identification andfunctional characterization tia-1 ... · vol. 89, pp. 8681-8685, september1992...
TRANSCRIPT

Proc. Nat!. Acad. Sci. USAVol. 89, pp. 8681-8685, September 1992Immunology
Identification and functional characterization of aTIA-1-related nucleolysin
(apoptosis/p d cell death/cytolytic lymphocytes/cytotoxic granules)
ATSUSHI KAWAKAMI*, QINGSHENG TIAN*, XIAOCHU DUAN*, MICHEL STREULI*, STUART F. SCHLOSSMAN*,AND PAUL ANDERSON*t*Division of Tumor Immunology, Dana-Farber Cancer Institute, 44 Binney Street, Boston, MA 02115; and tDepartment of Rheumatology and Immunology,Brigham and Women's Hospital, 75 Francis Street, Boston, MA 02115 and Harvard Medical School, Boston, MA 02115
Contributed by Stuart F. Schlossman, June 22, 1992
ABSTRACT We recently reported the molecular cloningof a cytotoxic granule-associated RNA-binding protein desig-nated TIA-1. The ability of recombinant TIA-1 to induce DNAfragmentation in permeabilized cells suggested that this proteinis the granule component responsible for inducing apoptosis incytolytic lymphocyte (CTL) targets. Here we report the char-acterization of a cDNA encoding a TIA-1-related proteindesignated TIAR. The deduced amino acid sequence of TIARreveals it to be a 42-kDa protein possessing three RNA-bindingdomains and a carboxyl-terminal auxiliary domain. Althoughthe RNA-binding domains of TIA-1 and TIAR share >85%amino acid homology, their carboxyl-terminal auxiliary do-mains are only 51% homologous. The carboxyl terminus ofTIAR contains a lysosome-targeting motif, indicating thatTUAR is probably a cytotoxic granule-associated protein. LikeTIA-1, purified recombinant TIAR induced DNA fragmenta-tion in permeabilized target cells. Although immunoblottinganalysis of post-nuclear supernatants revealed TIA-1 protein tobe restricted to CTLs, PCR analysis revealed the expression ofTIA-1 and TIAR mRNA transcripts in a wide variety of celltypes. Our data suggest that the granules of CTLs contain atleast two candidate nucleolysins involved in CTL killing.
Natural killer (NK) cells and cytotoxic lymphocytes (CTLs)can recognize and kill a wide variety of virus-infected ortransformed target cells (1, 2). Although the surface receptorsexpressed by these effector cells are distinct, they appear touse similar cytolytic mechanisms in the elimination of theirrespective target cells (3-6). Both NK cells and CTLspossess cytoplasmic granules containing putative effectormolecules, such as perforin and the serine proteases. Secre-tion of these granules in response to target cell recognition isthought necessary for the induction of target cell death.Indeed, isolated cytotoxic granules can render target cellspermeable to small ions such as Ca2+ and 51Cr (7). Thegranule component responsible for this permeabilization isperforin, a complement-related pore-forming protein. WithCa2+, perforin can insert into target cell membranes, whereit polymerizes to form nonspecific ion channels (8-10).Formation of these ion channels appears sufficient to inducethe lysis of certain cell types.
In addition to perforn-mediated cytolysis, CTLs can in-duce target cells to activate an endogenous pathway ofprogrammed cell death known as apoptosis (11). Central tothis autolytic pathway is the activation of an endogenousendonuclease that degrades target cell DNA (12, 13). Elec-trophoretic separation of degraded DNA produces a "lad-der" of oligonucleosome-sized fragments characteristic ofthis programmed suicide pathway. Isolated cytotoxic gran-
ules have been reported by some investigators (7, 14) toinduce DNA fragmentation in target cells, suggesting thatsome constituent of the cytotoxic granule possesses nucleo-lytic activity. Although pore-forming molecules such as per-forin and complement are generally not thought to triggerDNA fragmentation (15), prolonged exposure to these agentssometimes causes nonspecificDNA damage (16) that may notbe relevant to lymphocyte-mediated killing. Perforin may,however, be required for the nucleolytic activity of othergranule components. Recent results suggest that the combi-nation of a granule-associated serine protease (e.g., gran-zyme A or a rat natural killer protease 1-like protease), andperforin can induce DNA fragmentation in target cells (17,18). The further observation that granzyme A can induceDNA fragmentation in detergent-permeabilized cells (17)suggests that perforin is involved in the delivery of gran-zymes to target cells. The mechanisms by which serineproteases induce DNA fragmentation are completely un-known.We recently identified several molecular isoforms (15 kDa,
28 kDa, 40 kDa, and 53 kDa) of a cytotoxic granule-associated protein designated TIA-1. Molecular cloning ofthe 40-kDa TIA-1 isoform (p40-TIA-1) revealed it to beanother member of a family of RNA-binding proteins (19).p40-TIA-1 is composed of three 90-amino acid RNA-bindingdomains and a 102-amino acid glutamine-rich carboxyl-terminal auxiliary domain. The 15-kDa TIA-1 isoform, themajor species in cytotoxic granules, may be derived from thecarboxyl-terminal auxiliary domain of p40-TIA-1 by proteo-lytic processing (19). Because p15-TIA-1 possesses nucleo-lytic activity, release of this carboxyl-terminal fragment bygranule-associated proteases (e.g., granzymes) may serve toactivate the toxin before granule secretion. We postulatedthat the ability of serine proteases to induce DNA fragmen-tation in permeabilized cells might result from a similaractivation of TIA-1-related molecules expressed in the nu-cleus of all cells (19). Here we report the molecular cloningof a TIA-1-related cDNA that encodes a protein (TIAR)possessing nucleolytic activity against CTL target cells.t Ourresults suggest that a family of granule-associated RNA-binding proteins are candidate nucleolysins involved in CTL-mediated killing.
