heterotrophic planktonic microbes: virus, bacteria ... · heterotrophic planktonic microbes: virus,...
TRANSCRIPT

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
Heterotrophic Planktonic Microbes VirusBacteria Archaea and Protozoa
JED A FUHRMAN AND DAVID A CARON
422BACKGROUND AND HISTORICALDEVELOPMENT
Marine Microbial Ecology into the 1970sAlthough heterotrophic marine microorganisms in the opensea have been studied since the late 1800s and early 1900sby pioneers like Fischer Haeckel and Calkins (1ndash3) thecontribution of these diminutive species to the food websof oceanographic systems was not fully recognized untilnearly a century later Attention was drawn to the largerand more conspicuous taxa of photosynthetic protists (egdiatoms and many dinoflagellates) early in the history ofbiological oceanography In contrast little was known ofthe abundances or activities of bacteria in the ocean untilthe past 50 years and most early studies of marine hetero-trophic protists (the protozoa) focused on morphologicaldescriptions and natural history of larger species of thesetaxa rather than their functional roles in marine foodwebs Improvements in microscopy methods used to observebacteria in seawater during the 1930s to 1950s indicatedthat bacterial abundances were several orders of magni-tude greater than previously believed However confusionremained during regarding the ecological significance ofmarine bacteria because these early abundance estimateswere often hundreds of times greater than counts made bycultivation techniques (4)
During the 1960s and 1970s however the metabolicactivity of aquatic bacterial assemblages was demonstratedby the uptake of radioactive organic compounds in marineand freshwater samples (eg [5 6] and others) These studiesdemonstrated that organic compounds were readily turnedover by microorganisms in aquatic ecosystems and thatmicrobial communities appeared to be quite dynamic Dur-ing that same period abundant and diverse assemblages ofprotozoa were demonstrated from a wide array of aquatic eco-systems using more quantitative approaches for the collec-tion observation and enumeration of eukaryotes (7ndash10)Together these observations implied the presence of anactive and complex microbial community that might beresponsible for much of the metabolic activity in marineecosystems This hypothesis stood in contrast to broad oce-anographic models of the time that included the bacteriaonly as a sink for nonliving organic matter on the sea floorand largely ignored the potentially important roles of heter-otrophic protists (11)
The Microbial Loop RevolutionPomeroyrsquos prescient analysis ldquoThe oceanrsquos foodweb a chang-ing paradigmrdquo (12) had a significant impact toward trans-forming the field This publication is often cited by manymicrobial ecologists as a turning point in our understandingof the structure and function ofmarine ecosystems In it Pom-eroy pointed to lines of evidence that were starting to emergeshowing that the smallest members of the food web includingheterotrophic bacteria cyanobacteria and small protists(algae and protozoa lt20 microm) were probably responsible fora large fraction of important system activities such as overallrespiration photosynthesis and organic matter turnover(ie the ingestion of food particles by protozoa or the uptakeof dissolved substances by bacteria) Early studies of bacterialutilization of dissolved organic matter (eg 1) and subse-quent studies of community metabolism into the 1980s(eg 14) indicated that this process accounted for a surpris-ingly high percentage of total organic matter turnover Sim-ilarly Fenchel and Jorgensen (15) pointed out thatapproximately 10ndash30 of primary productivity might bereleased as dissolved organic matter which was then pre-sumed to be taken up by bacteria who respired a portionand passed the rest on to the next trophic level composedlargely of protozoa In this scenario protozoa constituted amechanism by which bacterial biomass reentered the ldquoclassi-calrdquo food web including the metazoan zooplankton and nek-ton and also served as a means of remineralizing some of thebacterial biomass back to inorganic nutrients and carbondioxide for subsequent utilization by primary producersAlthough these conclusions were later found to be generallycorrect the concepts were a radical departure from the estab-lished general biological oceanographic thinking of the time
The introductions of epifluorescence microscopy of fluo-rescently stained cells (16) and polycarbonate filters to posi-tion the cells all in a single optical plane (17) were significanttechnological advancements that allowed much more accu-rate estimates of the total number of microorganisms presentin natural water samples Using this method it was deter-mined that bacteria are typically present at abundances of 1million cells per ml in near-surface seawater This number issurprisingly constant around the world with most variationfalling within a factor of 10 worldwide Appreciation of theimportance of medium to large protozoa (ie gt20 microm) inoceanic food webs was made possible in the late 1960s andearly 1970s largely through the pioneering work of Beers
doi1011289781555818821ch422
422-1
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
et al (7 18) However as with marine bacteria epifluores-cence microscopy facilitated the observation of small single-celled eukaryotes during the 1970s and 1980s and enabledeasy discrimination of protists without chloroplasts (proto-zoa) from those with chloroplasts (algae) based on the auto-fluorescence of photosynthetic pigments (19ndash21) Thedevelopment and refinement of this approach for eukaryoticmicroorganisms was instrumental in establishing the standingstocks of small protozoa which typically occur at abundancesof tens to thousands per ml in most marine ecosystems Thedevelopment of microscopy approaches for larger protozoasuch as heterotrophic dinoflagellates and ciliates (10 22)has been equally important in documenting abundances ofthese taxa that range up to tens per ml in much of the worldrsquosoceans
Estimates of the overall biomass of various microbialassemblages and techniques to measure rate processes (egrates of growth substrate uptake prey consumption) beganto appear in the late 1970s and 1980s and the refinementof these estimates and measurements continue to the presentday Early attempts to measure bacterial growth rates in sea-water involved ldquoindirectrdquo methods such as relating the fre-quency of dividing cells to rates of division in culturedstrains (23) Isotope-uptake based approaches specificallythe incorporation of radioactively labeled thymidine intoDNA (24 25) andor the incorporation of leucine into pro-tein (26) have become the most commonly used methods(see ldquoEstimating lsquoBacterialrsquo Biomass and lsquoBacterial Produc-tionrsquordquo) These methods have indicated that bacterial dou-bling times can be on the order of one day in coastaltemperate waters Combined with estimates of bacterial bio-mass these results led to the conclusion that bacteria mustbe consuming a substantial proportionmdashon the order of50mdashof the total system primary productivity A similarconclusion was reached using direct estimation of microbialrespiration by careful measurements of oxygen concentrationchanges (micro-Winkler method) in seawater that had beenprefiltered through 5 microm pore filters to remove animals andmany of the protists (14)
During this same period small protozoa (primarily flagel-lates and ciliates) were gaining recognition as importantconsumers of bacteria in the marine plankton and benthos(27ndash29) An increasing volume of experimental work dem-onstrated a dominant role for small bacterivorous protozoaas a mechanism for removing bacterial production andrepackaging bacteria into larger particles that might be con-sumed by metazoan zooplankton Also it became recognizedaround this time that a significant fraction of the phytoplank-ton biomass and production was consumed directly by herbiv-orous protozoa rather than by metazoan zooplankton such ascopepods (30 31) Consequently heterotrophic protists wereacknowledged as an important food source for a variety ofmetazoan zooplankton and numerous experimental studiessubsequently demonstrated this trophic connection (32 33)
These observations were synthesized in a second bench-mark paper (34) The latter publication marked the begin-ning of the widespread recognition and use of the termldquomicrobial looprdquo in marine planktonic systems a conceptthat emphasizes the remarkable importance of the tiniestorganisms as well as dissolved organic matter as an intermedi-ate in material and energy transfer in aquatic ecosystems Anupdated illustration of this basic concept is shown in Fig 1
Definitions and ConceptsAbundance the number of individuals in a sample or a
population
Algae protists that exhibit phototrophic nutrition Likeprotozoa algae span a wide size range (lt1 to gt200microm) and have generally been referred to as phototrophicpico- nano- or microplankton
Amensalism interaction where members of a speciesinflicts harm to another species without any costs or ben-efits received by the other
Autotroph an organism that uses carbon dioxide as itssource of structural carbon
Biomass the mass of living organisms within a populationcommunity or ecosystem
Chemolithotroph an organism that uses reduced inorganicmolecules as its energy source
Chemoorganotroph an organism that uses reduced organiccarbon as its energy source
Commensalism interaction between species where onebenefits from the other but the other is not affected
Competition species-species interactions that have a nega-tive effect on both species
Cyanobacteria Prokaryotic photosynthetic organisms thatcontain chlorophyll a and generate oxygen during photo-synthesis The free-living ancestors of primary chloroplasts
Exploitation species-species interactions that have a nega-tive effect on one species while benefiting the othermdashcan include both parasitism and predation
Heterotroph an organism that uses preformed organic car-bon as its source of structural carbon also heterotrophicor heterotrophy when applied to metabolism
Mixotroph any of a number of types of organisms that com-bine (in one organism) multiple metabolic types asdescribed above For example a protist that consumes bac-teria as prey (heterotroph) but also contains functioningchloroplasts (phototroph) will often be referred to as amixotroph Similarly an archaeon that oxidizes ammoniaforenergy (chemolithotroph)butusesaminoacidstobuildproteins (heterotroph) could be considered a mixotroph
Mutualism interaction between species where both benefitfrom each other
Phototroph an organism that uses light as its energy sourcefor production of ATP (or to produce proton gradients inthe case of rhodoposin-based phototropy) and some-times also reducing power fromwater (in cyanobacteria)
Phytoplankton the photoautotrophic component of theplankton including cyanobacteria and a large numberof eukaryotic phyla that contain chloroplasts
Protists eukaryotic species that can exist as a single cellother than a spore gamete or zygote (although thereare many that form colonies)
Protozoa protists that exhibit heterotrophic nutrition Pro-tozoa span a wide size range (asymp2 to gt200 microm) and havegenerally been referred to as nano- or microzooplankton
Relative abundance (and the related term evenness) thecontribution of each species or operational taxonomicunit to a community
Species diversity a complex concept composed of speciesrichness and relative abundance
Species richness the number of different species or opera-tional taxonomic units present in a sample habitat orenvironment
Stoichiometry Studies that involve calculation of the rela-tive quantities of elements or compounds for exampleCNP ratios
422-2 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
Symbiosis the intimate living together of two kinds oforganisms especially if such an association is of mutualadvantagemdashtoo vague to be of use in quantitativedescriptions of population interactions but very usefulin indicating a close association among organisms
Syntrophy a metabolic mutualism where one species usesthe waste product produced by the other and in so doingallows both metabolic pathways to be energeticallyfeasible
Zooplankton Planktonic eukaryotes that consume otherplankton Includes single-celled organisms (protozoaor protists) and metazoans and some that are planktoniconly as larvae
Estimating ldquoBacterialrdquo Biomass and ldquoBacterialProductionrdquo Definitions and MethodsAquatic microbiologists tend to use the term ldquobacteriardquowith alowercase ldquobrdquo to describe organisms that appear to be prokary-otic by microscopymdashthat is organisms with no membrane-bound nucleus They include members of the taxonomicdomains Bacteria and Archaea (see ldquoBacteria and Archaeardquo)Organisms within and between these domains differ in manybiochemical and genetic aspects but they tend to look similarby traditional epifluorescence microscopy Special methodssuch as different versions of fluorescence in situ hybridization(FISH) are required to distinguish individual members ofthese domains microscopically (35ndash37)
The term ldquobacterial productionrdquo here refers to heterotro-phic production of biomass by bacteria It is meant to include
production of nonphotosynthetic bacterial biomass based onthe heterotrophic consumption of preformed organic matter(ie organic matter in various forms that has been producedprimarily by phytoplankton)
Bacterial biomass is usually determined by convertingdirect counts of bacteria using an estimate of the amount ofcarbon per cell Direct counts are most commonly done byepifluorescence microscopy with stains such as acridineorange 406-diamidino-2-phenylindole or SYBR green I(17 38 39) Special procedures are usually applied for sedi-ment samples and samples containing large numbers of bacte-ria attached to particles (40) SYBR green I also permits directvisualization and counts of viruses in the same preparationIncreasingly direct bacterial counts in seawater sampleshave been performed by flow cytometry of fluorochrome-stained cells (41 42) a method that allows separate countsof cyanobacteria such as Synechococcus and Prochlorococcuswhich have unique fluorescent signatures due to their photo-synthetic pigments and which can sometimes make up a sub-stantial fraction of the total number of bacteria (43) Flowcytometry is rapid and has a statistical advantage in that it typ-ically observes thousands of prokaryotic and minute photo-synthetic eukaryotic cells rather than the hundreds countedmicroscopically drawbacks include the cost of the instrumentand the fact that cells attached to each other or to other par-ticles are counted as one Bacterial carbon per cell has beenestimated in a variety of ways most commonly from a deter-mination of cell volume and carbon density per unit volumeThese numbers are difficult to obtain accurately for nativemarine bacteria which are very small typically 05 μm in
FIGURE1 An early vision of the ldquomicrobial looprdquo and its connections to the classical grazing food chain via dissolved organicmatter (DOM)flux and particulate trophic transfer with viruses included as a side loop Modified from (34) Large gray arrows indicate the flow of organiccarbon and energy into higher trophic levels of the food web with recognition of the important roles for heterotrophic microbes (bacteriaand protozoa) in this process Large stippled arrows indicate the production of DOM via excretion and trophic interactions (not all groupsare represented) Thin dotted arrows indicate mineralization of major nutrients contained in organic matter respired by consumers Whitearrows indicate bacteria lysis by viruses and DOM released by that process doi1011289781555818821ch422f1
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-3
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
diameter (range is about 02ndash1 μm for free-living unicells)Published estimates of bacterial carbon per cell vary widelyand probably constitute the greatest uncertainty with estimat-ing bacterial biomass in natural samples Typical estimates ofthe carbon content of a bacterium range from 7 to 50 fg C(1 fg is 10minus15 g) with most open ocean estimates near 10ndash20 fg C per cell and coastal ones about double that (44)Thus in a typical mesotrophic ocean environment with109 bacteria per liter and an average per cell C contentof 15 fg bacterial biomass would be approximately 109 times15 times 10minus14 = 15 times 10minus5 g C per liter or 15 μg C per liter
Bacterial production is most often measured by incorpora-tion of tritiated thymidine into DNA (24 45) or tritiated leu-cine into protein (26 46) Thymidine and leucine areintracellular precursors of DNA and protein respectively soincorporation of these precursors can be used to estimatethe total rates of synthesis of themacromolecules DNA is syn-thesized for cell division and protein synthesized roughly inproportion to total biomass someasuring their rates of synthe-sis is presumed to track production Both methods have beencalibrated on the basis of theoretical considerations as well aspurely empirical approaches and both methods yield similarresults (44) Leucine has a lower detection limit so it is pre-ferred for slower rates The results of thymidine incorporationare most often presented as cells produced per liter per hourwhich can be converted to a carbon production rate via esti-mates of carbon per cell as noted Leucine incorporation maybe used to calculate cell production as well as biomass produc-tion directly because protein is a major biomass constituent
Geographic and Temporal Distributions ofMicrobial Biomass and ActivityBacterial abundances by epifluorescence microscopy showthat bacteria are present in most marine euphotic zone
environments at approximately 106 cells per ml Samplesfrom around the world rarely vary by more than threefoldfrom this typical value (ie rarely lt3 times 105 or gt3 times 106)which is extraordinary compared to phytoplankton and zoo-plankton whichmay vary by several orders of magnitude overthe same spatial scales However despite this remarkable gen-eral predictability there is significant variation across bothspace and time More nutrient-rich eutrophic environmentstend to have more bacteria (sometimes gt107 per ml 47) andoligotrophic open ocean environments have less (summar-ized below) Although bacterial assemblages in warm temper-ate coastal waters may have doubling times as short as 1 daythis is at the rapid end of the spectrum of in situ growth ratesBacterial assemblages in the open sea especially in oligotro-phic environments have average generation times typicallyof a week or perhaps more (see Table 1) These abundancesgenerally apply to the euphotic zone and bacteria in colderdarker waters have substantially lower abundances and slowergrowth rates than those of surface waters (48) Benthic bacte-ria also exhibit fairly constant abundances across wide geo-graphic ranges but benthic bacteria occur at much higherdensities because of the organic-rich and particle-ladennature of the environment (an average of asymp109 per ml fluidvolume is typical 49) This value is three orders of magnitudegreater than abundances in the water column implying thatthe abundances of benthic bacterial assemblages are regulatedby a different set of parameters than planktonic assemblages
Bacterial biomass and productivity vary temporally on anumber of scales ranging from diel (50) to seasonal (51) orinterannual (52) On time scales of hours bacterial abun-dance and production have been shown to often peak inthe middle of the day and be low in the middle of the night(50 53ndash55) This pattern has been interpreted as a tight cou-pling between the production of labile organic compoundsvia photosynthesis and bacterial growth on one hand and
TABLE 1 Bacterioplankton properties in relation to phytoplankton in the open sea as compiled by Ducklow (44)
Property N Atlantica Eq Pac-Sprb Eq Pac-Fallc Sub N Pacd Arabiane Hawaiif Bermudag Ross Seah
Euphotic zone m 50 120 120 80 74 175 140 45
Biomass (mg C mminus2)
Bacteria 1000 1200 1467 1142 1448 1500 1317 217
Phytoplankton 4500 1700 1940 1274 1248 447 573 11450
BP 02 07 075 09 12 36 27 002
Production (mg C mminus2 dminus1)
Bacteria 275 285 176 56 257 Nd 70 55
Phytoplankton 1083 1083 1548 629 1165 486i 465 1248
BP 025 026 011 009 022 Nd 018 004
Growth rates (dminus1)
Bacteria 03 013 012 005 018 Nd 005 025
Phytoplankton 03 064 08 05 093 11 081 011
BP 1 02 015 01 019 Nd 006 23
Notes All bacterial biomass estimates based on 20 fg C per cell Data may overestimate heterotrophic bacterial biomass as a consequence of lower C per cell orinterference by Prochlorococcus and Archaea Production estimated from 3000 g C per mole leucine incorporated
aEastern North Atlantic spring phytoplankton bloom 47 N 20 W May 1989 n = 13bEquatorial Pacific 0 N 140 W MarchndashApril 1992 n = 8cEquatorial Pacific 0 N 140 W SeptemberndashOctober 1992 n = 19dSubarctic North Pacific 45 NeNorthwest Arabian Sea 10ndash20 N 165 E JanuaryndashDecember 1995 n = 21fHawaii Ocean Time Series (HOT) 1995ndash1997 n = 21 (httphahanasoesthawaiieduhothot_jgofshtml)gBermuda Atlantic Time Series (BATS) 1991ndash1998 n = 106 paired comparisons The ratios are means of the ratios not ratios of the means BP calculated
from thymidine incorporation (16 times 1018 cells per mole incorporated)hRoss Sea Antarctica 76 S 180 W 1994ndash1997i1989ndash1996 n = 64
422-4 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
bacterial mortality via grazers or viral lysis on the other handIt is consistent with some measurements of extremely rapidturnover sometimes several times per hour in rich coastalwaters of labile dissolved organic compounds such as dis-solved free amino acids (56) and might also indicate greaterpredation pressure by protozoa during the night On longertime scales of weeks to months bacteria show distinct sea-sonal patterns For example in temperate coastal waters bac-terial biomass and production increase considerably insummer months compared with winter However bacteriado not typically show a significant increase during earlyldquospringrdquo blooms in temperate waters when water is still verycold (51) It has been hypothesized that this phenomenonis the result of the suppression of the rate of substrate uptakeby temperate bacterial assemblages at low temperature (57)However while temperature probably has the effect of settinga limit on maximal growth rates (as for phytoplankton [58]and protozoa [59]) temperature alone does not appear to bethe main factor controlling growth of marine bacteria undermost circumstances (see ldquoLight Temperature and Pressurerdquo)
It would be overly simplistic to think that all the bacteriaand archaea in a sample or habitat have the same level ofactivity per cell but it is also easy to think of measured activ-ities as characteristic of all members of a microbial assem-blage So the question arises are most of the cells active ata similar level or are some hyperactive while others are com-pletely dead or moribund This question has been addressedseveral ways includingmicroautoradiography selective stain-ing ldquodirect viable countsrdquo (where nutrients are added to seewhat part of the community grows) and in situ hybridization(eg 60) Based on these contrasting approaches it appearsthat a continuum of activity exists within bacterial assemb-lages from truly dead (cannot be revived) to extremely activeA reasonable interpretation of the existing data is that undertypical conditions a small percentage of the marine bacterialcells perhaps 10ndash20 are generally inactive or dead the plu-rality or majority of cells perhaps 25ndash75 are intact andhave some moderate level of activity and a small percentageperhaps 5ndash20 are highly active It is useful to consider thisspectrum conceptually and numerically when modelingmicrobial processes
Comparisons of bacterial and phytoplankton biomasswithin planktonic ecosystems show that these are positivelycorrelated across broad scales Analyses of marine and fresh-water samples from several studies (61 62) have shown thatbacterial abundance increases with chlorophyll concentra-tion at least at the level of a log-log relationship Similarlybacterial abundances and the abundances of small protozoacorrelate over broad spatial and temporal scales (63) Theserelationships are sensible in that on the largest scale primaryproduction is the source of organic material that fuels hetero-trophic bacterial activity and bacteria constitute the prey ofmany small protozoa Individual data sets also have sometimesshown strong correlations between bacterial abundance andchlorophyll (eg 45) but variability in this relationshipover short temporal or spatial scales is to be expected It wouldpresumably be a consequence of rapid short-term changes inthe rate of substrate supply as well as the normal oscillatorynature of predator-prey relationships between bacteria andtheir consumers
Interestingly the extrapolation of the positive log-log rela-tionship between bacterial and phytoplankton biomassto environments with very low chlorophyll concentrations(eg ultra-oligotrophic oceans) indicates that bacterial bio-mass may exceed phytoplankton biomass in these situationsWhile this conclusion is obvious for the deep sea where there
is no photosynthesis this situation is also often true in oligo-trophic surface waters Measurements made in oligotrophicwaters bear out the high bacterial contribution to totalbiomass (64 65) Moreover Cho and Azam (66) confirmeda linear relationship between the log of chlorophyll and logof bacterial abundance but only at chlorophyll concen-trations above approximately 05 μg per liter Below thatconcentration bacterial abundance did not correlate signifi-cantly with chlorophyll It should be noted that subsequentanalyses have revealed that early epifluorescence measure-ments of bacterial biomass included the common cyanobacte-rium Prochlorococcus which can make up to 20 of totalbacterial numbers (67) Nonetheless heterotrophic bacterialbiomass is a major fraction of the living biomass of all plank-tonic ecosystems
The geographical and temporal distributions of marineprotozoa are much more varied than those of the bacteriaAs an all-inclusive group protozoa generally occur in plank-tonic ecosystems at abundances ranging from 10 s to 1000 sper ml Abundances in benthic ecosystems can be one tothree orders of magnitude higher commensurate with thehigher abundances of bacteria in those ecosystems Howeverit is important to remember that like the term ldquolsquobacteriardquo theterm ldquoprotozoardquo is a rather artificial conglomeration of evolu-tionarily and ecologically divergent taxa (see ldquoThe Changingand Complex World of Eukaryote Phylogenyrdquo) Thus theabundances of specific lineages of bacteria or protozoa mayshow spatial (or temporal) variability that is considerablygreater than the variability characteristic of these overarchinggroupings
The Changing and Complex World ofEukaryote PhylogenyNot that long ago textbooks still divided eukaryotic organ-isms into four major kingdoms (Animalia Plantae Fungiand Protista) while prokaryotic organisms were placed intoa single kingdom theMonera (68)Within this scheme pro-tists (eukaryotic organisms that can exist as single cells) weredivided into two subkingdoms (algae and protozoa) based ontheir basic nutritional mode a carryover from the historicaldistinction between single cells with ldquoanimal-likerdquo or ldquoplant-likerdquo nutrition This distinction presupposed a basic evolu-tionary divergence among protists into species that retaineda heterotrophic phagocytotic mode of life (protozoa) andthose that abandoned phagocytosis for a photosyntheticmode of life (algae)Moreover the presenceabsence of chlor-oplasts was a feature that could be easily distinguished by earlymicroscopists
The five-kingdom classification system of Whittaker wasrecognized as an improvement over previous classificationschemes but it posed a number of problems relating toprotists For example the distinction between single-celledand multicellular eukaryotes was somewhat arbitrary Moreimportant the division of protists based on whether theywere heterotrophic or photosynthetic was clearly not anappropriate feature if the classification was to recapitulateevolutionary relationships We now know that chloroplastacquisition and loss has occurred several times in the bio-logical history of our planet (69) giving rise to some closelyrelated protistan taxa that differ largely in the presence orabsence of a chloroplast Further complicating the matterwithin many protistan lineages there are species that possesschloroplasts and carry out photosynthesis (phototrophy)but also possess the ability to ingest and digest prey (hetero-trophy 70ndash72) Some heterotrophic protists even ingest
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-5
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
phytoplankton prey and retain the chloroplasts of their prey ina functional state for a limited amount of time (kleptidoplas-tidy73) Various forms and degrees of mixotrophy (mixedphototrophic and heterotrophic nutrition) are commonamong a number of algalprotozoan lineages (74ndash77) UnderWhittakerrsquos scheme phytoplankton ecologists studying a lin-eage of microalgae might have had little familiarity withclosely related heterotrophic species while protozoologistsstudying a particular protozoan groupmight have known littleabout closely related photosynthetic species
One might expect given these caveats that the termsldquoalgaerdquo and ldquoprotozoardquo are no longer used In fact the termldquoprotozoardquo is still commonly used (especially by ecologists)to recognize those eukaryotic species that exist as single cellsand whose nutrition is dependent on the uptake of preformedorganic substances (primarily via prey ingestion) while pro-tists possessing chloroplasts are still commonly called ldquoalgaerdquoSimilarly although the term ldquoprotistrdquo has been abandoned asa kingdom designation it is still widely employed to describeeukaryotic species that are capable of existence as single cells(ie algae and protozoa) The term ldquophagotrophic protistrdquohas also gained popularity in recent years because it recognizesthat many protistan species are capable of phagocytosis eventhough they may also possess their own chloroplasts and thusare technically ldquoalgaerdquo
Despite the shortcomings of Whittakerrsquos scheme it domi-nated the hierarchical organization of life for approximately aquarter century During the past few decades however thissystem has given way to a new organizational scheme that rec-ognizes three domains of life (Archaea Bacteria Eukarya [orEucarya] 78 Fig 2 upper panel) and is based onwhat is pres-ently believed to reflect amore realistic view of the evolution-ary distances that have developed between organisms in theasymp4 billion years that life has existed on our planet Withinthe Eukarya of Woesersquos scheme hypotheses regarding thephylogeny of ldquoprotistsrdquo have changed continuously and rap-idly during the past two decades reflecting new insightsinto eukaryote evolution provided largely by DNA sequenceinformation (Fig 2 lower panel from [79])
The former protistan phyla ofWhittakerrsquos systemhavenowbeen dispersed among candidate ldquosupergroupsrdquo within thedomain Eukarya to better reflect hypothesized phylogeneticrelationships For example the dinoflagellates (which encom-pass phototrophic heterotrophic and mixotrophic species)now form a single group and have been placed together withthe ciliates and apicomplexans (sporozoans) in themonophy-letic Alveolata (Fig 2 lower panel) On the other handeukaryotic heterotrophic single-celled species falling withinthe general description ldquoprotozoardquo are now widely distributedamong a number of protistan lineages In short nutrition hasbeen demoted as a phylogenetic character and other charac-ters (presumably more indicative of evolutionary relatedness)have ascended to address some long-standing contradictionsalthough the debate over the relationships among some line-ages is still very active at the present time 79)
Individual protozoan cells range in size from less than 2 micromto greater than 1 cm in diameter (gt4 orders of magnitude)(80 81) with some colonial radiolaria forming cylindricalgelatinous structures a centimeter in diameter and morethan a meter in length (82 83) Because they constitutesuch a large size range of organisms protozoa are often dividedinto size classes that very crudely correlate with their generalnutritional preferences A commonly used convention is thatof Sieburth et al (84) which groups planktonic microorgan-isms into order-of-magnitude size classes (02ndash20 microm =picoplankton 20ndash20 microm = nanoplankton 20ndash200 microm =
microplankton 02ndash2 mm =mesoplankton) Most protozoafall into the nanoplankton or microplankton size classesModeling microbial trophodynamics using this conventionassumes that protozoa in one size category generally consumeprey one order of magnitude smaller in size (34 85)Although this approach misses much of the detail and diver-sity of the trophic activities of individual protozoan taxa it isa necessary practical compromise for examining community-scale flows of energy and elements It also provides a usefulmechanism for summarizing and comparing the abundancesand biomasses of protozoa from different environments andto other microbial assemblages Protozoan abundance sum-marized in this way has been shown to contribute signifi-cantly to the living biomass of planktonic ecosystemsthroughout the world ocean (Fig 3)
Estimates of protozoan biomass such as those depicted inFig 3 typically do not include the contribution of mixotro-phic phytoflagellates to heterotrophy There is presently noeasy way to determine the abundances of small phagotrophicphytoflagellates in natural samples so these species are typi-cally counted as phytoplankton unless specific methods areemployed to identify the algae as consumers such as the useof fluorescently labeled particles (87ndash90) or through theexamination of food vacuole contents (91) On averagethese species appear to constitute a modest percentage ofthe phytoplankton assemblage (typically lt25) althoughthey may at times dominate the phototroph assemblages ofnatural plankton communities It is important to recognizethat their inclusion as functional heterotrophs rather thanphototrophs could significantly shift the relative contribu-tions of phototrophic and heterotrophic microbial biomassto total biomass within microbial assemblages and the flowof energy within plankton communities (92)
Heterotrophic protists that harbor photosynthetic pro-tists or their chloroplasts within their cytoplasm constituteanother complexity for estimating the contribution of pro-tozoa to total microbial biomass When bulk water samplesare analyzed the contribution of chlorophyll containedwithin those protozoa is generally assumed to come fromfree-living phytoplankton However studies have shownthat chloroplast-bearing ciliates can contribute up to halfthe total biomass of planktonic ciliates in ecosystems andchloroplast-retaining ciliates can sporadically dominate thechlorophyll and primary production of some planktonic eco-systems (93ndash96) The environmental conditions promotingthe success of these ciliates are poorly known Similarlymany species of planktonic foraminifera polycystine radio-laria and acantharia harbor large numbers (thousands perprotozoan) of endosymbiotic algae within their cytoplasm(97 98) Caron et al (99) have demonstrated that primaryproductivity within these species can contribute significantlyto total primary productivity in oceanic ecosystems and can bevery important locally in the convergences of Langmuir circu-lation cells (100)
BACTERIA AND ARCHAEAldquoCulturablerdquo versus ldquoNonculturablerdquo CellsMost conventional cultivation methods can grow only 1 orless of the bacteria that can be visualized by direct microscopytechniques (eg 4) This is true even though most can beshown to be active by techniques such as microautoradiogra-phy (25) These readily cultivable organisms appear to repre-sent a group of fast-growing so-called weeds that are adaptedto take advantage of rapid growth in rare organically enriched
422-6 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
environments This strategy contrasts with the numericallydominant bacteria that are adapted specifically for growthin the dilute nutrient conditions that characterize the vastmajority of the volume of the water column
The most common taxa readily cultured from seawaterwith standard nutrient broth media include the gamma pro-teobacterial genera Vibrio Alteromonas PseudoalteromonasMarinomonas Oceanospirillum Shewanella (usually isolatedfrom surfaces such as shellfish and sediments) the alpha pro-teobacterial generaRoseobacterSphingomonasmembers of thefamily Flavobacteriaceae and Planctomycetes as summarizedin Giovannoni and Rappeacute (101) and Fuhrman and Hagstrom(102) The cyanobacteria Synechococcus and Prochlorococcusare also now readily culturable but on low-nutrient inorganicmedia targeting photosynthetic forms as opposed to organicmedia used to cultivate the others listed above
Because of the low percentage of marine bacteria that canbe grown in standard media organisms that until recentlywere called ldquononculturablerdquo make up the large majority ofbacteria in the plankton Only during the pastsim20ndash25 yearshave molecular biological methods based on 16S rRNA genesequences been available to identify these organisms andthese powerful techniques have opened up a large area forexploration (see next section) Similar but more recent stud-ies use 18S rRNA sequences for characterizing protistandiversity as will be noted below
Molecular Phylogeny and MetagenomicsField ApplicationsModern phylogeny of microorganisms is based primarilyon genetic sequences the most well-studied gene being the
FIGURE 2 The three domains of life (upper left) as proposed by Woese et al (78) and a recent overview of modifications that have beenproposed by Adl et al (79) to higher-level phylogentic groups within the eukaryotic component of the tree (lower right) Domains figure fromWoese et al (78) eukaryotic tree figure from Adl et al (79) doi1011289781555818821ch422f2
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-7
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
small subunit ribosomal RNA gene (16S rRNA in Bacteriaand Archaea and its larger homolog 18S rRNA in eukar-yotes) This molecule is strongly conserved over evolutionarytime so this single molecule has been used for constructingphylogenetic trees of all living organisms (httptolweborgtreephylogenyhtml) Analysis of 16S18S rRNA genesequences has been used to evaluate deep evolutionary rela-tionships among organisms and was instrumental in point-ing out that Archaea Bacteria and Eukarya should beconsidered different Domains of equivalent phylogeneticrank above kingdoms (78) However there are sufficientdifferences in 16S18S rRNA gene sequences to demonstrate
relationships (ormake distinctions) at the genus or sometimesspecies level
The first phylogenetic studies based on 16S18S rRNAgenes used sequences derived from cultures However onedoes not need cultures to obtain rRNA gene sequences (orany other sequences for that matter) An idea developed inthe lab of Norman Pace in themid-1980s involved extractionof DNA directly from natural samples and then cloning andsequencing of the DNA as a means of assaying the microbespresent in the samples (103 104) The original protocolscalled for cloning by creating what are called ldquophage librariesrdquofrom the natural DNA but since 1986 PCR has been applied
FIGURE 3 (a b) Plankton biomass in the Arabian Sea during the 1995 southwest monsoon (a) and intermonsoon period (b) Areas of theboxes indicate the relative magnitudes of the biomass in each category Categories within the dashed boxes in a b c are composed of protozoaArrows indicate the direction of energymaterial flow in the food web thicker arrows depicting greater flow Redrawn from (86)(c) Depth-integrated biomass (mgm2) in the upper 100 m of the Sargasso Sea near Bermuda and in the upper 200 m of the equatorial Pacificat 175degE Thewidth of the bars indicates the biomass in each size category Heterotrophs have been separated by size class while phytoplanktonhave not Size classes delineated by the dotted box are comprised of protozoa Redrawn from (65) doi1011289781555818821ch422f3
422-8 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
extensively for cloning and related studies The targetsequence can be almost instantly ldquoidentifiedrdquo to its closestphylogenetic neighbor by what have come to be standardonline sequence comparisons
Beyond the study of targeted genes (like the 16S rRNAgene) shotgun metagenomic studies have examined theentire genetic repertoire of the microbes in a given sampleThe metagenome is the collective genome of all organismsin the sample Initially these studies extracted DNA fromall organisms in a sample (usually prefiltered through a filterapproximately 1 microm to remove most organisms larger thanbacteria) sheared it to produce fragments and clonedthem into standard vectors either as small (thousands ofbases) or large (to hundreds of thousands of bases) insertsthat is fragments of DNA from the environment nowcloned into the vectors in a form suitable for sequencingThe best known early marine study of the former type isthe Global Ocean Survey with initial results published byVenter et al (105) which generated more than a billionbases of DNA sequence and reported 12 million previouslyunsequenced genes estimated to come from at least 1800different genomic species cumulatively in the many samplesthey analyzed
With the advent of next-generation sequencing that gen-erates millions ormore sequences in a run (known by a varietyof acronyms including 454 Illumina SOLiD etc) cloningof genes has largely been replaced by clone-free sequencingThe extent and power of such sequencing has recentlybeen demonstrated by the ability to construct essentiallythe entire genome of an uncultivated marine Group II Eur-yarchaeon that constituted only approximately 2 of amicrobial community sample using SOLiD sequencingmade possible by high coverage and the use of mate pairsequencing ofsim3000 bp fragments (106) (this length isneeded to span repeats and highly conserved genes that oth-erwise make genome construction difficult)
For 16S rRNA gene studies ldquotag sequencingrdquo pioneeredby Sogin et al (107) consists of amplifying a suitable sizedpart of the gene with broadly conserved primers (choice isimportantmdashfew are truly universal for the groups intended)often ldquobarcodedrdquo to allow multiple samples to be combinedinto a single run The amplified products are then sequencedand sequences processed en masse Many thousands of partialSSU rRNA gene sequences per sample are economically ana-lyzed this way though they are usually short (currently a fewhundred bases each depending on the sequencing platform)This way of analyzing the composition of microbial commun-ities is now standard These approaches provide so muchinformation even about very rare sequences that the resultshave led to the important concept of the ldquorare biosphererdquoorganisms that may be active or dormant and constituting avery small proportion of the community (eg often muchless than 01) but are potentially important for dispersionadaptation to changing conditions or even critical activitieslike nitrogen fixation or vitamin production (108) Howeverdue to the potential for artifacts like error sequences thisapproach requires rigorous application of quality filteringand clustering algorithms to avoid erroneous taxa and overes-timation of species richness (109 110)
Next-generation sequencing approaches have also allowedwork to begin metagenomics studies of microbial eukaryotes(111) These studies are still constrained by the much largergenomes of eukaryotes and therefore the difficulties ofobtaining sufficient sequences to reconstruct the genome ofa particular species Along with advances in sequencing abil-ity studies of eukaryotic metagenomics have been facilitated
by advances in single-cell isolation which when coupledwith high-throughput sequencing approaches reduce the tre-mendous complexity present in natural complex eukaryoticcommunities to a manageable task (112) Such single-celltechniques are also quite valuable in studies of bacteria andarchaea though the amplification technique tends to bevery uneven and typically generates less than half the genomeof each isolated cell regardless of domain (113 114)
Additionally similar to DNA mRNA is amenable toextraction and sequencing although greater care must betaken during extraction and purification as RNA shows agreater susceptibility to degradation during processing Copy-ing of mRNA by reverse transcription of RNA into cDNAfollowed by DNA sequencing has allowed insights into themetatranscriptomes of environmental samples Metatran-scriptomic studies provide information on gene expressionin an ecosystem and thereby indicates ldquoactivityrdquo of themicro-bial community rather than simply ldquopotentialrdquo represented bythe genomic DNA present in the sample with many applica-tions from showing which processes are being carried outby which organisms to fine-scaled diel studies (115ndash118)Nevertheless given variations in the lifetimes of differenttranscripts and protein molecules the transcriptome maynot be fully representative of the current activity of anorganism
Limitations of these analyses include sequencing errorsPCR mismatches or biases clustering and bioinformaticschallenges and chimeras generated during PCR Also thephylogenetic resolution of short sequences is limited giventhe high conservation of rRNA sequences Even with clone-free shotgun metagenomics there may be biases such as non-random losses of DNA during extraction and preparationor biases (eg from G +C content or secondary structure)in the sequencing procedures Determination of speciesdiversity by these approaches provides an example of theselimitations Shakya et al (119) working with synthetic com-munities (purified genomicDNA from16Archaea represent-ing 3 phyla and 48 Bacteria representing 16 phyla remixed tosimulate an environmental DNA extract) applied both meta-genomic analysis (454 and Illumina platforms) and PCRamplification followed by 454 sequencing of 16S rRNA genesto determine both species richness and relative abundanceThey found that PCR amplification454 sequencing of 16SrRNA genes yielded an accurate measure of species richness(providing that appropriate data processing was applied)but that the relative abundance of up to 94 of the species(depending on domain and variable region amplified) wasover- or underestimated by at least 15-fold (values rangedfrom not detected to 103-fold overestimation) In contrastboth metagenomic approaches yielded relative abundancesthat were within the authorsrsquo 15-fold accuracy cutoff forsim50 of the species However they concluded that addressingrichness overestimation in metagenomic analyses that isdistinguishing rare but real OTUs from experimental andcomputational artifacts awaits further computational andclassification improvements More recently Parada et al(120) used mock communities composed of 16S rRNAclones from 27 common marine taxa (from nine Bacterialand two Archaeal phyla) to show that small differences inPCR primers (and different clustering methods) can yieldlarge differences in apparent relative abundances of reportedtaxa However one primer pair and informatics pipelinethey tested using a particular version of 515F-926R (V4ndashV5) provided accurate estimates of relative clone abundance(r2 = 095) when comparing observed versus expected cloneabundance
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-9
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
Culture-Independent Diversity Studies
The first groups to be identified using cultivation-independ-ent molecular techniques from the marine plankton (121)were the bacterial SAR11 cluster (a group of closely relatedgene sequences or phylotypes) and marine picoplanktonicunicellular cyanobacteria Synechococcus and Prochlorococcus)Of these the SAR11 cluster was completely unknown butthe cyanobacteria had previously been recognized by theirunique pigment fluorescence Waterbury et al (122) andJohnson et al (123) used epifluorescence to observe Synecho-coccus and Chisholm et al (124) discovered Prochlorococcususing flow cytometry These cyanobacteria were later isolatedand grown in phytoplankton culturemedia These two groupsare generally common in the euphotic zone with SAR11 typ-ically comprising one third of the planktonic bacteria (125)and the cyanobacteria common everywhere but polar waters
Probably the biggest surprise to come from the applicationof molecular tools was the discovery by Fuhrman et al (126)of abundant archaea in the deep sea The archaea were foundto be in a unique phylogenetic cluster that was only distantlyrelated to any previously known archaea but the ldquoclosest rel-ativesrdquo (not really close at all) were extreme thermophiles Asubsequent study also found archaea to be present in near-surface coastal waters albeit relatively rare (lt2 relativeabundance) This study used a PCR technique specificallytargeted archaea and found ldquomarine Crenarchaeardquo plus a sec-ond group belonging to the phylum Euryarchaea (127)
Up until this time all known Archaea were thought to beldquoextremophilesrdquomdashadapted for either very high temperatures(thermophiles) extremely salty conditions (halophiles) orstrictly anaerobic environments (methanogens) Yet theseorganisms were present in cold or cool water at ordinary salin-ity and high oxygen concentrations Fluorescent in situhybridization (FISH) measurements from deep-sea sampleshave since showed that the archaea may make up appro-ximately 40 of the total countable prokaryotes with thepercentage reaching to 60 at 200 m depth in theMediterra-nean (35 36) An extensive time series of FISH measure-ments near Hawaii confirmed that the archaea are indeedvery abundant throughout the year from below the photiczone to at least 4000 m and typically constituting 30ndash40of the total prokaryotes present in waters deeper than a fewhundred meters (128) with similar results found elsewhere(129 130) (Fig 4) They have been reported from many pla-ces including the Atlantic Pacific and Southern Oceansand are dynamic components of the plankton the most com-mon type by far being the ldquomarine Crenarcheaeardquo (128 129131) While the marine archaea have been reported to bedominated by a few major ldquophylotypesrdquo (132) they alsohave been shown to have a great deal of microdiversity withinthese phylotypes suggesting there are many kinds of close rel-atives coexisting (133) Interestingly this group of archaeamay likely be the most abundant kind of organism on Earthgiven the huge volume of the deep sea and their high abun-dance there (134) It has recently been proposed that ldquomarineCrenarchaeardquo be elevated to their own major phylum outsidethe Crenarchaeota called the Thaumarchaeota on the basisof deep phylogenetic branching and fundamental differencesbetween them and the Crenarchaeota (135) The Thau-marchaeota possess the uniquely archaeal membrane lipidcrenarchaeol and are nowalso known to be abundant in soils
The physiology of the Thaumarchaeota has been anintriguing area of study Initially an autoradiography-FISHcombination approach demonstrated that they take upamino acids (130 136) But subsequent data showed that
crenarchaeol was derived largely from CO2 suggesting auto-trophic metabolism (137 138) An experiment showinguptake of 13C-labeled bicarbonate into these archaeal lipids(139) directly pointed to autotrophy in this group Chemoli-thoautotrophywas first hinted at byVenter et al (105) whosemetagenomic analysis showed an apparent archaeal scaffoldthat contained genes suggestive of ammonia oxidation Fur-ther evidence came from Schleper et al (140) who detectedseveral ammonia oxidation genes in order directly adjacent toa Thaumarchaeota 16S rRNA gene in a soil-derived metage-nomic clone The issue was directly resolved when Konnekeet al (141) isolated a related marine archaeon from sedimentof a marine aquarium and this organism Candidatus Nitroso-pumilus maritimus was found to have a chemoautotrophicmetabolism quantitatively oxidize ammonia to nitrite andcontain an archaeal ammonia monooxygenase gene (amoA)and interestingly did not grow heterotrophically Cultivationexperiments showed this organism has a high affinity forammonium allowing it to outcompete bacterial nitrifiers atlow (submicromolar) concentrations as are typical in thesea (142) The complete genome of the organism has shownnovel adaptations for nitrification and autotrophy (143)
Even if the Thaumarchaea are primarily chemolitho-autotrophs field data suggest a level of mixotrophy in thatsome organic substrates are being incorporated into biomassA stable isotope study using cells collected at 670 m depthoff Hawaii estimated that about 80 of the carbon incor-porated into archaea-specific lipids came from inorganic sour-ces and about 20 from organic compounds (144 145)Interestingly it has also been claimed that the genomesof deep-sea (gt2000 m depth) members of the Thaumarch-aea as well as those living in equatorial waters rarely containthe amoA gene and thus may be primarily chemoorgano-trophs (48)
Major bacterial groups that have been documentedfrom seawater using 16S rRNA characterization includesome that are also known from culture (eg AlteromonasRoseobacter) and several that are phylogenetically distantfrom standard cultures The most common groups are inrough order of their relative abundance in clone librariesfrom most to least abundant SAR11 (relatives of Pelagibac-ter ubique) Roseobacter SAR86 cyanobacteria SAR116SAR202 SAR234 andMarine Group A The SAR designa-tion followed by a number is an arbitrary sequential cloneidentifier from Sargasso Sea cloning studies done by the Gio-vannoni lab that did most of the early systematic cataloging ofclones Summaries of the data and phylogenetic relationshipsof these groups can be found in Fuhrman andHagstrom (102)and Giovannoni et al (101)
Molecular Genetic Discoveries in Bacterial andArchaeal Marine BiologyAs described earlier metagenomics is an extension of theideas used in the 16S rRNA cloning studies in that all genesfrom the native microorganisms are separated and clonedwithout having cultivated the organisms These methodsearly on started to find unexpected and very interestingresults The best examples involve unexpected marine photo-trophy the first of which is discovery of a nonchlorophyllphotosynthetic bacterial pigment called proteorhodopsin(146) The gene was found on a large environmentallyderived fragment of DNA that also had a gene coding for16S rRNA from the so-called SAR86 group (one ofthe groups common in seawater) This pigment can act as alight-driven proton pump thought to permit cells to generate
422-10 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
FIGURE 4 Distribution of Bacteria Euryarchaea andmarine Crenarchaea (recently renamed Thaumarchaea) along a transect in theNorthAtlantic Ocean as measured by CARD FISH and expressed as of total bacteria + archaea counts via DAPI stained epifluorescence fromTeira et al (130) Top panel shows station locations that are shown on the top of the lower three panels with the distance in km reported alongthe transect from lower to higher station numbers doi1011289781555818821ch422f4
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-11
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ATP from sunlight Interestingly different versions of this pig-ment are found at different depths apparently ldquotuningrdquo theabsorption to match the ambient wavelengths of light asthey change with depth (147) The proteorhodopsin genesare very widespread and diverse perhaps in half or more ofall marine bacteria including SAR11 and Euryarchaeota(105 148ndash150) However almost all of several culturedorganisms with proteorhodopsin studied to date do not showa growth benefit from light (150) with the exception of amember of the Flavobacteria Dokdonia sp strain MED134that grows faster in the light only under moderately lownutrient conditions (151) and a marine Vibrio has beenshown to survive starvation longer in the light than in dark-ness (152) These observation may explain the wide phyloge-netic distribution and high abundance of this gene in thatproteorhodopsin may often assist long-term survival of bacte-ria under extreme energy-limiting conditions yet not havemucheffect on growthduringmore energy-replete conditionsInterestingly a recent report suggests the growth benefit fromlight in Dokdonia MED134 is from enhanced uptake of itsrequired growth factor vitamin B1 which is transported by atonB-dependent transporter powered by a proton gradient(153) This pigment might also have sensory or other rolesnot yet well examined (150)
The second kind of phototrophy found by moleculargenetic and fluorescence techniques to be unexpectedlyimportant in marine plankton is anoxygenic aerobic bacterialphototrophy based on the pigment bacteriochlorophyll aCulturable aerobic anoxygenic photosynthetic (AAP) bacte-ria have been known from seawater for several years (154) andare widespread (155) Initial recent reports of direct countsclaimed they represent about 11 of the total bacterial com-munity in the euphotic zone (156) but those early counts didnot correct for the presence of other bacteria and such correc-tionoften yields estimates averaging closer to 2(157)Directmeasurements show the bacteriochlorophyll pigment is rela-tively rare (158) However studies in various ocean locationsincluding ocean gyres report that these AAP bacteriacan indeed on occasionmake up a quarter of the total prokar-yotes (159 160) A recent energetic modeling of AAP andproteorhodopsin-containing bacteria (161) has suggestedthat proteorhodopsin-containingones cangain approximately02asmuchenergy from sunlight asSynechococcus (commonmarine cyanobacterium) and AAP bacteria can gain approxi-mately 13 as much energy from sunlight as SynechococcusThey concluded the AAP bacteria may gain energy sufficientto meet maintenance costs but proteorhodopsin-containingbacteria were not expected to do so except at high light inten-sities andwith largenumbersof proteorhodopsinmoleculespercell They concluded the ease and low cost of maintainingPR-based phototrophy (a few genes required) may explainthe high incidence of proteorhodopsin genes
Other metagenomic studies have yielded interestinginsights about organisms and processes that would otherwisebe difficult or impossible to establish For example thesequence of an archaeal gene fragment isolated from seawaterrevealed extensive evidence of genetic exchange with othertypes of archaea and even bacteria (162) Such geneticexchange was also quite evident from the complete genesequences of different strains of Prochlorococcus and Synecho-coccus which was attributed in part to virus-mediated geneflow (163ndash166)
Dilution Cultures of Bacteria and ArchaeaSome recent cultivation techniques that permit growth ofldquotypicalrdquo oligotrophic marine bacteria in pure culture are
based on the simple concept that bacteria living in seawateron dissolved organic matter might best be grown in the labo-ratory in ordinary filtered seawater This conclusion followsfrom similar techniques to grow mixed ldquoseawater culturesrdquoof marine bacteria (167) but the seawater inoculum in thiscase is diluted so that only one or a few bacteria are addedto the initial culture vessel (168) A rapid throughput versionof this method has been used to cultivate members of bacte-rial groups thought to be conventionally uncultivable such asthe SAR11 clade (169 170) albeit often at very low den-sities Such cultures permit focused studies of the propertiesof those organisms including genomic analysis (169 170)This valuable work provides insight into the roles and activ-ities of some of the most common bacterial phylotypesobserved in the ocean (171) showing for example that culti-vated SAR11 requires a source of reduced sulfur as well asproviding templates aiding the interpretation of metage-nomic analysis (172)
PROTOZOADiversity and DistributionHeterotrophic protists have been identified historically fromtheir morphological features that are apparent at the levelof the light or electronmicroscope Features of cell size shapetype and pattern of flagellationciliation skeletal structurescharacteristics of the nucleus and other cellular structuresand organelles have been used to differentiate themany thou-sands of described species A description of the extent of thisdiversity of form and function is well beyond the scope of thisbook (see [79] for more information) However much of thisdiversity can be reduced to three basic body plans that dictatethe broadest ecological roles of these cells amoeboid flagel-lated and ciliated forms (Fig 5) Among the lineages possess-ing one of these three forms only the last group the ciliatedprotists constitute a monophyletic group within the domainEukarya
One of the simplest body plans for protozoa is the amoe-boid cell exemplified by the gymnamoebae or ldquonakedrdquoamoebae Motile nonphotosynthetic cells lacking flagellaor cilia occur as life stages in a number of phylogeneticallydiverse taxa but for many free-living species of protozoathis form constitutes the only life stage Motility is largelyconfined to movement along surfaces by means of pseudopo-dia that can take on a variety of (species-specific) shapes Sig-nificant abundances of amoebae are largely confined tobenthic and epibiotic environments (174) and to suspendedparticulate material where they can occasionally be highlyenriched (27) Most amoebae consume bacteria and otherminute prokaryotes and eukaryotes
Several heterotrophic protistan groups possess complexamoeboid body plans most notably the foraminifera polycys-tine and phaeodarian radiolaria and the acantharia Plank-tonic forms are predominantly oceanic in their distributions(although a large number of benthic species of foraminiferaexist) These species are heterotrophic and many are visibleto the naked eye (individual cells can bege1 cm gelatinouscolonies can form ribbon-like structuresge1 m in length)They feed on a wide variety of bacterial protistan and meta-zoan prey using pseudopodial networks have rather longcomplex life cycles for individual cells (weeks to months)and are extensively used in studies of paleoclimatologicalreconstruction (82 175) In addition many of these speciespossess intracellular symbiotic usually eukaryotic algaeThe widespread occurrence of algal endosymbiosis among
422-12 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
these protozoa in oceanic pelagic environments implies thatstrong selective forces appear to give rise to these associations(97 98)
Flagellated forms of protozoa exist across many protistanlineages Eukaryotic flagella come in various sizes numbersand forms (eg with or without tiny hairs) that are character-istic of the different lineagesMany flagellate species are appa-rently purely phototrophic or heterotrophic (phagotrophic)but a significant (still poorly known) fraction of flagellates aremixotrophic combining both phototrophy and phagotrophy(70 88) Flagella in phagotrophic flagellates are employed formotility and prey capture Most free-living flagellates possessone to four flagella (typically one or two) that can be manytimes the length of the cell itself Although there is tremen-dous species diversity among flagellated protozoa many ofthese species have broadly overlapping ecologies As a groupflagellates are the most numerically abundant protozoa ofboth benthic and pelagic ecosystems and they are fundamen-tally important as consumers of bacteria cyanobacteriaand other eukaryotes Flagellated protozoa within the nano-plankton size class (2ndash20 microm) are often counted as a singleassemblage in plankton studies because morphological details
apparent using light microscope are insufficient to distinguishamong the many species This assemblage has been variouslyreferred to by a variety of names and acronyms including het-erotrophic nanoplankton (HNAN HN) heterotrophic flag-ellates (Hflags) heterotrophic nanoflagellates (HNF) andmicroflagellates Heterotrophic nanoplankton is the mostaccurate term for most methodologies employed to countthese cells because it does not require visualization of flagella(which are often lost from these small cells during preparationfor microscopy) The confusion over terminology is partly aconsequence of the different methodologies that have beenused to count these species and partly a consequence of thefact that these species were first studied prior to the wideacceptance of the size convention of Sieburth et al (84)
Heterotrophic flagellates in the microplanktonic size class(20ndash200 microm mostly dinoflagellates) are important consum-ers of phytoplankton in pelagic ecosystems Many of thesespecies are capable of the production of large pseudopodialnets the pallium that can engulf prey (particularly diatoms)significantly larger than the diameter of the dinoflagellatetheca (176) This behavior and its ecological significance forenergy flow in plankton communities have been recognized
FIGURE 5 Body plans and size ranges of protozoa These micrographs depict amoeboid (andashg) flagellated (hndashj) and ciliated (kndashm) forms ofprotozoa From Caron et al (173) Markers bars are 5 (i) 10 (h) 20 (c j l) 30 (a) 50 (k m) 100 (b f) 500 (d) and 1000 (e g) micromdoi1011289781555818821ch422f5
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-13
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
only within the past few decades (177) These species areoften abundant in waters where diatoms dominate (178179) In such situations heterotrophic dinoflagellate biomasscan be similar to that of ciliated protozoa (180)
Ciliates are generally the most recognizable form of pro-tozoa to nonspecialists Nonetheless ciliates are a diversegroup morphologically as well as ecologically (181 182)The degree of ciliature on these species can range from uni-formly ciliated to totally devoid of cilia during most of theirlife cycle to ciliature restricted to specific regions of thecell In addition cilia can fuse to form various complexstructures (eg ciliary membranes or veils cirri) that assistin locomotion food capture or attachmentCiliates are abun-dant and ecologically important species in both benthic (8)and pelagic (183) ecosystems and can consume a variety ofprokaryotic and eukaryotic prey Together with the heterotro-phic dinoflagellates these species are the dominant consum-ers of phytoplankton in many pelagic ecosystems (184) andas a consequence they form an important trophic link to met-azoan zooplankton (185 186)
Diversity and Biogeography of Protists FromMorphology to DNA SequencesUntil relatively recently the biodiversity of marine protistanassemblages was not generally considered a controversialtopic It was generally accepted that while all species of pro-tists certainly have not been identified representatives ofmost types of algae and protozoa had been observed anddescribed if not actually brought into culture in the labora-tory Direct sequencing of 18S rRNA genes from environ-mental samples (as described in ldquoMolecular Phylogeny andMetagenomics Field Applicationsrdquo) has changed that viewInitial forays into environmental DNA indicated a muchgreater diversity of protists than previously documented usingclassical approaches of culture and microscopy (81 187ndash191) Numerous publications over the past decades haveexpanded these findings (see [192] for a recent global analy-sis) which are highly analogous to discoveries in marine pro-karyote research as described already
Previously uncharacterized protistan diversity has nowbeen documented at virtually every level of eukaryoticorganismal classification Some of these findings couldhave been expected but some have been very unexpectedFor example a much greater diversity than noted previouslyhas been observed among small (lt10 microm) protists in plank-tonic ecosystems These species generally possess few distinc-tive morphological features and one could expect that manycryptic species might be present among these small morpho-types The molecular ldquodiscoveryrdquo of this eukaryotic diversityhas stimulated progress on the isolation and descriptionof new species and genera of minute algae and protozoa(193ndash197)
Analyses of environmental samples have also indicatedthe unanticipated existence of novel 18S rRNA gene sequen-ces that imply the presence of novel lineages of eukaryotes innatural protistan communities (198ndash200) These are sequen-ces for which there are apparently no known described orcultured species The degree to which these sequences dif-fer from sequences of known sequenced eukaryotes impliesthat some of these lineages may be distinct at the level ofphylum (201)
These findings have raised basic questions and somedebate concerning the true diversity and biogeography ofprotistan assemblages in natural ecosystems (202ndash204) thevalidity of the many novel phylotypes or cryptic species that
are being documented (205) the potential importance andsignificance of the many rare taxa that characterize thesecommunities (206) and the implications of these findingsfor the ecologicalbiogeochemical roles that protists playin aquatic ecosystems For example it was postulated andsubsequently confirmed that some novel alveolate lineagesreported from marine ecosystems make up a suite of parasiticprotozoa whose ecological importance may have been signifi-cantly underestimated in the past (207) Deciphering theidentity and significance of these many unknown phylotypeswill constitute a significant effort for protistologists in thefuture (80 85)
Life Histories and Ecological Strategies of ProtozoaAbundance prey type and life histories all vary tremendouslyamongmarine protozoa The smallest species (eg many flag-ellates) tend to be the most abundant and widely distributedin the world ocean Indeed many of these species may beglobally distributed (208) Most nanoplanktonic flagellateshave potentially rapid rates of grazing and growth Underoptimal conditions these species can divide by binary fissionseveral times a day and thus dramatic increases in their pop-ulations can take place in response to favorable conditionswithin a few days (209) Many of these species are capableof surviving for limited periods of time without food andhave developed a variety of physiological or life cycle strat-egies to cope with these events (209) However in contrastto some bacteria that may remain viable through long periodsof starvation (210) protozoa will expire or encyst in responseto low food abundance and thus ldquoboom-and-bustrdquo popula-tion changes are characteristic of small flagellates
