human gut bacteroidetes can utilize yeast mannan through a...
TRANSCRIPT

ARTICLEdoi:10.1038/nature13995
Human gut Bacteroidetes can utilize yeastmannan through a selfish mechanismFiona Cuskin1,2*, Elisabeth C. Lowe1*, Max J. Temple1*, Yanping Zhu1,2, Elizabeth A. Cameron3, Nicholas A. Pudlo3,Nathan T. Porter3, Karthik Urs3, Andrew J. Thompson4, Alan Cartmell5, Artur Rogowski1, Brian S. Hamilton6, Rui Chen7,Thomas J. Tolbert8, Kathleen Piens9, Debby Bracke9, Wouter Vervecken9, Zalihe Hakki5, Gaetano Speciale5,Jose L. Munoz-Munoz1, Andrew Day1, Maria J. Pena2, Richard McLean10, Michael D. Suits11, Alisdair B. Boraston11, Todd Atherly12,Cherie J. Ziemer12{, Spencer J. Williams5, Gideon J. Davies4, D. Wade Abbott2,10, Eric C. Martens3 & Harry J. Gilbert1,2
Yeasts, which have been a component of the human diet for at least 7,000 years, possess an elaborate cell wall a-mannan.The influence of yeast mannan on the ecology of the human microbiota is unknown. Here we show that yeast a-mannanis a viable food source for the Gram-negative bacterium Bacteroides thetaiotaomicron, a dominant member of themicrobiota. Detailed biochemical analysis and targeted gene disruption studies support a model whereby limited cleav-age of a-mannan on the surface generates large oligosaccharides that are subsequently depolymerized to mannose by theaction of periplasmic enzymes. Co-culturing studies showed that metabolism of yeast mannan by B. thetaiotaomicronpresents a ‘selfish’ model for the catabolism of this difficult to breakdown polysaccharide. Genomic comparison withB. thetaiotaomicron in conjunction with cell culture studies show that a cohort of highly successful members of themicrobiota has evolved to consume sterically-restricted yeast glycans, an adaptation that may reflect the incorporationof eukaryotic microorganisms into the human diet.
The microbial community in the human large bowel, the microbiota1,2,is central to the health and nutrition of its host3–6. Glycan utilization isa key evolutionary driver underpinning the structure1,2 of this micro-cosm1,2, with the Bacteroidetes playing a dominant role in this process.The genomes of Bacteroidetes contain polysaccharide utilization loci(PULs)7 that encode the apparatus required to utilize complex carbohy-drates, with each PUL orchestrating the degradation of a specific glycan.
The microbiota contains a cohort of bacteria that targeta-mannosidiclinkages (based on sequence similarity to glycoside hydrolase families(GHs) that are populated by a-mannosidases and a-mannanases)8,9,indicating thata-mannose-containing glycans, such as those from yeastand other fungala-mannans, are significant nutrients for these microbes(see Supplementary Information section 1.0). Furthermore, these glycansare implicated in the immunopathology of the inflammatory bowel disease,Crohn’s disease10,11 (Supplementary Information section 2.0). The genomeof Bacteroides thetaiotaomicron, a dominant member of the microbiota,encodes 36 proteins predicted to displaya-mannosidase ora-mannanaseactivity12. Here we unravel the mechanism by which B. thetaiotaomicronmetabolizes the major a-mannose-containing glycans presented to thelarge bowel. The data also show that B. thetaiotaomicron expresses aspecific yeasta-mannan degrading system that is distinct from the highmannose mammalian N-glycan (HMNG) depolymerizing apparatus.
Yeast a-mannan utilization by B. thetaiotaomicronB. thetaiotaomicron utilizesa-mannan as a sole carbon source and tran-scriptional studies identified three B. thetaiotaomicron PULs (MAN-PUL1,MAN-PUL2 and MAN-PUL3; Fig. 1a) that were activated bya-mannan
from Saccharomyces cerevisiae13, Schizosaccharomyces pombe and thepathogenic yeast Candida albicans (Fig. 1b and Extended Data Fig. 1a, b).B. thetaiotaomicron mutants lacking MAN-PUL2 or MAN-PUL1/2/3were unable to grow on yeast mannan in vitro, (Extended Data Fig. 1c).In gnotobiotic mice fed a diet lacking glycans the B. thetaiotaomicronmutantDMAN-PUL1/2/3 outcompeted the wild-type bacterium, whereaswild-type B. thetaiotaomicron was the dominant species in rodents feda yeast-mannan-rich diet, (Fig. 2a). These data underscore the impor-tance of MAN-PUL1/2/3 when B. thetaiotaomicron is exposed to yeastmannan in vivo, while the intriguing dominance of the mutant strainin animals fed a polysaccharide-free diet is further considered in Sup-plementary Information section 3.2. To explore whether B. thetaiotao-micron degradeda-mannan and HMNG13 by distinct enzyme systems,the PULs activated by a HMNG, Man8GlcNAc2, were evaluated. A singlePUL was activated by Man8GlcNAc2, which was distinct from MAN-PUL1/2/3 (Fig. 1c), demonstrating that degradation, and thus utiliza-tion, of a-mannan and HMNG are carried out by different PULs.
Analysis of the growth profiles of 29 human gut Bacteroidetes speciesrevealed that nine species metabolized S. cerevisiae a-mannan, with 33out of 34 strains of B. thetaiotaomicron growing on the glycan (Fig. 1d).These data show that B. thetaiotaomicron, along with some of its phy-logenetically related neighbours, dominates yeast mannan metabolismin the Bacteroidetes phylum of the microbiota. Within fully sequencedgenomes of 177 microbiota members, the presence of GHs integral toa-mannan degradation (GH38, GH92 and GH76) are restricted to fivespecies of Bacteroides and three of Parabacteroides; no Firmicutes con-tained this grouping of glycoside hydrolases8.
*These authors contributed equally to this work.{Deceased.
1Institute for Cell and Molecular Biosciences, Newcastle University, Newcastle-upon-Tyne NE2 4HH, UK. 2Complex Carbohydrate Research Center, The University of Georgia, 315 Riverbend Road, Athens,Georgia 30602,USA. 3Department of Microbiology and Immunology,University of Michigan Medical School, Ann Arbor, Michigan 48109 USA. 4Departmentof Chemistry, University of York, York YO10 5DD,UK. 5School of Chemistry and Bio21 Molecular Science and Biotechnology Institute, University of Melbourne, Parkville, Victoria 3010, Australia. 6Interdisciplinary Biochemistry Graduate Program, IndianaUniversity, 800 E. Kirkwood Avenue, Bloomington, Indiana 47405, USA. 7Department of Chemistry, Indiana University, 800 E. Kirkwood Avenue, Bloomington, Indiana 47405, USA. 8Department ofPharmaceutical Chemistry, University of Kansas School of Pharmacy, 2095 Constant Avenue, Lawrence, Kansas 66047, USA. 9Oxyrane, 9052 Ghent, Belgium. 10Agriculture and Agri-Food Canada,Lethbridge Research Centre, Lethbridge, Alberta T1J 4B1, Canada. 11Biochemistry and Microbiology, University of Victoria, Victoria, British Columbia V8P 5C2, Canada. 12USDA, Agricultural ResearchService, National Laboratory for Agriculture and the Environment, Ames, Iowa 50011, USA.
8 J A N U A R Y 2 0 1 5 | V O L 5 1 7 | N A T U R E | 1 6 5
Macmillan Publishers Limited. All rights reserved©2015

Structure of the B. thetaiotaomicron a-mannan PULsB. thetaiotaomicron MAN-PUL1 and MAN-PUL2 display significantsynteny (Supplementary Table 1) while MAN-PUL3 displays no organ-izational similarity to the other two loci (Fig. 1a). Characterization ofthe 15 enzymes encoded by the mannan PULs revealed that these lociorchestrate a-mannan degradation from diverse yeasts and possiblyother fungi. For example, MAN-PUL1 contains an a-galactosidase,BT2620, which targetsa-galactosyl linkages absent in S. cerevisiae mannanbut present in other fungal a-mannans such as the yeast Schizosac-charomyces pombe14 (Extended Data Fig. 1d), explaining why inactivationof MAN-PUL1 affects growth of B. thetaiotaomicron on this polysac-charide (Extended Data Fig. 1b). Functional diversity is also evident inMAN-PUL2, which, in addition to its catabolic role, encodes glycosyl-transferases that mediate synthesis of the trisaccharide Man-a-1,3-[Man-a-1,6]-Man (Extended Data Fig. 2). Thus, MAN-PUL2 comprisesa unique example of the co-regulation of related catabolic and biosyn-thetic functions within a single PUL13.
Mannan degradation at the cell surface and periplasmThe enzymatic degradation of a-mannan is restricted through stericconstraints imposed through the side chains appended to the back-bone (Supplementary Table 2 and Extended Data Figs 1e and 4a).Critically, the a-1,6-Man-backbone is not accessible to the endo-a-1,6-mannanases until the side chains have been removed, consistentwith the topography of the substrate binding cleft of these enzymes(Extended Data Fig. 3a, b). The MAN-PUL3-encoded endo-a-1,2-mannosidase BT3862 released a limited proportion of the terminalMan-a-1,3-Man disaccharides, thereby assisting in exposing the back-bone. The structural basis for the specificity displayed by BT3862 is
described in Extended Data Fig. 3c, d. In addition to BT3862, othersurface enzymes are required to partially expose thea-1,6-Man-backbone.Screening for such enzymes revealed an a-mannosidase, BT2199, thatmediates very limited extracellular removal of side chains (ExtendedData Fig. 4a) and thus facilitates limited cleavage of the mannan back-bone by the surface endo-a-1,6-mannanases BT2623 and BT3792. Theactivity profiles of BT2199 and the surface endo-acting mannanasesare consistent with the observed production of large oligosaccharidesby non-growing cells of B. thetaiotaomicron incubated with yeast man-nan (Extended Data Fig. 4b, c).
The cellular location of the key a-mannan hydrolysing enzymes(Fig. 2b, c) indicates that the polysaccharide was degraded primarilyin the periplasm where the side chains were removed by the synergisticaction ofa-mannosidases and sugar-6-monophosphatases15 (Fig. 2d, e,Extended Data Fig. 5, Supplementary Tables 2 and 3 and Supplemen-tary Information section 4.0). The broad specificity of BT3774 enabledthe a-mannosidase to play a key role in the removal of the unchargedside chains, being the only enzyme capable of removing the stericallyrestricteda-1,2-Man units linked to thea-mannan backbone (Fig. 3 andExtended Data Figs 4f and 5) as well as cleaving the Man-1-phosphatelinkage, a critical step in the removal of the phosphorylated branches(Fig. 2d). The importance of BT3774 ina-mannan catabolism was con-sistent with the growth profile of the B. thetaiotaomicron mutant Dbt3774(Extended Data Fig. 1d).
Hydrolysis of the a-1,6-Man backbone of a-mannan by B. thetaio-taomicron was mediated by surface and periplasmic ‘retaining’ endo-a-1,6-mannanases (Extended Data Fig. 4e). The surface endo-mannanasesBT2623 and BT3792 (Fig. 2b, c) generated large oligosaccharides, whilethe periplasmic mannanases BT2631 and BT3782 produced small limit
b
d
B. theta
iotao
micron
(34)
B. ova
tus(2
5)
B. saly
ersia
e (5)
B. xyla
nisolv
ens (1
9)
B. vulg
atus (2
9)
P. dist
ason
is(1
6)
P. mer
dae (3
)
P. john
sonii
(1)
B. cac
cae (1
2)
B. cell
ulosil
yticu
s (10)
B. clar
us(1
)
B. dor
ei(1
4)
B. egg
erthi
i (3)
B. fine
goldi
i (3)
B. flux
us(1
)
B. frag
ilis(5
6)
B. intes
tinali
s (1)
B. mas
silien
sis(3
)
B. nor
dii(4
)
B. olei
ciplen
us(1
)
B. pleb
eius (1
)
B. ster
coris
(7)
B. unif
ormis
(34)
B. intes
tiniho
minis (1
)
D. gad
ei(1
)
D. mos
sii(1
)
O. spla
nchn
icus (1
)
P. gold
steini
i (2)
P. gor
donii
(1)
1.25
1.00
0.75
0.50
0.25
0
No
rmaliz
ed
maxim
um
ab
so
rbance (600 n
M)
Asn
Mammalian high
mannose N-glycan
(HMNG)
P~150 residues
S. cerevisiaeα-mannan
P1
6
~100–150 residues
C. albicansα-mannan
S. pombeα-mannan
~60 residues
c
susC-like or other sentinel gene probed
10
20
BT3
983
BT2
626
BT3
788
BT3
854
BT2
952
BT4
088
BT3
519
BT1
040
BT1
042
BT1
047
BT2
107
BT1
875
BT2
202
BT3
297
BT4
081
BT0
455
BT4
958
BT1
774
30
40
50
60
0
1
10
100
1,000 Unbranched
α-1,6-mannan
C. albicans mannan
BT2626
PUL1
BT3788
PUL2
BT3854
PUL3
Glucose
Chitobiose
α-mannan
Man8GlcNAc2
Fo
ld c
hang
e v
s g
luco
se
BT3780 BT3792BT3791BT3784
BT3783
BT3782BT3781
GH76 GH92 GH76HTCS sensor/regulatorGH130 GH125
GH67 GH76 GH92 GH76
BT2620 BT2623BT2622 BT2629
BT2630
BT2631BT2632
GH125
MAN-PUL1 (BT2620–32)
BT3858 BT3862MAN-PUL3 (BT3853–62)
MAN-PUL2 (BT3773–92)
a
GH92 GH38/CBM32
GT32
BT3773 BT3774 BT3775–BT3778
GH97 HTCS regulator
GH92SARP/OmpR GH99
Glycoside hydrolaseSusC-like
SGBP Glycosyl transferaseSusD-like
Phosphatase
Unknown
Regulator
Mannose
α-1,2α-1,3α-1,6β -1,4
GlcNAcP Phosphate
Galactose
β -1,2
Linkage and sugar key:
B. xylanisolvens 100
98
97
96
95100
90
8196
99
56
100
B. ovatus B. finegoldii B. caccaeB. thetaiotaomicron B. fragilisB. salyersiaeB. nordiiB. intestinalisB. cellulosilyticusB. oleiciplenusB. fluxusB. uniformisB. eggerthiiB. stercorisB. clarusB. plebeiusB. massiliensisB. vulgatusB. doreiD. gadeiD. mossiiO. splanchnicusB. intestinihominisP. distasonisP. merdaeP. johnsoniiP. gordoniiP. goldsteiniiP. copri
100
100
100
100
100
100100100
100
100
100100
100
Figure 1 | B. thetaiotaomicron PULs involved inyeast a-mannan metabolism, and utilization ofthe glycan in Bacteroidetes. a, Genes encodingknown or predicted functionalities are colour-coded and, where appropriate, annotatedaccording to their CAZy family. SGBP; surfaceglycan binding protein. b, Structures of yeast(S. cerevisiae) mannan and HMNG. c, Cells weregrown on media containing glycans as the solecarbon source (n 5 3 separate cultures persubstrate), and susC transcripts derived from PULsthat encode GH76 and/or, GH92 enzymes werequantified by qPCR. The fold change relative toglucose-grown cells is shown on the y axis and thex axis shows the genes probed. Inset: levels of susCtranscripts derived from MAN-PUL1/2/3 whenB. thetaiotaomicron was cultured on mannancomprising only the a-1,6-Man backbone orCandida albicans mannan. Data are averages andstandard deviations of three biological replicates.d, Strains of 29 different bacterial species ofBacteroidetes (number of strains in parentheses)were inoculated into minimal media containingS. cerevisiae mannan as the sole carbon source(n 5 2 replicate cultures per strain). Growthmeasured at 48 h at A600nm. Inset: phylogeny ofBacteroidetes species with bootstrap numbersindicated; organisms growing on mannanconnected by red lines. The non-Bacteroides generawere Odoribacter (O), Parabacteroides (P) andDysgonomonas (D).
RESEARCH ARTICLE
1 6 6 | N A T U R E | V O L 5 1 7 | 8 J A N U A R Y 2 0 1 5
Macmillan Publishers Limited. All rights reserved©2015

