impact of mechanical stimulation on …1 impact of mechanical stimulation on tendon tissue in a...
TRANSCRIPT

1
IMPACT OF MECHANICAL STIMULATION ON TENDON TISSUE IN A
BIOREACTOR SYSTEM
TAO WANG This thesis is submitted to The University of Western Australia in fulfillment
of the requirement for degree of
DOCTOR OF PHILOSOPHY
IN
BIOMEDICAL ENGINEERING School of Surgery
The University of Western Australia
2014
The work presented in this thesis was performed in The University of Western
Australia, School of Surgery, Centre for Orthopaedic Translational Research

2
Declaration
This is to certify that all work contained herein was performed by myself, except where
indicated otherwise.
Tao Wang (PhD candidate)
W/Prof. Minghao Zheng (Supervisor)
A/Prof. Bruce Gardiner (Co-supervisor)
Prof. Brett Kirk (External supervisor)

3
TABLE OF CONTENTS
ABSTRACT 9
PUBLICATIONS 11
LIST OF AWARDS & PRESENTATIONS 12
ABBREVIATIONS 14
CHAPTER 1: INTRODUCTION 18
1.1 Tendon Biology 18 1.1.1 Anatomy of human Achilles tendon 19 1.1.2 Histology of healthy tendon 20 1.1.3 Aetiology of tendon injuries 24
1.1.3.1 Acute traumatic injury on Achilles tendon 24 1.1.3.2 Chronic Achilles tendinopathy 25
1.1.4 Pathology of tendon injuries 30 1.1.4.1 Pathology of tendinopathy 30 1.1.4.2 Healing process 31 1.1.4.3 Molecules involved in tendon remodeling 34
1.1.5 Treatments 43 1.1.5.1 Surgical treatment 43 1.1.5.2 Conservative treatment 44
1.1.6 Summary 56
1.2 Introduction to Bioreactor Design for Tendon/Ligament Engineering 56 1.2.1 Common key elements for tendon/ligament tissue engineering 57 1.2.2 Bioreactor design specific to tendon/ligament engineering 63
1.2.2.1 Actuator and culture chamber design 66 1.2.2.2 Environmental control and medium circulation systems 71 1.2.2.3 Monitoring and feedback systems 75
1.2.3 Commercial bioreactor systems for tendon/ligament engineering 79 1.2.4 Ideal bioreactors for tendon/ligament engineering 81 1.2.5 Previous work in this field 83 1.2.6 Conclusion 89
1.3 Hypothesis and aims 90 1.3.1 Hypothesis 90
1.3.1.1 Overall hypothesis 90 1.3.1.2 Specific hypothesis 91
1.3.2 Aims 91
CHAPTER 2 MATERIALS AND METHODS 94
2.1 Material 94 2.1 94 2.1.1 Tissue Culture reagents 94 2.1.2 Chemical reagents 95 2.1.3 Molecular products 96 2.1.4 Oligonucleotide Primers 96 2.1.5 Antibodies 98

4
2.1.6 Commercial kits 98 2.1.7 Other materials and equipment 99 2.1.8 Software 101 2.1.9 General solution 102
2.2 Tissue culture procedures 104 2.1 104 2.2.1 Isolation of rabbit Achilles tendon 104 2.2.2 Culture of rabbit Achilles tendon 104
2.3 RNA extraction 105 2.1 105 2.2 105 2.3.1 Extraction of total RNA 105 2.3.2 RNA concentration reading 106
2.4 Quantitative polymerase chain reaction (QPCR) 107 2.4.1 Reverse transcription of mRNA 107 2.4.2 Quantitative polymerase chain reaction (QPCR) 108
2.5 General histology 109 2.5.1 Tissue preparation 109 2.5.2 Haematoxylin and Eosin Staining 110 2.5.3 Histological scoring 112
2.6 Type III collagen immunohistochemistry 113
2.7 TUNEL assay 115
2.8 Statistics analyses 116
CHAPTER 3 (RESULTS): DESIGN AND CONSTRUCTION OF BIOREACTOR 118
3.1 Design criteria 118
3.2 Material selection 119
3.3 Overall bioreactor design 120 3.3.1 The Actuating System 124 3.3.2 The Culture Chamber 125 3.3.3 Tissue Fixation 126 3.3.4 The Control System 127
3.4 Bioreactor setup 128
3.5 Discussion 130
CHAPTER4 (RESULTS): PROGRAMMABLE MECHANICAL STIMULATION INFLUENCES TENDON HOMEOSTASIS IN A BIOREACTOR SYSTEM 133
4.1 Abstract 133
4.2 Introduction 134
4.3 Material and Method 136 4.3.1 Tissue harvest and distribution 136 4.3.2 Histological preparation and assessment 138

5
4.3.3 TUNEL assay 138 4.3.4 Immunostaining for type III collagen 138 4.3.5 Quantitative Real-time polymerase chain reaction (Q-PCR) 138 4.3.6 Statistical analyses 139
4.4 Results 139 4.4.1 Histological examination of tendon undergoing different cyclic tensile strains 139 4.4.2 Impact of cyclic tensile strain on apoptosis of tenocytes 144 4.4.3 Type III collagen turnover in Achilles tendon under dynamic culture 146 4.4.4 Impact of cyclic tensile strain on ECM remodeling gene expression 147
4.5 Discussion 148
4.6 Conclusion 154
CHAPTER 5 (RESULTS): CYCLIC MECHANICAL STIMULATION RESCUES RABBIT ACHILLES TENDON FROM DEGENERATION IN A BIOREACTOR SYSTEM 156
5.1 Abstract 156
5.2 Introduction 157
5.3 Material and method 158 5.3.1 Tissue preparation and programmable mechanical stimulation 158 5.3.2 Histological preparation and assessment 161 5.3.3 TUNEL assay 161 5.3.4 Biomechanics testing 161 5.3.5 Real-time polymerase chain reaction 162 5.3.6 Immunohistochemistry 162 5.3.7 Statistical analyses 162
5.4 Results 162 5.4.1 Cyclic mechanical stimulation improved the histological structure and cell viability of cultured tendon 162 5.4.2 Cyclic mechanical stimulation improved mechanical properties of cultured tendon 167 5.4.3 Rescue effect of cyclic mechanical stimulation on ECM remodeling gene expression 169 5.4.4 Cyclic mechanical stimulation decreased Type III collagen expression of cultured tendon 171
5.5 Discussion 172
5.6 Conclusion 178
CHAPTER 6 GENERAL DISCUSSION AND FUTURE DIRECTIONS 180
6.1 General discussion 180 6.1 181 6.1.1 The custom-made bioreactor (Chapter 3) 181 6.1.2 Mechanical stimulation in bioreactor system (Chapter 4 and 5) 183 6.1.3 Effect of various mechanical stimulation on tendon homeostasis (Chapter 4) 184 6.1.4 Rescue of degenerated tendon with mechanical stimulation. 187
6.2 Limitation of this work 188 6.2 188 6.3 188 6.2.1 Chapter 3 188 6.2.2 Chapter4 189 6.2.3 Chapter 5 190

6
6.3 Future directions 190 6.3.1 Gene profile of tenocyte subjected to 2D or 3D mechanical stimulation 190 6.3.2 The effect of mechanical stimulation on tendon nutrient infiltration 192 6.3.3 The effect of mechanical stimulation on stem cell tenogenic differentiation 193 6.3.4 Therapeutic guideline 194 6.3.5 Robot- driven orthosis 195 6.3.6 Tendon engineering 196
6.4 Conclusion 197
REFERENCE 198
APPENDIX 225

7
Dedicated to
My Awesome parents, Jihong Wang and Junhua Xu
and beautiful lovely wife, Xiang Jiang
for their love and support

8
ACKNOWLEDGEMENTS
The work described in this thesis was performed at the Center for Orthopaedic
Translational Research, School of Surgery, The University of Western Australia. I
would like to express my appreciation to my supervisor, Winthrop Professor
Minghao Zheng, co-supervisor Associate Professor Bruce Gardiner and External
supervisor Professor Brett Kirk. Without their support and guidance, this work
would not have been finished.
Sincere thanks to Dr. Zhen Lin and Ms. Euphemie Landao-Bassonga for their valuable
contributions, technical training and support.
I would love to thanks one of my best friend, Weiming Zeng, for the technical support
of bioreactor construction.
At last, I would also like to extend my appreciation to my colleagues and the
administrative officer for their assistance, friendship and encouragement.

9
ABSTRACT
Tendons are force-transmitting tissues connecting muscle to bone. Because of this
physiological function, biomechanics plays an essential role in maintaining tendon
homeostasis. Indeed tenocytes have the ability to sense and respond to different
mechanical loads by remodeling the tendon tissue. Studies reported that load
deprivation can cause tendinopathy-like morphology such as disorientated collagen
fibers and rounded cell nuclei, while mechanical overloading can result in tears and
rupture of the tendon. In tendon engineering studies, mechanical stimulation has been
shown to improve the cell viability, proliferation, and neo-tendon structure and
mechanical properties. Obviously, the biomechanical environment has various effects on
tendon tissue; and yet the specific effects of different loading conditions on tendon have
not been well elucidated. Mechanical loading is both a source of degradation (e.g.
through damage and indirectly through protease activity) and a stimulus for tissue
synthesis/repair. The hypothesis of this PhD project is that there is some optimal
mechanical load to maintain or restore functional tendon tissue in a bioreactor.
Firstly, in order to study the mechanical stimulation on tendon tissue, a functional
bioreactor is necessary. A uniaxial bioreactor was designed and fabricated. This
bioreactor system is able to provide pre-programmable mechanical simulation and
sterilized environment for a maximum 6 individual tendons simultaneously.
Secondly, rabbit Achilles tendon were cultured under different mechanical environments
including no loading, 3% strain, 6% strain and 9% strain at 0.25Hz for 8h/day for 6 days

10
in the bioreactor system developed in this study. Histological and gene assessment
showed that, of the strains applied, only at 6% strain was the rabbit model able to
maintain the tendon homeostasis, other loading regimes resulted in pathological changes
to the tendon as observed through histology testing.
Thirdly, due to the fact that loading deprivation of tendon can lead to progressive tendon
degeneration; static cultured rabbit Achilles tendons was used as a degradation model.
Application of 6% mechanical stimulation for 6 days followed by 6 days loading
deprivation culture significantly improved the morphology of degenerative tendon and
successfully restored the mechanical properties.
In summary, this study identified the effect of various loading intensities on tendon
homeostasis in bioreactor system, and proposed a hypothetical model that there is only a
narrow range of mechanical stimulation can maintain tendon homeostasis. We then
further estimated the therapeutic effect of mechanical stimulation on a degenerative
model, and found out accurate mechanical stimulation was able to reverse the
early-stage degradation of Achilles tendon.
This leads to the conclusion that proper mechanical stimulation may have therapeutic
benefits and clinical application.

11
Publications
Publications arising from this thesis
1. Wang, T., Gardiner, B. S., Lin, Z., Rubenson, J., Kirk, T. B., Wang, A., Xu, J., Smith,
D. W., Lloyd, D. G., Zheng, M. H., Bioreactor design for tendon/ligament
engineering. Tissue Eng Part B Rev 2013;19(2):133-46 (Appendix 1)
2. Wang, T., Lin, Z., Day, R. E., Gardiner B. S., Landao-Bassonga, E., Rubenson, J.,
Kirk, T. B., Smith, D. W., Lloyd, D. G.,Hardisty, G., Wang, A., Xu, J., Zheng, Q.
Zheng M. H. Programmable mechanical stimulation influences tendon homeostasis
in a bioreactor system. Biotechnol Bioeng 2013;110(5):1495-507 (Recommended
by Faculty 1000) (Appendix 2)
3. Wang, T., Lin, Z., Ni M., Day, R. E., Gardiner B. S., Landao-Bassonga, E.,
Rubenson, J., Kirk, T. B., Smith, D. W., Wang, A., Lloyd, D. G., Wang Y., Zheng Q.,
Zheng M. H., Cyclic mechanical stimulation rescues rabbit Achilles tendon from
degeneration in a bioreactor system. (manuscript submitted to Tissue engineering
Part A)

12
List of Awards & Presentations
Awards
1. “Peoples’ Choice Poster Award”, Combine Biology Science Meeting (CBSM), Perth,
Australia, 2012: Wang, T., Lin, Z., Landao-Bassonga, E., Zheng, M.H.,
Development and optimization of ex vivo bioreactor system for tendon tissue
engineering.
2. 2013 China Distinguished International student scholarship
Oral Presentations
1. Wang, T., Lin, Z., Landao-Bassonga, E., Zheng, M.H., Development and
optimization of ex vivo bioreactor system for tendon tissue engineering. In
Australian & New Zealand Orthopaedic Research Society 18th annual meeting.
Perth, Australia, 2012
2. Wang, T., Lin, Z., Landao-Bassonga, E., Zheng, M.H., Development and
optimization of ex vivo bioreactor system for tendon tissue engineering. In
Australian Society of Medical Research, 2012 annual meeting. Perth, Australia,
2012
3. Wang, T., Lin, Z., Day, R. E., Gardiner B. S., Rubenson, J., Kirk, T. B., Smith, D.
W., Lloyd, D. G., Wang, A., Zheng, M.H., The effect of programmable mechanical
stimulation on tendon repair in a bioreactor system. In Australian & New Zealand
Orthopaedic Research Society 19th annual meeting. Sydney, Australia, 2013
4. Wang, T., Lin, Z., Day, R. E., Gardiner B. S., Rubenson, J., Kirk, T. B., Smith, D.
W., Lloyd, D. G., Wang, A., Zheng, M.H., The effect of programmable mechanical

13
stimulation on tendon repair in a bioreactor system. In Combio 2013. Perth,
Australia, 2013
5. Wang, T., Lin, Z., Day, R. E., Gardiner B. S., Rubenson, J., Kirk, T. B., Smith, D.
W., Lloyd, D. G., Wang, A., Zheng, M.H., The effect of programmable mechanical
stimulation on tendon repair in a bioreactor system. In International Symposium on
Ligaments & Tendons. Arezzo, Italy, 2013
Poster Presentations
1. Wang, T., Lin, Z., Landao-Bassonga, E., Zheng, M.H.Minghao Zheng,
Development and optimization of ex vivo bioreactor system for tendon tissue
engineering. In Australian and New Zealand Bone and Mineral Society 22nd
annual meeting. Perth, Australia, 2012
2. Wang, T., Lin, Z., Day, R. E., Gardiner B. S.,Rubenson, J., Kirk, T. B., Smith, D. W.,
Lloyd, D. G., Wang, A.,Zheng, M.H., The effect of programmable mechanical
stimulation on tendon repair in a bioreactor system. In Australian and New Zealand
Bone and Mineral Society 23nd annual meeting. Perth, Australia, 2012

14
Abbreviations
ADAMTS A Disintegrin and Metalloproteinase with Thrombospondin Motifs
ADSCs Adipose-derived stem cells
AT Achilles tendon
ATT Autologous tenocyte therapy
bFGF Basic fibroblast growth factor
BMSCs Bone marrow-derived stem cells
BMP Bone morphogenetic proteins
COX-2 Prostaglandin-endoperoxide synthase 2
DMEM Dulbecco’s modified Eagle’s medium
ESCs Embryonic stem cells
ECM Extracellular matrix
GDF-5 Growth differentiation factor 5
GAG Glycosaminoglycan
H&E Haematoxylin-eosin
H2O2 Hydrogen peroxide
LE Lateral epicondylitis
IGF-I Insulin-like growth factor-I

15
IL-6 Interleukin-6
LVDT Linear variable differential transformers
MCP1 Monocyte chemotactic protein 1
MIF Macrophage inhibitory factor
MMP Matrix metalloproteinase
MSCs Mesenchymal stem cells
NSAID Non-steroid anti-inflammation drug
OCT Optical coherence tomography
PBS Phosphate buffered saline
PDGF Platelet-derived growth factor
PRP Platelet-rich plasma
PMMA Polymethylemethacrylate
PMS Programmable mechanical stimulation
PGE2 Prostaglandin E2
Scx Scleraxis
SDSCs Synovium-derived stem cells
SMBS Step motor-ball screw transmission system
SP Substance P

16
TDSCs Tendon-derived stem cells
TGF- Transforming growth factor
VEGF Vascular endothelial growth factor

17
Chapter 1
Introduction

18
CHAPTER 1: INTRODUCTION
1.1 Tendon Biology
The Achilles tendon (AT), also known as the calcaneal tendon, is the strongest and
thickest tendon in human body. It connects the gastrocnemius and soleus muscles to
calcaneus bone. As a force transferring tissue, AT serves the function of transferring the
power generated by the muscle to the calcaneus, allowing plantar flexion about the ankle
joint. It also helps store elastic energy while walking/running, increasing locomotion
efficiency (Bressel and McNair 2001).
AT injury is a serious and common injury. AT rupture was first reported in 1575 by
Ambrose Paré. The AT injuries rate is estimated at between 2 to 12 in 100,000 people
(So and Pollard 1997; Schepsis, Jones et al. 2002; Suchak, Bostick et al. 2005).
Typically, this injury affects males between 30 to 50 years old (Gebauer, Beil et al. 2007).
It is thought that due to the specific geometry of the AT, the mechanical weak point is 2
to 6 cm from the calcaneal insertion, that is the thinnest part of the AT and the common
rupture site(Wren, Lindsey et al. 2003). In addition, 11% of regular runners suffer from
Achilles tendinopathy(Rees, Wilson et al. 2006), which can cause significant pain and
restricts the activities in daily living, and potentially tendon rupture (Leppilahti and
Orava 1998; Smith 2000). Traditionally, overuse and/or overloading has been
considered to be the cause of tendinopathy, 30% to 50% of sports injuries were reported
to be an overuse injury, and tendon injuries comprise a large part of that(Schechtman
and Bader 1997; Khan and Cook 2003).

19
In the United States, injuries to tendons and ligaments represent about half of the 33
million musculoskeletal injuries(Huang, Qureshi et al. 2000). Each year more than
33,000 tendon reconstructions occur in U.S., costing $30 billion USD(Butler, Gooch et
al. 2010; Chen, Yin et al. 2010). In Australia, $250 million is spent annually just on
rotator cuff repair(Chen, Xu et al. 2009). However, despite the high prevalence of
tendon injury and associated tendinopathy worldwide, treatment options remain poorly
defined.
1.1.1 Anatomy of human Achilles tendon
The Achilles tendon is the extension of two muscles, the gastrocnemius and soleus
muscles (Suchak, Bostick et al. 2005). Depending on knee positioning, these separate
muscles have their own movement (Schepsis, Jones et al. 2002), but mainly control the
plantar flexion of the ankle, allowing for locomotion at the ankle (Suchak, Bostick et al.
2005). These muscles form together at the mid-calf region of the lower leg, where the
AT begins. The AT then merges into a single tendon at around 5 to 6 cm from the
calcaneal insertion (So and Pollard 1997). The AT is covered by a peritenon, a
single-cell layer tissue, which act as a lubricant during dynamic activity and provides
vascular supply to the tendon tissue (Schepsis, Jones et al. 2002). Beside the peritenon,
the main nutrient supply is from the muscle-tendon junction and bone tendon junction,
therefore, the nutrient delivery to the AT is very limited (Schepsis, Jones et al. 2002).
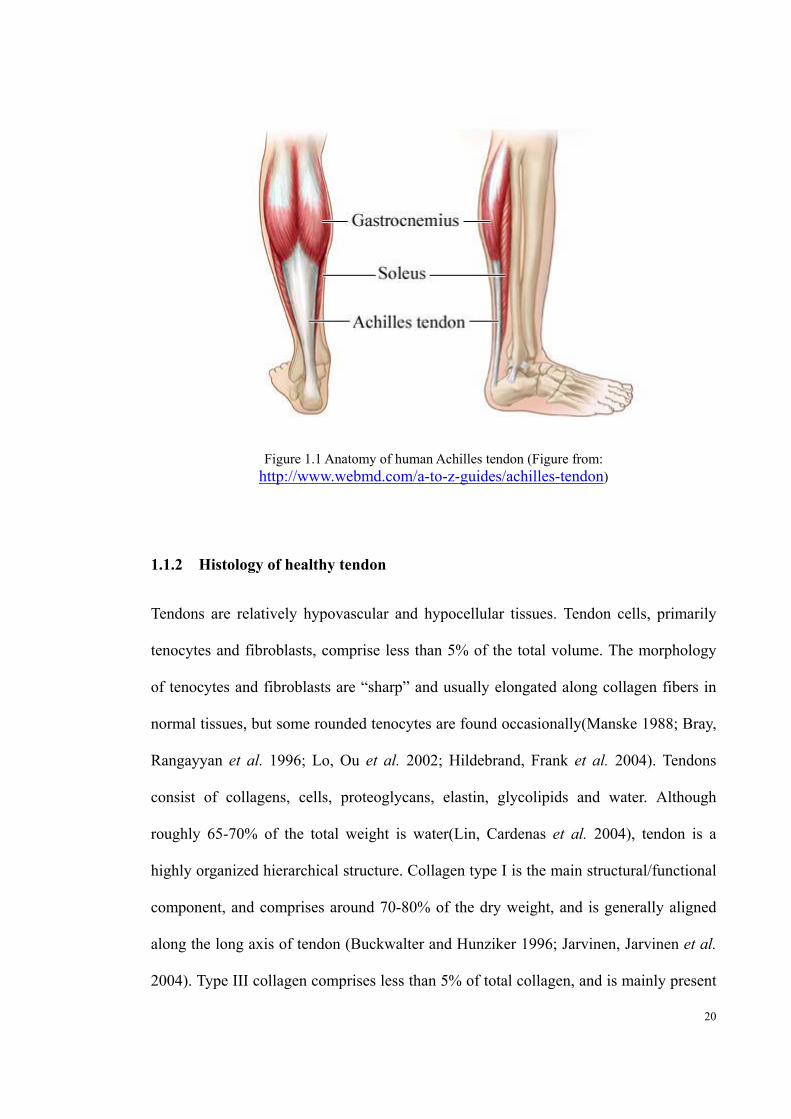
20
Figure 1.1 Anatomy of human Achilles tendon (Figure from: http://www.webmd.com/a-to-z-guides/achilles-tendon)
1.1.2 Histology of healthy tendon
Tendons are relatively hypovascular and hypocellular tissues. Tendon cells, primarily
tenocytes and fibroblasts, comprise less than 5% of the total volume. The morphology
of tenocytes and fibroblasts are “sharp” and usually elongated along collagen fibers in
normal tissues, but some rounded tenocytes are found occasionally(Manske 1988; Bray,
Rangayyan et al. 1996; Lo, Ou et al. 2002; Hildebrand, Frank et al. 2004). Tendons
consist of collagens, cells, proteoglycans, elastin, glycolipids and water. Although
roughly 65-70% of the total weight is water(Lin, Cardenas et al. 2004), tendon is a
highly organized hierarchical structure. Collagen type I is the main structural/functional
component, and comprises around 70-80% of the dry weight, and is generally aligned
along the long axis of tendon (Buckwalter and Hunziker 1996; Jarvinen, Jarvinen et al.
2004). Type III collagen comprises less than 5% of total collagen, and is mainly present

21
in the endotenon and epitenon, but is also present in the early phase of tendon repair
(Buckwalter and Hunziker 1996; Jarvinen, Jarvinen et al. 2004). The hierarchical
structure of tendon is shown in Figure 1.2, which is modified from the study of Kastelic
et al. (Kastelic, Galeski et al. 1978).
Type 1 collagen molecules synthesized by tenocytes bind together into a triple helix
tropocollagen (Kuhn 1969), which then self-assemble into microfibril and bind to
adjacent helixes by molecules such as decorin and biglycan, so forming collagen
fibrils(Tkocz and Kuhn 1969). This helixes structure is able to provide high resistance
to tensile strain while maintaining its flexibility (Screen, Lee et al. 2004). Due to this
structural characteristic, the tendon is able to efficiently transfer force from muscle to
bone (Magnusson, Hansen et al. 2003). The collagen fibrils align parallel to each other
assemble into collagen fiber with a waveform structure, enabling the tendon to absorb
the sudden initial force generate by muscle(Kastelic, Palley et al. 1980). As shown as
Figure 1.3, when unloaded the collagen fiber appears ‘wavy’. The wavy structure
disappears at around 4% strain, micro and partial rupture starts at ~8% strain, beyond
that, complete rupture is likely to occur (Maffulli 1998).
Tenocytes adhere to the collagen fiber; and sense the mechanical loading on the tendon
(Benjamin and Ralphs 1998; Murata, Nishizono et al. 2000). In the haematoxylin-eosin
(H&E) stained normal and healthy tendon section, tenocytes are evenly distributed
between the straight and parallel collagen fibrils with sharp and elongate nuclei (Chen,

22
Willers et al. 2007).
Although proteoglycans comprised only ~1% dry mass of tendon, they still play an
essential role in tendon biomechanics(Thompson 2013). Small proteoglycans such as
decorin control collagen fibril assembly and alignment(Parkinson, Samiric et al. 2011),
and the hydrophilic nature of the large proteoglycans has a strong effect on tissue
mechanical properties (Screen, Chhaya et al. 2006). Moreover, proteoglycan
concentration varies within and between tendon due to different local loading
characteristics(Birch 2007).
Elastin and microfibrils form elastic fibers, which comprise around 0.1% to 2% dry
mass of tendon(Kannus 2000). They plays a role in the tissue low strain and resilience
propertie by bridging between collagen fiber bundles with a network structure(Smith,
Vaughan-Thomas et al. 2011).

23
Figure 1.2 The structure of a healthy tendon. figure modified from Kastrlic et al. (Kastelic, Galeski et al. 1978)
Figure 1.3 Force versus strain curve of tendon (figure modified from the study of Wang et al.) (Wang 2006)

24
1.1.3 Aetiology of tendon injuries
Tendon injuries can be classified into acute and chronic. In acute traumatic injury
extrinsic factors predominate, whereas in chronic tendinopathy both intrinsic and
extrinsic factors interact (Williams 1993; Khan and Maffulli 1998).
1.1.3.1 Acute traumatic injury on Achilles tendon
Traumatic AT injuries are often associated with sport activities, although AT is the
strongest tendon in human body, sudden, excessive strain of AT from athletic or
recreational activities can cause tears or even rupture the AT (Kannus and Natri 1997).
Statistics show that 60-75% of AT ruptures happen during sport activities, especially in
soccer, badminton and basketball (Kannus and Natri 1997). Most of the AT ruptures are
associated with sudden acceleration and jumping, that is those activities which require
massive sudden force generation from muscle, and the rupture site tend to locate at 2 to
6 cm above the calcaneus-Achilles tendon junction. Interestingly the left tendon is more
commonly affected than the right tendon (Hattrup and Johnson 1985; Leppilahti and
Orava 1998).
In acute traumatic injuries, AT failure is mainly due to excessive strain caused by
sudden explosive power produced by the triceps surae (Langberg, Skovgaard et al.
1999). Hoffmeyer et al. found some pathological changes including increased lipid
droplet and ultrastructural muscle changes indicating partial ischemia in the triceps

25
surae muscle after AT rupture, and this might contribute to the abnormal stiffness in the
triceps muscle causing excessive muscle contraction. (Hoffmeyer, Freuler et al. 1990)
1.1.3.2 Chronic Achilles tendinopathy
Tendinopathy of the AT is a chronic non-inflammatory, degenerative condition, which
mainly affect male runners between 35 and 45 years old(Alfredson and Lorentzon 2000).
Tendinopathy is normally characterized by matrix disorganization, hypercellularity and
vascular hyperplasia (Teitz, Garrett et al. 1997). In some cases heterotopic
mineralization was considered as a feature of tendinopathy as well (Jarvinen, Jozsa et al.
1997). The clinical symptom of tendinopathy in the AT is pain, which generally happens
at the beginning and the end of training (Maffulli, Sharma et al. 2004). As the
pathological process progresses, it can affect daily activities. Approximately 3-10% of
chronic Achilles tendinopathy develop to AT ruptures (Khan, Cook et al. 1999).
Traditionally, overuse is considered as an extrinsic factor of Achilles tendinopathy.
Therefore, tendinopathy mainly affects runner, dancers and gymnasts, who constantly
and repetitively overload their tendon (Teitz, Garrett et al. 1997). During slow walking,
human AT is subjected to ~3kN force, and it can reach up to 9kN at a speed of 6m/s,
which corresponds to 12.5 times body weight (Komi, Fukashiro et al. 1992). Moreover,
the peak stress of AT is more than twice that of other tendons in human body(Wren,
Yerby et al. 2001). Although tendinopathy affects such a large population, the exact
aetiology remains uncertain beyond the catch-all ‘overuse’. There are several theories

26
proposed, including vascular supply, hypoxia, hyperthermia, apoptosis and abnormal
biomechanical environment.
1.1.3.2.1 Vascular supply
When tendon is subjected to mechanical loading, microruptures or damage of collagen
fibrils start to accumulate, and need to be repaired by tenocytes. However, the vascular
supply of the “high risk, high stress zone” (2-6 cm above the calcaneus insertion) is
relatively poor compared to the other parts of the tendon, presumably due to the hostile
mechanical environment (Hattrup and Johnson 1985; Carr and Norris 1989; Leppilahti
and Orava 1998). Under the situation of repetitive damage without enough blood supply,
the repair mechanism may be unable to maintain the tendon’s structural integrity, and
then the AT undergoes degeneration(Kannus and Jozsa 1991).
1.1.3.2.2 Hypoxia
The oxygen supply of tendon and ligament is only 13% that of skeletal muscles(Kannus
and Jozsa 1991). The degree and duration of the hypoxia plays a key role in cellular
viability(Nathan 2002). Several studies suggest that a hypoxic environment can induce
inflammatory cytokines, including interleukin-6 (IL-6), interleukin-8 (IL8), monocyte
chemotactic protein 1 (MCP1) and Platelet-derived growth factor (PDGF), and so might
significantly disrupt the balance between reparative and degenerative processes in the
tendon (Berse, Hunt et al. 1999; Zamara, Galastri et al. 2007; Millar, Reilly et al. 2012).

27
Moreover, hypoxia has been shown to increase the total collagen production but with a
‘shift’ in production to type III collagen instead of type I, thus disturbing the original
collagen composition ratio (Millar, Reilly et al. 2012). Lastly, when tendon repair
mechanisms are activated, tendon requires oxidative energy metabolism to maintain
cellular ATP levels. That is local tissue hypoxia may result in reduced repair capacity
and/or tenocyte death (Birch, Rutter et al. 1997).
1.1.3.2.3 Hyperthermia
During locomotion, 5% to 10% of energy stored in tendons is converted into heat (Ker
1981; Riemersma and Schamhardt 1985). Due to the hypovascular structure of tendon,
the blood supply to the structure is not sufficient to dissipate the heat generated. The
temperature of equine superficial digital flexor tendon has been recorded to reach 45℃
during galloping(Wilson and Goodship 1994). The in vitro study of Birch et al. indicates
that tenocytes appear to have high thermal tolerance; even under 45℃ culture condition
the viability of tenocyte is still unaffected. However, compromised cellular function
might be induced by repeated exposure to short periods of hyperthermia, as it has been
shown to be associated with reduced collagen synthesis and disturbed cell
metabolism(Arancia, Crateri Trovalusci et al. 1989; Birch, Wilson et al. 1997).

28
1.1.3.2.4 Apoptosis
Significant tenocyte apoptosis has been found in tendinopathy in various tendons,
including rotator cuff and AT(Yuan, Wang et al. 2003; Chen, Wang et al. 2010; Wang,
Lin et al. 2013). Deformation of the cytoskeleton of tenocytes can produce
stress-activated protein kinase that triggers apoptosis (Arnoczky, Tian et al. 2002;
Skutek, van Griensven et al. 2003). In ruptured tendon, apoptotic cell number is
elevated compared to normal tendon (Chen, Willers et al. 2007). For example,
quadriceps femoris tendons with tendinopathy exhibited a 1.6 times higher apoptosis
rate than normal tendon (Machner, Baier et al. 2003).
1.1.3.2.5 Biomechanical environment
As a force transferring tissue, it is unsurprising that the biomechanical environment is
essential to tendon homeostasis; overloading and underloading can both lead to tendon
abnormality. Repetitive and excessive loading may result in release of cytokines by
tenocytes leading to abnormal cellular activities(Leadbetter 1992). The release of
cytokines may induce the expression of MMPs, which can degrade the extracellular
matrix and eventually causes tendinopathy (Chen, Yu et al. 2011). However, loading
deprivation of tenocyte can cause direct upregulation of MMP-1(Choi, Kondo et al.
2002). It has been reported that mechanical stimulation is able to reduce MMP-1
expression (Lavagnino, Arnoczky et al. 2003). Due to the different protocols that have
been adopted in tendon studies on the effect of mechanical stimulation on tendon,
simple comparison is often difficult.

29
Compared to the tendon-muscle junction and the mid-tendon, the biomechanical
environment of the bone-tendon junction is more complicated. The attachment of
tendon is through fibrocartilage to mineralized fibrocartilage to bone in relatively a
short distance (<2mm), which is called as ‘the enthesis organ’ (Benjamin, Moriggl et al.
2004). Between the tendon and the bone is a bursa, and fibrocartilage is expressed on
the opposing bone and tendon surface to absorb the compression of the tendon against
the bone, as shown in Figure 1.4 (Cook and Purdam 2012). In most of the clinical cases,
the abnormal imaging findings and the reports of tendinopathy on tendon-bone insertion
are more likely at the site of compression proximal to the tendon insertion, which
indicate that abnormal compression might be able to trigger tendinopathy (Ohberg and
Alfredson 2003; Kong, Van der Vliet et al. 2007). The study of Grigg et al. suggests that
excessive loading on the tendon will cause the loss of bound water normally presented
in the transitional zones (Grigg, Wearing et al. 2009), which further increases the loads
carried by tenocytes located in these regions. To respond to the excessive compression,
tenocyte start to synthesis large water binding proteoglycans to reduce the permeability,
thereby protecting against further insult. Further loading on this swollen region may
aggravate this situation and potentially result in tendinopathy (Hamilton and Purdam
2004; Parkinson, Samiric et al. 2010).

30
Figure 1.4 Normal tendon enthesis, figure taken from Cook et al. (Cook and Purdam 2012)
1.1.4 Pathology of tendon injuries
1.1.4.1 Pathology of tendinopathy
“Tendinitis” has been used for definition of tendon degeneration in tendinopathy for
over 20 years (Puddu, Ippolito et al. 1976). Although the fundamental problem of
tendinopathy is collagen degeneration instead of inflammatory, many clinicians still
use the term of “tendinitis”. Most of the scientists nowadays have advocated the term
of “tendinopathy” for description of the clinical degeneration condition in and around
tendons caused by overuse, while the term “tendinitis” is only used after
histopathological examination (Maffulli, Khan et al. 1998).

31
Repetitive overloading on tendon was commonly considered as a cause of
tendinopathy. Due to repetitive loading during daily activities, Achilles tendon is one
of the most common tendons affected by tendinopathy. Histologically, tendinopathy
is mainly characterized by collagen fiber disorientation and thinning, scattered
vascular ingrowth, cell rounding, changes of cell density and increased expression of
type III collagen, in some cases, glycosaminoglycan (GAG) accumulation, lipid
droplets accumulation, ossification and increased cell apoptosis (Leadbetter 1992;
Khan and Maffulli 1998).
Various types of degeneration can be found in different tendons, however, in AT,
mucoid and lipoid are the most common types. In mucoid degeneration, vacuoles and
mucoid patches characterized by accumulation of proteoglycan/GAG are found
between the collagen fibers(Maffulli, Sharma et al. 2004). However, in lipoid
degeneration, the collagen structure of tendon is disrupted by the abnormal
accumulation of lipid(Maffulli, Sharma et al. 2004). An abnormally high expression
of type III collagen is found in degenerated tendon (Riley, Harrall et al. 1994).
1.1.4.2 Healing process
Tendon injuries can be divided into two categories, traumatic injury and chronic
tendinopathy. The tissue response to these two types of injuries are not quite the same
(Leadbetter 1992). The healing process of tendon has been studied in both human and
animals; however, most are concerned with traumatic injuries healing, whilst the

32
(inadequate) healing response in degenerative tendon is still poorly understood.
1.1.4.2.1 Acute traumatic injury healing
Tendon healing after acute traumatic injury requires reestablishment of the
connection between collagen fibers and the gliding mechanism between tendon and
its neighboring structure (Schneewind, Kline et al. 1964; Abrahamsson and
Gelberman 1994). Formation of scar tissue provides initial repair at the injury site
(Dunphy 1967). However, a lack of mechanical stimulation is thought to lead to an
excess of scar tissue, adhesions, and so compromises the normal tendon function.
Therefore, the rehabilitation procedure after initial immobilization of the injury site is
critical, as the mechanical loading is able to guide organized collagen fiber formation,
decrease the formation of postoperative adhesions and increase tendon
strength(James, Kesturu et al. 2008).
After acute tendon injury, the body initiates the healing process, which includes three
overlapping stages including: (1) acute inflammation; (2) proliferation and (3)
remodeling (Goodship, Birch et al. 1994).
The acute inflammation phase begins right after the injury and lasts 1 to 2 week
depending on the severity of the injury. Clinically, the inflammation phase is
characterized by well-known signs of inflammation such as heat, pain and swelling

33
(Goodship, Birch et al. 1994). Histologically, injuries on the tendon cause the
formation of hematomas in the tendon sheath, which release various chemotactic
factors and pro-inflammatory molecules. These molecules attract inflammatory
cells such as neutrophils, monocytes and macrophages to migrate from surrounding
tissue to the wound site where the cellular debris and foreign body matter are
engulfed and resorbed by phagocytosis(Maffulli, Sharma et al. 2004). Meanwhile,
tenocytes are recruited to the site and start to synthesize various extracellular matrix
components and reestablish vascular networks (Lindsay and Birch 1964; Myers and
Wolf 1974).
During the proliferation stage, tenocytes continue to be recruited and proliferate to
accelerate the repair process at the wound site. However, the newly synthesized ECM
is mainly a disorganized type III collagen(Garner, McDonald et al. 1989). An
extensive vascular network is formed and the wound exhibits scar-like tissue.
During the remodeling stage, 6-8 weeks after injury, cellular proliferation, matrix
synthesis, type III collagen expression start to decrease, and collagen type I synthesis
increases to replace the type III collagen (Liu, Yang et al. 1995). Type I collagen
fibers provide the long-term mechanical support of the regenerated tissue. However,
it is uncertain as to whether the repaired tissue achieves the strength of the original
tissue(James, Kesturu et al. 2008).

34
1.1.4.2.2 Chronic healing
Chronic healing processes in chronic tendinopathy are different from the acute
healing case described above. In normal tendon, type I collagen, the predominant
collagen, is well organized along the axis of the tendon. However, in tendinopathy,
the collagen structure is disorganized and type III collagen is highly expressed. In the
normal healing process, type III collagen is synthesized by tenocytes as a temporary
“band aid”(Maffulli, Khan et al. 1998). During the remodeling stage, type III
collagen is replaced by collagen type I which is more resistant to mechanical loading.
However, the repetitive and chronic damage in chronic tendinopathy keeps the high
expression of collagen type III in tendon without shifting to collagen type I (Riley,
Harrall et al. 1994). Due to the failure to complete this final remodeling stage, the
tendon is gradually weakened; and eventually leads to rupture even at ‘low’ daily
activities(Hamada, Okawara et al. 1994).
1.1.4.3 Molecules involved in tendon remodeling
1.1.4.3.1 Inflammatory mediators in tendon degeneration
Although tendinopathy is considered to be a degenerative process, rather than
inflammation, inflammatory mediators still play an important role. Interleukin (IL)-6
and IL-1 have been identified as two of the most important inflammatory mediators
and their functions are similar (Chen, Yu et al. 2011). IL-6 and IL-1 are both able to
induce Prostaglandin-endoperoxide synthase 2 (COX2), which stimulates the expression
of PGE2 and the acute phase of an inflammatory response in tendon (Chen, Yu et al.

35
2011). The inflammatory response alters the cell homeostasis causing apoptosis of
tenocytes which initiates the pathogenesis of tendon. IL-6 and IL-1have also been
shown to stimulate MMP1 and 3 expression (Archambault, Tsuzaki et al. 2002; Chen,
Yu et al. 2011), which can degrade collagen type I and collagen–associated small
proteoglycans respectively. Therefore, the upregulation of these inflammatory mediators
can cause matrix degradation.
The induction of IL-6 and IL-1 can be divided into chemical and mechanical
stimulation. Macrophage inhibitory factor (MIF) and Substance P (SP) have been
reported to upregulate IL-6 and IL-1 expression in tenocytes (Hart, Archambault et al.
1998; Nguyen, Lue et al. 2003; Morand, Leech et al. 2006). Tenocytes subjected to
cyclic strain increase the production of IL-6 and IL-1 (Skutek, van Griensven et al.
2001; Archambault, Tsuzaki et al. 2002; Tsuzaki, Bynum et al. 2003). The regulatory
network between inflammatory cytokines and tenocyte apoptosis remains unclear. It is
possible that in a routine process, normal loading induces cytokines in tissues that
trigger apoptotic cell death to remove the damaged cells. However, over-loading may
cause excessive production of cytokines and lead to excessive apoptosis and ultimate
tissue degeneration (Millar, Wei et al. 2009).
1.1.4.3.2 Enzyme in tendon degeneration
There are two kinds of metalloproteinase involved in tendon degeneration, which are
Matrix metalloproteinase (MMPs) and A Disintegrin and Metalloproteinase with

36
Thrombospondin Motifs (ADAMTS).
MMPs are zinc-dependent endopeptidases that are able to degrade all the components of
the extracellular matrix. MMPs can be classified into four main groups: collagenase,
gelatinases, stromelysins and membranes type MMPs(Bramono, Richmond et al. 2004).
Tendon degradation is initiated by MMPs (Riley, Curry et al. 2002). ADAMTS family,
which are also known as “aggrecanases”, are able to degrade proteoglycans, however,
the precise ADAMTSs that are involved in the degradation of tendon proteoglycans
remains unclear.
The regulation network of metalloproteinases is very complicated, but includes
transcription, activation and inhibition by tissue inhibitors of metalloproteinase (TIMPs)
(Nagase, Visse et al. 2006). There are 23 MMPs and 19 ADAMTS in humans, almost all
of which can be detected in Achilles tendon, although the expression levels vary widely.
These enzymes are essential regulators in cellular activities, matrix remodeling, and
pathologic processes (McCawley and Matrisian 2001). The main members of the MMP
family involved in tendon remodeling are summarized in Table 1.1(Magra and Maffulli
2005). In injured tendon, including acute tears and chronic tendinopathy, increased
expression of MMP1, 2, downregulation of MMP3 and TIMP2,3,4 are found(Ireland,
Harrall et al. 2001; Choi, Kondo et al. 2002; Riley, Curry et al. 2002; Lo, Marchuk et al.
2004). However, TIMP1, which is upregulated in acute tendon tears, is inhibited in
tendinopathy (Ireland, Harrall et al. 2001; Choi, Kondo et al. 2002; Lo, Marchuk et al.

37
2004). The balance of MMPs and TIMPs is important to tendon homeostasis. Over
expression of MMPs will cause the pathogenesis of tendinopathy(Riley, Curry et al.
2002; Lo, Marchuk et al. 2004; Jones, Corps et al. 2006).

38
Tabl
e 1.
1 T
he f
unct
ion
and
targ
ets
of th
e m
atri
x m
etal
lopr
otea
se (
MM
P)
fam
ily
Nam
eS
ynon
ym
Deg
rade
s O
ther
fun
ctio
nsR
efer
ence
MM
P1
Col
lage
nase
-1, I
nter
stiti
al c
olla
gena
se,
Fi
brob
last
col
lage
nase
C
olla
gen
type
I, I
I, I
II, V
II, V
III,
and
X
(G
oupi
lle,
Ja
yson
et
al
. 19
98;
Bra
mon
o,
Ric
hmon
d et
al
. 20
04;
Mag
ra
and
Maf
full
i 200
5)
MM
P2
72 k
Da
gela
tina
se A
ty
pe I
V g
elat
inas
e C
olla
gens
typ
e IV
, V, V
II, X
, and
XI,
Fib
rone
ctin
,ela
stin
, pro
teog
lyca
ns,
gela
tin
Syne
rgis
tic
wit
h M
MP
1
MM
P3
Str
omel
ysin
-1,T
rans
in
Col
lage
ns I
II, I
V, V
, and
IX
,pro
teog
lyca
ns, l
amin
in, f
ibro
nect
in, g
elat
in
Bro
adsu
bstr
ate
spec
ific
ity,
Act
ivat
es p
ro-M
MP
s
MM
P7
Pum
p-1
Gel
atin
, pro
teog
lyca
ns, f
ibro
nect
in, e
last
in, c
asei
n A
ctiv
ates
pro
-MM
P1
MM
P8
Neu
trop
hil c
olla
gena
se
Col
lage
ns ty
pe I
, II,
and
III
, agg
reca
n
MM
P9
92 k
Da
gela
tina
se-B
C
olla
gens
type
IV
, V, X
, XI,
gela
tin
MM
P10
S
trom
elys
in-2
,Tra
nsin
-2
Col
lage
ns ty
pe I
II, I
V, a
nd V
,gel
atin
, fib
rone
ctin
, A
ctiv
ates
pro
-MM
Ps
MM
P11
Stro
mel
ysin
-3
Agg
reca
n, f
ibro
nect
in, l
amin
in
MM
P12
M
acro
phag
e m
etal
loel
asta
se
C
olla
gen
type
s I
and
IV, a
ggre
can,
fib
rone
ctin
, lam
inin
, ent
acti
n, g
elat
in
type
I, v
itro
nect
in, f
ibri
llin,
e la
stin
, c
MM
P13
C
olla
gena
se-3
C
olla
gens
type
I, I
I an
d II
I, g
elat
in

39
1.1.4.3.3 Growth factor in tendon repair
PDGF- is another important cytokine in tendon and ligament repair. It is able to
promote chemotaxis, cell proliferation, ECM production, surface integrin expression,
and revascularization in tendon and ligament (Nakamura, Shino et al. 1998;
Nakamura, Timmermann et al. 1998; Harwood, Goomer et al. 1999). Treatment of
PDGF- in a small animal injury model of tendon significantly improved the enthesis
structure and biomechanical properties (Hildebrand, Woo et al. 1998; Chan, Fu et al.
2006). In a sheep rotator cuff injury model, scaffolds infused with PDGF-achieved
a better histologic score and mechanical properties than an empty scaffold group (Hee,
Dines et al. 2011).
The Transforming growth factor- (TGF-family is essential for tendon
development. It has been shown to stimulate collagen synthesis, cell proliferation and
migration. Furthermore, it is a key growth factor that modulates the scar tissue
following a wound (Chang, Most et al. 1997; Kashiwagi, Mochizuki et al. 2004;
Galatz, Rothermich et al. 2007; Kim, Galatz et al. 2011; Kovacevic, Fox et al. 2011).
In tendon healing, TGF-expression elevates during the inflammatory phase and
promotes collagen production and cell proliferation(Sporn, Roberts et al. 1986;
Kannus and Jozsa 1991), while TGF-is able to reduce scar tissue formation during
tendon repair(Galatz, Rothermich et al. 2007; Kim, Galatz et al. 2011; Kovacevic,
Fox et al. 2011). In a rat rotator cuff injury model, the tendon-bone junction displayed
better mechanical properties in the group supplemented with TGF-compared to

40
the TGF-group(Kim, Kang et al. 2007; Kim, Galatz et al. 2011; Manning, Kim et
al. 2011).
The Bone morphogenetic proteins (BMP) family is part of the TGF-superfamily.
Some BMPs are able to stimulate bone and cartilage formation and induce
mesenchymal stem cell to differentiate into the cartilage or bone linage(Ducy and
Karsenty 2000). However, instead of osteogenic and chondrogenic induction,
BMP-12 is able to induce tendon formation, and is important for tendon healing (Lou,
Tu et al. 2001). Treatment of BMP-12 in a tendon injury model showed a better
organized tendon tissue, higher volumes of collagen type I and improved mechanical
properties(Forslund, Rueger et al. 2003; Majewski, Betz et al. 2008)
The insulin-like growth factor IGF-1 expression increases during the initial
inflammatory phase of tendon healing and is able to stimulate the migration and
proliferation of fibroblasts and inflammatory cells to the wound site. Several small
animal studies have suggested that IGF-1 has the ability to accelerate the tendon
healing by promoting cell proliferation and ECM production (Abrahamsson 1991;
Abrahamsson, Lundborg et al. 1991; Dahlgren, Nixon et al. 2001; Dahlgren, van der
Meulen et al. 2002).
VEGF is not a common cytokine in human tendon, but is expressed in the areas with

41
high microvascular density during tendon healing (Petersen, Pufe et al. 2003). VEGF
treatment has been shown to increase the vessel formation from 1 to 8 weeks after
tendon rupture (Hou, Mao et al. 2009); however, the mechanical properties
improvement was not significant except for in the first week (Zhang, Liu et al. 2003).
bFGF is secreted by tenocytes and inflammatory cells at the injury site of tendon. It
can stimulate cell proliferation, cell migration, collagen synthesis and angiogenesis
(Chan, Chan et al. 1997; Chan, Fu et al. 2000; Thomopoulos, Harwood et al. 2005;
Tang, Cao et al. 2008). Increased cellularity associated with reduced collagen type I
and III expression in injury tendon was observed with additional bFGF in the early
healing process(Chan, Fu et al. 2008; Thomopoulos, Das et al. 2010), while collagen
synthesis levels increased after two weeks treatment, suggesting a prolonged phase of
proliferation can potentially lead to better tendon repair(Sahoo, Toh et al. 2010).
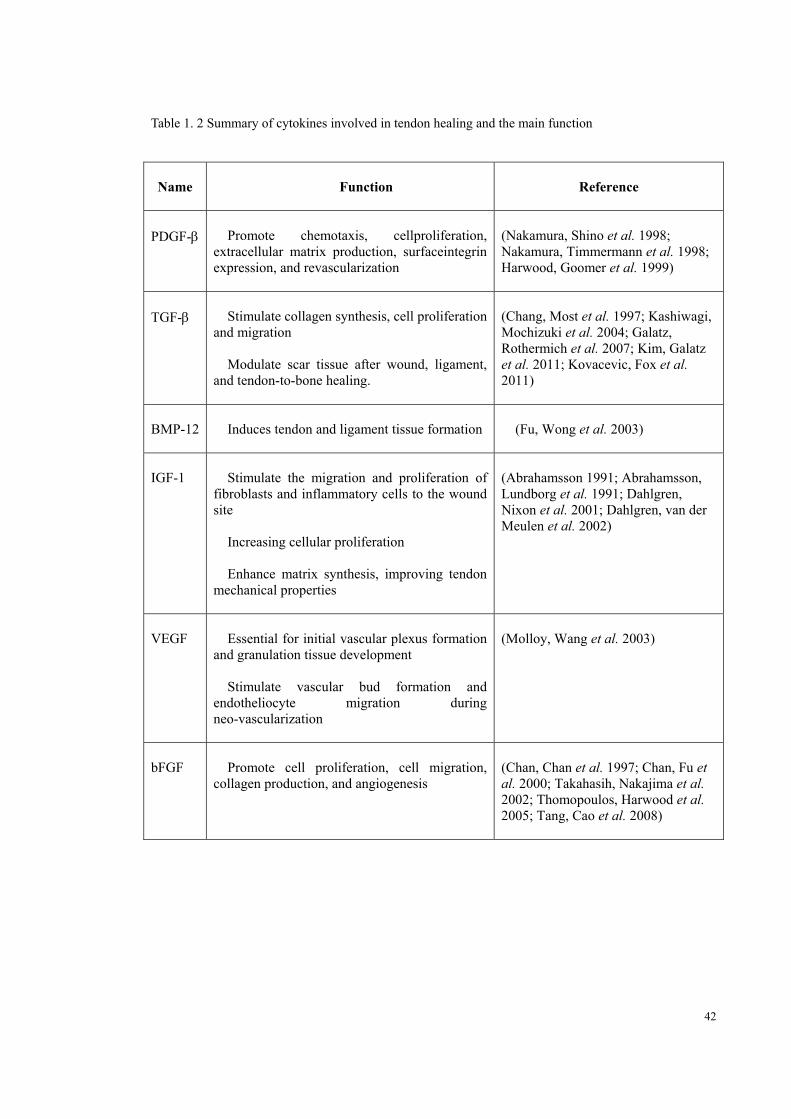
42
Table 1. 2 Summary of cytokines involved in tendon healing and the main function
Name Function Reference
PDGF- Promote chemotaxis, cellproliferation, extracellular matrix production, surfaceintegrin expression, and revascularization
(Nakamura, Shino et al. 1998; Nakamura, Timmermann et al. 1998; Harwood, Goomer et al. 1999)
TGF- Stimulate collagen synthesis, cell proliferation and migration
Modulate scar tissue after wound, ligament, and tendon-to-bone healing.
(Chang, Most et al. 1997; Kashiwagi, Mochizuki et al. 2004; Galatz, Rothermich et al. 2007; Kim, Galatz et al. 2011; Kovacevic, Fox et al. 2011)
BMP-12 Induces tendon and ligament tissue formation (Fu, Wong et al. 2003)
IGF-1 Stimulate the migration and proliferation of fibroblasts and inflammatory cells to the wound site
Increasing cellular proliferation
Enhance matrix synthesis, improving tendon mechanical properties
(Abrahamsson 1991; Abrahamsson, Lundborg et al. 1991; Dahlgren, Nixon et al. 2001; Dahlgren, van der Meulen et al. 2002)
VEGF Essential for initial vascular plexus formation and granulation tissue development
Stimulate vascular bud formation and endotheliocyte migration during neo-vascularization
(Molloy, Wang et al. 2003)
bFGF Promote cell proliferation, cell migration, collagen production, and angiogenesis
(Chan, Chan et al. 1997; Chan, Fu et al. 2000; Takahasih, Nakajima et al. 2002; Thomopoulos, Harwood et al. 2005; Tang, Cao et al. 2008)

