identification of a major pathogenic epitope in the human irbp
TRANSCRIPT

Identification of a Major Pathogenic Epitopein the Human IRBP Molecule Recognizedby Mice of the H-2r Haplotype
Phyllis B. Silver,* Luiz V. Rizzo,* Chi-Chao Chan,* Larry A. Donoso,~\Barbara Wiggert,% and Rachel R. Caspi*
Purpose. Mice of the H-2b, H-2k, and H-2r haplotypes develop experimental autoimmuneuveoretinitis (EAU) after immunization with interphotoreceptor retinoid-binding protein(IRBP) of bovine or monkey origin. The purpose of this study was to identify putative patho-genic epitope(s) of IRBP and to establish their immunodominance within the IRBP molecule.
Methods. Overlapping 20-amino acid peptides, spanning the entire human IRBP molecule,were synthesized and used to immunize C57BL/10 (H-2b), B10.BR (H-2k), and B10.RIII (H-2r) mice. Bovine IRBP was used as a positive control. Experimental autoimmune uveoretinitiswas examined by histopathology 21 days after immunization. Immunologic responses wereassessed by delayed-type hypersensitivity (DH) and lymphocyte proliferation assays.
Results. Peptide 161-180, spanning the sequence SGIPYIISYLHPGNTILHVD, was found tobe highly pathogenic for B10.RIII mice but not for the other strains. A dose-response curveshowed that peptide 161-180 was maximally pathogenic at 50 /xg, but incidence and scoreswere reduced at 10 ,ug. The truncated 13-mer 165-177 was also highly pathogenic (100 to200 /Ug), suggesting that it contained the pathogenic epitope. Mice immunized with thepeptide, or with whole IRBP, had positive DH and lymphocyte responses to the immunizingas well as to the reciprocal antigen. A cell line derived to peptide 161-180 was also pathogenicfor B10.RIII mice after adoptive transfer and responded (proliferation) to native IRBP.
Conclusions. High incidence and high severity scores, as well as immunologic cross-recognitionof peptide 161-180 and native IRBP in vivo and in vitro, suggest that this peptide containsa major epitope recognized as pathogenic by B10.RIII mice. Invest Ophthalmol Vis Sci.1995;36:946-954.
Jtixperimental autoimmune uveoretinitis (EAU) is anorgan-specific, T-cell mediated autoimmune diseasethat serves as a model for several human sight-threat-ening ocular disorders, among them sympathetic oph-thalmia, birdshot retinochoroidopathy, Behcet's dis-ease, and Vogt-Koyanagi-Harada syndrome. Pathol-ogy in the animal model resembles the human diseaseand is characterized by posterior retinal and choroidallesions, photoreceptor damage, vasculitis, vitritis,
From the * Ijiboratory of Immunology and. the %Laboratory of Cell and MolecularBiology, National Eye. Institute, Belhesda, Maryland, and the i Wills Eye Hospital,Philadelphia, Pennsylvania.Supported in part by a Research to Prevent Blindness Senior Scientific Investigatoraward (IJlD).Submitted far publication July 14, 1994; revised October 25, 1994; acceptedOctolier 28, 1994.Prof/rietaiy interest category: N.Reprint requests: Phyllis B. Silver, laboratory of Immunology, National EyeInstitute, NIH Building 10, Room 10N2I8, Bethesda, Ml) 20892.
granuloma formation, and varying degrees of anteriorchamber infiltration.1"3
Experimental autoimmune uveoretinitis can be in-duced in several animal models by immunization widiretinal antigens emulsified in complete Freund's adju-vant or by the adoptive transfer of retinal antigen-specificT lymphocytes into syngeneic recipients.'"7 We recentlydeveloped the mouse model for EAU using the inter-photoreceptor retinoid-binding protein (IRBP) as theuveitogen.8 The uveitogenic molecule IRBP is a 140-kd glycolipoprotein residing in the interphotoreceptormatrix between the neural retina and the retinal pig-ment epithelium. Interphotoreceptor retinoid-bindingprotein is thought to be involved in the transport ofvitamin A derivatives between the retinal pigment epi-thelium and the photoreceptors.9 It consists of a fourfoldrepeat structure with 30% to 40% amino acid sequencehomology shared between repeats.1" Interphotoreceptor
946Investigative Ophthalmology & Visual ScCopyright © Association for Research in
:e, April 1095, Vol. 36, No. 5ion and Ophthalmology
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933185/ on 04/12/2018
An IRBP Epitope Pathogenic for H-2r Mice 947
TABLE i. Peptide 161-180 Is Strongly Pathogenic for Mice of the H-21 Haplotype
ExperimentNumber
1
2
Mouse Strain
C57BL/10 (H-2b)B10.BR (H-2k)BIO.RIII (H-2r)BIO.RIII
Incidence*
0/50/55/5
20/20
161-180
Score ± SE\
00
1.8 ± 0.32.7 ± 0.09
hicidence*
5/55/55/5
20/20
IRBP
Score ±
1.2 ±2.0 ±2.85 ±3.2 ±
SE\
0.300.40.1
* Incidence = positive/iotal.f Average disease score ± standard error of animals that developed EAU. Mice were immunized either with 50 /ig of peptide 161-180 orwith 50 //g of bovine IRBP. EAU was scored by histopathology 21 days after immunization.EAU = experimental autoimmune uveoreiinitis; IRBP = interphotoreceptor reu'noid-binding protein.
