identification of a divalent metal cation binding site in herpes
TRANSCRIPT

Identification of a Divalent Metal Cation Binding Site in HerpesSimplex Virus 1 (HSV-1) ICP8 Required for HSV Replication
Kevin F. Bryant,a* Zhipeng Yan,a David H. Dreyfus,b* and David M. Knipea
Department of Microbiology and Immunobiology, Harvard Medical School, Boston, Massachusetts, USA,a and Clinical Faculty, Yale School of Medicine, New Haven,Connecticut, USAb
Herpes simplex virus 1 (HSV-1) ICP8 is a single-stranded DNA-binding protein that is necessary for viral DNA replication andexhibits recombinase activity in vitro. Alignment of the HSV-1 ICP8 amino acid sequence with ICP8 homologs from other her-pesviruses revealed conserved aspartic acid (D) and glutamic acid (E) residues. Amino acid residue D1087 was conserved in everyICP8 homolog analyzed, indicating that it is likely critical for ICP8 function. We took a genetic approach to investigate the func-tions of the conserved ICP8 D and E residues in HSV-1 replication. The E1086A D1087A mutant form of ICP8 failed to supportthe replication of an ICP8 mutant virus in a complementation assay. E1086A D1087A mutant ICP8 bound DNA, albeit with re-duced affinity, demonstrating that the protein is not globally misfolded. This mutant form of ICP8 was also recognized by a con-formation-specific antibody, further indicating that its overall structure was intact. A recombinant virus expressing E1086AD1087A mutant ICP8 was defective in viral replication, viral DNA synthesis, and late gene expression in Vero cells. A class ofenzymes called DDE recombinases utilize conserved D and E residues to coordinate divalent metal cations in their active sites.We investigated whether the conserved D and E residues in ICP8 were also required for binding metal cations and found that theE1086A D1087A mutant form of ICP8 exhibited altered divalent metal binding in an in vitro iron-induced cleavage assay. Theseresults identify a novel divalent metal cation-binding site in ICP8 that is required for ICP8 functions during viral replication.
Herpes simplex virus 1 (HSV-1) is a double-stranded DNAvirus that replicates its genome in the nuclei of infected cells.
HSV-1 encodes seven gene products that are directly involved inthe replication of viral DNA, all of which are essential for viralDNA replication (44). These proteins are the DNA polymerase(which consists of UL30, the catalytic subunit, and UL42, its pro-cessivity factor), an origin binding protein (UL9), a helicase-pri-mase complex (which consists of the UL5, UL52, and UL8 pro-teins), and the single-stranded DNA binding protein ICP8 (alsoknown as UL29), which is the focus of this report.
ICP8 is an early gene product that nonspecifically binds single-stranded DNA in a cooperative manner (22, 38). In addition to itsessential role in viral DNA replication, ICP8 has also been shownboth to repress transcription from the parental genome (13–15)and to stimulate late gene transcription (12). ICP8 is known tobind zinc (17), and several biochemical activities of ICP8 requiremagnesium cations (9, 24, 34), suggesting that the protein alsobinds magnesium. Zinc binding by ICP8 is thought to be involvedin its DNA binding activity, because altering residues in the ICP8zinc finger region results in an abolishment of DNA binding (11).The role of magnesium binding by ICP8 is much less well under-stood, and no mutations have been made that specifically disruptICP8 magnesium binding in order to directly assess its importanceto ICP8 function.
ICP8 has also been reported to mediate several activities in-volved in DNA recombination in vitro, including destabilizationof DNA duplexes (1), facilitation of the annealing of single-stranded DNA (9), mediation of strand exchange (2, 29, 34), andmediation of strand invasion (28). ICP8 interacts with the HSV-encoded alkaline nuclease UL12, and UL12 is proposed to play arole in the initiation and/or the resolution steps of the DNA re-combination mechanism (34, 36). The interaction of ICP8 withUL12 also stimulates the processivity of UL12-mediated digestionof DNA (35). Other nucleases, such as Escherichia coli exonuclease
III and lambda phage Red � exonuclease, can also stimulate thestrand exchange activity of ICP8 in vitro (36). The HSV-1 helicase-primase complex has also been reported to cooperate with ICP8 topromote strand exchange activity in vitro (29). Furthermore, ICP8is required for long-chain DNA synthesis in an in vitro recombi-nation-dependent replication assay (30, 31), although it is notclear that ICP8 recombinase activity is required in this assay. ICP8is a major component of HSV-1 replication compartments, whichare nuclear domains where viral DNA replication and late geneexpression occur. ICP8 also interacts with several cellular proteinsknown to be involved in recombination, including DNA-PKcs(DNA-dependent protein kinase, catalytic subunit), Rad50, andKu86, and recruits these proteins to viral replication compart-ments (40), where they may play important roles in mediating therecombination of the HSV-1 genome.
In this study, we report the identification of a conserved aspar-tic acid residue in ICP8 that is required for divalent metal cationbinding and viral DNA replication. ICP8 is thought to bind Mg2�,and Mg2� is required for ICP8 activity in some in vitro recombi-nase assays. Therefore, the conserved D residue in ICP8 may co-
Received 13 February 2012 Accepted 26 March 2012
Published ahead of print 4 April 2012
Address correspondence to David M. Knipe, [email protected].
* Present address: Kevin F. Bryand, Zymo Research Corporation, Irvine, California,USA; David H. Dreyfus, 488 Norton Parkway, New Haven, Connecticut, USA.
We dedicate this article to the memory of Bill Ruyechan, a friend and colleaguewhose work and advice on ICP8 continues to inspire us.
Supplemental material for this article may be found at http://jvi.asm.org/.
Copyright © 2012, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.00374-12
June 2012 Volume 86 Number 12 Journal of Virology p. 6825–6834 jvi.asm.org 6825
on February 13, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

ordinate Mg2�, and bound Mg2� at this site may be required forICP8 function during HSV-1 infection.
MATERIALS AND METHODSCells and viruses. Vero cells were obtained from the American Type Cul-ture Collection (Manassas, VA). The ICP8-complementing cell lines V529(5) and S2 (11) were generated as described previously. Cells were main-tained in Dulbecco’s modified Eagle’s medium (DMEM) supplementedwith 5% heat-inactivated fetal bovine serum (FBS) and 5% heat-inacti-vated newborn calf serum (NCS). The growth medium for the V529 andS2 cells was also supplemented with 500 �g/ml G418.
All experiments were performed with HSV-1 strain KOS as the wild-type (WT) virus (39) or with mutant virus 8lacZ (21), pm1.a (11), orKOS.8DDEm (described below). Viruses were propagated and titrated onVero or V529 cells by standard procedures.