On the other end of the size spectrum from the ratherubiquitous nanoflagellate species many of the largest proto-zoan species (polycystine and phaeodarian radiolaria plank-tonic foraminifera) are exclusively oceanic (ie do notsurvive in most coastal environments) andor have specificlatitudinal and depth distributions Thus the abundancesof these latter species may range from undetectable to max-imal abundances of gt105 individualsm3 Dramatic changesin the abundance of these protozoan taxa also can be relatedto changes in prey abundance physicalbehavioral aggrega-tion or to periodicity of life cycle events For example theplanktonic foraminifer Hastigerina pelagica reproduces on alunar cycle and thus abundances (and life stages) of this spe-cies in oceanic waters can vary considerably over the courseof a month (211) In general life cycles for the large amoe-boid protozoa are lengthy and complex (for single-celledorganisms) with life spans unknown for many species(attempts to culture them in the lab have so far been unsuc-cessful) but estimated to be on the order of months to per-haps years (175)
The abundance and activities of microplanktonic hetero-trophic protists (mostly ciliates and heterotrophic dinoflagel-lates) tend to be somewhat intermediate to those ofnanoplanktonic flagellates and the larger amoeboid formsThese species are present in the majority of marine ecosys-tems and collectively play an important role in the controlof phytoplankton biomass (and probably the abundance ofnanoplanktonic protozoa although there is little informa-tion on this topic) in waters throughout the world oceanLike small flagellates ciliates and dinoflagellates reproduceprimarily by binary fission but their maximal growth ratesare typically slower (one division a day is typical) and theirlife cycles often include sexual phases that allow for geneticrecombination
422-14 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
VIRUSESViral Abundance and General PropertiesViruses are simple biological agents typically 20ndash200 nm indiameter composed of a nucleic acid genome in a proteincoat that infect cells and ldquocommandeerrdquo the cellrsquos machineryto make more viruses which are released into the environ-ment when the host cell lyses or bursts A particular virus isthought to be capable of infecting only a narrow range of hosts(usually one species sometimes a genus rarely broader)Although some early studies had isolated viruses from thesea therewere no data prior to the 1980s showing such viruseswere very abundant and more significantly no evidence thatinfection was occurring in any important part of the planktoncommunity It wasnrsquot until the late 1980s that electronmicro-scopy with suitable concentration methods showed thatviruses are extremely abundant similar to or even higherthan bacterial abundance (212 213) Interestingly themost suitable transmission electron microscopy (TEM)approach used by Bergh et al (described in detail by [214])was actually similar to a direct centrifugation method devel-oped in 1949 (215)
TEM studies of viruses permit high-resolution images andobservation of viral morphology showing features such ashead diameter tails sheaths and tail fibers (Fig 6 collage
in upper left) Each type of virus has a fixed morphology(unlike bacteria that are potentially more plastic in theirappearance) and hence a coarse measure of viral diversityhas been possible by cataloging morphologies Studies thathave done so have found dozens or more different morpholo-gies of marine viruses in a given sample (217ndash219) A largeproportion resemble bacteriophages which are viruses thatinfect bacteria
The availability of brightly fluorescent nucleic acid stainsand high-porosity fine pore size (002 μm) filters made fromAl2O3 have made it easy to accurately count viruses by epi-fluorescence microscopy (38 220 221) Epifluorescence per-mits abundance estimates but does not allow observation ofviral morphology as viruses are below the resolution limitof light microscopy viruses are visible only as sources of light(like stars in the night sky see Fig 6 lower micrograph) Anextension of manual epifluorescence counts is the use of flowcytometry to count viruses now commonly employed in somelabs (222)
Virus abundance has been found to be closely related tobacterial abundance with a virusbacteria ratio typically10ndash301 A typical oceanic profile of bacterial and viral abun-dance is shown in (Fig 6 panel on right) This tight ratioand the strong correlation to bacterial abundance in relationto weaker correlations to chlorophyll have been cited as
FIGURE 6 Marine viruses (upper left collage) viewed by transmission electron microscopy These are cyanophages in the groups myoviridae(right and bottom) siphoviridae (left) and podoviridae (top) Scale bars represent 100 nm From Sullivan (216) Epifluorescence micrograph(lower left) of SYBR green I stained viruses (small fluorescing objects) and bacteria + archaea (large fluorescing objects) Depth distribution ofvirus and bacteria + archaea abundances obtained by epifluorescence microscopy of SYBRGreen stained cells in the central San Pedro BasinCalifornia (11 August 2000) doi1011289781555818821ch422f6
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-15
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
evidence that most marine viruses infect bacteria rather thaneukaryotic phytoplankton (217 218 223 224) though thereare also many important viruses of phytoplankton and otherorganisms with significant impacts on primary productivityblooms and ecosystem function (225ndash227)
The first demonstration of the activity of marine viruseswas presented by Proctor and Fuhrman (228) who showedby electron microscopic examination that viruses wereactually infecting marine bacteria and cyanobacteria at ameasurable rate Because only the final portion of the viruslife cycle is visible by electron microscopy (when the virusesare assembled and ready to lyse the host cell) only a very smallfraction of the infected cells can be counted as infected at anygiven time Proctor and Fuhrman (228) reported that only afew percent of the total bacteria from coastal waters and theSargasso Sea were visibly infected but they interpreted thedata with a model that implied the actual fraction of the totalcommunity that was infected was much higher They con-cluded that the total fraction of bacterial mortality attribut-able to viruses is roughly 10ndash40 Subsequent studies haveused refined versions of that same general approach andmodel (229 230) and numerous studies have used a varietyof alternative approaches to estimate viral activitymdashall yield-ing basically the same general conclusion (although themodel parameters need to be adjusted for cyanobacteriawhich on average seem to be infected less than bacteria)These approaches include (a) calculation of virus turnoverfrom decay estimates (231) (b) measurement of viral DNAsynthesis by incorporation of tritiated thymidine (232) (c)observation of effects of added viruses (233 234) (d) estima-tion of bacterial mortality in the absence of protists (e) use offluorescent viruses to estimate production by an approachanalogous to isotope dilution (38) and (f) dilution of virusesin filtered samples to estimate viral production (235) Theoverall consensus of these studies is that the initial estimateswere basically correct that is that viruses are responsiblefor about 10ndash40 of the bacterial mortality in most marinesystems (reviewed by [218 223 226 236 237]) Howeverthere is still some disagreement about the higher estimatesand it is likely that 40 mortality from viruses alone is nottypical for most marine systems
Most of the foregoing work has focused on the viral infec-tion of bacteria primarily thought to be heterotrophicalthough the original report by Proctor and Fuhrman (228)also noted the occurrence of cyanobacterial infection Sev-eral subsequent studies focused on phytoplankton includingthe potential effect of viruses on the termination of phyto-plankton blooms of Emiliania and Phaeocystis and the likelyimpact this might have on release of climate-active gasessuch as dimethyl sulfide (225 227 238ndash247) Althoughdetails are beyond the scope of this chapter viruses arethought to infect virtually all marine organisms with poten-tially significant impacts from zooplankton to whales (226)Detailed studies of viruses infecting cyanobacteria such asSynechococcus have shown differences in viral host specific-ity particularly toward coastal and oceanic host strainsand some occasional high virus abundances (to 105ml) asmeasured by most probable number (MPN) cultivation tech-niques in the Gulf of Mexico near Texas (248 249) Othervirus cultivation studies with Prochlorococcus and Synecho-coccus in oligotrophic waters of the Sargasso Sea showedan interesting pattern of cross-infection between these gen-era by some virus types (suggesting gene flow among theseorganisms) but generally low MPN estimates of abundanceto 103ml even when cyanobacterial abundance was near105ml (216 250)
Overall the consensus emerging from direct comparisonsof viral-mediated mortality and grazer-mediated (ie micro-zooplankton) mortality have indicated that viral lysis of bac-teria constitutes a significant fraction of total mortality ofthis assemblage while the mortality of phytoplanktonappears to be dominated bymicrozooplankton inmost instan-ces (251 252)
As mentioned viruses have significant morphologicaldiversity as observed in TEM studies It is also possible toinvestigate aspects of their genetic diversity In early workon this topic Wommack et al (253) and Steward et al(254) observed the diversity of viral genome lengths in a fieldsample by pulsed field gel electrophoresis with viral genomesranging 25 kb to gt300 kb in length Field results show thatthe viral community composition is dynamic in space andtime with clear changes in the banding patterns over seasonsand locations in Chesapeake Bay (253) between oceanbasins and subsequent to dinoflagellate blooms (254) andwith depths to 500 m at one location (255)
Although viruses do not all share a set of core genesthat allows a universal viral phylogeny (comparable to SSUrRNA gene in cellular organisms) genetic diversity amonga single group of viruses can be examined by sequence analysisof shared genes within the group An example is the g20 geneinT4-like cyanophages that has shown extremely high diver-sity even among very closely related viruses and has demon-strated geographic and seasonal variation (256) A secondexample is g23 which is found broadly in diverse and wide-spread T4-like phages (257) and can have seasonally repeat-ing patterns (258) as well as short-term rapid dynamics thatcorrelate to those in bacteria (259)
Viral MetagenomicsAs with cellular organisms metagenomics can be used toexamine viral diversity and genetics without themany restric-tions of cultivation Viruses can be collected by selective fil-tration and concentrated by tangential flow filtration orflocculated with iron chloride (260 261) then their collec-tive metagenome can be extracted linker amplified and ana-lyzed by sequencing (262 263) Viral metagenomic studiesare particularly challenging because the large majority ofsequences have no annotated matches in any databases butso far results from marine samples around the world haveshown extremely high diversity and variations with depthlocation and time presumably with highly dispersed typesselected by local conditions (262ndash266) Because metage-nomes are best interpreted when there are representative cul-tures available the best matches of viral metagenomesoriginally tended to be to the few viral isolates infecting trulycommonmarine bacteria like cyanophage infecting Synecho-coccus and Prochlorococcus (267) However the developmentof dilution-to-extinction cultures representing common het-erotrophic marine taxa like Peligibacter (a member of theSAR11 clade) and SAR116 has allowed isolation from sea-water of viruses infecting these common organisms and theseisolates have indeed been found to be highly abundant inmarine viral metagenomes (268 269) New approaches tointerpret the results include clustering the proteins independ-ent of known proteins to compare samples to each other andtry to find environmental factors driving viral communitychanges (270) Such analysis of a large global data set (TaraOceans expedition) has shown that extensive sampling hascome close to reaching the total diversity in tropical and tem-perate waters of such viral protein clusters (which essentiallyrepresent various viral protein motifs not all viral proteintypes) and that viruses appear to be directionally dispersed
422-16 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquodownstreamrdquo in major ocean currents as one may expectfrom first principles but also supporting the ldquoseed bankrdquohypothesis of viral biogeography (271)
Viruses and Host DiversityViruses are themselves thought to be instrumental in drivingincreased diversity of their microbial hosts via a hypothesizedprocess often called ldquokill the winnerrdquo This is because viralinfection is host-specific and density-dependent the latterbecause viruses diffuse from host to host so an abundanthost is more likely to pass on infection than a rare oneThis means that if an organism becomes abundant andblooms winning the competition for resources it becomesmore susceptible to a viral epidemic This would benefit therarer organisms and thus help foster diversity (223 272273) However bloom scenarios as described above involvesystems far from steady state and the formal theory of Thing-stad and Lignell (274) has interesting steady-state solutionswhere several viruses infect several hosts stably over time(via trade-offs between growth rates and viral susceptibility)which may occur at the strain or species level (274) There issome experimental evidence that viruses have effects on nat-ural marine microbial community composition weaker thanthe kill the winner (bloom version) hypothesis would sug-gest (275 276) so something resembling the steady-statecoexistence described in the model may in fact be commonThere also appear to be processes that foster coexistencebetween viruses and hosts but the mechanisms are largelyspeculative (223 277 278)
Viruses may also be directly involved in host genetic diver-sity because they can be the agents of genetic exchangebetween microorganisms (223 273) This often involvesthe viral lifestyle known as lysogeny whereby viruses survivewithin host cells as DNA only integrated into the host chro-mosome and being reproduced each time the host divides Ahost harboring such a genome is called a lysogen becauseunder conditions of stress to the host cell a genetic switchmay cause the viral genome to initiate the lytic process pro-ducing many progeny viruses and bursting from the hostLysogeny is a very common property occurring in a signifi-cant part of the bacterial community (279 280) althoughthe incidence of induction of the lytic phase in nature is appa-rently low (281) Overall lysogeny is poorly understood butthought to have both positive and negative impacts on themicrobial community (282) Recent results suggest that ben-efits of being lysogenic in highly seasonal polar seas leads tofundamental difference between polar and other marine viralcommunities (283)
Viruses and the Microbial LoopAs part of the food web viruses occupy a unique positionThey infect host cells that are mostly thought to be heterotro-phic bacteria and by doing so they typically burst the hosts torelease progeny viruses and cellular debris But what is the fateof this material Viruses themselves do not last indefinitelyand a simple steady-state assumption implies that from eachburst of viruses (typically 20ndash100 per lytic event) only onesuccessfully infects another cell The rest are inactivatedand broken down by sunlight (UV and visible exposure)and enzymatic attack (284) or consumed byminute phagotro-phic protists (285) thus reentering the food web as substratefor bacteria or food for protistan consumers Experiments incontrolled laboratory systems and field studies with radioac-tively labeled viral lysis products have supported the conclu-sion that most of the organic matter released by the viralinfection is either taken up by bacteria or respired (286 287)
Modeling this process as part of the microbial loop showsthat viral lysis represent a sort of side loop that has the neteffect of remineralizing a significant amount of the carbonand nutrients that enter the bacteria-protist part of themicro-bial loop (Fig 7) A theoretical numerical steady-state modelcomparing a systemwith no viral activity to onewhere virusesare responsible for 50 of bacterial mortality showed that thesystem with viruses had 33 more bacterial production andrespiration than the virus-free system implying that theviruses had the effect of permitting the bacteria to processmore of the primary production than they would otherwise(223) Although 50 is a high number unlikely to be com-mon in the sea this model nevertheless illustrates that virusescan reduce the amount of energy reaching higher trophic lev-els The implication is that viruses lead to increased bacterialactivity at the expense of the larger organisms
MAJOR ENVIRONMENTAL CONTROLSLight Temperature and PressureTemperature has an important potential influence on bio-chemical reactions and therefore on biological processesin general Most ocean waters fall in the range of ndash2degC to30degC with obvious exceptions in hydrothermally heatedareas Temperature has long been known to be a regulatingfactor for the growth of heterotrophic microbes
In temperate waters it has been established that microbialactivity is generally much higher in warm summer waters thanin winter (55) The relationship is not simple howeverbecause multiple factors act at the same time Some contro-versy still exists regarding the highest and lowest extremesfor marine bacterial growth although there is broad agree-ment that bacteria grow gt100degC at hydrothermal vents andltminus5degC in sea ice brines Pomeroy et al (57) noted the inter-esting observation that bacteria seem particularly inhibitednear the freezing point of seawater (ca ndash22degC) comparedto eukaryotic phytoplankton This effect results in polarspring phytoplankton blooms that accumulate organic car-bon in advance of the response of the bacterial communityand development of the microbial loop and perhaps lead toenhanced benthic-pelagic coupling (288)
The relationship between temperature and the growth rateof marine phytoplankton was described broadly in a now
FIGURE 7 Modification of the microbial loop concept that incor-porates the functional role of viruses Export can be via predation orsinking From Fuhrman (223)doi1011289781555818821ch422f7
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-17
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
classic paper by Eppley (58) and later Goldman and Carpen-ter (289) Temperature was shown to exert a strong and directeffect on themaximal growth rates of these species withmax-imal intrinsic growth rates at 0degC generally lt1 divisiondaywhile growth rates at 30degC may be gt4day These relation-ships indicate the maximal rate that might be attained byphytoplankton at these temperatures but they do not takeother factors into account (nutrients light) While warmtemperature ostensibly allows more rapid growth it createshydrographic conditions that typically give rise to nutrientlimitation of algal growth Thus some of the coldest watersin nature witness some of the most massive phytoplanktonblooms (290) while warm oceanic gyres represent some ofthe most oligotrophic areas of the ocean The multiple andoften conflicting effects of temperature on the growth of nat-ural phytoplankton assemblages limit the accuracy of presentpredictions regarding how primary producers will respond toclimate change (291)
An analysis of the effect of temperature on the growth ofheterotrophic protists has indicated that the response is qual-itatively similar to that of phytoplankton but with a twistThe growth rates of at least some protozoa can exceed thoseof phytoplankton at warmer environmental temperaturesbut the opposite effect is apparent at very low environmentaltemperature This differential effect of temperature on thegrowth of phototrophic and heterotrophic protists was basedon a large meta-analysis of published protistan growth (59)That analysis demonstrated that the maximal growth ratesattained by phototrophic protists could exceed the maximalgrowth rates attained by heterotrophic protists (all otherpotential growth-limiting factors not considered) Thereforeprotozoan growth rates may be constrained to a greater degreeat low environmental temperature than rates for phytoplank-ton If so then phytoplankton bloomsmay get a head start ongrazers during spring in polar ecosystems This scenario is con-sistent with information on seasonal biomass changes andmicrozooplankton herbivory in the Ross Sea Antarctica(292 293) but there are still too few data to fully vet thishypothesis
The importance of high pressure on bacterial growthgained considerable attention in the late 1960s when thedeep sea submersible Alvin was accidentally lost overboardwith its hatch openWhile no lives were lost in this accidentsome workmenrsquos lunches sank to the bottom (sim1500 m)inside the submersible Alvin was recovered after 10 monthsand interestingly there was a waterlogged lunch containingapples bologna sandwiches and broken vacuum bottleswith broth that all appeared hardly degraded and tasted palat-able Yet when placed in a refrigerator on the ship these itemsdegraded relatively quickly Initially pressure was thought tobe the preserving factor as the sea floor temperature was sim-ilar to the refrigerator temperature and the only major differ-ence would be pressure (294) Following this observationa series of experiments to measure degradation of variousorganic materials left in the deep sea for extended periodsindicated that degradationwas typically significantly reducedimplying that pressure reduces the degradation rates (295)Nonetheless changes in the protozoan community of naturaldetrital material sinking to the deep ocean floor indicate thatthe microbial community can respond relatively quickly insome situations (296) Deep sea microbiology has advancedconsiderably yet it is still difficult to interpret results withrespect to actual in situ rates of naturally occurring organicmatter Deep sea bacteria adapted to high pressures havebeen isolated that are barophilic (also called piezophilic)meaning that they prefer high pressures and have reduced
activity at lower pressures whereas others are barotolerant(piezotolerant) tolerating but not preferring high pressuresfor example see (297 298)
Few data are available on barotolerantbarophilic marineprotozoa Protozoa certainly exist and grow at great oceanicdepths but measurements of in situ growth rates for these spe-cies do not yet exist Measurable protozoan numbers havebeen documented in the deep-sea sediments for more than30 years (299 300) and viable protozoa have occasionallybeen cultured from these environments (301ndash305) butvery few direct measurements of the activities of these speciesin situ have been reported (306) A few protozoa have beenisolated that will grow at high pressure (302 303 307) andprotists that appear to be unique to the deep ocean havebeen observed either directly or through the analysis ofDNA sequences (308ndash311) but possibly the best direct evi-dence that protozoan activity takes place at the high pressurescharacteristics of the deep sea are experimental and observa-tional work noting the stimulatory effect that detrital deposi-tion has on some components of the protozoan community(296 312) These observations indicate a diverse and activeprotozoan fauna of the deep ocean although their biogeo-chemical significance is largely uncharacterized at this time
Dissolved and Particulate Organic MatterBacteria and archaea are thought to be by far the most impor-tant organisms with respect to the processing of dissolvedorganic matter (DOM) and nonliving particulate organicmatter (POM also called detritus) in the ocean While theremay be some uptake of DOM by protists particularly forgrowth factors needed in trace amounts such as vitamins(313) the bulk of this material is probably utilized by bacteriaand archaea (314) including the smallest cyanobacteriumProchlorococcus (315 316) which thus may be considered amixotroph Due to their small size bacteria have extremelyhigh surfacevolume ratios and combined with their over-whelmingly high abundance an extremely high integratedsurface area Protozoa tend to obtain the organic materialsthat they require for growth from their prey rather thanthrough the uptake of DOM Overall protozoa tend to besources of dissolved and detrital organic substances throughthe excretion of unassimilated prey biomass in expelledfood vacuoles
Particulate organic matter is not directly available as sub-strate to bacteria These substances must first be reduced tosmall molecules that can be transported into the cell Thisis accomplished by the production of extracellular enzymes(note that few if any large polymers are directly taken up bybacteria with the possible exception of DNA) Hydrolyticenzymes produced by bacteria (and almost certainly archaea)break down polymers like proteins polysaccharides andnucleic acids As with DOM POM is composed of a complexmixture of compounds that vary in their susceptibility to bac-terial degradation and utilization
Particulate material in the water column serves not only asbacterial substrate but also as substratum POM occurs in thewater column across a huge size spectrum from micrometersup to some detrital aggregates more than 1 m in diameter(317) Much of this particulate material is in a constant stateof flux with colloidal material constantly coalescing andaggregating to form new or larger particles (318) as microbialdegradation acts simultaneously to remineralize this materialDetrital particles that attain macroscopic size either by directformation (317 319 320) or via accretion and aggregation(321) and are often called marine snow or macroaggregatesMarine snow particles are readily colonized by bacteria and
422-18 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
many other microorganisms and their abundances on theseaggregates can be orders of magnitude greater than abundan-ces in an equivalent volume of the water surrounding theaggregates (320 322ndash324) and their diversity and highlyconcentrated abundances on detrital aggregates implies thatthere may be stronger trophic coupling and efficient energyelemental cycling relative to these processes among free-living microorganisms in the surrounding waters This specu-lation has resulted in a number of formalizations of the poten-tial impact of aggregate microbiology on biogeochemicalprocesses in the ocean (325 326) Most definitions of bio-films include aggregated particulate and these particles areoften studied within the biofilm conceptual model (see chap-ter by Lawrence et al in this section)
Inorganic Nutrients
Macronutrients (N P)Virtually all heterotrophic organisms in the ocean contributeto the pool of available macronutrients via the excretion ofmetabolic wastes and protozoa contribute significantly tothis process Macronutrients are often in excess within proto-zoan prey relative to the consumersrsquo needs because much ofthe prey carbon is respired to produce energy for metabolismand growth ldquoExcessrdquo nutrients are eliminated as eitherorganic compounds or more commonly as ammonium (forN) and phosphate (for P) The importance of the match(or mismatch) between predator and prey stoichiometry hasbeen demonstrated experimentally and repeatedly in marineand freshwater ecosystems and has been elevated to a funda-mental ecological principle governing elemental and energyflow in aquatic food webs (327) Based on this reasoningCaron et al (328) concluded that bacterivorous protozoaplay an important role in nutrient remineralization andrelease in the ocean Bacterial biomass is typically rich in Nand P relative to other organisms so bacterial predators expe-rience large excesses of these elements relative to their growthneedsMoreover bacterivorous protozoa should be dispropor-tionately important in nutrient remineralization amongsingle-celled eukaryotes because as the smallest protozoa inaquatic ecosystems they have high weight-specific metabolicrates (329) Thus both stoichiometric and allometric rela-tionships implicate small bacterivorous protozoa as impor-tant sources of remineralized nutrients in the plankton
The reasoning also holds true for bacterial utilization oforganic compounds and it has led to some interestingfindings and conclusions regarding the role of bacteria innutrient cycling in the ocean Bacteria are important sourcesremineralized nutrients when they are consuming N- orP-rich substrate Contrary to this traditional view howeverheterotrophic bacteria are often strong competitors for inor-ganic nutrients under many situations largely because bacte-rial biomass is richer in N and P than much of the organicmatter they consume in nature so they require additional Nand P to produce their biomass (summarized by [330]) Exper-imental studies have demonstrated the uptake of ammoniumby bacteria other than cyanobacteria using stable isotope 15N(331) and also the short-lived radioisotope 13N (332) oftenfinding that bacteria may be responsible for one third of the Nor P uptake Additionally there is evidence that bacteria insome oceans like the Sargasso Sea are limited by phosphorus(333 334) Growth limitation of bacteria by phosphorusappears to be a common phenomenon of freshwater ecosys-tems (335 336)
N2 fixation is restricted to prokaryotes and in the oceanwater column it was thought for many years to be done
primarily byTrichodesmium a warm-water colonial cyanobac-terium that blooms sporadically andRichelia a cyanobacterialsymbiont that lives within certain diatoms (337) Some uni-cellular cyanobacteria also contribute significantly to globalN2 fixation (338) and molecular biological data suggestthat a variety of other bacteriamdashnot phytoplanktonmdashmayalso be fixing nitrogen in seawater (339) An exciting recentdiscovery in this field is that a bacterial nitrogen fixer origi-nally known only from nifH sequences UCYN-A has hadits genome sequenced by parallel pyrosequencing and wasfound be a cyanobacterium (Candidatus Atelocyanobacteriumthalassa) lacking the apparatus to generate oxygen as well asother pathways (340) Further study showed it to be symbioticwith a marine alga (341)
Micronutrients (Trace Metals Growth Factors)Bacterial growth may often be limited by macronutrients asdescribed and sometimes by trace nutrients particularlyiron that occur at very low concentrations in seawater(342 343) The importance of Fe as an element limiting pri-mary production in some oceanic regions has come underclose scrutiny and extensive experimental investigation inrecent years Many bacteria have special uptake mechanismsto utilize extremely low concentrations of iron includingsiderophores (released compounds that bind iron and arethen specifically taken up) Even if bacteria are capable ofgrowth at low Fe concentrations growth may be less efficientKirchman et al (344) found that Fe limitation stronglyreduced bacterial growth efficiency in a cultured marine bac-terium (ca 50 efficiency Fe replete but lt10 when Fedeplete) Thus the effects may be complex Marine N2 fixa-tion also may be limited by the availability of Fe becausenitrogen fixers have high Fe requirements
Protozoa tend to be a source rather than a sink for Fe in theocean analogous to their role in macronutrient remineraliza-tion Protozoan grazing activity has been shown to releaseiron from bacterial biomass thereby relieving Fe limitationin co-occurring phytoplankton (345) a mechanistic demon-stration of the process using protozoa growing on Fe-repletebacteria However it has also been reported that Fe-limitedbacteria may contain insufficient Fe for the bacterivorous pro-tozoa that consume them thus leading to Fe limitation in theprotozoan as well (346)
OxygenHypoxic and anoxic regions of the ocean are expected to sig-nificantly expand in the coming decades as a consequence ofchanges in ocean stratification resulting from global climatechange (347) As oxygen becomes depleted bacteria havethe capability to utilize a series of alternate electron acceptorsCompositional changes in the bacterial assemblage accom-pany these changes in metabolic activity but are beyondthe scope of this chapter Many of the studies on benthicmicrobiology have focused on redox reactions that occurthere Oxygen is usually consumed by sediment microbialactivity much faster than the rate at which it can be replacedby diffusion so most sediments especially organically richones are anaerobic below a few mm (fine-grained muds) orcm (coarser sediments) from the sedimentwater interfaceexcept where animals ventilate the benthos through tubesWhen oxygen is absent bacteria and archaea use alternateelectron acceptors such as nitrate nitrite oxidized Fe orMn organic matter or sulfate The use of nitrate or nitriteas an electron acceptor with concomitant production of N2
gas (denitrification) results in net loss of biologically availablenitrogen Bacteria that perform this reaction are typically
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-19
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquofacultativerdquo organisms that can switch from oxygen to nitrateor nitrite Organic matter may be reduced by fermentationreactions in which part of an organic substrate is oxidizedwhile another is reduced fermenter organisms are also typi-cally facultative Sulfate is a particularly important alternateelectron acceptor in marine systems because it occurs as thesecond most abundant anion in seawater (behind chloride)typically 28 mM Its use as an electron acceptor leads to theproduction of elemental sulfur or sulfide The use of sulfateas an electron acceptor yields much less energy than oxygenor nitrate and sulfate-reducing bacteria are strict anaerobesunable to use oxygen (often strongly inhibited by oxygen)Methane can be produced by methanogenic archaea whichare all strict anaerobes using CO2 as an electron acceptor(see Findlay and Battin this section for further discussion)
Reduced compounds such as sulfide are produced by theuse of these alternate electron acceptors and they diffuse awayfrom their source When they diffuse to environments with astrong oxidant like oxygen they may be oxidized by chemo-trophic bacteria in energy-yielding reactions Similarlyammonium can be oxidized to nitrite and then nitrate by che-motrophic nitrifying bacteria and the nitrate can diffuse intoanaerobic layers and become denitrified to N2 Such coupledoxidation-reduction reactions can lead to accelerated biogeo-chemical cycling particularly in sediments
It was originally believed that relatively few free-livingspecies of protozoa inhabit anaerobic and hypoxic marineenvironments This view has changed based on (largely)molecular analyses of some of these environments (308348ndash351) A significant number of previously undescribedeukaryotic phylotypes have been observed in anoxic environ-ments High protozoan biodiversity in the absence of molec-ular oxygen is not completely unexpected because otheranaerobic environments such as some gut environments(eg cockroach guts and cattle rumen) have notoriouslyabundant and diverse protozoan fauna but marine anaerobicenvironments have been difficult to access and sample prop-erly Many anaerobic protozoa exist together with symbioticbacteria (352 353) or possess unique organelles such as thehydrogenosome (354)
MAJOR BIOTIC CONTROLSThe Trophic Activities of Protozoa Predation andTop-Down ControlsPredator-prey relationships are fundamental interactionswithin biological communities linking production of bio-mass on the one hand with removal on the other hand Estab-lishing the trophic relationships among microorganisms andquantifying the rates at which they take place constitutemajor endeavors within marine microbial ecology As notedpreviously protozoa consume a wide variety of prokaryoticand protistan prey Numerous laboratory and field measure-ments have been made throughout the past quarter centuryto attempt to quantify the rates of bacterial and phytoplank-ton consumption The methodology for accomplishing thesestudies has varied and a number of approaches are presentlyemployed
Methodologies for estimating microbial grazing are diversebut generally fall into one of two experimental approachesperturbation experiments or tracer experiments Perturbationexperiments (size fractionation metabolic inhibitors anddilution technique) rely on manipulation of the consumeror prey assemblages in somemanner such that the effect is evi-dent from changes in the abundance of the prey population
during incubations relative to control treatments that receiveno manipulation Tracer experiments (radioactively labeledprey fluorescently labeled prey stable isotopendashlabeled pray)attempt to use labeled prey or a prey proxy to quantitativelyfollow the movement of prey biomass into consumers Forsummaries of these methods see (74 355)
Perturbation experiments have entailed a variety of mech-anisms to ldquodecouplerdquo predators from prey in natural microbialcommunities For example size fractionation uses various sizefilters to remove an entire size class of grazers from awater sam-ple leaving the prey to grow in the relative absence of thosepredators during subsequent incubations Unfiltered samples(controls) indicate the growth of prey in the presence of pre-dation Predation impact is obtained by the difference ingrowth of the prey between these treatments This methodhas been used to examine predation on cyanobacteria heter-otrophic bacteria and minute phototrophic eukaryotesAlternatively metabolic inhibitors have been employed tohalt predation (or in some cases prokaryote growth) insteadof physical removal by filtration
The dilution technique (356) has become a commonmethod for examining grazing mortality of phytoplanktonby microzooplankton (which is comprised predominantly ofprotozoa) This method relies on the dilution of herbivorouszooplankton in a series of subsamples (and concomitantmeasurements of phytoplankton growth in each subsample)to estimate the grazing mortality of the herbivores
Tracer experiments have used both radioactively labeledprey and fluorescently labeled prey to examine predation onbacteria and phytoplankton Radioactive labeling has beenaccomplished using 14C- or 3H-labeled organic compounds(for bacteria) or 14C-bicarbonate (for phytoplankton) Theuse of radioisotope approaches has diminished with improve-ments in the number and types of fluorescent compoundsavailable for this work Fluorescently labeled prey havebeen labeled with a variety of compounds that allow theprey to be observed readily (even inside predators) by epi-fluorescencemicroscopy (357 358) Alternatively a decreasein the total number of fluorescent prey in a sample indicatesthe rate of removal of the prey assemblage
Both perturbation experiments and tracer experimentsdescribed have inherent advantages and disadvantages Forexample perturbation experiments are fairly straightforwardto conduct but they may introduce artifact by removingpotentially important sources of nutrients or organic com-pounds released by predators (359) On the other hand thedilution technique is very labor-intensive but one advantageof this method is that it provides the opportunity to determineboth phytoplankton growth and grazing mortality in a singleprocedure Most tracer experiments are fairly easy to conductand generally do not perturb the food web being studied butfeeding selectivity for or against the tracer (relative to naturalprey) may produce spurious results In addition some of thesemethods provide a measurement of the activity of the entiregrazer assemblage and others are applicable for examiningthe activity of specific taxa For example size fractionationapproaches examine the effect of the removal of an entiresize class of organisms (a community-levelrdquo measurement)whereas fluorescently labeled prey can be used to examinethe ingestion rate of a particular species by observing theuptake of labeled prey into that species (a species-levelmeasurement)
Predation rates of numerous marine protozoa have beenexamined in the laboratory From these experiments a gen-eral knowledge of the rates of consumption by these specieshas been accumulated Feeding in most protozoa shows a
422-20 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
functional response to the availability of prey that is similar tothe response of many organisms (although there are a greatdeal of species-specific differences) That is ingestion rateincreases rapidly as prey abundance increases until a maximalingestion rate is attained (360)Manyof these curves have theform of a Type II functional response curve (361)
Based on the features of these curves it is clear that proto-zoa have the ability to ingest substantial numbers of prokar-yote and eukaryote prey in natural ecosystems Somebacterivorous flagellates can consume bacteria (and cyano-bacteria) at rates up to 100 s per individual per hour whereassome bacterivorous ciliates can consume thousands of bacte-ria per individual per hour Rates of consumption of eukary-otic phytoplankton by herbivorous ciliates and largeflagellates are considerably more variable and highly depend-ent of the feeding mechanism (eg filtration in ciliates versuspseudopodial net in dinoflagellates) and the size of the prey
These lab studies demonstrate that rarely are prey suffi-ciently abundant in nature to support the maximal ingestionrates of which many protozoa are capable Nevertheless pre-dation rates of protozoa in nature are sufficient to constitute amajor factor controlling the standing stocks of bacteria andphytoplankton (63 180 184 355) For example these sum-maries indicate that on average protozoa consume 20ndash80ofthe standing stock of phytoplankton (measured as chloro-phyll) in the world ocean and 60ndash75 of daily primary pro-duction a departure from the classical view of copepods as theprimary consumers of phytoplankton This large role of her-bivorous protozoa infers a large role for copepods as consumersof planktonic protozoa (185) Similarly the role of protozoaas major consumers of bacterial biomass is indicated by therelative balance between bacterial production on the onehand and bacterivory on the other (Fig 8) The approximateone-to-one balance between these measurements over a largenumber of studies implies that protozoan bacterivory must bea major factor determining the fate of bacteria in marine (andfreshwater) plankton ecosystems
It is interesting to note that some types of bacteria includ-ing marine Bdellovibrio are known to prey on other marine
bacteria (362) At the time of this writing the importanceof bacterial predation by other bacteria within the contextof oceanographic processes is not known and presumed tobe small While Bdellovibrio may be cultivated from seawaterit can also be found among sequences from cold sediments(363) and the water column Bdellovibrio happens to be thebest studied predatory bacterium but there are no doubtothers that we simply have not yet identified (364)
Species-Specific Interactions (MutualismParasitism Commensalisms)Interactions between marine microorganisms such as compe-tition allelopathy and symbiosis are fundamental aspectscontrolling community composition and activity but atpresent most are poorly characterized in natural marine sys-tems Bacteria must compete for dissolved and particulatecompounds with other bacteria andwith protistan taxa whilephagotrophic protists must compete with each other andmetazoa for available prey These interactions manifest them-selves in many forms including true competition for availableresources resource partitioning such that some taxa specializeon the utilization of certain substrates or prey while othersspecialize on different ones (reducing competition) or eventhe cooperative use of compounds (eg one bacteriumrsquoswaste products being utilized as substrate by another bacte-rium) These metabolic consortia are probably common inthe ocean but can be difficult to identify and characterizeConsortia of bacteria working together to perform complexreactions especially in anaerobic systems are well known inmicrobiology
Consortia in which there is a very high degree of integra-tion between species (ie symbiosis) are very common in theocean These associations may be beneficial to both partners(mutualism) beneficial to one part and inconsequential forthe other (commensalism) or beneficial to one part and det-rimental to the other (parasitism) Many highly visible andwell-characterized marine mutualisms exist between chemo-synthetic bacteria and animals such as hydrothermal ventor methane seep tubeworms clams mussels or other inverte-brates (365) and also between luminescent bacteria and fishor squids (366) Similarly many protozoa form mutualisticassociations with photosynthetic protists (367) The mostconspicuous of these associations involved large amoeboidprotozoa of oceanic ecosystems (planktonic foraminiferanspolycystine radiolaria and acantharia) which contain largenumbers of intracellular algae (74) but associations betweensome heterotrophic dinoflagellates and cyanobacteria are alsocommon (368) Studies have indicated that efficient nutrientcycling in these protozoan-algal associations confers an eco-logical advantage for both partners in surfacewaters of the oli-gotrophic ocean
Parasitism is also a common symbiotic relationship amongmarine protozoa Protozoan parasites have been reported for adiverse array of protists and animals in the ocean Heterotro-phic dinoflagellates and presently uncultured alveolate cladesrelated to alveolates appear to be particularly successful atevolving thismode of existence (369ndash371) In a common par-asitic group Amoebophrya spp the parasite invades photo-synthetic dinoflagellates where it undergoes reproductioneventually lysing the host and releasing dozens of infectiveswarmers The parasitic protist Pirsonia spp is a parasite ofdiatoms (372) The overall importance of these parasitismsin the population dynamics of their hosts is not clearly knownbut may constitute a significant source of mortality for somespecies (373 374) The frequency and diversity of ldquonewrdquo
FIGURE 8 Meta-analysis of experimental studies that have com-pared rates of bacterial productivity (converted to population growthrate units of dminus1) with rates of bacterivory by phagotrophic protists(expressed as equivalent units of bacterial growth rate dminus1) Thesolid line indicates where measurements of bacterial removal by graz-ers were equal tomeasurements of bacterial production Data redrawnfrom Sanders et al (63) doi1011289781555818821ch422f8
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-21
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
et al (7 18) However as with marine bacteria epifluores-cence microscopy facilitated the observation of small single-celled eukaryotes during the 1970s and 1980s and enabledeasy discrimination of protists without chloroplasts (proto-zoa) from those with chloroplasts (algae) based on the auto-fluorescence of photosynthetic pigments (19ndash21) Thedevelopment and refinement of this approach for eukaryoticmicroorganisms was instrumental in establishing the standingstocks of small protozoa which typically occur at abundancesof tens to thousands per ml in most marine ecosystems Thedevelopment of microscopy approaches for larger protozoasuch as heterotrophic dinoflagellates and ciliates (10 22)has been equally important in documenting abundances ofthese taxa that range up to tens per ml in much of the worldrsquosoceans
Estimates of the overall biomass of various microbialassemblages and techniques to measure rate processes (egrates of growth substrate uptake prey consumption) beganto appear in the late 1970s and 1980s and the refinementof these estimates and measurements continue to the presentday Early attempts to measure bacterial growth rates in sea-water involved ldquoindirectrdquo methods such as relating the fre-quency of dividing cells to rates of division in culturedstrains (23) Isotope-uptake based approaches specificallythe incorporation of radioactively labeled thymidine intoDNA (24 25) andor the incorporation of leucine into pro-tein (26) have become the most commonly used methods(see ldquoEstimating lsquoBacterialrsquo Biomass and lsquoBacterial Produc-tionrsquordquo) These methods have indicated that bacterial dou-bling times can be on the order of one day in coastaltemperate waters Combined with estimates of bacterial bio-mass these results led to the conclusion that bacteria mustbe consuming a substantial proportionmdashon the order of50mdashof the total system primary productivity A similarconclusion was reached using direct estimation of microbialrespiration by careful measurements of oxygen concentrationchanges (micro-Winkler method) in seawater that had beenprefiltered through 5 microm pore filters to remove animals andmany of the protists (14)
During this same period small protozoa (primarily flagel-lates and ciliates) were gaining recognition as importantconsumers of bacteria in the marine plankton and benthos(27ndash29) An increasing volume of experimental work dem-onstrated a dominant role for small bacterivorous protozoaas a mechanism for removing bacterial production andrepackaging bacteria into larger particles that might be con-sumed by metazoan zooplankton Also it became recognizedaround this time that a significant fraction of the phytoplank-ton biomass and production was consumed directly by herbiv-orous protozoa rather than by metazoan zooplankton such ascopepods (30 31) Consequently heterotrophic protists wereacknowledged as an important food source for a variety ofmetazoan zooplankton and numerous experimental studiessubsequently demonstrated this trophic connection (32 33)
These observations were synthesized in a second bench-mark paper (34) The latter publication marked the begin-ning of the widespread recognition and use of the termldquomicrobial looprdquo in marine planktonic systems a conceptthat emphasizes the remarkable importance of the tiniestorganisms as well as dissolved organic matter as an intermedi-ate in material and energy transfer in aquatic ecosystems Anupdated illustration of this basic concept is shown in Fig 1
Definitions and ConceptsAbundance the number of individuals in a sample or a
population
Algae protists that exhibit phototrophic nutrition Likeprotozoa algae span a wide size range (lt1 to gt200microm) and have generally been referred to as phototrophicpico- nano- or microplankton
Amensalism interaction where members of a speciesinflicts harm to another species without any costs or ben-efits received by the other
Autotroph an organism that uses carbon dioxide as itssource of structural carbon
Biomass the mass of living organisms within a populationcommunity or ecosystem
Chemolithotroph an organism that uses reduced inorganicmolecules as its energy source
Chemoorganotroph an organism that uses reduced organiccarbon as its energy source
Commensalism interaction between species where onebenefits from the other but the other is not affected
Competition species-species interactions that have a nega-tive effect on both species
Cyanobacteria Prokaryotic photosynthetic organisms thatcontain chlorophyll a and generate oxygen during photo-synthesis The free-living ancestors of primary chloroplasts
Exploitation species-species interactions that have a nega-tive effect on one species while benefiting the othermdashcan include both parasitism and predation
Heterotroph an organism that uses preformed organic car-bon as its source of structural carbon also heterotrophicor heterotrophy when applied to metabolism
Mixotroph any of a number of types of organisms that com-bine (in one organism) multiple metabolic types asdescribed above For example a protist that consumes bac-teria as prey (heterotroph) but also contains functioningchloroplasts (phototroph) will often be referred to as amixotroph Similarly an archaeon that oxidizes ammoniaforenergy (chemolithotroph)butusesaminoacidstobuildproteins (heterotroph) could be considered a mixotroph
Mutualism interaction between species where both benefitfrom each other
Phototroph an organism that uses light as its energy sourcefor production of ATP (or to produce proton gradients inthe case of rhodoposin-based phototropy) and some-times also reducing power fromwater (in cyanobacteria)
Phytoplankton the photoautotrophic component of theplankton including cyanobacteria and a large numberof eukaryotic phyla that contain chloroplasts
Protists eukaryotic species that can exist as a single cellother than a spore gamete or zygote (although thereare many that form colonies)
Protozoa protists that exhibit heterotrophic nutrition Pro-tozoa span a wide size range (asymp2 to gt200 microm) and havegenerally been referred to as nano- or microzooplankton
Relative abundance (and the related term evenness) thecontribution of each species or operational taxonomicunit to a community
Species diversity a complex concept composed of speciesrichness and relative abundance
Species richness the number of different species or opera-tional taxonomic units present in a sample habitat orenvironment
Stoichiometry Studies that involve calculation of the rela-tive quantities of elements or compounds for exampleCNP ratios
422-2 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
Symbiosis the intimate living together of two kinds oforganisms especially if such an association is of mutualadvantagemdashtoo vague to be of use in quantitativedescriptions of population interactions but very usefulin indicating a close association among organisms
Syntrophy a metabolic mutualism where one species usesthe waste product produced by the other and in so doingallows both metabolic pathways to be energeticallyfeasible
Zooplankton Planktonic eukaryotes that consume otherplankton Includes single-celled organisms (protozoaor protists) and metazoans and some that are planktoniconly as larvae
Estimating ldquoBacterialrdquo Biomass and ldquoBacterialProductionrdquo Definitions and MethodsAquatic microbiologists tend to use the term ldquobacteriardquowith alowercase ldquobrdquo to describe organisms that appear to be prokary-otic by microscopymdashthat is organisms with no membrane-bound nucleus They include members of the taxonomicdomains Bacteria and Archaea (see ldquoBacteria and Archaeardquo)Organisms within and between these domains differ in manybiochemical and genetic aspects but they tend to look similarby traditional epifluorescence microscopy Special methodssuch as different versions of fluorescence in situ hybridization(FISH) are required to distinguish individual members ofthese domains microscopically (35ndash37)
The term ldquobacterial productionrdquo here refers to heterotro-phic production of biomass by bacteria It is meant to include
production of nonphotosynthetic bacterial biomass based onthe heterotrophic consumption of preformed organic matter(ie organic matter in various forms that has been producedprimarily by phytoplankton)
Bacterial biomass is usually determined by convertingdirect counts of bacteria using an estimate of the amount ofcarbon per cell Direct counts are most commonly done byepifluorescence microscopy with stains such as acridineorange 406-diamidino-2-phenylindole or SYBR green I(17 38 39) Special procedures are usually applied for sedi-ment samples and samples containing large numbers of bacte-ria attached to particles (40) SYBR green I also permits directvisualization and counts of viruses in the same preparationIncreasingly direct bacterial counts in seawater sampleshave been performed by flow cytometry of fluorochrome-stained cells (41 42) a method that allows separate countsof cyanobacteria such as Synechococcus and Prochlorococcuswhich have unique fluorescent signatures due to their photo-synthetic pigments and which can sometimes make up a sub-stantial fraction of the total number of bacteria (43) Flowcytometry is rapid and has a statistical advantage in that it typ-ically observes thousands of prokaryotic and minute photo-synthetic eukaryotic cells rather than the hundreds countedmicroscopically drawbacks include the cost of the instrumentand the fact that cells attached to each other or to other par-ticles are counted as one Bacterial carbon per cell has beenestimated in a variety of ways most commonly from a deter-mination of cell volume and carbon density per unit volumeThese numbers are difficult to obtain accurately for nativemarine bacteria which are very small typically 05 μm in
FIGURE1 An early vision of the ldquomicrobial looprdquo and its connections to the classical grazing food chain via dissolved organicmatter (DOM)flux and particulate trophic transfer with viruses included as a side loop Modified from (34) Large gray arrows indicate the flow of organiccarbon and energy into higher trophic levels of the food web with recognition of the important roles for heterotrophic microbes (bacteriaand protozoa) in this process Large stippled arrows indicate the production of DOM via excretion and trophic interactions (not all groupsare represented) Thin dotted arrows indicate mineralization of major nutrients contained in organic matter respired by consumers Whitearrows indicate bacteria lysis by viruses and DOM released by that process doi1011289781555818821ch422f1
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-3
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
diameter (range is about 02ndash1 μm for free-living unicells)Published estimates of bacterial carbon per cell vary widelyand probably constitute the greatest uncertainty with estimat-ing bacterial biomass in natural samples Typical estimates ofthe carbon content of a bacterium range from 7 to 50 fg C(1 fg is 10minus15 g) with most open ocean estimates near 10ndash20 fg C per cell and coastal ones about double that (44)Thus in a typical mesotrophic ocean environment with109 bacteria per liter and an average per cell C contentof 15 fg bacterial biomass would be approximately 109 times15 times 10minus14 = 15 times 10minus5 g C per liter or 15 μg C per liter
Bacterial production is most often measured by incorpora-tion of tritiated thymidine into DNA (24 45) or tritiated leu-cine into protein (26 46) Thymidine and leucine areintracellular precursors of DNA and protein respectively soincorporation of these precursors can be used to estimatethe total rates of synthesis of themacromolecules DNA is syn-thesized for cell division and protein synthesized roughly inproportion to total biomass someasuring their rates of synthe-sis is presumed to track production Both methods have beencalibrated on the basis of theoretical considerations as well aspurely empirical approaches and both methods yield similarresults (44) Leucine has a lower detection limit so it is pre-ferred for slower rates The results of thymidine incorporationare most often presented as cells produced per liter per hourwhich can be converted to a carbon production rate via esti-mates of carbon per cell as noted Leucine incorporation maybe used to calculate cell production as well as biomass produc-tion directly because protein is a major biomass constituent
Geographic and Temporal Distributions ofMicrobial Biomass and ActivityBacterial abundances by epifluorescence microscopy showthat bacteria are present in most marine euphotic zone
environments at approximately 106 cells per ml Samplesfrom around the world rarely vary by more than threefoldfrom this typical value (ie rarely lt3 times 105 or gt3 times 106)which is extraordinary compared to phytoplankton and zoo-plankton whichmay vary by several orders of magnitude overthe same spatial scales However despite this remarkable gen-eral predictability there is significant variation across bothspace and time More nutrient-rich eutrophic environmentstend to have more bacteria (sometimes gt107 per ml 47) andoligotrophic open ocean environments have less (summar-ized below) Although bacterial assemblages in warm temper-ate coastal waters may have doubling times as short as 1 daythis is at the rapid end of the spectrum of in situ growth ratesBacterial assemblages in the open sea especially in oligotro-phic environments have average generation times typicallyof a week or perhaps more (see Table 1) These abundancesgenerally apply to the euphotic zone and bacteria in colderdarker waters have substantially lower abundances and slowergrowth rates than those of surface waters (48) Benthic bacte-ria also exhibit fairly constant abundances across wide geo-graphic ranges but benthic bacteria occur at much higherdensities because of the organic-rich and particle-ladennature of the environment (an average of asymp109 per ml fluidvolume is typical 49) This value is three orders of magnitudegreater than abundances in the water column implying thatthe abundances of benthic bacterial assemblages are regulatedby a different set of parameters than planktonic assemblages
Bacterial biomass and productivity vary temporally on anumber of scales ranging from diel (50) to seasonal (51) orinterannual (52) On time scales of hours bacterial abun-dance and production have been shown to often peak inthe middle of the day and be low in the middle of the night(50 53ndash55) This pattern has been interpreted as a tight cou-pling between the production of labile organic compoundsvia photosynthesis and bacterial growth on one hand and
TABLE 1 Bacterioplankton properties in relation to phytoplankton in the open sea as compiled by Ducklow (44)
Property N Atlantica Eq Pac-Sprb Eq Pac-Fallc Sub N Pacd Arabiane Hawaiif Bermudag Ross Seah
Euphotic zone m 50 120 120 80 74 175 140 45
Biomass (mg C mminus2)
Bacteria 1000 1200 1467 1142 1448 1500 1317 217
Phytoplankton 4500 1700 1940 1274 1248 447 573 11450
BP 02 07 075 09 12 36 27 002
Production (mg C mminus2 dminus1)
Bacteria 275 285 176 56 257 Nd 70 55
Phytoplankton 1083 1083 1548 629 1165 486i 465 1248
BP 025 026 011 009 022 Nd 018 004
Growth rates (dminus1)
Bacteria 03 013 012 005 018 Nd 005 025
Phytoplankton 03 064 08 05 093 11 081 011
BP 1 02 015 01 019 Nd 006 23
Notes All bacterial biomass estimates based on 20 fg C per cell Data may overestimate heterotrophic bacterial biomass as a consequence of lower C per cell orinterference by Prochlorococcus and Archaea Production estimated from 3000 g C per mole leucine incorporated
aEastern North Atlantic spring phytoplankton bloom 47 N 20 W May 1989 n = 13bEquatorial Pacific 0 N 140 W MarchndashApril 1992 n = 8cEquatorial Pacific 0 N 140 W SeptemberndashOctober 1992 n = 19dSubarctic North Pacific 45 NeNorthwest Arabian Sea 10ndash20 N 165 E JanuaryndashDecember 1995 n = 21fHawaii Ocean Time Series (HOT) 1995ndash1997 n = 21 (httphahanasoesthawaiieduhothot_jgofshtml)gBermuda Atlantic Time Series (BATS) 1991ndash1998 n = 106 paired comparisons The ratios are means of the ratios not ratios of the means BP calculated
from thymidine incorporation (16 times 1018 cells per mole incorporated)hRoss Sea Antarctica 76 S 180 W 1994ndash1997i1989ndash1996 n = 64
422-4 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
bacterial mortality via grazers or viral lysis on the other handIt is consistent with some measurements of extremely rapidturnover sometimes several times per hour in rich coastalwaters of labile dissolved organic compounds such as dis-solved free amino acids (56) and might also indicate greaterpredation pressure by protozoa during the night On longertime scales of weeks to months bacteria show distinct sea-sonal patterns For example in temperate coastal waters bac-terial biomass and production increase considerably insummer months compared with winter However bacteriado not typically show a significant increase during earlyldquospringrdquo blooms in temperate waters when water is still verycold (51) It has been hypothesized that this phenomenonis the result of the suppression of the rate of substrate uptakeby temperate bacterial assemblages at low temperature (57)However while temperature probably has the effect of settinga limit on maximal growth rates (as for phytoplankton [58]and protozoa [59]) temperature alone does not appear to bethe main factor controlling growth of marine bacteria undermost circumstances (see ldquoLight Temperature and Pressurerdquo)
It would be overly simplistic to think that all the bacteriaand archaea in a sample or habitat have the same level ofactivity per cell but it is also easy to think of measured activ-ities as characteristic of all members of a microbial assem-blage So the question arises are most of the cells active ata similar level or are some hyperactive while others are com-pletely dead or moribund This question has been addressedseveral ways includingmicroautoradiography selective stain-ing ldquodirect viable countsrdquo (where nutrients are added to seewhat part of the community grows) and in situ hybridization(eg 60) Based on these contrasting approaches it appearsthat a continuum of activity exists within bacterial assemb-lages from truly dead (cannot be revived) to extremely activeA reasonable interpretation of the existing data is that undertypical conditions a small percentage of the marine bacterialcells perhaps 10ndash20 are generally inactive or dead the plu-rality or majority of cells perhaps 25ndash75 are intact andhave some moderate level of activity and a small percentageperhaps 5ndash20 are highly active It is useful to consider thisspectrum conceptually and numerically when modelingmicrobial processes
Comparisons of bacterial and phytoplankton biomasswithin planktonic ecosystems show that these are positivelycorrelated across broad scales Analyses of marine and fresh-water samples from several studies (61 62) have shown thatbacterial abundance increases with chlorophyll concentra-tion at least at the level of a log-log relationship Similarlybacterial abundances and the abundances of small protozoacorrelate over broad spatial and temporal scales (63) Theserelationships are sensible in that on the largest scale primaryproduction is the source of organic material that fuels hetero-trophic bacterial activity and bacteria constitute the prey ofmany small protozoa Individual data sets also have sometimesshown strong correlations between bacterial abundance andchlorophyll (eg 45) but variability in this relationshipover short temporal or spatial scales is to be expected It wouldpresumably be a consequence of rapid short-term changes inthe rate of substrate supply as well as the normal oscillatorynature of predator-prey relationships between bacteria andtheir consumers
Interestingly the extrapolation of the positive log-log rela-tionship between bacterial and phytoplankton biomassto environments with very low chlorophyll concentrations(eg ultra-oligotrophic oceans) indicates that bacterial bio-mass may exceed phytoplankton biomass in these situationsWhile this conclusion is obvious for the deep sea where there
is no photosynthesis this situation is also often true in oligo-trophic surface waters Measurements made in oligotrophicwaters bear out the high bacterial contribution to totalbiomass (64 65) Moreover Cho and Azam (66) confirmeda linear relationship between the log of chlorophyll and logof bacterial abundance but only at chlorophyll concen-trations above approximately 05 μg per liter Below thatconcentration bacterial abundance did not correlate signifi-cantly with chlorophyll It should be noted that subsequentanalyses have revealed that early epifluorescence measure-ments of bacterial biomass included the common cyanobacte-rium Prochlorococcus which can make up to 20 of totalbacterial numbers (67) Nonetheless heterotrophic bacterialbiomass is a major fraction of the living biomass of all plank-tonic ecosystems
The geographical and temporal distributions of marineprotozoa are much more varied than those of the bacteriaAs an all-inclusive group protozoa generally occur in plank-tonic ecosystems at abundances ranging from 10 s to 1000 sper ml Abundances in benthic ecosystems can be one tothree orders of magnitude higher commensurate with thehigher abundances of bacteria in those ecosystems Howeverit is important to remember that like the term ldquolsquobacteriardquo theterm ldquoprotozoardquo is a rather artificial conglomeration of evolu-tionarily and ecologically divergent taxa (see ldquoThe Changingand Complex World of Eukaryote Phylogenyrdquo) Thus theabundances of specific lineages of bacteria or protozoa mayshow spatial (or temporal) variability that is considerablygreater than the variability characteristic of these overarchinggroupings
The Changing and Complex World ofEukaryote PhylogenyNot that long ago textbooks still divided eukaryotic organ-isms into four major kingdoms (Animalia Plantae Fungiand Protista) while prokaryotic organisms were placed intoa single kingdom theMonera (68)Within this scheme pro-tists (eukaryotic organisms that can exist as single cells) weredivided into two subkingdoms (algae and protozoa) based ontheir basic nutritional mode a carryover from the historicaldistinction between single cells with ldquoanimal-likerdquo or ldquoplant-likerdquo nutrition This distinction presupposed a basic evolu-tionary divergence among protists into species that retaineda heterotrophic phagocytotic mode of life (protozoa) andthose that abandoned phagocytosis for a photosyntheticmode of life (algae)Moreover the presenceabsence of chlor-oplasts was a feature that could be easily distinguished by earlymicroscopists
The five-kingdom classification system of Whittaker wasrecognized as an improvement over previous classificationschemes but it posed a number of problems relating toprotists For example the distinction between single-celledand multicellular eukaryotes was somewhat arbitrary Moreimportant the division of protists based on whether theywere heterotrophic or photosynthetic was clearly not anappropriate feature if the classification was to recapitulateevolutionary relationships We now know that chloroplastacquisition and loss has occurred several times in the bio-logical history of our planet (69) giving rise to some closelyrelated protistan taxa that differ largely in the presence orabsence of a chloroplast Further complicating the matterwithin many protistan lineages there are species that possesschloroplasts and carry out photosynthesis (phototrophy)but also possess the ability to ingest and digest prey (hetero-trophy 70ndash72) Some heterotrophic protists even ingest
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-5
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
phytoplankton prey and retain the chloroplasts of their prey ina functional state for a limited amount of time (kleptidoplas-tidy73) Various forms and degrees of mixotrophy (mixedphototrophic and heterotrophic nutrition) are commonamong a number of algalprotozoan lineages (74ndash77) UnderWhittakerrsquos scheme phytoplankton ecologists studying a lin-eage of microalgae might have had little familiarity withclosely related heterotrophic species while protozoologistsstudying a particular protozoan groupmight have known littleabout closely related photosynthetic species
