inactivation, purification and stability of 146s antigens ... · inactivation, purification and...
TRANSCRIPT

Rev. sci. tech. Off. int. Epiz., 1984, 3 (2), 339-350.
Inactivation, purification and stability of 146S antigens of foot and mouth
disease virus for use as reagents in the complement fixation test
N.P. FERRIS, A.I. DONALDSON, I.T.R. BARNETT and R.W. OSBORNE*
Summary : The quantity of 146S antigen remaining after inactivation by acetylethyleneimine (AEI) or by formaldehyde plus AEI varied considerably between foot and mouth disease virus strains. Treatment with formaldehyde prior to AEI inactivation reduced the 146S recovery of several strains but improved I46S particle stabilisation during AEI inactivation of SAT 2 Kenya 183/74. Inactivated 146S antigens were found to be satisfactory for immunising guinea pigs and the minimum effective dose was around 0.64 µg. Concentrated 146S preparations were most stable when stored above liquid nitrogen.
INTRODUCTION
Traditionally, the complement fixation test (CF) has been used for the identification (typing) and classification (subtyping) of isolates of foot and mouth disease virus (FMDV) (4, 5, 11, 16). The reagents for such tests are prepared by a variety of methods in different laboratories around the world and there is a need for the development of standard procedures. In the case of antigens such methods would be for the preparation of reference sera and for use in subtyping. It would be advantageous if antigens were inactivated to permit safe handling within and transportat ion between laboratories and also if they were stable during long-term storage.
We have tested procedures for producing inactivated 146S antigens based on those proposed by Rowlands et al. (17). The suitability of these for use with different types and subtypes of FMDV, their stability during long-term storage at different temperatures, and their activity as immunogens for guinea pigs have been evaluated.
* The Animal Virus Research Institute, Pirbright, Woking, Surrey GU24 ONF, Great Britain.

— 340
M A T E R I A L S A N D M E T H O D S
Antigen production.
The following regimen was adopted for the production of inactivated 146S antigens :
1. Selected strains of the seven immunological types of FMDV (Table I) were grown in BHK-21 cells, the majority in monolayers and a few in suspension cultures, harvested when cultures showed maximal C P E and clarified by centrifugation.
2. The inactivation of infectivity was carried out by either of two procedures : fa) a 1:10 volume of 0 .5% formaldehyde in 0.04 M phosphate buffer ( P 0 4 buffer) was added (i.e. 0 .05% v/v final concentration) and suspensions maintained at 20°C for 72 hours, followed by the addition of a 1:10 volume of acetylethyleneimine (AEI; 0 .05% v/v final concentration) at either 26°C for 24 hours or 37°C for 6 hours (17). The inactivation reactions were stopped with excess sodium metabisulphite (solutions were immediately buffered with 8.8% sodium bicarbonate solution) and sodium thiosulphate, respectively, or (b) in the absence of the formaldehyde stage, AEI was added as previously described and virus cultures were maintained at 37 °C for an extended period of 24 hours (6).
Virus inactivation was verified by the method of Anderson et al. (1).
3. Concentration of virus and removal of lipid material was carried out using Polyethylene-glycol and trichloroethylene (TCE) (9). The aqueous suspensions resulting from TCE treatment were centrifuged at either 25,000 rpm for 3.5 hours or 30,000 rpm for 2 hours. Pellets were resuspended in P 0 4
buffer or 0.05 M tris buffer containing 0.01 M sodium chloride, p H 7.6. Nonidet P-40 was added to a final concentration of 1 % and samples layered on to 15-45% linear sucrose gradients buffered with the corresponding buffer and centrifuged at 30,000 rpm for 3 hours. Samples of the gradient were run through a flow cell of a Unicam SP 1800 ultraviolet spectrophotometer using a similar method to that described by Barteling and Meloen (3). The antigens were quantified by the method of Bachrach et al. (2), dispensed in glass containers treated with a water repellent (Repelcote)* and stored either at - 70°C or above liquid nitrogen as concentrated sucrose gradient material or diluted with buffer to contain either 5 or 10 µg/ml .
