in copyright - non-commercial use permitted rights / … · 2018-01-11 · parameters influencing...
TRANSCRIPT

Research Collection
Doctoral Thesis
Parameters influencing efficient T cell repertoire selection
Author(s): Martinic, Marianne M.A.
Publication Date: 2003
Permanent Link: https://doi.org/10.3929/ethz-a-004618228
Rights / License: In Copyright - Non-Commercial Use Permitted
This page was generated automatically upon download from the ETH Zurich Research Collection. For moreinformation please consult the Terms of use.
ETH Library

Diss. ETH No.: 15096
PARAMETERS INFLUENCING EFFICIENT
T CELL REPERTOIRE SELECTION
A dissertation submitted to the
SWISS FEDERAL INSTITUTE OF TECHNOLOGY ZURICH
for the degree of
Doctor of Natural Sciences
presented by
MARIANNE M. A. MARTINIC
Dipl. Natw. ETH
born 29.11.1974
from France
Accepted on the recommendation of
Prof. Dr. H. Hengartner, examiner
Prof. Dr. R.M. Zinkernagel, co-examiner
2003


Meinen Eltern


Table of Contents
- 5 -
Table of Contents
1 Summary..................................................................................................... 7
2 Zusammenfassung .................................................................................... 9
3 Abbreviations ........................................................................................... 11
4 Introduction .............................................................................................. 13
4.1 Thymus architecture and development ................................................. 13
4.2 T cell development................................................................................... 15
4.2.1 Commitment to the ααααββββ or γγγγδδδδ T cell lineage .................................. 18
4.2.2 T cell repertoire selection............................................................. 19
4.2.3 CD4/CD8 T cell lineage commitment ........................................... 21
4.2.4 NK / γγγγδδδδ / CD8αααααααα+ / CD4+CD25+ T cells........................................... 21
4.3 H-Y-specific TCR transgenic mice.......................................................... 23
4.4 Central Question ...................................................................................... 26
5 Results Part I:
Efficient T cell repertoire selection in tetraparental chimeric mice
independent of thymic epithelial MHC ....................................................27
6 Results Part II:
Influence of MHC class I H-2Db density on selection and survival of
H-Y-specific TCR transgenic T cells....................................................... 43
7 Results Part III
Selection of the H-Y-specific transgenic TCR in an athymic versus
euthymic environment............................................................................. 57
8 General Discussion ................................................................................. 73
9 References................................................................................................ 83
10 Curriculum Vitae ...................................................................................... 93
11 Bibliography ............................................................................................. 95
12 Danke, Merci, Thank You, Gracias ......................................................... 97


Summary
- 7 -
1 Summary
During T cell maturation, T cell precursors migrate from the bone marrow via the
bloodstream into the thymus. In the thymus, maturing thymocytes first rearrange their T cell
receptor (TCR)-chain genes followed by a stringent selection process. During this selection
process, only thymocytes expressing productively rearranged TCR with weak to intermediate
overall avidity to self-MHC (major histocompatibility complex)/self-peptide complex (useful
TCR) receive a survival signal (positive selection) whereas thymocytes expressing TCR with
high overall avidity to self-MHC/self-peptide complex (potentially self-reactive TCR) die via
TCR-induced apoptosis (negative selection). This selection process ensures survival of
exclusively self-MHC restricted and self-tolerant thymocytes. During the last maturation step,
thymocytes are committed to the CD4 or CD8 T cell lineage followed by emigration of now
mature and functional T cells into the periphery. Survival and expansion of these T cells,
however, is only guaranteed if they remain in continuous interaction with self-MHC.
The aim of this study is to obtain a better understanding of the requirements needed for
efficient T cell repertoire selection and survival. The first part of the results section addressed
the role of thymic epithelial (TE) versus non-TE MHC in T cell repertoire selection. In the
second and third part of the results section this thesis focused on selection and survival of
the H-Y-specific transgenic TCR, which is specific for a male antigen-derived peptide (H-Y)
presented on MHC class I H-2Db molecules. The influence of TCR-restricting H-2Db density
on H-Y-specific transgenic TCR selection and survival was analysed in the second part of the
results section. Finally, the last part compared selection of the H-Y-specific transgenic TCR
in a euthymic (optimal) versus an athymic (sub-optimal) environment.
The data obtained here showed that 1) non-TE MHC was sufficient to efficiently select a
mature and functional T cell repertoire, 2) efficiency of selection and survival of low-affinity
TCR was dependent on optimal TCR-restricting MHC density and 3) although under sub-
optimal conditions, protection against self-reactivity was still guaranteed, the efficiency of
positive selection, however, was too low to provide protective immunocompetence under
physiological conditions (i.e. in a non-TCR transgenic situation).


Zusammenfassung
- 9 -
2 Zusammenfassung
Während der T-Zell-Reifung wandern Vorläufer-T-Zellen aus dem Knochenmark über die
Blutbahn in den Thymus. Im Thymus beginnen die Thymozyten zuerst mit der
Rearrangierung ihrer T-Zell-Rezeptor (TCR)-Kettengene gefolgt von einem strikten
Selektionsprozess. Während dieses Selektionsprozesses werden nur diejenigen
Thymozyten, die TCR mit einer geringen bis mittleren Gesamtbindungsstärke für Selbst-
MHC (Haupthistokompatibilitätsantigen-Komplex)/Selbst-Peptid-Komplex exprimieren
(nützliche TCR), ein Überlebenssignal erhalten (positive Selektion). Thymozyten, deren TCR
eine hohe Gesamtbindungsstärke für Selbst-MHC/Selbst-Peptid Komplex aufweisen
(potentiell selbst-reaktive TCR), sterben durch TCR-vermittelte Apoptose (negative
Selektion). Dieser Selektionsprozess stellt somit sicher, dass ausschliesslich Selbst-MHC-
restringierte und selbst-tolerante Thymozyten überleben. Während der letzten Reifungs-
Etappe erfolgt der Entscheid zur CD4- oder zur CD8-positiven T Zelle mit anschliessender
Emigration in die Peripherie als reife und funktionelle T Zelle. Das weitere Überleben sowie
die Expansion dieser T Zelle sind nur dann garantiert, wenn sie in ständigem Kontakt mit
Selbst-MHC bleibt.
Das Ziel dieser Arbeit ist es, ein besseres Verständnis über die verschiedenen Bedingungen
zu erhalten, die zum Erreichen einer effizienten T Zell Repertoire-Selektion notwendig sind.
Im ersten Resultate-Teil wurde die Funktion von Thymusepithel (TE) versus Nicht-TE MHC
bei der T Zell Repertoire-Selektion untersucht. In den darauffolgenden Resultate-Teilen
wurden die Selektion und das Überleben des H-Y-spezifischen, transgenen TCR analysiert,
welcher ein männliches Peptidantigen auf MHC Klasse I Molekül H-2Db spezifisch erkennt.
Der Einfluss der TCR-restringierenden H-2Db-Dichte auf die Selektion und das Überleben
des H-Y-spezifischen transgenen TCR wurde im zweiten Resultate-Teil verfolgt. Zum
Abschluss wurde im dritten Resultate-Teil die Selektion des H-Y-spezifischen transgenen
TCR in einer athymischen (sub-optimal) und in einer normalen Umgebung verglichen.
Die hier erhaltenen Daten konnten zeigen, 1) dass Nicht-TE MHC ausreichend war, um ein
reifes und funktionelles T Zell Repertoire zu selektionieren, 2) dass die Effizienz der
Selektion und des Überlebens niedrig affiner TCR von einer optimalen Dichte an TCR-
restringierenden MHC-Molekülen abhing und 3) dass unter sub-optimalen Bedingungen der
Schutz gegen Selbst-Reaktivität zwar gegeben, die Effizienz der positiven Selektion dagegen
aber viel zu niedrig war, um unter physiologischen Bedingungen (z. B. in einer nicht-TCR
transgenen Situation) noch genügend schützende Immunkompetenz zu vermitteln.


Abbreviations
- 11 -
3 Abbreviations
APC Antigen presenting cells
BM Bonne marrow
CLP Common lymphoid precursors
cTEC Cortical TEC
CTL Cytotoxic T lymphocytes
DAG Diacylglycerol
DC Dendritic cells
DEC Dendritic epidermal cells
DN Double negative
DP Double positive
ED Embryonic day
ELISA Enzyme-linked immunosorbent assay
end. Endogenous
ERK Extracellular-signal-regulated kinase
FL Fetal liver
FLT3+ HSC Fms-like tyrosine kinase 3+ HSC
Gads SH2 and SH3 domain-containing adaptor
proteins, bind to tyrosine-phosphatase Shc
GPI Glucose-6-phosphate-isomerase
Grb2 Growth-factor receptor-bound protein 2
HSA Heat stable antigen, CD24
HSC Haematopoietic stem cells
H-Y Male antigen-derived peptide
IEL Intestinal intraepithelial lymphocytes
Ig Immunoglobulin
IgH Immunoglobulin heavy chain
IgL Immunoglobulin light chain
IP3 Inositol-1,4,5-trisphosphate
ISP Immature single positive
ITAM Immuno-tyrosine based activation motif
Itk Tec-family tyrosine kinase
JNK c-jun N-terminal kinase
LAT Linker for activation of T cells
LCMV Lymphocytic choriomeningitis virus

Abbreviations
- 12 -
LCMV-GP LCMV-glycoprotein
LCMV-NP LCMV-nucleoprotein
MFI Mean fluorescence intensity
MHC Major histocompatibility complex
MLP Myeloid- and lymphoid precursors
mTEC Medullary TEC
NF-κB Nuclear factor κB
NF-AT Nuclear factor of activated T cells
NK Natural killer cells
p38 MAP kinase
pfu Plaque-forming units
PKC Protein kinase C
PLC-γ1 Phospholipase C-γ1
PNAr Peanut agglutinin receptor
pTα Invariant pre-TCRα chain
Rag-1,2 Recombination-activating genes-1,2
Ras Protein products regulate cellular growth and
differentiation (family of proto-oncogenes)
RasGRP Ras activator with a DAG-binding C1 domain
Scid Severe combined immunodeficiency
Slp76 Cellular adaptor protein
SP Single positive
TCR T cell receptor
TCRα-CPM TCRα-chain connecting peptide motif
TE Thymic epithelial
TEC Thymic epithelial cells
tg Transgenic
TN Triple negative
VSV Vesicular stomatitis virus
VSV-IND VSV serotype Indiana
WHN Winged-helix nude
ZAP-70 Zeta-Associated Protein-70 Src-family
tyrosine kinase

Introduction
- 13 -
4 Introduction
4.1 Thymus architecture and development
In mice as well as in humans, the thymus is the organ where progenitor T cells
differentiate to mature T cells expressing a self-MHC-restricted and self-tolerant T
cell receptor (TCR). The thymus lies in the upper thorax of vertebrate animals,
resting on the heart and extending into the base of the neck. It is bilaterally
symmetrical, composed of two lobes, which join at the midline. It consists primarily of
T cells at different developmental stages (thymocytes) and a heterogeneous group of
supporting cells - thymic epithelial cells, fibroblasts, macrophages, dendritic cells and
B cells - forming the thymic stroma. A capsule of connective tissue, which repeatedly
invaginates to form septae leading to numerous lobules, surrounds the thymic lobes,
each filled with thymocytes and stromal cells (Figure 1). Each thymus lobule is
organized into three different regions: a subcapsular, cortical and medullary zone.
Baskets of epithelial reticular cells characterize the subcapsular region. Fibroblasts,
macrophages and a network of spider-shaped and sheetlike epithelial cells are
located in the cortical region. The medullary region consists of numerous dendritic
cells, macrophages and a network of stubby epithelial cells. B cells are mostly
present in the corticomedullary junction. The different cell-type compositions in each
region lead to distinct MHC-staining patterns: a weak staining in the subcapsular
region, a reticular pattern in the cortex (due to epithelial network) and a confluent
pattern in the medulla (due mainly to dendritic cells and a subset of medullary
epithelial cells) (Benoist and Mathis, 1999).
In mice, the beginning of thymus development starts at fetal day 9 with the
invagination of the ectoderm of the third branchial cleft and the endoderm of the third
or fourth pharyngeal pouch (Anderson et al., 1996). With continuous invagination, the
ectoderm surrounds the endoderm. The developing organ becomes surrounded by
mesenchymal cells initially of neural crest and later of mesodermal origin, playing an
inductive as well as structural role during thymus development (Le Lievre and Le
Douarin, 1975). At fetal day 11, invaginations close to form a thymic rudiment or
anlage. Cells of ectodermal origin give rise to cortical thymic epithelial cells (cTEC);
cells of endodermal origin give rise to medullary TEC (mTEC). Blood vessels
associated with mesoderm-derived mesenchyme permeate the thymus anlage

Introduction
- 14 -
allowing cells of haematopoietic origin (from the yolk sac or fetal liver) to enter. From
this point on, all progenitor T cells as well as stromal cells are present in the
developing thymus. Thymus architecture, however, is not completed yet. There is no
distinct division into a subcapsular, cortical and medullary region. Only few medullary
islets are present in the dominating cortical region. Just before birth a distinct
medullary region appears in parallel with the emergence of mature αβT cells. The
right three-dimensional thymus architecture as well as development of mature T cells
in the thymus is only achieved in the presence of a proper cross talk between
developing T cells and thymic stromal cells (Anderson and Jenkinson, 2001).
Nude mice have a loss-of-function mutation in the WHN (winged-helix nude) gene
encoding for the transcription factor WHN. The thymus anlage can still develop in
absence of WHN but it will only contain primitive epithelial cells unable to differentiate
into mature thymic epithelial cells. The nude thymic rudiment will therefore never
establish a functional three-dimensional epithelial network nor have the ability to
promote the development of progenitor T cells into mature T cells.
Figure 1: Thymus organization. A thymus section through a single lobule is shown. The different forms of stromal
cells in each region are indicated: cortical and medullary epithelial cells, dendritic cells, macrophages, and
fibroblasts. The maturation of thymocytes from blasts in the subcapsular zone, to small resting cells in the cortex,
to larger cells in the medulla is also illustrated (Benoist and Mathis, 1999).

Introduction
- 15 -
Figure 2: Development of TCRαβ+ T cells in the thymus. The major different stages and sites of thymocyte-
development are shown from the most immature stage on the left (DN stage) to the most mature stage on the
right (SP stage). Time-span of different TCR-chain gene rearrangements, expression of pre-TCR and TCRαβ,
degree of cell division, time spent at the various stages, percentage of thymocytes belonging to the different
stages out ot total thymocytes, anatomic location in which thymocytes reside during the different stages, and total
thymocyte numbers passing the thymus per day are shown from top to bottom. Adapted from (Benoist and
Mathis, 1999).
4.2 T cell development
Haematopoietic stem cells (HSC) are pluripotent and show a self-renewal capacity.
In the murine fetal liver, HSC differentiate to lymphoid precursors with myeloid- and
lymphoid- reconstitution potential (FL-MLP) (Akashi et al., 2000; Cumano et al.,
1992; Sagara et al., 1997) and then further on to either cells committed to the B or to
the T cell lineage (Kawamoto et al., 2000; Kawamoto et al., 1997) (Figure 2).
Whereas during embryonic development T lineage commitment occurs prior to
thymic entry, in adults, T lineage commitment occurs in the thymus and is Notch-1
signalling-dependent (Ashton-Rickardt et al., 1994; Osborne and Miele, 1999). In the
adult bone marrow (BM), HSC differentiate first to Flt3+ HSC (fms-like tyrosine kinase
3) with transient myeloid and long-term lymphoid reconstitution potential and then to
common lymphoid precursors (CLP) (Figure 2). These cells show only limited self-

Introduction
- 16 -
renewal capacity and have a potential of producing lymphoid but not myeloid cell
types such as T, B, natural killer (NK) and dendritic cells (DC) (Borowski et al., 2002).
In absence of Notch-1 signalling, CLP develop to the Pro-B cell stage (Borowski et
al., 2002). In presence of Notch-1 signalling, CLP migrate via the bloodstream into
the thymus to further develop to the Pro-T cell stage. CLP first entering the thymus at
the corticomedullary junction are CD4-CD8-TCR- triple negative (TN) (Figure 2). This
stage can further be divided into four different stages depending on expression levels
of CD44 and CD25. The most immature thymocytes are CD44+CD25- (DN1) (Figure
2). These cells still can be committed to the NK, DC or B cell lineage. In presence of
Notch-1 signalling, DN1 thymocytes up-regulate CD25 to become CD44+CD25+
(DN2) (Figure 2). DN2 thymocytes migrate to the subcapsular region of the thymus
and show an IL-7-dependent proliferation (Rodewald et al., 1997). DN2 cells still can
develop to DC but not any more to NK or B cells (Benoist and Mathis, 1999) (Figure
2). After down-regulating CD44, CD44loCD25+ DN3 thymocytes stop proliferating.
From this developmental stage on, all cells are committed to the T cell lineage
(Benoist and Mathis, 1999). DN3 thymocytes start to rearrange β-, γ- and δ-TCR
chain genes (Figure 2). A productive γδ-chain gene rearrangement followed by
expression of a functional γδTCR leads to commitment to the γδ T cell lineage
(MacDonald et al., 2001) (Figure 2). A productive β-chain gene rearrangement
followed by successful pairing with the invariant pre-TCRα chain (pTα) forming the
pre-TCR commits cells to the αβ T cell lineage (MacDonald et al., 2001). Further
thymocyte survival and differentiation along the appropriate lineage is dependent on
signals from the correctly assembled CD3+ γδTCR or CD3+ αβTCR (Starr et al.,
2002).
pTα is a type 1 transmembrane protein with a single immunoglobulin-like
extracellular domain (Saint-Ruf et al., 1994). Pre-TCR signalling in contrast to αβTCR
or γδTCR signalling occurs in a cell-autonomous or constitutive manner (Saint-Ruf et
al., 2000). Productive pre-TCR signalling - might occur in the absence of any
extrathymic ligand (Irving et al., 1998) - leads to thymocyte survival and down-
regulation of CD25 (DN4 stage) (Figure 2). CD44-CD25-pre-TCRlo DN4 thymocytes
express already some CD3 subunits. DN4 thymocytes still can develop to γδ T cells
(Petrie et al., 1992). However, it is not known whether these cells down-regulated
CD25 before or after commitment to the γδ T cell lineage and whether they are

Introduction
- 17 -
exactly the same CD44-CD25- DN4 thymocytes that develop later on to αβ T cells
(Benoist and Mathis, 1999) (Figure 2). DN4 thymocytes show a massive increase in
cell size and proliferation, allelic exclusion at the TCRβ locus and CD4 and CD8 co-
receptors up-regulation (Benoist and Mathis, 1999; Starr et al., 2002). Some α-chain
gene rearrangements are observed (Benoist and Mathis, 1999; Starr et al., 2002)
(Figure 2). Between DN4 and double-positive (DP) stage, thymocytes express low
levels of CD8 representing immature single positive (ISP) thymocytes. This
developmental stage cannot be seen in vitro (Benoist and Mathis, 1999) (Figure 2).
CD4+CD8+pre-TCRlo DP thymocytes show strongly up-regulated Rag gene
expression resulting in α-chain gene rearrangements. Productive α-chain gene
rearrangement does not prevent further Vα-Jα rearrangement within the same cell,
there is no feedback inhibition as seen in productively rearranged TCRβ- ,
immunoglobulin (Ig) heavy (H)- and Ig light (L)-chains (Borgulya et al., 1992; von
Boehmer, 1990). Whereas during B cell development, most properly assembled H/L
chain pairs trigger allelic exclusion and further differentiation (Casellas et al., 2001;
Nemazee, 2000), during T cell development TCRαβ assembly itself is not enough to
promote further differentiation and Rag gene repression (Starr et al., 2002). TCRα-
chain gene rearrangement stops only then when the newly rearranged TCRαβ has
been positively selected (Borgulya et al., 1992; Borowski et al., 2002; Brandle et al.,
1992). One third of mature T cells harbour two productively rearranged TCRα alleles
(Casanova et al., 1991), however, less than 30% of mature T cells express actually
two different TCRαβ complexes due to strong competition for binding to TCRβ-chain
(Heath et al., 1995; Kisielow et al., 1988b).
The development of DN thymocytes to DP thymocytes takes about 14 days and
comprises around 3% of the whole thymocyte fraction (Benoist and Mathis, 1999)
(Figure 2). DP thymocytes are located in the cortical region of the thymus (Benoist
and Mathis, 1999). With continuous maturation, large and cycling DP thymocytes
(CD4+CD8+pre-TCRlo) stop cycling, return to small cell size, up-regulate Rag gene
expression and rearrange α-chain genes. Productively rearranged α-chain
assembles with the already expressed TCRβ -chain to form TCRαβ
(CD4+CD8+TCRαβlo) (Figure 2). The TCRαβ of these DP thymocytes is subjected to
a selection process to allow only survival of DP thymocytes, which express a self-
MHC-restricted and self-tolerant TCR. DP thymocytes expressing a TCRαβ not able

Introduction
- 18 -
to recognize self-MHC are eliminated by glucocorticoid-induced apoptosis (death by
neglect) (Surh and Sprent, 1994). DP thymocytes expressing a TCRαβ, which
recognizes self-peptide/MHC complexes with too high avidity and therefore be
potentially auto-reactive, get eliminated by TCR-induced apoptosis (death by
negative selection) (Benoist and Mathis, 1999). During this selection process over
95% of DP thymocytes get deleted (see section 4.2.2) (Figure 2). Only positively
selected DP thymocytes (CD4+CD8+TCRαβhi) (Figure 2) are able to produce a signal
that leads to Rag-gene repression and therefore stop of further α-chain gene
rearrangement, long-term survival, migration into medullary region of the thymus and
further differentiation into mature T cells (Wilkinson et al., 1995). Thymocytes remain
3-4 days in the DP stage and comprise about 80% of the whole thymocyte fraction
(Benoist and Mathis, 1999) (Figure 2).
The last step of thymocyte maturation involves commitment to the CD4- or CD8 T
cell lineage. Positively selected DP thymocytes down-regulate one of either co-
receptors to become unreactive CD4 single positive (CD4-SP) or CD8-SP T cells
(TCR/CD3intHSAhiPNArhi) (Figure 2). The exact mechanisms involved during this
maturation step are not fully known yet. The three current models for CD4/CD8
lineage commitment are discussed in a later section (see section 4.2.3). SP
thymocytes remain 7-14 days in the medullary region and comprise about 15% of the
total thymocyte fraction (Benoist and Mathis, 1999) (Figure 2). They are exported
from the thymus as fully mature and competent SP cells (TCR/CD3hiHSAloPNArlo) via
blood and lymph vessels at the corticomedullary junction (Chaffin and Perlmutter,
1991) (Figure 2).
4.2.1 Commitment to the ααααββββ or γγγγδδδδ T cell lineage
Parameters involved for commitment to either αβ or γδ T cell lineage are not
completely known yet. The current favoured model is the competitive model (Benoist
and Mathis, 1999). This model predicts that αβ and γδ T cells derive from a common
precursor. During a certain period of thymocyte differentiation (mostly between the
DN3 and DN4 stage) until now unknown factor(s) will favour commitment to either αβ
or γδ T cell lineage. β-, γ- and δ-chain gene rearrangement begins at the DN3 stage.
Thymocytes with a productive γ- and δ-chain gene rearrangement followed by
expression of a functional γδTCR will develop along the γδ T cell lineage. Thymocytes

Introduction
- 19 -
with a productive β-chain gene rearrangement and successful pairing with pTα
before productive γδ-chain gene rearrangement and γδTCR expression will develop
along the αβ T cell lineage. Definitive commitment to the αβ T cell lineage, however,
is only given after α-chain gene rearrangement on both alleles followed by excision
of the δ-chain locus. Commitment to the αβ T cell lineage is probably the most
frequent because it only implies one productive β-chain gene rearrangement versus
two productive γ- and δ-chain gene rearrangements for the γδ pathway.
4.2.2 T cell repertoire selection
DP thymocytes expressing low levels of the newly rearranged TCRαβ
(CD4+CD8+TCRαβlo) go through a very strict selection process to ensure survival of
exclusively self-MHC-restricted and self-tolerant T cells. Over 95% of these DP
thymocytes will die during this selection process. Positive selection is defined as the
rescue of self-MHC-restricted DP thymocytes from programmed cell death (Sebzda
et al., 1999); negative selection is defined as deletion of potentially auto-reactive DP
thymocytes by TCR-induced apoptosis (Benoist and Mathis, 1999); DP thymocytes
which do not recognize the self-peptide/MHC complex are neglected and die by
glucocorticoid-induced apoptosis (Benoist and Mathis, 1999). The current model to
explain T cell repertoire selection on a molecular level is the differential TCR
signalling model (Figure 3) (Starr et al., 2002). DP thymocytes with an overall weak
to intermediate avidity interaction between their TCRαβ and self-peptide/MHC
complexes will receive a survival signal whereas DP thymocytes with an overall
strong avidity interaction between their TCRαβ and self-peptide/MHC complexes will
receive a signal to undergo programmed cell death (Figure 3) (Starr et al., 2002). The
overall avidity interaction between TCR and self-peptide/MHC complexes includes
the intrinsic affinity of the TCR (and its co-receptor) for self-peptide/MHC complex,
expression levels of TCR, co-receptor, MHC molecules and adhesion molecules,
peptide concentration, and duration of the contact between the TCR and self-
peptide/MHC complex (Ashton-Rickardt et al., 1994; Sebzda et al., 1999).
With continuous differentiation, DP thymocytes increase their TCR expression levels.
Potentially auto-reactive thymocytes, which at the DP-TCRαβlo developmental stage
still get a survival signal after contact with self-peptide/MHC, will receive at the DP-
TCRαβhi stage a stronger signal and therefore get negatively selected. The increase

Introduction
- 20 -
in TCR-expression levels therefore increases the safety net that prevents mature T
cell auto-reactivity (Sebzda et al., 1999).
Figure 3: Differential TCR Signalling Model. (A) Positive Selection. A weak to intermediate overall avidity
interaction between TCR and self-peptide/MHC complex stimulates the TCR, which transmits a signal via the
TCRα-CPM, CD3γ-ITAM and CD3δ activating ZAP-70, which will partially phosphorylate LAT. Partial LAT
phosphorylation recruits PLC-γ1 and Gads, Slp7, Itk, resulting in activation of PLC-γ1 and production of DAG and
IP3. RasGRP is turned on by DAG and stimulates sustained, but low-level ERK-activation leading to thymocyte
survival. NF-AT and NF-κB-activation leads to thymocyte proliferation and differentiation. (B) Negative selection.
A high overall avidity interaction between TCR and self-peptide/MHC complex results in fully phosphorylated LAT,