Plasmids. This section describes the construction of plasmids used inthe transient complementation assays. Plasmids used in other assays aredescribed in the relevant sections. Plasmid pFastBac HTa-ICP8 was con-structed by ligating the AvrII/EcoRI fragment containing ICP8 frompSV8.3 (12) into pFastBac HTa (Invitrogen) DNA digested with XbaI andEcoRI (all restriction enzymes were from New England Biolabs [NEB]).The pFastBac HTa-d105 plasmid was constructed in the same manner,except that the AvrII/EcoRI ICP8 fragment originated from pSVd105(12). The pFastBac HTa-UL29 and pFastBac HTa-UL29.d105 plasmidswere constructed by introducing an EcoRI site immediately upstream ofthe ICP8 initiation codon in pFastBac HTa-ICP8 and pFastBac HTa-d105, respectively, by site-directed mutagenesis, digestion of these plas-mids with EcoRI, and self-ligation to remove the ICP8 5= untranslatedregion (5= UTR) and vector sequences between the EcoRI site in pFastBacHTa and the ICP8-initiating ATG (newly introduced EcoRI site).pCI�.UL29 was constructed by ligating the ICP8-containing DNA frag-ment from pFastBac HTa-UL29 that was digested with HindIII, followedby blunting of the sticky ends with the Klenow fragment of E. coli DNApolymerase I (NEB) and digestion with EcoRI, into pCI� (27) that hadbeen digested with SmaI and EcoRI. pCI�.UL29.d105 was constructed inthe same manner, except that the HindIII/Klenow/EcoRI fragment con-taining the ICP8 gene originated from pFastBac HTa-d105. pCI�B wasgenerated by digesting pCI� with BamHI, blunting the sticky ends withKlenow fragment, and self-ligating (thus destroying the BamHI restric-tion site). pCI�B.UL29 was constructed by ligating the ICP8-containingDNA fragment from pFastBac HTa-UL29 that was digested with HindIII,followed by blunting of the sticky ends with Klenow fragment and diges-tion with EcoRI, into pCI�B that had been digested with SmaI and EcoRI.Several smaller regions of the ICP8 gene were subcloned into pBluescriptII SK(�) (pBS; Stratagene) to be used as the template for introducing D orE codon mutations into the ICP8 gene by performing PCR-based site-directed mutagenesis. pBS-ICP8-BS contains the BamHI/SalI fragment ofthe ICP8 gene cloned into the BamHI and SalI sites in pBS. pBS-ICP8-Ncontains the NotI fragment of the ICP8 gene cloned into the NotI site inpBS. The following oligonucleotides, and their complements, wereused to generate PCR-based site-specific mutations in the ICP8 gene(mutated nucleotides are in boldface): for D545A, 5=-ATGAACAGCATGTACAGCGCCTGCGACGTGCTGGGAAAC-3=; for D547A, 5=-AGCATGTACAGCGACTGCGCCGTGCTGGGAAACTACGCC-3=; forD625A, 5=-AACGTCAGGCAGGTCGTGGCCCGCGAGGTGGAGCAGCTG-3=; for E627A, 5=-AGGCAGGTCGTGGACCGCGCGGTGGAGCAGCTGATGCGC-3=; for D645A, 5=-GAGGGGAGGAACTTCAAGTTTCGCGCCGGTCTGGGCGAGGCCAACCACGCC-3=; for E735A, 5=-AAAACGCTGACGGTCGCGCTCTCGGCGGGGGCGGCTATCTGCGCCCCCAGC-3=; for E860A and D861A, 5=-CCGCCCGGCTCCTGTCGCGCGCGGCCATCGAGACCATCGCGTTCAA-3=; for E1086A and D1087A,5=-GCACCCAGCAGCTGCAGATCGCGGCCTGGCTGGCGCTCCTGGAGGA-3=; for E1086A, 5=-GCACCCAGCAGCTGCAGATCGCGGACTGGCTGGCGCTCCTGGAGGA-3=; for D1087A, 5=-GCACCCAGCAGCTGCAGATCGAGGCCTGGCTGGCGCTCCTGGAGGA-3=; and for
D1087C, 5=-GCACCCAGCTGCAGATCGAGTGCTGGCTGGCGCTCCTGGAGGA-3=.
The D545A, D547A, D625A, and E627A mutations were generated inpBS-ICP8-N, and the AfeI/AgeI DNA fragment from each resulting plas-mid, which contains the mutation, was ligated into AfeI- and AgeI-di-gested pCI�.UL29 to generate pCI�.UL29.D545A, pCI�.UL29.D547A,pCI�.UL29.D625A, and pCI�.UL29.E627A, respectively. The D645A mu-tation was generated in pBS-ICP8-BS, and the BamHI/SalI fragment fromthis plasmid was ligated into BamHI- and SalI-digested pCI�B.UL29 togenerate pCI�B.UL29.D645A. The E735A, E860A D861A, E1086AD1087A, E1086A, D1087A, and D1087C mutations were generated inpBS-ICP8-BS, and the AgeI/SalI DNA fragment containing the muta-tions was ligated into AgeI- and SalI-digested pCI�.UL29 to generatepCI�.UL29.E735A, pCI�.UL29.E860A/D861A, pCI�.UL29.E1086A/D1087A,pCI�.UL29.E1086A, pCI�.UL29.D1087A, and pCI�.UL29.D1087C, respec-tively. All mutated regions in the ICP8 gene were confirmed by DNA sequencing.
Transient complementation assay. The transient complementationassay was performed as a variation of our previous studies (33). HeLa cells,which do not complement the replication of the 8lacZ ICP8 mutant virus,were transfected with the indicated plasmids using Effectene transfectionreagents (Qiagen) according to the manufacturer’s instructions. At 24 hposttransfection, the transfected cells were infected with 8lacZ virus at amultiplicity of infection (MOI) of 10 PFU per cell. At 24 h postinfection(hpi), samples for determination of viral yield were harvested by scrapingthe infected-cell monolayer and collecting both the cells and the superna-tant. Samples were frozen at �80°C and were thawed, and cell-free super-natant was collected following centrifugation of the samples. The viralyield in each sample was determined by performing plaque assays on V529cells, which express ICP8 and thus complement the replication of theICP8 mutant 8lacZ. Complementation was expressed as the percentage ofviral yield relative to the viral yield observed in samples transfected withthe plasmid expressing wild-type ICP8, which was set at 100% comple-mentation.
Immunoblotting. Vero cells were infected with the indicated virus atan MOI of 10, or HeLa cells were transfected as described above. Cellmonolayers were washed with phosphate-buffered saline supplementedwith calcium and magnesium (PBS-ABC), and lysates were prepared byscraping the cells in 2� sodium dodecyl sulfate-polyacrylamide gel elec-trophoresis (SDS-PAGE) loading buffer and boiling for 5 min. Polypep-tides were resolved by SDS-PAGE and were transferred to a polyvi-nylidene difluoride (PVDF) membrane. Membranes were blocked for 1 hat room temperature with 5% milk in Tris-buffered saline with 0.1%Tween 20 (TBST). Blocked membranes were reacted with primary anti-bodies diluted in 5% milk in TBST. The primary antibodies used wereanti-ICP27 (�-ICP27; Abcam), �-ICP8 (antibody 3-83 [20]), �-gC (a giftfrom Gary Cohen), and anti-glyceraldehyde-3-phosphate dehydrogenase(�-GAPDH; from either Abcam or Applied Biological Materials Inc.).Horseradish peroxidase-conjugated secondary antibodies were fromSanta Cruz Biotechnology.
Generation of recombinant baculoviruses. To generate recombi-nant baculoviruses that express ICP8, we utilized the pFastBac.Dual-based vector (Invitrogen) pFBd.GFP, which expresses green fluores-cent protein (GFP) under the control of the p10 promoter. Weobtained pFBd.GFP.S2 (unpublished plasmid), which expresses thereovirus S2 protein, as a gift from Kenneth Murray (Florida Interna-tional University). pFastBac HTa-UL29 and pFastBac HTa-UL29.E1086A/D1087A were digested with EcoRI and HindIII, and theDNA fragment containing ICP8 was ligated into pFBd.GFP.S2 that hadbeen digested with EcoRI and HindIII, which removed the S2 codingsequence. The resulting plasmids were named pFBd.GFP.UL29 andpFBd.GFP.UL29.E1086A/D1087A. ICP8 was epitope tagged at its Nterminus with the 6� His epitope by ligating oligonucleotidesencoding six histidine residues into the EcoRI site upstream from theICP8-initiating ATG codon, generating pFBd.GFP.UL29.His andpFBd.GFP.UL29.E1086A/D1087A.His. The presence of the 6� His se-
Bryant et al.