Guinea pig anti-146S serum.
The minimum effective single dose of 146S antigen required to inoculate guinea pigs was investigated using virus strains O France 1/81 and Asia 1 Cambodia 9/80. Guinea pigs (400-500 gm) were inoculated subcutaneously with varying concentrations of 146S antigen in 1 ml of buffer with an equal
* Hopkin and Williams, Chadwell Heath, Essex, England.

— 341 —
volume of Freund 's complete adjuvant. After 4 weeks, the guinea pigs were killed and exsanguinated, the serum separated and heat-inactivated at 56°C for 30 minutes. Antisera were raised against 146S antigens of other strains by using an inoculum containing 10 µg per animal.
Antigen stability.
To test the influence of formaldehyde treatment on the long-term stability of virus, pools of strains O BFS 1860 and C Resende antigens were prepared by both inactivation procedures, held at 4°C and tested spectrophotometri-cally at intervals for 146S content. Percentage losses were determined by comparison with control preparations of non-formaldehyde/AEI treated 146S antigens stored under the same conditions. Other antigens which had been stored at - 7 0 ° C and above liquid nitrogen for long periods, were also retested for 146S concentration.
The immunogenicity of the materials stored at 4°C and of others from above liquid nitrogen was examined by determination of ant ibody responses of guinea pigs inoculated with 10 µg as previously described.
Complement fixation test.
The micro-complement fixation test (CF) technique was employed (8). Reagents were added in the following volumes : fi) antiserum dilution, 25 µl; (ii) three 50% haemolytic doses of complement, 50 (A; (Hi) antigen (either tissue culture material or inactivated 146S at 1 µg /ml , 25 (A), and (iv) haemolytic indicator system, 25 ¡A. C F titres were expressed as the reciprocal of the serum dilution producing 7 5 % haemolysis.
Micro-neutralisation test.
Serum neutralisation tests were carried out in microtitre plates using the method employed by Golding et al. (13). Equal volumes of a dilution series of sera were mixed with 100 T C I D 5 0 virus and incubated at 37°C with an IB-RS-2 cell suspension. Neutralisation titres were expressed as the reciprocal of the final dilution of serum that neutralised 50% virus.
R E S U L T S
FMDV antigen production.
All strains yielded working harvests in the infectivity range l o g 1 0