43
1.1.5 Treatments
Although our understanding of tendon biology and the healing mechanism has
improved tremendously recently, and various treatments have been adopted for
tendon injury, the outcomes remain unsatisfactory. The goal of treatment for Achilles
tendon diseases is to reduce pain, promote healing and restore its functional
mechanical properties. Surgical and so-called conservative treatments (e.g.
physiotherapy) are the most common procedures. However in recent years
achievements in tissue engineering in animal studies has begun to offer hope for an
alternative solution to tendon injury.
1.1.5.1 Surgical treatment
1.1.5.1.1 Chronic tendinopathy
Surgery is often considered as a last choice for treating tendinopathy. Patients who fail
to improve after a period of conservative treatments tend to be subjected to surgical
treatment, but the results are not uniformly satisfactory. Various surgical procedures
have been described in different studies. They can be divided into four categories: open
tenotomy with abnormal tissue removal, paratenon stripped (Paavola, Orava et al. 2000);
open tenotomy with abnormal tissue removal, paratenon not stripped(Leach, Schepsis et
al. 1992); open tenotomy with longitudinal tenotomy(Rolf and Movin 1997); and
percutaneous longitudinal tenotomy(Maffulli, Testa et al. 1997). In the study of Tallon
et al., of the1648 cases of surgically treated Achilles tendinopathies the success rate was
77.4% (Tallon, Coleman et al. 2001)

44
1.1.5.1.1 Acute rupture
In acute traumatic injuries such as Achilles tendon rupture, early surgery is preferred.
Reconstructive surgery is the common option for athletes, young people, and patients
with chronic ruptures (Christensen 1953; Krueger-Franke, Siebert et al. 1995).
However there is still no agreed protocol for the management of ruptured Achilles
tendons(Maffulli 1995; Nyyssonen and Luthje 2000) or which operative technique
gives the best outcomes (Bugg and Boyd 1968; Hogsaa, Nohr et al. 1990). Ruptured
Achilles tendons can be classified as open operative, percutaneous operative and
non-operative based on the modalities of management. In open surgery, some studies
reported high complication and skin healing problems (Gillespie and George 1969;
Nistor 1981; Bomler and Sturup 1989), while others show little or a low rerupture
rate(Goldman, Linscheid et al. 1969; Inglis and Sculco 1981; Cetti and Christensen
1983; Beskin, Sanders et al. 1987; Zell and Santoro 2000). Percutaneous repair is
another technique that is able to minimizes skin-healing problems under local
anesthetic(Ma and Griffith 1977). However, compared to open repair, percutaneously
repaired Achilles tendons had higher rerupture rates and thinner diameter(Bradley and
Tibone 1990; Cretnik, Kosanovic et al. 2005).
1.1.5.2 Conservative treatment
Conservative treatments are commonly adopted when the clinical symptoms,
including pain and restricted motion, first appear. Current conservative treatments
include rest, non-steroid anti-inflammation drug (NSAIDs), glucocorticosteroid

45
injection, platelet-rich plasma therapy, physiotherapies and physical modalities.
1.1.5.2.1 Rest
In the early stage of injury, rest with proper protection including cast is an effective
treatment to prevent repetitive injury caused by mechanical overload and allow the
self-repair of tendon (Darlington and Coomes 1977).
1.1.5.2.2 NSAIDs
NSAIDs are commonly used to reduce the pain and inflammation in soft-tissue
injuries. NSAIDs are able to block the acute inflammatory response through
non-specific cyclo-oxygenase inhibition. Although pain can be reduced by this
treatment, the use of NSAIDs in tendon injury remains controversial. Various studies
showed that they are potentially deleterious to tissue healing and might decrease the
mechanical properties of tendons (Kulick, Smith et al. 1986; Weiler, Unterhauser et
al. 2002; Forslund, Bylander et al. 2003).
1.1.5.2.3 Glucocorticosteroid Injection
Glucocorticosteroid injection has been used since the 1950s as a pain management
method for tendinopathy. The glucocorticosteroid is injected in and around the
chronic tendon injury. They are reported to reduce pain and recover the range of

46
motion to prevent stiffness (Darlington and Coomes 1977; Blair, Rokito et al. 1996).
Although the improvement following glucocorticosteroid injection is significant, the
effects are often temporary (Coombes, Bisset et al. 2010). There are also several
cases reported that tendon rupture followed corticosteroid injection, including
Achilles tendon rupture (Ford and DeBender 1979; Kleinman and Gross 1983).
Moreover, complications include weakened mechanical properties (Wiggins, Fadale
et al. 1995; Tillander, Franzen et al. 1999) and aggravated pain (Goldfarb, Gelberman
et al. 2007). In chronic tendinopathy, the absence of inflammation provides no basic
target to adopt steroid injection. It is worth mentioning that the study of Kabata et al.
showed steroid injection induced osteonecrosis-like lesions and cell apoptosis around
the lesions (Kabata, Kubo et al. 2000).
1.1.5.2.4 Platelet-rich Plasma (PRP) Therapy
Platelet-rich plasma (PRP) therapy is a technology that aims to deliver bioactive
agents to the injury site, to enable the activation of proliferative and anabolic cellular
response to then enhance the repair mechanism of the tissue(Anitua, Sanchez et al.
2006). PRP has been proposed as a treatment for various orthopaedic disorders and
conditions. Clinical use of PRP on tendon-related injuries and disorders is over a
decade, but the outcomes remained conflicting. There are 2 different Food and Drug
Administration (FDA) approved PRP extraction methods, SMARTPEP (SmartPREP,
Harvest Technologies Corp., Norwell, MA) and Platelet Concentrating Collection
Systems (3i/Implant Innovations, Palm Beach Gardens, FL)(Arora, Ramanayake et al.
2009). To obtain the PRP clinical trial data on human Achilles tendon, key words

47
‘PRP’ and ‘Achilles tendon’ were used in Pubmed, initially, 32 articles were
identified. Of these, 9 were clinical studies, which were summarized in Table 1.3. The
clinical outcome of PRP on Achilles tendon varied widely. In clinical studies,
Sanchez et al. reported that PRP injection followed by surgical repair of ruptured
Achilles tendon showed better and faster recovery at 6 months(Sanchez, Anitua et al.
2007), whilst Schepull et al. reported no significant improvement in biomechanical
tests and a worse clinical outcome compared to the non-injection group over 12
months(Schepull, Kvist et al. 2011). Several case studies suggested that patients
receiving PRP treatment for Achilles partial rupture and tendinopathy showed
significant reduction in pain and clinical improvement(Filardo, Presti et al. 2010;
Gaweda, Tarczynska et al. 2010; Finnoff, Fowler et al. 2011; Owens, Ginnetti et al.
2011; Monto 2012), whilst in randomized trials, de Vos et al. reported clinical
improvement in both groups but no significant different between the PRP injection
group and saline injection group(de Vos, Weir et al. 2010). Despite the fact that the
clinical effect of PRP treatment still remains controversial, no complication has been
reported from its clinical application(Mishra, Randelli et al. 2012). The conflicted
published data implied that the PRP treatment is still immature, but this technique
still has great potential for musculoskeletal medicine and orthopaedic surgery.
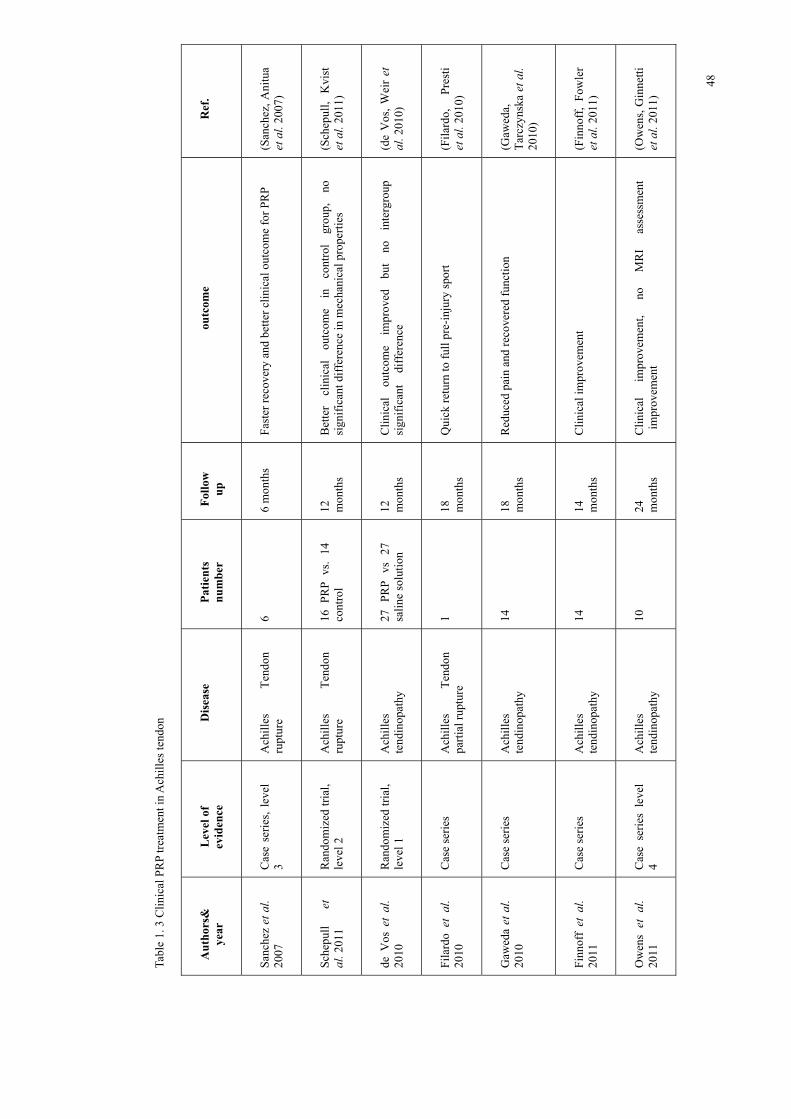
48
Tabl
e 1.
3 C
linic
al P
RP
trea
tmen
t in
Ach
illes
tend
on
Aut
hors
&
year
L
evel
of
evid
ence
D
isea
se
Patie
nts
num
ber
Follo
w
up
outc
ome
Ref
.
San
chez
et a
l. 20
07
Cas
e se
ries
, le
vel
3 A
chil
les
Ten
don
ru
ptur
e 6
6 m
onth
s F
aste
r re
cove
ry a
nd b
ette
r cl
inic
al o
utco
me
for
PRP
(San
chez
, Ani
tua
et a
l. 20
07)
Sch
epul
l et
al
. 201
1 R
ando
miz
ed tr
ial,
leve
l 2
Ach
ille
s T
endo
n
rupt
ure
16
PR
P
vs.
14
cont
rol
12
mon
ths
Bet
ter
clin
ical
ou
tcom
e in
co
ntro
l gr
oup,
no
si
gnif
ican
t dif
fere
nce
in m
echa
nica
l pro
pert
ies
(Sch
epul
l, K
vist
et
al.
2011
)
de V
os e
t al
. 20
10
Ran
dom
ized
tria
l, le
vel 1
A
chil
les
tend
inop
athy
27
P
RP
vs
27
sa
line
sol
utio
n 12
m
onth
s C
lini
cal
outc
ome
impr
oved
bu
t no
in
terg
roup
si
gnif
ican
t d
iffe
renc
e (d
e V
os,
Wei
r et
al
. 201
0)
Fil
ardo
et
al.
2010
C
ase
seri
es
Ach
ille
s T
endo
n pa
rtia
l rup
ture
1
18
mon
ths
Qui
ck r
etur
n to
ful
l pre
-inj
ury
spor
t (F
ilar
do,
Pre
sti
et a
l. 20
10)
Gaw
eda
et a
l. 20
10
Cas
e se
ries
A
chill
es
tend
inop
athy
14
18
m
onth
s R
educ
ed p
ain
and
reco
vere
d fu
ncti
on
(Gaw
eda,
T
arcz
ynsk
a et
al.
2010
)
Fin
noff
et
al.
2011
C
ase
seri
es
Ach
illes
te
ndin
opat
hy
14
14
mon
ths
Cli
nica
l im
prov
emen
t
(Fin
noff
, F
owle
r et
al.
2011
)
Ow
ens
et a
l. 20
11
Cas
e se
ries
lev
el
4 A
chil
les
tend
inop
athy
10
24
m
onth
s C
lini
cal
impr
ovem
ent,
no
MR
I as
sess
men
t im
prov
emen
t
(Ow
ens,
Gin
nett
i et
al.
2011
)

49
Tabl
e 1.
4 C
linic
al P
RP
trea
tmen
t in
Ach
illes
tend
on
Mon
to
et a
l. 20
12
Cas
e se
ries
, le
vel
4 A
chil
les
tend
inop
athy
30
24
m
onth
s S
igni
fica
nt c
lini
cal i
mpr
ovem
ent i
n 28
pat
ient
s (M
onto
201
2)
Fer
rero
et a
l
2012
Cas
e se
ries
A
chil
les
and
pate
llar
tend
inop
athy
30
A
chil
les
tend
on,
28
pate
llar
tend
ons
6 mon
ths
Sig
nifi
cant
cli
nica
l im
prov
emen
t (F
erre
ro,
Fab
bro
et a
l. 20
12)

50
1.1.5.2.5 Physical modalities
Physical modalities such as low-energy shock-wave (Rompe, Furia et al. 2008), laser
(Bjordal, Lopes-Martins et al. 2006), therapeutic ultrasound (Baysal, Bilsel et al.
2006) and heat (Giombini, Di Cesare et al. 2006) have also been used as a treatment
for tendon and ligament injuries. These modalities were suggested to be able to
relieve pain by altering the local vascular system and improve the mechanical
properties by simulating collagen production. Studies showed the success rate of
physical modalities varies widely; the range is from less than 50% to about 80%
(Samilson and Binder 1975; Chard, Sattelle et al. 1988; Hoying and Williams 1996;
Morrison, Frogameni et al. 1997; Goldberg, Nowinski et al. 2001). Due to
recurrence on longer follow-ups, the success rate was reported to drop to around 50%
in recent study (Baysal, Bilsel et al. 2006; Bjordal, Lopes-Martins et al. 2006; Rompe,
Furia et al. 2008). It was suggested that better results are achieved in patients with
minor symptoms.
1.1.5.2.6 Cell Therapy
Cell therapy is the procedure to deliver new cells into a tissue to stimulate the tissue
regeneration. Cell therapy for tendon repair has achieved great success in a rabbit model,
however, the argument about the best cell type in tendon therapy remains inconclusive.
Stem cells, dermal fibroblasts and tenocytes are the most commonly used cell types for
tendon healing (Obaid and Connell 2010). Mesenchymal stem cells are a potential
candidate due to their rapid proliferation, hypoimmunogenicity and multilineage

51
differentiation ability(Uccelli, Moretta et al. 2006). In the past decade promising results
have been achieved using bone marrow-derived stem cells (BMSCs) in tendon repair.
BMSCs therapy in a rabbit model have been reported to stimulate tendon regeneration,
achieve a better neo-tendon morphology and improve tendon biomechanical properties
(Awad, Butler et al. 1999; Ouyang, Goh et al. 2004; Chong, Ang et al. 2007;
Hankemeier, van Griensven et al. 2007). However, the potential risk of ectopic bone
formation needed to be considered(Ross, Duxson et al. 1987). Tendon-derived stem
cells (TDSCs) were first discovered by Bi et al. in 2007(Bi, Ehirchiou et al. 2007), and
they have multi-differentiation potential into musculoskeletal tissue. Ni et al. report that
TDSCs-engineered tendon is able to differentiate into tendon-like tissue and repair the
tendon defect (Ni, Lui et al. 2012; Ni, Rui et al. 2013). Adipose-derived stem cells
(ADSCs) have the advantage of wide availability and are easy to obtain (Obaid and
Connell 2010). Uysal et al. reported that the application of ADSCs in a rabbit tendon
repair model exhibited better tendon healing and biomechanical properties (Uysal and
Mizuno 2010; Uysal and Mizuno 2011; Uysal, Tobita et al. 2012). Synovium-derived
stem cells (SDSCs) were first identified by De Bari et al. in 2001 (De Bari, Dell'Accio
et al. 2001), and they have proven effective in a wide range of musculoskeletal
disorders (Fan, Varshney et al. 2009). The therapeutic effect of SMSCs in the
tendon-bone junction has been reported by Tomita et al. and Ju et al.(Tomita, Yasuda et
al. 2001; Ju, Muneta et al. 2008). Dermal fibroblasts have also been shown to form
tendon tissue (Liu, Chen et al. 2006; Deng, Liu et al. 2009; Woon, Kraus et al. 2011),
although further research has suggested that the healing process using skin-derived
fibroblasts is suppressed with a lack of tenocyte markers and histopathologic
correlations (Chen, Wang et al. 2010; Obaid and Connell 2010). Clinical trials of dermal

52
fibroblasts on lateral epicondylitis (LE) suggested that it a safe and effective treatment
and no significant complication in the majority of patients (Connell, Datir et al. 2009).
Being the native cell source, tenocytes and in situ fibroblasts are arguably the most ideal
cell sources for tendon and ligament therapy respectively. Preclinical and early clinical
studies using these native cell sources are promising, however, the potential morbidity
to the donor site needs to be considered(Cooper, Lu et al. 2005; Lee, Shin et al. 2005;
Webb, Hitchcock et al. 2006; Androjna, Spragg et al. 2007; Chen, Willers et al. 2007;
Freeman, Woods et al. 2007; Moffat, Kwei et al. 2009; Saber, Zhang et al. 2010). In the
most recent clinical trial of autologous tenocyte injection (ATI), patients with chronic
LE showed significant improved function and structural repair after ATI, and no adverse
event was reported at the biopsy sites(Wang, Breidahl et al. 2013).

53
Tabl
e 1.
5 A
sum
mar
y of
cel
l the
rapy
cel
l typ
e fo
r te
ndon
hea
ling
.
Cel
l Typ
e S
ourc
e A
dvan
tage
D
isad
vant
age
Mes
ench
ymal
S
tem
Cel
ls
(MS
C)
Bon
e m
arro
w-d
eriv
ed
1.
Mul
ti-d
iffe
rent
iati
on
abil
ity
incl
udin
g te
nocy
te
2.
Hig
h pr
olif
erat
ion
rate
3.
Hyp
oim
mun
ogen
icit
y
1.
Acc
eler
ate
tend
on h
ealin
g
2.
Impr
ove
mec
hani
cal p
rope
rtie
s of
tend
on
Dif
ficu
lt
to
man
ipul
ate
the
diff
eren
tiat
ion
of
stem
ce
ll
into
de
sira
ble
cell
type
. T
endo
n-de
rive
d 1.
B
ette
r te
noge
nic
diff
eren
tiat
ion
pote
ntia
l
2.
Ten
don-
like
tiss
ue f
orm
atio
n
Adi
pose
tiss
ue-d
eriv
ed
1.
Wid
e av
aila
bili
ty
2.
Eas
y to
ext
ract
3.
No
dam
age
to d
onor
sit
e
Syn
oviu
m-d
eriv
ed
Indu
ce b
one-
tend
on h
eali
ng
Fib
robl
asts
S
kin
1.
Wid
e av
aila
bili
ty
2.
No
dam
age
to d
onor
sit
e
3.
Non
-inv
asiv
e pr
oced
ure
for
cell
har
vest
1.
Dif
fere
nt
cell
type
2.
Unc
erta
in
beha
vior
in
tend
on

54
nich
e
Tabl
e 1.
6 A
sum
mar
y of
cel
l the
rapy
cel
l typ
e fo
r te
ndon
hea
ling
.
Ten
ocyt
e T
endo
n N
ativ
e ce
ll s
ourc
es
Pot
enti
al
mor
bidi
ty
to
dono
r si
te

55
1.1.5.2.7 Physiotherapies
Physiotherapy has been accepted as one of the mainstays of conservative treatment
for chronic tendon injuries. As a force transferring tissue, the biomechanical
environment is essential for tendon homeostasis. Eccentric overloading is a common
treatment for chronic Achilles tendinopathy. There are two types of eccentric
exercises used, maximize the loading on the calf muscle with a straight knee or
eccentrically load the soleus muscle with the knee bent (Alfredson, Pietila et al.
1998). Several studies reported positive effect on symptom relief(Stanish, Rubinovich
et al. 1986; Alfredson, Pietila et al. 1998; Silbernagel, Thomee et al. 2001; Alfredson
and Lorentzon 2003; Fahlstrom, Jonsson et al. 2003; Roos, Engstrom et al. 2004;
Shalabi, Kristoffersen-Wilberg et al. 2004), however, the study of Woodley et al.
showed no effect(Woodley, Newsham-West et al. 2007) and Rompe et al. report an
inferior results(Rompe, Furia et al. 2008). Although the eccentric overloading may be
effective for pain relief and tendon repair, the magnitude of loading and the
underlying mechanism remain unclear.
1.1.5.2.8 Tissue engineering
Tissue engineering’s aim is to grow tissue in the laboratory and usually involves a
procedure that cultures the engineered tissue using different cell sources combined with
various bioscaffolds in vitro. The development of tissue engineering is based on the idea
that instead of repairing the damaged tissue, it may be better to replace it. The detail is
discussed in 1.2.1

56
1.1.6 Summary
Achilles tendinopathy is a degenerative disease that results in the loss of the
functional mechanical properties of tendon and adversely affects the daily activities
of those afflicted. Despite the prevalence of those suffering from this condition, our
knowledge about tendinopathy remains relatively poor. Although the basic
characteristics of tendinopathy that we do understand enable diagnosis, the aetiology
of this degenerative process remains unclear. Moreover, none of the current
treatments, either conservative or surgical, are able to completely or reliably repair
the tendon integrity and restore the tendon mechanical properties. Therefore, further
studies are required to both better understand the disease and to develop effective
treatment strategies.
1.2 Introduction to Bioreactor Design for Tendon/Ligament
Engineering
(This section has been published in ‘Tissue Engineering Part B’ titled “Bioreactor
design for tendon/ligament engineering” PDF refer to appendix 1)
Autograph and allograft transplantations are a common surgical treatments for tendon
and ligaments injured or degenerated. However, the risks of damage to the donor site
from which the autograft are taken, and the potential immune reaction for allografts are
major concerns (Coupens, Yates et al. 1992; Cerullo, Puddu et al. 1995; Harner, Olson
et al. 1996). A promising translational approach to the treatment of tendon/ligament

57
injury or degeneration is through the use of engineered autologous grafts made available
through the development of bioreactors that generate tendon/ligament tissue in vitro.
One common view is that the key to a successful bioreactor is being able to recreate, in
vitro, the cell microenvironments that are experienced by cells in vivo. The cell
microenvironments can be defined using cell morphological information with data from
molecular biology, biochemistry and biomechanics. This review aims to clarify the
requirements for a ‘successful bioreactor’ that may be used for tendon/ligament
engineering, and to provide an overview of the range of components found in
tendon/ligament bioreactors, including custom-made and commercial products. We will
also discuss the studies that have involved the application of tendon/ligament
bioreactors.
1.2.1 Common key elements for tendon/ligament tissue engineering
Based on the composition and function of tendon/ligament tissue one must consider the
four basic elements for their successful regeneration: the cell source, the characteristics
of the scaffold matrix and establishing an appropriate chemical and physical cellular
microenvironment.
An ideal cell source should meet the following three requirements: availability, rapid
proliferation and the ability to differentiate into in situ cells(Arnsdorf, Jones et al. 2009).
The details refer to 1.1.5.2.6.

58
Apart from the selection of the cell source, cell seeding also plays an essential role in
the development of engineered tendon and ligament. Several reports indicate that
sufficient cell number and a uniform distribution throughout the scaffold is desirable for
achieving a homogeneous ECM deposition in vitro (Freed, Marquis et al. 1993; Freed,
Vunjak-Novakovic et al. 1993). Compared to lower initial cell seeding density, high
seeding density has been shown to result in increasing ECM deposition rate, a higher
final cell number and better cellular morphology (Awad, Butler et al. 2000; Wang,
Seshareddy et al. 2009). However, over-seeding also has potential for negative effects
on nutrient delivery, and consequently cellular metabolism and cell viability. Nutrient
depletion at high cell seeding densities could lead to spatially in homogenous ECM
production (Zhang, Gardiner et al. 2008). A study by Issa et al. showed
mechano-stimulated human umbilical veins seeded with 3 million cells/ml had better
cellular proliferation rates than other groups(Issa, Engebretson et al. 2011). However,
optimal seeding density will vary depending on the cell source and the bioscaffold
physical and chemical properties, as these properties will affect nutrient transport and
rates of consumption, as well as the mechanical and chemical stimuli provided to the
attached cells(Zhang, Gardiner et al. 2008).
The construct scaffold plays an important role in engineering the new tendon/ligament
tissue. Ideally the scaffold should be able to provide substantialinitial mechanical
strength for its immediate post-implantation functional role, while providing a suitable
biological environment for cell migration and proliferation. Furthermore, the
degradation rate of the biomaterial needs to be comparable with the rate of tissue

59
synthesis, to allow the eventual replacement of the starting scaffold with neo-tissue.
Both synthetic and natural biomaterials are commonly used in tendon/ligament
engineering. Synthetic polymer scaffolds have the advantage of reproducible
mechanical and chemical properties, and they are relatively easy to fabricate into
different sizes(Lee, Shin et al. 2005; Pham, Sharma et al. 2006; Sahoo, Ouyang et al.
2006; Sahoo, Cho-Hong et al. 2007; Moffat, Kwei et al. 2009). However, their rapid
degradation rate and potential risk of releasing acidic by-products or toxic polyesters
during degradation have limited their application in clinical trials. Given these
disadvantages, more researchers have turned their focus on exploring natural
biomaterials (Derwin, Baker et al. 2006; Juncosa-Melvin, Shearn et al. 2006; Chen,
Willers et al. 2007; Gilbert, Stewart-Akers et al. 2007; Nirmalanandhan, Rao et al. 2008;
Nirmalanandhan, Shearn et al. 2008; Fleming, Spindler et al. 2009; Kinneberg,
Nirmalanandhan et al.). Being the main component of native tendon, collagen type I is
the most obvious choice of material. Although the biocompatibility is excellent, the
poor mechanical properties of reconstituted type I collagen scaffolds has limited their
further development as a load bearing material. Silk fibroin, on the other hand, has
similar biocompatibility as collagen scaffolds and comparable mechanical properties as
native tendon/ligament. In several in vivo studies, silk fibroin-based engineered
ligaments have been proven their ability to restore the function of injured ligament
(Chen, Qi et al. 2008; Fan, Liu et al. 2008; Fan, Liu et al. 2009). Another option is the
decellularised tendon/ligament construct. Although the mechanical and biological
properties are a better match to native tissue than any other currently available scaffolds,

60
donor cells may remain in the allograft, even with strict sterilization and cleaning, and
thus they can potentially cause inflammatory responses (Malcarney, Bonar et al. 2005;
Zheng, Chen et al. 2005).
After choosing an appropriate cell source and scaffold type, the tissue needs to be
encouraged to develop the properties of native tissue by providing an appropriate
biochemical and biomechanical environment to stimulate ECM synthesis. Regarding the
biochemical environment, several growth factors have been found to play an important
role in tendon/ligament formation and healing. These include Insulin-like growth
factor-I (IGF-I), vascular endothelial growth factor (VEGF), platelet-derived growth
factor (PDGF), basic fibroblast growth factor (bFGF), transforming growth factor
(TGF) and growth differentiation factor 5 (GDF-5).The roles of TGFand GDF-5
seem to be particularly prominent. TGFremains active throughout tendon/ligament
healing(Chang, Thunder et al. 2000), and is able to regulate cell migration, proteinase
expression, fibronectin binding interactions, cell proliferation and collagen production
(Bennett and Schultz 1993; Wojciak and Crossan 1994; Marui, Niyibizi et al. 1997;
Chang, Thunder et al. 2000). Recent studies demonstrated that tendon and ligament
formation was impaired in TGF2 and TGF3 knockout mouse embryos, reinforce the
importance of TGFs in tendon development and homeostasis(Pryce, Watson et al.
2009). GDF-5 regulates cell growth and differentiation, with a lack of GDF-5 causing
delayed tendon healing, irregular collagen type I fibrils and weakened fibril mechanical
properties(Clark, Johnson et al. 2001; Mikic, Schalet et al. 2001; Chhabra, Tsou et al.
2003).

61
How these various biochemical factors should be introduced into the tissue bioreactor
system to shape the chemical environment is an extremely challenging and open
question. Specifically, at what concentrations, in what combinations and at what
sequence or timing should they be made available? Studies have shown that the
expression of these factors in tendon repair change over periods of days (Dahlgren,
Mohammed et al. 2005; Kobayashi, Itoi et al. 2006; Chamberlain, Crowley et al. 2009).
Presumably as the engineered tissue progresses, cells begin to control their own
biochemical environment, and the role of the bioreactor is to now provide the ‘building
block’ nutrients and expected systemic signals, along with mechanical stimulus. The
early stage in the tissue-engineered tendon is likely to be the most critical in
establishing tendon development along a pathway to resemble native tendon. Finding
the correct combination of factors is daunting due to the complexity arising from the
multitude of possible combinations and interactions. Systematically varying
experimental conditions, coupled with computational modeling of transport processes
and signaling molecule pathways leading to cell responses, provide the only
conceivable means to both understanding and efficiently optimizing the tissue bioreactor
system.
Tendon’s primary function is mechanical. It operates in a varying load environment,
both on short timescales (e.g. walking, running) and on longer timescales (e.g. changes
in body size with age). Tendon responds to its mechanical environment through changes
in ECM biosynthesis and degradation. Unsurprisingly, given its functional role, a
suitable mechanical stimulus is vital for tendon/ligament homeostasis. In fact, it has

62
been shown that after 4 weeks in a load-free culture environment tenocytes lose their
native elongated morphology, become increasingly rounded, and the collagen fiber
becomes more crimped (Hannafin, Arnoczky et al. 1995). In cell-free reconstructed
collagen fibril network systems, a tensile load has been shown to be protective to
degradation by MMP-8 (Flynn, Bhole et al. 2010). Furthermore, cyclic stretching has
been shown to produce an up to 9 fold increase in the cell number of an engineered
tendon compared with a static culture over a 2 week period(Abousleiman, Reyes et al.
2009). Finally, an appropriate mechanical environment could help guide collagen fiber
formation, i.e. along the direction of loading, which is able to enhance or optimize the
mechanical properties including stiffness, elastic modulus, maximum tensile stress and
maximum force(Juncosa-Melvin, Shearn et al. 2006; Saber, Zhang et al. 2010; Woon,
Kraus et al. 2011).
Rather than being two separate signals, there is crosstalk between mechanical and
chemical signals. Recent studies have shown that gradual and temporary loss of tensile
loading leads to reversible loss of Scleraxis (Scx) expression, which is a transcription
factor specific for tenocytes and their progenitors. In addition, it has been shown that
TGF directly induced the expression of Scx in cultured tenocytes isolated from mice
(Maeda, Sakabe et al. 2011). In Scx-/- mice, a disordered limb tendon phenotype was
observed (Lejard, Brideau et al. 2007), and similar phenomenon happened in TGF
type II receptor gene knock out (Tgfbr2-/-) mice with dramatic loss of Scx
expression(Pryce, Watson et al. 2009).

63
Providing a suitable biomechanical signal is clearly an important component for the
success of tendon/ligament engineering. Generating a suitable mechanical signal within
the bioreactor system is critically important for tendon/ligament tissue engineering.
1.2.2 Bioreactor design specific to tendon/ligament engineering
Despite the increasing appreciation of tendon/ligament biology and function,
conventional culture methods do not seem to meet the biochemical and biomechanical
requirements to generate bioengineered tendon/ligament in vitro. A bioreactor system
that subjects the ‘cell culture’ to programmable mechanical stimulation (PMS),
mimicking the physiological conditions of tendon/ligament in vivo, while allowing
cellular proliferation, differentiation and matrix production in a mechanical environment,
may provide a solution.
Bioreactors for tendon/ligament engineering are different to the systems that have been
used in various other tissue engineering fields in the past decades, i.e. systems for
muscle(Dennis, Smith et al. 2009), liver(Catapano, Patzer et al. 2010) and bone(Rauh,
Milan et al. 2011; Salter, Goh et al. 2012). Compared to other bioreactors, the main task
of the bioreactor for tendon/ligament engineering is to provide the proper biomechanical
and biochemical environment specific to tendon/ligament formation. In order to achieve
this, certain basic components are required, i.e. the actuating system and the culture
chamber, which can provide the construct’s PMS and controlled culture environment
respectively. Furthermore, the bioreactor may also include a medium circulation system,

64
monitoring system, feedback system, and a medium analysis system, depending on the
operational requirements (Figure 1.1). With these facts in mind, several custom-made
bioreactors have been developed for tendon/ligament engineering (see Table 1.1), and
the aforementioned components of these are now are discussed in detail.
Figure1.1 Schematic demonstration of connection between different components of tendon/ligament bioreactor system

65
Table 1.1 Components of custom-made bioreactor systems
Reference Actuating system
Culture chamber
Monitor system
Feedback system
Medium circulation system
Medium analysis system
(Altman, Lu et al. 2002)
Biaxial Multiple
(Webb, Hitchcock et al. 2006)
Uniaxial Single chamber, multiple samples
(Juncosa-Melvin, Shearn et al. 2006),(Nirmalanandhan, Rao et al. 2008),(Nirmalanandhan, Shearn et al. 2008),(Butler, Gooch et al. 2010)
Uniaxial Multiple Displacement
(Androjna, Spragg et al. 2007)
Uniaxial Multiple Force
(Nguyen, Liang et al. 2009)
Uniaxial Single Force
(Abousleiman, Reyes et al. 2009)
Uniaxial Multiple
(Butler, Hunter et al. 2009)
Uniaxial Single chamber, multiple samples
Displacement Force
(Chen, Yin et al. 2010)
Uniaxial Multiple
(Doroski, Levenston et al. 2010)
Uniaxial Multiple Displacement
(Parent, Huppe et al. 2011)
Uniaxial Multiple Displacement Force

66
1.2.2.1 Actuator and culture chamber design
The actuating system is the main component for providing different PMS to engineered
tissue. Pneumatic actuators(Juncosa-Melvin, Shearn et al. 2006; Nirmalanandhan, Rao
et al. 2008; Nirmalanandhan, Shearn et al. 2008), linear motors(Nguyen, Liang et al.
2009; Doroski, Levenston et al. 2010) and step motor-ball screws(Altman, Lu et al.
2002; Webb, Hitchcock et al. 2006; Chen, Yin et al. 2010) are the most common
actuators used in tendon/ligament bioreactors. Pneumatic actuators have several
advantages, including the ease of maintenance, cleanliness, low cost and high
power-to-weight ratio(Varseveld and Bone 1997). Unfortunately, by using air as a media,
pneumatic actuators are subject to high friction. The sensitivity and response to an input
signal is relatively slow because of the “dead band” and “dead time” caused by stiction
and air compressibility. Due to these nonlinearities, it is difficult to achieve accurate
position control with pneumatic actuators(Varseveld and Bone 1997). Typically, the
accuracy of pneumatic actuators is approximately0.1mm, which is not insignificant
when typical tendon bioreactors require <5% strain on 1-5cm tissues. Compared with
pneumatic actuators, electrical actuators are more expensive, but the level of accuracy in
their positional control is much higher. A direct-drive linear motor is able to provide
high-speed/high-accuracy linear motion by eliminating mechanical transmission(Alter
and Tsao 1994; Alter and Tsao 1996; Braembussche, Swevers et al. 1996), and the
accuracy is the highest of all three actuators; i.e. ~1µm. Step motor-ball screw
transmission systems (SMBS) are based on a ball-screw linkage and a crank-slider
mechanism. In general, SMBS transfer the rotation of the crank to a reciprocating
motion of the screw(Liu, Hsu et al. 2001). By choosing different ball screws, the
optimal speed range and output force can be selected using Equation (1), i.e.

67
vFlT2
(1)
whereT is torque applied to screw, F is linear force, l is ball screw lead, and ν is ball
screw efficiency. Although SMBS is not as accurate as a linear motor, a positional
accuracy of around 5µm can still be achieved. In a multi-chamber shared loading
system, SMBS is widely used given its high accuracy and relative high loading capacity
(Altman, Lu et al. 2002; Webb, Hitchcock et al. 2006; Androjna, Spragg et al. 2007). In
independent loading multi-chamber systems, linear motors are more popular due to their
small size and relatively simple mechanical arrangement(Nguyen, Liang et al. 2009;
Parent, Huppe et al. 2011).
In the addition to the actuator, the connection between the mechanical input and the
tissue is vital and often presents a major challenge. During PMS of the tissue in culture
an even distribution of force throughout the entire sample is critical, otherwise tissue
integrity is compromised by overloading the mechanical connection regions and/or by
inhomogeneous mechanical stimulation. Different strategies of applying PMS have been
adopted based on different construct dimensions. In a study by Chen et al., cell-seeded
knitted silk-collagen sponge scaffolds were fixed on stainless rings and connected to
sample hooks(Chen, Yin et al. 2010). The bioreactor system by Juncosa-Melvin et al.
applied two posts to fix the construct, punching through the scaffold as shown in Figure
1. 2(Juncosa-Melvin, Shearn et al. 2006). However, non-uniform construct deformation
is clearly apparent. Tensile force is focused on the side of constructs, which causes
uneven distribution of mechanical stimulation. Although tissue clamps are the most

68
popular and relatively effective method for holding tendon constructs(Webb, Hitchcock
et al. 2006; Nguyen, Liang et al. 2009; Parent, Huppe et al. 2011), the potential for
damage at the clamping region needs to be considered. The method of reproducibly
applying uniform loads to soft tissue without tissue damage or slippage is a critical
problem in need of a satisfactory solution. It is our opinion that a robust clamping
region should be designed along with the artificial bioscaffold to ensure the proper
connection between the sample and the PMS.

69
Figure 1.2 Stem cell seeded Collagen sponge deformation during mechanical stimulation. Modified
from reference 156
The culture chamber is an essential part of whole bioreactor system. The high humidity
of culturing conditions (99% humidity, 37℃ and 5% CO2) and chemistry of the culture
medium are corrosive to many materials. Corrosion products may in turn be toxic to the
tissue. Therefore, noncorrosive and autoclavable material such as stainless steel,
polymethylemethacrylate (PMMA), polyoxymethylene, polycarbonate, glass and silicon
are preferred and widely used in culture chamber design (Altman, Lu et al. 2002;

70
Abousleiman, Reyes et al. 2009; Butler, Hunter et al. 2009; Chen, Yin et al. 2010;
Doroski, Levenston et al. 2010).
The chamber structure is an important consideration in the bioreactor design, with most
currently available culture chambers divided into two groups: integrated(Webb,
Hitchcock et al. 2006; Butler, Hunter et al. 2009) and separated
chambers(Juncosa-Melvin, Shearn et al. 2006; Chen, Yin et al. 2010; Doroski,
Levenston et al. 2010) (Altman, Lu et al. 2002; Androjna, Spragg et al. 2007; Parent,
Huppe et al. 2011) (Butler, Gooch et al. 2010). In integrated chambers, multiple
samples are cultured while sharing the same culture medium. Conversely, separated
chambers can provide separate culture environments for each sample. Although the
complexity of design and manufacturing costs may be higher in a separated chamber
system, reduced cross-contamination and the option of independent environmental
control are distinct advantages.
During tissue culture, sufficient air exchange within the culture chamber is critical.
Air exchange in conventional cell culture incubators is through the integrated
hydrophobic filter of the culture flask and the gap between the leak and the culture
dish/well plate. Therefore, the ideal design for the culture chamber should be similar.
Like conventional cell culture, an unsealed chamber bioreactor connects to the
outside environment through various ways (Juncosa-Melvin, Shearn et al. 2006;
Androjna, Spragg et al. 2007; Nguyen, Liang et al. 2009; Doroski, Levenston et al.

71
2010), such as the hydrophobic filter leak(Webb, Hitchcock et al. 2006) and the
labyrinth channel (Figure 1.3)(Parent, Huppe et al. 2011). In a study by Webb et al.
(Webb, Hitchcock et al. 2006), a modified tissue culture flask was used as an
integrated culture chamber, and could culture up to four samples simultaneously.
Although there are potential risks of cross-contamination from different samples and
toxicity from autoclaving the culture flask, the integrated hydrophobic filter leak can
ensure adequate gas exchange without inducing contamination. In the bioreactor
system used in the research of Parent et al.(Parent, Huppe et al. 2011), a labyrinth
channel was added to improve the air exchange and eliminate contamination as
shown as Figure 1.3. However, in closed chambers air exchange mostly depends on
medium circulation, which is now discussed in the following section.
Figure 1.3 Schematic drawings of air exchange through the labyrinth channel in the culture chamber. Modified
from reference (Parent, Huppe et al. 2011)
1.2.2.2 Environmental control and medium circulation systems
Although to the best of our knowledge, no contamination has been reported in any
bioreactor study, transportation of the bioreactor and opening of chamber for medium
exchange every three days, especially for multi-chambers system, is still a potential

72
contamination risk. Therefore, a medium circulation system can be introduced to
improve the efficiency and minimize these risks. In addition to the advantages of
reduced contamination, a circulating medium may be better able to infiltrate into
cultured tissue. For example, perfusion bioreactors used in bone engineering circulate
culture medium for better nutrient delivery and subsequent improved cell numbers
(Cartmell, Porter et al. 2003; Uemura, Dong et al. 2003; Meinel, Karageorgiou et al.
2004; Holtorf, Jansen et al. 2005; Janssen, Oostra et al. 2006; Olivier, Hivart et al.
2007). Similarly, human umbilical vein cultured under a circulating medium had
approximately three times the cellular number as those cultured using a quiescent
medium(Abousleiman, Reyes et al. 2009).
A traditional incubator based bioreactor and/or independent bioreactor can be used for
tendon/ligament engineering. In an incubator bioreactor system, air exchange is through
the hydrophobic filter leak of the medium reservoir, and proper CO2 and temperature
level is controlled by the incubator. Then by circulating the culture medium, suitable
conditions can be applied to engineered tendon/ligament, as shown in Figure 1.4
(Abousleiman, Reyes et al. 2009) . However, an independent bioreactor system, as
shown Figure 1.5, does not rely on an incubator to control the culture environment. The
percentage of different gases (PO2, CO2 and N2) are control by air valves (Altman, Lu et
al. 2002), and the mixed gas is humidified first, and then passed into a medium heater.
Waste gas emission is transported through a filter in the case of contamination. The
prepared, warm culture medium is circulated through the bioreactor culture chamber.
For certain tissues such as cartilage, some specific culture conditions are required. For

73
example, under a low oxygen environment, engineered cartilage displays faster matrix
glycosaminoglycan deposition rate, and better cellular morphology but with less
dedifferentiation (Hansen, Schunke et al. 2001; Domm, Schunke et al. 2002; Saini and
Wick 2004). The environmental chamber allows researchers to manipulate different
culture conditions thereby enabling a systematic study of cell growth and differentiation
into functional tissue.

74
Figure 1.4 Schematic illustration of an incubator bioreactor system. Suitable temperature,
humidity and CO2 level of culture medium are maintained by the incubator in the medium reservoir.
The medium is circulated by a pump. The waste valve is closed normally and it will open during
medium exchange.

75
Figure 1.5 Schematic diagram of an independent bioreactor system. Suitable temperature, humidity
and CO2 level of the culture medium are control by the environmental chamber. Cold culture
medium is pumped to the environmental chamber for heating and then circulated through the culture
chamber. The waste valve is closed normally and it will open during medium exchange.
1.2.2.3 Monitoring and feedback systems
The biomechanical properties of the final engineered tendon/ligament should be a
central concern of bioreactor design, as the tendon/ligament’s functional role in the
body is primarily mechanical. The maximum load and elastic modulus are essential
mechanical properties to evaluate the suitability of engineered tendon/ligament.
However, in most studies, mechanical tests are performed only at the end of tissue
culture. For example, the stiffness of the constructs at different time points during

76
culture is rarely recorded or assessed. The correlation between stiffness and tissue
maturation may provide a better understanding about how the cell differentiates into
functional tissue, and for this reason ‘on-line’ stiffness monitoring is likely to be
invaluable.
In tendon/ligament bioreactors force measurement is enabled using sensors called load
cells (Butler, Juncosa-Melvin et al. 2008; Nguyen, Liang et al. 2009; Parent, Huppe et
al. 2011), which are located differently in various bioreactor designs. Load cells located
between the actuator and sample clamps(Butler, Hunter et al. 2009), shown in Figure
1.6A, requires high manufacturing accuracy, as the friction between the shaft and
culture chamber can induce error. For fragile material like biological tissues, this
friction might be higher than the applied load. Conversely, a load cell placed at the end
of the culture chamber, shown in Figure 1.6B, can minimize the fiction between the
actuator and culture chamber and also the fluid resistance during stimulation. In order to
acquire accurate data, load cell selection should be based on the initial mechanical
properties of the constructs. Working with the sensitivity of the load cell, extra attention
is needed when manipulating the construct so as to avoid causing damage to the load
cell through overloading.

77
Figure 1.6 Load cells position in different bioreactor systems. A. load cell is located between the actuator and sample clamps. B. a separate load cell placed at the end of culture chamber system.
In addition to force monitoring, tissue displacement is another important variable that
requires monitoring. Linear variable differential transformers (LVDT) and optical
decoders are commonly used position sensors(Juncosa-Melvin, Shearn et al. 2006;
Parent, Huppe et al. 2011). Although the motion of actuating systems is preprogrammed,
overload of the actuator and manual mis-operation can cause desynchronization
between the program and actual stimulation. The real time displacement monitor can
produce a full record of stimulation position, which then allows researchers to track if
there are any unusual features in the results. Another function of the position sensor is to
provide feedback of the displacement information to the actuator control system to
correct for any desynchronization(Parent, Huppe et al. 2011). Critically, by monitoring

78
load and displacement, real time stiffness during engineered tendon/ligament culture
can be estimated.
Lastly, an imaging component might be useful in the bioreactor system to monitor the
formation of the tissue construct at high resolution and possibly provide information to
the feedback system. Two imaging modalities, including the confocal microscope and
optical coherence tomography (OCT), are potential candidates (Drexler, Morgner et al.
1999; Goetz, Thomas et al. 2008). Both of these techniques are able to acquire
high-resolution image and might be adopted in the dynamic culture environment for
observation of cell and tissue morphology (Drexler, Morgner et al. 1999; Goetz, Thomas
et al. 2008). The magnification of the confocal microscopy may range from 500 to 2400
fold at a dynamic environment. A miniaturized confocal laser scanning probe has been
tested in rodents to provide the visualization of the vascular circulation and cell perfusion
Compared to the confocal microscope, OCT has a higher penetration depth up to 1~2
mm and longer working distance (the distance from the objective lens to the sample
surface) up to 10 mm, which is sufficient for distant imaging of the tissue constructs in
the bioreactor system(Smithpeter, Dunn et al. 1998; Drexler, Morgner et al. 2001;
Burkhardt, Walther et al. 2012) . However, there are two major issues to be addressed
before these systems could be integrated into the bioreactor design. Firstly, for distant
imaging, the optical light needs to penetrate3 layers of media (chamber cover, air and
culture medium) with different refraction index to obtain imaging. Secondly, sample
deformation induced by dynamic mechanical loading can cause image shift resulting in
poor focus or loss of the image. Imaging dynamic sample is extremely difficult in

79
conventional bioreactors, as features of interest (e.g. cells) may leave the field of view.
Techniques need to be developed to move the microscope alone with the sample to
achieve optimal focus on the view of interest. Other than distant imaging, minimally
invasive imaging technique might provide another potential strategy for this obstacle.
Recent studies show that confocal fluorescence microendoscopy and OCT could be
integrated into a hypodermic needle (Pillai, Lorenser et al. ; Quirk, McLaughlin et al.),
which might be inserted into the tissue constructs and thus creating a relatively static
imaging environment. However, the technique is invasive and could damage the tissue
integrity. In all, the current imaging technology is not mature enough to acquire the
dynamic sub-micrometer image, butan imaging system allows dynamic imaging at high
resolution is certainly worth to explore. Cell morphological and tissue structural change
coping to the PMS might provide a different respective for studying the effect of PMS
on tendon/ligament biology and tendon/ligament engineering.
1.2.3 Commercial bioreactor systems for tendon/ligament engineering
Recently, commercial tendon/ligament engineering bioreactor systems have become
available. As discussed above, these basic principles are applied to customized
bioreactors; however, with greater design input and advanced manufacturing techniques,
commercial products are able to provide more accurate and complex environments for
tendon/ligament culture. To our knowledge, two relatively complete commercial
bioreactor systems have been developed recently: The Bose® ElectroForce® BioDynamic®
system and the LigaGen system.

80
The Bose® ElectroForce® BioDynamic® test instrument provides an accurate
programmable uniaxial stretching stimulation and a controllable medium circulation
environment to engineered tendon/ligament, which, theoretically, can be adjusted to
mimic the in vivo biomechanical environment. With a load cell and optional laser
micrometer, this bioreactor system is able to monitor the force/strain curve of
engineered tendon/ligament during the culture period. Two different force and
displacement ranges are available. Single chamber and multiple chambers systems with
shared or independent loading are optional. As there are culture chambers compatible to
the Bose® testing devices such as ElectroForce 3200, biomechanical tests can be done
at different time points without disruption of the tissue culture
(www.bose-electroforce.com).
The LigaGen system is a lightweight (<3kg) incubator compatible bioreactor. It is
capable of applying a maximum force of 40N to the tissue sample, and simulate
complex, and presumably more physiologically realistic, loading patterns. Two systems
are available from LigaGen. L30-1X is a single culture model, which has a 23ml
internal volume chamber for single tissue culture, and the L30-4C is a multiple culture
model with an 80ml chamber for shared dynamic culture on two or four samples. The
standard medium circulation system can reduce contamination risk during medium
exchange. Rather than being a comprehensive system, the bioreactor is extensible to
suite individual needs, by adding various components to the (universal) basic model as
required. With an accessory tissue monitoring sensor, this system is able to achieve
real-time measurements of the sample stiffness during culture. If flow control is
necessary, extra control systems can be installed (http://www.tissuegrowth.com/).

81
However, there are some disadvantages in the commercial bioreactors. Firstly, the
fixation mechanism to stabilize the tendon tissue in the bioreactor cannot be adjusted.
Tissue clamps provided in the commercial bioreactor systems can become less effective
when it comes to the use of a cylindrical scaffold. Secondly, capacities of the chamber
and mechanical input are limited and they can only host small animal tissues. This may
restrict clinical development. Moreover, some ligaments, such as the anterior cruciate
ligament, are not only subjected to tensile force, but also to rotational loading, and none
of the commercial bioreactor systems provide the addition of torsional loading. Lastly,
full-scale commercial bioreactor systems are expensive, and for most of the time, not all
of the functions they provide are commonly used in every study.
1.2.4 Ideal bioreactors for tendon/ligament engineering
The ideal bioreactor should be able to culture tendon- and ligament-like constructs,
which are well-organized, cell-seeded, assemblies of collagen bundles with mechanical
properties functionally similar to the native tissue. Autologous cell-seeded constructs
are biological compatible, and able to provide mechanical support similar to native
tissue, and consequently they are widely studied in tendon/ligament engineering.
However, induction of cell directed collagen fiber reorganization and assembly of
collagen bundles are two important impediments to this approach. PMS can help
provide the necessary signals to cells to increase collagen synthesis, spatially organize
of the collagen along the primary stress direction, and stabilize collagen from
collagenase degradation (Lavagnino, Arnoczky et al. 2003; Yang, Im et al. 2005;
Nguyen, Liang et al. 2009). Moreover, proper PMS can upregulate different

82
proteoglycans, such as decorin, biglycan, fibromodulin and fibronectin, which help the
cells organize the parallel collagen fibrils forming bundles (Scott 1984; Cribb and Scott
1995; Vogel 2004; Webb, Hitchcock et al. 2006; Franchi, Trire et al. 2007).
The host body is in many ways the ultimate bioreactor for all engineered tissues. The
study of Juncosa-Melvin et al. indicated that the maximum force of engineered patellar
tendons increased more than 3000 times after 2 weeks implantation in rabbits
(Juncosa-Melvin, Shearn et al. 2006), and to date none of the bioreactors have been able
to accomplish this outcome. Therefore, an ideal bioreactor should aim to mimic the
dynamic biochemical and biophysical environments in vivo. In musculoskeletal tissue
engineering, various bioreactors have been developed. For instance, muscle tissues are
not only subjected to mechanical stretching but also able to receive the electrical
impulses to simulate inputs from the central nervous system (Ross, Duxson et al. 1987),
and mechanical and electrical stimulation bioreactors have been developed based on
mimicking the in vivo environment (Donnelly, Khodabukus et al. 2010; Sharifpoor,
Simmons et al. 2011). Compared to muscle, the in vivo environment is comparatively
less complex in tendon/ligament and appears to require only passive mechanical input.
Summarizing, the ideal tendon/ligament engineering bioreactor that enables systemic
research should integrate all aforementioned components (Figure 1.1). The bioreactor
should have culture chambers and an actuating system, but also be fitted with a medium
circulation system, an environmental system, a monitoring system, a feedback system,

83
and a waste medium analysis system. This bioreactor should first be able to provide not
only a multiple, suitably sized and sterilized chambers for tissue culture, but also
accurate and programmable mechanical stimulation. Tensile strain and rotation are
needed to mimic different in vivo tendon/ligament loads. Second, the circulated medium
should infiltrate into tissue better than a static medium configuration, and the circulation
system needs to reduce the risks of contamination during medium exchange and drug
delivery. Moreover, environmental control, such as PO2, CO2 and pH level, allows
researchers to explore the impact of different culture conditions on tissue maturation.
Third, a monitoring system should provide the real-time status of cultured
tendon/ligament, such as force and displacement, and based on these data, adjustment of
PMS by use of a feedback system. Fourth, through analysis of the waste medium,
nutrient consumption needs to be evaluated, which may enable the changing of the
medium base at different stages as required, rather than fixed, regular medium exchange
every 3 days. Finally and importantly, the best patterns of PMS for culturing engineered
tendon/ligaments need to be defined.
1.2.5 Previous work in this field
Since the first 3D engineered tendon/ligament bioreactor system published by Altman et
al. in 2002, the effect of PMS on the engineered tendon/ligament has drawn a lot of
research attention. In the past decade, dynamic loading of culture in bioreactor systems
has been proven to have been a significant development in producing an engineered
tendon/ligament. Indeed, various studies have been performed using bioreactor systems
and have met with considerable success, and these are summarized in Table 1.2.