MATERIALS AND METHODS
Immunoblotting. Immunoblotting analysis was done asdescribed (20). Immunoblots developed with a monoclonalantibody reactive with TIA-1 (1H10, IgGl) as described (19).
Abbreviations: CTL, cytotoxic lymphocyte; TIAR, TIA-1-relatedprotein; RP-PCR, reverse transcription PCR; NK, natural killer.*The sequence reported in this paper has been deposited in theGenBank data base (accession no. M%954).
8681
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

8682 Immunology: Kawakami et al.
Affinity Precipitations. Escherichia coli strain BL21 (DE3)was transformed with the pT7-7 plasmid vector containinginsert DNA encoding either TIA-1 or TIAR, and bacteriallysates were prepared as described (19). Recombinant pro-teins were affinity precipitated from E. coli lysates by usingeither poly(A)-Sepharose or poly(U)-Sepharose (Pharmacia).E. coli lysates [derived from 0.5 ml ofbacterial culture (OD600= 0.45)] were incubated with 50 Aul of a 50%o (vol/vol)suspension of Sepharose beads for 1 hr at 4TC. Sepharosebeads were then washed three times with Tris-bufferedsaline, eluted with SDS-sample buffer, and separated on anSDS/10% polyacrylamide gel. After transfer to nitrocellulose(Schleicher & Schuell), immunoblots were developed asdescribed (19).cDNA Cloning and Sequencing. A Agtll cDNA library
derived from phytohemagglutinin-activated T cells (Clon-tech) was plated at a density of 10,000 plaques per 150-mmPetri dish onto a lawn of E. coli strain Y1090. After 12 hr at37TC, plates were overlaid with nitrocellulose filters for 1 minbefore being processed for hybridization. Individual filterswere prehybridized in 50% (vol/vol) formamide, 5 X standardsaline citrate (SSC; lx SSC is 0.15 M sodium chloride/0.015M sodium citrate, pH 7)/50mM sodium phosphate buffer, pH7.0/5 x Denhardt's solution/0.1% SDS/denatured salmonsperm DNA at 100 jig/ml for 4 hr at 37TC. Probe DNAs were32P-labeled by nick translation, diluted in the above solution,and hybridized to nitrocellulose filters for 12-24 hr at 370C.Filters were then washed twice with 2x SSC/0.1% SDSbefore autoradiographic exposure. Positive plaques wereindividually selected and subcloned three times before ex-pansion. Phage DNA was extracted from plate lysates,digested with EcoRI to liberate inserts, and subcloned intopSP65 plasmid DNA. Dideoxynucleotide chain-terminationsequencing was performed using Sequenase (United StatesBiochemical).Southern Blotting. Human genomic DNA (Clontech) was
digested with the indicated restriction enzymes for 4 hr at37°C before electrophoretic separation on a 0.7% agarose gel.After alkali denaturation, DNA fragments were transferred tonitrocellulose by capillary blotting for 12 hr at room temper-
u EXco
cn m z
Ir_ 0
LU
* # w S w ~~~-12 kb
-7.1 kb
-5 kb
-4 kb
-3 kb
-2 kb
-1 kb
FIG. 1. Southern blot of TIA-1 genomic DNA. Total genomicDNA from human lymphocytes was digested with the indicated
restriction enzymes, separated on a 1% agarose gel, and transferred
to nitrocellulose. The blot was then probed with a 32P-labeled cDNA
encoding p40-TIA-1 (A2G9.4) and developed by autoradiography.Relative migration of molecular-size markers [in kilobases (kb)] is
indicated at right.
Proc. NatL. Acad. Sci. USA 89 (1992)
ature. Nitrocellulose filters were then treated with UV lightbefore prehybridization and hybridization using a 32P-labeledprobe encoding TIA-1.