6826 jvi.asm.org Journal of Virology
on February 13, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

quence in the correct reading frame was confirmed by DNA sequenc-ing. Recombinant baculovirus bacmids were generated by transform-ing chemically competent E. coli DH10Bac cells (Invitrogen) witheither pFBd.GFP.UL29.His or pFBd.GFP.UL29.E1086A/D1087A.Hisaccording to the manufacturer’s instructions.
Sf21 insect cells were transfected with recombinant bacmids that ex-press either wild-type ICP8 or the DDE mutant by using the Cellfectintransfection reagent (Invitrogen) according to the manufacturer’s in-structions. The supernatant from transfected cells was collected and wasdesignated the P1 stock. This was used to infect new Sf21 cells and togenerate the P2 stock. The P2 stock was used to infect Sf21 cells for theexpression and purification of recombinant ICP8 proteins.
Protein purification. Baculovirus-infected Sf21 cells were harvestedfor protein purification by washing once with PBS, and then cells werecollected by scraping from the flask into PBS. Cells were lysed by incubat-ing with a buffer consisting of 20 mM Tris (pH 7.5), 300 mM NaCl, 20%glycerol, and 0.5% NP-40 for 30 min on ice. The cell-free supernatantcontaining the soluble recombinant protein was collected following cen-trifugation of the samples, applied to a column containing Talon affinityresin (Clontech) that was equilibrated with lysis buffer, and allowed toproceed through the column by gravity flow. Bound protein was washedwith a buffer consisting of 20 mM Tris HCl (pH 7.5), 300 mM NaCl, 5%glycerol, and 0.1% NP-40 and was then eluted with a buffer consisting of50 mM Tris HCl (pH 7.5), 300 mM NaCl, 15% glycerol, and 150 mMimidazole. Ten fractions of the eluate were collected. The fractions con-taining the purified protein were identified by resolving a portion by SDS-PAGE and staining the gel with Coomassie stain, and those fractions werecombined. The pooled fractions were concentrated, and the buffer wasexchanged to ICP8 storage buffer (50 mM Tris HCl [pH 7.5], 150 mMNaCl, 1 mM EDTA, 1 mM dithiothreitol [DTT], 20% glycerol) usingVivaspin sample purification columns (Vivascience). Protein concentra-tions were measured using the Bio-Rad protein assay reagent (Bio-Rad)with bovine serum albumin (BSA) dilutions to generate the standardcurve.
DNA gel shift assay. A 25-mer oligo(dT) oligonucleotide [oligo(dT)25;synthesized by Integrated DNA Technologies] was phosphorylated at its 5=end with T4 polynucleotide kinase (NEB) using [�-32P]ATP (6,000 Ci/mmol,10 mCi/ml; Perkin-Elmer), and unincorporated radioactive nucleotides wereremoved using an illustra MicroSpin G-50 column (GE Healthcare). Approx-imately 2 pmol of oligo(dT)25 (based on the assumption of 100% recoveryfrom the MicroSpin column) was incubated on ice with the indicated con-centration of either wild-type ICP8 or the DDE mutant form of ICP8 in ICP8storage buffer for 30 min. The samples were resolved on a 5% native poly-acrylamide gel, and the gel was dried and exposed to a phosphor storagescreen.
Immunofluorescence. Indirect immunofluorescence was performedas described previously (4). Briefly, Vero cells on glass coverslips wereeither mock infected or infected with the indicated virus at an MOI of 10PFU/cell for 10 h prior to fixation with 3.65% formaldehyde. The anti-bodies used were 3-83, to detect total ICP8, and the conformation-specificantibody 39S, which detects correctly folded ICP8. The fixed and stainedcells were imaged using a Zeiss Axioplan 2 microscope, a PhotometricsCoolSNAP HQ2 charge-coupled device (CCD) camera, and Zeiss Axio-Vision 4 image acquisition software.
Construction of mutant viruses. Plasmid p8-8GFP, which encodesICP8 fused to GFP at its C terminus (41), was linearized by digestion withEcoRI and was cotransfected into S2 cells with 8lacZ infectious DNA togenerate the recombinant virus KOS.8GFP. Plaques expressing GFP wereidentified by fluorescence microscopy, and these recombinant viruseswere plaque purified 5 times prior to use in experiments. To generateKOS.8DDEm, KOS.8GFP infectious DNA was cotransfected into V529cells together with EcoRI-linearized pBS.8flank.8DDEm, which containsthe E1086A D1087A mutation in ICP8. To generate pBS.8flank.8DDEm,we constructed pBS.8flank by ligating a blunted BglII/NdeI fragmentfrom pSG18 (16), which contains the ICP8 locus and the surrounding
region, into EcoRV-digested pBS. pBS.8flank.8DDEm was then generatedby ligating a Bpu10I fragment containing the E1086A D1087A mutationfrom pCI�.UL29.E1086A/D1087A into Bpu10I-digested pBS-8flank.Plaques that did not express GFP were identified by fluorescence micros-copy and were purified 3 times prior to use in experiments. The presenceof the D1086A E1087A mutations in the ICP8 gene was confirmed bysequencing a PCR product from the appropriate region of ICP8. A recom-binant virus that rescued the ICP8 DDE mutation, KOS.8DDEm-R, wasalso generated in a similar manner. In this case, infectious KOS.8DDEmDNA was cotransfected into V529 cells together with EcoRI-linearizedpBS.8flank. Recombinant viruses were selected by their ability to formplaques on noncomplementing Vero cells and were plaque purified 4times prior to use in experiments. The presence of wild-type sequence wasconfirmed by sequencing a PCR product from the appropriate region ofICP8.
Viral yield assay. Vero or V529 cells were infected with the indicatedvirus at an MOI of 10 in PBS-ABC containing 1% bovine serum and 0.1%glucose for 1 h in a shaking incubator at 37°C. Following the 1-h adsorp-tion step, cells were washed twice with acid wash buffer (40 mM citric acid[pH 3], 10 mM KCl, 135 mM NaCl) and once with DMEM containing 1%FBS, and then DMEM containing 1% FBS was added. Progeny virus washarvested at the indicated times postinfection by scraping the infected-cellmonolayer and collecting the cells and supernatant. Samples were frozenat �80°C following harvesting. Viral yield was determined by performingplaque assays on V529 cells.
Viral DNA replication assay. Vero cells were infected with the indi-cated virus as described above for the viral yield assay. Following infectionfor the indicated time, samples were harvested by washing the cell mono-layers with PBS-ABC, and the cells were scraped in PBS-ABC and werethen collected by centrifugation. Total DNA (including both cellular andviral DNA) was purified by using the Generation Capture Column kit(Qiagen, formerly Gentra) according to the manufacturer’s instructions.Viral DNA was quantified by performing real-time PCR using primersspecific for the ICP27 promoter (3). Real-time PCR was performed usingPowerSYBR green reagents (Applied Biosystems) and an Applied Biosys-tems 7300 sequence detection system according to the manufacturer’sinstructions. The viral DNA levels were normalized to the levels of aGAPDH pseudogene in each sample (20).