One might expect given these caveats that the termsldquoalgaerdquo and ldquoprotozoardquo are no longer used In fact the termldquoprotozoardquo is still commonly used (especially by ecologists)to recognize those eukaryotic species that exist as single cellsand whose nutrition is dependent on the uptake of preformedorganic substances (primarily via prey ingestion) while pro-tists possessing chloroplasts are still commonly called ldquoalgaerdquoSimilarly although the term ldquoprotistrdquo has been abandoned asa kingdom designation it is still widely employed to describeeukaryotic species that are capable of existence as single cells(ie algae and protozoa) The term ldquophagotrophic protistrdquohas also gained popularity in recent years because it recognizesthat many protistan species are capable of phagocytosis eventhough they may also possess their own chloroplasts and thusare technically ldquoalgaerdquo
Despite the shortcomings of Whittakerrsquos scheme it domi-nated the hierarchical organization of life for approximately aquarter century During the past few decades however thissystem has given way to a new organizational scheme that rec-ognizes three domains of life (Archaea Bacteria Eukarya [orEucarya] 78 Fig 2 upper panel) and is based onwhat is pres-ently believed to reflect amore realistic view of the evolution-ary distances that have developed between organisms in theasymp4 billion years that life has existed on our planet Withinthe Eukarya of Woesersquos scheme hypotheses regarding thephylogeny of ldquoprotistsrdquo have changed continuously and rap-idly during the past two decades reflecting new insightsinto eukaryote evolution provided largely by DNA sequenceinformation (Fig 2 lower panel from [79])
The former protistan phyla ofWhittakerrsquos systemhavenowbeen dispersed among candidate ldquosupergroupsrdquo within thedomain Eukarya to better reflect hypothesized phylogeneticrelationships For example the dinoflagellates (which encom-pass phototrophic heterotrophic and mixotrophic species)now form a single group and have been placed together withthe ciliates and apicomplexans (sporozoans) in themonophy-letic Alveolata (Fig 2 lower panel) On the other handeukaryotic heterotrophic single-celled species falling withinthe general description ldquoprotozoardquo are now widely distributedamong a number of protistan lineages In short nutrition hasbeen demoted as a phylogenetic character and other charac-ters (presumably more indicative of evolutionary relatedness)have ascended to address some long-standing contradictionsalthough the debate over the relationships among some line-ages is still very active at the present time 79)
Individual protozoan cells range in size from less than 2 micromto greater than 1 cm in diameter (gt4 orders of magnitude)(80 81) with some colonial radiolaria forming cylindricalgelatinous structures a centimeter in diameter and morethan a meter in length (82 83) Because they constitutesuch a large size range of organisms protozoa are often dividedinto size classes that very crudely correlate with their generalnutritional preferences A commonly used convention is thatof Sieburth et al (84) which groups planktonic microorgan-isms into order-of-magnitude size classes (02ndash20 microm =picoplankton 20ndash20 microm = nanoplankton 20ndash200 microm =
microplankton 02ndash2 mm =mesoplankton) Most protozoafall into the nanoplankton or microplankton size classesModeling microbial trophodynamics using this conventionassumes that protozoa in one size category generally consumeprey one order of magnitude smaller in size (34 85)Although this approach misses much of the detail and diver-sity of the trophic activities of individual protozoan taxa it isa necessary practical compromise for examining community-scale flows of energy and elements It also provides a usefulmechanism for summarizing and comparing the abundancesand biomasses of protozoa from different environments andto other microbial assemblages Protozoan abundance sum-marized in this way has been shown to contribute signifi-cantly to the living biomass of planktonic ecosystemsthroughout the world ocean (Fig 3)
Estimates of protozoan biomass such as those depicted inFig 3 typically do not include the contribution of mixotro-phic phytoflagellates to heterotrophy There is presently noeasy way to determine the abundances of small phagotrophicphytoflagellates in natural samples so these species are typi-cally counted as phytoplankton unless specific methods areemployed to identify the algae as consumers such as the useof fluorescently labeled particles (87ndash90) or through theexamination of food vacuole contents (91) On averagethese species appear to constitute a modest percentage ofthe phytoplankton assemblage (typically lt25) althoughthey may at times dominate the phototroph assemblages ofnatural plankton communities It is important to recognizethat their inclusion as functional heterotrophs rather thanphototrophs could significantly shift the relative contribu-tions of phototrophic and heterotrophic microbial biomassto total biomass within microbial assemblages and the flowof energy within plankton communities (92)
Heterotrophic protists that harbor photosynthetic pro-tists or their chloroplasts within their cytoplasm constituteanother complexity for estimating the contribution of pro-tozoa to total microbial biomass When bulk water samplesare analyzed the contribution of chlorophyll containedwithin those protozoa is generally assumed to come fromfree-living phytoplankton However studies have shownthat chloroplast-bearing ciliates can contribute up to halfthe total biomass of planktonic ciliates in ecosystems andchloroplast-retaining ciliates can sporadically dominate thechlorophyll and primary production of some planktonic eco-systems (93ndash96) The environmental conditions promotingthe success of these ciliates are poorly known Similarlymany species of planktonic foraminifera polycystine radio-laria and acantharia harbor large numbers (thousands perprotozoan) of endosymbiotic algae within their cytoplasm(97 98) Caron et al (99) have demonstrated that primaryproductivity within these species can contribute significantlyto total primary productivity in oceanic ecosystems and can bevery important locally in the convergences of Langmuir circu-lation cells (100)
BACTERIA AND ARCHAEAldquoCulturablerdquo versus ldquoNonculturablerdquo CellsMost conventional cultivation methods can grow only 1 orless of the bacteria that can be visualized by direct microscopytechniques (eg 4) This is true even though most can beshown to be active by techniques such as microautoradiogra-phy (25) These readily cultivable organisms appear to repre-sent a group of fast-growing so-called weeds that are adaptedto take advantage of rapid growth in rare organically enriched
422-6 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
environments This strategy contrasts with the numericallydominant bacteria that are adapted specifically for growthin the dilute nutrient conditions that characterize the vastmajority of the volume of the water column
The most common taxa readily cultured from seawaterwith standard nutrient broth media include the gamma pro-teobacterial genera Vibrio Alteromonas PseudoalteromonasMarinomonas Oceanospirillum Shewanella (usually isolatedfrom surfaces such as shellfish and sediments) the alpha pro-teobacterial generaRoseobacterSphingomonasmembers of thefamily Flavobacteriaceae and Planctomycetes as summarizedin Giovannoni and Rappeacute (101) and Fuhrman and Hagstrom(102) The cyanobacteria Synechococcus and Prochlorococcusare also now readily culturable but on low-nutrient inorganicmedia targeting photosynthetic forms as opposed to organicmedia used to cultivate the others listed above
Because of the low percentage of marine bacteria that canbe grown in standard media organisms that until recentlywere called ldquononculturablerdquo make up the large majority ofbacteria in the plankton Only during the pastsim20ndash25 yearshave molecular biological methods based on 16S rRNA genesequences been available to identify these organisms andthese powerful techniques have opened up a large area forexploration (see next section) Similar but more recent stud-ies use 18S rRNA sequences for characterizing protistandiversity as will be noted below
Molecular Phylogeny and MetagenomicsField ApplicationsModern phylogeny of microorganisms is based primarilyon genetic sequences the most well-studied gene being the
FIGURE 2 The three domains of life (upper left) as proposed by Woese et al (78) and a recent overview of modifications that have beenproposed by Adl et al (79) to higher-level phylogentic groups within the eukaryotic component of the tree (lower right) Domains figure fromWoese et al (78) eukaryotic tree figure from Adl et al (79) doi1011289781555818821ch422f2
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-7
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
small subunit ribosomal RNA gene (16S rRNA in Bacteriaand Archaea and its larger homolog 18S rRNA in eukar-yotes) This molecule is strongly conserved over evolutionarytime so this single molecule has been used for constructingphylogenetic trees of all living organisms (httptolweborgtreephylogenyhtml) Analysis of 16S18S rRNA genesequences has been used to evaluate deep evolutionary rela-tionships among organisms and was instrumental in point-ing out that Archaea Bacteria and Eukarya should beconsidered different Domains of equivalent phylogeneticrank above kingdoms (78) However there are sufficientdifferences in 16S18S rRNA gene sequences to demonstrate
relationships (ormake distinctions) at the genus or sometimesspecies level
The first phylogenetic studies based on 16S18S rRNAgenes used sequences derived from cultures However onedoes not need cultures to obtain rRNA gene sequences (orany other sequences for that matter) An idea developed inthe lab of Norman Pace in themid-1980s involved extractionof DNA directly from natural samples and then cloning andsequencing of the DNA as a means of assaying the microbespresent in the samples (103 104) The original protocolscalled for cloning by creating what are called ldquophage librariesrdquofrom the natural DNA but since 1986 PCR has been applied
FIGURE 3 (a b) Plankton biomass in the Arabian Sea during the 1995 southwest monsoon (a) and intermonsoon period (b) Areas of theboxes indicate the relative magnitudes of the biomass in each category Categories within the dashed boxes in a b c are composed of protozoaArrows indicate the direction of energymaterial flow in the food web thicker arrows depicting greater flow Redrawn from (86)(c) Depth-integrated biomass (mgm2) in the upper 100 m of the Sargasso Sea near Bermuda and in the upper 200 m of the equatorial Pacificat 175degE Thewidth of the bars indicates the biomass in each size category Heterotrophs have been separated by size class while phytoplanktonhave not Size classes delineated by the dotted box are comprised of protozoa Redrawn from (65) doi1011289781555818821ch422f3
422-8 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
extensively for cloning and related studies The targetsequence can be almost instantly ldquoidentifiedrdquo to its closestphylogenetic neighbor by what have come to be standardonline sequence comparisons
Beyond the study of targeted genes (like the 16S rRNAgene) shotgun metagenomic studies have examined theentire genetic repertoire of the microbes in a given sampleThe metagenome is the collective genome of all organismsin the sample Initially these studies extracted DNA fromall organisms in a sample (usually prefiltered through a filterapproximately 1 microm to remove most organisms larger thanbacteria) sheared it to produce fragments and clonedthem into standard vectors either as small (thousands ofbases) or large (to hundreds of thousands of bases) insertsthat is fragments of DNA from the environment nowcloned into the vectors in a form suitable for sequencingThe best known early marine study of the former type isthe Global Ocean Survey with initial results published byVenter et al (105) which generated more than a billionbases of DNA sequence and reported 12 million previouslyunsequenced genes estimated to come from at least 1800different genomic species cumulatively in the many samplesthey analyzed
With the advent of next-generation sequencing that gen-erates millions ormore sequences in a run (known by a varietyof acronyms including 454 Illumina SOLiD etc) cloningof genes has largely been replaced by clone-free sequencingThe extent and power of such sequencing has recentlybeen demonstrated by the ability to construct essentiallythe entire genome of an uncultivated marine Group II Eur-yarchaeon that constituted only approximately 2 of amicrobial community sample using SOLiD sequencingmade possible by high coverage and the use of mate pairsequencing ofsim3000 bp fragments (106) (this length isneeded to span repeats and highly conserved genes that oth-erwise make genome construction difficult)
For 16S rRNA gene studies ldquotag sequencingrdquo pioneeredby Sogin et al (107) consists of amplifying a suitable sizedpart of the gene with broadly conserved primers (choice isimportantmdashfew are truly universal for the groups intended)often ldquobarcodedrdquo to allow multiple samples to be combinedinto a single run The amplified products are then sequencedand sequences processed en masse Many thousands of partialSSU rRNA gene sequences per sample are economically ana-lyzed this way though they are usually short (currently a fewhundred bases each depending on the sequencing platform)This way of analyzing the composition of microbial commun-ities is now standard These approaches provide so muchinformation even about very rare sequences that the resultshave led to the important concept of the ldquorare biosphererdquoorganisms that may be active or dormant and constituting avery small proportion of the community (eg often muchless than 01) but are potentially important for dispersionadaptation to changing conditions or even critical activitieslike nitrogen fixation or vitamin production (108) Howeverdue to the potential for artifacts like error sequences thisapproach requires rigorous application of quality filteringand clustering algorithms to avoid erroneous taxa and overes-timation of species richness (109 110)
Next-generation sequencing approaches have also allowedwork to begin metagenomics studies of microbial eukaryotes(111) These studies are still constrained by the much largergenomes of eukaryotes and therefore the difficulties ofobtaining sufficient sequences to reconstruct the genome ofa particular species Along with advances in sequencing abil-ity studies of eukaryotic metagenomics have been facilitated
by advances in single-cell isolation which when coupledwith high-throughput sequencing approaches reduce the tre-mendous complexity present in natural complex eukaryoticcommunities to a manageable task (112) Such single-celltechniques are also quite valuable in studies of bacteria andarchaea though the amplification technique tends to bevery uneven and typically generates less than half the genomeof each isolated cell regardless of domain (113 114)
Additionally similar to DNA mRNA is amenable toextraction and sequencing although greater care must betaken during extraction and purification as RNA shows agreater susceptibility to degradation during processing Copy-ing of mRNA by reverse transcription of RNA into cDNAfollowed by DNA sequencing has allowed insights into themetatranscriptomes of environmental samples Metatran-scriptomic studies provide information on gene expressionin an ecosystem and thereby indicates ldquoactivityrdquo of themicro-bial community rather than simply ldquopotentialrdquo represented bythe genomic DNA present in the sample with many applica-tions from showing which processes are being carried outby which organisms to fine-scaled diel studies (115ndash118)Nevertheless given variations in the lifetimes of differenttranscripts and protein molecules the transcriptome maynot be fully representative of the current activity of anorganism
Limitations of these analyses include sequencing errorsPCR mismatches or biases clustering and bioinformaticschallenges and chimeras generated during PCR Also thephylogenetic resolution of short sequences is limited giventhe high conservation of rRNA sequences Even with clone-free shotgun metagenomics there may be biases such as non-random losses of DNA during extraction and preparationor biases (eg from G +C content or secondary structure)in the sequencing procedures Determination of speciesdiversity by these approaches provides an example of theselimitations Shakya et al (119) working with synthetic com-munities (purified genomicDNA from16Archaea represent-ing 3 phyla and 48 Bacteria representing 16 phyla remixed tosimulate an environmental DNA extract) applied both meta-genomic analysis (454 and Illumina platforms) and PCRamplification followed by 454 sequencing of 16S rRNA genesto determine both species richness and relative abundanceThey found that PCR amplification454 sequencing of 16SrRNA genes yielded an accurate measure of species richness(providing that appropriate data processing was applied)but that the relative abundance of up to 94 of the species(depending on domain and variable region amplified) wasover- or underestimated by at least 15-fold (values rangedfrom not detected to 103-fold overestimation) In contrastboth metagenomic approaches yielded relative abundancesthat were within the authorsrsquo 15-fold accuracy cutoff forsim50 of the species However they concluded that addressingrichness overestimation in metagenomic analyses that isdistinguishing rare but real OTUs from experimental andcomputational artifacts awaits further computational andclassification improvements More recently Parada et al(120) used mock communities composed of 16S rRNAclones from 27 common marine taxa (from nine Bacterialand two Archaeal phyla) to show that small differences inPCR primers (and different clustering methods) can yieldlarge differences in apparent relative abundances of reportedtaxa However one primer pair and informatics pipelinethey tested using a particular version of 515F-926R (V4ndashV5) provided accurate estimates of relative clone abundance(r2 = 095) when comparing observed versus expected cloneabundance
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-9
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
Culture-Independent Diversity Studies
The first groups to be identified using cultivation-independ-ent molecular techniques from the marine plankton (121)were the bacterial SAR11 cluster (a group of closely relatedgene sequences or phylotypes) and marine picoplanktonicunicellular cyanobacteria Synechococcus and Prochlorococcus)Of these the SAR11 cluster was completely unknown butthe cyanobacteria had previously been recognized by theirunique pigment fluorescence Waterbury et al (122) andJohnson et al (123) used epifluorescence to observe Synecho-coccus and Chisholm et al (124) discovered Prochlorococcususing flow cytometry These cyanobacteria were later isolatedand grown in phytoplankton culturemedia These two groupsare generally common in the euphotic zone with SAR11 typ-ically comprising one third of the planktonic bacteria (125)and the cyanobacteria common everywhere but polar waters
Probably the biggest surprise to come from the applicationof molecular tools was the discovery by Fuhrman et al (126)of abundant archaea in the deep sea The archaea were foundto be in a unique phylogenetic cluster that was only distantlyrelated to any previously known archaea but the ldquoclosest rel-ativesrdquo (not really close at all) were extreme thermophiles Asubsequent study also found archaea to be present in near-surface coastal waters albeit relatively rare (lt2 relativeabundance) This study used a PCR technique specificallytargeted archaea and found ldquomarine Crenarchaeardquo plus a sec-ond group belonging to the phylum Euryarchaea (127)
Up until this time all known Archaea were thought to beldquoextremophilesrdquomdashadapted for either very high temperatures(thermophiles) extremely salty conditions (halophiles) orstrictly anaerobic environments (methanogens) Yet theseorganisms were present in cold or cool water at ordinary salin-ity and high oxygen concentrations Fluorescent in situhybridization (FISH) measurements from deep-sea sampleshave since showed that the archaea may make up appro-ximately 40 of the total countable prokaryotes with thepercentage reaching to 60 at 200 m depth in theMediterra-nean (35 36) An extensive time series of FISH measure-ments near Hawaii confirmed that the archaea are indeedvery abundant throughout the year from below the photiczone to at least 4000 m and typically constituting 30ndash40of the total prokaryotes present in waters deeper than a fewhundred meters (128) with similar results found elsewhere(129 130) (Fig 4) They have been reported from many pla-ces including the Atlantic Pacific and Southern Oceansand are dynamic components of the plankton the most com-mon type by far being the ldquomarine Crenarcheaeardquo (128 129131) While the marine archaea have been reported to bedominated by a few major ldquophylotypesrdquo (132) they alsohave been shown to have a great deal of microdiversity withinthese phylotypes suggesting there are many kinds of close rel-atives coexisting (133) Interestingly this group of archaeamay likely be the most abundant kind of organism on Earthgiven the huge volume of the deep sea and their high abun-dance there (134) It has recently been proposed that ldquomarineCrenarchaeardquo be elevated to their own major phylum outsidethe Crenarchaeota called the Thaumarchaeota on the basisof deep phylogenetic branching and fundamental differencesbetween them and the Crenarchaeota (135) The Thau-marchaeota possess the uniquely archaeal membrane lipidcrenarchaeol and are nowalso known to be abundant in soils
The physiology of the Thaumarchaeota has been anintriguing area of study Initially an autoradiography-FISHcombination approach demonstrated that they take upamino acids (130 136) But subsequent data showed that
crenarchaeol was derived largely from CO2 suggesting auto-trophic metabolism (137 138) An experiment showinguptake of 13C-labeled bicarbonate into these archaeal lipids(139) directly pointed to autotrophy in this group Chemoli-thoautotrophywas first hinted at byVenter et al (105) whosemetagenomic analysis showed an apparent archaeal scaffoldthat contained genes suggestive of ammonia oxidation Fur-ther evidence came from Schleper et al (140) who detectedseveral ammonia oxidation genes in order directly adjacent toa Thaumarchaeota 16S rRNA gene in a soil-derived metage-nomic clone The issue was directly resolved when Konnekeet al (141) isolated a related marine archaeon from sedimentof a marine aquarium and this organism Candidatus Nitroso-pumilus maritimus was found to have a chemoautotrophicmetabolism quantitatively oxidize ammonia to nitrite andcontain an archaeal ammonia monooxygenase gene (amoA)and interestingly did not grow heterotrophically Cultivationexperiments showed this organism has a high affinity forammonium allowing it to outcompete bacterial nitrifiers atlow (submicromolar) concentrations as are typical in thesea (142) The complete genome of the organism has shownnovel adaptations for nitrification and autotrophy (143)
Even if the Thaumarchaea are primarily chemolitho-autotrophs field data suggest a level of mixotrophy in thatsome organic substrates are being incorporated into biomassA stable isotope study using cells collected at 670 m depthoff Hawaii estimated that about 80 of the carbon incor-porated into archaea-specific lipids came from inorganic sour-ces and about 20 from organic compounds (144 145)Interestingly it has also been claimed that the genomesof deep-sea (gt2000 m depth) members of the Thaumarch-aea as well as those living in equatorial waters rarely containthe amoA gene and thus may be primarily chemoorgano-trophs (48)
Major bacterial groups that have been documentedfrom seawater using 16S rRNA characterization includesome that are also known from culture (eg AlteromonasRoseobacter) and several that are phylogenetically distantfrom standard cultures The most common groups are inrough order of their relative abundance in clone librariesfrom most to least abundant SAR11 (relatives of Pelagibac-ter ubique) Roseobacter SAR86 cyanobacteria SAR116SAR202 SAR234 andMarine Group A The SAR designa-tion followed by a number is an arbitrary sequential cloneidentifier from Sargasso Sea cloning studies done by the Gio-vannoni lab that did most of the early systematic cataloging ofclones Summaries of the data and phylogenetic relationshipsof these groups can be found in Fuhrman andHagstrom (102)and Giovannoni et al (101)
Molecular Genetic Discoveries in Bacterial andArchaeal Marine BiologyAs described earlier metagenomics is an extension of theideas used in the 16S rRNA cloning studies in that all genesfrom the native microorganisms are separated and clonedwithout having cultivated the organisms These methodsearly on started to find unexpected and very interestingresults The best examples involve unexpected marine photo-trophy the first of which is discovery of a nonchlorophyllphotosynthetic bacterial pigment called proteorhodopsin(146) The gene was found on a large environmentallyderived fragment of DNA that also had a gene coding for16S rRNA from the so-called SAR86 group (one ofthe groups common in seawater) This pigment can act as alight-driven proton pump thought to permit cells to generate
422-10 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
FIGURE 4 Distribution of Bacteria Euryarchaea andmarine Crenarchaea (recently renamed Thaumarchaea) along a transect in theNorthAtlantic Ocean as measured by CARD FISH and expressed as of total bacteria + archaea counts via DAPI stained epifluorescence fromTeira et al (130) Top panel shows station locations that are shown on the top of the lower three panels with the distance in km reported alongthe transect from lower to higher station numbers doi1011289781555818821ch422f4
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-11
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ATP from sunlight Interestingly different versions of this pig-ment are found at different depths apparently ldquotuningrdquo theabsorption to match the ambient wavelengths of light asthey change with depth (147) The proteorhodopsin genesare very widespread and diverse perhaps in half or more ofall marine bacteria including SAR11 and Euryarchaeota(105 148ndash150) However almost all of several culturedorganisms with proteorhodopsin studied to date do not showa growth benefit from light (150) with the exception of amember of the Flavobacteria Dokdonia sp strain MED134that grows faster in the light only under moderately lownutrient conditions (151) and a marine Vibrio has beenshown to survive starvation longer in the light than in dark-ness (152) These observation may explain the wide phyloge-netic distribution and high abundance of this gene in thatproteorhodopsin may often assist long-term survival of bacte-ria under extreme energy-limiting conditions yet not havemucheffect on growthduringmore energy-replete conditionsInterestingly a recent report suggests the growth benefit fromlight in Dokdonia MED134 is from enhanced uptake of itsrequired growth factor vitamin B1 which is transported by atonB-dependent transporter powered by a proton gradient(153) This pigment might also have sensory or other rolesnot yet well examined (150)
The second kind of phototrophy found by moleculargenetic and fluorescence techniques to be unexpectedlyimportant in marine plankton is anoxygenic aerobic bacterialphototrophy based on the pigment bacteriochlorophyll aCulturable aerobic anoxygenic photosynthetic (AAP) bacte-ria have been known from seawater for several years (154) andare widespread (155) Initial recent reports of direct countsclaimed they represent about 11 of the total bacterial com-munity in the euphotic zone (156) but those early counts didnot correct for the presence of other bacteria and such correc-tionoften yields estimates averaging closer to 2(157)Directmeasurements show the bacteriochlorophyll pigment is rela-tively rare (158) However studies in various ocean locationsincluding ocean gyres report that these AAP bacteriacan indeed on occasionmake up a quarter of the total prokar-yotes (159 160) A recent energetic modeling of AAP andproteorhodopsin-containing bacteria (161) has suggestedthat proteorhodopsin-containingones cangain approximately02asmuchenergy from sunlight asSynechococcus (commonmarine cyanobacterium) and AAP bacteria can gain approxi-mately 13 as much energy from sunlight as SynechococcusThey concluded the AAP bacteria may gain energy sufficientto meet maintenance costs but proteorhodopsin-containingbacteria were not expected to do so except at high light inten-sities andwith largenumbersof proteorhodopsinmoleculespercell They concluded the ease and low cost of maintainingPR-based phototrophy (a few genes required) may explainthe high incidence of proteorhodopsin genes
Other metagenomic studies have yielded interestinginsights about organisms and processes that would otherwisebe difficult or impossible to establish For example thesequence of an archaeal gene fragment isolated from seawaterrevealed extensive evidence of genetic exchange with othertypes of archaea and even bacteria (162) Such geneticexchange was also quite evident from the complete genesequences of different strains of Prochlorococcus and Synecho-coccus which was attributed in part to virus-mediated geneflow (163ndash166)
Dilution Cultures of Bacteria and ArchaeaSome recent cultivation techniques that permit growth ofldquotypicalrdquo oligotrophic marine bacteria in pure culture are
based on the simple concept that bacteria living in seawateron dissolved organic matter might best be grown in the labo-ratory in ordinary filtered seawater This conclusion followsfrom similar techniques to grow mixed ldquoseawater culturesrdquoof marine bacteria (167) but the seawater inoculum in thiscase is diluted so that only one or a few bacteria are addedto the initial culture vessel (168) A rapid throughput versionof this method has been used to cultivate members of bacte-rial groups thought to be conventionally uncultivable such asthe SAR11 clade (169 170) albeit often at very low den-sities Such cultures permit focused studies of the propertiesof those organisms including genomic analysis (169 170)This valuable work provides insight into the roles and activ-ities of some of the most common bacterial phylotypesobserved in the ocean (171) showing for example that culti-vated SAR11 requires a source of reduced sulfur as well asproviding templates aiding the interpretation of metage-nomic analysis (172)
PROTOZOADiversity and DistributionHeterotrophic protists have been identified historically fromtheir morphological features that are apparent at the levelof the light or electronmicroscope Features of cell size shapetype and pattern of flagellationciliation skeletal structurescharacteristics of the nucleus and other cellular structuresand organelles have been used to differentiate themany thou-sands of described species A description of the extent of thisdiversity of form and function is well beyond the scope of thisbook (see [79] for more information) However much of thisdiversity can be reduced to three basic body plans that dictatethe broadest ecological roles of these cells amoeboid flagel-lated and ciliated forms (Fig 5) Among the lineages possess-ing one of these three forms only the last group the ciliatedprotists constitute a monophyletic group within the domainEukarya
One of the simplest body plans for protozoa is the amoe-boid cell exemplified by the gymnamoebae or ldquonakedrdquoamoebae Motile nonphotosynthetic cells lacking flagellaor cilia occur as life stages in a number of phylogeneticallydiverse taxa but for many free-living species of protozoathis form constitutes the only life stage Motility is largelyconfined to movement along surfaces by means of pseudopo-dia that can take on a variety of (species-specific) shapes Sig-nificant abundances of amoebae are largely confined tobenthic and epibiotic environments (174) and to suspendedparticulate material where they can occasionally be highlyenriched (27) Most amoebae consume bacteria and otherminute prokaryotes and eukaryotes
Several heterotrophic protistan groups possess complexamoeboid body plans most notably the foraminifera polycys-tine and phaeodarian radiolaria and the acantharia Plank-tonic forms are predominantly oceanic in their distributions(although a large number of benthic species of foraminiferaexist) These species are heterotrophic and many are visibleto the naked eye (individual cells can bege1 cm gelatinouscolonies can form ribbon-like structuresge1 m in length)They feed on a wide variety of bacterial protistan and meta-zoan prey using pseudopodial networks have rather longcomplex life cycles for individual cells (weeks to months)and are extensively used in studies of paleoclimatologicalreconstruction (82 175) In addition many of these speciespossess intracellular symbiotic usually eukaryotic algaeThe widespread occurrence of algal endosymbiosis among
422-12 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
these protozoa in oceanic pelagic environments implies thatstrong selective forces appear to give rise to these associations(97 98)
Flagellated forms of protozoa exist across many protistanlineages Eukaryotic flagella come in various sizes numbersand forms (eg with or without tiny hairs) that are character-istic of the different lineagesMany flagellate species are appa-rently purely phototrophic or heterotrophic (phagotrophic)but a significant (still poorly known) fraction of flagellates aremixotrophic combining both phototrophy and phagotrophy(70 88) Flagella in phagotrophic flagellates are employed formotility and prey capture Most free-living flagellates possessone to four flagella (typically one or two) that can be manytimes the length of the cell itself Although there is tremen-dous species diversity among flagellated protozoa many ofthese species have broadly overlapping ecologies As a groupflagellates are the most numerically abundant protozoa ofboth benthic and pelagic ecosystems and they are fundamen-tally important as consumers of bacteria cyanobacteriaand other eukaryotes Flagellated protozoa within the nano-plankton size class (2ndash20 microm) are often counted as a singleassemblage in plankton studies because morphological details
apparent using light microscope are insufficient to distinguishamong the many species This assemblage has been variouslyreferred to by a variety of names and acronyms including het-erotrophic nanoplankton (HNAN HN) heterotrophic flag-ellates (Hflags) heterotrophic nanoflagellates (HNF) andmicroflagellates Heterotrophic nanoplankton is the mostaccurate term for most methodologies employed to countthese cells because it does not require visualization of flagella(which are often lost from these small cells during preparationfor microscopy) The confusion over terminology is partly aconsequence of the different methodologies that have beenused to count these species and partly a consequence of thefact that these species were first studied prior to the wideacceptance of the size convention of Sieburth et al (84)
Heterotrophic flagellates in the microplanktonic size class(20ndash200 microm mostly dinoflagellates) are important consum-ers of phytoplankton in pelagic ecosystems Many of thesespecies are capable of the production of large pseudopodialnets the pallium that can engulf prey (particularly diatoms)significantly larger than the diameter of the dinoflagellatetheca (176) This behavior and its ecological significance forenergy flow in plankton communities have been recognized
FIGURE 5 Body plans and size ranges of protozoa These micrographs depict amoeboid (andashg) flagellated (hndashj) and ciliated (kndashm) forms ofprotozoa From Caron et al (173) Markers bars are 5 (i) 10 (h) 20 (c j l) 30 (a) 50 (k m) 100 (b f) 500 (d) and 1000 (e g) micromdoi1011289781555818821ch422f5
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-13
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
only within the past few decades (177) These species areoften abundant in waters where diatoms dominate (178179) In such situations heterotrophic dinoflagellate biomasscan be similar to that of ciliated protozoa (180)
Ciliates are generally the most recognizable form of pro-tozoa to nonspecialists Nonetheless ciliates are a diversegroup morphologically as well as ecologically (181 182)The degree of ciliature on these species can range from uni-formly ciliated to totally devoid of cilia during most of theirlife cycle to ciliature restricted to specific regions of thecell In addition cilia can fuse to form various complexstructures (eg ciliary membranes or veils cirri) that assistin locomotion food capture or attachmentCiliates are abun-dant and ecologically important species in both benthic (8)and pelagic (183) ecosystems and can consume a variety ofprokaryotic and eukaryotic prey Together with the heterotro-phic dinoflagellates these species are the dominant consum-ers of phytoplankton in many pelagic ecosystems (184) andas a consequence they form an important trophic link to met-azoan zooplankton (185 186)
Diversity and Biogeography of Protists FromMorphology to DNA SequencesUntil relatively recently the biodiversity of marine protistanassemblages was not generally considered a controversialtopic It was generally accepted that while all species of pro-tists certainly have not been identified representatives ofmost types of algae and protozoa had been observed anddescribed if not actually brought into culture in the labora-tory Direct sequencing of 18S rRNA genes from environ-mental samples (as described in ldquoMolecular Phylogeny andMetagenomics Field Applicationsrdquo) has changed that viewInitial forays into environmental DNA indicated a muchgreater diversity of protists than previously documented usingclassical approaches of culture and microscopy (81 187ndash191) Numerous publications over the past decades haveexpanded these findings (see [192] for a recent global analy-sis) which are highly analogous to discoveries in marine pro-karyote research as described already
Previously uncharacterized protistan diversity has nowbeen documented at virtually every level of eukaryoticorganismal classification Some of these findings couldhave been expected but some have been very unexpectedFor example a much greater diversity than noted previouslyhas been observed among small (lt10 microm) protists in plank-tonic ecosystems These species generally possess few distinc-tive morphological features and one could expect that manycryptic species might be present among these small morpho-types The molecular ldquodiscoveryrdquo of this eukaryotic diversityhas stimulated progress on the isolation and descriptionof new species and genera of minute algae and protozoa(193ndash197)
Analyses of environmental samples have also indicatedthe unanticipated existence of novel 18S rRNA gene sequen-ces that imply the presence of novel lineages of eukaryotes innatural protistan communities (198ndash200) These are sequen-ces for which there are apparently no known described orcultured species The degree to which these sequences dif-fer from sequences of known sequenced eukaryotes impliesthat some of these lineages may be distinct at the level ofphylum (201)
These findings have raised basic questions and somedebate concerning the true diversity and biogeography ofprotistan assemblages in natural ecosystems (202ndash204) thevalidity of the many novel phylotypes or cryptic species that
are being documented (205) the potential importance andsignificance of the many rare taxa that characterize thesecommunities (206) and the implications of these findingsfor the ecologicalbiogeochemical roles that protists playin aquatic ecosystems For example it was postulated andsubsequently confirmed that some novel alveolate lineagesreported from marine ecosystems make up a suite of parasiticprotozoa whose ecological importance may have been signifi-cantly underestimated in the past (207) Deciphering theidentity and significance of these many unknown phylotypeswill constitute a significant effort for protistologists in thefuture (80 85)
Life Histories and Ecological Strategies of ProtozoaAbundance prey type and life histories all vary tremendouslyamongmarine protozoa The smallest species (eg many flag-ellates) tend to be the most abundant and widely distributedin the world ocean Indeed many of these species may beglobally distributed (208) Most nanoplanktonic flagellateshave potentially rapid rates of grazing and growth Underoptimal conditions these species can divide by binary fissionseveral times a day and thus dramatic increases in their pop-ulations can take place in response to favorable conditionswithin a few days (209) Many of these species are capableof surviving for limited periods of time without food andhave developed a variety of physiological or life cycle strat-egies to cope with these events (209) However in contrastto some bacteria that may remain viable through long periodsof starvation (210) protozoa will expire or encyst in responseto low food abundance and thus ldquoboom-and-bustrdquo popula-tion changes are characteristic of small flagellates
On the other end of the size spectrum from the ratherubiquitous nanoflagellate species many of the largest proto-zoan species (polycystine and phaeodarian radiolaria plank-tonic foraminifera) are exclusively oceanic (ie do notsurvive in most coastal environments) andor have specificlatitudinal and depth distributions Thus the abundancesof these latter species may range from undetectable to max-imal abundances of gt105 individualsm3 Dramatic changesin the abundance of these protozoan taxa also can be relatedto changes in prey abundance physicalbehavioral aggrega-tion or to periodicity of life cycle events For example theplanktonic foraminifer Hastigerina pelagica reproduces on alunar cycle and thus abundances (and life stages) of this spe-cies in oceanic waters can vary considerably over the courseof a month (211) In general life cycles for the large amoe-boid protozoa are lengthy and complex (for single-celledorganisms) with life spans unknown for many species(attempts to culture them in the lab have so far been unsuc-cessful) but estimated to be on the order of months to per-haps years (175)
The abundance and activities of microplanktonic hetero-trophic protists (mostly ciliates and heterotrophic dinoflagel-lates) tend to be somewhat intermediate to those ofnanoplanktonic flagellates and the larger amoeboid formsThese species are present in the majority of marine ecosys-tems and collectively play an important role in the controlof phytoplankton biomass (and probably the abundance ofnanoplanktonic protozoa although there is little informa-tion on this topic) in waters throughout the world oceanLike small flagellates ciliates and dinoflagellates reproduceprimarily by binary fission but their maximal growth ratesare typically slower (one division a day is typical) and theirlife cycles often include sexual phases that allow for geneticrecombination
422-14 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
VIRUSESViral Abundance and General PropertiesViruses are simple biological agents typically 20ndash200 nm indiameter composed of a nucleic acid genome in a proteincoat that infect cells and ldquocommandeerrdquo the cellrsquos machineryto make more viruses which are released into the environ-ment when the host cell lyses or bursts A particular virus isthought to be capable of infecting only a narrow range of hosts(usually one species sometimes a genus rarely broader)Although some early studies had isolated viruses from thesea therewere no data prior to the 1980s showing such viruseswere very abundant and more significantly no evidence thatinfection was occurring in any important part of the planktoncommunity It wasnrsquot until the late 1980s that electronmicro-scopy with suitable concentration methods showed thatviruses are extremely abundant similar to or even higherthan bacterial abundance (212 213) Interestingly themost suitable transmission electron microscopy (TEM)approach used by Bergh et al (described in detail by [214])was actually similar to a direct centrifugation method devel-oped in 1949 (215)
TEM studies of viruses permit high-resolution images andobservation of viral morphology showing features such ashead diameter tails sheaths and tail fibers (Fig 6 collage
in upper left) Each type of virus has a fixed morphology(unlike bacteria that are potentially more plastic in theirappearance) and hence a coarse measure of viral diversityhas been possible by cataloging morphologies Studies thathave done so have found dozens or more different morpholo-gies of marine viruses in a given sample (217ndash219) A largeproportion resemble bacteriophages which are viruses thatinfect bacteria
The availability of brightly fluorescent nucleic acid stainsand high-porosity fine pore size (002 μm) filters made fromAl2O3 have made it easy to accurately count viruses by epi-fluorescence microscopy (38 220 221) Epifluorescence per-mits abundance estimates but does not allow observation ofviral morphology as viruses are below the resolution limitof light microscopy viruses are visible only as sources of light(like stars in the night sky see Fig 6 lower micrograph) Anextension of manual epifluorescence counts is the use of flowcytometry to count viruses now commonly employed in somelabs (222)
Virus abundance has been found to be closely related tobacterial abundance with a virusbacteria ratio typically10ndash301 A typical oceanic profile of bacterial and viral abun-dance is shown in (Fig 6 panel on right) This tight ratioand the strong correlation to bacterial abundance in relationto weaker correlations to chlorophyll have been cited as
FIGURE 6 Marine viruses (upper left collage) viewed by transmission electron microscopy These are cyanophages in the groups myoviridae(right and bottom) siphoviridae (left) and podoviridae (top) Scale bars represent 100 nm From Sullivan (216) Epifluorescence micrograph(lower left) of SYBR green I stained viruses (small fluorescing objects) and bacteria + archaea (large fluorescing objects) Depth distribution ofvirus and bacteria + archaea abundances obtained by epifluorescence microscopy of SYBRGreen stained cells in the central San Pedro BasinCalifornia (11 August 2000) doi1011289781555818821ch422f6
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-15
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
evidence that most marine viruses infect bacteria rather thaneukaryotic phytoplankton (217 218 223 224) though thereare also many important viruses of phytoplankton and otherorganisms with significant impacts on primary productivityblooms and ecosystem function (225ndash227)
The first demonstration of the activity of marine viruseswas presented by Proctor and Fuhrman (228) who showedby electron microscopic examination that viruses wereactually infecting marine bacteria and cyanobacteria at ameasurable rate Because only the final portion of the viruslife cycle is visible by electron microscopy (when the virusesare assembled and ready to lyse the host cell) only a very smallfraction of the infected cells can be counted as infected at anygiven time Proctor and Fuhrman (228) reported that only afew percent of the total bacteria from coastal waters and theSargasso Sea were visibly infected but they interpreted thedata with a model that implied the actual fraction of the totalcommunity that was infected was much higher They con-cluded that the total fraction of bacterial mortality attribut-able to viruses is roughly 10ndash40 Subsequent studies haveused refined versions of that same general approach andmodel (229 230) and numerous studies have used a varietyof alternative approaches to estimate viral activitymdashall yield-ing basically the same general conclusion (although themodel parameters need to be adjusted for cyanobacteriawhich on average seem to be infected less than bacteria)These approaches include (a) calculation of virus turnoverfrom decay estimates (231) (b) measurement of viral DNAsynthesis by incorporation of tritiated thymidine (232) (c)observation of effects of added viruses (233 234) (d) estima-tion of bacterial mortality in the absence of protists (e) use offluorescent viruses to estimate production by an approachanalogous to isotope dilution (38) and (f) dilution of virusesin filtered samples to estimate viral production (235) Theoverall consensus of these studies is that the initial estimateswere basically correct that is that viruses are responsiblefor about 10ndash40 of the bacterial mortality in most marinesystems (reviewed by [218 223 226 236 237]) Howeverthere is still some disagreement about the higher estimatesand it is likely that 40 mortality from viruses alone is nottypical for most marine systems
Most of the foregoing work has focused on the viral infec-tion of bacteria primarily thought to be heterotrophicalthough the original report by Proctor and Fuhrman (228)also noted the occurrence of cyanobacterial infection Sev-eral subsequent studies focused on phytoplankton includingthe potential effect of viruses on the termination of phyto-plankton blooms of Emiliania and Phaeocystis and the likelyimpact this might have on release of climate-active gasessuch as dimethyl sulfide (225 227 238ndash247) Althoughdetails are beyond the scope of this chapter viruses arethought to infect virtually all marine organisms with poten-tially significant impacts from zooplankton to whales (226)Detailed studies of viruses infecting cyanobacteria such asSynechococcus have shown differences in viral host specific-ity particularly toward coastal and oceanic host strainsand some occasional high virus abundances (to 105ml) asmeasured by most probable number (MPN) cultivation tech-niques in the Gulf of Mexico near Texas (248 249) Othervirus cultivation studies with Prochlorococcus and Synecho-coccus in oligotrophic waters of the Sargasso Sea showedan interesting pattern of cross-infection between these gen-era by some virus types (suggesting gene flow among theseorganisms) but generally low MPN estimates of abundanceto 103ml even when cyanobacterial abundance was near105ml (216 250)
Overall the consensus emerging from direct comparisonsof viral-mediated mortality and grazer-mediated (ie micro-zooplankton) mortality have indicated that viral lysis of bac-teria constitutes a significant fraction of total mortality ofthis assemblage while the mortality of phytoplanktonappears to be dominated bymicrozooplankton inmost instan-ces (251 252)
As mentioned viruses have significant morphologicaldiversity as observed in TEM studies It is also possible toinvestigate aspects of their genetic diversity In early workon this topic Wommack et al (253) and Steward et al(254) observed the diversity of viral genome lengths in a fieldsample by pulsed field gel electrophoresis with viral genomesranging 25 kb to gt300 kb in length Field results show thatthe viral community composition is dynamic in space andtime with clear changes in the banding patterns over seasonsand locations in Chesapeake Bay (253) between oceanbasins and subsequent to dinoflagellate blooms (254) andwith depths to 500 m at one location (255)
Although viruses do not all share a set of core genesthat allows a universal viral phylogeny (comparable to SSUrRNA gene in cellular organisms) genetic diversity amonga single group of viruses can be examined by sequence analysisof shared genes within the group An example is the g20 geneinT4-like cyanophages that has shown extremely high diver-sity even among very closely related viruses and has demon-strated geographic and seasonal variation (256) A secondexample is g23 which is found broadly in diverse and wide-spread T4-like phages (257) and can have seasonally repeat-ing patterns (258) as well as short-term rapid dynamics thatcorrelate to those in bacteria (259)
Viral MetagenomicsAs with cellular organisms metagenomics can be used toexamine viral diversity and genetics without themany restric-tions of cultivation Viruses can be collected by selective fil-tration and concentrated by tangential flow filtration orflocculated with iron chloride (260 261) then their collec-tive metagenome can be extracted linker amplified and ana-lyzed by sequencing (262 263) Viral metagenomic studiesare particularly challenging because the large majority ofsequences have no annotated matches in any databases butso far results from marine samples around the world haveshown extremely high diversity and variations with depthlocation and time presumably with highly dispersed typesselected by local conditions (262ndash266) Because metage-nomes are best interpreted when there are representative cul-tures available the best matches of viral metagenomesoriginally tended to be to the few viral isolates infecting trulycommonmarine bacteria like cyanophage infecting Synecho-coccus and Prochlorococcus (267) However the developmentof dilution-to-extinction cultures representing common het-erotrophic marine taxa like Peligibacter (a member of theSAR11 clade) and SAR116 has allowed isolation from sea-water of viruses infecting these common organisms and theseisolates have indeed been found to be highly abundant inmarine viral metagenomes (268 269) New approaches tointerpret the results include clustering the proteins independ-ent of known proteins to compare samples to each other andtry to find environmental factors driving viral communitychanges (270) Such analysis of a large global data set (TaraOceans expedition) has shown that extensive sampling hascome close to reaching the total diversity in tropical and tem-perate waters of such viral protein clusters (which essentiallyrepresent various viral protein motifs not all viral proteintypes) and that viruses appear to be directionally dispersed
422-16 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquodownstreamrdquo in major ocean currents as one may expectfrom first principles but also supporting the ldquoseed bankrdquohypothesis of viral biogeography (271)
Viruses and Host DiversityViruses are themselves thought to be instrumental in drivingincreased diversity of their microbial hosts via a hypothesizedprocess often called ldquokill the winnerrdquo This is because viralinfection is host-specific and density-dependent the latterbecause viruses diffuse from host to host so an abundanthost is more likely to pass on infection than a rare oneThis means that if an organism becomes abundant andblooms winning the competition for resources it becomesmore susceptible to a viral epidemic This would benefit therarer organisms and thus help foster diversity (223 272273) However bloom scenarios as described above involvesystems far from steady state and the formal theory of Thing-stad and Lignell (274) has interesting steady-state solutionswhere several viruses infect several hosts stably over time(via trade-offs between growth rates and viral susceptibility)which may occur at the strain or species level (274) There issome experimental evidence that viruses have effects on nat-ural marine microbial community composition weaker thanthe kill the winner (bloom version) hypothesis would sug-gest (275 276) so something resembling the steady-statecoexistence described in the model may in fact be commonThere also appear to be processes that foster coexistencebetween viruses and hosts but the mechanisms are largelyspeculative (223 277 278)
Viruses may also be directly involved in host genetic diver-sity because they can be the agents of genetic exchangebetween microorganisms (223 273) This often involvesthe viral lifestyle known as lysogeny whereby viruses survivewithin host cells as DNA only integrated into the host chro-mosome and being reproduced each time the host divides Ahost harboring such a genome is called a lysogen becauseunder conditions of stress to the host cell a genetic switchmay cause the viral genome to initiate the lytic process pro-ducing many progeny viruses and bursting from the hostLysogeny is a very common property occurring in a signifi-cant part of the bacterial community (279 280) althoughthe incidence of induction of the lytic phase in nature is appa-rently low (281) Overall lysogeny is poorly understood butthought to have both positive and negative impacts on themicrobial community (282) Recent results suggest that ben-efits of being lysogenic in highly seasonal polar seas leads tofundamental difference between polar and other marine viralcommunities (283)
Viruses and the Microbial LoopAs part of the food web viruses occupy a unique positionThey infect host cells that are mostly thought to be heterotro-phic bacteria and by doing so they typically burst the hosts torelease progeny viruses and cellular debris But what is the fateof this material Viruses themselves do not last indefinitelyand a simple steady-state assumption implies that from eachburst of viruses (typically 20ndash100 per lytic event) only onesuccessfully infects another cell The rest are inactivatedand broken down by sunlight (UV and visible exposure)and enzymatic attack (284) or consumed byminute phagotro-phic protists (285) thus reentering the food web as substratefor bacteria or food for protistan consumers Experiments incontrolled laboratory systems and field studies with radioac-tively labeled viral lysis products have supported the conclu-sion that most of the organic matter released by the viralinfection is either taken up by bacteria or respired (286 287)
Modeling this process as part of the microbial loop showsthat viral lysis represent a sort of side loop that has the neteffect of remineralizing a significant amount of the carbonand nutrients that enter the bacteria-protist part of themicro-bial loop (Fig 7) A theoretical numerical steady-state modelcomparing a systemwith no viral activity to onewhere virusesare responsible for 50 of bacterial mortality showed that thesystem with viruses had 33 more bacterial production andrespiration than the virus-free system implying that theviruses had the effect of permitting the bacteria to processmore of the primary production than they would otherwise(223) Although 50 is a high number unlikely to be com-mon in the sea this model nevertheless illustrates that virusescan reduce the amount of energy reaching higher trophic lev-els The implication is that viruses lead to increased bacterialactivity at the expense of the larger organisms
MAJOR ENVIRONMENTAL CONTROLSLight Temperature and PressureTemperature has an important potential influence on bio-chemical reactions and therefore on biological processesin general Most ocean waters fall in the range of ndash2degC to30degC with obvious exceptions in hydrothermally heatedareas Temperature has long been known to be a regulatingfactor for the growth of heterotrophic microbes
In temperate waters it has been established that microbialactivity is generally much higher in warm summer waters thanin winter (55) The relationship is not simple howeverbecause multiple factors act at the same time Some contro-versy still exists regarding the highest and lowest extremesfor marine bacterial growth although there is broad agree-ment that bacteria grow gt100degC at hydrothermal vents andltminus5degC in sea ice brines Pomeroy et al (57) noted the inter-esting observation that bacteria seem particularly inhibitednear the freezing point of seawater (ca ndash22degC) comparedto eukaryotic phytoplankton This effect results in polarspring phytoplankton blooms that accumulate organic car-bon in advance of the response of the bacterial communityand development of the microbial loop and perhaps lead toenhanced benthic-pelagic coupling (288)
The relationship between temperature and the growth rateof marine phytoplankton was described broadly in a now
FIGURE 7 Modification of the microbial loop concept that incor-porates the functional role of viruses Export can be via predation orsinking From Fuhrman (223)doi1011289781555818821ch422f7
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-17
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
classic paper by Eppley (58) and later Goldman and Carpen-ter (289) Temperature was shown to exert a strong and directeffect on themaximal growth rates of these species withmax-imal intrinsic growth rates at 0degC generally lt1 divisiondaywhile growth rates at 30degC may be gt4day These relation-ships indicate the maximal rate that might be attained byphytoplankton at these temperatures but they do not takeother factors into account (nutrients light) While warmtemperature ostensibly allows more rapid growth it createshydrographic conditions that typically give rise to nutrientlimitation of algal growth Thus some of the coldest watersin nature witness some of the most massive phytoplanktonblooms (290) while warm oceanic gyres represent some ofthe most oligotrophic areas of the ocean The multiple andoften conflicting effects of temperature on the growth of nat-ural phytoplankton assemblages limit the accuracy of presentpredictions regarding how primary producers will respond toclimate change (291)
An analysis of the effect of temperature on the growth ofheterotrophic protists has indicated that the response is qual-itatively similar to that of phytoplankton but with a twistThe growth rates of at least some protozoa can exceed thoseof phytoplankton at warmer environmental temperaturesbut the opposite effect is apparent at very low environmentaltemperature This differential effect of temperature on thegrowth of phototrophic and heterotrophic protists was basedon a large meta-analysis of published protistan growth (59)That analysis demonstrated that the maximal growth ratesattained by phototrophic protists could exceed the maximalgrowth rates attained by heterotrophic protists (all otherpotential growth-limiting factors not considered) Thereforeprotozoan growth rates may be constrained to a greater degreeat low environmental temperature than rates for phytoplank-ton If so then phytoplankton bloomsmay get a head start ongrazers during spring in polar ecosystems This scenario is con-sistent with information on seasonal biomass changes andmicrozooplankton herbivory in the Ross Sea Antarctica(292 293) but there are still too few data to fully vet thishypothesis
The importance of high pressure on bacterial growthgained considerable attention in the late 1960s when thedeep sea submersible Alvin was accidentally lost overboardwith its hatch openWhile no lives were lost in this accidentsome workmenrsquos lunches sank to the bottom (sim1500 m)inside the submersible Alvin was recovered after 10 monthsand interestingly there was a waterlogged lunch containingapples bologna sandwiches and broken vacuum bottleswith broth that all appeared hardly degraded and tasted palat-able Yet when placed in a refrigerator on the ship these itemsdegraded relatively quickly Initially pressure was thought tobe the preserving factor as the sea floor temperature was sim-ilar to the refrigerator temperature and the only major differ-ence would be pressure (294) Following this observationa series of experiments to measure degradation of variousorganic materials left in the deep sea for extended periodsindicated that degradationwas typically significantly reducedimplying that pressure reduces the degradation rates (295)Nonetheless changes in the protozoan community of naturaldetrital material sinking to the deep ocean floor indicate thatthe microbial community can respond relatively quickly insome situations (296) Deep sea microbiology has advancedconsiderably yet it is still difficult to interpret results withrespect to actual in situ rates of naturally occurring organicmatter Deep sea bacteria adapted to high pressures havebeen isolated that are barophilic (also called piezophilic)meaning that they prefer high pressures and have reduced
activity at lower pressures whereas others are barotolerant(piezotolerant) tolerating but not preferring high pressuresfor example see (297 298)
Few data are available on barotolerantbarophilic marineprotozoa Protozoa certainly exist and grow at great oceanicdepths but measurements of in situ growth rates for these spe-cies do not yet exist Measurable protozoan numbers havebeen documented in the deep-sea sediments for more than30 years (299 300) and viable protozoa have occasionallybeen cultured from these environments (301ndash305) butvery few direct measurements of the activities of these speciesin situ have been reported (306) A few protozoa have beenisolated that will grow at high pressure (302 303 307) andprotists that appear to be unique to the deep ocean havebeen observed either directly or through the analysis ofDNA sequences (308ndash311) but possibly the best direct evi-dence that protozoan activity takes place at the high pressurescharacteristics of the deep sea are experimental and observa-tional work noting the stimulatory effect that detrital deposi-tion has on some components of the protozoan community(296 312) These observations indicate a diverse and activeprotozoan fauna of the deep ocean although their biogeo-chemical significance is largely uncharacterized at this time
Dissolved and Particulate Organic MatterBacteria and archaea are thought to be by far the most impor-tant organisms with respect to the processing of dissolvedorganic matter (DOM) and nonliving particulate organicmatter (POM also called detritus) in the ocean While theremay be some uptake of DOM by protists particularly forgrowth factors needed in trace amounts such as vitamins(313) the bulk of this material is probably utilized by bacteriaand archaea (314) including the smallest cyanobacteriumProchlorococcus (315 316) which thus may be considered amixotroph Due to their small size bacteria have extremelyhigh surfacevolume ratios and combined with their over-whelmingly high abundance an extremely high integratedsurface area Protozoa tend to obtain the organic materialsthat they require for growth from their prey rather thanthrough the uptake of DOM Overall protozoa tend to besources of dissolved and detrital organic substances throughthe excretion of unassimilated prey biomass in expelledfood vacuoles
Particulate organic matter is not directly available as sub-strate to bacteria These substances must first be reduced tosmall molecules that can be transported into the cell Thisis accomplished by the production of extracellular enzymes(note that few if any large polymers are directly taken up bybacteria with the possible exception of DNA) Hydrolyticenzymes produced by bacteria (and almost certainly archaea)break down polymers like proteins polysaccharides andnucleic acids As with DOM POM is composed of a complexmixture of compounds that vary in their susceptibility to bac-terial degradation and utilization
Particulate material in the water column serves not only asbacterial substrate but also as substratum POM occurs in thewater column across a huge size spectrum from micrometersup to some detrital aggregates more than 1 m in diameter(317) Much of this particulate material is in a constant stateof flux with colloidal material constantly coalescing andaggregating to form new or larger particles (318) as microbialdegradation acts simultaneously to remineralize this materialDetrital particles that attain macroscopic size either by directformation (317 319 320) or via accretion and aggregation(321) and are often called marine snow or macroaggregatesMarine snow particles are readily colonized by bacteria and
422-18 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
many other microorganisms and their abundances on theseaggregates can be orders of magnitude greater than abundan-ces in an equivalent volume of the water surrounding theaggregates (320 322ndash324) and their diversity and highlyconcentrated abundances on detrital aggregates implies thatthere may be stronger trophic coupling and efficient energyelemental cycling relative to these processes among free-living microorganisms in the surrounding waters This specu-lation has resulted in a number of formalizations of the poten-tial impact of aggregate microbiology on biogeochemicalprocesses in the ocean (325 326) Most definitions of bio-films include aggregated particulate and these particles areoften studied within the biofilm conceptual model (see chap-ter by Lawrence et al in this section)
Inorganic Nutrients
Macronutrients (N P)Virtually all heterotrophic organisms in the ocean contributeto the pool of available macronutrients via the excretion ofmetabolic wastes and protozoa contribute significantly tothis process Macronutrients are often in excess within proto-zoan prey relative to the consumersrsquo needs because much ofthe prey carbon is respired to produce energy for metabolismand growth ldquoExcessrdquo nutrients are eliminated as eitherorganic compounds or more commonly as ammonium (forN) and phosphate (for P) The importance of the match(or mismatch) between predator and prey stoichiometry hasbeen demonstrated experimentally and repeatedly in marineand freshwater ecosystems and has been elevated to a funda-mental ecological principle governing elemental and energyflow in aquatic food webs (327) Based on this reasoningCaron et al (328) concluded that bacterivorous protozoaplay an important role in nutrient remineralization andrelease in the ocean Bacterial biomass is typically rich in Nand P relative to other organisms so bacterial predators expe-rience large excesses of these elements relative to their growthneedsMoreover bacterivorous protozoa should be dispropor-tionately important in nutrient remineralization amongsingle-celled eukaryotes because as the smallest protozoa inaquatic ecosystems they have high weight-specific metabolicrates (329) Thus both stoichiometric and allometric rela-tionships implicate small bacterivorous protozoa as impor-tant sources of remineralized nutrients in the plankton
The reasoning also holds true for bacterial utilization oforganic compounds and it has led to some interestingfindings and conclusions regarding the role of bacteria innutrient cycling in the ocean Bacteria are important sourcesremineralized nutrients when they are consuming N- orP-rich substrate Contrary to this traditional view howeverheterotrophic bacteria are often strong competitors for inor-ganic nutrients under many situations largely because bacte-rial biomass is richer in N and P than much of the organicmatter they consume in nature so they require additional Nand P to produce their biomass (summarized by [330]) Exper-imental studies have demonstrated the uptake of ammoniumby bacteria other than cyanobacteria using stable isotope 15N(331) and also the short-lived radioisotope 13N (332) oftenfinding that bacteria may be responsible for one third of the Nor P uptake Additionally there is evidence that bacteria insome oceans like the Sargasso Sea are limited by phosphorus(333 334) Growth limitation of bacteria by phosphorusappears to be a common phenomenon of freshwater ecosys-tems (335 336)