Purification of Recombinant Proteins. Both recombinantTIA-1 and TIAR were purified by using a modification of thetwo-step affinity chromatography procedure described forTIA-1 purification (19). In the immunoaffinity chromatogra-phy step, we used monoclonal antibody 1H10instead ofclone2G9 (19).DNA Fragmentation. TIA-1 and TIAR were tested for their
ability to induce DNA fragmentation in digitonin-permeabi-lized human thymocytes, as described (19).PCR Analysis. Total cellular RNA was extracted from the
indicated cell types as described (19). cDNA was synthesized1 21 41ACC CTG CCC TCG GCC TTG TCC CGG GAT CGC TCC GTC GCA CCC ACC
61 81ATG ATG GAA GAC GAC GGG CAG CCC CGG ACT CTA TAC GTA GGT AAC
1: met met glu asp asp gly gin pro arg thr leu tyr val gly asn
101 121CTT TCC AGA GAT GTG ACA GAA GTC CTT ATA CTT CAG TTG TTC AGT
16:1eu ser arg asp val thr glu val leu 1le leu gin leu ph. ser141 161
CAG ATT GGA CCC TGT AAA AGC TGT AAA ATG ATA ACA GAG CAT ACA31:gln i11 gly pro cys lys ser cys lys met ile thr glu his thr
181 201 221AGC AAT GAC CCA TAT TGC TTT GTG GAA TTT TAT GAA CAC AGA GAT
46:ser asn asp pro tyr cys phe val glu phe tyr glu his arg asp241 261
GCA GCT GCT GCA TTA GCT GCT ATG AAT GGG AGA AAA ATT TTG GGA61:ala ala ala ala leu ala ala met asn gly arg lys le leu gly
281 301AAG GAG GTC AAA GTA AAC TGG GCA ACC ACA CCA AGT AGC CAG AAA
76:1ys glu val lys val asn trp ala thr thr pro s-r s-r gin lys321 341
AAA GAT ACT TCC AAT CAC TTC CAT GTG TTT GTT GGG GAT TTG AGT91:lys asp thr ser asn his phe his val phe val gly asp lou ser
361 381 401CCA GAA ATT ACA ACA GAA GAT ATC AAA TCA GCA TTT GCC CCC TTT
106pro glu ile thr thr glu asp ile lys sor ala phe ala pro ph.421 441
GGT AAA ATA TCG GAT GCC CGG GTA GTT AAA GAC ATG GCA ACT GGA131gly lys ile ser asp ala arg val val lys asp met ala thr gly
461 481AAA TCC AAA GGC TAT GGT TTT GTA TCT TTT TAT AAC AAA CTG GAT
1461ys ser lys gly tyr gly ph. val ser phe tyr asn lys leu asp501 521
GCA GAA AAT GCG ATT GTG CAT ATG GGC GGT CAG TGG TTG GGT GGT161ala glu asn ala ii1 val his met gly gly gin trp leu gly gly
541 561 581CGT CAA ATC CGA ACC AAT TGG GCC ACT CGT AAA CCA CCT GCA CCT
176arg gin ile arg thr asn trp ala thr arg lys pro pro ala pro601 621
AAA AGT ACA CAA GAA AAC AAC ACT AAG CAG TTG AGA TTT GAA GATl911ys ser thr gin glu asn asn thr lys gin leu arg phe glu asp
641 661GTA GTA AAC CAG TCA AGT CCA AAA AAT TGT ACT GTG TAC TGT GGA
206va1 val asn gin ser ser pro lys asn cys thr val tyr cys gly681 701
GGA ATT GCG TCT GGG TTA ACA GAT CAG CTT ATG AGA CAG ACA TTC216gly 11 ala sor gly leu thr asp gin leu met arg gin thr phe
721 741 761TCA CCA TTT GGA CAA ATT ATG GAA ATA AGA GTT TTC CCA GAA AAG
231ser pro ph. gly gin ile met glu ile arg val ph. pro glu lys781 801
GGC TAT TCA TTT GTC AGA TTT TCA ACC CAT GAA AGT GCA GCC CAT246gly tyr ser ph. val arg phe ser thr his glu ser ala ala his
821 841GCC ATT GTT TCG GTG AAC GGT ACT ACG ATT GAA GGA CAT GTG GTT
261ala ile val ser val asn gly thr thr ile glu gly his val val861 881
AAA TGC TAT TGG GGT AAA GAA TCT CCT GAT ATG ACT AM AAC TTC2761ys cys tyr trp gly lys glu ser pro asp met thr lys asn phe
901 921 941CAA CAG GTT GAC TAT AGT CAA TGG GGC CAA TGG AGC CAA GTG TAT
291gln gin val asp tyr ser gin trp gly gin trp ser gin val tyr961 981
GGA AAC CCA CAA CAG TAT GGA CAG TAT ATG GCA AAT GGG TGG CAA306gly asn pro gin gin tyr gly gin tyr met ala asn gly trp gin
1001 1021GTA CCG CCT TAT GGA GTA TAC GGG CAA CCA TGG AAT CAA CAA GGA
321val pro pro tyr gly val tyr gly gin pro trp asn gin gin gly1041 1061
TTT GGA GTA GAT CAA TCA CCT TCT GCT GCT TGG ATG GGT GGA TTT336phe gly val asp gin snr pro ser ala ala trp met gly gly phe
1081 1101 1121GGT GCT CAG CCT CCC CAA GGA CAA GCT CCT CCC CCT GTA ATA CCT
351gly ala gin pro pro gin gly gin ala pro pro pro val ile pro1141 1161
CCT CCT AAC CAA GCC GGA TAT GGT ATG GCA AGT TAC CAA ACA CAG366pro pro asn gin ala gly tyr gly met ala ner tyr gin thr gin
1181 1201TCA GCC GGG ACT CTA AAA AAA AAT TGT AAT TCA TGA TAG GCT TCG
1221 1241ATT TCC TGT GAC ACT CTG AAG ACA TGA AAG TAG ACA TCG GAA AAT1261 1281 1301GAA AAT ATT TAT TTT AAA AAT TGA AAT GTT TGG AAC CTT TAG CAC
1321 1341AGA TTT GCT TTG GTG AAG GAC ACG TGT CTT CTA GTT CTG CCT TTT
1361 1381TAA GTT TTT GTT CAT GAT GGA TAT GAA CAT GAT TTT TCT TTA TGTACA AAA
FIG. 2. Nucleotide sequence and predicted amino acid sequenceof TIAR.