retinoid-binding protein is an evolutionarily conservedmolecule with approximately 80% sequence homologyat the amino acid level between the bovine and thehuman protein.9'10
The mouse EAU model is rapidly becoming animportant tool for the study of uveitic disease. In itsclinical manifestations, murine EAU closely resemblessome types of human uveitis.8 Because the mouse iswithout a doubt the best-characterized species geneti-cally as well as immunologically, it offers a uniquemodel in which to study genetic cellular and molecu-lar aspects of ocular autoimmunity. As in some humanuveitic diseases that have strong HLA associations, sus-ceptibility to EAU of different mouse strains is in partcontrolled by major histocompatibility complexgenes."111 Provided that a "permissive" (e.g., B10)genetic background is present, mice of the haplotypesH-2', H2\ and H-2b develop EAU after immunizationwith IRBP, whereas many other haplotypes are resis-tant.12 In these and numerous subsequent studies us-ing different doses of whole IRBP (unpublished obser-vations), it has become evident that the hierarchy ofsusceptibility is, in descending order, H-2' > H-k >H-2b. However, to perform in-depth genetic and im-munologic studies, the uveitogenic epitopes of IRBP
TABLE 2. Uveitogenicity of Different Dosesof Peptide 161-180 in BIO.RIII Mice
Immunizing Antigen (fig)
Peptide50102
IRBP50
Incidence*
6/64/60/6
6/6
Score ± SEf
2.1 ± 0.31.1 ± 0.5
0
2.3 ± 0.2
* Incidence = positive/total.f Average disease score ± standard error of animals thatdeveloped EAU.Mice were immunized with the indicated doses of peptide 161-180 or bovine IRBP. EAU was scored by histopathology 21 daysafter immunization.EAU = experimental autoimmune uveoretinitis; IRBP =interphotoreceptor retinoid-binding protein.
recognized by the different haplotypes must be identi-fied and characterized.
In an earlier study,13 we tested recombinant pro-teins, expressed in bacteria, that contained the firsttwo or the second two repeats of bovine IRBP. Afteranalyzing the pattern of uveitic responses to these pro-teins in three strains representing the known EAU-susceptible haplotypes, i.e., C57BL/10 (H-2b), B10.BR(H-2k), and BIO.RIII (H-21), it was possible to predictthat a major pathogenic epitope for the H-2' haplo-type resided in the first repeat of bovine IRBP. In anattempt to identify the putative pathogenic site, wetested 20-amino acid peptides representing the entirefirst repeat of human IRBP that had been synthesizedpreviously.14 This approach relied on the assumptionthat the pathogenic sequence in question had beenevolutionarily preserved between the human and bo-vine IRBP molecules. We show that the human IRBPpeptide 161-180 (sequence SGIPYHSYLHPGN-TILHVD) is uveitogenic in BIO.RIII (H-21) mice andappears to contain a major pathogenic site immuno-dominant for this haplotype. This same sequence wasimmunogenic, but not pathogenic, in the other twohaplotypes. To our knowledge, this is the first descrip-tion of an immunodominant epitope of IRBP thatcauses EAU in mice.
MATERIALS AND METHODS
Animals
Six- to 8-week-old C57BL/10 (H-2b), B10.BR (H-2k),and BIO.RIII (H-21) mice were supplied by JacksonLaboratories (Bar Harbor, ME) or were bred at theNational Institutes of Health animal facility. All proce-dures adhered to the ARVO Statement for the Use ofAnimals in Ophthalmic and Vision Research.
Antigens
Human IRBP peptide sequence 161-180 was synthe-sized by conventional solid-phase techniques, as de-scribed.1415 Shorter peptides of human IRBP peptide161-180 and the bovine IRBP sequence 161-180 were
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933185/ on 04/12/2018

948 Investigative Ophthalmology &: Visual Science, April 1995, Vol. 36, No. 5
synthesized by Chiron Mimotopes (Emeryville, CA)using Fmoc chemistry. Whole bovine IRBP was puri-fied by concanavalin A Sepharose 4B (Pharmacia, Pis-cataway, NJ) affinity chromatography and fast-perfor-mance liquid chromatography.16
EAU Induction and Evaluation ofHistopathology
Peptides or IRBP were emulsified in completeFreund's adjuvant (1:1 wt/vol) containing 2.5 mg/ml Mycobacterium tuberculosis H37Ra (Sigma, St. Louis,MO). A total of 200 fi\ of emulsion was injected subcu-taneously, divided among three sites—the base of thetail and both thighs. Concurrent with immunization,1.0 fig of Bordetdla pertussis toxin (Sigma) was injectedintraperitoneally. Eyes were collected 21 days after im-munization, fixed in 4% phosphate-buffered glutaral-dehyde for 1 hour, and then transferred to 10% phos-phate-buffered formaldehyde until processing. Fixedand dehydrated tissue was embedded in methacrylate,and 4- to 6-/zm sections were stained by standard hema-toxylin and eosin. Eye sections cut at different planeswere scored in a masked fashion. Incidence and sever-ity of EAU were graded on a scale of 0 to 4 as describedpreviously,8 based on the size, number, and type oflesions present. Incidence is shown as the number ofpositive animals out of all animals in the group. Sever-ity of disease is the average score of eyes from thoseanimals that developed disease (if disease was unilat-eral, both eyes were averaged).