products (Extended Data Fig. 6a–c). Consistent with these product pro-files, the periplasmic enzymes were ,100- to 1,000-fold more activethan the surface mannanases against oligosaccharides with a degreeof polymerization (d.p.) ,6, whereas against large oligosaccharides(d.p. $ 6) the periplasmic and surface enzymes displayed broadlysimilar activities (Fig. 2f and Supplementary Table 4). The linear andhence unadorned a-1,6-mannooligosaccharides generated in the peri-plasm were hydrolyzed by the exo-a-1,6-mannosidases BT2632 andBT3781 (Extended Data Fig. 6d, e and Supplementary Table 2), as pre-viously proposed16. The inability of the backbone-cleaving enzymes toattack wild-type yeast a-mannan indicates that these glycoside hydro-lases cannot accommodatea-1,6-linked Man decorated at O2 (side chainsare appended a-1,2 to the mannan backbone) in the active site or prox-imal subsites, consistent with the structures of the substrate bindingclefts of BT3792 and BT3781 (Extended Data Fig. 3a–f).
A B. thetaiotaomicron PUL for HMNG catabolismHMNG-PUL encodes four enzymes and two surface glycan bindingproteins (Extended Data Fig. 7a). BT3990 and BT3991 targeta-1,2-Mananda-1,3-Man linkages, respectively, in HMNGs17. The terminal undec-orated a-1,6-Man exposed by BT3990 is hydrolysed by BT3994, whichrequires GlcNAc at the reducing end for activity, producing Man-a-1,6-Man-b1,4-GlcNAc (Extended Data Fig. 7b). BT3987 is a surface
endo-acting N-acetylglucosaminidase, which cleaves the oligosaccharidefrom its polypeptide (Extended Data Fig. 7c, d). The released HMNG isheld on the surface of B. thetaiotaomicron through the mannose-bindingprotein BT3986, while the SusD homologue BT3984, by recognizingGlcNAc at the reducing end of the glycan (Extended Data Fig. 7e, f andSupplementary Table 5), probably orientates the glycan into the outermembrane porin (SusC homologue BT3983) for transport into the peri-plasm, where the periplasmic a-mannosidases hydrolyse the oligosac-charide into a trisaccharide that is degraded by enzymes that are notencoded by HMNG-PUL. A model for the enzymatic degradation ofHMNG is displayed in Fig. 4. The importance of HMNG-PUL in themetabolism of the N-glycan is consistent with the reduced growth ofthe Dbt3993 mutant (lacking the extra-cellular s factor regulator ofHMNG-PUL) cultured on Man8GlcNAc2 (Extended Data Fig. 7g).
Yeast a-mannan degradation by B. thetaiotaomicronSubstrate promiscuity is a hallmark of thea-mannan degradation appa-ratus of B. thetaiotaomicron, as steric constraints limit enzymatic accessto the glycan side chains, and consequently the polysaccharide backbone.Accordingly, B. thetaiotaomicron uniquely uses surfacea-mannosidase(s),probably BT2199, to generate limited, but sufficient, side chain removalfor the surface endo-acting enzymes to mediate infrequent cleavage ofthe backbone. The B. thetaiotaomicron mannan-degrading apparatus
a) Man8-(P-Man)2
b) Man8-P-Man
c) Man7-P*-Man
d) Man6-(P-Man)2
e) Man6-P2
g) Man5-P-Man
f) Man6-P*-Man
h) Man5-P2
k) Man3-P
j) Man4-P
i) Man5-P
ih jk
Man5Man3 Man4Man6
BT3783 + Mnn4
uncapped
Mnn4 uncappedMnn4
BT3774 + Mnn4
BT2629 + Mnn4
Man9
Man5
Man7
Man6Man5
Man8
BT2629 +
RNaseB
RNaseB
a bc
h j
f
k
g
e
d
d
Rela
tive fl
uo
rescence u
nits
0 h 16 h RP
BT3792
BT3774
BT2630
BT3858
b ca
Wild typeΔMAN-PUL1/2/3
Days post-colonization0 10 20 30 40 0 10 20 30 40 0 10 20 30 40
0
25
50
75
100
Perc
enta
ge o
f
co
mp
eting
str
ain
**
Glycan-free diet
Glycan-free diet + mannan water
Bread diet
**
*
3 4 5 6 7 8
BT2623
BT2631
BT3782
BT3792
107
106
105
104
103
102
101
0
10–1
10–2
Manno-oligosaccharide D.P.
Lo
g10 k
cat/
KM
(M
–1 m
in–1)
e
BT3774 (GH38)
BT3792 (GH76)
Figure 2 | Mannan PULs enable colonization of gnotobiotic mice; keybiochemical and cellular features of the encoded enzymes. a, Colonizationof gnotobiotic mice (n 5 5) by wild type B. thetaiotaomicron (red) and amutant (black) lacking MAN-PUL1/2/3 (DMAN-PUL1/2/3). On day 0 micewere gavaged with ,108 colony-forming units (c.f.u.) of 50:50 of thetwo B. thetaiotaomicron strains and then fed a control diet lackingB. thetaiotaomicron-digestible glycans (green). Left, after 7 days the control dietwas supplemented with 1% YM in drinking water (pink shading). Middle,identical treatment as to the left panel except no mannan was included afterday 7 (shaded green). At day 21 mannan in the water was switched betweenthe groups in the left and middle panels, indicated by the colour panels(pink 5 mannan, green 5 no mannan). Right, mice fed the control diet (shadedgreen) were switched to a diet containing leavened bread (blue). The averageabundance of the mannan utilization mutant (black) in mice fed the bread dietcompared to the corresponding time points in mice fed the glycan-free diet(mean 6 s.e.m., 81% 6 1.2% in animals fed the bread diet versus 90% 6 1.7% inmice fed the glycan-free diet; P 5 0.00005 by unpaired Student’s t-test). Timepoints at which there was a significant difference between the mutant and wild
type B. thetaiotaomicron on the bread diet compared to the glycan-free diet(P # 0.05) are indicated with an asterisk . b, Fluorescent- and light-microscopyimages of B. thetaiotaomicron incubated with polyclonal antibodies againstBT3792 and BT3774. c, Western blots of B. thetaiotaomicron cells cultured onyeast mannan that were untreated with proteinase K (0 h) or incubated with2 mg ml21 proteinase K for 16 h. RP, recombinant protein. Blots wereprobed with antibodies against the B. thetaiotaomicron enzymes. Localizationblots and microscopy images (b, c) are representative data from three biologicalreplicates. d, Phosphorylated high mannose N-glycan MNN4 and RNAaseB incubated with BT2629 (GH92 a-mannosidase), BT3783 (phosphatase) orBT3774 (GH38 a-mannosidase) and the products analysed by capillaryelectrophoresis. Man5 to Man9 (subscript numbers refer to the number of Manunits in the N-glycans) lack phosphate and mannose-1-phosphate groups.P* indicates the N-glycan contains a single phosphate, which is in the followingstructure: Man-a-1-phosphate-a-6-Man-a-1,2-Man. e, Catalytic efficiencyof the endo-a-1,6-mannanases against a-1,6-mannooligosaccharides(see Supplementary Information Table 4 for full data). D.P., degree ofpolymerization of the mannooligosaccharides.
ARTICLE RESEARCH
8 J A N U A R Y 2 0 1 5 | V O L 5 1 7 | N A T U R E | 1 6 7
Macmillan Publishers Limited. All rights reserved©2015