Proc. Natl. Acad. Sci. USA 89 (1992) 8683
from an RNA template (3 ,ug) with Moloney murine leukemiavirus reverse transcriptase (200 units) (BRL) and randomhexanucleotide primers (150 pmol) (Pharmacia) in 75 mMKCl/50 mM Tris-HCl, pH 8.3/3 mM MgCl2/10 mM dithio-threitol/125 uM dNTP (total vol = 50 .ul) for 2 hr at 420C. PCRreactions were done in 100,ul of50mM KCl/15 mM Tris HCl,pH 8.3/2 mM MgCl2/0.001% gelatin (wt/vol)/200 gMdNTP/100 pmol ofoligonucleotide primers and 20 /l from theabove cDNA reaction. After the addition of 2.5 units ofAmpliTaq DNA polymerase (Perkin-Elmer/Cetus), 30 cy-cles of PCR consisting of 1 min at 94°C, 1 min at 55°C, 2 minat 72°C were completed before extracting with chloroformprior to electrophoretic separation on a 2% agarose gelcontaining ethidium bromide (0.5 ,g/ml). The 5' PCR primerused in the amplification of TIA-1/TIAR cDNAs was GG-TAACCTTTCCAGAGAT. The 3' primer used in these am-plifications was CCCCAATAGCATTT.
Sequence Analysis of PCR Products. PCR products wereseparated on a 1% low-melting-temperature agarose gel,identified by transillumination under UV light, and extractedusing Geneclean (Bio 101, La Jolla, CA). Extracted DNA wascloned into the plasmid vector pCR 1000 (Invitrogen, SanDiego), according to the manufacturer's instructions. Se-quence analysis was done with synthetic primers comple-mentary to the vector DNA at the junction of the cloning siteusing Sequenase (United States Biochemical).
RESULTSMolecular Cloning of a TIA-1-Related cDNA. The complex-
ity of Southern blots prepared from genomic DNA digestedwith several restriction enzymes and probed with a TIA-1cDNA (Fig. 1) suggested either that the TIA-1 gene was largeor that TIA-1-related genes were being detected by crosshybridization. To test the latter possibility, we screened aAgtll cDNA library derived from phytohemagglutinin-activated T cells with a 32P-labeled TIA-1 cDNA probe.Although most cDNAs thus isolated were derived from theTIA-1 gene, a distinct, but related, cDNA was also identified.A partial nucleotide sequence of this cDNA, designatedTIAR, is shown in Fig. 2. We were unable to sequence theentire 5' untranslated region ofTIAR because ofcompressionartifacts. The first methionine in the open reading frameconforms to the consensus sequence for eukaryotic initiationsites (21). The further observation that recombinant TIA-1and TIAR comigrate by SDS/PAGE suggests that the firstencountered methionine is the initiation site. Comparison ofthe predicted amino acid sequence of TIAR with that ofTIA-1 is shown in Fig. 3. Like TIA-1, TIAR is a member ofa family of RNA-binding proteins, possessing three RNA-binding domains and a carboxyl-terminal auxiliary domain.In their RNA-binding domains, TIA-1 and TIAR are highly
10 * 20 30 40
TIA-1 A G Y E TQ - COOH
TIAR A S Y Q T Q - COOH
LAMP-1 A G Y Q T I - COOH
LAMP-2 A GY E Q F - COOH
LAMP-3 S G Y E V M - COOH
Igp1 20 A G Y Q T I - COOH
Consensus: A G Y Q Q - COOK
FIG. 4. Comparison of carboxyl-terminal six amino acids ofTIA-1, TIAR, and several lysosome-associated membrane proteinsdefining a lysosome-targeting motif (1gpl2O, lysosomal glycopro-tein). The critical tyrosine residue located four amino acids from thecarboxyl terminus is shown in boldface type. A consensus sequencederived from this comparison is shown at bottom.
homologous (79o, 89%, and 91% amino acid homology indomains 1, 2, and 3, respectively). Although their carboxyl-terminal auxiliary domains are less related (50% amino acidhomology), both molecules possess a lyosome-targeting mo-tif (Fig. 4), suggesting that TIAR is also a cytotoxic granule-associated protein.
