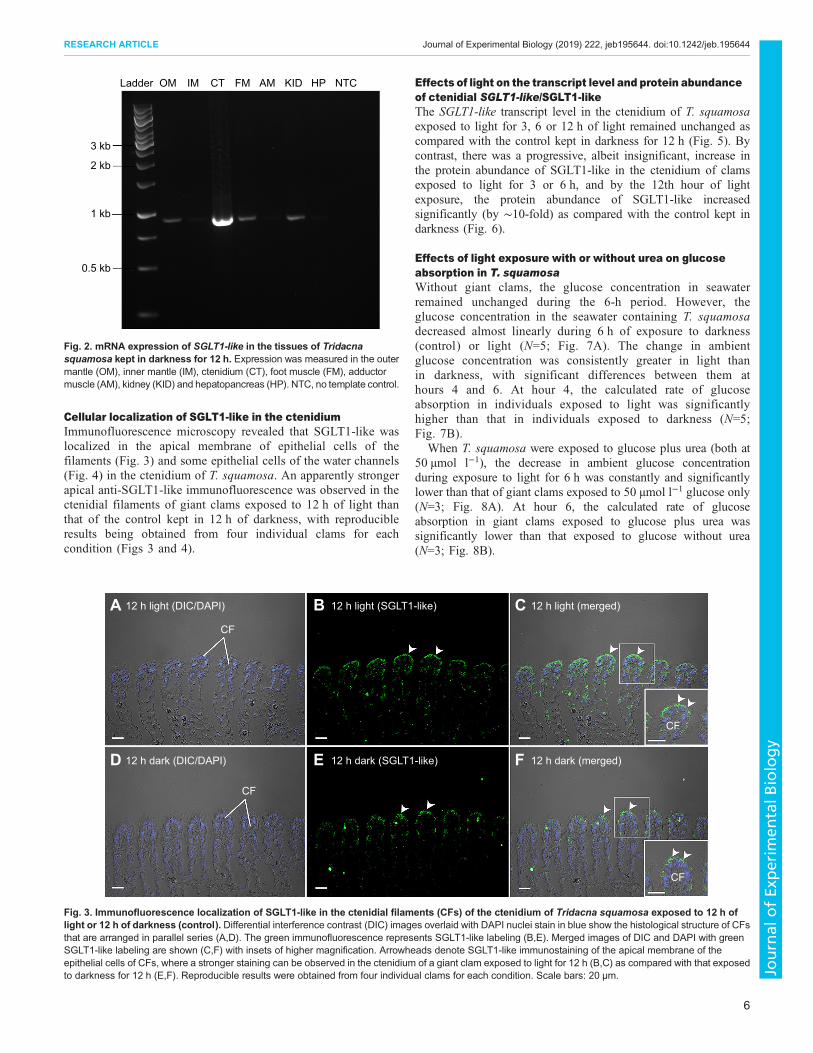
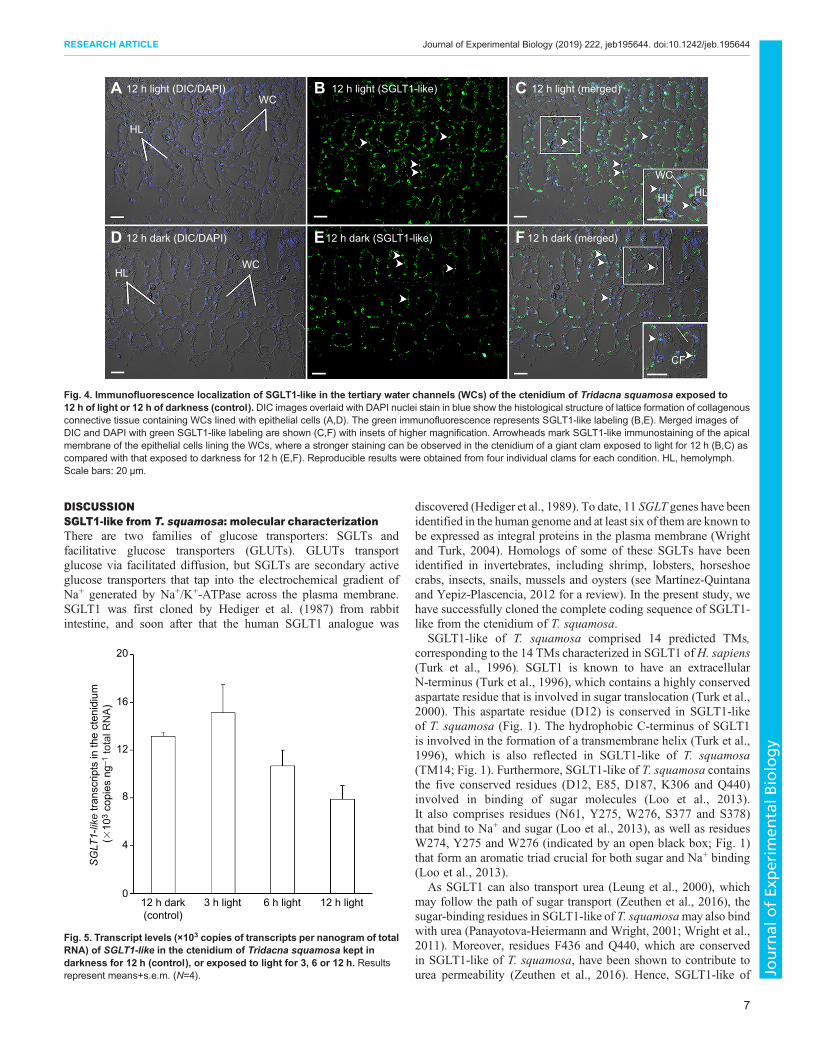
Cellular localization of SGLT1-like in the ctenidiumImmunofluorescence microscopy revealed that SGLT1-like waslocalized in the apical membrane of epithelial cells of thefilaments (Fig. 3) and some epithelial cells of the water channels(Fig. 4) in the ctenidium of T. squamosa. An apparently strongerapical anti-SGLT1-like immunofluorescence was observed in thectenidial filaments of giant clams exposed to 12 h of light thanthat of the control kept in 12 h of darkness, with reproducibleresults being obtained from four individual clams for eachcondition (Figs 3 and 4).
Effects of light on the transcript level and protein abundanceof ctenidial SGLT1-like/SGLT1-likeThe SGLT1-like transcript level in the ctenidium of T. squamosaexposed to light for 3, 6 or 12 h of light remained unchanged ascompared with the control kept in darkness for 12 h (Fig. 5). Bycontrast, there was a progressive, albeit insignificant, increase inthe protein abundance of SGLT1-like in the ctenidium of clamsexposed to light for 3 or 6 h, and by the 12th hour of lightexposure, the protein abundance of SGLT1-like increasedsignificantly (by ∼10-fold) as compared with the control kept indarkness (Fig. 6).