Introduction
- 21 -
recruitment of Grb2/Sos1, p38- and JNK-activation, and transient, but strong ERK-activation leading to thymocyte
apoptosis. Adapted from (Starr et al., 2002).
4.2.3 CD4/CD8 T cell lineage commitment
During the last step of thymocyte development, positively selected DP thymocytes
(CD4+CD8+TCRαβhi) will down-regulate either one of their co-receptors to exit the
thymus as mature single-positive T cells. Three models – “instructive model”,
“stochastic model” and “co-receptor reversal” or “Singer instructive model” – are
currently present to explain CD4/CD8 T cell lineage commitment (Borowski et al.,
2002; Singer, 2002). The first model, “instructive model”, predicts that the co-receptor
instructs the final phenotype of mature T cells. Recognition of the appropriate self-
peptide/MHC complex by the TCR allows co-ligation of the appropriate co-receptor,
which induces a co-receptor signal instructing the DP thymocyte to terminate
expression of the inappropriate co-receptor. The second model, “stochastic model”,
predicts that the DP thymocyte stochastically terminates expression of one co-
receptor. Only thymocytes expressing the co-receptor capable of binding the MHC
molecule for which their TCR is restricted to, will receive a survival signal. In the
latest model, “co-receptor reversal” or “Singer-instructive model”, recognition of the
appropriate MHC molecule by the TCR is followed by a pre-programmed CD8 down-
regulation (Brugnera et al., 2000). Interaction between MHC I-restricted TCR and
MHC I molecule results in a weak TCR-signal. In presence of IL-7, co-receptor
reversal leads to CD4-silencing, CD8 up-regulation and final maturation to the CD8-
SP stage. Interaction between MHC II-restricted TCR and MHC II molecule results in
a strong TCR-signal. Cells are now refractory to co-receptor reversal and develop
therefore to CD4-SP cells (Brugnera et al., 2000).
4.2.4 NK / γγγγδδδδ / CD8αααααααα+ / CD4+CD25+ T cells
The first γδ T cells appear at embryonic day (ED) 14 (Benoist and Mathis, 1999).
Between ED14 and ED17 γδ T cells expressing Vγ5+Vδ1+ TCR predominate. They
are defined as dendritic epidermal cells (DEC) due to their dendritic morphology and
location in the epidermis. DEC seem to be selectively activated by a product of
“stressed” keratinocytes leading to secretion of Th1-like inflammatory cytokines as
well as lymphotactin resulting in recruitment of conventional lymphocytes (Benoist
and Mathis, 1999). In addition, they support the growth of epithelial tissues by
secreting epithelial growth factors, especially during wound healing (Benoist and

Introduction
- 22 -
Mathis, 1999). Vγ6+ T cells are the second wave of γδ T cells. They are produced
between ED17 and birth and migrate to the reproductive tract and tongue where they
reside as intramucosal γδ T cells (Benoist and Mathis, 1999). After birth, a more
heterogeneous population of γδ T cells is produced. These cells populate the thymus,
gut, spleen and other secondary lymphoid organs. Comparable to conventional αβ T
cells, γδ T cells show cytotoxicity, provide B cell help via CD40-CD40L interaction,
and activate macrophages via IFN-γ secretion (Benoist and Mathis, 1999). In
addition, they are able to respond to microbiologic infections (Hiromatsu et al., 1992;
Ladel et al., 1995; Mombaerts et al., 1993) and to recognize non-peptidic antigens
(Constant et al., 1994; Pfeffer et al., 1990; Tanaka et al., 1994). They can even
directly recognize antigen without processing and presentation by MHC-like
molecules (Schild et al., 1994; Weintraub et al., 1994).
Natural Killer (NK) T cells, CD8αα+ intestinal intraepithelial lymphocytes (IEL) and
CD4+CD25+ regulatory T cells are produced during T cell development in the thymus.
They all show an activated phenotype, seem to exert regulatory functions and to
require a high-affinity interaction with self-antigen for proper development (Bendelac
et al., 1996; Capone et al., 2001; Curnow et al., 1995; Hayday et al., 2001; Levelt et
al., 1999; Rocha et al., 1992; Starr et al., 2002; Sydora et al., 1993).
NK T cells have a thymic precursor (DN TCRαβ+ NK1.1+), express NK1.1, are
predominantly Vα14+ and selected on the non-classical MHC I molecule CD1
(Bendelac et al., 1997; Benlagha et al., 2002; Guy-Grand et al., 2003). The natural
ligand is unknown. They regulate conventional T cell responses through cytokine
secretion (Bendelac et al., 2001).
CD8αα+ TCRαβ+ IEL are predominantly found in the gut epithelium (Starr et al.,
2002). Just recently it was suggested that DN TCRαβ+ NK1.1- thymocytes are the
thymic precursors of CD8αα+ IEL (Guy-Grand et al., 2003). These precursors reach
the gut epithelium via mesenteric lymph nodes and the thoracic duct lymph where
they up-regulate CD8αα-expression (Guy-Grand et al., 2003). CD8αα+ TCRαβ+ IEL
express classical or non-classical MHC I-restricted TCR, use both ζ- and γFcεRI-
chains as CD3-associated signal transmitting module, express Ly49 NK receptors
and display NK cytotoxic abilities (Guy-Grand et al., 2003; Guy-Grand et al., 1996;
Guy-Grand et al., 1994; Park et al., 1995; Starr et al., 2002).

Introduction
- 23 -
CD4+CD25+ regulatory T cells are able to prevent the development of gastritis, colitis
and diabetes in vivo and to inhibit T cell proliferation in vitro (Read and Powrie, 2001;
Starr et al., 2002). Whereas development of CD4+CD25+ regulatory T cells is directed
by TEC, BM-derived antigen presenting cells (APC) in the thymus direct development
of CD4+CD25- regulatory T cells (Apostolou et al., 2002; Jordan et al., 2001). In
contrast to conventional αβ T cells, which get negatively selected (clonal deletion)
after a high overall avidity interaction between their TCR and self-peptide/MHC
complexes, regulatory T cells require a high-affinity interaction with self-antigen for
their development. Differences in avidity and the type of APC presenting the self-
peptide/MHC complex might decide if clonal deletion (central tolerance) and/or
selection of regulatory T cells occurs during thymocyte development (Apostolou et
al., 2002; Bensinger et al., 2001; Jordan et al., 2001; Jordan et al., 2000; Modigliani
et al., 1996; Read and Powrie, 2001). Regulatory T cells present in the periphery
could inhibit auto-reactive T cells having escaped clonal deletion in the thymus and
therefore increase even further the safety net to prevent mature T cell auto-reactivity.
4.3 H-Y-specific TCR transgenic mice
H-Y-specific T cell receptor (TCR) transgenic (tg) mice express a transgenic TCRαβ
(Vα3+Vβ8.2+) specific for a male antigen-derived peptide (H-Y) presented on MHC
class I H-2Db molecules (Kisielow et al., 1988a). The minor histocompatibility (H)
male specific (Y) antigen (H-Y) (KCSRNRQYL), which is expressed in all male
tissues, is derived from a protein encoded by the SMCY gene located on the Y
chromosome (Simpson et al., 1997). The transgenic TCRαβ can be followed by flow
cytometry analysis using transgenic TCRα- and TCRβ-specific antibodies (T3.70
and Vβ8.1/2, respectively) (Teh et al., 1989).
In males, presence of the self-peptide H-Y together with the TCR-restricting MHC
molecule H-2Db leads to negative selection in the thymus of H-Y-specific TCR
transgenic CD8+ T cells (Table 1 and Figure 4, first row) (Kisielow et al., 1988a).
CD4-CD8- tg TCRαβhi cells which were shown to be aberrant γδ-lineage T cells, and
cells showing a down-regulation of their co-receptor and transgenic TCR, resulting in
CD8αβlo tg TCRαβ lo-hi cells, are also present in TCR tg male mice (Table 1 and
Figure 4, first row) (Bruno et al., 1996). These double negative (DN) tg TCRαβhi and
CD8αβlo tg TCRαβlo-hi cells are not H-Y-reactive anymore, can therefore escape

Introduction
- 24 -
negative selection and migrate into the periphery as mature self-MHC-restricted and
self-tolerant T cells. These cells do not recognize H-Y/H-2Db complex, do not
proliferate after contact with male stimulator cells but are functional against other
antigens (Kisielow et al., 1988a; Teh et al., 1989). In females, absence of the peptide
H-Y together with the presence of the TCR-restricting MHC molecule H-2Db leads to
positive selection in the thymus of H-Y-specific CD8αβhi tg TCRαβhi cells (Table 1
and Figure 4, second row) (Kisielow et al., 1988a). Because the tg TCRαβ is already
expressed at the DN stage during thymocyte maturation, few DN tg TCRαβ+ cells are
also found in the periphery of TCR transgenic female mice representing the aberrant
γδ-lineage cells (data not shown) (Bruno et al., 1996). However, CD8αβlo tg TCRαβlo-
hi cells as seen in TCR transgenic males are not found in TCR transgenic females
(Figure 4, second row). TCR transgenic mice, which do not express the TCR-
restricting MHC molecule H-2Db do not show positive selection of H-Y-specific
CD8αβhi tg TCRαβhi cells (Table 1 and Figure 4, third row) (Teh et al., 1988; von
Boehmer, 1990). Peripheral DN tg TCRαβ+ and CD8αβlo tg TCRαβ lo-hi cells are
absent in TCR transgenic mice on a non-selecting background (Figure 4 and data not
shown). H-Y-specific CD8αβhi tg TCRαβhi cells escape death from neglect via
endogenous rearrangement of TCRα-chain genes (Table 1 and Figure 4, third row
and data not shown) (Teh et al., 1988). In summary, positive selection of H-Y-specific
TCR transgenic CD8+ cells requires presence of TCR-restricting MHC class I
molecule H-2Db and absence of self-peptide H-Y. Transgenic TCRαβ does not get
positively selected by the presence of MHC class II molecules of the selecting H-2b
haplotype nor by the presence of MHC class I molecules of H-2b, d or k haplotype
(Table 1 and Figure 4 and data not shown) (Kisielow et al., 1988b; Teh et al., 1988;
von Boehmer, 1990).
Introduction
- 25 -
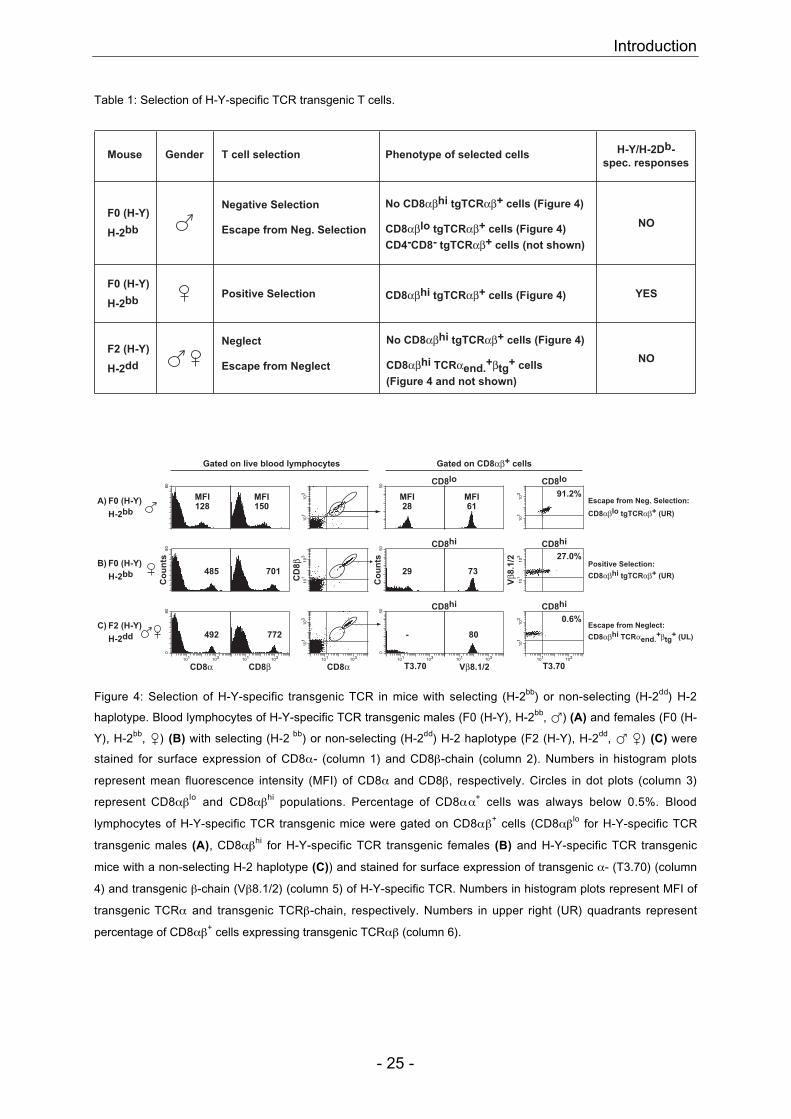
Table 1: Selection of H-Y-specific TCR transgenic T cells.
Mouse T cell selection
Positive Selection
Phenotype of selected cells
CD8αβhi tgTCRαβ+ cells (Figure 4)
H-Y/H-2Db-
spec. responses
YES
Negative Selection No CD8αβhi tgTCRαβ+ cells (Figure 4)
Escape from Neg. Selection
CD4-CD8- tgTCRαβ+ cells (not shown)
NOCD8αβlo tgTCRαβ+ cells (Figure 4)
Gender
F2 (H-Y)
H-2dd
F0 (H-Y)
H-2bb
F0 (H-Y)
H-2bb
No CD8αβhi tgTCRαβ+ cells (Figure 4)Neglect
NOEscape from Neglect
(Figure 4 and not shown)
CD8αβhi TCRαend.+βtg
+ cells
Gated on live blood lymphocytes Gated on CD8αβ+ cells
50
101
103
80
101
103 91.2%
A)
H-2bb
F0 (H-Y)
73701
50
29
Co
un
ts
101
103 27.0%
Vβ8
.1/2
CD
8β
101
103
Co
un
ts80
485B)
H-2bb
F0 (H-Y)
CD8α CD8αCD8β T3.70 T3.70Vβ8.1/2101 103
80
050
101 103
-
101 103
772
101
103
101 103
0.6%
101
103
101 103
080
101 103
492C)
H-2dd
F2 (H-Y)
CD8lo CD8lo
CD8hi
CD8hiCD8hi
CD8hi
128MFI
61MFI
28MFI
150MFI Escape from Neg. Selection:
CD8αβlo tgTCRαβ+ (UR)
Positive Selection:
CD8αβhi tgTCRαβ+ (UR)
Escape from Neglect:
CD8αβhi TCRαend.+βtg
+ (UL)
Figure 4: Selection of H-Y-specific transgenic TCR in mice with selecting (H-2bb) or non-selecting (H-2dd) H-2
haplotype. Blood lymphocytes of H-Y-specific TCR transgenic males (F0 (H-Y), H-2bb, �) (A) and females (F0 (H-
Y), H-2bb, �) (B) with selecting (H-2 bb) or non-selecting (H-2dd) H-2 haplotype (F2 (H-Y), H-2dd, � �) (C) were
stained for surface expression of CD8α- (column 1) and CD8β-chain (column 2). Numbers in histogram plots
represent mean fluorescence intensity (MFI) of CD8α and CD8β, respectively. Circles in dot plots (column 3)
represent CD8αβlo and CD8αβhi populations. Percentage of CD8αα+ cells was always below 0.5%. Blood
lymphocytes of H-Y-specific TCR transgenic mice were gated on CD8αβ+ cells (CD8αβlo for H-Y-specific TCR
transgenic males (A), CD8αβhi for H-Y-specific TCR transgenic females (B) and H-Y-specific TCR transgenic
mice with a non-selecting H-2 haplotype (C)) and stained for surface expression of transgenic α- (T3.70) (column
4) and transgenic β-chain (Vβ8.1/2) (column 5) of H-Y-specific TCR. Numbers in histogram plots represent MFI of
transgenic TCRα and transgenic TCRβ-chain, respectively. Numbers in upper right (UR) quadrants represent
percentage of CD8αβ+ cells expressing transgenic TCRαβ (column 6).

Introduction
- 26 -
4.4 Central Question
The key question of this thesis is which parameters are involved in selection and
survival of a functional and mature T cell repertoire. The first part of the Results
section addressed the role of thymic epithelial (TE) versus non-TE MHC in T cell
repertoire selection (Results Part I). The second part analysed the influence of TCR-
restricting MHC density on selection and survival of the above already described low-
affinity H-Y-specific transgenic TCR (Results Part II). The last part of the Results
section compared intra- and extrathymic selection of the H-Y-specific transgenic TCR
(Results Part III). The aim of this study and all performed experiments was to obtain
at the end a better understanding of the different requirements for efficient T cell
repertoire selection so that in a near future we might be able to improve therapies
against autoimmune diseases and for successful transplantations.

Results Part I
- 27 -
5 Results Part I
Efficient T cell repertoire selection in tetraparental chimeric mice
independent of thymic epithelial MHC
Marianne M. Martinic*¶||, Thomas Rülicke†||, Alana Althage*,**, Bernhard
Odermatt‡, Matthias Höchli§, Alain Lamarre*††, Tilman Dumrese*, Daniel E.
Speiser*‡‡, Diego Kyburz*§§, Hans Hengartner* and Rolf M. Zinkernagel*¶
*Institute of Experimental Immunology, Department of Pathology, University Hospital Zurich,
Schmelzbergstrasse 12, CH-8091 Zurich, Switzerland†Institute of Laboratory Animal Science, Central Biological Laboratory, University Hospital
Zurich, Sternwartstrasse 6, CH-8091 Zurich, Switzerland‡Laboratory of Molecular Diagnostics, Department of Pathology, University Hospital Zurich,
Schmelzbergstrasse 12, CH-8091 Zurich, Switzerland§Laboratory of Electron Microscopy, University of Zurich, Gloriastrasse 30, CH-8028 Zurich,
Switzerland¶To whom reprint requests should be addressed. Phone: +41-1-255 29 89, Fax: +41-1-255
44 20, E-mail: [email protected], [email protected].||These authors contributed equally to this work.**Present address: Department of Molecular and Experimental Medicine, The Scripps
Research Institute, 10550 North Torrey Pines Rd., 92037 La Jolla, CA††Present address: Institut National de la Recherche Scientifique-Institut Armand-Frappier,
531 Boul. des Prairies, H7V 1B7 Laval, Quebec, Canada‡‡Present address: Ludwig Institute for Cancer Research / CHUV, Division of Clinical Onco-
Immunology, Hôpital Orthopédique, Av. Pierre-Decker 4, CH-1005 Lausanne, Switzerland§§Present address: Rheumaklinik und Institut für Physikalische Medizin, University Hospital
Zurich, Gloriastrasse 25, CH-8091 Zurich, Switzerland
Abbreviations: LCMV, lymphocytic choriomeningitis virus; LCMV-GP, LCMV glycoprotein;
LCMV-NP, LCMV nucleoprotein; VSV, vesicular stomatitis virus; CTL, cytotoxic T
lymphocyte; pfu, plaque-forming units.

Results Part I
- 28 -
ABSTRACT
Non-thymic epithelial cells were compared to thymic epithelial cells for their
role in T cell repertoire selection. Tetraparental aggregation chimeras were
generated from T- and B cell-deficient mice (H-2d Scid or H-2b Rag-/-) and
thymus-deficient nude mice (H-2b or H-2d). These tetraparental mice showed
primary protective CD8+ T cell responses after lymphocytic choriomeningitis
virus (LCMV) infection, which were restricted to either thymic or non-thymic
epithelial MHC at comparable levels. These chimeras also mounted
neutralizing IgG responses dependent upon cognate CD4+ T helper cell activity
restricted to non-thymic epithelial MHC. Therefore, in contrast to earlier results
with irradiation or thymus chimeras, these relatively undisturbed tetraparental
mice reveal that the MHC of non-thymic epithelial cells efficiently selects a
functional T cell repertoire.
It is well established that the thymus is essential for T cell receptor rearrangement
and T cell maturation (Miller, 1961). It is also widely accepted that the MHC of radio-
resistant cells of the thymus – presumably thymic epithelial cells – selects the T cell
repertoire. This conclusion is based on a series of classical irradiation bone marrow
and thymus chimera experiments (reviewed in Moller, 1978; von Boehmer, 1990).
Several groups have shown that (H-2a x H-2b) F1-bone marrow cells reconstituting
lethally irradiated parental (H-2a)-mice generate H-2a-restricted but virtually no H-2b-
restricted virus-specific cytotoxic T lymphocytes (CTL) in a primary immune response
(Moller, 1978; von Boehmer, 1990). Although this view has since been accepted by
most immunologists and in textbooks, exceptions have been reported (Doherty and
Bennink, 1979; Longo and Schwartz, 1980; Matzinger and Mirkwood, 1978; Wagner
et al., 1980). However, examples of such non-thymic epithelial MHC-restricted T cells
have been rare and usually reflected weak responses that needed priming and
several rounds of in vitro restimulation before they could be measured. Surprisingly,
experiments with nude mice reconstituted with a completely allogeneic d14 fetal
thymus graft demonstrated that the T cell repertoire was almost exclusively specific
for the recipient MHC haplotype (Zinkernagel et al., 1980).
Therefore, to clarify the respective roles of the MHC of thymic epithelial versus non-
thymic epithelial cells in T cell repertoire selection, we have generated tetraparental
aggregation chimeras from T- and B-cell deficient mice (H-2d Scid or H-2b Rag-/-)

Results Part I
- 29 -
and thymus-deficient nude mice (H-2b or H-2d). In the resulting Scidd � nudeb
tetraparental mice, thymic epithelial cells are exclusively of Scid H-2d haplotype, and
T and B cells exclusively of nude H-2b haplotype. In the Rag-/-b ↔ nuded
tetraparental chimeras, thymic epithelial cells are exclusively of Rag-/- H-2b
haplotype, and T and B cells exclusively of nude H-2d haplotype. These relatively
undisturbed (i.e. non-irradiated and non-reconstituted) and well mixed tetraparental
adult chimeras allow a unique opportunity to study whether during a primary immune
response the T cell repertoire is restricted exclusively to the MHC of thymic epithelial
cells or to both parental haplotypes. Moreover, we analyzed whether these mice
could clear LCMV as efficiently as control mice and whether a protective CD4+ T
helper cell-dependent antibody response was present against LCMV or vesicular
stomatitis virus (VSV). Since B cells in these tetraparental mice express non-thymic
epithelial MHC, a protective CD4+ T helper cell-dependent antibody response would
be direct evidence for the presence of functional effector T cells restricted to non-
thymic epithelial MHC.
MATERIALS AND METHODS
Mice. C57BL/6 (H-2b), BALB/c (H-2d), C57BL/6-Rag1-/- or -Rag2-/- (Rag-/-b) were
obtained from the Institute of Laboratory Animal Science, University of Zurich,
Switzerland. C57BL/6-nude (nudeb) and BALB/c-nude (nuded) were purchased from
Biological Research Laboratories, Füllinsdorf, Switzerland. C.B-17-Scid (Scidd) were
purchased from IFFA CREDO, L'Arbresle, France. All mice were kept under specific
pathogen-free conditions.
Aggregation Chimeras. Mouse chimeras were generated by aggregation of 8-cell
embryos recovered from genetically immunodeficient mutants: Nuded and Scidd (both
albino, Gpi-1a, H-2d), nudeb and Rag-/-b (all black, Gpi-1b, H-2b). Embryos of each
strain were obtained by mating homozygous unreconstituted parents. Females were
super ovulated according to standard procedures with 5 iU PMSG/hCG and
additionally stimulated by the Whitten effect to improve their response. On day 3 of
gestation the embryos were flushed from the oviduct and their zona pellucida was
removed by brief incubation in pronase solution. After washing, embryos were
transferred in drops of M16 culture medium under liquid paraffin. Double embryo
aggregates of the following combinations were produced by gently pushing two
uncompacted morulae together: Scidd ↔ nudeb and Rag-/-b ↔ nuded. After 30 h of

Results Part I
- 30 -
incubation at 37°C and 5% CO2 in air, most aggregates formed early blastocysts.
Morphologically normal embryos were transferred into the uteri of pseudopregnant
histocompatible CB6F1 surrogate foster mothers under SPF conditions. Offspring
were born after 18 days of gestation and chimeras were recognized by the presence
of albino and pigmented skin patches a few days later. Additionally, several tissues
were tested for chimerism by GPI (glucose-6-phosphate isomerase)-isoenzyme gel
electrophoresis (Eppig et al., 1977). The contributions from both parental strains
were approximately equal, indicating that there was no strong strain-specific selective
advantage during embryonic development.
Cell Lines, ELISA, 51Cr-Release Assay, VSV-IND neutralization assay, and
Virus. EL-4 (H-2b) and P815 (H-2d) cells were obtained from the American Type
Culture Collection (Rockville, Maryland, USA). The LCMV nucleoprotein-specific
enzyme-linked immunosorbent assay (ELISA), the 51Cr-release assay, the VSV-IND
(VSV Indiana serotype) neutralization assay, LCMV-WE (LCMV strain WE) and VSV-
IND have been previously described (Bachmann et al., 1993; Battegay et al., 1993;
Charan and Zinkernagel, 1986; Kyburz et al., 1993; McCaren et al., 1959; Speiser et
al., 1992).
Immunohistology. Thymi were immersed in HBSS, snap-frozen, and 5-µm cryostat
sections were cut and fixed in acetone for 10 min. Sections were incubated with the
following antibodies: rat monoclonals against murine CD4 (YTS 191), CD8 (YTS
169), CD45R/B220 (RA3-6B2) and CD11b (M1/70), biotinylated mouse monoclonal
antibodies against murine MHC class I H-2Kb (AF6-88.5), H-2Kd (SF1-1.1), MHC
class II IAb (AF6-120.1) and IAd (AMS-32.1), followed by incubation with streptavidin-
alkaline phosphatase (all from Pharmingen). Primary antibodies were detected by
sequential incubation with goat antibodies against species-specific immunoglobulins,
followed by alkaline phosphatase labeled donkey anti-goat antibodies (Jackson
ImmunoResearch). Alkaline phosphatase was visualized using naphthol AS-BI (6-
bromo-2-hydroxy-3-naphtholic acid-2-methoxy anilide) phosphate and new fuchsin as
substrate, yielding a red color reaction product. Endogenous alkaline phosphatase
was blocked by levamisole. Sections were counterstained with hemalum.
Confocal Fluorescence Microscopy. Thymic epithelial cells were stained with a
polyclonal rabbit anti-cytokeratin antiserum (wide spectrum screening; dilution
1:1500; Dako). Primary rabbit antibodies were detected by sequential incubation with
affinity purified, rhodamine labeled goat anti-rabbit Ig antibodies followed by

