immunomodulatory spherical nucleic acids · 2015-03-27 · fig. 1. design and properties of...
TRANSCRIPT

Immunomodulatory spherical nucleic acidsAleksandar F. Radovic-Morenoa,1, Natalia Chernyakb,1, Christopher C. Madera, Subbarao Nallagatlaa, Richard S. Kanga,Liangliang Haob, David A. Walkerb, Tiffany L. Haloa, Timothy J. Merkelb, Clayton H. Rischea, Sagar Anantatmulaa,Merideth Burkharta, Chad A. Mirkinb,2, and Sergei M. Gryaznova,2
aAuraSense Therapeutics, LLC, Skokie, IL 60077; and bDepartment of Chemistry, International Institute for Nanotechnology, Northwestern University,Evanston, IL 60208
Contributed by Chad A. Mirkin, February 11, 2015 (sent for review January 6, 2015; reviewed by Marcin Kortylewski and Arthur Krieg)
Immunomodulatory nucleic acids have extraordinary promise fortreating disease, yet clinical progress has been limited by a lack oftools to safely increase activity in patients. Immunomodulatorynucleic acids act by agonizing or antagonizing endosomal toll-likereceptors (TLR3, TLR7/8, and TLR9), proteins involved in innateimmune signaling. Immunomodulatory spherical nucleic acids(SNAs) that stimulate (immunostimulatory, IS-SNA) or regulate(immunoregulatory, IR-SNA) immunity by engaging TLRs havebeen designed, synthesized, and characterized. Compared withfree oligonucleotides, IS-SNAs exhibit up to 80-fold increases inpotency, 700-fold higher antibody titers, 400-fold higher cellularresponses to a model antigen, and improved treatment of micewith lymphomas. IR-SNAs exhibit up to eightfold increases in po-tency and 30% greater reduction in fibrosis score in mice withnonalcoholic steatohepatitis (NASH). Given the clinical potentialof SNAs due to their potency, defined chemical nature, and goodtolerability, SNAs are attractive new modalities for developingimmunotherapies.
nanotechnology | vaccines | TLRs | immune regulation | oligonucleotides
Modulating immunity in a targeted and specific manner is anattractive approach for treating disease. Promising early
results have been observed in preclinical and clinical studies,particularly in infectious disease (1, 2), cancer (3–5), allergy (6–8), and autoimmune disease applications (9, 10). Several classesof agents are currently being evaluated for their ability to stim-ulate or repress immunity, but few are as versatile as nucleicacids. Nucleic acids can stimulate immunity by binding endo-somal toll-like receptors (TLR3, TLR7/8, and TLR9), a findingthat has been used to develop immunotherapies (11, 12). Nucleicacids can also antagonize endosomal TLRs to regulate aberrantimmunity and treat autoimmune disorders such as psoriasis (13)and systemic lupus erythematosus (14). Despite these advances,immunomodulatory nucleic acids have not yet reached their fullpotential in the clinic. A key challenge pertains to how thera-peutic biological activity in humans can be safely increased.Nanomaterials have shown potential for increasing immuno-
modulatory nucleic acid activity. Efforts have primarily focusedon constructs for immunostimulation (IS) and include com-plexing oligonucleotides with albumin (15), gold nanoparticleswith two-component external coronas consisting of IS-oligonu-cleotides together with protein antigens (16), gold nanoparticle–IS-oligonucleotide conjugates (17), IS-oligonucleotides conju-gated to polymeric nanoparticles (18), and IS-oligonucleotides incomplex with lipids (19). Although these studies have uncoveredthe promise of nanomaterials for immunomodulation, it is stillunclear what the dominant structural factors are. This informationis important not only to increase activity, but also to simplify andenhance manufacturability for effective clinical translation. Inaddition, there has been a relative lack of advances for nano-structured materials that use oligonucleotides to specifically de-activate immune cells, despite the promising potential of thisapproach to treat autoimmune diseases in humans.Our group has pioneered the development of spherical nucleic
acids (SNAs), a class of well-defined macromolecules, which are
formed by organizing nucleic acids radially around a nano-particle core (20, 21). These structures exhibit the ability to entercells without the need for auxiliary delivery vehicles or trans-fection reagents, by engaging scavenger receptors and lipidrafts (22). Various SNAs have been evaluated for their po-tential to treat a wide variety of diseases and conditions thatmay not be easily addressable with conventional therapeuticstrategies (23, 24). SNAs, especially ones containing naturalphosphodiester nucleic acids, are internalized into variouscells, mainly via caveolin-mediated endocytosis, leading toendosomal localization (25, 26). Once inside the cell, the nucleicacid components of SNAs resist nuclease degradation, leadingto longer intracellular lifetimes. Moreover, SNAs, due to theirmultifunctional chemical structures, have the ability to bind theirtargets in a high-affinity multivalent fashion (25, 26). Therefore,in addition to demonstrating many of the characteristics thatwould be considered preferred for designing immunomodulatorytherapies, SNAs also provide a chemical and structural scaffoldfor evaluating the structure–activity relationships for the twomajor components: the oligonucleotide shell and the nano-particle core. Indeed, SNAs can be assembled in many differentstructural forms, with their cores consisting of inorganic com-pounds, such as gold, or organic materials such as <50-nm lipo-somes (27, 28).Herein, we describe a previously unreported class of immu-
nomodulatory SNAs, which can be used either for stimulating(immunostimulatory, IS-SNAs) or regulating (immunoregulatory,IR-SNA) immune responses (Fig. 1). The functional activity ofthese SNAs is defined by the type of oligonucleotide pharmaco-phores used to assemble the SNA structures—oligonucleotide
Significance
We show that by organizing immunomodulatory nucleic acidsinto spherical nucleic acid (SNA) form, significant increases inactivity are observed. Treatment of mice with cancer usingimmunostimulatory SNAs and nonalcoholic steatohepatitis(NASH) using immunoregulatory SNAs leads to improved dis-ease outcomes vs. their unstructured counterparts. These im-provements derive from several key SNA properties, includingrapid cellular uptake, endosomal delivery, and multivalent bind-ing. Overall, this work underscores the importance of the spatialorientation and presentation of oligonucleotides in the design ofnovel immunomodulators.