Effects of light exposure with or without urea on glucoseabsorption in T. squamosaWithout giant clams, the glucose concentration in seawaterremained unchanged during the 6-h period. However, theglucose concentration in the seawater containing T. squamosadecreased almost linearly during 6 h of exposure to darkness(control) or light (N=5; Fig. 7A). The change in ambientglucose concentration was consistently greater in light thanin darkness, with significant differences between them athours 4 and 6. At hour 4, the calculated rate of glucoseabsorption in individuals exposed to light was significantlyhigher than that in individuals exposed to darkness (N=5;Fig. 7B).
When T. squamosa were exposed to glucose plus urea (both at50 µmol l−1), the decrease in ambient glucose concentrationduring exposure to light for 6 h was constantly and significantlylower than that of giant clams exposed to 50 µmol l−1 glucose only(N=3; Fig. 8A). At hour 6, the calculated rate of glucoseabsorption in giant clams exposed to glucose plus urea wassignificantly lower than that exposed to glucose without urea(N=3; Fig. 8B).
Ladder
3 kb
2 kb
1 kb
0.5 kb
OM IM CT FM AM KID HP NTC
Fig. 2. mRNA expression of SGLT1-like in the tissues of Tridacnasquamosa kept in darkness for 12 h. Expression was measured in the outermantle (OM), inner mantle (IM), ctenidium (CT), foot muscle (FM), adductormuscle (AM), kidney (KID) and hepatopancreas (HP). NTC, no template control.
12 h light (DIC/DAPI)A B C
D E F
CF
CF
CF
CF
12 h dark (DIC/DAPI) 12 h dark (SGLT1-like) 12 h dark (merged)
12 h light (SGLT1-like) 12 h light (merged)
Fig. 3. Immunofluorescence localization of SGLT1-like in the ctenidial filaments (CFs) of the ctenidium of Tridacna squamosa exposed to 12 h oflight or 12 h of darkness (control). Differential interference contrast (DIC) images overlaid with DAPI nuclei stain in blue show the histological structure of CFsthat are arranged in parallel series (A,D). The green immunofluorescence represents SGLT1-like labeling (B,E). Merged images of DIC and DAPI with greenSGLT1-like labeling are shown (C,F) with insets of higher magnification. Arrowheads denote SGLT1-like immunostaining of the apical membrane of theepithelial cells of CFs, where a stronger staining can be observed in the ctenidium of a giant clam exposed to light for 12 h (B,C) as compared with that exposedto darkness for 12 h (E,F). Reproducible results were obtained from four individual clams for each condition. Scale bars: 20 µm.
6
RESEARCH ARTICLE Journal of Experimental Biology (2019) 222, jeb195644. doi:10.1242/jeb.195644
Journal
ofEx
perim
entalB
iology
DISCUSSIONSGLT1-like from T. squamosa: molecular characterizationThere are two families of glucose transporters: SGLTs andfacilitative glucose transporters (GLUTs). GLUTs transportglucose via facilitated diffusion, but SGLTs are secondary activeglucose transporters that tap into the electrochemical gradient ofNa+ generated by Na+/K+-ATPase across the plasma membrane.SGLT1 was first cloned by Hediger et al. (1987) from rabbitintestine, and soon after that the human SGLT1 analogue was
discovered (Hediger et al., 1989). To date, 11 SGLT genes have beenidentified in the human genome and at least six of them are known tobe expressed as integral proteins in the plasma membrane (Wrightand Turk, 2004). Homologs of some of these SGLTs have beenidentified in invertebrates, including shrimp, lobsters, horseshoecrabs, insects, snails, mussels and oysters (see Martínez-Quintanaand Yepiz-Plascencia, 2012 for a review). In the present study, wehave successfully cloned the complete coding sequence of SGLT1-like from the ctenidium of T. squamosa.
SGLT1-like of T. squamosa comprised 14 predicted TMs,corresponding to the 14 TMs characterized in SGLT1 ofH. sapiens(Turk et al., 1996). SGLT1 is known to have an extracellularN-terminus (Turk et al., 1996), which contains a highly conservedaspartate residue that is involved in sugar translocation (Turk et al.,2000). This aspartate residue (D12) is conserved in SGLT1-likeof T. squamosa (Fig. 1). The hydrophobic C-terminus of SGLT1is involved in the formation of a transmembrane helix (Turk et al.,1996), which is also reflected in SGLT1-like of T. squamosa(TM14; Fig. 1). Furthermore, SGLT1-like of T. squamosa containsthe five conserved residues (D12, E85, D187, K306 and Q440)involved in binding of sugar molecules (Loo et al., 2013).It also comprises residues (N61, Y275, W276, S377 and S378)that bind to Na+ and sugar (Loo et al., 2013), as well as residuesW274, Y275 and W276 (indicated by an open black box; Fig. 1)that form an aromatic triad crucial for both sugar and Na+ binding(Loo et al., 2013).
As SGLT1 can also transport urea (Leung et al., 2000), whichmay follow the path of sugar transport (Zeuthen et al., 2016), thesugar-binding residues in SGLT1-like of T. squamosamay also bindwith urea (Panayotova-Heiermann and Wright, 2001; Wright et al.,2011). Moreover, residues F436 and Q440, which are conservedin SGLT1-like of T. squamosa, have been shown to contribute tourea permeability (Zeuthen et al., 2016). Hence, SGLT1-like of
12 h light (DIC/DAPI)A B C
D E F
WC
WC
HL
HLWC
CF
HLHL
12 h dark (DIC/DAPI) 12 h dark (SGLT1-like) 12 h dark (merged)
12 h light (SGLT1-like) 12 h light (merged)
Fig. 4. Immunofluorescence localization of SGLT1-like in the tertiary water channels (WCs) of the ctenidium of Tridacna squamosa exposed to12 h of light or 12 h of darkness (control). DIC images overlaid with DAPI nuclei stain in blue show the histological structure of lattice formation of collagenousconnective tissue containing WCs lined with epithelial cells (A,D). The green immunofluorescence represents SGLT1-like labeling (B,E). Merged images ofDIC and DAPI with green SGLT1-like labeling are shown (C,F) with insets of higher magnification. Arrowheads mark SGLT1-like immunostaining of the apicalmembrane of the epithelial cells lining the WCs, where a stronger staining can be observed in the ctenidium of a giant clam exposed to light for 12 h (B,C) ascompared with that exposed to darkness for 12 h (E,F). Reproducible results were obtained from four individual clams for each condition. HL, hemolymph.Scale bars: 20 µm.
12 h dark(control)
3 h light0
4
SG
LT1-
like
trans
crip
ts in
the
cten
idiu
m
(�10
3 co
pies
ng–
1 to
tal R
NA
)
8
12
16
20
6 h light 12 h light
Fig. 5. Transcript levels (×103 copies of transcripts per nanogram of totalRNA) of SGLT1-like in the ctenidium of Tridacna squamosa kept indarkness for 12 h (control), or exposed to light for 3, 6 or 12 h. Resultsrepresent means+s.e.m. (N=4).
7
RESEARCH ARTICLE Journal of Experimental Biology (2019) 222, jeb195644. doi:10.1242/jeb.195644
Journal
ofEx
perim
entalB
iology

T. squamosa can probably transport not only glucose, but alsourea, with Na+ being used as a motive force to drive theiractive uptake.
