hydrogen isotope fractionation during water uptake by woody xerophytes
TRANSCRIPT

Plant Soil (2007) 291:93–107
DOI 10.1007/s11104-006-9177-1ORIGINAL PAPER
Hydrogen isotope fractionation during water uptake by woody xerophytes
Patrick Z. Ellsworth · David G. Williams
Received: 19 September 2006 / Accepted: 30 November 2006 / Published online: 17 January 2007 Springer Science+Business Media B.V. 2007
Abstract Stable isotope measurements areemployed extensively in plant–water relationsresearch to investigate physiological and hydro-logical processes from whole plant to ecosystemscales. Stable isotopes of hydrogen and oxygenare routinely measured to identify plant sourcewater. This application relies on the assumptionthat no fractionation of oxygen and hydrogenisotopes in water occurs during uptake by roots.However, a large fraction of the water taken upthrough roots in halophytic and xerophyticplants transverses cell membranes in the endo-dermis before entering the root xylem. Passageof water through this symplastic pathway hasbeen hypothesized to cause fractionation leadingto a decrease in 2H of root xylem water relativeto that in the surrounding soil medium. Weexamined 16 woody halophytic and xerophyticplant species in controlled conditions for evi-dence of hydrogen isotope fractionation during
uptake at the root–soil interface. Isotopic separa-tion (�2H = �2Hsoil water ¡ �2Hxylem water) rangingfrom 3‰ to 9‰ was observed in 12 species. A sig-niWcant positive correlation between salinity tol-erance and the magnitude of �2H was observed.Water in whole stem segments, sapwood, androots had signiWcantly lower �2H values relativeto soil water in Prosopis velutina Woot., the spe-cies expressing the greatest �2H values among the16 species examined. Pressurized water Xowthrough intact root systems of Artemisia tridentataNutt. and Atriplex canescens (Pursh) Nutt. causedthe �2H values to decrease as Xow rate increased.This relationship was not observed in P. velutina.Destroying the plasma membranes of root cellsby excessive heat from boiling did not signiW-cantly alter the relationship between �2H ofexpressed water and Xow rate. In light of theseresults, care should be taken when using thestable isotope method to examine source-wateruse in halophytic and xerophytic species.
Keywords Hydrogen isotope fractionation · Roots · Xerophyte · Halophyte · Transpiration
Introduction
DiVerences in the physical properties of the stableisotopes of hydrogen and oxygen in water resultin isotopic separation or fractionation during
P. Z. Ellsworth (&)Department of Biology, University of Miami, 1301 Memorial Drive, Coral Gables, FL, 33124, USAe-mail: [email protected]
D. G. WilliamsDepartment of Renewable Resources, University of Wyoming, Laramie, USA
D. G. WilliamsDepartment of Botany, University of Wyoming, Laramie, USA
1 3

94 Plant Soil (2007) 291:93–107
physicochemical reactions (Chacko et al. 2001;Dansgaard 1964; McMillan 1985). These fraction-ation events provide important information aboutnumerous physical and biological processes. Pre-cipitation formed by the fractionating processesof evaporation and condensation producesisotopically distinguishable meteoric watersacross elevational gradients and between cold andwarm seasons (Craig 1961, Dansgaard 1964; Ken-dall and Caldwell 1998). Plant-available pools ofwater in the unsaturated soil and deeper ground-water therefore are often isotopically distinguish-able, establishing the means to use isotope ratiomeasurements to trace sources of water used byplants (Dawson 1996; Flanagan and Ehleringer1991; Williams and Ehleringer 2000). The isotopiccomposition of xylem water in the plant repre-sents a mixture of diVerent plant water sources.QuantiWcation of the proportion of water takenup from these sources is less complex providedthat there is no fractionation of water duringuptake at the root.
Several greenhouse and Weld-based studieshave veriWed that water is not altered isotopicallyduring uptake by roots (Dawson and Ehleringer1991; Dawson and Ehleringer 1993; Thorburnet al. 1993; Walker and Richardson 1991; Wash-burn and Smith 1934; Wershaw et al. 1966; Whiteet al. 1985; Zimmermann et al. 1967; Ziegler et al.1976). However, mangrove species (Laguncu-laria racemosa Gaert., Rhizophora mangle L., andAvicennia germinans L.) and the salt-tolerant(halophytic) plant, Conocarpus erecta L., do frac-tionate hydrogen isotopes during water uptake(Lin and Sternberg 1993). Fractionation by rootsin such species may lead to error in determinationof the proportional use of isotopically distinctwater sources (Lin and Sternberg 1992; Sternbergand Swart 1987).
The magnitude of hydrogen isotope fraction-ation during water uptake by roots in mangrovewas positively correlated with growth and transpi-ration rates (Lin and Sternberg 1993). Mangroveshave a highly developed Casparian strip on theradial cell walls of the root endodermis thatimpedes apoplastic movement of water, forcingwater to move across cell membranes (Atkinsonet al. 1966; Waisel et al. 1986). Lin and Sternberg(1993) hypothesized that the fractionation of
hydrogen isotopes in water occurred as watermoved symplastically through the endodermis.Several studies have observed fractionation ofisotopes of water across biological membranesand membranes composed of clay (Coplens andHanshaw 1973; Karan and Macey 1980; Phillipsand Bentley 1987). The energy required to disas-sociate individual water molecules from wateraggregates before passing through the plasmamembrane in the root endodermis is less for1H1H16O than for 1H2H16O (Chacko et al. 2001).Substitution of 18O for 16O in water causes only aslight relative diVerence in vibrational energybetween the isotopic species compared to the sub-stitution of 2H for 1H, such that fractionation ofisotopes in water during uptake by roots isexpected to be more apparent for hydrogen thanoxygen. If a large proportion of water enters theroot symplastically, root xylem water is likely tobe more depleted in 2H compared to the soilsolution. As the quantity of water entering theroots through the symplastic pathway increases,the magnitude of isotopic separation(�2H = �2Hsoil water ¡ �2Hxylem water) should alsoincrease. Under conditions where apoplasticmovement of water predominates, no fraction-ation is likely to occur because water aggregatescan enter the plant without the dissociation intosingle water molecules from water aggregates.
The hypothesis proposed by Lin and Sternberg(1993) for fractionation in mangroves predictsthat other species relying predominately on asymplastic pathway for water movement from soilto root xylem should fractionate hydrogen iso-topes of water. The degree of suberization andligniWcation of the root cell walls forming theCasparian strip determines the extent of symplas-tic water movement from the root cortex into theroot stele and xylem. Casparian strip develop-ment on the radial walls of the endodermis varieswidely among plant species, and is most extensivein halophytes (PoljakoV-Mayber 1975; Waisel1972). Glycophytic and mesophytic species havemuch less Casparian strip development, so sym-plastic movement of water is proportionally lessimportant in these species. The Casparian stripand suberization of epidermal and cortical cellwalls are also highly developed in woody xero-phytes (Nobel and Sanderson 1984; Passioura
1 3

Plant Soil (2007) 291:93–107 95
1981). Thus woody species that exhibit xerophyticand halophytic properties are likely to fractionatehydrogen isotopes during water uptake by roots.