is optimized to minimize nutrient loss, illustrated by the observation thatno oligosaccharides were released into the culture supernatant whenB. thetaiotaomicron was cultured ona-mannan (Extended Data Fig. 4c).
Endo-a-1,6-mannanases are central to the hierarchical degradationof yeast mannan, reflecting their location and substrate specificities.The surface endo-acting enzymes generate large oligosaccharides min-imizing extracellular metabolism and loss to other microbiota residents.The periplasmic endo-a-1,6-mannanases generate numerous short oli-gosaccharides, maximizing the substrate available to the periplasmicexo-acting a-1,6-mannosidases. This ‘selfish’ model is consistent withthe inability of B. thetaiotaomicron to support growth of mannose- and
mannan-backbone-using strains of Bacteroides on intact S. cerevisiaea-mannan (Fig. 5). It should be emphasized, however, that the degra-dation of at least some polysaccharides is mediated through synergisticinteractions between different members of the microbiota18,19, illustrat-ing the diverse mechanisms by which nutrients are used in this micro-bial community.
This work provide insights into the adaptation of the microbiota toyeast domestication in the human diet reflecting the regular consump-tion of yeast-leavened bread, fermented beverages and products suchas soy sauce. B. thetaiotaomicron, and a limited number of other micro-biota-derived Bacteroidetes, have evolved a complex machinery to digestand metabolize yeast cell-wall mannans. Phylogenetic analysis suggeststhat this trait penetrated the human gut Bacteroidetes at least twice,once each in the Bacteroides and Parabacteroides. It is also possible,however, that close relatives of B. thetaiotaomicron have gained parts ofthis complex catabolic trait via separate events. Analysis of 250 humanmetagenomic samples revealed a-mannan-degrading PULs closelyrelated to those from B. thetaiotaomicron in a majority (62%) of sub-jects (Extended Data Fig. 8) making it more common than culturally-restricted traits like red algal porphyran degradation20, but less commonthan plant cell-wall xyloglucan degrading PULs, identified in divergentBacteroides species21,22 (see Supplementary Information section 3.3).
The a-mannan degrading capacity of B. thetaiotaomicron is con-sistent with the bacterium’s ‘glycan generalist’ strategy7,12,23, enablingthe microorganism to thrive in the competitive environment of themicrobiota, where the omnivorous diet of the host requires rapid adap-tion to the nutrients presented to the distal gut. Many of the organismsin the microbiota produce enzymes that attack the major componentsof the human diet such as starch and pectins. An additional survivalstrategy for some members of the microbiota, such as B. thetaiotaomi-cron, is the targeting of low-abundance, highly complex dietary glycansthat are not metabolized by most other organisms, exemplified here byyeast mannan. With respect to the human host the degradation ofa-mannan by B. thetaiotaomicron may also be relevant to the health-promoting effects of the human microbiota24. This work providesinsights into a sophisticateda-mannan degrading apparatus that exists
Surface
endo-α-1,6-mannanases
BT3792 and BT2623 (GH76)
Constitutively expressed
α-mannosidases and endo-α-1,2
mannosidase BT3862 (GH99)
α-1,2/3-mannosidases
BT2629 and BT3784 (GH92)
BT3774 (GH38) and
mannose-6 phosphatases
BT2630 and BT3783
α-1,6-endo-mannanases
BT2631 and BT3782 (GH76)α-1,6-mannosidases
BT2632 and
BT3781 (GH125)
6
1
1
6P
P P
P1
6
1
6
P1
6
P1
6
P1
6
P1
6
BT2629/
BT3784
BT3862
α-1,2α-1,3
β-1,4α-1,6 P Phosphate
GlcNAcMannose
P1
6
P6
BT3774
P
Outer membrane
Periplasm
α-1,3-mannosidase
BT3858 (GH92)
BT3792/
BT2623
BT2630/
BT3783
BT2631/
BT3782
SusC
Figure 3 | Model of yeast mannandeconstruction by B. thetaiotaomicron. Boxesshow examples of bonds cleaved by the endo-a-1,2-mannosidase BT3862, by the a-1,3- and a-1,2-mannosidase activities displayed by BT2629 andBT3784, respectively, and the Man-1-P anda-1,2-Man linkages targeted by BT3774. ExtendedData Fig. 3 shows 3D structures (cartoons,colour-ramped from the amino terminus (blue) tothe carboxy terminus (red)) of the enzymes thatplay a key role in mannan degradation. In thismodel limited degradation occurs at the surfaceand the bulk of glycan degradation occurs in theperiplasm. SusC-like proteins mediate transportacross the outer membrane7.
α-1,2-mannosidase
GH92 (BT3990)
GH18 endo-β-N-
acetylglucosaminidase
(BT3987) Asn SGBP D
C
α-1,6-mannosidase
GH92 (BT3994)
α-1,3-mannosidase
GH92 (BT3991)
α-1,2α-1,3
β-1,4
α-1,6
GlcNAc
Mannose
Figure 4 | Model of HMNG depolymerization by B. thetaiotaomicron. Theinteraction of mannose with the surface glycan binding protein (SGBP) isshown, while the binding of the SusD homologue (D) to the reducing endGlcNAc directs the N-glycan into the SusC-like (C) porin. In both the yeastmannan and the HMNG models the majority of glycan degradation occurs inthe periplasm and not on the surface. The enzymes probably involved in theperiplasmic degradation of the trisaccharide generated by the HMNG-PUL-encoded system are addressed in Supplementary Information section 4.3.
RESEARCH ARTICLE
1 6 8 | N A T U R E | V O L 5 1 7 | 8 J A N U A R Y 2 0 1 5
Macmillan Publishers Limited. All rights reserved©2015

within widespread members of the human microbiota, thereby revealingthe impact made by the historical domestication of yeast and otherdietary fungi on the structure of this microbial consortium.
Online Content Methods, along with any additional Extended Data display itemsandSourceData, are available in the online version of the paper; references uniqueto these sections appear only in the online paper.
Received 24 January; accepted 22 October 2014.
1. Arumugam, M. et al. Enterotypes of the human gut microbiome. Nature 473,174–180 (2011).
2. Backhed, F., Ley, R. E., Sonnenburg, J. L., Peterson, D. A. & Gordon, J. I.Host-bacterial mutualism in the human intestine. Science 307, 1915–1920(2005).
3. Arpaia, N. et al. Metabolites produced by commensal bacteria promote peripheralregulatory T-cell generation. Nature 504, 451–455 (2013).
4. Flint, H. J., Bayer, E. A., Rincon, M. T., Lamed, R. & White, B. A. Polysaccharideutilization by gut bacteria: potential for new insights from genomic analysis.Nature Rev. Microbiol. 6, 121–131 (2008).
5. Kau, A. L., Ahern, P. P., Griffin, N. W., Goodman, A. L. & Gordon, J. I. Human nutrition,the gut microbiome and the immune system. Nature 474, 327–336 (2011).
6. Round, J. L. & Mazmanian, S. K. The gut microbiota shapes intestinal immuneresponses during health and disease. Nature Rev. Immunol. 9, 313–323 (2009).
7. Martens, E. C., Koropatkin, N. M., Smith, T. J. & Gordon, J. I. Complex glycancatabolism by the human gut microbiota: the Bacteroidetes Sus-like paradigm.J. Biol. Chem. 284, 24673–24677 (2009).
8. El Kaoutari, A., Armougom, F., Gordon, J. I., Raoult, D. & Henrissat, B. Theabundance and variety of carbohydrate-active enzymes in the human gutmicrobiota. Nature Rev. Microbiol. 11, 497–504 (2013).
9. Lombard, V., Golaconda Ramulu, H., Drula, E., Coutinho, P. M. & Henrissat, B. Thecarbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 42,D490–D495 (2014).
10. Konrad, A. et al. Immune sensitization to yeast antigens in ASCA-positive patientswith Crohn’s disease. Inflamm. Bowel Dis. 10, 97–105 (2004).
11. Mpofu, C. M. et al. Microbial mannan inhibits bacterial killing by macrophages: apossible pathogenic mechanism for Crohn’s disease. Gastroenterology 133,1487–1498 (2007).
12. Xu, J. et al. A genomic view of the human–Bacteroides thetaiotaomicron symbiosis.Science 299, 2074–2076 (2003).
13. Martens, E. C., Chiang, H. C. & Gordon, J. I. Mucosal glycan foraging enhancesfitness and transmission of a saccharolytic human gut bacterial symbiont. CellHost Microbe 4, 447–457 (2008).
14. Ballou, C. E., Ballou, L. & Ball, G. Schizosaccharomyces pombe glycosylation mutantwithalteredcell surfaceproperties.Proc.NatlAcad.Sci.USA91,9327–9331(1994).
15. Raschke,W.C., Kern,K.A., Antalis,C.&Ballou,C. E.Genetic control of yeastmannanstructure. Isolation and characterization of mannan mutants. J. Biol. Chem. 248,4660–4666 (1973).
16. Gregg, K. J. et al. Analysis of a new family of widely distributed metal-independentalpha-mannosidases provides unique insight into the processing of N-linkedglycans. J. Biol. Chem. 286, 15586–15596 (2011).
17. Zhu,Y.et al. Mechanistic insights intoa Ca21-dependent familyofa-mannosidasesin a human gut symbiont. Nature Chem. Biol. 6, 125–132 (2010).
18. Rakoff-Nahoum, S., Coyne, M. J. & Comstock, L. E. An ecological network ofpolysaccharide utilization among human intestinal symbionts. Curr. Biol. 24,40–49 (2014).
19. Ze, X., Duncan, S. H., Louis, P. & Flint, H. J. Ruminococcus bromii is a keystonespecies for the degradation of resistant starch in the human colon. ISME J. 6,1535–1543 (2012).
20. Hehemann, J. H. et al. Transfer of carbohydrate-active enzymes from marinebacteria to Japanese gut microbiota. Nature 464, 908–912 (2010).
21. Larsbrink, J. et al. A discrete genetic locus confers xyloglucan metabolism in selecthuman gut Bacteroidetes. Nature 506, 498–502 (2014).
22. Martens, E.C., Kelly, A.G., Tauzin, A. S. & Brumer, H. The devil lies in the details: howvariations in polysaccharide fine-structure impact the physiology and evolution ofgut microbes. J. Mol. Biol. 426, 3851–3865 (2014).
23. Martens, E. C. et al. Recognition and degradation of plant cell wall polysaccharidesby two human gut symbionts. PLoS Biol. 9, e1001221 (2011).
24. Everard, A., Matamoros, S., Geurts, L., Delzenne, N. M. & Cani, P. D. Saccharomycesboulardii administration changes gut microbiota and reduces hepatic steatosis,low-grade inflammation, and fat mass in obese and type 2 diabetic db/db mice.MBio. 5, e01011–e01014 (2014).
Supplementary Information is available in the online version of the paper.
Acknowledgements This work was supported by grants from the European ResearchCouncil (G.J.D., Glycopoise; H.J.G., No. 322820), The Wellcome Trust (H.J.G.,WT097907AIA), BBSRC (M.J.T., G.J.D.; BB/G016127/1), US Department of Energy(DOE) Bioenergy Research Center (BESC) supported by the Office of Biological andEnvironmental Research in the DOE Office of Science (M.J.P.) and the NationalInstitutes of Health (E.C.M. and T.J.T., GM090080). Gnotobiotic mouse experimentswere supported by a subsidy from the University of Michigan Medical School HostMicrobiome Initiative, Agriculture and Agri-Food Canada, AgriFlex (D.W.A., #2510),Canadian Institute of Health Research operating grant (A.B.B., MOP-68913), AustralianResearch Council; Mizutani Foundation (S.J.W.). We thank the staff of the DiamondLight Source for the provision of beamline facilities. We would also like to thank variousmembers of ICaMB for providing the yeast strains used in this work. We were greatlysaddened by the passing of C.Z. during the course of this work.
Author Contributions Enzyme characterization: F.C., M.J.T., J.L.M.-M. and D.W.A.Capillary electrophoresis: D.B., K.P. and W.V. E.C.L. created gene deletion strains anddetermined phenotypes with F.C. E.C.L., F.C. and A.R. performed enzyme localisation.F.C. and E.C.L. carried out the co-culturing experiments. Gene expression analysis:E.A.C.,N.A.P. andE.C.M.Growth analysis on purifiedmannans and HMNG:E.C.L., N.A.P.,K.U. and E.C.M.Characterization ofHMNG binding proteins:Y.Z. Characterizationof theDbt3774 mutant: A.D. Phylogenetic reconstruction and metagenomic analysis: E.C.M.Gnotobiotic mouse experiments: E.A.C., N.A.P., N.T.P. and E.C.M. Purification of HMNG:T.J.T., B.S.H. and R.C. Isolation and genomic analysis of pig gut strains: T.A., C.J.Z. A.C.and G.S. performed NMR experiments on GH76 and M.J.P. on GT products. Z.H. andG.S. synthesized substrates. Crystallographic studies by A.J.T., G.J.D., M.D.S., A.B.B. andR.M. Experiments designed by F.C., E.C.L., G.J.D., S.J.W., D.W.A., E.C.M. and H.J.G. Themanuscript was written primarily by H.J.G. and E.C.M. with contributions from S.J.W.,G.J.D. and D.W.A. E.C.L. and E.C.M. prepared the figures.
Author Information The protein crystal structures reported in this study have beendeposited under the following PDB accession codes: 4C1R (BT3783-Mg binarycomplex); 4C1S (BT3792); and 4UTF (BxGH99/Man-IFG/mannobiose ternarycomplex). Reprints and permissions information is available at www.nature.com/reprints. The authors declare no competing financial interests. Readers are welcometo comment on the online version of the paper. Correspondence and requestsfor materials should be addressed to H.J.G. ([email protected]),E.C.M. ([email protected]) or D.W.A. ([email protected]).
a
dc
b
Start
Early
exp
onen
tial
Late
exp
onen
tial
Statio
narySta
rt
Early
exp
onentia
l
Late
exponen
tial
Statio
nary
Start
Early
exp
onentia
l
Late
exponen
tial
Statio
nary
Bt in Bt/Bc mix
Start
Early
exp
onentia
l
Late
exponen
tial
Statio
nary
150
175
Start
Early
exp
onen
tial
Late
exp
onen
tial
Statio
nary
45
Start
Early
exp
onen
tial
Late
ex
ponen
tial
Statio
nary
Start
Early
exp
onen
tial
Late
exp
onen
tial
Statio
nary
Mannan Mannose
Bc alone
Bc in Bt/Bc mix
Bt in Bt/Bc mixBc in Bt/Bc mix
BtBc Bt
Bc
Bx in Bt/Bx mixBt in Bt/Bx mix
Bx in Bt/Bx mixBt in Bt/Bx mixBx alone
BtBx
BtBx
Fo
ld c
hang
e in c
.f.u
. m
l–1
Fo
ld c
hang
e in c
.f.u
. m
l–1
Fo
ld c
hang
e in c
.f.u
. m
l–1
Fo
ld c
hang
e in c
.f.u
. m
l–1
0
75
25
100
35
25
50
15
120
0
20
40
60
80
100
120
0
20
40
60
80
100
120
0
20
40
60
80
100120
0
20
40
60
80
100
125
Start
Early
exp
onen
tial
Late
exp
onen
tial
Statio
nary
0
50
100
150
0
25
50
75
100
Perc
enta
ge (%
)
Mannan Mannose
Perc
enta
ge (%
)
Perc
enta
ge (%
)
Perc
enta
ge (%
)
Figure 5 | Bacteroides co-culture sharing experiments. a–d, B.thetaiotaomicron was co-cultured with Bacteroides cellulosilyticus WH2 (a, b)or with Bacteroides xylanisolvens NLAE-zI-p352 (c, d) with either mannan (a, c)or mannose (b, d) as the sole carbon source. Each non-mannan user wasalso cultured on mannan independently. The upper graph in each panel depictsthe c.f.u. ml21 of each strain, relative to the c.f.u. ml21 at inoculation. Totalc.f.u. ml21 was determined by colony counts, and the proportion of each bacteriawas determined by qPCR of marker genes from genomic DNA (shown in thelower graph of each panel). Error bars represent s.d. of three biological replicates.Bc, B. cellulosilyticus; Bt, B. thetaiotaomicron; Bx, B. xylanisolvens.
ARTICLE RESEARCH
8 J A N U A R Y 2 0 1 5 | V O L 5 1 7 | N A T U R E | 1 6 9
Macmillan Publishers Limited. All rights reserved©2015