Iron-induced protein cleavage assay. The iron-induced proteincleavage assay was performed essentially as described previously (19).Briefly, 20 pmol of either wild-type ICP8 or the DDE mutant form of ICP8was either mock treated or incubated with ammonium iron(II) sulfatehexahydrate at a final concentration of 20 �M and ascorbic acid at a finalconcentration of 20 mM, either in the presence or in the absence of 20pmol of the oligo(dT)25 oligonucleotide, in ICP8 storage buffer for 30 minat room temperature. Hydrogen peroxide was added to the reaction mix-tures treated with ammonium iron(II) sulfate and ascorbic acid, at a finalconcentration of 0.2%, for 10 s at room temperature. The reactions werestopped by boiling the samples in SDS-PAGE loading buffer, and thepolypeptides were resolved by SDS-PAGE, transferred to PVDF mem-branes, and probed for ICP8 using rabbit polyclonal antibody 3-83.
RESULTSPresence of conserved aspartic acid and glutamic acid residuesin HSV-1 ICP8. To identify highly conserved regions (and there-fore potential new functional domains) in HSV-1 ICP8 and itshomologs in other herpesviruses, we performed an alignment ofamino acid sequences from nine ICP8 homologs, with represen-tatives from alpha-, beta-, and gammaherpesviruses. We discov-ered numerous aspartic acid (D) and glutamic acid (E) residuesthat were conserved in many or all of the ICP8 homologs (someconserved residues are shown in Fig. 1; a complete list is given inFig. S1 in the supplemental material), leading us to hypothesizethat these conserved residues are important for ICP8 function.
Divalent Metal Cation Binding Site of HSV-1 ICP8
June 2012 Volume 86 Number 12 jvi.asm.org 6827
on February 13, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

Several of these conserved residues were located in or near theDNA binding groove in the ICP8 crystal structure (25), suggestingthat they would be available to carry out enzymatic functions onbound DNA. Interestingly, members of a family of enzymes calledDDE recombinases, including transposases, RAG-1, and retrovi-ral integrases, also have conserved D and E residues that coordi-nate magnesium ions that are important for mediating the recom-bination reactions, leading us to speculate that ICP8 may sharebiochemical and pharmacological properties with these well-stud-ied proteins. Numerous ICP8 residues were further investigated,including D545 (amino acid residue positions are based on theKOS ICP8 sequence [10]), D547, D625, E627, D645, E735, E860,D861, E1086, and D1087.
An ICP8 molecule with an altered conserved D residue has adecreased ability to support HSV-1 replication. To determinewhether the conserved D and E residues in ICP8 were required forICP8 function during HSV-1 infection, specific D and E codonswere altered to encode alanine in a plasmid encoding HSV-1 ICP8.The ability of the encoded proteins to complement the replicationof an ICP8 mutant virus was then determined in a transient com-plementation assay. All D/E mutant forms of ICP8 were expressedto similar levels in transfected HeLa cells (Fig. 2A). At 24 h aftertransfection, the cells were infected with the ICP8-null mutantvirus 8lacZ at an MOI of 10. Samples were harvested at 24 hpostinfection (hpi), and the viral yield was determined by per-forming plaque assays on V529 cells, which contain the ICP8 geneand can complement the replication of the 8lacZ mutant virus.The viral yield observed in cells transfected with the plasmid ex-pressing wild-type ICP8 was designated 100% complementation,and complementation by the ICP8 mutants was compared to thatvalue. The d105 mutant form of ICP8, which is not able to com-plement the replication of an ICP8 mutant virus (12), did notcomplement 8lacZ replication to levels above background. TheE860A D861A mutant form of ICP8 complemented 8lacZ repli-cation to approximately 55% of the level of wild-type ICP8 (Fig.
FIG 1 Alignment of ICP8 homolog sequences. Amino acid sequences for HSV-1 ICP8 and ICP8 homologs from representative viruses in each of the three subfamiliesof herpesviruses (alphaherpesviruses, betaherpesviruses, and gammaherpesviruses) were aligned using the T-Coffee alignment algorithm (http://www.tcoffee.org [6]).The ICP8 homologs included in the analysis were from HSV-1 strain KOS (NCBI accession number P17470), HSV-2 (NP_044499), varicella-zoster virus (AEW89446),Marek’s disease virus (Q9E6P0), Epstein-Barr virus (P03227), human cytomegalovirus (P17147), murine cytomegalovirus (MCMV) (P30672), human herpesvirus 7(O56282), and Kaposi’s sarcoma-associated herpesvirus (ADQ57880). Sites with similar amino acids in 4 or more ICP8 homologs are in black letters highlighted in lightgray; sites with identical amino acids in 4 or more ICP8 homologs are in white letters highlighted in black; and sites with identical amino acids in all 9 ICP8 homologs arein white letters highlighted in dark gray. Two regions are shown, identifying conserved amino acids at positions 545 and 547 (based on their positions in HSV-1 ICP8)and the complete conservation of an aspartic acid residue at position 1087. A full alignment can be found in Fig. S1 in the supplemental material.
FIG 2 Transient complementation assay. HeLa cells were either mock trans-fected, transfected with an empty vector (pCI�), or transfected with plasmidsexpressing wild-type ICP8, the ICP8 d105 deletion mutant, or ICP8 with thecodons that encode the indicated amino acids mutated to encode alanine. At 24 hposttransfection, the cells were either harvested to prepare samples for immuno-blotting (A) or infected with the ICP8 mutant 8lacZ (B). Viral yield samples wereharvested at 24 h postinfection, and viral yield was determined by a plaque assay onICP8-complementing V529 cells. The reported values are percentages of the com-plementation by cells transfected with the plasmid encoding wild-type ICP8 andare averages for 4 independent experiments. Error bars represent standard devia-tions. Similar results were seen in four additional independent experiments inVero cells.
Bryant et al.
6828 jvi.asm.org Journal of Virology
on February 13, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

2B), indicating that residues 860 and 861 are important, but notessential, for wild-type activity of ICP8. The D645A mutant formof ICP8 also complemented 8lacZ replication to approximately50% of the level of wild-type ICP8 (Fig. 2B), indicating that resi-due 645 is also important, but not essential, for wild-type activityof ICP8. The E1086A D1087A mutant form of ICP8 did not com-plement the replication of 8lacZ to levels above the backgroundobserved when cells were transfected with the empty-vector plas-mid (Fig. 2B). These results indicated that either residue 1086 or1087, or both, is absolutely critical for ICP8 to function duringHSV-1 replication in this assay.
To determine whether both residues 1086 and 1087 are re-quired for ICP8 activity, we changed them each individually toalanine residues and assayed their abilities to complement 8lacZreplication. The E1086A mutant form of ICP8 complemented8lacZ replication to an extent similar to that of wild-type ICP8(Fig. 2B), indicating that this residue is not critical for ICP8 activ-ity. However, the D1087A mutant form of ICP8 did not comple-ment 8lacZ to levels above background (Fig. 2B), indicating thatresidue 1087 is absolutely essential for ICP8 to complement thereplication of an ICP8 mutant virus. Furthermore, the inability ofthe E1086A D1087A double point mutant to complement 8lacZreplication can be attributed entirely to the D1087A mutation.Cysteine residues are known to be able to coordinate divalentmetal cations in DDE recombinases; therefore, to investigatewhether divalent metal cation binding played a role in the D1087Aphenotype, we tested a mutant form of ICP8 in which residueD1087 was changed to cysteine. The D1087C mutant form of ICP8was able to complement 8lacZ to much higher levels than theD1087A mutant (Fig. 2B), suggesting that D1087 coordinates di-valent metal cations and that this activity is required for ICP8function. Other amino acid residue mutants (the D545A, D547A,D625A, E627A, E629A, and E735A mutants) complemented 8lacZreplication to near-wild-type levels when expressed in HeLa cells(Fig. 2B), indicating that these residues are not absolutely requiredfor ICP8 function in viral replication.