84
Compared with static culture, the tissue produced using PMS has better cell morphology,
including elongated cellular morphology and increased cell density. The mechanical
properties of engineered tendon/ligament, such as tensile strength and elastic modulus,
are also greatly improved by cyclic loading of the tissue culture, as is the microstructure
of the extracellular matrix, such as collagen fiber alignment. Gene expression is also
positively influenced by PMS. For instance, collagen type I expression under dynamic
loading is three times higher than static culture in 2 week(Butler, Hunter et al. 2009).
Although cyclic stretching has been proven to be an effective way to stimulate the
engineered tendon/ligament culture, the optimal stimulation pattern is still unknown.
Nirmalanandhan et al. revealed that a 2.4% strain cycle consisting of 3000 cycles per
day, produced the best linear stiffness in rabbit MSC seeded in type I collagen sponge
(Nirmalanandhan, Shearn et al. 2008). However from the perspective of Butler et al.,
the stimulation pattern should be adjusted based on maturation of the engineered
tendon/ligament, with higher dose loading applied at the later stage of tissue culture
(Butler, Hunter et al. 2009).
85
Tabl
e 1.
1 S
tudi
es u
sing
bio
reac
tor
syst
ems
for
tend
on/l
igam
ent e
ngin
eeri
ng
Fir
st a
utho
r B
iore
acto
r ty
pe
(Com
pany
) P
aram
eter
s of
m
echa
nica
l st
imul
atio
n
Sca
ffol
d m
ater
ial
(d
imen
sion
s)
Cel
l sou
rce
Eff
ect
Ref
eren
ce
Alt
man
(20
02)
Cus
tom
-mad
e st
ep
mot
or
bior
eact
or
wit
h en
viro
nmen
tal
cham
ber
Cyc
lic
stre
tchi
ng
2mm
, 90
o ro
tati
on,
0.01
67H
z,
21
days
Col
lage
n ty
pe
I ge
ls,
Bom
byxm
ori
silk
wor
m s
ilk
fibe
r m
atri
ces
(len
gth:
30 m
m)
Hum
an b
one
mar
row
st
rom
a ce
lls
(hB
MS
C)
Elo
ngat
ion
of
hBM
SC
, cr
oss-
sect
ion
cell
de
nsit
y
(Alt
man
, L
u et
al.
2002
)
Junc
osa-
Mel
vin
(200
6)
Cus
tom
-mad
e pn
eum
atic
cy
lind
er
bior
eact
or
wit
h LV
DT
fo
r di
spla
cem
ent
mon
itor
ing
Cyc
lic
stre
tchi
ng 2
.4%
st
rain
, 0.
0033
Hz,
8h
/day
fo
r 2
wee
ks
Type
I
coll
agen
sp
onge
(2
3
0.8
mm 9
0.
8mm 3
0.
1mm
)
Rab
bit M
SC
M
axim
um
forc
e
, li
near
st
iffn
ess
,
max
imum
st
ress
li
near
mod
ulus
(Jun
cosa
-Mel
vin,
S
hear
n et
al.
2006
)
Web
b (2
006)
C
usto
m-m
ade
step
m
otor
bio
reac
tor
Cyc
lic
stre
tchi
ng 1
0%
stra
in,
0.25
Hz,
8h
/day
fo
r 7d
ays
Pol
yure
than
e co
nstr
uct
(20 1
0 2
mm
)
Hum
an
trac
heal
fi
brob
last
Type
I
coll
agen
,
TG
F
-1
, C
TG
F,
Ela
stin
, al
pha
I ,
Pro
coll
agen
,
Fib
rone
ctin
,
MM
P-1
,
elas
tic
mod
ulus
(Web
b,
Hit
chco
ck
et a
l. 20
06)

86
Tabl
e 1.
2 S
tudi
es u
sing
bio
reac
tor
syst
ems
for
tend
on/l
igam
ent e
ngin
eeri
ng
And
rojn
a (2
007)
C
usto
m-m
ade
bior
eact
or
wit
h lo
ad-d
ispl
acem
ent
mea
sure
sys
tem
Cyc
lic
stre
tchi
ng
9%
stra
in,
twic
e da
ily
for
30m
in e
ach
peri
od s
epar
ated
by
8h r
est f
or 2
wee
ks
Sm
all-
inte
stin
e su
bmuc
osa
(3cm
5c
m 9
5
m)
Dog
te
nocy
te
Cel
l de
nsit
y
, st
iffn
ess
(A
ndro
jna,
Spr
agg
et a
l. 20
07)
Nir
mal
anan
dhan
(2
008)
C
usto
m-m
ade
pneu
mat
ic
cyli
nder
bi
orea
ctor
Cyc
lic
stre
tchi
ng
2.4%
an
d 1.
2%
stra
in,
1Hz,
st
imul
atio
n pe
riod
: 8h
/day
, 10
0 an
d 30
00 c
ycle
s/da
y fo
r 12
day
s
Type
I
coll
agen
sp
onge
R
abbi
t il
iac
cres
t M
SC
The
st
imul
atio
n pa
tter
n of
2.
4%
stra
in,
3000
cy
cles
/day
, 1H
z ha
s be
st e
ffec
t on
in
crea
sing
st
iffn
ess.
(Nir
mal
anan
dhan
, S
hear
n et
al.
2008
)
Nir
mal
anan
dhan
(2
008)
C
usto
m-m
ade
pneu
mat
ic
cyli
nder
bi
orea
ctor
Cyc
lic
stre
tchi
ng
2.4%
st
rain
, 0.
0033
Hz,
8h
/day
fo
r 12
day
s
Type
I
puri
fied
bo
vine
co
llag
en
gel,
type
I
coll
agen
sp
onge
s (l
engt
h: 1
1mm
and
51
mm
)
Rab
bit
MS
C
stif
fnes
s
(Nir
mal
anan
dhan
, R
ao e
t al.
2008
)
Ngu
yen
(200
9)
Cus
tom
-mad
e li
near
ac
tuat
or
bior
eact
or
wit
h lo
ad
cell
fo
r fo
rce
mea
sure
men
t
Pre
load
ed
wit
h 0.
05N
, cy
clic
st
retc
hing
10
%,
0.5H
z,
2h
stim
ulat
ion-
2h
rest
-2h
stim
ulat
ion-
18h
rest
fo
r 5
days
Por
cine
sm
all
inte
stin
e su
bmuc
osa
(len
gth:
2c
m
and
wid
th: 1
cm)
Rab
bit
MC
L
fibr
obla
st
Impr
oved
fi
ber
orie
ntat
ion,
fi
ber
angu
lar
disp
ersi
on
,
bett
er
orga
nize
d co
llag
en
fibe
r, el
onga
ted
cell
m
orph
olog
y,
(Ngu
yen,
Lia
ng e
t al
. 200
9)

87
Tabl
e 1.
3 S
tudi
es u
sing
bio
reac
tor
syst
ems
for
tend
on/l
igam
ent e
ngin
eeri
ng
Abo
usle
iman
(2
009)
C
usto
m-m
ade
line
ar
actu
ator
bi
orea
ctor
wit
h m
ediu
m
circ
ulat
ion
syst
em
Cyc
lic
stre
tchi
ng
2%
stra
in,
1h/d
ay,
0.01
67H
z fo
r 1
and
2 w
eeks
.
Hum
an u
mbi
lica
l vei
ns
(wal
l th
ickn
ess:
0.
75m
m,
oute
r di
amet
er:
6.75
0.
25m
m,
leng
th:
8.5c
m )
Wis
tar
Rat
bo
ne
mar
row
M
SC
bett
er c
ell
dist
ribu
tion
, m
ore
elon
gate
d ce
ll
mor
phol
ogy,
C
ell
prol
ifer
atio
n
, ul
tim
ate
stre
ss
, el
asti
c m
odul
us
(Abo
usle
iman
, R
eyes
et
al
. 20
09)
But
ler
2009
) C
usto
m-m
ade
pneu
mat
ic
cyli
nder
bi
orea
ctor
wit
h LV
DT
fo
r di
spla
cem
ent
mon
itor
ing
Cyc
lic
stre
tchi
ng 2
.4%
st
rain
, 0.
0033
Hz,
8h
/day
for
2 w
eeks
Type
I c
olla
gen
spon
ge
(94%
por
e vo
lum
e; 6
2 m
m
mea
n po
re
diam
eter
)
Mou
se
Mes
ench
ym
al
Ste
m
Cel
l
Type
I
coll
agen
,
line
ar s
tiff
ness
(B
utle
r, H
unte
r et
al.
2009
)
Che
n(20
10)
C
usto
m-m
ade
step
m
otor
bi
orea
ctor
Cyc
lic
stre
tchi
ng 1
0%
stra
in,
2h/d
ay,
1Hz
for
14 d
ays
Kni
tted
si
lk-c
olla
gen
spon
ge s
caff
old
(5cm
0.
5cm 0
.2cm
)
Hum
an
embr
yoni
c st
em c
ell
Col
lage
n I
, C
olla
gen
III
, E
pha4
,
Scx
,
Sox
9
, M
yosi
n
, In
tgri
n
1
, In
tgri
n
2
, In
tgri
n
1
,
Col
lage
n co
nten
t
, C
olla
gen
diam
eter
,
bett
er c
ell a
lign
men
t
(Che
n,
Yin
et
al
. 201
0)
Dor
oski
(2
010)
C
usto
m-m
ade
Lin
ear
mot
or
bior
eact
or
Cyc
lic
stre
tchi
ng 1
0%
stra
in (
5% o
ffse
t, 5%
am
plit
ude)
, 1H
z,
3h/d
ay,
1 7
14 a
nd 2
1 da
ys
Pol
y (e
thyl
ene
glyc
ol)-
base
d hy
drog
el
mat
eria
l ol
igo
(pol
y(et
hyle
ne
glyc
ol)
fum
arat
e)
(12.
5
9.5
1.6m
m)
MS
Cs
(PT-
2510
; L
onza
)
Col
lage
n I
, C
olla
gen
III
,
TN
C
, Te
nasc
in-C
(Dor
oski
, L
even
ston
et a
l. 20
10)

88
Tabl
e 1.
4 S
tudi
es u
sing
bio
reac
tor
syst
ems
for
tend
on/l
igam
ent e
ngin
eeri
ng
Sab
er (
2010
)
Lig
agen
L
30-4
C
(Tis
sue
Gro
wth
Te
chno
logi
es)
Cyc
lic
stre
tchi
ng
1.25
N,
1 cy
cle/
min
, 1h
/day
for
5 d
ays
Ace
llul
ar
rabb
it
hind
paw
te
ndon
(l
engt
h: 5
cm)
Rab
bit
teno
cyte
U
ltim
ate
tens
ile
stre
ss
(cl
osed
to
fl
eshl
y ha
rves
ted
tend
on)
, el
asti
c m
odul
us
(Sab
er,
Zha
ng
et a
l. 20
10)
Issa
(20
11)
C
usto
m-m
ade
bior
eact
or
Cyc
lic
stre
tchi
ng
2%
stra
in,
1h/d
ay,
0.01
67H
z fo
r 1
and
2 w
eeks
.
Hum
an u
mbi
lica
l ve
ins
(wal
l th
ickn
ess:
0.
41m
m)
wit
h W
hart
on’s
je
lly
mat
rix
as c
entr
al p
orti
on (
tota
l th
ickn
ess:
0.7
5mm
)
Wis
tar
rat
bone
m
arro
w
MS
C
Cel
l pr
olif
erat
ion
, te
nsil
e st
reng
th
(I
ssa,
E
ngeb
rets
on
et
al. 2
011)
Woo
n (2
011)
L
igag
en
L30
-1C
&
Lig
agen
L
30-4
C
(Tis
sue
Gro
wth
Te
chno
logi
es)
L30
-1C
: dy
nam
ic
load
ing,
10N
, 1h
/day
, 0.
0167
Hz
for
5 da
ys
L30
-4C
: dy
nam
ic
load
ing,
0.
625N
1.
25N
2.
5N,
1h/d
ay,
0.01
67 H
z fo
r 3
5 an
d 8
days
Ace
llul
ar h
uman
fle
xor
tend
on s
caff
olds
H
uman
D
erm
al
fibr
obla
st
Ult
imat
e te
nsil
e st
ress
, e
last
ic m
odul
us
(W
oon,
K
raus
et
al.
2011
)

89
1.2.6 Conclusion
The goal of tendon/ligament bioreactor design should be to create a construct with
similar microstructure and mechanical properties, as native tissue, using bioscaffolds
and autologous tenocytes. However, a tendon-like uni-orientated collagen structure
cannot be achieved without a mechanical stimulus within the culture environment.
Traditional culture techniques do not provide this PMS. The use of a bioreactor system
is able to bridge the gap between in vitro and in vivo systems by creating suitable
biochemical and biomechanical environments. Although it is clearly very difficult to
reproduce the in vivo microenvironmental conditions exactly within the bioreactor, the
goal is to mimic the in vivo biomechanical condition as closely as possible. The essential
components of a tendon/ligament engineering bioreactor are the actuating system and
culture chamber, which are responsible for the PMS and providing sterilized
environment for tissue culture. For better manipulation of the culture environment,
accurate stimulation and more precise reporting of mechanical maturation of engineered
tendon/ligament, an environmental control system, medium circulation system, monitor
and feedback system need to be included. As bioreactors nowadays are becoming more
focused on pre-clinical research, using these clinically in the future still presents a
substantial challenge. Tissue engineering requires substantial system optimization to
achieve a reproducible functional engineered tendon/ligament consistently. This process
is difficult and expensive in animal models, let alone in humans. However, there are
additional problems in moving from small animal models to humans related to the
physical size of human tissue samples. Issues of nutrient transport, cell source and
spatial heterogeneity of scaffold properties and cell stimulation become more prominent
in these larger tissues.

90
However, all the evidence suggests that by using bioreactors as described above it will
be possible to successfully produce engineered tendons and ligaments. When this occurs
we will be able to manufacture basic multi-chambers bioreactors with accurate and
appropriate patterns of PMS, which are affordable, and have low maintenance.
As one of the key elements of tissue engineering, biomechanical signal has been
introduced to generate engineered tendon for over a decade, and cyclic tensile strain is
commonly used. Positive effects, as summarized as figure 1.1, have been reported based
on various loading regimes with tensile strain ranged from 2% to 10% associated with
different frequencies and loading duration. These studies have demonstrated that
mechanical loading can improve cell proliferation, cell morphology, collagen I synthesis
and mechanical properties of engineered tendon. However, most of the studies only
examine one strain; the comparison of the effect caused by different mechanical strains
is poorly studied. The precise physiological levels of strain, frequency and duration to
affect such a response are not well understood.
1.3 Hypothesis and aims
1.3.1 Hypothesis
1.3.1.1 Overall hypothesis
I hypothesize that the biomechanical environment is essential for tendon
homeostasis. Further, the application of different mechanical stimulation on tendon

91
in a bioreactor system can induce different pathological changes and that the
biomechanical environment can help manipulate the repair mechanism of tendon
tissue.
1.3.1.2 Specific hypothesis
i. Different mechanical environments in a bioreactor have different impact on
tissue.
ii. There exists a mechanical strain regime that will maintain tendon homeostasis.
iii. There exists a mechanical strain regime that will lead to adverse changes in a
tendon
iv. A strain regime can be found which restores the functional and histological
properties of a tendon.
.
1.3.2 Aims
i. To design and construct a bioreactor system that is able to provide a
programmable biomechanical environment for in vitro tendon culture. (Study I,
Chapter 3)
ii. To evaluate the effect of different mechanical stimulation on tendon
homeostasis in a bioreactor system. (Study II, Chapter 4)

92
iii. To evaluate if the mechanical stimulation optimized in the previous study is
able to activate the self-repair mechanism to repair the tendon degeneration
caused by loading deprivation culture. (Study III, Chapter 5)

93
Chapter 2
Materials & Methods

94
CHAPTER 2 MATERIALS AND METHODS
There are a number of histological and biochemical analysis used throughout this thesis.
This chapter presents these techniques and methodology.
2.1 Material
All chemical reagents used during the course of this experiment were obtained through
the following manufacturers.
2.1.1 Tissue Culture reagents
REAGENT MANUFACTURER/SUPPLIER
DMEM/F-12 Gibco Life Technologies, USA
L-Glutamine 200mM 100X Gibco Life Technologies, USA
FBS (Fetal Bovine Serum) Invitrogen, Australia
Pen-Strep (Penicillin-streptomycin) Gibco Life Technologies, USA
Gentamicin Invitrogen, Auckland, New Zealand

95
2.1.2 Chemical reagents
REAGENT MANUFACTURER/SUPPLIER
Agarose LE, analytical grade Promega Corporation, USA
Baxter water Baxter Healthcare Pty.Ltd., Australia
Chloroform MERCK, Germany
Ethanol Hurst Scientific Pty. Ltd., Australia
Ethanol (molecular grade) Sigma-Aldrich, USA
Formaldehyde Sigma-Aldrich, USA
Paraformaldehyde (PFA) MERCK, Germany
TRIzol® reagent Invitrogen, Australia
Xylene Sigma-Aldrich, USA
Tissue embedding medium (Paraplast®
Regular) Paraffin wax Sigma-Aldrich, USA
Haematoxylin Sigma-Aldrich, USA
Eosin Sigma-Aldrich, USA
Diethyl pyrocarbonate Sigma-Aldrich, USA
proteinase K Qiagen, Germany

96
DPX Mountant Sigma-Aldrich, USA
2.1.3 Molecular products
PRODUCT MANUFACTURER/SUPPLIER
1kb DNA ladder Promega Corporation, USA
6x blue/orange loading dye Promega Corporation, USA
100bp DNA ladder Promega Corporation, USA
dNTPs (5nM) Promega Corporation, USA
Go-Taq® Green Master Mix 2x Promega Corporation, USA
M-MLV RT RNase (H-) Promega Corporation, USA
Oligo dT (100mM) Promega Corporation, USA
RT-PCR buffer (5x M-MLV RT) Promega Corporation, USA
SYBR® Safe DNA gel stain Invitrogen, USA
iQ™ SYBR® Green Supermix Bio-Rad Laboratories, USA
2.1.4 Oligonucleotide Primers
All oligonucleotide primers used in real-time polymerase chain reaction (PCR) were

97
purchased from GeneWorks Pty. Ltd. (Australia). The primers were reconstituted in
DEPC water to a concentration of 100M and stored at -20℃ until required.
PRIMER SEQUENCE (5’ to 3’) PRODUCT SIZE
COL1A2 Forward TTGCCCTTCCTTGATATTGC 241
COL1A2 Reverse CCTCTTTCGCCCACAATTTA
COL3A1 Forward AAGCCCCAGCAGAAAATTG 160
COL3A1 Reverse TGGTGGAACAGCAAAAATCA
MMP1 Forward GGCTCAGTTCGTCCTCACTC 246
MMP1 Reverse CAGGTCCATCAAAGGGAGAA
MMP3 Forward GCTTTGCTCAGCCTATCCAC 189
MMP3 Reverse ACCTCCAAGCCAAGGAACTT
MMP12 Forward ATGCCAGGGAGACCAGTATG 248
MMP12 Reverse AAAGCATGGGCTATGACACC
TIMP1 Forward CTTGTCATCAGGGCCAAGTT 213
TIMP1 Reverse TCCAGCGATGAGAAACTCCT
TIMP2 Forward GTGGACTCTGGGAACGACAT 223

98
TIMP2 Reverse AGTCACAGAGCGTGATGTGC
TGFB Forward TGCTTCAGCTCCACAGAGAA 181
TGFBReverse CCTTGCTGTACTGGGTGTCC
36B4 Forward ACCCAAAATGCTTCATCGTG 150
36B4 Reverse CAGGGTTGTTTTCCAGATGC
2.1.5 Antibodies
PRODUCT MANUFACTURER/SUPPLIER
Monoclonal anti-collagen type III Sigma-Aldrich, USA
Biotinylated goat anti-mouse IgG Sigma-Aldrich, USA
2.1.6 Commercial kits
PRODUCT MANUFACTURER/SUPP
LIER
RNeasy® minikit(250) QIAGEN, Germany
Liquid DAB+ Substrate Chromogen System DAKO, Denmark

99
In Situ Cell Death Detection Kit, POD Roche, USA
2.1.7 Other materials and equipment
MATERIAL/EQUIPMENT MANUFACTURER/SUPPLIER
AND EK-300i scientific weighing scale
A&D Mercury Pty. Ltd., China
Centrifuge 5810R Eppendorf, Germany
Corning high speed microcentrifuge Corning Incorporated, USA
Corning mini microcentrifuge Corning Incorporated, USA
CentriStarTM Cap centrifuge tubes:15ml and 50ml
Corning Incorporated, Mexico
Easypet® pipette dispenser Eppendorf, Germany
Forma Series II water jacker CO2 incubator
Thremo Scientific, Australia
Grant W-14 water bath Grant Instruments, UK
Homo-polymer, boil-proof microtubes: 0.6ml, 1.5ml and 2.0ml
Axygen® Scientific, USA
Hot plate model 209-1 IEC Australia Industrial Equipment & Control Pty. Ltd., Australia

100
KaterMaster catering foil Bunzl Limited, Australia
Laborlux S microscope Leitz, Germany
LabServTM incubator Biolabs (Aust) Pty. Ltd., Australia
Olympus CKX41 inverted microscope
Olympus Corporation, Japan
Nikon digital sight DS-5Mc Nikon Corporation, Japan
Nikon Eclopse E200 reflective microscope
Nikon Corporation, Japan
Parafilm “M” ® laboratory film Parafilm, USA
PCR-tube 0.5ml, thin wall Eppendorf, Germany
pH211 microprocessor pH meter Hanna Instruments, Australia
Ratek vortex mixer Rowe Scientific Pty. Ltd., Australia
Revco Elite Plus -80℃ freezer Thremo-Fisher Scientific, USA
Serological pipette: 2, 5, 10ml Sarstedt, France
SmartSpecTM 3000 Bio-Rad Laboratories, USA
SmartSpecTM 3000 cuvette quartz Bio-Rad Laboratories, USA
Tissue culture dish 100x20 mm Sarstedt, USA
Tissue culture flask: 25cm2 and Corning Incorporated, USA

101
75cm2
Leica TP 1020 tissue processor Leica, Germany
Sratfrost® microscope slides Knittel-Glaser, Germany
Superfrost ultra plus® slide Thremo Scientific, Australia
Microscope cover glass 22x50mm Hurst scientific Pty. Ltd., Australia
Leica 819 low profile microtome Blades
Leica, Germany
Jung Biocut 2035 Leica, Germany
Transferpipette 3.5ml Sarstedt, Germany
Veriti 96 well thermal cycler Applied BiosystemsTM, Singapore
CFX ConnectTM Real-time PCR detection system
Bio-Rad Laboratories, USA
Hard-Shell® PCR plates 96 well WHT/CLR
Bio-Rad Laboratories, USA
Labnet ExcelTM single electronic pipette
Labnet International, Inc.
2.1.8 Software
SOFTWARE MANUFACTURER/SUPPLIER

102
GraphPad Prism GraphPad Software Inc., USA
Image J National Institutes of Health, USA
NIS-C Elements Nikon Instruments Inc., USA
NIS-Elements BR 3.2 64bit Nikon Instruments Inc., USA
Solidworks 3D CAD Design Software
Solidworks Corp., USA
2.1.9 General solution
All general solution and buffers were prepared using MilliQ double distilled water,
unless otherwise indicated.
SOLUTION COMPOSITION
PBS 10x PBS stock solution:
30mM NaH2PO4, 70mM Na2HPO4, 1.3M NaCl
dissolved in MilliQ H2O
1x PBS pH 7.4
Dilution of 10x PBS with MilliQ H2O and adjusted to
pH 7.4

103
Paraformaldehyde 4% paraformaldehyde:
40g of Paraformaldehyde dissolved in prewarmed
0.75L MilliQ H2O ~ 60°C with stirring. Cover with foil
and stir with low heat, add 5-10 drops of 10M NaOH till
the solution become clear. Remove from heat and add
100mL 10x PBS and adjust pH to 7.4. Filter solution
through a 0.45m membrane filter before aliquoting and
storing at -20℃.
Hematoxylin working
solution (Harris)
Hematoxylin solution (Harris): 100g Postassium or
ammonium heat dissolves in 1L MilliQ H2O.
Add 50 ml of 10% alcoholic hematoxylin sol. and
heat to boil for 1min. Remove from heat and slowly add
2.5 g of mercuric oxide (red). Heat to the solution and
until it becomes dark purple color. Cool the solution in
cold water bath and add 20 ml of glacial acetic acid
(concentrated). Filter before use.
Eosin working solution Eosin stock solution: 1g Eosin Y dissolved in 100ml
MilliQ H2O.
Phloxine stock solution: 1g Phloxine B dissolved in

104
100ml MilliQ H2O
Eosin-Phloxine B working solution: Combine 100ml
Eosin stock solution, 10ml Phloxine stock solution, 780ml
95%[v/v] Ethanol and 4ml Glacial acetic acid
2.2 Tissue culture procedures
2.2.1 Isolation of rabbit Achilles tendon
Female New Zealand White rabbits (Oryctolagus cuniculus) aged between 14 and 15
weeks with body weight 3-5kg were sacrificed. The lower legs were wetted and
sterilized by 70%[v/v] ethanol. A transection was made between the Achilles tendon and
tibia to calcaneus using a sterilized scalpel. The Achilles tendon was then completely
exposed. A full length Achilles tendon was removed with part of the calcaneus and
approximately 1cm of the soleus muscle.
2.2.2 Culture of rabbit Achilles tendon
The Achilles tendons were briefly rinsed in 70%[v/v] ethanol and temporally stored in
serum-free DMEM-F12 supplemented with 50µg/ml Gentamicin after isolation from
rabbit. Before the tissue culture procedure, the Achilles tendons were washed in 40ml
1x PBS three times. In the loading deprivation culture group, tendons with muscle and
calcaneus were cultured in a T25 culture flask. In the dynamic culture group, the

105
Achilles tendons were cultured with muscle and calcaneus fixed in the bioreactor using
tissue clamps and hook. The whole culture procedure was processed in a standard
incubator at 37℃ and 5% CO2, and the culture media was DMEM/F-12 supplemented
with 10% fetal bovine serum and 50 µg/mL Gentamicin and changed every three days.
2.3 RNA extraction
2.3.1 Extraction of total RNA
Approximately 200mg of tendon tissue was dissected from the mid-substance of the
Achilles tendon, then washed with ice-cold 1x PBS for three times. The tissue was
placed into 5ml tube and snap frozen using liquid nitrogen and pulverized using mortar
and pestle. The tissue debris was then lysed using 1ml Trizol and homogenized by
pipetting in several times followed by 5 minutes incubation at room temperature. The
lysate was collected in a microcentrifuge tube, shaken by hand with 200m chloroform
for 15 seconds and incubated at room temperature for 2-3 minutes. The mixture was
spin at 12,000 × g for 15mins at 4℃, the RNA was separated in the aqueous phase. The
supernatant was transfer to a microcentrifuge tube, vortex mixed thoroughly with 1
volume of 70%[v/v] ethanol and transferred up to 700L to the spin cartridge (RNeasy®
minikit), and centrifuge at 12,000 × g for 15 seconds at room temperature. This
procedure was repeated several times till the entire samples has been processed. The
spin cartridge was then centrifuged at 12,000 × g for 15 seconds at room temperature
with 700L Wash Buffer I (RNeasy® minikit), and the flow-through and collection tube

106
were discarded. 500L Wash Buffer II (RNeasy® minikit) with ethanol was added into
the cartridge and centrifuged at 12,000 × g for 15 seconds at room temperature, and the
flow-through was discarded, this procedure was repeated once. The cartridge was
centrifuged at 12,000 × g for 1-2 minutes at room temperature to dry the membrane
with bound RNA; the collection tube was then discarded. Add 30–100L RNase-free
water to the center of the spin cartridge, incubated at room temperature for 1 minute
followed by 2 minutes centrifuge at ≥12,000 × g at room temperature to elute the RNA
from the membrane into the recovery tube. The purified RNA was stored at -80℃ until
required.
2.3.2 RNA concentration reading
200L of 1/50 dilution of purified RNA with Baxter water was prepared. The
SmartSpec™ 3000 (Bio-Rad) was blanked using 200L RNase-free water first, and the
samples was then injected into the Quartz Microvolume Cuvette, and read by the
Spectrophotometer. The absorbance ratio (A260/A280) provides an estimation of the
purity of the RNA. Generally, 1.8-2.0 is the acceptable range, where 2.0 represent
“pure”. All the purified RNA used in this study has A260/A280 ratio above 1.8.

107
2.4 Quantitative polymerase chain reaction (QPCR)
2.4.1 Reverse transcription of mRNA
A total 300ng of RNA was used to convert to equal amounts of cDNA using the
following protocol:
RNA 300ng
Oligo dT 0.25L
dNTP (5mM) 2L
RNase-free water final volume of 15L
Total volume 15L
The samples were incubated at 75℃ for 3 minutes, and then the following reagents
mixture was added:
5x MMLV RT-PCR Buffer 4L
RNase 0.5L
MMLV RT RNase 0.5L
Total volume 5L

108
The samples were then incubated at 42℃ for 60 minutes, followed by incubation at 92℃
for 10 minutes. The cDNA was stored at -20℃ until required
2.4.2 Quantitative polymerase chain reaction (QPCR)
Each of the QPCR reaction was performed using CFX ConnectTM Real-time PCR
detection system (Bio-Rad Laboratories, USA) and prepared according to the following
protocol:
iQ™ SYBR® Green Supermix 5L
cDNA 1L
Primer Forward (20M) 0.5L
Primer Reverse (20M) 0.5L
RNA free water 3L
Total volume 10L
The QPCR reaction primers were specified in 2.1.4 Oligonucleotide primer. The QPCR
conditions were carried out as per manufacturer’s instructions, each with 40 cycles.
Housing keeping gene 36B4 was used as internal control. A cycle threshold (Ct) value

109
was obtained from each reaction. The comparative 2-△△Ct method was used to calculate
the relative expression of each target gene. The RT-PCR were repeated in 3 individual
samples from each group and performed in triplicate in every RT-PCR setting. The
average expression levels were calculated and used for comparison.
2.5 General histology
2.5.1 Tissue preparation
The Achilles tendon was first washed by 1x PBS for three time after culture, and then
fixed using 35mL 4% PFA for overnight at 4℃ followed by 45 minutes wash with 1x
PBS. The samples were then placed in tissue cassettes, and put into the Leica TP 1020
processor, processed using the following protocol:
CHEMICAL CONDITION TIME
70%[v/v] Ethanol Vacuum 2 h
80%[v/v] Ethanol Vacuum 2 h
95%[v/v] Ethanol Vacuum 2 h
100%[v/v] Ethanol Vacuum 2 h
100%[v/v] Ethanol Vacuum 2 h
Xylene Vacuum 3 h

110
Xylene Vacuum 3 h
Xylene Vacuum 3 h
Paraffin Wax Vacuum 2h
Paraffin Wax Vacuum 2h
Paraffin Wax Vacuum 2h
TOTAL: 25h
After processing, the sample cassettes were removed from the tissue processor, and the
samples were taken out from the cassettes. A little wax was placed in the embedded tray,
allowing it to slightly cool. The sample then was placed down, and the tray was filled
with wax. The sample and tray was first cool down on the cooling station for 20 minutes
and then the embedded block was removed from the tray.
2.5.2 Haematoxylin and Eosin Staining
The paraffin-embedded tissue sample were cut into 5m thick section, placed onto the
microscope slider and processed by following protocol:
CHEMICAL TIME

111
Xylene 5min
Xylene 5min
100%[v/v] Ethanol 2min
90%[v/v] Ethanol 2min
70%[v/v] Ethanol 2min
Distill water 2min
Haematoxylin 2min
Washed in running water till clean
Lithium Carbonate 1min
Distill Water 1min
Eosin 30s
Washed in running water till clean
70%[v/v] Ethanol 2min
90%[v/v] Ethanol 2min
100%[v/v] Ethanol 2min
100%[v/v] Ethanol 3min
Xylene 5min

112
Xylene 5min
Mount in DPX
2.5.3 Histological scoring
Three tissue sections from different depth of the paraffin block of each sample were
selected. The H&E stained sections were viewed under the microscope, with at least 5
fields (20x magnifications) of each slice randomly selected for histological assessment
by 3 individuals blinded to group allocation. Four parameters, including fiber
arrangement, fiber structure cell roundness and cell density, were evaluated using a 4
point scoring scale from 0 to 3, where 0 being normal and 3 being severely abnormal.
The represented Figures of each score are shown as Figure 2.1 . The average score of
each parameter was calculated and used for comparison.

113
Figure 2.1 Histological scoring system modified from the study of Chen et al.(Chen, Yu et al. 2011)
2.6 Type III collagen immunohistochemistry
Paraffin-embedded sections were dewaxed and rehydrated using graded ethanol as
described in 2.5.2 Haematoxylin and Eosin Staining, and processed by the following
procedure.
CHEMICAL CONDITION TIME
3% hydrogen peroxide (H2O2)
Room temperature
10min
PBS 4min
5% fetal bovine serum in 1xPBS 30min
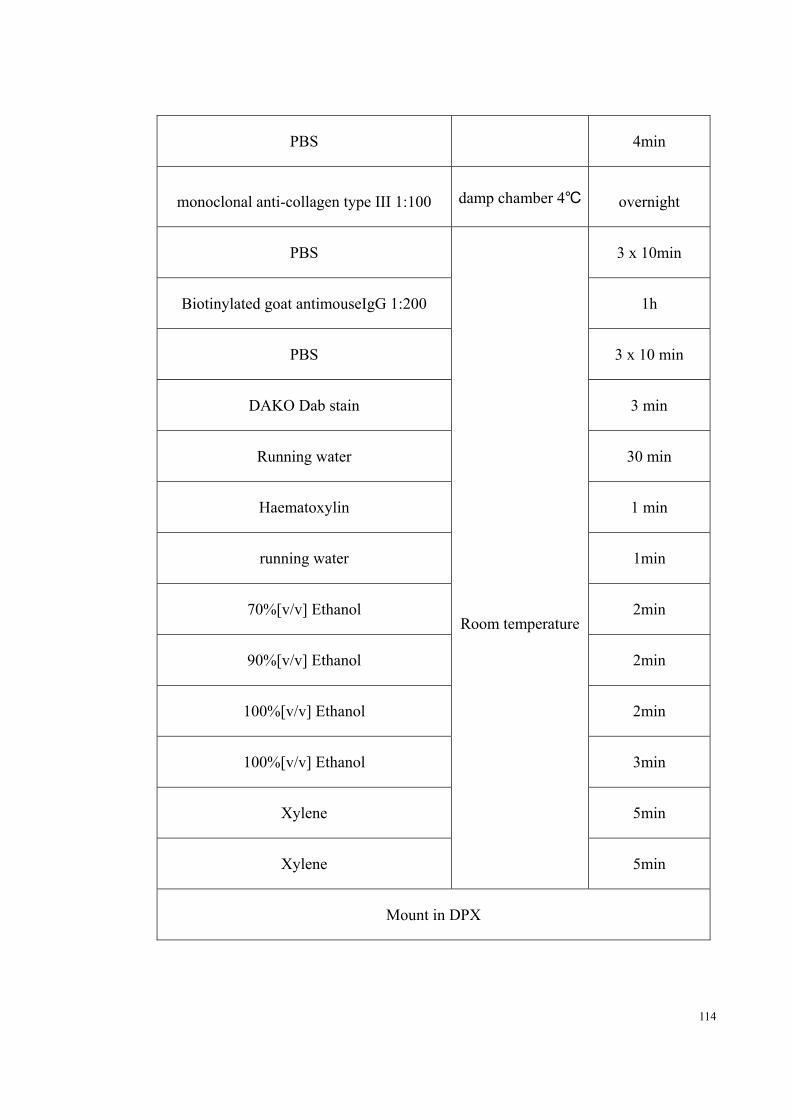
114
PBS 4min
monoclonal anti-collagen type III 1:100 damp chamber 4℃ overnight
PBS
Room temperature
3 x 10min
Biotinylated goat antimouseIgG 1:200 1h
PBS 3 x 10 min
DAKO Dab stain 3 min
Running water 30 min
Haematoxylin 1 min
running water 1min
70%[v/v] Ethanol 2min
90%[v/v] Ethanol 2min
100%[v/v] Ethanol 2min
100%[v/v] Ethanol 3min
Xylene 5min
Xylene 5min
Mount in DPX

115
2.7 TUNEL assay
Paraffin-embedded sections were dewaxed and rehydrated using graded ethanol as
described in 2.5.2 Haematoxylin and Eosin Staining, and processed by following
procedure.
CHEMICAL TIME
20g/mL proteinase K 15min
PBS 4min
TUNEL reaction mixture 1h
PBS 3 x 4min
convert-AP 30min
PBS 4min
DAKO Dab stain 5min
Running water 30 min
Haematoxylin 1 min
Running water 1min
70%[v/v] Ethanol 2min
90%[v/v] Ethanol 2min

116
100%[v/v] Ethanol 2min
100%[v/v] Ethanol 3min
Xylene 5min
Xylene 5min
Mount in DPX
Three tissue sections from different depth of the paraffin block of each sample were
selected. The TUNEL stained sections were viewed under the microscope, with at least
5 fields (20x magnifications) of each slice randomly selected for histological assessment
by 3 individuals blinded to group allocation. The positive staining cell and normal cell
were counted. The overall cell apoptotic rate was calculated and used for comparison.
2.8 Statistics analyses
All statistics have been analyzed using the prism 5.03 software package (GraphPad
Software 5.0, USA). Data are presented as mean ± SEM. Statistical analysis was
performed by one-way analysis of variance (ANOVA). P values less than 0.05 were
considered significant.

117
Chapter 3
Bioreactor Design and
Fabrication
This chapter is partly based on the paper
Programmable mechanical stimulation influences tendon homeostasis in a bioreactor system
Tao Wang, Zhen Lin, Robert E Day, Bruce Gardiner, Euphemie Landao-Bassonga, Jonas Rubenson, Thomas B. Kirk, David W. Smith, David G. Lloyd, Gerard Hardisty,
Allan Wang, Qiujian Zheng, Ming H Zheng
Biotechnology and Bioengineering
Volume 110, Issue 5, pages 1495-1507, May 2013

118
CHAPTER 3 (RESULTS): DESIGN AND CONSTRUCTION OF
BIOREACTOR
In this study we focus on the design of a tendon-engineering bioreactor, which is able to
provide programmable mechanical stimulation and a sterile culture environment. The
specific objective of the current study is to apply the design principles of mechanical
and electrical engineering to construct a functional bioreactor for tendon culture. To
fulfill this objective, a new bioreactor was designed using Solidworks 3D CAD software
(Solidworks Corp., Concord, MA), and constructed by an external mechanical vendor
under the supervision of the PhD candidate. This bioreactor system will assist in
evaluating the effect of various programmable mechanical stimulation regimes on
tendon homeostasis and repair in vitro, and ultimately enable optimization of the
mechanical stimulus for engineered tendon culture
3.1 Design criteria
To develop a functional bioreactor several design criteria were formulated:
1. The bioreactor must be able to process more than three independent culture
specimens simultaneously.
2. The system should have transparent covers, allowing visual monitoring.
3. The mechanical stimulation must be programmable.
4. The extension resolution should be better than ±100m with the load varying as

119
required to produce the set strain
5. The minimum expected load was 20N per sample, which is minimum 120N for the
bioreactor system.
6. The system must incorporate a means of holding the specimen ends to apply
stimulation without either damaging the specimen or allowing the construct to slip
relative to the grips.
7. The bioreactor must be able to operate inside a standard incubator, so all of the
electrical and mechanical components have to be able to function in a warm and
humid environment.
8. The parts in contact with the tissue culture should be autoclavable.
3.2 Material selection
In our design, the bioreactor is divided into two main components, the frame and the
culture chambers. The bioreactor is operated inside an incubator, and the culture
medium must be changed regularly. As this would require the operator to move the
bioreactor in and out of the incubator, the overall weight was a problem that needed to
be considered. For local sterilization, 70%[v/v] ethanol and UV treatment are
commonly used; therefore, both parts had to be chemical resistant and UV stable.
Moreover, the material used for culture chamber had to withstand a higher level of
sterilization such as autoclaving and also be resistant to humidity and chemical attack
from the culture medium.

120
In summary, the material used in the bioreactor had to meet the following requirements:
Good machinability.
Corrosion resistant (culture medium and warm humid air)
UV stable
Compatible with alcohol (up to 70%[v/v])
For the frame: as light as possible
For the culture chamber: compatible with autoclave sterilization
From the criteria listed above, a corrosion resistant metal would be preferred to other
materials due to its good machinability, high strength, UV stable and availability in the
market. We chose 6063 aluminum for the frame. Although its corrosion resistance is not
as good as stainless steel, it is relatively light compared to other metals. For the culture
chamber, AISI 316 stainless steel was chosen due to its good resistance to chemical
attack and humidity, moreover, it is suited to high pressure steam autoclaving, which is
the main sterilization method in our study.
3.3 Overall bioreactor design
After considering all the criteria and reviewing the published literature on both
custom-made and commercialized bioreactors, the design was finalized. The overall

121
dimensions of this bioreactor are 260mm×334mm×55mm, which is able to fit into a
standard incubator easily, and the overall weight is 6.9kg. Programmable mechanical
stimulation was generated with a single stepper motor and the motion transferred to the
cultured tissue through a ball-screw. This design is able to process dynamic cultures for
6 specimens simultaneously; and frequency, displacement and culture period can be
preprogrammed through a computer. Figure 3.1 shows the schematic illustration of the
bioreactor system, and Figure 3.2 shows the overview of the bioreactor.
Figure 3.2 Schematic illustration of the bioreactor system

122
Fig
ure
3.3
Des
ign
of 6
cha
mbe
rs b
iore
acto
r

123
Figure 3.4 Top view of bioreactor.

124
3.3.1 The Actuating System
The actuating system is the key of the whole bioreactor system, providing mechanical
stimulation to the cultured tissue. After reviewing the various designs of published
bioreactors, it was found that pneumatic actuators (Juncosa-Melvin, Shearn et al. 2006;
Nirmalanandhan, Rao et al. 2008; Nirmalanandhan, Shearn et al. 2008), linear
motors(Nguyen, Liang et al. 2009; Doroski, Levenston et al. 2010) and stepper
motor-ball screw combinations(Altman, Lu et al. 2002; Webb, Hitchcock et al. 2006;
Chen, Yin et al. 2010) are the most common actuating systems. Considering the position
accuracy and force output required in our study, a stepper motor-ball screw system was
chosen. There are two common types of stepper motors available in the market, 2-phase
and 5-phase stepper motors. A 2-phase motor only has 8 magnetic poles, whereas a 5
–phase motor has 10. The extra poles provide higher resolution, lower vibration, higher
acceleration and less synchronization loss than a 2-phase motor for little extra expense.
Therefore, a 5 phase stepper motor (PMC33A3, Oriental Motor, Japan) with 0.72°
resolution was selected for this bioreactor. To protect the stepper motor, an aluminum
cover was designed, which can eliminate the heat caused by the motor due to its good
heat conduction (Figure 3.3).
In order to transfer the rotation of the stepper motor to linear movement, a ball-screw is
necessary. Two common forms of ball-screw are commercially available, individual
ball-screws and guide actuator assemblies. Further machining and base design would be
required if a single ball-screw was chosen. The guide actuator consists of a
pre-assembled ball-screw, guide track and moving platform, which reduces the error

125
caused by further machining. Therefor the KR15 (THK, Japan) guide actuator was
chosen in our design. The lead of the KR15 is 1mm, and given the 500 steps per
revolution resolution of the stepper motor, this result in 1/500 or 2 micrometer per step
linear resolution.
3.3.2 The Culture Chamber
AISI 316 stainless steel was used for culture chamber construction due to its good
machinability and good resistance to chemical and high temperature. Multiple chambers
are necessary, considering that triplication is the minimum requirement for each
experiment. To balance the force distribution, the 6 chambers were uniformly located on
the base as shown as Figure 3.3. In order to simplify the operation, the culture chamber
was a separable individual design with 30mL capacity, shown as Figure 3.4. Each
culture chamber can be separated from the base and autoclaved. A removable glass
cover was designed to prevent contamination and enable better observation during the
culture period. A 1mm gap was reserved between the shaft of tissue clamps and the
glass cover, which allow the sufficient air exchange to the culture medium.

126
Figure 3.5 Design of culture chamber
3.3.3 Tissue Fixation
The method of tissue fixation is an important aspect of bioreactor designing needing
further consideration, as during mechanical stimulation in culture medium, sample
slippage can occur. Most bioreactor designs only have flat jawed tissue clamps that,
while suitable for flat membranes, cannot provide effective fixation with thick
cylindrical tissues such as tendon, leading to increased slippage. Therefore here two
fixation methods were designed for different sample types. The thin tissue or scaffold
can be clamped by the clamp cover and tightened using screws shown as figure (Figure
3.5A). Big and slippery tissues can be fixed by the tissue hook or tied to the tissue hook
using suture (Figure 3.5B).

127
Figure 3.6 Exploded view of the tissue clamps (A) and tissue hooks (B) assembly.
3.3.4 The Control System
The two main choices for control systems in other bioreactors are direct computer
control or a dedicated controller. Although programming a dedicated controller is more
complicated than using a computer, the dedicated controller is much more reliable.
Therefore, the selected control system was a programmable logic controller (MELSEC
FX1S, Mitsubishi, Japan) and a stepper motor driver (PMD03CA, Oriental Motor,

128
Japan). This driver is compatible with the PMC33A3 stepper motor. The motion control
program was written by the candidate himself using GX Developer (Mitsubishi, Japan),
and then downloaded to the PLC via RS232 cable. The PLC is able to provide
independent control on the stepper motor without any external requirements except
power.
3.4 Bioreactor setup
Before each experiment, the frame of the bioreactor was first sprayed using 70%[v/v]
ethanol and air dried in a laminar flow hood followed by sterilization under UV light for
30 minutes. The culture chambers and tissue clamps were double wrapped in paper bags
separately and autoclaved followed by drying in a 60℃ oven for 2 hours.
The bioreactor was placed in a standard incubator at 37℃and 5% CO2shown as Figure
3.6A. A 5-pin plug and cable connected the bioreactor to the control box outside of the
incubator through the standard hole at the back of the incubator (Figure 3.6B). Figure
3.7 illustrates the bioreactor setup in the incubator.

129
Figure 3.7 (A) Bioreactor in the incubator. (B) A five pin plug and cable connects the bioreactor to the control box through the hole at the back of the incubator.

130
Figure 3.8 illustration of bioreactor system setup in the incubator; the bioreactor is connected to the control box through a 5-core cable.
3.5 Discussion
In recent years, many studies of tendon engineering have been conducted using
custom-made bioreactor system, and the results have been promising(Juncosa-Melvin,
Shearn et al. 2006; Webb, Hitchcock et al. 2006; Androjna, Spragg et al. 2007;
Nirmalanandhan, Rao et al. 2008; Nirmalanandhan, Shearn et al. 2008; Abousleiman,
Reyes et al. 2009; Nguyen, Liang et al. 2009; Chen, Yin et al. 2010; Doroski, Levenston

131
et al. 2010; Saber, Zhang et al. 2010; Issa, Engebretson et al. 2011; Woon, Kraus et al.
2011). Several commercial tendon engineering bioreactors are available now, such as
Bose® ElectroForce® BioDynamic® and LigaGen system. To develop a functional
bioreactor system, several factors needed to be considered, including material,
mechanical design, tissue fixation method and operating environment. Compared with
commercial bioreactors in Bose and LigaGen, the current design has several advantages.
Firstly, our device can condition multiple samples simultaneously but separately. It can
be easily cleaned and sterilized in standard autoclaves. Being constructed from AISI 316
stainless steel it can go through many sterilization cycles without corrosion or
degradation. The device can produce a maximum force of 198N, which is sufficient
for most tendon scaffold and tendon tissue from small scale animals such as are
commonly used in tissue engineering research. Most commercial bioreactor systems
have problems in holding thick and slippery tissue during PMS, where as our novel
tissue hook design demonstrated excellent tissue fixation. Finally, the actuation system
is designed to operate inside a standard incubator. It is able to function in a humid
environment and the horizontal design is easy to move and will only occupy a small
space in the incubator. As the tendon’s main function is mechanical, this bioreactor
system is able to provide high-accuracy programmable mechanical stimulation (PMS),
enabling different loading regimes including displacement, frequency and stimulation
period. Once programmed it is able to operate reliably for weeks without computer
support.

132
Chapter 4
Programmable mechanical
stimulation influences tendon
homeostasis in a bioreactor
system
This chapter is partly based on the paper
Programmable mechanical stimulation influences tendon homeostasis in a bioreactor system
Tao Wang, Zhen Lin, Robert E Day, Bruce Gardiner, Euphemie Landao-Bassonga, Jonas Rubenson, Thomas B. Kirk, David W. Smith, David G. Lloyd, Gerard Hardisty,
Allan Wang, Qiujian Zheng, Ming H Zheng
Published in
Biotechnology and Bioengineering
Volume 110, Issue 5, pages 1495-1507, May 2013

133
CHAPTER4 (RESULTS): PROGRAMMABLE MECHANICAL
STIMULATION INFLUENCES TENDON HOMEOSTASIS IN A
BIOREACTOR SYSTEM
4.1 Abstract
Identification of functional programmable mechanical stimulation (PMS) on tendon not
only provides the insight of the tendon homeostasis under physical/pathological
conditions, but also guides a better engineering strategy for tendon regeneration. The
aims of the study are to design a bioreactor system with PMS to mimic the in vivo
loading conditions, and to define the impact of different cyclic tensile strains on tendon.
Rabbit Achilles tendons were loaded in the bioreactor with/without cyclic tensile
loading (0.25Hz for 8 hours/day, 0~9% for 6 days). Tendons without loading lost their
structural integrity, as evidenced by disorientated collagen fiber, increased type III
collagen expression, and increased cell apoptosis. Tendons with 3% of cyclic tensile
loading had moderate matrix deterioration and elevated expression levels of MMP-1, 3
and 12, whilst loading regime of 9% caused massive rupture of collagen bundles.
However, 6% of cyclic tensile strain was observed to maintain the structural integrity
and cellular function. Our data indicated that an optimal PMS exists to maintain tendon
homeostasis, although this is only a narrow range of tensile strain that can induce this
action. The suggested clinical impact of this study is that optimized eccentric training
program, based around targeting specific tendon strain, could be developed to achieve
beneficial effects on chronic tendinopathy management.

