in vivo nuclear magnetic resonance studies of the metabolism of methanol and pyruvate by...
TRANSCRIPT

FEMS Microbiology Reviews 87 (1990) 361-366 361 Published by Elsev/er
FEMSRE 00172
In vivo nuclear magnetic resonance studies of the metabolism of methanol and pyruvate by Methanosarcina barkeri
Helena Santos 1, Panla Fareleiia 2, Jean LeGall 3 and Antbnio V. Xavier 1
I Centro de Tecnologia .Qulmica e Biol6gica. CTQB/~VL ,-f:d 2 Esta~o Agron6mica Nacional. Quinta do Marquis. Oeiras. Portugal and 3 Department of Biochemistry. University of Georgia, Athens. GA. t_(SA.
Key words: Methanogenic bacteria; ~3C-NMR; 3~p-NMR; Bioenergetics
1. SUMMARY
The metabolism of methanol and pyruvate by cells of Methanosarcina barkeri was probed in vivo by NMR taking advantage of the non-inva- sire characteristics of this technique. Upon admin- istration of substrates, the kinetics of substrate consumption, the product formation and the en- ergetic state of the cells was monitored using carbon-13, phosphorus-31 or proton NMR. The effects of several inhibitors and uncouplers were investigated. Cells supplied with pyruvate devel- oped considerable levels of nucleotide triphos- phate; methane production was monitored, as well as CO2 and H2 formation. Most of the pyruvate was utilized for the synthesis of valine or inter- mediates of the valine pathway. The origin of the carbon atom in methane was elucidated using *3C-labelled pyruvate.
2. INTRODUCTION
Methanogenic bacteria are a group of anaerobic archaebacteria capable of coupling the formation
Correspondence to: H. Santos, Centro de Tecnologia Quimica e Biol6gica, CTQB/UNL, Apartado 127, 2780 Oeiras, Portugal.
of methane from a limited number of substrates with the synthesis of ATP [1,2]. The coupling mechanism between the formation of methane and the ATP synthesis remains controversial; however, a considerable amount of experimental data sup- porting a chemiosmotic type mechanism has been accumulated in recent years [3-7]. The results of experiments performed with Methanosarcina
barkeri are in accordance with the existence of a chemiosmotic mechanism for the synthesis of ATP from various substrates [4-7]. A hypothesis in- volving substrate level phosphorylation [8] has, so far, received no confirmation, but the possibility that this mechanism might be involved in the process of energy conservation in the oxidation of methanol to CO 2 is still regarded as plausible [9].
In recent years, in vivo nuclear magnetic reso- nance (NMR) has emerged as an attractive tech- nique to study cellular metabolism in a non-inva- sire way [10]. Phosphorus compounds can be de- tected by 31p-NMR if present in the cell in a concentration of approx. 0.5 mM or higher and if they are mobile. The measurement of the intracell- ular pH by in vivo 3~p-NMR is now straightfor- ward [11]; furthermore, information about the en- ergy status of the cell, rates of enzymatic reac- tions, the presence of unusual phosphorylated compounds, mobilization of phosphorus reserves,
0168-6445/90/$03.50 © 1990 Federation of European Microbiological Societies

362
etc, can be obtained without disturbing the cellu- lar processes [12]. In contrast to 3tp-NMR, in vivo t~C-NMR requires isotope enrichment and pro- rides information on the variations of metabolite levels and fluxes through biochemical pathways with no need for separative techniques. At pre- sent, the most serious disadvantage of in vivo NMR lies in the intrinsic low sensitivity of this technique.
Recently, we have used in vivo NMR, aiming to obtain additional information on the energy and carbon metabolisms of methanogenic bacteria. In this paper, we focus .on experiments performed with M. barkeri using methanol or pyruvate as substrates.