Results Part I
- 31 -
rhodamine labeled donkey anti-goat Ig antibodies (dilutions 1:100 in TBS containing
5% normal mouse serum; Jackson ImmunoResearch). MHC class II antigens were
revealed with biotinylated mouse anti-IAb antibodies (clone AF6-120.1; dilution 1:200)
or biotinylated mouse anti-IAd antibodies (clone AMS-32.1; dilution 1:60; both from
Pharmingen) and fluorescein streptavidin (dilution 1:200; Dako). The appropriate
primary and secondary reagents were mixed and incubated in three steps of 30 min
each; anti-MHC class II antibodies were added twice. Slides were mounted with
Dako medium. Images were recorded with a confocal laser scanning system TCS-
SP2 (Leica laser technique, Mannheim, Germany) and processed using Openlab
software (Improvision).
Flow Cytometric Analysis. Peripheral blood or splenic cells were stained with the
following antibodies: anti-CD8alpha-APC (53-6.7), anti-CD8b.2-PE (53-5.8), anti-
B220-PE (RA3-6B2), anti-CD11b-PE (M1/70), anti-H-2Dd-FITC (34-2-12), anti-H-2Db-
PE (KH95), anti-H-2Kb-Biotin (AF6-88.5), Streptavidin-PerCP and Streptavidin-APC
(all from Pharmingen). For double tetramer stains, peripheral blood or splenic cells
(7.5x105) were stained with equal amounts of APC-labeled LCMV-WE GP33 tetramer
(GP33-Db) and PE-labeled LCMV-WE NP118 tetramer (NP118-Ld) and incubated for
20 min at 37°C. One microgram of anti-CD8alpha-FITC antibody (53-6.7) was added
to each sample and incubated for another 20 minutes at 4°C. All samples were
acquired on a FACSCalibur and analyzed using CellQuest software (Becton
Dickinson).
RESULTS
Tetraparental aggregation chimeras show a well-mixed chimerism.
Tetraparental aggregation chimeric mice (chimeras) of the following combinations
were generated: Scidd ↔ nudeb (Figure 5A) and Rag-/-b ↔ nuded. All chimeras had a
well-mixed chimerism, shown by the presence of both GPI-isoforms (GPI-1A, GPI-
1B) in sections of spleen, kidney, liver, lung, thymus and heart (Figure 5B and Figure
5C). Blood lymphocytes of Rag-/-b ↔ nuded chimeras were tested for H-2d and H-2b
expression by FACS-analysis. Whereas CD8+ and B220+ cells expressed only the
parental BALB/c-nude H-2d, CD11b+ macrophages were distributed into two
populations, expressing either the parental BALB/c-nude H-2d or the parental Rag-/-
H-2b (Figure 5D). Like CD8+ T cells, CD4+ T cells expressed only H-2d (data not
shown). Equivalent expression patterns were observed with Scidd ↔ nudeb chimeras

Results Part I
- 32 -
(data not shown). Non-mucosal CD8+ T cells in these chimeras were all CD8αβ as
shown by co-staining with CD8α- and CD8β-specific antibodies (data not shown).
GPI-1B
GPI-1A
GPI-1B
GPI-1A
H H HThSp LuLiKi
C57B
L/6
BA
LB
/c-n
ud
e
C57B
L/6
+
BA
LB
/c-n
ud
e
Scidd ↔ nudeb
Rag-/-b ↔ nuded
1 2 3 4 5 6 7 8
1 2 3 4 5 6 7 8
H H HHSp LuLiKi
B
C
DA
101
103
101
103
101
103
101 103101 103
101
103
101 103
(Rag-/-b ↔ nuded)- Chimera #8
(Rag-/-b ↔ nuded)- Chimera #7
BALB/c (H-2d)
C57BL/6 (H-2b)
Blood lymphocytes gated on
CD8+ cells B220+ cells CD11b+ cells
H-2Db H-2Kb
H-2
Dd
Figure 5: Phenotypic analysis of tetraparental chimeric mice. (A) Picture of a six-week-old Scidd ↔ nudeb
chimera. Distribution of GPI (Glucose-6-phosphate isomerase)-isoforms (GPI-1A, GPI-1B) in different tissues (H =
Heart, Sp = Spleen, Ki = Kidney, Li = Liver, Lu = Lung, Th = Thymus) of a Scidd ↔ nudeb chimera (B, lanes 3-7)
and a Rag-/-b ↔ nuded chimera (C, lanes 3-7). As controls, heart preparations of a C57BL/6 (GPI-1B) (B and C,
lane 1), BALB/c-nude (GPI-1A) (B and C, lane 2) and a 50:50-mixture of both (B and C, lane 8) were used. The
experiment was repeated three times with similar results. Blood-FACS analysis of six-to-eight-week-old chimeric
and control mice (D). Blood lymphocytes of naïve C57BL/6, BALB/c and Rag-/-b ↔ nuded chimeras #7 and #8
were gated on CD8+, B220+ or CD11b+ cells and stained with H-2Db-, H-2Kb- and H-2Dd- specific antibodies. The
experiment was repeated five times with similar results.
No rescue of nude thymic rudiments in tetraparental chimeras. Expression of
MHC class I and II, CD4, CD8, B220 and CD11b in chimeric and control thymi was
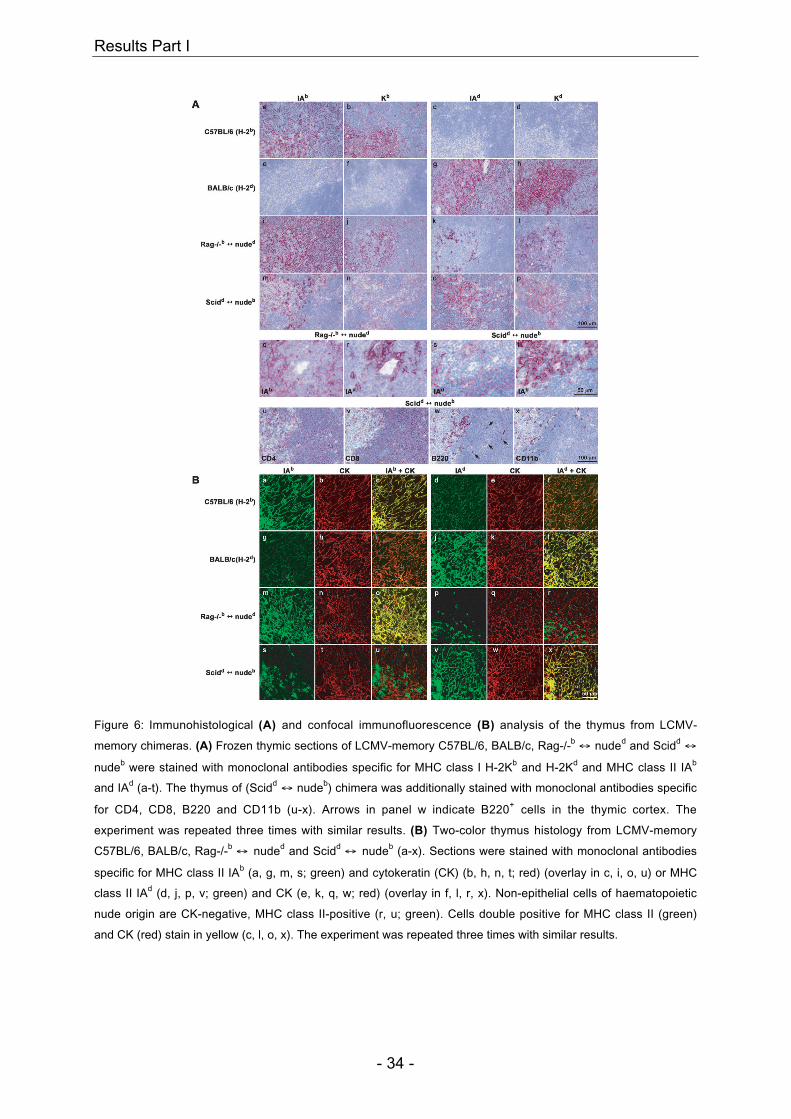
assessed by immunohistological analysis of frozen serial sections (Figure 6A, a-x).
Whereas analysis of MHC class I and II expression of the Rag-/- H-2b or Scid H-2d
haplotype (representing thymic epithelial haplotype) revealed a typical thymic
network (Figure 6A, i-j+q and Figure 6A, o-p+s), MHC class I and II expression
patterns of the nude haplotype revealed the presence of individual cells rather than
this thymic pattern (Figure 6A, k-l+r and Figure 6A, m-n+t). The presence of CD4+,
CD8+, B220+ and CD11b+ cells in Scidd ↔ nudeb chimeras was similar to control
mice (Figure 6A, u-x and data not shown). Some B220+ cells of C57BL/6-nude or
BALB/c-nude origin, respectively, were found in both the thymic medulla and in the
cortex (Figure 6A, w, arrows and data not shown).
In order to exclude eventual rescue of nude thymic rudiments (Blackburn et al., 1996;
Holub et al., 1975; Hsu et al., 1975; Kindred, 1979; Rodewald et al., 2001) and

Results Part I
- 33 -
confirm that thymic epithelial cells in these chimeras express exclusively the Rag-/-
or Scid- but not the nude haplotype, two-color thymus histology was performed
(Figure 6B, a-x). Chimeras and control mice were analyzed for expression of MHC
class II and cytokeratin (CK), the latter being a characteristic marker for epithelial
cells. Sections of chimeras and control mice revealed an intense yellow stain when
thymic epithelial MHC (Rag-/- or Scid-haplotype for chimeras) and cytokeratin stains
were superimposed, showing that both markers coincided on the thymic epithelial
network (Figure 6B, c+o and l+x). When nude MHC and cytokeratin stains were
superimposed, cytokeratin-negative, nude MHC class II-positive cells were found
(Figure 6B, r and u; green). Therefore, these cells must represent non-thymic
epithelial cells of haematopoietic nude origin, which have migrated into the thymus.
In summary, in all chimeras tested we found no evidence that nude thymic rudiments
were rescued as thymic epithelial cells were always of non-nude donor origin.
Results Part I
- 34 -
Figure 6: Immunohistological (A) and confocal immunofluorescence (B) analysis of the thymus from LCMV-
memory chimeras. (A) Frozen thymic sections of LCMV-memory C57BL/6, BALB/c, Rag-/-b ↔ nuded and Scidd ↔
nudeb were stained with monoclonal antibodies specific for MHC class I H-2Kb and H-2Kd and MHC class II IAb
and IAd (a-t). The thymus of (Scidd ↔ nudeb) chimera was additionally stained with monoclonal antibodies specific
for CD4, CD8, B220 and CD11b (u-x). Arrows in panel w indicate B220+ cells in the thymic cortex. The
experiment was repeated three times with similar results. (B) Two-color thymus histology from LCMV-memory
C57BL/6, BALB/c, Rag-/-b ↔ nuded and Scidd ↔ nudeb (a-x). Sections were stained with monoclonal antibodies
specific for MHC class II IAb (a, g, m, s; green) and cytokeratin (CK) (b, h, n, t; red) (overlay in c, i, o, u) or MHC
class II IAd (d, j, p, v; green) and CK (e, k, q, w; red) (overlay in f, l, r, x). Non-epithelial cells of haematopoietic
nude origin are CK-negative, MHC class II-positive (r, u; green). Cells double positive for MHC class II (green)
and CK (red) stain in yellow (c, l, o, x). The experiment was repeated three times with similar results.

Results Part I
- 35 -
Chimeras mount primary protective virus-specific CD8+ T cell responses
restricted to both thymic and non-thymic epithelial MHC. The effector function of
T cells of nude origin maturing in a thymic environment composed of epithelial cells
expressing non-matching MHC molecules was evaluated during an immune
response against LCMV. Chimeras aged 6-8 weeks showing comparable furry/nude
and pigmented/albino skin patches (Figure 5A) were infected intravenously with 200
plaque-forming units (pfu) of LCMV-WE. Eight days later, mice were
hemisplenectomized and cytotoxic CD8+ T cell activity was measured directly ex vivo
in a 5 hour 51Cr-release assay on H-2b (EL-4) and H-2d (P815) target cells prepulsed
with the immunodominant LCMV peptides LCMV-GP33-41 (H-2Db) and LCMV-NP118-
126 (H-2Ld) (Figure 7A) or the subdominant LCMV peptide LCMV-NP396-404 (H-2Db)
(data not shown). The chimeras exhibited strong primary CTL activity specific for all
three peptides tested. The CTL responses observed in chimeras were similar to
those of LCMV-infected C57BL/6 or BALB/c mice (Figure 7A). Chimeras were
efficiently protected against viral infection as indicated by the absence of detectable
virus in spleen and other organs eight days following infection (data not shown).
On day 36 after infection, lymph node cells from the hemisplenectomized mice were
restimulated in vitro for 5 days with peptide labeled H-2d or H-2b spleen cells (Fig.
3B). The strong CTL activity of chimeric lymph node cells was comparably restricted
to both thymic and non-thymic MHC (Figure 7B). As expected, no alloreactivity
against chimeric MHC was seen (Figure 7B). In contrast, in a standard mixed
lymphocyte culture assay, alloreactivity to third party H-2k was found for control and
chimeric effector cells (data not shown).

Results Part I
- 36 -
1 9 81
0
25
50
75
100
90 30 10 30
25
50
75
100
90 30 10 3
A
B
90 30 10 3 90 30 10 3
1 9 81 1 9 811 9 81
% S
pecif
ic L
ysis
% S
pecif
ic L
ysis
Effector : Target Ratio
Dilution of Standard Culture
Day 8 (directly ex vivo)
Day 36 (2° in vitro)
alloreactivity alloreactivity
Rag-/-b ↔ nuded
Scidd ↔ nudeb
BALB/c (H-2d)
C57BL/6 (H-2b)
EL-4 / ∅ EL-4 / GP33 P815 / ∅ P815 / NP118
EL-4 / ∅ EL-4 / GP33 P815 / ∅ P815 / NP118
(H-2b) (H-2
b) (H-2
d) (H-2
d)
(H-2b) (H-2
b) (H-2
d) (H-2
d)
naive C57BL/6
naive BALB/c
Figure 7: Primary ex vivo and secondary in vitro CTL response of LCMV-infected chimeras. Eight-week-old
C57BL/6 (�), BALB/c (�), Scidd ↔ nudeb (�) and Rag-/-b ↔ nuded (�) were infected intravenously with 200 pfu
of LCMV-WE. (A) On day 8 post infection, mice were hemisplenectomized and single cell suspensions were
tested directly ex vivo for 5h in a standard 51Cr-release assay on LCMV-GP33-loaded (EL-4 / GP33) or control
(EL-4 / ∅) EL-4 targets (H-2b) and on LCMV-NP118-loaded (P815 / NP118) or control (P815 / ∅) P815 targets
(H-2d). (B) On day 36 after infection, mice were sacrificed and pooled lymph node cells were restimulated in vitro
for 5 days with LCMV-GP33-loaded irradiated C57BL/6 splenocytes or LCMV-NP118-loaded irradiated BALB/c
splenocytes as stimulator cells. Cultures were tested for 5h in a standard 51Cr-release assay on LCMV-GP33-
loaded (EL-4 / GP33) or control (EL-4 / ∅) EL-4 targets and on LCMV-NP118-loaded (P815 / NP118) or control
(P815 / ∅) P815 targets. Similar results were obtained using restimulated splenocytes as effectors (data not
shown). Because of alloreactivity, C57BL/6 effectors lysed equally well peptide-loaded and control P815 targets.
The same was true for BALB/c effectors with peptide-loaded or control EL-4 targets. Therefore, results of
C57BL/6, or BALB/c effectors, with NP118-loaded P815 targets, or GP33-loaded EL-4 targets, are omitted for
clarity. The experiment was repeated six times with similar results.
To characterize the CD8+ T cell repertoire in more detail, day 8 and day 13 effector T
cells were also analyzed at the receptor level using tetramer staining (Figure 8 and
data not shown). Chimeras with strong primary CTL activity restricted to each of the
parental H-2 haplotypes always showed two distinct effector T cell populations, which
either bound LCMV-GP33 (H-2Db) tetramer or LCMV-NP118 (H-2Ld) tetramer, but
not both (Figure 8, 2nd-4th column and data not shown). While LCMV-NP118 (H-2Ld)

Results Part I
- 37 -
tetramer binding on day 8 was lower in chimeras than in BALB/c controls, by day 13,
LCMV-NP118 (H-2Ld) tetramer bindings were comparable (data not shown).
C57BL/6 (H-2b)
BALB/c (H-2d)
BALB/c-nude
(H-2d)
(Rag-/-b ↔ nuded)- Chimera #7
(Rag-/-b ↔ nuded)- Chimera #8
Gated on
CD8+ cells
NP
11
8-L
d
GP
33
-Db
Co
un
ts
NP
11
8-L
d
GP33-DbCD8
Gated on living splenocytes
Spleen (d8)
101
103
0.0
0.0
101
103
0.1
23
101
103
8.1
0.1
101
103
101 103
6.1
3.0
101
103 2.5
6.3
08
0
65.6
08
0
101 103
35.5
08
0
30.3
08
0
0.6
08
0
33.0
101
103 0.1
101
103 7.9
101
103 0.1
101
103
101 103
1.2
101
103 0.8
101
103 5.5
101
103 0.0
101
103 0.1
101
103 2.0
101
103
101 103
2.1
Figure 8: Tetramer analysis of LCMV-immune T cells of tetraparental chimeras. Eight-week-old C57BL/6, BALB/c,
BALB/c-nude and Rag-/-b ↔ nuded chimeras #7 and #8 were infected with 200 pfu of LCMV-WE intravenously.
Eight days after infection, mice were hemisplenectomized and 7.5x105 splenocytes were tested for binding to
LCMV-GP33 tetramer (GP33-Db) and LCMV-NP118 tetramer (NP118-Ld). Histogram plots show the percentage
of CD8+ cells amongst living splenocytes (1st column). Dot plots gated on living splenocytes show double staining
with a CD8-specific antibody and GP33-Db (2nd column) or double staining with a CD8-specific antibody and
NP118-Ld (3rd column). Numbers in upper right quadrants represent percentage of tetramer and CD8 double
positive splenocytes. Dot plots gated on CD8+ splenocytes show staining with equal amounts of GP33-Db and
NP118-Ld (4th column). Numbers in upper left (UL) and lower right (LR) quadrants represent percentage of CD8+
splenocytes binding to either NP118-Ld or to GP33-Db, respectively. The experiment was repeated three times
with similar results.
Non-thymic epithelial MHC-restricted CD4+ T cells co-operate efficiently with B
cells and mediate protective IgG responses. As B cells of tetraparental chimeras
express MHC class II of the non-thymic haplotype, co-operation is only possible with
CD4+ T cells being restricted to non-thymic MHC (Table 2). Therefore, to assess
whether CD4+ T cells restricted to non-thymic MHC are present and functional in
these chimeras, VSV- neutralizing IgG or LCMV-NP-specific IgG titers – which have
both been shown previously to be strictly dependent upon cognate MHC class II-
restricted CD4+ T helper cell activity (Leist et al., 1987; Ochsenbein et al., 2000) -
were monitored following VSV or LCMV infection, respectively. VSV-infected
chimeras generated protect ive neutral iz ing IgG responses (

Results Part I
- 38 -
Table 3) and LCMV-infected chimeras generated LCMV-NP-specific IgG responses
(Figure 9) comparable to control mice. These normal B cell responses confirm the
presence of functional CD4+ T cells restricted to non-thymic epithelial MHC.
Table 2: Productive T-B co-operation in tetraparental chimeras is only possible if CD4+ T cells express TCR
restricted to non-thymic MHC of B cells.
Scidd ↔ nudeb
Rag_/_b ↔ nuded
Tetraparental
ChimeraT / B cooperation
Thymic
epithelial
cells
H-2d
H-2b
T cells
B cells
H-2b
H-2d
H-2b-restricted TCR
Thb
TCR
Bb
Peptide
MHC II
H-2d-restricted TCR
Thb
TCR
Bb
Peptide
MHC II
H-2b-restricted TCR
ThThd
TCR
Bd
Peptide
MHC II
H-2d-restricted TCR
ThThd
TCR
Bd
Peptide
MHC II

Results Part I
- 39 -
Table 3: Non-thymic epithelial MHC-restricted CD4+ T cells mediate protective B cell IgG responses in
tetraparental chimeras following intravenous infection with 2x106 pfu VSV-IND.
Mouse*
VSV-IND neutralizing IgG titer** (serum dilution)
Rag-/- (H-2b)
Rag-/-b ↔ nuded
BALB/c (H-2d)
C57BL/6 (H-2b)
<1/40 <1/40
1/80'0001/80'000
1/40'0001/80'000
1/40'0001/40'000
Day 20Day 8
*Rag-/-b ↔ nuded tetraparental chimeras and control mice were infected intravenously with 2x106 pfu of VSV-IND.**VSV-IND-neutralizing IgG titers were monitored on day 8 and day 20 after infection. The highest serum dilution
that neutralizes 50% of input virus is expressed. At least four individual mice were tested in each group. Values
represent averages; variations were always less than one dilution step of two.
10
100
1000
10000
detection limit
An
ti-L
CM
V-N
P I
gG
[Re
cip
roc
al
of
Tit
er
Dil
uti
on
]
d1
4 C
57
BL
/6 (
H-2
b)
d1
4 B
AL
B/c
(H
-2d
)
d1
4 R
ag
-/-b
↔ n
ud
ed
d1
4 S
cid
d ↔
nu
de
b
na
ive
Scid
d ↔
nu
de
b
na
ive
C5
7B
L/6
(H
-2b
)
Figure 9: CD4+ T helper cells co-operate efficiently with B cells of non-thymic H-2 haplotype in LCMV-immune
chimeras. Eight-week-old C57BL/6, BALB/c, Scidd ↔ nudeb and Rag-/-b ↔ nuded were infected with 200 pfu of
LCMV-WE intravenously. On day 14 after infection, serum was prediluted 30-fold and tested for the presence of
LCMV-NP-specific IgG in a standard ELISA. Naïve Scidd ↔ nudeb and naïve C57BL/6 were used as negative
controls and signals were always below detection level. The same was true for non-infected Rag-/-b ↔ nuded-
chimeras or d14 LCMV-infected BALB/c-nude or for C.B-17-Scid or Rag-deficient mice (data not shown). The
experiment was repeated three times with similar results.

Results Part I
- 40 -
DISCUSSION
In summary, these chimeras showed protective virus-specific primary CD8+ T cell
responses restricted to both thymic and non-thymic MHC to comparable levels.
Virus-neutralizing IgG responses – strictly dependent on CD4+ T cell help restricted
to non-thymic epithelial MHC – were generated in these chimeras as efficiently as in
control mice. Taken together, these results demonstrate that cells other than thymic
epithelial cells are efficient in selecting a mature and functional T cell repertoire.
These findings challenge the widely accepted concept which postulates that MHC-
restriction is determined predominantly by the MHC of thymic epithelial cells (thymic
nurse cells) (Wekerle and Ketelsen, 1980) or the radio-resistant portion of the thymus
(reviewed in Moller, 1978; von Boehmer, 1990).
The present study was prompted by data obtained from experiments with nude mice
reconstituted with day 14 fetal thymus grafts from histoincompatible donors
(Zinkernagel and Althage, 1999; Zinkernagel et al., 1980). These nude thymus
chimeras only generated nude MHC- but not thymic MHC-restricted effector T cells.
The conclusion from these earlier studies was either that there was rescue of the
nude thymic rudiment or that cells other than thymic epithelia were efficient in and
essential for positive selection of MHC-restricted T cell specificities. More recently,
tetraparental chimeras between thymus-competent and nude donors of distinct MHC
haplotypes revealed that the thymic rudiment of the nude donor could not be rescued
anatomically in a tetraparental chimeric situation (Blackburn et al., 1996; Rodewald et
al., 2001), as we confirm here (Figure 6). In addition, histological analysis of 20
chimeric thymi showed no evidence of thymic epithelial cells, cysts or other
rudiments of nude origin in well-mixed chimeras.
The discrepancies between the present results and those obtained with F1(AxB) →
A-irradiation bone marrow chimeras and/or F1(AxB) nude grafted with an A-thymus
yielding virtually exclusively A-restricted but not B-restricted T cell responses are
particularly important (Matzinger, 1993; Moller, 1978; Singer, 1988). We believe they
are best explained as follows: it is possible that lethal or supralethal irradiation is not
capable of eliminating all host cells that contribute to T cell receptor interactions
resulting in survival of such T cells regardless of whether they are encountered only
in the thymus or also in the periphery. For example, radio-resistant follicular dendritic
cells in the spleen and lymph nodes, fibroblasts, or other mesenchymal cells would
fulfill such requirements. Also lymphohaemopoietically derived cells, including

Results Part I
- 41 -
macrophages and dendritic cells can probably not be eliminated completely and
current detection limits cannot exclude the presence of 0.5-2% of “contaminating”
cells. Therefore, precursor T cells migrating into the thymus of such F1(AxB) → A-
irradiation bone marrow chimeras will predominantly see A-expressing cells and
therefore will first and preferentially be restricted to A. Those A-restricted T cells will
strongly proliferate in the thymus and leave the thymus as A-restricted T cells. B-
expressing cells from F1(AxB) bone marrow first have to migrate into the thymus
leading to a time disadvantage compared to A-expressing cells already present in the
A-recipient. As the proliferation rate in the thymus is enormous, A-restricted T cells
will have a huge advantage over B-restricted T cells. For example, the numerical
advantage of A-restricted T cells in an A-thymus of an F1(AxB) → A-irradiation
chimera or in an F1(AxB) nude grafted with an A-thymus may readily reach factors of
10-30 (even after a subsequent second depletion of T cells) within 3-5 cell divisions
(corresponding to only 1-3 days!) (Lemischka et al., 1986; Longo and Schwartz,
1980; Zinkernagel, 1982).
In the case of nude mice reconstituted with a fully allogeneic fetal thymus, precursor
T cells migrating into the thymus predominantly encounter thymic epithelial MHC
(Zinkernagel et al., 1980). Only those T cells seeing non-thymic epithelial MHC on
cells from nude origin having migrated into the thymus will be restricted to non-thymic
MHC but will be at a numerical disadvantage compared to the former subset.
However, as soon as these hypothetical T cells would leave the thymus they will
contact cells expressing exclusively non-thymic MHC. Therefore, only T cells
restricted to non-thymic MHC will survive in these chimeras since it is now well
accepted that peripheral amplification and survival of mature T cells are strongly
MHC-dependent (Kirberg et al., 1997; Rocha and von Boehmer, 1991).
The advantage of the chimeras described here over F1(AxB) → A-irradiation bone
marrow chimeras or F1(AxB) nude grafted with an A-thymus is that cells expressing
either thymic epithelial or non-thymic epithelial MHC are present in the thymus and
the periphery in roughly equal numbers from the beginning, i.e. from the time-point
when precursor T cells migrate into the thymus. Therefore, selection and/or MHC-
dependent amplification and survival of a mature T cell repertoire restricted to either
thymic epithelial or non-thymic epithelial MHC is equivalent.
In conclusion, selection of a mature and functional T cell repertoire involves multiple
stages: Thymocyte maturation, followed by positive selection leading to self-MHC-

Results Part I
- 42 -
restricted T cells, and finally the continuous interaction between MHC-restricted
mature T cells and cells expressing the T cell restricting MHC molecules ensuring
their amplification and survival. Whereas thymic epithelium is crucial for T cell
maturation, the results presented here show that the MHC of thymic epithelial cells is
not required for the two latter stages of T cell repertoire selection.
We thank E. Horvath, K. Rappold, K. Osei-Boadum and N. Wey for technical help,
and Drs. A. Macpherson, K. McCoy and M. van den Broek for helpful comments on
the manuscript. This work was supported by the Swiss National Foundation for
Science and the Kanton of Zurich, Switzerland.