7.5-8.5 T C I D 5 0 / m l ; however, such levels were more difficult to obtain with SAT 1 Tanzania 155/71 and SAT 2 R1215.
The concentrations of 146S particles obtained after inactivation and purification expressed as a ratio to 108 cells, the average number of cells in a BHK Roux monolayer, are shown in Table I.
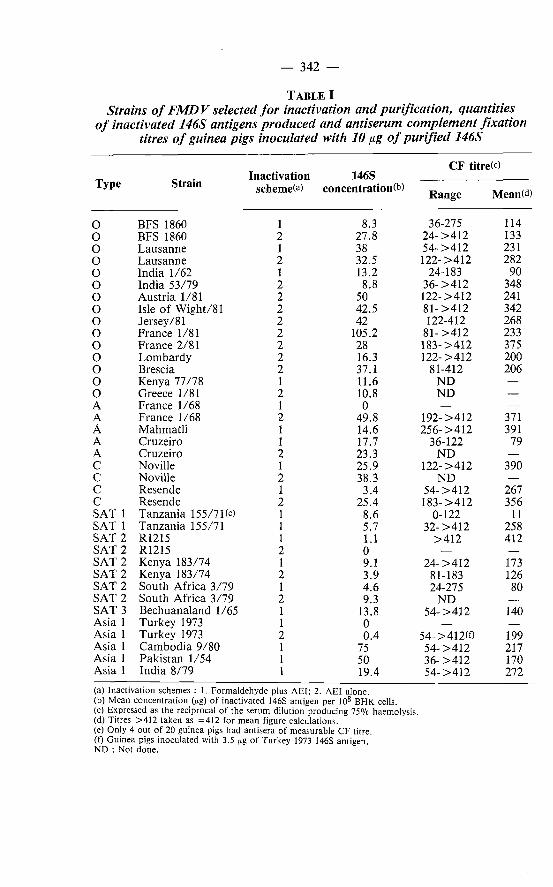
— 342
TABLE I Strains of FMDV selected for inactivation and purification, quantities
of inactivated 146S antigens produced and antiserum complement fixation titres of guinea pigs inoculated with 10 µg of purified 146S
CF titre(c)
Type Strain Inactivation 146S Type Strain schemed) concentration(b) Range Mean(d)
O BFS 1860 1 8.3 36-275 114 O BFS 1860 2 27.8 2 4 - > 4 1 2 133 O Lausanne 1 38 54 ->412 231 O Lausanne 2 32.5 122->412 282 O India 1/62 1 13.2 24-183 90 O India 53/79 2 8.8 36 ->412 348 O Austria 1/81 2 50 122->412 241 O Isle of Wight/81 2 42.5 81 ->412 342 O Jersey/81 2 42 122-412 268 O France 1/81 2 105.2 81 ->412 233 O France 2/81 2 28 183->412 375 O Lombardy 2 16.3 122->412 200 O Brescia 2 37.1 81-412 206 O Kenya 77/78 1 11.6 N D — o Greece 1/81 2 10.8 N D — A France 1/68 1 0 — — A France 1/68 2 49.8 192- >412 371 A Mahmatli 1 14.6 256->412 391 A Cruzeiro 1 17.7 36-122 79 A Cruzeiro 2 23.3 ND — C Noville 1 25.9 122->412 390 c Noville 2 38.3 N D — c Resende 1 3.4 5 4 -> 4 1 2 267 c Resende 2 25.4 183- >412 356 SAT 1 Tanzania 155/71(e) 1 8.6 0-122 11 SAT 1 Tanzania 155/71 1 5.7 3 2 -> 4 1 2 258 SAT 2 R1215 1 1.1 > 4 1 2 412 SAT 2 R1215 2 0 — — SAT 2 Kenya 183/74 1 9.1 2 4 -> 4 1 2 173 SAT 2 Kenya 183/74 2 3.9 81-183 126 SAT 2 South Africa 3/79 1 4.6 24-275 80 SAT 2 South Africa 3/79 2 9.3 ND SAT 3 Bechuanaland 1/65 1 13.8 54 ->412 140 Asia 1 Turkey 1973 1 0 — — Asia 1 Turkey 1973 2 0.4 5 4 - > 412(0 199 Asia 1 Cambodia 9/80 1 75 5 4 ->4 1 2 217 Asia 1 Pakistan 1/54 1 50 3 6 ->4 1 2 170 Asia 1 India 8/79 1 19.4 5 4 ->4 1 2 272
(a) Inactivation schemes : 1. Formaldehyde plus AEI; 2. AEI alone. (b) Mean concentration (µg) of inactivated 146S antigen per 10 8 BHK cells. (c) Expressed as the reciprocal of the serum dilution producing 75% haemolysis. (d) Titres > 4 1 2 taken as = 4 1 2 for mean figure calculations. (e) Only 4 out of 20 guinea pigs had antisera of measurable CF titre. (f) Guinea pigs inoculated with 3.5 µg of Turkey 1973 146S antigen. N D : Not done.