3. IN VlVO NMR STUDIES USING METH- ANOL AS SUBSTRATE; EFFECTS OF IN- HIBITORS AND UNCOUPLERS
M. barkeri, strain MS (DSM 800) was grown on methanol under the experimental conditions previously reported [13]. The consumption of [z3C]methanol was monitored by 13C-NMR. Reso- nances due to bicarbonate, methane and CO 2 were easily detected in addition to a resonance at 16.6 ppm, tentatively assigned to the C a of alanine; however, no resonances assignable to inter- mediates in the reduction of methanol to methane or the oxidation of methanol to CO 2 could be detected even when high concentrations of methanol were used, allowing longer times of acquisition. Furthermore, in the absence of inhibi- tors no resonances due to free formate or free formaldehyde were observed, in contrast to what was found with cells of aerobic methylotrophs [14]. This result is in accordance with the hypothe- sis that the single carbon units at the various stages of oxidation from methanol to CO 2 remain bound to carrier compounds [15]. In the presence of cyanide, accumulation of formate is observed (our unpublished results). Upon addition of in- hibitors of methanogenesis (chloroform or methyl viologen) the consumption of methanol and pro- duction of methane stopped immediately, but no new resonances were observed in the 13C-NMR spectra. The changes in the 3tp-NMR spectra of
~7
~P L. ~ . T P
c
s
NDP ~ N D P UDP-Hexose
A
l o 5 o -s . l o -15 -2o -2s PPM
Fig. 1. In vivo 31p-NMR spectra of a cell suspension of M. barkeri. Cells were suspended in a 50 mM MOPS buffer, pH 7.0. (A) Cell suspension under an argon atmosphere in starva- tion buffer. (B) Cells energized with methanol. (C) Cells after the addition of DCCD (300 nmol/mg protein). PME. phos- phomonoesters; Pl. inorganic phosphate; AcP, acetyl phos- phate; NTP. nucleotide triphosphates; NDP, nucleotide di-
phosphates.
M. barkeri cell suspensions upon energization with methanol are illustrated in Fig. 1. Prior to methanol addition (Fig. 1A) the spectrum is char- acterized by the presence of nucleotide diphos- phate (NDP) resonances, by a stron 3 phos- phomonoester peak (PME) and a resonance due to inorganic phosphate (Pi) In the phosphodiester region of the spectrum a resonance at -0 .5 ppm has been tentatively assigned to a phosphorylated cell wall or membrane component [16]. Addition of methanol leads to a build up of the phosphate peaks of nucleotide triphosphates (NTP) (Fig. 1B). It is interesting to note that the analysis of spectra of cell extracts shows that although ATP is the major contributor to the NTP resonances, about one third of the total NTP pool is due to uridine triphosphate. Other changes in the 31p-NMR spec- trum of the cells include the decreasing and broadening of the inorganic phosphate signal and the shift of the resonances due to inorganic phos- phate and phosphomonoester compounds, reflect- ing an acidification of the intracellular cell space. The intracellular pH changed from approx. 7.0 to a value lower than 6.5 in the steady state. In separate experiments, where the starting pH of the

external buffer was lower (6.5), a ApH of 0.5 was measured in the steady state, the intracellular pH being 6.1. The nucleotide triphosphate content as determined by in vivo NMR was 13 + 1 nmol/mg protein and a very high [NTP]/[NDP] ratio (> 10:1) for the free nucleotides was found.
The effects of the addition of the ionophores carbonylcyanide m-chlorophenylhydrazone (CCCP) and 3,3',4',5-tetrachlorosalicylanilide (TCS), and the ATP synthase inhibitor di- cyclohexylcarbodiimide (DCCD), to cell suspen- sions energized with methanol, were monitored by 13C-NMR to probe methanogenesis and by 31p. NMR to follow the changes in the levels of the phosphorylated metabolites. CCCP was unable to decrease the NTP level, probably due to imper- meability of the cell membrane to this compound. TCS stopped methanogenesis and abolished ApH as well as the resonances due to NTP, in agree- ment with observations by other authors [4]. These findings indicate the presence of a chemiosmotic type mechanism for the synthesis of ATE Ad- dition of DCCD in concentration of 30 nmol/mg protein had no effect on the levels of NTP and concentrations as high as 300 nmol/mg protein were necessary in order to reduce the NTP levels by 60~. These b.igh concentrations did not affect the rate of uptake of methanol (as judged from 13C-NMR), but the rate of methane production was slowed down (gas chromatography data) and a new resonance was observed in the alp-NMR spectrum of the cells (Fig. 1C). This resonance was assigned to acetyl phosphate on the basis of its chemical shift. This tentative assignment was also supported by the observation of small amounts of labelled acetate in the ~3C-NMR spec- tra of the cells, run after adding DCCD. The intensity of the resonance was variable in different experiments but reached values as high as those illustrated in Fig. 1C, for which a qualitative estimation of the concentration in the cell (0.5-1 mM) could be made by comparison with the levels of NTP (- -4 mM) determined in the absence of DCCD. The low efficiency of DCCD to decrease the NTP level is in agreement with observations of Sprott and Jarreii with an identical strain of M. barkeri [17]. The formation of acetyl phosphate in the presence of high concentrations of DCCD can
363
not be explained yet. Enzyme systems other than the ATP synthase might be affected by the high concentrations of DCCD, but the accumulation of a phosphorylatcd compound in a concentration high enough to allow detection by in vivo NMR is still surprising and deserves further investigation, since it might be connected with the presence of a subs*.rate level phosphorylation step.