Expression and Characterization of TIAR. The structuralsimilarity between TIA-1 and TIAR suggested that TIAR isprobably an RNA-binding protein. To confirm this, thecDNA encoding TIAR was subcloned into the pT7-7 induc-ible expression vector and used to transform E. coli BL21(DE3) cells. Affinity precipitation from bacterial lysates witheither poly(C)-agarose or poly(A)-agarose demonstrated thatboth TIA-1 and TIAR bind to poly(A) but not to poly(C) (Fig.SA). In E. coli lysates, two molecular species of TIA-1 andTIAR (p40 and p38) are reproducibly detected. The predom-inant 40-kDa species is likely to be the full-length recombi-nant protein. The 38-kDa species is probably a proteolyticdegradation product of the full-length recombinant protein ora translation product initiated from an internal methioninecodon. Recognition of TIAR by the monoclonal antibody1H10 (19) allowed us to purify this recombinant protein withthe two-step affinity chromatography procedure used inTIA-1 purification (19). Fig. SB shows the electrophoreticmigration of purified TIAR on a Coomassie blue-stainedSDS/polyacrylamide gel.The structural similarity between TIA-1 and TIAR sug-
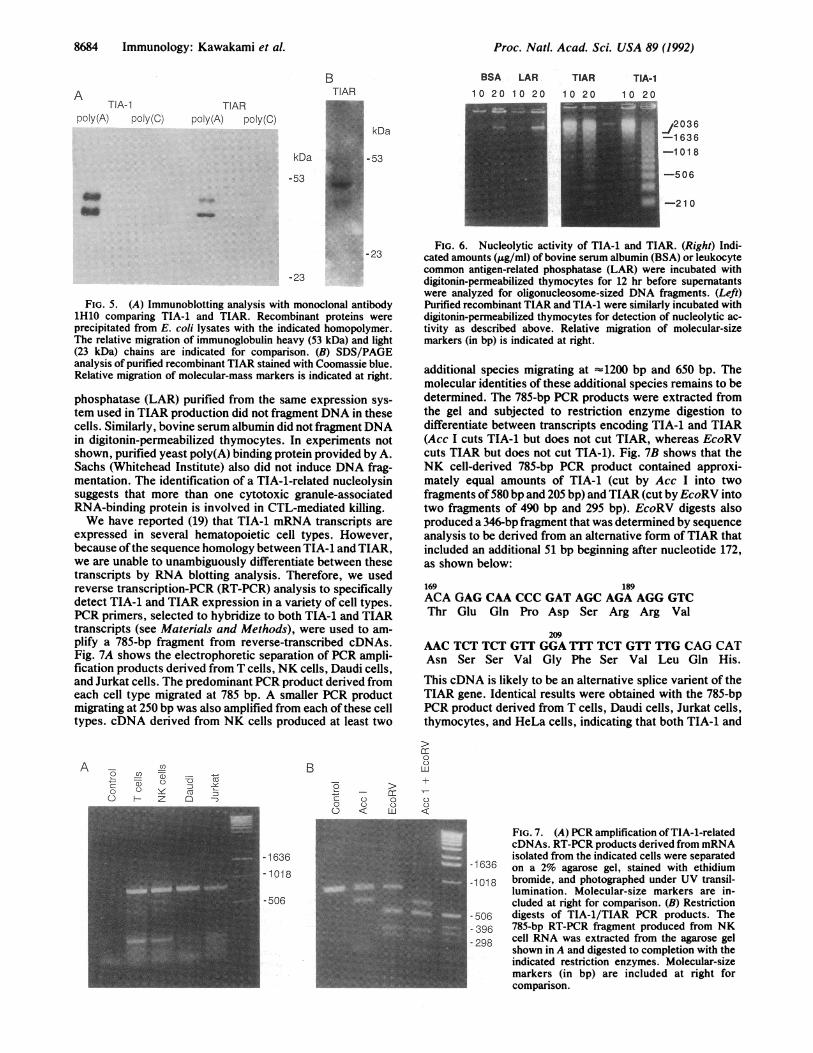
gested that these molecules function similarly. Therefore, wemeasured the nucleolytic activity of purified TIAR withdigitonin-permeabilized thymocytes. Fig. 6 shows that TIARalso fragments DNA in permeabilized thymocytes. Equalamounts of recombinant leukocyte common antigen-related
50 ** 60TIAR: MMEDDGQPRT LYVGNSRDV TEVLILQLFS QIGPCKSCKM ITEHTSNDPY CFVKFYEHRDTIA-1: .---E.M-K- ---------- --A------- ------N--- -MDTAG---- -----H--GH
70 80 90 100 * 110 120TIAR: AAAALAAMNG RKILGKEVKV NWATTPSSQK KDTSNHFHVF VGDLSPEITT EDIKSAFAPFTIA-1: ------L--- ---M------ ---------- ---T------ ---------- ----A-----
130 140 ** 150 160 170 180TIAR: GKISDARVVK DMATGKSKGY GFVSFYNKLD AENAIVHMGG QWLGGRQIRT NWATRKPPAPTIA-1: -R-------- ---------- -----F--W- -----QQ--- --------F- ----------
190 200 210 * 220 230 240TIAR: KSTQENNTKQ LRFEDVVNQS SPKNCTVYCG GIASGLTDQL MRQTFSPFGQ IMEIRVFPEKTIA-1: ---Y-S---- -SYDE----- --S------- -VT----E-- ---------- --------D-
** 250 260 270 280 290 300TIAR : GYSFVRFSTH ESAAHAIVSV NGTTIEGHVV KCYWGKESPD MTKNFQQVDY.... SQWGQWSQVYTIA-1: ---- NS- ---------- --------- - -------TL- -INPV--ONgigyPOPY---G-W-
310 320 330 340 350 360TIAR : GNPQQYGQYM ANGWQVPPYG VYGQPWNQQG FGVDQSPSAA WMGGF . GAQPP QGQAPPPVIPTIA-1: --A--IT---- P------A-- M---A----- -NOT--..--P ---PNY-V--- ---NGSL..