— 343 —
There was marked variation in the quantities of inactivated 146S obtained both between and within types of FMDV. Thus, two Asia 1 strains, Cambodia 9/80 and Pakistan 1/54, withstood formaldehyde pretreatment, judging by the high amounts of 146S antigen obtained, strain India 8/79 less well and strain Turkey 1973 not at all. Additionally, Turkey 1973 failed to withstand AEI inactivation alone. Although not all strains were inactivated by both regimens, direct comparisons can be made with some. O Lausanne and A Cruzeiro yielded similar levels of 146S by either procedure and C Noville produced slightly lower amounts by formaldehyde plus AEI treatment than by AEI inactivation alone (46.5% and 68 .9% of live 146S particle recovery respectively). In many cases inactivation by a combination of formaldehyde plus AEI significantly reduced the level of 146S particles that were recovered. From three separate experiments of inactivation of O BFS 1860, inactivation solely by AEI reduced the level of 146S particles by a mean of 3 4 . 1 % whereas formaldehyde plus AEI treatment reduced the yield by 8 0 . 5 % . Formaldehyde plus AEI inactivation reduced the 146S content of C Resende by 87% in comparison with the amount obtained by AEI treatment alone. Detectable quantities of A France 1/68 146S antigen only remained after AEI inactivation alone. SAT 2 Kenya 183/74 was the only strain which was stabilised by formaldehyde fixation.
In addition to Asia 1 Turkey 1973, difficulty was experienced in obtaining inactivated 146S antigens of SAT 1 Tanzania 155/71 and SAT 2 R1215. Five pools of Tanzania 155/71 were grown of high infectivity but only two produced measurable quantities of inactivated 146S particles. Similarly, one of six preparations of R1215 yielded 146S antigen and then in a small amount .
Guinea pig anti-146S serum.
The range and mean of the antiserum titres of guinea pigs inoculated with a single dose of 10 µg of antigen are listed in Table I. There was a wide variation in individual guinea pig antiserum titres although the majority of the strains generally stimulated a mean antiserum titre of >100 . The preparations of O India 1/62, A Cruzeiro and SAT 2 SAR 3/79 stimulated lower responses than others and it proved necessary to repeat the procedure with SAT 1 Tanzania 155/71 as the first antigen stimulated a response in only 20% of inoculated animals. The results in Table II illustrate that guinea pigs showed similar neutralising antibody responses with doses of 0.64 µg or more. Several individuals produced antiserum with detectable neutralising antibody at doses as low as 0.04 µg. There was no marked variation in antiserum CF titre when doses were reduced to 0.64 µg for O France 1/81, but CF titres were successively lower at each reduced dose of Asia 1 Cambodia 9/80.
Antigen stability.
Pools of O BFS 1860 antigens prepared by both inactivation procedures maintained the same 146S concentrations over the examination period of 19 weeks at 4°C. In contrast, C Resende 146S inactivated by formaldehyde
— 344 —
TA
BL
E
II
Ant
iser
um n
eutr
alis
atio
n an
d co
mpl
emen
t fix
atio
n tit
res
of g
uine
a pi
gs
inoc
ulat
ed w
ith v
aryi
ng d
oses
of
type
s O
Fra
nce
1/81
and
Asi
a 1
Cam
bodi
a 9/
80 1
46S
antig
ens
Ant
igen
N
eutr
alisa
tion
titre
* Pr
opor
tion
with
CF
titr
e**
Prop
ortio
n w
ith
Type
St
rain
d
ose (µ
g)
Ran
ge
Mea
n po
sitiv
e tit
re
Ran
ge
Mea
n po
sitiv
e tit
re
O
Fra
nce
1/81
10
.00
90-1
78 1
48
8/8
24-5
4 47
8/
8 2.
56
90-2
56 1
47
6/6
0-18
3 82
5/
6 0.
64
90-1
78 1
37
8/8
0-18
3 70
7/
8 0.
16
22-9
0 56
7/
7 0-
16
5 2/
7 0.
04
<6-
45
15
5/8
0 0
0/8
Asi
a 1
Cam
bodi
a 9/
80
10.0
0 25
6-70
8 37
4 8/
8 16
-275
88
8/
8 2.
56
128-
512
291
8/8
0-12
2 43
6/
8 0.
64
64-1
024
320
7/7
0-12
2 25
2/
7 0.
16
16-1
78 1
17
8/8
0-16
6
3/8
0.04
<
6-12
8 26
4/
7 0
0 0/
8
* E
xpre
ssed
as t
he
reci
proc
al o
f th
e se
rum
dil
utio
n th
at n
eutr
alis
ed 5
0% o
f vi
rus.