ICP8 molecules with altered conserved D/E residues are notglobally misfolded. To rule out that the possibility that theE1086A D1087A mutation in ICP8 reduced the activity of ICP8 bysimply destroying the overall folding of the protein, we investi-gated the ability of purified recombinant wild-type and E1086AD1087A (DDE mutant) ICP8 to bind DNA in vitro by performingelectrophoretic mobility shift assays. Both wild-type and DDEmutant forms of ICP8 were purified to near-homogeneity (Fig.3A). Increasing concentrations of both proteins in the bindingreactions resulted in a slower-migrating form of the labeled oli-go(dT)25 DNA (Fig. 3B), indicating that the DNA was bound byboth forms of purified ICP8 and therefore that the DDE mutantform of ICP8 is not globally misfolded. However, higher concen-trations of the DDE mutant form of ICP8 than of wild-type ICP8were required to bind the oligonucleotide, arguing that the mu-tant form bound DNA with decreased affinity.
To assess the folding of the mutant form of ICP8 by anothermethod, we investigated whether mutant ICP8 expressed duringinfection with the 8DDEm mutant virus was recognized by theconformation-specific 39S antibody (42). Mock-infected cellswere not stained with either the 39S antibody or the 3-83 �-ICP8antibody, which detects total ICP8, demonstrating the specificityof these antibodies (Fig. 4, top row). Cells infected with wild-typeHSV-1 were stained with both antibodies, and mature replication
compartments were observed (Fig. 4, second row). Cells infectedwith the ICP8 mutant virus pm1.a, which expresses an alteredform of ICP8 that is defective for DNA binding and is incorrectlyfolded (11), were not stained by antibody 39S but were stained byantibody 3-83 (Fig. 4, third row). 8DDEm-infected cells werestained by both 39S and 3-83 antibodies (Fig. 4, bottom row),further demonstrating that this mutant form of ICP8 maintainedthe correct conformation. In addition, we observed that the mu-tant form of ICP8 expressed during 8DDEm infection did notform mature replication compartments, indicating that the al-tered residues were required for this activity.
The conserved D/E residues in ICP8 are required for efficientHSV-1 replication and viral DNA replication. We constructed a
FIG 3 Effect of ICP8 DDE mutation on DNA binding. (A) His-tagged wild-type and DDE mutant ICP8 proteins were expressed from recombinant bacu-loviruses and were purified from infected Sf21 cells. Purified proteins wereresolved by SDS-PAGE, and the gel was stained with Coomassie blue stain. (B)The indicated concentration of each protein was incubated with radiolabeledoligo(dT)25 DNA, and protein-DNA complexes were resolved on a 5% nativepolyacrylamide gel.
Divalent Metal Cation Binding Site of HSV-1 ICP8
June 2012 Volume 86 Number 12 jvi.asm.org 6829
on February 13, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

mutant virus, 8DDEm, containing the E1086A D1087A mutationsin the ICP8 gene, to investigate whether this mutant form of ICP8affected HSV-1 replication when expressed from the viral ge-nome. We observed that replication of this mutant virus was de-creased 10- to 100-fold from that with wild-type HSV-1 strainKOS in noncomplementing Vero cells (Fig. 5A). However,8DDEm replicated better than the ICP8 mutant virus pm1.a (Fig.5A) (11) in Vero cells, suggesting that the replication functions ofthe altered ICP8 in the 8DDEm mutant virus were not completelydestroyed. A virus in which the E1086A D1087A mutation wasrestored to the wild-type sequence, 8DDEm-R, replicated simi-larly to the wild-type virus, indicating that the decrease in replica-tion observed for 8DDEm was likely due to the specific mutation.All viruses replicated to nearly wild type levels in V529 cells, whichexpress wild-type ICP8 and therefore complement ICP8 mutantviruses, suggesting that the replication defects of these viruses inVero cells were due to the changes in ICP8.
We also determined the plating efficiencies for the wild-typevirus, the 8DDEm mutant, and the 8DDEm-R rescued virus byplating serial dilutions of each virus on either noncomplementingVero cells or the complementing cell line V529. The wild-typevirus and the rescued virus 8DDEm-R had similar titers on bothcell lines, but the 8DDEm mutant virus had an approximately70-fold-lower titer on Vero cells than on V529 cells (Fig. 5B).These results are consistent with a replication defect of the8DDEm mutant virus.
We next investigated the levels of viral DNA replication in Verocells infected with either the wild-type virus, the 8DDEm mutantvirus, the 8DDEm-R rescued virus, or the pm1.a ICP8 mutantvirus, which is defective for DNA binding and viral DNA replica-tion (11). For the wild-type virus, we observed that levels of viralDNA increased �1,000-fold during the course of infection (Fig.6). We observed only a very small increase in viral DNA levelsbetween 4 and 12 h postinfection (�2-fold), and an approxi-mately 10-fold increase in viral DNA levels by 24 h postinfection,with the 8DDEm mutant virus (Fig. 6), indicating that DNA syn-thesis during 8DDEm infection was severely decreased relative tothat of the wild-type virus and strongly suggesting that the E1086and D1087 residues in ICP8 are essential for efficient HSV-1 DNAreplication. Viral DNA synthesis by the 8DDEm-R rescued viruswas similar to that by the wild type (Fig. 6). Viral DNA levels didnot increase at all during the course of infection with pm1.a (Fig.6). Because viral DNA levels in cells infected with 8DDEm weregreater than those in pm1.a-infected cells, it was clear that themutant ICP8 expressed by 8DDEm supported low levels of DNAreplication.
Effect of altered ICP8 D/E residues on viral gene expression.Because the 8DDEm mutant virus exhibited defects in viral repli-cation and DNA synthesis, we next investigated whether this mu-tation also had an effect on viral gene expression. We assayed theaccumulation of the immediate-early ICP27 gene product, theearly ICP8 gene product, and the late glycoprotein C (gC) gene
FIG 4 Effect of the ICP8 DDE mutation on recognition by the conformation-specific antibody 39S. Vero cells were either mock infected or infected with theindicated virus at an MOI of 10 PFU/cell. At 10 hpi, the cells were fixed and stained with either the �-ICP8 39S conformation-specific antibody (green; left) orthe 3-83 antibody, which detects total ICP8 (red; center).
Bryant et al.