METHODSProducing recombinant proteins for biochemical assays. DNAs encoding themature forms of the enzymes used in this study were amplified by PCR usingappropriate primers. The amplified DNAs were cloned into NcoI/XhoI, Nco/BamHI, NdeI/XhoI or NdeI/BamHI restricted pET21a or pET28a, as appropriate.The encoded recombinants generally contained a C-terminal His6-tag, although,where appropriate, the His-tag was located at the N-terminus of the protein. TheGH92 enzymes encoded by HMNG-PUL were cloned in a previous study17. Toexpress the recombinant genes encoding the mannan degrading enzymes, Escherichiacoli strains BL21(DE3) or TUNER, containing appropriate recombinant plasmids,were cultured to mid-exponential phase in Luria Bertani broth at 37 uC. This wasfollowed by the addition of 1 mM (strain BL21(DE3)) or 0.2 mM (TUNER) iso-propylb-D-galactopyranoside (IPTG) to induce recombinant gene expression, andthe culture was incubated for a further 5 h at 37 uC or 16 h at 16 uC, respectively.The recombinant proteins were purified to .90% electrophoretic purity by immo-bilized metal ion affinity chromatography using Talon, a cobalt-based matrix, andeluted with 100 mM imidazole, as described previously25.Producing recombinant proteins BT3783, BT3792 and BxGH99 for crystal-lization. pET28a expression vectors encoding mature BT3783 (residues 27 to 314)and BT3792 (residues 155 to 514) were cloned into pET28a via NheI and XhoIsites. The gene encoding the B. xylanisolvens homologue of GH99 (BxGH99) wasconstructed from synthesized oligonucleotide fragments (Genscript) and also sub-cloned into pET28a using NdeI and XhoI restriction sites. Plasmids encoding BT3783and BT3792 were transformed into E. coli BL21(DE3) Star chemically competentcells, and grown in LB broth at 37 uC supplemented with 50mg ml21 kanamycin.Production of recombinant BT3783 was induced by the addition of 0.2 mM IPTGat a culture OD600 5 0.6, and incubation at 16 uC for 16 h. BT3792 was producedusing the auto-induction method26 by shaking inoculated 0.5-l cultures supple-mented with 50mg ml21 kanamycin for 36 h at 37 uC and then 48 h at 20 uC. E. coliBL21 (DE3) cells harbouring the BxGH99-encoding plasmid were cultured in 0.5 lZYM-5052 auto-induction media26, supplemented with 50mg ml21 kanamycin, at37 uC for 8 h, with induction occurring overnight at 16 uC.
To purify BT3783 and BT3792, cells were collected by centrifugation and rup-tured by chemical lysis procedure at 4 uC. In brief, cells were resuspended in 25 mlof a solution consisting of 7% (w/v) sucrose, 50 mM Tris-HCl (pH 7.5), for 5 min.Lysozyme (Sigma-Aldrich; 10 mg) was then added and stirred for 10 min. A solu-tion (50 ml) consisting of 0.6% (w/v) Triton-X, 0.6% (w/v) deoxycholate, 20 mMTris-HCl (pH 7.5) was added and stirring continued, after which 5 mM MgCl2 wasadded followed by addition of DNase (Sigma-Aldrich) to a final concentration of8.5mg ml21. The resulting solutions were then centrifuged for 45 min at 13,000gSimilarly, cells harbouring BxGH99 were collected and resuspended in 50 mMNaH2PO4 (pH 8.0), 300 mM NaCl, and lysed by sonication.
BT3783 clarified cell lysates were purified by nickel Sepharose affinity chro-matography by stepwise elution with imidazole. Positive fractions identified bySDS gel electrophoresis were dialysed into 20 mM Tris (pH 8.0) and concentratedto 11.25 mg ml21 with an Amicon stirred ultrafiltration unit model 8200. BT3792clarified cell lysates were purified using nickel-affinity chromatography, andpositive fractions were buffer exchanged by dialysis into 25 mM Tris-HCl (pH 8.0)before further purification by anion exchange chromatography. Anion exchangepurified BT3792 buffered in 25 mM Tris-HCl (pH 8.0), 500 mM NaCl, and 2 mMdithiothreitol was concentrated using a stirred cell Amicon with a 10 kDa cutoff to20 mg ml21. For BxGH99 soluble lysate (isolated by centrifuging at 13,000g) wasapplied to a NiSO4-charged 5 ml HiTrap chelating column (GE Healthcare), pre-equilibrated in the same buffer. Protein was eluted in an imidazole gradient, dia-lysed, concentrated, and further purified on an S75 16/60 gel filtration column (GE)pre-equilibrated in 25 mM Na-HEPES (pH 7.0), 100 mM NaCl, 1 mM dithiothreitol.Binding studies. Affinity gel electrophoresis was used to screen for potentialglycan binding proteins, following the method described in ref. 27 with the targetpolysaccharide at 1 mg ml21. The binding of proteins to their ligands was quantifiedby isothermal titration calorimetry (ITC), as described previously27. Titrations werecarried out in 50 mM Na-HEPES buffer, pH 7.5 at 25 uC. The reaction cell containedprotein at 50mM, while the syringe contained either the oligosaccharide at 10 mMor polysaccharide at 3–5 mg ml21. The titrations were analysed using MicrocalOrigin version 7.0 software to derive n, Ka, andDH values, whileDS was calculatedusing the standard thermodynamic equation, RTlnKa 5DG 5DH 2 TDS.Glycoside hydrolase and phosphatase assays.a-Mannosidase activity was deter-mined by the continuous monitoring of mannose release using a linked enzymeassay system purchased from Megazyme International (mannose/fructose/glu-cose detection kit). The reaction was carried out at 37 uC in 50 mM Na-HEPESbuffer (pH 7.0) containing 2 mM MgCl2, 1 mM ATP and 1 mM NADP1, excessconcentrations of linker enzymes (hexokinase, phosphoglucose isomerase andglucose-6-phosphate dehydrogenase) and 1 mg ml21 BSA. Through linker enzymes
glucose-6-phosphate, generated from the released mannose, is oxidized by glucose-6-phosphate dehydrogenase with concomitant reduction of NADP1 to NADPH,which was monitored at 340 nm using an extinction coefficient of 6223 M21 cm21.The polysaccharide substrates used were S. cerevisiae a-mannans from wild-typeor appropriate mutant strains that produce variants of the polysaccharide (ExtendedData Fig. 5); these mannans were purified from stationary-phase cultures of theyeast grown in yeast extract peptone dextrose medium, as described previously15.The a-1,6-mannooligosaccharides, which were also used as substrates, were gen-erated as follows: 1 ga-mannan from the S. cerevisiae mutant mnn2 (comprises thea-1,6-mannan backbone with no side chains) was digested to completion with theendo-mannanase BT3792. The products were freeze-dried and the small mannoo-ligosacchardes (d.p. 2 to 5) were purified on two P2 Bio-gel columns set up in series,while the large oligosaccharides, with a d.p. of 6 to 8, were fractionated on two P4Bio-gel columns also in series. The columns were run at 0.2 ml min21 in distilledwater. The 5-ml fractions were evaluated by thin-layer chromatography (TLC) andthose containing the same oligosaccharide were pooled. The activity of the endo-mannanases against a-mannans was determined in 50 mM Na-HEPES buffer(pH 7.5) containing an appropriate concentration of the polysaccharide (rangingfrom 0.1–6 mg ml21) and 1 mg ml21 BSA. Reactions were incubated for 30 min at37 uC and, at regular time intervals, 500-ml aliquots were removed and the amountof reducing sugar was quantified deploying the dinitrosalicylic acid reagent28 and astandard curve of mannose in the reaction conditions used. TLC was also used toprovide a qualitative profile of the mannooligosaccharides generated by the GH76endo-mannanases from these reactions. Around 4ml of the reaction was spotted onsilica gel TLC plates and the plates were developed in butanol:acetic acid:water2:1:1 and carbohydrate products detected by spraying with 0.5% orcinol in 10%sulfuric acid and heating to 100 uC for 10 min. Substrate-depletion assays wereused to determine the activity of the endo-mannanases against a-1,6-mannooli-gosaccharides. In brief, 50mM of the oligosaccharides in 50 mM sodium phosphatebuffer (pH 7.5), containing 0.1 mg ml21 BSA (NaP buffer), was incubated with anappropriate concentration of enzyme (10 to 500 nM). Aliquots were removed atregular intervals for up to 1 h and, after boiling for 10 min to inactivate the enzyme,the amount of the substrate mannooligosaccharide remaining was quantifiedby high-performance anion-exchange chromatography (HPAEC) using standardmethodology. In brief, the reaction products were bound to a Dionex CarboPacPA-200 column equilibrated with 100 mM NaOH. Mannose and mannooligosac-charides were eluted with a 0–200 mM sodium acetate gradient in 100 mM NaOHat a flow rate of 0.25 ml min21, using pulsed amperometric detection. The datawere used to determine catalytic efficiency (kcat/KM) as described previously29. Todetermine the activity of the GH99 endo-a-1,2-mannosidase BT3862, the enzymewas incubated with Man-a-1,3-Man-a-1,2-Man-a-1,2-Man-1-CH3. and, in com-bination with the a-1,2-mannosidase BT3990, mannose release was monitoredusing the continuous assay described above (BT3990 is active on the product ofBT3862, Man-a-1,2-Man-a-1,2-Man-1-CH3). The activity of BT3862 against aryl-mannosidases was as described previously30. A strategy deploying 2-aminobenzamide(2-AB)-labelled glycans at the reducing end, using the Sigma GlycoProfile 2-ABlabelling kit as per the manufacturer’s instructions, was used to determine thecatalytic efficiency of BT3994. The enzyme (100 nM) was incubated with 5mMMan5GlcNAc2 (generated by treating Man9GlcNAc2 with BT3990 an a-1,2-mannosidases) or Man3GlcNAc2 (generated from Man5GlcNAc2 by treating withBT3991, a-1,3-mannosidase17) in NaP buffer for up to 1 h. Aliquots were removedat regular intervals and the amount of glycan remaining was determined usingHPAEC and a fluorometric detector. A similar strategy, employing labelled glycansin combination with HPAEC and fluorescence detection was used to measure theendo-b-N-acetylglucosaminidase activity of BT3987 against Man9GlcNAc2. Theactivity of the enzyme was also evaluated against glycoproteins using matrix-assistedlaser desorption/ionization time of flight (MALDI-TOF) mass spectrometry asdescribed below. BT2630 and BT3783 were assayed for phosphatase activity usingthe EnzChek Phosphate Assay kit to detect the release of phosphate from appro-priate phosphorylated sugars. Prior to these assays the two proteins were treatedwith EDTA and the chelating agent was removed using a PD10 column. The assayswere carried out in 50 mM Na-HEPES buffer (pH 7.5) containing 1 mM of MgCl2or another divalent ion. In this discontinuous assay aliquots were removed atregular intervals, the enzyme was inactivated by boiling and phosphate releasedetermined.
The specificity of enzymes against phosphorylated and neutral high mannoseN-glycans was assessed by capillary electrophoresis. The methodology was essen-tially that described in ref. 31. The two substrates were MNN4 (phosphorylatedhigh mannose N-glycans released from glycoproteins expressed by a geneticallymodified strain of Yarrowia lipolytica) and N-glycan from RNAase B (primarilyhigh mannose N-glycans), which were released from their respective glycopro-teins with PNGase F. The N-glycans were labelled at the reducing end with 8-aminopyrene-1,3,6-trisulfonic acid32, and incubated overnight at 30 uC with
RESEARCH ARTICLE
Macmillan Publishers Limited. All rights reserved©2015