Delayed-type Hypersensitivity
Twenty micrograms of IRBP or peptide in 20 fj\ phos-phate-buffered saline was injected into the ear pinna.Ear thickness was measured 48 hours later with aspring-loaded micrometer. The response was calcu-lated as the difference between ear thickness beforeand after challenge. Statistical analysis was by indepen-dent Rest in comparison to naive controls. Probabilityvalues of < 0.05 were considered significant. Eachgroup was composed of five to eight mice.
Lymphocyte Proliferation Assay
Each group was composed of six mice. Inguinal andiliac lymph nodes were collected after 14 days, and thecells were pooled within the group. Triplicate cultureswere seeded in 96-well, round-bottom plates at a con-centration of 5 X 105 cells/well in RPMI 1640 supple-mented as described*5 with 2.5 //.g/ml peptide 161 —180 or 50 /zg/ml IRBP. The cultures were incubatedfor a total of 60 hours, with a pulse of [3H] thymidine(1.0 /xCi/10 (i\ per well) administered during the last18 hours. The data are presented as ACPM (ACPM= mean cpm in cultures with stimulus — mean cpmin control cultures without stimulus).
FIGURE l. (A) Experimental autoimmune uveoretinitis pa-thology in B10.RIII mice immunized vvir.li 50 //g IRBP (dis-ease grade 3). Note thickened choroid (C), infiltrating in-flammatory cells [small arrows) in the vitreous (V), retina(R) and choroid, granuloma {open arrow), retinal folding,and detachment, (B) EAU pathology in B10.RI1I mice im-munized with 50 fig peptide 161-180 (disease grade 3).Note thickened choroid, inflammatory cells in the vitreousand choroid, retinal detachment, granulomas, subretinalhemorrhage (asterisk), destruction of photoreceptor archi-tecture. (C) Normal retinal structure of an eye of a B10.BRmouse immunized with 50 fj,g peptide 161-180. Hematoxy-lin and eosin; original magnification, X20.
Derivation of T Cell Lines Specific to Peptide161-180B10.RIII mice were immunized with 50 fig peptide161-180 by the usual protocol. Inguinal and iliaclymph nodes were collected after 6 days. Cells wereseeded at 1 X 10(i cells per well in 24-well Linbro plates
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933185/ on 04/12/2018

An IRBP Epitope Pathogenic for H-2r Mice
TABLE 3. Uveitogenicity of Truncated Peptides Spanning Positions161-180
949
Position
161-180163-177165-179165-177168-180
Sequence
SGIPYIISYLHPGNTILHVDIPYIISYLHPGNTIL
YIISYLHPGNTILHVYIISYLHPGNTIL
SYLHPGNTILHVD
Incidence*
8/84/55/63/70/8
EAU Score ± SE\
2.7 ± 0.22.1 ± 0.52.4 ± 0.10.2 ± 0.3
0
* Incidence = positive/total.f Average disease score ± standard error of animals that developed EAU.B10.R1II mice were immunized with 50 /xg of the indicated peptides. EAU was scored byhistopathology 21 days after immunization.EAU = experimental autoimmune uveoretinitis.
r
(ICN, Irvine, CA) in Dulbecco's minimal essential me-dium (Hyclone, Logan, UT) containing 1% mouseserum and 1.0 ^xg/ml peptide. Some wells were supple-mented with anti-IL-4 (11B11 hybridoma, generouslyprovided by Dr. W. Paul, National Institutes of Health)(line A) or with 0.05 p.g/ml rIL-12 (generously pro-vided by Stanley Wolf, Genetics Institute, Boston, MA)(line B) to encourage development of TH 1 type cells.After 48 hours, cells were suspended in expansionmedium consisting of Dulbecco's minimal essentialmedium, 10% fetal calf serum, and 5% to 10% IL-2-containing ConA conditioned medium, and then cul-tured for 5 days. During the first month, the devel-oping lines were restimulated weekly with antigen for2 days (in the presence of 2.5 X 10fi syngeneic spleno-cytes per milliliter as antigen-presenting cells) andthen cultured for 5 days in IL-2-containing expansionmedium. Thereafter, the lines were restimulated onceevery 2 to 3 weeks. To test pathogenicity of the Tcell lines, freshly restimulated cells were harvested,washed, and injected intravenously into naive B10.RIIImice. Experimental autoimmune uveoretinitis was as-sessed by histopathology 10 to 12 days after adoptivetransfer.