SGLT1-like is expressed predominantly in the ctenidium ofT. squamosa, and has an apical localization in the ctenidialepithelial cellsMany mollusks, including bivalves, cephalopods and aquaticgastropods, possess a pair of ctenidia inside their mantle cavities.A ctenidium is primarily a respiratory organ in water, but can alsoparticipate in filter feeding, ionoregulation and acid–base balance.In T. squamosa, the ctenidia arewhitish in color, and each ctenidiumconsists of two demibranches (dorsal and ventral). The ctenidium iscomb-shaped, and has a central part from which many filamentsprotrude and line up in a row to increase the surface area for variousphysiological functions (Norton and Jones, 1992). The surface areais further increased by numerous water channels found below thefilaments inside the ctenidium.The ctenidium of T. squamosa is known to express transporters
and enzymes (Ip et al., 2015) related to nitrogen transport andassimilation (DUR3-like, Chan et al., 2018; AmmoniaTransporter 1, Boo et al., 2018; Glutamine Synthetase, Hionget al., 2017a), inorganic carbon absorption (Dual DomainCarbonic Anhydrase; Koh et al., 2018) and proton excretion(Na+/H+ Exchanger 3-like, Hiong et al., 2017b; Vacuolar-typeH+-ATPase subunit A, Ip et al., 2018). In the present study, wedemonstrated that SGLT1-like was expressed predominantly inthe ctenidium of T. squamosa. Importantly, SGLT1-like had anapical localization in epithelial cells covering the ctenidialfilament and lining the tertiary water channels. These resultsdenote that SGLT1-like is positioned to transport Na+ togetherwith glucose (or urea) from the ambient seawater into thectenidial epithelial cells, and suggest the ctenidium as the site ofactive glucose uptake.
Light-enhanced expression of SGLT1-like in the ctenidiumindicates that T. squamosa can conduct light-enhancedglucose absorptionIt has been reported that the protein expression level of SGLT1 inthe intestinal brush border of mammals can be upregulated byglucose (Wood et al., 2000), and protein kinases (PKA and PKC)regulate the transport of SGLT1 by rapidly inserting it into theplasma membrane (Turk and Wright, 1997). Similarly, SGLT1-likeof oyster (C. gigas) is regulated according to trophic conditions,including food abundance and quality (Hanquet et al., 2011).Nevertheless, this is the first report on the upregulation of the proteinabundance of SGLT1-like in the ctenidium of T. squamosa inresponse to light. Our results indicate that SGLT1-like was regulatedprincipally through translation, as light exposure did not have asignificant effect on its transcript level. This differs from DualDomain Carbonic Anhydrase (Koh et al., 2018), GlutamineSynthetase (Hiong et al., 2017a) and Na+/H+ Exchanger 3-like(Hiong et al., 2017b), of which both the transcript level and theprotein abundance increase significantly in the ctenidium ofT. squamosa during 12 h of light exposure. Overall, these resultscorroborate the proposition that T. squamosa could perform light-enhanced glucose/urea absorption.
Light-enhanced glucose uptake in T. squamosaThe hemolymph of giant clams is known to have very highconcentrations of glucose (500–600 mmol l−1; Deane and O’Brien,1980; Rees et al., 1993a) owing to the donation of photosynthatesfrom zooxanthellae. Yet, T. squamosa could reduce the ambientglucose concentration (50 mmol l−1) by 45% during 6 h of lightexposure despite such a huge concentration gradient. Hence, it canbe concluded that T. squamosa could actively absorb glucose formthe environment. Notably, the observation on the rate of glucoseuptake being higher in light than in darkness is novel. Althoughglucose uptake is known to occur in non-symbiotic mussels
120 kDa
12 hdark
(control)
3 hlight
6 hlight
12 hlight
100 kDa
80 kDa
60 kDa
50 kDa
40 kDa
120 kDa
LadderA
0
Rel
ativ
e no
rmal
ized
SG
LT1-
like
prot
ein
abun
danc
e in
the
cten
idiu
m (a
rbitr
ary
units
)
12 h dark(control)
3 h light 6 h light 12 h light
a
a,b a,b
b
0.2
0.1
0.3
0.4
0.5
B12 hlight
(PCA)Ladder
100 kDa
80 kDa
60 kDa No band
Tubulin
50 kDa
40 kDa
50 kDa
SGLT1-like
Fig. 6. Protein abundance of SGLT1-like in the ctenidium of Tridacna squamosa kept in darkness for 12 h (control), or exposed to light for 3,6 or 12 h. (A) Examples of immunoblot of SGLT1-like, with or without the anti-SGLT1-like antibody neutralized by the immunizing peptide, and with tubulinas the reference protein. (B) The intensity of the SGLT1-like band for 20 µg protein was normalized with respect to that of tubulin. Results represent means+s.e.m.(N=4). Means not sharing the same letter are significantly different (P<0.05).
8
RESEARCH ARTICLE Journal of Experimental Biology (2019) 222, jeb195644. doi:10.1242/jeb.195644
Journal
ofEx
perim
entalB
iology

(Péquignat, 1973) and oysters (Bamford and Gingles, 1974), there isno indication of it being a light-dependent process. Scleractiniancorals (e.g. Fungia) can also absorb glucose from the externalmedium (Stephens, 1960, 1962), but the possibility of it beingenhanced by light has not been explored.Natural seawater contains dissolved organic molecules that
comprise a wide variety of carbohydrates, lipids, amino acids, aswell as vitamins and hormones (Duursma, 1965). Specifically,glucose is the major monosaccharide in seawater (Ittekkot et al.,1981), derived largely from oligosaccharide hydrolysis.Phytoplankton produces and stores glucan oligosaccharides (Lewin,1974), which can be released to the ambient seawater throughexudation or cell lysis. The degradation of these glucanoligosaccharides in seawater produces monosaccharides includingglucose. Generally, the concentration of glucose in open seawaterranges between 10−6 and 10−8 mol l−1 (Vaccaro et al., 1968).Although the glucose concentration can be higher in waters aroundcoral reefs owing to both algal and coral exudations that containsaccharides, proteins and lipids (Haas and Wild, 2010; Nelson et al.,2013), it is unlikely to reach the concentration of 0.05 mmol l−1
adopted in this study. Therefore, it is possible that the rate of glucose
uptake obtained for T. squamosa is non-physiological, althoughits ability to absorb glucose is affirmed. However, in effect, theabsorption rate of glucose should be defined by the concentrations ofglucose in the unstirredmucus layer covering the ctenidial epithelium,and not the glucose concentrations in the ambient seawater.
In scleractinian corals, the oral ectoderm, consisting ofmultiple specialized cells such as cnidocytes, mucocytes andepitheliomuscular cells (Fautin and Mariscal, 1991), is involved inthe transport ofCa2+ into calicoblastic cells associatedwith calcification(Tambutté et al., 2007).During insolation, theCa2+ concentration (14.5±0.7 mmol l−1) in the 10–20 μm thick mucus layer coating the oralectoderm is consistently higher than that (11.3±0.3 mmol l−1) in theambient seawater (Clode andMarshall, 2002).Hence, the existence of amucus boundary between the seawater and oral ectoderm couldfacilitate the efficient absorption of Ca2+ into the oral ectodermal cells.Similarly, type 2 and type 3 mucin gland cells have been identified inthe ctenidium of T. squamosa, and these cells can secrete mucus ontosurfaces of the epithelial cells lining the filaments and water channels(Norton and Jones, 1992). If themicrobiomeof the ctenidial surface canperform oligosaccharide degradation, the glucose concentration in themucus layer could be higher than that in the seawater, but theconfirmation of this proposition awaits future study.