6830 jvi.asm.org Journal of Virology
on February 13, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
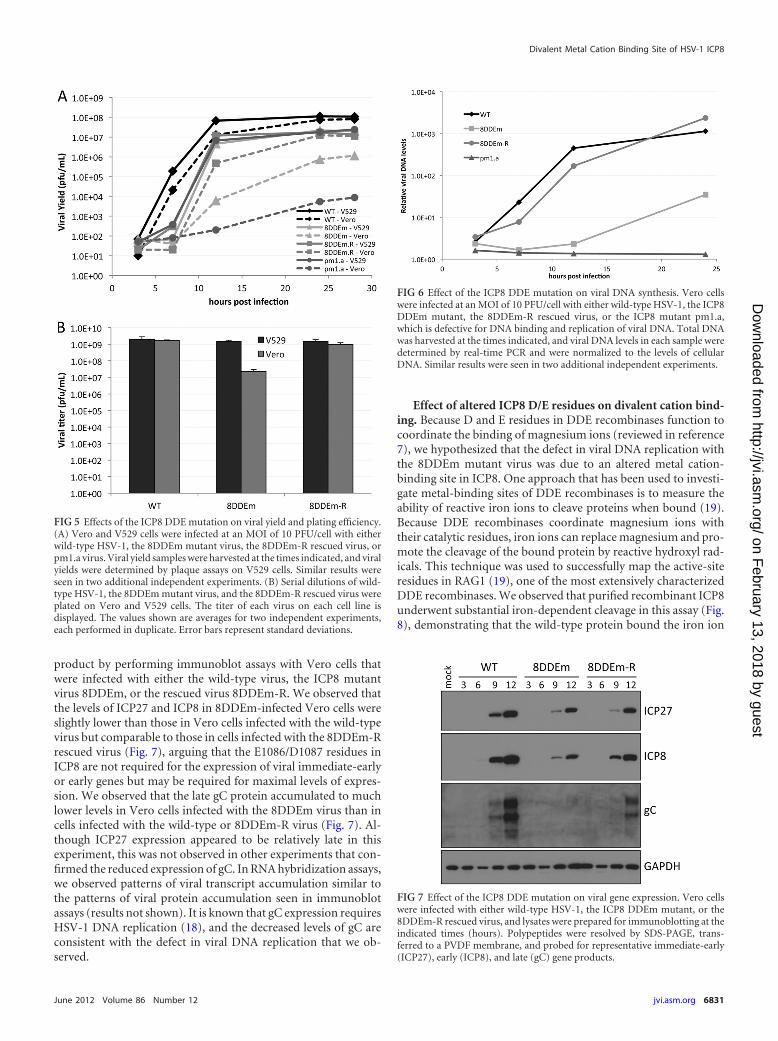
product by performing immunoblot assays with Vero cells thatwere infected with either the wild-type virus, the ICP8 mutantvirus 8DDEm, or the rescued virus 8DDEm-R. We observed thatthe levels of ICP27 and ICP8 in 8DDEm-infected Vero cells wereslightly lower than those in Vero cells infected with the wild-typevirus but comparable to those in cells infected with the 8DDEm-Rrescued virus (Fig. 7), arguing that the E1086/D1087 residues inICP8 are not required for the expression of viral immediate-earlyor early genes but may be required for maximal levels of expres-sion. We observed that the late gC protein accumulated to muchlower levels in Vero cells infected with the 8DDEm virus than incells infected with the wild-type or 8DDEm-R virus (Fig. 7). Al-though ICP27 expression appeared to be relatively late in thisexperiment, this was not observed in other experiments that con-firmed the reduced expression of gC. In RNA hybridization assays,we observed patterns of viral transcript accumulation similar tothe patterns of viral protein accumulation seen in immunoblotassays (results not shown). It is known that gC expression requiresHSV-1 DNA replication (18), and the decreased levels of gC areconsistent with the defect in viral DNA replication that we ob-served.
Effect of altered ICP8 D/E residues on divalent cation bind-ing. Because D and E residues in DDE recombinases function tocoordinate the binding of magnesium ions (reviewed in reference7), we hypothesized that the defect in viral DNA replication withthe 8DDEm mutant virus was due to an altered metal cation-binding site in ICP8. One approach that has been used to investi-gate metal-binding sites of DDE recombinases is to measure theability of reactive iron ions to cleave proteins when bound (19).Because DDE recombinases coordinate magnesium ions withtheir catalytic residues, iron ions can replace magnesium and pro-mote the cleavage of the bound protein by reactive hydroxyl rad-icals. This technique was used to successfully map the active-siteresidues in RAG1 (19), one of the most extensively characterizedDDE recombinases. We observed that purified recombinant ICP8underwent substantial iron-dependent cleavage in this assay (Fig.8), demonstrating that the wild-type protein bound the iron ion
FIG 5 Effects of the ICP8 DDE mutation on viral yield and plating efficiency.(A) Vero and V529 cells were infected at an MOI of 10 PFU/cell with eitherwild-type HSV-1, the 8DDEm mutant virus, the 8DDEm-R rescued virus, orpm1.a virus. Viral yield samples were harvested at the times indicated, and viralyields were determined by plaque assays on V529 cells. Similar results wereseen in two additional independent experiments. (B) Serial dilutions of wild-type HSV-1, the 8DDEm mutant virus, and the 8DDEm-R rescued virus wereplated on Vero and V529 cells. The titer of each virus on each cell line isdisplayed. The values shown are averages for two independent experiments,each performed in duplicate. Error bars represent standard deviations.
FIG 6 Effect of the ICP8 DDE mutation on viral DNA synthesis. Vero cellswere infected at an MOI of 10 PFU/cell with either wild-type HSV-1, the ICP8DDEm mutant, the 8DDEm-R rescued virus, or the ICP8 mutant pm1.a,which is defective for DNA binding and replication of viral DNA. Total DNAwas harvested at the times indicated, and viral DNA levels in each sample weredetermined by real-time PCR and were normalized to the levels of cellularDNA. Similar results were seen in two additional independent experiments.
FIG 7 Effect of the ICP8 DDE mutation on viral gene expression. Vero cellswere infected with either wild-type HSV-1, the ICP8 DDEm mutant, or the8DDEm-R rescued virus, and lysates were prepared for immunoblotting at theindicated times (hours). Polypeptides were resolved by SDS-PAGE, trans-ferred to a PVDF membrane, and probed for representative immediate-early(ICP27), early (ICP8), and late (gC) gene products.
Divalent Metal Cation Binding Site of HSV-1 ICP8
June 2012 Volume 86 Number 12 jvi.asm.org 6831
on February 13, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

and therefore suggesting that it is also competent for binding mag-nesium ions. The amount of iron-dependent cleavage of theE1086A D1087A mutant form of ICP8 was dramatically reducedrelative to that of the wild-type protein, indicating that this mu-tant protein was defective for iron ion-induced cleavage and thuswould also be defective for magnesium binding.
DISCUSSION
In this report, we have identified a divalent metal cation-bindingsite in HSV-1 ICP8 that is essential for efficient viral replication.Identification of this site was based on strong conservation withICP8 homologs. Plasmids encoding ICP8 with E1086A D1087Aand D1087A mutational alterations failed to complement the rep-lication of an ICP8-null virus, arguing that the 1087 residue isessential for ICP8 function and viral replication. A mutant virus,8DDEm, containing the E1086A D1087A mutations in the UL29gene, which encodes ICP8, exhibits a severe defect in viral replica-tion and viral DNA synthesis. The mutant virus also shows re-duced late gene expression. The 8DDEm mutant ICP8 shows ev-idence of reduced divalent cation binding, consistent with thenotion that this is a DDE recombinase enzymatic domain. ICP8has been proposed to be a DDE recombinase (7, 8), a member of aclass of enzymes that catalyze recombination using a catalytic triadof aspartic acid and glutamic acid residues that coordinate diva-lent metal cations, usually Mg2�. The 8DDEm mutant virus couldprovide a new reagent for the functional analysis of ICP8 recom-binase activity during infection.
Nature of the functional defect in 8DDEm ICP8. ICP8 is amultifunctional protein that binds to single-stranded DNA, facil-itates the unwinding of DNA at viral origins of DNA replication,and interacts with the UL9 origin-binding protein and the viralhelicase-primase complex to promote viral DNA replication. Ad-ditionally, ICP8 exhibits recombinase activities in vitro (1, 2, 28–31, 34, 36). The 8DDEm mutant ICP8 could have defects in one ormore of these activities. The 8DDEm ICP8 is capable of binding toDNA in a gel shift assay, demonstrating that it is not completelyfunctionally destroyed. The 8DDEm ICP8 is also recognized bythe conformation-specific 39S monoclonal antibody, evidencethat its conformation and interaction with other HSV DNA rep-lication proteins are normal (42). However, our initial results in-dicate that the affinity of DNA binding by 8DDEm ICP8 is lowerthan that of WT ICP8. Thus, the reduced DNA replication couldbe due to reduced DNA binding and promotion of viral DNAreplication. Alternatively, the recombinase function could be de-
fective, and this defect could be responsible for the defect in viralreplication. Further studies of the DNA binding properties andrecombinase activities of this and other ICP8 mutant proteins willbe necessary in order to parse out the precise roles of DNA bindingand recombinase activity in viral replication.