TABLE 4. Uveitogenicity of Increasing Dosesof Peptide 165-177 in B10.RIII Mice
Immunizing Antigen (fig)
Peptide50
100200
IRBP50
Incidence*
2/44/58/8
4/5
Score ± SE\
0.5 ± 01.6 ± 0.42.4 ± 0.4
2.5 ± 0.3
* Incidence = positive/total.f Average disease score ± standard error of animals thatdeveloped EAU.Mice were immunized with the indicated doses of peptide 165-177 or bovine IRBP. EAU was scored by histopathology 21 daysafter immunization.EAU = experimental autoimmune uveoretinits; IRBP =interphotoreceptor retinoid-binding protein.
RESULTS
Human IRBP Peptide 161-180 Contains aMajor Pathogenic Epitope for B10.RIII MiceB10.RIII mice, as well as C57BL/10 and B10.BR mice,were immunized with 50 .̂g of each of 30 peptidesrepresenting the first repeat of human IRBP. BovineIRBP was used as control. Peptide 161-180 inducedEAU in B10.RIII mice (H-2r) with high scores andhigh incidence; C57BL/10 (H-2b) and B10.BR (H-2k)mice, though susceptible to uveitis induced with wholeIRBP, did not develop EAU after immunization withthe peptide (Table 1). The dose response showed that10 fig of peptide 161-180 was sufficient to inducedisease in B10R.III mice, though score and incidencewere reduced compared to the 50-fj.g dose. A dose of2.0 /xg of peptide was insufficient to induce EAU (Ta-ble 2). Low-grade disease was induced with peptide1-20 (GPTHLFQPSLVLDMAKVLLD) in the C57BL/10 mice and with peptide 171-190 (HPGNTILHVD-TIYNRPSNTT) in B10.RIII mice. These responses arebeing characterized and will be the subject of a subse-quent report. None of the other peptides derived fromthe first repeat of human IRBP were uveitogenic un-der the conditions described.
Figure 1 illustrates the ocular pathology resultingfrom active immunization with peptide 161-180 (Fig.IB). As in IRBP-induced uveitis (Fig. 1A), inflamma-tion was localized at the choroid and the retina of theeye. Subretinal hemorrhages and extensive destruc-tion of the photoreceptor architecture were observed.Retinal vasculitis and perivasculitis, retinal folds, andgranuloma formation were evident. Mild to moderatevitritis also occurred. Figure 1C shows a healthy eyefrom a B10.BR mouse (resistant to peptide 161-180-induced EAU) after immunization with the peptide.
The 13-mer Peptide 165-177 Contains theCore Epitope of 161-180
To define more precisely the pathogenic site of pep-tide 161-180, truncated peptides were synthesizedand their uveitogenicity in B10.RIII mice was evalu-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933185/ on 04/12/2018
950 Investigative Ophthalmology & Visual Science, April 1995, Vol. 36, No. 5
ated. The shortest peptides to induce EAU at the50-fJ.g dose, with incidence and scores comparableto the 20-mer peptide 161-180, were the 15-merpeptides 163-177 and 165-179 (Table 3). Fromthese results, it is apparent that the isoleucine andproline residues on 163-177 and the histidine andvaline residues on 165-179 were not required fordisease induction. Therefore, peptide 165-177 wassynthesized. This 13-mer peptide was the shortestpeptide to induce any EAU at the standard 50-fj.gdose. Because smaller peptides may require higherdoses to induce comparable disease, we tested theuveitogenicity of graded doses of peptide 165-177.Results showed that a progressively increasing im-munization dose resulted in increased disease inci-dence and scores. Disease scores at the 200-/Ug doseapproached that of the IRBP-immunized mice (Ta-ble 4). These results suggested that peptide 165-177 contains the core epitope of 161-180.
Immunodominance of Peptide 161-180
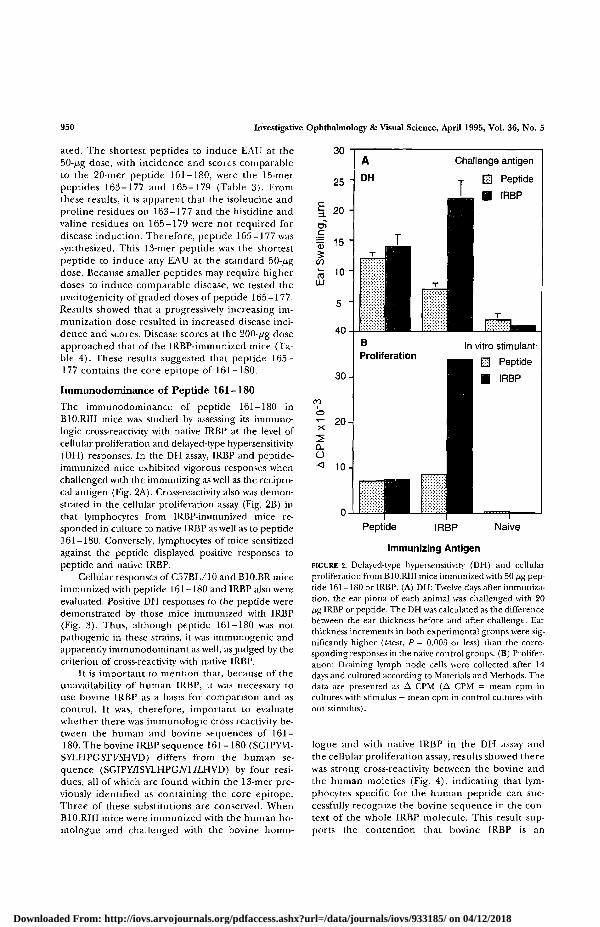
The immunodominance of peptide 161-180 inB10.RIII mice was studied by assessing its immuno-logic cross-reactivity with native IRBP at the level ofcellular proliferation and delayed-type hypersensitivity(DH) responses. In the DH assay, IRBP and peptide-immunized mice exhibited vigorous responses whenchallenged with the immunizing as well as the recipro-cal antigen (Fig. 2A). Cross-reactivity also was demon-strated in the cellular proliferation assay (Fig. 2B) inthat lymphocytes from IRBP-immunized mice re-sponded in culture to native IRBP as well as to peptide161-180. Conversely, lymphocytes of mice sensitizedagainst the peptide displayed positive responses topeptide and native IRBP.