** C
F t
est
usin
g 1
fig/
ml
of i
nact
ivat
ed a
ntig
en.

— 345 —
plus AEI was reduced by 15% after 9 weeks. When inactivated by AEI alone, however, there was no reduction. AEI inactivated antigens stored at - 7 0 ° C lost from 6 .5% to 2 8 % of 146S particles over a period of 9 months (Table III), whereas antigens stored above liquid nitrogen showed either no or very little breakdown over periods up to 59 weeks.
The antibody responses of guinea pigs inoculated with antigens before and after storage are shown in Table IV. The results obtained with C Resende indicate that formaldehyde had reduced the quality of the antigen as both neutralisation and CF titres were depressed and a higher dose of formaldehyde-fixed material was required to provoke an adequate response (results not shown). However, the antigen appeared to maintain this same level of immunogenicity upon storage at 4°C while that prepared from AEI treatment alone induced a slightly lower neutralising antibody response than previously obtained. Guinea pigs inoculated with preparations of O BFS 1860 antigens stored at 4°C for 19 weeks produced antisera of similar neutralising and CF titre to those obtained with other freshly prepared O BFS 1860 antigens. Each of the strains retested from liquid nitrogen maintained immunogenicity upon storage.
DISCUSSION
Current methods for preparing reagents for C F tests for F M D diagnosis and subtyping vary at different laboratories and some can be criticised on the grounds of being less than optimal. For example, hyperimmune guinea pig sera is often used in both tests and the necessity of adapting FMDV strains to guinea pigs exerts a species-selective pressure which could result in sera of misleading reactivity. Fur thermore, the use of crude concentrated tissue culture harvests of virus in subtyping can reduce the specificity of CF reactions through the presence of 75S and 12S subunits (18, 7).
To eliminate some of these disadvantages it was considered that it might be of value to examine the suitability of inactivated 146S preparations of FMDV as immunogens for producing antiserum for typing and subtyping tests and as known positive controls for typing reactions. As a first step, procedures for producing inactivated 146S antigens were evaluated and a method involving combined formaldehyde and AEI treatment was compared with AEI treatment alone. Pretreatment with formaldehyde has been shown to prevent disruption by AEI (17) and to improve vaccine potency (15). It was considered that formaldehyde might, therefore, have a stabilising influence on other FMDV strains and protect against proteolytic degradation that has been shown to occur during storage of concentrated antigens (10).
While formaldehyde improved the stabilisation of SAT 2 Kenya 183/74 146S antigen, it had either little influence on the 146S recovery rates of some strains (e.g. O Lausanne), produced reduction of others (e.g. O BFS 1860, C Resende) or degraded others (e.g. A France 1/68). In several cases pretreat-
— 346
TA
BL
E I
II
Per
cent
age
redu
ctio
n in
146
S co
nten
t of
ant
igen
s st
ored
at
- 70
°C
and
abov
e liq
uid
nitr
ogen
(-
180°
C)
Stor
age
tem
pera
ture
Inac
ti-
-70°
C
-180
°C
Stra
in
vatio
n sc
hem
e*
Stor
age
time
% l
oss
of
Stor
age
time
% l
oss o
f (w
eeks
) 14
6S c
onte
nt
(wee
ks) 1
46S
cont
ent
1.5
0 1.5
4.1 1.9
0 3.2
3.1
2.3
0 0 0 0 0 0 2.0
1.1
0
41
41
41
41
41
58
38
41
41
58 6 6 45
11
32
58
59
59
55
14.3
13
.7
10.4
27
.9
6.5 0 N
D
ND
N
D
0 ND
N
D
ND
N
D
ND
0 0 0 0
36
36
34
35
35 4
ND
N
D
ND
2
ND
N
D
ND
N
D
ND
4 6 5 3
2 2 2 2 2
Aus
tria
1/8
1 Je
rsey
/81
Bre
scia
L
omba
rdy
Fran
ce 1
/81
Indi
a 1/
62
BFS
186
0 In
dia
53/7
9 K
enya
77/
78
Cru
zeir
o F
ranc
e 1/
68
Mah
mat
li R
esen
de
Nov
ille
T
anza
nia
155/
71
Bec
huan
alan
d 1/
65
Cam
bodi
a 9/
80
Paki
stan
1/5
4 In
dia
8/79
Type
O
O
O
o o o o o o A
A
- A
C
C
SAT
1
SAT
3
Asi
a 1
Asi
a 1
Asi
a 1
* ln
acli
vali
on s
chem
es :
1
. F
orm
alde
hyde
plu
s A
E1;
2.