Results Part II
- 43 -
6 Results Part II
Influence of MHC class I H-2Db density on selection and survival of
H-Y-specific TCR transgenic T cells
Marianne M. Martinic, Hans Hengartner and Rolf M. Zinkernagel
Institute of Experimental Immunology, Department of Pathology, University Hospital
Zurich, Schmelzbergstrasse 12, CH-8091 Zurich, Switzerland

Results Part II
- 44 -
Abstract
H-Y-specific T cell receptor (TCR) transgenic mice express a transgenic TCR
specific for a male antigen-derived peptide (H-Y) presented on MHC class I H-
2Db molecules. The influence of different parameters on overall positive
selection of these transgenic T cells was analysed. Selection was
independent of age. However, positive selection of H-Y-specific TCR
transgenic T cells in females and escape from negative selection in males
were both strongly dependent on the density of H-2Db molecules on antigen
presenting cells. The higher the density of H-2Db molecules, the higher the
percentage of peripheral CD8+ T cells expressing transgenic TCR in females
and the stronger the down-regulation of the CD8 co-receptor in males. Thus
minor changes in the density of TCR-restricting H-2Db influence selection of
the H-Y-specific TCR.
Introduction
The minor histocompatibility (H) male specific (Y) antigen (H-Y) (KCSRNRQYL),
which is expressed in male tissues, is derived from a protein encoded by the SMCY
gene located on the Y chromosome (Simpson et al., 1997). The H-Y-specific
transgenic (tg) T cell receptor (TCR) binds specifically to the male antigen-derived
peptide (H-Y) presented on MHC class I molecules H-2Db (Kisielow et al., 1988a).
In males, presence of the self-peptide H-Y together with TCR-restricting MHC class
I molecules H-2Db leads to negative selection in the thymus of H-Y-specific CD8αβhi
tg TCRαβ+ cells (Table 4 and Figure 10, first row) (Kisielow et al., 1988a). CD4-
CD8- double negative (DN) tg TCRαβ+ cells which were shown to be aberrant γδ-
lineage T cells, and cells showing a down-regulation of their co-receptor and
transgenic TCR, resulting in CD8αβlo tg TCRαβ+ cells, are also present in TCR
transgenic males (Table 4 and Figure 10, first row and data not shown) (Bruno et
al., 1996). These DN tg TCRαβ+ and CD8αβlo tg TCRαβ+ cells are not measurably
H-Y-reactive anymore and are not negatively selected (Bruno et al., 1996). CD8αβlo
tg TCRαβ+ cells where binding to self-peptide/MHC complex probably is of low
avidity apparently have escaped negative selection in the thymus. They migrate into
the periphery as mature self-MHC-restricted, H-Y non-reactive T cells, do not
proliferate after contact with male stimulator cells but are functional against non H-Y

Results Part II
- 45 -
antigens (Kisielow et al., 1988a; Teh et al., 1989). In females, absence of the
peptide H-Y together with the presence of TCR-restricting MHC class I molecules
H-2Db leads to positive selection in the thymus of H-Y-specific CD8αβhi tg TCRαβ+
cells (Table 4 and Figure 10, second row) (Kisielow et al., 1988a). However, only
maximally 40% of peripheral CD8+ cells express the actual H-Y-specific transgenic
TCR (Figure 10, second row). The remaining CD8+ population have recombined the
endogenous TCRα loci and therefore have changed TCR specificity during
thymocyte selection (Figure 10, second row and data not shown) (Buch et al., 2002;
Huesmann et al., 1991; Merkenschlager et al., 1994). Because the transgenic
TCRαβ is already expressed at the DN stage during thymocyte maturation, few DN
tg TCRαβ+ cells are found also in the periphery of TCR transgenic females
representing the aberrant γδ-lineage cells (data not shown) (Bruno et al., 1996).
CD8αβlo tg TCRαβ+ cells as seen in TCR transgenic males are not found in TCR
transgenic females (Figure 10, second row). H-Y-specific TCR transgenic mice,
which do not express TCR-restricting MHC class I molecules H-2Db do not show
positive selection of H-Y-specific CD8αβhi tg TCRαβ+ cells (Table 4 and Figure 10,
third row) (Fink and McMahan, 2000; Teh et al., 1988; von Boehmer, 1990). These
cells, however, can escape death from neglect via endogenous rearrangement of
TCRα-chain genes (Table 4 and Figure 10, third row and data not shown) (Fink and
McMahan, 2000; Teh et al., 1988; von Boehmer, 1990). Peripheral DN tg TCRαβ+
and CD8αβlo tg TCRαβ+ cells are absent in H-Y-specific TCR transgenic mice with a
non-selecting H-2 haplotype (Figure 10, third row and data not shown). It also has
been shown that neither the presence of MHC class II molecules of the selecting H-
2b haplotype nor MHC class I molecules of the H-2d or H-2k haplotype were
sufficient to rescue H-Y-specific transgenic T cells from programmed cell death
(Table 4 and Figure 10 and data not shown) (Kisielow et al., 1988b; Teh et al.,
1988; von Boehmer, 1990). In summary, positive selection of H-Y-specific TCR
transgenic T cells seemed to be exclusively dependent on the presence of TCR-
restricting MHC class I molecules H-2Db and escape from negative selection was
modulated via CD8αβ co-receptor down-regulation. We analysed here the influence
of different H-2Db densities on selection of these TCR transgenic T cells and their
survival in the periphery.
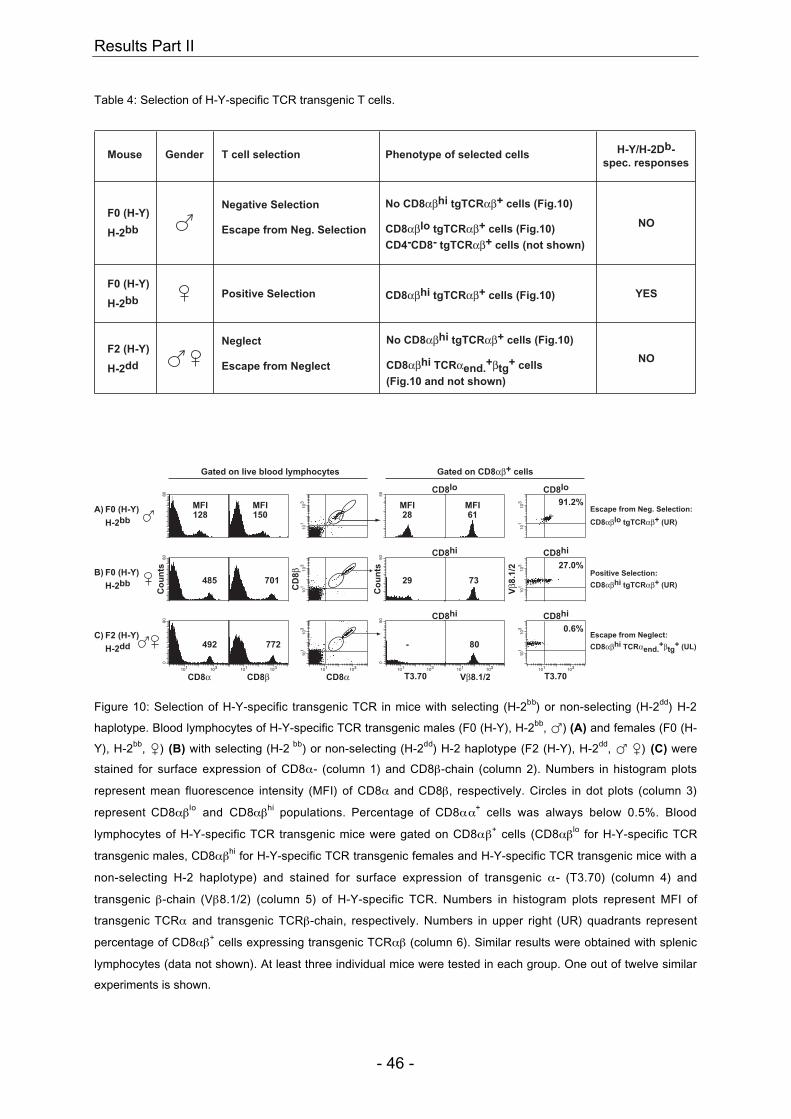
Results Part II
- 46 -
Table 4: Selection of H-Y-specific TCR transgenic T cells.
Mouse T cell selection
Positive Selection
Phenotype of selected cells
CD8αβhi tgTCRαβ+ cells (Fig.10)
H-Y/H-2Db-
spec. responses
YES
Negative Selection No CD8αβhi tgTCRαβ+ cells (Fig.10)
Escape from Neg. Selection
CD4-CD8- tgTCRαβ+ cells (not shown)
NOCD8αβlo tgTCRαβ+ cells (Fig.10)
Gender
F2 (H-Y)
H-2dd
F0 (H-Y)
H-2bb
F0 (H-Y)
H-2bb
No CD8αβhi tgTCRαβ+ cells (Fig.10)Neglect
NOEscape from Neglect
(Fig.10 and not shown)
CD8αβhi TCRαend.+βtg
+ cells
Gated on live blood lymphocytes Gated on CD8αβ+ cells
50
101
103
80
101
103 91.2%
A)
H-2bb
F0 (H-Y)
73701
50
29
Co
un
ts
101
103 27.0%
Vβ8
.1/2
CD
8β
101
103
Co
un
ts80
485B)
H-2bb
F0 (H-Y)
CD8α CD8αCD8β T3.70 T3.70Vβ8.1/2101 103
80
050
101 103
-
101 103
772
101
103
101 103
0.6%
101
103
101 103
080
101 103
492C)
H-2dd
F2 (H-Y)
CD8lo CD8lo
CD8hi
CD8hiCD8hi
CD8hi
128MFI
61MFI
28MFI
150MFI Escape from Neg. Selection:
CD8αβlo tgTCRαβ+ (UR)
Positive Selection:
CD8αβhi tgTCRαβ+ (UR)
Escape from Neglect:
CD8αβhi TCRαend.+βtg
+ (UL)
Figure 10: Selection of H-Y-specific transgenic TCR in mice with selecting (H-2bb) or non-selecting (H-2dd) H-2
haplotype. Blood lymphocytes of H-Y-specific TCR transgenic males (F0 (H-Y), H-2bb, �) (A) and females (F0 (H-
Y), H-2bb, �) (B) with selecting (H-2 bb) or non-selecting (H-2dd) H-2 haplotype (F2 (H-Y), H-2dd, � �) (C) were
stained for surface expression of CD8α- (column 1) and CD8β-chain (column 2). Numbers in histogram plots
represent mean fluorescence intensity (MFI) of CD8α and CD8β, respectively. Circles in dot plots (column 3)
represent CD8αβlo and CD8αβhi populations. Percentage of CD8αα+ cells was always below 0.5%. Blood
lymphocytes of H-Y-specific TCR transgenic mice were gated on CD8αβ+ cells (CD8αβlo for H-Y-specific TCR
transgenic males, CD8αβhi for H-Y-specific TCR transgenic females and H-Y-specific TCR transgenic mice with a
non-selecting H-2 haplotype) and stained for surface expression of transgenic α- (T3.70) (column 4) and
transgenic β-chain (Vβ8.1/2) (column 5) of H-Y-specific TCR. Numbers in histogram plots represent MFI of
transgenic TCRα and transgenic TCRβ-chain, respectively. Numbers in upper right (UR) quadrants represent
percentage of CD8αβ+ cells expressing transgenic TCRαβ (column 6). Similar results were obtained with splenic
lymphocytes (data not shown). At least three individual mice were tested in each group. One out of twelve similar
experiments is shown.
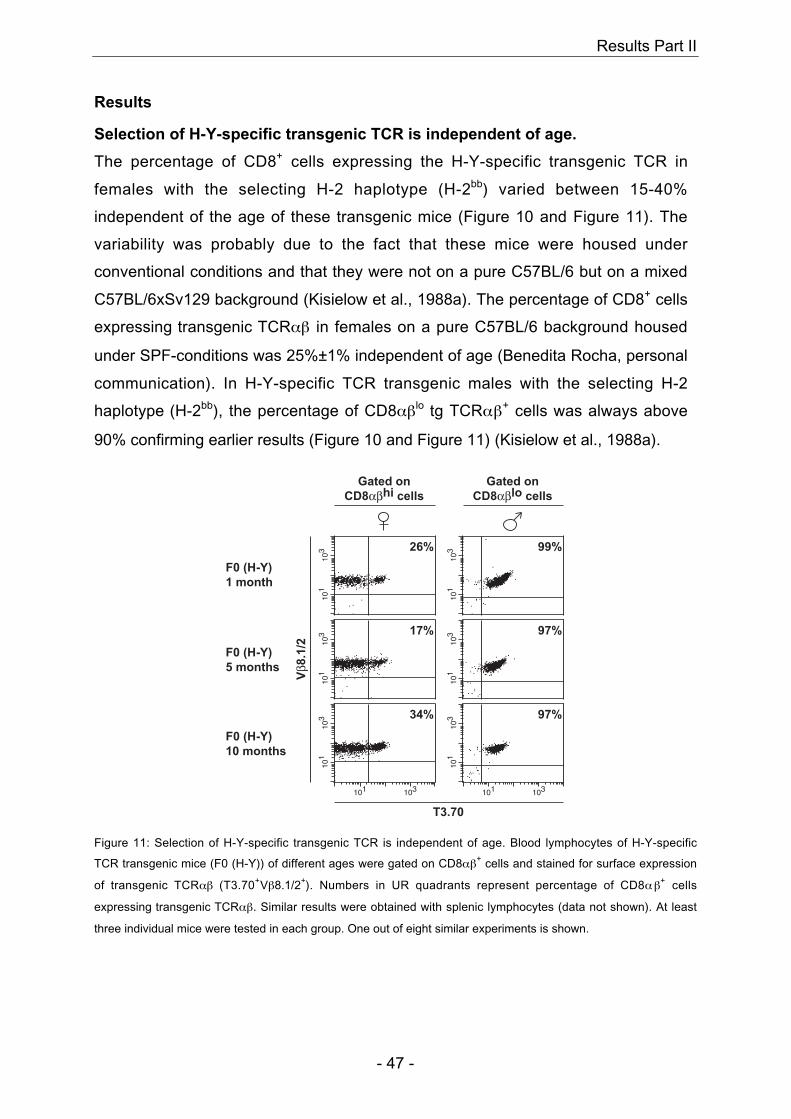
Results Part II
- 47 -
Results
Selection of H-Y-specific transgenic TCR is independent of age.
The percentage of CD8+ cells expressing the H-Y-specific transgenic TCR in
females with the selecting H-2 haplotype (H-2bb) varied between 15-40%
independent of the age of these transgenic mice (Figure 10 and Figure 11). The
variability was probably due to the fact that these mice were housed under
conventional conditions and that they were not on a pure C57BL/6 but on a mixed
C57BL/6xSv129 background (Kisielow et al., 1988a). The percentage of CD8+ cells
expressing transgenic TCRαβ in females on a pure C57BL/6 background housed
under SPF-conditions was 25%±1% independent of age (Benedita Rocha, personal
communication). In H-Y-specific TCR transgenic males with the selecting H-2
haplotype (H-2bb), the percentage of CD8αβlo tg TCRαβ+ cells was always above
90% confirming earlier results (Figure 10 and Figure 11) (Kisielow et al., 1988a).
T3.70
Vβ8
.1/2
F0 (H-Y)
1 month
F0 (H-Y)
10 months
F0 (H-Y)
5 months
Gated on
CD8αβhi cells
Gated on
CD8αβlo cells
101
103 26%
101
103 97%
101
103 99%
101
103 17%
101
103
101 103
101
103
101 103
97%34%
Figure 11: Selection of H-Y-specific transgenic TCR is independent of age. Blood lymphocytes of H-Y-specific
TCR transgenic mice (F0 (H-Y)) of different ages were gated on CD8αβ+ cells and stained for surface expression
of transgenic TCRαβ (T3.70+Vβ8.1/2+). Numbers in UR quadrants represent percentage of CD8αβ+ cells
expressing transgenic TCRαβ. Similar results were obtained with splenic lymphocytes (data not shown). At least
three individual mice were tested in each group. One out of eight similar experiments is shown.

Results Part II
- 48 -
The higher the H-2Db density, the higher the percentage of CD8+ cells
expressing H-Y-specific transgenic TCR in females.
To analyse percentage of CD8+ cells expressing transgenic TCRαβ in mice with half
H-2Db density, we crossed H-Y-specific TCR transgenic mice (F0 (H-Y), H-2bb) with
either BALB/c (H-2dd), CBA (H-2kk) or C57B10.G (H-2qq) mice obtaining TCR
transgenic mice with a heterozygous H-2 haplotype (F1 (H-Y), H-2bd, H-2bk or H-2bq,
respectively) (Figure 12 and Figure 13). Additionally we crossed F1 (H-Y), H-2bd
and F1 (H-Y), H-2bk mice with BALB/c or CBA mice, respectively, to obtain F2 mice
with a non-selecting H-2 haplotype (F2 (H-Y), H-2dd or H-2kk) (Figure 12 and Figure
13 and data not shown). All F1 (H-Y) transgenic females exhibited a strong
reduction in their percentage of peripheral CD8+ cells expressing transgenic TCRαβ
resulting in only 5-10% (Figure 12). The remaining CD8+ population still expressed
the transgenic TCRβ-chain but paired with endogenous TCRα-chains (Figure 12
and data not shown). The drastic decrease in percentage of CD8+ cells expressing
transgenic TCRαβ was mainly observed in the periphery because analysis of F1 (H-
Y) versus F0 (H-Y) CD8-single positive (SP) thymocytes revealed only a slight
decrease in the percentage of CD8-SP thymocytes expressing transgenic TCRαβ
(80% CD8-SP thymocytes in F0 (H-Y) versus 60% in F1 (H-Y) TCR transgenic
females expressed transgenic TCRαβ, data not shown). A possible explanation for
the latter observation could be the excellent three-dimensional thymic
microenvironment offering cytokines and chemokines, which could overcome the
lack of sufficient H-2Db density for efficient positive selection of transgenic
thymocytes. In the periphery, however, these conditions are not available optimally
for further survival and expansion of all positively selected transgenic T cells
(Kirberg et al., 1997; Rocha and von Boehmer, 1991). The reduced H-2Db density in
the periphery of F1 (H-Y) transgenic mice seemed not sufficient for survival of all
positively selected T cells and therefore only about one fourth to half of the once
positively selected T cells survived and expanded in the periphery (Figure 12 and
Benedita Rocha, personal communication). CD8+ cells from F2 (H-Y) transgenic
females with a non-selecting H-2 haplotype (H-2dd or H-2kk) did not show positive
selection of H-Y-specific transgenic TCR but expressed high levels of the
transgenic TCRβ-chain paired with endogenous TCRα-chains confirming previous
observations (Figure 12 and data not shown) (Fink and McMahan, 2000; Teh et al.,

Results Part II
- 49 -
1988; von Boehmer, 1990). In summary, in H-Y-specific TCR transgenic females on
a mixed C57BL/6xSv129 background, kept under conventional conditions, a
reduction in H-2Db density by 50% (F1 (H-Y) females) resulted in a 2 to 4-fold
decrease in percentage of peripheral CD8+ cells expressing transgenic TCRαβ. In
H-Y-specific TCR transgenic females on a pure C57BL/6-background, kept under
SPF-conditions, reduction in H-2Db density by 50% resulted in a 50% decrease in
percentage of peripheral CD8+ cells expressing transgenic TCRαβ and almost no
decrease in percentage of thymic CD8+ cells expressing transgenic TCRαβ
(Benedita Rocha, personal communication). Overall, the efficiency of positive
selection and survival in the periphery for a low-affinity TCR such as the H-Y-
specific TCR seemed therefore to depend on high TCR-restricting H-2Db densities
and high CD8αβ-expression levels (Figure 12) (Cruz et al., 1998; Podd et al., 2001;
Zerrahn et al., 1999). The latter results were already observed by Cruz et al.
showing that only CD8αβ+ cells expressing transgenic TCRαβ were positively
selected in females whereas CD8αα+ cells expressing transgenic TCRαβ were of
too low overall avidity to get positively selected (Cruz et al., 1998). The observation
that H-Y-specific TCR transgenic females with a mixed C57BL/6xSv129
background, housed under conventional conditions, showed a different percentage
of CD8+ cells expressing transgenic TCRαβ than TCR transgenic females with a
pure C57BL/6 background, housed under SPF conditions, showed that not only H-
2Db density but also other parameters such as the background of these mice and
chronic infections could directly influence positive selection and peripheral survival
of TCR transgenic T cells.
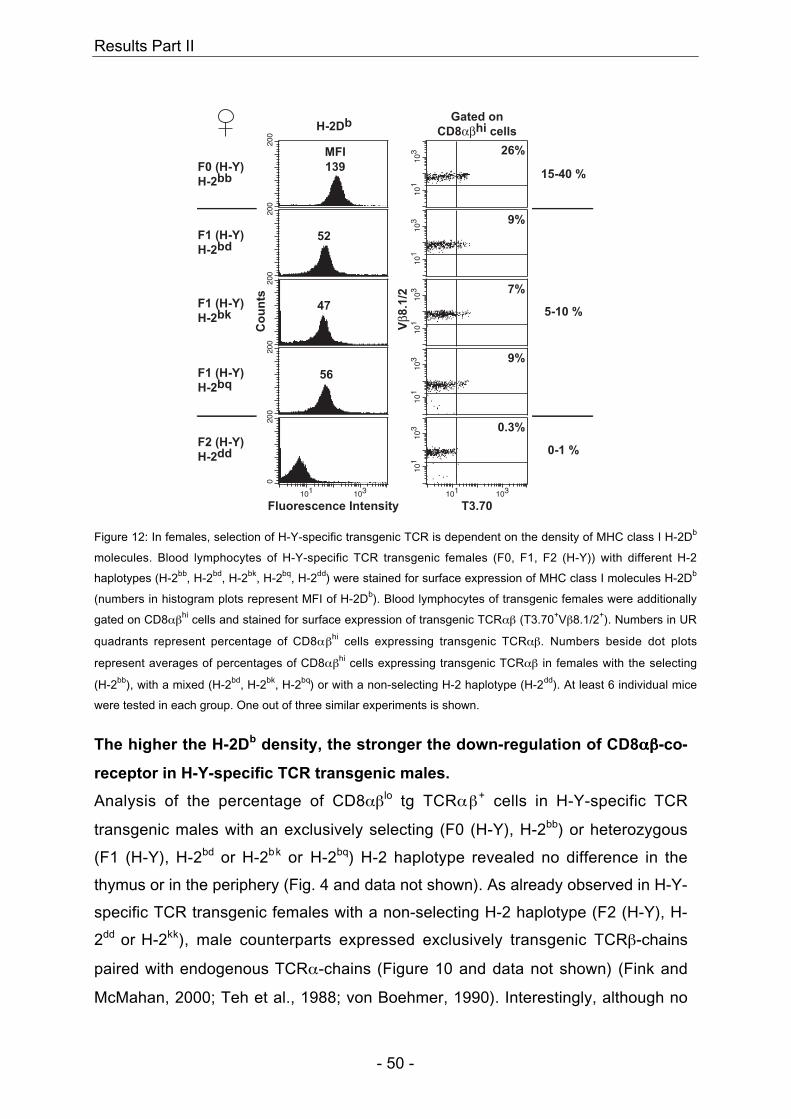
Results Part II
- 50 -
H-2DbGated on
CD8αβhi cells
T3.70
Vβ8
.1/2
Fluorescence Intensity
Co
un
ts200
200
52
0200
101 103
200
56
200
47
F0 (H-Y)
H-2bb
F2 (H-Y)
H-2dd
F1 (H-Y)
H-2bq
F1 (H-Y)
H-2bk
F1 (H-Y)
H-2bd
0-1 %
15-40 %
5-10 %
101
103
101
103
101
103
101
103
101
103
101 103
0.3%
9%
7%
9%
26%
139
MFI
Figure 12: In females, selection of H-Y-specific transgenic TCR is dependent on the density of MHC class I H-2Db
molecules. Blood lymphocytes of H-Y-specific TCR transgenic females (F0, F1, F2 (H-Y)) with different H-2
haplotypes (H-2bb, H-2bd, H-2bk, H-2bq, H-2dd) were stained for surface expression of MHC class I molecules H-2Db
(numbers in histogram plots represent MFI of H-2Db). Blood lymphocytes of transgenic females were additionally
gated on CD8αβhi cells and stained for surface expression of transgenic TCRαβ (T3.70+Vβ8.1/2+). Numbers in UR
quadrants represent percentage of CD8αβhi cells expressing transgenic TCRαβ. Numbers beside dot plots
represent averages of percentages of CD8αβhi cells expressing transgenic TCRαβ in females with the selecting
(H-2bb), with a mixed (H-2bd, H-2bk, H-2bq) or with a non-selecting H-2 haplotype (H-2dd). At least 6 individual mice
were tested in each group. One out of three similar experiments is shown.
The higher the H-2Db density, the stronger the down-regulation of CD8ααααββββ-co-
receptor in H-Y-specific TCR transgenic males.
Analysis of the percentage of CD8αβlo tg TCRαβ+ cells in H-Y-specific TCR
transgenic males with an exclusively selecting (F0 (H-Y), H-2bb) or heterozygous
(F1 (H-Y), H-2bd or H-2bk or H-2bq) H-2 haplotype revealed no difference in the
thymus or in the periphery (Fig. 4 and data not shown). As already observed in H-Y-
specific TCR transgenic females with a non-selecting H-2 haplotype (F2 (H-Y), H-
2dd or H-2kk), male counterparts expressed exclusively transgenic TCRβ-chains
paired with endogenous TCRα-chains (Figure 10 and data not shown) (Fink and
McMahan, 2000; Teh et al., 1988; von Boehmer, 1990). Interestingly, although no