Cellular responses of C57BL/10 and B10.BR miceimmunized with peptide 161-180 and IRBP also wereevaluated. Positive DH responses to the peptide weredemonstrated by those mice immunized with IRBP(Fig. 3). Thus, although peptide 161-180 was notpathogenic in these strains, it was immunogenic andapparently immunodominant as well, as judged by thecriterion of cross-reactivity with native IRBP.
It is important to mention that, because of theunavailability of human IRBP, it was necessary touse bovine IRBP as a basis for comparison and ascontrol. It was, therefore, important to evaluatewhether there was immunologic cross-reactivity be-tween the human and bovine sequences of 161 —180. The bovine IRBP sequence 161-180 (SGIPYW-SYLHPGSTVSHVD) differs from the human se-quence (SGIPY7ISYLHPGM7LHVD) by four resi-dues, all of which are found within the 13-mer pre-viously identified as containing the core epitope.Three of these substitutions are conserved. WhenB10.RIII mice were immunized with the human ho-mologue and challenged with the bovine homo-
Peptide IRBP Naive
Immunizing Antigen
FIGURE 2. Delayed-type hypersensitivity (DH) and cellularproliferation from B10.RIII mice immunized with 50 /zg pep-tide 161-180 or IRBP. (A) DH: Twelve days after immuniza-tion, the ear pinna of each animal was challenged with 20/ig IRBP or peptide. The DH was calculated as the differencebetween the ear thickness before and after challenge. Earthickness increments in both experimental groups were sig-nificantly higher (/-test, P = 0.003 or less) than the corre-sponding responses in the naive control groups. (B) Prolifer-ation: Draining lymph node cells were collected after 14days and cultured according to Materials and Methods. Thedata are presented as A CPM (A CPM = mean cpm incultures with stimulus — mean cpm in control cultures with-out stimulus).
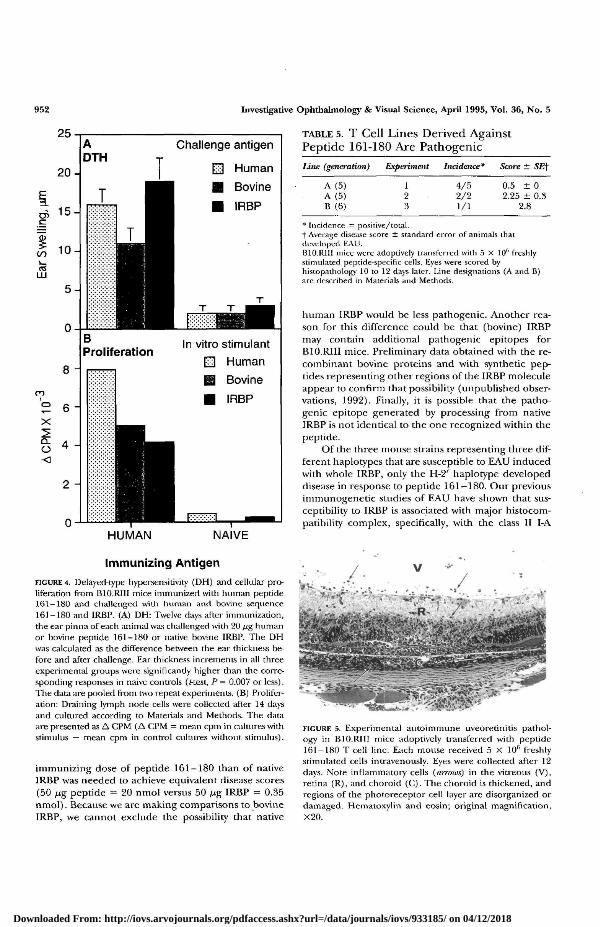
logue and with native IRBP in the DH assay andthe cellular proliferation assay, results showed therewas strong cross-reactivity between the bovine andthe human moieties (Fig. 4), indicating that lym-phocytes specific for the human peptide can suc-cessfully recognize the bovine sequence in the con-text of the whole IRBP molecule. This result sup-ports the contention that bovine IRBP is an
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933185/ on 04/12/2018

An IRBP Epitope Pathogenic for H-2r Mice
50
40-
E 30
! 2 0 'CO
S io-l
ImmunizingAntigen
Strain
951
- - T -
flrl
Challenge
mm
T
antigen
Peptide
IRBP
IRBP IRBP IRBP Peptide Peptide Peptide Naive
B10.RIII C57BL B10.BR B10.RIII C57BL B10.BR (All)
FIGURE 3. Delayed-type hypersensitivity (DH) from B10.RIII, C57BL/10, and B10.BR miceimmunized with 50 /xg peptide 161-180 or IRBP. Twelve days after immunization, the earpinna of each animal was challenged with 20 fj,g IRBP or peptide. The DH was calculatedas the difference between the ear thickness before and after challenge. Ear thickness incre-ments in all experimental groups were significantly higher than the corresponding responsesin the naive control groups (f-test, P = 0.004 or less). The data are pooled from two repeatexperiments.