Author contributions: A.F.R.-M., N.C., C.C.M., S.N., R.S.K., C.A.M., and S.M.G. designedresearch; A.F.R.-M., N.C., C.C.M., S.N., R.S.K., L.H., T.L.H., T.J.M., C.H.R., S.A., and M.B.performed research; A.F.R.-M., N.C., C.C.M., S.N., R.S.K., D.A.W., C.H.R., C.A.M., andS.M.G. analyzed data; and A.F.R.-M., N.C., C.A.M., and S.M.G. wrote the paper.
Reviewers: M.K., City of Hope; and A.K., Checkmate Pharmaceuticals.
Conflict of interest statement: C.A.M. is a cofounder and A.F.R.-M., S.N., R.S.K., C.H.R.,S.A., M.B., and S.M.G. are employees of AuraSense Therapeutics, LLC.1A.F.R.-M. and N.C. contributed equally to this work.2To whom correspondence may be addressed. Email: [email protected] [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1502850112/-/DCSupplemental.
3892–3897 | PNAS | March 31, 2015 | vol. 112 | no. 13 www.pnas.org/cgi/doi/10.1073/pnas.1502850112
Dow
nloa
ded
by g
uest
on
June
14,
202
0

TLR agonists or antagonists, respectively. In addition to thismodularity, we demonstrate that the properties of SNAs leadto significant activity improvements compared with free oligo-nucleotide counterparts for both IS-SNAs and IR-SNAs in vitroand in vivo. Furthermore, through systematic modification ofmultiple components that comprise the SNA structure, in-cluding oligonucleotide sequence, sugar-phosphate backbonechemistry, orientation, and core template, we are able to elucidatethe key structural factors that provide for enhanced immunomod-ulatory function.
Results and DiscussionCellular Internalization Pathway for IS- and IR-SNAs. Similarly toprevious observations involving other cell lines (25, 26), immu-nomodulatory SNAs (Fig. 1A) are taken up by RAW 264.7macrophages, a model antigen-presenting cell (APC) line, viaendocytosis (Fig. 1B). Using confocal fluorescence microscopy,we observed rapid internalization of fluorophore-labeled SNAsbearing immunostimulatory motifs in RAW 264.7 macrophages(Fig. 1 C and D and Fig. S1). We also found that the SNAconstructs associate with cells via class A scavenger receptors(SRA), as IS-SNA internalization was dramatically decreasedin the presence of an SRA inhibitor, fucoidan (Fig. 1 D and E).Furthermore, immunostaining studies show that the SNAs arefound primarily in the endosomes at early time points (Fig.1F). These data suggest that IS-SNAs enter immune cells viareceptor-mediated endocytosis and accumulate primarily inendosomes.
SNA 3D Structure Leads to Enhanced Immunomodulatory Activity inVitro. We next sought to evaluate whether 3D organization of ISoligonucleotides into SNAs substantially changes their baselineactivity in vitro. To do this, we synthesized a family of compoundsbased on the TLR9 agonist oligonucleotide, CpG 1826, witheither phosphodiester (PO) or phosphorothioate (PS) back-bones, in free or SNA forms, using the conventional 13-nm goldcore template (Au-SNA) (21). Following an overnight incu-bation of IS-SNAs with RAW-Blue macrophages, an 80-foldincrease in potency (as measured by EC50 values) was observedfor PO SNA CpG 1826 compared with the free oligonucleotidecounterpart (Fig. 2A). For PS CpG 1826, the differences in ac-tivity of IS-SNAs compared with the corresponding free oligo-nucleotides were highly time-dependent and in all cases higherthan the PO counterparts, as expected (29). At relatively shortincubation times (30 min), a 94-fold increase in potency (EC50value) was observed, which decreased to a 1.8-fold higher potencyafter 4 h of the treatment, ultimately becoming comparable inpotency after overnight incubation (Fig. 2 A and B and Fig. S2).This marked kinetic difference, which likely is the result of dif-ferences in uptake rates, may be beneficial with regard to in vivo(vide infra) applications of these SNA constructs. We also syn-thesized the corresponding CpG 1826-based compounds with
Fig. 1. Design and properties of immunomodulatory spherical nucleic acids.(A) IS-SNAs, which contain a shell of TLR agonist oligonucleotides or IR-SNAs,which contain a shell of TLR antagonist oligonucleotides, (B) are both takenup into immune cells and reside in endosomes where they modulate toll-likereceptor (TLR) activity. (C) Confocal microscopy image showing Cy-5–labeledAu-SNAs have been internalized. (D and E) Probing the importance of classA scavenger receptors in the uptake pathway for Cy-5–labeled (red signal)L-SNAs: (D) SNAs freely enter cells in the absence of SR-A receptor ligandfucoidan. (E) Cellular uptake of the same SNAs is inhibited in the presence offucoidan. (F) Confocal microscopy image of Cy-5–labeled L-SNAs (red signal)colocalizing (yellow signal) with Alexa Flour 488–labeled EEA-1 marker(green signal) after 1 h of uptake. Cells were fixed before immunostainingand imaging.