The objectives of this study were to investigatethe extent of hydrogen isotope fractionationduring water uptake in woody plants of arid andsemi-arid regions and identify the mechanismresponsible for this fractionation. We hypothe-sized that symplastic water movement fraction-ates hydrogen isotopes in water, while apoplasticwater movement does not. We predicted thatsalinity tolerant desert xerophytes with a well-developed Casparian strip in the root endodermisshould fractionate hydrogen isotopes duringwater uptake to a greater extent than those spe-cies with lower salinity tolerance. We further pre-dicted that if isotopic fractionation during uptakefrom soil occurred, the isotopic composition ofroot xylem water should be similar to that ofshoots reXecting the location of fractionation atthe root–soil interface. Finally, the isotopic sepa-ration (�2Hsoil ¡ �2Hxylem water) should increase asXow rate through the root system increases.Destroying the plasma membranes with excessiveheat by boiling should remove the relationshipbetween �2H and Xow rate. Sixteen woody speciesfrom arid and semi-arid zones of the AmericanSouthwest varying in salinity tolerance wereexamined for hydrogen isotope fractionation.Prosopis velutina Woot., a widespread warm-des-ert shrub, was used in most of the experimentsdescribed here because prior studies (Ellsworth,unpublished results) suggested that it fractionatedhydrogen isotopes in water during uptake byroots. P. velutina was solely used in two green-house experiments to locate the tissue wherehydrogen isotope fractionation originated and todetermine if oxygen isotope fractionation alsooccurred. A root pressurization experiment wasconducted to identify the degree that Xow ratethrough the symplastic and apoplastic pathwaysaVected isotopic separation.
Methods
Several pot experiments were conducted to evalu-ate the magnitude, pattern, and extent of fraction-ation in desert shrubs and trees.
Extent of hydrogen isotope fractionation among woody xerophytes
Sixteen shrub and tree species native to semi-aridor arid areas of southwestern North Americawere evaluated for their capacity to fractionatehydrogen isotopes in water during uptake(Table 1). All but Ephedra viridis Coville, a gym-nosperm, were woody angiosperms. In addition,one mesophytic species, Lycopersicon esculen-tum L. (tomato), was included for comparison asthis species was shown not to fractionate eitherhydrogen or oxygen isotopes in an earlier study(Lin and Sternberg 1993). Salt-intolerant and tol-erant taxa were included (Table 1). Every indi-vidual of each species except Tamarix chinensisLuor and Prosopis spp. was 2 years old and nurs-ery grown in 4-L pots. The three Prosopis specieswere 3-year-old and nursery grown in 20-L pots.Cloned Tamarix stems from parent plants ofunknown age were propogated and grown in 20-L pots. The three Prosopis species were 1.5–2 mtall and had basal stem diameters of 1.5–3 cm.Larrea tridentata (Sessé & Moc. ex DC.) Covilleand Artemisia ludoviciana Nutt. were 50 cm talland had multiple basal stems with diametersranging from 0.4 to 0.7 cm. The stems were her-baceous except at the base of the main stems. Allother species were 50–70 cm tall and single-stemmed with basal stem diameters of 1–1.5 cm.The tomato plants were 40 cm tall and had basalstem diameters of 1 cm. The plants were grown ina greenhouse in full sun with a 16 h photoperiod.The mean daytime and nighttime temperatureswere 25 and 21°C, respectively, and daytime rela-tive humidity was 19%.
The original potting soil used in the nurserywhere the plants were purchased was carefullyremoved from the root system and the plantswere repotted in with sandy loam soil with 0.8%organic matter. The pH of the soil was 7.8, andthe electrical conductivity was 0.69 dS m¡1. A 20–20–20 NPK fertilizer was applied weekly in theirrigation water to ensure adequate nutrientswere available for plant growth. Three weeksprior to the collection of stems and soils, the potswere completely covered with plastic to preventevaporation from the soil surface. At this time thepots were thoroughly Xushed with isotopically
1 3

96 Plant Soil (2007) 291:93–107
uniform water (�2H = ¡131‰) and watered regu-larly with the same water.
To minimize contamination of transpiration-derived, 2H and 18O-enriched water in the stems,all stems were collected distal from leaves andherbaceous material. Unlike all other species,stems collected from L. tridentata and A. ludovici-ana were from the base of the main stems andwere close to transpiring tissue. Numerous greenleaves were removed from the collected twig seg-ments of Artemisia tridentata Nutt. The rootcrown of the tomato was collected to avoid tissuesaVected by evaporative enrichment of 2H and 18O
(Thorburn and Mensforth 1993). A single soilsample was collected from each pot. Each specieswas represented by 3–6 individuals (Table 1). Thecollection order of the individuals of each speciesand the species was randomized.
Magnitude of isotopic separation(�2H = �2Hsoil water ¡ �2Hxylem water) was calculatedfor all the species and correlated with salinity tol-erance. Salinity tolerance (dS m¡1) was deWnedquantitatively as the electrical conductivity thatbegins to adversely aVect plant growth. Values ofsalinity tolerance were obtained from the litera-ture for 12 of the 16 species. Values of salinity tol-erance for Wve species (Artemisia luduviciana,Acacia greggii Gray, A. constricta Benth., Olneyatesota Gray, and Flourensia cernua DC.) were notavailable.
Variation in �2H and �18O among tissues and organs in Prosopis velutina
Prosopis velutina was identiWed as a species thatfractionated hydrogen isotopes in water and wasused to investigate the isotopic variation amongdiVerent plant tissues and organs. Five P. velu-tina plants were nursery grown in 20-L pots oforganic soil, consisting of wood chips, decom-posed leaf litter, and a small amount of sand.Plants were 3 years old and had a basal stemdiameter of 1–2 cm and a height of 1 m. Theplants were grown outdoors with daily maximumtemperatures of 38°C, minimum nighttime tem-peratures of 19°C, and 15% minimum daytimerelative humidity. The pots were completely cov-ered and sealed with black plastic and wateredwith isotopically uniform water (�2H = ¡65‰,�18O = ¡9.4‰) daily for 10 days prior to samplecollection. Covering the pots 10 days prior to col-lection prevented evaporation, and any isotopi-cally heavy water was Xushed from the pot withsubsequent irrigations. Whole stem segments(including bark) and soils were collected from1100 to 1300 h when the trees were likely tran-spiring at maximum rates. Leaves, young stems,old stems, and soil from the top and bottom ofeach pot were collected in vials and kept frozenuntil the water was extracted from the samplesby cryogenic vacuum distillation. The young,green stems were probably not completely
Table 1 Species examined for fractionation of hydrogenisotopes in water during uptake by roots, the number ofindividuals (n) examined for each species, and the salinitytolerance of each species reported as electrical conductivity(EC: dS m¡1)
The EC values represent the threshold level after whichplant growth is adversely aVected. EC values were obtainedfrom published studies.a Feucht (2001)b Forti (1986)c El-Ghonemy et al. (1980)d Maas (1985)e Jackson et al. (1990). The values used are threshold val-ues when the plant is signiWcantly damaged by the salinityconcentrationf Houerou (1986)
ScientiWc name n EC (dS m¡1)
Acacia constricta Benth. 4 N/AAcacia greggii Gray 3 N/AArtemisia ludoviciana Nutt. 6 N/AArtemisia tridentata Nutt. 4 6a
Atriplex canescens (Pursh) Nutt. 6 20b
Chrysothamnus nauseosus (Pallas ex Pursh) Britt.