AF
I al
one.
N
D :
N
ot
don
e.

- 347 -
TA
BL
E I
V
Antis
erum
neu
tralis
atio
n an
d co
mpl
emen
t fix
atio
n tit
res o
f gui
nea
pigs
in
ocul
ated
with
ant
igen
s befo
re a
nd a
fter
stora
ge
Neu
tral
isatio
n tit
re r
ange
C
F tit
re r
ange
**
. In
activ
atio
n St
orag
e St
orag
e tim
e ra
,n
sche
me*
te
mpe
ratu
re
(wee
ks)
Before storage A
fter storage
Bef
ore
stora
ge
Aft
er s
tora
ge
C
Res
ende
1
4°C
9
22
-128
(66
)***
16
-128
(78
) 0-
122
(51)
0-
275
(67)
C
Res
ende
2
4°C
9
90
-512
(26
9)
90-2
56 (
130)
81
-412
(17
8)
16-4
12 (
131)
A
Cru
zeir
o 1
-1
80
DC
46
N
T
128-
512
(276
) 36
-122
(79
) 81
-275
(14
4)
O
Fra
nce
1/81
2
-\80°C
63
25
6-10
8(43
8)
512-
1400
(93
4)
24-5
4 (4
7)
183-
>41
2 (
348)
Asi
a 1
Cam
bodi
a 9/
80 1
-1
80°C
51
25
6-70
8 (3
74)
355-
1400
(90
2)
16-2
75 (
88)
122-
>41
2 (3
54)
* In
acti
vati
on
sch
emes
: 1
. F
orm
alde
hyde
plu
s A
IH;
2.
AF
I al
one.
** C
F l
est
usin
g I
fig/
ml
of
inac
tiva
ted
anti
gen.
***
Fig
ures
in
bra
cket
s re
pres
ent
litr
e m
eans
, C
F l
itre
s >
4I2
tak
en a
s -4
12
.
NT :
Not
tes
ted.

— 348 —
ment with formaldehyde resulted in antigens of reduced immunogenicity for guinea pigs. This was not surprising as it is recognised that both the quality and mass of an antigen are important in influencing immunogenic potency (15, 9, 10).
The long-term storage stability of a range of 146S antigens at different temperatures was assessed both in terms of quantity by direct spectrophoto-metric measurement and of quality by guinea pig immunogenic response. The latter was taken as an indirect indication of the preservation of antigenic integrity. Investigations by Doel and Collen (10) of the practicality of storing F M D vaccines as concentrated preparations have highlighted the importance of preserving the antigenic polypeptide VP1 against proteolytic degradation.
It was found that only C Resende antigen prepared by combined formaldehyde and AEI treatment degraded at 4°C, judged by lower spectrophoto-metric readings and depressed antiserum titres. There was a variation in 146S degradation of strains stored at - 70°C, whilst those maintained above liquid nitrogen for long periods showed very little deterioration and their immunogenic properties were not greatly altered.
The minimum amount of antigen necessary to produce sera with workable antibody titres was found to be 0.64 µg, in agreement with Morgan et al. (14) and Forman (12). The specificity and performance of the guinea pig anti-146S sera in serological tests will be presented in a subsequent paper.