Fig. 2. IS-SNAs demonstrate increased potency vs. matched unformulatedoligonucleotides in vitro. IS-SNAs formed with oligonucleotides (as in-dicated) and a 13-nm Au nanoparticle core were tested for their ability toinduce NF-κB following incubation with RAW-Blue macrophages: (A) over-night or (B) 30 min incubation. (C) A similar set of IS-SNAs with small uni-lamellar liposomal vesicle cores (L-SNA) were tested following overnightincubation. (D) Production of TNF-α following 48-h incubation with RAW264.7 cells at 0.4 μM concentration. (E) Evaluation of IS-SNA activity inRamos-Blue cells compared with liposomes with TLR9 agonist oligonucleo-tides facing inward, with or without a d-A20 all-PO oligonucleotide (5′-d-A20-3′).Results are representative of two or more independent experiments; pointsshow mean ± SD (F) Immunoregulatory SNAs (IR-SNA) demonstrate en-hanced potency vs. matched unformulated oligonucleotides in vitro. IR-SNAsformed using the TLR9 antagonist sequence (4084F) were tested for theirability to inhibit activation of RAW-Blue macrophages in Au-SNA form(mean ± SD).
Radovic-Moreno et al. PNAS | March 31, 2015 | vol. 112 | no. 13 | 3893
CHEM
ISTR
YIM
MUNOLO
GYAND
INFLAMMATION
Dow
nloa
ded
by g
uest
on
June
14,
202
0

a liposomal core (L-SNA) instead of a gold core and found thatthe IS activity was nearly identical and independent of core,for both PO and PS backbones (Fig. 2C). We also found thatfree 3′-tocopherol oligonucleotides showed similar activity to theSNAs (Fig. S2G); this led us to hypothesize that the 3′-tocopherololigonucleotides were forming small micellar structures underthese conditions. We confirmed formation of the micellularstructures in PBS by dynamic light scattering (see SI Materials andMethods for details).We corroborated our findings of enhanced NF-κB activation
in the APC reporter system by IS-SNAs by demonstrating that(i) IS-SNAs in RAW 264.7 murine macrophages generally inducehigher levels of several proinflammatory cytokines regulatedby NF-κB (TNF-α, IL-12, and IL-6) compared with free oligo-nucleotides at the same dose (Fig. 2D and Fig. S2 B, C, and F);(ii) NF-κB activation by IS-SNAs was dependent on the presenceof functional TLR9 (Fig. S2 D and E); (iii) TLR9 activation byIS-SNAs could be replicated in a similar human B-cell reportersystem (Ramos-Blue cells; Fig. S2 D and G); (iv) inversion of theoligonucleotide orientation relative to SNA core by placement ofthe functionally important 5′-terminus (30) internally and imme-diately adjacent to the SNA core greatly reduced the compound’sactivity (Fig. S2H); (v) use of a different TLR9 agonist oligonu-cleotides, all-PO CpG 7909 (more optimized for interacting withhuman TLR9), against human PBMCs, resulted in markedlyhigher cytokine activation in SNA form (∼1,400-fold higher pro-duction of TNF-α) compared with linear either all-PO or all-PSoligonucleotides (Fig. S2F). This improvement in activity in hu-man cells (for CpG 7909 constructs) vs. murine cells (for CpG1826 counterparts) may be due to the previously described dif-ferences in TLR receptor affinities and the preferences of oligo-nucleotide phosphodiester vs. phosphorothioate backbones forhuman and murine TLR ligands, respectively (31); and (vi) thestudied IS-SNAs did not induce significant production of IFN-α inhPBMCs in vitro under the conditions studied (Fig. S2 J–L). Takentogether, these results strongly suggest that the 3D structured
oligonucleotide pharmacophore shell, and not the core, are themain determinant of IS activity.Next, we sought to evaluate whether it is advantageous to or-
ganize IS-oligonucleotides into IS-SNAs vs. encapsulating themin liposomes of comparable size. To do this, we prepared an ISL-SNA (external CpG 7909-PO L-SNA) and a correspondingliposome with the IS-oligonucleotides encapsulated (internalCpG 7909-PO liposome) and compared them in terms of theirability to activate signaling in the NF-κB reporter cells. Ourresults show that the outward-facing L-SNA structure led to anapproximate threefold improvement in potency following over-night incubation (Fig. 2E). To test whether this improvement wassimply the result of greater SNA-mediated uptake, we took theinternal CpG 7909-PO liposome structures and postfunction-alized them with immunologically inactive d-A20 all-PO oligo-nucleotide and tested their activity. The internal CpG/externald-A20 L-SNAs showed no difference compared with the oligo-nucleotide-encapsulated liposome, suggesting that external pre-sentation of oligonucleotides in SNA format confers otheradvantages in addition to uptake. These advantages may includepresentation of the oligonucleotide pharmacophore in a moreavailable conformation for interactions with the target receptor.Next, we tested the IR-SNAs (both Au-SNA and L-SNA) as
potential negative regulators of TLR activity in vitro. IR-SNAswere designed based on the TLR9 antagonist oligonucleotide,4084F (32). To evaluate TLR antagonist activity, cells were firstactivated by incubation with PS CpG 1826 (at 0.5 μM) for 2 h.The cells were then treated with various antagonists: 4084F-derivatized Au-SNA, free 4084F all-PS oligonucleotide, or mis-matched oligonucleotide controls. The results demonstrate that4084F-derivatized Au-SNAs show an approximate eightfold in-crease in potency compared with the free 4084F all-PS counterpart(Fig. 2F). Similar trend in the activity was observed with L-SNAantagonist constructs designed with 4084F sequences (Fig. S3A).These data further suggest that the outer oligonucleotide shell, andnot the SNA core, is the defining factor for the increased IRproperties. Last, we confirmed that the administration of IR-SNAs
Fig. 3. IS-SNAs induce systemic responses. IS-SNAs (Au-SNA) were administered into the footpad and the draining (popliteal) LN was collected aseptically at(A) t = 1 h and (B) t = 24 h and placed into RPMI-1640 growth medium for overnight incubation. The LNs were silver stained to reveal the presence of the Aunanoparticle core and counterstained with eosin. (C) The supernatants from LNs collected at t = 4 h and incubated overnight were probed for cytokines,including IL-12 at multiple doses. (D) Full cytokine panel showing specificity for TLR9 oligonucleotide motifs. (E) Cytokine panel collected from serum at t = 4 hpost footpad administration. (F) Elevated and prolonged induction of IL-12 following systemic administration (n = 6 mice per group pooled to n = 3 toincrease sample volume, points or bars show mean ± SD).