6 6a
Ephedra viridis Coville 4 10a
Flourensia cernua DC. 6 N/AKrascheninnikovia lanata
(Pursh) A.D.J. Meeuse & Smit 3 10a
Larrea tridentata (Sessé & Moc. ex DC.) Coville
5 1.84c
Lycopersicon esculentum L. 4 2.5d
Olneya tesota Gray 6 N/AProsopis glandulosa Torr. 3 8e
Prosopis pubescens Benth. 6 9.4e
Prosopis velutina Woot. 5 15b
Simmondsia chinensis (Link) C. Schneider
4 10b
Tamarix chinensis Luor 4 25f
Total 79
1 3

Plant Soil (2007) 291:93–107 97
suberized and likely subject to evaporativeenrichment of 2H and 18O.
An additional experiment was conducted toverify that fractionation occurred in roots ratherthan down stream in the stem and to determinethe contribution of evaporatively enriched waterin the bark to xylem water of sapwood. Five 2-year-old P. velutina plants were grown in 4-L potsconsisting of a mixture of Wne organic matter,wood chips, and sand. The plants had a basaldiameter of approximately 1 cm and height of 50–75 cm. Plants were soaked in a bucket of water for1 h to Xush existing water from the soil 5 daysbefore the tissues were collected for isotopicanalysis. Plants were then removed from thewater bath, and the soil medium was Xushed withan additional 8 L of water. Water used to irrigateand Xush the root system of the plants was isoto-pically uniform (�2H = ¡134‰, �18O = ¡17.0‰).The pots were wrapped and sealed with plastic toprevent evaporation from the soil surface. Afterthe pots were allowed to drain, the holes in thebottom of the pots were sealed. The plants wereXushed again with water of the same isotopiccomposition three more times in 5 days prior toharvesting tissue. The plants were kept in the sunthroughout the day in a greenhouse at 31°Cmaximum daytime and 19°C minimum nighttimetemperatures. Minimum daytime relative humid-ity was 17%.
Whole stem segments containing both sap-wood and bark, sapwood only, bark only, androots were collected separately from the Wveplants. Tissue samples were collected from fourplants between 0930 and 1730 h. Tissues from theWfth plant were collected at 400 h when the plantwas expected to be transpiring minimally. Barkwas quickly removed from the sapwood, and eachtissue component was placed in separate vials forwater extraction and isotopic analysis. Anotherstem was collected intact with bark and sapwoodtogether. The taproot was quickly cleaned, blot-ted dry of any external soil and moisture andplaced in a vial. A single soil sample was collectedfrom each pot. The plants were small, so all thestems were in close proximity to leaves and youngherbaceous stems. None of the collected tissueswas from large stems with extensive bark devel-opment. The isotopic composition of the leaf
water was calculated using the Craig and Gordonmodel, which was modiWed to calculate theisotopic composition of leaf water (Yakir andSternberg 2000). The magnitude of isotopicseparation (�2H = �2Hsoil water ¡ �2Hxylem water and�18O = �18Osoil water ¡ �18Oxylem water) was calcu-lated for all plant parts.
Hydrogen isotope fractionation in live and heat-killed intact root systems
Three individuals of three species (P. velutina, A.tridentata, and Atriplex canescens) were used in aroot pressurization experiment to examine therelationship between water Xow rate and �2H ofthe expressed water for live and heat-killed rootsystems. The role of symplastic water movementin hydrogen isotope fractionation was evaluatedby immersing the root systems in boiling water for35 min to destroy the root cell membranes andcomparing the �2H of expressed water to that inlive root systems. Isotopic fractionation shouldnot be apparent in the heat-killed root systems,which allows only for apoplastic water movement.Whole root systems were placed in a Scholander-type pressure chamber Wlled with water. Flow rateof water exuding from the stem was maintainedby regulation of pressure applied to the chamber.
Three weeks prior to the pressurization experi-ment, each pot was covered with plastic to pre-vent evaporation as was done in the previousexperiments. The root system was Xushed severaltimes then soaked for 2 h with water isotopicallyidentical with the water used in the pressurizationchamber (�2H = ¡132‰), ensuring isotopic uni-formity within the pressurization chamber. AfterXushing, the root system was completely sub-merged in water in the pressure chamber withonly the cut stem protruding out of the top of thelid. Water was collected for at least 1 h at eachpressure, so that at least 3 ml of water exudedfrom root system to ensure that the water col-lected from the root system for isotopic analysiswas water that entered the root system under thatpressure. Water exuding from the stem wascollected in pre-weighed micro-centrifuge tubesWlled with oven-dried glass wool. The tubeswere weighed again after water collection to cal-culate the volume of water collected. Flow rate
1 3

98 Plant Soil (2007) 291:93–107
measurements (ml h¡1) were made by collectingthe water exuding from the stem in 1 min intervals.Only water collected in the last 5 min at each pres-sure was analyzed for isotopic composition. Alltreatments were placed under a series of pressuresin the following order: 0.3, 0.5, 0.7, 0.9, 0.7, 0.3 MPa.Each root system was placed in boiling water for35 min after the Wrst pressurization on the live rootsystem. The same pressurization method was fol-lowed on the heat-killed root system.
A composite sample consisting of Wve consecu-tive 1-min water samples was used for isotopicanalysis. The water from the micro-centrifugetubes with glass wool was distilled using a cryo-genic vacuum line. Water from the pressurizationchamber also was collected to measure the initialisotopic composition of the water in the pressuri-zation chamber.
To test the extent that experimental error hadon the isotopic composition of the samples duringcollection and distillation, a mock pressurizationexperiment was conducted on samples of knownisotopic composition. The mock pressurizationexperiment was conducted and these watersamples were distilled and analyzed for isotopiccomposition in the same manner as above. Theisotopic composition of the water used in themock pressurization experiments was comparedto the original �2H value of the water used in thisexperiment to calculate the eVect of evaporationand experimental error.
�2H and �18O analyses
Stem, root, and soil samples were stored frozenuntil the water was quantitatively extracted bycryogenic vacuum distillation (Ehleringer andOsmond 1989). Extraction eYciency for all sam-ples, deWned as the proportion of water extractedfrom the sample, was greater than 97%. All watersamples collected were prepared for hydrogenisotope analysis by reacting 2 �l of extracted waterwith 100 mg of zinc at 500°C for 1 h in sealed Pyrextubes (method modiWed from Coleman et al. 1982).The �2H value of the resulting H2 gas was analyzedon a dual-inlet stable isotope ratio mass spectrome-ter (Optima, Micromass UK Ltd., Manchester,UK) at the University of Wyoming Stable IsotopeFacility (UWSIF), Department of Renewable
Resources, University of Wyoming, Laramie, WY,USA, or at the University of Arizona Laboratoryof Isotope Geochemistry, using a dual-inlet stableisotope ratio mass spectrometer (Delta S, FinniganMAT, San Jose, CA, USA) and an on-line chro-mium reduction furnace (HDevice, FinniganMAT, San Jose, CA, USA; Nelson and Dettman2001). Precision of the analysis using chromiumreduction calculated from lab internal workingstandards also was 0.8‰. Oxygen isotopic composi-tion of all samples was measured at UWSIF byCO2 equilibration using a Gas Bench II connectedto a DeltaPlus XP mass spectrometer (ThermoFinn-igan, San Jose, CA, USA) or at the University ofArizona Laboratory of Isotope Geochemistry byCO2 equilibration (Epstein and Mayeda 1953). Theprecision of the analysis for �18O was 0.08‰ atUWSIF and 0.15‰ at the University of ArizonaLaboratory of Isotope Geochemistry. Standard labwaters were calibrated against the internationalstandard V-SMOW and SLAP (InternationalAtomic Energy Agency (IAEA) 1995), whichwere used in linear corrections of values obtainedfrom the mass spectrometer. Reported values arein per mil (‰) relative to V-SMOW.