acceptable substitute for h u m a n IRBP as control inthese exper iments .
A T Cell Line Specific to Peptide 161-180Induces EAU and Proliferates to Native IRBP
Two T cell lines, A and B, specific to 161-180, werederived from primed lymph node cells of B10.RIIImice and were maintained by periodic restimulationwith antigen, as described in Materials and Methods.Recipient mice were injected with 5 X 106 freshly re-stimulated cells, and the eyes were tested for pathol-ogy. Table 5 shows that the lines were invariably patho-genic, although the disease induced with differentbatches of cells varied in severity. By histopathology,the disease induced by adoptive transfer of the T celllines closely resembled that induced by active immuni-zation. Features included serous retinal detachment,infiltrating inflammatory cells in the subretinal spaceand retina, and multiple areas of photoreceptor dam-age (Fig. 5). Both cell lines also exhibited strong pro-liferative responses to 20 Mg/ml IRBP and 2 fig/mlIRBP in culture in a dose-response manner (data notshown). The recognition of peptide 161-180 withinwhole IRBP by the pathogenic T cell lines providesadditional support for an immunodominant status ofthe epitope contained within this peptide.
DISCUSSION
This study has shown that the 20-amino acid peptideof human IRBP, 161-180, contains a major site that
is pathogenic for B10.R1II (H-21) mice, but not forB10.BR (H-2k) or C57BL/10 (H-2b) mice. The histopa-thology induced with this pathogenic site was severeand closely mimicked disease induced by native IRBP,i.e. ocular inflammation with focal lesions, retinal de-tachment, vasculitis and perivasculitis, granuloma for-mation in the retina and choroid, and destructionof the photoreceptor layer. A shorter 13-amino acidsequence, 165-177, possessed the requisite aminoacids for induction of disease and was identified ascontaining the core epitope of 161-180.
Our contention that 161-180 contains a majorpathogenic site is based on the severity of the diseaseinduced by relatively low doses of this peptide and itsimmunodominance. Immunodominance, evidencedby immunologic cross-reactivity between the peptideand the native IRBP, was observed at the level of DH,lymph node cell responses, and by recognition ofIRBP by the long-term pathogenic T cell lines derivedagainst 161-180. However, had we not observed im-munologic cross-reactivity, it would still not constituteevidence that the site is not immunodominant. Thisis because the comparison was to bovine IRBP; humanIRBP was unavailable for testing. Although cross-reac-tivity between the bovine and the human moieties of161-180 strongly suggest that the same site is recog-nized in IRBP of both species, we cannot exclude thepossibility that the fine specificity was not for the samecore sequence because our experiments were per-formed with the 20-mer peptides.
It is notable that, on a molar basis, a much higher
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933185/ on 04/12/2018
952 Investigative Ophthalmology & Visual Science, April 1995, Vol. 36, No. 5
25
o>
inmLU
20-
1 5 -
10-
ADTH
Challenge antigen
E2] Human
Bovine
IRBP
ProliferationIn vitro stimulant
Human
Bovine
IRBP
TABLE 5. T Cell Lines Derived AgainstPeptide 161-180 Are Pathogenic
Line (generation) Experiment Incidence* Score ± SEf
A (5)A (5)B (6)
4/52/21/1
0.5 ± 02.25 ± 0.3
2.8
HUMAN NAIVE
* Incidence — positive/total.f Average disease score ± standard error of animals thatdeveloped EAU.B10.R11I mice were adoptively transferred with 5 X 106 freshlystimulated peptide-specific cells. Eyes were scored byhistopathology 10 to 12 days later. Line designations (A and B)are described in Materials and Methods.
human IRBP would be less pathogenic. Another rea-son for this difference could be that (bovine) IRBPmay contain additional pathogenic epitopes forB10.RJII mice. Preliminary data obtained with the re-combinant bovine proteins and with synthetic pep-tides representing other regions of the IRBP moleculeappear to confirm that possibility (unpublished obser-vations, 1992). Finally, it is possible that the patho-genic epitope generated by processing from nativeIRBP is not identical to the one recognized within thepeptide.