400 2 4
Time (h)6
0
1
2
3
4
2 4Time (h)
6
50
60
70
Cha
nge
in g
luco
se c
once
ntra
tion
in th
e ex
tern
al m
ediu
m (%
)R
ate
of g
luco
se u
ptak
e(�
10–2
µm
ol g
–1 h
–1)
80
90
100
A
B
*
*
*
Fig. 7. Effects of light on the absorption of glucose in Tridacnasquamosa. (A) The change in glucose concentration (%) in the externalmedium, containing glucose at an initial concentration of 0.05 mmol l−1,without clams (▵), or with clams exposed to light (○) or darkness (●; control)for 2, 4 or 6 h. (B) The rate of glucose absorption (×10−2 µmol g−1 h−1) byT. squamosa at hour 2, 4 or 6 of exposure to darkness (closed bars; control)or light (open bars). Results represent means±s.e.m. (N=5). *Significantlydifferent from clams kept in darkness at the specific time point (P<0.05).
100A
B
**
*
*
80
60
40
Cha
nge
in g
luco
se c
once
ntra
tion
in th
e ex
tern
al m
ediu
m (%
)R
ate
of g
luco
se u
ptak
e(�
10–2
µm
ol g
–1 h
–1)
20
00
4
3
2
1
0
2 4Time (h)
6
2 4Time (h)
6
Fig. 8. Effects of urea on the absorption of glucose in Tridacna squamosain light. (A) The change in glucose concentration (%) in the external medium,containing glucose at an initial concentration of 0.05 mmol l−1, without clams(▵), with clams (○), or with clams and 0.05 mmol l−1 urea (●) for 2, 4 or 6 h.(B) The rate of glucose absorption (×10−2 µmol g−1 h−1) by T. squamosa athour 2, 4 or 6 in seawater containing 0.05 mmol l−1 glucose (open bars) or0.05 mmol l−1 glucose+0.05 mmol l−1 urea (closed bars). Results representmeans+s.e.m. (N=3). *Significantly different from clams exposed to0.05 mmol l−1 glucose at the specific time point (P<0.05).
9
RESEARCH ARTICLE Journal of Experimental Biology (2019) 222, jeb195644. doi:10.1242/jeb.195644
Journal
ofEx
perim
entalB
iology

Possible relationships between nutrient cycling by thesurface microbiome of the ctenidium and glucose uptakeThe assimilation of dissolved organic matter by sponge holobiontsis known to facilitate the cycling of dissolved organic matters inbenthic habitats (de Goeij et al., 2013). Microbes contribute to thisassimilation process (Rix et al., 2016), accounting for ∼90% of theholobiont’s total heterotrophic carbon uptake (Morganti et al.,2017; Hoer et al., 2018). In corals, the associated microbialcommunities are also important components of the coralholobionts (Leggat et al., 2007; Rosenberg et al., 2007), as theyparticipate in nutrient cycling and influence coral health anddiseases (Rosenberg and Loya, 2004; Rosenberg et al., 2007).Kimes et al. (2010) analyzed the microbiome associated withthe scleractinian coral Montastraea faveolata and detected134 ribulose-1,5-bisphosphate carboxylase/oxygenase genes(RuBisCO) of archaea and bacteria origins, indicating that thesemicrobes could fix CO2 and produce organic compounds throughphotosynthesis (Berg et al., 2002). Furthermore, Kimes et al.(2010) detected 825 microbial genes involved in polysaccharidedegradation, including bacterial, archaeal and fungal cellulases(359 sequences), chitinases (206 sequences), mannanases (55sequences) and polygalactases (63 sequences). Degradation ofpolysaccharide by these microbes could release glucose and othermonosaccharides close to the surface of the host. In fact, theability of microbes to metabolize nutrients, which can then betranslocated to their host, is likely a driver in the establishment ofcoral-associated microbial assemblages. Assuming that the surfacemicrobiome of giant clams can do the same, the concentration ofglucose in the mucus layer covering the ctenidial epithelium couldbe higher than that in the ambient seawater, rendering an effectiveuptake of glucose though SGLT1-like.However, the contribution of light-enhanced glucose
absorption through ctenidial SGLT1-like to the overallmetabolic needs of the host clam could be inconsequentialwhen compared with the donation of carbohydrates, includingglucose, from the photosynthesizing zooxanthellae (Rees et al.,1993a; Ishikura et al., 1999). In fact, it has been establishedthat the photosynthates transferred from the symbionts to thehost clam can satisfy ∼100% of the host’s energy requirements(Fisher et al., 1985; Klumpp et al., 1992). Therefore, thephysiological significance of active glucose absorption insymbiotic T. squamosa is obscure. Nonetheless, there couldbe three physiological reasons, related separately to themetabolic needs of the aposymbiotic larva, the ctenidium andthe zooxanthellae.
Exogenous glucose uptakemay be essential for the survivalof aposymbiotic larvaeGiant clam larvae are aposymbiotic, and they acquire symbioticzooxanthellae only during the veliger stage through filter feeding(Fitt and Trench, 1981; Heslinga et al., 1984; Mies and Sumida,2012). It has been reported that veliger larvae of oyster (C. gigas)and red abalone (Haliotis rufescens) absorb glucose and someother complex sugars from the external medium, which allowsthem to utilize a greater part of the dissolved organic material inthe sea as a source of nutrition (Welborn and Manahan, 1990).Therefore, it is probable that the absorption of exogenous glucosethrough SGLT1-like in the rudimentary ctenidium contributes tothe successful survival of giant clam larvae, which needs to beconfirmed in future studies. Nevertheless, this could lead to theretention of SGLT1-like expression in the ctenidium of symbioticT. squamosa.
Exogenous glucose uptake may support the metabolicneeds of the ctenidiumThe ctenidium of T. squamosa is metabolically active owing to itsimportant roles in, for example, inorganic carbon absorption (Kohet al., 2018), excretion of excess H+ (Hiong et al., 2017b; Ip et al.,2018), and nitrogen transport and assimilation (Chan et al., 2018;Boo et al., 2018; Hiong et al., 2017a). It contains few zooxanthellae,and is located inside the mantle cavity with no direct exposure tosunlight. Although the ctenidium can undoubtedly receive somephotosynthates from the outer mantle through the hemolymph,these two organs are separated far apart. Importantly, other tissuessuch as the inner mantle, which adjoins the outer mantle andconducts light-enhanced shell formation, would also compete fornutrients, especially during insolation. Therefore, the expression ofSGLT1-like in the ctenidium could be advantageous to symbioticT. squamosa, as the absorbed glucose can specifically fuel themetabolic needs of this organ. It is possible that the problemconcerning low ambient glucose concentrations could be remediedby the activities of its surface microbiome.