Paired student’s t-tests were used to determine ifthe � values of stem xylem water and soil weresigniWcantly diVerent (P < 0.05). Paired student’st-tests in the pot experiments and linear regressionsin the pressurization experiment were calculatedusing SigmaPlot for Windows (Version 8.02, SPSSInc. 2002). The slopes and y-intercepts of all linearregressions were compared by covariance analysisof the two groups where y was the dependent vari-able and x the co-variable (Armitage 1980).
Results
Extent of hydrogen isotope fractionation among woody xerophytes
Isotopic separation (�2H = �2Hsoil ¡ �2Hplant) washighest in P. velutina (9 § 1‰) and lowest inChrysothamnus nauseosus (Pallas ex Pursh) Britt.(3 § 1‰; Fig. 1). Isotopic separation for Wvespecies (L. esculentum, A. tridentata, Prosopispubescens Benth., E. viridis, and L. tridentata)was not signiWcantly diVerent than zero.
1 3
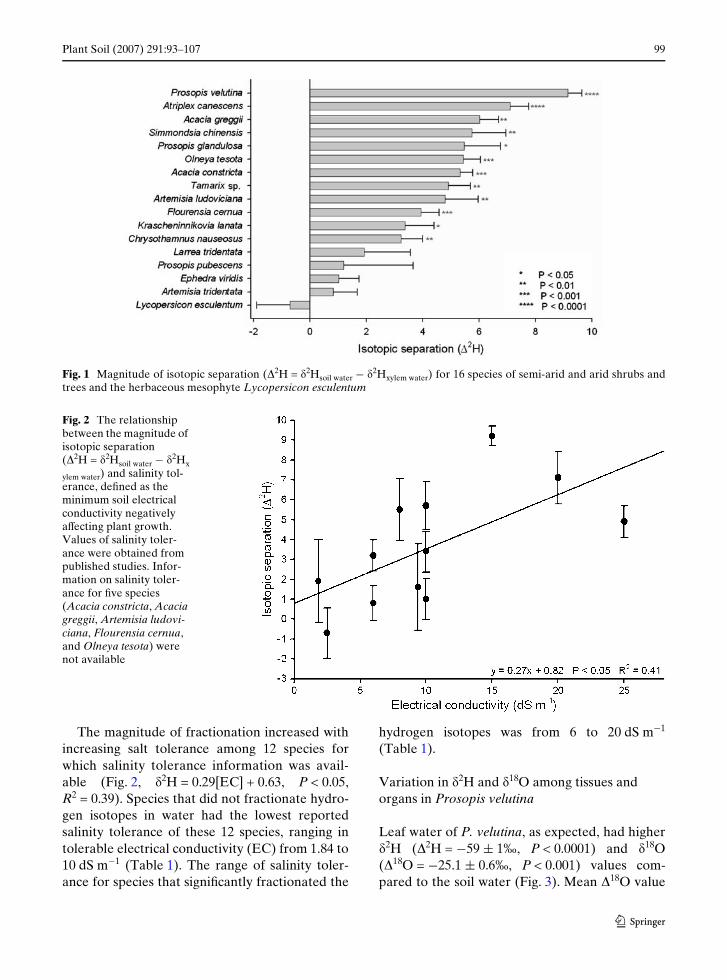
Plant Soil (2007) 291:93–107 99
The magnitude of fractionation increased withincreasing salt tolerance among 12 species forwhich salinity tolerance information was avail-able (Fig. 2, �2H = 0.29[EC] + 0.63, P < 0.05,R2 = 0.39). Species that did not fractionate hydro-gen isotopes in water had the lowest reportedsalinity tolerance of these 12 species, ranging intolerable electrical conductivity (EC) from 1.84 to10 dS m¡1 (Table 1). The range of salinity toler-ance for species that signiWcantly fractionated the
hydrogen isotopes was from 6 to 20 dS m¡1
(Table 1).
Variation in �2H and �18O among tissues and organs in Prosopis velutina
Leaf water of P. velutina, as expected, had higher�2H (�2H = ¡59 § 1‰, P < 0.0001) and �18O(�18O = ¡25.1 § 0.6‰, P < 0.001) values com-pared to the soil water (Fig. 3). Mean �18O value
Fig. 1 Magnitude of isotopic separation (�2H = �2Hsoil water ¡ �2Hxylem water) for 16 species of semi-arid and arid shrubs andtrees and the herbaceous mesophyte Lycopersicon esculentum
Fig. 2 The relationship between the magnitude of isotopic separation (�2H = �2Hsoil water ¡ �2Hx
ylem water) and salinity tol-erance, deWned as the minimum soil electrical conductivity negatively aVecting plant growth. Values of salinity toler-ance were obtained from published studies. Infor-mation on salinity toler-ance for Wve species (Acacia constricta, Acacia greggii, Artemisia ludovi-ciana, Flourensia cernua, and Olneya tesota) were not available
1 3

100 Plant Soil (2007) 291:93–107
was ¡0.3 § 0.3‰ for woody, green (young) stems(P = 0.36) and 0.4 § 0.2‰ for older stems(P = 0.07). Mean �2H value of water in young andold stems was 5 § 1‰ (P < 0.01) and 8 § 1‰(P < 0.001), respectively. The �2H and �18Ovalues of xylem water of the young stems wereintermediate between that of the xylem water ofthe old stems and that of the leaves (Fig. 3). Thetwo soil samples collected from each pot weresimilar isotopically.
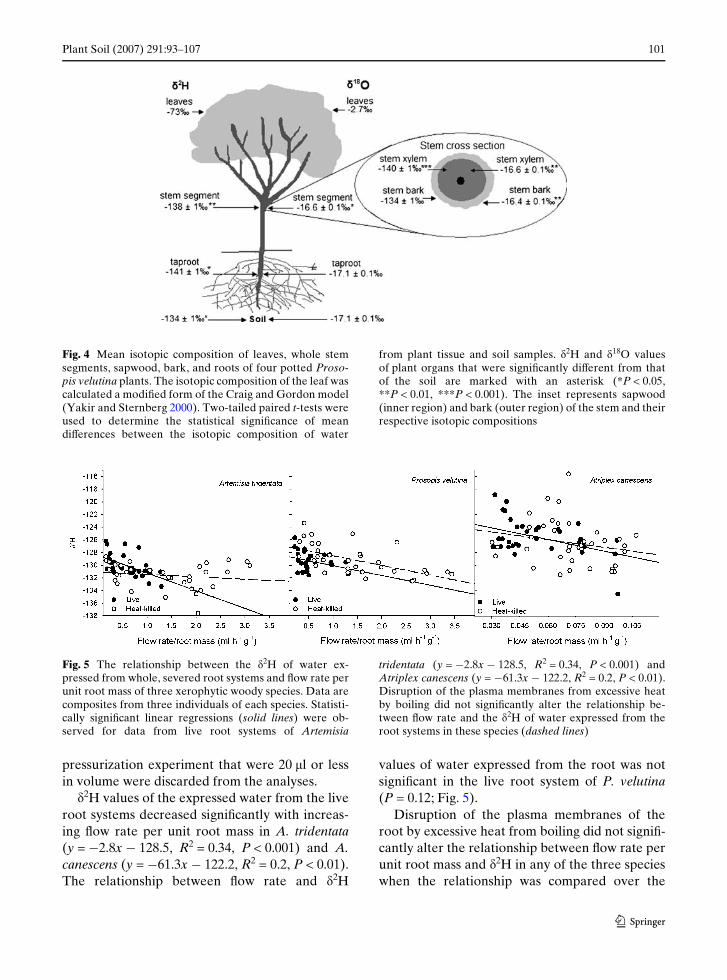
The pattern of variation among plant tissuesand organs of P. velutina was similar for hydrogenand oxygen isotopes, with the water in bark hav-ing the highest �2H and �18O values and water inroots having the lowest �2H and �18O values rela-tive to soil (Fig. 4). The �2H value of water in barkwas higher by 6 § 1‰ (P < 0.01) than that of thesapwood. However, the �2H value of water inbark was not signiWcantly higher than that ofsource water (P = 0.82), while the �18O value ofwater in bark was higher by 0.7 § 0.1‰ (P < 0.01).The �2H value of water in whole stem segmentswas only slightly higher than that of sapwood(2 § 1‰, P = 0.06). Although the bark was thin, itwas 33 to 43% of the water in the entire stem, con-tributing substantially to the isotopic compositionof the stem. The plant organ exhibiting the largest
positive isotopic separation in hydrogen isotopevalues compared to soil water was the root(�2H = 7 § 1‰, P < 0.05), but the mean �18Ovalue of water in roots was identical to that of soil(i.e., �18O = 0). Water in whole stem segmentsand sapwood had signiWcantly lower �2H valuesthan the soil source water (P < 0.01 and P < 0.001,respectively). The �18O values of water in wholestem segments and sapwood collected separatelywere higher than that of the soil water, having�18O values of ¡0.4 § 0.1‰ (P < 0.05) and¡0.4 § 0.1‰ (P < 0.01), respectively.