,2mM of enzyme in 10 mM Na-Hepes buffer (pH 7.0) containing 2 mM CaCl2.The reactions were analysed by capillary electrophoresis with laser-induced fluor-escence detection (CE-LIF) using an ABI 3130 capillary DNA sequencer as describedpreviously32.Mannosyl-transferase assays. BT3775 and BT3776 at 30mM were screened foractivity using mannose and all possible a-mannobiose acceptors at a concentra-tion of 10 mM. Reactions were performed in 10 mM Na-Hepes buffer (pH 7.5),10 mM MnCl2, 50 mM GDP-Mannose at 37 uC over time course intervals of0 min, 5 min, 15 min, 30 min, 60 min and 16 h. Reactions were stopped by boilingfor 10 min. To explore the synergy between the two GT32 glycosyltransferases,reactions were performed with BT3775 and BT3776 simultaneously using man-nose as an acceptor, and in a step-wise progression with BT3775 and mannose asthe acceptor, followed by BT3776 on the purified products of the initial BT3775reaction. Enzyme concentrations were maintained in both conditions at 30 mM.MALDI-TOF mass spectrometry of reaction products. To permethylate N-glycanfragments and mannooligosaccharides appropriate enzyme reactions were freeze-dried and suspended in dry dimethylsulfoxide (DMSO; 200ml). Oligoglycans wereper-O-methylated using standard methods33. Freshly prepared sodium hydroxidebase in dry DMSO (300ml) and then iodomethane (150ml) was added to eachsample. The tube was purged with nitrogen and vortexed vigorously. Permethylatedoligosaccharides (PMOs) were extracted in 2 ml water. After removing excessiodomethane (CH3I), 2 ml dichloromethane was added and vortexed to extractlipophilic components. The aqueous layer was removed after centrifugation. Thiswas repeated five more times. After the final rinse, the dichloromethane layer wastransferred into a fresh tube and dried down under N2 stream. The samples werethen dehydrated and dissolved in 50% aqueous methanol and loaded onto C18reverse phase resin (100 mg ml21, Resprep SPE Cartridges) and eluted with acet-onitrile (AcN). AcN washes were dried down under N2 and dissolved in 20mlmethanol. Equal volumes of the sample and 2% DHBA in aqueous methanol, whichwas used as the matrix, were combined in a separate microfuge tube and loaded onthe MALDI-TOF mass spectrometry plate. MALDI-TOF mass spectrometry ana-lyses were performed with an AB4700 MS instrument (Applied Biosystems) inpositive ion, reflector mode with 100 shots taken per spectrum. PMO mass valueswere queried online (http://www.expasy.org/tools/glycomod/) by GlycoMod toolthat can predict possible oligosaccharide structures that occur on proteins fromtheir experimentally determined masses.NMR analysis of glycosyltransferase reaction products. The mannooligosac-charide products were separated by size exclusion chromatography on a Superdex-75 HR10/30 column using a Dionex Ultimate 3000 HPLC equipped with a ShodexRI-101 refractive index detector. The column was eluted with water and fractions werecollected and freeze-dried. The lyophilized fraction containing the trisaccharidewas dissolved in D2O (0.3 ml, 99.9%; Cambridge Isotope Laboratories) and one-and two-dimensional NMR spectra were recorded at 298 K with a Varian InovaNMR spectrometer operating at 600 MHz equipped with a 3 mm NMR cold probe.The homonuclear (gCOSY, TOCSY and NOESY) and the hetereonuclear (gHSQCand gHMBC) experiments were recorded using standard Varian pulse programs.Chemical shifts were measured relative to internal acetone (dH 2.225). Data wereprocessed using MestReNova software (Universidad de Santiago de Compostela,Spain). Interglycosidic scalar couplings observed in the gHMBC spectra were usedto determine the sequence and glycosidic linkages of the mannosyl residues in thea-Man-(1,3)-[a-Man-(1,6)]-Man trisaccharide product.Linkage analysis of GT32 glycosyltransferase mannooligosaccharide pro-ducts. Overnight incubations were boiled for 5 min and then centrifuged to pelletdenatured protein. A 200mg aliquot of the sample was suspended in 200ml DMSOand magnetically stirred for 5 days. The sample was then permethylated accord-ing to the method in ref. 34. Two hundred microlitres of the NaOH base wereadded and, after 10 min, 100ml CH3I were added and the sample was stirred for40 min. An additional 200ml base and 100mL CH3I were then added and stirringwas continued for 40 min. The reaction was worked up by addition of 2 ml H2O,removal of excess CH3I by sparging with N2, and CH2Cl2 extraction. The per-methylated material was hydrolyzed using 2 M trifluroacetic acid (TFA) (2 h in asealed tube at 121 uC), reduced with NaBD4, and acetylated using Ac2O/TFA. Theresulting partially methylated alditol acetates were analysed on a Hewlett Packard5890 GC interfaced to a 5970 MSD (mass selective detector, electron impact ion-ization mode); separation was performed on a 30 m Supelco 2330 bonded phasefused silica capillary column.Stereochemistry of endo-a-1,6-mannanases. A solution of BT3792 (25 mg,approximately 400mmole) in buffered D2O (0.25 ml, 50 mM phosphate-citrate,200 mM NaCl, pD 6.0 (deuterium ion concentration is 1 mM)) was added to asolution of 4-nitrophenyla-D-mannopyranosyl-1,6-a-D-mannopyranoside (4.1 mg,0.0086 mmol) in D2O (0.75 ml, 50 mM phosphate-citrate, 200 mM NaCl, pD 6.0)at 22 uC. The course of the reaction was monitored by 1H NMR (500 MHz) toidentify the stereochemistry of the reaction by analysing the relative timing of the
appearance of the anomerica- andb-proton signals. Signals were assigned by two-dimensional NMR analysis (HSQC).Bacteroides culture and whole-cell assays. B. thetaiotaomicron was culturedanaerobically at 37 uC in minimal media containing an appropriate carbon source,or in TYG (tryptone yeast extract glucose medium) as described previously21.Growth curves presented in the paper are averages of six biological replicates.B. thetaiotaomicron was grown in 5 ml minimal media on 0.5% w/v S. cerevisiaemannan (Sigma) or glucose as the sole source to mid exponential phase (OD600nm
0.6–0.8). Cells were harvested by centrifugation, 5000g for 10 min at room tem-perature and washed in 5 ml PBS (pH 7.1) before being resuspended in 500ml PBS.Cells (50ml) were assayed against yeast mannan (10 mg ml21) at 37 uC for 16 h.Assays were analysed by thin layer chromatography, 5ml of each sample was spottedonto silica plates and resolved in butanol/acetic acid/water buffer. The plates weredried and sugars visualized by orcinol/sulfuric acid heated to 70 uC.Cellular localization. Cultures of B. thetaiotaomicron (100 ml) were grown inminimal media on yeast mannan (0.5% w/v) as a sole carbon source, to midexponential growth phase (OD600nm 0.6–0.8). Cells were harvested by centrifu-gation and washed in 10 ml PBS before being resuspended in 5 ml of the buffer.The cells were split into four 1 ml aliquots. To 3 of the aliquots 2 mg ml21 ProteinaseK was added and incubated at 37 uC for 1–16 h, the fourth sample was left as anuntreated control also for 16 h. Following incubation with the protease the sampleswere centrifuged at 5000g for 10 min and the supernatant discarded. The cellpellets were resuspended in 1 ml PBS and the proteins precipitated by the additionof 200ml trichloroacetic acid and incubation on ice for 30 min. The precipitatedproteins were pelleted by centrifugation and washed 4 times in 1 ml ice-coldacetone. The protein pellets were resuspended in 250ml Laemmli buffer and sub-jected to SDS/PAGE. Gels were transferred to Whatman Protran BA 85 nitrocel-lulose membrane. Proteins of interest were detected using anti-sera raised againstthe corresponding protein. The secondary antibody used was a chicken anti-ratconjugated to horseradish peroxidase. Antibodies were detected by chemi-lumin-escence using Biorad Clarity Western ECL Substrate.Immunofluorescence microscopy. B. thetaiotaomicron suspensions (OD600nm 5
0.8) in PBS (pH 7.0) were applied to clean Eppendorf tubes, fixed by an equalvolume of 23 formalin (9% formaldehyde in PBS), and rocked for 90 min at 25 uC.The cells were then pelleted by centrifugation for 3 min at 7000g and washed twicewith 1 ml of PBS. The bacterial cell pellet was resuspended in 1 ml of blockingsolution (2% goat serum, 0.02% NaN3 in PBS) and incubated at 4 uC for 16 h. Afterincubation cells were centrifuged again at 7000g and the supernatant discarded.For labelling, the bacteria were incubated with 0.5 ml of primary rat IgG (1/500dilution of IgG in blocking solution) for 2 h at 25 uC. The cells were then pelleted,by centrifugation, washed in 1 ml of PBS and resuspended in 0.4 ml goat anti-ratIgG Alexa-Fluor 488 (Sigma), diluted 1/500 in blocking solution, and incubated 1 hat 25 uC in the dark. The cells were then pelleted, washed with PBS and resus-pended in 50ml of PBS containing ProLong Gold antifade reagent (Life Tech-nologies). Labelled bacterial cells were mounted onto glass slides and secured withcoverslips. Fluorescence was visualized using a Leica SP2 UV microscope (LeicaMicrosystems, Heidelberg, GmbH) with 363 NA 1.32 lens. Alexa-Fluor 488-labelledbacteria were viewed under an ultraviolet microscope view and compared withbright-field phase-contrast of the same image.Constructing mutants of the yeast mannan and HMNG PULs in B. thetaio-taomicron. The inactivation of the HMNG-PUL extra-cellular factor regulatorwas described previously13. The other mutants deployed in this study, PUL knock-outs and single gene clean deletions, were introduced by allelic exchange using thepExchange vector as described in ref. 35 The use of quantitative reverse transcrip-tase PCR to quantify appropriate transcripts followed the methods described inref. 13.Gnotobiotic mice experiments. All animal experiments were approved by theUniversity Committee on Use and Care of Animals at the University of Michiganand were supervised by a veterinarian. Germ-free mouse experiments were con-ducted in a total of 24, 6–8-week-old male and female Swiss Webster mice (eachwas considered to be a single biological replicate in its respective experiment andtreatment group). Mice were randomly assigned into groups by a technician whowas not familiar with the project. The investigators were not blinded to the iden-tities of the treatment groups during the experiment and no data were excludedfrom the final analysis. For in vivo gene expression studies, mice were randomlygrouped into three groups containing three animals each, and then subjected tothree different dietary regiments. Two groups were fed a gamma-irradiated customdiet (glycan-free diet) that contained only glucose as the available carbohydrate aswell as cellulose, a polysaccharide that cannot be degraded by B. thetaiotaomicron,as a non-digestible fibre supplement36. One of the two groups maintained on thisdiet was provided with purified a-mannan (1% w/v) in drinking water (Harlan-Teklad). The third group was fed a custom version of the glycan-free diet in whichthe glucose was replaced with dried/crumbed bread (50% w/w of final diet) that
ARTICLE RESEARCH
Macmillan Publishers Limited. All rights reserved©2015