Ctenidial SGLT1-like may also participate in urea absorptionin symbiotic T. squamosa to benefit the nitrogen-deficientzooxanthellaeIt has been established that SGLT1 can transport urea in addition toglucose (Leung et al., 2000; Panayotova-Heiermann and Wright,2001), with the coupling of urea transport to its cotransport cycle(Zeuthen et al., 2001). This could explain why urea interfered withlight-enhanced glucose absorption in T. squamosa, and the rate ofglucose uptake in individuals exposed to glucose+urea waslower than that in individuals exposed to glucose only. Althoughthe host clam benefits from photosynthates donated by symbioticzooxanthellae (Streamer et al., 1993), the symbionts require asupply of nutrients from the host (Furla et al., 2005). Specifically,the host clam would need to absorb exogenous nitrogen andsupply it to the symbiotic zooxanthellae, which are nitrogendeficient. Dissolved inorganic nitrogen is present in seawateras ammonium, nitrite and nitrate, whereas dissolved organicnitrogen is represented mainly by urea and amino acids. In reefenvironments, urea concentrations range between 0.2 and2.0 µmol N l−1 (Wafar et al., 1986; Beauregard, 2004), but theurea concentration in the mucus covering the ctenidial epitheliumof T. squamosa could be higher, attributable to the activities of itsmicrobiome. Indeed, nitrogen cycling is often cited as a probablerole filled by the microbial community of coral reefs (Rosenberget al., 2007), and bacteria are known to produce and excrete urea(Pedersen et al., 1993).
Recently, it has been demonstrated that T. squamosa can performlight-enhanced urea absorption, and its ctenidium expresses a urea-active (energy-dependent) transporter, DUR3-like, of animal origin(Chan et al., 2018). Urea can be a good nitrogen source for thesymbiotic zooxanthellae because it contains two nitrogen atoms andone carbon atom (H2NCONH2), and zooxanthellae possess urease,which can catabolize urea. The degradation of urea by urease inzooxanthellae releases NH3 to support amino acid metabolism andCO2 to sustain photosynthesis. Therefore, it would be beneficial forT. squamosa to possess multiple mechanisms in the ctenidium,including DUR3-like and SGLT1-like, to absorb urea from theambient seawater. Notably, the competition between glucose andurea for transport by SGLT1-like would be defined by thetransporter’s kinetic properties towards these two substrates, aswell as the effective concentrations of these two substrates in thevicinity of the transporter.
10
RESEARCH ARTICLE Journal of Experimental Biology (2019) 222, jeb195644. doi:10.1242/jeb.195644
Journal
ofEx
perim
entalB
iology

ConclusionsTridacna squamosa expresses SGLT1-like in its ctenidia, and theprotein abundance of SGLT1-like is upregulated by light exposure. Italso increases the absorption of glucose from the ambient seawater inresponse to light. When taken together with information in theliterature, it can be concluded that the ctenidium of T. squamosa is notsimply a respiratory organ, as it also participates in various absorptiveand excretory processes. Many of these processes are light dependentand they involve enzymes and transporters that can respond to lightthrough transcriptional and/or translational changes. Such phenomenaprobably stem from the symbiotic association of the clam host withphototrophic zooxanthellae, as the light-dependent properties of theseenzymes and transporters would allow the host to react to light insynchrony with the photosynthetic activity of its symbionts.
Competing interestsThe authors declare no competing or financial interests.
Author contributionsConceptualization: Y.K.I.; Methodology: Y.K.I.; Validation: C.Y.L. Chan, K.C.H.,C.Y.L. Choo, M.V.B.; Formal analysis: C.Y.L. Chan, K.C.H., C.Y.L. Choo, M.V.B.,S.F.C., Y.K.I.; Investigation: C.Y.L. Chan, K.C.H., C.Y.L. Choo, M.V.B.; Resources:W.P.W., S.F.C.; Data curation: C.Y.L. Chan, K.C.H., C.Y.L. Choo, M.V.B., W.P.W.,Y.K.I.; Writing - original draft: C.Y.L. Chan, Y.K.I.; Writing - review & editing:C.Y.L. Chan, K.C.H., C.Y.L. Choo, M.V.B., S.F.C., Y.K.I.; Visualization: C.Y. Chan,K.C.H., C.Y.L. Choo, M.V.B., S.F.C.; Supervision: W.P.W., S.F.C., Y.K.I.; Projectadministration: Y.K.I.; Funding acquisition: YK.I.
FundingThis study was supported by the Ministry of Education - Singapore through a grant(R-154-000-A37-114) to Y.K.I.
ReferencesAhearn, G. A. and Gomme, J. (1975). Transport of exogenous D-glucose by theintegument of a polychaete worm (Nereis diversicolor Muller). J. Exp. Biol. 62,243-264.
Baillie, B. K. and Yellowlees, D. (1998). Characterization and function of carbonicanhydrase in the zooxanthellae–giant clam symbiosis.Proc. Biol. Sci. 265, 465-473.
Bamford, D. R. and Gingles, R. (1974). Absorption of sugar in the gill of theJapanese oyster, Crassostrea gigas. Comp. Biochem. Physiol. A. 49, 637-646.
Bankir, L. and Yang, B. (2012). New insights into urea and glucose handling by thekidney, and the urine concentrating mechanism. Kidney Int. 81, 1179-1198.
Beauregard, A. Y. (2004). Biogeochemical cycling of carbon and nitrogen by thecoral–zooxanthellae symbiosis. PhD thesis, University of Delaware.
Berg, J. M., Tymoczko, J. L. and Stryer, L. (2002). Biochemistry. New York: W.H.Freeman and Company.
Bergmeyer, H. U., Bernt, E., Schmidt, F. and Stork, H. (1974). D-Glucose. InMethods of Enzymatic Analysis, Vol. 3 (ed. H. U. Bergmeyer and K. Gawehn), pp.1196-1201. Weinheim: Verlag Chemie.
Boo, M. V., Hiong, K. C., Choo, C. Y. L., Cao-Pham, A. H., Wong,W. P., Chew, S. F.and Ip, Y. K. (2017). The inner mantle of the giant clam, Tridacna squamosa,expresses a basolateral Na+/K+-ATPase α-subunit, which displays light-dependentgene and protein expression along the shell-facing epithelium. PLoS ONE 12,e0186865.
Boo, M. V., Hiong, K. C., Goh, E. J. K., Choo, C. Y. L., Wong, W. P., Chew, S. F.and Ip, Y. K. (2018). The ctenidium of the giant clam, Tridacna squamosa,expresses an ammonium transporter 1 that displays light-suppressed gene andprotein expression and may be involved in ammonia excretion. J. Comp. Physiol.B 188, 765-777.
Chan,C. Y. L., Hiong,K.C., Boo,M.V., Choo, C.Y. L.,Wong,W. P., Chew, S. F. andIp, Y. K. (2018). Light exposure enhances urea absorption in the fluted giant clam,Tridacna squamosa, and up-regulates the protein abundance of a light-dependenturea active transporter, DUR3-like, in its ctenidium. J. Exp. Biol. 221, jeb176313.