4. IN VIVO NMR STUDIES ON THE METAB- OLISM OF PYRUVATE
Pyruvate is a key intermediate in the break- down of various fermentable substrates by anaerobic bacteria, in which it is converted into acetyl-CoA plus carbon dioxide and hydrogen. Part of the acetyl-CoA is converted into acetate via acetyl phosphate. All the enzyme systems re- quired for these conversions (pyruvate dehydro- genase, acetate kinase and phosphotr~sacetylase) have been detected in extracts of Methanosarcina spp. [18-20]. Pyruvate is also an important pre- cursor of cell material; in methanogens, both the autotrophic and methylotrophic carbon assimila- tion seem to involve the acetyl-CoA pathway [21]. Acetyl-CoA is then reductively carboxylated to pyp-'~'e and from acetyl-CoA and/or pyruvate all cell compounds are formed [15].
The in vivo NMR technique was used to study the metabolism of pyruvate by M. barkeri. When sodium pyravate (100 mM) was supplied to cell suspensions of 3t. barkeri, treated as previously described [13], methane was produced (~3C-NMR and IH-NMR) and nucleotide triphosphate synthesis was detected (31p-NMR). The NTP con- tent determined in the steady state was about 10 nmol/mg protein. Pyruvate was consumed at a rate of 20-50 nmol/min/mg protein. Mass spec- trometry was used as a complementary technique to monitor gas evolution derived from the metabolism of pyruvate (Santos, Fareleira, Berlier, Toci, LeGall and Xavier, manuscript in prepara- tion). H2, CO 2 and methane could be monitored in this way. Methane was produced from pyruvate at a rate of 6-8 nmol /min/mg protein and the total amount of methane produced corresoonded to about 10~ of the initial amount of pyruvate.
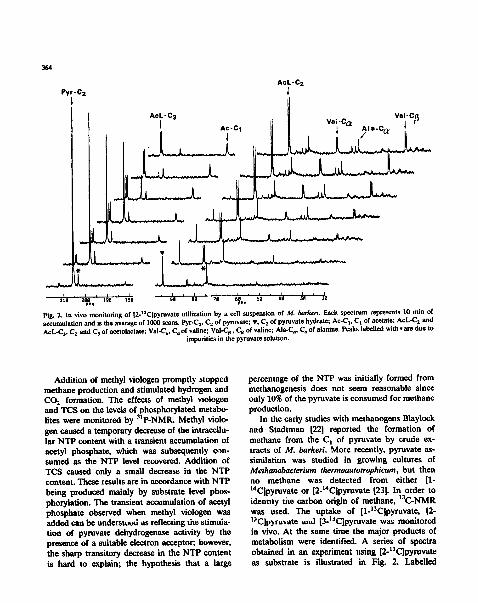
364
A c L - C 2 Pyr-C 2
II
A c L - C 3 ~ . - Va l - C/?,
Fig. 2. In vivo monitoring of [2-13C]pyruvate utilization by a cell suspension of M. barkeri. Each spectrum ~'epresents l0 rain of accumulation and is the average of 1000 scans. Pyr-C 2. C2 of pyruvate; v. C 2 of pyruvate hydrate; Ac-C I, Ct of acetate; AcL-C 2 and AcL-C3, C 2 and C 3 of acetolactate; VaI-C,,, C= of ratine; Val-C0, C o of valine; Ala-C,,, Ca of alanine. Peaks labelled with * are due to
impurities in the pyruvate solution.
Addition of methyl viologen promptly stopped methane production and stimulated hydrogen and CO 2 formation. The effects of methyl viologen and TCS on the levels of phosphorylated metabo- rites were monitored by 3~P-NMR. Methyl violo- gen caused a temporary decrease of the intraceilu- lar NTP content with a transient accumulation of acetyl phosphate, which was subsequently con- sumed as the NTP level recovered. Addition of TCS caused only a small decrease in the NTP content. These results are in accordance with NTP being produced mainly by substrate level pbos- phorylation. The transient accumulation of acetyl phosphate observed when methyl vioiogen was added can be understood as reflecting the stimula- tion of pyruvate dehydrogenase activity by the presence of a suitable electron acceptor; however, the sharp transitory decrease in the NTP content is hard to explain; the hypothesis that a large
percentage of the NTP was initially formed from methanogenesis does not seem reasonable since only 10% of the pyr)lvate is consumed for methane production.