134
4.2 Introduction
Tissue engineering methods have great potential in the repair of damaged tendons.
However, these methods are still very much in their infancy, due to the lack of an
integrated understanding of the key conditions required for successful tendinopathy
intervention and repair. Recent tendon research has focused on the development of
tendon tissue engineering, with the aim of constructing a bio-substitute material for
tendon/ligament repair(Juncosa-Melvin, Shearn et al. 2006; Sahoo, Ouyang et al. 2006;
Webb, Hitchcock et al. 2006; Chen, Willers et al. 2007; Chen, Qi et al. 2008;
Nirmalanandhan, Rao et al. 2008; Saber, Zhang et al. 2010; Stoll, John et al. 2010;
Peach, James et al. 2012; Wang, Gardner et al. 2012; Woon, Farnebo et al. 2012); This
strategy involves an integration of technologies from cell biology, material science and
mechanical engineering.
Engineered bioreactor systems have been demonstrated to be effective tools for
constructing various biological tissues and organs such as muscle(Dennis, Smith et al.
2009), liver (Catapano, Patzer et al. 2010) and bone(Rauh, Milan et al. 2011; Rauh,
Milan et al. 2012). Several bioreactor systems have been developed specifically for
engineered tendon/ligament formation (Altman, Lu et al. 2002; Juncosa-Melvin, Shearn
et al. 2006; Webb, Hitchcock et al. 2006; Nirmalanandhan, Rao et al. 2008;
Nirmalanandhan, Shearn et al. 2008; Issa, Engebretson et al. 2011). Owing to force
transmission being the primary function role of tendon/ligament, mechanical
stimulation plays an essential role in the maintenance of tendon structure and

135
function(Hannafin, Arnoczky et al. 1995). Achilles tendons endure tensile loading in
vivo during daily physical activities. More complex structures, like the anterior cruciate
ligament, undergo both tensile and torsion loading during weight-bearing knee
flexion(Li, Defrate et al. 2005). To restore tendon function after injury, engineered
tendon constructs should be compatible with the in vivo environment both biologically
and mechanically(Calve, Lytle et al. 2010). Cell-scaffold constructs may be ‘exercised’
in an artificial mechanical environment before implantation. Indeed, programmable
mechanical stimulation (PMS) is considered to be one of the key components for
tendon/ligament forming bioreactor system (Altman, Lu et al. 2002; Juncosa-Melvin,
Shearn et al. 2006; Webb, Hitchcock et al. 2006; Nirmalanandhan, Rao et al. 2008;
Nirmalanandhan, Shearn et al. 2008; Issa, Engebretson et al. 2011). In this study, a
bioreactor system was designed to apply different cyclic tensile strain on isolated rabbit
Achilles tendon to establish an ex vivo model (see previous Chapter). The present aim is
to evaluate the effect of different mechanical stimulation on tendon tissue using this
developed bioreactor system.
The mechanical stimulation of engineered tendon constructs can result in different
structures. Non-loading of engineered neo-tendons results in loose, disorganized
matrices with reduced collagen quantity(Jiang, Liu et al. 2011). Static mechanical stress
has been shown to improve the fiber arrangement of engineered tendon constructs, but
did not produce a compacted tendon-like collagen structure in vitro(Chen, Willers et al.
2007). More promisingly, engineered tendon under cyclic tensile strain exhibited a
well-organized collagen fiber arrangement and structure and enhanced tensile strength,

136
suggesting that PMS is preferable for engineered tendon formation(Jiang, Liu et al.
2011). Even though the importance of providing PMS in tendon bioreactor has been
recognized previously (Brown and Carson ; Wang, Gardner et al. 2012), the optimal
cyclic tensile stimulation parameters have not been defined. It is generally assumed that
mimicking the dynamic mechanical loading created by daily physical activity will give
the optimal loading regime in tendon bioreactor system, but this is not yet proven.
The goals of this study were to investigate the effect of different mechanical stimulation
regimes on tendon integrity. I hypothesized that there is an optimum range of
mechanical stimulation required for tendon homeostasis and that under or
over-stimulation would reduce tendon integrity.
4.3 Material and Method
4.3.1 Tissue harvest and distribution
Achilles tendons were dissected from the hindlimbs of female New Zealand White
rabbits (Oryctolaguscuniculus) aged between 14 and 15 weeks with body weight 3-5kg.
The tissue was briefly rinsed in 70%[v/v] ethanol, washed in 1x phosphate buffered
saline (PBS) 3 times, and submerged in serum free Dulbecco’s modified Eagle’s
medium (DMEM)/F-12 (GIBCO, Invitrogen, Auckland, New Zealand) supplemented
with 50 µg/mL Gentamicin before further processing.

137
In total 24 Achilles tendons from 12 rabbits were allocated to 3 groups (Figure 4.1A): a
loading deprivation group (8), a dynamic culture (cyclic tensile strain) group (12) and a
control group (4). In the loading deprivation group, the Achilles tendons were cultured
in petri dish in growth medium (DMEM/F-12, 10% fetal bovine serum and 50 µg/mL
Gentamicin) without mechanical loading for either 6 days (4 tendons) or 2 weeks (4
tendons). In the dynamic strain group, the tendons were fixed on the tissue hooks using
suture and cultured under 3%, 6% or 9% cyclic tensile strain for 6 days (8 hours per day,
0.25Hz, Figure 4.1A). The stimulating cycle was as demonstrated in Figure 4.1B.This
regimen was based on previously published studies and data on rabbit activity levels
(West, Juncosa et al. 2004). The samples were sliced into two half, ones was fixed in 4%
paraformaldehyde overnight for histological analysis and the other half directly
subjected to RNA extraction. The control group was made up of 4 tendons directly
dissected from the animal and immediately fixed.
Figure 4.1 (A) Number and distribution of samples for each of the 12 rabbits. (B) Daily
mechanical stimulation pattern used in this study. (C) Mechanical input signal from the control
system

138
4.3.2 Histological preparation and assessment
Refer to 2.5
4.3.3 TUNEL assay
Refer to 2.7
4.3.4 Immunostaining for type III collagen
Refer to 2.6
4.3.5 Quantitative Real-time polymerase chain reaction (Q-PCR)
Refer to 2.3 and 2.4

139
Table 4.5 Rabbit specific primer sequences for the genes used.
Target Genes Forward (5’ to 3’) Reverse (3’ to 5’)
COL1A2 TTGCCCTTCCTTGATATTGC CCTCTTTCGCCCACAATTTA
COL3A1 AAGCCCCAGCAGAAAATTG TGGTGGAACAGCAAAAATCA
MMP1 GGCTCAGTTCGTCCTCACTC CAGGTCCATCAAAGGGAG
AA
MMP3 GCTTTGCTCAGCCTATCCAC ACCTCCAAGCCAAGGAACTT
MMP12 ATGCCAGGGAGACCAGTATG AAAGCATGGGCTATGACACC
36B4 ACCCAAAATGCTTCATCGTG CAGGGTTGTTTTCCAGATGC
4.3.6 Statistical analyses
Refer to 2.8
4.4 Results
4.4.1 Histological examination of tendon undergoing different cyclic tensile
strains
Compared to the native tendon (Figure 4.2 A), the collagen fiber arrangement changed
from parallel to moderately wavy after 6 days in the absence of applied loading. Cell
density was increased with slightly rounded nuclei. After 2 weeks without loading,
progressive extracellular matrix (ECM) disruption was observed. Collagen fiber

140
arrangement had no discernible pattern. Fragmentation of collagen fibers and rounded
tenocyte nuclei were observed microscopically (Figure 4.2C). Histological assessment
showed a time dependent increasing grading in fiber arrangement, fiber structure and
cell roundness, indicating progressive divergence from the native (control) tissue. Cell
density was also increased after 6 days without loading (Figure 4.2D-G).

141
Figure 4.2 Loading deprivation culture group. (A) Native Achilles tendon structure (day 0). (B) Achilles tendon histology after 6 days loading deprivation culture with minor wavy collagen structure. (C) Achilles tendon histology after 2 weeks culture displays disorganized collagen
structure with round tenocytes. (D) Fiber arrangement, scale of 0-3, 0 represents compacted and parallel, and 3 represents no identifiable pattern. (E) Fiber structure, scale of 0-3, 0 represents
continuous, long fiber, and 3 represents severely fragmented. (F) Cell roundness, scale of 0-3, 0 represents long spindle shape, and 3 represents severe rounding. (G) Cell density, scale of 0-3, 0
represents normal pattern, and 3 represents severely increase Result are expressed as the mean±S.D. (*p<0.05, **p<0.01, **p<0.001). Scale bar=50 micrometers.

142
In the dynamic culture (cyclic tensile strain) group, histology revealed that the 3% strain
stimulated group had slightly disrupted extracellular matrix structure and significant
cellular morphology changes (Figure 4.3B). These tendon samples graded 1.5 in fiber
arrangement, 1.1 in fiber structure and 1.3 in cell roundness (Figure 4.3E-G). However,
in the tendons that underwent 6% strain, the extracellular matrix orientation and cellular
morphology appeared normal (Figure 4.3A, C). That is, there was no statistical
difference of the histology scoring between the 6% group and native tissue (Figure
4.3E-H). In the 9% strain stimulated group, the tendon samples were found to be
partially torn and graded 1.45 in fiber structure and 0.8 in fiber arrangement (Figure
4.3E, F). Increased cell number and slightly rounded tenocyte nuclei were observed
around the rupture site (Figure 4.3D, G, H).

143
Figure 4.3 6 days dynamic culture group. (A) Native Achilles tendon structure. (B) 3% stimulated group with wavy collagen structure and minor rounded tenocytes. (C) 6% stimulated group with similar
morphology compared with the native group. (D) 9% stimulated group with severe tear collagen structure, and rounded nuclei around the tear region (arrow). (E) Fiber arrangement, scale of 0-3, 0 represents compacted and parallel, and 3 represents no identifiable pattern. (F) Fiber structure, scale of 0-3, 0
represents continuous, long fiber, and 3 represents severely fragmented. (G) Cell roundness, scale of 0-3, 0 represents long spindle shape, and 3 represents severe rounding. (H) Cell density, scale of 0-3, 0
represents normal pattern, and 3 represents severely increase. Result are expressed as the mean±S.D. (*p<0.05, **p<0.01, **p<0.001). Scale bar=50 micrometers.

144
4.4.2 Impact of cyclic tensile strain on apoptosis of tenocytes
To investigate the impact of mechanical force on the apoptosis of tenocytes, I used the
TUNEL assay to examine the cellular apoptosis rate in specimens. In the native tendons,
the average cell apoptosis fraction (as a proportion of total cell number) was less than
10% (Figure 4.4A, G). The apoptosis fraction progressively increased over time when
the tissues were cultured without loading (35% in 6 days and 95% in 2 weeks) (Figure
4.4G). Cell viability in 3% and 6% stimulated Achilles tendon was similar to that in
native tendon (Figure 4.4D, E) without statistical significance (Figure 4.4G), whilst cell
apoptosis rate increased to 45% when the stimulation strain reach 9% (Figure 4.4F, G).

145
Figure 4.4TUNEL Assay (A) Native Achilles tendon with less than 10% cell apoptosis. (B) 6 days loading deprivation group has increased apoptotic tenocytes pointed out by arrows. (C) In 2 weeks
loading deprivation group, almost all tenocytes are under apoptosis (D) 3% stimulated group with minor increased apoptotic cells (arrow). (E) 6% stimulated group with similar cell apoptosis rate as native
sample (F) 9% stimulated group with massive cell apoptosis around the damaged site (pointed by arrows). (G) Quantified cell apoptosis fraction of each group. Result are expressed as the mean±S.D. (*p<0.05,
**p<0.01, **p<0.001). Scale bar=50 micrometers

146
4.4.3 Type III collagen turnover in Achilles tendon under dynamic culture
To investigate the extracellular matrix turnover in tendon undergoing different cyclic
tensile strain, type III collagen immunohistochemistry was done. No positive type III
collagen was observed in the core of healthy tendon using immunochemistry (Figure
4.5A). After 6 days of either no loading or 3% strain, the Achilles tendons expressed
only minor and moderate collagen type III respectively (Figure 4.5B,C), whilst
relatively higher positive staining were found in the 9% strain group (Figure 4.5E). No
type III collagen expression was observed in the 6% cyclic strain stimulated group
(Figure 4.5D), which was similar to the native tendon.
Figure 4.5Collagen type III immunohistochemistry (A) Native Achilles tendon with minor collagen type III expression. (B) Loading deprivation group has minor increased collagen type III expression and some positive staining tenocytes pointed out by arrow. (C) 3% stimulated group with increased collagen
type III expression. (D) 6% stimulated group with similar collagen type III expression as native sample (E) 9% stimulated group with massive increased collagen type III expression and positive staining tenocytes
(pointed by arrows). Scale bar=50 micrometers

147
4.4.4 Impact of cyclic tensile strain on ECM remodeling gene expression
Q-PCR was carried out to examine the gene expression of type I (COL1A2), type III
collagen (COL3A1) and MMP1, 3, 12 (matrix metalloproteinases). As shown in Figure 8,
the tissues subjected to 6% cyclic tensile strain had the highest expression of COL1A2
(Figure 8A), whereas 3% stimulated group had the lowest expression. Type III collagen
(COL3A1) was suppressed by minor and moderate loading (3% and 6%), yet
upregulated in the 9% strain stimulated group (Figure 8B). Gene expression of MMP1,
3 and 12 were highly upregulated by 3% strain stimulation compared to the other
groups.
Figure 4.6Quantitative PCR results. A: COL1A2 expression in 6% stimulated group is significantly higher than other groups. B: COL3A1 expression in 9% stimulated group is
significantly higher than other groups. Three percent and 6% group had much lower COL3A1 expression compared to the other groups. C–E: Three percent stimulated group has significantly high
expression on MMP1 MMP3 and MMP12, respectively. Result is expressed as the mean±S.D (*p<0.05, **p<0.01, **p<0.001)

148
4.5 Discussion
A previous study has suggested that engineered neo-tendons in a non-loading
environment result in loose, disorganized and less collagen-rich matrices (Jiang, Liu et
al. 2011). Static mechanical loading has been shown to improve the fiber arrangement
of engineered tendon constructs, but did not produce a compacted tissue structure in
vitro (Chen, Willers et al. 2007). Engineered neo-tendons with PMS had a
well-organized fiber arrangement and structure and enhanced tensile strength,
suggesting that dynamic mechanical loading is preferable for engineered tendon
formation (Jiang, Liu et al. 2011). Although PMS has been recognized as a critical
regulator for tendon formation (Brown and Carson et al. 1999), the optimal PMS
parameters are yet to be defined.
In this study we designed a uni-axial bioreactor system with a programmable
stimulation regimen. Achilles tendons from rabbits were used to evaluate the impact of
different PMS conditions on tendon tissue. Our result showed that 3% cyclic tensile
strain in the bioreactor did not prevent matrix deterioration, whilst at 6% cyclic tensile
strain structural integrity and cellular function was maintained. At 9% cyclic tensile
strain there was massive rupture of the collagen bundles. However, these results are for
an already mature tendon and the loading regimes may be different for engineered
constructs at various stages of growth. Furthermore, the frequency of loading may also
have an important role in maintaining tendon homeostasis and this need to be better
qualified by in vivo measurements of Achilles tendon loading. Nevertheless, the current

149
results demonstrate that PMS can be achieved in this bioreactor, and that an optimal
cyclic tensile loading (around 6% cyclic tensile strain) is required to maintain healthy
tendon homeostasis, at least over the time period of the cultures used in this study.
It has been well known that tensile mechanical loading is essential for tendon integrity,
and loading deprivation can cause tendinopathy-like morphology such as disorientated
collagen fibers and rounded cell nuclei(Hannafin, Arnoczky et al. 1995). Yet the specific
effect of different loading conditions on tendon has not been well elucidated. To study
the effect of PMS on tendon/ligament, both in vitro and in vivo models have been used.
The Flexcell® system has been applied to examine the in vitro gene expression profiles
of tenocytes under tensile or shear force (Skutek, van Griensven et al. 2001; Wang, Jia
et al. 2003; Yang, Im et al. 2005). These systems can precisely control the mechanical
regimen and monitor the cell behaviors, but only use a monolayer of cells, not an
organized tissue. Cells isolated from their extracellular niche and cultivated in a
monolayer may not necessarily replicate their original behavior in vivo. In vivo models,
such as Achilles tendon in rats or rabbit, have been commonly used as study models.
Tendon loading is mainly induced by electrical stimulation on muscle(Nakama, King et
al. 2005; Asundi, King et al. 2008), treadmill training on animal and partial tendon
transaction(Smith, Sakurai et al. 2008). However, mechanical stimulation on the tendon
varies between individuals, making precise quantification of load next to impossible. In
this study, we have demonstrated a bioreactor system that not only can be used as an
device for tendon/ligament engineering, but also serves as an ex vivo culture model for
the study of tendon biology. Although the biomechanics of Achilles tendon is more

150
complicated under in vivo environment (West, Juncosa et al. 2004), by applying
programmable mechanical stimulation, the effect of different cyclic tensile strain on
tendon tissue could be systematically analyzed on a tissue level.
The present study has shown for the first time that only a narrow range of PMS can
stimulate the anabolic effect of tendon tissue. This suggests a proposed model
describing the effect of mechanical loading, from under-load to over-load, on tendon
homeostasis. This model is illustrated in Figure 4.7 in which the black curve presents
the typical mechanical response caused by increasing tensile strain in tendon. The
breakage of collagen fibers increases with increasing strain, leading to eventual rupture
at the failure point. The red curve, on the other hand, proposes the biological repair
response of the tendon induced by tensile strain. Mechanical loading is required for the
biological activity of tenocytes, yet the regime needs to be within a certain range to
promote matrix product and net tissue repair. By combining these two curves, three
crossover areas are seen, which can be divided according to their metabolic status into:
A (anabolic) zone and C (catabolic) zones. In the A zone, where the tensile loading is
within a certain range, the biological repair response caused by PMS exceeds the
induced damage. In the C zone, there is either too little load to stimulate matrix
production or too much mechanical damage caused by excess tensile loading. In our
study the 3% strain stimulated group (under-loaded) was located in C zone. When the
tendons were subjected to insufficient tensile strain, early tendinopathologic changes
(including increased cellular number, type III collagen expression and disorientated
collagen fibers) could be observed. These morphological changes may be attributed by

151
the up-regulation of MMPs expression in tendon under 3% tensile loading. MMPs are
zinc-dependent endopeptidases that able to degrade all the components of the
extracellular matrix.MMP-1 is primarily responsible for the cleavage of collagen,
including type I, II and III(Krane, Byrne et al. 1996). MMP-3 and MMP-12 are other
members of the MMP family that have a wide range of substrate specificities. Moreover,
MMP-12 is able to activate MMP-3, and MMP-3 has the ability to activate other MMPs
such as MMP-1, MMP-7 and MMP-9 (Knauper, Lopez-Otin et al. 1996; Chen 2004).
Studies have shown that decreased mechanical force on tendon fiber would lead to the
deprivation of cytoskeletal tension in tenocytes and alter the mechanostransduction in the
cells (Arnoczky 2008). Insufficient loading of tenocyte can initiate the degenerative
cascade by expressing a pattern of catabolic gene, including MMPs(Lavagnino,
Arnoczky et al. 2003; Arnoczky, Tian et al. 2004; Lavagnino, Arnoczky et al. 2006;
Arnoczky, Lavagnino et al. 2008; Egerbacher, Arnoczky et al. 2008).In this study, we
have confirmed that under the underloading condition, the phenotype of the tenocytes
changed from spindle to round, and the expression levels of catabolic genes, such as
MMP-1, 2, and 12, were upregulated. The elevated catabolic factors may contribute to the
further micro-damage of the collagen fiber. Conversely, although the tendons in the 6%
strain stimulated group were subjected to more mechanical damage, the repair
mechanism was able to maintain tissue homeostasis (i.e. there is a narrow ‘net anabolic
window’ between two net catabolic zones). In the 9% group (overloaded), mechanical
damage overwhelmed the tendon tissue repair and caused partial rupture, which is
consistent with our proposed model.

152
The identification of an optimal PMS range for tendon will not only guide us to a better
tissue-engineering pathway for tendon regeneration, but also provide an insight into
tendon homeostasis under both physiological and pathological conditions. Clinically,
chronic tendinopathy of Achilles tendon is usually conservatively managed by eccentric
overload training. The calf muscle is eccentrically loaded with the knee held straight or
the soleus muscle is maximally loaded with the knee bent(Alfredson, Pietila et al. 1998).
Although positive impacts of eccentric overload training on tendinopathy treatment
have been recorded, the outcomes have varied widely (Stanish, Rubinovich et al. 1986;
Alfredson, Pietila et al. 1998; Silbernagel, Thomee et al. 2001; Alfredson and Lorentzon
2003; Fahlstrom, Jonsson et al. 2003; Roos, Engstrom et al. 2004; Shalabi,
Kristoffersen-Wilberg et al. 2004). In the study of Alfredson et al.,(Alfredson, Pietila et
al. 1998) the overall improvement in pain score was up to 94%, though in the study of
Silbernag el et al. reported only 29% (Alfredson and Lorentzon). Although the
mechanism of eccentric overload training on tendinopathy management remains unclear,
the present study has provided a possible explanation of the mechanism of eccentric
loading on chronic tendinopathy. The high variation of clinical outcomes might be due
to poor control of the loading regimen. Given that there is only a narrow range of tensile
strain where eccentric exercise could provide an anabolic effect on tendon, clinical
treatment protocols will need to accurately control the magnitude of load applied
through the injured tendon.
Finally, it is noted that nutrient supply might be altered by advective transport
associated with mechanical stimulation. Although a previous study on canine flexor

153
tendons suggested that mechanical stimulation did not alter small nutrient uptake
compared to static culture, and the tendon repair improvements derived from passive
motion is not related to an increase of nutrient transport (Hannafin and Arnoczky 1994),
under different loading regimes, or in different tendon types, this may not be the case.
Moreover, the transport of large molecules, such as growth factors and matrix
components, may be affected by mechanical stimulation, as has been shown in studies
in related tissues (Bonassar, Grodzinsky et al. 2000; Zhang, Gardiner et al. 2007).
Therefore, the tendon stimulated by dynamic mechanical loading in this study might be
attributed to the enhanced nutrient transport, compared to the static control.
Consequently it is difficult to isolate the direct effect of mechanical stimulation on
tendon response from potential enhanced advective transport processes. Further study
would be required to measure the nutrition transport in tendon under PMS

154
Figure 4.7 Matrix damage (black curve) and production (red broken) affected by mechanical stimulation. A zone indicates a small range of anabolic effect caused by mechanical stimulation. Matrix
production stimulated by mechanical loading overcomes the matrix damage that the tendon is able to maintain its structural integrity. C zone indicates two catabolic zones that matrix damage caused by
mechanical stretching overcomes the matrix production result in tendinopathy.
4.6 Conclusion
We have developed and validated a uniaxial bioreactor system. The results in our study
showed tendons subjected to 6% of cyclic tensile strain were able to maintain their
structural integrity and cellular function. Our data suggested that tendon is a
mechano-sensing tissue where the net anabolic action of cyclic tensile strain only occurs
over a narrow range. In our study, mechanical underloading (3% cyclic tensile strain)
and overloading (9% cyclic tensile strain) produced a net negative impact on the rabbit
Achilles tendon tissue.

155
Chapter 5
Cyclic mechanical stimulation
rescues rabbit Achilles tendon in
a bioreactor system
This chapter is based on the paper
Cyclic mechanical stimulation rescues rabbit Achilles tendon from degeneration in a bioreactor system
Tao Wang, Zhen Lin, Ming Ni, Robert E Day, Bruce Gardiner, Euphemie Landao-Bassonga, Jonas Rubenson, Thomas B. Kirk, David W. Smith, David G. Lloyd,
Yan Wang, Qiujian Zheng, Ming H Zheng
(Submitted to Tissue Engineering Part A)

156
CHAPTER 5 (RESULTS): CYCLIC MECHANICAL
STIMULATION RESCUES RABBIT ACHILLES TENDON FROM
DEGENERATION IN A BIOREACTOR SYSTEM
5.1 Abstract
Mechanical stimulation has been identified as an essential factor for maintaining tendon
homeostasis, and various studies have reported that Achilles tendons subjected to
long-term in vivo immobilization display pathological changes while mechanical
properties degrade. Our previous study showed that 6% strain cyclic mechanical
stimulation is able to maintain the tendon homeostasis in the bioreactor system.
Therefore, in this study, we hypothesize that 6% strain cyclic mechanical stimulation
can rescue morphological and mechanical degeneration similar to tendinopathy ex vivo.
New Zealand white rabbits and divided into 4 groups: native control group, 6 days and
12 days loading deprivation groups, and stimulation group. In loading deprivation
groups, the Achilles tendons were cultured without loading for either 6 days or 12 days.
In stimulation group, tendons were subject to 6% cyclic strain at 0.25Hz for 8 h/d for 6
days after 6 days no loading culture. Tendons in both loading deprivation groups
developed progressive histological degradation, increased tenocytes apoptosis and
weakened mechanical properties after loading deprivation. However, in stimulation
group, the histological and mechanical properties substantially improved. Our results
indicate that a 6% strain cyclic mechanical stimulation was able to rescue early-stage
pathological changes from loading deprivation and restore Achilles tendon ex vivo.

157
5.2 Introduction
Clinical findings have demonstrated tendon degeneration and compromised
biomechanical properties occurs after load deprivation, including in situations ranging
from bedridden patients to astronauts(Reeves, Maganaris et al. 2005). Studies have
reported that early-stage pathological alterations in Achilles tendon following long term
immobilization in both human and rat (Larsen, Forwood et al. 1987; Nakagawa, Totsuka
et al. 1989; Kannus, Jozsa et al. 1997; Trudel, Koike et al. 2007). The previous study
(Chapter 4) observed that rabbit Achilles tendon after load deprivation exhibited
pathological changes that included: (i) wavy and ruptured collagen fibers, (ii) rounded
cell morphology and (iii) increased cell apoptosis (Wang, Lin et al. 2013). Accumulated
evidence, including Chapter 4, shows that in order to maintain normal tendon
homeostasis, mechanical stimulation is required(Arnoczky, Lavagnino et al. 2007;
Egerbacher, Arnoczky et al. 2008; Smith, Sakurai et al. 2008; Thornton, Shao et al.
2008; Wang, Gardiner et al. 2013). The key question is can functional tendon be
restored after pathological changes due to unloading?
In the previous study, appearing in Chapter 4, it was found that a relatively narrow
range of mechanical stimulation is able to maintain rabbit Achilles tendon in the
bioreactor, whereas too little strain results in protease degradation while too much strain
results in mechanically induced fatigue damage. For Achilles rabbit tendon it was found
that a 6% strain set in a cyclic mechanical stimulation device is able to maintain tendon
integrity in a bioreactor system. This current study examined if it was possible to rescue

158
the early pathological changes caused by 6 days of loading deprivation, similar to that
observed in tendinopathy, by then applying 6% strain cyclic loading for a further 6 days.
The positive effects of mechanical loading following load deprivation reported here
have implications for all kinds of situations involving tendon degeneration, ranging
from physical treatment following immobilization to the tendon healing process
following surgery, and even to planning the most effective exercises for astronauts.
5.3 Material and method
5.3.1 Tissue preparation and programmable mechanical stimulation
Full-length Achilles tendons, including the bone-tendon and tendon-muscle junction
were dissected from the hindlimbs of 15 weeks old female New Zealand white rabbits
(Oryctolagus cuniculus). Animals were obtained immediately post-euthanasia.
By-products from tendon samples were washed by 1x phosphate buffered saline (PBS)
3 times, and submerged in serum free Dulbecco’s modified Eagle’s medium
(DMEM/F-12; GIBCO, Invitrogen, Auckland, New Zealand) supplemented with 50
µg/mL Gentamicin (Invitrogen, Auckland, New Zealand) before further processing.
In total, 72 Achilles tendons from 36 rabbits were evenly allocated to 4 groups (see
Figure 1). In the native control group 18 Achilles tendons were dissected from the
rabbits and stored for further experiments. In the 6 and 12 day load deprivation groups
(LD6, LD12), 18 tendons each were cultured in the growth medium (DMEM/F-12, 10%

159
fetal bovine serum and 50 µg/ml Gentamicin) on a petri dish without mechanical
loading at 37℃ in a humid environment with 5% CO2 for either 6 or 12 days. In the
stimulation group an additional 18 rabbit Achilles tendon were cultured on petri dish
without mechanical loading for 6 days, followed by 6 days in the bioreactor chambers
undergoing 6% cyclic tensile loading.
Figure 5. 1 Flowchart of experimental plan and rabbit Achilles tendon allocation (LD: loading
deprivation)
In the bioreactor chambers, the Achilles tendons were secured to the tissue hook using
surgical sutures, as shown in Figure 5.2A, and then placed into the bioreactor chamber,
which was in turn placed in a standard incubator at 37℃ in a humid environment with

160
5% CO2 (Figure 5.2B). During the dynamic culture process, the tendons were subjected
to 6% cyclic strain mechanical stimulation at 0.25Hz for 8 hours followed by 16 hours
rest (7200 cycles/day) for total 6 days (see Figure 5.2C).
Figure 5. 2 (A) Rabbit Achilles tendon was fixed in a bioreactor culture chamber; (B) Bioreactor setup in the incubator; (C) mechanical stimulation regime applied in the stimulation group; (D) Biomechanical testing with a custom cryogrips. The arrow indicates the rupture of
Achilles tendon happened at the middle portion.
In each group, 6 samples were fixed in 4% paraformaldehyde overnight for
histological analysis, 6 samples were directly subjected to RNA extraction, and the rest
were wrapped by PBS soaked gauze and stored at -20℃ for further mechanical testing.

161
5.3.2 Histological preparation and assessment
Refer to 2.5
5.3.3 TUNEL assay
Refer to 2.6
5.3.4 Biomechanics testing
To examine the effect of mechanical stimulation on the biomechanical properties of
tendon, we performed mechanical testing using a modified dual frozen technique (Chen,
Yu et al. 2011). Nine tendons from each group were used for the testing. The samples
were thawed at room temperature on the day of testing. Both bone-tendon and
tendon-muscle ends were placed in a pair of custom-designed cry grips. To prevent
tissue sliding, both ends were snap-frozen by liquid nitrogen. A thermometer was used
to monitor the temperature of the tissue between two cryogrips, to avoid the testing
substrate freezing. The grips were then fixed into the Instron mechanical testing system
(model 5566, Instron, China), and the middle portion of the Achilles tendon were
subjected to tensile loading to failure (Chen, Yu et al. 2011). The samples were
preloaded at 2N, then gradually pulled at a speed of 2mm/second until it reach the
ultimate force and dropped to failure force. Failure force was preprogrammed as 70% of
ultimate force. During the experiment, the samples were closely monitored. To confirm
that each tested tendon was broken at the middle portion but not the point near the grip

162
(Figure 2A). A force versus displacement curve was recorded, and the ultimate load was
recorded. Mean stiffness was calculated based on the linear region of the loading curve.
5.3.5 Real-time polymerase chain reaction
Refer to 2.3 and 2.4
5.3.6 Immunohistochemistry
Refer to 2.6
5.3.7 Statistical analyses
Refer to 2.8
5.4 Results
5.4.1 Cyclic mechanical stimulation improved the histological structure and cell
viability of cultured tendon
The Achilles tendons cultured under loading deprivation exhibit minor wavy and loose
collagen structure (Figure 5.3B), compared to native tendon (Figure 5.3A), more gaps
between the collagen fiber were observed, and the tenocytes started to lose their

163
elongated morphology (Figure 5.3F). When the culture period extended to 12 days,
disorientated fibers with progressive extracellular matrix (ECM) disruption and rounded
cell nuclei were observed (Figure 5.3C, G). These morphological changes are consistent
with the pathology of tendinopathy. Interestingly, when 6% strain cyclic mechanical
loading was applied on the Achilles tendon after 6 days loading deprivation culture, the
tendon morphology seems to be similar to native tendon. Collagen fibers were found to
be straight and well organized (Figure 5.3D), with no gap was observed between
collagen fibers, while long spindle-shape cell morphology was successfully restored
(Figure 5.3H).
Figure 5. 3 Comparison of morphological features of Achilles tendons. (A) Native Achilles tendon tissue; (B) 6 days loading deprivation culture; (C) 12 days loading deprivation culture; (D) 6 days loading deprivation culture followed by 6% strain dynamic culture; (E) Elongated spindle cell morphology in native tendon tissue; (F) Tenocytes started to lose their original morphology and turn
round after 6 days loading-deprived culture; (G) Round tenocytes; (H) 6% strain stimulation successfully restore the long spindle morphology of tenocytes. Scale bar is 20m.
Using the quantitative scoring system (Figure 5.4), the 6 days loading deprivation group

164
expressed a slight abnormality in fiber arrangement, i.e. fiber structure, cell roundness,
and a moderate increase of cell density. In addition, the overall score further increased
as the loading deprivation culture period was extended to 12 days, whereby every aspect
increased significantly except for cell density. For the stimulation group, the tendon
successfully regained normality, in both the overall score and the individual scores with
no significant differences observed when compared to native tendon.
Figure 5. 4Histological score of three groups. (A) Overall histology score, and the 6% rescue group is significantly better than the other groups; (B) fiber arrangement; (C) fiber structure; (D) cell roundness; (E) cell density. Results are expressed as the mean±SEM. One way ANOVA significance values were *p
< 0.05, **p < 0.01, and ***p < 0.001
In regard to cell apoptosis, the 6 day loading deprivation group exhibited numerous

165
positive TUNEL stained tenocytes, as can be observed under the microscope image
(Figure 5.5B), while for the 12 day loading deprivation group, large numbers of
apoptotic cells were observed (Figure 5.5C). However, there were only a few apoptotic
cells detected in the stimulation group (Figure 5.5D), similar to that seen in the native
tendon (Figure 5.5A). Quantification of these observations showed that cell apoptosis
rate was around 30.6% for the 6 days loading deprivation group, which further
increased to 91.3% in the 12 days loading deprivation group. However, the stimulation
group’s tissue had a 17.8% cell apoptosis rate that was significantly lower than either
load deprivation groups (Figure 5.5E), and only slightly higher than the native group.

166
Figure 5. 5TUNEL assay (A) Apoptotic cell was rarely detected in native Achilles tendon tissue (B) In 6 days loading deprivation group, around 30% of tenocytes were under apoptosis, pointed by arrows; (C) In 12 days loading deprivation group, almost all the tenocytes were undergo apoptosis;
(D) apoptotic tenocytes significantly decreased compared to the other experimental groups; (E) Quantitative cell apoptosis rate. Results are expressed as the mean±SEM. One way ANOVA
significance values were *p < 0.05, **p < 0.01, and ***p < 0.001 Scale bar is 20mm.

167
5.4.2 Cyclic mechanical stimulation improved mechanical properties of cultured
tendon
As the force transferring tissue, mechanical properties are the fundamental characteristic
of tendon, although the pathological changes in the tendon microstructure were
observed, the impact on the overall mechanical properties is still unclear. Therefore,
mechanical testing was performed to examine the strength of the Achilles tendon after
the ex vivo culture. In the mechanical tests, all samples failed at the mid-tendon region,
which was the region of interest in this study (Figure 5.2D). As shown in Figure 5.6A,
the average ultimate tensile force after 6 days of loading deprivation was 613.6±49.7 N,
which was significantly higher than 12 days loading deprivation group (449.8±55.9 N)
but lower than stimulation group (773.3±89.6 N). However, all the culture groups had a
significantly lower ultimate tensile force compared to the native Achilles tendon (896.1
±96.1 N).
The mean stiffness (Figure 5.6B) of the stimulation group was 165.6±39.1 N/mm,
which was significantly higher than 6 days and 12 days loading deprivation groups
(99.2 ± 22.4 N/mm, 52.5 ± 12.8 N/mm), while there was no statistical difference
compared to the native controls (193.1±26.8 N/mm).

168
Figure 5. 6 (A) The maximum load of Achilles tendon tissue. The maximum loads decreased with extended cultured time in static cultured group, however, after 6 days static culture, 6% stimulation group is able to restored part of mechanical properties. Compared to native control tissue, the maximum loads in
cultured group are significantly lower. (B) The mean stiffness of Achilles tendon tissue. The mean stiffness of static cultured group dropped dramatically with extended culture period. However, in 6% rescue group, no significant difference was observed compared to native control tendon. Results are
expressed as the mean±SD. One way ANOVA significance values were *p < 0.05, **p < 0.01, and ***p < 0.001
The force versus displacement curves of native Achilles tendons, 6 days loading
deprivation group and stimulation group are similar (Figure 5.7). In these the force
increased dramatically for the first 4mm displacement, and then moderately increased,
which was followed by a sudden drop in the tensile force as the tendons began to fail.
However, in the 12 days loading deprivation group, the force climbed moderately for
the first 8mm and slowly decreased to 70% of ultimate force, at which point the tests
were terminated.

169
Figure 5. 7 Force versus displacement curves for the native Achilles tendon (Control), rescue group (0% 6d+6% 6d), 6 days static cultured and 12 days static cultured Achilles tendon. 6% rescued cultured tendon displays similar mechanical character with native Achilles tendon. After 12 days static culture,
Achilles tendon lost its original mechanical character became less stiff and soft.
5.4.3 Rescue effect of cyclic mechanical stimulation on ECM remodeling gene
expression
Although the histological and mechanical changes have been identified, the underlying
molecular mechanism still remains unclear. The homeostasis of tendon required the
balance of the ECM generation and degradation. To estimate the generative process, the
gene expression of COL1A1, COL3A1, TGFbwere tested, whiles MMP1, MMP3,
TIMP1, and TIMP2 were considered to be the predominant cytokine in tendon
remodeling. All the gene expression level was examined using RT-PCR. As shown in
Figure. 5.8, although there was a slight decrease on COL1A1 expression of the loading

170
deprivation groups compared to native control, no significant difference was detected;
however, COL1A1 expression increased dramatically in the stimulation group. A similar
trend was observed in TGFband MMP3 expression. For the first 6 days no loading
culture, MMP 1 was highly expressed, yet it dropped with extended culture period. In
the stimulation group, MMP 1 was less expressed compared to 6 days loading
deprivation group, however, no statistical difference was detected compared to the 12
day loading deprivation group. On the other hand, TIMPs expressions of culture tendon
were significant higher than the native control except for the 6 day loading deprivation
group on TIMP2. Extensive increase on TIMPs expressions were detected on
stimulation group, especially TIMP1. However, no significant difference between
stimulation group and 12 days loading deprivation groups on TIMP2.
Figure 5. 8 Comparison of gene expression fold change normalized by native tissue gene expression amount in the 6 day and 12 day loading deprivation group, the stimulation group and the native control group, the gene expression level is normalized against 36B4. Result are expressed as
the mean±SEM. One way ANOVA significance values were *p < 0.05, **p < 0.01, and ***p < 0.001

171
5.4.4 Cyclic mechanical stimulation decreased Type III collagen expression of
cultured tendon
Increased collagen type III content is an index of tendon degeneration and tendinopathy.
Tendon cultured under loading deprivation condition for 6 days started to express
moderate collagen type III turnover and several positive stained tenocytes were detected
(as pointed by arrow in Figure 5.9B). Furthermore, almost all the tenocytes in the 12
days loading deprivation group were expressing type III collagen, and substantial type
III collagen was observed histologically (Figure 5.9C). Interestingly, in the stimulation
group, collagen type III expression was only minimally elevated, but at a level (Figure
5.9D) similar to native tendon (Figure 5.9A).
Figure 5. 9 Type III collagen immunohistochemistry, positive staining is indicated by arrows (A) Type III collagen rarely detected in native Achilles tendon tissue (B) In 6 days loading deprivation group, type III collagen was upregulated, positive staining were pointed by arrow; (C) In 12 days
loading deprivation group with massive increased type III collagen expression ; (D) No obvious type III collagen expression detected in 6% rescue group. Scale bar is 20m.

172
5.5 Discussion
It is well known that mechanical stimulation plays a fundamental role in maintaining
tendon homeostasis, and that the absence of mechanical stimulation can cause
morphological changes such as rounded cell nuclei and disorientated collagen fibers
associated with tissue degeneration (Hannafin, Arnoczky et al. 1995; Wang, Lin et al.
2013). “Tendon exercise” such as eccentric overload training or concentric training
seems to be effective in patient rehabilitation, however, the mechanism still remains
unclear(Kingma, de Knikker et al. 2007). In Chapter 3, a bioreactor system was
developed that could be used to mimic the in vivo tensile mechanical loading on
Achilles tendon and in Chapter 4 it was verified that cyclic mechanical stimulation
indeed was essential for the maintenance of the tissue integrity of tendon. We also
suggested that a narrow range of mechanical stimulation was required to create a net
anabolic effect in Achilles tendon, and identified that 6% strain cyclic mechanical
stimulation was optimal to maintain Achilles tendon homeostasis in the bioreactor
system(Wang, Lin et al. 2013). However, the therapeutic effect of mechanical
stimulation on early-stage of pathological change in Achilles tendon was yet to be
observed.
In this study, this dynamic bioreactor culture system was combined with the unloaded
culture condition to examine the effect of cyclic mechanical stimulation on deteriorated
tissue caused by loading deprivation. Loading deprivation was achieved by culturing
the rabbit Achilles tendons freely on the petri dish with adequate growth medium. After

173
6 days of loading deprivation, early-stage degenerative changes were observed,
including wavy collagen fiber, increased type III collagen expression, and slightly
rounded nuclei. After 12 days of culture without loading, further severe pathological
changes such as disorientated collagen fiber pattern, ruptured fibers, rounded nuclei and
substantial increased type III collagen expression developed evenly throughout the
tendon. The deterioration of the tendon appeared to be no different at the center and the
edge of the tissue, suggesting that inadequate nutrition penetration was not the sole
cause of the damage. In the stimulation group, the tendons were transferred into the
bioreactor system after 6 days loading deprivation, and 6% strain cyclic mechanical
strain stimulation was applied for a further 6 days. The stimulation tendons successfully
regained their original morphology, exhibiting as straight, well-organized collagen
fibers and cells with elongated nuclei, whilst the type III collagen expression had
reduced to the normal level. The result suggested that the previously identified optimal
cyclic tensile loading (6%) was able to repair the early pathological change of tendon
caused by load deprivation.
Programmed cell death (apoptosis) has been identified as a common feature of tendon
degeneration and injury (Yuan, Wang et al. 2003; Hosaka, Teraoka et al. 2005; Lian,
Scott et al. 2007). I previously showed that mechanical loading was essential for
tenocyte viability, where increased cell apoptosis was observed in the tendon tissues that
has no mechanical loading (Wang, Lin et al. 2013). In this study, the cell apoptosis rate
in the 6 and 12 days loading deprivation groups were 30.6±3.9% and 91.3±3.5%
respectively, which might explain why in the 12 day loading deprivation group, no

174
increased cell density was detected. When the tendon developed early degeneration, cell
proliferation increased, presumably to activate repair. However, in the absence of
mechanical stimulation, the observed high cell apoptosis rate inhibited this repair
cascade then reducing the cell population to a “normal” level. The stimulation group,
on the other hand, expressed an apoptosis rate of 17.8±3.9%, which was closed to that
of healthy tendon tissue, suggesting that 6% strain cyclic mechanical stimulation
eliminated cell apoptosis in early-stage degradation and may thus contribute to the
restoration of the homeostasis of tendon.
The microstructural organization of the tendon’s extracellular matrix, particularly
collagen type I, determines the mechanical properties of tendon tissue. Although the
disturbed collagen structure and increased collagen type III turnover were observed in
loading deprivation culture groups, this was rescued by 6% strain cyclic loading
stimulation. Critically mechanical testing confirmed that the mechanical stimulation
restored normal mechanical properties of the Achilles tendon.
The biomechanical characteristics of tendon have not been well documented in
literature as failure often occurs at sites other than the middle portion of tendon, such as
muscle-tendon junction, tendon-bone junction or surgical repair site (Thermann,
Frerichs et al. 2001; Matsumoto, Trudel et al. 2003; Gebauer, Beil et al. 2007; Chen, Yu
et al. 2011). To ensure the efficacy of the testing, a pair of modified cryogrips and the
freezing technique using liquid nitrogen was adopted in the study to avoid sliding of the

175
tissue during testing. During the mechanical loading, the tissue was closely monitored
to ensure it broke at the middle portion rather than the ends near the grips. The peak
load of native control tissue tested for the whole Achilles tendon in this study was 896.1
±96.1 N, which is comparable to the findings reported in the study by Trudel et
al.(Trudel, Koike et al. 2007). The results of biomechanical testing showed significantly
reduced mechanical strength and stiffness of Achilles tendon cultured for 6 days and 12
days load deprivation. The mechanical characteristic could be better described from the
force versus displacement curves. Due to ruptured and disorientated collagen fibers, the
force increased more slowly in 12 days loading deprivation group compared to the other
groups, for a given strain. However, in the stimulation group, the rabbit Achilles tendons
regained their mechanical properties to levels close to that of normal tissue. This was
evidenced by there being no significant difference in the mean stiffness of the
stimulation group compared to that of the native control group, and the
force-displacement curve in stimulation group appearing similar to that of the native
control, apart from a lower ultimate tensile strength. Thus, the testing demonstrated a
positive effect of mechanical stimulation in preventing further tendon degeneration, and
indeed in promoting tendon healing.
The homeostasis of tendon requires constant remodeling of the collagen through the
generation of new collagen in response to the degradation of collagen by proteases and
mechanical loading. Matrix metalloproteinase (MMPs) are zinc-dependent
endopeptidases that are able to degrade all the components of the extracellular matrix.
MMP-1 is primarily responsible for cleavage the collagen type I, the dominant collagen

176
of tendon (Knauper, Lopez-Otin et al. 1996; Krane, Byrne et al. 1996). MMP-3 is
another subset of the MMP family with a wide range of substrate specificity. Moreover,
it has the ability to activate other MMPs such as MMP-1, MMP-7 and MMP-9(Ye,
Eriksson et al. 1996). TIMPs, on the other hand, are the inhibitors of MMPs. The ratio
of MMPs to TIMPs is the key to understanding the rate of protease degradation, which
then impacts on tendon homeostasis. Over expression of MMPs or under expression of
TIMPs will cause the pathogenesis of tendon degradation (Riley, Curry et al. 2002; Lo,
Marchuk et al. 2004; Jones, Corps et al. 2006). In our study, in the first 6 days of
loading deprivation culture, MMP1 and MMP3 were significantly up-regulated, while
TIMP1 and 2 were down-regulated. The shift in balance between MMPs and TIMPs
may explain the early-stage pathological changes observed in the tendon. However, in
the stimulation group, the up-regulation of TIMP1 and down-regulation of MMP1 may
have suppressed the protease degeneration of collagen fibers. Upon mechanical
stimulation, expression of TGF- and COL1A2 promoted the formation of collagen
fiber. The RT-PCR results are consistent with our observation that 6% cyclic tensile
strain mechanical stimulation was optimal for creating a net anabolic effect that could
reverse the early-stage degenerated changes.
Immobilization is a common postoperative treatment for bone fracture and tendon injury.
For example, fracture of the lower end of the tibia or rupture of the Achilles tendon
commonly require immobilization for up to 6 weeks (Cetti, Henriksen et al. 1994)
(Ruedi and Allgower 1979), resulting in the deprivation of physical dynamic loading of
the tendon. Early-stage structural degradation of Achilles tendon has been reported after

177
long-term immobilization in both humans and rats (Larsen, Forwood et al. 1987;
Nakagawa, Totsuka et al. 1989; Kannus, Jozsa et al. 1997; Trudel, Koike et al. 2007).
However, clinical rehabilitation guideline for Achilles tendon weakness after a period of
immobilization has not been well established. In this study, we have demonstrated that
loading deprivation in a bioreactor tendon culture led to tendon structural deterioration
compromised mechanical properties and elevated expression levels of catabolic markers.
By applying 6% strain cyclic mechanical stimulation, the histomorphology and
biomechanical properties of rabbit Achilles tendon were successfully restored. However,
all characteristics of the preferred cyclic strain profiles need to be defined, i.e.
magnitude, frequency and dosage, and these may vary between species. Interestingly, a
recent review suggests that about 5% strain may be the value that affects human
Achilles tendons in vivo(Obst, Newsham-West et al.). Even though all strain profile
characteristics are to be elucidated, the findings of this current study suggest that
precisely calibrated mechanical stimulation is able to reverse early-stage tendon
degradation. It has been reported that compared to long episodes of early loading, short
periods of mechanical stimulation during the inflammatory phase could improve tendon
healing with histological signs of repeated damage in the form of increased bleeding
(Enwemeka, Spielholz et al. 1988; Eliasson, Andersson et al. 2011). Taken together,
accurate and controllable mechanical stimulation on injury tendon may be considered as
a potential therapeutic approach for patient rehabilitation. A specially designed orthotic
device could be developed to apply an optimal titration of strain on tendon tissue and
facilitate tendon repair.

178
5.6 Conclusion
In summary, our findings suggest that loading deprivation induced degenerated
pathogenesis in rabbit Achilles tendon could be rescued by 6% cyclic tensile loading
stimulation. The findings suggest that loading deprivation, such as during long-term
immobilization, should be avoided when possible to reduce the complication of tendon
weakness in patients. In addition, the benefit of dynamic loading may be an inspiration
to develop a rehabilitation device for tendon weakness after long-term immobilization.

179
Chapter 6
General Discussion and Future
Directions

180
CHAPTER 6 GENERAL DISCUSSION AND FUTURE
DIRECTIONS
6.1 General discussion
Tendon is a soft connective tissue between muscle and bone and functions to transfer
forces. Tendons are also responsible for storing elastic energy. Tendons are subjected to
high mechanical loading during daily activities and must undergo continuous repair to
maintain its functional properties. Owing to its specific mechanical function, it is
unsurprising that its biomechanical environment is essential for maintaining tendon
homeostasis.
In 1995, Hannafin et al. demonstrated that even without mechanical loading, tendons
undergo tendinopathy-like morphological changes characterized by disorientated
collagen fibers and rounded cell nuclei(Hannafin, Arnoczky et al. 1995). Manipulation
of loading on the tendon is able to regulate tendon metabolism and activate self-repair
mechanisms. Eccentric overload is one of the known effective interventions for patients
with chronic Achilles tendinopathy. By maximizing the activation of soleus or
gastrocnemius muscles, mechanical loading is able to be applied on tendon(Kingma, de
Knikker et al. 2007). Traditionally, immobilization is a standard recommendation
post-surgical tendon repair. However, studies have shown that short periods of
mechanical stimulation during the inflammatory phase could improve tendon healing
(Enwemeka, Spielholz et al. 1988; Eliasson, Andersson et al. 2011). This is also
consistent with the results present in this thesis (Chapter 5). Various studies on tendon

181
engineering suggest that mechanical stimulation is able to increase cell proliferation,
collagen deposition and improve the mechanical properties of neo-tendon tissue(Altman,
Lu et al. 2002; Juncosa-Melvin, Shearn et al. 2006; Webb, Hitchcock et al. 2006;
Androjna, Spragg et al. 2007; Nirmalanandhan, Rao et al. 2008; Nirmalanandhan,
Shearn et al. 2008; Butler, Hunter et al. 2009; Nguyen, Liang et al. 2009; Chen, Yin et
al. 2010; Doroski, Levenston et al. 2010; Saber, Zhang et al. 2010; Issa, Engebretson et
al. 2011; Woon, Kraus et al. 2011). Previous evidence suggests that mechanical
stimulation is able to confer beneficial effects on tendon regeneration, although the
reported range of improvements is varied. Silbernagel et al. showed that in 12 weeks of
eccentric training, the overall improvement on pain relief was 29%(Silbernagel, Thomee
et al. 2001), however, in a study by Alfredson et al., 94% improvement was reported
(Alfredson, Pietila et al. 1998). Owing to limited understanding of the biomechanical
environment in tendon tissue and the tissues’ repair and remodeling response, the
explanation of the present results within a physiological framework was difficult. It is
well known that sudden and excessive loading on the tendon can cause tear or rupture
and loading deprivation can cause tendon degradation. However, the physiology of
loading and underloading is rarely studied. Therefore, the investigation of various
loading conditions on the tendon is essential for understanding tendon biology and the
development of treatments for tendon repair.
6.1.1 The custom-made bioreactor (Chapter 3)
In the past decade, various methodologies have been employed to study the effect of
mechanical loading on tendon such as in vitro monolayer cellular models and in vivo

182
animal models. In the cellular studies, the Flexcell® system has been widely used
(Skutek, van Griensven et al. 2001; Wang, Jia et al. 2003; Yang, Im et al. 2005).
Although precise control and monitoring systems are allowed at the cellular level, the
application is on a monolayer of cells, not an organized tissue. Cells isolated from their
extracellular niche and cultivated in a monolayer may not necessarily replicate their
behavior in vivo. In vivo models, on the other hand, allow the systemic study of the
biomechanical environment of the tendon. However, mechanical stimulation on the
tendon varies between individuals, making precise quantification of load difficult.
Therefore, in order to study the effect of various mechanical stimulations on tendon, a
bioreactor that can provide a sterilized and programmable biomechanical environment
for tendon culture is necessary.
In the first study, we adopted a relatively simple “incubator-based” design, in which the
culture conditions such as temperature, humidity and CO2 are controlled by the
incubator. An actuation system and a culture chamber are the key components of the
bioreactor system, as they provide the mechanical output and the sterilized culture
environment, respectively. A step motor-ball screw transmission system (SMBS) was
adopted as the actuator in our bioreactor. Compared to the other actuator systems such
as the pneumatic actuators(Juncosa-Melvin, Shearn et al. 2006; Nirmalanandhan, Rao et
al. 2008; Nirmalanandhan, Shearn et al. 2008) and linear motors(Nguyen, Liang et al.
2009; Doroski, Levenston et al. 2010), SMBS has the advantages of high resolution,
highloading capacity and good resistance to humid and warm environment(Liu, Hsu et
al. 2001; Altman, Lu et al. 2002; Webb, Hitchcock et al. 2006; Androjna, Spragg et al.