Chew, S. F., Koh, C. Z. Y., Hiong,K. C., Choo, C. Y. L.,Wong,W. P., Neo,M. L. andIp, Y. K. (2019). Light-enhanced expression of Carbonic Anhydrase 4-like supportsshell formation in the fluted giant clam Tridacna squamosa. Gene 683, 101-112.
Clode, P. L. and Marshall, A. T. (2002). X-ray microanalysis of calcium in ascleractinian coral. J. Exp. Biol. 205, 3543-3553.
Davis, J. P. and Stephens, G. C. (1984). Uptake of free amino acids by bacteria-free larvae of the sand dollar Dendraster excentricus. Am J. Physiol. 247,R733-R739.
Deane, E. M. and O’Brien, R. W. (1980). Composition of the hemolymph ofTridacna maxima (Mollusca: Bivalvia). Comp. Biochem. Physiol. 66A, 339-341.
de Goeij, J. M., van Oevelen, D., Vermeij, M. J. A., Osinga, R., Middelburg, J. J.,de Goeij, A. F. P. M. and Admiraal, W. (2013). Surviving in a marine desert: thesponge loop retains resources within coral reefs. Science 342, 108-110.
Duursma, E. K. (1965). The dissolved organic constituents of seawater. InChemical Oceanography, Vol. 1 (ed. J. P. Riley and G. Skirrow), pp. 433-475.New York: Academic Press.
Fankboner, P. V. and Reid, R. G. B. (1986). Nutrition in giant clams (Tridacnidae).In The Bivalvia-Proceedings of a Memorial Symposium in Honour of Sir CharlesMaurice Yonge, Edinburgh, 1986 (ed. B. Morton), pp. 195-209. Hong Kong: HongKong University Press.
Fautin, D. G. and Mariscal, R. N. (1991). Cnidaria: Anthozoa. In MicroscopicAnatomy of Invertebrates, Vol. 2 (ed. F. W. Harrison and J. A. Westfall), pp.267-358. New York: Wiley-Liss.
Ferguson, J. C. (1967). An autoradiographic study of the utilization of freeexogenous amino acids by starfishes. Biol. Bull. 33, 317-329.
Fisher, C. R., Fitt, W. K. and Trench, R. K. (1985). Photosynthesis and respirationin Tridacna gigas as a function of irradiance and size. Biol. Bull. 169, 230-245.
Fitt, W. K. (1993). Nutrition of giant clams. In Biology and Mariculture of GiantClams, ACIARProceedings no. 47 (ed.W. K. Fitt), pp. 31-40. Canberra: AustralianCentre for International Agricultural Research.
Fitt, W. K. and Trench, R. K. (1981). Spawning, development, and acquisition ofzooxanthellae byTridacna squamosa (Mollusca: Bivalvia).Biol. Bull. 161, 213-235.
Furla, P., Allemand, D., Shick, J. M., Ferrier-Pages, C., Richier, S., Plantivaux,A., Merle, P. L. and Tambutte, S. (2005). The symbiotic anthozoan: aphysiological chimera between alga and animal. Integr. Comp. Biol. 45, 595-604.
Gorboulev, V. I., Schurmann, A., Vallon, V., Kipp, H., Jaschke, A., Klessen, D.,Friedrich, A., Scherneck, S., Rieg, T., Cunard, R. et al. (2012). Na+-D-glucosecotransporter SGLT1 is pivotal for intestinal glucose absorption and glucose-dependent incretin secretion. Diabetes 61, 187-196.
Griffiths, C. L. Klumpp, D. W. (1996). Relationships between size, mantle area andzooxanthellae numbers in five species of giant clam (Tridacnidae). Mar. Ecol.Prog. Ser. 137, 139-147.
Haas, A. F. andWild, C. (2010). Composition analysis of organic matter released bycosmopolitan coral reef-associated green algae. Aquatic. Biol. 10, 131-138.
Hanquet, A.-C., Jouaux, A., Heude, C., Mathieu, M. and Kellner, K. (2011). Asodium glucose co-transporter (SGLT) for glucose transport intoCrassostrea gigasvesicular cells: impact of alimentation on its expression. Aquaculture 313, 123-128.
Hediger, M. A., Coady, M. J., Ikeda, T. S. and Wright, E. M. (1987). Expressioncloning and cDNA sequencing of the Na+/glucose co-transporter. Nature 330,379-381.
Hediger, M. A., Budarf, M. L., Emanuel, B. S., Mohandas, T. K. andWright, E. M.(1989). Assignment of the human intestinal Na+/glucose cotransporter gene(SGLT1) to the q11.2→qter region of chromosome 22. Genomics 4, 297-300.
Hernawan, U. E. (2008). Symbiosis between the giant clams (Bivalvia: Cardiidae)and zooxanthallae (Dinophyceae). Biodiversitas 9, 53-58.
Heslinga, G. A., Perron, F. E. and Orak, O. (1984). Mass culture of giant clams(f. Tridacnidae) in Palau. Aquaculture 39, 197-215.
Hiong, K. C., Choo, C. Y. L., Boo, M. V., Ching, B., Wong, W. P., Chew, S. F. andIp, Y. K. (2017a). A light-dependent ammonia-assimilating mechanism in thectenidia of a giant clam. Coral Reefs 36, 311-323.
Hiong, K. C., Cao-Pham, A. H., Choo, C. Y. L., Boo, M. V., Wong, W. P., Chew,S. F. and Ip, Y. K. (2017b). Light-dependent expression of a Na+/H+ exchanger3-like transporter in the ctenidium of the giant clam, Tridacna squamosa, can berelated to increased H+ excretion during light-enhanced calcification. Physiol.Rep. 5, e13209.
Hoer, D. R., Gibson, P. J., Tommerdahl, J. P., Lindquist, N. L. andMartens, C. S.(2018). Consumption of dissolved organic carbon by Caribbean reef sponges.Limnol. Oceanogr. 63, 337-351.
Huvet, A., Herpin, A., Degremont, L., Labreuche, Y., Samain, J.-F. andCunningham, C. (2004). The identification of genes from the oysterCrassostrea gigas that are differentially expressed in progeny exhibitingopposed susceptibility to summer mortality. Gene 343, 211-220.
Ip, Y. K., Ching, B., Hiong, K. C., Choo, C. Y., Boo, M. V., Wong, W. P. and Chew,S. F. (2015). Light induces changes in activities of Na+/K+-ATPase, H+/K+-ATPaseand glutamine synthetase in tissues involved directly or indirectly in light-enhancedcalcification in the giant clam, Tridacna squamosa. Front. Physiol. 6, 68.
Ip, Y. K., Hiong, K. C., Goh, E. J. K., Boo, M. V., Choo, C. Y. L., Ching, B., Wong,W. P. andChew, S. F. (2017a). Thewhitish inner mantle of the giant clam,Tridacnasquamosa, expresses an apical plasma membrane Ca2+-ATPase (PMCA) whichdisplays light-dependent gene and protein expressions. Front. Physiol. 8, 781.