Potential function of ICP8-mediated recombination in rep-lication. The recombinase activity of ICP8 could play any of sev-eral roles in HSV-1 biology. First, recombination could be in-volved in the circularization of the viral genome upon entry intothe nucleus. However, ICP8 is expressed as a delayed early protein,and ICP8 has not been demonstrated to be present within thevirion, so it is difficult to postulate a role for ICP8 this early in theviral replication process. Second, recombination could be in-volved in the initiation of viral DNA replication or in the transi-tion from theta-form replication to rolling-circle replication (37,43). This would be consistent with our observation of a limitedamount of viral DNA replication by the 8DDEm mutant virus.Third, recombinase activity may also be required to resolve thebranched DNA intermediates that have been observed duringHSV-1 infection, perhaps to generate the linear genomes that arepackaged into virions. In support of this, ICP8 is associated withprogeny viral DNA (23). Fourth, the recombinase activity of ICP8could play a role in the isomerization of the viral genome, whichresults in inversion of the genome at the repeated sequences. Therole of genome isomerization is not known; however, viruses inwhich the internal repeated sequences are deleted and which aretherefore noninverting retain their ability to replicate (32). Fifth,the ICP8 recombinase activity could lead to conformationalchanges in viral progeny DNA that allow efficient late gene tran-scription. cis-acting changes in viral genomes following viral DNAreplication are known to activate late gene transcription (26).ICP8 is known to bind to viral progeny DNA (23), and the recom-binase activity of ICP8 could act to resolve concatemers and in-crease viral late gene transcription. This would be consistent withthe strong defect in gC expression observed with the 8DDEm mu-tant virus. Finally, general recombination could enhance the ex-change of genetic information to allow new genotypes that couldbe more evolutionarily fit; however, it is not apparent how generalrecombination could enhance replication in cultured cells otherthan through the roles described above.
Identification of a potential new drug target. The HIV inte-grase, a DDE recombinase, has been exploited as a very effectiveantiviral target, and multiple small molecules and compoundsthat bind the DDE active site and inhibit the activity of this viralenzyme have been designed. HSV-1 ICP8 may have a DDE activesite similar to the HIV integrase active site, and one of the putativeDDE residues in ICP8 (D1087) was required for efficient HSV-1DNA replication, suggesting that this site in ICP8 may represent anovel antiviral target. We are currently working to test this hy-pothesis and to develop molecules that bind to ICP8 and inhibit itsactivity. If these drugs do target a domain including the D1087 resi-due, the conservation of this residue in human alphaherpesviruses(including HSV-1, HSV-2, and varicella-zoster virus [VZV]), humanbetaherpesviruses (including human cytomegalovirus [HCMV] andhuman herpesvirus 7 [HHV7]), and human gammaherpesviruses(including Epstein-Barr virus [EBV] and Kaposi’s sarcoma-associ-ated herpesvirus [KSHV]) (Fig. 1) indicates that this class of drugsmight be applicable to multiple human herpesviruses.
It has been known for several years that ICP8 has recombinaseactivity in vitro, and this study provides evidence that ICP8 shares
FIG 8 Effect of the DDE mutation on iron-dependent cleavage of ICP8. Pu-rified His-tagged wild-type ICP8 or ICP8 DDE mutant protein, either in thepresence or in the absence of oligo(dT)25 DNA, was either mock treated ortreated with the iron/hydroxyl radical reagents. Following the iron-dependentcleavage reaction, samples were resolved by SDS-PAGE, transferred to a PVDFmembrane, and probed for ICP8.
Bryant et al.
6832 jvi.asm.org Journal of Virology
on February 13, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

some similarities with enzymes in the DDE family of recombi-nases. Additionally, while it is well established that recombinationof the HSV-1 genome occurs at a high frequency, the viral andcellular factors involved in mediating this recombination, as wellas the role of recombination in viral replication, remain unclear.This report is the first description of an amino acid residue in ICP8that may be involved in its recombinase activity, and this studyprovides reagents with which to investigate this residue in themechanism of ICP8-mediated recombination during HSV-1 rep-lication.
We have identified amino acid residues in ICP8 that are im-portant for divalent metal cation binding, and we provide evi-dence that metal ion binding by ICP8 is required for efficient viralreplication. More work is clearly required to define the remainingresidues involved in magnesium binding and to determinewhether these residues constitute a bona fide DDE recombinaseactive site in ICP8, as well as to define the other proteins that maybe involved in ICP8-dependent recombination. Furthermore, theprecise mechanism of recombination is still unclear, as is the spe-cific role of recombination during viral DNA replication. The re-sults in this report provide experimental evidence consistent withthe hypothesis that ICP8 is a DDE recombinase.
ACKNOWLEDGMENTS
This work was funded by NIH grant AI063106 (to D.M.K.). K.F.B. wassupported by NIH institutional NRSA training grant AI007245 and NIHindividual NRSA fellowship AI081477.
We thank members of the Knipe laboratory for advice and helpfuldiscussions, Kenneth Murray for providing the pFBd.GFP.S2 plasmid,Gary Cohen for providing the �-gC antibody, and the NERCE Biomol-ecule Production Core Laboratory for providing Sf21 cells and media.
REFERENCES1. Boehmer PE, Lehman IR. 1993. Herpes simplex virus type 1 ICP8: helix-
destabilizing properties. J. Virol. 67:711–715.2. Bortner C, Hernandez TR, Lehman IR, Griffith J. 1993. Herpes simplex
virus 1 single-strand DNA-binding protein (ICP8) will promote homol-ogous pairing and strand transfer. J. Mol. Biol. 231:241–250.
3. Bryant K, Colgrove R, Knipe DM. 2011. Cellular SNF2H chromatin-remodeling factor promotes herpes simplex virus 1 immediate-early geneexpression and replication. mBio 2(1):e00330 –10. doi:10.1128/mBio.00330 –10.
4. Chang L, et al. 2011. Herpesviral replication compartments move andcoalesce at nuclear speckles to enhance export of viral late mRNA. Proc.Natl. Acad. Sci. U. S. A. 108:E136 –E144.
5. Da Costa XJ, Kramer MF, Zhu J, Brockman MA, Knipe DM. 2000.Construction, phenotypic analysis, and immunogenicity of a UL5/UL29double deletion mutant of herpes simplex virus 2. J. Virol. 74:7963–7971.
6. Di Tommaso P, et al. 2011. T-Coffee: a web server for the multiplesequence alignment of protein and RNA sequences using structural infor-mation and homology extension. Nucleic Acids Res. 39(Web Serverissue):W13–W17.
7. Dreyfus DH. 2006. The DDE recombinases: diverse roles in acquired andinnate immunity. Ann. Allergy Asthma Immunol. 97:567–578, 602.
8. Dreyfus DH. 2009. Paleo-immunology: evidence consistent with inser-tion of a primordial herpes virus-like element in the origins of acquiredimmunity. PLoS One 4:e5778. doi:10.1371/journal.pone.0005778.
9. Dutch RE, Lehman IR. 1993. Renaturation of complementary DNAstrands by herpes simplex virus type 1 ICP8. J. Virol. 67:6945– 6949.
10. Gao M, Bouchey J, Curtin K, Knipe DM. 1988. Genetic identification ofa portion of the herpes simplex virus ICP8 protein required for DNA-binding. Virology 163:319 –329.