183
2007). AISI 316 stainless steel, which has good machinability and resistance to
chemicals and high temperature, was used for the construction of the culture chamber.
The labyrinth channel integrated in the chamber ensured sufficient air exchange without
inducing contamination, and the glass covers allowed for visual monitoring. Several
commercial bioreactor systems have been developed such as The Bose® ElectroForce
BioDynamic® system and the LigaGen system. Compared to these products, our system
has an advantage in that it provides two methods for tissue fixation, tissue clamp and
tissue hook. Tissue clamps have been found to be more effective at stabilizing
membrane-like tissue, while tissue hooks are designed for fixing thick and
irregular-shaped tissue.
In the present studies, full-length rabbit Achilles tendons, including the bone-tendon and
tendon-muscle junction, were used as an Achilles tendon tissue model and cultured in
the bioreactor system. Tissue hooks were found to be able to provide excellent fixation
without slippage, which was essential for this study. Slippage can cause inconsistent
mechanical loading and can comprise the results.
6.1.2 Mechanical stimulation in bioreactor system (Chapter 4 and 5)
The present studies utilized the novel approach of applying mechanical stimulation on
fresh-harvest rabbit Achilles tendon in a bioreactor system. The application of the
bioreactor in evaluating the effect of mechanical stimulation on tendon has several
advantages:

184
Unlike the monolayer cellular study, the tenocytes were in their native niche
embedded in the collagen matrix. During mechanical stimulation, tenocytes were
not only subjected to tensile strain, but also compression from adjacent collagen
fibers (Caliari, Weisgerber et al. 2012). Therefore, mechanical stimulation applied
by the bioreactor system can be better replicated in in vivo loading environments
compared to a monolayer system.
Unlike animal studies such as treadmill training, the bioreactor system used in this
study is able to provide accurate, programmable and reproducible mechanical
stimulation, allowing the systematic study of different intensity loading on the
tendon.
The use of a bioreactor system allows the chemical environment in the culture to be
manipulated. Therefore, it offers the potential for systematically studying the effects
of nutrients, growth factors and cytokines under different loading regimes.
6.1.3 Effect of various mechanical stimulation on tendon homeostasis (Chapter 4)
Full-length rabbit Achilles tendons were dissected and cultured under different
mechanical environments: 0% 3% 6% and 9% strain at 0.25Hz 8h/day for 6 days.
Histology, immunohistochemistry, TUNEL assay and RT-PCR were carried out to
evaluate tissue integrity.
Histologically, tendons subjected to 6% strain mechanical loading exhibited similar
morphology as native tendon. However, loading deprivation group, 3% strain and 9%

185
strain groups showed pathological changes.
Loading deprivation
Tendon exhibited minor pathological changes including loose collagen fiber and a
moderately increased cell apoptosis rate after 6 days of loading deprivation culture
(Figure 4.2 B). The morphology further degenerated when the culture period was
extended to 2 weeks. Typical tendinopathy morphologies were observed such as
massive cellular apoptosis, disorientated collagen fiber and rounded tenocyte nuclei
(Figure 4.2C, 5.4C). The severe pathological changes are initiated by high cell
death. The remaining cells then fail to maintain the tissue integrity.
3% strain
In the 3% strain group, wavy collagen fiber, minor rounded cell nuclei, loose ECM
structure and increased collagen type III synthesis were observed (Figure 4.3B,
5.5C). The elevated MMPs expression and downregulation of COL1A2 are
consistent with these pathological changes. In 2007, Arnoczky et al. hypothesized
that pathological development of tendinopathy maybe the result of under
stimulation rather than overloading(Arnoczky, Lavagnino et al. 2007). Our study
provided further evidence in support of this theory. It is suggested in this thesis that
increase protease activity acting on underloaded collagen and reduced anabolic
activity of tenocytes due to reduced mechanical stimulus, despite a reduced
mechanical damage, can explain the observed pathological changes in this 3%
strain group.

186
6% strain
In the 6% loading group, tendons were able to maintain their normal morphology,
type III collagen content and apoptosis rate. RT-PCT results showed an increased
expression of COL1A2. The rate of micro-damage caused by mechanical loading is
matched by the rate of repair such that the structural integrity was able to be
maintained.
9% strain
In this group, an acute injury model was successfully established. Tendon subjected
to excessive loading caused rupture of collagen fiber (i.e. mechanical damage) and
increased cell apoptosis close to the injury site, which are the typical morphology of
acute injury on tendon. Type III collagen was highly expressed in RT-PCR results
and was confirmed by immunohistochemistry.
Although pathological changes were detected in three of four loading conditions, the
underlying causes are different. From the different loading groups and the pathological
changes and protein expressions, we proposed a hypothetical model for tendon damage
and repair, as shown in Figure 4.7. An increased intensity of mechanical stimulation
increases the mechanical damage. Protease (chemical) damage was active in the low
strain groups. Only at intermediate strains of 6% was the repair able to match the rate of
mechanical and proteolytic damage, and hence the ability to maintain structural integrity.

187
Therefore, the conclusion drawn from this study is that a narrow range of mechanical
stimulation in this bioreactor system was able to maintain tendon homeostasis; with
mechanical underloading and overloading both detrimental to tendon.
6.1.4 Rescue of degenerated tendon with mechanical stimulation.
In the previous study, we demonstrated that 6% strain mechanical stimulation was able
to maintain the tendon homeostasis in the bioreactor system. Following this, the next
hypothesis was whether or not 6% strain can also rescue tendon following load
deprivation. Therefore, in this study, rabbit Achilles tendon was first cultured without
loading for 6 days, and then followed by either an additional 6 days without load or 6
days at 6% strain at 0.25Hz 8hr/day for. The tendons then were analyzed by
biomechanical testing and histological assessment.
Compared to native rabbit Achilles tendons, tendons under static culture for 6 days and
12 days showed 30% and 50% reduction in their mechanical properties, respectively.
However, the rescue group showed significant improvement compared to these static
culture groups. Indeed tendons in the rescue group exhibited similar morphology,
apoptosis rate and collagen type III content to native tendons. RT-PCR results showed,
application of 6% strain mechanical stimulation significantly reduced MMPs expression
and increases COL1A2 and TIMPs expression, which was consistent with a suppression
of tissue catabolic processes and an increase in those associated with tissue repair.

188
Immobilization is a common postoperative treatment of bone fracture and tendon
injuries(Ruedi and Allgower 1979; Cetti, Henriksen et al. 1994). In various studies,
early-stage degradation in Achilles tendon was reported after long-term immobilization
in both humans and rats(Larsen, Forwood et al. 1987; Nakagawa, Totsuka et al. 1989;
Kannus, Jozsa et al. 1997; Trudel, Koike et al. 2007). However, there is no clinical
rehabilitation guideline for Achilles tendon weakness after a period of immobilization.
The results of this study suggests that proper mechanical loading may activate a
self-repair mechanism to repair early stage degenerative pathological changes in tendon.
6.2 Limitation of this work
It is important to mention several limitations of the work described in this PhD thesis.
6.2.1 Chapter 3
In chapter 3, the bioreactor designed is only able to provide some basic function; there
are still several limitations in this design, with the benefit of hindsight could be
improved.
1. Failure warning: although the bioreactor system can provide preprogrammed
automatic mechanical stimulation on the tissue sample, some accidents like power
failure and maloperation might still happen. A complete computer monitoring
system would be ideal, but even a basic warning light should be installed and
preprogrammed in the system to report the error.

189
2. Program interface: The operational system was programed using the basic PLC
language, which might not be familiar to the other users. A more user-friendly
‘common’ programming language should be designed for the convenience of the
other users.
3. Bioreactor connection: in the present design, the bioreactor is connected to the
control box through a cable, which required specific incubator model with a cable
hole back panel. However, a cable hole is not a standard configuration in all
incubators; therefore, this bioreactor can only be used in some special incubator.
6.2.2 Chapter4
1. Mechanical stimulation accuracy: Achilles tendons from different individuals
show variations in length. Although the tissue hook of the bioreactor can provide
an effective way to stabilize the tendon and the stimulation distant are fixed at
23mm, some error might still be induced.
2. Medium infiltration: During the in vitro culture, nutrient delivery is depended on
medium infiltration. In loading deprivation group, medium infiltration might not
sufficient for the whole tendon. Although the histological section exhibited
consistent morphology in the center and the edge of the tendon, the nutrient
transport still needs to be considered when interpreting the results.
3. Statistical analyze: In the histological scoring, scores should have been analyzed
using a nonparametric statistical procedure as the data is ordinal, not continuous.

190
6.2.3 Chapter 5
Biomechanical testing: Achilles tendons from different individuals show
variations in initial mechanical properties. Although in this experiment, the
Achilles tendon from same individual we distributed in different groups to
minimize error, it is still needed to be mentioned; especially as the sample size was
relatively small.
6.3 Future directions
6.3.1 Gene profile of tenocyte subjected to 2D or 3D mechanical stimulation
As demonstrated by the results in this thesis, tendons are able to sense and respond to
mechanical loading. Although they are mainly subjected to tensile strain on the
macroscale (i.e. the whole tissue scale), the micromechanical environment is much more
complicated. Tendon is a dense connective tissue that comprised by well-organized
collagen fiber with tenocyte adhered on it. Subjected to overall tensile strain, tenocytes
not only receive tensile but also the shear force and compression from the surrounding
collagen fibers and proteoglycans as well as potential fluid shear stress (Figure 7.1)
(Khan and Scott 2009). However, the in vitro studies that evaluate the effect of
mechanical stimulation on tenocyte or fibroblast mostly focus on tensile strain only. The
Flexcell® system was commonly used in the studies of in vitro gene expression profiles
of tenocytes under mechanical stimulation (Skutek, van Griensven et al. 2001; Wang,

191
Jia et al. 2003; Yang, Im et al. 2005). These systems can precisely control the
mechanical regimen and monitor the cell behaviors, but only can be used for 2D
monolayer cell culture, not the organized tissue. Cells isolated from their extracellular
niche and cultured in a monolayer may not necessarily replicate their original behavior
in vivo. The recent study of Ni et al. successfully generated a scaffold-free engineered
tendon from tendon-derived stem cell by stimulation with connective tissue growth
factor (Ni, Rui et al. 2013). This model allows the better study on the effect of
mechanical stimulation on tenocyte. As the structure of scaffold-free tendon is more
similar to native tendon, when compared to monolayer cell cultures, the in vivo
mechanical stimulation can be better mimicked in our bioreactor system using this
scaffold-free tendon model. The gene profile of tenocyte subjected to 2D and 3D
mechanical stimulation can lead to the better understanding how tenocyte respond to in
vivo mechanical stimulation.
Figure 7. 1 Tenocyte undergoing (A,B) shear and (C) compression during tendon subjected to tensile strain. Figures taken from Khan et al.(Khan and Scott 2009)

192
6.3.2 The effect of mechanical stimulation on tendon nutrient infiltration
Nutrient supply during in vitro culture of tendon mainly depends on passive infiltration.
However, during mechanical stimulation, advective nutrient transport would be induced.
Although Hannafin et al. reported that mechanical stimulation did not alter small
nutrient uptake, compared to static culture, and that the tendon repair improvements
derived from passive motion is not related to an increase of nutrient transport (Hannafin
and Arnoczky 1994), under different loading regimes, or in different tendon types, this
may not be the case. Moreover, the transport of large molecules, such as growth factors
and matrix components, may be affected by mechanical stimulation, as has been shown
in studies in cartilage (Bonassar, Grodzinsky et al. 2000; Zhang, Gardiner et al. 2007).
Although unlike cartilage, the in vivo nutrient supply is dependent on limited blood
vessels instead of synovial fluid, the nutrient infiltration in vitro is essentially the same
as cartilage, especially for large scale animal engineered tendon generation. Therefore,
rabbit Achilles tendon culture in a bioreactor system can be used as a nutrient transport
model for study. Different sizes of dye that are compatible to target cytokines can be
used in the bioreactor system (Day, Megson et al. 2005).Nutrient delivery could then be
assessed based on the dye infiltration. Furthermore, nutrient delivery is often a key
obstacle in tissue engineering. If the nutrient delivery induced by tensile cyclic strain in
the bioreactor is not sufficient, medium circulation system could be designed based on
the current model of bioreactor to enhance nutrient infiltration.

193
6.3.3 The effect of mechanical stimulation on stem cell tenogenic differentiation
Stem cells have been suggested to have great potential in regenerative medicine
application due to its self-renewal capacity, hypoimmunogenicity and multi-lineage
differentiation ability(Uccelli, Moretta et al. 2006). However, the complexity and
reliability of manipulating stem cell to differentiate into a target cell type limits the
clinical application. In vitro induction of stem cell differentiation is commonly achieved
by chemical modulations. Recent studies suggested that mechanical stimulation acted as
an important regulator in stem cell differentiation induced by cytokines (Rui, Lui et al.
2011), and might be essential for lineage selection and cell differentiation. Khayat et al.
suggested low frequency mechanical stimulation inhibited MSC adipogenic
differentiation (Khayat, Rosenzweig et al. 2012), and a similar result was reported by
Sen et al.(Sen, Xie et al. 2008). Continuous cyclic mechanical tension has been
indicated to inhibit osteogenic differentiation of MSC (Shi, Li et al. 2011), whilst short
term cyclic tensile strain has been reported to stimulate osteogenic differentiation (Rui,
Lui et al. 2011). Furthermore, dynamic compression can induce the chondrogenic
differentiation of MSC (Huang, Farrell et al. 2010; Kupcsik, Stoddart et al. 2010). The
study of Chen et al. reported that static mechanical stimulation was able to stimulate
tenogenic differentiation of human embryonic stem cells (Chen, Song et al. 2009).
Although the mechanical stimulation has been suggested as a key regulator of stem cell
differentiates into musculoskeletal cell type, tenogenic differentiation induced by
mechanical loading is relatively rarely recorded. As a mechano-transduction tissue, it is
reasonable to speculate that dynamic tensile strain is able to simulate the tenogenic
differentiation of stem cells, although the magnitude, frequency and duration remain
unclear. The present study showed that 6% strain, 0.25Hz for 8h/day was able to

194
increase collagen type I expression of dissected rabbit Achilles tendon, which was one
of the tenogenic market of tenocytes. Therefore, the mechanical stimulation regime used
in this study should be tested for tenogenic differentiation of stem cells.
6.3.4 Therapeutic guideline
One commonly used conservative treatment for chronic Achilles tendinopathy is
eccentric overload. The basic principle is to maximize the activation of the calf or
soleus muscle using body weight, so generate a tensile strain on Achilles tendon
(Alfredson, Pietila et al. 1998). The treatment is instructed to be continued unless the
patient experience disabling pain, however, if the patient can perform the exercise
without experiencing any pain, extra weight is required(Alfredson, Pietila et al. 1998).
The treatment is normally performed 2 times daily, 7 days/week for 12 weeks, and for
each treatment including 3 sets of loading, 15 repetitions each set(Alfredson, Pietila et
al. 1998). Several studies reported positive effect on symptoms relief(Stanish,
Rubinovich et al. 1986; Alfredson, Pietila et al. 1998; Silbernagel, Thomee et al. 2001;
Alfredson and Lorentzon 2003; Fahlstrom, Jonsson et al. 2003; Roos, Engstrom et al.
2004; Shalabi, Kristoffersen-Wilberg et al. 2004) however, the study of Woodley et al.
showed no effect(Woodley, Newsham-West et al. 2007) and Rompe et al. reported an
inferior results(Rompe, Furia et al. 2008). The present study indicated that there was
only a narrow range of tensile strain stimulation that could achieve a net anabolic or
even therapeutic effect, which was 6% strain in the rabbit Achilles tendon model.
Overloading and underloading would trigger degenerative cascade through different
pathways. This study provides a cue to explain the clinical outcome variety from the

195
eccentric overloading training. As the treatment is based on the sense of patients, the
strain applied on Achilles tendon may not be sufficient to achieve the optimal clinical
outcome; moreover, the treatment is not consistence with different patients. Although
rabbit tissue is different from human tissue, similar principles can be applied in human.
Physiological soleus and calf muscle force during gait, running and jumping can be
measured by the inverse dynamics algorithm, and then the physiological strain on
Achilles tendon can be calculated. Base on the physiological strain range of Achilles
tendon, more experiments are required to evaluate the optimal loading regime for the
rehabilitation program. However, even knowing the optimal loading strain on Achilles
tendon, the monitoring of tendon strain is still a big challenge, which requires various
expertise from sport science.
6.3.5 Robot- driven orthosis
Immobilization is a standard postoperative procedure after tendon surgery to eliminate
the loading on the surgical site then prevent the rerupture during recovery period;
however, several studies have reported that the immobilization clinically caused
degenerative changes in tendon tissue (Amiel, Woo et al. 1982; Loitz, Zernicke et al.
1989). Recent study reported that short-episode of mechanical stimulation at the
inflammatory stage could improve healing with histological signs of repeated damage in
the form of increased bleeding (Enwemeka, Spielholz et al. 1988; Eliasson, Andersson
et al. 2011). Therefore, early intervention seems to be beneficial for the tendon healing;
however, self-initiative training might be difficult to control and might damage the
surgical site. Robot-driven orthosis, in the other hand, can control the activity level and

196
protect the surgical site from overloading. Although no robot-driven orthosis is currently
being used for tendon rehabilitation, a clinical trial on patient suffered from multiple
sclerosis and stroke seems to be promising (Husemann, Muller et al. 2007; Lo and
Triche 2008; Schwartz, Sajin et al. 2012; Straudi, Benedetti et al. 2013). Our studies
indicate that proper mechanical stimulation has therapeutic effect; therefore,
robot-driven orthosis could be developed based on this model as a postoperative
rehabilitation. The combination of surgical treatment and conservative treatment at the
early inflammatory phase might provide a better clinical outcome.
6.3.6 Tendon engineering
Tendon’s primary function is force transmission. It operates in a varying load
environment, both on short timescales (e.g. walking, running) and on longer timescales
(e.g. changes in body size with age). In fact, it has been shown that after 4 weeks in a
load-free culture environment tenocytes lost their native elongated morphology, became
increasingly rounded, and the collagen fiber became more crimped (Hannafin, Arnoczky
et al. 1995). Unsurprisingly, given its functional role, a suitable mechanical stimulus is
vital for engineered tendon/ligament formation. Cyclic stretching has been shown to
produce an up to 9 folds increase in the cell number of an engineered tendon compared
with a static culture over a 2 week period (Abousleiman, Reyes et al. 2009).
Furthermore, mechanical stimulation could help guide collagen fiber formation, i.e.
along the direction of loading, which is able to enhance or optimize the mechanical
properties including stiffness, elastic modulus, maximum tensile stress and maximum
force(Juncosa-Melvin, Shearn et al. 2006; Saber, Zhang et al. 2010; Woon, Kraus et al.

197
2011). However, most mechanical stimulation applied on the engineered tendon
cultured is lack of systemic optimization, the magnitude and frequency varied from
studies. Some studies used up to 10% strain for tendon stimulation, which might excess
the tolerance of native tendon (our study indicated 9% strain could cause partial tear of
rabbit Achilles tendon). In the present study, rabbit Achilles tendon was used as a model
to evaluation the effect of different mechanical stimulation on tendon homeostasis.
Knowing the fact that 6% strain was able to create anabolic effect and increased tendon
formation, the further application of this study is to combine the optimal mechanical
stimulation and the engineered tendon culture to create a better tendon construct for
implantation.
6.4 Conclusion
In the present PhD project, a functional bioreactor that can provide programmable
mechanical stimulation and sterilized culture environment was designed and
constructed. The application of this bioreactor allowed the evaluation on the effect
of different mechanical stimulation on rabbit Achilles tendon homeostasis. It was
observed that only a narrow range of mechanical stimulation can maintain the
tendon integrity, mechanical underloading and overloading were both detrimental
to tendon tissue. It was observed that successful restoration of the tendon’s
structural integrity and mechanical properties was available by the application of 6%
tensile cyclic loading from early degenerative load deprivation model. The current
results suggest that mechanical stimulation may provide therapeutic benefits.

198
REFERENCE
Abousleiman, R. I., Y. Reyes, et al. (2009). "Tendon tissue engineering using cell-seeded umbilical veins cultured in a mechanical stimulator." Tissue Eng Part A 15(4): 787-795.
Abrahamsson, S. O. (1991). "Matrix metabolism and healing in the flexor tendon. Experimental studies on rabbit tendon." Scand J Plast Reconstr Surg Hand Surg Suppl 23: 1-51.
Abrahamsson, S. O. and R. Gelberman (1994). "Maintenance of the gliding surface of tendon autografts in dogs." Acta Orthop Scand 65(5): 548-552.
Abrahamsson, S. O., G. Lundborg, et al. (1991). "Long-term explant culture of rabbit flexor tendon: effects of recombinant human insulin-like growth factor-I and serum on matrix metabolism." J Orthop Res 9(4): 503-515.
Alfredson, H. and R. Lorentzon (2000). "Chronic Achilles tendinosis: recommendations for treatment and prevention." Sports Med 29(2): 135-146.
Alfredson, H. and R. Lorentzon (2003). "Intratendinous glutamate levels and eccentric training in chronic Achilles tendinosis: a prospective study using microdialysis technique." Knee Surg Sports Traumatol Arthrosc 11(3): 196-199.
Alfredson, H., T. Pietila, et al. (1998). "Heavy-load eccentric calf muscle training for the treatment of chronic Achilles tendinosis." Am J Sports Med 26(3): 360-366.
Alter, D. and T. Tsao (1994). Dynamic stiffness enhancement of direct linear motor feed drives for machining. Proceedings of the American Control Conference.
Alter, D. and T. Tsao (1996). "Control of linear motors for machine tool feed drives: design and implementation of H1 optimal feedback control." ASME J Dyn Syst, Meas, Control 118: 649-656.
Altman, G. H., H. H. Lu, et al. (2002). "Advanced bioreactor with controlled application of multi-dimensional strain for tissue engineering." J Biomech Eng 124(6): 742-749.
Amiel, D., S. L. Woo, et al. (1982). "The effect of immobilization on collagen turnover in connective tissue: a biochemical-biomechanical correlation." Acta Orthop Scand 53(3): 325-332.
Androjna, C., R. K. Spragg, et al. (2007). "Mechanical conditioning of cell-seeded small intestine submucosa: a potential tissue-engineering strategy for tendon repair." Tissue Eng 13(2): 233-243.
Anitua, E., M. Sanchez, et al. (2006). "New insights into and novel applications for platelet-rich fibrin therapies." Trends Biotechnol 24(5): 227-234.

199
Arancia, G., P. Crateri Trovalusci, et al. (1989). "Ultrastructural changes induced by hyperthermia in Chinese hamster V79 fibroblasts." Int J Hyperthermia 5(3): 341-350.
Archambault, J., M. Tsuzaki, et al. (2002). "Stretch and interleukin-1beta induce matrix metalloproteinases in rabbit tendon cells in vitro." J Orthop Res 20(1): 36-39.
Arnoczky, S. P., M. Lavagnino, et al. (2007). "The mechanobiological aetiopathogenesis of tendinopathy: is it the over-stimulation or the under-stimulation of tendon cells?" Int J Exp Pathol 88(4): 217-226.
Arnoczky, S. P., M. Lavagnino, et al. (2008). "Loss of homeostatic strain alters mechanostat "set point" of tendon cells in vitro." Clin Orthop Relat Res 466(7): 1583-1591.
Arnoczky, S. P., T. Tian, et al. (2004). "Ex vivo static tensile loading inhibits MMP-1 expression in rat tail tendon cells through a cytoskeletally based mechanotransduction mechanism." J Orthop Res 22(2): 328-333.
Arnoczky, S. P., T. Tian, et al. (2002). "Activation of stress-activated protein kinases (SAPK) in tendon cells following cyclic strain: the effects of strain frequency, strain magnitude, and cytosolic calcium." J Orthop Res 20(5): 947-952.
Arnsdorf, E. J., L. M. Jones, et al. (2009). "The periosteum as a cellular source for functional tissue engineering." Tissue Eng Part A 15(9): 2637-2642.
Arora, N. S., T. Ramanayake, et al. (2009). "Platelet-rich plasma: a literature review." Implant Dent 18(4): 303-310.
Asundi, K. R., K. B. King, et al. (2008). "Evaluation of gene expression through qRT-PCR in cyclically loaded tendons: an in vivo model." Eur J Appl Physiol 102(3): 265-270.
Awad, H. A., D. L. Butler, et al. (1999). "Autologous mesenchymal stem cell-mediated repair of tendon." Tissue Eng 5(3): 267-277.
Awad, H. A., D. L. Butler, et al. (2000). "In vitro characterization of mesenchymal stem cell-seeded collagen scaffolds for tendon repair: effects of initial seeding density on contraction kinetics." J Biomed Mater Res 51(2): 233-240.
Baysal, A., S. Bilsel, et al. (2006). "Comparison of the usage of intravenous iloprost and nitroglycerin for pulmonary hypertension during valvular heart surgery." Heart Surg Forum 9(1): E536-542.
Benjamin, M., B. Moriggl, et al. (2004). "The "enthesis organ" concept: why enthesopathies may not present as focal insertional disorders." Arthritis Rheum 50(10): 3306-3313.
Benjamin, M. and J. R. Ralphs (1998). "Fibrocartilage in tendons and ligaments--an

200
adaptation to compressive load." J Anat 193 ( Pt 4): 481-494.
Bennett, N. T. and G. S. Schultz (1993). "Growth factors and wound healing: biochemical properties of growth factors and their receptors." Am J Surg 165(6): 728-737.
Berse, B., J. A. Hunt, et al. (1999). "Hypoxia augments cytokine (transforming growth factor-beta (TGF-beta) and IL-1)-induced vascular endothelial growth factor secretion by human synovial fibroblasts." Clin Exp Immunol 115(1): 176-182.
Beskin, J. L., R. A. Sanders, et al. (1987). "Surgical repair of Achilles tendon ruptures." Am J Sports Med 15(1): 1-8.
Bi, Y., D. Ehirchiou, et al. (2007). "Identification of tendon stem/progenitor cells and the role of the extracellular matrix in their niche." Nat Med 13(10): 1219-1227.
Birch, H. L. (2007). "Tendon matrix composition and turnover in relation to functional requirements." Int J Exp Pathol 88(4): 241-248.
Birch, H. L., G. A. Rutter, et al. (1997). "Oxidative energy metabolism in equine tendon cells." Research in Veterinary Science 62(2): 93-97.
Birch, H. L., A. M. Wilson, et al. (1997). "The effect of exercise-induced localised hyperthermia on tendon cell survival." J Exp Biol 200(Pt 11): 1703-1708.
Bjordal, J. M., R. A. Lopes-Martins, et al. (2006). "A randomised, placebo controlled trial of low level laser therapy for activated Achilles tendinitis with microdialysis measurement of peritendinous prostaglandin E2 concentrations." Br J Sports Med 40(1): 76-80; discussion 76-80.
Blair, B., A. S. Rokito, et al. (1996). "Efficacy of injections of corticosteroids for subacromial impingement syndrome." J Bone Joint Surg Am 78(11): 1685-1689.
Bomler, J. and J. Sturup (1989). "Achilles tendon rupture. An 8-year follow up." Acta Orthop Belg 55(3): 307-310.
Bonassar, L. J., A. J. Grodzinsky, et al. (2000). "Mechanical and physicochemical regulation of the action of insulin-like growth factor-I on articular cartilage." Arch Biochem Biophys 379(1): 57-63.
Bradley, J. P. and J. E. Tibone (1990). "Percutaneous and open surgical repairs of Achilles tendon ruptures. A comparative study." Am J Sports Med 18(2): 188-195.
Braembussche, P., J. Swevers, et al. (1996). "Accurate tracking control of linear synchronous motor machine tool axes." Mechatronics 6(5): 507-521.
Bramono, D. S., J. C. Richmond, et al. (2004). "Matrix metalloproteinases and their clinical applications in orthopaedics." Clin Orthop Relat Res(428): 272-285.

201
Bray, R. C., R. M. Rangayyan, et al. (1996). "Normal and healing ligament vascularity: a quantitative histological assessment in the adult rabbit medial collateral ligament." J Anat 188 ( Pt 1): 87-95.
Bressel, E. and P. J. McNair (2001). "Biomechanical behavior of the plantar flexor muscle-tendon unit after an Achilles tendon rupture." Am J Sports Med 29(3): 321-326.
Brown, C. H., Jr. and E. W. Carson (1999). Revision anterior cruciate ligament surgery.
Buckwalter, J. A. and E. B. Hunziker (1996). "Orthopaedics. Healing of bones, cartilages, tendons, and ligaments: a new era." Lancet 348 Suppl 2: sII18.
Bugg, E. I., Jr. and B. M. Boyd (1968). "Repair of neglected rupture or laceration of the Achilles tendon." Clin Orthop Relat Res 56: 73-75.
Burkhardt, A., J. Walther, et al. (2012). "Endoscopic optical coherence tomography device for forward imaging with broad field of view." J Biomed Opt 17(7): 071302.
Butler, D. L., C. Gooch, et al. (2010). "The use of mesenchymal stem cells in collagen-based scaffolds for tissue-engineered repair of tendons." Nat Protoc 5(5): 849-863.
Butler, D. L., S. A. Hunter, et al. (2009). "Using functional tissue engineering and bioreactors to mechanically stimulate tissue-engineered constructs." Tissue Eng Part A 15(4): 741-749.
Butler, D. L., N. Juncosa-Melvin, et al. (2008). "Functional tissue engineering for tendon repair: A multidisciplinary strategy using mesenchymal stem cells, bioscaffolds, and mechanical stimulation." J Orthop Res 26(1): 1-9.
Caliari, S. R., D. W. Weisgerber, et al. (2012). "The influence of collagen-glycosaminoglycan scaffold relative density and microstructural anisotropy on tenocyte bioactivity and transcriptomic stability." J Mech Behav Biomed Mater 11: 27-40.
Calve, S., I. F. Lytle, et al. (2010). "Implantation increases tensile strength and collagen content of self-assembled tendon constructs." J Appl Physiol 108(4): 875-881.
Carr, A. J. and S. H. Norris (1989). "The blood supply of the calcaneal tendon." J Bone Joint Surg Br 71(1): 100-101.
Cartmell, S. H., B. D. Porter, et al. (2003). "Effects of medium perfusion rate on cell-seeded three-dimensional bone constructs in vitro." Tissue Eng 9(6): 1197-1203.
Catapano, G., J. F. Patzer, 2nd, et al. (2010). "Transport advances in disposable bioreactors for liver tissue engineering." Adv Biochem Eng Biotechnol 115:

202
117-143.
Cerullo, G., G. Puddu, et al. (1995). "Anterior cruciate ligament patellar tendon reconstruction: it is probably better to leave the tendon defect open!" Knee Surg Sports Traumatol Arthrosc 3(1): 14-17.
Cetti, R. and S. E. Christensen (1983). "Surgical treatment under local anesthesia of Achilles tendon rupture." Clin Orthop Relat Res(173): 204-208.
Cetti, R., L. O. Henriksen, et al. (1994). "A new treatment of ruptured Achilles tendons. A prospective randomized study." Clin Orthop Relat Res(308): 155-165.
Chamberlain, C. S., E. Crowley, et al. (2009). "The spatio-temporal dynamics of ligament healing." Wound Repair Regen 17(2): 206-215.
Chan, B. P., K. M. Chan, et al. (1997). "Effect of basic fibroblast growth factor. An in vitro study of tendon healing." Clin Orthop Relat Res(342): 239-247.
Chan, B. P., S. Fu, et al. (2000). "Effects of basic fibroblast growth factor (bFGF) on early stages of tendon healing: a rat patellar tendon model." Acta Orthop Scand 71(5): 513-518.
Chan, B. P., S. C. Fu, et al. (2006). "Supplementation-time dependence of growth factors in promoting tendon healing." Clin Orthop Relat Res 448: 240-247.
Chan, K. M., S. C. Fu, et al. (2008). "Expression of transforming growth factor beta isoforms and their roles in tendon healing." Wound Repair Regen 16(3): 399-407.
Chang, J., D. Most, et al. (1997). "Gene expression of transforming growth factor beta-1 in rabbit zone II flexor tendon wound healing: evidence for dual mechanisms of repair." Plast Reconstr Surg 100(4): 937-944.
Chang, J., R. Thunder, et al. (2000). "Studies in flexor tendon wound healing: neutralizing antibody to TGF-beta1 increases postoperative range of motion." Plast Reconstr Surg 105(1): 148-155.
Chard, M. D., L. M. Sattelle, et al. (1988). "The long-term outcome of rotator cuff tendinitis--a review study." Br J Rheumatol 27(5): 385-389.
Chen, J., A. Wang, et al. (2010). "In chronic lateral epicondylitis, apoptosis and autophagic cell death occur in the extensor carpi radialis brevis tendon." J Shoulder Elbow Surg 19(3): 355-362.
Chen, J., J. Xu, et al. (2009). "Scaffolds for tendon and ligament repair: review of the efficacy of commercial products." Expert Rev Med Devices 6(1): 61-73.
Chen, J., Q. Yu, et al. (2011). "Autologous tenocyte therapy for experimental Achilles tendinopathy in a rabbit model." Tissue Eng Part A 17(15-16): 2037-2048.

203
Chen, J. L., Z. Yin, et al. (2010). "Efficacy of hESC-MSCs in knitted silk-collagen scaffold for tendon tissue engineering and their roles." Biomaterials 31(36): 9438-9451.
Chen, J. M., C. Willers, et al. (2007). "Autologous tenocyte therapy using porcine-derived bioscaffolds for massive rotator cuff defect in rabbits." Tissue Eng 13(7): 1479-1491.
Chen, X., Y. Y. Qi, et al. (2008). "Ligament regeneration using a knitted silk scaffold combined with collagen matrix." Biomaterials 29(27): 3683-3692.
Chen, X., X. H. Song, et al. (2009). "Stepwise differentiation of human embryonic stem cells promotes tendon regeneration by secreting fetal tendon matrix and differentiation factors." Stem Cells 27(6): 1276-1287.
Chen, Y. E. (2004). "MMP-12, an old enzyme plays a new role in the pathogenesis of rheumatoid arthritis?" Am J Pathol 165(4): 1069-1070.
Chhabra, A., D. Tsou, et al. (2003). "GDF-5 deficiency in mice delays Achilles tendon healing." J Orthop Res 21(5): 826-835.
Choi, H. R., S. Kondo, et al. (2002). "Expression and enzymatic activity of MMP-2 during healing process of the acute supraspinatus tendon tear in rabbits." J Orthop Res 20(5): 927-933.
Chong, A. K., A. D. Ang, et al. (2007). "Bone marrow-derived mesenchymal stem cells influence early tendon-healing in a rabbit achilles tendon model." J Bone Joint Surg Am 89(1): 74-81.
Christensen, I. (1953). "Rupture of the Achilles tendon; analysis of 57 cases." Acta Chir Scand 106(1): 50-60.
Clark, R. T., T. L. Johnson, et al. (2001). "GDF-5 deficiency in mice leads to disruption of tail tendon form and function." Connect Tissue Res 42(3): 175-186.
Connell, D., A. Datir, et al. (2009). "Treatment of lateral epicondylitis using skin-derived tenocyte-like cells." Br J Sports Med 43(4): 293-298.
Cook, J. L. and C. Purdam (2012). "Is compressive load a factor in the development of tendinopathy?" Br J Sports Med 46(3): 163-168.
Coombes, B. K., L. Bisset, et al. (2010). "Efficacy and safety of corticosteroid injections and other injections for management of tendinopathy: a systematic review of randomised controlled trials." Lancet 376(9754): 1751-1767.
Cooper, J. A., H. H. Lu, et al. (2005). "Fiber-based tissue-engineered scaffold for ligament replacement: design considerations and in vitro evaluation." Biomaterials 26(13): 1523-1532.

204
Coupens, S. D., C. K. Yates, et al. (1992). "Magnetic resonance imaging evaluation of the patellar tendon after use of its central one-third for anterior cruciate ligament reconstruction." Am J Sports Med 20(3): 332-335.
Cretnik, A., M. Kosanovic, et al. (2005). "Percutaneous versus open repair of the ruptured Achilles tendon: a comparative study." Am J Sports Med 33(9): 1369-1379.
Cribb, A. M. and J. E. Scott (1995). "Tendon response to tensile stress: an ultrastructural investigation of collagen:proteoglycan interactions in stressed tendon." J Anat 187 ( Pt 2): 423-428.
Dahlgren, L. A., H. O. Mohammed, et al. (2005). "Temporal expression of growth factors and matrix molecules in healing tendon lesions." J Orthop Res 23(1): 84-92.
Dahlgren, L. A., A. J. Nixon, et al. (2001). "Effects of beta-aminopropionitrile on equine tendon metabolism in vitro and on effects of insulin-like growth factor-I on matrix production by equine tenocytes." Am J Vet Res 62(10): 1557-1562.
Dahlgren, L. A., M. C. van der Meulen, et al. (2002). "Insulin-like growth factor-I improves cellular and molecular aspects of healing in a collagenase-induced model of flexor tendinitis." J Orthop Res 20(5): 910-919.
Darlington, L. G. and E. N. Coomes (1977). "The effects of local steroid injection for supraspinatus tears." Rheumatol Rehabil 16(3): 172-179.
Day, R. E., S. Megson, et al. (2005). "Iontophoresis as a means of delivering antibiotics into allograft bone." J Bone Joint Surg Br 87(11): 1568-1574.
De Bari, C., F. Dell'Accio, et al. (2001). "Multipotent mesenchymal stem cells from adult human synovial membrane." Arthritis Rheum 44(8): 1928-1942.
de Vos, R. J., A. Weir, et al. (2010). "Platelet-rich plasma injection for chronic Achilles tendinopathy: a randomized controlled trial." JAMA 303(2): 144-149.
Deng, D., W. Liu, et al. (2009). "Engineering human neo-tendon tissue in vitro with human dermal fibroblasts under static mechanical strain." Biomaterials 30(35): 6724-6730.
Dennis, R. G., B. Smith, et al. (2009). "Bioreactors for guiding muscle tissue growth and development." Adv Biochem Eng Biotechnol 112: 39-79.
Derwin, K. A., A. R. Baker, et al. (2006). "Commercial extracellular matrix scaffolds for rotator cuff tendon repair. Biomechanical, biochemical, and cellular properties." J Bone Joint Surg Am 88(12): 2665-2672.
Domm, C., M. Schunke, et al. (2002). "Redifferentiation of dedifferentiated bovine articular chondrocytes in alginate culture under low oxygen tension."

205
Osteoarthritis Cartilage 10(1): 13-22.
Donnelly, K., A. Khodabukus, et al. (2010). "A novel bioreactor for stimulating skeletal muscle in vitro." Tissue Eng Part C Methods 16(4): 711-718.
Doroski, D. M., M. E. Levenston, et al. (2010). "Cyclic tensile culture promotes fibroblastic differentiation of marrow stromal cells encapsulated in poly(ethylene glycol)-based hydrogels." Tissue Eng Part A 16(11): 3457-3466.
Drexler, W., U. Morgner, et al. (2001). "Ultrahigh-resolution ophthalmic optical coherence tomography." Nat Med 7(4): 502-507.
Drexler, W., U. Morgner, et al. (1999). "In vivo ultrahigh-resolution optical coherence tomography." Opt Lett 24(17): 1221-1223.
Ducy, P. and G. Karsenty (2000). "The family of bone morphogenetic proteins." Kidney Int 57(6): 2207-2214.
Dunphy, J. E. (1967). "The healing of wounds." Can J Surg 10(3): 281-287.
Egerbacher, M., S. P. Arnoczky, et al. (2008). "Loss of homeostatic tension induces apoptosis in tendon cells: an in vitro study." Clin Orthop Relat Res 466(7): 1562-1568.
Eliasson, P., T. Andersson, et al. (2011). "Achilles tendon healing in rats is improved by intermittent mechanical loading during the inflammatory phase." J Orthop Res 30(2): 274-279.
Enwemeka, C. S., N. I. Spielholz, et al. (1988). "The effect of early functional activities on experimentally tenotomized Achilles tendons in rats." Am J Phys Med Rehabil 67(6): 264-269.
Fahlstrom, M., P. Jonsson, et al. (2003). "Chronic Achilles tendon pain treated with eccentric calf-muscle training." Knee Surg Sports Traumatol Arthrosc 11(5): 327-333.
Fan, H., H. Liu, et al. (2009). "Anterior cruciate ligament regeneration using mesenchymal stem cells and silk scaffold in large animal model." Biomaterials 30(28): 4967-4977.
Fan, H., H. Liu, et al. (2008). "In vivo study of anterior cruciate ligament regeneration using mesenchymal stem cells and silk scaffold." Biomaterials 29(23): 3324-3337.
Fan, J., R. R. Varshney, et al. (2009). "Synovium-derived mesenchymal stem cells: a new cell source for musculoskeletal regeneration." Tissue Eng Part B Rev 15(1): 75-86.
Ferrero, G., E. Fabbro, et al. (2012). "Ultrasound-guided injection of platelet-rich

206
plasma in chronic Achilles and patellar tendinopathy." J Ultrasound 15(4): 260-266.
Filardo, G., M. L. Presti, et al. (2010). "Nonoperative biological treatment approach for partial Achilles tendon lesion." Orthopedics 33(2): 120-123.
Finnoff, J. T., S. P. Fowler, et al. (2011). "Treatment of chronic tendinopathy with ultrasound-guided needle tenotomy and platelet-rich plasma injection." PM R 3(10): 900-911.
Fleming, B. C., K. P. Spindler, et al. (2009). "Collagen-platelet composites improve the biomechanical properties of healing anterior cruciate ligament grafts in a porcine model." Am J Sports Med 37(8): 1554-1563.
Flynn, B. P., A. P. Bhole, et al. (2010). "Mechanical strain stabilizes reconstituted collagen fibrils against enzymatic degradation by mammalian collagenase matrix metalloproteinase 8 (MMP-8)." PLoS One 5(8): e12337.
Ford, L. T. and J. DeBender (1979). "Tendon rupture after local steroid injection." South Med J 72(7): 827-830.
Forslund, C., B. Bylander, et al. (2003). "Indomethacin and celecoxib improve tendon healing in rats." Acta Orthop Scand 74(4): 465-469.
Forslund, C., D. Rueger, et al. (2003). "A comparative dose-response study of cartilage-derived morphogenetic protein (CDMP)-1, -2 and -3 for tendon healing in rats." J Orthop Res 21(4): 617-621.
Franchi, M., A. Trire, et al. (2007). "Collagen structure of tendon relates to function." ScientificWorldJournal 7: 404-420.
Freed, L. E., J. C. Marquis, et al. (1993). "Neocartilage formation in vitro and in vivo using cells cultured on synthetic biodegradable polymers." J Biomed Mater Res 27(1): 11-23.
Freed, L. E., G. Vunjak-Novakovic, et al. (1993). "Cultivation of cell-polymer cartilage implants in bioreactors." J Cell Biochem 51(3): 257-264.
Freeman, J. W., M. D. Woods, et al. (2007). "Tissue engineering of the anterior cruciate ligament using a braid-twist scaffold design." J Biomech 40(9): 2029-2036.
Fu, S. C., Y. P. Wong, et al. (2003). "The roles of bone morphogenetic protein (BMP) 12 in stimulating the proliferation and matrix production of human patellar tendon fibroblasts." Life Sci 72(26): 2965-2974.
Galatz, L., S. Rothermich, et al. (2007). "Development of the supraspinatus tendon-to-bone insertion: localized expression of extracellular matrix and growth factor genes." J Orthop Res 25(12): 1621-1628.

207
Garner, W. L., J. A. McDonald, et al. (1989). "Identification of the collagen-producing cells in healing flexor tendons." Plast Reconstr Surg 83(5): 875-879.
Gaweda, K., M. Tarczynska, et al. (2010). "Treatment of Achilles tendinopathy with platelet-rich plasma." Int J Sports Med 31(8): 577-583.
Gebauer, M., F. T. Beil, et al. (2007). "Mechanical evaluation of different techniques for Achilles tendon repair." Arch Orthop Trauma Surg 127(9): 795-799.
Gilbert, T. W., A. M. Stewart-Akers, et al. (2007). "Degradation and remodeling of small intestinal submucosa in canine Achilles tendon repair." J Bone Joint Surg Am 89(3): 621-630.
Gillespie, H. S. and E. A. George (1969). "Results of surgical repair of spontaneous rupture of the Achilles tendon." J Trauma 9(3): 247-249.
Giombini, A., A. Di Cesare, et al. (2006). "Short-term effectiveness of hyperthermia for supraspinatus tendinopathy in athletes: a short-term randomized controlled study." Am J Sports Med 34(8): 1247-1253.
Goetz, M., S. Thomas, et al. (2008). "Dynamic in vivo imaging of microvasculature and perfusion by miniaturized confocal laser microscopy." Eur Surg Res 41(3): 290-297.
Goldberg, B. A., R. J. Nowinski, et al. (2001). "Outcome of nonoperative management of full-thickness rotator cuff tears." Clin Orthop Relat Res(382): 99-107.
Goldfarb, C. A., R. H. Gelberman, et al. (2007). "Extra-articular steroid injection: early patient response and the incidence of flare reaction." J Hand Surg Am 32(10): 1513-1520.
Goldman, S., R. L. Linscheid, et al. (1969). "Disruptions of the tendo Achillis: analysis of 33 cases." Mayo Clin Proc 44(1): 28-35.
Goodship, A. E., H. L. Birch, et al. (1994). "The pathobiology and repair of tendon and ligament injury." Vet Clin North Am Equine Pract 10(2): 323-349.
Goupille, P., M. I. Jayson, et al. (1998). "Matrix metalloproteinases: the clue to intervertebral disc degeneration?" Spine (Phila Pa 1976) 23(14): 1612-1626.
Grigg, N. L., S. C. Wearing, et al. (2009). "Eccentric calf muscle exercise produces a greater acute reduction in Achilles tendon thickness than concentric exercise." Br J Sports Med 43(4): 280-283.
Hamada, K., Y. Okawara, et al. (1994). "Localization of mRNA of procollagen alpha 1 type I in torn supraspinatus tendons. In situ hybridization using digoxigenin labeled oligonucleotide probe." Clin Orthop Relat Res(304): 18-21.
Hamilton, B. and C. Purdam (2004). "Patellar tendinosis as an adaptive process: a new

208
hypothesis." Br J Sports Med 38(6): 758-761.
Hankemeier, S., M. van Griensven, et al. (2007). "Tissue engineering of tendons and ligaments by human bone marrow stromal cells in a liquid fibrin matrix in immunodeficient rats: results of a histologic study." Arch Orthop Trauma Surg 127(9): 815-821.
Hannafin, J. A. and S. P. Arnoczky (1994). "Effect of cyclic and static tensile loading on water content and solute diffusion in canine flexor tendons: an in vitro study." J Orthop Res 12(3): 350-356.
Hannafin, J. A., S. P. Arnoczky, et al. (1995). "Effect of stress deprivation and cyclic tensile loading on the material and morphologic properties of canine flexor digitorum profundus tendon: an in vitro study." J Orthop Res 13(6): 907-914.
Hansen, U., M. Schunke, et al. (2001). "Combination of reduced oxygen tension and intermittent hydrostatic pressure: a useful tool in articular cartilage tissue engineering." J Biomech 34(7): 941-949.
Harner, C. D., E. Olson, et al. (1996). "Allograft versus autograft anterior cruciate ligament reconstruction: 3- to 5-year outcome." Clin Orthop Relat Res(324): 134-144.
Hart, D. A., J. M. Archambault, et al. (1998). "Gender and neurogenic variables in tendon biology and repetitive motion disorders." Clin Orthop Relat Res(351): 44-56.
Harwood, F. L., R. S. Goomer, et al. (1999). "Regulation of alpha(v)beta3 and alpha5beta1 integrin receptors by basic fibroblast growth factor and platelet-derived growth factor-BB in intrasynovial flexor tendon cells." Wound Repair Regen 7(5): 381-388.
Hattrup, S. J. and K. A. Johnson (1985). "A review of ruptures of the Achilles tendon." Foot Ankle 6(1): 34-38.
Hee, C. K., J. S. Dines, et al. (2011). "Augmentation of a rotator cuff suture repair using rhPDGF-BB and a type I bovine collagen matrix in an ovine model." Am J Sports Med 39(8): 1630-1639.
Hildebrand, K. A., C. B. Frank, et al. (2004). "Gene intervention in ligament and tendon: current status, challenges, future directions." Gene Ther 11(4): 368-378.
Hildebrand, K. A., S. L. Woo, et al. (1998). "The effects of platelet-derived growth factor-BB on healing of the rabbit medial collateral ligament. An in vivo study." Am J Sports Med 26(4): 549-554.
Hoffmeyer, P., C. Freuler, et al. (1990). "Pathological changes in the triceps surae muscle after rupture of the Achilles tendon." Int Orthop 14(2): 183-188.

209
Hogsaa, B., M. Nohr, et al. (1990). "Surgical treatment of Achilles tendon ruptures." Unfallchirurg 93(1): 40-43.
Holtorf, H. L., J. A. Jansen, et al. (2005). "Flow perfusion culture induces the osteoblastic differentiation of marrow stroma cell-scaffold constructs in the absence of dexamethasone." J Biomed Mater Res A 72(3): 326-334.
Hosaka, Y., H. Teraoka, et al. (2005). "Mechanism of cell death in inflamed superficial digital flexor tendon in the horse." J Comp Pathol 132(1): 51-58.
Hou, Y., Z. Mao, et al. (2009). "Effects of transforming growth factor-beta1 and vascular endothelial growth factor 165 gene transfer on Achilles tendon healing." Matrix Biol 28(6): 324-335.
Hoying, J. B. and S. K. Williams (1996). "Effects of basic fibroblast growth factor on human microvessel endothelial cell migration on collagen I correlates inversely with adhesion and is cell density dependent." J Cell Physiol 168(2): 294-304.
Huang, A. H., M. J. Farrell, et al. (2010). "Long-term dynamic loading improves the mechanical properties of chondrogenic mesenchymal stem cell-laden hydrogel." Eur Cell Mater 19: 72-85.
Huang, H. H., A. A. Qureshi, et al. (2000). "Sports and other soft tissue injuries, tendinitis, bursitis, and occupation-related syndromes." Curr Opin Rheumatol 12(2): 150-154.
Husemann, B., F. Muller, et al. (2007). "Effects of locomotion training with assistance of a robot-driven gait orthosis in hemiparetic patients after stroke: a randomized controlled pilot study." Stroke 38(2): 349-354.
Inglis, A. E. and T. P. Sculco (1981). "Surgical repair of ruptures of the tendo Achillis." Clin Orthop Relat Res(156): 160-169.
Ireland, D., R. Harrall, et al. (2001). "Multiple changes in gene expression in chronic human Achilles tendinopathy." Matrix Biol 20(3): 159-169.
Issa, R. I., B. Engebretson, et al. (2011). "The effect of cell seeding density on the cellular and mechanical properties of a mechanostimulated tissue-engineered tendon." Tissue Eng Part A 17(11-12): 1479-1487.
James, R., G. Kesturu, et al. (2008). "Tendon: biology, biomechanics, repair, growth factors, and evolving treatment options." J Hand Surg Am 33(1): 102-112.
Janssen, F. W., J. Oostra, et al. (2006). "A perfusion bioreactor system capable of producing clinically relevant volumes of tissue-engineered bone: in vivo bone formation showing proof of concept." Biomaterials 27(3): 315-323.
Jarvinen, M., L. Jozsa, et al. (1997). "Histopathological findings in chronic tendon disorders." Scand J Med Sci Sports 7(2): 86-95.

210
Jarvinen, T. A., T. L. Jarvinen, et al. (2004). "Collagen fibres of the spontaneously ruptured human tendons display decreased thickness and crimp angle." J Orthop Res 22(6): 1303-1309.
Jiang, Y., H. Liu, et al. (2011). "A proteomic analysis of engineered tendon formation under dynamic mechanical loading in vitro." Biomaterials 32(17): 4085-4095.
Jones, G. C., A. N. Corps, et al. (2006). "Expression profiling of metalloproteinases and tissue inhibitors of metalloproteinases in normal and degenerate human achilles tendon." Arthritis Rheum 54(3): 832-842.
Ju, Y. J., T. Muneta, et al. (2008). "Synovial mesenchymal stem cells accelerate early remodeling of tendon-bone healing." Cell Tissue Res 332(3): 469-478.
Juncosa-Melvin, N., J. T. Shearn, et al. (2006). "Effects of mechanical stimulation on the biomechanics and histology of stem cell-collagen sponge constructs for rabbit patellar tendon repair." Tissue Eng 12(8): 2291-2300.
Kabata, T., T. Kubo, et al. (2000). "Apoptotic cell death in steroid induced osteonecrosis: an experimental study in rabbits." J Rheumatol 27(9): 2166-2171.
Kannus, P. (2000). "Structure of the tendon connective tissue." Scand J Med Sci Sports 10(6): 312-320.
Kannus, P. and L. Jozsa (1991). "Histopathological changes preceding spontaneous rupture of a tendon. A controlled study of 891 patients." J Bone Joint Surg Am 73(10): 1507-1525.
Kannus, P., L. Jozsa, et al. (1997). "Effects of training, immobilization and remobilization on tendons." Scand J Med Sci Sports 7(2): 67-71.
Kannus, P. and A. Natri (1997). "Etiology and pathophysiology of tendon ruptures in sports." Scand J Med Sci Sports 7(2): 107-112.
Kashiwagi, K., Y. Mochizuki, et al. (2004). "Effects of transforming growth factor-beta 1 on the early stages of healing of the Achilles tendon in a rat model." Scand J Plast Reconstr Surg Hand Surg 38(4): 193-197.
Kastelic, J., A. Galeski, et al. (1978). "The multicomposite structure of tendon." Connect Tissue Res 6(1): 11-23.
Kastelic, J., I. Palley, et al. (1980). "A structural mechanical model for tendon crimping." J Biomech 13(10): 887-893.
Ker, R. F. (1981). "Dynamic tensile properties of the plantaris tendon of sheep (Ovis aries)." J Exp Biol 93: 283-302.
Khan, K. and J. Cook (2003). "The painful nonruptured tendon: clinical aspects." Clin Sports Med 22(4): 711-725.