Isotopic separation in whole stem segments,sapwood, and root of the plant collected at pre-dawn was less than that measured in the sameorgans and tissues of plants collected at midday(�2H at predawn = 3, 2, and 5, respectively).Bark, collected at predawn, had a higher �2Hvalue than bark collected from plants at midday(�2H = ¡6). For plants collected at predawn andmidday, bark had higher �2H and �18O valuesthan all other plant parts except leaves.
Isotopic variation in the whole stem segments,sapwood, and bark was a mixture of xylem waterand phloem enriched in 2H and 18O by cuticularevaporation in the stems and backXow from theleaves. The �2H values of water in whole stemsegments, sapwood, and bark were intermediatebetween the �2H values of the roots and the calcu-lated �2H value of the leaf (Fig. 4). The �18O val-ues of water in roots, whole stem segments,sapwood, and bark were intermediate betweenthe �18O values of the soil water and the calcu-lated �18O value of the leaf (Fig. 4).
Hydrogen isotope fractionation in live and heat-killed intact root systems
Evaporation during sample collection and frac-tionation during sample distillation and isotopicanalysis did not have any signiWcant eVect on theisotopic composition of the water samples. Themean �2H value of Wve 20 �l samples of waterused to test the eVect of sample collection andprocessing was only 2 § 1‰ diVerent than controlwater not subjected to the collection and extrac-tion processes, and that of samples greater than20 �l in volume was 1 § 1‰ diVerent than controlsamples. Water samples collected during the
Fig. 3 Mean isotopic composition of water extracted fromwoody, green stems (young), woody stems with well-devel-oped bark (old), leaves and soil from Wve potted individualsof P. velutina. Two-tailed paired t-tests were used to deter-mine the statistical signiWcance of mean diVerences be-tween the isotopic composition of water from stems andsoil samples. �2H and �18O values of plant organs that weresigniWcantly diVerent from that of the soil are marked withan asterisk (*P < 0.01, **P < 0.001, ***P < 0.0001). Mean�18O values of water extracted from old stems were slightlymore positive than that of soil water (P = 0.07)
1 3
Plant Soil (2007) 291:93–107 101
pressurization experiment that were 20 �l or lessin volume were discarded from the analyses.
�2H values of the expressed water from the liveroot systems decreased signiWcantly with increas-ing Xow rate per unit root mass in A. tridentata(y = ¡2.8x ¡ 128.5, R2 = 0.34, P < 0.001) and A.canescens (y = ¡61.3x ¡ 122.2, R2 = 0.2, P < 0.01).The relationship between Xow rate and �2H
values of water expressed from the root was notsigniWcant in the live root system of P. velutina(P = 0.12; Fig. 5).
Disruption of the plasma membranes of theroot by excessive heat from boiling did not signiW-cantly alter the relationship between Xow rate perunit root mass and �2H in any of the three specieswhen the relationship was compared over the
Fig. 4 Mean isotopic composition of leaves, whole stemsegments, sapwood, bark, and roots of four potted Proso-pis velutina plants. The isotopic composition of the leaf wascalculated a modiWed form of the Craig and Gordon model(Yakir and Sternberg 2000). Two-tailed paired t-tests wereused to determine the statistical signiWcance of meandiVerences between the isotopic composition of water
from plant tissue and soil samples. �2H and �18O valuesof plant organs that were signiWcantly diVerent from thatof the soil are marked with an asterisk (*P < 0.05,**P < 0.01, ***P < 0.001). The inset represents sapwood(inner region) and bark (outer region) of the stem and theirrespective isotopic compositions
Fig. 5 The relationship between the �2H of water ex-pressed from whole, severed root systems and Xow rate perunit root mass of three xerophytic woody species. Data arecomposites from three individuals of each species. Statisti-cally signiWcant linear regressions (solid lines) were ob-served for data from live root systems of Artemisia
tridentata (y = ¡2.8x ¡ 128.5, R2 = 0.34, P < 0.001) andAtriplex canescens (y = ¡61.3x ¡ 122.2, R2 = 0.2, P < 0.01).Disruption of the plasma membranes from excessive heatby boiling did not signiWcantly alter the relationship be-tween Xow rate and the �2H of water expressed from theroot systems in these species (dashed lines)
1 3

102 Plant Soil (2007) 291:93–107
same range of Xow rates in both live and heat-killed root systems (Fig. 5). The relationshipbetween Xow rate and �2H was signiWcantly nega-tive in the heat-killed root system of A. tridentata(y = ¡2.1x ¡ 129.9, R2 = 0.22, P < 0.05) and inP. velutina (y = ¡2.9x ¡ 125.8, R2 = 0.24, P < 0.05),but neither relationship was signiWcantly diVerentthan the relationship produced in the correspond-ing live root systems. The relationship betweenXow rate per unit root mass and �2H was not sig-niWcant in heat-treated root systems of A. canes-cens, nor was the relationship signiWcantlydiVerent from the relationship produced from thecorresponding live root system.
Discussion
Extent of hydrogen isotope fractionation among woody xerophytes
All 12 species that fractionated hydrogen isotopesin water during uptake in this study are adaptedto saline or xeric environments. Such speciesoften develop highly suberized and ligniWed cellwalls in their roots, especially the Casparian stripon the endodermis cell walls, to prevent waterloss to dry or highly saline soils and to excludesalts from entering the root (Craig et al. 1990;Nobel and Sanderson 1984; PoljakoV-Mayber1975; Reinoso et al. 2004; Serrato Valenti et al.1991; Serrato Valenti et al. 1992; Sharma 1982;Vartanian 1981). Both salinity and droughttolerance result in part from suberization of theendodermis and exodermis, which proportionallyincreases symplastic movement of water in theroot (Feder and Taube 1952; Passioura 1981;Stewart and Friedman 1975). Though some plantssuch as A. canescens and Tamarix chinensis usesalt secretion from salt glands and storage in tis-sues to remove excess salt, the adaptive featuresof salt exclusion such as suberization of the rootendodermis and exodermis are still present inmost halophytic species to reduce salt concentra-tions entering the root (Scholander et al. 1966;Shannon 1997). The water movement through thesymplastic pathway potentially fractionateshydrogen isotopes in water (Karan and Macey1980).