3894 | www.pnas.org/cgi/doi/10.1073/pnas.1502850112 Radovic-Moreno et al.
Dow
nloa
ded
by g
uest
on
June
14,
202
0
(Au-SNA) inhibited production of a key proinflammatory proteindownstream of NF-κB, TNF-α (Fig. S3B).
IS-SNA 3D Structure Leads to More Potent and Durable Activation ofInnate Immune Cells in Vivo. Next, we sought to evaluate the po-tential of PS CpG 1826 Au-SNA (IS-SNA) constructs to activateinnate immune cells in vivo. We first confirmed that Au-basedIS-SNAs accumulate in draining lymph nodes in mice (LNs) atshort time points following intradermal injection (t = 1, 4, and24 h postinjection). The histological sections show evidence ofsignificant accumulation of Au-based IS-SNAs in the cortical andmedullary LN regions as early as 1 h postinjection (Fig. 3A).Progressive enlargement of lymph nodes with time was also ob-served, which is consistent with IS-SNA–induced local cellularactivation (Fig. 3B and Fig. S4).To test which formulation of nucleic acid induces more robust
draining LN responses, we compared the levels of cytokinesproduced by the aseptically collected LNs at varying oligonu-cleotide doses. The results demonstrate that IS-SNA formulationof PS CpG 1826 produces a ∼10-fold increase in the amounts ofseveral secreted proinflammatory cytokines compared with freeIS-oligonucleotide, particularly for IL-12 and IFN-γ (Fig. 3 Cand D and Fig. S5). Importantly, the increased IS potency didnot appear to be an intrinsic feature of the SNA 3D structureitself, because a control SNA (containing TLR9 inactive GpCreplacing CpG motif) was essentially inactive (Fig. 3D); giventhe robust local LN cytokine response, we sought to evaluatewhether this led to elevations in a representative set of cytokinesin the serum. Despite the profound IS effect in the draining LN,we saw no notable increases in serum levels for a panel of 12different cytokines (Fig. 3E).Next, we sought to determine whether IS-SNA structures
could be used to increase the activity of systemically adminis-tered (i.v. administration) oligonucleotides. To evaluate the ef-fect of IS-SNA compounds, we administered PO or PS CpG 1826in either free or in Au-SNA form, and then measured the serumcytokines over a 6-h period. The results show that SNAs, par-ticularly PS-containing SNAs, induce up to 10-fold higher levelsof serum proinflammatory cytokines, particularly for IL-12 (Fig.3F and Fig. S6). In addition, the duration of the stimulatoryeffect appeared to be extended by several hours compared withfree oligonucleotides.
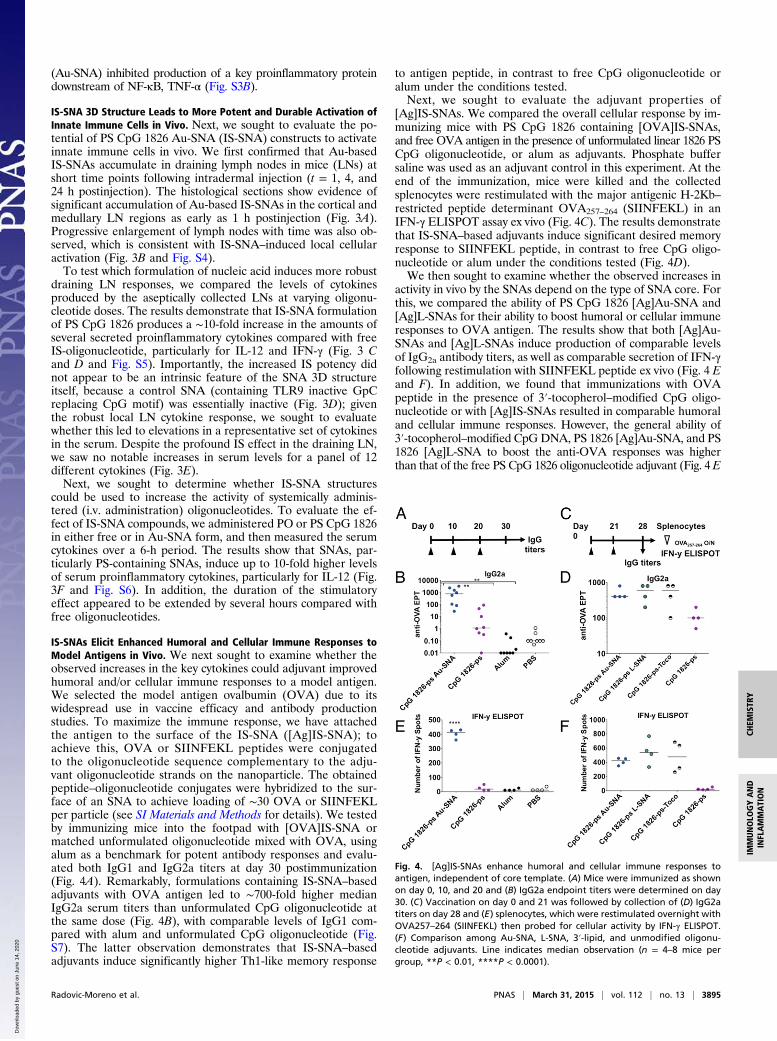
IS-SNAs Elicit Enhanced Humoral and Cellular Immune Responses toModel Antigens in Vivo. We next sought to examine whether theobserved increases in the key cytokines could adjuvant improvedhumoral and/or cellular immune responses to a model antigen.We selected the model antigen ovalbumin (OVA) due to itswidespread use in vaccine efficacy and antibody productionstudies. To maximize the immune response, we have attachedthe antigen to the surface of the IS-SNA ([Ag]IS-SNA); toachieve this, OVA or SIINFEKL peptides were conjugatedto the oligonucleotide sequence complementary to the adju-vant oligonucleotide strands on the nanoparticle. The obtainedpeptide–oligonucleotide conjugates were hybridized to the sur-face of an SNA to achieve loading of ∼30 OVA or SIINFEKLper particle (see SI Materials and Methods for details). We testedby immunizing mice into the footpad with [OVA]IS-SNA ormatched unformulated