In the early studies with methanogens Biaylock and Stadtman [22] reported the formation of methane from the C I of pyruvate by crude ex- tracts of M. barkeri. More recently, pyruvate as- similation was studied in growing cultures of Methanobacterium thermoautotrophicum, but then no methane was detected from either [1- 14C]pyruvate or [2-14C]pyruvate [23]. In order to identity tile carbon origin of methane, 13C-NMR was used. The uptake of [1-13C]pyruvate, [2- 13C]pyruvate a , d [3-~3C]pyruvate was monitored in vivo. At the same time the major products of metabolism were identified. A series of spectra obtained in an experiment using [2-13C]pyruvate as substrate is illustrated in Fig. 2. Labelled
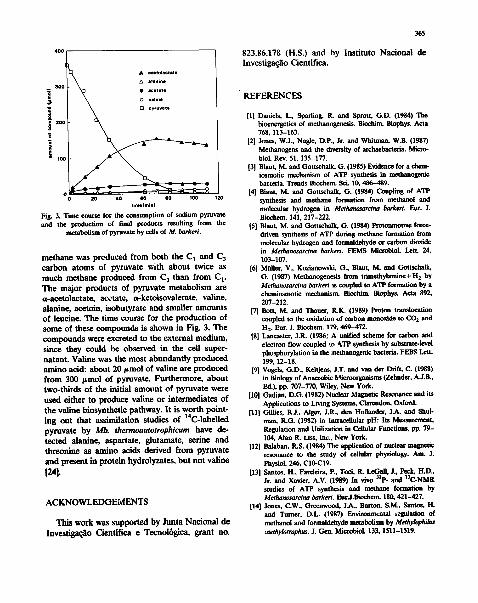
400~ • acetolsctate alanine
3OO o • aceta le
o va.ne D pyruvate
s l o ~
timllminl Fig. 3. Time course for the consumption of sodium pyruvate end the production of final products resulting from the
metabolism of pyruvate by cells of M. barkeri.
methane was produced from both the C 1 and C 3 carbon atoms of pyruvate with about twice as much methane produced from C 3 than from C i. The major products of pyruvate metabol i sm are a-acetolactate, acetate, a-ketoisovalerate, rat ine, alanine, acetoin, isobutyrate and smaller amount s of leucine. The t ime course for the p roduc t ion of some of these compounds is shown in Fig. 3. The c o m p o u n d s were excreted to the external medium, since they could be observed in the cell super- na tant . Valine was the most abundant ly p roduced amino acid: about 20 ~tmol o f ra t ine are p roduced f rom 300 t tmol of pyruvate. Fur thermore , about two-thi rds o f the initial amotmt of pyruvate were used either to p roduce valine or in termedia tes of the ra t ine biosynthet ic pathway. I t is wor th poin t - ing out that assimilat ion studies of 14C-labelled pyruvate by Mb. thermoautotrophicum have de- tec ted alanine, aspartate, glutamate, serine and threonine as amino acids derived f rom pyrnva te and present in prote in hydrolyzates, bu t no t valine
[241.
A C K N O W L E D G E M E N T S
This work was suppor ted by Junta Naciona l de I n v e s f i g a ~ o Cientifica e Tecnol6gica, grant no.
365
823.86.178 (H.S.) and by lns t i tu to Nacional de Invest igas~o Cientifica.
"REFERENCES
[I] Daniels, L., Sparring, R. and Sprott, G.D. (1984) The bioenergetics of methanogenesis. Bioehim. Biophys. A m 768,113-163.
[2] Jones, WJ., Nasle, D.P., Jr. and Whitman, W.B. (1987) Methanogens and the diversity of archaebacteria. Micro- biol. Rev. 51,135-177.
[3] Blaut* M. and Gottschalk" G. (1985) Evidence for a chenv iosmotic mechanism of ATP synthesis in mcthano$enic bacteria. Trends Biuchem. Sci. 10, 486-489.
[4] B|aut, M. and Gottschalk, G. (1984) Coupling of ATP synthesis and methane formation from methanol and molecular hydrogen in Methanosarcina barkeri. Eor. J. Biuchem. 141, 217-222.
[5] Blaut, M. and Gottschalk, G. (1984) Protonmotive force- driven synthesis of ATP during methane formation from molecular hydrogen and formaldehyde or carbon dioxide in Methanosarcina barkeri. FEMS Microbiol. Len. 24, 103-107.