With 12 of the 16 desert species examined inthis experiment exhibiting hydrogen isotope frac-tionation, halophytic and xerophytic species mustbe examined for fractionation of hydrogen iso-topes in water during uptake by roots beforeplant water source studies that use isotopic com-position of xylem water are conducted. Only asmall quantity of desert species were included inthis study, yet numerous species were found tofractionate hydrogen isotopes in water, having�2H values from 3 § 1‰ in Chrysothamnusnauseosus to 9 § 1‰ in P. velutina (Fig. 1). Otherhalophytic and xerophytic species, yet to beexamined, are likely also to fractionate hydrogenisotopes in water during uptake from soil, poten-tially leading to errors in water source calcula-tions using linear mixing models. For example, ifthe diVerences in the �2H values of the plantwater sources are small and hydrogen isotopefractionation in water during uptake by rootsresults in a large �2H values, such as 9‰ inP. velutina, miscalculations in water source usewould be quite large and would lead to misinter-pretations of plant water-source use and the rolethe species plays in hydrologic processes at theecosystem or larger scales.
Since anatomical features of roots conferringsalinity tolerance cause symplastic movement ofwater in the root, salinity tolerance was used as anindicator of the predominance of the symplasticpathway in water movement (Fig. 2). Species withhigh salinity tolerance tended to fractionatehydrogen isotopes more so than those with lim-ited salinity tolerance. The relationship betweensalt tolerance and apparent fractionationreported here provides more evidence that thehalophytic properties of salt tolerant plants, suchas the reliance on the symplastic pathway forwater movement into the root, contributes to thefractionation of hydrogen isotopes in water.
Another mechanism potentially accountingfor the lower observed �2H values of xylemwater than the source water is hydrogen ionexchange between water entering the root andapoplastic H+ pumped into the apoplast andfrom organic acids secreted from the root. Frac-tionation of the apoplastic H+ is high (670‰;Luo et al. 1991). Assuming the apoplastic pH is5.5 and incoming water had an extremely high
1 3

Plant Soil (2007) 291:93–107 103
pH of 9.0, the quantity of H+ needed to changethe pH from 5.5 to 9.0 would cause a decrease ofthe hydrogen isotope ratio of apoplastic waterby only 0.002‰. The decrease in 2H of waterentering the root by exchange with apoplastic H+
apparently is not suYcient to produce the low�2H values of stem xylem water measured in thisexperiment.
Variation in �2H and �18O among tissues and organs in Prosopis velutina
Low �2H values in xylem water in young and oldstem classes relative to source water likely wasthe result of an ultraWltration process occurring aswater entered the root through a symplastic path-way. Low �2H values in xylem water in young andold stems was not a result of cuticular evaporationor isotopic contamination from leaf waterbecause these processes would lead to higherrather than lower �2H values in xylem water rela-tive to source water. Isotopic variation in the soildid not account for the isotopic variationexpressed in the stems as the two soil samplescollected from each pot showed no signiWcantdiVerence in either 2H or 18O. The old stems hadmarginally lower �2H values (P = 0.07) thansource water. One would expect some depletionof 18O, albeit small, as water enters the plantaccording to our hypothesis of isotopic fraction-ation occurring as water passes through theplasma membranes in the roots. However,because of the relative mass diVerences, it isexpected that the depletion of the heavy isotopein stem xylem water compared to that of the soilwas principally found for hydrogen (Fig. 3).
Isotopic composition of young stems, wholestem segments, bark, and sapwood in both experi-ments represented a mixture of 2H-enrichedphloem water from the leaves and 2H-depletedroot xylem water. Leaf water, enriched in 2H and18O by evaporation during transpiration is thelikely source for the enrichment in 18O and 2H ofwater in young stems with respect that in the old,suberized stems (Fig. 3). Isotopically heavy leafwater is also the cause of 18O and 2H enrichmentin water of bark, whole stem segments, andsapwood relative to root xylem water (Fig. 4,Dawson and Ehleringer 1993). On the other
hand, root xylem water depleted in 2H, caused�2H values in young and old stems, whole stemsegments, sapwood, and roots to be lower thanthe source water for both experiments. The older,woody stems of these P. velutina plants had fullydeveloped protective bark and were likely suber-ized, so that evaporative enrichment in 2H and18O was minimal. Mature stems with developedbark generally have a lower proportion of theirwater in the phloem, so the eVects of contamina-tion from isotopically heavy phloem water aresmaller than in young stems (Dawson and Ehle-ringer 1993; Thorburn et al. 1993). As a result,evaporation from the young stems and backXowin the phloem from the leaves caused greaterenrichment in 2H than in the older stems (Fig. 3).Also suYcient exchange occurred between isoto-pically heavy phloem water and xylem water tocause the xylem water in the sapwood to beenriched in 18O with respect to the soil water andto be enriched in 2H with respect to the rootxylem water (Fig. 4).
Fractionation of hydrogen isotopes duringuptake in P. velutina occurred in the root. Themost negative �2H values of water observed inpotted P. velutina plants was from root xylemwater samples, indicating that the root–soil inter-face was the likely location of fractionation(Fig. 4). However, the �18O of xylem water inroots was not signiWcantly lower than that ofsource water, indicating minimal fractionation ofoxygen isotopes in water during uptake. Thegenus Prosopis is reported to exclude salt in rootsto tolerate high salinity (Reinoso et al. 2004).Prosopis species develop ligniWed and suberizedroot endodermis and exodermis cell walls and aCasparian strip that develops faster and closer tothe root tip than in salt-intolerant species, forcingwater to move via the symplastic pathway (Rein-oso et al. 2004; Serrato Valenti et al. 1991; SerratoValenti et al. 1992).
The magnitude of isotopic fractionation of 2Hin water during uptake by roots in P. velutina var-ied between midday and predawn periods. Atpredawn, during a period with minimal transpira-tion, a low �2H value was observed, but did notreach isotopic equilibrium with the soil. Bark col-lected at predawn was enriched in 2H with respectto the soil water. The Wrst possibility suggests that
1 3

104 Plant Soil (2007) 291:93–107
when transpiration dropped to low levels at pre-dawn, isotopically light xylem water moving upfrom the root contributed less to the �2H and�18O values of the water in the plant tissues, mak-ing the overall contribution of isotopically heavywater in the phloem proportionally larger, result-ing in higher �18O and �2H values. The secondpossibility, as hypothesized Lin and Sternberg(1993), is that the fractionating process of ultraWl-tration can occurs as water enters the root fromthe rhizosphere and as water leaves the root. Atlow transpiration rates, the fractionation event ofwater entering the root would be only slightlyhigher than the fractionation event of waterleaking from the root, therefore the magnitude ofisotopic separation would be negligible.
Hydrogen isotope fractionation in live and heat-killed intact root systems
The hypothesis that water movement through thesymplastic pathway fractionates hydrogenisotopes in water during uptake by roots whensymplastic water movement predominates isconsistent with observations that the �2H valuesof the water expressed from the root systemdecreased with increasing Xow rate in A. canes-cens and A. tridentata. A. tridentata apparently didnot fractionate hydrogen isotopes in water in thesurvey study using 16 woody xerophyte and halo-phyte taxa reported above. Stems collected fromA. tridentata in the survey study were attached toseveral green leaves. It is likely that the xylemwater was enriched in 2H and 18O by evaporationand back Xow through the xylem as whole stemsegments were sampled.