had been produced using yeast as a leavening agent (Zingerman’s Bake House,Ann Arbor, MI). Mice were pre-fed on each dietary condition for five days, colo-nized with B. thetaiotaomicron by oral gavage (,108 c.f.u. per animal) and main-tained for an additional 5 days before euthanizing and collection of caecal contentsfor gene expression analysis. For the in vivo competition experiment, wild-type andmannan PUL triple mutant strains were each labelled with a unique 24 base pair(bp) oligonucleotide tag, which is contained in a pNBU2-based chromosomalintegration vector and quantifiable by qPCR, as previously described13. Each strainwas grown overnight in TYG medium and combined at approximately equal amountsbefore being gavaged into mice as described above. For competition experiments,five mice were used in each group (chosen based on the sample size used inprevious studies to observe significant changes in competitive index18,29) and fedvarying regimens of the three diets described above for a total period of 38 days. Allgermfree mice used in the competition experiment were pre-fed the glycan-freediet for 1 week before colonization.Bacteroides co-culturing experiments. B. thetaiotaomicron, B. xylanisolvensNLAE-zl p352 and B. cellulosilyticus WH2 were cultured in minimal media con-taining glucose (0.5% w/v) as a carbon source to mid exponential growth phase(OD600 0.7–0.8) . Cells were collected by centrifugation and washed twice with5 ml of PBS to remove any residual glucose. The washed cells were resuspended in5 ml of fresh minimal media with no carbon source. Minimal media (10 ml)containing mannose or yeast mannan as the carbon source was inoculated withequal volumes of B. thetaiotaomicron/B. xylanisolvens or B. thetaiotaomicron/B.cellulosilyticus. A control culture of minimal media containing yeast mannan wasalso inoculated with either B. xylanisolvens or B. cellulosilyticus. All cultures wereset up in triplicate. Samples (1 ml) were taken at the point of inoculation, earlyexponential, late exponential and stationary phase of growth. Serial dilutions ofeach sample were plated on to rich media and incubated for 2 days before colonycounts were recorded. Proportions of B. thetaiotaomicron, B. xylanisolvens andB.cellulosilyticus per sample were determined by quantitative PCR from genomicDNA using unique marker genes for each strain.Searches of human gut metagenomic data sets for mannan PULs. A search of250 different healthy and diseased (Crohn’s disease and ulcerative colitis) humanmetagenomic samples was conducted exactly as described previously36. Probescorresponding to the three B. thetaiotaomicron mannan PULs were first checkedagainst database of complete and draft microbial genome sequences to identifyregions that were present in species other than B. thetaiotaomicron, or the porcineB. xylanisolvens strains in the case of MAN-PUL1. In the case of MAN-PUL2 andMAN-PUL3, the entire sequence was determined to be specific for sequencedB. thetaiotaomicron isolates; whereas, MAN-PUL1 was trimmed to a region con-taining 15.059 kilobases (kb) that were unique to B. thetaiotaomicron and porcineB. xylanisolvens. The three probes used correspond to the following base pair coor-dinates in the B. thetaiotaomicron VPI-5482 genome sequences: MAN-PUL1(3262908..3277966), MAN-PUL2 (4893415..4928241) and MAN-PUL3 (5012915..5031551).Crystallization. BT3783 and BT3792 were crystallized using the sitting drop vapourdiffusion method at 18 uC. BT3783 crystals were obtained by mixing equal volumesof purified, recombinant BT3783 protein at a concentration of 11.25 mg ml21 withmother liquor solution consisting of 15% (w/v) polytheylene glycol 4,000, 5% (1/-)-2-methyl-2,4-pentanediol (v/v), 3 mM D-mannose, and 200 mM MgCl2. Crystalsof BT3792 were obtained by mixing equal volumes of purified recombinant BT3792at a concentration of 20 mg ml21 buffered in in 25 mM Tris-HCl (pH 8.0), 500 mMNaCl, 2 mM dithiothreitol with a reservoir solution consisting of 5% glycerol (v/v),24% (w/v) polyethylene glycol monomethyl ether (average Mw 2,000), 0.25 M sodiumacetate, and Bis-Tris-HCl (pH 5.5). Large, thin plate crystals of BT3792 developedover a period of 5 days to a week. BT3792 crystals were cryoprotected in crystallizationsolutions supplemented with 25% ethylene glycol and cryo-cooled directly in a N2
stream at 2160 uC before diffraction experiments.BxGH99 crystals were grown at 19 uC using hanging-drop vapour diffusion
with equal volumes of protein (30 mg ml21) and reservoir solution consisting of0.1 M sodium acetate pH 5.1, 20% w/v polyethylene glycol monomethyl ether(average Mw 2,000), 2.0% low molecular weight poly-c-glutamic acid. Ligandcomplex formation as a ternary with Man-isofagamine and a-1,2-mannobiose,was achieved by soaking native BxGH99 crystals in mother liquor supplementedwith approximately 10 mM of respective ligand solutions for a period of 30 minbefore flash cooling in liquid N2. Crystals were cryo-protected by the stepwiseaddition of ethylene glycol, supplemented with appropriate ligand(s), to a finalconcentration of 20% v/v.Data collection, structure solution and refinement. Diffraction data from bothBT3783 and BT3792 crystals were collected at the Canadian Lightsource beam-line 08B1-1 (CMCF-BM) in oscillation mode (l5 0.97961 and 0.98005 A, withdata collected at 100 K, respectively). The BT3783 and BT3792 data sets wereprocessed with iMosflm37,38 and XDS39, respectively, and both were scaled with
SCALA38. BT3783 and BT3792 structures were solved by molecular replacementwith the program Phaser MR40 using the coordinates of apo-BT3783 (PDB:3MPR) and the coordinates of Lin0763 from Listeria innocua (PDB: 3K7X).The asymmetric unit of BT3783 and BT3792 contain four and two protein mole-cules, respectively. The structures were subsequently improved with cycles ofmanual building with COOT41 and refinement with REFMAC38. Five per centof the reflections were flagged as ‘free’ to monitor the refinement process. Thestructure of BT3783 extends from residues 26–309 with a magnesium ion coor-dinated in the putative active centre. The structure of BT3792 runs continuouslyfrom residues 151–525. Due to a lack of continuous density corresponding to thispolypeptide region, residues 274–290 are absent in the model of BT3783. BT3792structural refinement required amplitude correction for twinning. The final refinedstructures were validated with the aid of Molprobity42. Final BT3783 and BT3792structures featured, respectively, 94.8 and 99.6% of all modelled residues within theRamachandran ‘favoured’ region, with a further 4.2 and 0.4% within the ‘additionalallowed’ region.
Diffraction data for the BxGH99-Man-IFG-mannobiose ternary complex werecollected at beamline I03 (l5 0.97625 A, data collected at 100 K) of the DiamondLight Source, Didcot, UK. The data sets were processed using XDS39 and AIMLESS43
(also within CCP438). Phases were derived from a previously solved (native) BxGH99atomic model, with initial refinement conducted using REFMAC38. Calculatedelectron density maps were visually assessed for evidence of ligand-binding, withsubsequent completed atomic models, featuring appropriately assigned ligandcoordinates, refined to convergence through numerous cycles of REFMAC38 andadditional manual correction using COOT41. The final BxGH99 ternary structurefeatured, respectively, 96.5 and 96.9% of all modelled residues within the Ramachandran‘favoured’ region, with a further 2.3 and 1.9% within the ‘additional allowed’ region.Data collection and refinement statistics for all structures are presented in Sup-plementary Table 6.Experimental group size and statistical analysis of data. For all quantitativeenzyme assays at least three technical replicates were performed unless specif-ically stated in the Table legends. Standard errors of the mean are stated for kineticparameters in Supplementary Tables 2–4. For Supplementary Table 5 the kineticparameters are reported 6 the error of the fit of the data using linear regression.For isothermal titration calorimetry experiments three technical replicates wereperformed for each titration. Standard errors of the mean are shown in Sup-plementary Table 5. In the gnotobiotic mice experiments displayed in Fig. 2a fivemice were assigned to each diet and the error bars for the two bacterial strainsrepresent standard errors of the mean. Statistical significance of the persistence ofdifferent strains of B. thetaiotaomicron in the same mice was analysed by unpairedStudent’s t-test. For the microbial sharing experiment detailed in Fig. 4 the errorbars represent standard deviations of the mean from three biological replicates.
25. Charnock, S. J. et al. Key residues in subsite F play a critical role in the activityof Pseudomonas fluorescens subspecies cellulosa xylanase A againstxylooligosaccharides but not against highly polymeric substrates such as xylan.J. Biol. Chem. 272, 2942–2951 (1997).
26. Studier, F. W. Protein production by auto-induction in high density shakingcultures. Protein Expr. Purif. 41, 207–234 (2005).
27. Szabo, L. et al. Structure of a family 15 carbohydrate-binding module in complexwith xylopentaose. Evidence that xylan binds in an approximate 3-fold helicalconformation. J. Biol. Chem. 276, 49061–49065 (2001).
28. Miller, G. L. Useofdinitrosalicylic acid reagent for determination of reducing sugar.Anal. Chem. 31, 426–428 (1959).
29. Charnock, S. J. et al. The topology of the substrate binding clefts of glycosylhydrolase family 10 xylanases are not conserved. J. Biol. Chem. 273,32187–32199 (1998).
30. Thompson, A. J. et al. Structural and mechanistic insight into N-glycan processingby endo-a-mannosidase. Proc. Natl Acad. Sci. USA 109, 781–786 (2012).
31. Stewart, T. S., Mendershausen, P. B. & Ballou, C. E. Preparation of amannopentaose, mannohexaose, and mannoheptaose from Saccharomycescerevisiae mannan. Biochemistry 7, 1843–1854 (1968).
32. Laroy, W., Contreras, R. & Callewaert, N. Glycome mapping on DNA sequencingequipment. Nature Protocols 1, 397–405 (2006).
33. Ciucanu, I. Per-O-methylation reaction for structural analysis of carbohydrates bymass spectrometry. Anal. Chim. Acta 576, 147–155 (2006).
34. Anumula, K. R. & Taylor, P. B. A comprehensive procedure for preparation ofpartially methylated alditol acetates from glycoprotein carbohydrates. Anal.Biochem. 203, 101–108 (1992).
35. Koropatkin, N. M., Martens, E. C., Gordon, J. I. & Smith, T. J. Starch catabolism by aprominent human gut symbiont is directed by the recognition of amylose helices.Structure 16, 1105–1115 (2008).
36. Larsbrink, J. et al. A discrete genetic locus confers xyloglucan metabolism in selecthuman gut Bacteroidetes. Nature 506, 498–502 (2014).
37. Leslie, A. W. & Powell, H. in Evolving Methods for Macromolecular Crystallography(eds Read, J. R. & Sussman, J. L.) Ch. 4, 41–51 (Springer, 2007).
38. Winn, M. D. et al. Overview of the CCP4 suite and current developments. ActaCrystallogr. D 67, 235–242 (2011).
39. Kabsch, W. Xds. Acta Crystallogr. D 66, 125–132 (2010).
RESEARCH ARTICLE
Macmillan Publishers Limited. All rights reserved©2015

40. McCoy, A. J.et al.Phasercrystallographic software. J. Appl. Crystallogr.40, 658–674(2007).
41. Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development ofCoot. Acta Crystallogr. D 66, 486–501 (2010).
42. Chen, V. B. et al. MolProbity: all-atom structure validation for macromolecularcrystallography. Acta Crystallogr. D 66, 12–21 (2010).
43. Evans, P. R. & Murshudov, G. N. How good are my data and what is the resolution?Acta Crystallogr. D 69, 1204–1214 (2013).
ARTICLE RESEARCH
Macmillan Publishers Limited. All rights reserved©2015

Extended Data Figure 1 | The role of specific B. thetaiotaomicron PULs andenzymes in utilization of mannan from S. cerevisiae and other yeast species.a, Growth of wild-type B. thetaiotaomicron on Candida albicans mannan andglucose. b, Growth of wild-type B. thetaiotaomicron and the mutant lackingMAN-PUL1 and MAN-PUL3 (DMAN-PUL1/3) on Schizosaccharomycespombe a-mannan. c, Growth of wild-type B. thetaiotaomicron, and theB. thetaiotaomicron mutants lacking MAN-PUL2 (DMAN-PUL2), or all threemannan PULs (DMAN-PUL1/2/3) on S. cerevisiae a-mannan. d, The growthprofile of wild-type B. thetaiotaomicron and the B. thetaiotaomicron mutantlacking bt3774 (Dbt3774) on S. cerevisiae mannan. In panels a, b, c and d, eachpoint on the growth curve represents the mean of three biological replicates.
e, Enzymes at 1 mM at 37 uC were incubated with either undecorated a-1,6-mannan (derived from mnn2 mutant of S. cerevisiae) (lanes 1–3) or mannanfrom S. pombe (lanes 4–9). Lanes 1 and 4, the mannans incubated in the absenceof the enzymes; lanes 2 and 6, mannans incubated with the periplasmicmannanase BT3782; lanes 3 and 7, mannans incubated with the surfacemannanase BT3792; lane 5, S. pombe mannan incubated with the GH97a-galactosidase BT2620; lanes 8 and 9, S. pombe mannan incubated withBT2620/BT3782 and BT2620/BT3792, respectively. Lane 10 galactosestandard; lane 11 a-1,6-mannooligosaccharides: mannose (M1), mannobiose(M2), mannotriose (M3) and mannotetraose (M4).
RESEARCH ARTICLE
Macmillan Publishers Limited. All rights reserved©2015