370TIAR: PPNQAGYGMA SYQTQTIA-1: -NOPS--RV- G-E--
FIG. 3. Amino acid sequence compari-son of TIA-1 and TIAR. Identical aminoacids are depicted by -, and gaps aredepicted by *. Ribonucleoprotein 2(RNP2) hexamer (*) and ribonucleopro-tein 1 (RNP1) octamer (**) sequencescorresponding to the three RNA-bindingdomains are in boldface type. The car-boxyl-terminal auxiliary domain isunderlined.
Immunology: Kawakami et al.
8684 Immunology: Kawakami et al.
ATIA-1
poly(A) poly(C)
BTIAR
TIARpoly(A) poly(C)
kDa
-53
BSA LAR TIAR TIA-110 20 10 20
kDa
-53
am.
I236-1636-1018
-506
-23
-23
FIG. 5. (A) Immunoblotting analysis with monoclonal antibody1H10 comparing TIA-1 and TIAR. Recombinant proteins wereprecipitated from E. coli lysates with the indicated homopolymer.The relative migration of immunoglobulin heavy (53 kDa) and light(23 kDa) chains are indicated for comparison. (B) SDS/PAGEanalysis of purified recombinant TIAR stained with Coomassie blue.Relative migration of molecular-mass markers is indicated at right.
phosphatase (LAR) purified from the same expression sys-tem used in TIAR production did not fragment DNA in thesecells. Similarly, bovine serum albumin did not fragment DNAin digitonin-permeabilized thymocytes. In experiments notshown, purified yeast poly(A) binding protein provided by A.Sachs (Whitehead Institute) also did not induce DNA frag-mentation. The identification of a TIA-1-related nucleolysinsuggests that more than one cytotoxic granule-associatedRNA-binding protein is involved in CTL-mediated killing.We have reported (19) that TIA-1 mRNA transcripts are
expressed in several hematopoietic cell types. However,because ofthe sequence homology between TIA-1 and TIAR,we are unable to unambiguously differentiate between thesetranscripts by RNA blotting analysis. Therefore, we usedreverse transcription-PCR (RT-PCR) analysis to specificallydetect TIA-1 and TIAR expression in a variety of cell types.PCR primers, selected to hybridize to both TIA-1 and TIARtranscripts (see Materials and Methods), were used to am-plify a 785-bp fragment from reverse-transcribed cDNAs.Fig. 7A shows the electrophoretic separation of PCR ampli-fication products derived from T cells, NK cells, Daudi cells,and Jurkat cells. The predominant PCR product derived fromeach cell type migrated at 785 bp. A smaller PCR productmigrating at 250 bp was also amplified from each of these celltypes. cDNA derived from NK cells produced at least two
A -_ n-5
O Q) Y0 Z
B
: =o >
- ccC C.0 v oC) < Lu
FIG. 6. Nucleolytic activity of TIA-1 and TIAR. (Right) Indi-cated amounts (j.g/ml) of bovine serum albumin (BSA) or leukocytecommon antigen-related phosphatase (LAR) were incubated withdigitonin-permeabilized thymocytes for 12 hr before supernatantswere analyzed for oligonucleosome-sized DNA fragments. (Left)Purified recombinant TIAR and TIA-1 were similarly incubated withdigitonin-permeabilized thymocytes for detection of nucleolytic ac-tivity as described above. Relative migration of molecular-sizemarkers (in bp) is indicated at right.
additional species migrating at -1200 bp and 650 bp. Themolecular identities of these additional species remains to bedetermined. The 785-bp PCR products were extracted fromthe gel and subjected to restriction enzyme digestion todifferentiate between transcripts encoding TIA-1 and TIAR(Acc I cuts TIA-1 but does not cut TIAR, whereas EcoRVcuts TIAR but does not cut TIA-1). Fig. 7B shows that theNK cell-derived 785-bp PCR product contained approxi-mately equal amounts of TIA-1 (cut by Acc I into twofragments of580 bp and 205 bp) and TIAR (cut by EcoRV intotwo fragments of 490 bp and 295 bp). EcoRV digests alsoproduced a 346-bp fragment that was determined by sequenceanalysis to be derived from an alternative form of TIAR thatincluded an additional 51 bp beginning after nucleotide 172,as shown below:
169 189ACA GAG CAA CCC GAT AGC AGA AGG GTCThr Glu Gln Pro Asp Ser Arg Arg Val
209AAC TCT TCT GTT GGA TTT TCT GTT TTG CAG CATAsn Ser Ser Val Gly Phe Ser Val Leu Gln His.