The relationship between Xow rate and �2H ofexpressed xylem water was not signiWcant in liveroot systems of P. velutina, although hydrogenisotope fractionation was observed in the experi-ments described above. The range of pressuresused in this experiment to produce a gradient inXow rate resulted in only a small range of Xowrates of water expressed from the live root systemof P. velutina. A larger range of Xow rates mayhave resulted in greater variation in �2H values ofthe expressed water from the live root system anda signiWcant relationship between Xow rate and�2H of the expressed water from the root.
Destroying the plasma membranes by exces-sive heat did not alter the relationship betweenXow rate and �2H of the water expressed from theroot. Flow rates of water moving through theheat-killed root systems of A. tridentata andP. velutina increased with respect to the live rootsystems, showing that resistance to Xow decreasedafter the root system was immersed in boilingwater. However, this increase in Xow rate did notalter the relationship between Xow rate and �2H,contrary to our prediction. Immersing the rootsystems of A. canescens in boiling water to disruptthe root plasma membranes did not increase Xowrates or alter the relationship between �2H ofwater expressed from the root system and Xowrate. Possibly the plasma membranes were notfully destroyed by excessive heat.
Evaporation during collection of watersamples and fractionation during distillation andisotopic analysis of water samples did not haveany signiWcant eVect on the isotopic compositionof the samples. Therefore these results were notthe result of fractionation due to experimentaldesign. Water collected from the pressurizationchamber before the pressurization experimentwas conducted did not vary isotopically fromwater samples collected from the pressurizationexperiment after the experiment was concluded;therefore, isotopic heterogeneity was not respon-sible for the isotopic variation of water samplescollected from the root system during the pres-surization experiment.
DiVerences between positive pressure and ten-sion could explain the results that �2H values ofthe water expressed from root systems werehigher at lower Xow rates with respect to thesource water in the pressurization chamber. Asthe Xow rate increased, �2H values became moresimilar to the �2H value of the source water(�2H = ¡132‰). The positive pressure needed topush water through the xylem conduits is of thesame magnitude but opposite sign as the tensionrequired to pull water through the xylem (Tyreeand Sperry 1989). Though the response of pres-sure to Xow is the same if the pressure is positiveor negative, the eVect that positive pressure hason the ultraWltration process of water passingthrough a membrane is unknown. This higher �2Hvalues with respect to the source water at lower
1 3

Plant Soil (2007) 291:93–107 105
Xow rates may be due to the positive pressureinstead of tension that was exerted on theroot system. Positive pressure might have elimi-nated the fractionating process of ultraWltration.If water leaking from the root still underwent thefractionating process of ultraWltration, the eVectof water entering and leaving the root wouldcause water inside the root to be enriched in 2Hwith respect to soil water. As the net quantity ofwater entering the plant increases, the propor-tional importance of water leaking from theroot would decrease. As a result, the �2H valuesof the water entering the root would approachthat of the source water. In this case theexpected results would be similar to the resultsobtained in this experiment where the waterexpressed from the severed root system wouldhave lower �2H values than the source water atlow Xow rates and similar to the source water athigher Xow rates.
Conclusions
Although this study did not fully resolve the mech-anism responsible for fractionation, it does providemore evidence to support the hypothesis thatsymplastic movement of water during uptakefractionates hydrogen isotopes. Halophytic andxerophytic species rely more heavily on thesymplastic water transport pathway in the rootsthan the apoplastic pathway due to the high degreeof endodermal and exodermal cell wall develop-ment associated with salt and drought tolerance,and consequently fractionate hydrogen isotopes inwater. Twelve of the 16 xerophytic and halophyticspecies examined in this study exhibited fraction-ation of hydrogen isotopes in water during uptakeby roots. The magnitude of isotopic separation waspositively correlated with salinity tolerance, show-ing that adaptations to salinity tolerance result inhydrogen isotope fractionation. Roots, having thelargest �2H values, were the apparent location offractionation, and the isotopic composition ofwhole stem segments was a mixture of 2H-enrichedleaf water transported through the phloem and 2H-depleted xylem water from the root. The �2H val-ues of xylem water did not accurately reXect the�2H values of soil water. However, �18O values of
xylem water of the same species that fractionatedhydrogen isotopes in water did accurately reXectthe �18O values of the soil water. Also, as pre-dicted, the �2H values of water expressed from liveroot systems decreased as Xow rate through theroots increased. However, disrupting root plasmamembranes by excessive heat did not have a con-sistent eVect on the relationship between Xow rateand �2H among three species examined.
Acknowledgements We thank Enrico Yepez, Victor Re-sco, Rico Gazal, and Ayme Ahrens for their help with thegreenhouse experiments. We also thank Ann Hild andBrent Ewers for their help during the manuscriptpreparation.
References
Armitage P (1980) Statistical methods in medical re-search. Blackwell ScientiWc Publications, OxfordUK, pp 279–301
Atkinson MR, Findlay GP, Hope AB, Pitman MG, SaddlerHDW, West KR (1966) Salt regulation in the man-groves Rhizophora mucronata Lam. and Aegialitisannulata R.Br. Aust J Biol Sci 20:589–599
Chacko T, Cole DR, Horita J (2001) Equilibrium oxygen,hydrogen, and carbon isotope fractionation factorsapplicable to geologic systems. In: Valley JW, ColeDR (eds) Reviews in mineralogy and geochemistry,vol 43. Stable isotope geochemistry. MineralogicalSociety of America. Washington, DC pp 1–82
Coleman ML, Shepard TJ, Durham JJ, Rouse JE, MooreGR (1982) Reduction of water with zinc for hydrogenisotope analysis. Anal Chem 54:993–995
Coplens TB, Hanshaw BB (1973) Ultratitration by acompacted clay membrane-I. Oxygen and hydrogenisotopic fractionation. Geochim Cosmochim Acta37:2295–2310
Craig GF, Bell DT, Atkins CA (1990) Response to salt andwaterlogging stress of ten taxa of Acacia selected fromnaturally saline areas of Western Australia. Aust J Bot38:619–630
Craig H (1961) Isotopic variations in meteoric water. Sci-ence 133:1702–1703
Dansgaard W (1964) Stable isotopes in precipitation. Tel-lus 16:436–468
Dawson TE (1996) Determining water use by trees and for-ests from isotopic, energy balance and transpirationanalyses—the roles of tree size and hydraulic lift. TreePhysiol 16:263–272