ppm4.854.904.955.005.055.105.155.20
β-Manr
α-ManrT-α-Man-(1→3)-
T-α-Man-(1→6)-
0
100
200
300
400
500
600
700
156
174
192
210
228
246
264
282
300
318
336
354
372
390
408
426
444
462
480
498
516
534
552
570
588
606
624
642
660
678
696
714
732
750
768
786
804
Sign
al (n
C)
Retention time (sec)
Man
BT3775
BT3775+BT3776
0
100
200
300
400
500
600
70015
617
519
421
323
225
127
028
930
832
734
636
538
440
342
244
146
047
949
851
753
655
557
459
361
263
165
066
968
870
772
674
576
478
380
2
Sign
al (n
C)
Retention time (sec)
Man1,3
BT3775
BT3775+BT3776
αα-Man-(1→3)-[αα -Man-(1→6)]-Manαα-Man-(1→3)-Man
12
3 12
3
αα3 αα6
4
β-Manr
α-Manr
ppm4.854.904.955.005.055.105.155.20
T-α-Man-(1→3)-
8.00 9.00 10.00 11.00 12.00 13.00 14.00 15.00 16.00
2000000
4000000
6000000
8000000
1e+07
1.2e+07
1.4e+07
Time
Abun
danc
e
9.00 9.50 10.00 10.50 11.00 11.50 12.00 12.50
200000
400000
600000
800000
1000000
1200000
1400000
1600000
Time
Abun
danc
e
αα3 Terminal MannoseTerminal Mannose
3-linked Mannose
3,6-linked Mannose
a b
c d
fe
g h
Extended Data Figure 2 | Product profiling of GT32 glycosyltransferasesencoded by MAN-PUL2. a, HPAEC of biosynthetic reactions using mannoseas an acceptor and GDP-a-Man as the donor. Mannobiose is formed in thepresence of BT3775 (black) and mannotriose with BT3775 and BT3776 (red).The blue trace is a mannose standard. b, HPAEC of biosynthetic reactions witha-1,3-mannobiose as the acceptor and GDP-a-Man as the donor. BT3775 isnot capable of extending mannobiose (black). In the presence of BT3775 andBT3776 mannotriose is produced (red). The blue trace is an a-1,3-mannobiosestandard. c, d, MALDI-TOF analysis of the reaction products ofBT3775 1 BT3776 using mannose (c) and a-1,3-mannobiose (d) as anacceptor. e, NMR analysis of the a-1,3-mannobiose substrate. Peaks 1 and 3
correspond to thea-anomer andb-anomer of the mannose at the reducing end,respectively; peak 2 corresponds to the terminal a-mannosyl residue linked toO3 of the mannose at the reducing end. f, NMR analysis of the a-1,3,( a-1,6)-mannotriose BT3776 product. The numbering of the peaks are the same asin e. Peak 4 corresponds to the terminala-mannosyl residue linked to O6 of the3,6-linked mannose at the reducing end. g, Alditol-acetate linkage analysis ofmannobiose produced by BT3775 from mannose. h, Alditol-acetate linkageanalysis of branched (a-1,3),(a-1,6)-mannotriose produced by BT3776 froma-1,3-mannobiose. The green circles indicate the mannose residues present incarbohydrates identified by HPAEC, MALDI-TOF and NMR.
ARTICLE RESEARCH
Macmillan Publishers Limited. All rights reserved©2015

g h
D178/D272
E51/E162
H142/H236
D280/H359
F214/W307
BT3792 GH76 endo-α1,6-mannanase
a b
D258
D259
BT3781 GH125 exo- α 1,6-mannosidase
e f
R234/R188
D241/D195
K176/K131
E174/E129
E174/E129
R109/R64
W107/W62 D110/D65
N348/N302
R481/R405
E439/E393H396/H350
W126/W125W126/W125
R295/R291
H63/H64
E333/E329
E336/E332
+1
+2-1
-2
c dBT3862 GH99 endo-α 1,2-mannosidase
BT3783 mannose-6-phosphatase
RESEARCH ARTICLE
Macmillan Publishers Limited. All rights reserved©2015

Extended Data Figure 3 | The structures of enzymes that play a key role inyeast mannan degradation. a, Overlay of the hydrophobic conserved residuesin the predicted substrate-binding cleft of BT3792 (yellow), BT2949 (cyan) andthe Listeria protein Lin0763 (green; PDB code 3K7X), and the predictedcatalytic aspartates. b, Solvent representation of BT3792 in which the predictedcatalytic residues, Asp258 and Asp259, are coloured green. c, Overlay ofBT3862 (cyan) with a homologue of the enzyme from B. xylanisolvens, BxGH99(green; PDB code 4UTF) in complex with Man-a-1,3-isofagomine and a-1,2-mannobiose (Man residues coloured yellow and isofagomine pink). d, Solvent-exposed surface of the substrate binding cleft of the BxGH99 (teal) ligandcomplex overlaid with BT3862 (grey). The subsites are numbered with thecatalytic residues, Glu 333 and Glu 336, coloured red and the solvent exposedO2 of Man bound at the 22 subsite and O1 and O6 of the Man located at the
12 subsite are coloured bright green. e, Overlay of BT3781 (green; PDB code2P0V) with the substrate and catalytic residues of the Clostridium perfringensGH125 a-mannosidase CpGH125 (cyan; PDB code 3QT9), in which theligand 6-S-a-D-mannopyranosyl-6-thio-a-D-mannopyranose (Man-S-Man) isshown in yellow. f, Solvent-exposed surface of BT3781 in the vicinity of theactive site in which the catalytic residues (Glu 174 and Glu 439) are depictedin green. The position of Man-S-Man is based on the overlay shown in e.g, Overlay of BT3783 with the catalytic and substrate binding residues of atyrosyl-DNA phosphodiesterase (PDB code 4GYZ) in complex with Mg21
(slate-blue sphere) and phosphate (coloured orange). h, A region of the solvent-accessible surface of BT3783 in which the catalytic residues are coloured green.The figure was prepared using PyMOL. A detailed description of the structuresof these proteins is provided in Supplementary Information section 5.0.
ARTICLE RESEARCH
Macmillan Publishers Limited. All rights reserved©2015

aGH92 GH92/GH76
M1
M2
M4
1 2 3 1 2 3
M3
M1
1 32
M2
M4M3
M2
M4M3
M2M2M2M2MMM2M2
M4M4M4M4M4M3M3M3MMM3M
M1
M2
M4
M3
0h 2h 6h 8h 10h 12h 4h 24h 31h 36h 48h 0 10 20 30 40 50 600.0
0.5
1.0
1.5
2.0
T im e (h )A
bs
orb
an
ce
60
0 n
m
M1
M2
M4
M3
1 2 3 4
b
dc
HOOHO
OHOHO
OHHO
O
OpNP
OH
GH76
endo-α-1,6-mannosidaseBt3792 HO
OHO
OHOHO
OHHO
O
OH
OH
HOOHO
OHOHO
OHHO
O OH
OH
productα-anomer (P-α)
H1'
H1
H1'
H1
H1'
H1substrate, S product
β-anomer (P-β)
0 min 8 min 13 min 18 min 30 min 1 day
SH1α
PH1α PH1’α
PH1β
PH1’β
fe
RESEARCH ARTICLE
Macmillan Publishers Limited. All rights reserved©2015

Extended Data Figure 4 | The degradation of yeast mannan byB. thetaiotaomicron in culture and the selected enzymes expressed by thebacterium, and the stereochemistry of the reaction catalysed by GH76 endo-a-1,6-mannanases. a, GH92 a-mannosidases at high concentrations (50mM)were incubated with yeast mannan for 5 h in the absence (labelled GH92)or in the presence (GH92/GH76) of the endo-a-1,6-mannanase BT3782.The GH92 a-mannosidases in this example were BT2199 (1), BT2130(2) and BT3773 (3). The GH76 endo-a-1,6-mannanase only releasesmannooligosaccharides in the presence of BT2199; see also SupplementaryInformation section 4.1. b, B. thetaiotaomicron was grown on yeastmannan or glucose. Yeast mannan was incubated with no bacterium (1),B. thetaiotaomicron previously cultured on yeast mannan (2) andB. thetaiotaomicron grown on glucose (3). The cells were incubated for 5 h at37 uC with the polysaccharide without a nitrogen source and thus were notgrowing. The products released by the B. thetaiotaomicron cells, analysed byTLC, were mediated by the activity of enzymes presented on the surface ofB. thetaiotaomicron, and not through the action of periplasmic mannanasesand mannosidases. The black box highlights very low levels of high molecularweight mannooligosaccharides generated by the cells incubated in yeastmannan. c, B. thetaiotaomicron was cultured for up to 48 h (stationary phase)on yeast mannan. The supernatant of the culture at the time points indicated
were analysed by TLC. In all panels the samples were chromatographed withthe following a-1,6-mannooligosaccharides: mannose, M1; mannobiose, M2;mannotriose, M3; mannotetraose, M4. d, The absorbance of the culture usedin c. e, BT3792 (GH76) endo-a-1,6-mannosidase is a retaining glycosidehydrolase. Enzymatic hydrolysis of 4-nitrophenyl a-D-mannopyranosyl-1,6-a-D-mannopyranoside (S) was monitored by 1H-NMR spectroscopy (500 MHz).The stacked spectra show the reaction progress over time. SH1a is the anomericproton of the reducing end mannopyranoside of the substrate, and SH19a is theanomeric proton of the non-reducing end mannopyranoside. The reactionproceeds with the initial formation of the product, the a-anomer of a-1,6-mannobiose (P-a, peaks PH1a and PH19a), which slowly mutarotates to theb-anomer (P-b, peaks PH1b and PH19b). f, TLC analysis of S. cerevisiae mannanincubated without enzyme (lane 1), BT3774 (lane 2), BT3792 (lane 3), andBT3774 and BT3792 (lane 4). M1–M4 area-mannnooligosaccharide standardsnumbered according to their d.p. GH76 mannanase BT3792 does not attack thebackbone of S. cerevisiae mannan unless the side chains are first removed bythe GH38 a-mannosidase BT3774, confirming that this enzyme cleaves themannose a-1,2-linked to the mannan backbone. The data in a, b and c arerepresentative of two biological replicates, while the data in f are representativeof two technical replicates.
ARTICLE RESEARCH
Macmillan Publishers Limited. All rights reserved©2015
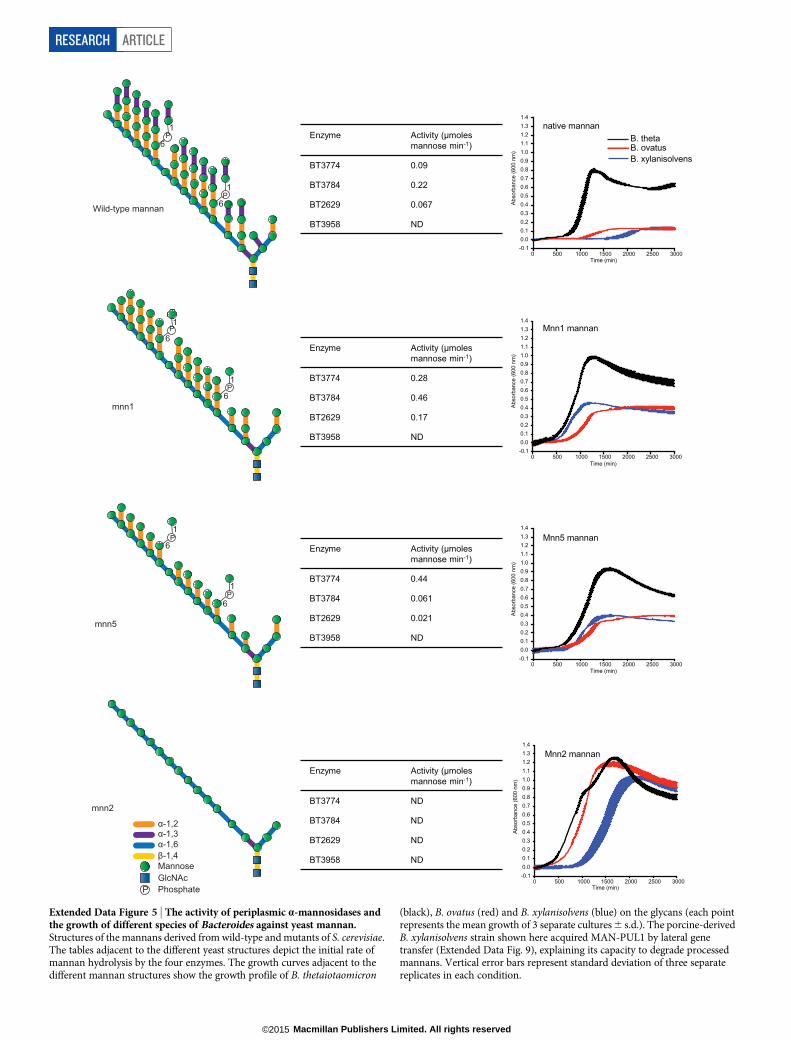
1
6P
6
1P
6
1
1
6P
P
α-1,2α-1,3
β-1,4α-1,6
P PhosphateGlcNAcMannose
0 500 1000 1500 2000 2500 3000-0.10.00.10.20.30.40.50.60.70.80.91.01.11.21.31.4
Time (min)
Abso
rban
ce (6
00 n
m)
native mannanB. thetaB. ovatusB. xylanisolvens
0 500 1000 1500 2000 2500 3000-0.10.00.10.20.30.40.50.60.70.80.91.01.11.21.31.4
Time (min)
Abso
rban
ce (6
00 n
m)
Mnn1 mannan
0 500 1000 1500 2000 2500 3000-0.10.00.10.20.30.40.50.60.70.80.91.01.11.21.31.4
Time (min)
Abso
rban
ce (6
00 n
m)
Mnn5 mannan
0 500 1000 1500 2000 2500 3000-0.10.00.10.20.30.40.50.60.70.80.91.01.11.21.31.4
Time (min)
Abso
rban
ce (6
00 n
m)
Mnn2 mannan
Wild-type mannan
mnn2
mnn5
mnn16
1
1
6P
PP
P
Enzyme Activity (μmoles mannose min-1)
BT3774 0.09
BT3784 0.22
BT2629 0.067
BT3958 ND
Enzyme Activity (μmoles mannose min-1)
BT3774 0.28
BT3784 0.46
BT2629 0.17
BT3958 ND
Enzyme Activity (μmoles mannose min-1)
BT3774 0.44
BT3784 0.061
BT2629 0.021
BT3958 ND
Enzyme Activity (μmoles mannose min-1)
BT3774 ND
BT3784 ND
BT2629 ND
BT3958 ND
Extended Data Figure 5 | The activity of periplasmic a-mannosidases andthe growth of different species of Bacteroides against yeast mannan.Structures of the mannans derived from wild-type and mutants of S. cerevisiae.The tables adjacent to the different yeast structures depict the initial rate ofmannan hydrolysis by the four enzymes. The growth curves adjacent to thedifferent mannan structures show the growth profile of B. thetaiotaomicron
(black), B. ovatus (red) and B. xylanisolvens (blue) on the glycans (each pointrepresents the mean growth of 3 separate cultures 6 s.d.). The porcine-derivedB. xylanisolvens strain shown here acquired MAN-PUL1 by lateral genetransfer (Extended Data Fig. 9), explaining its capacity to degrade processedmannans. Vertical error bars represent standard deviation of three separatereplicates in each condition.
RESEARCH ARTICLE
Macmillan Publishers Limited. All rights reserved©2015