211
Khan, K. M., J. L. Cook, et al. (1999). "Histopathology of common tendinopathies. Update and implications for clinical management." Sports Med 27(6): 393-408.
Khan, K. M. and N. Maffulli (1998). "Tendinopathy: an Achilles' heel for athletes and clinicians." Clin J Sport Med 8(3): 151-154.
Khan, K. M. and A. Scott (2009). "Mechanotherapy: how physical therapists' prescription of exercise promotes tissue repair." Br J Sports Med 43(4): 247-252.
Khayat, G., D. H. Rosenzweig, et al. (2012). "Low frequency mechanical stimulation inhibits adipogenic differentiation of C3H10T1/2 mesenchymal stem cells." Differentiation 83(4): 179-184.
Kim, H. J., S. W. Kang, et al. (2007). "The role of transforming growth factor-beta and bone morphogenetic protein with fibrin glue in healing of bone-tendon junction injury." Connect Tissue Res 48(6): 309-315.
Kim, H. M., L. M. Galatz, et al. (2011). "The role of transforming growth factor beta isoforms in tendon-to-bone healing." Connect Tissue Res 52(2): 87-98.
Kingma, J. J., R. de Knikker, et al. (2007). "Eccentric overload training in patients with chronic Achilles tendinopathy: a systematic review." Br J Sports Med 41(6): e3.
Kinneberg, K. R., V. S. Nirmalanandhan, et al. (2010). "Chondroitin-6-sulfate incorporation and mechanical stimulation increase MSC-collagen sponge construct stiffness." J Orthop Res 28(8): 1092-1099.
Kleinman, M. and A. E. Gross (1983). "Achilles tendon rupture following steroid injection. Report of three cases." J Bone Joint Surg Am 65(9): 1345-1347.
Knauper, V., C. Lopez-Otin, et al. (1996). "Biochemical characterization of human collagenase-3." J Biol Chem 271(3): 1544-1550.
Kobayashi, M., E. Itoi, et al. (2006). "Expression of growth factors in the early phase of supraspinatus tendon healing in rabbits." J Shoulder Elbow Surg 15(3): 371-377.
Komi, P. V., S. Fukashiro, et al. (1992). "Biomechanical loading of Achilles tendon during normal locomotion." Clin Sports Med 11(3): 521-531.
Kong, A., A. Van der Vliet, et al. (2007). "MRI and US of gluteal tendinopathy in greater trochanteric pain syndrome." Eur Radiol 17(7): 1772-1783.
Kovacevic, D., A. J. Fox, et al. (2011). "Calcium-phosphate matrix with or without TGF-beta3 improves tendon-bone healing after rotator cuff repair." Am J Sports Med 39(4): 811-819.
Krane, S. M., M. H. Byrne, et al. (1996). "Different collagenase gene products have different roles in degradation of type I collagen." J Biol Chem 271(45): 28509-28515.

212
Krueger-Franke, M., C. H. Siebert, et al. (1995). "Surgical treatment of ruptures of the Achilles tendon: a review of long-term results." Br J Sports Med 29(2): 121-125.
Kuhn, K. (1969). "The structure of collagen." Essays Biochem 5: 59-87.
Kulick, M. I., S. Smith, et al. (1986). "Oral ibuprofen: evaluation of its effect on peritendinous adhesions and the breaking strength of a tenorrhaphy." J Hand Surg Am 11(1): 110-120.
Kupcsik, L., M. J. Stoddart, et al. (2010). "Improving chondrogenesis: potential and limitations of SOX9 gene transfer and mechanical stimulation for cartilage tissue engineering." Tissue Eng Part A 16(6): 1845-1855.
Langberg, H., D. Skovgaard, et al. (1999). "Type I collagen synthesis and degradation in peritendinous tissue after exercise determined by microdialysis in humans." J Physiol 521 Pt 1: 299-306.
Larsen, N. P., M. R. Forwood, et al. (1987). "Immobilization and retraining of cruciate ligaments in the rat." Acta Orthop Scand 58(3): 260-264.
Lavagnino, M., S. P. Arnoczky, et al. (2006). "Isolated fibrillar damage in tendons stimulates local collagenase mRNA expression and protein synthesis." J Biomech 39(13): 2355-2362.
Lavagnino, M., S. P. Arnoczky, et al. (2003). "Effect of amplitude and frequency of cyclic tensile strain on the inhibition of MMP-1 mRNA expression in tendon cells: an in vitro study." Connect Tissue Res 44(3-4): 181-187.
Leach, R. E., A. A. Schepsis, et al. (1992). "Long-term results of surgical management of Achilles tendinitis in runners." Clin Orthop Relat Res(282): 208-212.
Leadbetter, W. B. (1992). "Cell-matrix response in tendon injury." Clin Sports Med 11(3): 533-578.
Lee, C. H., H. J. Shin, et al. (2005). "Nanofiber alignment and direction of mechanical strain affect the ECM production of human ACL fibroblast." Biomaterials 26(11): 1261-1270.
Lejard, V., G. Brideau, et al. (2007). "Scleraxis and NFATc regulate the expression of the pro-alpha1(I) collagen gene in tendon fibroblasts." J Biol Chem 282(24): 17665-17675.
Leppilahti, J. and S. Orava (1998). "Total Achilles tendon rupture. A review." Sports Med 25(2): 79-100.
Li, G., L. E. Defrate, et al. (2005). "In vivo kinematics of the ACL during weight-bearing knee flexion." J Orthop Res 23(2): 340-344.
Lian, O., A. Scott, et al. (2007). "Excessive apoptosis in patellar tendinopathy in

213
athletes." Am J Sports Med 35(4): 605-611.
Lin, T. W., L. Cardenas, et al. (2004). "Biomechanics of tendon injury and repair." J Biomech 37(6): 865-877.
Lindsay, W. K. and J. R. Birch (1964). "The Fibroblast in Flexor Tendon Healing." Plast Reconstr Surg 34: 223-232.
Liu, J., M. Hsu, et al. (2001). "On the design of rotating speed functions to improve the acceleration peak value of ball-screw transmission mechanism." Mechanism and Machine Theory 36(9): 1035-1049.
Liu, S. H., R. S. Yang, et al. (1995). "Collagen in tendon, ligament, and bone healing. A current review." Clin Orthop Relat Res(318): 265-278.
Liu, W., B. Chen, et al. (2006). "Repair of tendon defect with dermal fibroblast engineered tendon in a porcine model." Tissue Eng 12(4): 775-788.
Lo, A. C. and E. W. Triche (2008). "Improving gait in multiple sclerosis using robot-assisted, body weight supported treadmill training." Neurorehabil Neural Repair 22(6): 661-671.
Lo, I. K., L. L. Marchuk, et al. (2004). "Matrix metalloproteinase and tissue inhibitor of matrix metalloproteinase mRNA levels are specifically altered in torn rotator cuff tendons." Am J Sports Med 32(5): 1223-1229.
Lo, I. K., Y. Ou, et al. (2002). "The cellular networks of normal ovine medial collateral and anterior cruciate ligaments are not accurately recapitulated in scar tissue." J Anat 200(Pt 3): 283-296.
Loitz, B. J., R. F. Zernicke, et al. (1989). "Effects of short-term immobilization versus continuous passive motion on the biomechanical and biochemical properties of the rabbit tendon." Clin Orthop Relat Res(244): 265-271.
Lou, J., Y. Tu, et al. (2001). "BMP-12 gene transfer augmentation of lacerated tendon repair." J Orthop Res 19(6): 1199-1202.
Ma, G. W. and T. G. Griffith (1977). "Percutaneous repair of acute closed ruptured achilles tendon: a new technique." Clin Orthop Relat Res(128): 247-255.
Machner, A., A. Baier, et al. (2003). "Higher susceptibility to Fas ligand induced apoptosis and altered modulation of cell death by tumor necrosis factor-alpha in periarticular tenocytes from patients with knee joint osteoarthritis." Arthritis Res Ther 5(5): R253-261.
Maeda, T., T. Sakabe, et al. (2011). "Conversion of mechanical force into TGF-beta-mediated biochemical signals." Curr Biol 21(11): 933-941.
Maffulli, N. (1995). "Ultrasound of the Achilles tendon after surgical repair:

214
morphology and function." Br J Radiol 68(816): 1372-1373.
Maffulli, N. (1998). "Current concepts in the management of subcutaneous tears of the Achilles tendon." Bull Hosp Jt Dis 57(3): 152-158.
Maffulli, N., K. M. Khan, et al. (1998). "Overuse tendon conditions: time to change a confusing terminology." Arthroscopy 14(8): 840-843.
Maffulli, N., P. Sharma, et al. (2004). "Achilles tendinopathy: aetiology and management." J R Soc Med 97(10): 472-476.
Maffulli, N., V. Testa, et al. (1997). "Results of percutaneous longitudinal tenotomy for Achilles tendinopathy in middle- and long-distance runners." Am J Sports Med 25(6): 835-840.
Magnusson, S. P., P. Hansen, et al. (2003). "Tendon properties in relation to muscular activity and physical training." Scand J Med Sci Sports 13(4): 211-223.
Magra, M. and N. Maffulli (2005). "Matrix metalloproteases: a role in overuse tendinopathies." Br J Sports Med 39(11): 789-791.
Majewski, M., O. Betz, et al. (2008). "Ex vivo adenoviral transfer of bone morphogenetic protein 12 (BMP-12) cDNA improves Achilles tendon healing in a rat model." Gene Ther 15(16): 1139-1146.
Malcarney, H. L., F. Bonar, et al. (2005). "Early inflammatory reaction after rotator cuff repair with a porcine small intestine submucosal implant: a report of 4 cases." Am J Sports Med 33(6): 907-911.
Manning, C. N., H. M. Kim, et al. (2011). "Sustained delivery of transforming growth factor beta three enhances tendon-to-bone healing in a rat model." J Orthop Res 29(7): 1099-1105.
Manske, P. R. (1988). "Flexor tendon healing." J Hand Surg Br 13(3): 237-245.
Marui, T., C. Niyibizi, et al. (1997). "Effect of growth factors on matrix synthesis by ligament fibroblasts." J Orthop Res 15(1): 18-23.
Matsumoto, F., G. Trudel, et al. (2003). "Mechanical effects of immobilization on the Achilles' tendon." Arch Phys Med Rehabil 84(5): 662-667.
McCawley, L. J. and L. M. Matrisian (2001). "Matrix metalloproteinases: they're not just for matrix anymore!" Curr Opin Cell Biol 13(5): 534-540.
Meinel, L., V. Karageorgiou, et al. (2004). "Bone tissue engineering using human mesenchymal stem cells: effects of scaffold material and medium flow." Ann Biomed Eng 32(1): 112-122.
Mikic, B., B. J. Schalet, et al. (2001). "GDF-5 deficiency in mice alters the

215
ultrastructure, mechanical properties and composition of the Achilles tendon." J Orthop Res 19(3): 365-371.
Millar, N. L., J. H. Reilly, et al. (2012). "Hypoxia: a critical regulator of early human tendinopathy." Ann Rheum Dis 71(2): 302-310.
Millar, N. L., A. Q. Wei, et al. (2009). "Cytokines and apoptosis in supraspinatus tendinopathy." J Bone Joint Surg Br 91(3): 417-424.
Mishra, A., P. Randelli, et al. (2012). "Platelet-rich plasma and the upper extremity." Hand Clin 28(4): 481-491.
Moffat, K. L., A. S. Kwei, et al. (2009). "Novel nanofiber-based scaffold for rotator cuff repair and augmentation." Tissue Eng Part A 15(1): 115-126.
Molloy, T., Y. Wang, et al. (2003). "The roles of growth factors in tendon and ligament healing." Sports Med 33(5): 381-394.
Monto, R. R. (2012). "Platelet rich plasma treatment for chronic Achilles tendinosis." Foot Ankle Int 33(5): 379-385.
Morand, E. F., M. Leech, et al. (2006). "MIF: a new cytokine link between rheumatoid arthritis and atherosclerosis." Nat Rev Drug Discov 5(5): 399-410.
Morrison, D. S., A. D. Frogameni, et al. (1997). "Non-operative treatment of subacromial impingement syndrome." J Bone Joint Surg Am 79(5): 732-737.
Murata, H., H. Nishizono, et al. (2000). "Postnatal development of structure and arrangement of tendon cells. A scanning and transmission electron microscope study in the rat calcaneal tendon." Okajimas Folia Anat Jpn 77(2-3): 77-86.
Myers, B. and M. Wolf (1974). "Vascularization of the healing wound." Am Surg 40(12): 716-722.
Nagase, H., R. Visse, et al. (2006). "Structure and function of matrix metalloproteinases and TIMPs." Cardiovasc Res 69(3): 562-573.
Nakagawa, Y., M. Totsuka, et al. (1989). "Effect of disuse on the ultrastructure of the achilles tendon in rats." Eur J Appl Physiol Occup Physiol 59(3): 239-242.
Nakama, L. H., K. B. King, et al. (2005). "Evidence of tendon microtears due to cyclical loading in an in vivo tendinopathy model." J Orthop Res 23(5): 1199-1205.
Nakamura, N., K. Shino, et al. (1998). "Early biological effect of in vivo gene transfer of platelet-derived growth factor (PDGF)-B into healing patellar ligament." Gene Ther 5(9): 1165-1170.
Nakamura, N., S. A. Timmermann, et al. (1998). "A comparison of in vivo gene delivery methods for antisense therapy in ligament healing." Gene Ther 5(11):

216
1455-1461.
Nathan, C. (2002). "Points of control in inflammation." Nature 420(6917): 846-852.
Nguyen, M. T., H. Lue, et al. (2003). "The cytokine macrophage migration inhibitory factor reduces pro-oxidative stress-induced apoptosis." J Immunol 170(6): 3337-3347.
Nguyen, T. D., R. Liang, et al. (2009). "Effects of cell seeding and cyclic stretch on the fiber remodeling in an extracellular matrix-derived bioscaffold." Tissue Eng Part A 15(4): 957-963.
Ni, M., P. P. Lui, et al. (2012). "Tendon-derived stem cells (TDSCs) promote tendon repair in a rat patellar tendon window defect model." J Orthop Res 30(4): 613-619.
Ni, M., Y. F. Rui, et al. (2013). "Engineered scaffold-free tendon tissue produced by tendon-derived stem cells." Biomaterials 34(8): 2024-2037.
Nirmalanandhan, V. S., M. Rao, et al. (2008). "Effect of scaffold material, construct length and mechanical stimulation on the in vitro stiffness of the engineered tendon construct." J Biomech 41(4): 822-828.
Nirmalanandhan, V. S., J. T. Shearn, et al. (2008). "Improving linear stiffness of the cell-seeded collagen sponge constructs by varying the components of the mechanical stimulus." Tissue Eng Part A 14(11): 1883-1891.
Nistor, L. (1981). "Surgical and non-surgical treatment of Achilles Tendon rupture. A prospective randomized study." J Bone Joint Surg Am 63(3): 394-399.
Nyyssonen, T. and P. Luthje (2000). "Achilles tendon ruptures in South-East Finland between 1986-1996, with special reference to epidemiology, complications of surgery and hospital costs." Ann Chir Gynaecol 89(1): 53-57.
Obaid, H. and D. Connell (2010). "Cell therapy in tendon disorders: what is the current evidence?" Am J Sports Med 38(10): 2123-2132.
Obst, S. J., R. Newsham-West, et al. "In Vivo Measurement of Human Achilles Tendon Morphology Using Freehand 3-D Ultrasound." Ultrasound Med Biol.
Ohberg, L. and H. Alfredson (2003). "Sclerosing therapy in chronic Achilles tendon insertional pain-results of a pilot study." Knee Surg Sports Traumatol Arthrosc 11(5): 339-343.
Olivier, V., P. Hivart, et al. (2007). "In vitro culture of large bone substitutes in a new bioreactor: importance of the flow direction." Biomed Mater 2(3): 174-180.
Ouyang, H. W., J. C. Goh, et al. (2004). "Viability of allogeneic bone marrow stromal cells following local delivery into patella tendon in rabbit model." Cell

217
Transplant 13(6): 649-657.
Owens, R. F., Jr., J. Ginnetti, et al. (2011). "Clinical and magnetic resonance imaging outcomes following platelet rich plasma injection for chronic midsubstance Achilles tendinopathy." Foot Ankle Int 32(11): 1032-1039.
Paavola, M., S. Orava, et al. (2000). "Chronic Achilles tendon overuse injury: complications after surgical treatment. An analysis of 432 consecutive patients." Am J Sports Med 28(1): 77-82.
Parent, G., N. Huppe, et al. (2011). "Low stress tendon fatigue is a relatively rapid process in the context of overuse injuries." Ann Biomed Eng 39(5): 1535-1545.
Parkinson, J., T. Samiric, et al. (2010). "Change in proteoglycan metabolism is a characteristic of human patellar tendinopathy." Arthritis Rheum 62(10): 3028-3035.
Parkinson, J., T. Samiric, et al. (2011). "Involvement of proteoglycans in tendinopathy." J Musculoskelet Neuronal Interact 11(2): 86-93.
Peach, M. S., R. James, et al. (2012). "Polyphosphazene functionalized polyester fiber matrices for tendon tissue engineering: in vitro evaluation with human mesenchymal stem cells." Biomed Mater 7(4): 045016.
Petersen, W., T. Pufe, et al. (2003). "Hypoxia and PDGF have a synergistic effect that increases the expression of the angiogenetic peptide vascular endothelial growth factor in Achilles tendon fibroblasts." Arch Orthop Trauma Surg 123(9): 485-488.
Pham, Q. P., U. Sharma, et al. (2006). "Electrospinning of polymeric nanofibers for tissue engineering applications: a review." Tissue Eng 12(5): 1197-1211.
Pillai, R. S., D. Lorenser, et al. "Deep-tissue access with confocal fluorescence microendoscopy through hypodermic needles." Opt Express 19(8): 7213-7221.
Pryce, B. A., S. S. Watson, et al. (2009). "Recruitment and maintenance of tendon progenitors by TGFbeta signaling are essential for tendon formation." Development 136(8): 1351-1361.
Puddu, G., E. Ippolito, et al. (1976). "A classification of Achilles tendon disease." Am J Sports Med 4(4): 145-150.
Quirk, B. C., R. A. McLaughlin, et al. "In situ imaging of lung alveoli with an optical coherence tomography needle probe." J Biomed Opt 16(3): 036009.
Rauh, J., F. Milan, et al. (2011). "Bioreactor systems for bone tissue engineering." Tissue Eng Part B Rev 17(4): 263-280.
Rauh, J., F. Milan, et al. (2012). "Bioreactor systems for bone tissue engineering."

218
Tissue Eng Part B Rev 17(4): 263-280.
Rees, J. D., A. M. Wilson, et al. (2006). "Current concepts in the management of tendon disorders." Rheumatology (Oxford) 45(5): 508-521.
Reeves, N. D., C. N. Maganaris, et al. (2005). "Influence of 90-day simulated microgravity on human tendon mechanical properties and the effect of resistive countermeasures." J Appl Physiol 98(6): 2278-2286.
Riemersma, D. J. and H. C. Schamhardt (1985). "In vitro mechanical properties of equine tendons in relation to cross-sectional area and collagen content." Research in Veterinary Science 39(3): 263-270.
Riley, G. P., V. Curry, et al. (2002). "Matrix metalloproteinase activities and their relationship with collagen remodelling in tendon pathology." Matrix Biol 21(2): 185-195.
Riley, G. P., R. L. Harrall, et al. (1994). "Tendon degeneration and chronic shoulder pain: changes in the collagen composition of the human rotator cuff tendons in rotator cuff tendinitis." Ann Rheum Dis 53(6): 359-366.
Rolf, C. and T. Movin (1997). "Etiology, histopathology, and outcome of surgery in achillodynia." Foot Ankle Int 18(9): 565-569.
Rompe, J. D., J. Furia, et al. (2008). "Eccentric loading compared with shock wave treatment for chronic insertional achilles tendinopathy. A randomized, controlled trial." J Bone Joint Surg Am 90(1): 52-61.
Roos, E. M., M. Engstrom, et al. (2004). "Clinical improvement after 6 weeks of eccentric exercise in patients with mid-portion Achilles tendinopathy -- a randomized trial with 1-year follow-up." Scand J Med Sci Sports 14(5): 286-295.
Ross, J. J., M. J. Duxson, et al. (1987). "Neural determination of muscle fibre numbers in embryonic rat lumbrical muscles." Development 100(3): 395-409.
Ruedi, T. P. and M. Allgower (1979). "The operative treatment of intra-articular fractures of the lower end of the tibia." Clin Orthop Relat Res(138): 105-110.
Rui, Y. F., P. P. Lui, et al. (2011). "Mechanical loading increased BMP-2 expression which promoted osteogenic differentiation of tendon-derived stem cells." J Orthop Res 29(3): 390-396.
Saber, S., A. Y. Zhang, et al. (2010). "Flexor tendon tissue engineering: bioreactor cyclic strain increases construct strength." Tissue Eng Part A 16(6): 2085-2090.
Sahoo, S., J. G. Cho-Hong, et al. (2007). "Development of hybrid polymer scaffolds for potential applications in ligament and tendon tissue engineering." Biomed Mater 2(3): 169-173.

219
Sahoo, S., H. Ouyang, et al. (2006). "Characterization of a novel polymeric scaffold for potential application in tendon/ligament tissue engineering." Tissue Eng 12(1): 91-99.
Sahoo, S., S. L. Toh, et al. (2010). "A bFGF-releasing silk/PLGA-based biohybrid scaffold for ligament/tendon tissue engineering using mesenchymal progenitor cells." Biomaterials 31(11): 2990-2998.
Saini, S. and T. M. Wick (2004). "Effect of low oxygen tension on tissue-engineered cartilage construct development in the concentric cylinder bioreactor." Tissue Eng 10(5-6): 825-832.
Salter, E., B. Goh, et al. (2012). "Bone tissue engineering bioreactors: a role in the clinic?" Tissue Eng Part B Rev 18(1): 62-75.
Samilson, R. L. and W. F. Binder (1975). "Symptomatic full thickness tears of rotator cuff. An analysis of 292 shoulders in 276 patients." Orthop Clin North Am 6(2): 449-466.
Sanchez, M., E. Anitua, et al. (2007). "Comparison of surgically repaired Achilles tendon tears using platelet-rich fibrin matrices." Am J Sports Med 35(2): 245-251.
Schechtman, H. and D. L. Bader (1997). "In vitro fatigue of human tendons." J Biomech 30(8): 829-835.
Schepsis, A. A., H. Jones, et al. (2002). "Achilles tendon disorders in athletes." Am J Sports Med 30(2): 287-305.
Schepull, T., J. Kvist, et al. (2011). "Autologous platelets have no effect on the healing of human achilles tendon ruptures: a randomized single-blind study." Am J Sports Med 39(1): 38-47.
Schneewind, J. H., I. K. Kline, et al. (1964). "The Role of Paratenon in Healing of Experimental Tendon Transplants." J Occup Med 6: 429-436.
Schwartz, I., A. Sajin, et al. (2012). "Robot-assisted gait training in multiple sclerosis patients: a randomized trial." Mult Scler 18(6): 881-890.
Scott, J. E. (1984). "The periphery of the developing collagen fibril. Quantitative relationships with dermatan sulphate and other surface-associated species." Biochem J 218(1): 229-233.
Screen, H. R., V. H. Chhaya, et al. (2006). "The influence of swelling and matrix degradation on the microstructural integrity of tendon." Acta Biomater 2(5): 505-513.
Screen, H. R., D. A. Lee, et al. (2004). "An investigation into the effects of the hierarchical structure of tendon fascicles on micromechanical properties." Proc

220
Inst Mech Eng H 218(2): 109-119.
Sen, B., Z. Xie, et al. (2008). "Mechanical strain inhibits adipogenesis in mesenchymal stem cells by stimulating a durable beta-catenin signal." Endocrinology 149(12): 6065-6075.
Shalabi, A., M. Kristoffersen-Wilberg, et al. (2004). "Eccentric training of the gastrocnemius-soleus complex in chronic Achilles tendinopathy results in decreased tendon volume and intratendinous signal as evaluated by MRI." Am J Sports Med 32(5): 1286-1296.
Sharifpoor, S., C. A. Simmons, et al. (2011). "Functional characterization of human coronary artery smooth muscle cells under cyclic mechanical strain in a degradable polyurethane scaffold." Biomaterials 32(21): 4816-4829.
Shi, Y., H. Li, et al. (2011). "Continuous cyclic mechanical tension inhibited Runx2 expression in mesenchymal stem cells through RhoA-ERK1/2 pathway." J Cell Physiol 226(8): 2159-2169.
Silbernagel, K. G., R. Thomee, et al. (2001). "Eccentric overload training for patients with chronic Achilles tendon pain--a randomised controlled study with reliability testing of the evaluation methods." Scand J Med Sci Sports 11(4): 197-206.
Skutek, M., M. van Griensven, et al. (2001). "Cyclic mechanical stretching enhances secretion of Interleukin 6 in human tendon fibroblasts." Knee Surg Sports Traumatol Arthrosc 9(5): 322-326.
Skutek, M., M. van Griensven, et al. (2001). "Cyclic mechanical stretching modulates secretion pattern of growth factors in human tendon fibroblasts." Eur J Appl Physiol 86(1): 48-52.
Skutek, M., M. van Griensven, et al. (2003). "Cyclic mechanical stretching of human patellar tendon fibroblasts: activation of JNK and modulation of apoptosis." Knee Surg Sports Traumatol Arthrosc 11(2): 122-129.
Smith, K. D., A. Vaughan-Thomas, et al. (2011). "The organisation of elastin and fibrillins 1 and 2 in the cruciate ligament complex." J Anat 218(6): 600-607.
Smith, L. L. (2000). "Cytokine hypothesis of overtraining: a physiological adaptation to excessive stress?" Med Sci Sports Exerc 32(2): 317-331.
Smith, M. M., G. Sakurai, et al. (2008). "Modulation of aggrecan and ADAMTS expression in ovine tendinopathy induced by altered strain." Arthritis Rheum 58(4): 1055-1066.
Smithpeter, C. L., A. K. Dunn, et al. (1998). "Penetration depth limits of in vivo confocal reflectance imaging." Appl Opt 37(13): 2749-2754.
So, V. and H. Pollard (1997). "Management of Achilles tendon disorders. A case

221
review." Australas Chiropr Osteopathy 6(2): 58-62.
Sporn, M. B., A. B. Roberts, et al. (1986). "Transforming growth factor-beta: biological function and chemical structure." Science 233(4763): 532-534.
Stanish, W. D., R. M. Rubinovich, et al. (1986). "Eccentric exercise in chronic tendinitis." Clin Orthop Relat Res(208): 65-68.
Stoll, C., T. John, et al. (2010). "Extracellular matrix expression of human tenocytes in three-dimensional air-liquid and PLGA cultures compared with tendon tissue: implications for tendon tissue engineering." J Orthop Res 28(9): 1170-1177.
Straudi, S., M. G. Benedetti, et al. (2013). "Does robot-assisted gait training ameliorate gait abnormalities in multiple sclerosis? A pilot randomized-control trial." NeuroRehabilitation.
Suchak, A. A., G. Bostick, et al. (2005). "The incidence of Achilles tendon ruptures in Edmonton, Canada." Foot Ankle Int 26(11): 932-936.
Takahasih, S., M. Nakajima, et al. (2002). "Effect of recombinant basic fibroblast growth factor (bFGF) on fibroblast-like cells from human rotator cuff tendon." Tohoku J Exp Med 198(4): 207-214.
Tallon, C., B. D. Coleman, et al. (2001). "Outcome of surgery for chronic Achilles tendinopathy. A critical review." Am J Sports Med 29(3): 315-320.
Tang, J. B., Y. Cao, et al. (2008). "Adeno-associated virus-2-mediated bFGF gene transfer to digital flexor tendons significantly increases healing strength. an in vivo study." J Bone Joint Surg Am 90(5): 1078-1089.
Teitz, C. C., W. E. Garrett, Jr., et al. (1997). "Tendon problems in athletic individuals." Instr Course Lect 46: 569-582.
Thermann, H., O. Frerichs, et al. (2001). "Healing of the Achilles tendon: an experimental study." Foot Ankle Int 22(6): 478-483.
Thomopoulos, S., R. Das, et al. (2010). "bFGF and PDGF-BB for tendon repair: controlled release and biologic activity by tendon fibroblasts in vitro." Ann Biomed Eng 38(2): 225-234.
Thomopoulos, S., F. L. Harwood, et al. (2005). "Effect of several growth factors on canine flexor tendon fibroblast proliferation and collagen synthesis in vitro." J Hand Surg Am 30(3): 441-447.
Thompson, M. S. (2013). "Tendon mechanobiology: experimental models require mathematical underpinning." Bull Math Biol 75(8): 1238-1254.
Thornton, G. M., X. Shao, et al. (2008). "Changes in mechanical loading lead to tendonspecific alterations in MMP and TIMP expression: influence of stress

222
deprivation and intermittent cyclic hydrostatic compression on rat supraspinatus and Achilles tendons." Br J Sports Med 44(10): 698-703.
Tillander, B., L. E. Franzen, et al. (1999). "Effect of steroid injections on the rotator cuff: an experimental study in rats." J Shoulder Elbow Surg 8(3): 271-274.
Tkocz, C. and K. Kuhn (1969). "The formation of triple-helical collagen molecules from alpha-1 or alpha-2 polypeptide chains." Eur J Biochem 7(4): 454-462.
Tomita, F., K. Yasuda, et al. (2001). "Comparisons of intraosseous graft healing between the doubled flexor tendon graft and the bone-patellar tendon-bone graft in anterior cruciate ligament reconstruction." Arthroscopy 17(5): 461-476.
Trudel, G., Y. Koike, et al. (2007). "Mechanical alterations of rabbit Achilles' tendon after immobilization correlate with bone mineral density but not with magnetic resonance or ultrasound imaging." Arch Phys Med Rehabil 88(12): 1720-1726.
Tsuzaki, M., D. Bynum, et al. (2003). "ATP modulates load-inducible IL-1beta, COX 2, and MMP-3 gene expression in human tendon cells." J Cell Biochem 89(3): 556-562.
Uccelli, A., L. Moretta, et al. (2006). "Immunoregulatory function of mesenchymal stem cells." Eur J Immunol 36(10): 2566-2573.
Uemura, T., J. Dong, et al. (2003). "Transplantation of cultured bone cells using combinations of scaffolds and culture techniques." Biomaterials 24(13): 2277-2286.
Uysal, A. C. and H. Mizuno (2010). "Tendon regeneration and repair with adipose derived stem cells." Curr Stem Cell Res Ther 5(2): 161-167.
Uysal, A. C. and H. Mizuno (2011). "Differentiation of adipose-derived stem cells for tendon repair." Methods Mol Biol 702: 443-451.
Uysal, C. A., M. Tobita, et al. (2012). "Adipose-derived stem cells enhance primary tendon repair: biomechanical and immunohistochemical evaluation." J Plast Reconstr Aesthet Surg 65(12): 1712-1719.
Varseveld, R. B. v. and G. M. Bone (1997). "Accurate Position Control of a Pneumatic Actuator Using On/Off Solenoid Valves." TRANSACTIONS ON MECHATRONICS 2: 195-204.
Vogel, K. G. (2004). "What happens when tendons bend and twist? Proteoglycans." J Musculoskelet Neuronal Interact 4(2): 202-203.
Wang, A., W. Breidahl, et al. (2013). "Autologous Tenocyte Injection for the Treatment of Severe, Chronic Resistant Lateral Epicondylitis: A Pilot Study." Am J Sports Med.

223
Wang, J. H. (2006). "Mechanobiology of tendon." J Biomech 39(9): 1563-1582.
Wang, J. H., F. Jia, et al. (2003). "Cyclic mechanical stretching of human tendon fibroblasts increases the production of prostaglandin E2 and levels of cyclooxygenase expression: a novel in vitro model study." Connect Tissue Res 44(3-4): 128-133.
Wang, L., K. Seshareddy, et al. (2009). "Effect of initial seeding density on human umbilical cord mesenchymal stromal cells for fibrocartilage tissue engineering." Tissue Eng Part A 15(5): 1009-1017.
Wang, T., B. S. Gardiner, et al. (2013). "Bioreactor design for tendon/ligament engineering." Tissue Eng Part B Rev 19(2): 133-146.
Wang, T., B. Gardner, et al. (2012). "Bioreactor design for tendon/ligament engineering." Tissue Eng Part B Rev.
Wang, T., Z. Lin, et al. (2013). "Programmable mechanical stimulation influences tendon homeostasis in a bioreactor system." Biotechnol Bioeng 110(5): 1495-1507.
Webb, K., R. W. Hitchcock, et al. (2006). "Cyclic strain increases fibroblast proliferation, matrix accumulation, and elastic modulus of fibroblast-seeded polyurethane constructs." J Biomech 39(6): 1136-1144.
Weiler, A., F. N. Unterhauser, et al. (2002). "Alpha-smooth muscle actin is expressed by fibroblastic cells of the ovine anterior cruciate ligament and its free tendon graft during remodeling." J Orthop Res 20(2): 310-317.
West, J. R., N. Juncosa, et al. (2004). "Characterization of in vivo Achilles tendon forces in rabbits during treadmill locomotion at varying speeds and inclinations." J Biomech 37(11): 1647-1653.
Wiggins, M. E., P. D. Fadale, et al. (1995). "Effects of local injection of corticosteroids on the healing of ligaments. A follow-up report." J Bone Joint Surg Am 77(11): 1682-1691.
Williams, J. G. (1993). "Achilles tendon lesions in sport." Sports Med 16(3): 216-220.
Wilson, A. M. and A. E. Goodship (1994). "Exercise-induced hyperthermia as a possible mechanism for tendon degeneration." J Biomech 27(7): 899-905.
Wojciak, B. and J. F. Crossan (1994). "The effects of T cells and their products on in vitro healing of epitenon cell microwounds." Immunology 83(1): 93-98.
Woodley, B. L., R. J. Newsham-West, et al. (2007). "Chronic tendinopathy: effectiveness of eccentric exercise." Br J Sports Med 41(4): 188-198; discussion 199.

224
Woon, C. Y., S. Farnebo, et al. (2012). "Human Flexor Tendon Tissue Engineering: Revitalization of Biostatic Allograft Scaffolds." Tissue Eng Part A.
Woon, C. Y., A. Kraus, et al. (2011). "Three-dimensional-construct bioreactor conditioning in human tendon tissue engineering." Tissue Eng Part A 17(19-20): 2561-2572.
Woon, C. Y., A. Kraus, et al. (2011). "Three-Dimensional-Construct Bioreactor Conditioning in Human Tendon Tissue Engineering." Tissue Eng Part A.
Wren, T. A., D. P. Lindsey, et al. (2003). "Effects of creep and cyclic loading on the mechanical properties and failure of human Achilles tendons." Ann Biomed Eng 31(6): 710-717.
Wren, T. A., S. A. Yerby, et al. (2001). "Mechanical properties of the human achilles tendon." Clin Biomech (Bristol, Avon) 16(3): 245-251.
Yang, G., H. J. Im, et al. (2005). "Repetitive mechanical stretching modulates IL-1beta induced COX-2, MMP-1 expression, and PGE2 production in human patellar tendon fibroblasts." Gene 363: 166-172.
Ye, S., P. Eriksson, et al. (1996). "Progression of coronary atherosclerosis is associated with a common genetic variant of the human stromelysin-1 promoter which results in reduced gene expression." J Biol Chem 271(22): 13055-13060.
Yuan, J., M. X. Wang, et al. (2003). "Cell death and tendinopathy." Clin Sports Med 22(4): 693-701.
Zamara, E., S. Galastri, et al. (2007). "Prevention of severe toxic liver injury and oxidative stress in MCP-1-deficient mice." J Hepatol 46(2): 230-238.
Zell, R. A. and V. M. Santoro (2000). "Augmented repair of acute Achilles tendon ruptures." Foot Ankle Int 21(6): 469-474.
Zhang, F., H. Liu, et al. (2003). "Effect of vascular endothelial growth factor on rat Achilles tendon healing." Plast Reconstr Surg 112(6): 1613-1619.
Zhang, L., B. S. Gardiner, et al. (2007). "The effect of cyclic deformation and solute binding on solute transport in cartilage." Arch Biochem Biophys 457(1): 47-56.
Zhang, L., B. S. Gardiner, et al. (2008). "A fully coupled poroelastic reactive-transport model of cartilage." Mol Cell Biomech 5(2): 133-153.
Zheng, M. H., J. Chen, et al. (2005). "Porcine small intestine submucosa (SIS) is not an acellular collagenous matrix and contains porcine DNA: possible implications in human implantation." J Biomed Mater Res B Appl Biomater 73(1): 61-67.

225
APPENDIX

Bioreactor Design for Tendon/Ligament Engineering
Tao Wang, BS,1 Bruce S. Gardiner, PhD,2 Zhen Lin, PhD,1,3 Jonas Rubenson, PhD,4
Thomas B. Kirk, PhD,5 Allan Wang, PhD,6 Jiake Xu, PhD,7 David W. Smith, PhD,2
David G. Lloyd, PhD,4,8 and Ming H. Zheng, MD, PhD, FRCPath, FRCPA1
Tendon and ligament injury is a worldwide health problem, but the treatment options remain limited. Tendonand ligament engineering might provide an alternative tissue source for the surgical replacement of injuredtendon. A bioreactor provides a controllable environment enabling the systematic study of specific biological,biochemical, and biomechanical requirements to design and manufacture engineered tendon/ligament tissue.Furthermore, the tendon/ligament bioreactor system can provide a suitable culture environment, which mimicsthe dynamics of the in vivo environment for tendon/ligament maturation. For clinical settings, bioreactors alsohave the advantages of less-contamination risk, high reproducibility of cell propagation by minimizing manualoperation, and a consistent end product. In this review, we identify the key components, design preferences, andcriteria that are required for the development of an ideal bioreactor for engineering tendons and ligaments.
Introduction
Tendons and ligaments have an important function intransferring force from muscle to bone or bone to bone.
Tendons also help store elastic energy while walking, in-creasing locomotion efficiency. However, sudden, excessivestrain of ligaments and tendons from athletic or recreationalactivities can cause acute traumatic injury to these tissues,which can range from small, partial tears to complete rup-tures. Furthermore, repetitive loadings over long periods oftime can lead to similar tissue injuries, when the fatiguedamage within the tendon exceeds the capacity of the tissueto repair itself. In the United States, injuries to tendons andligaments represent about half of the 33 million musculo-skeletal injuries.1 Each year more than 33,000 tendon recon-structions occur in the United States, costing $30 billionUSD2,3 and in Australia, $250 million is spent annually juston rotator cuff repair.4 However, despite the high prevalenceof tendon injury and associated tendinopathy worldwide,treatment options remain poorly defined.
Autograft and allograft transplantations are the commonsurgical treatments for tendon and ligaments that are injured ordegenerated. However, the risks of damage to the donor sitefrom which the autografts are taken, and the potential immune
reactions for allografts are the major concerns.5–7 A promisingtranslational approach to the treatment of tendon/ligamentinjury or degeneration is through the use of engineered autol-ogous grafts made available through the development of bio-reactors that generate tendon/ligament tissue in vitro. Onecommon view is that the key to a successful bioreactor is beingable to recreate, in vitro, the cell microenvironments that areexperienced by cells in vivo. The cell microenvironments can bedefined using cellular morphological information with datafrom molecular biology, biochemistry, and biomechanics. Thisreview aims to clarify the requirements for a successful biore-actor that may be used for tendon/ligament engineering, andto provide an overview of the range of components found intendon/ligament bioreactors, including custom-made andcommercial products. We will also discuss the studies that haveinvolved the application of tendon/ligament bioreactors.
Key Elements for Tendon/Ligament Formationand Regeneration
Tendons and ligaments are force-transferring tissues frommuscle to bone and bone to bone, respectively. They consistof collagens, cells, proteoglycans, elastin, glycolipids, andwater. Although roughly 65%–70% of the total weight is
1Centre for Orthopaedic Translational Research, School of Surgery, University of Western Australia, Crawley, Australia.2School of Computer Science and Software Engineering, University of Western Australia, Crawley, Australia.3Division of Orthopaedic Surgery, Department of Surgery, Guangdong General Hospital, Guangdong Academy of Medical Sciences,
Guangzhou, Guangdong, China.4School of Sport Science, Exercise and Health, University of Western Australia, Crawley, Australia.5Department of Mechanical Engineering, Curtin University, Bentley, Australia.6Sir Charles Gairdner Hospital, Perth, Australia.7School of Pathology and Laboratory Medicine, University of Western Australia, Crawley, Australia.8Centre for Musculoskeletal Research, Griffith Health Institute, Griffith University, Gold Coast, Australia.
TISSUE ENGINEERING: Part BVolume 19, Number 2, 2013ª Mary Ann Liebert, Inc.DOI: 10.1089/ten.teb.2012.0295
133
226

water,8 tendon is a highly organized structure. Collagen typeI is the main structural/functional component and comprisesaround 70%–80% of the dry weight. Type III collagen ismainly present in the endotenon and epitenon, but is alsopresent in the early phase of tendon repair.9,10 Tendons andligaments are relatively hypovascular and hypocellular tis-sues, with their cells (tenocytes and fibroblasts) comprising< 5% of the total volume. The morphology of tenocytes andfibroblasts is sharp and usually elongated along collagen fi-bers in normal tissues, but some rounded tenoblasts are foundoccasionally.11–14 Based on the composition and function oftendon/ligament tissue, one must consider the four basic el-ements for their successful regeneration: the cell source, thecharacteristics of the scaffold matrix, and establishing an ap-propriate chemical and physical cellular microenvironment.
An ideal cell source should meet the following three re-quirements: availability, rapid proliferation, and the abilityto differentiate into in situ cells.15 Stem cells, dermal fibro-blasts, and tenocytes have all been tested as potential cellsources for tendon repair. In the past decade, promising re-sults have been achieved using mesenchymal stem cells(MSCs) in tendon/ligament engineering.16–23 However, thepotential for MSCs to also differentiate into osteoblastic cellsleading to ectopic bone formation raises concerns for theirapplication in tendon repair. Likewise, bone marrow-derivedMSCs that have been used in various studies24–28 also pose apotential risk of ectopic bone formation.29 A recent studydemonstrated that embryonic stem cells (ESCs) can be in-duced to exhibit a tenocyte-like morphology and expresstendon-related gene markers if exposed to an appropriatebiomechanical environment.2 Unfortunately, the clinical ap-plication of ESCs is restricted due to their limited availability(depending on the policy of different countries) and thecomplexity of cell manipulation required. Dermal fibroblastshave also been shown to form tendon tissue,30–32 althoughfurther research has suggested that the healing process usingskin-derived fibroblasts is suppressed with a lack of tenocytemarkers and histopathologic correlations.33,34 Being nativecell sources, tenocytes and in situ fibroblasts are perhaps themost ideal cell sources for engineered tendon and ligamenttissue, respectively. Preclinical and early clinical studies us-ing these native cell sources are promising.35–42 In the mostrecent clinical trial of autologous tenocyte therapy, a total of25 patients with recalcitrant lateral epicondylitis were treatedwith fibroblasts isolated from biopsied anterior cruciate lig-ament tissue using a patented technique developed by ourlaboratory. We found a 60% or greater improvement in boththe Quick DASH and pain assessment scores.43,44
Apart from the selection of the cell source, cell seeding alsoplays an essential role in the development of engineeredtendon and ligament. Several reports indicate that sufficientcell number and a uniform distribution throughout thescaffold are desirable for achieving a homogeneous extra-cellular matrix (ECM) deposition in vitro.45,46 Compared tolower initial cell-seeding density, high seeding density hasbeen shown to result in increasing ECM deposition rate, ahigher final cell number and better cellular morphology.47,48
However, overseeding also has potential for negative effectson nutrient delivery, and consequently cellular metabolismand cell viability. Nutrient depletion at high cell-seedingdensities could lead to spatially in homogenous ECM pro-duction.49 A study by Issa et al. showed that mechan-
ostimulated human umbilical veins seeded with 3 millioncells/mL had better cellular proliferation rates than othergroups.28 However, optimal seeding density will vary de-pending on the cell source and the bioscaffold physical andchemical properties, as these properties will affect nutrienttransport and rates of consumption, as well as the mechan-ical and chemical stimuli provided to the attached cells.49
The construct scaffold plays on important role in engi-neering the new tendon/ligament tissue. Ideally, the scaffoldshould be able to provide substantial initial mechanicalstrength for its immediate postimplantation functional role,while providing a suitable biological environment for cellmigration and proliferation. Furthermore, the degradationrate of the biomaterial needs to be comparable with the rateof tissue synthesis, to allow the eventual replacement of thestarting scaffold with neotissue.
Both synthetic and natural biomaterials are commonlyused in tendon/ligament engineering. Synthetic polymerscaffolds have the advantage of reproducible mechanical andchemical properties, and they are relatively easy to fabricateinto different sizes.24,25,40,41,50 However, their rapid degra-dation rate and potential risk of releasing acidic byproductsor toxic polyesters during degradation have limited theirapplication in clinical trials. Given these disadvantages, moreresearchers have turned their focus on exploring naturalbiomaterials.19–21,37,51–54 Being the main component of nativetendon, collagen type I is the most obvious choice of mate-rial. Although the biocompatibility is excellent, the poormechanical properties of reconstituted type I collagen scaf-folds has limited their further development as a load-bearingmaterial. Silk fibroin, on the other hand, has similar bio-compatibility as collagen scaffolds and comparable me-chanical properties as native tendon/ligament. In severalin vivo studies, silk fibroin-based engineered ligaments havebeen proven their ability to restore the function of injuredligament.16,17,26 Another option is the decellularised tendon/ligament construct. Although the mechanical and biologicalproperties are a better match to native tissue than any othercurrently available scaffolds, donor cells may remain in theallograft, even with strict sterilization and cleaning, and thusthey can potentially cause inflammatory responses.55,56
After choosing an appropriate cell source and scaffold type,the tissue needs to be encouraged to develop the properties ofnative tissue by providing an appropriate biochemical andbiomechanical environment to stimulate ECM synthesis. Re-garding the biochemical environment, several growth factorshave been found to play an important role in tendon/ligamentformation and healing. These include Insulin-like growth fac-tor-I, vascular endothelial growth factor, platelet-derivedgrowth factor, basic fibroblast growth factor, transforminggrowth factor (TGFb), and growth differentiation factor 5(GDF-5). The roles of TGFb and GDF-5 seem to be particularlyprominent. TGFb remains active throughout tendon/ligamenthealing,57 and is able to regulate cell migration, proteinaseexpression, fibronectin-binding interactions, cell proliferation,and collagen production.57–60 Recent studies demonstratedthat tendon and ligament formation was impaired in TGFb2and TGFb3 knockout mouse embryos, reinforce the importanceof TGFbs in tendon development and homeostasis.61 GDF-5regulates cell growth and differentiation, with a lack of GDF-5causing delayed tendon healing, irregular collagen type I fi-brils, and weakened fibril mechanical properties.62–64
134 WANG ET AL.
227