oligonucleotide mixed with OVA, usingalum as a benchmark for potent antibody responses and evalu-ated both IgG1 and IgG2a titers at day 30 postimmunization(Fig. 4A). Remarkably, formulations containing IS-SNA–basedadjuvants with OVA antigen led to ∼700-fold higher medianIgG2a serum titers than unformulated CpG oligonucleotide atthe same dose (Fig. 4B), with comparable levels of IgG1 com-pared with alum and unformulated CpG oligonucleotide (Fig.S7). The latter observation demonstrates that IS-SNA–basedadjuvants induce significantly higher Th1-like memory response
to antigen peptide, in contrast to free CpG oligonucleotide oralum under the conditions tested.Next, we sought to evaluate the adjuvant properties of
[Ag]IS-SNAs. We compared the overall cellular response by im-munizing mice with PS CpG 1826 containing [OVA]IS-SNAs,and free OVA antigen in the presence of unformulated linear 1826 PSCpG oligonucleotide, or alum as adjuvants. Phosphate buffersaline was used as an adjuvant control in this experiment. At theend of the immunization, mice were killed and the collectedsplenocytes were restimulated with the major antigenic H-2Kb–restricted peptide determinant OVA257–264 (SIINFEKL) in anIFN-γ ELISPOT assay ex vivo (Fig. 4C). The results demonstratethat IS-SNA–based adjuvants induce significant desired memoryresponse to SIINFEKL peptide, in contrast to free CpG oligo-nucleotide or alum under the conditions tested (Fig. 4D).We then sought to examine whether the observed increases in
activity in vivo by the SNAs depend on the type of SNA core. Forthis, we compared the ability of PS CpG 1826 [Ag]Au-SNA and[Ag]L-SNAs for their ability to boost humoral or cellular immuneresponses to OVA antigen. The results show that both [Ag]Au-SNAs and [Ag]L-SNAs induce production of comparable levelsof IgG2a antibody titers, as well as comparable secretion of IFN-γfollowing restimulation with SIINFEKL peptide ex vivo (Fig. 4 Eand F). In addition, we found that immunizations with OVApeptide in the presence of 3′-tocopherol–modified CpG oligo-nucleotide or with [Ag]IS-SNAs resulted in comparable humoraland cellular immune responses. However, the general ability of3′-tocopherol–modified CpGDNA, PS 1826 [Ag]Au-SNA, and PS1826 [Ag]L-SNA to boost the anti-OVA responses was higherthan that of the free PS CpG 1826 oligonucleotide adjuvant (Fig. 4 E
Fig. 4. [Ag]IS-SNAs enhance humoral and cellular immune responses toantigen, independent of core template. (A) Mice were immunized as shownon day 0, 10, and 20 and (B) IgG2a endpoint titers were determined on day30. (C) Vaccination on day 0 and 21 was followed by collection of (D) IgG2atiters on day 28 and (E) splenocytes, which were restimulated overnight withOVA257–264 (SIINFEKL) then probed for cellular activity by IFN-γ ELISPOT.(F) Comparison among Au-SNA, L-SNA, 3′-lipid, and unmodified oligonu-cleotide adjuvants. Line indicates median observation (n = 4–8 mice pergroup, **P < 0.01, ****P < 0.0001).
Radovic-Moreno et al. PNAS | March 31, 2015 | vol. 112 | no. 13 | 3895
CHEM
ISTR
YIM
MUNOLO
GYAND
INFLAMMATION
Dow
nloa
ded
by g
uest
on
June
14,
202
0

and F). Again we confirmed that the 3′-tocopherol–modified CpGoligonucleotides were forming SNA-like micellar structures in situvia dynamic light scattering. These data provide further evidencethat the oligonucleotide spatial 3D organization and the phar-macophore orientation play key roles in providing the highlybeneficial immunologic properties of IS-SNAs.