N2 fixation is restricted to prokaryotes and in the oceanwater column it was thought for many years to be done
primarily byTrichodesmium a warm-water colonial cyanobac-terium that blooms sporadically andRichelia a cyanobacterialsymbiont that lives within certain diatoms (337) Some uni-cellular cyanobacteria also contribute significantly to globalN2 fixation (338) and molecular biological data suggestthat a variety of other bacteriamdashnot phytoplanktonmdashmayalso be fixing nitrogen in seawater (339) An exciting recentdiscovery in this field is that a bacterial nitrogen fixer origi-nally known only from nifH sequences UCYN-A has hadits genome sequenced by parallel pyrosequencing and wasfound be a cyanobacterium (Candidatus Atelocyanobacteriumthalassa) lacking the apparatus to generate oxygen as well asother pathways (340) Further study showed it to be symbioticwith a marine alga (341)
Micronutrients (Trace Metals Growth Factors)Bacterial growth may often be limited by macronutrients asdescribed and sometimes by trace nutrients particularlyiron that occur at very low concentrations in seawater(342 343) The importance of Fe as an element limiting pri-mary production in some oceanic regions has come underclose scrutiny and extensive experimental investigation inrecent years Many bacteria have special uptake mechanismsto utilize extremely low concentrations of iron includingsiderophores (released compounds that bind iron and arethen specifically taken up) Even if bacteria are capable ofgrowth at low Fe concentrations growth may be less efficientKirchman et al (344) found that Fe limitation stronglyreduced bacterial growth efficiency in a cultured marine bac-terium (ca 50 efficiency Fe replete but lt10 when Fedeplete) Thus the effects may be complex Marine N2 fixa-tion also may be limited by the availability of Fe becausenitrogen fixers have high Fe requirements
Protozoa tend to be a source rather than a sink for Fe in theocean analogous to their role in macronutrient remineraliza-tion Protozoan grazing activity has been shown to releaseiron from bacterial biomass thereby relieving Fe limitationin co-occurring phytoplankton (345) a mechanistic demon-stration of the process using protozoa growing on Fe-repletebacteria However it has also been reported that Fe-limitedbacteria may contain insufficient Fe for the bacterivorous pro-tozoa that consume them thus leading to Fe limitation in theprotozoan as well (346)
OxygenHypoxic and anoxic regions of the ocean are expected to sig-nificantly expand in the coming decades as a consequence ofchanges in ocean stratification resulting from global climatechange (347) As oxygen becomes depleted bacteria havethe capability to utilize a series of alternate electron acceptorsCompositional changes in the bacterial assemblage accom-pany these changes in metabolic activity but are beyondthe scope of this chapter Many of the studies on benthicmicrobiology have focused on redox reactions that occurthere Oxygen is usually consumed by sediment microbialactivity much faster than the rate at which it can be replacedby diffusion so most sediments especially organically richones are anaerobic below a few mm (fine-grained muds) orcm (coarser sediments) from the sedimentwater interfaceexcept where animals ventilate the benthos through tubesWhen oxygen is absent bacteria and archaea use alternateelectron acceptors such as nitrate nitrite oxidized Fe orMn organic matter or sulfate The use of nitrate or nitriteas an electron acceptor with concomitant production of N2
gas (denitrification) results in net loss of biologically availablenitrogen Bacteria that perform this reaction are typically
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-19
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquofacultativerdquo organisms that can switch from oxygen to nitrateor nitrite Organic matter may be reduced by fermentationreactions in which part of an organic substrate is oxidizedwhile another is reduced fermenter organisms are also typi-cally facultative Sulfate is a particularly important alternateelectron acceptor in marine systems because it occurs as thesecond most abundant anion in seawater (behind chloride)typically 28 mM Its use as an electron acceptor leads to theproduction of elemental sulfur or sulfide The use of sulfateas an electron acceptor yields much less energy than oxygenor nitrate and sulfate-reducing bacteria are strict anaerobesunable to use oxygen (often strongly inhibited by oxygen)Methane can be produced by methanogenic archaea whichare all strict anaerobes using CO2 as an electron acceptor(see Findlay and Battin this section for further discussion)
Reduced compounds such as sulfide are produced by theuse of these alternate electron acceptors and they diffuse awayfrom their source When they diffuse to environments with astrong oxidant like oxygen they may be oxidized by chemo-trophic bacteria in energy-yielding reactions Similarlyammonium can be oxidized to nitrite and then nitrate by che-motrophic nitrifying bacteria and the nitrate can diffuse intoanaerobic layers and become denitrified to N2 Such coupledoxidation-reduction reactions can lead to accelerated biogeo-chemical cycling particularly in sediments
It was originally believed that relatively few free-livingspecies of protozoa inhabit anaerobic and hypoxic marineenvironments This view has changed based on (largely)molecular analyses of some of these environments (308348ndash351) A significant number of previously undescribedeukaryotic phylotypes have been observed in anoxic environ-ments High protozoan biodiversity in the absence of molec-ular oxygen is not completely unexpected because otheranaerobic environments such as some gut environments(eg cockroach guts and cattle rumen) have notoriouslyabundant and diverse protozoan fauna but marine anaerobicenvironments have been difficult to access and sample prop-erly Many anaerobic protozoa exist together with symbioticbacteria (352 353) or possess unique organelles such as thehydrogenosome (354)
MAJOR BIOTIC CONTROLSThe Trophic Activities of Protozoa Predation andTop-Down ControlsPredator-prey relationships are fundamental interactionswithin biological communities linking production of bio-mass on the one hand with removal on the other hand Estab-lishing the trophic relationships among microorganisms andquantifying the rates at which they take place constitutemajor endeavors within marine microbial ecology As notedpreviously protozoa consume a wide variety of prokaryoticand protistan prey Numerous laboratory and field measure-ments have been made throughout the past quarter centuryto attempt to quantify the rates of bacterial and phytoplank-ton consumption The methodology for accomplishing thesestudies has varied and a number of approaches are presentlyemployed
Methodologies for estimating microbial grazing are diversebut generally fall into one of two experimental approachesperturbation experiments or tracer experiments Perturbationexperiments (size fractionation metabolic inhibitors anddilution technique) rely on manipulation of the consumeror prey assemblages in somemanner such that the effect is evi-dent from changes in the abundance of the prey population
during incubations relative to control treatments that receiveno manipulation Tracer experiments (radioactively labeledprey fluorescently labeled prey stable isotopendashlabeled pray)attempt to use labeled prey or a prey proxy to quantitativelyfollow the movement of prey biomass into consumers Forsummaries of these methods see (74 355)
Perturbation experiments have entailed a variety of mech-anisms to ldquodecouplerdquo predators from prey in natural microbialcommunities For example size fractionation uses various sizefilters to remove an entire size class of grazers from awater sam-ple leaving the prey to grow in the relative absence of thosepredators during subsequent incubations Unfiltered samples(controls) indicate the growth of prey in the presence of pre-dation Predation impact is obtained by the difference ingrowth of the prey between these treatments This methodhas been used to examine predation on cyanobacteria heter-otrophic bacteria and minute phototrophic eukaryotesAlternatively metabolic inhibitors have been employed tohalt predation (or in some cases prokaryote growth) insteadof physical removal by filtration
The dilution technique (356) has become a commonmethod for examining grazing mortality of phytoplanktonby microzooplankton (which is comprised predominantly ofprotozoa) This method relies on the dilution of herbivorouszooplankton in a series of subsamples (and concomitantmeasurements of phytoplankton growth in each subsample)to estimate the grazing mortality of the herbivores
Tracer experiments have used both radioactively labeledprey and fluorescently labeled prey to examine predation onbacteria and phytoplankton Radioactive labeling has beenaccomplished using 14C- or 3H-labeled organic compounds(for bacteria) or 14C-bicarbonate (for phytoplankton) Theuse of radioisotope approaches has diminished with improve-ments in the number and types of fluorescent compoundsavailable for this work Fluorescently labeled prey havebeen labeled with a variety of compounds that allow theprey to be observed readily (even inside predators) by epi-fluorescencemicroscopy (357 358) Alternatively a decreasein the total number of fluorescent prey in a sample indicatesthe rate of removal of the prey assemblage
Both perturbation experiments and tracer experimentsdescribed have inherent advantages and disadvantages Forexample perturbation experiments are fairly straightforwardto conduct but they may introduce artifact by removingpotentially important sources of nutrients or organic com-pounds released by predators (359) On the other hand thedilution technique is very labor-intensive but one advantageof this method is that it provides the opportunity to determineboth phytoplankton growth and grazing mortality in a singleprocedure Most tracer experiments are fairly easy to conductand generally do not perturb the food web being studied butfeeding selectivity for or against the tracer (relative to naturalprey) may produce spurious results In addition some of thesemethods provide a measurement of the activity of the entiregrazer assemblage and others are applicable for examiningthe activity of specific taxa For example size fractionationapproaches examine the effect of the removal of an entiresize class of organisms (a community-levelrdquo measurement)whereas fluorescently labeled prey can be used to examinethe ingestion rate of a particular species by observing theuptake of labeled prey into that species (a species-levelmeasurement)
Predation rates of numerous marine protozoa have beenexamined in the laboratory From these experiments a gen-eral knowledge of the rates of consumption by these specieshas been accumulated Feeding in most protozoa shows a
422-20 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
functional response to the availability of prey that is similar tothe response of many organisms (although there are a greatdeal of species-specific differences) That is ingestion rateincreases rapidly as prey abundance increases until a maximalingestion rate is attained (360)Manyof these curves have theform of a Type II functional response curve (361)
Based on the features of these curves it is clear that proto-zoa have the ability to ingest substantial numbers of prokar-yote and eukaryote prey in natural ecosystems Somebacterivorous flagellates can consume bacteria (and cyano-bacteria) at rates up to 100 s per individual per hour whereassome bacterivorous ciliates can consume thousands of bacte-ria per individual per hour Rates of consumption of eukary-otic phytoplankton by herbivorous ciliates and largeflagellates are considerably more variable and highly depend-ent of the feeding mechanism (eg filtration in ciliates versuspseudopodial net in dinoflagellates) and the size of the prey
These lab studies demonstrate that rarely are prey suffi-ciently abundant in nature to support the maximal ingestionrates of which many protozoa are capable Nevertheless pre-dation rates of protozoa in nature are sufficient to constitute amajor factor controlling the standing stocks of bacteria andphytoplankton (63 180 184 355) For example these sum-maries indicate that on average protozoa consume 20ndash80ofthe standing stock of phytoplankton (measured as chloro-phyll) in the world ocean and 60ndash75 of daily primary pro-duction a departure from the classical view of copepods as theprimary consumers of phytoplankton This large role of her-bivorous protozoa infers a large role for copepods as consumersof planktonic protozoa (185) Similarly the role of protozoaas major consumers of bacterial biomass is indicated by therelative balance between bacterial production on the onehand and bacterivory on the other (Fig 8) The approximateone-to-one balance between these measurements over a largenumber of studies implies that protozoan bacterivory must bea major factor determining the fate of bacteria in marine (andfreshwater) plankton ecosystems
It is interesting to note that some types of bacteria includ-ing marine Bdellovibrio are known to prey on other marine
bacteria (362) At the time of this writing the importanceof bacterial predation by other bacteria within the contextof oceanographic processes is not known and presumed tobe small While Bdellovibrio may be cultivated from seawaterit can also be found among sequences from cold sediments(363) and the water column Bdellovibrio happens to be thebest studied predatory bacterium but there are no doubtothers that we simply have not yet identified (364)
Species-Specific Interactions (MutualismParasitism Commensalisms)Interactions between marine microorganisms such as compe-tition allelopathy and symbiosis are fundamental aspectscontrolling community composition and activity but atpresent most are poorly characterized in natural marine sys-tems Bacteria must compete for dissolved and particulatecompounds with other bacteria andwith protistan taxa whilephagotrophic protists must compete with each other andmetazoa for available prey These interactions manifest them-selves in many forms including true competition for availableresources resource partitioning such that some taxa specializeon the utilization of certain substrates or prey while othersspecialize on different ones (reducing competition) or eventhe cooperative use of compounds (eg one bacteriumrsquoswaste products being utilized as substrate by another bacte-rium) These metabolic consortia are probably common inthe ocean but can be difficult to identify and characterizeConsortia of bacteria working together to perform complexreactions especially in anaerobic systems are well known inmicrobiology
Consortia in which there is a very high degree of integra-tion between species (ie symbiosis) are very common in theocean These associations may be beneficial to both partners(mutualism) beneficial to one part and inconsequential forthe other (commensalism) or beneficial to one part and det-rimental to the other (parasitism) Many highly visible andwell-characterized marine mutualisms exist between chemo-synthetic bacteria and animals such as hydrothermal ventor methane seep tubeworms clams mussels or other inverte-brates (365) and also between luminescent bacteria and fishor squids (366) Similarly many protozoa form mutualisticassociations with photosynthetic protists (367) The mostconspicuous of these associations involved large amoeboidprotozoa of oceanic ecosystems (planktonic foraminiferanspolycystine radiolaria and acantharia) which contain largenumbers of intracellular algae (74) but associations betweensome heterotrophic dinoflagellates and cyanobacteria are alsocommon (368) Studies have indicated that efficient nutrientcycling in these protozoan-algal associations confers an eco-logical advantage for both partners in surfacewaters of the oli-gotrophic ocean
Parasitism is also a common symbiotic relationship amongmarine protozoa Protozoan parasites have been reported for adiverse array of protists and animals in the ocean Heterotro-phic dinoflagellates and presently uncultured alveolate cladesrelated to alveolates appear to be particularly successful atevolving thismode of existence (369ndash371) In a common par-asitic group Amoebophrya spp the parasite invades photo-synthetic dinoflagellates where it undergoes reproductioneventually lysing the host and releasing dozens of infectiveswarmers The parasitic protist Pirsonia spp is a parasite ofdiatoms (372) The overall importance of these parasitismsin the population dynamics of their hosts is not clearly knownbut may constitute a significant source of mortality for somespecies (373 374) The frequency and diversity of ldquonewrdquo
FIGURE 8 Meta-analysis of experimental studies that have com-pared rates of bacterial productivity (converted to population growthrate units of dminus1) with rates of bacterivory by phagotrophic protists(expressed as equivalent units of bacterial growth rate dminus1) Thesolid line indicates where measurements of bacterial removal by graz-ers were equal tomeasurements of bacterial production Data redrawnfrom Sanders et al (63) doi1011289781555818821ch422f8
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-21
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
Symbiosis the intimate living together of two kinds oforganisms especially if such an association is of mutualadvantagemdashtoo vague to be of use in quantitativedescriptions of population interactions but very usefulin indicating a close association among organisms
Syntrophy a metabolic mutualism where one species usesthe waste product produced by the other and in so doingallows both metabolic pathways to be energeticallyfeasible
Zooplankton Planktonic eukaryotes that consume otherplankton Includes single-celled organisms (protozoaor protists) and metazoans and some that are planktoniconly as larvae
Estimating ldquoBacterialrdquo Biomass and ldquoBacterialProductionrdquo Definitions and MethodsAquatic microbiologists tend to use the term ldquobacteriardquowith alowercase ldquobrdquo to describe organisms that appear to be prokary-otic by microscopymdashthat is organisms with no membrane-bound nucleus They include members of the taxonomicdomains Bacteria and Archaea (see ldquoBacteria and Archaeardquo)Organisms within and between these domains differ in manybiochemical and genetic aspects but they tend to look similarby traditional epifluorescence microscopy Special methodssuch as different versions of fluorescence in situ hybridization(FISH) are required to distinguish individual members ofthese domains microscopically (35ndash37)
The term ldquobacterial productionrdquo here refers to heterotro-phic production of biomass by bacteria It is meant to include
production of nonphotosynthetic bacterial biomass based onthe heterotrophic consumption of preformed organic matter(ie organic matter in various forms that has been producedprimarily by phytoplankton)
Bacterial biomass is usually determined by convertingdirect counts of bacteria using an estimate of the amount ofcarbon per cell Direct counts are most commonly done byepifluorescence microscopy with stains such as acridineorange 406-diamidino-2-phenylindole or SYBR green I(17 38 39) Special procedures are usually applied for sedi-ment samples and samples containing large numbers of bacte-ria attached to particles (40) SYBR green I also permits directvisualization and counts of viruses in the same preparationIncreasingly direct bacterial counts in seawater sampleshave been performed by flow cytometry of fluorochrome-stained cells (41 42) a method that allows separate countsof cyanobacteria such as Synechococcus and Prochlorococcuswhich have unique fluorescent signatures due to their photo-synthetic pigments and which can sometimes make up a sub-stantial fraction of the total number of bacteria (43) Flowcytometry is rapid and has a statistical advantage in that it typ-ically observes thousands of prokaryotic and minute photo-synthetic eukaryotic cells rather than the hundreds countedmicroscopically drawbacks include the cost of the instrumentand the fact that cells attached to each other or to other par-ticles are counted as one Bacterial carbon per cell has beenestimated in a variety of ways most commonly from a deter-mination of cell volume and carbon density per unit volumeThese numbers are difficult to obtain accurately for nativemarine bacteria which are very small typically 05 μm in
FIGURE1 An early vision of the ldquomicrobial looprdquo and its connections to the classical grazing food chain via dissolved organicmatter (DOM)flux and particulate trophic transfer with viruses included as a side loop Modified from (34) Large gray arrows indicate the flow of organiccarbon and energy into higher trophic levels of the food web with recognition of the important roles for heterotrophic microbes (bacteriaand protozoa) in this process Large stippled arrows indicate the production of DOM via excretion and trophic interactions (not all groupsare represented) Thin dotted arrows indicate mineralization of major nutrients contained in organic matter respired by consumers Whitearrows indicate bacteria lysis by viruses and DOM released by that process doi1011289781555818821ch422f1
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-3
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
diameter (range is about 02ndash1 μm for free-living unicells)Published estimates of bacterial carbon per cell vary widelyand probably constitute the greatest uncertainty with estimat-ing bacterial biomass in natural samples Typical estimates ofthe carbon content of a bacterium range from 7 to 50 fg C(1 fg is 10minus15 g) with most open ocean estimates near 10ndash20 fg C per cell and coastal ones about double that (44)Thus in a typical mesotrophic ocean environment with109 bacteria per liter and an average per cell C contentof 15 fg bacterial biomass would be approximately 109 times15 times 10minus14 = 15 times 10minus5 g C per liter or 15 μg C per liter
Bacterial production is most often measured by incorpora-tion of tritiated thymidine into DNA (24 45) or tritiated leu-cine into protein (26 46) Thymidine and leucine areintracellular precursors of DNA and protein respectively soincorporation of these precursors can be used to estimatethe total rates of synthesis of themacromolecules DNA is syn-thesized for cell division and protein synthesized roughly inproportion to total biomass someasuring their rates of synthe-sis is presumed to track production Both methods have beencalibrated on the basis of theoretical considerations as well aspurely empirical approaches and both methods yield similarresults (44) Leucine has a lower detection limit so it is pre-ferred for slower rates The results of thymidine incorporationare most often presented as cells produced per liter per hourwhich can be converted to a carbon production rate via esti-mates of carbon per cell as noted Leucine incorporation maybe used to calculate cell production as well as biomass produc-tion directly because protein is a major biomass constituent
Geographic and Temporal Distributions ofMicrobial Biomass and ActivityBacterial abundances by epifluorescence microscopy showthat bacteria are present in most marine euphotic zone
environments at approximately 106 cells per ml Samplesfrom around the world rarely vary by more than threefoldfrom this typical value (ie rarely lt3 times 105 or gt3 times 106)which is extraordinary compared to phytoplankton and zoo-plankton whichmay vary by several orders of magnitude overthe same spatial scales However despite this remarkable gen-eral predictability there is significant variation across bothspace and time More nutrient-rich eutrophic environmentstend to have more bacteria (sometimes gt107 per ml 47) andoligotrophic open ocean environments have less (summar-ized below) Although bacterial assemblages in warm temper-ate coastal waters may have doubling times as short as 1 daythis is at the rapid end of the spectrum of in situ growth ratesBacterial assemblages in the open sea especially in oligotro-phic environments have average generation times typicallyof a week or perhaps more (see Table 1) These abundancesgenerally apply to the euphotic zone and bacteria in colderdarker waters have substantially lower abundances and slowergrowth rates than those of surface waters (48) Benthic bacte-ria also exhibit fairly constant abundances across wide geo-graphic ranges but benthic bacteria occur at much higherdensities because of the organic-rich and particle-ladennature of the environment (an average of asymp109 per ml fluidvolume is typical 49) This value is three orders of magnitudegreater than abundances in the water column implying thatthe abundances of benthic bacterial assemblages are regulatedby a different set of parameters than planktonic assemblages
Bacterial biomass and productivity vary temporally on anumber of scales ranging from diel (50) to seasonal (51) orinterannual (52) On time scales of hours bacterial abun-dance and production have been shown to often peak inthe middle of the day and be low in the middle of the night(50 53ndash55) This pattern has been interpreted as a tight cou-pling between the production of labile organic compoundsvia photosynthesis and bacterial growth on one hand and
TABLE 1 Bacterioplankton properties in relation to phytoplankton in the open sea as compiled by Ducklow (44)
Property N Atlantica Eq Pac-Sprb Eq Pac-Fallc Sub N Pacd Arabiane Hawaiif Bermudag Ross Seah
Euphotic zone m 50 120 120 80 74 175 140 45
Biomass (mg C mminus2)
Bacteria 1000 1200 1467 1142 1448 1500 1317 217
Phytoplankton 4500 1700 1940 1274 1248 447 573 11450
BP 02 07 075 09 12 36 27 002
Production (mg C mminus2 dminus1)
Bacteria 275 285 176 56 257 Nd 70 55
Phytoplankton 1083 1083 1548 629 1165 486i 465 1248
BP 025 026 011 009 022 Nd 018 004
Growth rates (dminus1)
Bacteria 03 013 012 005 018 Nd 005 025
Phytoplankton 03 064 08 05 093 11 081 011
BP 1 02 015 01 019 Nd 006 23
Notes All bacterial biomass estimates based on 20 fg C per cell Data may overestimate heterotrophic bacterial biomass as a consequence of lower C per cell orinterference by Prochlorococcus and Archaea Production estimated from 3000 g C per mole leucine incorporated
aEastern North Atlantic spring phytoplankton bloom 47 N 20 W May 1989 n = 13bEquatorial Pacific 0 N 140 W MarchndashApril 1992 n = 8cEquatorial Pacific 0 N 140 W SeptemberndashOctober 1992 n = 19dSubarctic North Pacific 45 NeNorthwest Arabian Sea 10ndash20 N 165 E JanuaryndashDecember 1995 n = 21fHawaii Ocean Time Series (HOT) 1995ndash1997 n = 21 (httphahanasoesthawaiieduhothot_jgofshtml)gBermuda Atlantic Time Series (BATS) 1991ndash1998 n = 106 paired comparisons The ratios are means of the ratios not ratios of the means BP calculated
from thymidine incorporation (16 times 1018 cells per mole incorporated)hRoss Sea Antarctica 76 S 180 W 1994ndash1997i1989ndash1996 n = 64
422-4 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
bacterial mortality via grazers or viral lysis on the other handIt is consistent with some measurements of extremely rapidturnover sometimes several times per hour in rich coastalwaters of labile dissolved organic compounds such as dis-solved free amino acids (56) and might also indicate greaterpredation pressure by protozoa during the night On longertime scales of weeks to months bacteria show distinct sea-sonal patterns For example in temperate coastal waters bac-terial biomass and production increase considerably insummer months compared with winter However bacteriado not typically show a significant increase during earlyldquospringrdquo blooms in temperate waters when water is still verycold (51) It has been hypothesized that this phenomenonis the result of the suppression of the rate of substrate uptakeby temperate bacterial assemblages at low temperature (57)However while temperature probably has the effect of settinga limit on maximal growth rates (as for phytoplankton [58]and protozoa [59]) temperature alone does not appear to bethe main factor controlling growth of marine bacteria undermost circumstances (see ldquoLight Temperature and Pressurerdquo)
It would be overly simplistic to think that all the bacteriaand archaea in a sample or habitat have the same level ofactivity per cell but it is also easy to think of measured activ-ities as characteristic of all members of a microbial assem-blage So the question arises are most of the cells active ata similar level or are some hyperactive while others are com-pletely dead or moribund This question has been addressedseveral ways includingmicroautoradiography selective stain-ing ldquodirect viable countsrdquo (where nutrients are added to seewhat part of the community grows) and in situ hybridization(eg 60) Based on these contrasting approaches it appearsthat a continuum of activity exists within bacterial assemb-lages from truly dead (cannot be revived) to extremely activeA reasonable interpretation of the existing data is that undertypical conditions a small percentage of the marine bacterialcells perhaps 10ndash20 are generally inactive or dead the plu-rality or majority of cells perhaps 25ndash75 are intact andhave some moderate level of activity and a small percentageperhaps 5ndash20 are highly active It is useful to consider thisspectrum conceptually and numerically when modelingmicrobial processes
Comparisons of bacterial and phytoplankton biomasswithin planktonic ecosystems show that these are positivelycorrelated across broad scales Analyses of marine and fresh-water samples from several studies (61 62) have shown thatbacterial abundance increases with chlorophyll concentra-tion at least at the level of a log-log relationship Similarlybacterial abundances and the abundances of small protozoacorrelate over broad spatial and temporal scales (63) Theserelationships are sensible in that on the largest scale primaryproduction is the source of organic material that fuels hetero-trophic bacterial activity and bacteria constitute the prey ofmany small protozoa Individual data sets also have sometimesshown strong correlations between bacterial abundance andchlorophyll (eg 45) but variability in this relationshipover short temporal or spatial scales is to be expected It wouldpresumably be a consequence of rapid short-term changes inthe rate of substrate supply as well as the normal oscillatorynature of predator-prey relationships between bacteria andtheir consumers
Interestingly the extrapolation of the positive log-log rela-tionship between bacterial and phytoplankton biomassto environments with very low chlorophyll concentrations(eg ultra-oligotrophic oceans) indicates that bacterial bio-mass may exceed phytoplankton biomass in these situationsWhile this conclusion is obvious for the deep sea where there
is no photosynthesis this situation is also often true in oligo-trophic surface waters Measurements made in oligotrophicwaters bear out the high bacterial contribution to totalbiomass (64 65) Moreover Cho and Azam (66) confirmeda linear relationship between the log of chlorophyll and logof bacterial abundance but only at chlorophyll concen-trations above approximately 05 μg per liter Below thatconcentration bacterial abundance did not correlate signifi-cantly with chlorophyll It should be noted that subsequentanalyses have revealed that early epifluorescence measure-ments of bacterial biomass included the common cyanobacte-rium Prochlorococcus which can make up to 20 of totalbacterial numbers (67) Nonetheless heterotrophic bacterialbiomass is a major fraction of the living biomass of all plank-tonic ecosystems
The geographical and temporal distributions of marineprotozoa are much more varied than those of the bacteriaAs an all-inclusive group protozoa generally occur in plank-tonic ecosystems at abundances ranging from 10 s to 1000 sper ml Abundances in benthic ecosystems can be one tothree orders of magnitude higher commensurate with thehigher abundances of bacteria in those ecosystems Howeverit is important to remember that like the term ldquolsquobacteriardquo theterm ldquoprotozoardquo is a rather artificial conglomeration of evolu-tionarily and ecologically divergent taxa (see ldquoThe Changingand Complex World of Eukaryote Phylogenyrdquo) Thus theabundances of specific lineages of bacteria or protozoa mayshow spatial (or temporal) variability that is considerablygreater than the variability characteristic of these overarchinggroupings
The Changing and Complex World ofEukaryote PhylogenyNot that long ago textbooks still divided eukaryotic organ-isms into four major kingdoms (Animalia Plantae Fungiand Protista) while prokaryotic organisms were placed intoa single kingdom theMonera (68)Within this scheme pro-tists (eukaryotic organisms that can exist as single cells) weredivided into two subkingdoms (algae and protozoa) based ontheir basic nutritional mode a carryover from the historicaldistinction between single cells with ldquoanimal-likerdquo or ldquoplant-likerdquo nutrition This distinction presupposed a basic evolu-tionary divergence among protists into species that retaineda heterotrophic phagocytotic mode of life (protozoa) andthose that abandoned phagocytosis for a photosyntheticmode of life (algae)Moreover the presenceabsence of chlor-oplasts was a feature that could be easily distinguished by earlymicroscopists
The five-kingdom classification system of Whittaker wasrecognized as an improvement over previous classificationschemes but it posed a number of problems relating toprotists For example the distinction between single-celledand multicellular eukaryotes was somewhat arbitrary Moreimportant the division of protists based on whether theywere heterotrophic or photosynthetic was clearly not anappropriate feature if the classification was to recapitulateevolutionary relationships We now know that chloroplastacquisition and loss has occurred several times in the bio-logical history of our planet (69) giving rise to some closelyrelated protistan taxa that differ largely in the presence orabsence of a chloroplast Further complicating the matterwithin many protistan lineages there are species that possesschloroplasts and carry out photosynthesis (phototrophy)but also possess the ability to ingest and digest prey (hetero-trophy 70ndash72) Some heterotrophic protists even ingest
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-5
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
phytoplankton prey and retain the chloroplasts of their prey ina functional state for a limited amount of time (kleptidoplas-tidy73) Various forms and degrees of mixotrophy (mixedphototrophic and heterotrophic nutrition) are commonamong a number of algalprotozoan lineages (74ndash77) UnderWhittakerrsquos scheme phytoplankton ecologists studying a lin-eage of microalgae might have had little familiarity withclosely related heterotrophic species while protozoologistsstudying a particular protozoan groupmight have known littleabout closely related photosynthetic species
One might expect given these caveats that the termsldquoalgaerdquo and ldquoprotozoardquo are no longer used In fact the termldquoprotozoardquo is still commonly used (especially by ecologists)to recognize those eukaryotic species that exist as single cellsand whose nutrition is dependent on the uptake of preformedorganic substances (primarily via prey ingestion) while pro-tists possessing chloroplasts are still commonly called ldquoalgaerdquoSimilarly although the term ldquoprotistrdquo has been abandoned asa kingdom designation it is still widely employed to describeeukaryotic species that are capable of existence as single cells(ie algae and protozoa) The term ldquophagotrophic protistrdquohas also gained popularity in recent years because it recognizesthat many protistan species are capable of phagocytosis eventhough they may also possess their own chloroplasts and thusare technically ldquoalgaerdquo
Despite the shortcomings of Whittakerrsquos scheme it domi-nated the hierarchical organization of life for approximately aquarter century During the past few decades however thissystem has given way to a new organizational scheme that rec-ognizes three domains of life (Archaea Bacteria Eukarya [orEucarya] 78 Fig 2 upper panel) and is based onwhat is pres-ently believed to reflect amore realistic view of the evolution-ary distances that have developed between organisms in theasymp4 billion years that life has existed on our planet Withinthe Eukarya of Woesersquos scheme hypotheses regarding thephylogeny of ldquoprotistsrdquo have changed continuously and rap-idly during the past two decades reflecting new insightsinto eukaryote evolution provided largely by DNA sequenceinformation (Fig 2 lower panel from [79])
The former protistan phyla ofWhittakerrsquos systemhavenowbeen dispersed among candidate ldquosupergroupsrdquo within thedomain Eukarya to better reflect hypothesized phylogeneticrelationships For example the dinoflagellates (which encom-pass phototrophic heterotrophic and mixotrophic species)now form a single group and have been placed together withthe ciliates and apicomplexans (sporozoans) in themonophy-letic Alveolata (Fig 2 lower panel) On the other handeukaryotic heterotrophic single-celled species falling withinthe general description ldquoprotozoardquo are now widely distributedamong a number of protistan lineages In short nutrition hasbeen demoted as a phylogenetic character and other charac-ters (presumably more indicative of evolutionary relatedness)have ascended to address some long-standing contradictionsalthough the debate over the relationships among some line-ages is still very active at the present time 79)
Individual protozoan cells range in size from less than 2 micromto greater than 1 cm in diameter (gt4 orders of magnitude)(80 81) with some colonial radiolaria forming cylindricalgelatinous structures a centimeter in diameter and morethan a meter in length (82 83) Because they constitutesuch a large size range of organisms protozoa are often dividedinto size classes that very crudely correlate with their generalnutritional preferences A commonly used convention is thatof Sieburth et al (84) which groups planktonic microorgan-isms into order-of-magnitude size classes (02ndash20 microm =picoplankton 20ndash20 microm = nanoplankton 20ndash200 microm =
microplankton 02ndash2 mm =mesoplankton) Most protozoafall into the nanoplankton or microplankton size classesModeling microbial trophodynamics using this conventionassumes that protozoa in one size category generally consumeprey one order of magnitude smaller in size (34 85)Although this approach misses much of the detail and diver-sity of the trophic activities of individual protozoan taxa it isa necessary practical compromise for examining community-scale flows of energy and elements It also provides a usefulmechanism for summarizing and comparing the abundancesand biomasses of protozoa from different environments andto other microbial assemblages Protozoan abundance sum-marized in this way has been shown to contribute signifi-cantly to the living biomass of planktonic ecosystemsthroughout the world ocean (Fig 3)
Estimates of protozoan biomass such as those depicted inFig 3 typically do not include the contribution of mixotro-phic phytoflagellates to heterotrophy There is presently noeasy way to determine the abundances of small phagotrophicphytoflagellates in natural samples so these species are typi-cally counted as phytoplankton unless specific methods areemployed to identify the algae as consumers such as the useof fluorescently labeled particles (87ndash90) or through theexamination of food vacuole contents (91) On averagethese species appear to constitute a modest percentage ofthe phytoplankton assemblage (typically lt25) althoughthey may at times dominate the phototroph assemblages ofnatural plankton communities It is important to recognizethat their inclusion as functional heterotrophs rather thanphototrophs could significantly shift the relative contribu-tions of phototrophic and heterotrophic microbial biomassto total biomass within microbial assemblages and the flowof energy within plankton communities (92)
Heterotrophic protists that harbor photosynthetic pro-tists or their chloroplasts within their cytoplasm constituteanother complexity for estimating the contribution of pro-tozoa to total microbial biomass When bulk water samplesare analyzed the contribution of chlorophyll containedwithin those protozoa is generally assumed to come fromfree-living phytoplankton However studies have shownthat chloroplast-bearing ciliates can contribute up to halfthe total biomass of planktonic ciliates in ecosystems andchloroplast-retaining ciliates can sporadically dominate thechlorophyll and primary production of some planktonic eco-systems (93ndash96) The environmental conditions promotingthe success of these ciliates are poorly known Similarlymany species of planktonic foraminifera polycystine radio-laria and acantharia harbor large numbers (thousands perprotozoan) of endosymbiotic algae within their cytoplasm(97 98) Caron et al (99) have demonstrated that primaryproductivity within these species can contribute significantlyto total primary productivity in oceanic ecosystems and can bevery important locally in the convergences of Langmuir circu-lation cells (100)
BACTERIA AND ARCHAEAldquoCulturablerdquo versus ldquoNonculturablerdquo CellsMost conventional cultivation methods can grow only 1 orless of the bacteria that can be visualized by direct microscopytechniques (eg 4) This is true even though most can beshown to be active by techniques such as microautoradiogra-phy (25) These readily cultivable organisms appear to repre-sent a group of fast-growing so-called weeds that are adaptedto take advantage of rapid growth in rare organically enriched
422-6 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
environments This strategy contrasts with the numericallydominant bacteria that are adapted specifically for growthin the dilute nutrient conditions that characterize the vastmajority of the volume of the water column
The most common taxa readily cultured from seawaterwith standard nutrient broth media include the gamma pro-teobacterial genera Vibrio Alteromonas PseudoalteromonasMarinomonas Oceanospirillum Shewanella (usually isolatedfrom surfaces such as shellfish and sediments) the alpha pro-teobacterial generaRoseobacterSphingomonasmembers of thefamily Flavobacteriaceae and Planctomycetes as summarizedin Giovannoni and Rappeacute (101) and Fuhrman and Hagstrom(102) The cyanobacteria Synechococcus and Prochlorococcusare also now readily culturable but on low-nutrient inorganicmedia targeting photosynthetic forms as opposed to organicmedia used to cultivate the others listed above
Because of the low percentage of marine bacteria that canbe grown in standard media organisms that until recentlywere called ldquononculturablerdquo make up the large majority ofbacteria in the plankton Only during the pastsim20ndash25 yearshave molecular biological methods based on 16S rRNA genesequences been available to identify these organisms andthese powerful techniques have opened up a large area forexploration (see next section) Similar but more recent stud-ies use 18S rRNA sequences for characterizing protistandiversity as will be noted below
Molecular Phylogeny and MetagenomicsField ApplicationsModern phylogeny of microorganisms is based primarilyon genetic sequences the most well-studied gene being the
FIGURE 2 The three domains of life (upper left) as proposed by Woese et al (78) and a recent overview of modifications that have beenproposed by Adl et al (79) to higher-level phylogentic groups within the eukaryotic component of the tree (lower right) Domains figure fromWoese et al (78) eukaryotic tree figure from Adl et al (79) doi1011289781555818821ch422f2
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-7
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
small subunit ribosomal RNA gene (16S rRNA in Bacteriaand Archaea and its larger homolog 18S rRNA in eukar-yotes) This molecule is strongly conserved over evolutionarytime so this single molecule has been used for constructingphylogenetic trees of all living organisms (httptolweborgtreephylogenyhtml) Analysis of 16S18S rRNA genesequences has been used to evaluate deep evolutionary rela-tionships among organisms and was instrumental in point-ing out that Archaea Bacteria and Eukarya should beconsidered different Domains of equivalent phylogeneticrank above kingdoms (78) However there are sufficientdifferences in 16S18S rRNA gene sequences to demonstrate
relationships (ormake distinctions) at the genus or sometimesspecies level
The first phylogenetic studies based on 16S18S rRNAgenes used sequences derived from cultures However onedoes not need cultures to obtain rRNA gene sequences (orany other sequences for that matter) An idea developed inthe lab of Norman Pace in themid-1980s involved extractionof DNA directly from natural samples and then cloning andsequencing of the DNA as a means of assaying the microbespresent in the samples (103 104) The original protocolscalled for cloning by creating what are called ldquophage librariesrdquofrom the natural DNA but since 1986 PCR has been applied
FIGURE 3 (a b) Plankton biomass in the Arabian Sea during the 1995 southwest monsoon (a) and intermonsoon period (b) Areas of theboxes indicate the relative magnitudes of the biomass in each category Categories within the dashed boxes in a b c are composed of protozoaArrows indicate the direction of energymaterial flow in the food web thicker arrows depicting greater flow Redrawn from (86)(c) Depth-integrated biomass (mgm2) in the upper 100 m of the Sargasso Sea near Bermuda and in the upper 200 m of the equatorial Pacificat 175degE Thewidth of the bars indicates the biomass in each size category Heterotrophs have been separated by size class while phytoplanktonhave not Size classes delineated by the dotted box are comprised of protozoa Redrawn from (65) doi1011289781555818821ch422f3
422-8 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
extensively for cloning and related studies The targetsequence can be almost instantly ldquoidentifiedrdquo to its closestphylogenetic neighbor by what have come to be standardonline sequence comparisons
Beyond the study of targeted genes (like the 16S rRNAgene) shotgun metagenomic studies have examined theentire genetic repertoire of the microbes in a given sampleThe metagenome is the collective genome of all organismsin the sample Initially these studies extracted DNA fromall organisms in a sample (usually prefiltered through a filterapproximately 1 microm to remove most organisms larger thanbacteria) sheared it to produce fragments and clonedthem into standard vectors either as small (thousands ofbases) or large (to hundreds of thousands of bases) insertsthat is fragments of DNA from the environment nowcloned into the vectors in a form suitable for sequencingThe best known early marine study of the former type isthe Global Ocean Survey with initial results published byVenter et al (105) which generated more than a billionbases of DNA sequence and reported 12 million previouslyunsequenced genes estimated to come from at least 1800different genomic species cumulatively in the many samplesthey analyzed
With the advent of next-generation sequencing that gen-erates millions ormore sequences in a run (known by a varietyof acronyms including 454 Illumina SOLiD etc) cloningof genes has largely been replaced by clone-free sequencingThe extent and power of such sequencing has recentlybeen demonstrated by the ability to construct essentiallythe entire genome of an uncultivated marine Group II Eur-yarchaeon that constituted only approximately 2 of amicrobial community sample using SOLiD sequencingmade possible by high coverage and the use of mate pairsequencing ofsim3000 bp fragments (106) (this length isneeded to span repeats and highly conserved genes that oth-erwise make genome construction difficult)
For 16S rRNA gene studies ldquotag sequencingrdquo pioneeredby Sogin et al (107) consists of amplifying a suitable sizedpart of the gene with broadly conserved primers (choice isimportantmdashfew are truly universal for the groups intended)often ldquobarcodedrdquo to allow multiple samples to be combinedinto a single run The amplified products are then sequencedand sequences processed en masse Many thousands of partialSSU rRNA gene sequences per sample are economically ana-lyzed this way though they are usually short (currently a fewhundred bases each depending on the sequencing platform)This way of analyzing the composition of microbial commun-ities is now standard These approaches provide so muchinformation even about very rare sequences that the resultshave led to the important concept of the ldquorare biosphererdquoorganisms that may be active or dormant and constituting avery small proportion of the community (eg often muchless than 01) but are potentially important for dispersionadaptation to changing conditions or even critical activitieslike nitrogen fixation or vitamin production (108) Howeverdue to the potential for artifacts like error sequences thisapproach requires rigorous application of quality filteringand clustering algorithms to avoid erroneous taxa and overes-timation of species richness (109 110)
Next-generation sequencing approaches have also allowedwork to begin metagenomics studies of microbial eukaryotes(111) These studies are still constrained by the much largergenomes of eukaryotes and therefore the difficulties ofobtaining sufficient sequences to reconstruct the genome ofa particular species Along with advances in sequencing abil-ity studies of eukaryotic metagenomics have been facilitated
by advances in single-cell isolation which when coupledwith high-throughput sequencing approaches reduce the tre-mendous complexity present in natural complex eukaryoticcommunities to a manageable task (112) Such single-celltechniques are also quite valuable in studies of bacteria andarchaea though the amplification technique tends to bevery uneven and typically generates less than half the genomeof each isolated cell regardless of domain (113 114)
Additionally similar to DNA mRNA is amenable toextraction and sequencing although greater care must betaken during extraction and purification as RNA shows agreater susceptibility to degradation during processing Copy-ing of mRNA by reverse transcription of RNA into cDNAfollowed by DNA sequencing has allowed insights into themetatranscriptomes of environmental samples Metatran-scriptomic studies provide information on gene expressionin an ecosystem and thereby indicates ldquoactivityrdquo of themicro-bial community rather than simply ldquopotentialrdquo represented bythe genomic DNA present in the sample with many applica-tions from showing which processes are being carried outby which organisms to fine-scaled diel studies (115ndash118)Nevertheless given variations in the lifetimes of differenttranscripts and protein molecules the transcriptome maynot be fully representative of the current activity of anorganism
Limitations of these analyses include sequencing errorsPCR mismatches or biases clustering and bioinformaticschallenges and chimeras generated during PCR Also thephylogenetic resolution of short sequences is limited giventhe high conservation of rRNA sequences Even with clone-free shotgun metagenomics there may be biases such as non-random losses of DNA during extraction and preparationor biases (eg from G +C content or secondary structure)in the sequencing procedures Determination of speciesdiversity by these approaches provides an example of theselimitations Shakya et al (119) working with synthetic com-munities (purified genomicDNA from16Archaea represent-ing 3 phyla and 48 Bacteria representing 16 phyla remixed tosimulate an environmental DNA extract) applied both meta-genomic analysis (454 and Illumina platforms) and PCRamplification followed by 454 sequencing of 16S rRNA genesto determine both species richness and relative abundanceThey found that PCR amplification454 sequencing of 16SrRNA genes yielded an accurate measure of species richness(providing that appropriate data processing was applied)but that the relative abundance of up to 94 of the species(depending on domain and variable region amplified) wasover- or underestimated by at least 15-fold (values rangedfrom not detected to 103-fold overestimation) In contrastboth metagenomic approaches yielded relative abundancesthat were within the authorsrsquo 15-fold accuracy cutoff forsim50 of the species However they concluded that addressingrichness overestimation in metagenomic analyses that isdistinguishing rare but real OTUs from experimental andcomputational artifacts awaits further computational andclassification improvements More recently Parada et al(120) used mock communities composed of 16S rRNAclones from 27 common marine taxa (from nine Bacterialand two Archaeal phyla) to show that small differences inPCR primers (and different clustering methods) can yieldlarge differences in apparent relative abundances of reportedtaxa However one primer pair and informatics pipelinethey tested using a particular version of 515F-926R (V4ndashV5) provided accurate estimates of relative clone abundance(r2 = 095) when comparing observed versus expected cloneabundance
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-9
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
Culture-Independent Diversity Studies
The first groups to be identified using cultivation-independ-ent molecular techniques from the marine plankton (121)were the bacterial SAR11 cluster (a group of closely relatedgene sequences or phylotypes) and marine picoplanktonicunicellular cyanobacteria Synechococcus and Prochlorococcus)Of these the SAR11 cluster was completely unknown butthe cyanobacteria had previously been recognized by theirunique pigment fluorescence Waterbury et al (122) andJohnson et al (123) used epifluorescence to observe Synecho-coccus and Chisholm et al (124) discovered Prochlorococcususing flow cytometry These cyanobacteria were later isolatedand grown in phytoplankton culturemedia These two groupsare generally common in the euphotic zone with SAR11 typ-ically comprising one third of the planktonic bacteria (125)and the cyanobacteria common everywhere but polar waters
Probably the biggest surprise to come from the applicationof molecular tools was the discovery by Fuhrman et al (126)of abundant archaea in the deep sea The archaea were foundto be in a unique phylogenetic cluster that was only distantlyrelated to any previously known archaea but the ldquoclosest rel-ativesrdquo (not really close at all) were extreme thermophiles Asubsequent study also found archaea to be present in near-surface coastal waters albeit relatively rare (lt2 relativeabundance) This study used a PCR technique specificallytargeted archaea and found ldquomarine Crenarchaeardquo plus a sec-ond group belonging to the phylum Euryarchaea (127)
Up until this time all known Archaea were thought to beldquoextremophilesrdquomdashadapted for either very high temperatures(thermophiles) extremely salty conditions (halophiles) orstrictly anaerobic environments (methanogens) Yet theseorganisms were present in cold or cool water at ordinary salin-ity and high oxygen concentrations Fluorescent in situhybridization (FISH) measurements from deep-sea sampleshave since showed that the archaea may make up appro-ximately 40 of the total countable prokaryotes with thepercentage reaching to 60 at 200 m depth in theMediterra-nean (35 36) An extensive time series of FISH measure-ments near Hawaii confirmed that the archaea are indeedvery abundant throughout the year from below the photiczone to at least 4000 m and typically constituting 30ndash40of the total prokaryotes present in waters deeper than a fewhundred meters (128) with similar results found elsewhere(129 130) (Fig 4) They have been reported from many pla-ces including the Atlantic Pacific and Southern Oceansand are dynamic components of the plankton the most com-mon type by far being the ldquomarine Crenarcheaeardquo (128 129131) While the marine archaea have been reported to bedominated by a few major ldquophylotypesrdquo (132) they alsohave been shown to have a great deal of microdiversity withinthese phylotypes suggesting there are many kinds of close rel-atives coexisting (133) Interestingly this group of archaeamay likely be the most abundant kind of organism on Earthgiven the huge volume of the deep sea and their high abun-dance there (134) It has recently been proposed that ldquomarineCrenarchaeardquo be elevated to their own major phylum outsidethe Crenarchaeota called the Thaumarchaeota on the basisof deep phylogenetic branching and fundamental differencesbetween them and the Crenarchaeota (135) The Thau-marchaeota possess the uniquely archaeal membrane lipidcrenarchaeol and are nowalso known to be abundant in soils
The physiology of the Thaumarchaeota has been anintriguing area of study Initially an autoradiography-FISHcombination approach demonstrated that they take upamino acids (130 136) But subsequent data showed that
crenarchaeol was derived largely from CO2 suggesting auto-trophic metabolism (137 138) An experiment showinguptake of 13C-labeled bicarbonate into these archaeal lipids(139) directly pointed to autotrophy in this group Chemoli-thoautotrophywas first hinted at byVenter et al (105) whosemetagenomic analysis showed an apparent archaeal scaffoldthat contained genes suggestive of ammonia oxidation Fur-ther evidence came from Schleper et al (140) who detectedseveral ammonia oxidation genes in order directly adjacent toa Thaumarchaeota 16S rRNA gene in a soil-derived metage-nomic clone The issue was directly resolved when Konnekeet al (141) isolated a related marine archaeon from sedimentof a marine aquarium and this organism Candidatus Nitroso-pumilus maritimus was found to have a chemoautotrophicmetabolism quantitatively oxidize ammonia to nitrite andcontain an archaeal ammonia monooxygenase gene (amoA)and interestingly did not grow heterotrophically Cultivationexperiments showed this organism has a high affinity forammonium allowing it to outcompete bacterial nitrifiers atlow (submicromolar) concentrations as are typical in thesea (142) The complete genome of the organism has shownnovel adaptations for nitrification and autotrophy (143)
Even if the Thaumarchaea are primarily chemolitho-autotrophs field data suggest a level of mixotrophy in thatsome organic substrates are being incorporated into biomassA stable isotope study using cells collected at 670 m depthoff Hawaii estimated that about 80 of the carbon incor-porated into archaea-specific lipids came from inorganic sour-ces and about 20 from organic compounds (144 145)Interestingly it has also been claimed that the genomesof deep-sea (gt2000 m depth) members of the Thaumarch-aea as well as those living in equatorial waters rarely containthe amoA gene and thus may be primarily chemoorgano-trophs (48)
Major bacterial groups that have been documentedfrom seawater using 16S rRNA characterization includesome that are also known from culture (eg AlteromonasRoseobacter) and several that are phylogenetically distantfrom standard cultures The most common groups are inrough order of their relative abundance in clone librariesfrom most to least abundant SAR11 (relatives of Pelagibac-ter ubique) Roseobacter SAR86 cyanobacteria SAR116SAR202 SAR234 andMarine Group A The SAR designa-tion followed by a number is an arbitrary sequential cloneidentifier from Sargasso Sea cloning studies done by the Gio-vannoni lab that did most of the early systematic cataloging ofclones Summaries of the data and phylogenetic relationshipsof these groups can be found in Fuhrman andHagstrom (102)and Giovannoni et al (101)
Molecular Genetic Discoveries in Bacterial andArchaeal Marine BiologyAs described earlier metagenomics is an extension of theideas used in the 16S rRNA cloning studies in that all genesfrom the native microorganisms are separated and clonedwithout having cultivated the organisms These methodsearly on started to find unexpected and very interestingresults The best examples involve unexpected marine photo-trophy the first of which is discovery of a nonchlorophyllphotosynthetic bacterial pigment called proteorhodopsin(146) The gene was found on a large environmentallyderived fragment of DNA that also had a gene coding for16S rRNA from the so-called SAR86 group (one ofthe groups common in seawater) This pigment can act as alight-driven proton pump thought to permit cells to generate
422-10 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
FIGURE 4 Distribution of Bacteria Euryarchaea andmarine Crenarchaea (recently renamed Thaumarchaea) along a transect in theNorthAtlantic Ocean as measured by CARD FISH and expressed as of total bacteria + archaea counts via DAPI stained epifluorescence fromTeira et al (130) Top panel shows station locations that are shown on the top of the lower three panels with the distance in km reported alongthe transect from lower to higher station numbers doi1011289781555818821ch422f4
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-11
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ATP from sunlight Interestingly different versions of this pig-ment are found at different depths apparently ldquotuningrdquo theabsorption to match the ambient wavelengths of light asthey change with depth (147) The proteorhodopsin genesare very widespread and diverse perhaps in half or more ofall marine bacteria including SAR11 and Euryarchaeota(105 148ndash150) However almost all of several culturedorganisms with proteorhodopsin studied to date do not showa growth benefit from light (150) with the exception of amember of the Flavobacteria Dokdonia sp strain MED134that grows faster in the light only under moderately lownutrient conditions (151) and a marine Vibrio has beenshown to survive starvation longer in the light than in dark-ness (152) These observation may explain the wide phyloge-netic distribution and high abundance of this gene in thatproteorhodopsin may often assist long-term survival of bacte-ria under extreme energy-limiting conditions yet not havemucheffect on growthduringmore energy-replete conditionsInterestingly a recent report suggests the growth benefit fromlight in Dokdonia MED134 is from enhanced uptake of itsrequired growth factor vitamin B1 which is transported by atonB-dependent transporter powered by a proton gradient(153) This pigment might also have sensory or other rolesnot yet well examined (150)
The second kind of phototrophy found by moleculargenetic and fluorescence techniques to be unexpectedlyimportant in marine plankton is anoxygenic aerobic bacterialphototrophy based on the pigment bacteriochlorophyll aCulturable aerobic anoxygenic photosynthetic (AAP) bacte-ria have been known from seawater for several years (154) andare widespread (155) Initial recent reports of direct countsclaimed they represent about 11 of the total bacterial com-munity in the euphotic zone (156) but those early counts didnot correct for the presence of other bacteria and such correc-tionoften yields estimates averaging closer to 2(157)Directmeasurements show the bacteriochlorophyll pigment is rela-tively rare (158) However studies in various ocean locationsincluding ocean gyres report that these AAP bacteriacan indeed on occasionmake up a quarter of the total prokar-yotes (159 160) A recent energetic modeling of AAP andproteorhodopsin-containing bacteria (161) has suggestedthat proteorhodopsin-containingones cangain approximately02asmuchenergy from sunlight asSynechococcus (commonmarine cyanobacterium) and AAP bacteria can gain approxi-mately 13 as much energy from sunlight as SynechococcusThey concluded the AAP bacteria may gain energy sufficientto meet maintenance costs but proteorhodopsin-containingbacteria were not expected to do so except at high light inten-sities andwith largenumbersof proteorhodopsinmoleculespercell They concluded the ease and low cost of maintainingPR-based phototrophy (a few genes required) may explainthe high incidence of proteorhodopsin genes
Other metagenomic studies have yielded interestinginsights about organisms and processes that would otherwisebe difficult or impossible to establish For example thesequence of an archaeal gene fragment isolated from seawaterrevealed extensive evidence of genetic exchange with othertypes of archaea and even bacteria (162) Such geneticexchange was also quite evident from the complete genesequences of different strains of Prochlorococcus and Synecho-coccus which was attributed in part to virus-mediated geneflow (163ndash166)
Dilution Cultures of Bacteria and ArchaeaSome recent cultivation techniques that permit growth ofldquotypicalrdquo oligotrophic marine bacteria in pure culture are
based on the simple concept that bacteria living in seawateron dissolved organic matter might best be grown in the labo-ratory in ordinary filtered seawater This conclusion followsfrom similar techniques to grow mixed ldquoseawater culturesrdquoof marine bacteria (167) but the seawater inoculum in thiscase is diluted so that only one or a few bacteria are addedto the initial culture vessel (168) A rapid throughput versionof this method has been used to cultivate members of bacte-rial groups thought to be conventionally uncultivable such asthe SAR11 clade (169 170) albeit often at very low den-sities Such cultures permit focused studies of the propertiesof those organisms including genomic analysis (169 170)This valuable work provides insight into the roles and activ-ities of some of the most common bacterial phylotypesobserved in the ocean (171) showing for example that culti-vated SAR11 requires a source of reduced sulfur as well asproviding templates aiding the interpretation of metage-nomic analysis (172)
PROTOZOADiversity and DistributionHeterotrophic protists have been identified historically fromtheir morphological features that are apparent at the levelof the light or electronmicroscope Features of cell size shapetype and pattern of flagellationciliation skeletal structurescharacteristics of the nucleus and other cellular structuresand organelles have been used to differentiate themany thou-sands of described species A description of the extent of thisdiversity of form and function is well beyond the scope of thisbook (see [79] for more information) However much of thisdiversity can be reduced to three basic body plans that dictatethe broadest ecological roles of these cells amoeboid flagel-lated and ciliated forms (Fig 5) Among the lineages possess-ing one of these three forms only the last group the ciliatedprotists constitute a monophyletic group within the domainEukarya
One of the simplest body plans for protozoa is the amoe-boid cell exemplified by the gymnamoebae or ldquonakedrdquoamoebae Motile nonphotosynthetic cells lacking flagellaor cilia occur as life stages in a number of phylogeneticallydiverse taxa but for many free-living species of protozoathis form constitutes the only life stage Motility is largelyconfined to movement along surfaces by means of pseudopo-dia that can take on a variety of (species-specific) shapes Sig-nificant abundances of amoebae are largely confined tobenthic and epibiotic environments (174) and to suspendedparticulate material where they can occasionally be highlyenriched (27) Most amoebae consume bacteria and otherminute prokaryotes and eukaryotes
Several heterotrophic protistan groups possess complexamoeboid body plans most notably the foraminifera polycys-tine and phaeodarian radiolaria and the acantharia Plank-tonic forms are predominantly oceanic in their distributions(although a large number of benthic species of foraminiferaexist) These species are heterotrophic and many are visibleto the naked eye (individual cells can bege1 cm gelatinouscolonies can form ribbon-like structuresge1 m in length)They feed on a wide variety of bacterial protistan and meta-zoan prey using pseudopodial networks have rather longcomplex life cycles for individual cells (weeks to months)and are extensively used in studies of paleoclimatologicalreconstruction (82 175) In addition many of these speciespossess intracellular symbiotic usually eukaryotic algaeThe widespread occurrence of algal endosymbiosis among
422-12 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
these protozoa in oceanic pelagic environments implies thatstrong selective forces appear to give rise to these associations(97 98)
Flagellated forms of protozoa exist across many protistanlineages Eukaryotic flagella come in various sizes numbersand forms (eg with or without tiny hairs) that are character-istic of the different lineagesMany flagellate species are appa-rently purely phototrophic or heterotrophic (phagotrophic)but a significant (still poorly known) fraction of flagellates aremixotrophic combining both phototrophy and phagotrophy(70 88) Flagella in phagotrophic flagellates are employed formotility and prey capture Most free-living flagellates possessone to four flagella (typically one or two) that can be manytimes the length of the cell itself Although there is tremen-dous species diversity among flagellated protozoa many ofthese species have broadly overlapping ecologies As a groupflagellates are the most numerically abundant protozoa ofboth benthic and pelagic ecosystems and they are fundamen-tally important as consumers of bacteria cyanobacteriaand other eukaryotes Flagellated protozoa within the nano-plankton size class (2ndash20 microm) are often counted as a singleassemblage in plankton studies because morphological details
apparent using light microscope are insufficient to distinguishamong the many species This assemblage has been variouslyreferred to by a variety of names and acronyms including het-erotrophic nanoplankton (HNAN HN) heterotrophic flag-ellates (Hflags) heterotrophic nanoflagellates (HNF) andmicroflagellates Heterotrophic nanoplankton is the mostaccurate term for most methodologies employed to countthese cells because it does not require visualization of flagella(which are often lost from these small cells during preparationfor microscopy) The confusion over terminology is partly aconsequence of the different methodologies that have beenused to count these species and partly a consequence of thefact that these species were first studied prior to the wideacceptance of the size convention of Sieburth et al (84)
Heterotrophic flagellates in the microplanktonic size class(20ndash200 microm mostly dinoflagellates) are important consum-ers of phytoplankton in pelagic ecosystems Many of thesespecies are capable of the production of large pseudopodialnets the pallium that can engulf prey (particularly diatoms)significantly larger than the diameter of the dinoflagellatetheca (176) This behavior and its ecological significance forenergy flow in plankton communities have been recognized
FIGURE 5 Body plans and size ranges of protozoa These micrographs depict amoeboid (andashg) flagellated (hndashj) and ciliated (kndashm) forms ofprotozoa From Caron et al (173) Markers bars are 5 (i) 10 (h) 20 (c j l) 30 (a) 50 (k m) 100 (b f) 500 (d) and 1000 (e g) micromdoi1011289781555818821ch422f5
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-13