How these various biochemical factors should be intro-duced into the tissue bioreactor system to shape the chemicalenvironment is an extremely challenging and open question.Specifically, at what concentrations, in what combinations,and at what sequence or timing should they be made avail-able? Studies have shown that the expression of these factorsin tendon repair changes over periods of days.65–67 Pre-sumably, as the engineered tissue progresses, cells begin tocontrol their own biochemical environment, and the role of thebioreactor is to now provide the building block nutrients andexpected systemic signals, along with a mechanical stimulus.The early stage in the tissue-engineered tendon is likely to bethe most critical in establishing tendon development along apathway to resemble native tendon. Finding the correctcombination of factors is daunting due to the complexityarising from the multitude of possible combinations and in-teractions. Systematically, varying experimental conditions,coupled with computational modeling of transport processesand signaling molecule pathways leading to cell responses,provide the only conceivable means to both understandingand efficiently optimizing the tissue bioreactor system.
Tendon’s primary function is mechanical. It operates in avarying load environment, both on short timescales (e.g.,walking and running) and on longer timescales (e.g., changes inbody size with age). Tendon responds to its mechanical envi-ronment through changes in ECM biosynthesis and degrada-tion. Unsurprisingly, given its functional role, a suitablemechanical stimulus is vital for tendon/ligament homeostasis.In fact, it has been shown that after 4 weeks in a load-free cultureenvironment, tenocytes lose their native elongated morphology,become increasingly rounded, and the collagen fiber becomesmore crimped.68 In cell-free reconstructed collagen fibril net-work systems, a tensile load has been shown to be protective todegradation by MMP-8.69 Furthermore, cyclic stretching hasbeen shown to produce an up to nine-fold increase in the cellnumber of an engineered tendon compared with a static cultureover a 2-week period.23 Finally, an appropriate mechanical en-vironment could help guide collagen fiber formation, that is,along the direction of loading, which is able to enhance or op-timize the mechanical properties, including stiffness, elasticmodulus, maximum tensile stress, and maximum force.19,31,36
Rather than being two separate signals, there is a crosstalkbetween mechanical and chemical signals. Recent studieshave shown that gradual and temporary loss of tensileloading leads to reversible loss of Scleraxis (Scx) expression,which is a transcription factor specific for tenocytes and theirprogenitors. In addition, it has been shown that TGFb di-rectly induced the expression of Scx in cultured tenocytesisolated from mice.70 In Scx - / - mice, a disordered limbtendon phenotype was observed,71 and a similar phenome-non happened in TGFb type II receptor gene knockout(Tgfbr2 - / - ) mice with dramatic loss of Scx expression.61
Providing a suitable biomechanical signal is clearly animportant component for the success of tendon/ligamentengineering. Generating a suitable mechanical signal withinthe bioreactor system is critically important for tendon/lig-ament tissue engineering.
Bioreactor Design for Tendon/Ligament Engineering
Despite the increasing appreciation of tendon/ligamentbiology and function, conventional culture methods do not
seem to meet the biochemical and biomechanical require-ments to generate bioengineered tendon/ligament in vitro. Abioreactor system that subjects the cell culture to dynamicloading, mimicking the physiological conditions of tendon/ligament in vivo, while allowing cellular proliferation, dif-ferentiation, and matrix production in a mechanical envi-ronment, may provide a solution.
Bioreactors for tendon/ligament engineering are differentto the systems that have been used in various other tissue-engineering fields in the past decades, that is, systems for themuscle,72 liver,73 and bone.74,75 Compared to other bioreac-tors, the main task of the bioreactor for tendon/ligamentengineering is to provide the proper biomechanical andbiochemical environment specific to tendon/ligament for-mation. To achieve this, certain basic components are re-quired, that is, the actuating system and the culture chamber,which can provide the construct’s mechanical stimulationand controlled culture environment, respectively. Further-more, the bioreactor may also include a medium circulationsystem, monitoring system, feedback system, and a mediumanalysis system, depending on the operational requirements(Fig. 1). With these facts in mind, several custom-made bio-reactors have been developed for tendon/ligament engi-neering (see Table 1), and the aforementioned components ofthese are now are discussed in detail.
Actuator and Culture Chamber Design
The actuating system is the main component for providingdifferent mechanical stimulation to engineered tissue.Pneumatic actuators,19–21 linear motors,22,27 and step motor-ball screws (SMBSs)2,42,76 are the most common actuatorsused in tendon/ligament bioreactors. Pneumatic actuatorshave several advantages, including the ease of maintenance,cleanliness, low cost, and high power-to-weight ratio.77 Un-fortunately, by using air as a medium, pneumatic actuatorsare subject to high friction. The sensitivity and response to aninput signal are relatively slow because of the dead band anddead time caused by stiction and air compressibility. Due tothese nonlinearities, it is difficult to achieve accurate positioncontrol with pneumatic actuators.77 Typically, the accuracyof pneumatic actuators is *– 0.1 mm, which is not insignif-icant when typical tendon bioreactors require < 5% strain on1–5-cm tissues. Compared with pneumatic actuators, elec-trical actuators are more expensive, but the level of accuracyin their positional control is much higher. A direct-drivelinear motor is able to provide high-speed/high-accuracylinear motion by eliminating mechanical transmission,78–80
and the accuracy is the highest of all three actuators, that is,*– 1mm. SMBS transmission systems are based on a ball-screw linkage and a crank-slider mechanism. In general,SMBSs transfer the rotation of the crank to a reciprocatingmotion of the screw.81 By choosing different ball screws, theoptimal speed range and output force can be selected usingEquation (1), that is,
T¼ Fl
2p�(1)
where T is torque applied to screw; F is linear force; l is ballscrew lead; and n is ball screw efficiency. Although SMBS isnot as accurate as a linear motor, a positional accuracy ofaround – 5mm can still be achieved. In a multichamber-shared
BIOREACTOR DESIGN FOR TENDON/LIGAMENT ENGINEERING 135
228

loading system, SMBS is widely used given its high accuracyand relative high loading capacity.35,42,76 In independentloading multichamber systems, linear motors are morepopular due to their small size and relatively simple me-chanical arrangement.22,82
In the addition to the actuator, the connection between themechanical input and the tissue is vital and often presents amajor challenge. During dynamic loading of the tissue inculture, an even distribution of force throughout the entiresample is critical, otherwise tissue integrity is compromisedby overloading the mechanical connection regions and/or byinhomogeneous mechanical stimulation. Different strategiesof applying mechanical loads have been adopted based ondifferent construct dimensions. In a study by Chen et al., cell-seeded knitted silk–collagen sponge scaffolds were fixed onstainless rings and connected to sample hooks.2 The biore-actor system by Juncosa-Melvin et al. applied two posts to fixthe construct, punching through the scaffold as shownin Figure 2.19 However, nonuniform construct deformationis clearly apparent. Tensile force is focused on the side ofconstructs, which causes uneven distribution of mechanical
stimulation. Although tissue clamps are the most popularand relatively effective method for holding tendon con-tructs,22,42,82 the potential for damage at the clamping regionneeds to be considered. The method of reproducibly ap-plying uniform loads to soft tissue without tissue damageor slippage is a critical problem in need of a satisfactorysolution. It is our opinion that a robust clamping regionshould be designed along with the artificial bioscaffold toensure the proper connection between the sample and themechanical load.
The culture chamber is an essential part of whole biore-actor system. The high humidity of culturing conditions(99% humidity, 37�C, and 5% CO2) and chemistry of theculture medium are corrosive to many materials. Corrosionproducts may in turn be toxic to the tissue. Therefore,noncorrosive and autoclavable materials such as stainlesssteel, polymethylemethacrylate, polyoxymethylene, poly-carbonate, glass, and silicon are preferred and widely used inculture chamber design.2,23,27,76,83
The chamber structure is an important consideration in thebioreactor design, with most currently available culture
FIG. 1. Schematicdemonstration of connectionbetween differentcomponents of tendon/ligament bioreactor system.Color images available onlineat www.liebertpub.com/teb
136 WANG ET AL.
229

chambers divided into two groups: integrated42,83 and sep-arated chambers.2,3,19–27,35,76,82 In integrated chambers, mul-tiple samples are cultured while sharing the same culturemedium. Conversely, separated chambers can provide sep-arate culture environments for each sample. Although thecomplexity of design and manufacturing costs may be higherin a separated chamber system, reduced cross-contaminationand the option of independent environmental control aredistinct advantages.
During tissue culture, sufficient air exchange within theculture chamber is critical. Air exchange in conventional cellculture incubators is through the integrated hydrophobicfilter of the culture flask and the gap between the leak andthe culture dish/well plate. Therefore, the ideal design forthe culture chamber should be similar. Like conventional cellculture, an unsealed chamber bioreactor connects to theoutside environment through various ways,19,22,27,35 such asthe hydrophobic filter leak42 and the labyrinth channel (Fig.3).82 In a study by Webb et al.,42 a modified tissue cultureflask was used as an integrated culture chamber, and could
culture up to four samples simultaneously. Although thereare potential risks of cross-contamination from differentsamples and toxicity from autoclaving the culture flask, theintegrated hydrophobic filter leak can ensure adequate gasexchange without inducing contamination. In the bioreactorsystem used in Parent et al.’s research,82 a labyrinth channelwas added to improve the air exchange and eliminate con-tamination as shown as Figure 3. However, in closedchambers, air exchange mostly depends on medium circu-lation, which is now discussed in the following section.
Environmental Control and MediumCirculation Systems
Although to the best of our knowledge, no contaminationhas been reported in any bioreactor study, transportation ofthe bioreactor and opening of chamber for medium exchangeevery 3 days, especially for multichamber systems, are still apotential contamination risk. Therefore, a medium circula-tion system can be introduced to improve the efficiency andminimize these risks. In addition to the advantages of re-duced contamination, a circulating medium may be betterable to infiltrate into cultured tissue. For example, perfusionbioreactors used in bone engineering circulate the culturemedium for better nutrient delivery and subsequent im-proved cell numbers.84–89 Similarly, human umbilical veincultured under a circulating medium had approximatelythree times the cellular number as those cultured using aquiescent medium.23
FIG. 2. Stem cell-seeded Collagen sponge deformationduring mechanical stimulation. Modified from reference 19.Color images available online at www.liebertpub.com/teb
Table 1. Components of Custom-Made Bioreactor Systems
ReferenceActuating
system Culture chamber Monitor systemFeedbacksystem
Mediumcirculation system
Mediumanalysis system
76 Biaxial Multiple · · X ·42 Uniaxial Single chamber, multiple
samples· · · ·
19,20,21,3 Uniaxial Multiple Displacement · · ·35 Uniaxial Multiple Force · · ·22 Uniaxial Single Force · · ·23 Uniaxial Multiple · · X ·83 Uniaxial Single chamber, multiple
samplesDisplacement Force · · X
2 Uniaxial Multiple · · · ·27 Uniaxial Multiple Displacement · · ·82 Uniaxial Multiple Displacement Force X · ·
FIG. 3. Schematic drawings of air exchange through thelabyrinth channel in the culture chamber. Modified fromreference 82. Color images available online at www.liebertpub.com/teb
BIOREACTOR DESIGN FOR TENDON/LIGAMENT ENGINEERING 137
230

A traditional incubator-based bioreactor and/or inde-pendent bioreactor can be used for tendon/ligament engi-neering. In an incubator bioreactor system, air exchange isthrough the hydrophobic filter leak of the medium reservoir,and proper CO2 and temperature level is controlled by theincubator. Then, by circulating the culture medium, suitableconditions can be applied to engineered tendon/ligament, asshown in Figure 4.23 However, an independent bioreactorsystem, as shown Figure 5, does not rely on an incubator tocontrol the culture environment. The percentage of differentgases (PO2, CO2, and N2) are controlled by air valves,76 andthe mixed gas is humidified first, and then passed into amedium heater. Waste gas emission is transported through afilter in the case of contamination. The prepared, warm cul-ture medium is circulated through the bioreactor culturechamber. For certain tissues such as cartilage, some specificculture conditions are required. For example, under a low-oxygen environment, engineered cartilage displays fastermatrix glycosaminoglycan deposition rate and better cellularmorphology, but with less dedifferentiation.90–92 The envi-ronmental chamber allows researchers to manipulate differ-ent culture conditions, thereby enabling a systematic studyof cell growth and differentiation into functional tissue.
Monitoring and Feedback Systems
The biomechanical properties of the final engineered ten-don/ligament should be a central concern of bioreactor de-sign, as the tendon/ligament’s functional role in the body isprimarily mechanical. The maximum load and elastic mod-
ulus are essential mechanical properties to evaluate thesuitability of engineered tendon/ligament. However, in moststudies, mechanical tests are performed only at the end oftissue culture. For example, the stiffness of the constructs atdifferent time points during culture is rarely recorded orassessed. The correlation between stiffness and tissue mat-uration may provide a better understanding about how thecell differentiates into functional tissue, and for this reason,online stiffness monitoring is likely to be invaluable.
In tendon/ligament bioreactors, force measurement isenabled using sensors called load cells,22,82,93 which are lo-cated differently in various bioreactor designs. Load cellslocated between the actuator and sample clamps,83 shown inFigure 6A, requires high manufacturing accuracy, as thefriction between the shaft and culture chamber can induceerror. For fragile materials such as biological tissues, thisfriction might be higher than the applied load. Conversely, aload cell placed at the end of the culture chamber, shown inFigure 6B, can minimize the fiction between the actuator andculture chamber and also the fluid resistance during stimu-lation. To acquire accurate data, load cell selection should bebased on the initial mechanical properties of the constructs.Working with the sensitivity of the load cell, extra attentionis needed when manipulating the construct so as to avoidcausing damage to the load cell through overloading.
In addition to force monitoring, tissue displacement isanother important variable that requires monitoring. Linearvariable differential transformers and optical decoders arecommonly used position sensors.19,82 Although the motionof actuating systems is preprogrammed, overload of theactuator and manual misoperation can cause desynchroni-zation between the program and actual stimulation. The real-time displacement monitor can produce a full record ofstimulation position, which then allows researchers to trackif there are any unusual features in the results. Anotherfunction of the position sensor is to provide feedback of thedisplacement information to the actuator control system tocorrect for any desynchronization.82 Critically, by monitor-ing load and displacement, real-time stiffness during en-gineered tendon/ligament culture can be estimated.
Lastly, imaging the construct might be a potentially im-portant monitoring method that can be adopted in bioreactorsystems. Two imaging modalities, the confocal microscopeand optical coherence tomography, are potential candi-dates.94,95 However, sample deformation induced by me-chanical stimulation can cause image shift resulting in poorfocus or loss of image. Imaging dynamic sample is extremelydifficult in conventional bioreactors, as features of interest(e.g., cells) soon leave the field of view. Techniques need tobe developed to move the sample and microscope together.This can become difficult, especially, when doing in situimaging of a tendon tissue culture.
Commercial Bioreactor Systems for Tendon/Ligament Engineering
Recently, commercial tendon-/ligament-engineering bio-reactor systems have become available. As discussed above,these basic principles are applied to customized bioreactors;however, with greater design input and advancedmanufacturing techniques, commercial products are ableto provide more accurate and complex environments for
FIG. 4. Schematic illustration of an incubator bioreactorsystem. Suitable temperature, humidity, and CO2 level of theculture medium are maintained by the incubator in the me-dium reservoir. The medium is circulated by a pump. Thewaste valve is closed normally, and it will open duringmedium exchange. Color images available online at www.liebertpub.com/teb
138 WANG ET AL.
231

FIG. 5. Schematic diagram of an independent bioreactor system. Suitable temperature, humidity, and CO2 level of theculture medium are controlled by the environmental chamber. Cold culture medium is pumped to the environmentalchamber for heating and then circulated through the culture chamber. The waste valve is closed normally, and it will openduring medium exchange. Color images available online at www.liebertpub.com/teb
FIG. 6. Load cell position indifferent bioreactor systems. (A)Load cell is located between theactuator and sample clamps. (B) Aseparate load cell placed at the endof culture chamber system. Colorimages available online at www.liebertpub.com/teb
BIOREACTOR DESIGN FOR TENDON/LIGAMENT ENGINEERING 139
232

Ta
ble
2.S
tu
dies
Usin
gB
io
rea
cto
rS
ystem
sfo
rT
en
do
n/
Lig
am
en
tE
ng
in
eer
in
g
Fir
stau
thor
Bio
reac
tor
type
(Com
pan
y)
Par
amet
ers
ofm
echan
ical
stim
ula
tion
Sca
ffol
dm
ater
ial
(dim
ensi
ons)
Cel
lso
urc
eE
ffec
tR
efer
ence
Alt
man
(200
2)C
ust
om
-mad
est
epm
oto
rb
iore
acto
rw
ith
env
iro
nm
enta
lch
amb
erC
ycl
icst
retc
hin
g2-
mm
,90
o
rota
tio
n,
0.01
67H
z,21
day
sC
oll
agen
typ
eI
gel
s,B
omby
xm
ori
silk
wo
rmsi
lkfi
ber
mat
rice
s(l
eng
th:3
0m
m)
Hu
man
bo
ne
mar
row
stro
mal
cell
s(h
BM
SC
)
Elo
ng
atio
no
fh
BM
SC
s,cr
oss
-sec
tio
nce
lld
ensi
ty[
76
Jun
cosa
-Mel
vin
(200
6)C
ust
om
-mad
ep
neu
mat
iccy
lin
der
bio
reac
tor
wit
hL
VD
Tfo
rd
isp
lace
men
tm
on
ito
rin
g
Cy
clic
stre
tch
ing
2.4%
stra
in,
0.00
33H
z,8
h/
day
for
2w
eek
sT
yp
eI
coll
agen
spo
ng
e(2
3–
0.8
mm
·9
–0.
8m
m·
3–
0.1
mm
)
Rab
bit
MS
Cs
Max
imu
mfo
rce[
,li
nea
rst
iffn
ess[
,m
axim
um
stre
ss[
,li
nea
rm
od
ulu
s[
19
Web
b(2
006)
Cu
sto
m-m
ade
step
mo
tor
bio
reac
tor
Cy
clic
stre
tch
ing
10%
stra
in,
0.25
Hz,
8h
/d
ayfo
r7
day
sP
oly
ure
than
eco
nst
ruct
(20
·10
·2
mm
)H
um
antr
ach
eal
fib
rob
last
sT
yp
eI
coll
agen
[,
TG
Fb-
1[,
CT
GF
,E
last
in[
,al
ph
aI[
,P
roco
llag
en[
,F
ibro
nec
tin
[,
MM
P-1
[,
elas
tic
mo
du
lus[
42
An
dro
jna
(200
7)C
ust
om
-mad
eb
iore
acto
rw
ith
load
-d
isp
lace
men
tm
easu
resy
stem
Cy
clic
stre
tch
ing
9%st
rain
,tw
ice
dai
lyfo
r30
min
each
per
iod
sep
arat
edb
y8-
hre
stfo
r2
wee
ks
Sm
all-
inte
stin
esu
bm
uco
sa(3
·5
·95
mm)
Do
gte
no
cyte
sC
ell
den
sity
[,
stif
fnes
s[35
Nir
mal
anan
dh
an(2
008)
Cu
sto
m-m
ade
pn
eum
atic
cyli
nd
erb
iore
acto
rC
ycl
icst
retc
hin
g2.
4%an
d1.
2%st
rain
,1
Hz,
stim
ula
tio
np
erio
d:
8h
/d
ay,
100
and
3000
cycl
es/
day
for
12d
ays
Ty
pe
Ico
llag
ensp
on
ge
Rab
bit
ilia
c-cr
est
MS
Cs
Th
est
imu
lati
on
pat
tern
of
2.4%
stra
in,
3000
cycl
es/
day
,1
Hz
has
the
bes
tef
fect
on
incr
easi
ng
stif
fnes
s.
21
Nir
mal
anan
dh
an(2
008)
Cu
sto
m-m
ade
pn
eum
atic
cyli
nd
erb
iore
acto
rC
ycl
icst
retc
hin
g2.
4%st
rain
,0.
0033
Hz,
8h
/d
ayfo
r12
day
sT
yp
eI
pu
rifi
edb
ov
ine
coll
agen
gel
,ty
pe
Ico
llag
ensp
on
ges
(len
gth
:11
and
51m
m)
Rab
bit
MS
Cs
Sti
ffn
ess[
20
Ng
uy
en(2
009)
Cu
sto
m-m
ade
lin
ear
actu
ato
rb
iore
acto
rw
ith
load
cell
for
forc
em
easu
rem
ent
Pre
load
edw
ith
0.05
N,
cycl
icst
retc
hin
g10
%,
0.5
Hz,
2h
stim
ula
tio
n—
2h
rest
—2
hst
imu
lati
on
—18
hre
stfo
r5
day
s
Po
rcin
esm
all
inte
stin
esu
bm
uco
sa(l
eng
th:
2cm
and
wid
th:
1cm
)
Rab
bit
MC
Lfi
bro
bla
sts
Imp
rov
edfi
ber
ori
enta
tio
n,
fib
eran
gu
lar
dis
per
sio
nY
,b
ette
ro
rgan
ized
coll
agen
fib
er,
elo
ng
ated
cell
mo
rph
olo
gy
,
22
Ab
ou
slei
man
(200
9)C
ust
om
-mad
eli
nea
rac
tuat
or
bio
reac
tor
wit
hm
ediu
mci
rcu
lati
on
syst
em
Cy
clic
stre
tch
ing
2%st
rain
,1
h/
day
,0.
0167
Hz
for
1an
d2
wee
ks.
Hu
man
um
bil
ical
vei
ns
(wal
lth
ick
nes
s:0.
75m
m,
ou
ter
dia
met
er:
6.75
–0.
25m
m,
len
gth
:8.
5cm
)
Wis
tar
Rat
bo
ne
mar
row
MS
Cs
bet
ter
cell
dis
trib
uti
on
,m
ore
elo
ng
ated
cell
mo
rph
olo
gy
,ce
llp
roli
fera
tio
n[
,u
ltim
ate
stre
ss[
,el
asti
cm
od
ulu
s[
23
Bu
tler
2009
)C
ust
om
-mad
ep
neu
mat
iccy
lin
der
bio
reac
tor
wit
hL
VD
Tfo
rd
isp
lace
men
tm
on
ito
rin
g
Cy
clic
stre
tch
ing
2.4%
stra
in,
0.00
33H
z,8
h/
day
for
2w
eek
sT
yp
eI
coll
agen
spo
ng
e(9
4%p
ore
vo
lum
e;62
-mm
mea
np
ore
dia
met
er)
Mo
use
mes
ench
ym
alst
emce
llT
yp
eI
coll
agen
[,
lin
ear
stif
fnes
s[83
(con
tin
ued
)
140
233

Ta
bl
e2.
(Co
nt
in
ue
d)
Fir
stau
thor
Bio
reac
tor
type
(Com
pan
y)
Par
amet
ers
ofm
echan
ical
stim
ula
tion
Sca
ffol
dm
ater
ial
(dim
ensi
ons)
Cel
lso
urc
eE
ffec
tR
efer
ence
Ch
en(2
010)
Cu
sto
m-m
ade
step
mo
tor
bio
reac
tor
Cy
clic
stre
tch
ing
10%
stra
in,
2h
/d
ay,
1H
zfo
r14
day
sK
nit
ted
silk
-co
llag
ensp
on
ge
scaf
fold
(5·
0.5
·0.
2cm
)H
um
anem
bry
on
icst
emce
llC
oll
agen
I[,
Co
llag
enII
I[,
Ep
ha4
[,
Scx
[,
So
x9Y
,M
yo
sin
[,
Inte
gri
na1
[,
Inte
gri
na2
[,
Inte
gri
nb1
[,
Co
llag
enco
nte
nt[
,C
oll
agen
dia
met
er[
,b
ette
rce
llal
ign
men
t
2
Do
rosk
i(2
010)
Cu
sto
m-m
ade
lin
ear
mo
tor
bio
reac
tor
Cy
clic
stre
tch
ing
10%
stra
in(5
%o
ffse
t,5%
amp
litu
de)
,1
Hz,
3h
/d
ay,
1.7,
14,
and
21d
ays
Po
ly(e
thy
len
eg
lyco
l)-b
ased
hy
dro
gel
mat
eria
lo
lig
o(p
oly
(eth
yle
ne
gly
col)
fum
arat
e)(1
2.5
·9.
5·
1.6
mm
)
MS
Cs
(PT
-251
0;L
on
za)
Co
llag
enI[
,C
oll
agen
III[
,T
NC
[,
Ten
asci
n-C
[27
Sab
er(2
010)
Lig
agen
L30
-4C
(Tis
sue
Gro
wth
Tec
hn
olo
gie
s)C
ycl
icst
retc
hin
g1.
25N
,1
cycl
e/m
in,
1h
/d
ayfo
r5
day
sA
cell
ula
rra
bb
ith
ind
paw
ten
do
n(l
eng
th:
5cm
)R
abb
itte
no
cyte
sU
ltim
ate
ten
sile
stre
ss[
(clo
sed
tofl
esh
lyh
arv
este
dte
nd
on
),el
asti
cm
od
ulu
s[
36
Issa
(201
1)C
ust
om
-mad
eb
iore
acto
rC
ycl
icst
retc
hin
g2%
stra
in,
1h
/d
ay,
0.01
67H
zfo
r1
and
2w
eek
s.H
um
anu
mb
ilic
alv
ein
s(w
all
thic
kn
ess:
0.41
mm
)w
ith
Wh
arto
n’s
jell
ym
atri
xas
cen
tral
po
rtio
n(t
ota
lth
ick
nes
s:0.
75m
m)
Wis
tar
rat
bo
ne
mar
row
MS
Cs
Cel
lp
roli
fera
tio
n[
,te
nsi
lest
ren
gth
[28
Wo
on
(201
1)L
igag
enL
30-1
C&
Lig
agen
L30
-4C
(Tis
sue
Gro
wth
Tec
hn
olo
gie
s)L
30-1
C:
dy
nam
iclo
adin
g,
10N
,1
h/
day
,0.
0167
Hz
for
5d
ays
L30
-4C
:d
yn
amic
load
ing
,0.
625
N1.
25N
2.5
N,
1h
/d
ay,
0.01
67H
zfo
r3,
5,an
d8
day
s
Ace
llu
lar
hu
man
flex
or
ten
do
nsc
affo
lds
Hu
man
der
mal
fib
rob
last
sU
ltim
ate
ten
sile
stre
ss[
,el
asti
cm
od
ulu
s[31
LV
DT
,li
nea
rv
aria
ble
dif
fere
nti
altr
ansf
orm
ers;
[,
up
reg
ula
tio
n;
Y,
do
wn
reg
ula
tio
n.
141
234

tendon/ligament culture. To our knowledge, two relativelycomplete commercial bioreactor systems have been devel-oped recently: The Bose� ElectroForce� BioDynamic� sys-tem and the LigaGen system.
The Bose ElectroForce BioDynamic test instrument provi-des an accurate programmable uniaxial stretching stimula-tion and a controllable medium circulation environment toengineered tendon/ligament, which theoretically can beadjusted to mimic the in vivo biomechanical environment.With a load cell and optional laser micrometer, this biore-actor system is able to monitor the force/strain curve ofengineered tendon/ligament during the culture period. Twodifferent force and displacement ranges are available. Single-chamber and multiple-chamber systems with shared orindependent loading are optional. As there are culturechambers compatible to the Bose testing devices such asElectroForce 3200, biomechanical tests can be done at dif-ferent time points without disruption of the tissue culture(www.bose-electroforce.com).
The LigaGen system is a lightweight (< 3 kg) incubator-compatible bioreactor. It is capable of applying a maximumforce of 40 N to the tissue sample and simulating complex,and presumably more physiologically realistic, loading pat-terns. Two systems are available from LigaGen. L30-1 · is asingle-culture model, which has a 23-mL-internal-volumechamber for single-tissue culture, and the L30-4C is a multi-ple-culture model with an 80-mL chamber for shared dynamicculture on two or four samples. The standard medium circu-lation system can reduce contamination risk during mediumexchange. Rather than being a comprehensive system, thebioreactor is extensible to suit individual needs, by addingvarious components to the (universal) basic model as re-quired. With an accessory tissue-monitoring sensor, this sys-tem is able to achieve real-time measurements of the samplestiffness during culture. If flow control is necessary, extracontrol systems can be installed (www.tissuegrowth.com/).
However, there are some disadvantages in the commercialbioreactors. First, the fixation mechanism to stabilize thetendon tissue in the bioreactor cannot be adjusted. Tissueclamps provided in the commercial bioreactor systems canbecome less effective when it comes to the use of a cylindricalscaffold. Secondly, capacities of the chamber and mechanicalinput are limited, and they can only host small-animal tis-sues. This may restrict clinical development. Moreover, someligaments, such as the anterior cruciate ligament, are not onlysubjected to tensile force, but also to rotational loading, andnone of the commercial bioreactor systems provide the ad-dition of torsional loading. Lastly, full-scale commercialbioreactor systems are expensive, and for most of the time,not all of the functions they provide are commonly used inevery study.
Ideal Bioreactors for Tendon/Ligament Engineering
The ideal bioreactor should be able culture tendon- andligament-like constructs, which are well-organized, cell-seeded assemblies of collagen bundles with mechanical prop-erties functionally similar to the native tissue. Autologouscell-seeded constructs are biologically compatible, and ableto provide mechanical support similar to native tissue, andconsequently, they are widely studied in tendon/ligamentengineering. However, induction of cell-directed collagen
fiber reorganization and assembly of collagen bundles aretwo important impediments to this approach. Mechanicaltensile loads can help provide the necessary signals to cells toincrease collagen synthesis, spatially organize of the collagenalong the primary stress direction, and stabilize collagenfrom collagenase degradation.22,96,97 Moreover, proper me-chanical stimulation can upregulate different proteoglycans,such as decorin, biglycan, fibromodulin, and fibronectin,which help the cells organize the parallel collagen fibrilsforming bundles.42,98–101
The host body is in many ways the ultimate bioreactor forall engineered tissues. The study of Juncosa-Melvin et al.indicated that the maximum force of engineered patellartendons increased more than 3000 times after 2 weeks ofimplantation in rabbits,19 and to date, none of the bioreactorshave been able to accomplish this outcome. Therefore, anideal bioreactor should aim to mimic the dynamic bio-chemical and biophysical environments in vivo. In musculo-skeletal tissue engineering, various bioreactors have beendeveloped. For instance, muscle tissues are not only sub-jected to mechanical stretching, but also able to receive theelectrical impulses to simulate inputs from the central ner-vous system,29 and mechanical and electrical stimulationbioreactors have been developed based on mimicking thein vivo environment.102,103 Compared to muscle, the in vivoenvironment is comparatively less complex in tendon/liga-ment and appears to require only passive mechanical input.
Summarizing, the ideal tendon-/ligament-engineeringbioreactor that enables systemic research should integrate allaforementioned components (Fig. 1). The bioreactor shouldhave culture chambers and an actuating system, but also befitted with a medium circulation system, an environmentalsystem, a monitoring system, a feedback system, and a wastemedium analysis system. This bioreactor should first be ableto provide not only a multiple, suitably sized. and sterilizedchambers for tissue culture, but also accurate and program-mable mechanical stimulation. Tensile strain and rotation areneeded to mimic different in vivo tendon/ligament loads.Second, the circulated medium should infiltrate into tissuebetter than a static medium configuration, and the circulationsystem needs to reduce the risks of contamination duringmedium exchange and drug delivery. Moreover, environ-mental control, such as PO2, CO2, and pH level, allows re-searchers to explore the impact of different cultureconditions on tissue maturation. Third, a monitoring systemshould provide the real-time status of cultured tendon/lig-ament, such as force and displacement, and based on thesedata, adjustment of mechanical stimulation by use of afeedback system. Fourth, through analysis of the waste me-dium, nutrient consumption needs to be evaluated, whichmay enable the changing of the medium base at differentstages as required, rather than fixed, regular medium ex-change every 3 days. Finally and importantly, the best pat-terns of mechanical stimulation for culturing engineeredtendon/ligaments need to be defined.
Investigation of Evidence
Since the first 3D engineered tendon/ligament bioreactorsystem published by Altman et al. in 2002, the effect of me-chanical stimulation on the engineered tendon/ligament hasdrawn a lot of research attention. In the past decade,
142 WANG ET AL.
235

dynamic loading of culture in bioreactor systems has beenproven to have been a significant development in producingan engineered tendon/ligament. Indeed, various studieshave been performed using bioreactor systems and have metwith considerably success, and these are summarized inTable 2. Compared with static culture, the tissue producedusing dynamic mechanical stimulation has a better cellmorphology, including elongated cellular morphology andincreased cell density. The mechanical properties of en-gineered tendon/ligament, such as tensile strength andelastic modulus, are also greatly improved by cyclic loadingof the tissue culture, as is the microstructure of the extra-cellular matrix, such as collagen fiber alignment. Gene ex-pression is also positively influenced by cyclic mechanicalstimulation. For instance, collagen I expression under dy-namic loading is three times higher than static culture in 2weeks.83 Although cyclic stretching has been proven to be aneffective way to stimulate the engineered tendon/ligamentculture, the optimal stimulation pattern is still unknown.Nirmalanandhan et al. revealed that a 2.4% strain cycleconsisting of 3000 cycles per day produced the best linearstiffness in rabbit MSCs seeded in type I collagen sponge.21
However, from the perspective of Butler et al., the stimula-tion pattern should be adjusted based on maturation of theengineered tendon/ligament, with higher dose loading ap-plied at the later stage of tissue culture.83
Conclusion
The goal should be to create a construct with similar mi-crostructure and mechanical properties, as native tissue, us-ing bioscaffolds and autologous tenocytes. However, atendon-like uniorientated collagen structure cannot beachieved without a mechanical stimulus within the cultureenvironment. Traditional culture techniques do not providethis mechanical stimulation. The use of a bioreactor system isable to bridge the gap between in vitro and in vivo systems bycreating suitable biochemical and biomechanical environ-ments. Although it is clearly very difficult to reproduce thein vivo microenvironmental conditions exactly within thebioreactor, the goal is to mimic the in vivo biomechanicalcondition as closely as possible. The essential components ofa tendon-/ligament-engineering bioreactor are the actuatingsystem and culture chamber, which are responsible for themechanical stimulation and providing sterilized environ-ment for tissue culture. For better manipulation of the cultureenvironment, accurate stimulation and more precise report-ing of mechanical maturation of engineered tendon/liga-ment, an environmental control system, medium circulationsystem, monitor, and feedback system need to be included.As bioreactors nowadays are becoming more focused onpreclinical research, using these clinically in the future stillpresents a substantial challenge. Tissue engineering requiressubstantial system optimization to achieve a reproduciblefunctional engineered tendon/ligament consistently. Thisprocess is difficult and expensive in animal models, let alonein humans. However, there are additional problems inmoving from small-animal models to humans related to thephysical size of human tissue samples. Issues of nutrienttransport, cell source, and spatial heterogeneity of scaffoldproperties and cell stimulation become more prominent inthese larger tissues.
However, all the evidence suggests that by using biore-actors as described above, it will be possible to successfullyproduce engineered tendons and ligaments. When this oc-curs, we will be able to manufacture basic multichamberbioreactors with accurate and appropriate patterns of me-chanical stimulation, which are affordable, and have lowmaintenance costs that can be used in commercial settings.
Acknowledgments
This work is supported by the Australia Research CouncilLinkage Grant (LP110100581). We would like to thank ourfellow group members, in particular Ms. Euphemie Landaoand Mr. Robert Day for helpful discussion.
Disclosure Statement
No competing financial interests exist.
References
1. Huang, H.H., Qureshi, A.A., and Biundo, J.J., Jr. Sports andother soft tissue injuries, tendinitis, bursitis, and occupation-related syndromes. Curr Opin Rheumatol 12, 150, 2000.
2. Chen, J.L., Yin, Z., Shen, W.L., Chen, X., Heng, B.C., Zou,X.H., and Ouyang, H.W. Efficacy of hESC-MSCs in knittedsilk-collagen scaffold for tendon tissue engineering andtheir roles. Biomaterials 31, 9438, 2010.
3. Butler, D.L., Gooch, C., Kinneberg, K.R., Boivin, G.P.,Galloway, M.T., Nirmalanandhan, V.S., Shearn, J.T.,Dyment, N.A., and Juncosa-Melvin, N. The use of mesen-chymal stem cells in collagen-based scaffolds for tissue-engineered repair of tendons. Nat Protoc 5, 849, 2010.
4. Chen, J., Xu, J., Wang, A., and Zheng, M. Scaffolds fortendon and ligament repair: review of the efficacy ofcommercial products. Expert Rev Med Devices 6, 61,2009.
5. Cerullo, G., Puddu, G., Gianni, E., Damiani, A., and Pi-gozzi, F. Anterior cruciate ligament patellar tendon recon-struction: it is probably better to leave the tendon defectopen! Knee Surg Sports Traumatol Arthrosc 3, 14, 1995.
6. Coupens, S.D., Yates, C.K., Sheldon, C., and Ward, C.Magnetic resonance imaging evaluation of the patellartendon after use of its central one-third for anterior cruciateligament reconstruction. Am J Sports Med 20, 332, 1992.
7. Harner, C.D., Olson, E., Irrgang, J.J., Silverstein, S., Fu, F.H.,and Silbey, M. Allograft versus autograft anterior cruciateligament reconstruction: 3- to 5-year outcome. Clin OrthopRelat Res 324, 134, 1996.
8. Lin, T.W., Cardenas, L., and Soslowsky, L.J. Biomechanicsof tendon injury and repair. J Biomech 37, 865, 2004.
9. Jarvinen, T.A., Jarvinen, T.L., Kannus, P., Jozsa, L., andJarvinen, M. Collagen fibres of the spontaneously rupturedhuman tendons display decreased thickness and crimpangle. J Orthop Res 22, 1303, 2004.
10. Buckwalter, J.A. and Hunziker, E.B. Orthopaedics. Healingof bones, cartilages, tendons, and ligaments: a new era.Lancet 348 Suppl 2, sII18, 1996.
11. Hildebrand, K.A., Frank, C.B., and Hart, D.A. Gene inter-vention in ligament and tendon: current status, challenges,future directions. Gene Ther 11, 368, 2004.
12. Bray, R.C., Rangayyan, R.M., and Frank, C.B. Normal andhealing ligament vascularity: a quantitative histologicalassessment in the adult rabbit medial collateral ligament. JAnat 188 (Pt 1), 87, 1996.
BIOREACTOR DESIGN FOR TENDON/LIGAMENT ENGINEERING 143
236

13. Lo, I.K., Ou, Y., Rattner, J.P., Hart, D.A., Marchuk, L.L., Frank,C.B., and Rattner, J.B. The cellular networks of normal ovinemedial collateral and anterior cruciate ligaments are not ac-curately recapitulated in scar tissue. J Anat 200, 283, 2002.
14. Manske, P.R. Flexor tendon healing. J Hand Surg Br 13, 237,1988.
15. Arnsdorf, E.J., Jones, L.M., Carter, D.R., and Jacobs, C.R.The periosteum as a cellular source for functional tissueengineering. Tissue Eng Part A 15, 2637, 2009.
16. Fan, H., Liu, H., Wong, E.J., Toh, S.L., and Goh, J.C. In vivostudy of anterior cruciate ligament regeneration usingmesenchymal stem cells and silk scaffold. Biomaterials 29,
3324, 2008.17. Fan, H., Liu, H., Toh, S.L., and Goh, J.C. Anterior cruciate
ligament regeneration using mesenchymal stem cells and silkscaffold in large animal model. Biomaterials 30, 4967, 2009.
18. Garcia-Fuentes, M., Meinel, A.J., Hilbe, M., Meinel, L., andMerkle, H.P. Silk fibroin/hyaluronan scaffolds for humanmesenchymal stem cell culture in tissue engineering. Bio-materials 30, 5068, 2009.
19. Juncosa-Melvin, N., Shearn, J.T., Boivin, G.P., Gooch, C.,Galloway, M.T., West, J.R., Nirmalanandhan, V.S., Bradica,G., and Butler, D.L. Effects of mechanical stimulation on thebiomechanics and histology of stem cell-collagen spongeconstructs for rabbit patellar tendon repair. Tissue Eng 12,
2291, 2006.20. Nirmalanandhan, V.S., Rao, M., Shearn, J.T., Juncosa-
Melvin, N., Gooch, C., and Butler, D.L. Effect of scaffoldmaterial, construct length and mechanical stimulation onthe in vitro stiffness of the engineered tendon construct. JBiomech 41, 822, 2008.
21. Nirmalanandhan, V.S., Shearn, J.T., Juncosa-Melvin, N.,Rao, M., Gooch, C., Jain, A., Bradica, G., and Butler, D.L.Improving linear stiffness of the cell-seeded collagensponge constructs by varying the components of the me-chanical stimulus. Tissue Eng Part A 14, 1883, 2008.
22. Nguyen, T.D., Liang, R., Woo, S.L., Burton, S.D., Wu, C.,Almarza, A., Sacks, M.S., and Abramowitch, S. Effects ofcell seeding and cyclic stretch on the fiber remodeling in anextracellular matrix-derived bioscaffold. Tissue Eng Part A15, 957, 2009.
23. Abousleiman, R.I., Reyes, Y., McFetridge, P., and Sika-vitsas, V. Tendon tissue engineering using cell-seededumbilical veins cultured in a mechanical stimulator. TissueEng Part A 15, 787, 2009.
24. Sahoo, S., Ouyang, H., Goh, J.C., Tay, T.E., and Toh, S.L.Characterization of a novel polymeric scaffold for potentialapplication in tendon/ligament tissue engineering. TissueEng 12, 91, 2006.
25. Sahoo, S., Cho-Hong, J.G., and Siew-Lok, T. Development ofhybrid polymer scaffolds for potential applications in ligamentand tendon tissue engineering. Biomed Mater 2, 169, 2007.
26. Chen, X., Qi, Y.Y., Wang, L.L., Yin, Z., Yin, G.L., Zou, X.H.,and Ouyang, H.W. Ligament regeneration using a knittedsilk scaffold combined with collagen matrix. Biomaterials29, 3683, 2008.
27. Doroski, D.M., Levenston, M.E., and Temenoff, J.S. Cyclictensile culture promotes fibroblastic differentiation ofmarrow stromal cells encapsulated in poly(ethylene gly-col)-based hydrogels. Tissue Eng Part A 16, 3457, 2010.
28. Issa, R.I., Engebretson, B., Rustom, L., McFetridge, P.S., andSikavitsas, V.I. The effect of cell seeding density on thecellular and mechanical properties of a mechanostimulatedtissue-engineered tendon. Tissue Eng Part A 17, 1479, 2011.
29. Ross, J.J., Duxson, M.J., and Harris, A.J. Neural determi-nation of muscle fibre numbers in embryonic rat lumbricalmuscles. Development 100, 395, 1987.
30. Liu, W., Chen, B., Deng, D., Xu, F., Cui, L., and Cao, Y.Repair of tendon defect with dermal fibroblast engineeredtendon in a porcine model. Tissue Eng 12, 775, 2006.
31. Woon, C.Y., Kraus, A., Raghavan, S.S., Pridgen, B.C., Me-gerle, K., Pham, H., and Chang, J. Three-dimensional-construct bioreactor conditioning in human tendon tissueengineering. Tissue Eng Part A 17, 2561, 2011.
32. Deng, D., Liu, W., Xu, F., Yang, Y., Zhou, G., Zhang, W.J.,Cui, L., and Cao, Y. Engineering human neo-tendon tissuein vitro with human dermal fibroblasts under static me-chanical strain. Biomaterials 30, 6724, 2009.
33. Obaid, H. and Connell, D. Cell therapy in tendon disorders:what is the current evidence? Am J Sports Med 38, 2123, 2010.
34. Chen, J., Wang, A., Xu, J., and Zheng, M. In chronic lateralepicondylitis, apoptosis and autophagic cell death occur inthe extensor carpi radialis brevis tendon. J Shoulder ElbowSurg 19, 355, 2010.
35. Androjna, C., Spragg, R.K., and Derwin, K.A. Mechanicalconditioning of cell-seeded small intestine submucosa: apotential tissue-engineering strategy for tendon repair.Tissue Eng 13, 233, 2007.
36. Saber, S., Zhang, A.Y., Ki, S.H., Lindsey, D.P., Smith, R.L.,Riboh, J., Pham, H., and Chang, J. Flexor tendon tissueengineering: bioreactor cyclic strain increases constructstrength. Tissue Eng Part A 16, 2085, 2010.
37. Chen, J.M., Willers, C., Xu, J., Wang, A., and Zheng, M.H.Autologous tenocyte therapy using porcine-derived bios-caffolds for massive rotator cuff defect in rabbits. TissueEng 13, 1479, 2007.
38. Cooper, J.A., Lu, H.H., Ko, F.K., Freeman, J.W., and Laur-encin, C.T. Fiber-based tissue-engineered scaffold for liga-ment replacement: design considerations and in vitroevaluation. Biomaterials 26, 1523, 2005.
39. Freeman, J.W., Woods, M.D., and Laurencin, C.T. Tissueengineering of the anterior cruciate ligament using a braid-twist scaffold design. J Biomech 40, 2029, 2007.
40. Lee, C.H., Shin, H.J., Cho, I.H., Kang, Y.M., Kim, I.A., Park,K.D., and Shin, J.W. Nanofiber alignment and direction ofmechanical strain affect the ECM production of humanACL fibroblast. Biomaterials 26, 1261, 2005.
41. Moffat, K.L., Kwei, A.S., Spalazzi, J.P., Doty, S.B., Levine,W.N., and Lu, H.H. Novel nanofiber-based scaffold for rotatorcuff repair and augmentation. Tissue Eng Part A 15, 115, 2009.
42. Webb, K., Hitchcock, R.W., Smeal, R.M., Li, W., Gray, S.D.,and Tresco, P.A. Cyclic strain increases fibroblast prolifera-tion, matrix accumulation, and elastic modulus of fibroblast-seeded polyurethane constructs. J Biomech 39, 1136, 2006.
43. Zheng, M., and Wang, A., Autologous Cell Therapy for Ten-don Tissue Reconstruction, in Sino-American Symposium onClinical and Translational Medicine. Washington, DC, 2011.
44. Zheng, M., Tenocyte cell culturing method, 2009.US7985408 (Issued date July 26, 2011).
45. Freed, L.E., Vunjak-Novakovic, G., and Langer, R. Culti-vation of cell-polymer cartilage implants in bioreactors. JCell Biochem 51, 257, 1993.
46. Freed, L.E., Marquis, J.C., Nohria, A., Emmanual, J., Mikos,A.G., and Langer, R. Neocartilage formation in vitro and invivo using cells cultured on synthetic biodegradable poly-mers. J Biomed Mater Res 27, 11, 1993.
47. Awad, H.A., Butler, D.L., Harris, M.T., Ibrahim, R.E., Wu,Y., Young, R.G., Kadiyala, S., and Boivin, G.P. In vitro
144 WANG ET AL.
237

characterization of mesenchymal stem cell-seeded collagenscaffolds for tendon repair: effects of initial seeding densityon contraction kinetics. J Biomed Mater Res 51, 233, 2000.
48. Wang, L., Seshareddy, K., Weiss, M.L., and Detamore, M.S.Effect of initial seeding density on human umbilical cordmesenchymal stromal cells for fibrocartilage tissue engi-neering. Tissue Eng Part A 15, 1009, 2009.
49. Zhang, L., Gardiner, B.S., Smith, D.W., Pivonka, P., andGrodzinsky, A. A fully coupled poroelastic reactive-trans-port model of cartilage. Mol Cell Biomech 5, 133, 2008.
50. Pham, Q.P., Sharma, U., and Mikos, A.G. Electrospinningof polymeric nanofibers for tissue engineering applications:a review. Tissue Eng 12, 1197, 2006.
51. Derwin, K.A., Baker, A.R., Spragg, R.K., Leigh, D.R., andIannotti, J.P. Commercial extracellular matrix scaffolds forrotator cuff tendon repair. Biomechanical, biochemical, andcellular properties. J Bone Joint Surg Am 88, 2665, 2006.
52. Kinneberg, K.R., Nirmalanandhan, V.S., Juncosa-Melvin,N., Powell, H.M., Boyce, S.T., Shearn, J.T., and Butler, D.L.Chondroitin-6-sulfate incorporation and mechanical stim-ulation increase MSC-collagen sponge construct stiffness. JOrthop Res 28, 1092, 2010.
53. Gilbert, T.W., Stewart-Akers, A.M., Simmons-Byrd, A., andBadylak, S.F. Degradation and remodeling of small intes-tinal submucosa in canine Achilles tendon repair. J BoneJoint Surg Am 89, 621, 2007.
54. Fleming, B.C., Spindler, K.P., Palmer, M.P., Magarian, E.M.,and Murray, M.M. Collagen-platelet composites improvethe biomechanical properties of healing anterior cruciateligament grafts in a porcine model. Am J Sports Med 37,
1554, 2009.55. Malcarney, H.L., Bonar, F., and Murrell, G.A. Early in-
flammatory reaction after rotator cuff repair with a porcinesmall intestine submucosal implant: a report of 4 cases. AmJ Sports Med 33, 907, 2005.
56. Zheng, M.H., Chen, J., Kirilak, Y., Willers, C., Xu, J., andWood, D. Porcine small intestine submucosa (SIS) is not anacellular collagenous matrix and contains porcine DNA:possible implications in human implantation. J BiomedMater Res B Appl Biomater 73, 61, 2005.
57. Chang, J., Thunder, R., Most, D., Longaker, M.T., andLineaweaver, W.C. Studies in flexor tendon wound heal-ing: neutralizing antibody to TGF-beta1 increases postop-erative range of motion. Plast Reconstr Surg 105, 148, 2000.
58. Bennett, N.T., and Schultz, G.S. Growth factors and woundhealing: biochemical properties of growth factors and theirreceptors. Am J Surg 165, 728, 1993.
59. Wojciak, B., and Crossan, J.F. The effects of T cells and theirproducts on in vitro healing of epitenon cell microwounds.Immunology 83, 93, 1994.
60. Marui, T., Niyibizi, C., Georgescu, H.I., Cao, M., Kavalk-ovich, K.W., Levine, R.E., and Woo, S.L. Effect of growthfactors on matrix synthesis by ligament fibroblasts. J Or-thop Res 15, 18, 1997.
61. Pryce, B.A., Watson, S.S., Murchison, N.D., Staverosky, J.A.,Dunker, N., and Schweitzer, R. Recruitment and mainte-nance of tendon progenitors by TGFbeta signaling are es-sential for tendon formation. Development 136, 1351, 2009.
62. Clark, R.T., Johnson, T.L., Schalet, B.J., Davis, L., Gaschen,V., Hunziker, E.B., Oldberg, A., and Mikic, B. GDF-5 defi-ciency in mice leads to disruption of tail tendon form andfunction. Connect Tissue Res 42, 175, 2001.
63. Mikic, B., Schalet, B.J., Clark, R.T., Gaschen, V., and Hun-ziker, E.B. GDF-5 deficiency in mice alters the ultrastruc-
ture, mechanical properties and composition of the Achillestendon. J Orthop Res 19, 365, 2001.
64. Chhabra, A., Tsou, D., Clark, R.T., Gaschen, V., Hunziker,E.B., and Mikic, B. GDF-5 deficiency in mice delays Achillestendon healing. J Orthop Res 21, 826, 2003.
65. Dahlgren, L.A., Mohammed, H.O., and Nixon, A.J. Tem-poral expression of growth factors and matrix molecules inhealing tendon lesions. J Orthop Res 23, 84, 2005.
66. Kobayashi, M., Itoi, E., Minagawa, H., Miyakoshi, N., Taka-hashi, S., Tuoheti, Y., Okada, K., and Shimada, Y. Expressionof growth factors in the early phase of supraspinatus tendonhealing in rabbits. J Shoulder Elbow Surg 15, 371, 2006.
67. Chamberlain, C.S., Crowley, E., and Vanderby, R. Thespatio-temporal dynamics of ligament healing. WoundRepair Regen 17, 206, 2009.
68. Hannafin, J.A., Arnoczky, S.P., Hoonjan, A., and Torzilli,P.A. Effect of stress deprivation and cyclic tensile loadingon the material and morphologic properties of canine flexordigitorum profundus tendon: an in vitro study. J OrthopRes 13, 907, 1995.
69. Flynn, B.P., Bhole, A.P., Saeidi, N., Liles, M., Dimarzio,C.A., and Ruberti, J.W. Mechanical strain stabilizes recon-stituted collagen fibrils against enzymatic degradation bymammalian collagenase matrix metalloproteinase 8 (MMP-8). PLoS One 5, e12337, 2010.
70. Maeda, T., Sakabe, T., Sunaga, A., Sakai, K., Rivera, A.L.,Keene, D.R., Sasaki, T., Stavnezer, E., Iannotti, J., Schweit-zer, R., Ilic, D., Baskaran, H., and Sakai, T. Conversion ofmechanical force into TGF-beta-mediated biochemical sig-nals. Curr Biol 21, 933, 2011.
71. Lejard, V., Brideau, G., Blais, F., Salingcarnboriboon, R.,Wagner, G., Roehrl, M.H., Noda, M., Duprez, D., Houillier,P., and Rossert, J. Scleraxis and NFATc regulate the ex-pression of the pro-alpha1(I) collagen gene in tendon fi-broblasts. J Biol Chem 282, 17665, 2007.
72. Dennis, R.G., Smith, B., Philp, A., Donnelly, K., and Baar,K. Bioreactors for guiding muscle tissue growth and de-velopment. Adv Biochem Eng Biotechnol 112, 39, 2009.
73. Catapano, G., Patzer, J.F., 2nd, and Gerlach, J.C. Transportadvances in disposable bioreactors for liver tissue engi-neering. Adv Biochem Eng Biotechnol 115, 117, 2010.
74. Rauh, J., Milan, F., Gunther, K.P., and Stiehler, M. Bior-eactor systems for bone tissue engineering. Tissue Eng PartB Rev 17, 263, 2011.
75. Salter, E., Goh, B., Hung, B., Hutton, D., Ghone, N., andGrayson, W.L. Bone tissue engineering bioreactors: a role inthe clinic? Tissue Eng Part B Rev 18, 62, 2012.
76. Altman, G.H., Lu, H.H., Horan, R.L., Calabro, T., Ryder, D.,Kaplan, D.L., Stark, P., Martin, I., Richmond, J.C., andVunjak-Novakovic, G. Advanced bioreactor with con-trolled application of multi-dimensional strain for tissueengineering. J Biomech Eng 124, 742, 2002.
77. Varseveld, R.B.v. and Bone, G.M. Accurate Position Con-trol of a Pneumatic Actuator Using On/Off SolenoidValves. Trans Mechatronics 2, 195, 1997.
78. Alter, D. and Tsao, T. Control of linear motors for machine toolfeed drives: design and implementation of H1 optimal feed-back control. ASME J Dyn Syst, Meas, Control. 118, 649, 1996.
79. Alter, D., and Tsao, T., Dynamic stiffness enhancement ofdirect linear motor feed drives for machining, in Proceedingsof the American Control Conference, Baltimore, MD, 1994.
80. Braembussche, P., Swevers, J., Brussel, H., and Vanherck, P.Accurate tracking control of linear synchronous motormachine tool axes. Mechatronics 6, 507, 1996.
BIOREACTOR DESIGN FOR TENDON/LIGAMENT ENGINEERING 145
238

81. Liu, J., Hsu, M., and Chen, F. On the design of rotatingspeed functions to improve the acceleration peak value ofball-screw transmission mechanism. Mech Machine Theory36, 1035, 2001.
82. Parent, G., Huppe, N., and Langelier, E. Low stress tendonfatigue is a relatively rapid process in the context of over-use injuries. Ann Biomed Eng 39, 1535, 2011.
83. Butler, D.L., Hunter, S.A., Chokalingam, K., Cordray, M.J.,Shearn, J., Juncosa-Melvin, N., Nirmalanandhan, S., andJain, A. Using functional tissue engineering and bioreactorsto mechanically stimulate tissue-engineered constructs.Tissue Eng Part A 15, 741, 2009.
84. Janssen, F.W., Oostra, J., Oorschot, A., and van Blitterswijk,C.A. A perfusion bioreactor system capable of producingclinically relevant volumes of tissue-engineered bone: invivo bone formation showing proof of concept. Biomater-ials 27, 315, 2006.
85. Cartmell, S.H., Porter, B.D., Garcia, A.J., and Guldberg, R.E.Effects of medium perfusion rate on cell-seeded three-dimensional bone constructs in vitro. Tissue Eng 9, 1197, 2003.
86. Meinel, L., Karageorgiou, V., Fajardo, R., Snyder, B.,Shinde-Patil, V., Zichner, L., Kaplan, D., Langer, R., andVunjak-Novakovic, G. Bone tissue engineering using hu-man mesenchymal stem cells: effects of scaffold materialand medium flow. Ann Biomed Eng 32, 112, 2004.
87. Uemura, T., Dong, J., Wang, Y., Kojima, H., Saito, T., Iejima,D., Kikuchi, M., Tanaka, J., and Tateishi, T. Transplantationof cultured bone cells using combinations of scaffolds andculture techniques. Biomaterials 24, 2277, 2003.
88. Holtorf, H.L., Jansen, J.A., and Mikos, A.G. Flow perfusionculture induces the osteoblastic differentiation of marrowstroma cell-scaffold constructs in the absence of dexa-methasone. J Biomed Mater Res A 72, 326, 2005.
89. Olivier, V., Hivart, P., Descamps, M., and Hardouin, P. Invitro culture of large bone substitutes in a new bioreactor:importance of the flow direction. Biomed Mater 2, 174,2007.
90. Saini, S. and Wick, T.M. Effect of low oxygen tension ontissue-engineered cartilage construct development in theconcentric cylinder bioreactor. Tissue Eng 10, 825, 2004.
91. Domm, C., Schunke, M., Christesen, K., and Kurz, B. Re-differentiation of dedifferentiated bovine articular chon-drocytes in alginate culture under low oxygen tension.Osteoarthritis Cartilage 10, 13, 2002.
92. Hansen, U., Schunke, M., Domm, C., Ioannidis, N.,Hassenpflug, J., Gehrke, T., and Kurz, B. Combination ofreduced oxygen tension and intermittent hydrostatic pres-sure: a useful tool in articular cartilage tissue engineering.J Biomech 34, 941, 2001.
93. Butler, D.L., Juncosa-Melvin, N., Boivin, G.P., Galloway,M.T., Shearn, J.T., Gooch, C., and Awad, H. Functionaltissue engineering for tendon repair: a multidisciplinarystrategy using mesenchymal stem cells, bioscaffolds, andmechanical stimulation. J Orthop Res 26, 1, 2008.
94. Goetz, M., Thomas, S., Heimann, A., Delaney, P., Schnei-der, C., Relle, M., Schwarting, A., Galle, P.R., Kempski, O.,Neurath, M.F., and Kiesslich, R. Dynamic in vivo imagingof microvasculature and perfusion by miniaturized confo-cal laser microscopy. Eur Surg Res 41, 290, 2008.
95. Drexler, W., Morgner, U., Kartner, F.X., Pitris, C., Boppart,S.A., Li, X.D., Ippen, E.P., and Fujimoto, J.G. In vivo ul-trahigh-resolution optical coherence tomography. Opt Lett24, 1221, 1999.
96. Yang, G., Im, H.J., and Wang, J.H. Repetitive mechanicalstretching modulates IL-1beta induced COX-2, MMP-1 ex-pression, and PGE2 production in human patellar tendonfibroblasts. Gene 363, 166, 2005.
97. Lavagnino, M., Arnoczky, S.P., Tian, T., and Vaupel, Z.Effect of amplitude and frequency of cyclic tensile strain onthe inhibition of MMP-1 mRNA expression in tendon cells:an in vitro study. Connect Tissue Res 44, 181, 2003.
98. Cribb, A.M. and Scott, J.E. Tendon response to tensile stress:an ultrastructural investigation of collagen:proteoglycan in-teractions in stressed tendon. J Anat 187 (Pt 2), 423, 1995.
99. Scott, J.E. The periphery of the developing collagen fibril.Quantitative relationships with dermatan sulphate andother surface-associated species. Biochem J 218, 229, 1984.
100. Vogel, K.G. What happens when tendons bend andtwist? Proteoglycans. J Musculoskelet Neuronal Interact 4,
202, 2004.101. Franchi, M., Trire, A., Quaranta, M., Orsini, E., and Ottani,
V. Collagen structure of tendon relates to function. SciWorld J 7, 404, 2007.
102. Sharifpoor, S., Simmons, C.A., Labow, R.S., and Paul Santerre,J. Functional characterization of human coronary arterysmooth muscle cells under cyclic mechanical strain in a de-gradable polyurethane scaffold. Biomaterials 32, 4816, 2011.
103. Donnelly, K., Khodabukus, A., Philp, A., Deldicque, L.,Dennis, R.G., and Baar, K. A novel bioreactor for stimu-lating skeletal muscle in vitro. Tissue Eng Part C Methods16, 711, 2010.
Address correspondence to:Ming H. Zheng, MD, PhD, FRCPath, FRCPA
Centre for Orthopaedic ResearchSchool of Surgery
The University of Western AustraliaM Block, QE2 Medical Centre
Nedlands, WA 6009Australia
E-mail: [email protected]
David G. Lloyd, PhDCentre for Musculoskeletal Research
Griffith Health InstituteClinical Science 1 (G02) Room 2.40
Griffith UniversityGold Coast Campus, QLD 4222
Australia
E-mail: [email protected]
David W. Smith, PhDSchool of Computer Science and Software Engineering
The University of Western Australia (M002)35 Stirling Highway
Crawley, WA 6009Australia
E-mail: [email protected]
Received: May 17, 2012Accepted: September 25, 2012
Online Publication Date: November 19, 2012
146 WANG ET AL.
239