Of the three mouse strains representing three dif-ferent haplotypes that are susceptible to EAU inducedwith whole IRBP, only the H-2' haplotype developeddisease in response to peptide 161-180. Our previousimmunogenetic studies of EAU have shown that sus-ceptibility to IRBP is associated with major histocom-patibility complex, specifically, with the class II I-A
Immunizing Antigen
FIGURE 4. Detayed-type hypersensitivity (DH) and cellular pro-liferation from B10.RIII mice immunized with human peptide161-180 and challenged with human and bovine sequence161-180 and IRBP. (A) DH: Twelve days after immunization,the ear pinna of each animal was challenged with 20 /ig humanor bovine peptide 161-180 or native bovine IRBP. The DHwas calculated as die difference between the ear thickness be-fore and after challenge. Ear thickness increments in all threeexperimental groups were significantly higher than the corre-sponding responses in naive controls (Rest, P= 0.007 or less).The data are pooled from two repeat experiments. (B) Prolifer-ation: Draining lymph node cells were collected after 14 daysand cultured according to Materials and Methods. The dataare presented as A CPM (A CPM = mean cpm in cultures withstimulus — mean cpm in control cultures without stimulus).
immunizing dose of peptide 161-180 than of nativeIRBP was needed to achieve equivalent disease scores(50 fig peptide = 20 nmol versus 50 ^g IRBP = 0.35nmol). Because we are making comparisons to bovineIRBP, we cannot exclude the possibility that native
FIGURE 5. Experimental autoimmune uveoretinitis pathol-ogy in B10.RIII mice adoptively transferred with peptide161-180 T cell line. Each mouse received 5 X 10fi freshlystimulated cells intravenously. Eyes were collected after 12days. Note inflammatory cells (mrows) in the vitreous (V),retina (R), and choroid (C). The choroid is thickened, andregions of the photoreceptor cell layer are disorganized ordamaged. Hematoxylin and eosin; original magnification,X20.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933185/ on 04/12/2018

An IRBP Epitope Pathogenic for H-2r Mice 953
subregion, suggesting that class II-restricted presenta-tion of an epitope is connected to pathogenicity. Thepresent results indicate that different sites of the IRBPmolecule must function as immunopathogenic epi-topes for the three EAU-susceptible haplotypes. Thisconclusion is in line with our previous findings thatidentified human IRBP peptide 461-480 as a minorepitope for the C57BL/10 strain (H-2b), but not forB10.BR (H-2k) or B10.RIII (H-21) strains.17 Parallelresults had been reported by other investigators inthe rat EAU model. Hu et al's showed that differentepitopes of IRBP were uveitogenic in different strainsof rats. Hirose and coworkers19'20 reported that pep-tide M of S-Ag, uveitopathogenic in Lewis (RT11) andWKAH.1L (RT11) rats, is nonpathogenic in WKAH rats(RTlk). These investigators concluded that develop-ment of EAU in rats was in part dependent on themajor histocompatibility complex haplotype.
Interestingly, peptide 161-180 was immunogenicand apparently immunodominant as well in the H-2k
and H-2b haplotypes. This observation raises a ques-tion: Why, of three haplotypes that respond immuno-logically to the peptide, was the response pathogenicin only one? One explanation is that the three haplo-types in fact do not recognize the same epitope withinthe peptide. In vivo processing of 161-180 might re-sult in several different products, and the class II mole-cules of the three H-2 haplotypes could preferentiallybind and present different fragments, resulting in dif-ferent fine specificity of the recognition. According tothis line of reasoning, and assuming that all threehaplotypes process antigen similarly, only the particu-lar fragment(s) presented by H-2' class II moleculeswould be sufficiently similar to the fragment(s) gener-ated in vivo from autologous IRBP to elicit pathogenic-ity. This notion is supported by our recent observationthat the autologous mouse homologue of 161-180 isimmunogenic as well as pathogenic in B10.RIII mice,but not in the other two strains, and that it cross-reactsimmunologically with the human and bovine peptidesequences as well as with native bovine IRBP (unpub-lished observations, 1994). Immunogenicity withoutpathogenicity of peptides also has been seen in otheruveitogenic antigens and in other autoimmune dis-ease models. Fling and coworkers21 demonstrated adissociation between pathogenicity and immunoge-nicity of epitopes derived from the retinal soluble anti-gen (S-Ag). Bovine S-Ag peptides that were nonpatho-genic in Lewis rats were able to stimulate lymph nodecells in vitro. Conversely, when the autologous rat se-quence was synthesized and used as the immunogen,disease occurred but in vitro cellular proliferation wasabsent. Similarly, lymph node cells from mice immu-nized with certain peptides from myelin basic proteinwere able to proliferate in vitro to the peptide, butthe animal failed to develop encephalitis.22
Further study of the relationships between se-
quence, immunogenicity, and pathogenicity of closelyrelated homologues of evolutionarily conserved au-toantigens might help to predict autologous se-quences that could be immunopathogenic for differ-ent HLA types in human autoimmune disease.