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
only within the past few decades (177) These species areoften abundant in waters where diatoms dominate (178179) In such situations heterotrophic dinoflagellate biomasscan be similar to that of ciliated protozoa (180)
Ciliates are generally the most recognizable form of pro-tozoa to nonspecialists Nonetheless ciliates are a diversegroup morphologically as well as ecologically (181 182)The degree of ciliature on these species can range from uni-formly ciliated to totally devoid of cilia during most of theirlife cycle to ciliature restricted to specific regions of thecell In addition cilia can fuse to form various complexstructures (eg ciliary membranes or veils cirri) that assistin locomotion food capture or attachmentCiliates are abun-dant and ecologically important species in both benthic (8)and pelagic (183) ecosystems and can consume a variety ofprokaryotic and eukaryotic prey Together with the heterotro-phic dinoflagellates these species are the dominant consum-ers of phytoplankton in many pelagic ecosystems (184) andas a consequence they form an important trophic link to met-azoan zooplankton (185 186)
Diversity and Biogeography of Protists FromMorphology to DNA SequencesUntil relatively recently the biodiversity of marine protistanassemblages was not generally considered a controversialtopic It was generally accepted that while all species of pro-tists certainly have not been identified representatives ofmost types of algae and protozoa had been observed anddescribed if not actually brought into culture in the labora-tory Direct sequencing of 18S rRNA genes from environ-mental samples (as described in ldquoMolecular Phylogeny andMetagenomics Field Applicationsrdquo) has changed that viewInitial forays into environmental DNA indicated a muchgreater diversity of protists than previously documented usingclassical approaches of culture and microscopy (81 187ndash191) Numerous publications over the past decades haveexpanded these findings (see [192] for a recent global analy-sis) which are highly analogous to discoveries in marine pro-karyote research as described already
Previously uncharacterized protistan diversity has nowbeen documented at virtually every level of eukaryoticorganismal classification Some of these findings couldhave been expected but some have been very unexpectedFor example a much greater diversity than noted previouslyhas been observed among small (lt10 microm) protists in plank-tonic ecosystems These species generally possess few distinc-tive morphological features and one could expect that manycryptic species might be present among these small morpho-types The molecular ldquodiscoveryrdquo of this eukaryotic diversityhas stimulated progress on the isolation and descriptionof new species and genera of minute algae and protozoa(193ndash197)
Analyses of environmental samples have also indicatedthe unanticipated existence of novel 18S rRNA gene sequen-ces that imply the presence of novel lineages of eukaryotes innatural protistan communities (198ndash200) These are sequen-ces for which there are apparently no known described orcultured species The degree to which these sequences dif-fer from sequences of known sequenced eukaryotes impliesthat some of these lineages may be distinct at the level ofphylum (201)
These findings have raised basic questions and somedebate concerning the true diversity and biogeography ofprotistan assemblages in natural ecosystems (202ndash204) thevalidity of the many novel phylotypes or cryptic species that
are being documented (205) the potential importance andsignificance of the many rare taxa that characterize thesecommunities (206) and the implications of these findingsfor the ecologicalbiogeochemical roles that protists playin aquatic ecosystems For example it was postulated andsubsequently confirmed that some novel alveolate lineagesreported from marine ecosystems make up a suite of parasiticprotozoa whose ecological importance may have been signifi-cantly underestimated in the past (207) Deciphering theidentity and significance of these many unknown phylotypeswill constitute a significant effort for protistologists in thefuture (80 85)
Life Histories and Ecological Strategies of ProtozoaAbundance prey type and life histories all vary tremendouslyamongmarine protozoa The smallest species (eg many flag-ellates) tend to be the most abundant and widely distributedin the world ocean Indeed many of these species may beglobally distributed (208) Most nanoplanktonic flagellateshave potentially rapid rates of grazing and growth Underoptimal conditions these species can divide by binary fissionseveral times a day and thus dramatic increases in their pop-ulations can take place in response to favorable conditionswithin a few days (209) Many of these species are capableof surviving for limited periods of time without food andhave developed a variety of physiological or life cycle strat-egies to cope with these events (209) However in contrastto some bacteria that may remain viable through long periodsof starvation (210) protozoa will expire or encyst in responseto low food abundance and thus ldquoboom-and-bustrdquo popula-tion changes are characteristic of small flagellates
On the other end of the size spectrum from the ratherubiquitous nanoflagellate species many of the largest proto-zoan species (polycystine and phaeodarian radiolaria plank-tonic foraminifera) are exclusively oceanic (ie do notsurvive in most coastal environments) andor have specificlatitudinal and depth distributions Thus the abundancesof these latter species may range from undetectable to max-imal abundances of gt105 individualsm3 Dramatic changesin the abundance of these protozoan taxa also can be relatedto changes in prey abundance physicalbehavioral aggrega-tion or to periodicity of life cycle events For example theplanktonic foraminifer Hastigerina pelagica reproduces on alunar cycle and thus abundances (and life stages) of this spe-cies in oceanic waters can vary considerably over the courseof a month (211) In general life cycles for the large amoe-boid protozoa are lengthy and complex (for single-celledorganisms) with life spans unknown for many species(attempts to culture them in the lab have so far been unsuc-cessful) but estimated to be on the order of months to per-haps years (175)
The abundance and activities of microplanktonic hetero-trophic protists (mostly ciliates and heterotrophic dinoflagel-lates) tend to be somewhat intermediate to those ofnanoplanktonic flagellates and the larger amoeboid formsThese species are present in the majority of marine ecosys-tems and collectively play an important role in the controlof phytoplankton biomass (and probably the abundance ofnanoplanktonic protozoa although there is little informa-tion on this topic) in waters throughout the world oceanLike small flagellates ciliates and dinoflagellates reproduceprimarily by binary fission but their maximal growth ratesare typically slower (one division a day is typical) and theirlife cycles often include sexual phases that allow for geneticrecombination
422-14 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
VIRUSESViral Abundance and General PropertiesViruses are simple biological agents typically 20ndash200 nm indiameter composed of a nucleic acid genome in a proteincoat that infect cells and ldquocommandeerrdquo the cellrsquos machineryto make more viruses which are released into the environ-ment when the host cell lyses or bursts A particular virus isthought to be capable of infecting only a narrow range of hosts(usually one species sometimes a genus rarely broader)Although some early studies had isolated viruses from thesea therewere no data prior to the 1980s showing such viruseswere very abundant and more significantly no evidence thatinfection was occurring in any important part of the planktoncommunity It wasnrsquot until the late 1980s that electronmicro-scopy with suitable concentration methods showed thatviruses are extremely abundant similar to or even higherthan bacterial abundance (212 213) Interestingly themost suitable transmission electron microscopy (TEM)approach used by Bergh et al (described in detail by [214])was actually similar to a direct centrifugation method devel-oped in 1949 (215)
TEM studies of viruses permit high-resolution images andobservation of viral morphology showing features such ashead diameter tails sheaths and tail fibers (Fig 6 collage
in upper left) Each type of virus has a fixed morphology(unlike bacteria that are potentially more plastic in theirappearance) and hence a coarse measure of viral diversityhas been possible by cataloging morphologies Studies thathave done so have found dozens or more different morpholo-gies of marine viruses in a given sample (217ndash219) A largeproportion resemble bacteriophages which are viruses thatinfect bacteria
The availability of brightly fluorescent nucleic acid stainsand high-porosity fine pore size (002 μm) filters made fromAl2O3 have made it easy to accurately count viruses by epi-fluorescence microscopy (38 220 221) Epifluorescence per-mits abundance estimates but does not allow observation ofviral morphology as viruses are below the resolution limitof light microscopy viruses are visible only as sources of light(like stars in the night sky see Fig 6 lower micrograph) Anextension of manual epifluorescence counts is the use of flowcytometry to count viruses now commonly employed in somelabs (222)
Virus abundance has been found to be closely related tobacterial abundance with a virusbacteria ratio typically10ndash301 A typical oceanic profile of bacterial and viral abun-dance is shown in (Fig 6 panel on right) This tight ratioand the strong correlation to bacterial abundance in relationto weaker correlations to chlorophyll have been cited as
FIGURE 6 Marine viruses (upper left collage) viewed by transmission electron microscopy These are cyanophages in the groups myoviridae(right and bottom) siphoviridae (left) and podoviridae (top) Scale bars represent 100 nm From Sullivan (216) Epifluorescence micrograph(lower left) of SYBR green I stained viruses (small fluorescing objects) and bacteria + archaea (large fluorescing objects) Depth distribution ofvirus and bacteria + archaea abundances obtained by epifluorescence microscopy of SYBRGreen stained cells in the central San Pedro BasinCalifornia (11 August 2000) doi1011289781555818821ch422f6
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-15
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
evidence that most marine viruses infect bacteria rather thaneukaryotic phytoplankton (217 218 223 224) though thereare also many important viruses of phytoplankton and otherorganisms with significant impacts on primary productivityblooms and ecosystem function (225ndash227)
The first demonstration of the activity of marine viruseswas presented by Proctor and Fuhrman (228) who showedby electron microscopic examination that viruses wereactually infecting marine bacteria and cyanobacteria at ameasurable rate Because only the final portion of the viruslife cycle is visible by electron microscopy (when the virusesare assembled and ready to lyse the host cell) only a very smallfraction of the infected cells can be counted as infected at anygiven time Proctor and Fuhrman (228) reported that only afew percent of the total bacteria from coastal waters and theSargasso Sea were visibly infected but they interpreted thedata with a model that implied the actual fraction of the totalcommunity that was infected was much higher They con-cluded that the total fraction of bacterial mortality attribut-able to viruses is roughly 10ndash40 Subsequent studies haveused refined versions of that same general approach andmodel (229 230) and numerous studies have used a varietyof alternative approaches to estimate viral activitymdashall yield-ing basically the same general conclusion (although themodel parameters need to be adjusted for cyanobacteriawhich on average seem to be infected less than bacteria)These approaches include (a) calculation of virus turnoverfrom decay estimates (231) (b) measurement of viral DNAsynthesis by incorporation of tritiated thymidine (232) (c)observation of effects of added viruses (233 234) (d) estima-tion of bacterial mortality in the absence of protists (e) use offluorescent viruses to estimate production by an approachanalogous to isotope dilution (38) and (f) dilution of virusesin filtered samples to estimate viral production (235) Theoverall consensus of these studies is that the initial estimateswere basically correct that is that viruses are responsiblefor about 10ndash40 of the bacterial mortality in most marinesystems (reviewed by [218 223 226 236 237]) Howeverthere is still some disagreement about the higher estimatesand it is likely that 40 mortality from viruses alone is nottypical for most marine systems
Most of the foregoing work has focused on the viral infec-tion of bacteria primarily thought to be heterotrophicalthough the original report by Proctor and Fuhrman (228)also noted the occurrence of cyanobacterial infection Sev-eral subsequent studies focused on phytoplankton includingthe potential effect of viruses on the termination of phyto-plankton blooms of Emiliania and Phaeocystis and the likelyimpact this might have on release of climate-active gasessuch as dimethyl sulfide (225 227 238ndash247) Althoughdetails are beyond the scope of this chapter viruses arethought to infect virtually all marine organisms with poten-tially significant impacts from zooplankton to whales (226)Detailed studies of viruses infecting cyanobacteria such asSynechococcus have shown differences in viral host specific-ity particularly toward coastal and oceanic host strainsand some occasional high virus abundances (to 105ml) asmeasured by most probable number (MPN) cultivation tech-niques in the Gulf of Mexico near Texas (248 249) Othervirus cultivation studies with Prochlorococcus and Synecho-coccus in oligotrophic waters of the Sargasso Sea showedan interesting pattern of cross-infection between these gen-era by some virus types (suggesting gene flow among theseorganisms) but generally low MPN estimates of abundanceto 103ml even when cyanobacterial abundance was near105ml (216 250)
Overall the consensus emerging from direct comparisonsof viral-mediated mortality and grazer-mediated (ie micro-zooplankton) mortality have indicated that viral lysis of bac-teria constitutes a significant fraction of total mortality ofthis assemblage while the mortality of phytoplanktonappears to be dominated bymicrozooplankton inmost instan-ces (251 252)
As mentioned viruses have significant morphologicaldiversity as observed in TEM studies It is also possible toinvestigate aspects of their genetic diversity In early workon this topic Wommack et al (253) and Steward et al(254) observed the diversity of viral genome lengths in a fieldsample by pulsed field gel electrophoresis with viral genomesranging 25 kb to gt300 kb in length Field results show thatthe viral community composition is dynamic in space andtime with clear changes in the banding patterns over seasonsand locations in Chesapeake Bay (253) between oceanbasins and subsequent to dinoflagellate blooms (254) andwith depths to 500 m at one location (255)
Although viruses do not all share a set of core genesthat allows a universal viral phylogeny (comparable to SSUrRNA gene in cellular organisms) genetic diversity amonga single group of viruses can be examined by sequence analysisof shared genes within the group An example is the g20 geneinT4-like cyanophages that has shown extremely high diver-sity even among very closely related viruses and has demon-strated geographic and seasonal variation (256) A secondexample is g23 which is found broadly in diverse and wide-spread T4-like phages (257) and can have seasonally repeat-ing patterns (258) as well as short-term rapid dynamics thatcorrelate to those in bacteria (259)
Viral MetagenomicsAs with cellular organisms metagenomics can be used toexamine viral diversity and genetics without themany restric-tions of cultivation Viruses can be collected by selective fil-tration and concentrated by tangential flow filtration orflocculated with iron chloride (260 261) then their collec-tive metagenome can be extracted linker amplified and ana-lyzed by sequencing (262 263) Viral metagenomic studiesare particularly challenging because the large majority ofsequences have no annotated matches in any databases butso far results from marine samples around the world haveshown extremely high diversity and variations with depthlocation and time presumably with highly dispersed typesselected by local conditions (262ndash266) Because metage-nomes are best interpreted when there are representative cul-tures available the best matches of viral metagenomesoriginally tended to be to the few viral isolates infecting trulycommonmarine bacteria like cyanophage infecting Synecho-coccus and Prochlorococcus (267) However the developmentof dilution-to-extinction cultures representing common het-erotrophic marine taxa like Peligibacter (a member of theSAR11 clade) and SAR116 has allowed isolation from sea-water of viruses infecting these common organisms and theseisolates have indeed been found to be highly abundant inmarine viral metagenomes (268 269) New approaches tointerpret the results include clustering the proteins independ-ent of known proteins to compare samples to each other andtry to find environmental factors driving viral communitychanges (270) Such analysis of a large global data set (TaraOceans expedition) has shown that extensive sampling hascome close to reaching the total diversity in tropical and tem-perate waters of such viral protein clusters (which essentiallyrepresent various viral protein motifs not all viral proteintypes) and that viruses appear to be directionally dispersed
422-16 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquodownstreamrdquo in major ocean currents as one may expectfrom first principles but also supporting the ldquoseed bankrdquohypothesis of viral biogeography (271)
Viruses and Host DiversityViruses are themselves thought to be instrumental in drivingincreased diversity of their microbial hosts via a hypothesizedprocess often called ldquokill the winnerrdquo This is because viralinfection is host-specific and density-dependent the latterbecause viruses diffuse from host to host so an abundanthost is more likely to pass on infection than a rare oneThis means that if an organism becomes abundant andblooms winning the competition for resources it becomesmore susceptible to a viral epidemic This would benefit therarer organisms and thus help foster diversity (223 272273) However bloom scenarios as described above involvesystems far from steady state and the formal theory of Thing-stad and Lignell (274) has interesting steady-state solutionswhere several viruses infect several hosts stably over time(via trade-offs between growth rates and viral susceptibility)which may occur at the strain or species level (274) There issome experimental evidence that viruses have effects on nat-ural marine microbial community composition weaker thanthe kill the winner (bloom version) hypothesis would sug-gest (275 276) so something resembling the steady-statecoexistence described in the model may in fact be commonThere also appear to be processes that foster coexistencebetween viruses and hosts but the mechanisms are largelyspeculative (223 277 278)
Viruses may also be directly involved in host genetic diver-sity because they can be the agents of genetic exchangebetween microorganisms (223 273) This often involvesthe viral lifestyle known as lysogeny whereby viruses survivewithin host cells as DNA only integrated into the host chro-mosome and being reproduced each time the host divides Ahost harboring such a genome is called a lysogen becauseunder conditions of stress to the host cell a genetic switchmay cause the viral genome to initiate the lytic process pro-ducing many progeny viruses and bursting from the hostLysogeny is a very common property occurring in a signifi-cant part of the bacterial community (279 280) althoughthe incidence of induction of the lytic phase in nature is appa-rently low (281) Overall lysogeny is poorly understood butthought to have both positive and negative impacts on themicrobial community (282) Recent results suggest that ben-efits of being lysogenic in highly seasonal polar seas leads tofundamental difference between polar and other marine viralcommunities (283)
Viruses and the Microbial LoopAs part of the food web viruses occupy a unique positionThey infect host cells that are mostly thought to be heterotro-phic bacteria and by doing so they typically burst the hosts torelease progeny viruses and cellular debris But what is the fateof this material Viruses themselves do not last indefinitelyand a simple steady-state assumption implies that from eachburst of viruses (typically 20ndash100 per lytic event) only onesuccessfully infects another cell The rest are inactivatedand broken down by sunlight (UV and visible exposure)and enzymatic attack (284) or consumed byminute phagotro-phic protists (285) thus reentering the food web as substratefor bacteria or food for protistan consumers Experiments incontrolled laboratory systems and field studies with radioac-tively labeled viral lysis products have supported the conclu-sion that most of the organic matter released by the viralinfection is either taken up by bacteria or respired (286 287)
Modeling this process as part of the microbial loop showsthat viral lysis represent a sort of side loop that has the neteffect of remineralizing a significant amount of the carbonand nutrients that enter the bacteria-protist part of themicro-bial loop (Fig 7) A theoretical numerical steady-state modelcomparing a systemwith no viral activity to onewhere virusesare responsible for 50 of bacterial mortality showed that thesystem with viruses had 33 more bacterial production andrespiration than the virus-free system implying that theviruses had the effect of permitting the bacteria to processmore of the primary production than they would otherwise(223) Although 50 is a high number unlikely to be com-mon in the sea this model nevertheless illustrates that virusescan reduce the amount of energy reaching higher trophic lev-els The implication is that viruses lead to increased bacterialactivity at the expense of the larger organisms
MAJOR ENVIRONMENTAL CONTROLSLight Temperature and PressureTemperature has an important potential influence on bio-chemical reactions and therefore on biological processesin general Most ocean waters fall in the range of ndash2degC to30degC with obvious exceptions in hydrothermally heatedareas Temperature has long been known to be a regulatingfactor for the growth of heterotrophic microbes
In temperate waters it has been established that microbialactivity is generally much higher in warm summer waters thanin winter (55) The relationship is not simple howeverbecause multiple factors act at the same time Some contro-versy still exists regarding the highest and lowest extremesfor marine bacterial growth although there is broad agree-ment that bacteria grow gt100degC at hydrothermal vents andltminus5degC in sea ice brines Pomeroy et al (57) noted the inter-esting observation that bacteria seem particularly inhibitednear the freezing point of seawater (ca ndash22degC) comparedto eukaryotic phytoplankton This effect results in polarspring phytoplankton blooms that accumulate organic car-bon in advance of the response of the bacterial communityand development of the microbial loop and perhaps lead toenhanced benthic-pelagic coupling (288)
The relationship between temperature and the growth rateof marine phytoplankton was described broadly in a now
FIGURE 7 Modification of the microbial loop concept that incor-porates the functional role of viruses Export can be via predation orsinking From Fuhrman (223)doi1011289781555818821ch422f7
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-17
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
classic paper by Eppley (58) and later Goldman and Carpen-ter (289) Temperature was shown to exert a strong and directeffect on themaximal growth rates of these species withmax-imal intrinsic growth rates at 0degC generally lt1 divisiondaywhile growth rates at 30degC may be gt4day These relation-ships indicate the maximal rate that might be attained byphytoplankton at these temperatures but they do not takeother factors into account (nutrients light) While warmtemperature ostensibly allows more rapid growth it createshydrographic conditions that typically give rise to nutrientlimitation of algal growth Thus some of the coldest watersin nature witness some of the most massive phytoplanktonblooms (290) while warm oceanic gyres represent some ofthe most oligotrophic areas of the ocean The multiple andoften conflicting effects of temperature on the growth of nat-ural phytoplankton assemblages limit the accuracy of presentpredictions regarding how primary producers will respond toclimate change (291)
An analysis of the effect of temperature on the growth ofheterotrophic protists has indicated that the response is qual-itatively similar to that of phytoplankton but with a twistThe growth rates of at least some protozoa can exceed thoseof phytoplankton at warmer environmental temperaturesbut the opposite effect is apparent at very low environmentaltemperature This differential effect of temperature on thegrowth of phototrophic and heterotrophic protists was basedon a large meta-analysis of published protistan growth (59)That analysis demonstrated that the maximal growth ratesattained by phototrophic protists could exceed the maximalgrowth rates attained by heterotrophic protists (all otherpotential growth-limiting factors not considered) Thereforeprotozoan growth rates may be constrained to a greater degreeat low environmental temperature than rates for phytoplank-ton If so then phytoplankton bloomsmay get a head start ongrazers during spring in polar ecosystems This scenario is con-sistent with information on seasonal biomass changes andmicrozooplankton herbivory in the Ross Sea Antarctica(292 293) but there are still too few data to fully vet thishypothesis
The importance of high pressure on bacterial growthgained considerable attention in the late 1960s when thedeep sea submersible Alvin was accidentally lost overboardwith its hatch openWhile no lives were lost in this accidentsome workmenrsquos lunches sank to the bottom (sim1500 m)inside the submersible Alvin was recovered after 10 monthsand interestingly there was a waterlogged lunch containingapples bologna sandwiches and broken vacuum bottleswith broth that all appeared hardly degraded and tasted palat-able Yet when placed in a refrigerator on the ship these itemsdegraded relatively quickly Initially pressure was thought tobe the preserving factor as the sea floor temperature was sim-ilar to the refrigerator temperature and the only major differ-ence would be pressure (294) Following this observationa series of experiments to measure degradation of variousorganic materials left in the deep sea for extended periodsindicated that degradationwas typically significantly reducedimplying that pressure reduces the degradation rates (295)Nonetheless changes in the protozoan community of naturaldetrital material sinking to the deep ocean floor indicate thatthe microbial community can respond relatively quickly insome situations (296) Deep sea microbiology has advancedconsiderably yet it is still difficult to interpret results withrespect to actual in situ rates of naturally occurring organicmatter Deep sea bacteria adapted to high pressures havebeen isolated that are barophilic (also called piezophilic)meaning that they prefer high pressures and have reduced
activity at lower pressures whereas others are barotolerant(piezotolerant) tolerating but not preferring high pressuresfor example see (297 298)
Few data are available on barotolerantbarophilic marineprotozoa Protozoa certainly exist and grow at great oceanicdepths but measurements of in situ growth rates for these spe-cies do not yet exist Measurable protozoan numbers havebeen documented in the deep-sea sediments for more than30 years (299 300) and viable protozoa have occasionallybeen cultured from these environments (301ndash305) butvery few direct measurements of the activities of these speciesin situ have been reported (306) A few protozoa have beenisolated that will grow at high pressure (302 303 307) andprotists that appear to be unique to the deep ocean havebeen observed either directly or through the analysis ofDNA sequences (308ndash311) but possibly the best direct evi-dence that protozoan activity takes place at the high pressurescharacteristics of the deep sea are experimental and observa-tional work noting the stimulatory effect that detrital deposi-tion has on some components of the protozoan community(296 312) These observations indicate a diverse and activeprotozoan fauna of the deep ocean although their biogeo-chemical significance is largely uncharacterized at this time
Dissolved and Particulate Organic MatterBacteria and archaea are thought to be by far the most impor-tant organisms with respect to the processing of dissolvedorganic matter (DOM) and nonliving particulate organicmatter (POM also called detritus) in the ocean While theremay be some uptake of DOM by protists particularly forgrowth factors needed in trace amounts such as vitamins(313) the bulk of this material is probably utilized by bacteriaand archaea (314) including the smallest cyanobacteriumProchlorococcus (315 316) which thus may be considered amixotroph Due to their small size bacteria have extremelyhigh surfacevolume ratios and combined with their over-whelmingly high abundance an extremely high integratedsurface area Protozoa tend to obtain the organic materialsthat they require for growth from their prey rather thanthrough the uptake of DOM Overall protozoa tend to besources of dissolved and detrital organic substances throughthe excretion of unassimilated prey biomass in expelledfood vacuoles
Particulate organic matter is not directly available as sub-strate to bacteria These substances must first be reduced tosmall molecules that can be transported into the cell Thisis accomplished by the production of extracellular enzymes(note that few if any large polymers are directly taken up bybacteria with the possible exception of DNA) Hydrolyticenzymes produced by bacteria (and almost certainly archaea)break down polymers like proteins polysaccharides andnucleic acids As with DOM POM is composed of a complexmixture of compounds that vary in their susceptibility to bac-terial degradation and utilization
Particulate material in the water column serves not only asbacterial substrate but also as substratum POM occurs in thewater column across a huge size spectrum from micrometersup to some detrital aggregates more than 1 m in diameter(317) Much of this particulate material is in a constant stateof flux with colloidal material constantly coalescing andaggregating to form new or larger particles (318) as microbialdegradation acts simultaneously to remineralize this materialDetrital particles that attain macroscopic size either by directformation (317 319 320) or via accretion and aggregation(321) and are often called marine snow or macroaggregatesMarine snow particles are readily colonized by bacteria and
422-18 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
many other microorganisms and their abundances on theseaggregates can be orders of magnitude greater than abundan-ces in an equivalent volume of the water surrounding theaggregates (320 322ndash324) and their diversity and highlyconcentrated abundances on detrital aggregates implies thatthere may be stronger trophic coupling and efficient energyelemental cycling relative to these processes among free-living microorganisms in the surrounding waters This specu-lation has resulted in a number of formalizations of the poten-tial impact of aggregate microbiology on biogeochemicalprocesses in the ocean (325 326) Most definitions of bio-films include aggregated particulate and these particles areoften studied within the biofilm conceptual model (see chap-ter by Lawrence et al in this section)
Inorganic Nutrients
Macronutrients (N P)Virtually all heterotrophic organisms in the ocean contributeto the pool of available macronutrients via the excretion ofmetabolic wastes and protozoa contribute significantly tothis process Macronutrients are often in excess within proto-zoan prey relative to the consumersrsquo needs because much ofthe prey carbon is respired to produce energy for metabolismand growth ldquoExcessrdquo nutrients are eliminated as eitherorganic compounds or more commonly as ammonium (forN) and phosphate (for P) The importance of the match(or mismatch) between predator and prey stoichiometry hasbeen demonstrated experimentally and repeatedly in marineand freshwater ecosystems and has been elevated to a funda-mental ecological principle governing elemental and energyflow in aquatic food webs (327) Based on this reasoningCaron et al (328) concluded that bacterivorous protozoaplay an important role in nutrient remineralization andrelease in the ocean Bacterial biomass is typically rich in Nand P relative to other organisms so bacterial predators expe-rience large excesses of these elements relative to their growthneedsMoreover bacterivorous protozoa should be dispropor-tionately important in nutrient remineralization amongsingle-celled eukaryotes because as the smallest protozoa inaquatic ecosystems they have high weight-specific metabolicrates (329) Thus both stoichiometric and allometric rela-tionships implicate small bacterivorous protozoa as impor-tant sources of remineralized nutrients in the plankton
The reasoning also holds true for bacterial utilization oforganic compounds and it has led to some interestingfindings and conclusions regarding the role of bacteria innutrient cycling in the ocean Bacteria are important sourcesremineralized nutrients when they are consuming N- orP-rich substrate Contrary to this traditional view howeverheterotrophic bacteria are often strong competitors for inor-ganic nutrients under many situations largely because bacte-rial biomass is richer in N and P than much of the organicmatter they consume in nature so they require additional Nand P to produce their biomass (summarized by [330]) Exper-imental studies have demonstrated the uptake of ammoniumby bacteria other than cyanobacteria using stable isotope 15N(331) and also the short-lived radioisotope 13N (332) oftenfinding that bacteria may be responsible for one third of the Nor P uptake Additionally there is evidence that bacteria insome oceans like the Sargasso Sea are limited by phosphorus(333 334) Growth limitation of bacteria by phosphorusappears to be a common phenomenon of freshwater ecosys-tems (335 336)
N2 fixation is restricted to prokaryotes and in the oceanwater column it was thought for many years to be done
primarily byTrichodesmium a warm-water colonial cyanobac-terium that blooms sporadically andRichelia a cyanobacterialsymbiont that lives within certain diatoms (337) Some uni-cellular cyanobacteria also contribute significantly to globalN2 fixation (338) and molecular biological data suggestthat a variety of other bacteriamdashnot phytoplanktonmdashmayalso be fixing nitrogen in seawater (339) An exciting recentdiscovery in this field is that a bacterial nitrogen fixer origi-nally known only from nifH sequences UCYN-A has hadits genome sequenced by parallel pyrosequencing and wasfound be a cyanobacterium (Candidatus Atelocyanobacteriumthalassa) lacking the apparatus to generate oxygen as well asother pathways (340) Further study showed it to be symbioticwith a marine alga (341)
Micronutrients (Trace Metals Growth Factors)Bacterial growth may often be limited by macronutrients asdescribed and sometimes by trace nutrients particularlyiron that occur at very low concentrations in seawater(342 343) The importance of Fe as an element limiting pri-mary production in some oceanic regions has come underclose scrutiny and extensive experimental investigation inrecent years Many bacteria have special uptake mechanismsto utilize extremely low concentrations of iron includingsiderophores (released compounds that bind iron and arethen specifically taken up) Even if bacteria are capable ofgrowth at low Fe concentrations growth may be less efficientKirchman et al (344) found that Fe limitation stronglyreduced bacterial growth efficiency in a cultured marine bac-terium (ca 50 efficiency Fe replete but lt10 when Fedeplete) Thus the effects may be complex Marine N2 fixa-tion also may be limited by the availability of Fe becausenitrogen fixers have high Fe requirements
Protozoa tend to be a source rather than a sink for Fe in theocean analogous to their role in macronutrient remineraliza-tion Protozoan grazing activity has been shown to releaseiron from bacterial biomass thereby relieving Fe limitationin co-occurring phytoplankton (345) a mechanistic demon-stration of the process using protozoa growing on Fe-repletebacteria However it has also been reported that Fe-limitedbacteria may contain insufficient Fe for the bacterivorous pro-tozoa that consume them thus leading to Fe limitation in theprotozoan as well (346)
OxygenHypoxic and anoxic regions of the ocean are expected to sig-nificantly expand in the coming decades as a consequence ofchanges in ocean stratification resulting from global climatechange (347) As oxygen becomes depleted bacteria havethe capability to utilize a series of alternate electron acceptorsCompositional changes in the bacterial assemblage accom-pany these changes in metabolic activity but are beyondthe scope of this chapter Many of the studies on benthicmicrobiology have focused on redox reactions that occurthere Oxygen is usually consumed by sediment microbialactivity much faster than the rate at which it can be replacedby diffusion so most sediments especially organically richones are anaerobic below a few mm (fine-grained muds) orcm (coarser sediments) from the sedimentwater interfaceexcept where animals ventilate the benthos through tubesWhen oxygen is absent bacteria and archaea use alternateelectron acceptors such as nitrate nitrite oxidized Fe orMn organic matter or sulfate The use of nitrate or nitriteas an electron acceptor with concomitant production of N2
gas (denitrification) results in net loss of biologically availablenitrogen Bacteria that perform this reaction are typically
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-19
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquofacultativerdquo organisms that can switch from oxygen to nitrateor nitrite Organic matter may be reduced by fermentationreactions in which part of an organic substrate is oxidizedwhile another is reduced fermenter organisms are also typi-cally facultative Sulfate is a particularly important alternateelectron acceptor in marine systems because it occurs as thesecond most abundant anion in seawater (behind chloride)typically 28 mM Its use as an electron acceptor leads to theproduction of elemental sulfur or sulfide The use of sulfateas an electron acceptor yields much less energy than oxygenor nitrate and sulfate-reducing bacteria are strict anaerobesunable to use oxygen (often strongly inhibited by oxygen)Methane can be produced by methanogenic archaea whichare all strict anaerobes using CO2 as an electron acceptor(see Findlay and Battin this section for further discussion)
Reduced compounds such as sulfide are produced by theuse of these alternate electron acceptors and they diffuse awayfrom their source When they diffuse to environments with astrong oxidant like oxygen they may be oxidized by chemo-trophic bacteria in energy-yielding reactions Similarlyammonium can be oxidized to nitrite and then nitrate by che-motrophic nitrifying bacteria and the nitrate can diffuse intoanaerobic layers and become denitrified to N2 Such coupledoxidation-reduction reactions can lead to accelerated biogeo-chemical cycling particularly in sediments
It was originally believed that relatively few free-livingspecies of protozoa inhabit anaerobic and hypoxic marineenvironments This view has changed based on (largely)molecular analyses of some of these environments (308348ndash351) A significant number of previously undescribedeukaryotic phylotypes have been observed in anoxic environ-ments High protozoan biodiversity in the absence of molec-ular oxygen is not completely unexpected because otheranaerobic environments such as some gut environments(eg cockroach guts and cattle rumen) have notoriouslyabundant and diverse protozoan fauna but marine anaerobicenvironments have been difficult to access and sample prop-erly Many anaerobic protozoa exist together with symbioticbacteria (352 353) or possess unique organelles such as thehydrogenosome (354)
MAJOR BIOTIC CONTROLSThe Trophic Activities of Protozoa Predation andTop-Down ControlsPredator-prey relationships are fundamental interactionswithin biological communities linking production of bio-mass on the one hand with removal on the other hand Estab-lishing the trophic relationships among microorganisms andquantifying the rates at which they take place constitutemajor endeavors within marine microbial ecology As notedpreviously protozoa consume a wide variety of prokaryoticand protistan prey Numerous laboratory and field measure-ments have been made throughout the past quarter centuryto attempt to quantify the rates of bacterial and phytoplank-ton consumption The methodology for accomplishing thesestudies has varied and a number of approaches are presentlyemployed
Methodologies for estimating microbial grazing are diversebut generally fall into one of two experimental approachesperturbation experiments or tracer experiments Perturbationexperiments (size fractionation metabolic inhibitors anddilution technique) rely on manipulation of the consumeror prey assemblages in somemanner such that the effect is evi-dent from changes in the abundance of the prey population
during incubations relative to control treatments that receiveno manipulation Tracer experiments (radioactively labeledprey fluorescently labeled prey stable isotopendashlabeled pray)attempt to use labeled prey or a prey proxy to quantitativelyfollow the movement of prey biomass into consumers Forsummaries of these methods see (74 355)
Perturbation experiments have entailed a variety of mech-anisms to ldquodecouplerdquo predators from prey in natural microbialcommunities For example size fractionation uses various sizefilters to remove an entire size class of grazers from awater sam-ple leaving the prey to grow in the relative absence of thosepredators during subsequent incubations Unfiltered samples(controls) indicate the growth of prey in the presence of pre-dation Predation impact is obtained by the difference ingrowth of the prey between these treatments This methodhas been used to examine predation on cyanobacteria heter-otrophic bacteria and minute phototrophic eukaryotesAlternatively metabolic inhibitors have been employed tohalt predation (or in some cases prokaryote growth) insteadof physical removal by filtration
The dilution technique (356) has become a commonmethod for examining grazing mortality of phytoplanktonby microzooplankton (which is comprised predominantly ofprotozoa) This method relies on the dilution of herbivorouszooplankton in a series of subsamples (and concomitantmeasurements of phytoplankton growth in each subsample)to estimate the grazing mortality of the herbivores
Tracer experiments have used both radioactively labeledprey and fluorescently labeled prey to examine predation onbacteria and phytoplankton Radioactive labeling has beenaccomplished using 14C- or 3H-labeled organic compounds(for bacteria) or 14C-bicarbonate (for phytoplankton) Theuse of radioisotope approaches has diminished with improve-ments in the number and types of fluorescent compoundsavailable for this work Fluorescently labeled prey havebeen labeled with a variety of compounds that allow theprey to be observed readily (even inside predators) by epi-fluorescencemicroscopy (357 358) Alternatively a decreasein the total number of fluorescent prey in a sample indicatesthe rate of removal of the prey assemblage
Both perturbation experiments and tracer experimentsdescribed have inherent advantages and disadvantages Forexample perturbation experiments are fairly straightforwardto conduct but they may introduce artifact by removingpotentially important sources of nutrients or organic com-pounds released by predators (359) On the other hand thedilution technique is very labor-intensive but one advantageof this method is that it provides the opportunity to determineboth phytoplankton growth and grazing mortality in a singleprocedure Most tracer experiments are fairly easy to conductand generally do not perturb the food web being studied butfeeding selectivity for or against the tracer (relative to naturalprey) may produce spurious results In addition some of thesemethods provide a measurement of the activity of the entiregrazer assemblage and others are applicable for examiningthe activity of specific taxa For example size fractionationapproaches examine the effect of the removal of an entiresize class of organisms (a community-levelrdquo measurement)whereas fluorescently labeled prey can be used to examinethe ingestion rate of a particular species by observing theuptake of labeled prey into that species (a species-levelmeasurement)
Predation rates of numerous marine protozoa have beenexamined in the laboratory From these experiments a gen-eral knowledge of the rates of consumption by these specieshas been accumulated Feeding in most protozoa shows a
422-20 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
functional response to the availability of prey that is similar tothe response of many organisms (although there are a greatdeal of species-specific differences) That is ingestion rateincreases rapidly as prey abundance increases until a maximalingestion rate is attained (360)Manyof these curves have theform of a Type II functional response curve (361)
Based on the features of these curves it is clear that proto-zoa have the ability to ingest substantial numbers of prokar-yote and eukaryote prey in natural ecosystems Somebacterivorous flagellates can consume bacteria (and cyano-bacteria) at rates up to 100 s per individual per hour whereassome bacterivorous ciliates can consume thousands of bacte-ria per individual per hour Rates of consumption of eukary-otic phytoplankton by herbivorous ciliates and largeflagellates are considerably more variable and highly depend-ent of the feeding mechanism (eg filtration in ciliates versuspseudopodial net in dinoflagellates) and the size of the prey
These lab studies demonstrate that rarely are prey suffi-ciently abundant in nature to support the maximal ingestionrates of which many protozoa are capable Nevertheless pre-dation rates of protozoa in nature are sufficient to constitute amajor factor controlling the standing stocks of bacteria andphytoplankton (63 180 184 355) For example these sum-maries indicate that on average protozoa consume 20ndash80ofthe standing stock of phytoplankton (measured as chloro-phyll) in the world ocean and 60ndash75 of daily primary pro-duction a departure from the classical view of copepods as theprimary consumers of phytoplankton This large role of her-bivorous protozoa infers a large role for copepods as consumersof planktonic protozoa (185) Similarly the role of protozoaas major consumers of bacterial biomass is indicated by therelative balance between bacterial production on the onehand and bacterivory on the other (Fig 8) The approximateone-to-one balance between these measurements over a largenumber of studies implies that protozoan bacterivory must bea major factor determining the fate of bacteria in marine (andfreshwater) plankton ecosystems
It is interesting to note that some types of bacteria includ-ing marine Bdellovibrio are known to prey on other marine
bacteria (362) At the time of this writing the importanceof bacterial predation by other bacteria within the contextof oceanographic processes is not known and presumed tobe small While Bdellovibrio may be cultivated from seawaterit can also be found among sequences from cold sediments(363) and the water column Bdellovibrio happens to be thebest studied predatory bacterium but there are no doubtothers that we simply have not yet identified (364)
Species-Specific Interactions (MutualismParasitism Commensalisms)Interactions between marine microorganisms such as compe-tition allelopathy and symbiosis are fundamental aspectscontrolling community composition and activity but atpresent most are poorly characterized in natural marine sys-tems Bacteria must compete for dissolved and particulatecompounds with other bacteria andwith protistan taxa whilephagotrophic protists must compete with each other andmetazoa for available prey These interactions manifest them-selves in many forms including true competition for availableresources resource partitioning such that some taxa specializeon the utilization of certain substrates or prey while othersspecialize on different ones (reducing competition) or eventhe cooperative use of compounds (eg one bacteriumrsquoswaste products being utilized as substrate by another bacte-rium) These metabolic consortia are probably common inthe ocean but can be difficult to identify and characterizeConsortia of bacteria working together to perform complexreactions especially in anaerobic systems are well known inmicrobiology
Consortia in which there is a very high degree of integra-tion between species (ie symbiosis) are very common in theocean These associations may be beneficial to both partners(mutualism) beneficial to one part and inconsequential forthe other (commensalism) or beneficial to one part and det-rimental to the other (parasitism) Many highly visible andwell-characterized marine mutualisms exist between chemo-synthetic bacteria and animals such as hydrothermal ventor methane seep tubeworms clams mussels or other inverte-brates (365) and also between luminescent bacteria and fishor squids (366) Similarly many protozoa form mutualisticassociations with photosynthetic protists (367) The mostconspicuous of these associations involved large amoeboidprotozoa of oceanic ecosystems (planktonic foraminiferanspolycystine radiolaria and acantharia) which contain largenumbers of intracellular algae (74) but associations betweensome heterotrophic dinoflagellates and cyanobacteria are alsocommon (368) Studies have indicated that efficient nutrientcycling in these protozoan-algal associations confers an eco-logical advantage for both partners in surfacewaters of the oli-gotrophic ocean
Parasitism is also a common symbiotic relationship amongmarine protozoa Protozoan parasites have been reported for adiverse array of protists and animals in the ocean Heterotro-phic dinoflagellates and presently uncultured alveolate cladesrelated to alveolates appear to be particularly successful atevolving thismode of existence (369ndash371) In a common par-asitic group Amoebophrya spp the parasite invades photo-synthetic dinoflagellates where it undergoes reproductioneventually lysing the host and releasing dozens of infectiveswarmers The parasitic protist Pirsonia spp is a parasite ofdiatoms (372) The overall importance of these parasitismsin the population dynamics of their hosts is not clearly knownbut may constitute a significant source of mortality for somespecies (373 374) The frequency and diversity of ldquonewrdquo
FIGURE 8 Meta-analysis of experimental studies that have com-pared rates of bacterial productivity (converted to population growthrate units of dminus1) with rates of bacterivory by phagotrophic protists(expressed as equivalent units of bacterial growth rate dminus1) Thesolid line indicates where measurements of bacterial removal by graz-ers were equal tomeasurements of bacterial production Data redrawnfrom Sanders et al (63) doi1011289781555818821ch422f8
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-21
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-
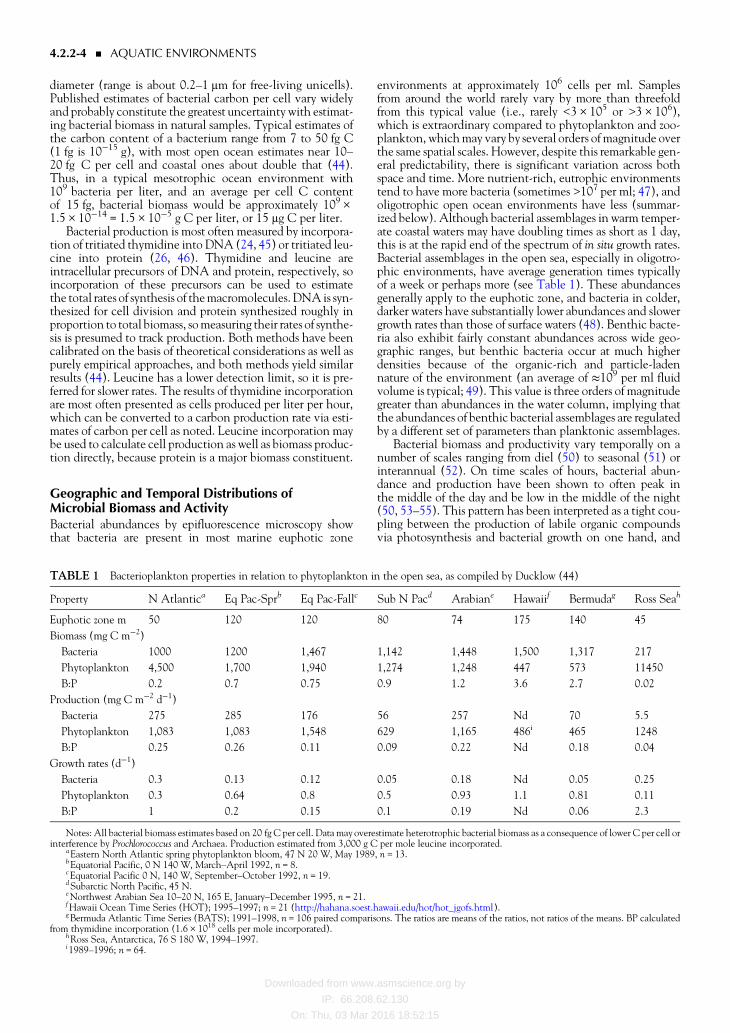
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
diameter (range is about 02ndash1 μm for free-living unicells)Published estimates of bacterial carbon per cell vary widelyand probably constitute the greatest uncertainty with estimat-ing bacterial biomass in natural samples Typical estimates ofthe carbon content of a bacterium range from 7 to 50 fg C(1 fg is 10minus15 g) with most open ocean estimates near 10ndash20 fg C per cell and coastal ones about double that (44)Thus in a typical mesotrophic ocean environment with109 bacteria per liter and an average per cell C contentof 15 fg bacterial biomass would be approximately 109 times15 times 10minus14 = 15 times 10minus5 g C per liter or 15 μg C per liter
Bacterial production is most often measured by incorpora-tion of tritiated thymidine into DNA (24 45) or tritiated leu-cine into protein (26 46) Thymidine and leucine areintracellular precursors of DNA and protein respectively soincorporation of these precursors can be used to estimatethe total rates of synthesis of themacromolecules DNA is syn-thesized for cell division and protein synthesized roughly inproportion to total biomass someasuring their rates of synthe-sis is presumed to track production Both methods have beencalibrated on the basis of theoretical considerations as well aspurely empirical approaches and both methods yield similarresults (44) Leucine has a lower detection limit so it is pre-ferred for slower rates The results of thymidine incorporationare most often presented as cells produced per liter per hourwhich can be converted to a carbon production rate via esti-mates of carbon per cell as noted Leucine incorporation maybe used to calculate cell production as well as biomass produc-tion directly because protein is a major biomass constituent
Geographic and Temporal Distributions ofMicrobial Biomass and ActivityBacterial abundances by epifluorescence microscopy showthat bacteria are present in most marine euphotic zone
environments at approximately 106 cells per ml Samplesfrom around the world rarely vary by more than threefoldfrom this typical value (ie rarely lt3 times 105 or gt3 times 106)which is extraordinary compared to phytoplankton and zoo-plankton whichmay vary by several orders of magnitude overthe same spatial scales However despite this remarkable gen-eral predictability there is significant variation across bothspace and time More nutrient-rich eutrophic environmentstend to have more bacteria (sometimes gt107 per ml 47) andoligotrophic open ocean environments have less (summar-ized below) Although bacterial assemblages in warm temper-ate coastal waters may have doubling times as short as 1 daythis is at the rapid end of the spectrum of in situ growth ratesBacterial assemblages in the open sea especially in oligotro-phic environments have average generation times typicallyof a week or perhaps more (see Table 1) These abundancesgenerally apply to the euphotic zone and bacteria in colderdarker waters have substantially lower abundances and slowergrowth rates than those of surface waters (48) Benthic bacte-ria also exhibit fairly constant abundances across wide geo-graphic ranges but benthic bacteria occur at much higherdensities because of the organic-rich and particle-ladennature of the environment (an average of asymp109 per ml fluidvolume is typical 49) This value is three orders of magnitudegreater than abundances in the water column implying thatthe abundances of benthic bacterial assemblages are regulatedby a different set of parameters than planktonic assemblages
Bacterial biomass and productivity vary temporally on anumber of scales ranging from diel (50) to seasonal (51) orinterannual (52) On time scales of hours bacterial abun-dance and production have been shown to often peak inthe middle of the day and be low in the middle of the night(50 53ndash55) This pattern has been interpreted as a tight cou-pling between the production of labile organic compoundsvia photosynthesis and bacterial growth on one hand and
TABLE 1 Bacterioplankton properties in relation to phytoplankton in the open sea as compiled by Ducklow (44)
Property N Atlantica Eq Pac-Sprb Eq Pac-Fallc Sub N Pacd Arabiane Hawaiif Bermudag Ross Seah
Euphotic zone m 50 120 120 80 74 175 140 45
Biomass (mg C mminus2)
Bacteria 1000 1200 1467 1142 1448 1500 1317 217
Phytoplankton 4500 1700 1940 1274 1248 447 573 11450
BP 02 07 075 09 12 36 27 002
Production (mg C mminus2 dminus1)
Bacteria 275 285 176 56 257 Nd 70 55
Phytoplankton 1083 1083 1548 629 1165 486i 465 1248
BP 025 026 011 009 022 Nd 018 004
Growth rates (dminus1)
Bacteria 03 013 012 005 018 Nd 005 025
Phytoplankton 03 064 08 05 093 11 081 011
BP 1 02 015 01 019 Nd 006 23
Notes All bacterial biomass estimates based on 20 fg C per cell Data may overestimate heterotrophic bacterial biomass as a consequence of lower C per cell orinterference by Prochlorococcus and Archaea Production estimated from 3000 g C per mole leucine incorporated
aEastern North Atlantic spring phytoplankton bloom 47 N 20 W May 1989 n = 13bEquatorial Pacific 0 N 140 W MarchndashApril 1992 n = 8cEquatorial Pacific 0 N 140 W SeptemberndashOctober 1992 n = 19dSubarctic North Pacific 45 NeNorthwest Arabian Sea 10ndash20 N 165 E JanuaryndashDecember 1995 n = 21fHawaii Ocean Time Series (HOT) 1995ndash1997 n = 21 (httphahanasoesthawaiieduhothot_jgofshtml)gBermuda Atlantic Time Series (BATS) 1991ndash1998 n = 106 paired comparisons The ratios are means of the ratios not ratios of the means BP calculated
from thymidine incorporation (16 times 1018 cells per mole incorporated)hRoss Sea Antarctica 76 S 180 W 1994ndash1997i1989ndash1996 n = 64
422-4 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
bacterial mortality via grazers or viral lysis on the other handIt is consistent with some measurements of extremely rapidturnover sometimes several times per hour in rich coastalwaters of labile dissolved organic compounds such as dis-solved free amino acids (56) and might also indicate greaterpredation pressure by protozoa during the night On longertime scales of weeks to months bacteria show distinct sea-sonal patterns For example in temperate coastal waters bac-terial biomass and production increase considerably insummer months compared with winter However bacteriado not typically show a significant increase during earlyldquospringrdquo blooms in temperate waters when water is still verycold (51) It has been hypothesized that this phenomenonis the result of the suppression of the rate of substrate uptakeby temperate bacterial assemblages at low temperature (57)However while temperature probably has the effect of settinga limit on maximal growth rates (as for phytoplankton [58]and protozoa [59]) temperature alone does not appear to bethe main factor controlling growth of marine bacteria undermost circumstances (see ldquoLight Temperature and Pressurerdquo)
It would be overly simplistic to think that all the bacteriaand archaea in a sample or habitat have the same level ofactivity per cell but it is also easy to think of measured activ-ities as characteristic of all members of a microbial assem-blage So the question arises are most of the cells active ata similar level or are some hyperactive while others are com-pletely dead or moribund This question has been addressedseveral ways includingmicroautoradiography selective stain-ing ldquodirect viable countsrdquo (where nutrients are added to seewhat part of the community grows) and in situ hybridization(eg 60) Based on these contrasting approaches it appearsthat a continuum of activity exists within bacterial assemb-lages from truly dead (cannot be revived) to extremely activeA reasonable interpretation of the existing data is that undertypical conditions a small percentage of the marine bacterialcells perhaps 10ndash20 are generally inactive or dead the plu-rality or majority of cells perhaps 25ndash75 are intact andhave some moderate level of activity and a small percentageperhaps 5ndash20 are highly active It is useful to consider thisspectrum conceptually and numerically when modelingmicrobial processes
Comparisons of bacterial and phytoplankton biomasswithin planktonic ecosystems show that these are positivelycorrelated across broad scales Analyses of marine and fresh-water samples from several studies (61 62) have shown thatbacterial abundance increases with chlorophyll concentra-tion at least at the level of a log-log relationship Similarlybacterial abundances and the abundances of small protozoacorrelate over broad spatial and temporal scales (63) Theserelationships are sensible in that on the largest scale primaryproduction is the source of organic material that fuels hetero-trophic bacterial activity and bacteria constitute the prey ofmany small protozoa Individual data sets also have sometimesshown strong correlations between bacterial abundance andchlorophyll (eg 45) but variability in this relationshipover short temporal or spatial scales is to be expected It wouldpresumably be a consequence of rapid short-term changes inthe rate of substrate supply as well as the normal oscillatorynature of predator-prey relationships between bacteria andtheir consumers
Interestingly the extrapolation of the positive log-log rela-tionship between bacterial and phytoplankton biomassto environments with very low chlorophyll concentrations(eg ultra-oligotrophic oceans) indicates that bacterial bio-mass may exceed phytoplankton biomass in these situationsWhile this conclusion is obvious for the deep sea where there
is no photosynthesis this situation is also often true in oligo-trophic surface waters Measurements made in oligotrophicwaters bear out the high bacterial contribution to totalbiomass (64 65) Moreover Cho and Azam (66) confirmeda linear relationship between the log of chlorophyll and logof bacterial abundance but only at chlorophyll concen-trations above approximately 05 μg per liter Below thatconcentration bacterial abundance did not correlate signifi-cantly with chlorophyll It should be noted that subsequentanalyses have revealed that early epifluorescence measure-ments of bacterial biomass included the common cyanobacte-rium Prochlorococcus which can make up to 20 of totalbacterial numbers (67) Nonetheless heterotrophic bacterialbiomass is a major fraction of the living biomass of all plank-tonic ecosystems
The geographical and temporal distributions of marineprotozoa are much more varied than those of the bacteriaAs an all-inclusive group protozoa generally occur in plank-tonic ecosystems at abundances ranging from 10 s to 1000 sper ml Abundances in benthic ecosystems can be one tothree orders of magnitude higher commensurate with thehigher abundances of bacteria in those ecosystems Howeverit is important to remember that like the term ldquolsquobacteriardquo theterm ldquoprotozoardquo is a rather artificial conglomeration of evolu-tionarily and ecologically divergent taxa (see ldquoThe Changingand Complex World of Eukaryote Phylogenyrdquo) Thus theabundances of specific lineages of bacteria or protozoa mayshow spatial (or temporal) variability that is considerablygreater than the variability characteristic of these overarchinggroupings
The Changing and Complex World ofEukaryote PhylogenyNot that long ago textbooks still divided eukaryotic organ-isms into four major kingdoms (Animalia Plantae Fungiand Protista) while prokaryotic organisms were placed intoa single kingdom theMonera (68)Within this scheme pro-tists (eukaryotic organisms that can exist as single cells) weredivided into two subkingdoms (algae and protozoa) based ontheir basic nutritional mode a carryover from the historicaldistinction between single cells with ldquoanimal-likerdquo or ldquoplant-likerdquo nutrition This distinction presupposed a basic evolu-tionary divergence among protists into species that retaineda heterotrophic phagocytotic mode of life (protozoa) andthose that abandoned phagocytosis for a photosyntheticmode of life (algae)Moreover the presenceabsence of chlor-oplasts was a feature that could be easily distinguished by earlymicroscopists
The five-kingdom classification system of Whittaker wasrecognized as an improvement over previous classificationschemes but it posed a number of problems relating toprotists For example the distinction between single-celledand multicellular eukaryotes was somewhat arbitrary Moreimportant the division of protists based on whether theywere heterotrophic or photosynthetic was clearly not anappropriate feature if the classification was to recapitulateevolutionary relationships We now know that chloroplastacquisition and loss has occurred several times in the bio-logical history of our planet (69) giving rise to some closelyrelated protistan taxa that differ largely in the presence orabsence of a chloroplast Further complicating the matterwithin many protistan lineages there are species that possesschloroplasts and carry out photosynthesis (phototrophy)but also possess the ability to ingest and digest prey (hetero-trophy 70ndash72) Some heterotrophic protists even ingest
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-5
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
phytoplankton prey and retain the chloroplasts of their prey ina functional state for a limited amount of time (kleptidoplas-tidy73) Various forms and degrees of mixotrophy (mixedphototrophic and heterotrophic nutrition) are commonamong a number of algalprotozoan lineages (74ndash77) UnderWhittakerrsquos scheme phytoplankton ecologists studying a lin-eage of microalgae might have had little familiarity withclosely related heterotrophic species while protozoologistsstudying a particular protozoan groupmight have known littleabout closely related photosynthetic species
One might expect given these caveats that the termsldquoalgaerdquo and ldquoprotozoardquo are no longer used In fact the termldquoprotozoardquo is still commonly used (especially by ecologists)to recognize those eukaryotic species that exist as single cellsand whose nutrition is dependent on the uptake of preformedorganic substances (primarily via prey ingestion) while pro-tists possessing chloroplasts are still commonly called ldquoalgaerdquoSimilarly although the term ldquoprotistrdquo has been abandoned asa kingdom designation it is still widely employed to describeeukaryotic species that are capable of existence as single cells(ie algae and protozoa) The term ldquophagotrophic protistrdquohas also gained popularity in recent years because it recognizesthat many protistan species are capable of phagocytosis eventhough they may also possess their own chloroplasts and thusare technically ldquoalgaerdquo
Despite the shortcomings of Whittakerrsquos scheme it domi-nated the hierarchical organization of life for approximately aquarter century During the past few decades however thissystem has given way to a new organizational scheme that rec-ognizes three domains of life (Archaea Bacteria Eukarya [orEucarya] 78 Fig 2 upper panel) and is based onwhat is pres-ently believed to reflect amore realistic view of the evolution-ary distances that have developed between organisms in theasymp4 billion years that life has existed on our planet Withinthe Eukarya of Woesersquos scheme hypotheses regarding thephylogeny of ldquoprotistsrdquo have changed continuously and rap-idly during the past two decades reflecting new insightsinto eukaryote evolution provided largely by DNA sequenceinformation (Fig 2 lower panel from [79])
The former protistan phyla ofWhittakerrsquos systemhavenowbeen dispersed among candidate ldquosupergroupsrdquo within thedomain Eukarya to better reflect hypothesized phylogeneticrelationships For example the dinoflagellates (which encom-pass phototrophic heterotrophic and mixotrophic species)now form a single group and have been placed together withthe ciliates and apicomplexans (sporozoans) in themonophy-letic Alveolata (Fig 2 lower panel) On the other handeukaryotic heterotrophic single-celled species falling withinthe general description ldquoprotozoardquo are now widely distributedamong a number of protistan lineages In short nutrition hasbeen demoted as a phylogenetic character and other charac-ters (presumably more indicative of evolutionary relatedness)have ascended to address some long-standing contradictionsalthough the debate over the relationships among some line-ages is still very active at the present time 79)
Individual protozoan cells range in size from less than 2 micromto greater than 1 cm in diameter (gt4 orders of magnitude)(80 81) with some colonial radiolaria forming cylindricalgelatinous structures a centimeter in diameter and morethan a meter in length (82 83) Because they constitutesuch a large size range of organisms protozoa are often dividedinto size classes that very crudely correlate with their generalnutritional preferences A commonly used convention is thatof Sieburth et al (84) which groups planktonic microorgan-isms into order-of-magnitude size classes (02ndash20 microm =picoplankton 20ndash20 microm = nanoplankton 20ndash200 microm =
microplankton 02ndash2 mm =mesoplankton) Most protozoafall into the nanoplankton or microplankton size classesModeling microbial trophodynamics using this conventionassumes that protozoa in one size category generally consumeprey one order of magnitude smaller in size (34 85)Although this approach misses much of the detail and diver-sity of the trophic activities of individual protozoan taxa it isa necessary practical compromise for examining community-scale flows of energy and elements It also provides a usefulmechanism for summarizing and comparing the abundancesand biomasses of protozoa from different environments andto other microbial assemblages Protozoan abundance sum-marized in this way has been shown to contribute signifi-cantly to the living biomass of planktonic ecosystemsthroughout the world ocean (Fig 3)
Estimates of protozoan biomass such as those depicted inFig 3 typically do not include the contribution of mixotro-phic phytoflagellates to heterotrophy There is presently noeasy way to determine the abundances of small phagotrophicphytoflagellates in natural samples so these species are typi-cally counted as phytoplankton unless specific methods areemployed to identify the algae as consumers such as the useof fluorescently labeled particles (87ndash90) or through theexamination of food vacuole contents (91) On averagethese species appear to constitute a modest percentage ofthe phytoplankton assemblage (typically lt25) althoughthey may at times dominate the phototroph assemblages ofnatural plankton communities It is important to recognizethat their inclusion as functional heterotrophs rather thanphototrophs could significantly shift the relative contribu-tions of phototrophic and heterotrophic microbial biomassto total biomass within microbial assemblages and the flowof energy within plankton communities (92)