IS-SNAs Reduce Tumor Growth Rates and Improve Animal Survivalin Lymphoma Model. We next sought to determine whether[Ag]IS-SNAs could demonstrate improved efficacy in cancer im-munotherapy disease model. For this purpose, we used thewell-established E.G7-OVA lymphoma syngeneic flank modelas a model for a cancer expressing a well-defined antigen (OVA).To produce OVA-expressing tumors, we injected 2 × 106 E.G7-OVA lymphoma cells into the flank of C57BL/6 mice. Followingthe tumor cell injection (on day 0), we treated mice with either[Ag]IS-SNAs or appropriate controls on days 3, 5, and 7 (Fig.5A). The results show that SNA-boosted anti-OVA responseswere able to induce profound and durable tumor growth re-mission (Fig. 5B) that translated into a doubling of mouse sur-vival rates (Fig. 5C). Use of the full OVA protein or the majorantigenic determinant SIINFEKL peptide in vaccination ap-peared to show no difference in either tumor growth rates orsurvival under these conditions (Fig. S8). Interestingly, OVA-free formulations also exhibited a significant reduction in tumorgrowth rates and improvements in mouse survival. The latterobservation could be attributed to the induction of innate im-mune responses. Moreover, significant differences were ob-served in tumor growth rates in vaccinations containing L-SNAsand 3′-tocopherol–modified oligonucleotides. Thus, results showa significant reduction in tumor growth rates by the 3D struc-tured L-SNAs compared with free 3′-tocopherol–modified CpGoligonucleotides, which also correlates with positive differencesin mouse survival. Importantly, for all adjuvant formulations, andin all animal groups, no signs of gross toxicity were observed(Fig. S8C).
IR-SNAs Demonstrate Enhanced Antifibrotic Activity in a MouseModel of NASH. To further elucidate therapeutic potential ofIR-SNAs, we sought to determine whether SNA structure ex-hibited advantages over sequence- and chemistry-matched free
unformulated TLR antagonist oligonucleotides in vivo. Non-alcoholic steatohepatitis (NASH) is a fatty liver disease withprevalence as high as 3–5% in the United States. It is charac-terized by pathologic changes in livers of patients, includingsteatosis and inflammation, which ultimately progress in a subset ofpatients to nodular fibrosis, full-blown cirrhosis, and hepatocellularcarcinoma (HCC). Currently, there are no FDA-approved treat-ments for this disease.The STAM mouse model has been developed as a small ani-
mal model that recapitulates important features of the humandisease (33). Nearly 100% of STAM mice follow disease pro-gression from steatosis to NASH to fibrosis to HCC, making thisa model that is well suited to evaluate the ability of compoundsto prevent disease progression. In this model, mice that are 6 wkold demonstrate evidence of NASH, and from 6 to 9 wk oldprogress to histological evidence of fibrosis.To assess whether IR-SNAs could affect disease progression,
we began treatment at week 6 with IR-SNAs (L-SNA-4084F-Ext,L-SNA-4084F), sequence-matched unformulated free oligonu-cleotides (4084F-Ext, 4084F), or controls (L-SNA-CTL, PBS)and continued treatment following an every-other-day schedulefor the 3-wk period to week 9 (Fig. 5D). After 9 wk, the mice inall groups were killed, and their livers were collected, processedfor histology (H&E and Sirius Red), and scored with regards tofibrosis (Fig. 5E and Fig. S9) and NASH (Fig. 5F and Fig. S9).The results demonstrate that IR-SNA administration leads toa 40–51% reduction in fibrosis relative to control; this is incontrast to unformulated free oligonucleotides, which showedmore modest antifibrotic effects (21–25% reduction). NeitherIR-SNAs nor unformulated oligonucleotides showed any signif-icant effect on nonalcoholic fatty liver disease score in this study.These results suggest that IR-SNAs can potentially be developedinto compounds that modify rates of NASH progression to fi-brosis, an advance that could reduce mortality from this disease.
ConclusionThe results presented in this manuscript are significant for thefollowing reasons. The data convincingly show that SNAs can beused as constructs for sequence-specific, potent, and therapeu-tically relevant immunostimulation and immunoregulation.Importantly, such constructs outperform linear nonlipidated
Fig. 5. SNAs demonstrate immunomodulatory prop-erties in vivo. (A) [Ag]IS-SNAs enhance tumor clear-ance. Mice were inoculated with 2 × 106 E.G7-OVAcells into the right flank on day 0, then treated asindicated with one treatment on days 3, 5, and 7.(B) Tumor growth curves (n = 11 mice per group, errorbars show mean ± SEM) and (C) corresponding sur-vival endpoints. (D) IR-SNAs show enhanced treatmentof liver fibrosis in mice with NASH. STAM mice weretreated every other day from 6 to 9 wk of age. At9 wk, the liver was collected and processed for(E) fibrosis score by Sirius Red staining and (F) non-alcoholic fatty liver disease activity (NAS) score by H&E(n = 8 mice per group; points show mean ± SEM).
3896 | www.pnas.org/cgi/doi/10.1073/pnas.1502850112 Radovic-Moreno et al.
Dow
nloa
ded
by g
uest
on
June
14,
202
0

phosphodiester deoxynucleic acids due to a rapid cellular up-take, predominant accumulation in endosomes, and increasedresistance to nucleases. Moreover, the data suggest that many ofthe attractive SNA properties are defined primarily by the 3Darrangement and orientation of the oligonucleotide shell forboth IS and IR applications. Taken together, the presented dataunderscore the importance of spatial orientation of the immu-nomodulatory pharmacophores, and highlight the modular na-ture of the SNA therapeutic constructs.