Results Part II
- 51 -
change could be observed in the percentage of CD8αβlo tg TCRαβ+ cells in F0 (H-
Y) versus F1 (H-Y) transgenic males, a drastic difference was seen in CD8α- and
CD8β-chain expression levels on both thymic and peripheral CD8+ cells (Figure 13
and data not shown). CD8+ cells from F0 (H-Y) transgenic males showed the
strongest down-regulation for both CD8α- and CD8β-chains (Figure 13). CD8+ cells
from F1 (H-Y) transgenic males had less down-regulated CD8α- and CD8β-chains
(Figure 13) and CD8+ cells from F2 (H-Y) transgenic males exhibited the same
CD8αβ-expression levels as their female counterparts (Figure 10 and Figure 13).
Taken together, these results showed that TCR transgenic T cells in males actively
modulated their CD8αβ-expression levels during development in order to escape
clonal deletion. High H-2Db density in H-Y-specific TCR transgenic males (F0 (H-Y),
H-2bb) resulted in a too high overall avidity interaction between transgenic TCR and
self-peptide/self-MHC (H-Y/H-2Db) complexes on APC leading to negative selection
of H-Y-specific CD8αβhi tg TCRαβ+ cells (Table 4 and Figure 10) (Kisielow et al.,
1988a). Through strong down-regulation of both CD8α- and CD8β-chains, the
overall avidity was decreased and therefore CD8αβlo tg TCRαβ+ cells could escape
negative selection and migrate into the periphery (Table 4 and Figure 10 and Figure
13) (von Boehmer, 1990). Although decreased H-2Db density in TCR transgenic
mice with a heterozygous H-2 haplotype (F1 (H-Y), H-2bd, H-2bk or H-2bq) lead to a
lower overall avidity interaction between transgenic TCR and H-Y/H-2Db complexes,
clonal deletion of all H-Y-specific CD8αβhi tg TCRαβ+ cells was still observed
(Figure 13 and data not shown). Escape from negative selection in these mice,
however, was already achieved by a less strong down-regulation of both CD8α- and
CD8β-chains than the one observed in F0 (H-Y) transgenic males (Figure 13).
This study shows that the density of TCR-restricting MHC class I molecules H-2Db
is an important parameter in determining the fate of H-Y-specific TCR transgenic T
cells. In females, positive selection and survival of peripheral H-Y-specific CD8αβhi
tg TCRαβ+ cells was most efficient at high H-2Db density (Figure 12). In males,
negative selection of H-Y-specific CD8αβhi tg TCRαβ+ cells was independent of H-
2Db density (Figure 10 and Figure 13). Escape from negative selection, however,
was dependent on H-2Db density; the higher the H-2Db density, the stronger the
down-regulation of CD8αβ co-receptor (Figure 13).
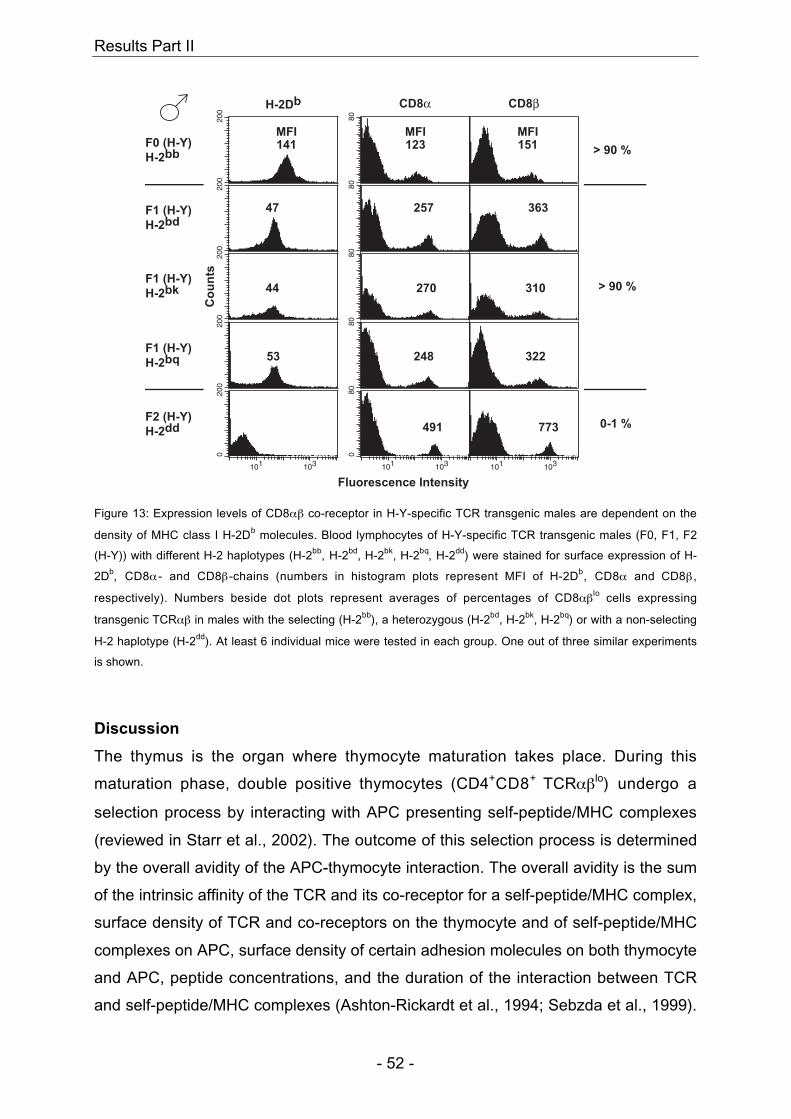
Results Part II
- 52 -
H-2Db CD8β
Fluorescence Intensity
Co
un
ts
F0 (H-Y)
H-2bb
F2 (H-Y)
H-2dd
F1 (H-Y)
H-2bq
F1 (H-Y)
H-2bk
F1 (H-Y)
H-2bd
> 90 %
0-1 %
> 90 %
CD8α
200
0200
101 103
200
53
200
44
200
47
080
101 103
491
80248
80
270
80
257
80
101 103
773
322
310
363
141MFI
151MFI
123MFI
Figure 13: Expression levels of CD8αβ co-receptor in H-Y-specific TCR transgenic males are dependent on the
density of MHC class I H-2Db molecules. Blood lymphocytes of H-Y-specific TCR transgenic males (F0, F1, F2
(H-Y)) with different H-2 haplotypes (H-2bb, H-2bd, H-2bk, H-2bq, H-2dd) were stained for surface expression of H-
2Db, CD8α - and CD8β-chains (numbers in histogram plots represent MFI of H-2Db, CD8α and CD8β ,
respectively). Numbers beside dot plots represent averages of percentages of CD8αβlo cells expressing
transgenic TCRαβ in males with the selecting (H-2bb), a heterozygous (H-2bd, H-2bk, H-2bq) or with a non-selecting
H-2 haplotype (H-2dd). At least 6 individual mice were tested in each group. One out of three similar experiments
is shown.
Discussion
The thymus is the organ where thymocyte maturation takes place. During this
maturation phase, double positive thymocytes (CD4+CD8+ TCRαβlo) undergo a
selection process by interacting with APC presenting self-peptide/MHC complexes
(reviewed in Starr et al., 2002). The outcome of this selection process is determined
by the overall avidity of the APC-thymocyte interaction. The overall avidity is the sum
of the intrinsic affinity of the TCR and its co-receptor for a self-peptide/MHC complex,
surface density of TCR and co-receptors on the thymocyte and of self-peptide/MHC
complexes on APC, surface density of certain adhesion molecules on both thymocyte
and APC, peptide concentrations, and the duration of the interaction between TCR
and self-peptide/MHC complexes (Ashton-Rickardt et al., 1994; Sebzda et al., 1999).

Results Part II
- 53 -
An overall weak to intermediate avidity interaction transmits a survival signal to the
maturing thymocyte (positive selection) whereas an overall strong avidity interaction
transmits a signal to the maturing thymocyte to undergo programmed cell death
(negative selection) (reviewed in Starr et al., 2002). This selection process ensures
therefore only survival of self-MHC-restricted (useful) and self-tolerant T cells, which
leave the thymus as mature T cells (reviewed in Ohashi, 2002; Starr et al., 2002).
Survival and further expansion of each T cell in the periphery, however, is only
achieved if the T cell remains in continuous interaction between its TCR and TCR-
restring MHC molecules on APC (Kirberg et al., 1997; Rocha and von Boehmer,
1991).
In this study we showed that positive selection and subsequent peripheral survival of
the low-affinity H-Y-specific transgenic TCR was less efficient with reduced TCR-
restricting MHC density (Figure 12 and Benedita Rocha, personal communication). In
contrast, this is not the case for high-affinity TCR such as the 318 or 2C transgenic
TCR (Pircher et al., 1989; Sha et al., 1988). In 318 TCR transgenic mice with
exclusively selecting (H-2bb) or heterozygous H-2 haplotype (H-2bd, H-2bk or H-2bq),
the percentage of peripheral CD8+ cells expressing transgenic TCR always remains
above 50% independent of reduced TCR-restricting H-2Db density (M. Martinic,
unpublished observation). Moreover, Zerrahn et al. show that only 2C TCR but not H-
Y-specific TCR transgenic cells are positively selected and survive in the periphery
when exclusively haematopoietic cells express TCR-restricting MHC molecules
(Zerrahn et al., 1999). Further, under sub-optimal conditions as given in Oncostatin M
(OM) transgenic mice, which show a profound thymic atrophy but massive
extrathymic T cell development (Boileau et al., 2000; Clegg et al., 1996), positive
selection and survival of the H-Y-specific but not of the 2C transgenic TCR is
reduced (Terra et al., 2002).
Taken together, these results show that as soon as environmental conditions are
sub-optimal – reduced TCR-restricting MHC density, different microenvironment,
different cytokine milieu – efficiency of positive selection and survival of low-affinity
but not of high-affinity TCR is decreased. A possible explanation could be that even
under sub-optimal conditions the intrinsic affinity of the high-affinity TCR but not of
the low-affinity TCR for self-peptide/MHC complexes is still high enough to provide
sufficient survival signal.

Results Part II
- 54 -
Negative selection of self-reactive T cells, however, is always complete. Even under
sub-optimal conditions, as already shown in H-Y/LckOM and 2C TCR/LckOM double
transgenic mice, all self-reactive T cells are absent (Terra et al., 2002). We confirmed
that observation, showing absence of all H-Y-specific cells (CD8αβhi tg TCRαβ+) in
TCR transgenic males with normal or with reduced H-2Db density (Figure 13 and
Benedita Rocha, personal communication). Additionally, even in a complete athymic
environment as is the case for nude H-Y-specific TCR transgenic males, we do not
detect any self-reactive T cells (Martinic et al., Manuscript in preparation-b).
Finally, escape from negative selection is probably only possible with low-affinity
TCR as seen with the H-Y-specific transgenic TCR (Figure 10 and Figure 13). There,
already a small modulation of the co-receptor was sufficient to fall under the overall
avidity threshold for negative selection. High-affinity TCR, in contrast, have under
physiological conditions most probably no chance to escape negative selection
because even by modulating co-receptors or other molecules on their cell-surface,
the intrinsic affinity of the high-affinity TCR itself remains too high.
In summary, selection and survival of a low-affinity TCR is most efficient under
optimal conditions, for example like in this study when restricting MHC antigens are
optimally present both in the thymus for initial positive selection and in the periphery
to ensure further survival. Furthermore, it is crucial that even under sub-optimal
conditions, negative selection remains tight to avoid potential auto-reactivity.

Results Part II
- 55 -
Materials and Methods
Mice
H-Y-specific TCR transgenic mice (F0 (H-Y), H-2bb) were a generous gift from Jörg
Kirberg. C57BL/6 (H-2bb), BALB/c (H-2dd), CBA (H-2kk) and C57B10.G (H-2qq) were
obtained from the Institute of Laboratory Animal Science, University of Zurich,
Switzerland. H-Y-specific TCR transgenic mice were bred with BALB/c, CBA or
C57B10.G mice yielding H-Y-specific TCR transgenic mice (F1 (H-Y)) with a
heterozygous H-2 haplotype (H-2bd, H-2bk or H-2b q, respectively). To obtain
transgenic mice with a non-selecting H-2 haplotype (H-2dd), H-Y-specific TCR
transgenic mice were bred twice with BALB/c (F2 (H-Y)) and tested with PCR and
FACS analysis for expression of transgenic TCR, presence of H-2dd and absence of
H-2bb MHC haplotype. All mice were kept under conventional conditions.
PCR
DNA was prepared from mouse-tail. 2 µl of mouse-tail-DNA were used for PCR
analysis. Primers used for amplification were specific for the H-Y-specific transgenic
TCR (Vβ8.2-fwd (5’ ACA AGG TGG CAG TAA CAG GA 3’) and Jβ2.3-rev (5’ ACA
GTC AGT CTG GTT CCT GA 3’) primers), for both H-2bb and H-2dd MHC haplotypes
(Ea5’-fwd (5’ AGT CTT CCC AGC CTT CAC ACT CAG AGG TAC 3’) and Ea3’-rev
(5’ CAT AGC CCC AAA TGT CTG ACC TCT GGA GAG 3’) primers) and for the H-
2dd MHC haplotype (K5’-fwd (5’ CAT GGG CAT AGA AAG GGC AGT CTT TGA ACT
3’) and Ea3’-rev primers). Expected bands were 302 bp for presence of H-Y-specific
transgenic TCR, 155 bp and 700 bp for presence of H-2b and H-2d MHC haplotype,
respectively, and 210 bp for presence of H-2d MHC haplotype.
Flow Cytometric Analysis
Peripheral blood cells were stained with the following antibodies: anti-CD8α-APC
(53-6.7), anti-CD8β.2-PE (53-5.8), T3.70-Biotin (specific for transgenic TCRα-chain
of H-Y-specific TCR) (generous gift from Benedita Rocha), anti-Vβ8.1/2-FITC
(specific for transgenic TCRβ-chain of H-Y-specific TCR) (MR5-2) and anti-H-2Db-
Biotin (KH95). Except for T3.70-Biotin, all antibodies were purchased from
Pharmingen. Streptavidin-PerCP and Streptavidin-Tricolor were purchased from

Results Part II
- 56 -
Pharmingen and Caltag Laboratories, respectively. All samples were acquired on a
FACSCalibur and analysed using CellQuest software (Becton Dickinson).
Acknowledgments
We thank Dr. Benedita Rocha for the generous gift of T3.70 mAb, and Drs. M. van
den Broek for helpful comments on the manuscript. This work was supported by the
Swiss National Foundation for Science and the Kanton of Zurich, Switzerland.

Results Part III
- 57 -
7 Results Part III
Selection of the H-Y-specific transgenic TCR in an athymic versus
euthymic environment
Marianne M. Martinic, Hans Hengartner and Rolf M. Zinkernagel
Institute of Experimental Immunology, Department of Pathology, University Hospital
Zurich, Schmelzbergstrasse 12, CH-8091 Zurich, Switzerland

Results Part III
- 58 -
Abstract
To compare intrathymic and extrathymic T cell selection, H-Y-specific T cell
receptor (TCR) transgenic mice (H-2bb) were crossed onto athymic C57BL/6- (H-
2bb) or BALB/c-nudes (H-2dd). Athymic and euthymic offspring were analysed
for selecting (H-2bb, H-2bd) or non-selecting (H-2dd) H-2 haplotype, and for
expression of transgenic TCR. Athymic mice had much lower absolute
numbers of CD8+ cells expressing transgenic TCR than euthymic controls. The
percentage of positively selected H-Y-specific transgenic CD8+ cells in females,
however, was comparable to euthymic controls. Negative selection of
transgenic TCR in athymic males was complete, as has been known for
euthymic males. Also escape from negative selection via down-regulation of
CD8ααααββββ co-receptor was observed in athymic males. In contrast, in athymic
mice escape from neglect via endogenous Vαααα-chain gene rearrangement was
less efficient than in euthymic mice. These results indicate that the thymic
microenvironment is 1) not essential for but much improves measurable
maturation and 2) is not necessary for negative selection of highly frequent
transgenic TCR expressing T cells.
Introduction
Our aim was to compare intrathymic and extrathymic T cell selection to analyse
whether the periphery could substitute a non-functional thymus by providing an
environment capable of selecting a functional self-MHC-restricted and self-tolerant T
cell repertoire. As it is well known that the thymus is absolutely required for T cell
receptor (TCR) rearrangement (Miller, 1961), we took advantage of a TCR transgenic
(tg) mouse system to compare selection of this specific transgenic TCR in an intra-
versus extrathymic environment. We analysed selection of the H-Y-specific
transgenic TCR, which is specific for a male antigen-derived peptide (H-Y) presented
on MHC class I H-2Db molecules (Kisielow et al., 1988a). The advantage of this
transgenic TCR is that without need of any external manipulation and by solely
comparing transgenic females with transgenic males, various parameters of T cell
selection can be directly assessed. In H-Y-specific TCR transgenic females (H-2bb),
presence of self-MHC (H-2Db) and absence of self-antigen (H-Y) lead to positive
selection of self-MHC-restricted H-Y-specific CD8αβhi tg TCRαβ+ cells (Kisielow et
al., 1988a). Maximally only 40% of CD8+ cells express the actual transgenic TCR, the

Results Part III
- 59 -
remaining CD8+ population recombined the endogenous TCRα loci and changed
TCR specificity during thymocyte selection (Buch et al., 2002; Huesmann et al.,
1991; Merkenschlager et al., 1994). In H-Y-specific TCR transgenic males (H-2bb),
presence of self-MHC (H-2Db) and of self-antigen (H-Y) clonally deletes all H-Y-
specific CD8αβhi tg TCRαβ+ cells (Kisielow et al., 1988a). Escape from negative
selection occurs through down-regulation of CD8αβ co-receptor resulting in H-Y-
unreactive CD8αβlo tg TCRαβ+ cells (Kisielow et al., 1988a; Teh et al., 1989). In H-Y-
specific TCR transgenic mice with non-selecting H-2 haplotype (H-2dd), H-Y-specific
CD8αβhi tg TCRαβ+ cells do not get positively selected and escape death from
neglect via endogenous rearrangement of TCRα-chain genes (Fink and McMahan,
2000; Teh et al., 1988; von Boehmer, 1990). To compare positive and negative
selection, escape from negative selection and escape from neglect in an intra- versus
extrathymic environment, euthymic H-Y-specific TCR transgenic mice (H-2bb) were
crossed twice with athymic C57BL/6- (H-2bb) or BALB/c-nudes (H-2dd) yielding
euthymic and athymic offspring with a selecting (H-2bb or H-2bd) or non-selecting H-2
haplotype (H-2dd).
Results
Athymic TCR transgenic mice show a strong reduction in absolute numbers of
transgenic cells compared to euthymic counterparts.
The biggest and striking difference between euthymic and athymic TCR transgenic
mice was the drastic reduction in absolute cell numbers (Table 5). Euthymic males
always had at least 40-fold more CD8αβlo tg TCRαβ+ splenocytes than athymic
males (Table 5). The difference between H-Y-specific CD8αβhi tg TCRαβ+ and
CD8αβhi tg TCRαend.+βtg
+ splenocytes in euthymic and athymic females was even
more striking, reaching a 120-fold decrease for athymic females (Table 5). Similar
reductions in absolute numbers were observed in liver and blood of athymic TCR
transgenic mice (data not shown). A possible explanation for the higher decrease in
absolute numbers in athymic females versus males may be that in athymic males
presence of self-MHC and self-antigen may lead to a primary survival signal for the
transgenic T cell via the TCR, allowing this T cell even under sub-optimal conditions
to move on to the next maturation step, which would be escape from negative
selection through CD8αβ co-receptor down-regulation. In presence of self-MHC but

Results Part III
- 60 -
absence of self-antigen - representing the athymic female environment – not all TCR
transgenic T cells may receive a sufficient high and long survival signal to move on to
the next maturation step, which would either be positive selection of H-Y-specific
CD8αβhi tg TCRαβ+ cells or internalisation of transgenic TCRα-chain followed by
endogenous Vα-chain gene rearrangement and expression of a TCR with new
specificity. A thymic environment, however, provides optimal conditions via the
perfect three-dimensional micro-architecture, its professional antigen presenting cells
and the specialized chemokine and cytokine milieu allowing immature thymocytes
not only to survive longer and therefore giving the thymocyte the chance to move on
to the next maturation step but also to increase their proliferation rate resulting in
higher absolute output cell numbers. Finally, thymocytes thanks to the thymus/blood
barrier are protected against peripheral chemokines such as TNF-α, which is
secreted during an immune response and is toxic for maturing cells, giving
thymocytes an additional survival advantage over extra-thymically maturing T cells
(Martin and Bevan, 1997; Orange et al., 1995; Wang et al., 1994).
Additionally, we analysed athymic TCR transgenic mice for presence of CD8αα+ tg
TCRαβ+ cells in the gut, blood, spleen and liver (data not shown). As observed
previously for euthymic TCR transgenic mice (Cruz et al., 1998), CD8αα+ tg TCRαβ+
cells were found exclusively in the gut of euthymic and athymic males but not at all in
euthymic or athymic females (data not shown).
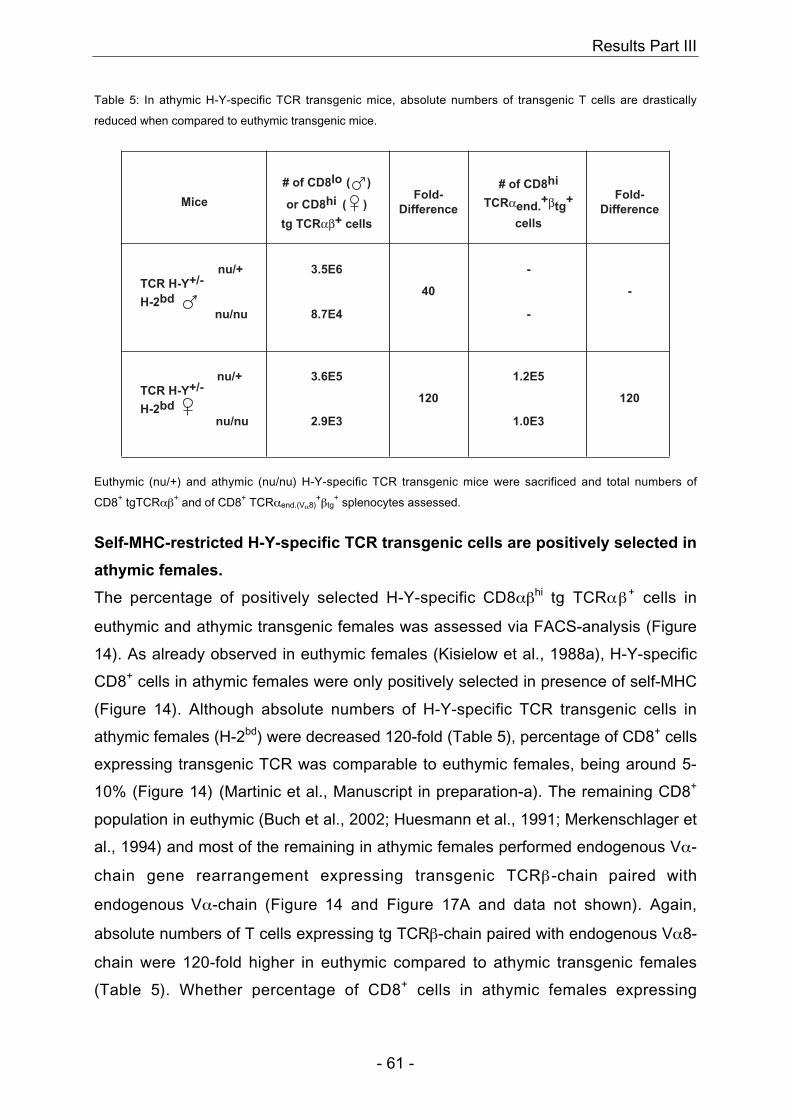
Results Part III
- 61 -
Table 5: In athymic H-Y-specific TCR transgenic mice, absolute numbers of transgenic T cells are drastically
reduced when compared to euthymic transgenic mice.
TCR H-Y+/-
H-2bd
TCR H-Y+/-
H-2bd
nu/+
nu/+
nu/nu
nu/nu
MiceFold-
Difference
40
120
3.5E6
2.9E3
3.6E5
8.7E4
tg TCRαβ+ cells
# of CD8lo ( )
or CD8hi ( )Fold-
Difference
120
-
1.0E3
1.2E5
# of CD8hi
TCRαend.+βtg
+
cells
-
-
Euthymic (nu/+) and athymic (nu/nu) H-Y-specific TCR transgenic mice were sacrificed and total numbers of
CD8+ tgTCRαβ+ and of CD8+ TCRαend.(Vα8)+βtg
+ splenocytes assessed.
Self-MHC-restricted H-Y-specific TCR transgenic cells are positively selected in
athymic females.
The percentage of positively selected H-Y-specific CD8αβhi tg TCRαβ+ cells in
euthymic and athymic transgenic females was assessed via FACS-analysis (Figure
14). As already observed in euthymic females (Kisielow et al., 1988a), H-Y-specific
CD8+ cells in athymic females were only positively selected in presence of self-MHC
(Figure 14). Although absolute numbers of H-Y-specific TCR transgenic cells in
athymic females (H-2bd) were decreased 120-fold (Table 5), percentage of CD8+ cells
expressing transgenic TCR was comparable to euthymic females, being around 5-
10% (Figure 14) (Martinic et al., Manuscript in preparation-a). The remaining CD8+
population in euthymic (Buch et al., 2002; Huesmann et al., 1991; Merkenschlager et
al., 1994) and most of the remaining in athymic females performed endogenous Vα-
chain gene rearrangement expressing transgenic TCRβ-chain paired with
endogenous Vα-chain (Figure 14 and Figure 17A and data not shown). Again,
absolute numbers of T cells expressing tg TCRβ-chain paired with endogenous Vα8-
chain were 120-fold higher in euthymic compared to athymic transgenic females
(Table 5). Whether percentage of CD8+ cells in athymic females expressing

Results Part III
- 62 -
transgenic TCRαβ is also dependent on MHC-density levels as observed in euthymic
females remains to be proven (Martinic et al., Manuscript in preparation-a). H-Y-
specific CD8αβhi tg TCRαβ+ cells in euthymic females with a non-selecting H-2
haplotype (TCR H-Y+/-, H-2d d) escaped death from neglect via endogenous
rearrangement of TCRα-chain genes (Figure 14 and data not shown) (Fink and
McMahan, 2000; Teh et al., 1988). Athymic females with a non-selecting H-2
haplotype (TCR H-Y+/-, H-2dd), however, did not escape death from neglect via
endogenous Vα-chain gene rearrangement to detectable levels (Figure 14 and data
not shown). The latter observation may reflect again the disadvantage of a sub-
optimal athymic environment where absence of both self-MHC and self-antigen may
result in a too low and short survival signal for the maturing T cell.
101
103
101 103
101
103
101
103
101
103
Gated on CD8αβhi cells
nu/+ nu/nu
T3.70
Vβ8
.1/2
74%
90%
98%
18%
25.9%
9.4%
0.4%
0.0%
101
103
101
103
101 103
101
103 10% 0.7%
3% 0.0%
69% 7.4%
TCR H-Y+/-
H-2dd
TCR H-Y-/-
H-2bd
TCR H-Y+/-
H-2bb
TCR H-Y+/-
H-2bd
Figure 14: In presence of self-MHC (H-2b), positive selection of H-Y-specific transgenic TCR is seen in both
euthymic and athymic females. Blood lymphocytes of euthymic (nu/+) and splenocytes of athymic (nu/nu) H-Y-
specific TCR transgenic females (TCR H-Y+/-) expressing the selecting (H-2bb), a mixed (H-2bd) or non-selecting
H-2 haplotype (H-2dd) were gated on CD8αβhi cells and stained for surface expression of transgenic TCRαβ
(T3.70+Vβ8.1/2+). Transgenic negative littermates (TCR H-Y-/-, H-2bd) were used as negative controls. Numbers in
upper left (UL) and upper right (UR) quadrants represent percentage of CD8+ cells expressing exclusively
transgenic TCRβ-chain, and transgenic TCRαβ, respectively. Similar results were obtained with liver lymphocytes
of euthymic and athymic mice (data not shown). At least three individual mice were tested in each group. One out
of three similar experiments is shown.