Dawson TE, Ehleringer JR (1991) Streamside trees that donot use stream water. Nature 350:335–337
Dawson TE, Ehleringer JR (1993) Isotopic enrichment ofwater in the ‘woody’ tissues: Implications for plant wa-ter source, water uptake, and other studies which usethe stable isotopic composition of cellulose. GeochimCosmochim Acta 57:3487–3492
1 3

106 Plant Soil (2007) 291:93–107
Ehleringer JR, Osmond CB (1989) Stable isotopes. In: Pe-arcy RW, Ehleringer JR, Mooney HA, Rundel PW(eds) Plant physiological ecology: Weld techniques andinstrumentation. Chapman & Hall, London, UK, pp281–300
El-Ghonemy AA, Wallace A, Romney EM (1980) Socio-ecological and soil-plant studies of the natural vegeta-tion in the northern Mojave desert-Great Basin desertinterface. Gt Basin Nat Mem 4:73–88
Epstein S, Mayeda T (1953) Variation of 18O content of wa-ter from natural sources. Geochim Cosmochim Acta4:213–224
Feder M, Taube H (1952) Ionic hydration: an isotopic frac-tionation technique. J Chem Phys 20:1335–1336
Feucht JR (2001) Trees and shrubs for high alkaline andhigh salt conditions. Cooperative Extension Service,Tri River Area, Colorado State University
Flanagan LB, Ehleringer JR (1991) Stable isotope compo-sition of stem and leaf water: applications to the studyof plant water use. Funct Ecol 5:270–277
Forti M (1986) Salt tolerant and halophytic plants in Israel.Reclam Reveg Res 5:83–96
Houerou Le (1986) Salt tolerant plants of economic valuein the Mediterranean Basin. Reclam Reveg Res 5:319–341
Jackson J, Ball JT, Rose MR (1990) Assessment of thesalinity tolerance of eight Sonoran desert ripariantrees and shrubs. USBR Contract No. 9-CP-30-017170.University of Nevada System
Karan DM, Macey RI (1980) The permeability of the hu-man red cell to deuterium oxide (heavy water). J CellPhysiol 104:209–214
Kendall C, Caldwell EA (1998) Fundamentals of isotopegeochemistry. In: Kendall C, McDonnell JJ (eds) Iso-tope tracers in catchment hydrology. Elsevier SciencePublishers, New York, pp 51–86
Lin G, da Sternberg LSL (1993) Hydrogen isotopic frac-tionation by plant roots during water uptake in coastalwetland plants. In: Ehleringer JR, Hall AE, FarquharGD (eds) Stable isotopes and plant carbon-water rela-tions. Academic Press Inc., New York, pp 497–510
Lin G, da Sternberg LSL (1992) Comparative study of wa-ter uptake and photosynthetic gas exchange betweenscrub and fringe red mangroves, Rhizophora mangleL. Oecologia 90:399–403
Luo Y-H, da Sternberg LSL, Suda S, Kumazawa S, MitsuiA (1991) Extremely low D/H ratios of photopro-duced hydrogen by cyanobacteria. Plant Cell Physiol32:897–900
Maas EV (1985) Crop tolerance to saline sprinkling water.Plant Soil 89:273–284
McMillan P (1985) Vibrational spectroscopy in the mineralsciences. In: KieVer SW, Navrotsky A (eds) Reviews inmineralogy and geochemistry, vol 14. Macroscopic tomicroscopic: Atomic environments to mineral thermo-dynamics. Mineralogical Society of America, Wash-ington, DC, pp 9–64
Nelson ST, Dettman D (2001) Improving hydrogen isotoperatio measurements for on-line chromium reductionsystems. Rapid Commun Mass Spectrom 15:2301–2306
Nobel PS, Sanderson J (1984) RectiWer-like activities ofroots of two desert succulents. J Exp Bot 35:727–737
Passioura JB (1981) Water collection by roots. In: PalegLG, Aspinell D (eds) The physiology and biochemis-try of drought resistance in plants. Academic Press,New York, pp 39–53
Phillips FM, Bentley HW (1987) Isotopic fractionation dur-ing ion Wltration: I. Theory. Geochim CosmochimActa 51:683–695
PoljakoV-Mayber A (1975) Morphological and anatomicalchanges in plants as a response to salinity stress. In:PoljakoV-Mayber A, Gale J (eds) Plants in saline envi-ronments. Springer-Verlag, New York
Reinoso H, Soas L, Ramírez L, Luna V (2004) Salt-in-duced changes in the vegetative anatomy ofProsopis strombulifera (Leguminosae). Can J Bot82:618–628
Scholander PF, Bradstreet ED, Hammel HT, HemmingsenEA (1966) Sap concentrations in halophytes and someother plants. Plant Physiol 41:529–532
Serrato Valenti G, Ferro M, Ferraro D, Riveros F (1991)Anatomical changes in Prosopis tamarugo Phil. Seed-lings growing at diVerent levels of NaCl salinity. AnnBot London 68:47–53
Serrato Valenti G, Melone L, Orsi O, Riveros F (1992)Anatomical changes in Prosopis cineraria (L.) Duce-hil. Seedlings growing at diVerent levels of NaCl salin-ity. Ann Bot London 70:399–404
Shannon MC (1997) Adaptation of plants to salinity. AdvAgron 60:75–120
Sharma ML (1982) Aspects of salinity and water relationsof Australian chenopods. In: Sen DN, Rajpurohit KS(eds) Task for vegetation science. 2. Contribution tothe biology of halophyte. Junk, The Hague, pp 155–172
Sternberg LSL, Swart PK (1987) Utilization of freshwaterand ocean water by coastal plants of southern Florida.Ecology 68:1898–1905
Stewart MK, Friedman I (1975) Deuterium fractionationbetween aqueous salt solutions and water vapor.J Geophys Res 80:3812–3818
Thorburn PJ, Mensforth LS (1993) Sampling waterfrom alfalfa (Medicago sativa) for analysis ofstable isotopes of water. Commun Soil Sci Plan24:549–557
Thorburn PJ, Walker GR, Brunel J-P (1993) Extraction ofwater from Eucalyptus trees for analysis of deuteriumand oxygen-18: laboratory and Weld techniques. PlantCell Environ 16:269–277
Tyree MT, Sperry JS (1989) The vulnerability of xylem tocavitation and embolism. Annu Rev Plant Phys40:19–38
Vartanian N (1981) Some aspects of structural andfunctional modiWcations induced by drought in rootsystems. Plant Soil 63:83–92
Waisel Y (1972) Biology of halophytes. Academic Press,New York
Waisel Y, Eshel A, Agami M (1986) Salt balance of leavesof the mangrove Avicennia marina. Physiol Plantarum67:67–72
1 3

Plant Soil (2007) 291:93–107 107
Walker CD, Richardson SB (1991) The use of stable iso-topes of water in characterizing the source of water invegetation. Chem Geol (Iso Geosci) 94:145–158
Washburn EW, Smith ER (1934) The isotopic fraction-ation of water by physiological processes. Science79:188–189
Wershaw RL, Friedman I, Heller SJ, Frank PA (1966)Hydrogen isotopic fractionation of water passingthrough trees. In: Hobson GD, Speers GC, IndersonDE (eds) Advances in organic geochemistry. Interna-tional series of monographs on earth sciences, vol 32.Pergamon Press, New York, pp 55–67
White JWC, Cook ER, Lawrence JR, Broecker WS (1985)The D/H ratios of sap in tree: Implications for watersources and tree ring D/H ratios. Geochim Cosmo-chim Acta 49:237–246
Williams DG, Ehleringer JR (2000) Intra- and interspeciWcvariation for summer precipitation use in pinyon-juni-per woodlands. Ecol Monogr 70:517–537
Yakir D, da Sternberg LSL (2000) The use of stable iso-topes to study ecosystem gas exchange. Oecologia123:297–311
Ziegler H, Osmond CB, Stichler W, Trimborn P (1976)Hydrogen isotope discrimination in higher plants:Correlations with photosynthetic pathway and envi-ronment. Planta 128:85–92
Zimmermann U, Ehhalt E, Munnich KO (1967) Soil-watermovement and evapotranspiration: changes in the iso-topic composition of the water. In: Proceedings of thesymposium on isotopes in hydrology, 14–18 November1966. IAEA, Vienna, pp 567–585
1 3