Materials and MethodsSpherical Nucleic Acids, Adjuvants, Antigens, and Reagents. Oligonucleotideswere synthesized using automated solid support phosphoramidite syn-thesis (sequences for activating and regulating immunity are included in SIMaterials and Methods). Ovalbumin (Sigma-Aldrich) or SIINFEKL (GenScript)were purchased at their highest purity and used as-is without additionalpurification. The 13-nm Au-SNAs were prepared as described (21) with im-portant modifications (SI Materials and Methods) and formulated to con-tain 5.2% (wt/wt) (L-SNA) or 6.3% (wt/wt) (Au-SNA) of oligonucleotide.L-SNAs were synthesized as described (27) with modifications (SI Materialsand Methods and Fig. S10).
Antibody Titers. All animal studies were conducted according to protocolsapproved by the local Institutional Animal Care and Use Committees ofExplora BioLabs, SRI Biosciences, Stelic Institute & Co., Inc., and Ricerca Bio-sciences, and by the local Ethical Review Process committee of KWS BioTest.Anti-OVA antibody endpoint titers were determined by ELISA. In brief, micewere bled as indicated in the figure captions either on day 28 or day 30following the first immunization. Either OVA323–339 peptide (Fig. 5B and Fig.S6) or ovalbumin protein (Fig. 5E) was used as the capture antigen, and anIgG or IgG isotype-specific (IgG1 or IgG2) was used as the secondary antibodylinked to HRP. The endpoint titer was determined via linear regressionanalysis using a curve with six or more points.
Ex Vivo ELISPOT. The OVA-specific cellular response was evaluated by IFN-γELISPOT. In brief, at the conclusion of the in-life portion (day 28), mice werekilled and spleens were processed to splenocytes by mechanical disruption ofthe spleen and removal of RBC by hypotonic lysis. Splenocytes were seeded
at 4 × 105 cells per well of IFN-γ ELISPOT plates together with media alone(negative control), 1 μM OVA257–264, or PMA/ionomycin (positive control).Following incubation overnight, the number of IFN-γ–secreting cells wasquantified by an automated ELISPOT counter.
In Vivo E.G7-OVA Tumor Growth Study. The 2 × 106 E.G7-OVA (American TypeCell Culture Collection) cells were inoculated into the right flank of 4- to6-wk-old C57BL/6 mice on day 0. On days 3, 5, and 7 post tumor induction,200 μL of compound were administered. Tumor volumes were measuredtwice a week by quantifying the length and width of the tumor and ap-plying the formula V = L × W × W/2. If a second tumor occurred in a givenmouse, the second tumor was measured via the same method and the twovolumes were added together. The general health of animals was monitoreddaily. Mice were euthanized if and when tumor volume reached or exceeded2,000 mm3 or if found to be moribund.
Liver Fibrosis Prevention Study. The activity of IR-SNAs was tested in the STAMmouse model (Stelic Institute & Co., Inc.) of NASH. In brief, pathogen-free,15-d pregnant C57BL/6 mice were obtained from Japan SLC. NASH was in-duced by s.c. injection of streptozotocin (Sigma-Aldrich) after birth followedby feeding with a high fat diet (CLEA Japan) ad libitum after 4 wk of age. At6 wk old, mice were randomized into groups of eight and treated everyother day from 6 to 9 wk old (3 wk). Compounds or control (PBS) wereadministered (40 μM oligonucleotide equivalent concentration) in 100 μL byi.p. administration. The impact of treatment administration on NASH andliver fibrosis was determined by collecting livers at 9 wk of age and assessingNASH disease and fibrosis histologically in blinded fashion. In brief, H&E-stained liver sections were scored for steatosis (0–3), lobular inflammation(0–3), and hepatocellular ballooning (0–2), after which a composite non-alcoholic fatty liver disease score was assigned (scale: 0–8). Sections were alsostained with Sirius Red for fibrosis and scored as percent positive areaaccording to established protocols.
ACKNOWLEDGMENTS. We thank Andrew Schook, Blake Tutterow, andShweta Iyer for technical assistance. This material is based upon worksupported by the Center for Cancer Nanotechnology Excellence initiative ofthe National Institutes of Health Award U54 CA151880 and the DefenseAdvanced Research Projects Agency Grant HR0011-13-2-0018.
1. Krieg AM (2006) Therapeutic potential of Toll-like receptor 9 activation. Nat Rev DrugDiscov 5(6):471–484.
2. Autran B, Carcelain G, Combadiere B, Debre P (2004) Therapeutic vaccines for chronicinfections. Science 305(5681):205–208.
3. Cheever MA, Higano CS (2011) PROVENGE (Sipuleucel-T) in prostate cancer: The firstFDA-approved therapeutic cancer vaccine. Clin Cancer Res 17(11):3520–3526.
4. Robert C, et al. (2015) Nivolumab in previously untreated melanoma without BRAFmutation. N Engl J Med 372(4):320–330.
5. Melero I, et al. (2014) Therapeutic vaccines for cancer: An overview of clinical trials.Nat Rev Clin Oncol 11(9):509–524.
6. Anderson RJ, et al. (2014) A self-adjuvanting vaccine induces cytotoxic T lymphocytesthat suppress allergy. Nat Chem Biol 10(11):943–949.
7. Fraser CC (2010) Nanoparticle therapy for allergic and inflammatory disease. Anti-Inflammatory Anti-Allergy Agents Med Chem 9(1):54–70.