Results Part III
- 63 -
Athymic TCR transgenic males show absence of all H-Y-specific transgenic
cells. Escape from negative selection occurs via down-regulation of CD8ααααββββ co-
receptor.
To compare negative selection in an athymic versus euthymic environment, H-Y-
specific TCR transgenic males were analysed for transgenic TCRαβ expression
(Figure 15 and Figure 16). Without exception, all males with selecting H-2 haplotype
(H-2bb, H-2bd) showed complete absence of H-Y-specific CD8αβhi tg TCRαβ+ cells
(Figure 15 and Figure 16 and data not shown). In addition, as already observed in
euthymic males, athymic males showed escape from negative selection via down-
regulation of CD8αβ co-receptor (Figure 16) leading to H-Y unreactive CD8αβlo tg
TCRαβ+ cells (Figure 15) (Kisielow et al., 1988a; Teh et al., 1989). The degree of
CD8αβ co-receptor down-regulation was strongly dependent on the density of MHC
class I H-2Db molecules; the higher the density of H-2Db, the stronger the down-
regulation of CD8αβ co-receptor in both euthymic and athymic males (Figure 16)
(Martinic et al., Manuscript in preparation-a). Escape from neglect via endogenous
Vα-chain gene rearrangement as observed in euthymic males with non-selecting H-2
haplotype (H-2dd) was also seen in athymic males (Figure 15 and Figure 17) (Fink
and McMahan, 2000; Teh et al., 1988). Interestingly, only athymic males but not
females with a non-selecting H-2 haplotype (H-2dd) were able to escape death from
neglect via endogenous Vα-chain gene rearrangement (Figure 14, Figure 15 and
Figure 17 and data not shown). One possible explanation may be that the H-Y-
specific transgenic TCR in athymic males with non-selecting H-2 haplotype (H-2dd)
cross-reacted with a male-specific antigen presented on H-2d leading to a high and
long enough survival signal to perform subsequent endogenous gene rearrangement
even in absence of a thymic environment. The resulting TCR (TCRαend.+βtg
+) with
new specificity and restriction to self-MHC H-2d would then enable the transgenic T
cell to survive and expand through continuous peripheral interaction with self-MHC
(Kirberg et al., 1997; Rocha and von Boehmer, 1991).

Results Part III
- 64 -
101
103
101
103
101
103
101 103
101
103
101
103
101 103
101
103
101
103
Gated on CD8αβlo cells
nu/+ nu/nu
T3.70
Vβ8
.1/2
95.9%
90.1%
1.4%
0.0%
1.2%
0.0%
92.4%
2.9%
92.9%
4.3%
17.4%
97.0%
7.7%
1.7%
TCR H-Y+/-
H-2dd
TCR H-Y-/-
H-2bd
TCR H-Y+/-
H-2bb
TCR H-Y+/-
H-2bd
Figure 15: In presence of self-MHC (H-2b) and self-peptide (H-Y), H-Y-specific transgenic T cells are absent in
both euthymic and athymic males. Blood lymphocytes of euthymic (nu/+) and athymic (nu/nu) H-Y-specific TCR
transgenic males (TCR H-Y+/-) expressing the selecting (H-2bb), a mixed (H-2bd) or non-selecting H-2 haplotype
(H-2dd) were gated on CD8αβlo cells and stained for surface expression of transgenic TCRαβ (T3.70+Vβ8.1/2+).
Transgenic negative littermates (TCR H-Y-/-, H-2bd) were used as negative controls. Numbers in UL and UR
corners represent percentage of CD8+ cells expressing exclusively transgenic TCRβ-chain, and transgenic
TCRαβ, respectively. Similar results were obtained with splenocytes and liver lymphocytes of euthymic and
athymic mice (data not shown). At least three individual mice were tested in each group. One out of three similar
experiments is shown.
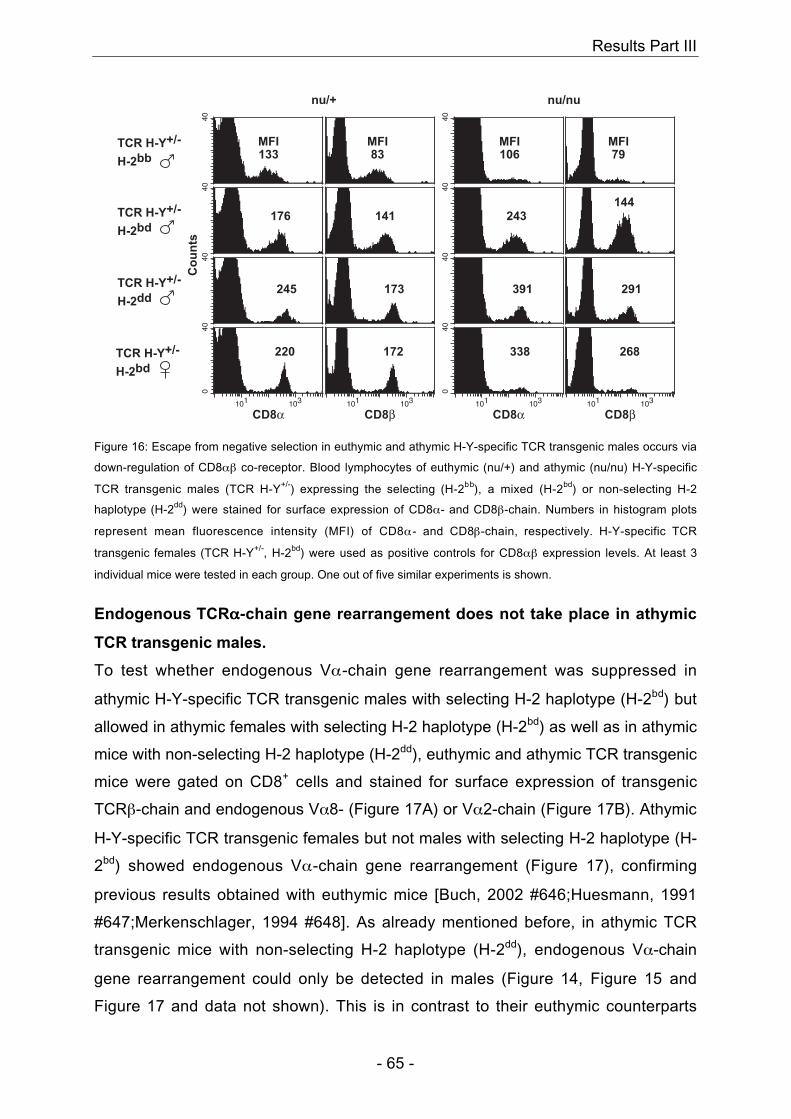
Results Part III
- 65 -
040
101 103
4040
40
101 103
040
101 103
4040
40
101 103
nu/+ nu/nu
MFI133
268
291
144
MFI79
338
391
243
MFI106
MFI83
141
173
172220
245
176
Co
un
ts
CD8α CD8α CD8βCD8β
TCR H-Y+/-
H-2dd
TCR H-Y+/-
H-2bd
TCR H-Y+/-
H-2bb
TCR H-Y+/-
H-2bd
Figure 16: Escape from negative selection in euthymic and athymic H-Y-specific TCR transgenic males occurs via
down-regulation of CD8αβ co-receptor. Blood lymphocytes of euthymic (nu/+) and athymic (nu/nu) H-Y-specific
TCR transgenic males (TCR H-Y+/-) expressing the selecting (H-2bb), a mixed (H-2bd) or non-selecting H-2
haplotype (H-2dd) were stained for surface expression of CD8α- and CD8β-chain. Numbers in histogram plots
represent mean fluorescence intensity (MFI) of CD8α- and CD8β-chain, respectively. H-Y-specific TCR
transgenic females (TCR H-Y+/-, H-2bd) were used as positive controls for CD8αβ expression levels. At least 3
individual mice were tested in each group. One out of five similar experiments is shown.
Endogenous TCRαααα-chain gene rearrangement does not take place in athymic
TCR transgenic males.
To test whether endogenous Vα-chain gene rearrangement was suppressed in
athymic H-Y-specific TCR transgenic males with selecting H-2 haplotype (H-2bd) but
allowed in athymic females with selecting H-2 haplotype (H-2bd) as well as in athymic
mice with non-selecting H-2 haplotype (H-2dd), euthymic and athymic TCR transgenic
mice were gated on CD8+ cells and stained for surface expression of transgenic
TCRβ-chain and endogenous Vα8- (Figure 17A) or Vα2-chain (Figure 17B). Athymic
H-Y-specific TCR transgenic females but not males with selecting H-2 haplotype (H-
2bd) showed endogenous Vα-chain gene rearrangement (Figure 17), confirming
previous results obtained with euthymic mice [Buch, 2002 #646;Huesmann, 1991
#647;Merkenschlager, 1994 #648]. As already mentioned before, in athymic TCR
transgenic mice with non-selecting H-2 haplotype (H-2dd), endogenous Vα-chain
gene rearrangement could only be detected in males (Figure 14, Figure 15 and
Figure 17 and data not shown). This is in contrast to their euthymic counterparts

Results Part III
- 66 -
where both females and males performed endogenous Vα -chain gene
rearrangement (Figure 14, Figure 15 and Figure 17 and data not shown) (Fink and
McMahan, 2000; Teh et al., 1988).
101
103
101
103
101
103
101
103
101
103
101
103
101
103
101 103
101
103
101 103
nu/+ nu/nu
Vα8
Vβ8
.1/2
96%
97%
96%
15%
0.0%
1.9%
3.1%
0.8%
7.2% 1.0%
2.0% 0.0%
45% 8.4%
CD8αβhiCD8αβhi
CD8αβloCD8αβlo
78% 0.0%
TCR H-Y+/-
H-2bd
TCR H-Y+/-
H-2dd
TCR H-Y-/-
H-2bd
TCR H-Y+/-
H-2bd
TCR H-Y+/-
H-2dd
TCR H-Y+/-
H-2bd
Vα2
CD8αβhiCD8αβhi
CD8αβloCD8αβlo
Vβ8
.1/2
101
103
101 103
101
103
101
103
101 103
101
103 99%
92%
0.0%
4.8% 96% 1.6%
98% 0.0%
A
B
Figure 17: Endogenous Vα-chain gene rearrangement is less efficient in athymic H-Y-specific TCR transgenic
mice. Splenocytes of euthymic (nu/+) and athymic (nu/nu) H-Y-specific TCR transgenic mice (TCR H-Y+/-) with a
mixed (H-2bd) or non-selecting H-2 haplotype (H-2dd) were gated on CD8αβlo or CD8αβhi cells and stained for
surface expression of endogenous Vα8-chain (Vα8) and transgenic TCRβ-chain (Vβ8.1/2) (A) or endogenous
Vα2-chain (Vα2) and transgenic TCRβ-chain (Vβ8.1/2) (B). Transgenic negative females (TCR H-Y-/-, H-2bd) were
used as negative controls. Numbers in UL and UR quadrants represent percentage of CD8+ cells expressing
exclusively transgenic TCRβ-chain, and endogenous Vα- and transgenic TCRβ-chain, respectively. Similar

Results Part III
- 67 -
results were obtained with blood lymphocytes of euthymic and athymic mice (data not shown). At least three
individual mice were tested in each group. One out of three similar experiments is shown.
In summary, selection of the H-Y-specific transgenic TCR in an athymic versus
euthymic environment was mostly qualitatively (Figure 14, Figure 15, Figure 16,
Figure 17 and Table 6) but not quantitatively (Table 5) comparable. Athymic TCR
transgenic females with selecting H-2 haplotype (H-2bd) showed positive selection of
H-Y-specific CD8αβhi tg TCRαβ+ cells (Figure 14) whereas these cells were
completely absent in males (Figure 15 and Figure 16). Escape from negative
selection via CD8αβ co-receptor down-regulation was as efficient as in euthymic
males (Figure 16) (Kisielow et al., 1988a). The level of CD8αβ co-receptor down-
regulation was dependent on TCR-restricting H-2Db density confirming previous
observations with euthymic TCR transgenic males (Figure 16) (Martinic et al.,
Manuscript in preparation-a). Escape from neglect via endogenous Vα-chain gene
rearrangement in athymic TCR transgenic mice with non-selecting H-2 haplotype (H-
2dd) was only observed in males whereas euthymic controls showed escape from
neglect in both females and males (Figure 14, Figure 15 and Figure 17 and data not
shown) (Fink and McMahan, 2000; Teh et al., 1988). Finally, absolute numbers of H-
Y-specific CD8αβhi tg TCRαβ+ cells and of CD8αβhi TCRαend.+βtg
+ cells in athymic
females with selecting H-2 haplotype (H-2bd) were reduced 120-fold, absolute
numbers of CD8αβlo tg TCRαβ+ cells in athymic males having escaped negative
selection were reduced 40-fold when compared to their euthymic counterparts (Table
5).
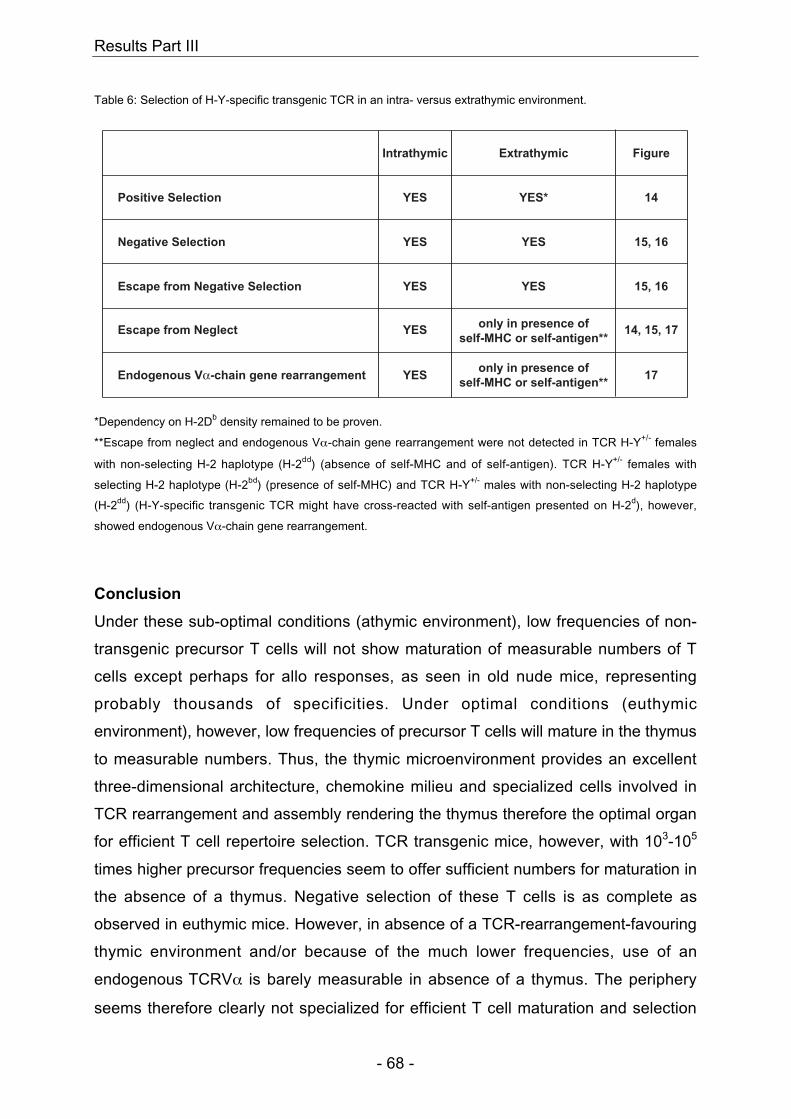
Results Part III
- 68 -
Table 6: Selection of H-Y-specific transgenic TCR in an intra- versus extrathymic environment.
Intrathymic FigureExtrathymic
Escape from Negative Selection YES 15, 16YES
Negative Selection YES 15, 16YES
Positive Selection YES 14YES*
Endogenous Vα-chain gene rearrangement YES 17only in presence of
self-MHC or self-antigen**
Escape from Neglect YES 14, 15, 17only in presence of
self-MHC or self-antigen**
*Dependency on H-2Db density remained to be proven.
**Escape from neglect and endogenous Vα-chain gene rearrangement were not detected in TCR H-Y+/- females
with non-selecting H-2 haplotype (H-2dd) (absence of self-MHC and of self-antigen). TCR H-Y+/- females with
selecting H-2 haplotype (H-2bd) (presence of self-MHC) and TCR H-Y+/- males with non-selecting H-2 haplotype
(H-2dd) (H-Y-specific transgenic TCR might have cross-reacted with self-antigen presented on H-2d), however,
showed endogenous Vα-chain gene rearrangement.
Conclusion
Under these sub-optimal conditions (athymic environment), low frequencies of non-
transgenic precursor T cells will not show maturation of measurable numbers of T
cells except perhaps for allo responses, as seen in old nude mice, representing
probably thousands of specificities. Under optimal conditions (euthymic
environment), however, low frequencies of precursor T cells will mature in the thymus
to measurable numbers. Thus, the thymic microenvironment provides an excellent
three-dimensional architecture, chemokine milieu and specialized cells involved in
TCR rearrangement and assembly rendering the thymus therefore the optimal organ
for efficient T cell repertoire selection. TCR transgenic mice, however, with 103-105
times higher precursor frequencies seem to offer sufficient numbers for maturation in
the absence of a thymus. Negative selection of these T cells is as complete as
observed in euthymic mice. However, in absence of a TCR-rearrangement-favouring
thymic environment and/or because of the much lower frequencies, use of an
endogenous TCRVα is barely measurable in absence of a thymus. The periphery
seems therefore clearly not specialized for efficient T cell maturation and selection

Results Part III
- 69 -
but as already known is essential for further survival, expansion and differentiation
(Kirberg et al., 1997; Rocha and von Boehmer, 1991; Zinkernagel and Althage,
1999). Thus, T cell maturation is probably 103-104 times more efficient in a thymus
but can occur rarely in its absence. Biologically, however, this numbers game
separates immunocompetence from general immunodeficiency as experienced by
thymus-deficient nude mice or humans.

Results Part III
- 70 -
Materials and Methods
Mice
H-Y-specific TCR transgenic mice (TCR H-Y+/+, H-2bb) were a generous gift from Jörg
Kirberg. BALB/c- (H-2dd) and C57BL/6-nudes (H-2bb) were purchased from the
Institute of Laboratory Animal Science, University of Zurich, Switzerland, and from
RCC Biotech, Füllinsdorf, Switzerland, respectively. H-Y-specific TCR transgenic
mice were bred with C57BL/6- or BALB/c-nudes for two generations yielding
euthymic (nu/+) and athymic (nu/nu) H-Y-specific TCR transgenic mice expressing
different H-2 haplotypes (TCR H-Y+/-, H-2bb, H-2bd or H-2dd). The H-2 haplotype of
these mice and expression of transgenic TCR were tested using PCR- and FACS-
analysis.
PCR
DNA was prepared from mouse-tail. 2 µl of mouse-tail-DNA were used for PCR
analysis. Primers used for amplification were specific for the H-Y-specific transgenic
TCR (Vβ8.2-fwd (5’ ACA AGG TGG CAG TAA CAG GA 3’) and Jβ2.3-rev (5’ ACA
GTC AGT CTG GTT CCT GA 3’) primers), for both H-2bb and H-2dd MHC haplotypes
(Ea5’-fwd (5’ AGT CTT CCC AGC CTT CAC ACT CAG AGG TAC 3’) and Ea3’-rev
(5’ CAT AGC CCC AAA TGT CTG ACC TCT GGA GAG 3’) primers) and for the H-
2dd MHC haplotype (K5’-fwd (5’ CAT GGG CAT AGA AAG GGC AGT CTT TGA ACT
3’) and Ea3’-rev primers). Expected bands were 302 bp for presence of H-Y-specific
transgenic TCR, 155 bp and 700 bp for presence of H-2b and H-2d MHC haplotype,
respectively, and 210 bp for presence of H-2d MHC haplotype.
Flow Cytometric Analysis
Peripheral blood cells or splenocytes were stained with the following antibodies: anti-
CD8α-APC (53-6.7), anti-CD8α-Tricolor (53-6.7), anti-CD8β.2-PE (53-5.8), T3.70-
Biotin (specific for transgenic TCRα-chain of H-Y-specific TCR) (generous gift from
Benedita Rocha), anti-Vβ8.1/2-FITC (MR5-2) (specific for transgenic TCRβ-chain of
H-Y-specific TCR), anti-Vα8-Biotin (KT50) and anti-Vα2-PE (B20.1). Except for
T3.70-Biotin antibody, all antibodies were purchased from Pharmingen. Streptavidin-
PerCP was purchased from Pharmingen, Streptavidin-Tricolor from Caltag

Results Part III
- 71 -
Laboratories. All samples were acquired on a FACScan or FACSCalibur and
analysed using CellQuest software (Becton Dickinson).
Acknowledgments
We thank Dr. Benedita Rocha for the generous gift of T3.70 mAb. This work was
supported by the Swiss National Foundation for Science and the Kanton of Zurich,
Switzerland.


General Discussion
- 73 -
8 General Discussion
An optimal T-cell mediated immune response is achieved in the presence of a
mature, functional and diverse T cell repertoire, which consists of self-MHC-restricted
and self-tolerant T cells (reviewed in Sebzda et al., 1999; Stefanova et al., 2003; von
Boehmer et al., 2003). Based on the established data and results obtained here,
selection and survival of such an efficient T cell repertoire can be summarized as
follows with respect to what defines this T cell repertoire, why is it needed, which
cells are involved, when and where does it occur and which are the different
parameters influencing it (Figure 18).
T cell repertoire
selection and survival
WHAT?
WHERE?
HOW?
WHEN?WHO?
WHY?
Figure 18: On T cell repertoire selection and survival.

General Discussion
- 74 -
WHAT and WHY?
The immune system of humans and other vertebrates is an adaptive defence system
that has evolved to protect from invading pathogenic microorganisms (bacteria, virus,
fungi) and cancer (Goldsby et al., 2000). It consists of both non-specific (innate
immunity) and specific (adaptive immunity) components. Just after contact to a
pathogen, innate immunity provides the first line of defence (i.e. via phagocytic cells,
release of interferon, complement activation). If the invading pathogen evades the
innate immunity or is not cleared by it (as observed with many co-evolved
pathogens), the specific branch of the immune system gets triggered. The advantage
of the adaptive immunity is the ability of specific recognition and selective elimination
of foreign microorganisms and molecules resulting from a tremendous diversity in its
recognition molecules together with the capability to distinguish between self and
non-self (Goldsby et al., 2000). Failure of the latter results in auto-immunity and
destruction of the host. Therefore, with emergence of the adaptive immune system it
is an absolute prerequisite that the T cell repertoire is able to discriminate between
self and non-self (Stefanova et al., 2003; von Boehmer et al., 2003). To ensure this
self-non-self discrimination, thymocytes go through a strict selection process during
their maturation. Thymocytes expressing productively rearranged TCR with weak to
intermediate overall avidity to self-peptide/self-MHC complex receive a survival signal
(positive selection) whereas TCR with high overall avidity to self-peptide/self-MHC
complex are deleted via TCR-induced apoptosis (negative selection) (reviewed in
Benoist and Mathis, 1999). This selection process ensures that all T cells leaving the
thymus are self-MHC-restricted and self-tolerant. Finally, for further survival in the
periphery, it is essential that T cells remain in continuous interaction with self-MHC
(Kirberg et al., 1997; Rocha and von Boehmer, 1991). Furthermore, self-MHC-
restriction gives T cells a greater chance of being activated by non-self-peptide/self-
MHC complex than by non-self-peptide alone or together with non-self-MHC (Bevan,
1977; von Boehmer et al., 1978; Zinkernagel et al., 1978). These observations are
supported by recent data using new methods analysing TCR-MHC interactions and
TCR signalling events (Davis, 2002; Stefanova et al., 2002). Mark Davis et al. show
that MHC surface amino acids accessible for the TCR contribute significantly to the
on-rate of the TCR-MHC interaction whereas the peptide contributes to the off-rate
(Davis, 2002). Thus, TCR self-MHC interaction positions the TCR already in the right
position for future peptide binding and therefore increases the strength and duration

General Discussion
- 75 -
of the future TCR peptide/MHC interaction. Stefanova et al. show that the TCR self-
MHC interaction itself increases the sensitivity of the TCR towards foreign antigen by
constantly transmitting a signalling threshold through partial phosphorylation of the
TCRζ chain, therefore preparing the T cell to respond to a low density of foreign
peptide already early in an infection (Stefanova et al., 2002; Stefanova et al., 2003).
In summary, evolution of a T cell repertoire capable of distinguishing between self
and non-self made it possible to increase the diversity of the T cell repertoire and
therefore the quality of the immune response helping humans and other vertebrates
to eliminate more efficiently even highly evolved pathogens.
WHO?
During the past twenty to thirty years, many experiments have been performed to
analyse which cells are involved in T cell repertoire selection. Until recently the
predominant point of view was that the MHC of radio-resistant cells of the thymus
(presumably thymic epithelial (TE) cells) is responsible for efficient T cell repertoire
selection. This conclusion is based on a series of classical irradiation bone marrow
and thymus chimera experiments (Fink and Bevan, 1978; Kappler and Marrack,
1978; Kisielow et al., 1988b; von Boehmer et al., 1978; Zinkernagel et al., 1978) and
reviewed in Moller, 1978; von Boehmer et al., 2003). For example, reconstitution of
lethally irradiated A-recipients with F1 (AxB) bone marrow and/or F1 (AxB) nude
grafted with a fetal thymus A yield A-restricted but virtually no B-restricted virus-
specific cytotoxic T lymphocytes (CTL) in a primary immune response (Table 7) (Fink
and Bevan, 1978; Zinkernagel et al., 1978). In parallel to this view, some researchers
suggested that the MHC of non-TE cells could also be involved in T cell repertoire
selection (Bix and Raulet, 1992; Doherty and Bennink, 1979; Hugo et al., 1993;
Longo and Schwartz, 1980; Matzinger and Mirkwood, 1978; Wagner et al., 1980;
Zinkernagel et al., 1980). However, these earlier studies were often based on relative
complicated protocols - in vivo filtration experiments, several adoptive transfers into
lethally irradiated recipients (Doherty and Bennink, 1979) – or required several
rounds of in vitro restimulation before detecting any non-TE MHC-restricted
responses (Bix and Raulet, 1992; Hugo et al., 1992; Hugo et al., 1993; Longo and
Schwartz, 1980; Matzinger and Mirkwood, 1978; Pawlowski et al., 1993; Vukmanovic
et al., 1992; Wagner et al., 1980). In addition, the specificity of non-TE MHC-
restricted T cells was not defined at the peptide level, but rather involved activity