The choice of inactivation procedure, therefore, presents a problem in that while formaldehyde pretreatment may reduce 146S concentration prior to AEI inactivation and affect antigenic sites, the residual antigen is likely to maintain immunogenicity for longer at 4°C than without it. Its use, however, might be limited to strains which are susceptible to degradation during inactivation. An investigation to find alternative procedures for improving the antigen stability of labile strains would be worth pursuing.
ACKNOWLEDGEMENTS
We are grateful to Mr. A. R. Samuel for help in the preparation of strains type A France 1/68 and Mahmatl i , Mrs . J. Oxtoby for carrying out some of the complement fixation tests, Mr. K. Beamish and Mrs. T. Quigley for performing serum neutralisation tests and to Mr. L. Pullen and Miss S. M. Heugh for much assistance with guinea pigs.
* * *
INACTIVATION, PURIFICATION ET STABILITÉ DES ANTIGÈNES 146S DU VIRUS APHTEUX DESTINÉS A SERVIR DE RÉACTIFS POUR L'ÉPREUVE DE FIXATION DU COMPLÉMENT. — N.P. Ferris, A.I. Donaldson, I.T.R. Barnett et R.W. Osborne.
Résumé : La quantité d'antigène 146S restant présente après inactivation du virus aphteux par l'acétyléthylèneimine (AEI) ou par le formol

— 349 —
plus l'AEI s'est révélée extrêmement variable selon les souches étudiées. L'application de formol avant l'inactivation par l'AEI a réduit la quantité d'antigène 146S récupéré pour un certain nombre de souches mais a amélioré la stabilisation des particules 146S au cours de l'inactivation par l'AEI pour la souche SAT 2 Kenya 183/74. Les antigènes 146S inactivés se sont montrés satisfaisants pour l'immunisation des cobayes; la dose active minimale se situait autour de 0,64 µg. La meilleure stabilité des préparations concentrées de 146S a été obtenue par leur conservation sur azote liquide.
* * *
INACTIVACIÓN, PURIFICACIÓN Y ESTABILIDAD DE LOS ANTÍGENOS 146S DEL VIRUS AFTOSO PARA SERVIR DE REACTIVOS EN LA PRUEBA DE FIJACIÓN DEL COMPLEMENTO. — N.P. Ferris, A.I. Donaldson, I.T.R. Barnett y R.W. Osborne.
Resumen : Ha resultado sumamente variable la cantidad de antígeno 146S que permanece presente trás la inactivación del virus aftoso con acetiletileneimina (AEI) o con formol más AEI, según las cepas estudiadas. La aplicación de formol antes de la inactivación con AEI redujo la cantidad de antígeno 146S recuperado para algunas cepas, aunque mejoró la estabilización de las partículas 146S en el transcurso de la inactivación con AEI para la cepa SAT 2 Kenia 183/74. Los antí-genos 146S resultaron satisfactorios para inmunizar a los cobayos; se situaba la dosis mínima en torno a 0,64 µg. Se logró la mejor estabilidad de las preparaciones concentradas de 146S conservando las mismas en nitrógeno líquido.
* * *
REFERENCES
1. ANDERSON E . C . , CAPSTICK P . B . , M O W A T G . N . and LEECH F . B . (1970). — In vitro method for safety testing of foot and mouth disease vaccines. J. Hyg., Camb., 68, 159-172.
2. BACHRACH H . L . , T R A U T M A N R. and BREESE S.S. (1964). — Chemical and physical properties of virtually pure foot and mou th disease virus. Amer. J. vet. Res., 25, 333-342.
3. BARTELING S .J . and M E L O E N R .H . (1974). — A simple method for the quantification of 140S particles of foot and mouth disease virus (FMDV). Arch. ges. Virus-forsch., 45, 362-364.
4. BROOKSBY J . B . (1952). — The technique of complement fixation in foot and mouth disease research. A.R.C. Report Series No. 12. H M S O , London .