11. Gao M, Knipe DM. 1989. Genetic evidence for multiple nuclear functionsof the herpes simplex virus ICP8 DNA-binding protein. J. Virol. 63:5258 –5267.
12. Gao M, Knipe DM. 1991. Potential role for herpes simplex virus ICP8
DNA replication protein in stimulation of late gene expression. J. Virol.65:2666 –2675.
13. Godowski PJ, Knipe DM. 1985. Identification of a herpes simplex virusfunction that represses late gene expression from parental viral genomes. J.Virol. 55:357–365.
14. Godowski PJ, Knipe DM. 1983. Mutations in the major DNA-bindingprotein gene of herpes simplex virus type 1 result in increased levels of viralgene expression. J. Virol. 47:478 – 486.
15. Godowski PJ, Knipe DM. 1986. Transcriptional control of herpesvirusgene expression: gene functions required for positive and negative regu-lation. Proc. Natl. Acad. Sci. U. S. A. 83:256 –260.
16. Goldin AL, Sandri-Goldin RM, Levine M, Glorioso JC. 1981. Cloning ofherpes simplex virus type 1 sequences representing the whole genome. J.Virol. 38:50 –58.
17. Gupte SS, Olson JW, Ruyechan WT. 1991. The major herpes simplexvirus type-1 DNA-binding protein is a zinc metalloprotein. J. Biol. Chem.266:11413–11416.
18. Jones PC, Roizman B. 1979. Regulation of herpesvirus macromolecularsynthesis. VIII. The transcription program consists of three phases duringwhich both extent of transcription and accumulation of RNA in the cyto-plasm are regulated. J. Virol. 31:299 –314.
19. Kim DR, Dai Y, Mundy CL, Yang W, Oettinger MA. 1999. Mutations ofacidic residues in RAG1 define the active site of the V(D)J recombinase.Genes Dev. 13:3070 –3080.
20. Knipe DM, Senechek D, Rice SA, Smith JL. 1987. Stages in the nuclearassociation of the herpes simplex virus transcriptional activator proteinICP4. J. Virol. 61:276 –284.
21. LaVail JH, et al. 2005. Genetic and molecular in vivo analysis of herpessimplex virus assembly in murine visual system neurons. J. Virol. 79:11142–11150.
22. Lee CK, Knipe DM. 1985. An immunoassay for the study of DNA-binding activities of herpes simplex virus protein ICP8. J. Virol. 54:731–738.
23. Lee CK, Knipe DM. 1983. Thermolabile in vivo DNA-binding activityassociated with a protein encoded by mutants of herpes simplex virus type1. J. Virol. 46:909 –919.
24. Makhov AM, Griffith JD. 2006. Visualization of the annealing of com-plementary single-stranded DNA catalyzed by the herpes simplex virustype 1 ICP8 SSB/recombinase. J. Mol. Biol. 355:911–922.
25. Mapelli M, Panjikar S, Tucker PA. 2005. The crystal structure of theherpes simplex virus 1 ssDNA-binding protein suggests the structural ba-sis for flexible, cooperative single-stranded DNA binding. J. Biol. Chem.280:2990 –2997.
26. Mavromara-Nazos P, Roizman B. 1989. Delineation of regulatory do-mains of early (beta) and late (gamma 2) genes by construction of chime-ric genes expressed in herpes simplex virus 1 genomes. Proc. Natl. Acad.Sci. U. S. A. 86:4071– 4075.
27. Murphy CG, et al. 2000. Vaccine protection against simian immunode-ficiency virus by recombinant strains of herpes simplex virus. J. Virol.74:7745–7754.
28. Nimonkar AV, Boehmer PE. 2003. The herpes simplex virus type-1single-strand DNA-binding protein (ICP8) promotes strand invasion. J.Biol. Chem. 278:9678 –9682.
29. Nimonkar AV, Boehmer PE. 2002. In vitro strand exchange promoted bythe herpes simplex virus type-1 single strand DNA-binding protein(ICP8) and DNA helicase-primase. J. Biol. Chem. 277:15182–15189.
30. Nimonkar AV, Boehmer PE. 2003. Reconstitution of recombination-dependent DNA synthesis in herpes simplex virus 1. Proc. Natl. Acad. Sci.U. S. A. 100:10201–10206.
31. Nimonkar AV, Boehmer PE. 2004. Role of protein-protein interactionsduring herpes simplex virus type 1 recombination-dependent replication.J. Biol. Chem. 279:21957–21965.
32. Poffenberger KL, Tabares E, Roizman B. 1983. Characterization of aviable, noninverting herpes simplex virus 1 genome derived by insertionand deletion of sequences at the junction of components L and S. Proc.Natl. Acad. Sci. U. S. A. 80:2690 –2694.
33. Quinlan MP, Knipe DM. 1985. Stimulation of expression of a herpessimplex virus DNA-binding protein by two viral functions. Mol. Cell. Biol.5:957–963.
34. Reuven NB, Staire AE, Myers RS, Weller SK. 2003. The herpes simplexvirus type 1 alkaline nuclease and single-stranded DNA binding proteinmediate strand exchange in vitro. J. Virol. 77:7425–7433.
35. Reuven NB, Weller SK. 2005. Herpes simplex virus type 1 single-
Divalent Metal Cation Binding Site of HSV-1 ICP8
June 2012 Volume 86 Number 12 jvi.asm.org 6833
on February 13, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

strand DNA binding protein ICP8 enhances the nuclease activity of theUL12 alkaline nuclease by increasing its processivity. J. Virol. 79:9356 –9358.
36. Reuven NB, Willcox S, Griffith JD, Weller SK. 2004. Catalysis of strandexchange by the HSV-1 UL12 and ICP8 proteins: potent ICP8 recombi-nase activity is revealed upon resection of dsDNA substrate by nuclease. J.Mol. Biol. 342:57–71.
37. Roizman B, Jacob RJ, Knipe DM, Morse LS, Ruyechan WT. 1979. Onthe structure, functional equivalence, and replication of the four arrange-ments of herpes simplex virus DNA. Cold Spring Harb. Symp. Quant Biol.43(Pt. 2):809 – 826.
38. Ruyechan WT. 1983. The major herpes simplex virus DNA-binding pro-tein holds single-stranded DNA in an extended configuration. J. Virol.46:661– 666.
39. Schaffer P, Vonka V, Lewis R, Benyesh-Melnick M. 1970. Temperature-sensitive mutants of herpes simplex virus. Virology 42:1144 –1146.
40. Taylor TJ, Knipe DM. 2004. Proteomics of herpes simplex virus replica-tion compartments: association of cellular DNA replication, repair, re-combination, and chromatin remodeling proteins with ICP8. J. Virol.78:5856 –5866.
41. Taylor TJ, McNamee EE, Day C, Knipe DM. 2003. Herpes simplex virusreplication compartments can form by coalescence of smaller compart-ments. Virology 309:232–247.
42. Uprichard SL, Knipe DM. 2003. Conformational changes in the herpessimplex virus ICP8 DNA-binding protein coincident with assembly inviral replication structures. J. Virol. 77:7467–7476.
43. Ward SA, Weller SK. 2011. HSV-1 DNA replication, p 89 –112. In WellerSK (ed), Alphaherpesviruses: molecular virology. Caister Academic Press,Norfolk, United Kingdom.
44. Wu CA, Nelson NJ, McGeoch DJ, Challberg MD. 1988. Identification ofherpes simplex virus type 1 genes required for origin-dependent DNAsynthesis. J. Virol. 62:435– 443.
Bryant et al.
6834 jvi.asm.org Journal of Virology
on February 13, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from