Key Words
uveitis, EAU, mouse, autoimmune disease, peptides
References
1. Fame JP. Autoimmunity and the retina. Curr Top EyeRes. 1980; 2:215-301.
2. Gery I, Mochizuki M, Nussenblatt RB. Retinal specificantigens and immunopathogenic processes they pro-voke. Prog Retinal Res. 1986;5:75-109.
3. Caspi RR. Basic mechanisms in immune-mediateduveitic disease. In: Lightman SL, ed. Immunology of EyeDisease. Lancaster, UK: Kluwer Academic Publishers.1989:61-86.
4. Mochizuki M, Kuwabara T, McAllister C, NussenblattRB, Gery I. Adoptive transfer of experimental autoim-mune uveoretinitis in rats: Immunopathogenic mech-anisms and histologic features. Invest Ophthalmol VisSci. 1985; 26:1-9.
5. Caspi RR, Roberge FG, McAllister CG, et al. T celllines mediating experimental autoimmune uveoretin-itis (EAU) in the rat. JImmunol. 1986; 136:1928-1933.
6. Gregerson DS, Obritsch WF, Fling SP, Cameron JD.T cell lines recognize peptide fragments of S-antigenand mediate experimental autoimmune uveoretinitisand pinealitis. / Immunol. 1986; 136:2875-2882.
7. Rizzo LV, Silver PB, Hakim F, Chan C, Wiggert B,Caspi RR. Establishment and characterization of anIRBP-specific T cell line that induces EAU in B10.Amice. Invest Ophthalmol Vis Sci. ARVO Abstracts.1993;34:1143.
8. Caspi RR, Roberge FG, Chan CC, et al. Experimentalautoimmune uveoretinitis induced in mice with twodifferent retinal antigens, f Immunol. 1988; 140:1490-1495.
9. Pepperberg DR, Okajima TL, Wiggert B, Ripps H,Crouch RK, Chader GJ. Molecular biology and physio-logical role in the visual cycle of rhodopsin. MolNeuro-biol. 1993; 7:61-85.
10. Borst DE, Redmond TM, Elser JE, et al. lnterphotore-ceptor retinoid-binding protein: Gene characteriza-tion, protein repeat structure, and its evolution. JBiolChem. 1989; 264:1115-1123.
11. Caspi R. Immunogenedc aspects of clinical and exper-imental uveitis. Reg Immunol. 1993;4:321-330.
12. Caspi RR, Grubbs BG, Chan CC, Chader GJ, WiggertB. Genetic control of susceptibility to experimentalautoimmune uveoretinitis in the mouse model: Con-comitant regulation by MHC and non-MHC genes. /Immunol. 1992; 148:2384-2389.
13. Caspi RR, Silver PB, Chan CC, Wiggert B, RedmondTM, Donoso L. Immunogenetics of experimental au-toimmune uveoretinitis (EAU). Reg Immunol. 1994(suppl); 6:20-23.
14. Donoso LA, Merryman CF, Sery T, Sanders R, VrabecT, Fong S-L. Human interstitial retinoid binding pro-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933185/ on 04/12/2018

954 Investigative Ophthalmology & Visual Science, April 1995, Vol. 36, No. 5
tein: A potent uveitopathogenic agent for the induc-tion of experimental autoimmune uveitis. J Immunol.1989; 143:79-83.
15. Donoso LA, Merryman CF, Sery TW, et al. S-antigen:Characterization of a pathogenic epitope which medi-ates experimental autoimmune uveitis and pinealitisin Lewis rats. CurrEye Res. 1987;6:1151-1159.
16. Pepperberg DR, Okajima TL, Ripps H, Chader GJ,Wiggert B. Functional properties of interphotorecep-tor retinoid-binding protein. Photochem Photbiol1991;54:1057-1060.
17. Silver PB, Rizzo LV, Chan CC, Donoso LA, WiggertB, Caspi RR. Identification of a putative epitope inthe IRBP molecule that is uveitogenic for mice of theH-2b haplotype. Invest Ophlhalmol Vis Sti. ARVO Ab-stracts. 1993; 34:1482.
18. Hu L-H, Wiggert B, Caspi R, et al. Different epitopesof IRBP are immunodominant and immunopatho-
genic in different strains of rats and mice. ARVO Ab-stracts. Invest Ophthalmol Vis Sti. 1989; 30:82.
19. Hirose S, Singh VK, Donoso LA, et al. An 18-merpeptide derived from the retinal S antigen inducesuveitis and pinealitis in primates. Clin Exp Immunol.1989;77:106-lll.
20. Hirose S, Ogasawara K, Natori T, et al. Regulation ofexperimental autoimmune uveitis in rats—separationof MHC and non-MHC gene effects. Clin Exp Immunol.1991;86:419-425.
21. Fling S, Donoso L, Gregerson DS. In vitro unrespon-siveness to autologous sequences of the immunopa-thogenic autoantigen, S-antigen. J Immunol. 1991;147:483-489.
22. Pettinelli CB, Fritz RB, Chou C-HJ, McFarlin DE. En-cephalitogenic activity of guinea pig myelin basic pro-tein in the SJL mouse. J Immunol. 1982; 129:1209-1211.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933185/ on 04/12/2018