ARTICLE
Programmable Mechanical Stimulation InfluencesTendon Homeostasis in a Bioreactor System
Tao Wang,1 Zhen Lin,1,2 Robert E. Day,3 Bruce Gardiner,4 Euphemie Landao-Bassonga,1
Jonas Rubenson,5 Thomas B. Kirk,6 David W. Smith,4 David G. Lloyd,7 Gerard Hardisty,8
Allan Wang,9 Qiujian Zheng,2 Ming H. Zheng1
1Centre for Orthopaedic Translational Research, School of Surgery, University of Western
Australia, M Block, QE2 Medical Centre, Nedlands, Crawley, Western Australia 6009,
Australia; telephone: 61-8-93464050; fax: 61-8-93463210;
e-mail: [email protected] of Orthopaedic Surgery, Department of Surgery, Guangdong General Hospital,
Guangdong Academy of Medicine Science, Guangzhou, Guangdong, China3Department of Medical Engineering and Physics, Royal Perth Hospital, Perth, Western
Australia, Australia4School of Computer Science and Software Engineering, University of Western Australia
(M002), Crawley, Western Australia, Australia5School of Sport Science, Exercise and Health, Musculoskeletal Tissue Mechanics
and Muscle Energetics University of Western Australia (M408), Crawley,
Western Australia, Australia6Curtin University, Bentley, Chancellory, Western Australia, Australia7Centre for Musculoskeletal Research, Griffith Health Institute, Griffith University,
Gold Coast Campus, Queensland, Australia8St John of God Medical Clinic, Perth, Subiaco, Western Australia, Australia9Sir Charles Gairdner Hospital, Perth, Nedlands, Western Australia, Australia
ABSTRACT: Identification of functional programmable me-chanical stimulation (PMS) on tendon not only provides theinsight of the tendon homeostasis under physical/patholog-ical condition, but also guides a better engineering strategyfor tendon regeneration. The aims of the study are to designa bioreactor system with PMS to mimic the in vivo loadingconditions, and to define the impact of different cyclictensile strain on tendon. Rabbit Achilles tendons wereloaded in the bioreactor with/without cyclic tensile loading(0.25Hz for 8 h/day, 0–9% for 6 days). Tendons withoutloading lost its structure integrity as evidenced by disor-ientated collagen fiber, increased type III collagen expres-sion, and increased cell apoptosis. Tendons with 3% of cyclictensile loading had moderate matrix deterioration andelevated expression levels of MMP-1, 3, and 12, whilst
exceeded loading regime of 9% caused massive rupture ofcollagen bundle. However, 6% of cyclic tensile strain wasable to maintain the structural integrity and cellular func-tion. Our data indicated that an optimal PMS is required tomaintain the tendon homeostasis and there is only a narrowrange of tensile strain that can induce the anabolic action.The clinical impact of this study is that optimized eccentrictraining program is needed to achieve maximum beneficialeffects on chronic tendinopathy management.
Biotechnol. Bioeng. 2013;110: 1495–1507.
� 2012 Wiley Periodicals, Inc.
KEYWORDS: bioreactor; programmable mechanical stimu-lation; tendon; collagen
Introduction
Tendinopathy is common in both young athletes and theelderly and is thought to be due to overuse and/oroverloading. Tendinopathy causes significant pain andcan lead to chronic degenerative changes in the tendon
Correspondence to: M. H. Zheng
Contract grant sponsor: Australia Research Council Linkage
Contract grant number: LP110100581
Received 28 September 2012; Revision received 27 November 2012;
Accepted 7 December 2012
Accepted manuscript online 14 December 2012;
Article first published online 4 February 2013 in Wiley Online Library
(http://onlinelibrary.wiley.com/doi/10.1002/bit.24809/abstract)
DOI 10.1002/bit.24809
� 2012 Wiley Periodicals, Inc. Biotechnology and Bioengineering, Vol. 110, No. 5, May, 2013 1495
240

matrix (Smith, 2000), and potentially tendon rupture (Leaand Smith, 1972; Leppilahti and Orava, 1998). Achillestendinopathy is one of the most common tendinopathiesand has the greatest potential to restrict the activities in dailyliving. Treatment outcomes are often poor. These poorresults are largely due to the underlying physiologicalprocesses within tendons, which are not well understood.
Tissue engineering methods such as cell-seeded bioscaf-folds have great potential in the repair of damaged tendons.However, these methods are still very much in their infancy,due to the lack of an integrated understanding of the keyconditions required for successful tendinopathy interven-tion and repair. Recent tendon research has focused on thedevelopment of tendon tissue engineering, with the aim ofconstructing a bio-substitute material for tendon/ligamentrepair (Chen et al., 2007, 2008; Juncosa-Melvin et al., 2006;Nirmalanandhan et al., 2008a; Peach et al., 2012; Saber et al.,2010; Sahoo et al., 2006; Stoll et al., 2010; Wang et al., 2012;Webb et al., 2006; Woon et al., 2012); This strategy involvesan integration of technologies from cell biology, materialscience, and mechanical engineering.
Engineered bioreactor systems have been demonstrated tobe effective tools for constructing various biological tissuesand organs such as muscle (Dennis et al., 2009), liver(Catapano et al., 2010), and bone (Rauh et al., 2011, 2012).Several bioreactor systems have been developed specificallyfor engineered tendon/ligament formation (Altman et al.,2002; Issa et al., 2011; Juncosa-Melvin et al., 2006;Nirmalanandhan et al., 2008a,b; Webb et al., 2006).Owing to force transmission being the primary functionrole of tendon/ligament, mechanical stimulation plays anessential role in the maintenance of tendon structure andfunction (Hannafin et al., 1995). Achilles tendons enduretensile loading in vivo during daily physical activities. Morecomplex structures, like the anterior cruciate ligament,undergo both tensile and torsion loading during weight-bearing knee flexion (Li et al., 2005). To restore tendonfunction after injury, engineered tendon constructs shouldbe compatible with the in vivo environment bothbiologically and mechanically (Calve et al., 2010). Cell-scaffold constructs may be ‘‘exercised’’ in an artificialmechanical environment before implantation. Indeed,programmable mechanical stimulation (PMS) is consideredto be one of the key components for tendon/ligamentforming bioreactor system (Altman et al., 2002; Issa et al.,2011; Juncosa-Melvin et al., 2006; Nirmalanandhan et al.,2008a,b; Webb et al., 2006). In this study, a bioreactorsystem was designed to apply different cyclic tensile strain onisolated rabbit Achilles tendon to establish an ex vivo model.The present aim is to evaluate the effect of differentmechanical stimulation on tendon tissue.
The mechanical stimulation of engineered tendonconstructs can result in different structures. Non-loadingof engineered neo-tendons results in loose, disorganizedmatrices with reduced collagen quantity (Jiang et al., 2011).Static mechanical stress has been shown to improve the fiberarrangement of engineered tendon constructs, but did not
produce a compacted tendon-like collagen structure in vitro(Chen et al., 2007). More promisingly, engineered tendonunder cyclic tensile strain exhibited a well-organizedcollagen fiber arrangement and structure and enhancedtensile strength, suggesting that PMS is preferable forengineered tendon formation (Jiang et al., 2011). Eventhough the importance of providing PMS in tendonbioreactor has been recognized previously (Brown andCarson, 1999; Wang et al., 2012), the optimal cyclic tensilestimulation parameters have not been defined. It is generallyassumed that mimicking the dynamic mechanical loadingcreated by daily physical activity will give the optimalloading regime in tendon bioreactor system, but this is notyet proven.
The goals of this study were: (1) to develop and validate abioreactor system; (2) to investigate the effect of differentmechanical stimulation regimes on tendon integrity. Wehypothesized that there is an optimum range of mechanicalstimulation required for tendon homeostasis and that underor over-stimulation would reduce tendon integrity.
Materials and Methods
Bioreactor Design Criteria
To develop a functional bioreactor, several design criteriawere formulated. Firstly, the bioreactor must be able toprocess more than three individual cultures simultaneouslywith transparent covers allowing visual monitoring.Secondly, the extension accuracy should be better than�100mm with the load varying as required to produce theset strain (the minimum expected load was 20N persample). Third, a construct fixation method must preventslippage during stimulation. Moreover, as the bioreactorsystem is operated in an incubator, all of the electrical andmechanical components have to be able to function in awarm, humid environment. Finally, but not least, the partsinvolved with the tissue culture should be autoclavable.
To achieve these design criteria two structural materialswere used, AISI 316 stainless steel and 6063 aluminum. Themain bioreactor components are the culture chamber, tissueclamps and hooks made of stainless steel (which can be easilysterilized) and frame. The frame was made of aluminum toreduce the weight and 2mm thick glass was used fortransparent cover. The actuator was a motor-ball screwtransmission design using a step motor and precision ballscrew to maximize the load output and positional accuracy.A programmable logic control was chosen, rather than directcomputer control, to reduce cost and allow reliableoperation in uncertain experimental environments.
Tissue Harvest and Distribution
Achilles tendons were dissected from the hindlimbs offemale New Zealand White rabbits (Oryctolagus cuniculus)
1496 Biotechnology and Bioengineering, Vol. 110, No. 5, May, 2013
241

aged between 14 and 15 weeks with body weight 3–5kg. Thetissue was briefly rinsed in 70% ethanol, washed in 1�phosphate-buffered saline (PBS) three times, and submergedin serum free Dulbecco’s modified Eagle’s medium (DMEM)/F-12 (GIBCO, Invitrogen, Auckland, New Zealand) supple-mented with 50mg/mL Gentamicin before further processing.
In total 24 Achilles tendons from 12 rabbits were allocatedto three groups (Fig. 1A): a loading deprivation group (8), adynamic culture (cyclic tensile strain) group (12), and acontrol group (4). In the loading deprivation group, theAchilles tendons were cultured ex vivo in growth medium(DMEM/F-12, 10% fetal bovine serum and 50mg/mLGentamicin) without mechanical loading for either 6 days(four tendons) or 2 weeks (four tendons). In the dynamicstrain group, the tendons were cultured under 3%, 6%, or 9%cyclic tensile strain for 6 days (8 h per day, 0.25Hz, Fig. 1A).The stimulating cycle was as demonstrated in Figure 1B. Thisregimen was based on previously published studies and dataon rabbit activity levels (West et al., 2004). The samples werethen fixed in 4% paraformaldehyde overnight for histologicalanalysis or directly subjected to RNA extraction. The controlgroup was made up of four tendons directly dissected fromthe animal and immediately fixed.
Histological Preparation and Assessment
For histological assessment, the tissue samples were fixed in4% paraformaldehyde overnight, dehydrated throughgraded alcohol, cleaned with xylene and embedded inparaffin blocks. Histological sections (5mm) were cut on amicrotome and stained with Haematoxylin & Eosin. Blindgeneral histological scoring was performed by three
individuals as previously described (Chen et al., 2011).Four elements (cell density, cell roundness, fiber structure,and fiber arrangement) were assessed using scoring scale of0–3, where 0 is normal and 3 severely abnormal. The averagescore of each parameter was used for comparison.
TUNEL Assay
The terminal deoxynucleotidyl transferase-mediated deox-yuridine triphosphate nick end labeling (TUNEL) assay(Cell Death Detection-AP Kit, Roche, Switzerland) wasperformed to identify apoptotic cells by labeling the nuclearDNA fragments. Paraffin-embedded sections were dewaxedand rehydrated using graded ethanol (100%, 95%, 80%, and70%). The tissue was digested with 20mg/mL proteinase K(Qiagen, Hilden, Germany) for 15min, incubated withTUNEL reaction mixture for 60min and the convert-AP(alkaline phosphatase) reagent for another 30min, accord-ing to the manufacturer’s instruction. After rinsing withPBS, the sections were visualized with Diaminobenzidine(DAB) kit (DAKO, Glostrup, Denmark) for 5min, whichstained the positive cells brown; and counterstained withhematoxylin.
Immunostaining for Type III Collagen
To evaluate the extracellular matrix (ECM) turnover intendons with and without dynamic tensile strain, proteinexpression of type III collagen was evaluated. Paraffin-embedded sections were dewaxed and rehydrated. Theendogenous peroxidase was blocked with 3% hydrogenperoxide (H2O2) for 10min, and the sections were then
Figure 1. A: Number and distribution of samples for each of the 12 rabbits. B: Daily mechanical stimulation pattern used in this study. C: Mechanical input signal from the
control system.
Wang et al.: Programmable Mechanical Stimulation 1497
Biotechnology and Bioengineering
242

incubated with 5% fetal bovine serum in 1� PBS for 30min,followed by monoclonal anti-collagen type III (036K4782,Sigma, St Louis, MO; 1:100) in a damp chamber overnight at48C. Biotinylated goat antimouse IgG at a dilution of 1:200(Sigma) was used as secondary type III antibody. PBSwas usedto wash the sections between solutions and reagents. Sectionswere developed to visualize the positive staining using theDAB kit, and counterstained with hematoxylin solution.
Quantitative Real-Time Polymerase ChainReaction (Q-PCR)
The effect of mechanical stimulation on collagen geneexpression was examined in Achilles tendon after theprogrammed dynamic training. Approximately 200mg oftendon tissue was dissected from the middle region, and thetotal ribonucleic acid (RNA) was extracted using TRIzol1
reagent (Invitrogen, Carlsbad, CA). The RNA concentrationwas quantified and using SmartSpecTM 3000 (Bio-Rad,Hercules, CA), and A260/A280 was measured to evaluate theRNA quality. The RNA (500ng) was then reverse-transcribedinto complementary deoxyribonucleic acid (cDNA) usingSuperScript1 III First-Strand Synthesis System (Invitrogen,USA) according to the manufacturer’s instructions.
The cDNA was amplified and quantified by Q-PCR. Forthis reaction, 5mL of iQTM SYBR1 Green Supermix (Bio-Rad), 1mL of cDNA, 0.5mL of each forward and reverseprimer and 3mL of DEPC water were combined and mixed.The real-time PCR was performed as previously described(Lin et al., 2008). The primer sequences are listed in Table I.
Statistical Analyses
All statistics have been analyzed using the prism 5.03 softwarepackage (GraphPad Software, Inc., San Diego, CA). Data arepresented as mean� standard deviation (SD). Statisticalanalysis was performed by one-way analysis of variance(ANOVA). P values less than 0.05 were considered significant.
Results
Bioreactor for Tendon Engineering
By constructing a bioreactor to the design criteria, we havegenerated a bioreactor that is able to simultaneously apply
pre-programmed uniaxial stimulation on a maximum of sixtendons, cultured separately in their own chamber (toprevent cross contamination) (Fig. 2A). As Achilles tendonsrequire only tensile stimulation (Louis-Ugbo et al., 2004),system performance was tested with rabbit Achilles tendons.The system was actuated by a step motor-ball screwtransmission system (SMBS) using VEXTA step motor(PX245M-01AA, Oriental Motor, Kabushiki Kaisha, Tokyo,Japan) and precision ball screw (SG0601, KSS Co., Ltd.,Ojiya, Niigata, Japan), which produced 198N peak force anda positional accuracy of �5mm. Using programmable logiccontrollers (MELSEC FX1S, Mitsubishi, Marunouchi,Chiyoda-ku, Tokyo, Japan), as shown as Figure 2A,displacement, frequency, and stimulation period are allprogrammable. The culture chamber contains six indepen-dent vessels with individual capacity of 20mL (width¼2 cm, length¼ 9 cm). Individual transparent glass coversminimized the risk of cross contamination between testedsamples, and provided a window for monitoring eachtendon (Fig. 3). This system provides two samplestabilization methods: tissue clamps (Fig. 2C) and hooks(Fig. 2D). Soft and flat scaffolds, like a collagen and silkconstruct, can be fixed using the tissue clamps, whereasrobust and slippery tissue, such as bone/tendon and muscle/tendon junctions, can be tied to the hooks using suture. Thewhole system was placed inside a water-jacketed incubator(Forma Scientific, Marietta, OH) at 378C with 5% CO2, and95% air.
Using the final bioreactor design, Achilles tendonsisolated from rabbits were placed into the system tooptimize of condition of dynamic culture. Cyclic tensilestrain at 3% 6%, and 9% (0.25Hz for 8 h per day for 6 days)were applied, to a total of 12 rabbit Achilles tendons. Overallculture period was over 3 weeks with no contaminationfound during the culture period. Clear vision was achievedthrough transparent glass cover. Both the tissue clamp andtissue hook worked effectively with no slippage observed.
Histological Examination of Tendon UndergoingDifferent Cyclic Tensile Strains
Compared with the native tendon (Fig. 4A), the collagenfiber arrangement changed from parallel to moderately wavyafter 6 days in the absence of applied loading. Cell densitywas increased with slightly rounded nuclei. After 2 weekswithout loading, progressive ECM disruption was observed.
Table I. Rabbit specific primer sequences for the genes used.
Target genes Forward (50–30) Reverse (30–50)
COL1A1 TTGCCCTTCCTTGATATTGC CCTCTTTCGCCCACAATTTA
COL3A1 AAGCCCCAGCAGAAAATTG TGGTGGAACAGCAAAAATCA
MMP-1 GGCTCAGTTCGTCCTCACTC CAGGTCCATCAAAGGGAGAA
MMP-3 GCTTTGCTCAGCCTATCCAC ACCTCCAAGCCAAGGAACTT
MMP-12 ATGCCAGGGAGACCAGTATG AAAGCATGGGCTATGACACC
36B4 ACCCAAAATGCTTCATCGTG CAGGGTTGTTTTCCAGATGC
1498 Biotechnology and Bioengineering, Vol. 110, No. 5, May, 2013
243

Collagen fiber arrangement had no discernible pattern.Fragmentation of collagen fibers and rounded tenocytenuclei were observed microscopically (Fig. 4C). Histologicalassessment showed a time dependent increasing grading infiber arrangement, fiber structure and cell roundness,indicating progressive divergence from the native (control)tissue. Cell density was also increased after 6 days withoutloading (Fig. 4D–G).
In the dynamic culture (cyclic tensile strain) group,histology revealed that the 3% strain stimulated group hadslightly disrupted ECM structure and significant cellularmorphology changes (Fig. 5B). These tendon samplesgraded 1.5 in fiber arrangement, 1.1 in fiber structure and1.3 in cell roundness (Fig. 5E–G). However, in the tendonsthat underwent 6% strain, the ECM orientation and cellular
morphology appeared normal (Fig. 5A and C). That is, therewas no statistical difference of the histology scoring betweenthe 6% group and native tissue (Fig. 5E–H). In the 9% strainstimulated group, the tendon samples were found to bepartially torn and graded 1.45 in fiber structure and 0.8 infiber arrangement (Fig. 5E and F). Increased cell numberand slightly rounded tenocyte nuclei were observed aroundthe rupture site (Fig. 5D, G, and H).
Impact of Cyclic Tensile Strain on Apoptosis of Tenocytes
To investigate the impact of mechanical force on theapoptosis of tenocytes, we used the TUNEL assay to examinethe cellular apoptosis rate in specimens. In the nativetendons, the average cell apoptosis fraction (as a proportionof total cell number) was less than 10% (Fig. 6A and G). Theapoptosis fraction progressively increased over time whenthe tissues were cultured without loading (35% in 6 days and95% in 2 weeks) (Fig. 6G). Cell viability in 3% and 6%stimulated Achilles tendon was similar to that in nativetendon (Fig. 6D and E) without statistical significance(Fig. 6G), whilst cell apoptosis rate increased to 45% whenthe stimulation strain reach 9% (Fig. 6F and G).
Type III Collagen Turnover in Achilles Tendon UnderDynamic Culture
To investigate the ECM turnover in tendon undergoingdifferent cyclic tensile strain, type III collagen immunohis-tochemistry was done. No positive type III collagen wasobserved in the core of healthy tendon using
Figure 2. A: The over view of the bioreactor system. B: Schematic illustration of the bioreactor system. C: Exploded view of the tissue clamps assembly. D: Exploded view of
the tissue hooks assembly.
Figure 3. Bioreactor chamber with three rabbit Achilles’ tendons cultured
inside.
Wang et al.: Programmable Mechanical Stimulation 1499
Biotechnology and Bioengineering
244

Figure 4. H&E staining assessment and histological scoring of loading deprivation culture group. A: Native Achilles tendon structure (day 0). B: Achilles tendon histology after
6 days loading deprivation culture with minor wavy collagen structure. C: Achilles tendon histology after 2 weeks culture displays disorganized collagen structure with round
tenocytes. D: Fiber arrangement, scale of 0–3, 0 represents compacted and parallel, and 3 represents no identifiable pattern. E: Fiber structure, scale of 0–3, 0 represents continue,
long fiber, and 3 represents severe fragmented. F: Cell roundness, scale of 0–3, 0 represents long spindle shape, and 3 represents severely rounding. G: Cell density, scale of 0–3, 0
represents normal pattern, and 3 represents severely increase Result are expressed as the mean� SD (�P< 0.05, ��P< 0.01, ���P< 0.001). Scale bar¼ 50mm.
1500 Biotechnology and Bioengineering, Vol. 110, No. 5, May, 2013
245

Figure 5. H&E staining assessment and histological scoring of 6 days dynamic culture group. A: Native Achilles tendon structure. B: Three percent stimulated group with
wavy collagen structure and minor rounded tenocytes. C: Six percent stimulated group with similar morphology compared with the native group. D: Nine percent stimulated group
with severe tear collagen structure, and rounded nuclei around the tear region (arrow). E: Fiber arrangement, scale of 0–3, 0 represents compacted and parallel, and 3 represents no
identifiable pattern. F: Fiber structure, scale of 0–3, 0 represents continue, long fiber, and 3 represents severely fragmented. G: Cell roundness, scale of 0–3, 0 represents long
spindle shape, and 3 represents severely rounding. H: Cell density, scale of 0–3, 0 represents normal pattern, and 3 represents severely increase. Result is expressed as the
mean�SD (�P< 0.05, ��P< 0.01, ���P< 0.001). Scale bar¼ 50mm.
Wang et al.: Programmable Mechanical Stimulation 1501
Biotechnology and Bioengineering
246

Figure 6. TUNEL assay. A: Native Achilles tendon with less than 10% cell apoptosis. B: Six days loading deprivation group has increased apoptotic tenocytes pointed out by
arrows. C: In 2 weeks loading deprivation group, almost all tenocytes are under apoptosis D: Three percent stimulated group with minor increased apoptotic cells (arrow). E: Six
percent stimulated group with similar cell apoptosis rate as native sample. F: Nine percent stimulated group with massive cell apoptosis around the damaged site (pointed by
arrows). G: Quantified cell apoptosis fraction of each group. Result are expressed as the mean�SD (�P< 0.05, ��P< 0.01, ���P< 0.001). Scale bar¼ 50mm.
1502 Biotechnology and Bioengineering, Vol. 110, No. 5, May, 2013
247

immunochemistry (Fig. 7A). After 6 days of either noloading or 3% strain, the Achilles tendons expressed onlyminor and moderate collagen type III respectively(Fig. 7B and C), whilst relatively higher positive stainingwere found in the 9% strain group (Fig. 7E). No type IIIcollagen expression was observed in the 6% cyclic strainstimulated group (Fig. 7D), which was similar to the nativetendon.
Impact of Cyclic Tensile Strain on ECM RemodelingGene Expression
Q-PCR was carried out to examine the gene expression oftype I (COL1A1), type III collagen (COL3A1), and MMP-1,3, 12 (matrix metalloproteinases). As shown in Figure 8, thetissues subjected to 6% cyclic tensile strain had the highestexpression of COL1A1 (Fig. 8A), whereas 3% stimulatedgroup had the lowest expression. Type III collagen(COL3A1) was suppressed by minor and moderate loading(3% and 6%), yet upregulated in the 9% strain stimulatedgroup (Fig. 8B). Gene expression of MMP1, 3, and 12 werehighly upregulated by 3% strain stimulation compared tothe other groups.
Discussion
A previous study has suggested that engineered neo-tendonsin a non-loading environment result in loose, disorganizedand less collagen-rich matrices (Jiang et al., 2011). Staticmechanical loading has been shown to improve the fiberarrangement of engineered tendon constructs, but did notproduce a compacted tissue structure in vitro (Chen et al.,2007). Engineered neo-tendons with PMS had a well-organized fiber arrangement and structure and enhancedtensile strength, suggesting that dynamic mechanicalloading is preferable for engineered tendon formation(Jiang et al., 2011). Although PMS has been recognized as acritical regulator for tendon formation (Brown and Carson,1999), the optimal PMS parameters are yet to be defined.
In this study we designed a uni-axial bioreactor systemwith a programmable stimulation regimen. Achilles tendonsfrom rabbits were used to evaluate the impact of differentPMS conditions on tendon tissue. Our result showed that3% cyclic tensile strain in the bioreactor did not preventmatrix deterioration, whilst at 6% cyclic tensile strainstructural integrity and cellular function was maintained. At9% cyclic tensile strain there was massive rupture of thecollagen bundles. However, these results are for an already
Figure 7. Collagen type III immunohistochemistry. A: Native Achilles tendon with minor collagen type III expression. B: Loading deprivation group has minor increased
collagen type III expression and some positive staining tenocytes pointed out by arrow. C: Three percent stimulated group with increased collagen type III expression. D: Six
percent stimulated group with similar collagen type III expression as native sample. E: Nine percent stimulated group with massive increased collagen type III expression and
positive staining tenocytes (pointed by arrows). Scale bars¼ 50mm.
Wang et al.: Programmable Mechanical Stimulation 1503
Biotechnology and Bioengineering
248

mature tendon and the loading regimes may be different forengineered constructs at various stages of growth.Furthermore, the frequency of loading may also have animportant role in maintaining tendon homeostasis and thisneed to be better qualified by in vivo measurements ofAchilles tendon loading. Nevertheless, the current resultsdemonstrate that PMS can be achieved in this bioreactor,and that an optimal cyclic tensile loading (around 6% cyclictensile strain) is required to maintain healthy tendonhomeostasis, at least over the time period of the culturesused in this study.
In recent years, many studies of tendon engineering havebeen conducted using custom-made bioreactor system, andthe results have been promising (Abousleiman et al., 2009;Androjna et al., 2007; Chen et al., 2010; Doroski et al., 2010;Issa et al., 2011; Juncosa-Melvin et al., 2006; Nguyen et al.,2009; Nirmalanandhan et al., 2008a,b; Saber et al., 2010;Webb et al., 2006; Woon et al., 2011). Several commercialtendon engineering bioreactors are available now, such asBose1 ElectroForce1 BioDynamic1 and LigaGen system.To develop a functional bioreactor system, several factors
needed to be considered, including material, mechanicaldesign, tissue fixation method, and operating environment(Wang et al., 2012). Compared with commercial bioreactorsin Bose and LigaGen, the current design has severaladvantages. Firstly, our device can condition multiplesamples simultaneously but separately. It can be easilycleaned and sterilized in standard autoclaves. Beingconstructed from AISI 316 stainless steel it can go throughmany sterilization cycles without corrosion or degradation.The device can produce a maximum force of 198N, which issufficient for most tendon scaffold and tendon tissue fromsmall scale animals such as are commonly used in tissueengineering research. Most commercial bioreactor systemshave problems in holding thick and slippery tissue duringPMS, where as our novel tissue hook design demonstratedexcellent tissue fixation. Finally, the actuation system isdesigned to operate inside a standard incubator. It is able tofunction in a humid environment and the horizontal designis easy to move and will only occupy a small space in theincubator. As the tendon’s main function is mechanical, thisbioreactor system is able to provide high-accuracy PMS,
Figure 8. Quantitative PCR results. A: COL1A1 expression in 6% stimulated group is significantly higher than other groups. B: COL3A1 expression in 9% stimulated group is
significantly higher than other groups. Three percent and 6% group had much lower COL3A1 expression compared to the other groups. C–E: Three percent stimulated group has
significantly high expression on MMP1 MMP3 and MMP12, respectively. Result is expressed as the mean�SD (�P< 0.05, ��P< 0.01, ���P< 0.001). Scale bar¼ 50mm.
1504 Biotechnology and Bioengineering, Vol. 110, No. 5, May, 2013
249

enabling different loading regimes including displacement,frequency and stimulation period. Once programmed it isable to operate reliably for weeks without computer support.
It has been well known that tensile mechanical loading isessential for tendon integrity, and loading deprivation cancause tendinopathy-like morphology such as disorientatedcollagen fibers and rounded cell nuclei (Hannafin et al.,1995). Yet the specific effect of different loading conditionson tendon has not been well elucidated. To study the effectof PMS on tendon/ligament, both in vitro and in vivomodels have been used. The Flexcell1 system has beenapplied to examine the in vitro gene expression profiles oftenocytes under tensile or shear force (Skutek et al., 2001;Wang et al., 2003; Yang et al., 2005). These systems canprecisely control the mechanical regimen and monitor thecell behaviors, but only use a monolayer of cells, not anorganized tissue. Cells isolated from their extracellular nicheand cultivated in a monolayer may not necessarily replicatetheir original behavior in vivo. In vivo models, such asAchilles tendon in rats or rabbit, have been commonly usedas study models. Tendon loading is mainly induced byelectrical stimulation on muscle (Asundi et al., 2008;Nakama et al., 2005), treadmill training on animal andpartial tendon transaction (Smith et al., 2008). However,mechanical stimulation on the tendon varies betweenindividuals, making precise quantification of load next toimpossible. In this study, we have demonstrated a bioreactorsystem that not only can be used as an device for tendon/ligament engineering, but also serves as an ex vivo culturemodel for the study of tendon biology. Although thebiomechanics of Achilles tendon is more complicated underin vivo environment (West et al., 2004), by applying PMS,the effect of different cyclic tensile strain on tendon tissuecould be systematically analyzed on a tissue level.
The present study has shown for the first time that only anarrow range of PMS can stimulate the anabolic effect oftendon tissue. This suggests a proposed model describingthe effect of mechanical loading, from under-load to over-load, on tendon homeostasis. This model is illustrated inFigure 9 in which the black curve presents the typicalmechanical response caused by increasing tensile strain intendon. The breakage of collagen fibers increases withincreasing strain, leading to eventual rupture at the failurepoint. The red curve, on the other hand, proposes thebiological repair response of the tendon induced by tensilestrain. Mechanical loading is required for the biologicalactivity of tenocytes, yet the regime needs to be within acertain range to promote matrix product and net tissuerepair. By combining these two curves, three crossover areasare seen, which can be divided according to their metabolicstatus into: A (anabolic) zone and C (catabolic) zones. In theA zone, where the tensile loading is within a certain range,the biological repair response caused by PMS exceeds theinduced damage. In the C zone, there is either too little loadto stimulate matrix production or too much mechanicaldamage caused by excess tensile loading. In our study the 3%strain stimulated group (under-loaded) was located in C
zone. When the tendons were subjected to insufficienttensile strain, early tendinopathologic changes (includingincreased cellular number, type III collagen expression anddisorientated collagen fibers) could be observed. Thesemorphological changes may be attributed by the upregula-tion of MMPs expression in tendon under 3% tensileloading. MMPs are zinc-dependent endopeptidases that ableto degrade all the components of the ECM. MMP-1 isprimarily responsible for the cleavage of collagen, includingtype I, II, and III (Krane et al., 1996). MMP-3 and MMP-12are other members of the MMP family that have a widerange of substrate specificities. Moreover, MMP-12 is able toactivate MMP-3, and MMP-3 has the ability to activateother MMPs such as MMP-1, MMP-7, and MMP-9 (Chen,2004; Knauper et al., 1996). Studies have shown thatdecreased mechanical force on tendon fiber would lead tothe deprivation of cytoskeletal tension in tenocytes and alterthe mechanostransduction in the cells (Arnoczky et al.,2008). Insufficient loading of tenocyte can initiate thedegenerative cascade by expressing a pattern of catabolicgene, including MMPs (Arnoczky et al., 2004, 2008;Egerbacher et al., 2008; Lavagnino et al., 2003, 2006). Inthis study, we have confirmed that under the underloadingcondition, the phenotype of the tenocytes changed fromspindle to round, and the expression levels of catabolicgenes, such as MMP-1, 2, and 12, were upregulated. Theelevated catabolic factors may contribute to the furthermicro-damage of the collagen fiber. Conversely, althoughthe tendons in the 6% strain stimulated group weresubjected to more mechanical damage, the repair mecha-nism was able to maintain tissue homeostasis (i.e., there is anarrow ‘‘net anabolic window’’ between two net cataboliczones). In the 9% group (overloaded), mechanical damageoverwhelmed the tendon tissue repair and caused partialrupture, which is consistent with our proposed model.
The identification of an optimal PMS range for tendonwill not only guide us to a better tissue-engineering pathway
Figure 9. Matrix damage (black curve) and production (red broken) affected by
mechanical stimulation. A zone indicates a small range of anabolic effect caused
by mechanical stimulation. Matrix production stimulated by mechanical loading
overcomes the matrix damage that the tendon is able to maintain its structural
integrity. C zone indicates two catabolic zones that matrix damage caused by
mechanical stretching overcomes the matrix production result in tendinopathy.
Wang et al.: Programmable Mechanical Stimulation 1505
Biotechnology and Bioengineering
250

for tendon regeneration, but also provide an insight intotendon homeostasis under both physiological and patho-logical conditions. Clinically, chronic tendinopathy ofAchilles tendon is usually conservatively managed byeccentric overload training. The calf muscle is eccentricallyloaded with the knee held straight or the soleus muscle ismaximally loaded with the knee bent (Alfredson et al.,1998). Although positive impacts of eccentric overloadtraining on tendinopathy treatment have been recorded, theoutcomes have varied widely (Alfredson and Lorentzon,2003; Alfredson et al., 1998; Fahlstrom et al., 2003; Rooset al., 2004; Shalabi et al., 2004; Silbernagel et al., 2001;Stanish et al., 1986). In the study of Alfredson et al. (1998)the overall improvement in pain score was up to 94%,though in the study of Silbernagel et al. reported only 29%(Silbernagel et al., 2001). Although the mechanism ofeccentric overload training on tendinopathy managementremains unclear, the present study has provided a possibleexplanation of the mechanism of eccentric loading onchronic tendinopathy. The high variation of clinicaloutcomes might be due to poor control of the loadingregimen. Given that there is only a narrow range of tensilestrain where eccentric exercise could provide an anaboliceffect on tendon, clinical treatment protocols will need toaccurately control the magnitude of load applied throughthe injured tendon.
Finally, it is noted that nutrient supply might be altered byadvective transport associated with mechanical stimulation.Although a previous study on canine flexor tendonssuggested that mechanical stimulation did not alter smallnutrient uptake compared to static culture, and the tendonrepair improvements derived from passive motion is notrelated to an increase of nutrient transport (Hannafin andArnoczky, 1994), under different loading regimes, or indifferent tendon types, this may not be the case. Moreover,the transport of large molecules, such as growth factors andmatrix components, may be affected by mechanicalstimulation, as has been shown in studies in related tissues(Bonassar et al., 2000; Zhang et al., 2007). Therefore, thetendon stimulated by dynamic mechanical loading in thisstudy might be attributed to the enhanced nutrient transport,compared to the static control. Consequently it is difficult toisolate the direct effect of mechanical stimulation on tendonresponse from potential enhanced advective transportprocesses. Further study would be required to measure thenutrition transport in tendon under PMS.
Conclusion
We have developed and validated a uniaxial bioreactorsystem. The results in our study showed tendons subjectedto 6% of cyclic tensile strain were able to maintain theirstructural integrity and cellular function. Our data suggestedthat tendon is a mechano-sensing tissue where the netanabolic action of cyclic tensile strain only occurs over anarrow range. In our study, mechanical underloading (3%
cyclic tensile strain) and overloading (9% cyclic tensilestrain) produced a net negative impact on the rabbit Achillestendon tissue.
References
Abousleiman R.I., Reyes Y, McFetridge P, Sikavitsas V. 2009. Tendon tissue
engineering using cell-seeded umbilical veins cultured in a mechanical
stimulator. Tissue Eng Part A 15(4):787–795.
Alfredson H, Lorentzon R. 2003. Intratendinous glutamate levels and
eccentric training in chronic Achilles tendinosis: A prospective study
using microdialysis technique. Knee Surg Sports Traumatol Arthrosc
11(3):196–199.
Alfredson H, Pietila T, Jonsson P, Lorentzon R. 1998. Heavy-load eccentric
calf muscle training for the treatment of chronic Achilles tendinosis.
Am J Sports Med 26(3):360–366.
Altman GH, Lu HH, Horan RL, Calabro T, Ryder D, Kaplan DL, Stark P,
Martin I, Richmond JC, Vunjak-Novakovic G. 2002. Advanced biore-
actor with controlled application of multi-dimensional strain for tissue
engineering. J Biomech Eng 124(6):742–749.
Androjna C, Spragg RK, Derwin KA. 2007. Mechanical conditioning of cell-
seeded small intestine submucosa: A potential tissue-engineering strat-
egy for tendon repair. Tissue Eng 13(2):233–243.
Arnoczky SP, Lavagnino M, Egerbacher M, Caballero O, Gardner K,
Shender MA. 2008. Loss of homeostatic strain alters mechanostat
‘‘set point’’ of tendon cells in vitro. Clin Orthop Relat Res
466(7):1583–1591.
Arnoczky SP, Tian T, Lavagnino M, Gardner K. 2004. Ex vivo static tensile
loading inhibits MMP-1 expression in rat tail tendon cells through a
cytoskeletally based mechanotransduction mechanism. J Orthop Res
22(2):328–333.
Asundi KR, King KB, Rempel DM. 2008. Evaluation of gene expression
through qRT-PCR in cyclically loaded tendons: An in vivo model. Eur J
Appl Physiol 102(3):265–270.
Bonassar LJ, Grodzinsky AJ, Srinivasan A, Davila SG, Trippel SB. 2000.
Mechanical and physicochemical regulation of the action of insulin-like
growth factor-I on articular cartilage. Arch Biochem Biophys
379(1):57–63.
Brown CH, Jr., Carson EW. 1999. Revision anterior cruciate ligament
surgery. Clin Sports Med 18(1):109–107.
Calve S, Lytle IF, Grosh K, Brown DL, Arruda EM. 2010. Implantation
increases tensile strength and collagen content of self-assembled tendon
constructs. J Appl Physiol 108(4):875–881.
Catapano G, Patzer JF 2nd, Gerlach JC. 2010. Transport advances in
disposable bioreactors for liver tissue engineering. Adv Biochem Eng
Biotechnol 115:117–143.
Chen J, Yu Q,WuB, Lin Z, Pavlos NJ, Xu J, OuyangH,Wang A, ZhengMH.
2011. Autologous tenocyte therapy for experimental Achilles tendino-
pathy in a rabbit model. Tissue Eng Part A 17(15–16):2037–2048.
Chen JL, Yin Z, Shen WL, Chen X, Heng BC, Zou XH, Ouyang HW. 2010.
Efficacy of hESC-MSCs in knitted silk-collagen scaffold for tendon
tissue engineering and their roles. Biomaterials 31(36):9438–9451.
Chen JM, Willers C, Xu J, Wang A, Zheng MH. 2007. Autologous tenocyte
therapy using porcine-derived bioscaffolds for massive rotator cuff
defect in rabbits. Tissue Eng 13(7):1479–1491.
Chen X, Qi YY, Wang LL, Yin Z, Yin GL, Zou XH, Ouyang HW. 2008.
Ligament regeneration using a knitted silk scaffold combined with
collagen matrix. Biomaterials 29(27):3683–3692.
Chen YE. 2004. MMP-12, an old enzyme plays a new role in the pathogen-
esis of rheumatoid arthritis? Am J Pathol 165(4):1069–1070.
Dennis RG, Smith B, Philp A, Donnelly K, Baar K. 2009. Bioreactors for
guiding muscle tissue growth and development. Adv Biochem Eng
Biotechnol 112:39–79.
Doroski DM, Levenston ME, Temenoff JS. 2010. Cyclic tensile culture
promotes fibroblastic differentiation of marrow stromal cells encapsu-
lated in poly(ethylene glycol)-based hydrogels. Tissue Eng Part A
16(11):3457–3466.
1506 Biotechnology and Bioengineering, Vol. 110, No. 5, May, 2013
251

Egerbacher M, Arnoczky SP, Caballero O, Lavagnino M, Gardner KL. 2008.
Loss of homeostatic tension induces apoptosis in tendon cells: An in
vitro study. Clin Orthop Relat Res 466(7):1562–1568.
FahlstromM, Jonsson P, Lorentzon R, Alfredson H. 2003. Chronic Achilles
tendon pain treated with eccentric calf-muscle training. Knee Surg
Sports Traumatol Arthrosc 11(5):327–333.
Hannafin JA, Arnoczky SP. 1994. Effect of cyclic and static tensile loading on
water content and solute diffusion in canine flexor tendons: An in vitro
study. J Orthop Res 12(3):350–356.
Hannafin JA, Arnoczky SP, Hoonjan A, Torzilli PA. 1995. Effect of stress
deprivation and cyclic tensile loading on the material and morphologic
properties of canine flexor digitorum profundus tendon: An in vitro
study. J Orthop Res 13(6):907–914.
Issa RI, Engebretson B, Rustom L, McFetridge PS, Sikavitsas VI. 2011. The
effect of cell seeding density on the cellular and mechanical properties
of a mechanostimulated tissue-engineered tendon. Tissue Eng Part A
17(11–12):1479–1487.
Jiang Y, Liu H, Li H,Wang F, Cheng K, Zhou G, ZhangW, YeM, Cao Y, Liu
W, Zou H. 2011. A proteomic analysis of engineered tendon formation
under dynamic mechanical loading in vitro. Biomaterials 32(17):4085–
4095.
Juncosa-Melvin N, Shearn JT, Boivin GP, Gooch C, Galloway MT, West JR,
Nirmalanandhan VS, Bradica G, Butler DL. 2006. Effects of mechanical
stimulation on the biomechanics and histology of stem cell-collagen
sponge constructs for rabbit patellar tendon repair. Tissue Eng
12(8):2291–2300.
Knauper V, Lopez-Otin C, Smith B, Knight G, Murphy G. 1996. Biochemical
characterization of human collagenase-3. J Biol Chem 271(3):1544–1550.
Krane SM, Byrne MH, Lemaitre V, Henriet P, Jeffrey JJ, Witter JP, Liu X,
Wu H, Jaenisch R, Eeckhout Y. 1996. Different collagenase gene
products have different roles in degradation of type I collagen. J
Biol Chem 271(45):28509–28515.
Lavagnino M, Arnoczky SP, Egerbacher M, Gardner KL, Burns ME. 2006.
Isolated fibrillar damage in tendons stimulates local collagenase mRNA
expression and protein synthesis. J Biomech 39(13):2355–2362.
LavagninoM, Arnoczky SP, Tian T, Vaupel Z. 2003. Effect of amplitude and
frequency of cyclic tensile strain on the inhibition of MMP-1 mRNA
expression in tendon cells: An in vitro study. Connect Tissue Res
44(3–4):181–187.
Lea RB, Smith L. 1972. Non-surgical treatment of tendo achillis rupture. J
Bone Joint Surg Am 54(7):1398–1407.
Leppilahti J, Orava S. 1998. Total Achilles tendon rupture. A review. Sports
Med 25(2):79–100.
Li G, Defrate LE, Rubash HE, Gill TJ. 2005. In vivo kinematics of the
ACL during weight-bearing knee flexion. J Orthop Res 23(2):
340–344.
Lin Z, Fitzgerald JB, Xu J, Willers C, Wood D, Grodzinsky AJ, Zheng MH.
2008. Gene expression profiles of human chondrocytes during passaged
monolayer cultivation. J Orthop Res 26(9):1230–1237.
Louis-Ugbo J, Leeson B, Hutton WC. 2004. Tensile properties of fresh
human calcaneal (Achilles) tendons. Clin Anat 17(1):30–35.
Nakama LH, King KB, Abrahamsson S, Rempel DM. 2005. Evidence of
tendon microtears due to cyclical loading in an in vivo tendinopathy
model. J Orthop Res 23(5):1199–1205.
Nguyen TD, Liang R, Woo SL, Burton SD, Wu C, Almarza A, Sacks MS,
Abramowitch S. 2009. Effects of cell seeding and cyclic stretch on the
fiber remodeling in an extracellular matrix-derived bioscaffold. Tissue
Eng Part A 15(4):957–963.
Nirmalanandhan VS, Rao M, Shearn JT, Juncosa-Melvin N, Gooch C,
Butler DL. 2008a. Effect of scaffold material, construct length and
mechanical stimulation on the in vitro stiffness of the engineered
tendon construct. J Biomech 41(4):822–828.
Nirmalanandhan VS, Shearn JT, Juncosa-Melvin N, Rao M, Gooch C, Jain
A, Bradica G, Butler DL. 2008b. Improving linear stiffness of the cell-
seeded collagen sponge constructs by varying the components of the
mechanical stimulus. Tissue Eng Part A 14(11):1883–1891.
Peach MS, James R, Toti US, Deng M, Morozowich NL, Allcock HR,
Laurencin CT, Kumbar SG. 2012. Polyphosphazene functionalized
polyester fiber matrices for tendon tissue engineering: In vitro evalua-
tion with human mesenchymal stem cells. Biomed Mater 7(4):045016.
Rauh J, Milan F, Gunther KP, Stiehler M. 2011. Bioreactor systems for bone
tissue engineering. Tissue Eng Part B Rev 17(4):263–280.
Rauh J, Milan F, Gunther KP, Stiehler M. 2012. Bioreactor systems for bone
tissue engineering. Tissue Eng Part B Rev 17(4):263–280.
Roos EM, Engstrom M, Lagerquist A, Soderberg B. 2004. Clinical improve-
ment after 6 weeks of eccentric exercise in patients with mid-portion
Achilles tendinopathy—A randomized trial with 1-year follow-up.
Scand J Med Sci Sports 14(5):286–295.
Saber S, Zhang AY, Ki SH, Lindsey DP, Smith RL, Riboh J, Pham H, Chang
J. 2010. Flexor tendon tissue engineering: Bioreactor cyclic strain
increases construct strength. Tissue Eng Part A 16(6):2085–2090.
Sahoo S, Ouyang H, Goh JC, Tay TE, Toh SL. 2006. Characterization of a
novel polymeric scaffold for potential application in tendon/ligament
tissue engineering. Tissue Eng 12(1):91–99.
Shalabi A, Kristoffersen-Wilberg M, Svensson L, Aspelin P, Movin T. 2004.
Eccentric training of the gastrocnemius-soleus complex in chronic Achilles
tendinopathy results in decreased tendon volume and intratendinous
signal as evaluated by MRI. Am J Sports Med 32(5):1286–1296.
Silbernagel KG, Thomee R, Thomee P, Karlsson J. 2001. Eccentric overload
training for patients with chronic Achilles tendon pain—A randomised
controlled study with reliability testing of the evaluation methods.
Scand J Med Sci Sports 11(4):197–206.
Skutek M, van Griensven M, Zeichen J, Brauer N, Bosch U. 2001. Cyclic
mechanical stretching modulates secretion pattern of growth factors in
human tendon fibroblasts. Eur J Appl Physiol 86(1):48–52.
Smith LL. 2000. Cytokine hypothesis of overtraining: A physiological
adaptation to excessive stress? Med Sci Sports Exerc 32(2):317–331.
Smith MM, Sakurai G, Smith SM, Young AA, Melrose J, Stewart CM,
Appleyard RC, Peterson JL, Gillies RM, Dart AJ, Sonnabend DH, Little
CB. 2008. Modulation of aggrecan and ADAMTS expression in ovine
tendinopathy induced by altered strain. Arthritis Rheum 58(4):1055–1066.
StanishWD, Rubinovich RM, Curwin S. 1986. Eccentric exercise in chronic
tendinitis. Clin Orthop Relat Res (208): 65–68.
Stoll C, John T, Endres M, Rosen C, Kaps C, Kohl B, Sittinger M, Ertel W,
Schulze-Tanzil. 2010. Extracellular matrix expression of human teno-
cytes in three-dimensional air-liquid and PLGA cultures compared
with tendon tissue: Implications for tendon tissue engineering. J
Orthop Res 28(9):1170–1177.
Wang JH, Jia F, Yang G, Yang S, Campbell BH, Stone D, Woo SL. 2003. Cyclic
mechanical stretching of human tendon fibroblasts increases the produc-
tion of prostaglandin E2 and levels of cyclooxygenase expression: A novel
in vitro model study. Connect Tissue Res 44(3–4):128–133.
Wang T, Gardner B, Lin Z, Rubenson J, Wang A, Kirk TB, Xu J, Smith D,
Lloyd D, Zheng MH. 2012. Bioreactor design for tendon/ligament
engineering. Tissue Eng Part B Rev. [Epub ahead of print] DOI:
10.1089/ten.teb.2012.0295
Webb K, Hitchcock RW, Smeal RM, LiW, Gray SD, Tresco PA. 2006. Cyclic
strain increases fibroblast proliferation, matrix accumulation, and
elastic modulus of fibroblast-seeded polyurethane constructs. J Bio-
mech 39(6):1136–1144.
West JR, Juncosa N, Galloway MT, Boivin GP, Butler DL. 2004. Characteriza-
tion of in vivo Achilles tendon forces in rabbits during treadmill locomo-
tion at varying speeds and inclinations. J Biomech 37(11):1647–1653.
Woon CY, Farnebo S, Schmitt T, Kraus A, Megerle K, Pham H, Yan X,
Gambhir SS, Chang J. 2012. Human Flexor Tendon Tissue Engineering:
Revitalization of Biostatic Allograft Scaffolds. Tissue Eng Part A
18(23–24):2406–2417.
Woon CY, Kraus A, Raghavan SS, Pridgen B.C., Megerle K, PhamH, Chang J.
2011. Three-dimensional-construct bioreactor conditioning in human
tendon tissue engineering. Tissue Eng Part A 17(19–20):2561–2572.
Yang G, ImHJ,Wang JH. 2005. Repetitive mechanical stretchingmodulates
IL-1beta induced COX-2, MMP-1 expression, and PGE2 production in
human patellar tendon fibroblasts. Gene 363:166–172.
Zhang L, Gardiner BS, Smith DW, Pivonka P, Grodzinsky A. 2007. The
effect of cyclic deformation and solute binding on solute transport in
cartilage. Arch Biochem Biophys 457(1):47–56.
Wang et al.: Programmable Mechanical Stimulation 1507
Biotechnology and Bioengineering
252