Ip, Y. K., Koh, C. Z. Y., Hiong, K. C., Choo, C. Y. L., Boo, M. V., Wong, W. P., Neo,M. L. and Chew, S. F. (2017b). Carbonic anhydrase 2-like in the giant clam,Tridacna squamosa: characterization, localization, response to light, and possiblerole in the transport of inorganic carbon from the host to its symbionts. Physiol.Rep. 5, e13494.
Ip, Y. K., Hiong, K. C., Lim, L. J. Y., Choo, C. Y. L., Boo, M. V., Wong, W. P., Neo,M. L. and Chew, S. F. (2018). Molecular characterization, light-dependentexpression, and cellular localization of a host vacuolar-type H+-ATPase (VHA)subunit A in the giant clam, Tridacna squamosa, indicate the involvement of the
11
RESEARCH ARTICLE Journal of Experimental Biology (2019) 222, jeb195644. doi:10.1242/jeb.195644
Journal
ofEx
perim
entalB
iology

host VHA in the uptake of inorganic carbon and its supply to the symbioticzooxanthellae. Gene 659, 137-148.
Ishikura, M., Adchi, K. andMaruyama, T. (1999). Zooxanthellae release glucose inthe tissue of a giant clam, Tridacna crocea. Mar. Biol. 133, 665-673.
Ittekkot, V., Brockmann, U., Michaelis, W. and Degens, E. (1981). Dissolved freeand combined carbohydrates during a phytoplankton bloom in the North Sea.MarEcol. Prog. Ser. 4, 299-305.
Kimes, N. E., Van Nostrand, J. D., Weil, E., Zhou, J. Z. and Morris, P. J. (2010).Microbial functional structure of Montastraea faveolata, an important Caribbeanreef-building coral, differs between healthy and yellow-band diseased colonies.Environ. Microbiol. 12, 541-556.
Klumpp, D. W., Bayne, B. L. and Hawkins, A. J. S. (1992). Nutrition of the giantclam Tridacna gigas (L.) I. Contribution of filter feeding and photosynthates torespiration and growth. J. Exp. Mar. Biol. Ecol. 155, 105-122.
Koh, C. Z. Y., Hiong, K. C., Choo, C. Y. L., Boo, M. V., Wong, W. P., Chew, S. F.,Neo, M. L. and Ip, Y. K. (2018). Molecular characterization of a Dual DomainCarbonic Anhydrase from the ctenidium of the giant clam, Tridacna squamosa,and its expression levels after light exposure, cellular localization, and possiblerole in the uptake of exogenous inorganic carbon. Front. Physiol. 9, 281.
LaJeunesse, T. C., Parkinson, J. E., Gabrielson, P. W., Jeong, H. J., Reimer,J. D., Voolstra, C. R. and Santos, S. R. (2018). Systematic revision ofSymbiodiniaceae highlights the antiquity and diversity of coral endosymbionts.Curr. Biol. 28, 2570-2580.
Leggat,W., Marendy, E. M., Baillie, B., Whitney, S. M., Ludwig, M., Badger, M. R.and Yellowlees, D. (2002). Dinoflagellate symbioses: strategies and adaptationsfor the acquisition and fixation of inorganic carbon. Funct. Plant Biol. 29, 309-322.
Leggat, W., Dixon, R., Saleh, S. and Yellowlees, D. (2005). A novel carbonicanhydrase from the giant clam Tridacna gigas contains two carbonic anhydrasedomains. FEBS J. 272, 3297-3305.
Leggat, W., Ainsworth, T., Bythell, J. C., Dove, S., Gates, R. D., Hoegh-Guldberg, O., Iglesias-Prieto, R. and Yellowlees, D. (2007). The hologenometheory disregards the coral holobiont. Nat. Rev. Microbiol. 5, 826.
Leung, D.W., Loo, D. D. F., Hirayama, B. A., Zeuthen, T. andWright, E. M. (2000).Urea transport by cotransporters. J. Physiol. 528, 251-257.
Lewin, R. A. (1974). Biochemical taxonomy. In Algal Physiology and Biochemistry(ed. D. W. P. Stewart), pp. 1-39. Oxford: Blackwell Scientific Publications.
Loo, D. D. F., Jiang, X., Gorraitz, E., Hirayama, B. A. and Wright, E. M. (2013).Functional identification and characterization of sodium binding sites in Nasymporters. Proc. Natl. Acad. Sci. USA 110, 4557-4566.
Lucas, J. S., Nash,W. J., Crawford, C. M. and Braley, R. D. (1989). Environmentalinfluences on growth and survival during the ocean-nursery rearing of giant clams,Tridacna gigas. Aquaculture 80, 45-61.
Manahan, D. T. (1989). Amino acid fluxes to and from seawater in axenic veligerlarvae of a bivalve (Crassostrea gigas). Mar. Ecol. Prog. Ser. 53, 247-255.
Martınez-Quintana, J. A. and Yepiz-Plascencia, G. (2012). Glucose and otherhexoses transporters in marine invertebrates: a mini review. ElectronJ. Biotechnol. 15.
Mies, M. and Sumida, P. Y. G. (2012). Giant clam aquaculture: a review on inducedspawning and larval rearing. Int. J. Mar. Sci. 2, 62-69.
Mopper, K., Dawson, R., Liebezeit, G. and Ittekkot, V. (1980). Themonosaccharide spectra of natural waters. Mar. Chem. 10, 55-66.
Morganti, T.,Coma,R.,Yahel,G.andRibes,M. (2017). Trophicnicheseparation thatfacilitates co-existenceof high and lowmicrobial abundance sponges is revealed byin situ study of carbon and nitrogen fluxes. Limnol. Oceanogr. 62, 1963-1983.
Nelson, C. E., Goldberg, S. J., Kelly, L. W., Haas, A. F., Smith, J. E., Rohwer, F.and Carlson, C. A. (2013). Coral and macroalgal exudates vary in neutral sugarcomposition and differentially enrich reef bacterioplankton lineages. ISME J. 7,962-979.
Neo, M. L., Wabnitz, C. C. C., Braley, R. D., Heslinga, G. A., Fauvelot, C.,Wynsberge, S. V., Andrefouet, S., Waters, C., Tan, A. S. H., Gomez, E. et al.(2017). Giant clams (Bivalvia: Cardiidae: Tridacninae): a comprehensive updateof species and their distribution, current threats and conservation status.Oceanogr. Mar. Biol. 55, 87-387.
Norton, J. H. and Jones, G. W. (1992). The Giant Clam: an Anatomical andHistological Atlas. Canberra: Australian Centre for International AgriculturalResearch.
Norton, J. H., Shepherd, M. A., Long, H. M. and Fitt, W. K. (1992). Thezooxanthellal tubular system in the giant clam. Biol. Bull. 183, 503-506.
Pajor, A. M., Moon, D. A. and Wright, S. H. (1989). Sodium D-glucose cotransportin the gill of marine mussels: studies with intact tissue and brush bordermembrane vesicles. J. Membr. Biol. 107, 77-88.
Panayotova-Heiermann, M., Loo, D. D. F., Kong, C. T., Lever, J. E., and Wright,E. M. (1996). Sugar binding to Na+/glucose cotransporters is determined by thecarboxyl-terminal half of the protein. J. Biol. Chem. 271, 10029-10034.