General Discussion
- 76 -
against minor histocompatibility antigens (Bix and Raulet, 1992; Matzinger and
Mirkwood, 1978), allo-antigens (Bix and Raulet, 1992; Longo and Schwartz, 1980;
Wagner et al., 1980) and/or synthetic polymers (Bix and Raulet, 1992; Hugo et al.,
1992; Hugo et al., 1993) detected via secondary CTL, mixed lymphocyte reaction
and/or proliferation assays, respectively. In another set of experiments, injection of
allogeneic MHC class II-positive A-fibroblasts or A-thymic epithelial cells into the
thymus of lethally irradiated B-mice reveals restriction to both donor and host MHC
haplotype (Hugo et al., 1992; Hugo et al., 1993; Pawlowski et al., 1993; Vukmanovic
et al., 1992). These results suggest that even fibroblasts not specialized to process
and present antigens in the context of MHC class II molecules can mediate positive
selection when transfected with the appropriate MHC molecule (Hugo et al., 1993).
However, it has to be kept in mind that before intrathymic injection of A-positive cells,
lethally irradiated B-mice were reconstituted with (AxB)-fetal liver cells and therefore
A-positive cells are present both in the thymus and in the periphery allowing positive
selection and further survival of A-restricted T cells. Surprisingly, Zinkernagel et al.
show that nude mice reconstituted with a completely allogeneic day-14 fetal thymus
graft develop virus-specific primary immune response restricted almost exclusively to
the nude- but not to the thymus graft-MHC haplotype (Table 7) (Zinkernagel et al.,
1980). These results were again not entirely accepted because of potential rescue of
the nude thymic rudiment although recent data using tetraparental chimeras between
euthymic and nude donors of distinct MHC haplotype reveals that the thymic
rudiment of the nude donor cannot be rescued anatomically to form mature thymic
epithelial cells (Blackburn et al., 1996; Rodewald et al., 2001). Taken together, the
above-discussed experiments were not able to define satisfactorily the role of non-TE
MHC in T cell repertoire selection and survival. Therefore, in order to clarify the role
of TE versus non-TE MHC in T cell repertoire selection and survival, tetraparental
aggregation chimeras from T and B cell-deficient mice (H-2d Scid or H-2b Rag-/-) and
thymus-deficient nude mice (H-2b or H-2d) were generated (Results Part I) (Martinic
et al., 2003). Analysis of these relatively undisturbed (non-irradiated and non-
reconstituted) tetraparental aggregation chimeras confirmed the absence of rescue of
nude thymic rudiment (Results Part I) (Martinic et al., 2003). After viral infection,
tetraparental aggregation chimeras revealed primary protective CD8+ T cell
responses that were peptide-specifically restricted to either TE or non-TE MHC at
comparable levels (Results Part I) (Martinic et al., 2003). Furthermore, protective

General Discussion
- 77 -
virus-neutralizing IgG responses dependent on cognate CD4+ T helper cells
restricted to non-TE MHC were also mounted (Results Part I) (Martinic et al., 2003).
Therefore, these results showed convincingly that non-TE MHC was sufficient to
select a functional T cell repertoire (Results Part I) (Martinic et al., 2003).
Table 7: On the role of thymic epithelial (TE) versus non-TE MHC in T cell repertoire selection.
WHERE and WHEN?
In addition to the now obtained knowledge with respect to which cells are involved in
T cell repertoire selection, the next step is to analyse whether there exists a specific
location and a time-frame for most efficient T cell repertoire selection. Unfortunately it
was not possible to design any experiment where the periphery could be observed
independently from the thymus, therefore the focus is set on both thymus and
periphery simultaneously. Based on previously published data and results obtained
here, T cell repertoire selection and survival of positively selected T cells is most
efficient when TCR-restricting MHC-expressing cells are present both in the thymus
and in the periphery at all times (Table 7). This is shown by the following examples:
Reconstitution of lethally irradiated A-mice with (AxB)-bone marrow leads to survival

General Discussion
- 78 -
of A- but not B-restricted cells (Table 7) (Fink and Bevan, 1978; Longo and Schwartz,
1980; Zinkernagel et al., 1978). In these mice, MHCA is present in both the thymus
and the periphery at all times, MHCB however is present initially exclusively in the
periphery and only after a few months BM-derived MHCB-positive cells will have
migrated into the thymus (Longo and Schwartz, 1980). By that time it is too late for
selection and survival of B-restricted T cells because A-restricted T cells have
already outnumbered B-restricted T cells and therefore only A-restricted T cells are
detected in the periphery (Lemischka et al., 1986; Longo and Schwartz, 1980;
Zinkernagel, 1982). Interestingly, this exclusive restriction to MHCA will change if
these mice are depleted of thymocytes and mature T cells eight months after BM-
reconstitution and allowed to replenish the T cell pool. Analysis of these mice three
months after T cell depletion reveals an A- and B-restricted T cell repertoire (Longo
and Schwartz, 1980) confirming the here developed hypothesis that survival of MHC-
restricted T cells is only guaranteed if TCR-restricting MHC-expressing cells are
present both in the thymus and in the periphery from the start. However, it remains to
say that Longo et al. do not use assays analysing defined antigen-specificity but
rather proliferation measurements or mixed lymphocyte reaction assays. In addition,
A- and B-restricted responses are never obtained ex vivo but only after in vitro
restimulation. Therefore, it might be interesting to repeat above-discussed
experiments but this time using defined antigens and more sensitive assays (i.e.
tetramer staining). Finally, with the here obtained results, it might be interesting to
readdress the question whether MHC class II positive haematopoietic cells can
mediate positive selection in MHC class II deficient mice (Markowitz et al., 1993).
Markowitz et al. show that lethally irradiated MHC class II-deficient mice reconstituted
with bone marrow from MHC class II-positive donors or nude mice grafted with
irradiated MHC class II-deficient thymus do not show positive selection and survival
of CD4+ cells. Positive selection and survival of CD4+ cells is only observed when TE
cells express MHC class II molecules (Markowitz et al., 1993). It would be interesting
to analyse what would happen if donor-MHC would be present both in the thymus
and in the periphery. This can be easily achieved if eight months after reconstitution
with bone marrow from MHC class II-positive donors, the peripheral T cell pool and
all thymocytes would be eliminated by cortisone and anti-thymocyte-serum treatment
(similar to the experiments performed by Longo et al. (Longo and Schwartz, 1980).
Three months after T cell repertoire depletion, newly emerging T cells would

General Discussion
- 79 -
encounter donor-APC (MHC class II-positive) already in the thymus, could therefore
be positively selected on donor-MHC, followed by commitment to the CD4 T cell
lineage and emigration into the periphery. There, further survival would be
guaranteed through continuous interaction with donor-MHC. Another way to test for
the ability of bone marrow-derived MHC class II-positive cells to positively select DP
thymocytes would be analysis of H-2b Rag-/- MHC class II-/- ↔ nuded tetraparental
aggregation chimeras. In these mice, all TE cells are MHC class II-deficient and only
professional APC from nuded donor are MHC class II-positive. In these mice but not
in the irradiation chimeras from Markowitz et al, MHC class II-positive cells are
present from the start both in the thymus and in the periphery. If after LCMV-infection
or VSV-infection, tetraparental aggregation chimeras are able to mount virus-specific
CD4-dependent IgG responses, haematopoietic cells are efficient in selecting CD4+ T
cells. If, however, these mice show a reduced CD4+ T cell response this might be
explained by the overall reduced density of MHC class II molecules in these
tetraparental aggregation chimeras as already observed by different labs for MHC
class I-deficient irradiation chimeras (Bix and Raulet, 1992; Terra et al., 2002;
Zerrahn et al., 1999).
Another example with nudeA mice grafted with fetal thymusB reveals survival of A-
restricted T cells (Table 7) (Zinkernagel et al., 1980). Again, MHCA-expressing cells
are the only cells present, both in the periphery and in the thymus, whereas MHCB-
expressing cells are only present in the thymus and therefore MHCB-restricted T cells
have no chance to survive further on in the periphery.
Finally, analysis of Rag-/-A ↔ nudeB and ScidB ↔ nudeA tetraparental aggregation
chimeras where both MHCA and MHCB-expressing cells are present from the start in
the thymus and in the periphery show survival of both A- and B-restricted T cells
confirming the above formulated hypothesis (Table 7) (Martinic et al., 2003).
Furthermore, these results are in agreement with data obtained from transplantation
studies. Mice with a high degree of haematopoietic chimerism showing host- and
donor-derived cells, both in the thymus and in the periphery, reveal complete allograft
tolerance and show both host- and donor-restricted T cell response after LCMV- or
VSV-infection (Adams et al., 2001; Williams et al., 2003). In another study, mice
receiving syngeneic thymus graft show presence of syngeneic MHC class II-
restricted CD4+ cells only if syngeneic MHC class II is found also in the periphery
(Rodriguez-Barbosa et al., 2002). Surprisingly, mice receiving xenogeneic porcine

General Discussion
- 80 -
thymus graft show presence of porcine MHC class II-restricted CD4+ cells even in
absence of porcine MHC class II in the periphery. However, this is only possible
because of the presence of murine MHC class II IAb in the periphery, which shows
highly conserved amino acid sequence with porcine MHC class II DQd probably
substituting therefore the need for porcine MHC class II (Rodriguez-Barbosa et al.,
2002).
HOW?
It is well known and accepted that the most efficient site for initial TCR
rearrangement and maturation is the thymus itself (Bach, 1979; Goldstein and White,
1971; Miller, 1961). The thymus provides an excellent three-dimensional
microenvironment including compartmentalization of maturing DP and SP
thymocytes and therefore optimal cytokine milieu for each individual compartment, a
physical barrier between the thymus and the periphery protecting maturing
thymocytes from the peripheral environment (optimised for efficient T cell activation
and differentiation but not for maturation) and preventing the re-entry of mature
thymocytes into the thymus (impeding therefore possible competition for certain
cytokines), and finally presence of professional APC with the unique capability of
promiscuous expression of peripheral self-antigens enabling negative selection of
thymocytes with high overall avidity against peripheral self-antigens (Anderson et al.,
2002). It was therefore interesting to analyse the efficiency of T cell repertoire
selection and survival under sub-optimal conditions, for example in the presence of
reduced TCR-restricting MHC density (Results Part II) or in the absence of a thymus
(Results Part III). Selection and survival of the low-affinity H-Y-specific transgenic
TCR but not of the high-affinity 2C transgenic TCR were less efficient under reduced
TCR-restricting MHC density (Results Part II) (Zerrahn et al., 1999). In an athymic
environment, positive selection and survival of the H-Y-specific transgenic TCR in
females were even more drastically reduced (Results Part III). Absence of self-
reactive H-Y-specific transgenic TCR in athymic males, however, was complete
(Results Part III). Other labs studying T cell repertoire selection and survival under
sub-optimal conditions have obtained similar results (Terra et al., 2002) and Perreault
et al., manuscript in preparation). Perreault et al, for example, show reduced positive
selection and survival of the H-Y-specific transgenic TCR in double transgenic H-
Y/LckOM females but unimpaired positive selection and survival of the 2C transgenic

General Discussion
- 81 -
TCR in 2C/LckOM double transgenic mice (Terra et al., 2002). However, the overall
TCR signalling strength in these mice represented by CD5 expression levels in DP
and SP mesenteric lymph node cells is reduced when compared to the ones in DP
and SP thymocytes of H-Y or 2C TCR single transgenic mice (Terra et al., 2002). To
compare the immune response of thymically derived with the one of extrathymically
derived T cells, C57BL/6 control mice and adult thymectomized lethally irradiated
Rag2-/- mice reconstituted with fetal liver of LckOM transgenic mice were infected
with LCMV and VSV (Perreault et al., manuscript in preparation). Only few of the
extrathymically derived T cells were able to show some cytotoxic activity against
LCMV-derived epitopes after in vitro restimulation and only few were able to produce
partial VSV-specific IgM and very low IgG titers. Nevertheless, none of the mice with
extrathymically derived T cells were able to clear LCMV or VSV-infection (Perreault
et al., manuscript in preparation). Taken together, these results show that although
protection against self-reactivity is still guaranteed under sub-optimal extrathymic
conditions, efficiency of positive selection is so reduced that it is not sufficient for full
immunocompetence. Just recently Guy-Grand et al. analysed extrathymic
lymphopoiesis in detail in athymic and euthymic mice (Guy-Grand et al., 2003). As
already seen in LckOM transgenic mice, mesenteric lymph nodes (mLN) are the
major site and Peyer’s patches (PP) a minor site of extrathymic lymphopoiesis (Guy-
Grand et al., 2003). However, in contradiction to previous observations (Poussier and
Julius, 1994; Rocha et al., 1991; Saito et al., 1998), the cryptopatches and the gut
epithelium prove rather inefficient (Guy-Grand et al., 2003). Guy-Grand also
concludes that the reduced efficiency of selection and survival of an extrathymically
derived T cell repertoire is due to the sub-optimal athymic environment. As an
example, Guy-Grand shows that the absence of CD8αα+ cells in athymic mice is due
to the lack of the right self-peptide/MHC complexes and the absence of the right
cytokine milieu (lack of IL-7 and IL-15) for survival and expansion of CD8αα+
precursors (DN TCRαβ+ NK1.1- cells which are present in the thymus of euthymic
mice) in the mLN (Guy-Grand et al., 2003). In euthymic mice, both specific
peptide/MHC complexes and the right cytokine milieu are provided in the thymus and
therefore CD8αα+ cells are present (Guy-Grand et al., 2003). In conclusion, in
healthy euthymic mice, extrathymic lymphopoiesis is absent. It can resume, however,
in conditions of severe lymphocyte depletion, absence of TCRαβ+ cells or marked

General Discussion
- 82 -
thymic atrophy to ensure at least some T cell repertoire selection and survival (Guy-
Grand et al., 2003).
In summary, optimal conditions (presence of a thymus and high TCR-restricting MHC
density present at all times throughout the whole body) lead to efficient selection and
survival of a self-MHC-restricted and self-tolerant T cell repertoire capable to
generate immune responses against all kinds of pathogens. Trying to understand
more in detail the exact mechanisms governing this selection and survival process,
might help us to envisage new strategies for improved therapies against autoimmune
diseases and for successful organ transplantations.

References
- 83 -
9 References
Adams, A. B., Durham, M. M., Kean, L., Shirasugi, N., Ha, J., Williams, M. A., Rees,P. A., Cheung, M. C., Mittelstaedt, S., Bingaman, A. W., et al. (2001). Costimulationblockade, busulfan, and bone marrow promote titratable macrochimerism, inducetransplantation tolerance, and correct genetic hemoglobinopathies with minimalmyelosuppression. J Immunol 167, 1103-1111.
Akashi, K., Reya, T., Dalma-Weiszhausz, D., and Weissman, I. L. (2000). Lymphoidprecursors. Curr Opin Immunol 12, 144-150.
Anderson, G., and Jenkinson, E. J. (2001). Lymphostromal interactions in thymicdevelopment and function. Nat Rev Immunol 1, 31-40.
Anderson, G., Moore, N. C., Owen, J. J., and Jenkinson, E. J. (1996). Cellularinteractions in thymocyte development. Annu Rev Immunol 14, 73-99.
Anderson, M. S., Venanzi, E. S., Klein, L., Chen, Z., Berzins, S. P., Turley, S. J., vonBoehmer, H., Bronson, R., Dierich, A., Benoist, C., and Mathis, D. (2002). Projectionof an immunological self shadow within the thymus by the aire protein. Science 298,1395-1401.
Apostolou, I., Sarukhan, A., Klein, L., and von Boehmer, H. (2002). Origin ofregulatory T cells with known specificity for antigen. Nat Immunol 3, 756-763.
Ashton-Rickardt, P. G., Bandeira, A., Delaney, J. R., Van Kaer, L., Pircher, H. P.,Zinkernagel, R. M., and Tonegawa, S. (1994). Evidence for a differential aviditymodel of T cell selection in the thymus. Cell 76, 651-663.
Bach, J. F. (1979). Thymic hormones. J Immunopharmacol 1, 277-310.
Bachmann, M. F., Kundig, T. M., Kalberer, C. P., Hengartner, H., and Zinkernagel, R.M. (1993). Formalin inactivation of vesicular stomatitis virus impairs T-cell- but not T-help-independent B-cell responses. J Virol 67, 3917-3922.
Battegay, M., Moskophidis, D., Waldner, H., Brundler, M. A., Fung-Leung, W. P.,Mak, T. W., Hengartner, H., and Zinkernagel, R. M. (1993). Impairment and delay ofneutralizing antiviral antibody responses by virus-specific cytotoxic T cells [publishederratum appears in J Immunol 1994 Mar 1;152(5):1635]. J Immunol 151, 5408-5415.
Bendelac, A., Bonneville, M., and Kearney, J. F. (2001). Autoreactivity by design:innate B and T lymphocytes. Nat Rev Immunol 1, 177-186.
Bendelac, A., Hunziker, R. D., and Lantz, O. (1996). Increased interleukin 4 andimmunoglobulin E production in transgenic mice overexpressing NK1 T cells. J ExpMed 184, 1285-1293.
Bendelac, A., Rivera, M. N., Park, S. H., and Roark, J. H. (1997). Mouse CD1-specific NK1 T cells: development, specificity, and function. Annu Rev Immunol 15,535-562.
Benlagha, K., Kyin, T., Beavis, A., Teyton, L., and Bendelac, A. (2002). A thymicprecursor to the NK T cell lineage. Science 296, 553-555.

References
- 84 -
Benoist, C., and Mathis, D. (1999). T-lymphocyte differentiation and biology. InFundamental Immunology. W. E. Paul, ed., Fourth edn (Philadelphia, Lippincott-Raven Publishers).
Bensinger, S. J., Bandeira, A., Jordan, M. S., Caton, A. J., and Laufer, T. M. (2001).Major histocompatibility complex class II-positive cortical epithelium mediates theselection of CD4(+)25(+) immunoregulatory T cells. J Exp Med 194, 427-438.
Bevan, M. J. (1977). In a radiation chimaera, host H-2 antigens determine immuneresponsiveness of donor cytotoxic cells. Nature 269, 417-418.
Bix, M., and Raulet, D. (1992). Inefficient positive selection of T cells directed byhaematopoietic cells. Nature 359, 330-333.
Blackburn, C. C., Augustine, C. L., Li, R., Harvey, R. P., Malin, M. A., Boyd, R. L.,Miller, J. F., and Morahan, G. (1996). The nu gene acts cell-autonomously and isrequired for differentiation of thymic epithelial progenitors. Proc Natl Acad Sci U S A93, 5742-5746.
Boileau, C., Houde, M., Dulude, G., Clegg, C. H., and Perreault, C. (2000).Regulation of extrathymic T cell development and turnover by oncostatin M. JImmunol 164, 5713-5720.
Borgulya, P., Kishi, H., Uematsu, Y., and von Boehmer, H. (1992). Exclusion andinclusion of alpha and beta T cell receptor alleles. Cell 69, 529-537.
Borowski, C., Martin, C., Gounari, F., Haughn, L., Aifantis, I., Grassi, F., and vonBoehmer, H. (2002). On the brink of becoming a T cell. Curr Opin Immunol 14, 200-206.
Brandle, D., Muller, C., Rulicke, T., Hengartner, H., and Pircher, H. (1992).Engagement of the T-cell receptor during positive selection in the thymus down-regulates RAG-1 expression. Proc Natl Acad Sci U S A 89, 9529-9533.
Brugnera, E., Bhandoola, A., Cibotti, R., Yu, Q., Guinter, T. I., Yamashita, Y.,Sharrow, S. O., and Singer, A. (2000). Coreceptor reversal in the thymus: signaledCD4+8+ thymocytes initially terminate CD8 transcription even when differentiatinginto CD8+ T cells. Immunity 13, 59-71.
Bruno, L., Fehling, H. J., and von Boehmer, H. (1996). The alpha beta T cell receptorcan replace the gamma delta receptor in the development of gamma delta lineagecells. Immunity 5, 343-352.
Buch, T., Rieux-Laucat, F., Forster, I., and Rajewsky, K. (2002). Failure of HY-specific thymocytes to escape negative selection by receptor editing. Immunity 16,707-718.
Capone, M., Troesch, M., Eberl, G., Hausmann, B., Palmer, E., and MacDonald, H.R. (2001). A critical role for the T cell receptor alpha-chain connecting peptidedomain in positive selection of CD1-independent NKT cells. Eur J Immunol 31, 1867-1875.
Casanova, J. L., Romero, P., Widmann, C., Kourilsky, P., and Maryanski, J. L.(1991). T cell receptor genes in a series of class I major histocompatibility complex-restricted cytotoxic T lymphocyte clones specific for a Plasmodium bergheinonapeptide: implications for T cell allelic exclusion and antigen-specific repertoire. JExp Med 174, 1371-1383.

References
- 85 -
Casellas, R., Shih, T. A., Kleinewietfeld, M., Rakonjac, J., Nemazee, D., Rajewsky,K., and Nussenzweig, M. C. (2001). Contribution of receptor editing to the antibodyrepertoire. Science 291, 1541-1544.
Chaffin, K. E., and Perlmutter, R. M. (1991). A pertussis toxin-sensitive processcontrols thymocyte emigration. Eur J Immunol 21, 2565-2573.
Charan, S., and Zinkernagel, R. M. (1986). Antibody mediated suppression ofsecondary IgM response in nude mice against vesicular stomatitis virus. J Immunol136, 3057-3061.
Clegg, C. H., Rulffes, J. T., Wallace, P. M., and Haugen, H. S. (1996). Regulation ofan extrathymic T-cell development pathway by oncostatin M. Nature 384, 261-263.
Constant, P., Davodeau, F., Peyrat, M. A., Poquet, Y., Puzo, G., Bonneville, M., andFournie, J. J. (1994). Stimulation of human gamma delta T cells by nonpeptidicmycobacterial ligands. Science 264, 267-270.
Cruz, D., Sydora, B. C., Hetzel, K., Yakoub, G., Kronenberg, M., and Cheroutre, H.(1998). An opposite pattern of selection of a single T cell antigen receptor in thethymus and among intraepithelial lymphocytes. J Exp Med 188, 255-265.
Cumano, A., Paige, C. J., Iscove, N. N., and Brady, G. (1992). Bipotential precursorsof B cells and macrophages in murine fetal liver. Nature 356, 612-615.
Curnow, S. J., Boyer, C., Buferne, M., and Schmitt-Verhulst, A. M. (1995). TCR-associated zeta-Fc epsilon RI gamma heterodimers on CD4-CD8- NK1.1+ T cellsselected by specific class I MHC antigen. Immunity 3, 427-438.
Davis, M. (2002). Gene expression and signaling in the immunal system. Paperpresented at: Cold Spring Harbour Symposium (Cold Spring Harbour).
Doherty, P. C., and Bennink, J. C. (1979). Vaccinia-specific cytotoxic T-cellresponses in the context of H-2 antigens not encountered in thymus may reflectaberrant recognition of a virus-H-2 complex. J Exp Med 149, 150-157.
Eppig, J. J., Kozak, L. P., Eicher, E. M., and Stevens, L. C. (1977). Ovarianteratomas in mice are derived from oocytes that have completed the first meioticdivision. Nature 269, 517-518.
Fink, P. J., and Bevan, M. J. (1978). H-2 antigens of the thymus determinelymphocyte specificity. J Exp Med 148, 766-775.
Fink, P. J., and McMahan, C. J. (2000). Lymphocytes rearrange, edit and revise theirantigen receptors to be useful yet safe. Immunol Today 21, 561-566.
Goldsby, R. A., Kindt, T. J., and Osborne, B. A. (2000). Kuby immunology, Fourthedn (New York, W. H. Freeman and Company).
Goldstein, A. L., and White, A. (1971). Role of thymosin and other thymic factors inthe development, maturation, and functions of lymphoid tissue. Curr Top ExpEndocrinol 1, 121-149.
Guy-Grand, D., Azogui, O., Celli, S., Darche, S., Nussenzweig, M. C., Kourilsky, P.,and Vassalli, P. (2003). Extrathymic T cell lymphopoiesis: ontogeny and contributionto gut intraepithelial lymphocytes in athymic and euthymic mice. J Exp Med 197, 333-341.

References
- 86 -
Guy-Grand, D., Cuenod-Jabri, B., Malassis-Seris, M., Selz, F., and Vassalli, P.(1996). Complexity of the mouse gut T cell immune system: identification of twodistinct natural killer T cell intraepithelial lineages. Eur J Immunol 26, 2248-2256.
Guy-Grand, D., Rocha, B., Mintz, P., Malassis-Seris, M., Selz, F., Malissen, B., andVassalli, P. (1994). Different use of T cell receptor transducing modules in twopopulations of gut intraepithelial lymphocytes are related to distinct pathways of T celldifferentiation. J Exp Med 180, 673-679.
Hayday, A., Theodoridis, E., Ramsburg, E., and Shires, J. (2001). Intraepitheliallymphocytes: exploring the Third Way in immunology. Nat Immunol 2, 997-1003.
Heath, W. R., Carbone, F. R., Bertolino, P., Kelly, J., Cose, S., and Miller, J. F.(1995). Expression of two T cell receptor alpha chains on the surface of normalmurine T cells. Eur J Immunol 25, 1617-1623.
Hiromatsu, K., Yoshikai, Y., Matsuzaki, G., Ohga, S., Muramori, K., Matsumoto, K.,Bluestone, J. A., and Nomoto, K. (1992). A protective role of gamma/delta T cells inprimary infection with Listeria monocytogenes in mice. J Exp Med 175, 49-56.
Holub, M., Rossmann, P., Tlaskalova, H., and Vidmarova, H. (1975). Thymusrudiment of the athymic nude mouse. Nature 256, 491-493.
Hsu, C., Whitney, R. A., Jr., and Hansen, C. T. (1975). Thymus-like lymph node innude mice. Nature 257, 681-682.
Huesmann, M., Scott, B., Kisielow, P., and von Boehmer, H. (1991). Kinetics andefficacy of positive selection in the thymus of normal and T cell receptor transgenicmice. Cell 66, 533-540.
Hugo, P., Kappler, J. W., Godfrey, D. I., and Marrack, P. C. (1992). A cell line thatcan induce thymocyte positive selection. Nature 360, 679-682.
Hugo, P., Kappler, J. W., McCormack, J. E., and Marrack, P. (1993). Fibroblasts caninduce thymocyte positive selection in vivo. Proc Natl Acad Sci U S A 90, 10335-10339.
Irving, B. A., Alt, F. W., and Killeen, N. (1998). Thymocyte development in theabsence of pre-T cell receptor extracellular immunoglobulin domains. Science 280,905-908.
Jordan, M. S., Boesteanu, A., Reed, A. J., Petrone, A. L., Holenbeck, A. E., Lerman,M. A., Naji, A., and Caton, A. J. (2001). Thymic selection of CD4+CD25+ regulatory Tcells induced by an agonist self-peptide. Nat Immunol 2, 301-306.
Jordan, M. S., Riley, M. P., von Boehmer, H., and Caton, A. J. (2000). Anergy andsuppression regulate CD4(+) T cell responses to a self peptide. Eur J Immunol 30,136-144.
Kappler, J. W., and Marrack, P. (1978). The role of H-2 linked genes in helper T-cellfunction. IV. Importance of T-cell genotype and host environment in I-region and Irgene expression. J Exp Med 148, 1510-1522.
Kawamoto, H., Ikawa, T., Ohmura, K., Fujimoto, S., and Katsura, Y. (2000). T cellprogenitors emerge earlier than B cell progenitors in the murine fetal liver. Immunity12, 441-450.