5. BROOKSBY J . B . (1968). — Variants and immunity : definitions for serological investigation. International Symposium on Foot and Mouth Disease : Variants and

— 350 —
Immunity, Lyon, 1 9 6 7 . Symposia Series in Immunological Standardization, 8, 1-1 0 .
6 . BROWN F . , HYSLOP N . S t . G . , CRICK J . and MORROW A . W . ( 1 9 6 3 ) . — The use of acetylethyleneimine in the production of inactivated foot and mouth disease vaccines. J. Hyg., Camb., 61, 3 3 7 - 3 4 4 .
7 . CARTWRIGHT B. , C H A P M A N W . G . and BROWN F . ( 1 9 8 0 ) . — Serological and immunological relationships between the 1 4 6 S and 1 2 S particles of foot and mouth disease virus. J. gen. Virol., 50, 3 6 9 - 3 7 5 .
8 . CASEY H . L . ( 1 9 6 5 ) . — Standardised diagnostic complement fixation method and adaptation to the micro test. Publ. Hlth. Monogr., 74, 1 -34 .
9 . D O E L T.R. and BACCARINI P . J . ( 1 9 8 1 ) . — Thermal stability of foot and mouth disease virus. Arch. Virol., 70, 2 1 - 3 2 .
1 0 . D O E L T .R. and COLLEN T. ( 1 9 8 2 ) . — Quantitative assessment of 1 4 6 S particles of foot and mouth disease virus in preparations destined for vaccines. J. Biol. Stand., 10, 6 9 - 8 1 .
1 1 . FONTAINE J . , MACKOWIAK C . and ROUMIANTZEFF M. ( 1 9 6 8 ) . — Types, sous-types et variantes du virus aphteux. Etude des variants. International Symposium on Foot and Mouth Disease : Variants and Immunity, Lyon, 1 9 6 7 . Symposia Series in Immunological Standardization, 8, 1 3 - 6 4 .
1 2 . FORMAN A . J . ( 1 9 7 5 ) . — A comparison of some immunological methods for the differentiation of strains of foot and mouth disease virus. J. Hyg., Camb., 74, 2 1 5 - 2 2 5 .
1 3 . GOLDING S . M . , HEDGER R . S . , TALBOT P . and W A T S O N J . ( 1 9 7 6 ) . — Radial immuno-diffusion and serum-neutralisation techniques for the assay of antibodies to swine vesicular disease. Res. vet. Sci., 20, 1 4 2 - 1 4 7 .
1 4 . MORGAN D .O . , BACHRACH H . L . and MCKERCHER P . D . ( 1 9 6 9 ) . — Immunogenicity of nanogram to milligram quantities of inactivated foot and mouth disease virus. 1. Relative virus-neutralizing potency of guinea pig sera. App. Microbiol., 17, 4 4 1 - 4 4 5 .
15 . MOWAT G . N . , MASTERS R . C . and PRINCE M . J . ( 1 9 7 3 ) . — Enhancement of immunizing potency of a foot and mouth disease vaccine for cattle by treatment of the antigen with formaldehyde. Arch. ges. Virusforsch., 41, 3 6 5 - 3 7 0 .
1 6 . PEREIRA H . G . ( 1 9 7 7 ) . — Subtyping of foot and mouth disease virus. Internatio. nal Symposium on Foot and Mouth Disease, Lyon, 1 9 7 6 . Dev. Biol. Stand., 35,
1 6 7 - 1 7 4 (S. Karger, Basel).
17 . ROWLANDS D . J . , SANGAR D.V. and BROWN F . ( 1 9 7 2 ) . — Stabilizing the immunizing antigen of foot and mouth disease virus by fixation with formaldehyde. Arch. ges. Virusforsch., 39, 2 7 4 - 2 8 3 .
1 8 . RWEYEMAMU M.M. , BOOTH J . C . , H E A D M. and P A Y T . W . F . ( 1 9 7 8 ) . — Micro-neutralization tests for serological typing and subtyping of foot and mouth disease virus strains. J. Hyg., Camb., 81, 1 0 7 - 1 2 3 .