Panayotova-Heiermann, M. and Wright, E. M. (2001). Mapping the urea channelthrough the rabbit Na+-glucose cotransporter SGLT1. J. Physiol. 535, 419-425.
Pedersen, H., Lomstein, B. A. and Blackburn, T. H. (1993). Evidence for bacterialurea production in marine sediments. FEMS Microbiol. Ecol. 12, 51-59.
Pequignat, E. (1973). A kinetic and autoradiographic study of the direct assimilationof amino acids and glucose by organs of the mussel Mytilus edulis.Mar. Biol. 19,227-244.
Putter, A. F. R. (1909). Die Ernahrung der Wassertiere und der Stoffhaushaft derGewasser. Jena: Fischer.
Rees, T. A. V., Fitt, W. K., Baillie, B. and Yellowlees, D. (1993a). A method fortemporal measurement of hemolymph composition in the giant clam symbiosisand its application to glucose and glycerol levels during a diel cycle. Limnol.Oceanogr. 38, 213-217.
Rees, T. A. V., Fitt, W. K. and Yellowlees, D. (1993b). The haemolymph and itstemporal relationship with zooxanthellae metabolism in the giant clam symbiosis.In Biology and Mariculture of Giant Clams (ed. W. K. Fitt), pp. 41-45. Canberra:Pirie Printers.
Reid, R. G. B., Fankboner, P. V. and Brand, D. G. (1984). Studies on thephysiology of the giant clam Tridacna gigas Linne—I. Feeding and digestions.Comp. Biochem. Physiol. 78, 95-101.
Rix, L., de Goeij, J. M., van Oevelen, D., Struck, U., Al-horani, F. A., Wild, C. andNaumann, M. S. (2016). Differential recycling of coral and algal dissolved organicmatter via the sponge loop. Funct. Ecol. 31, 778-789.
Rosenberg, E., Koren, O., Reshef, L., Efrony, R. and Zilber-Rosenberg, I. (2007).The role ofmicroorganisms in coral health, disease and evolution.Nature 5, 355-362.
Rosenberg, E. and Loya, Y. (2004). Coral Health and Disease. New York:Springer-Verlag.
Sano, Y., Kobayashi, S., Shirai, K., Takahata, N., Matsumoto, K., Watanabe, T.,Sowa, K. and Iwai, K. (2012). Past daily light cycle recorded in the strontium/calcium ratios of giant clam shells. Nat. Commun. 3, 761.
Sigrist, C. J., Cerutti, L., Hulo, N., Gattiker, A., Falquet, L., Pagni, M., Bairoch, A.and Bucher, P. (2002). PROSITE: a documented database using patterns andprofiles as motif descriptors. Brief. Bioinform. 3, 265-274.
Stephens, G. C. (1960). Uptake of glucose from solution by the solitary coral,Fungia. Science 131, 1532.
Stephens,G. C. (1962). Uptake of organicmaterial byaquatic invertebrates. I. Uptakeof glucose by the solitary coral, Fungia scutaria. Biol. Bull. 23, 648-660.
Stephens, G. C. and Schinske, R. A. (1961). Uptake of amino acids by marineinvertebrates. Limnol. Oceanogr. 6, 175-181.
Streamer, M., McNeil, Y. R. and Yellowlees, D. (1993). Photosynthetic carbondioxide fixation in zooxanthellae. Mar. Biol. 115, 195-198.
Takabayashi, M., Santos, S. R. and Cook, C. B. (2004). Mitochondrial DNAphylogeny of the symbiotic dinoflagellates (Symbiodinium, Dinophyta). J. Phycol.40, 160-164.
Tambutte, E., Allemand, D., Zoccola, D., Meibom, A., Lotto, S., Caminiti, N. andTambutte, S. (2007). Observations of the tissue-skeleton interface in thescleractinian coral Stylophora pistillata. Coral Reefs 26, 517-529.
Trench, R. K. (1987). Dinoflagellates in non-parasitic symbiosis. In The Biology ofDinoflagellates (ed. F. J. R. Taylor), pp. 530-570. Oxford: Blackwell Scientific.
Turk, E. and Wright, E. M. (1997). Membrane topology motifs in the SGLTcotransporter family. J. Membr. Biol. 159, 1-20.
Turk, E., Kerner, C. J., Lostao, M. P. andWright, E.M. (1996). Membrane topologyof the human Na+/glucose cotransporter SGLT1. J. Biol. Chem. 271, 1925-1934.
Turk, E., Kim, O., le Coutre, J., Whitelegge, J. P., Eskandari, S., Lam, J. T.,Kreman, M., Zampighi, G., Faull, K. F. and Wright, E. M. (2000). Molecularcharacterization of Vibrio parahaemolyticus vSGLT: a model for sodium-coupledsugar cotransporters. J. Biol. Chem. 275, 25711-25716.
Vaccaro, R. F., Hicks, S. E., Jannasch, H. W. and Carey, F. G. (1968). Theoccurrence and role of glucose in seawater. Limnol. Oceanogr. 13, 356-360.
Wafar, M. V. M., Wafar, S. and Devassy, V. P. (1986). Nitrogenous nutrients andprimary production in a tropical oceanic environment. Bull. Mar. Sci. 38, 273-284.
Welborn, J. R. and Manahan, D. T. (1990). Direct measurements of sugar uptakefrom seawater into molluscan larvae. Mar. Ecol. Prog. Ser. 65, 233-239.
Wilkerson, F. P. and Trench, R. K. (1986). Uptake of dissolved inorganic nitrogen bythe symbiotic clam Tridacna gigas and the coralAcropora sp.Mar. Biol. 93, 237-246.
Wood, I. S., Dyer, J., Hofmann, R. R. and Shirazi-Beechey, S. P. (2000).Expression of the Na+/glucose co-transporter (SGLT1) in the intestine of domesticand wild ruminants. Pflugers Arch. 441, 155-162.
Wright, E. M., Loo, D. D. F. and Hirayama, B. A. (2011). Biology of human sodiumglucose transporters. Physiol. Rev. 91, 733-794.
Wright, E. M. and Turk, E. (2004). The sodium glucose cotransport family SLC5.Pflugers Arch. 447, 510-518.
Yellowlees, D., Rees, T. A. V. and Leggat, W. (2008). Metabolic interactionsbetween algal symbionts and invertebrate hosts. Plant Cell Environ. 31, 679-694.
Zeuthen, T., Meinild, A.-K., Loo, D. D. F., Wright, E. M. and Klaerke, D. A. (2001).Isotonic transport by the Na+–glucose cotransporter SGLT1 from humans andrabbit. J. Physiol. 531, 631-644.
Zeuthen, T., Gorraitz, E., Her, K., Wright, E. M. and Loo, D. D. F. (2016). Structuraland functional significance of water permeation through cotransporters. Proc.Natl. Acad. Sci. USA 113, E6887-E6894.
12
RESEARCH ARTICLE Journal of Experimental Biology (2019) 222, jeb195644. doi:10.1242/jeb.195644
Journal
ofEx
perim
entalB
iology