Heterotrophic protists that harbor photosynthetic pro-tists or their chloroplasts within their cytoplasm constituteanother complexity for estimating the contribution of pro-tozoa to total microbial biomass When bulk water samplesare analyzed the contribution of chlorophyll containedwithin those protozoa is generally assumed to come fromfree-living phytoplankton However studies have shownthat chloroplast-bearing ciliates can contribute up to halfthe total biomass of planktonic ciliates in ecosystems andchloroplast-retaining ciliates can sporadically dominate thechlorophyll and primary production of some planktonic eco-systems (93ndash96) The environmental conditions promotingthe success of these ciliates are poorly known Similarlymany species of planktonic foraminifera polycystine radio-laria and acantharia harbor large numbers (thousands perprotozoan) of endosymbiotic algae within their cytoplasm(97 98) Caron et al (99) have demonstrated that primaryproductivity within these species can contribute significantlyto total primary productivity in oceanic ecosystems and can bevery important locally in the convergences of Langmuir circu-lation cells (100)
BACTERIA AND ARCHAEAldquoCulturablerdquo versus ldquoNonculturablerdquo CellsMost conventional cultivation methods can grow only 1 orless of the bacteria that can be visualized by direct microscopytechniques (eg 4) This is true even though most can beshown to be active by techniques such as microautoradiogra-phy (25) These readily cultivable organisms appear to repre-sent a group of fast-growing so-called weeds that are adaptedto take advantage of rapid growth in rare organically enriched
422-6 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
environments This strategy contrasts with the numericallydominant bacteria that are adapted specifically for growthin the dilute nutrient conditions that characterize the vastmajority of the volume of the water column
The most common taxa readily cultured from seawaterwith standard nutrient broth media include the gamma pro-teobacterial genera Vibrio Alteromonas PseudoalteromonasMarinomonas Oceanospirillum Shewanella (usually isolatedfrom surfaces such as shellfish and sediments) the alpha pro-teobacterial generaRoseobacterSphingomonasmembers of thefamily Flavobacteriaceae and Planctomycetes as summarizedin Giovannoni and Rappeacute (101) and Fuhrman and Hagstrom(102) The cyanobacteria Synechococcus and Prochlorococcusare also now readily culturable but on low-nutrient inorganicmedia targeting photosynthetic forms as opposed to organicmedia used to cultivate the others listed above
Because of the low percentage of marine bacteria that canbe grown in standard media organisms that until recentlywere called ldquononculturablerdquo make up the large majority ofbacteria in the plankton Only during the pastsim20ndash25 yearshave molecular biological methods based on 16S rRNA genesequences been available to identify these organisms andthese powerful techniques have opened up a large area forexploration (see next section) Similar but more recent stud-ies use 18S rRNA sequences for characterizing protistandiversity as will be noted below
Molecular Phylogeny and MetagenomicsField ApplicationsModern phylogeny of microorganisms is based primarilyon genetic sequences the most well-studied gene being the
FIGURE 2 The three domains of life (upper left) as proposed by Woese et al (78) and a recent overview of modifications that have beenproposed by Adl et al (79) to higher-level phylogentic groups within the eukaryotic component of the tree (lower right) Domains figure fromWoese et al (78) eukaryotic tree figure from Adl et al (79) doi1011289781555818821ch422f2
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-7
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
small subunit ribosomal RNA gene (16S rRNA in Bacteriaand Archaea and its larger homolog 18S rRNA in eukar-yotes) This molecule is strongly conserved over evolutionarytime so this single molecule has been used for constructingphylogenetic trees of all living organisms (httptolweborgtreephylogenyhtml) Analysis of 16S18S rRNA genesequences has been used to evaluate deep evolutionary rela-tionships among organisms and was instrumental in point-ing out that Archaea Bacteria and Eukarya should beconsidered different Domains of equivalent phylogeneticrank above kingdoms (78) However there are sufficientdifferences in 16S18S rRNA gene sequences to demonstrate
relationships (ormake distinctions) at the genus or sometimesspecies level
The first phylogenetic studies based on 16S18S rRNAgenes used sequences derived from cultures However onedoes not need cultures to obtain rRNA gene sequences (orany other sequences for that matter) An idea developed inthe lab of Norman Pace in themid-1980s involved extractionof DNA directly from natural samples and then cloning andsequencing of the DNA as a means of assaying the microbespresent in the samples (103 104) The original protocolscalled for cloning by creating what are called ldquophage librariesrdquofrom the natural DNA but since 1986 PCR has been applied
FIGURE 3 (a b) Plankton biomass in the Arabian Sea during the 1995 southwest monsoon (a) and intermonsoon period (b) Areas of theboxes indicate the relative magnitudes of the biomass in each category Categories within the dashed boxes in a b c are composed of protozoaArrows indicate the direction of energymaterial flow in the food web thicker arrows depicting greater flow Redrawn from (86)(c) Depth-integrated biomass (mgm2) in the upper 100 m of the Sargasso Sea near Bermuda and in the upper 200 m of the equatorial Pacificat 175degE Thewidth of the bars indicates the biomass in each size category Heterotrophs have been separated by size class while phytoplanktonhave not Size classes delineated by the dotted box are comprised of protozoa Redrawn from (65) doi1011289781555818821ch422f3
422-8 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
extensively for cloning and related studies The targetsequence can be almost instantly ldquoidentifiedrdquo to its closestphylogenetic neighbor by what have come to be standardonline sequence comparisons
Beyond the study of targeted genes (like the 16S rRNAgene) shotgun metagenomic studies have examined theentire genetic repertoire of the microbes in a given sampleThe metagenome is the collective genome of all organismsin the sample Initially these studies extracted DNA fromall organisms in a sample (usually prefiltered through a filterapproximately 1 microm to remove most organisms larger thanbacteria) sheared it to produce fragments and clonedthem into standard vectors either as small (thousands ofbases) or large (to hundreds of thousands of bases) insertsthat is fragments of DNA from the environment nowcloned into the vectors in a form suitable for sequencingThe best known early marine study of the former type isthe Global Ocean Survey with initial results published byVenter et al (105) which generated more than a billionbases of DNA sequence and reported 12 million previouslyunsequenced genes estimated to come from at least 1800different genomic species cumulatively in the many samplesthey analyzed
With the advent of next-generation sequencing that gen-erates millions ormore sequences in a run (known by a varietyof acronyms including 454 Illumina SOLiD etc) cloningof genes has largely been replaced by clone-free sequencingThe extent and power of such sequencing has recentlybeen demonstrated by the ability to construct essentiallythe entire genome of an uncultivated marine Group II Eur-yarchaeon that constituted only approximately 2 of amicrobial community sample using SOLiD sequencingmade possible by high coverage and the use of mate pairsequencing ofsim3000 bp fragments (106) (this length isneeded to span repeats and highly conserved genes that oth-erwise make genome construction difficult)
For 16S rRNA gene studies ldquotag sequencingrdquo pioneeredby Sogin et al (107) consists of amplifying a suitable sizedpart of the gene with broadly conserved primers (choice isimportantmdashfew are truly universal for the groups intended)often ldquobarcodedrdquo to allow multiple samples to be combinedinto a single run The amplified products are then sequencedand sequences processed en masse Many thousands of partialSSU rRNA gene sequences per sample are economically ana-lyzed this way though they are usually short (currently a fewhundred bases each depending on the sequencing platform)This way of analyzing the composition of microbial commun-ities is now standard These approaches provide so muchinformation even about very rare sequences that the resultshave led to the important concept of the ldquorare biosphererdquoorganisms that may be active or dormant and constituting avery small proportion of the community (eg often muchless than 01) but are potentially important for dispersionadaptation to changing conditions or even critical activitieslike nitrogen fixation or vitamin production (108) Howeverdue to the potential for artifacts like error sequences thisapproach requires rigorous application of quality filteringand clustering algorithms to avoid erroneous taxa and overes-timation of species richness (109 110)
Next-generation sequencing approaches have also allowedwork to begin metagenomics studies of microbial eukaryotes(111) These studies are still constrained by the much largergenomes of eukaryotes and therefore the difficulties ofobtaining sufficient sequences to reconstruct the genome ofa particular species Along with advances in sequencing abil-ity studies of eukaryotic metagenomics have been facilitated
by advances in single-cell isolation which when coupledwith high-throughput sequencing approaches reduce the tre-mendous complexity present in natural complex eukaryoticcommunities to a manageable task (112) Such single-celltechniques are also quite valuable in studies of bacteria andarchaea though the amplification technique tends to bevery uneven and typically generates less than half the genomeof each isolated cell regardless of domain (113 114)
Additionally similar to DNA mRNA is amenable toextraction and sequencing although greater care must betaken during extraction and purification as RNA shows agreater susceptibility to degradation during processing Copy-ing of mRNA by reverse transcription of RNA into cDNAfollowed by DNA sequencing has allowed insights into themetatranscriptomes of environmental samples Metatran-scriptomic studies provide information on gene expressionin an ecosystem and thereby indicates ldquoactivityrdquo of themicro-bial community rather than simply ldquopotentialrdquo represented bythe genomic DNA present in the sample with many applica-tions from showing which processes are being carried outby which organisms to fine-scaled diel studies (115ndash118)Nevertheless given variations in the lifetimes of differenttranscripts and protein molecules the transcriptome maynot be fully representative of the current activity of anorganism
Limitations of these analyses include sequencing errorsPCR mismatches or biases clustering and bioinformaticschallenges and chimeras generated during PCR Also thephylogenetic resolution of short sequences is limited giventhe high conservation of rRNA sequences Even with clone-free shotgun metagenomics there may be biases such as non-random losses of DNA during extraction and preparationor biases (eg from G +C content or secondary structure)in the sequencing procedures Determination of speciesdiversity by these approaches provides an example of theselimitations Shakya et al (119) working with synthetic com-munities (purified genomicDNA from16Archaea represent-ing 3 phyla and 48 Bacteria representing 16 phyla remixed tosimulate an environmental DNA extract) applied both meta-genomic analysis (454 and Illumina platforms) and PCRamplification followed by 454 sequencing of 16S rRNA genesto determine both species richness and relative abundanceThey found that PCR amplification454 sequencing of 16SrRNA genes yielded an accurate measure of species richness(providing that appropriate data processing was applied)but that the relative abundance of up to 94 of the species(depending on domain and variable region amplified) wasover- or underestimated by at least 15-fold (values rangedfrom not detected to 103-fold overestimation) In contrastboth metagenomic approaches yielded relative abundancesthat were within the authorsrsquo 15-fold accuracy cutoff forsim50 of the species However they concluded that addressingrichness overestimation in metagenomic analyses that isdistinguishing rare but real OTUs from experimental andcomputational artifacts awaits further computational andclassification improvements More recently Parada et al(120) used mock communities composed of 16S rRNAclones from 27 common marine taxa (from nine Bacterialand two Archaeal phyla) to show that small differences inPCR primers (and different clustering methods) can yieldlarge differences in apparent relative abundances of reportedtaxa However one primer pair and informatics pipelinethey tested using a particular version of 515F-926R (V4ndashV5) provided accurate estimates of relative clone abundance(r2 = 095) when comparing observed versus expected cloneabundance
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-9
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
Culture-Independent Diversity Studies
The first groups to be identified using cultivation-independ-ent molecular techniques from the marine plankton (121)were the bacterial SAR11 cluster (a group of closely relatedgene sequences or phylotypes) and marine picoplanktonicunicellular cyanobacteria Synechococcus and Prochlorococcus)Of these the SAR11 cluster was completely unknown butthe cyanobacteria had previously been recognized by theirunique pigment fluorescence Waterbury et al (122) andJohnson et al (123) used epifluorescence to observe Synecho-coccus and Chisholm et al (124) discovered Prochlorococcususing flow cytometry These cyanobacteria were later isolatedand grown in phytoplankton culturemedia These two groupsare generally common in the euphotic zone with SAR11 typ-ically comprising one third of the planktonic bacteria (125)and the cyanobacteria common everywhere but polar waters
Probably the biggest surprise to come from the applicationof molecular tools was the discovery by Fuhrman et al (126)of abundant archaea in the deep sea The archaea were foundto be in a unique phylogenetic cluster that was only distantlyrelated to any previously known archaea but the ldquoclosest rel-ativesrdquo (not really close at all) were extreme thermophiles Asubsequent study also found archaea to be present in near-surface coastal waters albeit relatively rare (lt2 relativeabundance) This study used a PCR technique specificallytargeted archaea and found ldquomarine Crenarchaeardquo plus a sec-ond group belonging to the phylum Euryarchaea (127)
Up until this time all known Archaea were thought to beldquoextremophilesrdquomdashadapted for either very high temperatures(thermophiles) extremely salty conditions (halophiles) orstrictly anaerobic environments (methanogens) Yet theseorganisms were present in cold or cool water at ordinary salin-ity and high oxygen concentrations Fluorescent in situhybridization (FISH) measurements from deep-sea sampleshave since showed that the archaea may make up appro-ximately 40 of the total countable prokaryotes with thepercentage reaching to 60 at 200 m depth in theMediterra-nean (35 36) An extensive time series of FISH measure-ments near Hawaii confirmed that the archaea are indeedvery abundant throughout the year from below the photiczone to at least 4000 m and typically constituting 30ndash40of the total prokaryotes present in waters deeper than a fewhundred meters (128) with similar results found elsewhere(129 130) (Fig 4) They have been reported from many pla-ces including the Atlantic Pacific and Southern Oceansand are dynamic components of the plankton the most com-mon type by far being the ldquomarine Crenarcheaeardquo (128 129131) While the marine archaea have been reported to bedominated by a few major ldquophylotypesrdquo (132) they alsohave been shown to have a great deal of microdiversity withinthese phylotypes suggesting there are many kinds of close rel-atives coexisting (133) Interestingly this group of archaeamay likely be the most abundant kind of organism on Earthgiven the huge volume of the deep sea and their high abun-dance there (134) It has recently been proposed that ldquomarineCrenarchaeardquo be elevated to their own major phylum outsidethe Crenarchaeota called the Thaumarchaeota on the basisof deep phylogenetic branching and fundamental differencesbetween them and the Crenarchaeota (135) The Thau-marchaeota possess the uniquely archaeal membrane lipidcrenarchaeol and are nowalso known to be abundant in soils
The physiology of the Thaumarchaeota has been anintriguing area of study Initially an autoradiography-FISHcombination approach demonstrated that they take upamino acids (130 136) But subsequent data showed that
crenarchaeol was derived largely from CO2 suggesting auto-trophic metabolism (137 138) An experiment showinguptake of 13C-labeled bicarbonate into these archaeal lipids(139) directly pointed to autotrophy in this group Chemoli-thoautotrophywas first hinted at byVenter et al (105) whosemetagenomic analysis showed an apparent archaeal scaffoldthat contained genes suggestive of ammonia oxidation Fur-ther evidence came from Schleper et al (140) who detectedseveral ammonia oxidation genes in order directly adjacent toa Thaumarchaeota 16S rRNA gene in a soil-derived metage-nomic clone The issue was directly resolved when Konnekeet al (141) isolated a related marine archaeon from sedimentof a marine aquarium and this organism Candidatus Nitroso-pumilus maritimus was found to have a chemoautotrophicmetabolism quantitatively oxidize ammonia to nitrite andcontain an archaeal ammonia monooxygenase gene (amoA)and interestingly did not grow heterotrophically Cultivationexperiments showed this organism has a high affinity forammonium allowing it to outcompete bacterial nitrifiers atlow (submicromolar) concentrations as are typical in thesea (142) The complete genome of the organism has shownnovel adaptations for nitrification and autotrophy (143)
Even if the Thaumarchaea are primarily chemolitho-autotrophs field data suggest a level of mixotrophy in thatsome organic substrates are being incorporated into biomassA stable isotope study using cells collected at 670 m depthoff Hawaii estimated that about 80 of the carbon incor-porated into archaea-specific lipids came from inorganic sour-ces and about 20 from organic compounds (144 145)Interestingly it has also been claimed that the genomesof deep-sea (gt2000 m depth) members of the Thaumarch-aea as well as those living in equatorial waters rarely containthe amoA gene and thus may be primarily chemoorgano-trophs (48)
Major bacterial groups that have been documentedfrom seawater using 16S rRNA characterization includesome that are also known from culture (eg AlteromonasRoseobacter) and several that are phylogenetically distantfrom standard cultures The most common groups are inrough order of their relative abundance in clone librariesfrom most to least abundant SAR11 (relatives of Pelagibac-ter ubique) Roseobacter SAR86 cyanobacteria SAR116SAR202 SAR234 andMarine Group A The SAR designa-tion followed by a number is an arbitrary sequential cloneidentifier from Sargasso Sea cloning studies done by the Gio-vannoni lab that did most of the early systematic cataloging ofclones Summaries of the data and phylogenetic relationshipsof these groups can be found in Fuhrman andHagstrom (102)and Giovannoni et al (101)
Molecular Genetic Discoveries in Bacterial andArchaeal Marine BiologyAs described earlier metagenomics is an extension of theideas used in the 16S rRNA cloning studies in that all genesfrom the native microorganisms are separated and clonedwithout having cultivated the organisms These methodsearly on started to find unexpected and very interestingresults The best examples involve unexpected marine photo-trophy the first of which is discovery of a nonchlorophyllphotosynthetic bacterial pigment called proteorhodopsin(146) The gene was found on a large environmentallyderived fragment of DNA that also had a gene coding for16S rRNA from the so-called SAR86 group (one ofthe groups common in seawater) This pigment can act as alight-driven proton pump thought to permit cells to generate
422-10 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
FIGURE 4 Distribution of Bacteria Euryarchaea andmarine Crenarchaea (recently renamed Thaumarchaea) along a transect in theNorthAtlantic Ocean as measured by CARD FISH and expressed as of total bacteria + archaea counts via DAPI stained epifluorescence fromTeira et al (130) Top panel shows station locations that are shown on the top of the lower three panels with the distance in km reported alongthe transect from lower to higher station numbers doi1011289781555818821ch422f4
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-11
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ATP from sunlight Interestingly different versions of this pig-ment are found at different depths apparently ldquotuningrdquo theabsorption to match the ambient wavelengths of light asthey change with depth (147) The proteorhodopsin genesare very widespread and diverse perhaps in half or more ofall marine bacteria including SAR11 and Euryarchaeota(105 148ndash150) However almost all of several culturedorganisms with proteorhodopsin studied to date do not showa growth benefit from light (150) with the exception of amember of the Flavobacteria Dokdonia sp strain MED134that grows faster in the light only under moderately lownutrient conditions (151) and a marine Vibrio has beenshown to survive starvation longer in the light than in dark-ness (152) These observation may explain the wide phyloge-netic distribution and high abundance of this gene in thatproteorhodopsin may often assist long-term survival of bacte-ria under extreme energy-limiting conditions yet not havemucheffect on growthduringmore energy-replete conditionsInterestingly a recent report suggests the growth benefit fromlight in Dokdonia MED134 is from enhanced uptake of itsrequired growth factor vitamin B1 which is transported by atonB-dependent transporter powered by a proton gradient(153) This pigment might also have sensory or other rolesnot yet well examined (150)
The second kind of phototrophy found by moleculargenetic and fluorescence techniques to be unexpectedlyimportant in marine plankton is anoxygenic aerobic bacterialphototrophy based on the pigment bacteriochlorophyll aCulturable aerobic anoxygenic photosynthetic (AAP) bacte-ria have been known from seawater for several years (154) andare widespread (155) Initial recent reports of direct countsclaimed they represent about 11 of the total bacterial com-munity in the euphotic zone (156) but those early counts didnot correct for the presence of other bacteria and such correc-tionoften yields estimates averaging closer to 2(157)Directmeasurements show the bacteriochlorophyll pigment is rela-tively rare (158) However studies in various ocean locationsincluding ocean gyres report that these AAP bacteriacan indeed on occasionmake up a quarter of the total prokar-yotes (159 160) A recent energetic modeling of AAP andproteorhodopsin-containing bacteria (161) has suggestedthat proteorhodopsin-containingones cangain approximately02asmuchenergy from sunlight asSynechococcus (commonmarine cyanobacterium) and AAP bacteria can gain approxi-mately 13 as much energy from sunlight as SynechococcusThey concluded the AAP bacteria may gain energy sufficientto meet maintenance costs but proteorhodopsin-containingbacteria were not expected to do so except at high light inten-sities andwith largenumbersof proteorhodopsinmoleculespercell They concluded the ease and low cost of maintainingPR-based phototrophy (a few genes required) may explainthe high incidence of proteorhodopsin genes
Other metagenomic studies have yielded interestinginsights about organisms and processes that would otherwisebe difficult or impossible to establish For example thesequence of an archaeal gene fragment isolated from seawaterrevealed extensive evidence of genetic exchange with othertypes of archaea and even bacteria (162) Such geneticexchange was also quite evident from the complete genesequences of different strains of Prochlorococcus and Synecho-coccus which was attributed in part to virus-mediated geneflow (163ndash166)
Dilution Cultures of Bacteria and ArchaeaSome recent cultivation techniques that permit growth ofldquotypicalrdquo oligotrophic marine bacteria in pure culture are
based on the simple concept that bacteria living in seawateron dissolved organic matter might best be grown in the labo-ratory in ordinary filtered seawater This conclusion followsfrom similar techniques to grow mixed ldquoseawater culturesrdquoof marine bacteria (167) but the seawater inoculum in thiscase is diluted so that only one or a few bacteria are addedto the initial culture vessel (168) A rapid throughput versionof this method has been used to cultivate members of bacte-rial groups thought to be conventionally uncultivable such asthe SAR11 clade (169 170) albeit often at very low den-sities Such cultures permit focused studies of the propertiesof those organisms including genomic analysis (169 170)This valuable work provides insight into the roles and activ-ities of some of the most common bacterial phylotypesobserved in the ocean (171) showing for example that culti-vated SAR11 requires a source of reduced sulfur as well asproviding templates aiding the interpretation of metage-nomic analysis (172)
PROTOZOADiversity and DistributionHeterotrophic protists have been identified historically fromtheir morphological features that are apparent at the levelof the light or electronmicroscope Features of cell size shapetype and pattern of flagellationciliation skeletal structurescharacteristics of the nucleus and other cellular structuresand organelles have been used to differentiate themany thou-sands of described species A description of the extent of thisdiversity of form and function is well beyond the scope of thisbook (see [79] for more information) However much of thisdiversity can be reduced to three basic body plans that dictatethe broadest ecological roles of these cells amoeboid flagel-lated and ciliated forms (Fig 5) Among the lineages possess-ing one of these three forms only the last group the ciliatedprotists constitute a monophyletic group within the domainEukarya
One of the simplest body plans for protozoa is the amoe-boid cell exemplified by the gymnamoebae or ldquonakedrdquoamoebae Motile nonphotosynthetic cells lacking flagellaor cilia occur as life stages in a number of phylogeneticallydiverse taxa but for many free-living species of protozoathis form constitutes the only life stage Motility is largelyconfined to movement along surfaces by means of pseudopo-dia that can take on a variety of (species-specific) shapes Sig-nificant abundances of amoebae are largely confined tobenthic and epibiotic environments (174) and to suspendedparticulate material where they can occasionally be highlyenriched (27) Most amoebae consume bacteria and otherminute prokaryotes and eukaryotes
Several heterotrophic protistan groups possess complexamoeboid body plans most notably the foraminifera polycys-tine and phaeodarian radiolaria and the acantharia Plank-tonic forms are predominantly oceanic in their distributions(although a large number of benthic species of foraminiferaexist) These species are heterotrophic and many are visibleto the naked eye (individual cells can bege1 cm gelatinouscolonies can form ribbon-like structuresge1 m in length)They feed on a wide variety of bacterial protistan and meta-zoan prey using pseudopodial networks have rather longcomplex life cycles for individual cells (weeks to months)and are extensively used in studies of paleoclimatologicalreconstruction (82 175) In addition many of these speciespossess intracellular symbiotic usually eukaryotic algaeThe widespread occurrence of algal endosymbiosis among
422-12 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
these protozoa in oceanic pelagic environments implies thatstrong selective forces appear to give rise to these associations(97 98)
Flagellated forms of protozoa exist across many protistanlineages Eukaryotic flagella come in various sizes numbersand forms (eg with or without tiny hairs) that are character-istic of the different lineagesMany flagellate species are appa-rently purely phototrophic or heterotrophic (phagotrophic)but a significant (still poorly known) fraction of flagellates aremixotrophic combining both phototrophy and phagotrophy(70 88) Flagella in phagotrophic flagellates are employed formotility and prey capture Most free-living flagellates possessone to four flagella (typically one or two) that can be manytimes the length of the cell itself Although there is tremen-dous species diversity among flagellated protozoa many ofthese species have broadly overlapping ecologies As a groupflagellates are the most numerically abundant protozoa ofboth benthic and pelagic ecosystems and they are fundamen-tally important as consumers of bacteria cyanobacteriaand other eukaryotes Flagellated protozoa within the nano-plankton size class (2ndash20 microm) are often counted as a singleassemblage in plankton studies because morphological details
apparent using light microscope are insufficient to distinguishamong the many species This assemblage has been variouslyreferred to by a variety of names and acronyms including het-erotrophic nanoplankton (HNAN HN) heterotrophic flag-ellates (Hflags) heterotrophic nanoflagellates (HNF) andmicroflagellates Heterotrophic nanoplankton is the mostaccurate term for most methodologies employed to countthese cells because it does not require visualization of flagella(which are often lost from these small cells during preparationfor microscopy) The confusion over terminology is partly aconsequence of the different methodologies that have beenused to count these species and partly a consequence of thefact that these species were first studied prior to the wideacceptance of the size convention of Sieburth et al (84)
Heterotrophic flagellates in the microplanktonic size class(20ndash200 microm mostly dinoflagellates) are important consum-ers of phytoplankton in pelagic ecosystems Many of thesespecies are capable of the production of large pseudopodialnets the pallium that can engulf prey (particularly diatoms)significantly larger than the diameter of the dinoflagellatetheca (176) This behavior and its ecological significance forenergy flow in plankton communities have been recognized
FIGURE 5 Body plans and size ranges of protozoa These micrographs depict amoeboid (andashg) flagellated (hndashj) and ciliated (kndashm) forms ofprotozoa From Caron et al (173) Markers bars are 5 (i) 10 (h) 20 (c j l) 30 (a) 50 (k m) 100 (b f) 500 (d) and 1000 (e g) micromdoi1011289781555818821ch422f5
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-13
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
only within the past few decades (177) These species areoften abundant in waters where diatoms dominate (178179) In such situations heterotrophic dinoflagellate biomasscan be similar to that of ciliated protozoa (180)
Ciliates are generally the most recognizable form of pro-tozoa to nonspecialists Nonetheless ciliates are a diversegroup morphologically as well as ecologically (181 182)The degree of ciliature on these species can range from uni-formly ciliated to totally devoid of cilia during most of theirlife cycle to ciliature restricted to specific regions of thecell In addition cilia can fuse to form various complexstructures (eg ciliary membranes or veils cirri) that assistin locomotion food capture or attachmentCiliates are abun-dant and ecologically important species in both benthic (8)and pelagic (183) ecosystems and can consume a variety ofprokaryotic and eukaryotic prey Together with the heterotro-phic dinoflagellates these species are the dominant consum-ers of phytoplankton in many pelagic ecosystems (184) andas a consequence they form an important trophic link to met-azoan zooplankton (185 186)
Diversity and Biogeography of Protists FromMorphology to DNA SequencesUntil relatively recently the biodiversity of marine protistanassemblages was not generally considered a controversialtopic It was generally accepted that while all species of pro-tists certainly have not been identified representatives ofmost types of algae and protozoa had been observed anddescribed if not actually brought into culture in the labora-tory Direct sequencing of 18S rRNA genes from environ-mental samples (as described in ldquoMolecular Phylogeny andMetagenomics Field Applicationsrdquo) has changed that viewInitial forays into environmental DNA indicated a muchgreater diversity of protists than previously documented usingclassical approaches of culture and microscopy (81 187ndash191) Numerous publications over the past decades haveexpanded these findings (see [192] for a recent global analy-sis) which are highly analogous to discoveries in marine pro-karyote research as described already
Previously uncharacterized protistan diversity has nowbeen documented at virtually every level of eukaryoticorganismal classification Some of these findings couldhave been expected but some have been very unexpectedFor example a much greater diversity than noted previouslyhas been observed among small (lt10 microm) protists in plank-tonic ecosystems These species generally possess few distinc-tive morphological features and one could expect that manycryptic species might be present among these small morpho-types The molecular ldquodiscoveryrdquo of this eukaryotic diversityhas stimulated progress on the isolation and descriptionof new species and genera of minute algae and protozoa(193ndash197)
Analyses of environmental samples have also indicatedthe unanticipated existence of novel 18S rRNA gene sequen-ces that imply the presence of novel lineages of eukaryotes innatural protistan communities (198ndash200) These are sequen-ces for which there are apparently no known described orcultured species The degree to which these sequences dif-fer from sequences of known sequenced eukaryotes impliesthat some of these lineages may be distinct at the level ofphylum (201)
These findings have raised basic questions and somedebate concerning the true diversity and biogeography ofprotistan assemblages in natural ecosystems (202ndash204) thevalidity of the many novel phylotypes or cryptic species that
are being documented (205) the potential importance andsignificance of the many rare taxa that characterize thesecommunities (206) and the implications of these findingsfor the ecologicalbiogeochemical roles that protists playin aquatic ecosystems For example it was postulated andsubsequently confirmed that some novel alveolate lineagesreported from marine ecosystems make up a suite of parasiticprotozoa whose ecological importance may have been signifi-cantly underestimated in the past (207) Deciphering theidentity and significance of these many unknown phylotypeswill constitute a significant effort for protistologists in thefuture (80 85)
Life Histories and Ecological Strategies of ProtozoaAbundance prey type and life histories all vary tremendouslyamongmarine protozoa The smallest species (eg many flag-ellates) tend to be the most abundant and widely distributedin the world ocean Indeed many of these species may beglobally distributed (208) Most nanoplanktonic flagellateshave potentially rapid rates of grazing and growth Underoptimal conditions these species can divide by binary fissionseveral times a day and thus dramatic increases in their pop-ulations can take place in response to favorable conditionswithin a few days (209) Many of these species are capableof surviving for limited periods of time without food andhave developed a variety of physiological or life cycle strat-egies to cope with these events (209) However in contrastto some bacteria that may remain viable through long periodsof starvation (210) protozoa will expire or encyst in responseto low food abundance and thus ldquoboom-and-bustrdquo popula-tion changes are characteristic of small flagellates
On the other end of the size spectrum from the ratherubiquitous nanoflagellate species many of the largest proto-zoan species (polycystine and phaeodarian radiolaria plank-tonic foraminifera) are exclusively oceanic (ie do notsurvive in most coastal environments) andor have specificlatitudinal and depth distributions Thus the abundancesof these latter species may range from undetectable to max-imal abundances of gt105 individualsm3 Dramatic changesin the abundance of these protozoan taxa also can be relatedto changes in prey abundance physicalbehavioral aggrega-tion or to periodicity of life cycle events For example theplanktonic foraminifer Hastigerina pelagica reproduces on alunar cycle and thus abundances (and life stages) of this spe-cies in oceanic waters can vary considerably over the courseof a month (211) In general life cycles for the large amoe-boid protozoa are lengthy and complex (for single-celledorganisms) with life spans unknown for many species(attempts to culture them in the lab have so far been unsuc-cessful) but estimated to be on the order of months to per-haps years (175)
The abundance and activities of microplanktonic hetero-trophic protists (mostly ciliates and heterotrophic dinoflagel-lates) tend to be somewhat intermediate to those ofnanoplanktonic flagellates and the larger amoeboid formsThese species are present in the majority of marine ecosys-tems and collectively play an important role in the controlof phytoplankton biomass (and probably the abundance ofnanoplanktonic protozoa although there is little informa-tion on this topic) in waters throughout the world oceanLike small flagellates ciliates and dinoflagellates reproduceprimarily by binary fission but their maximal growth ratesare typically slower (one division a day is typical) and theirlife cycles often include sexual phases that allow for geneticrecombination
422-14 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
VIRUSESViral Abundance and General PropertiesViruses are simple biological agents typically 20ndash200 nm indiameter composed of a nucleic acid genome in a proteincoat that infect cells and ldquocommandeerrdquo the cellrsquos machineryto make more viruses which are released into the environ-ment when the host cell lyses or bursts A particular virus isthought to be capable of infecting only a narrow range of hosts(usually one species sometimes a genus rarely broader)Although some early studies had isolated viruses from thesea therewere no data prior to the 1980s showing such viruseswere very abundant and more significantly no evidence thatinfection was occurring in any important part of the planktoncommunity It wasnrsquot until the late 1980s that electronmicro-scopy with suitable concentration methods showed thatviruses are extremely abundant similar to or even higherthan bacterial abundance (212 213) Interestingly themost suitable transmission electron microscopy (TEM)approach used by Bergh et al (described in detail by [214])was actually similar to a direct centrifugation method devel-oped in 1949 (215)
TEM studies of viruses permit high-resolution images andobservation of viral morphology showing features such ashead diameter tails sheaths and tail fibers (Fig 6 collage
in upper left) Each type of virus has a fixed morphology(unlike bacteria that are potentially more plastic in theirappearance) and hence a coarse measure of viral diversityhas been possible by cataloging morphologies Studies thathave done so have found dozens or more different morpholo-gies of marine viruses in a given sample (217ndash219) A largeproportion resemble bacteriophages which are viruses thatinfect bacteria
The availability of brightly fluorescent nucleic acid stainsand high-porosity fine pore size (002 μm) filters made fromAl2O3 have made it easy to accurately count viruses by epi-fluorescence microscopy (38 220 221) Epifluorescence per-mits abundance estimates but does not allow observation ofviral morphology as viruses are below the resolution limitof light microscopy viruses are visible only as sources of light(like stars in the night sky see Fig 6 lower micrograph) Anextension of manual epifluorescence counts is the use of flowcytometry to count viruses now commonly employed in somelabs (222)
Virus abundance has been found to be closely related tobacterial abundance with a virusbacteria ratio typically10ndash301 A typical oceanic profile of bacterial and viral abun-dance is shown in (Fig 6 panel on right) This tight ratioand the strong correlation to bacterial abundance in relationto weaker correlations to chlorophyll have been cited as
FIGURE 6 Marine viruses (upper left collage) viewed by transmission electron microscopy These are cyanophages in the groups myoviridae(right and bottom) siphoviridae (left) and podoviridae (top) Scale bars represent 100 nm From Sullivan (216) Epifluorescence micrograph(lower left) of SYBR green I stained viruses (small fluorescing objects) and bacteria + archaea (large fluorescing objects) Depth distribution ofvirus and bacteria + archaea abundances obtained by epifluorescence microscopy of SYBRGreen stained cells in the central San Pedro BasinCalifornia (11 August 2000) doi1011289781555818821ch422f6
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-15
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
evidence that most marine viruses infect bacteria rather thaneukaryotic phytoplankton (217 218 223 224) though thereare also many important viruses of phytoplankton and otherorganisms with significant impacts on primary productivityblooms and ecosystem function (225ndash227)
The first demonstration of the activity of marine viruseswas presented by Proctor and Fuhrman (228) who showedby electron microscopic examination that viruses wereactually infecting marine bacteria and cyanobacteria at ameasurable rate Because only the final portion of the viruslife cycle is visible by electron microscopy (when the virusesare assembled and ready to lyse the host cell) only a very smallfraction of the infected cells can be counted as infected at anygiven time Proctor and Fuhrman (228) reported that only afew percent of the total bacteria from coastal waters and theSargasso Sea were visibly infected but they interpreted thedata with a model that implied the actual fraction of the totalcommunity that was infected was much higher They con-cluded that the total fraction of bacterial mortality attribut-able to viruses is roughly 10ndash40 Subsequent studies haveused refined versions of that same general approach andmodel (229 230) and numerous studies have used a varietyof alternative approaches to estimate viral activitymdashall yield-ing basically the same general conclusion (although themodel parameters need to be adjusted for cyanobacteriawhich on average seem to be infected less than bacteria)These approaches include (a) calculation of virus turnoverfrom decay estimates (231) (b) measurement of viral DNAsynthesis by incorporation of tritiated thymidine (232) (c)observation of effects of added viruses (233 234) (d) estima-tion of bacterial mortality in the absence of protists (e) use offluorescent viruses to estimate production by an approachanalogous to isotope dilution (38) and (f) dilution of virusesin filtered samples to estimate viral production (235) Theoverall consensus of these studies is that the initial estimateswere basically correct that is that viruses are responsiblefor about 10ndash40 of the bacterial mortality in most marinesystems (reviewed by [218 223 226 236 237]) Howeverthere is still some disagreement about the higher estimatesand it is likely that 40 mortality from viruses alone is nottypical for most marine systems
Most of the foregoing work has focused on the viral infec-tion of bacteria primarily thought to be heterotrophicalthough the original report by Proctor and Fuhrman (228)also noted the occurrence of cyanobacterial infection Sev-eral subsequent studies focused on phytoplankton includingthe potential effect of viruses on the termination of phyto-plankton blooms of Emiliania and Phaeocystis and the likelyimpact this might have on release of climate-active gasessuch as dimethyl sulfide (225 227 238ndash247) Althoughdetails are beyond the scope of this chapter viruses arethought to infect virtually all marine organisms with poten-tially significant impacts from zooplankton to whales (226)Detailed studies of viruses infecting cyanobacteria such asSynechococcus have shown differences in viral host specific-ity particularly toward coastal and oceanic host strainsand some occasional high virus abundances (to 105ml) asmeasured by most probable number (MPN) cultivation tech-niques in the Gulf of Mexico near Texas (248 249) Othervirus cultivation studies with Prochlorococcus and Synecho-coccus in oligotrophic waters of the Sargasso Sea showedan interesting pattern of cross-infection between these gen-era by some virus types (suggesting gene flow among theseorganisms) but generally low MPN estimates of abundanceto 103ml even when cyanobacterial abundance was near105ml (216 250)
Overall the consensus emerging from direct comparisonsof viral-mediated mortality and grazer-mediated (ie micro-zooplankton) mortality have indicated that viral lysis of bac-teria constitutes a significant fraction of total mortality ofthis assemblage while the mortality of phytoplanktonappears to be dominated bymicrozooplankton inmost instan-ces (251 252)
As mentioned viruses have significant morphologicaldiversity as observed in TEM studies It is also possible toinvestigate aspects of their genetic diversity In early workon this topic Wommack et al (253) and Steward et al(254) observed the diversity of viral genome lengths in a fieldsample by pulsed field gel electrophoresis with viral genomesranging 25 kb to gt300 kb in length Field results show thatthe viral community composition is dynamic in space andtime with clear changes in the banding patterns over seasonsand locations in Chesapeake Bay (253) between oceanbasins and subsequent to dinoflagellate blooms (254) andwith depths to 500 m at one location (255)
Although viruses do not all share a set of core genesthat allows a universal viral phylogeny (comparable to SSUrRNA gene in cellular organisms) genetic diversity amonga single group of viruses can be examined by sequence analysisof shared genes within the group An example is the g20 geneinT4-like cyanophages that has shown extremely high diver-sity even among very closely related viruses and has demon-strated geographic and seasonal variation (256) A secondexample is g23 which is found broadly in diverse and wide-spread T4-like phages (257) and can have seasonally repeat-ing patterns (258) as well as short-term rapid dynamics thatcorrelate to those in bacteria (259)
Viral MetagenomicsAs with cellular organisms metagenomics can be used toexamine viral diversity and genetics without themany restric-tions of cultivation Viruses can be collected by selective fil-tration and concentrated by tangential flow filtration orflocculated with iron chloride (260 261) then their collec-tive metagenome can be extracted linker amplified and ana-lyzed by sequencing (262 263) Viral metagenomic studiesare particularly challenging because the large majority ofsequences have no annotated matches in any databases butso far results from marine samples around the world haveshown extremely high diversity and variations with depthlocation and time presumably with highly dispersed typesselected by local conditions (262ndash266) Because metage-nomes are best interpreted when there are representative cul-tures available the best matches of viral metagenomesoriginally tended to be to the few viral isolates infecting trulycommonmarine bacteria like cyanophage infecting Synecho-coccus and Prochlorococcus (267) However the developmentof dilution-to-extinction cultures representing common het-erotrophic marine taxa like Peligibacter (a member of theSAR11 clade) and SAR116 has allowed isolation from sea-water of viruses infecting these common organisms and theseisolates have indeed been found to be highly abundant inmarine viral metagenomes (268 269) New approaches tointerpret the results include clustering the proteins independ-ent of known proteins to compare samples to each other andtry to find environmental factors driving viral communitychanges (270) Such analysis of a large global data set (TaraOceans expedition) has shown that extensive sampling hascome close to reaching the total diversity in tropical and tem-perate waters of such viral protein clusters (which essentiallyrepresent various viral protein motifs not all viral proteintypes) and that viruses appear to be directionally dispersed
422-16 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquodownstreamrdquo in major ocean currents as one may expectfrom first principles but also supporting the ldquoseed bankrdquohypothesis of viral biogeography (271)
Viruses and Host DiversityViruses are themselves thought to be instrumental in drivingincreased diversity of their microbial hosts via a hypothesizedprocess often called ldquokill the winnerrdquo This is because viralinfection is host-specific and density-dependent the latterbecause viruses diffuse from host to host so an abundanthost is more likely to pass on infection than a rare oneThis means that if an organism becomes abundant andblooms winning the competition for resources it becomesmore susceptible to a viral epidemic This would benefit therarer organisms and thus help foster diversity (223 272273) However bloom scenarios as described above involvesystems far from steady state and the formal theory of Thing-stad and Lignell (274) has interesting steady-state solutionswhere several viruses infect several hosts stably over time(via trade-offs between growth rates and viral susceptibility)which may occur at the strain or species level (274) There issome experimental evidence that viruses have effects on nat-ural marine microbial community composition weaker thanthe kill the winner (bloom version) hypothesis would sug-gest (275 276) so something resembling the steady-statecoexistence described in the model may in fact be commonThere also appear to be processes that foster coexistencebetween viruses and hosts but the mechanisms are largelyspeculative (223 277 278)
Viruses may also be directly involved in host genetic diver-sity because they can be the agents of genetic exchangebetween microorganisms (223 273) This often involvesthe viral lifestyle known as lysogeny whereby viruses survivewithin host cells as DNA only integrated into the host chro-mosome and being reproduced each time the host divides Ahost harboring such a genome is called a lysogen becauseunder conditions of stress to the host cell a genetic switchmay cause the viral genome to initiate the lytic process pro-ducing many progeny viruses and bursting from the hostLysogeny is a very common property occurring in a signifi-cant part of the bacterial community (279 280) althoughthe incidence of induction of the lytic phase in nature is appa-rently low (281) Overall lysogeny is poorly understood butthought to have both positive and negative impacts on themicrobial community (282) Recent results suggest that ben-efits of being lysogenic in highly seasonal polar seas leads tofundamental difference between polar and other marine viralcommunities (283)
Viruses and the Microbial LoopAs part of the food web viruses occupy a unique positionThey infect host cells that are mostly thought to be heterotro-phic bacteria and by doing so they typically burst the hosts torelease progeny viruses and cellular debris But what is the fateof this material Viruses themselves do not last indefinitelyand a simple steady-state assumption implies that from eachburst of viruses (typically 20ndash100 per lytic event) only onesuccessfully infects another cell The rest are inactivatedand broken down by sunlight (UV and visible exposure)and enzymatic attack (284) or consumed byminute phagotro-phic protists (285) thus reentering the food web as substratefor bacteria or food for protistan consumers Experiments incontrolled laboratory systems and field studies with radioac-tively labeled viral lysis products have supported the conclu-sion that most of the organic matter released by the viralinfection is either taken up by bacteria or respired (286 287)
Modeling this process as part of the microbial loop showsthat viral lysis represent a sort of side loop that has the neteffect of remineralizing a significant amount of the carbonand nutrients that enter the bacteria-protist part of themicro-bial loop (Fig 7) A theoretical numerical steady-state modelcomparing a systemwith no viral activity to onewhere virusesare responsible for 50 of bacterial mortality showed that thesystem with viruses had 33 more bacterial production andrespiration than the virus-free system implying that theviruses had the effect of permitting the bacteria to processmore of the primary production than they would otherwise(223) Although 50 is a high number unlikely to be com-mon in the sea this model nevertheless illustrates that virusescan reduce the amount of energy reaching higher trophic lev-els The implication is that viruses lead to increased bacterialactivity at the expense of the larger organisms
MAJOR ENVIRONMENTAL CONTROLSLight Temperature and PressureTemperature has an important potential influence on bio-chemical reactions and therefore on biological processesin general Most ocean waters fall in the range of ndash2degC to30degC with obvious exceptions in hydrothermally heatedareas Temperature has long been known to be a regulatingfactor for the growth of heterotrophic microbes
In temperate waters it has been established that microbialactivity is generally much higher in warm summer waters thanin winter (55) The relationship is not simple howeverbecause multiple factors act at the same time Some contro-versy still exists regarding the highest and lowest extremesfor marine bacterial growth although there is broad agree-ment that bacteria grow gt100degC at hydrothermal vents andltminus5degC in sea ice brines Pomeroy et al (57) noted the inter-esting observation that bacteria seem particularly inhibitednear the freezing point of seawater (ca ndash22degC) comparedto eukaryotic phytoplankton This effect results in polarspring phytoplankton blooms that accumulate organic car-bon in advance of the response of the bacterial communityand development of the microbial loop and perhaps lead toenhanced benthic-pelagic coupling (288)
The relationship between temperature and the growth rateof marine phytoplankton was described broadly in a now
FIGURE 7 Modification of the microbial loop concept that incor-porates the functional role of viruses Export can be via predation orsinking From Fuhrman (223)doi1011289781555818821ch422f7
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-17
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
classic paper by Eppley (58) and later Goldman and Carpen-ter (289) Temperature was shown to exert a strong and directeffect on themaximal growth rates of these species withmax-imal intrinsic growth rates at 0degC generally lt1 divisiondaywhile growth rates at 30degC may be gt4day These relation-ships indicate the maximal rate that might be attained byphytoplankton at these temperatures but they do not takeother factors into account (nutrients light) While warmtemperature ostensibly allows more rapid growth it createshydrographic conditions that typically give rise to nutrientlimitation of algal growth Thus some of the coldest watersin nature witness some of the most massive phytoplanktonblooms (290) while warm oceanic gyres represent some ofthe most oligotrophic areas of the ocean The multiple andoften conflicting effects of temperature on the growth of nat-ural phytoplankton assemblages limit the accuracy of presentpredictions regarding how primary producers will respond toclimate change (291)
An analysis of the effect of temperature on the growth ofheterotrophic protists has indicated that the response is qual-itatively similar to that of phytoplankton but with a twistThe growth rates of at least some protozoa can exceed thoseof phytoplankton at warmer environmental temperaturesbut the opposite effect is apparent at very low environmentaltemperature This differential effect of temperature on thegrowth of phototrophic and heterotrophic protists was basedon a large meta-analysis of published protistan growth (59)That analysis demonstrated that the maximal growth ratesattained by phototrophic protists could exceed the maximalgrowth rates attained by heterotrophic protists (all otherpotential growth-limiting factors not considered) Thereforeprotozoan growth rates may be constrained to a greater degreeat low environmental temperature than rates for phytoplank-ton If so then phytoplankton bloomsmay get a head start ongrazers during spring in polar ecosystems This scenario is con-sistent with information on seasonal biomass changes andmicrozooplankton herbivory in the Ross Sea Antarctica(292 293) but there are still too few data to fully vet thishypothesis
The importance of high pressure on bacterial growthgained considerable attention in the late 1960s when thedeep sea submersible Alvin was accidentally lost overboardwith its hatch openWhile no lives were lost in this accidentsome workmenrsquos lunches sank to the bottom (sim1500 m)inside the submersible Alvin was recovered after 10 monthsand interestingly there was a waterlogged lunch containingapples bologna sandwiches and broken vacuum bottleswith broth that all appeared hardly degraded and tasted palat-able Yet when placed in a refrigerator on the ship these itemsdegraded relatively quickly Initially pressure was thought tobe the preserving factor as the sea floor temperature was sim-ilar to the refrigerator temperature and the only major differ-ence would be pressure (294) Following this observationa series of experiments to measure degradation of variousorganic materials left in the deep sea for extended periodsindicated that degradationwas typically significantly reducedimplying that pressure reduces the degradation rates (295)Nonetheless changes in the protozoan community of naturaldetrital material sinking to the deep ocean floor indicate thatthe microbial community can respond relatively quickly insome situations (296) Deep sea microbiology has advancedconsiderably yet it is still difficult to interpret results withrespect to actual in situ rates of naturally occurring organicmatter Deep sea bacteria adapted to high pressures havebeen isolated that are barophilic (also called piezophilic)meaning that they prefer high pressures and have reduced
activity at lower pressures whereas others are barotolerant(piezotolerant) tolerating but not preferring high pressuresfor example see (297 298)
Few data are available on barotolerantbarophilic marineprotozoa Protozoa certainly exist and grow at great oceanicdepths but measurements of in situ growth rates for these spe-cies do not yet exist Measurable protozoan numbers havebeen documented in the deep-sea sediments for more than30 years (299 300) and viable protozoa have occasionallybeen cultured from these environments (301ndash305) butvery few direct measurements of the activities of these speciesin situ have been reported (306) A few protozoa have beenisolated that will grow at high pressure (302 303 307) andprotists that appear to be unique to the deep ocean havebeen observed either directly or through the analysis ofDNA sequences (308ndash311) but possibly the best direct evi-dence that protozoan activity takes place at the high pressurescharacteristics of the deep sea are experimental and observa-tional work noting the stimulatory effect that detrital deposi-tion has on some components of the protozoan community(296 312) These observations indicate a diverse and activeprotozoan fauna of the deep ocean although their biogeo-chemical significance is largely uncharacterized at this time
Dissolved and Particulate Organic MatterBacteria and archaea are thought to be by far the most impor-tant organisms with respect to the processing of dissolvedorganic matter (DOM) and nonliving particulate organicmatter (POM also called detritus) in the ocean While theremay be some uptake of DOM by protists particularly forgrowth factors needed in trace amounts such as vitamins(313) the bulk of this material is probably utilized by bacteriaand archaea (314) including the smallest cyanobacteriumProchlorococcus (315 316) which thus may be considered amixotroph Due to their small size bacteria have extremelyhigh surfacevolume ratios and combined with their over-whelmingly high abundance an extremely high integratedsurface area Protozoa tend to obtain the organic materialsthat they require for growth from their prey rather thanthrough the uptake of DOM Overall protozoa tend to besources of dissolved and detrital organic substances throughthe excretion of unassimilated prey biomass in expelledfood vacuoles
Particulate organic matter is not directly available as sub-strate to bacteria These substances must first be reduced tosmall molecules that can be transported into the cell Thisis accomplished by the production of extracellular enzymes(note that few if any large polymers are directly taken up bybacteria with the possible exception of DNA) Hydrolyticenzymes produced by bacteria (and almost certainly archaea)break down polymers like proteins polysaccharides andnucleic acids As with DOM POM is composed of a complexmixture of compounds that vary in their susceptibility to bac-terial degradation and utilization
Particulate material in the water column serves not only asbacterial substrate but also as substratum POM occurs in thewater column across a huge size spectrum from micrometersup to some detrital aggregates more than 1 m in diameter(317) Much of this particulate material is in a constant stateof flux with colloidal material constantly coalescing andaggregating to form new or larger particles (318) as microbialdegradation acts simultaneously to remineralize this materialDetrital particles that attain macroscopic size either by directformation (317 319 320) or via accretion and aggregation(321) and are often called marine snow or macroaggregatesMarine snow particles are readily colonized by bacteria and
422-18 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
many other microorganisms and their abundances on theseaggregates can be orders of magnitude greater than abundan-ces in an equivalent volume of the water surrounding theaggregates (320 322ndash324) and their diversity and highlyconcentrated abundances on detrital aggregates implies thatthere may be stronger trophic coupling and efficient energyelemental cycling relative to these processes among free-living microorganisms in the surrounding waters This specu-lation has resulted in a number of formalizations of the poten-tial impact of aggregate microbiology on biogeochemicalprocesses in the ocean (325 326) Most definitions of bio-films include aggregated particulate and these particles areoften studied within the biofilm conceptual model (see chap-ter by Lawrence et al in this section)
Inorganic Nutrients
Macronutrients (N P)Virtually all heterotrophic organisms in the ocean contributeto the pool of available macronutrients via the excretion ofmetabolic wastes and protozoa contribute significantly tothis process Macronutrients are often in excess within proto-zoan prey relative to the consumersrsquo needs because much ofthe prey carbon is respired to produce energy for metabolismand growth ldquoExcessrdquo nutrients are eliminated as eitherorganic compounds or more commonly as ammonium (forN) and phosphate (for P) The importance of the match(or mismatch) between predator and prey stoichiometry hasbeen demonstrated experimentally and repeatedly in marineand freshwater ecosystems and has been elevated to a funda-mental ecological principle governing elemental and energyflow in aquatic food webs (327) Based on this reasoningCaron et al (328) concluded that bacterivorous protozoaplay an important role in nutrient remineralization andrelease in the ocean Bacterial biomass is typically rich in Nand P relative to other organisms so bacterial predators expe-rience large excesses of these elements relative to their growthneedsMoreover bacterivorous protozoa should be dispropor-tionately important in nutrient remineralization amongsingle-celled eukaryotes because as the smallest protozoa inaquatic ecosystems they have high weight-specific metabolicrates (329) Thus both stoichiometric and allometric rela-tionships implicate small bacterivorous protozoa as impor-tant sources of remineralized nutrients in the plankton
The reasoning also holds true for bacterial utilization oforganic compounds and it has led to some interestingfindings and conclusions regarding the role of bacteria innutrient cycling in the ocean Bacteria are important sourcesremineralized nutrients when they are consuming N- orP-rich substrate Contrary to this traditional view howeverheterotrophic bacteria are often strong competitors for inor-ganic nutrients under many situations largely because bacte-rial biomass is richer in N and P than much of the organicmatter they consume in nature so they require additional Nand P to produce their biomass (summarized by [330]) Exper-imental studies have demonstrated the uptake of ammoniumby bacteria other than cyanobacteria using stable isotope 15N(331) and also the short-lived radioisotope 13N (332) oftenfinding that bacteria may be responsible for one third of the Nor P uptake Additionally there is evidence that bacteria insome oceans like the Sargasso Sea are limited by phosphorus(333 334) Growth limitation of bacteria by phosphorusappears to be a common phenomenon of freshwater ecosys-tems (335 336)
N2 fixation is restricted to prokaryotes and in the oceanwater column it was thought for many years to be done
primarily byTrichodesmium a warm-water colonial cyanobac-terium that blooms sporadically andRichelia a cyanobacterialsymbiont that lives within certain diatoms (337) Some uni-cellular cyanobacteria also contribute significantly to globalN2 fixation (338) and molecular biological data suggestthat a variety of other bacteriamdashnot phytoplanktonmdashmayalso be fixing nitrogen in seawater (339) An exciting recentdiscovery in this field is that a bacterial nitrogen fixer origi-nally known only from nifH sequences UCYN-A has hadits genome sequenced by parallel pyrosequencing and wasfound be a cyanobacterium (Candidatus Atelocyanobacteriumthalassa) lacking the apparatus to generate oxygen as well asother pathways (340) Further study showed it to be symbioticwith a marine alga (341)
Micronutrients (Trace Metals Growth Factors)Bacterial growth may often be limited by macronutrients asdescribed and sometimes by trace nutrients particularlyiron that occur at very low concentrations in seawater(342 343) The importance of Fe as an element limiting pri-mary production in some oceanic regions has come underclose scrutiny and extensive experimental investigation inrecent years Many bacteria have special uptake mechanismsto utilize extremely low concentrations of iron includingsiderophores (released compounds that bind iron and arethen specifically taken up) Even if bacteria are capable ofgrowth at low Fe concentrations growth may be less efficientKirchman et al (344) found that Fe limitation stronglyreduced bacterial growth efficiency in a cultured marine bac-terium (ca 50 efficiency Fe replete but lt10 when Fedeplete) Thus the effects may be complex Marine N2 fixa-tion also may be limited by the availability of Fe becausenitrogen fixers have high Fe requirements
Protozoa tend to be a source rather than a sink for Fe in theocean analogous to their role in macronutrient remineraliza-tion Protozoan grazing activity has been shown to releaseiron from bacterial biomass thereby relieving Fe limitationin co-occurring phytoplankton (345) a mechanistic demon-stration of the process using protozoa growing on Fe-repletebacteria However it has also been reported that Fe-limitedbacteria may contain insufficient Fe for the bacterivorous pro-tozoa that consume them thus leading to Fe limitation in theprotozoan as well (346)
OxygenHypoxic and anoxic regions of the ocean are expected to sig-nificantly expand in the coming decades as a consequence ofchanges in ocean stratification resulting from global climatechange (347) As oxygen becomes depleted bacteria havethe capability to utilize a series of alternate electron acceptorsCompositional changes in the bacterial assemblage accom-pany these changes in metabolic activity but are beyondthe scope of this chapter Many of the studies on benthicmicrobiology have focused on redox reactions that occurthere Oxygen is usually consumed by sediment microbialactivity much faster than the rate at which it can be replacedby diffusion so most sediments especially organically richones are anaerobic below a few mm (fine-grained muds) orcm (coarser sediments) from the sedimentwater interfaceexcept where animals ventilate the benthos through tubesWhen oxygen is absent bacteria and archaea use alternateelectron acceptors such as nitrate nitrite oxidized Fe orMn organic matter or sulfate The use of nitrate or nitriteas an electron acceptor with concomitant production of N2
gas (denitrification) results in net loss of biologically availablenitrogen Bacteria that perform this reaction are typically
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-19
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquofacultativerdquo organisms that can switch from oxygen to nitrateor nitrite Organic matter may be reduced by fermentationreactions in which part of an organic substrate is oxidizedwhile another is reduced fermenter organisms are also typi-cally facultative Sulfate is a particularly important alternateelectron acceptor in marine systems because it occurs as thesecond most abundant anion in seawater (behind chloride)typically 28 mM Its use as an electron acceptor leads to theproduction of elemental sulfur or sulfide The use of sulfateas an electron acceptor yields much less energy than oxygenor nitrate and sulfate-reducing bacteria are strict anaerobesunable to use oxygen (often strongly inhibited by oxygen)Methane can be produced by methanogenic archaea whichare all strict anaerobes using CO2 as an electron acceptor(see Findlay and Battin this section for further discussion)
Reduced compounds such as sulfide are produced by theuse of these alternate electron acceptors and they diffuse awayfrom their source When they diffuse to environments with astrong oxidant like oxygen they may be oxidized by chemo-trophic bacteria in energy-yielding reactions Similarlyammonium can be oxidized to nitrite and then nitrate by che-motrophic nitrifying bacteria and the nitrate can diffuse intoanaerobic layers and become denitrified to N2 Such coupledoxidation-reduction reactions can lead to accelerated biogeo-chemical cycling particularly in sediments