This cDNA is likely to be an alternative splice varient of theTIAR gene. Identical results were obtained with the 785-bpPCR product derived from T cells, Daudi cells, Jurkat cells,thymocytes, and HeLa cells, indicating that both TIA-1 and
cc00LU
v-u
FIG. 7. (A) PCR amplification ofTIA-1-relatedcDNAs. RT-PCR products derived from mRNAisolated from the indicated cells were separated
-1636 on a 2% agarose gel, stained with ethidium-1018 bromide, and photographed under UV transil-
lumination. Molecular-size markers are in-cluded at right for comparison. (B) Restriction
-506 digests of TIA-1/TIAR PCR products. The- 396 785-bp RT-PCR fragment produced from NK298 cell RNA was extracted from the agarose gel
- shown in A and digested to completion with theindicated restriction enzymes. Molecular-sizemarkers (in bp) are included at right forcomparison.
-21 0
-1636-1018
-506
Proc. Natl. Acad. Sci. USA 89 (1992)

Proc. NatL. Acad. Sci. USA 89 (1992) 8685
TIAR are widely expressed in hematopoietic and nonhe-matopoietic cell types.
DISCUSSIONWe recently described the cDNA cloning and functionalcharacterization of a cytotoxic granule-associated proteindesignated TIA-1 (19). Purified recombinant TIA-1 was foundto induce DNA fragmentation in permeabilized target cells,suggesting that TIA-1 is the granule component responsiblefor inducing programmed cell death, or apoptosis, in CTLtargets. In the present report, we describe the identificationand characterization of a TIA-1-related cDNA encoding anRNA-binding protein designated TIAR. Both TIA-1 andTIAR possess three highly homologous RNA-binding do-mains at their amino termini. Both recombinant proteins bindspecifically to poly(A) but not to poly(C) homopolymers. Thecarboxyl-terminal auxiliary domains of TIA-1 and TIARincluded a lysosome-targeting motif that has been shown tolocalize lamp-1 (lysosome-associated membrane protein),lamp-2, and Igp-120 (lysosomal glycoprotein) to the mem-branes of cytotoxic granules and lysosomes (22). RT-PCRanalysis indicates that both TIA-1 and TIAR are expressed ina wide variety of human cell types. Purified recombinantTIAR induces DNA fragmentation in digitonin-permeabi-lized target cells. Taken together, our results suggest that twoor more cytotoxic granule-associated RNA-binding proteinsmay be involved in inducing apoptosis in CTL target cells.Due to their structural similarity and immunologic cross-
reactivity, we cannot easily distinguish TIA-1 from TIAR inimmunoblots or RNA blots. RT-PCR analysis indicates thatboth species are expressed in cytotoxic and noncytotoxic celltypes. Expression of TIA-1-related mRNAs in noncytotoxiccells previously led us to hypothesize that certain isoforms ofthis molecule served an essential regulatory function in allcells (19). Although both TIA-1 and TIAR can degrade DNAin permeabilized cells, neither molecule can degrade nativeDNA (unpublished observations), suggesting that these pro-teins activate target cell endonucleases. It is possible thatTIA-1-related molecules found in noncytotoxic cells mightregulate endonucleases normally involved in DNA replica-tion or repair. In this regard, CTLs could have redirectedsuch an endonuclease regulator to their cytotoxic granules tofunction as a cellular toxin. Although TIA-1 (and presumablyTIAR) is associated with cytotoxic granule membranes, it isnot a transmembrane protein, nor does TIA-1 possess a signalpeptide that would account for its delivery to the secretorypathway. However, both TIA-1 and TIAR possess a car-boxyl-terminal lysosome-targeting motif that has been shown(23, 24) to retain lamp-1 and lamp-2 in cytotoxic granules andlysosomes. In lamp-1 and lamp-2, this targeting motif isoriented toward the cytoplasmic face of the outer-granulemembrane, where it presumably interacts with a granule-associated receptor. Recognition of the lysosome-targetingmotifofTIA-1 by this putative granule receptor might explainits localization to cytotoxic-granule membranes. Cytotoxicgranules contain many small internal vesicles that are thoughtto form by budding from the outer limiting membrane. It ispossible that TIA-1 and TIAR are delivered to the intersticesofthese internal vesicles, a localization that would allow theirrelease during granule exocytosis.We previously showed that the 15-kDa isoform of TIA-1
can be derived from the 40-kDa RNA-binding protein byproteolytic processing (19). Both TIA-1 and TIAR share apotential serine protease-cleavage site (Phe-Val-Arg, aminoacids 242-244) localized at the junction between their RNA-binding domains and their carboxyl-terminal auxiliary do-mains. The 15-kDa isoform ofTIA-1 appears sufficient for theinduction of DNA fragmentation in permeabilized cells,suggesting that the glutamine-rich carboxyl terminus is re-
sponsible for interacting with an endogenous endonuclease orwith a proximal regulatory protein in an activation cascade.Such a function would be consistent with the propensity forthe carboxyl-terminal auxiliary domains of RNA-bindingproteins to interact with other proteins (25).