[6] MUller, V., Kozianowski, G., Blaut, M. and Gottschalk, G. (1987) Methanogenesis from ~ y l a m i n e + H 2 by Methanosarcina barkeri is coupled to ATP formation by a chemiosmotic mechanism. Bioehim. Biophys. Acta 892, 207-212.
[7] Bott, M. and Thauer, R.K. (1989) Proton translocation coupled to the oxidation of carbon monoxide to CO 2 and H 2. Eur. J. Biochem. 179, 469-472.
[8] Lancaster, J.R. (1986) A unified scheme for carbon and electron flow coupled ~o ATP synthesis by substrate-level phosphorylation in the methanogenic bacteria. FEBS Lett. 199,12-18.
[9] Vogels, G.D., Keitjens, J.T. and van der Drift. C. (1988) in Biology of Anaerobic Microorganisms (Zehnder, A.J.B., Ed.), pp. 707-770, Wiley, New York.
[10] Gadian, D.G. (1982) Nuclear Magnetic Resonance and its Applications to Living Systems, Clarendon, Oxford.
[11] Gillies, R.J., Alger, J.R., den Hollander, J.A. and Shut- man, R.G. (1982) in Intracellular pH: Its M ~ t . Regulation and Utilization in Cellular Functions, pp. 79- 104, Alan R. Liss, Inc., New York.
[12] Balaban, R.S. (1984) The application of nuclear magnetic resonance to the study of cellular physiology. Am. J. Physiol. 246, C10-C19.
[13] Santos, H., Fareleira, P., To,A, R. LeGall, J., Peck, H.D., Jr. and Xavier, A.V. (1989) In vivo Sip. and 13C-NMR studies of ATP synthe~Jis and methane formation by Methanosarcina barkeri. EurJ.Biuchem. 180, 421--427.
[14] Jones, C.W., Greenwood, J.A., Burton, S.M., Santos, H. and Turner, D.L. (1987) Enviromn~ntal ~qgulation of methanol and formaldehyde metabolism by Methy/oph//us ,.nethyl6trophus. J. Gen. Microbiol. 133,1511-1519.

366
[15] Zeikus, J.G. (1983) Metabolism of one-carbon compounds by chemotrophic anaerobes. Adv. Microb. Physiol. 25, 215-299.
[16] Tolman, CJ., Kanodia, S., Roberts, M.F. and Daniels, L. (1986) 31p-NMR spectra of methanogens: 2,3.cyclopyro- phosphoi0ycerate is detectable only in methanobecteria strains. Biochim. Biophys. Acta 886, 345-352.
[17] Sprott, G.D. and Jarrell, K.F. (1982) Sensitivity of methanosenic bacteria to dicyclohexylcarbodiimide. Can. J. Microbiol. 28, 982-986.
[18] Kl'zycki, J.A., Woikin, R.H. and Zeikus, J.G. (1982) Com- parison of unltrophic and mixotrophlc substrate metabo- lism by an acetate-adapted strain of Methanosarcina barkeri. J. Bacteriol. 149, 247-254.
[19] Kenealy, W.R. and Zeikus, J.G. (1982) One-carbon metabofism in methanogens: evidence for synthesis of a two-carbon cellular intermediate and unification of catabolism and anabolism in Methanosarcina barkeri. J. Bacteriol. 151, 932-941.
[20] Blant, M. and Gottschalk, G. (1982) Effect of trimethyl- amine on acetate utilization by Methanosarci,a barkeri. Arch. Microbiol. 133, 230-235.
[21] Wood, H.G., Ragsdale, S.W. and Pezacka, EL. (1986) The acetyI-CoA pathway of autotrophic growth. FEMS Micro- biol. Rev. 39, 345-362.
[22] Blaylock, B.A. and Stadtman, T.C. (1966) Methane bio- synthesis by Methanosarcina barkeri. Properties of the soluble enzyme systems. Arch. Biochem. Biophys. 116, 138-158.
[23] Httster, R. and Thauer, R.K. (1983) Pyruvate assimilation by Metkanobacterium thermoautotrophicum. FEMS Micro- biol. Left. 19, 207-209.
[24] Holder, U., Schmidt, D.-E., Stupperich, E. and Fuchs, G. (1985) Autotrophic synthesis of activated acetic acid from two CO 2 in Methanobacterium thermoautotrophicum. IlL Evidence for common one-carbon precursor pool and the role of corrinoid. Arch. Microbiol. 1,,1, 229-238.