8. Creticos PS, et al.; Immune Tolerance Network Group (2006) Immunotherapywith a ragweed-toll-like receptor 9 agonist vaccine for allergic rhinitis. N Engl J Med355(14):1445–1455.
9. Getts DR, et al. (2014) Therapeutic inflammatory monocyte modulation using immune-modifying microparticles. Sci Transl Med 6(219):ra7.
10. Kontos S, Kourtis IC, Dane KY, Hubbell JA (2013) Engineering antigens for in situerythrocyte binding induces T-cell deletion. Proc Natl Acad Sci USA 110(1):E60–E68.
11. Cooper C, Mackie D (2011) Hepatitis B surface antigen-1018 ISS adjuvant-containing vaccine:A review of HEPLISAV™ safety and efficacy. Expert Rev Vaccines 10(4):417–427.
12. Zhu X, et al. (2007) Toll like receptor-3 ligand poly-ICLC promotes the efficacy ofperipheral vaccinations with tumor antigen-derived peptide epitopes in murine CNStumor models. J Transl Med 5:10–25.
13. Jiang W, et al. (2013) A Toll-like receptor 7, 8, and 9 antagonist inhibits Th1 and Th17responses and inflammasome activation in a model of IL-23-induced psoriasis. J InvestDermatol 133(7):1777–1784.
14. Zhu FG, et al. (2013) A novel antagonist of Toll-like receptors 7, 8 and 9 suppresseslupus disease-associated parameters in NZBW/F1 mice. Autoimmunity 46(7):419–428.
15. Liu H, et al. (2014) Structure-based programming of lymph-node targeting in mo-lecular vaccines. Nature 507(7493):519–522.
16. Lee IH, et al. (2012) Imageable antigen-presenting gold nanoparticle vaccines foreffective cancer immunotherapy in vivo. Angew Chem Int Ed Engl 51(35):8800–8805.
17. Lin AY, et al. (2013) Gold nanoparticle delivery of modified CpG stimulates macrophagesand inhibits tumor growth for enhanced immunotherapy. PLoS ONE 8(5):e63550.
18. de Titta A, et al. (2013) Nanoparticle conjugation of CpG enhances adjuvancy for cellularimmunity and memory recall at low dose. Proc Natl Acad Sci USA 110(49):19902–19907.
19. Nguyen DN, et al. (2012) Lipid-derived nanoparticles for immunostimulatory RNAadjuvant delivery. Proc Natl Acad Sci USA 109(14):E797–E803.
20. Mirkin CA, Letsinger RL, Mucic RC, Storhoff JJ (1996) A DNA-based method for rationallyassembling nanoparticles into macroscopic materials. Nature 382(6592):607–609.
21. Cutler JI, Auyeung E, Mirkin CA (2012) Spherical nucleic acids. J Am Chem Soc 134(3):1376–1391.
22. Patel PC, et al. (2010) Scavenger receptors mediate cellular uptake of polyvalentoligonucleotide-functionalized gold nanoparticles. Bioconjug Chem 21(12):2250–2256.
23. Jensen SA, et al. (2013) Spherical nucleic acid nanoparticle conjugates as an RNAi-based therapy for glioblastoma. Sci Transl Med 5(209):ra152.
24. Zheng D, et al. (2012) Topical delivery of siRNA-based spherical nucleic acid nano-particle conjugates for gene regulation. Proc Natl Acad Sci USA 109(30):11975–11980.
25. Choi CH, Hao L, Narayan SP, Auyeung E, Mirkin CA (2013) Mechanism for the endo-cytosis of spherical nucleic acid nanoparticle conjugates. Proc Natl Acad Sci USA110(19):7625–7630.
26. Wu XA, Choi CH, Zhang C, Hao L, Mirkin CA (2014) Intracellular fate of sphericalnucleic acid nanoparticle conjugates. J Am Chem Soc 136(21):7726–7733.
27. Banga RJ, Chernyak N, Narayan SP, Nguyen ST, Mirkin CA (2014) Liposomal sphericalnucleic acids. J Am Chem Soc 136(28):9866–9869.
28. Lee JS, Lytton-Jean AK, Hurst SJ, Mirkin CA (2007) Silver nanoparticle-oligonucleotideconjugates based on DNA with triple cyclic disulfide moieties. Nano Lett 7(7):2112–2115.
29. Krieg AM, et al. (1995) CpG motifs in bacterial DNA trigger direct B-cell activation.Nature 374(6522):546–549.
30. Yu D, Zhao Q, Kandimalla ER, Agrawal S (2000) Accessible 5′-end of CpG-containingphosphorothioate oligodeoxynucleotides is essential for immunostimulatory activity.Bioorg Med Chem Lett 10(23):2585–2588.
31. Hartmann G, Krieg AM (2000) Mechanism and function of a newly identified CpGDNA motif in human primary B cells. J Immunol 164(2):944–953.
32. Lenert P, et al. (2009) DNA-like class R inhibitory oligonucleotides (INH-ODNs) pref-erentially block autoantigen-induced B-cell and dendritic cell activation in vitro andautoantibody production in lupus-prone MRL-Fas(lpr/lpr) mice in vivo. Arthritis ResTher 11(3):R79–R95.
33. Fujii M, et al. (2013) A murine model for non-alcoholic steatohepatitis showingevidence of association between diabetes and hepatocellular carcinoma. Med MolMorphol 46(3):141–152.
Radovic-Moreno et al. PNAS | March 31, 2015 | vol. 112 | no. 13 | 3897
CHEM
ISTR
YIM
MUNOLO
GYAND
INFLAMMATION
Dow
nloa
ded
by g
uest
on
June
14,
202
0