Expression of TIA-1 and TIAR in cells other than CTLssuggests that these molecules are involved in apoptosisinduced by stimuli other than cytotoxic granules. Indeed,apoptotic cell death can be induced in a variety of cell typesby multiple exogenous stimuli (26). The receptor-mediatedactivation ofT cells (27), B cells, and thymocytes (28) inducesthe expression of unknown intermediaries that activate anendogenous endonuclease. As described above, TIAR oranother TIA-1-related molecule is a candidate for such aregulatory intermediate. It will be important to determinewhether stimuli that induce apoptosis also induce the expres-sion of TIA-1 or TIAR. The identification of a putativeTIAR-derived alternative splice product suggests anotherway that these proteins might be regulated during the induc-tion of programmed cell death. Apoptotic cell death can alsobe induced by the passive withdrawal of required growthfactors. Again, the availability of cDNA probes encodingTIA-1 and TIAR will allow us to quantitate the expression ofthese genes after growth factor depletion.
This work was supported, in part, by grants from the NationalInstitutes of Health (CA53595 and A112069). P.A. was supported bya Pew Scholar Award and by a Carl J. Herzog Foundation Investi-gator Award from the Cancer Research Institute.
1. Trinchieri, G. (1989) Adv. Immunol. 47, 187-376.2. Ritz, J., Schmidt, R. E., Michon, J., Hercend, T. & Schlossman,
S. F. (1988) Adv. Immunol. 42, 181-211.3. Henkart, P. A. (1985) Annu. Rev. Immunol. 3, 31-58.4. Podack, E. R., Hengartner, H. & Lichtenheld, M. G. (1991) Annu.
Rev. Immunol. 9, 129-157.5. Tschopp, J. & Nabholz, M. (1990) Annu. Rev. Immunol. 8, 279.6. Young, J. D. E. & Liu, C. C. (1988) Immunol. Today 9, 140-144.7. Allbritton, N. L., Verret, C. R., Wolley, R. C. & Eisen, H. N.
(1988) J. Exp. Med. 167, 514-527.8. Young, J. D. E., Nathan, C. F., Podack, E. R., Palladino, M. A. &
Cohn, Z. (1986) Proc. Natd. Acad. Sci. USA 83, 150-154.9. Yue, C. C., Reynolds, C. W. & Henkart, P. A. (1987) Mol. Immu-
nol. 24, 647-653.10. Tschopp, J., Schafer, S., Masson, D., Peitch, M. C. & Heusser, C.
(1989) Nature (London) 337, 272-274.11. Russell, J. H. (1983) Immunol. Rev. 72, 97-118.12. Wyllie, A. H. (1980) Nature (London) 284, 555-556.13. Duke, R. C., Chervenak, R. & Cohen, J. J. (1983) Proc. Nat!. Acad.
Sci. USA 80, 6361-6365.14. Podack, E. R. (1986) J. Cell. Biochem. 30, 127-164.15. Duke, R. C., Persechini, P. M., Chang, S., Liu, C. C., Cohen, J. J.
& Young, J. D. E. (1989) J. Exp. Med. 170, 1451-1456.16. Hameed, A., Osen, K. J., Lee, M. K., Lichtenheld, M. G. &
Podack, E. R. (1989) J. Exp. Med. 169, 765-777.17. Hayes, M. P., Berrebi, G. A. & Henkart, P. A. (1989) J. Exp. Med.
170, 933-946.18. Shi, L., Kraut, R. P., Aebersold, R. & Greenberg, A. H. (1992) J.
Exp. Med. 175, 553-566.19. Tian, Q., Streuli, M., Saito, H., Schlossman, S. F. & Anderson, P.
(1991) Cell 67, 629-639.20. Anderson, P., Nagler-Anderson, C., O'Brien, C., Levine, H.,
Watkins, S., Slayter, H. S., Blue, M.-L. & Schlossman, S. F. (1990)J. Immunol. 144, 574-582.
21. Kozak, M. (1984) Nucleic Acids Res. 12, 857-872.22. Fukuda, M. (1991) J. Biol. Chem. 266, 21327-21330.23. Burkhardt, J. K., Hester, S., Lapham, C. K. & Argon, Y. (1990) J.
Cell Biol. 111, 1227-1240.24. Peters, P. J., Borst, J., Oorschot, V., Fukuda, M., Krahenbuhl, O.,
Tschopp, J., Slot, J. W. & Geuze, H. J. (1991) J. Exp. Med. 173,1099-1109.
25. Bandziulis, R. J., Swanson, M. S. & Dreyfuss, G. (1989) GenesDev. 3, 431-437.
26. Martz, E. & Howell, D. M. (1989) Immunol. Today 10, 79-86.27. Ucker, D. S. (1987) Nature (London) 327, 62-64.28. Smith, C. A., Williams, G. T., Kingston, R., Jenkinson, E. J. &
Owen, J. J. T. (1989) Nature (London) 337, 181-183.
Immunology: Kawakami et al.