It was originally believed that relatively few free-livingspecies of protozoa inhabit anaerobic and hypoxic marineenvironments This view has changed based on (largely)molecular analyses of some of these environments (308348ndash351) A significant number of previously undescribedeukaryotic phylotypes have been observed in anoxic environ-ments High protozoan biodiversity in the absence of molec-ular oxygen is not completely unexpected because otheranaerobic environments such as some gut environments(eg cockroach guts and cattle rumen) have notoriouslyabundant and diverse protozoan fauna but marine anaerobicenvironments have been difficult to access and sample prop-erly Many anaerobic protozoa exist together with symbioticbacteria (352 353) or possess unique organelles such as thehydrogenosome (354)
MAJOR BIOTIC CONTROLSThe Trophic Activities of Protozoa Predation andTop-Down ControlsPredator-prey relationships are fundamental interactionswithin biological communities linking production of bio-mass on the one hand with removal on the other hand Estab-lishing the trophic relationships among microorganisms andquantifying the rates at which they take place constitutemajor endeavors within marine microbial ecology As notedpreviously protozoa consume a wide variety of prokaryoticand protistan prey Numerous laboratory and field measure-ments have been made throughout the past quarter centuryto attempt to quantify the rates of bacterial and phytoplank-ton consumption The methodology for accomplishing thesestudies has varied and a number of approaches are presentlyemployed
Methodologies for estimating microbial grazing are diversebut generally fall into one of two experimental approachesperturbation experiments or tracer experiments Perturbationexperiments (size fractionation metabolic inhibitors anddilution technique) rely on manipulation of the consumeror prey assemblages in somemanner such that the effect is evi-dent from changes in the abundance of the prey population
during incubations relative to control treatments that receiveno manipulation Tracer experiments (radioactively labeledprey fluorescently labeled prey stable isotopendashlabeled pray)attempt to use labeled prey or a prey proxy to quantitativelyfollow the movement of prey biomass into consumers Forsummaries of these methods see (74 355)
Perturbation experiments have entailed a variety of mech-anisms to ldquodecouplerdquo predators from prey in natural microbialcommunities For example size fractionation uses various sizefilters to remove an entire size class of grazers from awater sam-ple leaving the prey to grow in the relative absence of thosepredators during subsequent incubations Unfiltered samples(controls) indicate the growth of prey in the presence of pre-dation Predation impact is obtained by the difference ingrowth of the prey between these treatments This methodhas been used to examine predation on cyanobacteria heter-otrophic bacteria and minute phototrophic eukaryotesAlternatively metabolic inhibitors have been employed tohalt predation (or in some cases prokaryote growth) insteadof physical removal by filtration
The dilution technique (356) has become a commonmethod for examining grazing mortality of phytoplanktonby microzooplankton (which is comprised predominantly ofprotozoa) This method relies on the dilution of herbivorouszooplankton in a series of subsamples (and concomitantmeasurements of phytoplankton growth in each subsample)to estimate the grazing mortality of the herbivores
Tracer experiments have used both radioactively labeledprey and fluorescently labeled prey to examine predation onbacteria and phytoplankton Radioactive labeling has beenaccomplished using 14C- or 3H-labeled organic compounds(for bacteria) or 14C-bicarbonate (for phytoplankton) Theuse of radioisotope approaches has diminished with improve-ments in the number and types of fluorescent compoundsavailable for this work Fluorescently labeled prey havebeen labeled with a variety of compounds that allow theprey to be observed readily (even inside predators) by epi-fluorescencemicroscopy (357 358) Alternatively a decreasein the total number of fluorescent prey in a sample indicatesthe rate of removal of the prey assemblage
Both perturbation experiments and tracer experimentsdescribed have inherent advantages and disadvantages Forexample perturbation experiments are fairly straightforwardto conduct but they may introduce artifact by removingpotentially important sources of nutrients or organic com-pounds released by predators (359) On the other hand thedilution technique is very labor-intensive but one advantageof this method is that it provides the opportunity to determineboth phytoplankton growth and grazing mortality in a singleprocedure Most tracer experiments are fairly easy to conductand generally do not perturb the food web being studied butfeeding selectivity for or against the tracer (relative to naturalprey) may produce spurious results In addition some of thesemethods provide a measurement of the activity of the entiregrazer assemblage and others are applicable for examiningthe activity of specific taxa For example size fractionationapproaches examine the effect of the removal of an entiresize class of organisms (a community-levelrdquo measurement)whereas fluorescently labeled prey can be used to examinethe ingestion rate of a particular species by observing theuptake of labeled prey into that species (a species-levelmeasurement)
Predation rates of numerous marine protozoa have beenexamined in the laboratory From these experiments a gen-eral knowledge of the rates of consumption by these specieshas been accumulated Feeding in most protozoa shows a
422-20 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
functional response to the availability of prey that is similar tothe response of many organisms (although there are a greatdeal of species-specific differences) That is ingestion rateincreases rapidly as prey abundance increases until a maximalingestion rate is attained (360)Manyof these curves have theform of a Type II functional response curve (361)
Based on the features of these curves it is clear that proto-zoa have the ability to ingest substantial numbers of prokar-yote and eukaryote prey in natural ecosystems Somebacterivorous flagellates can consume bacteria (and cyano-bacteria) at rates up to 100 s per individual per hour whereassome bacterivorous ciliates can consume thousands of bacte-ria per individual per hour Rates of consumption of eukary-otic phytoplankton by herbivorous ciliates and largeflagellates are considerably more variable and highly depend-ent of the feeding mechanism (eg filtration in ciliates versuspseudopodial net in dinoflagellates) and the size of the prey
These lab studies demonstrate that rarely are prey suffi-ciently abundant in nature to support the maximal ingestionrates of which many protozoa are capable Nevertheless pre-dation rates of protozoa in nature are sufficient to constitute amajor factor controlling the standing stocks of bacteria andphytoplankton (63 180 184 355) For example these sum-maries indicate that on average protozoa consume 20ndash80ofthe standing stock of phytoplankton (measured as chloro-phyll) in the world ocean and 60ndash75 of daily primary pro-duction a departure from the classical view of copepods as theprimary consumers of phytoplankton This large role of her-bivorous protozoa infers a large role for copepods as consumersof planktonic protozoa (185) Similarly the role of protozoaas major consumers of bacterial biomass is indicated by therelative balance between bacterial production on the onehand and bacterivory on the other (Fig 8) The approximateone-to-one balance between these measurements over a largenumber of studies implies that protozoan bacterivory must bea major factor determining the fate of bacteria in marine (andfreshwater) plankton ecosystems
It is interesting to note that some types of bacteria includ-ing marine Bdellovibrio are known to prey on other marine
bacteria (362) At the time of this writing the importanceof bacterial predation by other bacteria within the contextof oceanographic processes is not known and presumed tobe small While Bdellovibrio may be cultivated from seawaterit can also be found among sequences from cold sediments(363) and the water column Bdellovibrio happens to be thebest studied predatory bacterium but there are no doubtothers that we simply have not yet identified (364)
Species-Specific Interactions (MutualismParasitism Commensalisms)Interactions between marine microorganisms such as compe-tition allelopathy and symbiosis are fundamental aspectscontrolling community composition and activity but atpresent most are poorly characterized in natural marine sys-tems Bacteria must compete for dissolved and particulatecompounds with other bacteria andwith protistan taxa whilephagotrophic protists must compete with each other andmetazoa for available prey These interactions manifest them-selves in many forms including true competition for availableresources resource partitioning such that some taxa specializeon the utilization of certain substrates or prey while othersspecialize on different ones (reducing competition) or eventhe cooperative use of compounds (eg one bacteriumrsquoswaste products being utilized as substrate by another bacte-rium) These metabolic consortia are probably common inthe ocean but can be difficult to identify and characterizeConsortia of bacteria working together to perform complexreactions especially in anaerobic systems are well known inmicrobiology
Consortia in which there is a very high degree of integra-tion between species (ie symbiosis) are very common in theocean These associations may be beneficial to both partners(mutualism) beneficial to one part and inconsequential forthe other (commensalism) or beneficial to one part and det-rimental to the other (parasitism) Many highly visible andwell-characterized marine mutualisms exist between chemo-synthetic bacteria and animals such as hydrothermal ventor methane seep tubeworms clams mussels or other inverte-brates (365) and also between luminescent bacteria and fishor squids (366) Similarly many protozoa form mutualisticassociations with photosynthetic protists (367) The mostconspicuous of these associations involved large amoeboidprotozoa of oceanic ecosystems (planktonic foraminiferanspolycystine radiolaria and acantharia) which contain largenumbers of intracellular algae (74) but associations betweensome heterotrophic dinoflagellates and cyanobacteria are alsocommon (368) Studies have indicated that efficient nutrientcycling in these protozoan-algal associations confers an eco-logical advantage for both partners in surfacewaters of the oli-gotrophic ocean
Parasitism is also a common symbiotic relationship amongmarine protozoa Protozoan parasites have been reported for adiverse array of protists and animals in the ocean Heterotro-phic dinoflagellates and presently uncultured alveolate cladesrelated to alveolates appear to be particularly successful atevolving thismode of existence (369ndash371) In a common par-asitic group Amoebophrya spp the parasite invades photo-synthetic dinoflagellates where it undergoes reproductioneventually lysing the host and releasing dozens of infectiveswarmers The parasitic protist Pirsonia spp is a parasite ofdiatoms (372) The overall importance of these parasitismsin the population dynamics of their hosts is not clearly knownbut may constitute a significant source of mortality for somespecies (373 374) The frequency and diversity of ldquonewrdquo
FIGURE 8 Meta-analysis of experimental studies that have com-pared rates of bacterial productivity (converted to population growthrate units of dminus1) with rates of bacterivory by phagotrophic protists(expressed as equivalent units of bacterial growth rate dminus1) Thesolid line indicates where measurements of bacterial removal by graz-ers were equal tomeasurements of bacterial production Data redrawnfrom Sanders et al (63) doi1011289781555818821ch422f8
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-21
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
bacterial mortality via grazers or viral lysis on the other handIt is consistent with some measurements of extremely rapidturnover sometimes several times per hour in rich coastalwaters of labile dissolved organic compounds such as dis-solved free amino acids (56) and might also indicate greaterpredation pressure by protozoa during the night On longertime scales of weeks to months bacteria show distinct sea-sonal patterns For example in temperate coastal waters bac-terial biomass and production increase considerably insummer months compared with winter However bacteriado not typically show a significant increase during earlyldquospringrdquo blooms in temperate waters when water is still verycold (51) It has been hypothesized that this phenomenonis the result of the suppression of the rate of substrate uptakeby temperate bacterial assemblages at low temperature (57)However while temperature probably has the effect of settinga limit on maximal growth rates (as for phytoplankton [58]and protozoa [59]) temperature alone does not appear to bethe main factor controlling growth of marine bacteria undermost circumstances (see ldquoLight Temperature and Pressurerdquo)
It would be overly simplistic to think that all the bacteriaand archaea in a sample or habitat have the same level ofactivity per cell but it is also easy to think of measured activ-ities as characteristic of all members of a microbial assem-blage So the question arises are most of the cells active ata similar level or are some hyperactive while others are com-pletely dead or moribund This question has been addressedseveral ways includingmicroautoradiography selective stain-ing ldquodirect viable countsrdquo (where nutrients are added to seewhat part of the community grows) and in situ hybridization(eg 60) Based on these contrasting approaches it appearsthat a continuum of activity exists within bacterial assemb-lages from truly dead (cannot be revived) to extremely activeA reasonable interpretation of the existing data is that undertypical conditions a small percentage of the marine bacterialcells perhaps 10ndash20 are generally inactive or dead the plu-rality or majority of cells perhaps 25ndash75 are intact andhave some moderate level of activity and a small percentageperhaps 5ndash20 are highly active It is useful to consider thisspectrum conceptually and numerically when modelingmicrobial processes
Comparisons of bacterial and phytoplankton biomasswithin planktonic ecosystems show that these are positivelycorrelated across broad scales Analyses of marine and fresh-water samples from several studies (61 62) have shown thatbacterial abundance increases with chlorophyll concentra-tion at least at the level of a log-log relationship Similarlybacterial abundances and the abundances of small protozoacorrelate over broad spatial and temporal scales (63) Theserelationships are sensible in that on the largest scale primaryproduction is the source of organic material that fuels hetero-trophic bacterial activity and bacteria constitute the prey ofmany small protozoa Individual data sets also have sometimesshown strong correlations between bacterial abundance andchlorophyll (eg 45) but variability in this relationshipover short temporal or spatial scales is to be expected It wouldpresumably be a consequence of rapid short-term changes inthe rate of substrate supply as well as the normal oscillatorynature of predator-prey relationships between bacteria andtheir consumers
Interestingly the extrapolation of the positive log-log rela-tionship between bacterial and phytoplankton biomassto environments with very low chlorophyll concentrations(eg ultra-oligotrophic oceans) indicates that bacterial bio-mass may exceed phytoplankton biomass in these situationsWhile this conclusion is obvious for the deep sea where there
is no photosynthesis this situation is also often true in oligo-trophic surface waters Measurements made in oligotrophicwaters bear out the high bacterial contribution to totalbiomass (64 65) Moreover Cho and Azam (66) confirmeda linear relationship between the log of chlorophyll and logof bacterial abundance but only at chlorophyll concen-trations above approximately 05 μg per liter Below thatconcentration bacterial abundance did not correlate signifi-cantly with chlorophyll It should be noted that subsequentanalyses have revealed that early epifluorescence measure-ments of bacterial biomass included the common cyanobacte-rium Prochlorococcus which can make up to 20 of totalbacterial numbers (67) Nonetheless heterotrophic bacterialbiomass is a major fraction of the living biomass of all plank-tonic ecosystems
The geographical and temporal distributions of marineprotozoa are much more varied than those of the bacteriaAs an all-inclusive group protozoa generally occur in plank-tonic ecosystems at abundances ranging from 10 s to 1000 sper ml Abundances in benthic ecosystems can be one tothree orders of magnitude higher commensurate with thehigher abundances of bacteria in those ecosystems Howeverit is important to remember that like the term ldquolsquobacteriardquo theterm ldquoprotozoardquo is a rather artificial conglomeration of evolu-tionarily and ecologically divergent taxa (see ldquoThe Changingand Complex World of Eukaryote Phylogenyrdquo) Thus theabundances of specific lineages of bacteria or protozoa mayshow spatial (or temporal) variability that is considerablygreater than the variability characteristic of these overarchinggroupings
The Changing and Complex World ofEukaryote PhylogenyNot that long ago textbooks still divided eukaryotic organ-isms into four major kingdoms (Animalia Plantae Fungiand Protista) while prokaryotic organisms were placed intoa single kingdom theMonera (68)Within this scheme pro-tists (eukaryotic organisms that can exist as single cells) weredivided into two subkingdoms (algae and protozoa) based ontheir basic nutritional mode a carryover from the historicaldistinction between single cells with ldquoanimal-likerdquo or ldquoplant-likerdquo nutrition This distinction presupposed a basic evolu-tionary divergence among protists into species that retaineda heterotrophic phagocytotic mode of life (protozoa) andthose that abandoned phagocytosis for a photosyntheticmode of life (algae)Moreover the presenceabsence of chlor-oplasts was a feature that could be easily distinguished by earlymicroscopists
The five-kingdom classification system of Whittaker wasrecognized as an improvement over previous classificationschemes but it posed a number of problems relating toprotists For example the distinction between single-celledand multicellular eukaryotes was somewhat arbitrary Moreimportant the division of protists based on whether theywere heterotrophic or photosynthetic was clearly not anappropriate feature if the classification was to recapitulateevolutionary relationships We now know that chloroplastacquisition and loss has occurred several times in the bio-logical history of our planet (69) giving rise to some closelyrelated protistan taxa that differ largely in the presence orabsence of a chloroplast Further complicating the matterwithin many protistan lineages there are species that possesschloroplasts and carry out photosynthesis (phototrophy)but also possess the ability to ingest and digest prey (hetero-trophy 70ndash72) Some heterotrophic protists even ingest
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-5
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
phytoplankton prey and retain the chloroplasts of their prey ina functional state for a limited amount of time (kleptidoplas-tidy73) Various forms and degrees of mixotrophy (mixedphototrophic and heterotrophic nutrition) are commonamong a number of algalprotozoan lineages (74ndash77) UnderWhittakerrsquos scheme phytoplankton ecologists studying a lin-eage of microalgae might have had little familiarity withclosely related heterotrophic species while protozoologistsstudying a particular protozoan groupmight have known littleabout closely related photosynthetic species
One might expect given these caveats that the termsldquoalgaerdquo and ldquoprotozoardquo are no longer used In fact the termldquoprotozoardquo is still commonly used (especially by ecologists)to recognize those eukaryotic species that exist as single cellsand whose nutrition is dependent on the uptake of preformedorganic substances (primarily via prey ingestion) while pro-tists possessing chloroplasts are still commonly called ldquoalgaerdquoSimilarly although the term ldquoprotistrdquo has been abandoned asa kingdom designation it is still widely employed to describeeukaryotic species that are capable of existence as single cells(ie algae and protozoa) The term ldquophagotrophic protistrdquohas also gained popularity in recent years because it recognizesthat many protistan species are capable of phagocytosis eventhough they may also possess their own chloroplasts and thusare technically ldquoalgaerdquo
Despite the shortcomings of Whittakerrsquos scheme it domi-nated the hierarchical organization of life for approximately aquarter century During the past few decades however thissystem has given way to a new organizational scheme that rec-ognizes three domains of life (Archaea Bacteria Eukarya [orEucarya] 78 Fig 2 upper panel) and is based onwhat is pres-ently believed to reflect amore realistic view of the evolution-ary distances that have developed between organisms in theasymp4 billion years that life has existed on our planet Withinthe Eukarya of Woesersquos scheme hypotheses regarding thephylogeny of ldquoprotistsrdquo have changed continuously and rap-idly during the past two decades reflecting new insightsinto eukaryote evolution provided largely by DNA sequenceinformation (Fig 2 lower panel from [79])
The former protistan phyla ofWhittakerrsquos systemhavenowbeen dispersed among candidate ldquosupergroupsrdquo within thedomain Eukarya to better reflect hypothesized phylogeneticrelationships For example the dinoflagellates (which encom-pass phototrophic heterotrophic and mixotrophic species)now form a single group and have been placed together withthe ciliates and apicomplexans (sporozoans) in themonophy-letic Alveolata (Fig 2 lower panel) On the other handeukaryotic heterotrophic single-celled species falling withinthe general description ldquoprotozoardquo are now widely distributedamong a number of protistan lineages In short nutrition hasbeen demoted as a phylogenetic character and other charac-ters (presumably more indicative of evolutionary relatedness)have ascended to address some long-standing contradictionsalthough the debate over the relationships among some line-ages is still very active at the present time 79)
Individual protozoan cells range in size from less than 2 micromto greater than 1 cm in diameter (gt4 orders of magnitude)(80 81) with some colonial radiolaria forming cylindricalgelatinous structures a centimeter in diameter and morethan a meter in length (82 83) Because they constitutesuch a large size range of organisms protozoa are often dividedinto size classes that very crudely correlate with their generalnutritional preferences A commonly used convention is thatof Sieburth et al (84) which groups planktonic microorgan-isms into order-of-magnitude size classes (02ndash20 microm =picoplankton 20ndash20 microm = nanoplankton 20ndash200 microm =
microplankton 02ndash2 mm =mesoplankton) Most protozoafall into the nanoplankton or microplankton size classesModeling microbial trophodynamics using this conventionassumes that protozoa in one size category generally consumeprey one order of magnitude smaller in size (34 85)Although this approach misses much of the detail and diver-sity of the trophic activities of individual protozoan taxa it isa necessary practical compromise for examining community-scale flows of energy and elements It also provides a usefulmechanism for summarizing and comparing the abundancesand biomasses of protozoa from different environments andto other microbial assemblages Protozoan abundance sum-marized in this way has been shown to contribute signifi-cantly to the living biomass of planktonic ecosystemsthroughout the world ocean (Fig 3)
Estimates of protozoan biomass such as those depicted inFig 3 typically do not include the contribution of mixotro-phic phytoflagellates to heterotrophy There is presently noeasy way to determine the abundances of small phagotrophicphytoflagellates in natural samples so these species are typi-cally counted as phytoplankton unless specific methods areemployed to identify the algae as consumers such as the useof fluorescently labeled particles (87ndash90) or through theexamination of food vacuole contents (91) On averagethese species appear to constitute a modest percentage ofthe phytoplankton assemblage (typically lt25) althoughthey may at times dominate the phototroph assemblages ofnatural plankton communities It is important to recognizethat their inclusion as functional heterotrophs rather thanphototrophs could significantly shift the relative contribu-tions of phototrophic and heterotrophic microbial biomassto total biomass within microbial assemblages and the flowof energy within plankton communities (92)
Heterotrophic protists that harbor photosynthetic pro-tists or their chloroplasts within their cytoplasm constituteanother complexity for estimating the contribution of pro-tozoa to total microbial biomass When bulk water samplesare analyzed the contribution of chlorophyll containedwithin those protozoa is generally assumed to come fromfree-living phytoplankton However studies have shownthat chloroplast-bearing ciliates can contribute up to halfthe total biomass of planktonic ciliates in ecosystems andchloroplast-retaining ciliates can sporadically dominate thechlorophyll and primary production of some planktonic eco-systems (93ndash96) The environmental conditions promotingthe success of these ciliates are poorly known Similarlymany species of planktonic foraminifera polycystine radio-laria and acantharia harbor large numbers (thousands perprotozoan) of endosymbiotic algae within their cytoplasm(97 98) Caron et al (99) have demonstrated that primaryproductivity within these species can contribute significantlyto total primary productivity in oceanic ecosystems and can bevery important locally in the convergences of Langmuir circu-lation cells (100)
BACTERIA AND ARCHAEAldquoCulturablerdquo versus ldquoNonculturablerdquo CellsMost conventional cultivation methods can grow only 1 orless of the bacteria that can be visualized by direct microscopytechniques (eg 4) This is true even though most can beshown to be active by techniques such as microautoradiogra-phy (25) These readily cultivable organisms appear to repre-sent a group of fast-growing so-called weeds that are adaptedto take advantage of rapid growth in rare organically enriched
422-6 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
environments This strategy contrasts with the numericallydominant bacteria that are adapted specifically for growthin the dilute nutrient conditions that characterize the vastmajority of the volume of the water column
The most common taxa readily cultured from seawaterwith standard nutrient broth media include the gamma pro-teobacterial genera Vibrio Alteromonas PseudoalteromonasMarinomonas Oceanospirillum Shewanella (usually isolatedfrom surfaces such as shellfish and sediments) the alpha pro-teobacterial generaRoseobacterSphingomonasmembers of thefamily Flavobacteriaceae and Planctomycetes as summarizedin Giovannoni and Rappeacute (101) and Fuhrman and Hagstrom(102) The cyanobacteria Synechococcus and Prochlorococcusare also now readily culturable but on low-nutrient inorganicmedia targeting photosynthetic forms as opposed to organicmedia used to cultivate the others listed above
Because of the low percentage of marine bacteria that canbe grown in standard media organisms that until recentlywere called ldquononculturablerdquo make up the large majority ofbacteria in the plankton Only during the pastsim20ndash25 yearshave molecular biological methods based on 16S rRNA genesequences been available to identify these organisms andthese powerful techniques have opened up a large area forexploration (see next section) Similar but more recent stud-ies use 18S rRNA sequences for characterizing protistandiversity as will be noted below
Molecular Phylogeny and MetagenomicsField ApplicationsModern phylogeny of microorganisms is based primarilyon genetic sequences the most well-studied gene being the
FIGURE 2 The three domains of life (upper left) as proposed by Woese et al (78) and a recent overview of modifications that have beenproposed by Adl et al (79) to higher-level phylogentic groups within the eukaryotic component of the tree (lower right) Domains figure fromWoese et al (78) eukaryotic tree figure from Adl et al (79) doi1011289781555818821ch422f2
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-7
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
small subunit ribosomal RNA gene (16S rRNA in Bacteriaand Archaea and its larger homolog 18S rRNA in eukar-yotes) This molecule is strongly conserved over evolutionarytime so this single molecule has been used for constructingphylogenetic trees of all living organisms (httptolweborgtreephylogenyhtml) Analysis of 16S18S rRNA genesequences has been used to evaluate deep evolutionary rela-tionships among organisms and was instrumental in point-ing out that Archaea Bacteria and Eukarya should beconsidered different Domains of equivalent phylogeneticrank above kingdoms (78) However there are sufficientdifferences in 16S18S rRNA gene sequences to demonstrate
relationships (ormake distinctions) at the genus or sometimesspecies level
The first phylogenetic studies based on 16S18S rRNAgenes used sequences derived from cultures However onedoes not need cultures to obtain rRNA gene sequences (orany other sequences for that matter) An idea developed inthe lab of Norman Pace in themid-1980s involved extractionof DNA directly from natural samples and then cloning andsequencing of the DNA as a means of assaying the microbespresent in the samples (103 104) The original protocolscalled for cloning by creating what are called ldquophage librariesrdquofrom the natural DNA but since 1986 PCR has been applied
FIGURE 3 (a b) Plankton biomass in the Arabian Sea during the 1995 southwest monsoon (a) and intermonsoon period (b) Areas of theboxes indicate the relative magnitudes of the biomass in each category Categories within the dashed boxes in a b c are composed of protozoaArrows indicate the direction of energymaterial flow in the food web thicker arrows depicting greater flow Redrawn from (86)(c) Depth-integrated biomass (mgm2) in the upper 100 m of the Sargasso Sea near Bermuda and in the upper 200 m of the equatorial Pacificat 175degE Thewidth of the bars indicates the biomass in each size category Heterotrophs have been separated by size class while phytoplanktonhave not Size classes delineated by the dotted box are comprised of protozoa Redrawn from (65) doi1011289781555818821ch422f3
422-8 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
extensively for cloning and related studies The targetsequence can be almost instantly ldquoidentifiedrdquo to its closestphylogenetic neighbor by what have come to be standardonline sequence comparisons
Beyond the study of targeted genes (like the 16S rRNAgene) shotgun metagenomic studies have examined theentire genetic repertoire of the microbes in a given sampleThe metagenome is the collective genome of all organismsin the sample Initially these studies extracted DNA fromall organisms in a sample (usually prefiltered through a filterapproximately 1 microm to remove most organisms larger thanbacteria) sheared it to produce fragments and clonedthem into standard vectors either as small (thousands ofbases) or large (to hundreds of thousands of bases) insertsthat is fragments of DNA from the environment nowcloned into the vectors in a form suitable for sequencingThe best known early marine study of the former type isthe Global Ocean Survey with initial results published byVenter et al (105) which generated more than a billionbases of DNA sequence and reported 12 million previouslyunsequenced genes estimated to come from at least 1800different genomic species cumulatively in the many samplesthey analyzed
With the advent of next-generation sequencing that gen-erates millions ormore sequences in a run (known by a varietyof acronyms including 454 Illumina SOLiD etc) cloningof genes has largely been replaced by clone-free sequencingThe extent and power of such sequencing has recentlybeen demonstrated by the ability to construct essentiallythe entire genome of an uncultivated marine Group II Eur-yarchaeon that constituted only approximately 2 of amicrobial community sample using SOLiD sequencingmade possible by high coverage and the use of mate pairsequencing ofsim3000 bp fragments (106) (this length isneeded to span repeats and highly conserved genes that oth-erwise make genome construction difficult)
For 16S rRNA gene studies ldquotag sequencingrdquo pioneeredby Sogin et al (107) consists of amplifying a suitable sizedpart of the gene with broadly conserved primers (choice isimportantmdashfew are truly universal for the groups intended)often ldquobarcodedrdquo to allow multiple samples to be combinedinto a single run The amplified products are then sequencedand sequences processed en masse Many thousands of partialSSU rRNA gene sequences per sample are economically ana-lyzed this way though they are usually short (currently a fewhundred bases each depending on the sequencing platform)This way of analyzing the composition of microbial commun-ities is now standard These approaches provide so muchinformation even about very rare sequences that the resultshave led to the important concept of the ldquorare biosphererdquoorganisms that may be active or dormant and constituting avery small proportion of the community (eg often muchless than 01) but are potentially important for dispersionadaptation to changing conditions or even critical activitieslike nitrogen fixation or vitamin production (108) Howeverdue to the potential for artifacts like error sequences thisapproach requires rigorous application of quality filteringand clustering algorithms to avoid erroneous taxa and overes-timation of species richness (109 110)
Next-generation sequencing approaches have also allowedwork to begin metagenomics studies of microbial eukaryotes(111) These studies are still constrained by the much largergenomes of eukaryotes and therefore the difficulties ofobtaining sufficient sequences to reconstruct the genome ofa particular species Along with advances in sequencing abil-ity studies of eukaryotic metagenomics have been facilitated
by advances in single-cell isolation which when coupledwith high-throughput sequencing approaches reduce the tre-mendous complexity present in natural complex eukaryoticcommunities to a manageable task (112) Such single-celltechniques are also quite valuable in studies of bacteria andarchaea though the amplification technique tends to bevery uneven and typically generates less than half the genomeof each isolated cell regardless of domain (113 114)
Additionally similar to DNA mRNA is amenable toextraction and sequencing although greater care must betaken during extraction and purification as RNA shows agreater susceptibility to degradation during processing Copy-ing of mRNA by reverse transcription of RNA into cDNAfollowed by DNA sequencing has allowed insights into themetatranscriptomes of environmental samples Metatran-scriptomic studies provide information on gene expressionin an ecosystem and thereby indicates ldquoactivityrdquo of themicro-bial community rather than simply ldquopotentialrdquo represented bythe genomic DNA present in the sample with many applica-tions from showing which processes are being carried outby which organisms to fine-scaled diel studies (115ndash118)Nevertheless given variations in the lifetimes of differenttranscripts and protein molecules the transcriptome maynot be fully representative of the current activity of anorganism
Limitations of these analyses include sequencing errorsPCR mismatches or biases clustering and bioinformaticschallenges and chimeras generated during PCR Also thephylogenetic resolution of short sequences is limited giventhe high conservation of rRNA sequences Even with clone-free shotgun metagenomics there may be biases such as non-random losses of DNA during extraction and preparationor biases (eg from G +C content or secondary structure)in the sequencing procedures Determination of speciesdiversity by these approaches provides an example of theselimitations Shakya et al (119) working with synthetic com-munities (purified genomicDNA from16Archaea represent-ing 3 phyla and 48 Bacteria representing 16 phyla remixed tosimulate an environmental DNA extract) applied both meta-genomic analysis (454 and Illumina platforms) and PCRamplification followed by 454 sequencing of 16S rRNA genesto determine both species richness and relative abundanceThey found that PCR amplification454 sequencing of 16SrRNA genes yielded an accurate measure of species richness(providing that appropriate data processing was applied)but that the relative abundance of up to 94 of the species(depending on domain and variable region amplified) wasover- or underestimated by at least 15-fold (values rangedfrom not detected to 103-fold overestimation) In contrastboth metagenomic approaches yielded relative abundancesthat were within the authorsrsquo 15-fold accuracy cutoff forsim50 of the species However they concluded that addressingrichness overestimation in metagenomic analyses that isdistinguishing rare but real OTUs from experimental andcomputational artifacts awaits further computational andclassification improvements More recently Parada et al(120) used mock communities composed of 16S rRNAclones from 27 common marine taxa (from nine Bacterialand two Archaeal phyla) to show that small differences inPCR primers (and different clustering methods) can yieldlarge differences in apparent relative abundances of reportedtaxa However one primer pair and informatics pipelinethey tested using a particular version of 515F-926R (V4ndashV5) provided accurate estimates of relative clone abundance(r2 = 095) when comparing observed versus expected cloneabundance
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-9
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
Culture-Independent Diversity Studies
The first groups to be identified using cultivation-independ-ent molecular techniques from the marine plankton (121)were the bacterial SAR11 cluster (a group of closely relatedgene sequences or phylotypes) and marine picoplanktonicunicellular cyanobacteria Synechococcus and Prochlorococcus)Of these the SAR11 cluster was completely unknown butthe cyanobacteria had previously been recognized by theirunique pigment fluorescence Waterbury et al (122) andJohnson et al (123) used epifluorescence to observe Synecho-coccus and Chisholm et al (124) discovered Prochlorococcususing flow cytometry These cyanobacteria were later isolatedand grown in phytoplankton culturemedia These two groupsare generally common in the euphotic zone with SAR11 typ-ically comprising one third of the planktonic bacteria (125)and the cyanobacteria common everywhere but polar waters
Probably the biggest surprise to come from the applicationof molecular tools was the discovery by Fuhrman et al (126)of abundant archaea in the deep sea The archaea were foundto be in a unique phylogenetic cluster that was only distantlyrelated to any previously known archaea but the ldquoclosest rel-ativesrdquo (not really close at all) were extreme thermophiles Asubsequent study also found archaea to be present in near-surface coastal waters albeit relatively rare (lt2 relativeabundance) This study used a PCR technique specificallytargeted archaea and found ldquomarine Crenarchaeardquo plus a sec-ond group belonging to the phylum Euryarchaea (127)
Up until this time all known Archaea were thought to beldquoextremophilesrdquomdashadapted for either very high temperatures(thermophiles) extremely salty conditions (halophiles) orstrictly anaerobic environments (methanogens) Yet theseorganisms were present in cold or cool water at ordinary salin-ity and high oxygen concentrations Fluorescent in situhybridization (FISH) measurements from deep-sea sampleshave since showed that the archaea may make up appro-ximately 40 of the total countable prokaryotes with thepercentage reaching to 60 at 200 m depth in theMediterra-nean (35 36) An extensive time series of FISH measure-ments near Hawaii confirmed that the archaea are indeedvery abundant throughout the year from below the photiczone to at least 4000 m and typically constituting 30ndash40of the total prokaryotes present in waters deeper than a fewhundred meters (128) with similar results found elsewhere(129 130) (Fig 4) They have been reported from many pla-ces including the Atlantic Pacific and Southern Oceansand are dynamic components of the plankton the most com-mon type by far being the ldquomarine Crenarcheaeardquo (128 129131) While the marine archaea have been reported to bedominated by a few major ldquophylotypesrdquo (132) they alsohave been shown to have a great deal of microdiversity withinthese phylotypes suggesting there are many kinds of close rel-atives coexisting (133) Interestingly this group of archaeamay likely be the most abundant kind of organism on Earthgiven the huge volume of the deep sea and their high abun-dance there (134) It has recently been proposed that ldquomarineCrenarchaeardquo be elevated to their own major phylum outsidethe Crenarchaeota called the Thaumarchaeota on the basisof deep phylogenetic branching and fundamental differencesbetween them and the Crenarchaeota (135) The Thau-marchaeota possess the uniquely archaeal membrane lipidcrenarchaeol and are nowalso known to be abundant in soils
The physiology of the Thaumarchaeota has been anintriguing area of study Initially an autoradiography-FISHcombination approach demonstrated that they take upamino acids (130 136) But subsequent data showed that
crenarchaeol was derived largely from CO2 suggesting auto-trophic metabolism (137 138) An experiment showinguptake of 13C-labeled bicarbonate into these archaeal lipids(139) directly pointed to autotrophy in this group Chemoli-thoautotrophywas first hinted at byVenter et al (105) whosemetagenomic analysis showed an apparent archaeal scaffoldthat contained genes suggestive of ammonia oxidation Fur-ther evidence came from Schleper et al (140) who detectedseveral ammonia oxidation genes in order directly adjacent toa Thaumarchaeota 16S rRNA gene in a soil-derived metage-nomic clone The issue was directly resolved when Konnekeet al (141) isolated a related marine archaeon from sedimentof a marine aquarium and this organism Candidatus Nitroso-pumilus maritimus was found to have a chemoautotrophicmetabolism quantitatively oxidize ammonia to nitrite andcontain an archaeal ammonia monooxygenase gene (amoA)and interestingly did not grow heterotrophically Cultivationexperiments showed this organism has a high affinity forammonium allowing it to outcompete bacterial nitrifiers atlow (submicromolar) concentrations as are typical in thesea (142) The complete genome of the organism has shownnovel adaptations for nitrification and autotrophy (143)
Even if the Thaumarchaea are primarily chemolitho-autotrophs field data suggest a level of mixotrophy in thatsome organic substrates are being incorporated into biomassA stable isotope study using cells collected at 670 m depthoff Hawaii estimated that about 80 of the carbon incor-porated into archaea-specific lipids came from inorganic sour-ces and about 20 from organic compounds (144 145)Interestingly it has also been claimed that the genomesof deep-sea (gt2000 m depth) members of the Thaumarch-aea as well as those living in equatorial waters rarely containthe amoA gene and thus may be primarily chemoorgano-trophs (48)
Major bacterial groups that have been documentedfrom seawater using 16S rRNA characterization includesome that are also known from culture (eg AlteromonasRoseobacter) and several that are phylogenetically distantfrom standard cultures The most common groups are inrough order of their relative abundance in clone librariesfrom most to least abundant SAR11 (relatives of Pelagibac-ter ubique) Roseobacter SAR86 cyanobacteria SAR116SAR202 SAR234 andMarine Group A The SAR designa-tion followed by a number is an arbitrary sequential cloneidentifier from Sargasso Sea cloning studies done by the Gio-vannoni lab that did most of the early systematic cataloging ofclones Summaries of the data and phylogenetic relationshipsof these groups can be found in Fuhrman andHagstrom (102)and Giovannoni et al (101)
Molecular Genetic Discoveries in Bacterial andArchaeal Marine BiologyAs described earlier metagenomics is an extension of theideas used in the 16S rRNA cloning studies in that all genesfrom the native microorganisms are separated and clonedwithout having cultivated the organisms These methodsearly on started to find unexpected and very interestingresults The best examples involve unexpected marine photo-trophy the first of which is discovery of a nonchlorophyllphotosynthetic bacterial pigment called proteorhodopsin(146) The gene was found on a large environmentallyderived fragment of DNA that also had a gene coding for16S rRNA from the so-called SAR86 group (one ofthe groups common in seawater) This pigment can act as alight-driven proton pump thought to permit cells to generate
422-10 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
FIGURE 4 Distribution of Bacteria Euryarchaea andmarine Crenarchaea (recently renamed Thaumarchaea) along a transect in theNorthAtlantic Ocean as measured by CARD FISH and expressed as of total bacteria + archaea counts via DAPI stained epifluorescence fromTeira et al (130) Top panel shows station locations that are shown on the top of the lower three panels with the distance in km reported alongthe transect from lower to higher station numbers doi1011289781555818821ch422f4
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-11
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ATP from sunlight Interestingly different versions of this pig-ment are found at different depths apparently ldquotuningrdquo theabsorption to match the ambient wavelengths of light asthey change with depth (147) The proteorhodopsin genesare very widespread and diverse perhaps in half or more ofall marine bacteria including SAR11 and Euryarchaeota(105 148ndash150) However almost all of several culturedorganisms with proteorhodopsin studied to date do not showa growth benefit from light (150) with the exception of amember of the Flavobacteria Dokdonia sp strain MED134that grows faster in the light only under moderately lownutrient conditions (151) and a marine Vibrio has beenshown to survive starvation longer in the light than in dark-ness (152) These observation may explain the wide phyloge-netic distribution and high abundance of this gene in thatproteorhodopsin may often assist long-term survival of bacte-ria under extreme energy-limiting conditions yet not havemucheffect on growthduringmore energy-replete conditionsInterestingly a recent report suggests the growth benefit fromlight in Dokdonia MED134 is from enhanced uptake of itsrequired growth factor vitamin B1 which is transported by atonB-dependent transporter powered by a proton gradient(153) This pigment might also have sensory or other rolesnot yet well examined (150)
The second kind of phototrophy found by moleculargenetic and fluorescence techniques to be unexpectedlyimportant in marine plankton is anoxygenic aerobic bacterialphototrophy based on the pigment bacteriochlorophyll aCulturable aerobic anoxygenic photosynthetic (AAP) bacte-ria have been known from seawater for several years (154) andare widespread (155) Initial recent reports of direct countsclaimed they represent about 11 of the total bacterial com-munity in the euphotic zone (156) but those early counts didnot correct for the presence of other bacteria and such correc-tionoften yields estimates averaging closer to 2(157)Directmeasurements show the bacteriochlorophyll pigment is rela-tively rare (158) However studies in various ocean locationsincluding ocean gyres report that these AAP bacteriacan indeed on occasionmake up a quarter of the total prokar-yotes (159 160) A recent energetic modeling of AAP andproteorhodopsin-containing bacteria (161) has suggestedthat proteorhodopsin-containingones cangain approximately02asmuchenergy from sunlight asSynechococcus (commonmarine cyanobacterium) and AAP bacteria can gain approxi-mately 13 as much energy from sunlight as SynechococcusThey concluded the AAP bacteria may gain energy sufficientto meet maintenance costs but proteorhodopsin-containingbacteria were not expected to do so except at high light inten-sities andwith largenumbersof proteorhodopsinmoleculespercell They concluded the ease and low cost of maintainingPR-based phototrophy (a few genes required) may explainthe high incidence of proteorhodopsin genes
Other metagenomic studies have yielded interestinginsights about organisms and processes that would otherwisebe difficult or impossible to establish For example thesequence of an archaeal gene fragment isolated from seawaterrevealed extensive evidence of genetic exchange with othertypes of archaea and even bacteria (162) Such geneticexchange was also quite evident from the complete genesequences of different strains of Prochlorococcus and Synecho-coccus which was attributed in part to virus-mediated geneflow (163ndash166)
Dilution Cultures of Bacteria and ArchaeaSome recent cultivation techniques that permit growth ofldquotypicalrdquo oligotrophic marine bacteria in pure culture are
based on the simple concept that bacteria living in seawateron dissolved organic matter might best be grown in the labo-ratory in ordinary filtered seawater This conclusion followsfrom similar techniques to grow mixed ldquoseawater culturesrdquoof marine bacteria (167) but the seawater inoculum in thiscase is diluted so that only one or a few bacteria are addedto the initial culture vessel (168) A rapid throughput versionof this method has been used to cultivate members of bacte-rial groups thought to be conventionally uncultivable such asthe SAR11 clade (169 170) albeit often at very low den-sities Such cultures permit focused studies of the propertiesof those organisms including genomic analysis (169 170)This valuable work provides insight into the roles and activ-ities of some of the most common bacterial phylotypesobserved in the ocean (171) showing for example that culti-vated SAR11 requires a source of reduced sulfur as well asproviding templates aiding the interpretation of metage-nomic analysis (172)
PROTOZOADiversity and DistributionHeterotrophic protists have been identified historically fromtheir morphological features that are apparent at the levelof the light or electronmicroscope Features of cell size shapetype and pattern of flagellationciliation skeletal structurescharacteristics of the nucleus and other cellular structuresand organelles have been used to differentiate themany thou-sands of described species A description of the extent of thisdiversity of form and function is well beyond the scope of thisbook (see [79] for more information) However much of thisdiversity can be reduced to three basic body plans that dictatethe broadest ecological roles of these cells amoeboid flagel-lated and ciliated forms (Fig 5) Among the lineages possess-ing one of these three forms only the last group the ciliatedprotists constitute a monophyletic group within the domainEukarya
One of the simplest body plans for protozoa is the amoe-boid cell exemplified by the gymnamoebae or ldquonakedrdquoamoebae Motile nonphotosynthetic cells lacking flagellaor cilia occur as life stages in a number of phylogeneticallydiverse taxa but for many free-living species of protozoathis form constitutes the only life stage Motility is largelyconfined to movement along surfaces by means of pseudopo-dia that can take on a variety of (species-specific) shapes Sig-nificant abundances of amoebae are largely confined tobenthic and epibiotic environments (174) and to suspendedparticulate material where they can occasionally be highlyenriched (27) Most amoebae consume bacteria and otherminute prokaryotes and eukaryotes
Several heterotrophic protistan groups possess complexamoeboid body plans most notably the foraminifera polycys-tine and phaeodarian radiolaria and the acantharia Plank-tonic forms are predominantly oceanic in their distributions(although a large number of benthic species of foraminiferaexist) These species are heterotrophic and many are visibleto the naked eye (individual cells can bege1 cm gelatinouscolonies can form ribbon-like structuresge1 m in length)They feed on a wide variety of bacterial protistan and meta-zoan prey using pseudopodial networks have rather longcomplex life cycles for individual cells (weeks to months)and are extensively used in studies of paleoclimatologicalreconstruction (82 175) In addition many of these speciespossess intracellular symbiotic usually eukaryotic algaeThe widespread occurrence of algal endosymbiosis among
422-12 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
these protozoa in oceanic pelagic environments implies thatstrong selective forces appear to give rise to these associations(97 98)
Flagellated forms of protozoa exist across many protistanlineages Eukaryotic flagella come in various sizes numbersand forms (eg with or without tiny hairs) that are character-istic of the different lineagesMany flagellate species are appa-rently purely phototrophic or heterotrophic (phagotrophic)but a significant (still poorly known) fraction of flagellates aremixotrophic combining both phototrophy and phagotrophy(70 88) Flagella in phagotrophic flagellates are employed formotility and prey capture Most free-living flagellates possessone to four flagella (typically one or two) that can be manytimes the length of the cell itself Although there is tremen-dous species diversity among flagellated protozoa many ofthese species have broadly overlapping ecologies As a groupflagellates are the most numerically abundant protozoa ofboth benthic and pelagic ecosystems and they are fundamen-tally important as consumers of bacteria cyanobacteriaand other eukaryotes Flagellated protozoa within the nano-plankton size class (2ndash20 microm) are often counted as a singleassemblage in plankton studies because morphological details
apparent using light microscope are insufficient to distinguishamong the many species This assemblage has been variouslyreferred to by a variety of names and acronyms including het-erotrophic nanoplankton (HNAN HN) heterotrophic flag-ellates (Hflags) heterotrophic nanoflagellates (HNF) andmicroflagellates Heterotrophic nanoplankton is the mostaccurate term for most methodologies employed to countthese cells because it does not require visualization of flagella(which are often lost from these small cells during preparationfor microscopy) The confusion over terminology is partly aconsequence of the different methodologies that have beenused to count these species and partly a consequence of thefact that these species were first studied prior to the wideacceptance of the size convention of Sieburth et al (84)
Heterotrophic flagellates in the microplanktonic size class(20ndash200 microm mostly dinoflagellates) are important consum-ers of phytoplankton in pelagic ecosystems Many of thesespecies are capable of the production of large pseudopodialnets the pallium that can engulf prey (particularly diatoms)significantly larger than the diameter of the dinoflagellatetheca (176) This behavior and its ecological significance forenergy flow in plankton communities have been recognized
FIGURE 5 Body plans and size ranges of protozoa These micrographs depict amoeboid (andashg) flagellated (hndashj) and ciliated (kndashm) forms ofprotozoa From Caron et al (173) Markers bars are 5 (i) 10 (h) 20 (c j l) 30 (a) 50 (k m) 100 (b f) 500 (d) and 1000 (e g) micromdoi1011289781555818821ch422f5
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-13
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
only within the past few decades (177) These species areoften abundant in waters where diatoms dominate (178179) In such situations heterotrophic dinoflagellate biomasscan be similar to that of ciliated protozoa (180)
Ciliates are generally the most recognizable form of pro-tozoa to nonspecialists Nonetheless ciliates are a diversegroup morphologically as well as ecologically (181 182)The degree of ciliature on these species can range from uni-formly ciliated to totally devoid of cilia during most of theirlife cycle to ciliature restricted to specific regions of thecell In addition cilia can fuse to form various complexstructures (eg ciliary membranes or veils cirri) that assistin locomotion food capture or attachmentCiliates are abun-dant and ecologically important species in both benthic (8)and pelagic (183) ecosystems and can consume a variety ofprokaryotic and eukaryotic prey Together with the heterotro-phic dinoflagellates these species are the dominant consum-ers of phytoplankton in many pelagic ecosystems (184) andas a consequence they form an important trophic link to met-azoan zooplankton (185 186)
Diversity and Biogeography of Protists FromMorphology to DNA SequencesUntil relatively recently the biodiversity of marine protistanassemblages was not generally considered a controversialtopic It was generally accepted that while all species of pro-tists certainly have not been identified representatives ofmost types of algae and protozoa had been observed anddescribed if not actually brought into culture in the labora-tory Direct sequencing of 18S rRNA genes from environ-mental samples (as described in ldquoMolecular Phylogeny andMetagenomics Field Applicationsrdquo) has changed that viewInitial forays into environmental DNA indicated a muchgreater diversity of protists than previously documented usingclassical approaches of culture and microscopy (81 187ndash191) Numerous publications over the past decades haveexpanded these findings (see [192] for a recent global analy-sis) which are highly analogous to discoveries in marine pro-karyote research as described already
Previously uncharacterized protistan diversity has nowbeen documented at virtually every level of eukaryoticorganismal classification Some of these findings couldhave been expected but some have been very unexpectedFor example a much greater diversity than noted previouslyhas been observed among small (lt10 microm) protists in plank-tonic ecosystems These species generally possess few distinc-tive morphological features and one could expect that manycryptic species might be present among these small morpho-types The molecular ldquodiscoveryrdquo of this eukaryotic diversityhas stimulated progress on the isolation and descriptionof new species and genera of minute algae and protozoa(193ndash197)
Analyses of environmental samples have also indicatedthe unanticipated existence of novel 18S rRNA gene sequen-ces that imply the presence of novel lineages of eukaryotes innatural protistan communities (198ndash200) These are sequen-ces for which there are apparently no known described orcultured species The degree to which these sequences dif-fer from sequences of known sequenced eukaryotes impliesthat some of these lineages may be distinct at the level ofphylum (201)
These findings have raised basic questions and somedebate concerning the true diversity and biogeography ofprotistan assemblages in natural ecosystems (202ndash204) thevalidity of the many novel phylotypes or cryptic species that
are being documented (205) the potential importance andsignificance of the many rare taxa that characterize thesecommunities (206) and the implications of these findingsfor the ecologicalbiogeochemical roles that protists playin aquatic ecosystems For example it was postulated andsubsequently confirmed that some novel alveolate lineagesreported from marine ecosystems make up a suite of parasiticprotozoa whose ecological importance may have been signifi-cantly underestimated in the past (207) Deciphering theidentity and significance of these many unknown phylotypeswill constitute a significant effort for protistologists in thefuture (80 85)
Life Histories and Ecological Strategies of ProtozoaAbundance prey type and life histories all vary tremendouslyamongmarine protozoa The smallest species (eg many flag-ellates) tend to be the most abundant and widely distributedin the world ocean Indeed many of these species may beglobally distributed (208) Most nanoplanktonic flagellateshave potentially rapid rates of grazing and growth Underoptimal conditions these species can divide by binary fissionseveral times a day and thus dramatic increases in their pop-ulations can take place in response to favorable conditionswithin a few days (209) Many of these species are capableof surviving for limited periods of time without food andhave developed a variety of physiological or life cycle strat-egies to cope with these events (209) However in contrastto some bacteria that may remain viable through long periodsof starvation (210) protozoa will expire or encyst in responseto low food abundance and thus ldquoboom-and-bustrdquo popula-tion changes are characteristic of small flagellates
On the other end of the size spectrum from the ratherubiquitous nanoflagellate species many of the largest proto-zoan species (polycystine and phaeodarian radiolaria plank-tonic foraminifera) are exclusively oceanic (ie do notsurvive in most coastal environments) andor have specificlatitudinal and depth distributions Thus the abundancesof these latter species may range from undetectable to max-imal abundances of gt105 individualsm3 Dramatic changesin the abundance of these protozoan taxa also can be relatedto changes in prey abundance physicalbehavioral aggrega-tion or to periodicity of life cycle events For example theplanktonic foraminifer Hastigerina pelagica reproduces on alunar cycle and thus abundances (and life stages) of this spe-cies in oceanic waters can vary considerably over the courseof a month (211) In general life cycles for the large amoe-boid protozoa are lengthy and complex (for single-celledorganisms) with life spans unknown for many species(attempts to culture them in the lab have so far been unsuc-cessful) but estimated to be on the order of months to per-haps years (175)
The abundance and activities of microplanktonic hetero-trophic protists (mostly ciliates and heterotrophic dinoflagel-lates) tend to be somewhat intermediate to those ofnanoplanktonic flagellates and the larger amoeboid formsThese species are present in the majority of marine ecosys-tems and collectively play an important role in the controlof phytoplankton biomass (and probably the abundance ofnanoplanktonic protozoa although there is little informa-tion on this topic) in waters throughout the world oceanLike small flagellates ciliates and dinoflagellates reproduceprimarily by binary fission but their maximal growth ratesare typically slower (one division a day is typical) and theirlife cycles often include sexual phases that allow for geneticrecombination
422-14 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
VIRUSESViral Abundance and General PropertiesViruses are simple biological agents typically 20ndash200 nm indiameter composed of a nucleic acid genome in a proteincoat that infect cells and ldquocommandeerrdquo the cellrsquos machineryto make more viruses which are released into the environ-ment when the host cell lyses or bursts A particular virus isthought to be capable of infecting only a narrow range of hosts(usually one species sometimes a genus rarely broader)Although some early studies had isolated viruses from thesea therewere no data prior to the 1980s showing such viruseswere very abundant and more significantly no evidence thatinfection was occurring in any important part of the planktoncommunity It wasnrsquot until the late 1980s that electronmicro-scopy with suitable concentration methods showed thatviruses are extremely abundant similar to or even higherthan bacterial abundance (212 213) Interestingly themost suitable transmission electron microscopy (TEM)approach used by Bergh et al (described in detail by [214])was actually similar to a direct centrifugation method devel-oped in 1949 (215)
TEM studies of viruses permit high-resolution images andobservation of viral morphology showing features such ashead diameter tails sheaths and tail fibers (Fig 6 collage
in upper left) Each type of virus has a fixed morphology(unlike bacteria that are potentially more plastic in theirappearance) and hence a coarse measure of viral diversityhas been possible by cataloging morphologies Studies thathave done so have found dozens or more different morpholo-gies of marine viruses in a given sample (217ndash219) A largeproportion resemble bacteriophages which are viruses thatinfect bacteria
The availability of brightly fluorescent nucleic acid stainsand high-porosity fine pore size (002 μm) filters made fromAl2O3 have made it easy to accurately count viruses by epi-fluorescence microscopy (38 220 221) Epifluorescence per-mits abundance estimates but does not allow observation ofviral morphology as viruses are below the resolution limitof light microscopy viruses are visible only as sources of light(like stars in the night sky see Fig 6 lower micrograph) Anextension of manual epifluorescence counts is the use of flowcytometry to count viruses now commonly employed in somelabs (222)
Virus abundance has been found to be closely related tobacterial abundance with a virusbacteria ratio typically10ndash301 A typical oceanic profile of bacterial and viral abun-dance is shown in (Fig 6 panel on right) This tight ratioand the strong correlation to bacterial abundance in relationto weaker correlations to chlorophyll have been cited as
FIGURE 6 Marine viruses (upper left collage) viewed by transmission electron microscopy These are cyanophages in the groups myoviridae(right and bottom) siphoviridae (left) and podoviridae (top) Scale bars represent 100 nm From Sullivan (216) Epifluorescence micrograph(lower left) of SYBR green I stained viruses (small fluorescing objects) and bacteria + archaea (large fluorescing objects) Depth distribution ofvirus and bacteria + archaea abundances obtained by epifluorescence microscopy of SYBRGreen stained cells in the central San Pedro BasinCalifornia (11 August 2000) doi1011289781555818821ch422f6
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-15
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
evidence that most marine viruses infect bacteria rather thaneukaryotic phytoplankton (217 218 223 224) though thereare also many important viruses of phytoplankton and otherorganisms with significant impacts on primary productivityblooms and ecosystem function (225ndash227)
The first demonstration of the activity of marine viruseswas presented by Proctor and Fuhrman (228) who showedby electron microscopic examination that viruses wereactually infecting marine bacteria and cyanobacteria at ameasurable rate Because only the final portion of the viruslife cycle is visible by electron microscopy (when the virusesare assembled and ready to lyse the host cell) only a very smallfraction of the infected cells can be counted as infected at anygiven time Proctor and Fuhrman (228) reported that only afew percent of the total bacteria from coastal waters and theSargasso Sea were visibly infected but they interpreted thedata with a model that implied the actual fraction of the totalcommunity that was infected was much higher They con-cluded that the total fraction of bacterial mortality attribut-able to viruses is roughly 10ndash40 Subsequent studies haveused refined versions of that same general approach andmodel (229 230) and numerous studies have used a varietyof alternative approaches to estimate viral activitymdashall yield-ing basically the same general conclusion (although themodel parameters need to be adjusted for cyanobacteriawhich on average seem to be infected less than bacteria)These approaches include (a) calculation of virus turnoverfrom decay estimates (231) (b) measurement of viral DNAsynthesis by incorporation of tritiated thymidine (232) (c)observation of effects of added viruses (233 234) (d) estima-tion of bacterial mortality in the absence of protists (e) use offluorescent viruses to estimate production by an approachanalogous to isotope dilution (38) and (f) dilution of virusesin filtered samples to estimate viral production (235) Theoverall consensus of these studies is that the initial estimateswere basically correct that is that viruses are responsiblefor about 10ndash40 of the bacterial mortality in most marinesystems (reviewed by [218 223 226 236 237]) Howeverthere is still some disagreement about the higher estimatesand it is likely that 40 mortality from viruses alone is nottypical for most marine systems
Most of the foregoing work has focused on the viral infec-tion of bacteria primarily thought to be heterotrophicalthough the original report by Proctor and Fuhrman (228)also noted the occurrence of cyanobacterial infection Sev-eral subsequent studies focused on phytoplankton includingthe potential effect of viruses on the termination of phyto-plankton blooms of Emiliania and Phaeocystis and the likelyimpact this might have on release of climate-active gasessuch as dimethyl sulfide (225 227 238ndash247) Althoughdetails are beyond the scope of this chapter viruses arethought to infect virtually all marine organisms with poten-tially significant impacts from zooplankton to whales (226)Detailed studies of viruses infecting cyanobacteria such asSynechococcus have shown differences in viral host specific-ity particularly toward coastal and oceanic host strainsand some occasional high virus abundances (to 105ml) asmeasured by most probable number (MPN) cultivation tech-niques in the Gulf of Mexico near Texas (248 249) Othervirus cultivation studies with Prochlorococcus and Synecho-coccus in oligotrophic waters of the Sargasso Sea showedan interesting pattern of cross-infection between these gen-era by some virus types (suggesting gene flow among theseorganisms) but generally low MPN estimates of abundanceto 103ml even when cyanobacterial abundance was near105ml (216 250)
Overall the consensus emerging from direct comparisonsof viral-mediated mortality and grazer-mediated (ie micro-zooplankton) mortality have indicated that viral lysis of bac-teria constitutes a significant fraction of total mortality ofthis assemblage while the mortality of phytoplanktonappears to be dominated bymicrozooplankton inmost instan-ces (251 252)