References
- 87 -
Kawamoto, H., Ohmura, K., and Katsura, Y. (1997). Direct evidence for thecommitment of hematopoietic stem cells to T, B and myeloid lineages in murine fetalliver. Int Immunol 9, 1011-1019.
Kindred, B. (1979). Nude mice in immunology. Prog Allergy 26, 137-238.
Kirberg, J., Berns, A., and von Boehmer, H. (1997). Peripheral T cell survival requirescontinual ligation of the T cell receptor to major histocompatibility complex-encodedmolecules. J Exp Med 186, 1269-1275.
Kisielow, P., Bluthmann, H., Staerz, U. D., Steinmetz, M., and von Boehmer, H.(1988a). Tolerance in T-cell-receptor transgenic mice involves deletion of nonmatureCD4+8+ thymocytes. Nature 333, 742-746.
Kisielow, P., Teh, H. S., Bluthmann, H., and von Boehmer, H. (1988b). Positiveselection of antigen-specific T cells in thymus by restricting MHC molecules. Nature335, 730-733.
Kyburz, D., Aichele, P., Speiser, D. E., Hengartner, H., Zinkernagel, R. M., andPircher, H. (1993). T cell immunity after a viral infection versus T cell toleranceinduced by soluble viral peptides. Eur J Immunol 23, 1956-1962.
Ladel, C. H., Blum, C., Dreher, A., Reifenberg, K., and Kaufmann, S. H. (1995).Protective role of gamma/delta T cells and alpha/beta T cells in tuberculosis. Eur JImmunol 25, 2877-2881.
Le Lievre, C. S., and Le Douarin, N. M. (1975). Mesenchymal derivatives of theneural crest: analysis of chimaeric quail and chick embryos. J Embryol Exp Morphol34, 125-154.
Leist, T. P., Cobbold, S. P., Waldmann, H., Aguet, M., and Zinkernagel, R. M. (1987).Functional analysis of T lymphocyte subsets in antiviral host defense. J Immunol 138,2278-2281.
Lemischka, I. R., Raulet, D. H., and Mulligan, R. C. (1986). Developmental potentialand dynamic behavior of hematopoietic stem cells. Cell 45, 917-927.
Levelt, C. N., de Jong, Y. P., Mizoguchi, E., O'Farrelly, C., Bhan, A. K., Tonegawa,S., Terhorst, C., and Simpson, S. J. (1999). High- and low-affinity single-peptide/MHC ligands have distinct effects on the development of mucosalCD8alphaalpha and CD8alphabeta T lymphocytes. Proc Natl Acad Sci U S A 96,5628-5633.
Longo, D. L., and Schwartz, R. H. (1980). T-cell specificity for H-2 and Ir genephenotype correlates with the phenotype of thymic antigen-presenting cells. Nature287, 44-46.
MacDonald, H. R., Radtke, F., and Wilson, A. (2001). T cell fate specification andalphabeta/gammadelta lineage commitment. Curr Opin Immunol 13, 219-224.
Markowitz, J. S., Auchincloss, H., Jr., Grusby, M. J., and Glimcher, L. H. (1993).Class II-positive hematopoietic cells cannot mediate positive selection of CD4+ Tlymphocytes in class II-deficient mice. Proc Natl Acad Sci U S A 90, 2779-2783.
Martin, S., and Bevan, M. J. (1997). Antigen-specific and nonspecific deletion ofimmature cortical thymocytes caused by antigen injection. Eur J Immunol 27, 2726-2736.

References
- 88 -
Martinic, M. M., Hengartner, H., and Zinkernagel, R. M. (Manuscript in preparation-a).Influence of MHC class I H-2Db density on selection and survival of H-Y-specificTCR transgenic T cells.
Martinic, M. M., Hengartner, H., and Zinkernagel, R. M. (Manuscript in preparation-b).Selection of the H-Y-specific transgenic TCR in an athymic versus euthymicenvironment.
Martinic, M. M., Rulicke, T., Althage, A., Odermatt, B., Hochli, M., Lamarre, A.,Dumrese, T., Speiser, D. E., Kyburz, D., Hengartner, H., and Zinkernagel, R. M.(2003). Efficient T cell repertoire selection in tetraparental chimeric mice independentof thymic epithelial MHC. Proc Natl Acad Sci U S A 100, 1861-1866.
Matzinger, P. (1993). Why positive selection? Immunol Rev 135, 81-117.
Matzinger, P., and Mirkwood, G. (1978). In a fully H-2 incompatible chimera, T cellsof donor origin can respond to minor histocompatibility antigens in association witheither donor or host H-2 type. J Exp Med 148, 84-92.
McCaren, L. C., Holland, J. J., and Syverton, J. T. (1959). The mammalian cell-virusrelationship: I. Attachment of poliovirus to cultivated cells of primate and non-primateorigin. J Exp Med 109, 475-485.
Merkenschlager, M., Benoist, C., and Mathis, D. (1994). Evidence for a single-nichemodel of positive selection. Proc Natl Acad Sci U S A 91, 11694-11698.
Miller, J. (1961). Immunological function of the thymus. Lancet 2, 748-749.
Modigliani, Y., Bandeira, A., and Coutinho, A. (1996). A model for developmentallyacquired thymus-dependent tolerance to central and peripheral antigens. ImmunolRev 149, 155-120.
Moller, G. e. (1978). Acquisition of the T cell Repertoire. Immunol Rev 42, 3-270.
Mombaerts, P., Arnoldi, J., Russ, F., Tonegawa, S., and Kaufmann, S. H. (1993).Different roles of alpha beta and gamma delta T cells in immunity against anintracellular bacterial pathogen. Nature 365, 53-56.
Nemazee, D. (2000). Receptor selection in B and T lymphocytes. Annu Rev Immunol18, 19-51.
Ochsenbein, A. F., Pinschewer, D. D., Odermatt, B., Ciurea, A., Hengartner, H., andZinkernagel, R. M. (2000). Correlation of T cell independence of antibody responseswith antigen dose reaching secondary lymphoid organs: implications forsplenectomized patients and vaccine design. J Immunol 164, 6296-6302.
Ohashi, P. S. (2002). T-cell signalling and autoimmunity: molecular mechanisms ofdisease. Nat Rev Immunol 2, 427-438.
Orange, J. S., Salazar-Mather, T. P., Opal, S. M., Spencer, R. L., Miller, A. H.,McEwen, B. S., and Biron, C. A. (1995). Mechanism of interleukin 12-mediatedtoxicities during experimental viral infections: role of tumor necrosis factor andglucocorticoids. J Exp Med 181, 901-914.
Osborne, B., and Miele, L. (1999). Notch and the immune system. Immunity 11, 653-663.

References
- 89 -
Park, S. Y., Arase, H., Wakizaka, K., Hirayama, N., Masaki, S., Sato, S., Ravetch, J.V., and Saito, T. (1995). Differential contribution of the FcR gamma chain to thesurface expression of the T cell receptor among T cells localized in epithelia: analysisof FcR gamma-deficient mice. Eur J Immunol 25, 2107-2110.
Pawlowski, T., Elliott, J. D., Loh, D. Y., and Staerz, U. D. (1993). Positive selection ofT lymphocytes on fibroblasts. Nature 364, 642-645.
Petrie, H. T., Scollay, R., and Shortman, K. (1992). Commitment to the T cellreceptor-alpha beta or -gamma delta lineages can occur just prior to the onset ofCD4 and CD8 expression among immature thymocytes. Eur J Immunol 22, 2185-2188.
Pfeffer, K., Schoel, B., Gulle, H., Kaufmann, S. H., and Wagner, H. (1990). Primaryresponses of human T cells to mycobacteria: a frequent set of gamma/delta T cellsare stimulated by protease-resistant ligands. Eur J Immunol 20, 1175-1179.
Pircher, H., Burki, K., Lang, R., Hengartner, H., and Zinkernagel, R. M. (1989).Tolerance induction in double specific T-cell receptor transgenic mice varies withantigen. Nature 342, 559-561.
Podd, B. S., Aberg, C., Kudla, K. L., Keene, L., Tobias, E., and Camerini, V. (2001).MHC class I allele dosage alters CD8 expression by intestinal intraepitheliallymphocytes. J Immunol 167, 2561-2568.
Poussier, P., and Julius, M. (1994). Thymus independent T cell development andselection in the intestinal epithelium. Annu Rev Immunol 12, 521-553.
Read, S., and Powrie, F. (2001). CD4(+) regulatory T cells. Curr Opin Immunol 13,644-649.
Rocha, B., Vassalli, P., and Guy-Grand, D. (1991). The V beta repertoire of mousegut homodimeric alpha CD8+ intraepithelial T cell receptor alpha/beta + lymphocytesreveals a major extrathymic pathway of T cell differentiation. J Exp Med 173, 483-486.
Rocha, B., and von Boehmer, H. (1991). Peripheral selection of the T cell repertoire.Science 251, 1225-1228.
Rocha, B., von Boehmer, H., and Guy-Grand, D. (1992). Selection of intraepitheliallymphocytes with CD8 alpha/alpha co-receptors by self-antigen in the murine gut.Proc Natl Acad Sci U S A 89, 5336-5340.
Rodewald, H. R., Ogawa, M., Haller, C., Waskow, C., and DiSanto, J. P. (1997). Pro-thymocyte expansion by c-kit and the common cytokine receptor gamma chain isessential for repertoire formation. Immunity 6, 265-272.
Rodewald, H. R., Paul, S., Haller, C., Bluethmann, H., and Blum, C. (2001). Thymusmedulla consisting of epithelial islets each derived from a single progenitor. Nature414, 763-768.
Rodriguez-Barbosa, J. I., Zhao, Y., Zhao, G., Ezquerra, A., and Sykes, M. (2002).Murine CD4 T cells selected in a highly disparate xenogeneic porcine thymus graftdo not show rapid decay in the absence of selecting MHC in the periphery. JImmunol 169, 6697-6710.

References
- 90 -
Sagara, S., Sugaya, K., Tokoro, Y., Tanaka, S., Takano, H., Kodama, H., Nakauchi,H., and Takahama, Y. (1997). B220 expression by T lymphoid progenitor cells inmouse fetal liver. J Immunol 158, 666-676.
Saint-Ruf, C., Panigada, M., Azogui, O., Debey, P., von Boehmer, H., and Grassi, F.(2000). Different initiation of pre-TCR and gammadeltaTCR signalling. Nature 406,524-527.
Saint-Ruf, C., Ungewiss, K., Groettrup, M., Bruno, L., Fehling, H. J., and vonBoehmer, H. (1994). Analysis and expression of a cloned pre-T cell receptor gene.Science 266, 1208-1212.
Saito, H., Kanamori, Y., Takemori, T., Nariuchi, H., Kubota, E., Takahashi-Iwanaga,H., Iwanaga, T., and Ishikawa, H. (1998). Generation of intestinal T cells fromprogenitors residing in gut cryptopatches. Science 280, 275-278.
Schild, H., Mavaddat, N., Litzenberger, C., Ehrich, E. W., Davis, M. M., Bluestone, J.A., Matis, L., Draper, R. K., and Chien, Y. H. (1994). The nature of majorhistocompatibility complex recognition by gamma delta T cells. Cell 76, 29-37.
Sebzda, E., Mariathasan, S., Ohteki, T., Jones, R., Bachmann, M. F., and Ohashi, P.S. (1999). Selection of the T cell repertoire. Annu Rev Immunol 17, 829-874.
Sha, W. C., Nelson, C. A., Newberry, R. D., Kranz, D. M., Russell, J. H., and Loh, D.Y. (1988). Selective expression of an antigen receptor on CD8-bearing Tlymphocytes in transgenic mice. Nature 335, 271-274.
Simpson, E., Scott, D., and Chandler, P. (1997). The male-specific histocompatibilityantigen, H-Y: a history of transplantation, immune response genes, sexdetermination and expression cloning. Annu Rev Immunol 15, 39-61.
Singer, A. (1988). Experimentation and thymic selection. J Immunol 140, 2481-2483.
Singer, A. (2002). New perspectives on a developmental dilemma: the kineticsignaling model and the importance of signal duration for the CD4/CD8 lineagedecision. Curr Opin Immunol 14, 207-215.
Speiser, D. E., Stubi, U., and Zinkernagel, R. M. (1992). Extrathymic positiveselection of alpha beta T-cell precursors in nude mice. Nature 355, 170-172.
Starr, T. K., Jameson, S. C., and Hogquist, K. A. (2002). Positive and Negative T CellSelection. Annu Rev Immunol.
Stefanova, I., Dorfman, J. R., and Germain, R. N. (2002). Self-recognition promotesthe foreign antigen sensitivity of naive T lymphocytes. Nature 420, 429-434.
Stefanova, I., Dorfman, J. R., Tsukamoto, M., and Germain, R. N. (2003). On the roleof self-recognition in T cell responses to foreign antigen. Immunol Rev 191, 97-106.
Surh, C. D., and Sprent, J. (1994). T-cell apoptosis detected in situ during positiveand negative selection in the thymus. Nature 372, 100-103.
Sydora, B. C., Mixter, P. F., Holcombe, H. R., Eghtesady, P., Williams, K., Amaral, M.C., Nel, A., and Kronenberg, M. (1993). Intestinal intraepithelial lymphocytes areactivated and cytolytic but do not proliferate as well as other T cells in response tomitogenic signals. J Immunol 150, 2179-2191.

References
- 91 -
Tanaka, Y., Sano, S., Nieves, E., De Libero, G., Rosa, D., Modlin, R. L., Brenner, M.B., Bloom, B. R., and Morita, C. T. (1994). Nonpeptide ligands for human gammadelta T cells. Proc Natl Acad Sci U S A 91, 8175-8179.
Teh, H. S., Kishi, H., Scott, B., and Von Boehmer, H. (1989). Deletion of autospecificT cells in T cell receptor (TCR) transgenic mice spares cells with normal TCR levelsand low levels of CD8 molecules. J Exp Med 169, 795-806.
Teh, H. S., Kisielow, P., Scott, B., Kishi, H., Uematsu, Y., Bluthmann, H., and vonBoehmer, H. (1988). Thymic major histocompatibility complex antigens and the alphabeta T-cell receptor determine the CD4/CD8 phenotype of T cells. Nature 335, 229-233.
Terra, R., Labrecque, N., and Perreault, C. (2002). Thymic and extrathymic T celldevelopment pathways follow different rules. J Immunol 169, 684-692.
von Boehmer, H. (1990). Developmental biology of T cells in T cell-receptortransgenic mice. Annu Rev Immunol 8, 531-556.
von Boehmer, H., Aifantis, I., Gounari, F., Azogui, O., Haughn, L., Apostolou, I.,Jaeckel, E., Grassi, F., and Klein, L. (2003). Thymic selection revisited: how essentialis it? Immunol Rev 191, 62-78.
von Boehmer, H., Haas, W., and Jerne, N. K. (1978). Major histocompatibilitycomplex-linked immune-responsiveness is acquired by lymphocytes of low-responder mice differentiating in thymus of high-responder mice. Proc Natl Acad SciU S A 75, 2439-2442.
Vukmanovic, S., Grandea, A. G., 3rd, Faas, S. J., Knowles, B. B., and Bevan, M. J.(1992). Positive selection of T-lymphocytes induced by intrathymic injection of athymic epithelial cell line. Nature 359, 729-732.
Wagner, H., Rollinghoff, M., Rodt, H., and Thierfelder, S. (1980). T cell-mediatedcytotoxic immune responsiveness of chimeric mice bearing a thymus graft fullyallogeneic to the graft of lymphoid stem cells. Eur J Immunol 10, 521-525.
Wang, S. D., Huang, K. J., Lin, Y. S., and Lei, H. Y. (1994). Sepsis-inducedapoptosis of the thymocytes in mice. J Immunol 152, 5014-5021.
Weintraub, B. C., Jackson, M. R., and Hedrick, S. M. (1994). Gamma delta T cellscan recognize nonclassical MHC in the absence of conventional antigenic peptides. JImmunol 153, 3051-3058.
Wekerle, H., and Ketelsen, U. P. (1980). Thymic nurse cells--Ia-bearing epitheliuminvolved in T-lymphocyte differentiation? Nature 283, 402-404.
Wilkinson, R. W., Anderson, G., Owen, J. J., and Jenkinson, E. J. (1995). Positiveselection of thymocytes involves sustained interactions with the thymicmicroenvironment. J Immunol 155, 5234-5240.
Williams, M. A., Adams, A. B., Walsh, M. B., Shirasugi, N., Onami, T. M., Pearson, T.C., Ahmed, R., and Larsen, C. P. (2003). Primary and secondaryimmunocompetence in mixed allogeneic chimeras. J Immunol 170, 2382-2389.
Zerrahn, J., Volkmann, A., Coles, M. C., Held, W., Lemonnier, F. A., and Raulet, D.H. (1999). Class I MHC molecules on hematopoietic cells can support intrathymicpositive selection of T cell receptor transgenic T cells. Proc Natl Acad Sci U S A 96,11470-11475.

References
- 92 -
Zinkernagel, R. M. (1982). Selection of restriction specificities of virus-specificcytotoxic T cells in the thymus: no evidence for a crucial role of antigen-presentingcells. J Exp Med 156, 1842-1847.
Zinkernagel, R. M., and Althage, A. (1999). On the role of thymic epithelium vs. bonemarrow-derived cells in repertoire selection of T cells. Proc Natl Acad Sci U S A 96,8092-8097.
Zinkernagel, R. M., Althage, A., Waterfield, E., Kindred, B., Welsh, R. M., Callahan,G., and Pincetl, P. (1980). Restriction specificities, alloreactivity, and allotoleranceexpressed by T cells from nude mice reconstituted with H-2-compatible or -incompatible thymus grafts. J Exp Med 151, 376-399.
Zinkernagel, R. M., Callahan, G. N., Althage, A., Cooper, S., Klein, P. A., and Klein,J. (1978). On the thymus in the differentiation of "H-2 self-recognition" by T cells:evidence for dual recognition? J Exp Med 147, 882-896.

Curriculum Vitae
- 93 -
10 Curriculum Vitae
Name: Marianne M. A. Martinic
Work address: Institute of Experimental Immunology
Department of Pathology
University Hospital Zurich
Schmelzbergstrasse 12
8091 Zurich, Switzerland
Tel: +41 1 255 29 89
Fax: +41 1 255 44 20
Home address: Haldenbachstrasse 22
8006 Zurich, Switzerland
Tel: +41-79 407 20 83
Date of birth: 29.11.1974
Place of birth: La Paz, Bolivia
Nationality: Bolivian and French
Marital status: Single
Education:
1980-1982 Godehard Grundschule, Göttingen/Germany
1982-1992 Deutsche Schule “Mariscal Braun”, La Paz/Bolivia
1993-1995 Undergraduate study of biochemistry, Tübingen/Germany
1995-1998 Studies in biochemistry, immunology, genetics, neurobiology and physiology,
molecular biology and biophysics (Abt. XAe)
Swiss Federal Institute of Technology (ETH), Zurich/Switzerland
1996-1997 Diploma thesis at the Institute of Experimental Immunology.
Supervisors: Dr. M. van den Broek and Prof. H. Hengartner
Subject: “Establishment of a transgenic mouse model for inducible, tissue-
specific expression of neo-self antigens.”
1998 Start of PhD thesis at the Institute of Experimental Immunology.
Supervisors: Prof. H. Hengartner and Prof. R. M. Zinkernagel
Subject: “Parameters influencing efficient T cell repertoire selection.”


Bibliography
- 95 -
11 Bibliography
Martinic M. M., Rülicke T., Althage A., Odermatt B., Höchli M., Lamarre A., Dumrese
T., Speiser D. E., Kyburz D., Hengartner H., Zinkernagel R. M. (2003). Efficient T cell
repertoire selection in tetraparental chimeric mice independent of thymic epithelial
MHC. Proc Natl Acad Sci USA. 2003 Feb 18;100(4):1861-6.
Martinic M. M., Hengartner H., Zinkernagel R. M. (2003). Influence of MHC class I
H-2Db density on selection and survival of H-Y-specific TCR transgenic T cells.
Manuscript in preparation.
Martinic M. M., Hengartner H., Zinkernagel R. M. (2003). Selection of the H-Y-
specific transgenic TCR in an athymic versus euthymic environment. Manuscript in
preparation.
Macpherson A. J., Martinic M. M., Harris N. (2002). The functions of mucosal T cells
in containing the indigenous commensal flora of the intestine. Cell Mol Life Sci. 2002
Dec;59(12):2088-96. Review.
Hunziker L., Recher M., Ciurea A., Martinic M. M., Odermatt B., Hengartner H.,
Zinkernagel R. M. (2002). Antagonistic variant virus prevents wild-type virus-induced
lethal immunopathology. J Exp Med 2002 Oct 21;196(8):1039-46.
Medana I., Martinic M. M., Wekerle H., Neumann H. (2001). Transection of major
histocompatibility complex class I-induced neuritis by cytotoxic T lymphocytes. Am J
Pathol. 2001 Sep;159(3):809-15.
Ciurea A., Hunziker L., Martinic M. M., Oxenius A., Hengartner H., Zinkernagel R. M.
(2001). CD4+ T-cell-epitope escape mutant virus selected in vivo. Nat. Med. 2001
Jul;7(7):795-800.

Bibliography
- 96 -
Medana I. M., Gallimore A., Oxenius A., Martinic M. M., Wekerle H., Neumann H.
(2000). MHC class I-restricted killing of neurons by virus-specific CD8+ T
lymphocytes is effected through the Fas/FasL, but not the perforin pathway. Eur J
Immunol. 2000 Dec;30(12):3623-33.
Oxenius A., Martinic M. M., Hengartner H., Zinkernagel R. M. (1999). CpG-
containing oligonucleotides are efficient adjuvants for induction of protective antiviral
immune responses with T-cell peptide vaccines. J Virol. 1999 May;73(5):4120-6.

Acknowledgements
- 97 -
12 Danke, Merci, Thank You, Gracias
Zuallererst möchte ich mich ganz herzlich bei Hans Hengartner und Rolf Zinkernagel
für die Unterstützung und Anregungen während der gesamten Diss-Zeit bedanken.
Ich möchte mich insbesondere auch für die Freiheit bedanken, die ich bei den
einzelnen Projekten, beim Ausprobieren neuer Methoden und bei meiner eigenen
Meinungsbildung bezüglich bestimmter Themen jeweils hatte. Zuletzt möchte ich
mich nochmals speziell bei Rolf Zinkernagel für seine Geduld bei unseren ewigen
Diskussionen bedanken, die mich zwar manchmal zur Weissglut gebracht haben,
dafür aber doch immer sehr hilfreich und bereichernd waren.
J’aimerai aussi envoyer un très grand merci à Alain Lamarre, qui m’a toujours
soutenue et aidée pendant ces cinq dernières années - tu étais même d’accord de
discuter avec moi le sujet de T cell selection et ça en étant un 100% B cell type :-).
Ich möchte mich ganz fest bei Edit Horvath für ihre grosse Hilfe bei den
verschiedensten assays und ihrer immer anhaltenden guten Laune bedanken, sowie
für die schönen und sehr leckeren Scrabble-Abende zusammen mit Karin, Kathrin
und Co.
A big, big, big thank you to Kathy McCoy, Nathalie Oetiker, Veronika Pochanke,
Nicola Harris, Chiara Nembrini and Régine Dayer from the G43-lab for all their
support during the whole time inside and outside of the lab. I really enjoyed our
dinner-partys, snowboard-weekends, Christmas-celebrations and coffee- and lunch-
breaks talking about everything and nothing resulting in a perfect relaxing and
energy-re-loading atmosphere. In addition, I would like to thank Alana Althage and
Kathy McCoy for their constant support during the first year of my PhD-thesis, which
really helped me a lot.
Ganz besonders möchte ich mich beim gesamten Labor für die immer vorhandene
Unterstützung bedanken. Bei Elisabeth Hörhager, Simone Schmaderer und Daniela
Schuppisser möchte ich mich recht herzlich für die Beantwortung aller möglichen und
unmöglichen und vor allem dringenden Fragen sowie für die moralische

Acknowledgements
- 98 -
Unterstützung während der letzten paar Monate bedanken. Bei allen Laborantinnen
sowie Ljiljana Milojevic, Ioannis Chantziaras und Aytac Altuncevahir bedanke ich
mich für die perfekte Organisation unseres Labors, die es jedem ermöglicht, innert
kürzester Zeit die unmöglichsten Sachen wiederzufinden. Un gran agradecimiento va
al “feo, peludo y gordo” Lars Hangartner por su paciencia y gigante ayuda con todos
mis problemas de computación. Ein grosser Dank gilt allen Doktoranden für den
guten Zusammenhalt und für die chaotische, aber eben doch gemütliche Atmosphäre
im PhD-Raum. Zu guter letzt möchte ich mich noch bei Maries van den Broek für ihre
fortwährende Unterstützung und aufschlussreichen Diskussionen während der
gesamten Zeit hier bedanken.
Ein grosser Dank geht an Bernhard Odermatt und an sein gesamtes Histologie-Team
für die hervorragenden histologischen Schnitte. Ganz speziell möchte ich Bernhard
Odermatt für die sehr geduldige Einführung in die Thymus-Histologie und für das
Beantworten all meiner Fragen bedanken.
Ich möchte mich ganz herzlich bei Norbert Wey für seine ausserordentliche Geduld
und Hilfsbereitschaft beim Erklären und Vorführen aller möglichen Mikroskopier-
Computer-Techniken bedanken sowie für die enorme Hilfe beim Zusammenstellen
der Figuren.
Matthias Höchli vom Elektronischen Mikroskopie-Labor möchte ich danken für seine
grosse Unterstützung beim Erklären und Auswerten der Fluoreszenzmikroskopie-
Daten.
Ein grosser Dank gebührt auch Thomas Rülicke, dem ich mein ganzes Wissen über
die Herstellung der tetraparentalen Aggregationschimären zu verdanken habe.
Zum Schluss möchte ich meinen Freunden ausserhalb des Labors für ihre immer
anhaltende Unterstützung während der gesamten Diss-Zeit bedanken. Ganz speziell
möchte ich mich bei Familie Bock dafür bedanken, dass ich mich bei ihnen immer
daheim fühlen durfte und somit jedes Mal nach einem Wochenende in Tübingen
wieder völlig entspannt und voller frischer Energien nach Zürich zurückfahren konnte.
Hier in Zürich gilt mein ganz, ganz grosser Dank Sabine Bösch, die alle meine “Diss-

Acknowledgements
- 99 -
Launen“ miterleben durfte, und es doch jedes Mal wieder geschafft hat, mich zum
lachen zu bringen und das Leben in der Haldenbachstrasse wunderschön zu
gestalten.
Finalmente quisiera agradecer a mi hermana, Julie Martinic, por el lindo e
interesante tiempo que pasamos juntas en Zürich y por su constante apoyo como
también por sus consejos durante todo este tiempo.
Am allermeisten aber möchte ich mich ganz, ganz herzlich bei meinen Eltern
bedanken, dafür, dass sie immer, wirklich immer, für mich da waren und sind, für ihre
permanente Unterstützung während meiner gesamten Zeit hier in Europa und vor
allem dafür, dass es sie gibt.