6 0 m in
4 8 m in
3 6 m in
2 4 m in
1 2 m in
0 m in
IP M 1 M 2 M 3 M 4
6 0 m in
4 8 m in
3 6 m in
2 4 m in
1 2 m in
0 m in
IP M 1 M 2 M 3 M 4a
b
M1
M4
M3
M2
BT3792 (Surface)
BT3782 (Periplasmic)
No enzyme
cM1
M2
M3
M4
d
BT2632 BT3781 No enzyme
0 10 20 30 40 50 60 700.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
Time (min)ln
(S0
/St)
0 10 20 30 40 50 60 700.0
0.5
1.0
1.5
2.0
Time (min)
ln (S
0/S
t)
Mannose (100 μM)e
Untreated mnn2 mannan
BT3781 vs mnn2 mannan
BT2632 vs mnn2 mannan
Extended Data Figure 6 | The activity of GH76 a-mannanases and GH125a-mannosidases a, b, BT3792 and BT3782, respectively, were incubated witha-1,6-mannotetraose at a concentration=KM. Substrate depletion wasmeasured using HPAEC and the rate (right of a and b) enabled kcat/KM to bedetermined. c, BT3792 and BT3782 were incubated with unbranched yeastmannan (derived from the S. cerevisiae mutant MNN2). The yeast mannan at0.1% was incubated with the two GH76 a-1,6-mannanases for 1 h at 37 uC in
50 mM sodium phosphate buffer, pH 7.0. The limit products were analysed byTLC. a-1,6-Mannooligosaccharides are identified by their degree ofpolymerization (M1, mannose; M2, mannobiose; M3, mannotriose; M4,mannotetraose); IP, injection peak. d, e, BT2632 and BT3781 at 100 nM wereincubated with 1 mg ml21 of the debranched mannan for 1 h in the bufferdescribed above. TLC analysis (d) and HPAEC traces (e) of the reactions areshown. The data in c and d are representative of two technical replicates.
ARTICLE RESEARCH
Macmillan Publishers Limited. All rights reserved©2015

Man5GlcNAc Man6GlcNAc
a
0 min10 min20 min30 min40 min60 min
0 min10 min20 min30 min40 min60 min
100 nM BT3994 + Man GlcNAc -AB 10 μM BT3994 + Man -ABb
BT3990
1
2
cd
g
0h 16h RP
e
0 250 500 750 1000 1250 1500 1750 2000 2250 25000.0
0.1
0.2
0.3
0.4
0.5
0.6
Time (minutes)
Abso
rban
ce (6
00nm
)
WT BtΔbt3993
BT3993BT3992
BT3991BT3990BT3989BT3988BT3987BT3986BT3985BT3983 BT3984 BT3994GH92GH92GH92GH18 anti-s ECF-s
Glycoside hydrolase SusD-like SusC-likeSGBP Regulator Unknown
f
Extended Data Figure 7 | HMNG deconstruction by B. thetaiotaomicron.a, Structure of the HMNG PUL. Genes drawn to scale with their orientationindicated. Genes encoding known or predicted functionalities are colour-codedand, where appropriate, are also annotated according to their CAZy glycosidehydrolase (GH) family number. SGBP represents a surface glycan bindingprotein. b, BT3994 was incubated with a-1,6-mannotetraose (Man4) or thehigh mannose N-glycan Man5GlcNAc2, with both oligosaccharides labelledwith 2-aminobenzamide (AB). At the indicated time points aliquots wereremoved and analysed by HPAEC using a fluorescence detection system. WhileMan5GlcNac2-AB was hydrolysed by BT3994, the enzyme was not activeagainst Man4-AB. c, Chicken ovalbumin was incubated with buffer (1) or 1 mMof BT3987 (2) in 20 mM Na-HEPES buffer, pH 7.5, for 5 h at 37 uC, and thesoluble material was permethylated and analysed by MALDI-TOF massspectrometry. The high mannose N-glycans released are labelled. d, Westernblot of B. thetaiotaomicron cells cultured on yeast mannan that were untreated
with proteinase K (0 h) or incubated with 2 mg ml21 proteinase K for 16 h(16 h). The lane labelled RP contained a purified recombinant form of BT3990.The blots were probed with antibodies against BT3990. The data in d arerepresentative of two biological replicates. e, f, Representative isothermalcalorimetry titrations for BT3984 titrated with Gal-b1,4-GlcNAc (LacNAc;25 mM) (e), and for BT3986 titrated with mannose (50 mM) (f). The top half ofeach panel shows the raw isothermal calorimetry titration heats; the bottomhalf, the integrated peak areas fitted using a single binding model by MicroCalOrigin software. ITC was carried out in 50 mM Na-HEPES, pH 7.5 at 25 uC.The affinities and thermodynamic parameters of binding are shown inSupplementary Table 5. g, Growth profile of wild-type B. thetaiotaomicron(WT Bt; black) and the mutant Dbt3993 (red), which lacks the extra-cellularfactors regulator gene of HMNG-PUL, cultured on Man8GlcNAc2 (each curveshows the mean 6 s.d. of 3 separate cultures).
RESEARCH ARTICLE
Macmillan Publishers Limited. All rights reserved©2015

Extended Data Figure 8 | Metagenomic analysis of the occurrence of theyeast mannan PULs in humans. Abundance of Bacteroides mannan PULs inhumans from a survey of metagenomic sequencing data from a total of 250adult human samples (211 healthy, 27 ulcerative colitis, 12 Crohn’s disease; seeMethods for references). Data sets were individually queried by BLASTusing either each entire mannan PUL (PULs 2,3) or a sub-fragment that wastrimmed to eliminate cross-detection of other species genomes beyondB. thetaiotaomicron and porcine B. xylanisolvens (PUL1; see Methods foradditional search details). Each horizontal line represents the presence of a hit
in a single individual. The leftmost column summarizes the total mannan PULcontent in each person (annotated according to the colour key in the upperright corner). The mannan PUL frequency across all 250 samples is shown atthe bottom for each condition and is compared to the frequency of several otherPULs implicated in xyloglucan and porphyran utilization. Graph at far rightillustrates the variation in sequencing depth for each sample/study; black linesshow the average depth in megabase pairs (Mbp) for each study; the light greyline shows the depth for each individual sample.
ARTICLE RESEARCH
Macmillan Publishers Limited. All rights reserved©2015

Extended Data Figure 9 | Presence of a conjugative transposon (cTn) thatcontains MAN-PUL1 in the genomes of porcine B. xylanisolvens strainsand the mannan presented to these organisms. a, Shown across the top is aschematic of a cTn that has been integrated into the 39 end of a tRNAphe genein B. thetaiotaomicron strain VPI-5482. Integration is mediated by a 22 bpdirect repeat sequence that is contained in tRNAphe and repeated again at theother side of the cTn (right insertion site). The location of B. thetaiotaomicronMAN-PUL1 is denoted within the larger cTn element using a colour schemeidentical to Fig. 1a. The lower panel shows an expanded view of the MAN-PUL1 locus in five sequenced strains of B. xylanisolvens from the faeces of pigsfed a diet enriched with distillers grains that were fermented with yeast.A nearly identical copy (both by amino acid homology and syntenicorganization) of this genomic region is present in B. thetaiotaomicron and the
porcine B. xylanisolvens strains. Although the draft genomes of the B.xylanisolvens strains contains gaps in all five assemblies at the left side of theMAN-PUL1, the right side insertion site was resolved in all genomes,suggesting that the B. xylanisolvens loci were also transferred by lateral transferat some point in the history of these strains. b, Forty-three different strainsfrom five Bacteroidetes isolated from animal guts (each indicated with a solidcircle) were inoculated into minimal media containing S. cerevisiae mannan asthe sole carbon source. The growth of the cultures was measured over 48 hby recording the optical density at 600 nm. c, TLC analysis of the productsgenerated by incubating BT3774 and BT3792 with a-mannan extracted fromthe distillers grain fed to the pigs from which the B. xylanisolvens were isolated.The data are representative of two technical replicates.
RESEARCH ARTICLE
Macmillan Publishers Limited. All rights reserved©2015

Extended Data Figure 10 | In vivo and in vitro expression of the mannanPULs. a, Level of susC-like transcripts derived from MAN-PUL1 (BT2626),MAN-PUL2 (BT3788) and MAN-PUL3 (BT3854) from B. thetaiotaomicronin monocolonized gnotobiotic mice fed a glycan-free diet deficient inB. thetaiotaomicron-digestible glycans (red), the same diet with added yeastmannan (1% w/v in drinking water) as the only usable polysaccharide (green),and a diet containing 50% bread (blue). The levels of the susC transcripts werequantified relative to the same mRNA species in B. thetaiotaomicron culturedin vitro on glucose minimal medium (MM-G). Note that in all cases, expressionof MAN-PUL genes is equally high in vivo. b, Transcription of the samemannan susC-like genes in response to increasing concentrations of yeast
mannan in the media after 30 min exposure. The prototypic susC (BT3701)involved in starch metabolism is shown for comparison. c, An identicalexposure experiment to that shown in b, except that glucose-grownB. thetaiotaomicron cells were exposed to aqueous extracts of the cereal graindiet (natural diet) fed to mice before the experiment, or the digestible glycan-free control diet (glycan-free diet) used as a base for all feeding treatments.Exposure was conducted for 30 min to determine if any diet extract containedcontaminating levels of mannan that could be detected by B. thetaiotaomicroncells; inclusion of purified mannan (5 mg ml21) in addition to theglycan-free diet served as positive controls. In all panels, the results representthe mean of 3 biological replicates and error bars represent s.d.
ARTICLE RESEARCH
Macmillan Publishers Limited. All rights reserved©2015

CORRECTIONS & AMENDMENTS
CORRIGENDUMdoi:10.1038/nature14334
Corrigendum: Human gutBacteroidetes can utilize yeastmannan through a selfishmechanismFiona Cuskin, Elisabeth C. Lowe, Max J. Temple, Yanping Zhu,Elizabeth A. Cameron, Nicholas A. Pudlo, Nathan T. Porter,Karthik Urs, Andrew J. Thompson, Alan Cartmell,Artur Rogowski, Brian S. Hamilton, Rui Chen,Thomas J. Tolbert, Kathleen Piens, Debby Bracke,Wouter Vervecken, Zalihe Hakki, Gaetano Speciale,Jose L. Munoz-Munoz, Andrew Day, Maria J. Pena,Richard McLean, Michael D. Suits, Alisdair B. Boraston,Todd Atherly, Cherie J. Ziemer, Spencer J. Williams,Gideon J. Davies, D. Wade Abbott, Eric C. Martens& Harry J. Gilbert
Nature 517, 165–169 (2015); doi:10.1038/nature13995
In this Article focusing on the selfish metabolism of yeast mannan byBacteroidetes, we also described a polysaccharide utilization locus (PUL)responsible for the degradation of high mannose mammalian N-glycan(HMNG) but omitted to cite two relevant papers1,2, for which we apol-ogise. Both studies describe a model for the degradation of complexbiantennary N-glycans by Bacteroidetes in which the degradative enzymesare encoded by PULs. These studies1,2 provide examples of how PULscan orchestrate N-glycan metabolism in addition to the HMNG PULwe describe in this Article. In all three papers it is proposed that N-glycandepolymerization occurs primarily in the periplasm.
1. Renzi, F. et al. The N-glycan glycoprotein deglycosylation complex (Gpd) fromCapnocytophaga canimorsus deglycosylates human IgG. PLoS Pathog. 7,e1002118 (2011).
2. Nihira, T. et al. Discovery of b-1,4-D-mannosyl-N-acetyl-D-glucosaminephosphorylase involved in the metabolism of N-glycans. J. Biol. Chem. 288,27366–27374 (2013).
G2015 Macmillan Publishers Limited. All rights reserved
3 8 8 | N A T U R E | V O L 5 2 0 | 1 6 A P R I L 2 0 1 5