As mentioned viruses have significant morphologicaldiversity as observed in TEM studies It is also possible toinvestigate aspects of their genetic diversity In early workon this topic Wommack et al (253) and Steward et al(254) observed the diversity of viral genome lengths in a fieldsample by pulsed field gel electrophoresis with viral genomesranging 25 kb to gt300 kb in length Field results show thatthe viral community composition is dynamic in space andtime with clear changes in the banding patterns over seasonsand locations in Chesapeake Bay (253) between oceanbasins and subsequent to dinoflagellate blooms (254) andwith depths to 500 m at one location (255)
Although viruses do not all share a set of core genesthat allows a universal viral phylogeny (comparable to SSUrRNA gene in cellular organisms) genetic diversity amonga single group of viruses can be examined by sequence analysisof shared genes within the group An example is the g20 geneinT4-like cyanophages that has shown extremely high diver-sity even among very closely related viruses and has demon-strated geographic and seasonal variation (256) A secondexample is g23 which is found broadly in diverse and wide-spread T4-like phages (257) and can have seasonally repeat-ing patterns (258) as well as short-term rapid dynamics thatcorrelate to those in bacteria (259)
Viral MetagenomicsAs with cellular organisms metagenomics can be used toexamine viral diversity and genetics without themany restric-tions of cultivation Viruses can be collected by selective fil-tration and concentrated by tangential flow filtration orflocculated with iron chloride (260 261) then their collec-tive metagenome can be extracted linker amplified and ana-lyzed by sequencing (262 263) Viral metagenomic studiesare particularly challenging because the large majority ofsequences have no annotated matches in any databases butso far results from marine samples around the world haveshown extremely high diversity and variations with depthlocation and time presumably with highly dispersed typesselected by local conditions (262ndash266) Because metage-nomes are best interpreted when there are representative cul-tures available the best matches of viral metagenomesoriginally tended to be to the few viral isolates infecting trulycommonmarine bacteria like cyanophage infecting Synecho-coccus and Prochlorococcus (267) However the developmentof dilution-to-extinction cultures representing common het-erotrophic marine taxa like Peligibacter (a member of theSAR11 clade) and SAR116 has allowed isolation from sea-water of viruses infecting these common organisms and theseisolates have indeed been found to be highly abundant inmarine viral metagenomes (268 269) New approaches tointerpret the results include clustering the proteins independ-ent of known proteins to compare samples to each other andtry to find environmental factors driving viral communitychanges (270) Such analysis of a large global data set (TaraOceans expedition) has shown that extensive sampling hascome close to reaching the total diversity in tropical and tem-perate waters of such viral protein clusters (which essentiallyrepresent various viral protein motifs not all viral proteintypes) and that viruses appear to be directionally dispersed
422-16 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquodownstreamrdquo in major ocean currents as one may expectfrom first principles but also supporting the ldquoseed bankrdquohypothesis of viral biogeography (271)
Viruses and Host DiversityViruses are themselves thought to be instrumental in drivingincreased diversity of their microbial hosts via a hypothesizedprocess often called ldquokill the winnerrdquo This is because viralinfection is host-specific and density-dependent the latterbecause viruses diffuse from host to host so an abundanthost is more likely to pass on infection than a rare oneThis means that if an organism becomes abundant andblooms winning the competition for resources it becomesmore susceptible to a viral epidemic This would benefit therarer organisms and thus help foster diversity (223 272273) However bloom scenarios as described above involvesystems far from steady state and the formal theory of Thing-stad and Lignell (274) has interesting steady-state solutionswhere several viruses infect several hosts stably over time(via trade-offs between growth rates and viral susceptibility)which may occur at the strain or species level (274) There issome experimental evidence that viruses have effects on nat-ural marine microbial community composition weaker thanthe kill the winner (bloom version) hypothesis would sug-gest (275 276) so something resembling the steady-statecoexistence described in the model may in fact be commonThere also appear to be processes that foster coexistencebetween viruses and hosts but the mechanisms are largelyspeculative (223 277 278)
Viruses may also be directly involved in host genetic diver-sity because they can be the agents of genetic exchangebetween microorganisms (223 273) This often involvesthe viral lifestyle known as lysogeny whereby viruses survivewithin host cells as DNA only integrated into the host chro-mosome and being reproduced each time the host divides Ahost harboring such a genome is called a lysogen becauseunder conditions of stress to the host cell a genetic switchmay cause the viral genome to initiate the lytic process pro-ducing many progeny viruses and bursting from the hostLysogeny is a very common property occurring in a signifi-cant part of the bacterial community (279 280) althoughthe incidence of induction of the lytic phase in nature is appa-rently low (281) Overall lysogeny is poorly understood butthought to have both positive and negative impacts on themicrobial community (282) Recent results suggest that ben-efits of being lysogenic in highly seasonal polar seas leads tofundamental difference between polar and other marine viralcommunities (283)
Viruses and the Microbial LoopAs part of the food web viruses occupy a unique positionThey infect host cells that are mostly thought to be heterotro-phic bacteria and by doing so they typically burst the hosts torelease progeny viruses and cellular debris But what is the fateof this material Viruses themselves do not last indefinitelyand a simple steady-state assumption implies that from eachburst of viruses (typically 20ndash100 per lytic event) only onesuccessfully infects another cell The rest are inactivatedand broken down by sunlight (UV and visible exposure)and enzymatic attack (284) or consumed byminute phagotro-phic protists (285) thus reentering the food web as substratefor bacteria or food for protistan consumers Experiments incontrolled laboratory systems and field studies with radioac-tively labeled viral lysis products have supported the conclu-sion that most of the organic matter released by the viralinfection is either taken up by bacteria or respired (286 287)
Modeling this process as part of the microbial loop showsthat viral lysis represent a sort of side loop that has the neteffect of remineralizing a significant amount of the carbonand nutrients that enter the bacteria-protist part of themicro-bial loop (Fig 7) A theoretical numerical steady-state modelcomparing a systemwith no viral activity to onewhere virusesare responsible for 50 of bacterial mortality showed that thesystem with viruses had 33 more bacterial production andrespiration than the virus-free system implying that theviruses had the effect of permitting the bacteria to processmore of the primary production than they would otherwise(223) Although 50 is a high number unlikely to be com-mon in the sea this model nevertheless illustrates that virusescan reduce the amount of energy reaching higher trophic lev-els The implication is that viruses lead to increased bacterialactivity at the expense of the larger organisms
MAJOR ENVIRONMENTAL CONTROLSLight Temperature and PressureTemperature has an important potential influence on bio-chemical reactions and therefore on biological processesin general Most ocean waters fall in the range of ndash2degC to30degC with obvious exceptions in hydrothermally heatedareas Temperature has long been known to be a regulatingfactor for the growth of heterotrophic microbes
In temperate waters it has been established that microbialactivity is generally much higher in warm summer waters thanin winter (55) The relationship is not simple howeverbecause multiple factors act at the same time Some contro-versy still exists regarding the highest and lowest extremesfor marine bacterial growth although there is broad agree-ment that bacteria grow gt100degC at hydrothermal vents andltminus5degC in sea ice brines Pomeroy et al (57) noted the inter-esting observation that bacteria seem particularly inhibitednear the freezing point of seawater (ca ndash22degC) comparedto eukaryotic phytoplankton This effect results in polarspring phytoplankton blooms that accumulate organic car-bon in advance of the response of the bacterial communityand development of the microbial loop and perhaps lead toenhanced benthic-pelagic coupling (288)
The relationship between temperature and the growth rateof marine phytoplankton was described broadly in a now
FIGURE 7 Modification of the microbial loop concept that incor-porates the functional role of viruses Export can be via predation orsinking From Fuhrman (223)doi1011289781555818821ch422f7
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-17
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
classic paper by Eppley (58) and later Goldman and Carpen-ter (289) Temperature was shown to exert a strong and directeffect on themaximal growth rates of these species withmax-imal intrinsic growth rates at 0degC generally lt1 divisiondaywhile growth rates at 30degC may be gt4day These relation-ships indicate the maximal rate that might be attained byphytoplankton at these temperatures but they do not takeother factors into account (nutrients light) While warmtemperature ostensibly allows more rapid growth it createshydrographic conditions that typically give rise to nutrientlimitation of algal growth Thus some of the coldest watersin nature witness some of the most massive phytoplanktonblooms (290) while warm oceanic gyres represent some ofthe most oligotrophic areas of the ocean The multiple andoften conflicting effects of temperature on the growth of nat-ural phytoplankton assemblages limit the accuracy of presentpredictions regarding how primary producers will respond toclimate change (291)
An analysis of the effect of temperature on the growth ofheterotrophic protists has indicated that the response is qual-itatively similar to that of phytoplankton but with a twistThe growth rates of at least some protozoa can exceed thoseof phytoplankton at warmer environmental temperaturesbut the opposite effect is apparent at very low environmentaltemperature This differential effect of temperature on thegrowth of phototrophic and heterotrophic protists was basedon a large meta-analysis of published protistan growth (59)That analysis demonstrated that the maximal growth ratesattained by phototrophic protists could exceed the maximalgrowth rates attained by heterotrophic protists (all otherpotential growth-limiting factors not considered) Thereforeprotozoan growth rates may be constrained to a greater degreeat low environmental temperature than rates for phytoplank-ton If so then phytoplankton bloomsmay get a head start ongrazers during spring in polar ecosystems This scenario is con-sistent with information on seasonal biomass changes andmicrozooplankton herbivory in the Ross Sea Antarctica(292 293) but there are still too few data to fully vet thishypothesis
The importance of high pressure on bacterial growthgained considerable attention in the late 1960s when thedeep sea submersible Alvin was accidentally lost overboardwith its hatch openWhile no lives were lost in this accidentsome workmenrsquos lunches sank to the bottom (sim1500 m)inside the submersible Alvin was recovered after 10 monthsand interestingly there was a waterlogged lunch containingapples bologna sandwiches and broken vacuum bottleswith broth that all appeared hardly degraded and tasted palat-able Yet when placed in a refrigerator on the ship these itemsdegraded relatively quickly Initially pressure was thought tobe the preserving factor as the sea floor temperature was sim-ilar to the refrigerator temperature and the only major differ-ence would be pressure (294) Following this observationa series of experiments to measure degradation of variousorganic materials left in the deep sea for extended periodsindicated that degradationwas typically significantly reducedimplying that pressure reduces the degradation rates (295)Nonetheless changes in the protozoan community of naturaldetrital material sinking to the deep ocean floor indicate thatthe microbial community can respond relatively quickly insome situations (296) Deep sea microbiology has advancedconsiderably yet it is still difficult to interpret results withrespect to actual in situ rates of naturally occurring organicmatter Deep sea bacteria adapted to high pressures havebeen isolated that are barophilic (also called piezophilic)meaning that they prefer high pressures and have reduced
activity at lower pressures whereas others are barotolerant(piezotolerant) tolerating but not preferring high pressuresfor example see (297 298)
Few data are available on barotolerantbarophilic marineprotozoa Protozoa certainly exist and grow at great oceanicdepths but measurements of in situ growth rates for these spe-cies do not yet exist Measurable protozoan numbers havebeen documented in the deep-sea sediments for more than30 years (299 300) and viable protozoa have occasionallybeen cultured from these environments (301ndash305) butvery few direct measurements of the activities of these speciesin situ have been reported (306) A few protozoa have beenisolated that will grow at high pressure (302 303 307) andprotists that appear to be unique to the deep ocean havebeen observed either directly or through the analysis ofDNA sequences (308ndash311) but possibly the best direct evi-dence that protozoan activity takes place at the high pressurescharacteristics of the deep sea are experimental and observa-tional work noting the stimulatory effect that detrital deposi-tion has on some components of the protozoan community(296 312) These observations indicate a diverse and activeprotozoan fauna of the deep ocean although their biogeo-chemical significance is largely uncharacterized at this time
Dissolved and Particulate Organic MatterBacteria and archaea are thought to be by far the most impor-tant organisms with respect to the processing of dissolvedorganic matter (DOM) and nonliving particulate organicmatter (POM also called detritus) in the ocean While theremay be some uptake of DOM by protists particularly forgrowth factors needed in trace amounts such as vitamins(313) the bulk of this material is probably utilized by bacteriaand archaea (314) including the smallest cyanobacteriumProchlorococcus (315 316) which thus may be considered amixotroph Due to their small size bacteria have extremelyhigh surfacevolume ratios and combined with their over-whelmingly high abundance an extremely high integratedsurface area Protozoa tend to obtain the organic materialsthat they require for growth from their prey rather thanthrough the uptake of DOM Overall protozoa tend to besources of dissolved and detrital organic substances throughthe excretion of unassimilated prey biomass in expelledfood vacuoles
Particulate organic matter is not directly available as sub-strate to bacteria These substances must first be reduced tosmall molecules that can be transported into the cell Thisis accomplished by the production of extracellular enzymes(note that few if any large polymers are directly taken up bybacteria with the possible exception of DNA) Hydrolyticenzymes produced by bacteria (and almost certainly archaea)break down polymers like proteins polysaccharides andnucleic acids As with DOM POM is composed of a complexmixture of compounds that vary in their susceptibility to bac-terial degradation and utilization
Particulate material in the water column serves not only asbacterial substrate but also as substratum POM occurs in thewater column across a huge size spectrum from micrometersup to some detrital aggregates more than 1 m in diameter(317) Much of this particulate material is in a constant stateof flux with colloidal material constantly coalescing andaggregating to form new or larger particles (318) as microbialdegradation acts simultaneously to remineralize this materialDetrital particles that attain macroscopic size either by directformation (317 319 320) or via accretion and aggregation(321) and are often called marine snow or macroaggregatesMarine snow particles are readily colonized by bacteria and
422-18 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
many other microorganisms and their abundances on theseaggregates can be orders of magnitude greater than abundan-ces in an equivalent volume of the water surrounding theaggregates (320 322ndash324) and their diversity and highlyconcentrated abundances on detrital aggregates implies thatthere may be stronger trophic coupling and efficient energyelemental cycling relative to these processes among free-living microorganisms in the surrounding waters This specu-lation has resulted in a number of formalizations of the poten-tial impact of aggregate microbiology on biogeochemicalprocesses in the ocean (325 326) Most definitions of bio-films include aggregated particulate and these particles areoften studied within the biofilm conceptual model (see chap-ter by Lawrence et al in this section)
Inorganic Nutrients
Macronutrients (N P)Virtually all heterotrophic organisms in the ocean contributeto the pool of available macronutrients via the excretion ofmetabolic wastes and protozoa contribute significantly tothis process Macronutrients are often in excess within proto-zoan prey relative to the consumersrsquo needs because much ofthe prey carbon is respired to produce energy for metabolismand growth ldquoExcessrdquo nutrients are eliminated as eitherorganic compounds or more commonly as ammonium (forN) and phosphate (for P) The importance of the match(or mismatch) between predator and prey stoichiometry hasbeen demonstrated experimentally and repeatedly in marineand freshwater ecosystems and has been elevated to a funda-mental ecological principle governing elemental and energyflow in aquatic food webs (327) Based on this reasoningCaron et al (328) concluded that bacterivorous protozoaplay an important role in nutrient remineralization andrelease in the ocean Bacterial biomass is typically rich in Nand P relative to other organisms so bacterial predators expe-rience large excesses of these elements relative to their growthneedsMoreover bacterivorous protozoa should be dispropor-tionately important in nutrient remineralization amongsingle-celled eukaryotes because as the smallest protozoa inaquatic ecosystems they have high weight-specific metabolicrates (329) Thus both stoichiometric and allometric rela-tionships implicate small bacterivorous protozoa as impor-tant sources of remineralized nutrients in the plankton
The reasoning also holds true for bacterial utilization oforganic compounds and it has led to some interestingfindings and conclusions regarding the role of bacteria innutrient cycling in the ocean Bacteria are important sourcesremineralized nutrients when they are consuming N- orP-rich substrate Contrary to this traditional view howeverheterotrophic bacteria are often strong competitors for inor-ganic nutrients under many situations largely because bacte-rial biomass is richer in N and P than much of the organicmatter they consume in nature so they require additional Nand P to produce their biomass (summarized by [330]) Exper-imental studies have demonstrated the uptake of ammoniumby bacteria other than cyanobacteria using stable isotope 15N(331) and also the short-lived radioisotope 13N (332) oftenfinding that bacteria may be responsible for one third of the Nor P uptake Additionally there is evidence that bacteria insome oceans like the Sargasso Sea are limited by phosphorus(333 334) Growth limitation of bacteria by phosphorusappears to be a common phenomenon of freshwater ecosys-tems (335 336)
N2 fixation is restricted to prokaryotes and in the oceanwater column it was thought for many years to be done
primarily byTrichodesmium a warm-water colonial cyanobac-terium that blooms sporadically andRichelia a cyanobacterialsymbiont that lives within certain diatoms (337) Some uni-cellular cyanobacteria also contribute significantly to globalN2 fixation (338) and molecular biological data suggestthat a variety of other bacteriamdashnot phytoplanktonmdashmayalso be fixing nitrogen in seawater (339) An exciting recentdiscovery in this field is that a bacterial nitrogen fixer origi-nally known only from nifH sequences UCYN-A has hadits genome sequenced by parallel pyrosequencing and wasfound be a cyanobacterium (Candidatus Atelocyanobacteriumthalassa) lacking the apparatus to generate oxygen as well asother pathways (340) Further study showed it to be symbioticwith a marine alga (341)
Micronutrients (Trace Metals Growth Factors)Bacterial growth may often be limited by macronutrients asdescribed and sometimes by trace nutrients particularlyiron that occur at very low concentrations in seawater(342 343) The importance of Fe as an element limiting pri-mary production in some oceanic regions has come underclose scrutiny and extensive experimental investigation inrecent years Many bacteria have special uptake mechanismsto utilize extremely low concentrations of iron includingsiderophores (released compounds that bind iron and arethen specifically taken up) Even if bacteria are capable ofgrowth at low Fe concentrations growth may be less efficientKirchman et al (344) found that Fe limitation stronglyreduced bacterial growth efficiency in a cultured marine bac-terium (ca 50 efficiency Fe replete but lt10 when Fedeplete) Thus the effects may be complex Marine N2 fixa-tion also may be limited by the availability of Fe becausenitrogen fixers have high Fe requirements
Protozoa tend to be a source rather than a sink for Fe in theocean analogous to their role in macronutrient remineraliza-tion Protozoan grazing activity has been shown to releaseiron from bacterial biomass thereby relieving Fe limitationin co-occurring phytoplankton (345) a mechanistic demon-stration of the process using protozoa growing on Fe-repletebacteria However it has also been reported that Fe-limitedbacteria may contain insufficient Fe for the bacterivorous pro-tozoa that consume them thus leading to Fe limitation in theprotozoan as well (346)
OxygenHypoxic and anoxic regions of the ocean are expected to sig-nificantly expand in the coming decades as a consequence ofchanges in ocean stratification resulting from global climatechange (347) As oxygen becomes depleted bacteria havethe capability to utilize a series of alternate electron acceptorsCompositional changes in the bacterial assemblage accom-pany these changes in metabolic activity but are beyondthe scope of this chapter Many of the studies on benthicmicrobiology have focused on redox reactions that occurthere Oxygen is usually consumed by sediment microbialactivity much faster than the rate at which it can be replacedby diffusion so most sediments especially organically richones are anaerobic below a few mm (fine-grained muds) orcm (coarser sediments) from the sedimentwater interfaceexcept where animals ventilate the benthos through tubesWhen oxygen is absent bacteria and archaea use alternateelectron acceptors such as nitrate nitrite oxidized Fe orMn organic matter or sulfate The use of nitrate or nitriteas an electron acceptor with concomitant production of N2
gas (denitrification) results in net loss of biologically availablenitrogen Bacteria that perform this reaction are typically
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-19
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquofacultativerdquo organisms that can switch from oxygen to nitrateor nitrite Organic matter may be reduced by fermentationreactions in which part of an organic substrate is oxidizedwhile another is reduced fermenter organisms are also typi-cally facultative Sulfate is a particularly important alternateelectron acceptor in marine systems because it occurs as thesecond most abundant anion in seawater (behind chloride)typically 28 mM Its use as an electron acceptor leads to theproduction of elemental sulfur or sulfide The use of sulfateas an electron acceptor yields much less energy than oxygenor nitrate and sulfate-reducing bacteria are strict anaerobesunable to use oxygen (often strongly inhibited by oxygen)Methane can be produced by methanogenic archaea whichare all strict anaerobes using CO2 as an electron acceptor(see Findlay and Battin this section for further discussion)
Reduced compounds such as sulfide are produced by theuse of these alternate electron acceptors and they diffuse awayfrom their source When they diffuse to environments with astrong oxidant like oxygen they may be oxidized by chemo-trophic bacteria in energy-yielding reactions Similarlyammonium can be oxidized to nitrite and then nitrate by che-motrophic nitrifying bacteria and the nitrate can diffuse intoanaerobic layers and become denitrified to N2 Such coupledoxidation-reduction reactions can lead to accelerated biogeo-chemical cycling particularly in sediments
It was originally believed that relatively few free-livingspecies of protozoa inhabit anaerobic and hypoxic marineenvironments This view has changed based on (largely)molecular analyses of some of these environments (308348ndash351) A significant number of previously undescribedeukaryotic phylotypes have been observed in anoxic environ-ments High protozoan biodiversity in the absence of molec-ular oxygen is not completely unexpected because otheranaerobic environments such as some gut environments(eg cockroach guts and cattle rumen) have notoriouslyabundant and diverse protozoan fauna but marine anaerobicenvironments have been difficult to access and sample prop-erly Many anaerobic protozoa exist together with symbioticbacteria (352 353) or possess unique organelles such as thehydrogenosome (354)
MAJOR BIOTIC CONTROLSThe Trophic Activities of Protozoa Predation andTop-Down ControlsPredator-prey relationships are fundamental interactionswithin biological communities linking production of bio-mass on the one hand with removal on the other hand Estab-lishing the trophic relationships among microorganisms andquantifying the rates at which they take place constitutemajor endeavors within marine microbial ecology As notedpreviously protozoa consume a wide variety of prokaryoticand protistan prey Numerous laboratory and field measure-ments have been made throughout the past quarter centuryto attempt to quantify the rates of bacterial and phytoplank-ton consumption The methodology for accomplishing thesestudies has varied and a number of approaches are presentlyemployed
Methodologies for estimating microbial grazing are diversebut generally fall into one of two experimental approachesperturbation experiments or tracer experiments Perturbationexperiments (size fractionation metabolic inhibitors anddilution technique) rely on manipulation of the consumeror prey assemblages in somemanner such that the effect is evi-dent from changes in the abundance of the prey population
during incubations relative to control treatments that receiveno manipulation Tracer experiments (radioactively labeledprey fluorescently labeled prey stable isotopendashlabeled pray)attempt to use labeled prey or a prey proxy to quantitativelyfollow the movement of prey biomass into consumers Forsummaries of these methods see (74 355)
Perturbation experiments have entailed a variety of mech-anisms to ldquodecouplerdquo predators from prey in natural microbialcommunities For example size fractionation uses various sizefilters to remove an entire size class of grazers from awater sam-ple leaving the prey to grow in the relative absence of thosepredators during subsequent incubations Unfiltered samples(controls) indicate the growth of prey in the presence of pre-dation Predation impact is obtained by the difference ingrowth of the prey between these treatments This methodhas been used to examine predation on cyanobacteria heter-otrophic bacteria and minute phototrophic eukaryotesAlternatively metabolic inhibitors have been employed tohalt predation (or in some cases prokaryote growth) insteadof physical removal by filtration
The dilution technique (356) has become a commonmethod for examining grazing mortality of phytoplanktonby microzooplankton (which is comprised predominantly ofprotozoa) This method relies on the dilution of herbivorouszooplankton in a series of subsamples (and concomitantmeasurements of phytoplankton growth in each subsample)to estimate the grazing mortality of the herbivores
Tracer experiments have used both radioactively labeledprey and fluorescently labeled prey to examine predation onbacteria and phytoplankton Radioactive labeling has beenaccomplished using 14C- or 3H-labeled organic compounds(for bacteria) or 14C-bicarbonate (for phytoplankton) Theuse of radioisotope approaches has diminished with improve-ments in the number and types of fluorescent compoundsavailable for this work Fluorescently labeled prey havebeen labeled with a variety of compounds that allow theprey to be observed readily (even inside predators) by epi-fluorescencemicroscopy (357 358) Alternatively a decreasein the total number of fluorescent prey in a sample indicatesthe rate of removal of the prey assemblage
Both perturbation experiments and tracer experimentsdescribed have inherent advantages and disadvantages Forexample perturbation experiments are fairly straightforwardto conduct but they may introduce artifact by removingpotentially important sources of nutrients or organic com-pounds released by predators (359) On the other hand thedilution technique is very labor-intensive but one advantageof this method is that it provides the opportunity to determineboth phytoplankton growth and grazing mortality in a singleprocedure Most tracer experiments are fairly easy to conductand generally do not perturb the food web being studied butfeeding selectivity for or against the tracer (relative to naturalprey) may produce spurious results In addition some of thesemethods provide a measurement of the activity of the entiregrazer assemblage and others are applicable for examiningthe activity of specific taxa For example size fractionationapproaches examine the effect of the removal of an entiresize class of organisms (a community-levelrdquo measurement)whereas fluorescently labeled prey can be used to examinethe ingestion rate of a particular species by observing theuptake of labeled prey into that species (a species-levelmeasurement)
Predation rates of numerous marine protozoa have beenexamined in the laboratory From these experiments a gen-eral knowledge of the rates of consumption by these specieshas been accumulated Feeding in most protozoa shows a
422-20 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
functional response to the availability of prey that is similar tothe response of many organisms (although there are a greatdeal of species-specific differences) That is ingestion rateincreases rapidly as prey abundance increases until a maximalingestion rate is attained (360)Manyof these curves have theform of a Type II functional response curve (361)
Based on the features of these curves it is clear that proto-zoa have the ability to ingest substantial numbers of prokar-yote and eukaryote prey in natural ecosystems Somebacterivorous flagellates can consume bacteria (and cyano-bacteria) at rates up to 100 s per individual per hour whereassome bacterivorous ciliates can consume thousands of bacte-ria per individual per hour Rates of consumption of eukary-otic phytoplankton by herbivorous ciliates and largeflagellates are considerably more variable and highly depend-ent of the feeding mechanism (eg filtration in ciliates versuspseudopodial net in dinoflagellates) and the size of the prey
These lab studies demonstrate that rarely are prey suffi-ciently abundant in nature to support the maximal ingestionrates of which many protozoa are capable Nevertheless pre-dation rates of protozoa in nature are sufficient to constitute amajor factor controlling the standing stocks of bacteria andphytoplankton (63 180 184 355) For example these sum-maries indicate that on average protozoa consume 20ndash80ofthe standing stock of phytoplankton (measured as chloro-phyll) in the world ocean and 60ndash75 of daily primary pro-duction a departure from the classical view of copepods as theprimary consumers of phytoplankton This large role of her-bivorous protozoa infers a large role for copepods as consumersof planktonic protozoa (185) Similarly the role of protozoaas major consumers of bacterial biomass is indicated by therelative balance between bacterial production on the onehand and bacterivory on the other (Fig 8) The approximateone-to-one balance between these measurements over a largenumber of studies implies that protozoan bacterivory must bea major factor determining the fate of bacteria in marine (andfreshwater) plankton ecosystems
It is interesting to note that some types of bacteria includ-ing marine Bdellovibrio are known to prey on other marine
bacteria (362) At the time of this writing the importanceof bacterial predation by other bacteria within the contextof oceanographic processes is not known and presumed tobe small While Bdellovibrio may be cultivated from seawaterit can also be found among sequences from cold sediments(363) and the water column Bdellovibrio happens to be thebest studied predatory bacterium but there are no doubtothers that we simply have not yet identified (364)
Species-Specific Interactions (MutualismParasitism Commensalisms)Interactions between marine microorganisms such as compe-tition allelopathy and symbiosis are fundamental aspectscontrolling community composition and activity but atpresent most are poorly characterized in natural marine sys-tems Bacteria must compete for dissolved and particulatecompounds with other bacteria andwith protistan taxa whilephagotrophic protists must compete with each other andmetazoa for available prey These interactions manifest them-selves in many forms including true competition for availableresources resource partitioning such that some taxa specializeon the utilization of certain substrates or prey while othersspecialize on different ones (reducing competition) or eventhe cooperative use of compounds (eg one bacteriumrsquoswaste products being utilized as substrate by another bacte-rium) These metabolic consortia are probably common inthe ocean but can be difficult to identify and characterizeConsortia of bacteria working together to perform complexreactions especially in anaerobic systems are well known inmicrobiology
Consortia in which there is a very high degree of integra-tion between species (ie symbiosis) are very common in theocean These associations may be beneficial to both partners(mutualism) beneficial to one part and inconsequential forthe other (commensalism) or beneficial to one part and det-rimental to the other (parasitism) Many highly visible andwell-characterized marine mutualisms exist between chemo-synthetic bacteria and animals such as hydrothermal ventor methane seep tubeworms clams mussels or other inverte-brates (365) and also between luminescent bacteria and fishor squids (366) Similarly many protozoa form mutualisticassociations with photosynthetic protists (367) The mostconspicuous of these associations involved large amoeboidprotozoa of oceanic ecosystems (planktonic foraminiferanspolycystine radiolaria and acantharia) which contain largenumbers of intracellular algae (74) but associations betweensome heterotrophic dinoflagellates and cyanobacteria are alsocommon (368) Studies have indicated that efficient nutrientcycling in these protozoan-algal associations confers an eco-logical advantage for both partners in surfacewaters of the oli-gotrophic ocean
Parasitism is also a common symbiotic relationship amongmarine protozoa Protozoan parasites have been reported for adiverse array of protists and animals in the ocean Heterotro-phic dinoflagellates and presently uncultured alveolate cladesrelated to alveolates appear to be particularly successful atevolving thismode of existence (369ndash371) In a common par-asitic group Amoebophrya spp the parasite invades photo-synthetic dinoflagellates where it undergoes reproductioneventually lysing the host and releasing dozens of infectiveswarmers The parasitic protist Pirsonia spp is a parasite ofdiatoms (372) The overall importance of these parasitismsin the population dynamics of their hosts is not clearly knownbut may constitute a significant source of mortality for somespecies (373 374) The frequency and diversity of ldquonewrdquo
FIGURE 8 Meta-analysis of experimental studies that have com-pared rates of bacterial productivity (converted to population growthrate units of dminus1) with rates of bacterivory by phagotrophic protists(expressed as equivalent units of bacterial growth rate dminus1) Thesolid line indicates where measurements of bacterial removal by graz-ers were equal tomeasurements of bacterial production Data redrawnfrom Sanders et al (63) doi1011289781555818821ch422f8
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-21
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
phytoplankton prey and retain the chloroplasts of their prey ina functional state for a limited amount of time (kleptidoplas-tidy73) Various forms and degrees of mixotrophy (mixedphototrophic and heterotrophic nutrition) are commonamong a number of algalprotozoan lineages (74ndash77) UnderWhittakerrsquos scheme phytoplankton ecologists studying a lin-eage of microalgae might have had little familiarity withclosely related heterotrophic species while protozoologistsstudying a particular protozoan groupmight have known littleabout closely related photosynthetic species
One might expect given these caveats that the termsldquoalgaerdquo and ldquoprotozoardquo are no longer used In fact the termldquoprotozoardquo is still commonly used (especially by ecologists)to recognize those eukaryotic species that exist as single cellsand whose nutrition is dependent on the uptake of preformedorganic substances (primarily via prey ingestion) while pro-tists possessing chloroplasts are still commonly called ldquoalgaerdquoSimilarly although the term ldquoprotistrdquo has been abandoned asa kingdom designation it is still widely employed to describeeukaryotic species that are capable of existence as single cells(ie algae and protozoa) The term ldquophagotrophic protistrdquohas also gained popularity in recent years because it recognizesthat many protistan species are capable of phagocytosis eventhough they may also possess their own chloroplasts and thusare technically ldquoalgaerdquo
Despite the shortcomings of Whittakerrsquos scheme it domi-nated the hierarchical organization of life for approximately aquarter century During the past few decades however thissystem has given way to a new organizational scheme that rec-ognizes three domains of life (Archaea Bacteria Eukarya [orEucarya] 78 Fig 2 upper panel) and is based onwhat is pres-ently believed to reflect amore realistic view of the evolution-ary distances that have developed between organisms in theasymp4 billion years that life has existed on our planet Withinthe Eukarya of Woesersquos scheme hypotheses regarding thephylogeny of ldquoprotistsrdquo have changed continuously and rap-idly during the past two decades reflecting new insightsinto eukaryote evolution provided largely by DNA sequenceinformation (Fig 2 lower panel from [79])
The former protistan phyla ofWhittakerrsquos systemhavenowbeen dispersed among candidate ldquosupergroupsrdquo within thedomain Eukarya to better reflect hypothesized phylogeneticrelationships For example the dinoflagellates (which encom-pass phototrophic heterotrophic and mixotrophic species)now form a single group and have been placed together withthe ciliates and apicomplexans (sporozoans) in themonophy-letic Alveolata (Fig 2 lower panel) On the other handeukaryotic heterotrophic single-celled species falling withinthe general description ldquoprotozoardquo are now widely distributedamong a number of protistan lineages In short nutrition hasbeen demoted as a phylogenetic character and other charac-ters (presumably more indicative of evolutionary relatedness)have ascended to address some long-standing contradictionsalthough the debate over the relationships among some line-ages is still very active at the present time 79)
Individual protozoan cells range in size from less than 2 micromto greater than 1 cm in diameter (gt4 orders of magnitude)(80 81) with some colonial radiolaria forming cylindricalgelatinous structures a centimeter in diameter and morethan a meter in length (82 83) Because they constitutesuch a large size range of organisms protozoa are often dividedinto size classes that very crudely correlate with their generalnutritional preferences A commonly used convention is thatof Sieburth et al (84) which groups planktonic microorgan-isms into order-of-magnitude size classes (02ndash20 microm =picoplankton 20ndash20 microm = nanoplankton 20ndash200 microm =
microplankton 02ndash2 mm =mesoplankton) Most protozoafall into the nanoplankton or microplankton size classesModeling microbial trophodynamics using this conventionassumes that protozoa in one size category generally consumeprey one order of magnitude smaller in size (34 85)Although this approach misses much of the detail and diver-sity of the trophic activities of individual protozoan taxa it isa necessary practical compromise for examining community-scale flows of energy and elements It also provides a usefulmechanism for summarizing and comparing the abundancesand biomasses of protozoa from different environments andto other microbial assemblages Protozoan abundance sum-marized in this way has been shown to contribute signifi-cantly to the living biomass of planktonic ecosystemsthroughout the world ocean (Fig 3)
Estimates of protozoan biomass such as those depicted inFig 3 typically do not include the contribution of mixotro-phic phytoflagellates to heterotrophy There is presently noeasy way to determine the abundances of small phagotrophicphytoflagellates in natural samples so these species are typi-cally counted as phytoplankton unless specific methods areemployed to identify the algae as consumers such as the useof fluorescently labeled particles (87ndash90) or through theexamination of food vacuole contents (91) On averagethese species appear to constitute a modest percentage ofthe phytoplankton assemblage (typically lt25) althoughthey may at times dominate the phototroph assemblages ofnatural plankton communities It is important to recognizethat their inclusion as functional heterotrophs rather thanphototrophs could significantly shift the relative contribu-tions of phototrophic and heterotrophic microbial biomassto total biomass within microbial assemblages and the flowof energy within plankton communities (92)
Heterotrophic protists that harbor photosynthetic pro-tists or their chloroplasts within their cytoplasm constituteanother complexity for estimating the contribution of pro-tozoa to total microbial biomass When bulk water samplesare analyzed the contribution of chlorophyll containedwithin those protozoa is generally assumed to come fromfree-living phytoplankton However studies have shownthat chloroplast-bearing ciliates can contribute up to halfthe total biomass of planktonic ciliates in ecosystems andchloroplast-retaining ciliates can sporadically dominate thechlorophyll and primary production of some planktonic eco-systems (93ndash96) The environmental conditions promotingthe success of these ciliates are poorly known Similarlymany species of planktonic foraminifera polycystine radio-laria and acantharia harbor large numbers (thousands perprotozoan) of endosymbiotic algae within their cytoplasm(97 98) Caron et al (99) have demonstrated that primaryproductivity within these species can contribute significantlyto total primary productivity in oceanic ecosystems and can bevery important locally in the convergences of Langmuir circu-lation cells (100)
BACTERIA AND ARCHAEAldquoCulturablerdquo versus ldquoNonculturablerdquo CellsMost conventional cultivation methods can grow only 1 orless of the bacteria that can be visualized by direct microscopytechniques (eg 4) This is true even though most can beshown to be active by techniques such as microautoradiogra-phy (25) These readily cultivable organisms appear to repre-sent a group of fast-growing so-called weeds that are adaptedto take advantage of rapid growth in rare organically enriched
422-6 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
environments This strategy contrasts with the numericallydominant bacteria that are adapted specifically for growthin the dilute nutrient conditions that characterize the vastmajority of the volume of the water column
The most common taxa readily cultured from seawaterwith standard nutrient broth media include the gamma pro-teobacterial genera Vibrio Alteromonas PseudoalteromonasMarinomonas Oceanospirillum Shewanella (usually isolatedfrom surfaces such as shellfish and sediments) the alpha pro-teobacterial generaRoseobacterSphingomonasmembers of thefamily Flavobacteriaceae and Planctomycetes as summarizedin Giovannoni and Rappeacute (101) and Fuhrman and Hagstrom(102) The cyanobacteria Synechococcus and Prochlorococcusare also now readily culturable but on low-nutrient inorganicmedia targeting photosynthetic forms as opposed to organicmedia used to cultivate the others listed above
Because of the low percentage of marine bacteria that canbe grown in standard media organisms that until recentlywere called ldquononculturablerdquo make up the large majority ofbacteria in the plankton Only during the pastsim20ndash25 yearshave molecular biological methods based on 16S rRNA genesequences been available to identify these organisms andthese powerful techniques have opened up a large area forexploration (see next section) Similar but more recent stud-ies use 18S rRNA sequences for characterizing protistandiversity as will be noted below
Molecular Phylogeny and MetagenomicsField ApplicationsModern phylogeny of microorganisms is based primarilyon genetic sequences the most well-studied gene being the
FIGURE 2 The three domains of life (upper left) as proposed by Woese et al (78) and a recent overview of modifications that have beenproposed by Adl et al (79) to higher-level phylogentic groups within the eukaryotic component of the tree (lower right) Domains figure fromWoese et al (78) eukaryotic tree figure from Adl et al (79) doi1011289781555818821ch422f2
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-7
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
small subunit ribosomal RNA gene (16S rRNA in Bacteriaand Archaea and its larger homolog 18S rRNA in eukar-yotes) This molecule is strongly conserved over evolutionarytime so this single molecule has been used for constructingphylogenetic trees of all living organisms (httptolweborgtreephylogenyhtml) Analysis of 16S18S rRNA genesequences has been used to evaluate deep evolutionary rela-tionships among organisms and was instrumental in point-ing out that Archaea Bacteria and Eukarya should beconsidered different Domains of equivalent phylogeneticrank above kingdoms (78) However there are sufficientdifferences in 16S18S rRNA gene sequences to demonstrate
relationships (ormake distinctions) at the genus or sometimesspecies level
The first phylogenetic studies based on 16S18S rRNAgenes used sequences derived from cultures However onedoes not need cultures to obtain rRNA gene sequences (orany other sequences for that matter) An idea developed inthe lab of Norman Pace in themid-1980s involved extractionof DNA directly from natural samples and then cloning andsequencing of the DNA as a means of assaying the microbespresent in the samples (103 104) The original protocolscalled for cloning by creating what are called ldquophage librariesrdquofrom the natural DNA but since 1986 PCR has been applied
FIGURE 3 (a b) Plankton biomass in the Arabian Sea during the 1995 southwest monsoon (a) and intermonsoon period (b) Areas of theboxes indicate the relative magnitudes of the biomass in each category Categories within the dashed boxes in a b c are composed of protozoaArrows indicate the direction of energymaterial flow in the food web thicker arrows depicting greater flow Redrawn from (86)(c) Depth-integrated biomass (mgm2) in the upper 100 m of the Sargasso Sea near Bermuda and in the upper 200 m of the equatorial Pacificat 175degE Thewidth of the bars indicates the biomass in each size category Heterotrophs have been separated by size class while phytoplanktonhave not Size classes delineated by the dotted box are comprised of protozoa Redrawn from (65) doi1011289781555818821ch422f3
422-8 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
extensively for cloning and related studies The targetsequence can be almost instantly ldquoidentifiedrdquo to its closestphylogenetic neighbor by what have come to be standardonline sequence comparisons
Beyond the study of targeted genes (like the 16S rRNAgene) shotgun metagenomic studies have examined theentire genetic repertoire of the microbes in a given sampleThe metagenome is the collective genome of all organismsin the sample Initially these studies extracted DNA fromall organisms in a sample (usually prefiltered through a filterapproximately 1 microm to remove most organisms larger thanbacteria) sheared it to produce fragments and clonedthem into standard vectors either as small (thousands ofbases) or large (to hundreds of thousands of bases) insertsthat is fragments of DNA from the environment nowcloned into the vectors in a form suitable for sequencingThe best known early marine study of the former type isthe Global Ocean Survey with initial results published byVenter et al (105) which generated more than a billionbases of DNA sequence and reported 12 million previouslyunsequenced genes estimated to come from at least 1800different genomic species cumulatively in the many samplesthey analyzed
With the advent of next-generation sequencing that gen-erates millions ormore sequences in a run (known by a varietyof acronyms including 454 Illumina SOLiD etc) cloningof genes has largely been replaced by clone-free sequencingThe extent and power of such sequencing has recentlybeen demonstrated by the ability to construct essentiallythe entire genome of an uncultivated marine Group II Eur-yarchaeon that constituted only approximately 2 of amicrobial community sample using SOLiD sequencingmade possible by high coverage and the use of mate pairsequencing ofsim3000 bp fragments (106) (this length isneeded to span repeats and highly conserved genes that oth-erwise make genome construction difficult)
For 16S rRNA gene studies ldquotag sequencingrdquo pioneeredby Sogin et al (107) consists of amplifying a suitable sizedpart of the gene with broadly conserved primers (choice isimportantmdashfew are truly universal for the groups intended)often ldquobarcodedrdquo to allow multiple samples to be combinedinto a single run The amplified products are then sequencedand sequences processed en masse Many thousands of partialSSU rRNA gene sequences per sample are economically ana-lyzed this way though they are usually short (currently a fewhundred bases each depending on the sequencing platform)This way of analyzing the composition of microbial commun-ities is now standard These approaches provide so muchinformation even about very rare sequences that the resultshave led to the important concept of the ldquorare biosphererdquoorganisms that may be active or dormant and constituting avery small proportion of the community (eg often muchless than 01) but are potentially important for dispersionadaptation to changing conditions or even critical activitieslike nitrogen fixation or vitamin production (108) Howeverdue to the potential for artifacts like error sequences thisapproach requires rigorous application of quality filteringand clustering algorithms to avoid erroneous taxa and overes-timation of species richness (109 110)
Next-generation sequencing approaches have also allowedwork to begin metagenomics studies of microbial eukaryotes(111) These studies are still constrained by the much largergenomes of eukaryotes and therefore the difficulties ofobtaining sufficient sequences to reconstruct the genome ofa particular species Along with advances in sequencing abil-ity studies of eukaryotic metagenomics have been facilitated
by advances in single-cell isolation which when coupledwith high-throughput sequencing approaches reduce the tre-mendous complexity present in natural complex eukaryoticcommunities to a manageable task (112) Such single-celltechniques are also quite valuable in studies of bacteria andarchaea though the amplification technique tends to bevery uneven and typically generates less than half the genomeof each isolated cell regardless of domain (113 114)
Additionally similar to DNA mRNA is amenable toextraction and sequencing although greater care must betaken during extraction and purification as RNA shows agreater susceptibility to degradation during processing Copy-ing of mRNA by reverse transcription of RNA into cDNAfollowed by DNA sequencing has allowed insights into themetatranscriptomes of environmental samples Metatran-scriptomic studies provide information on gene expressionin an ecosystem and thereby indicates ldquoactivityrdquo of themicro-bial community rather than simply ldquopotentialrdquo represented bythe genomic DNA present in the sample with many applica-tions from showing which processes are being carried outby which organisms to fine-scaled diel studies (115ndash118)Nevertheless given variations in the lifetimes of differenttranscripts and protein molecules the transcriptome maynot be fully representative of the current activity of anorganism
Limitations of these analyses include sequencing errorsPCR mismatches or biases clustering and bioinformaticschallenges and chimeras generated during PCR Also thephylogenetic resolution of short sequences is limited giventhe high conservation of rRNA sequences Even with clone-free shotgun metagenomics there may be biases such as non-random losses of DNA during extraction and preparationor biases (eg from G +C content or secondary structure)in the sequencing procedures Determination of speciesdiversity by these approaches provides an example of theselimitations Shakya et al (119) working with synthetic com-munities (purified genomicDNA from16Archaea represent-ing 3 phyla and 48 Bacteria representing 16 phyla remixed tosimulate an environmental DNA extract) applied both meta-genomic analysis (454 and Illumina platforms) and PCRamplification followed by 454 sequencing of 16S rRNA genesto determine both species richness and relative abundanceThey found that PCR amplification454 sequencing of 16SrRNA genes yielded an accurate measure of species richness(providing that appropriate data processing was applied)but that the relative abundance of up to 94 of the species(depending on domain and variable region amplified) wasover- or underestimated by at least 15-fold (values rangedfrom not detected to 103-fold overestimation) In contrastboth metagenomic approaches yielded relative abundancesthat were within the authorsrsquo 15-fold accuracy cutoff forsim50 of the species However they concluded that addressingrichness overestimation in metagenomic analyses that isdistinguishing rare but real OTUs from experimental andcomputational artifacts awaits further computational andclassification improvements More recently Parada et al(120) used mock communities composed of 16S rRNAclones from 27 common marine taxa (from nine Bacterialand two Archaeal phyla) to show that small differences inPCR primers (and different clustering methods) can yieldlarge differences in apparent relative abundances of reportedtaxa However one primer pair and informatics pipelinethey tested using a particular version of 515F-926R (V4ndashV5) provided accurate estimates of relative clone abundance(r2 = 095) when comparing observed versus expected cloneabundance
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-9
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
Culture-Independent Diversity Studies
The first groups to be identified using cultivation-independ-ent molecular techniques from the marine plankton (121)were the bacterial SAR11 cluster (a group of closely relatedgene sequences or phylotypes) and marine picoplanktonicunicellular cyanobacteria Synechococcus and Prochlorococcus)Of these the SAR11 cluster was completely unknown butthe cyanobacteria had previously been recognized by theirunique pigment fluorescence Waterbury et al (122) andJohnson et al (123) used epifluorescence to observe Synecho-coccus and Chisholm et al (124) discovered Prochlorococcususing flow cytometry These cyanobacteria were later isolatedand grown in phytoplankton culturemedia These two groupsare generally common in the euphotic zone with SAR11 typ-ically comprising one third of the planktonic bacteria (125)and the cyanobacteria common everywhere but polar waters
Probably the biggest surprise to come from the applicationof molecular tools was the discovery by Fuhrman et al (126)of abundant archaea in the deep sea The archaea were foundto be in a unique phylogenetic cluster that was only distantlyrelated to any previously known archaea but the ldquoclosest rel-ativesrdquo (not really close at all) were extreme thermophiles Asubsequent study also found archaea to be present in near-surface coastal waters albeit relatively rare (lt2 relativeabundance) This study used a PCR technique specificallytargeted archaea and found ldquomarine Crenarchaeardquo plus a sec-ond group belonging to the phylum Euryarchaea (127)
Up until this time all known Archaea were thought to beldquoextremophilesrdquomdashadapted for either very high temperatures(thermophiles) extremely salty conditions (halophiles) orstrictly anaerobic environments (methanogens) Yet theseorganisms were present in cold or cool water at ordinary salin-ity and high oxygen concentrations Fluorescent in situhybridization (FISH) measurements from deep-sea sampleshave since showed that the archaea may make up appro-ximately 40 of the total countable prokaryotes with thepercentage reaching to 60 at 200 m depth in theMediterra-nean (35 36) An extensive time series of FISH measure-ments near Hawaii confirmed that the archaea are indeedvery abundant throughout the year from below the photiczone to at least 4000 m and typically constituting 30ndash40of the total prokaryotes present in waters deeper than a fewhundred meters (128) with similar results found elsewhere(129 130) (Fig 4) They have been reported from many pla-ces including the Atlantic Pacific and Southern Oceansand are dynamic components of the plankton the most com-mon type by far being the ldquomarine Crenarcheaeardquo (128 129131) While the marine archaea have been reported to bedominated by a few major ldquophylotypesrdquo (132) they alsohave been shown to have a great deal of microdiversity withinthese phylotypes suggesting there are many kinds of close rel-atives coexisting (133) Interestingly this group of archaeamay likely be the most abundant kind of organism on Earthgiven the huge volume of the deep sea and their high abun-dance there (134) It has recently been proposed that ldquomarineCrenarchaeardquo be elevated to their own major phylum outsidethe Crenarchaeota called the Thaumarchaeota on the basisof deep phylogenetic branching and fundamental differencesbetween them and the Crenarchaeota (135) The Thau-marchaeota possess the uniquely archaeal membrane lipidcrenarchaeol and are nowalso known to be abundant in soils
The physiology of the Thaumarchaeota has been anintriguing area of study Initially an autoradiography-FISHcombination approach demonstrated that they take upamino acids (130 136) But subsequent data showed that
crenarchaeol was derived largely from CO2 suggesting auto-trophic metabolism (137 138) An experiment showinguptake of 13C-labeled bicarbonate into these archaeal lipids(139) directly pointed to autotrophy in this group Chemoli-thoautotrophywas first hinted at byVenter et al (105) whosemetagenomic analysis showed an apparent archaeal scaffoldthat contained genes suggestive of ammonia oxidation Fur-ther evidence came from Schleper et al (140) who detectedseveral ammonia oxidation genes in order directly adjacent toa Thaumarchaeota 16S rRNA gene in a soil-derived metage-nomic clone The issue was directly resolved when Konnekeet al (141) isolated a related marine archaeon from sedimentof a marine aquarium and this organism Candidatus Nitroso-pumilus maritimus was found to have a chemoautotrophicmetabolism quantitatively oxidize ammonia to nitrite andcontain an archaeal ammonia monooxygenase gene (amoA)and interestingly did not grow heterotrophically Cultivationexperiments showed this organism has a high affinity forammonium allowing it to outcompete bacterial nitrifiers atlow (submicromolar) concentrations as are typical in thesea (142) The complete genome of the organism has shownnovel adaptations for nitrification and autotrophy (143)
Even if the Thaumarchaea are primarily chemolitho-autotrophs field data suggest a level of mixotrophy in thatsome organic substrates are being incorporated into biomassA stable isotope study using cells collected at 670 m depthoff Hawaii estimated that about 80 of the carbon incor-porated into archaea-specific lipids came from inorganic sour-ces and about 20 from organic compounds (144 145)Interestingly it has also been claimed that the genomesof deep-sea (gt2000 m depth) members of the Thaumarch-aea as well as those living in equatorial waters rarely containthe amoA gene and thus may be primarily chemoorgano-trophs (48)
Major bacterial groups that have been documentedfrom seawater using 16S rRNA characterization includesome that are also known from culture (eg AlteromonasRoseobacter) and several that are phylogenetically distantfrom standard cultures The most common groups are inrough order of their relative abundance in clone librariesfrom most to least abundant SAR11 (relatives of Pelagibac-ter ubique) Roseobacter SAR86 cyanobacteria SAR116SAR202 SAR234 andMarine Group A The SAR designa-tion followed by a number is an arbitrary sequential cloneidentifier from Sargasso Sea cloning studies done by the Gio-vannoni lab that did most of the early systematic cataloging ofclones Summaries of the data and phylogenetic relationshipsof these groups can be found in Fuhrman andHagstrom (102)and Giovannoni et al (101)
Molecular Genetic Discoveries in Bacterial andArchaeal Marine BiologyAs described earlier metagenomics is an extension of theideas used in the 16S rRNA cloning studies in that all genesfrom the native microorganisms are separated and clonedwithout having cultivated the organisms These methodsearly on started to find unexpected and very interestingresults The best examples involve unexpected marine photo-trophy the first of which is discovery of a nonchlorophyllphotosynthetic bacterial pigment called proteorhodopsin(146) The gene was found on a large environmentallyderived fragment of DNA that also had a gene coding for16S rRNA from the so-called SAR86 group (one ofthe groups common in seawater) This pigment can act as alight-driven proton pump thought to permit cells to generate
422-10 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
FIGURE 4 Distribution of Bacteria Euryarchaea andmarine Crenarchaea (recently renamed Thaumarchaea) along a transect in theNorthAtlantic Ocean as measured by CARD FISH and expressed as of total bacteria + archaea counts via DAPI stained epifluorescence fromTeira et al (130) Top panel shows station locations that are shown on the top of the lower three panels with the distance in km reported alongthe transect from lower to higher station numbers doi1011289781555818821ch422f4
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-11
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ATP from sunlight Interestingly different versions of this pig-ment are found at different depths apparently ldquotuningrdquo theabsorption to match the ambient wavelengths of light asthey change with depth (147) The proteorhodopsin genesare very widespread and diverse perhaps in half or more ofall marine bacteria including SAR11 and Euryarchaeota(105 148ndash150) However almost all of several culturedorganisms with proteorhodopsin studied to date do not showa growth benefit from light (150) with the exception of amember of the Flavobacteria Dokdonia sp strain MED134that grows faster in the light only under moderately lownutrient conditions (151) and a marine Vibrio has beenshown to survive starvation longer in the light than in dark-ness (152) These observation may explain the wide phyloge-netic distribution and high abundance of this gene in thatproteorhodopsin may often assist long-term survival of bacte-ria under extreme energy-limiting conditions yet not havemucheffect on growthduringmore energy-replete conditionsInterestingly a recent report suggests the growth benefit fromlight in Dokdonia MED134 is from enhanced uptake of itsrequired growth factor vitamin B1 which is transported by atonB-dependent transporter powered by a proton gradient(153) This pigment might also have sensory or other rolesnot yet well examined (150)
The second kind of phototrophy found by moleculargenetic and fluorescence techniques to be unexpectedlyimportant in marine plankton is anoxygenic aerobic bacterialphototrophy based on the pigment bacteriochlorophyll aCulturable aerobic anoxygenic photosynthetic (AAP) bacte-ria have been known from seawater for several years (154) andare widespread (155) Initial recent reports of direct countsclaimed they represent about 11 of the total bacterial com-munity in the euphotic zone (156) but those early counts didnot correct for the presence of other bacteria and such correc-tionoften yields estimates averaging closer to 2(157)Directmeasurements show the bacteriochlorophyll pigment is rela-tively rare (158) However studies in various ocean locationsincluding ocean gyres report that these AAP bacteriacan indeed on occasionmake up a quarter of the total prokar-yotes (159 160) A recent energetic modeling of AAP andproteorhodopsin-containing bacteria (161) has suggestedthat proteorhodopsin-containingones cangain approximately02asmuchenergy from sunlight asSynechococcus (commonmarine cyanobacterium) and AAP bacteria can gain approxi-mately 13 as much energy from sunlight as SynechococcusThey concluded the AAP bacteria may gain energy sufficientto meet maintenance costs but proteorhodopsin-containingbacteria were not expected to do so except at high light inten-sities andwith largenumbersof proteorhodopsinmoleculespercell They concluded the ease and low cost of maintainingPR-based phototrophy (a few genes required) may explainthe high incidence of proteorhodopsin genes
Other metagenomic studies have yielded interestinginsights about organisms and processes that would otherwisebe difficult or impossible to establish For example thesequence of an archaeal gene fragment isolated from seawaterrevealed extensive evidence of genetic exchange with othertypes of archaea and even bacteria (162) Such geneticexchange was also quite evident from the complete genesequences of different strains of Prochlorococcus and Synecho-coccus which was attributed in part to virus-mediated geneflow (163ndash166)
Dilution Cultures of Bacteria and ArchaeaSome recent cultivation techniques that permit growth ofldquotypicalrdquo oligotrophic marine bacteria in pure culture are
based on the simple concept that bacteria living in seawateron dissolved organic matter might best be grown in the labo-ratory in ordinary filtered seawater This conclusion followsfrom similar techniques to grow mixed ldquoseawater culturesrdquoof marine bacteria (167) but the seawater inoculum in thiscase is diluted so that only one or a few bacteria are addedto the initial culture vessel (168) A rapid throughput versionof this method has been used to cultivate members of bacte-rial groups thought to be conventionally uncultivable such asthe SAR11 clade (169 170) albeit often at very low den-sities Such cultures permit focused studies of the propertiesof those organisms including genomic analysis (169 170)This valuable work provides insight into the roles and activ-ities of some of the most common bacterial phylotypesobserved in the ocean (171) showing for example that culti-vated SAR11 requires a source of reduced sulfur as well asproviding templates aiding the interpretation of metage-nomic analysis (172)
PROTOZOADiversity and DistributionHeterotrophic protists have been identified historically fromtheir morphological features that are apparent at the levelof the light or electronmicroscope Features of cell size shapetype and pattern of flagellationciliation skeletal structurescharacteristics of the nucleus and other cellular structuresand organelles have been used to differentiate themany thou-sands of described species A description of the extent of thisdiversity of form and function is well beyond the scope of thisbook (see [79] for more information) However much of thisdiversity can be reduced to three basic body plans that dictatethe broadest ecological roles of these cells amoeboid flagel-lated and ciliated forms (Fig 5) Among the lineages possess-ing one of these three forms only the last group the ciliatedprotists constitute a monophyletic group within the domainEukarya
One of the simplest body plans for protozoa is the amoe-boid cell exemplified by the gymnamoebae or ldquonakedrdquoamoebae Motile nonphotosynthetic cells lacking flagellaor cilia occur as life stages in a number of phylogeneticallydiverse taxa but for many free-living species of protozoathis form constitutes the only life stage Motility is largelyconfined to movement along surfaces by means of pseudopo-dia that can take on a variety of (species-specific) shapes Sig-nificant abundances of amoebae are largely confined tobenthic and epibiotic environments (174) and to suspendedparticulate material where they can occasionally be highlyenriched (27) Most amoebae consume bacteria and otherminute prokaryotes and eukaryotes
Several heterotrophic protistan groups possess complexamoeboid body plans most notably the foraminifera polycys-tine and phaeodarian radiolaria and the acantharia Plank-tonic forms are predominantly oceanic in their distributions(although a large number of benthic species of foraminiferaexist) These species are heterotrophic and many are visibleto the naked eye (individual cells can bege1 cm gelatinouscolonies can form ribbon-like structuresge1 m in length)They feed on a wide variety of bacterial protistan and meta-zoan prey using pseudopodial networks have rather longcomplex life cycles for individual cells (weeks to months)and are extensively used in studies of paleoclimatologicalreconstruction (82 175) In addition many of these speciespossess intracellular symbiotic usually eukaryotic algaeThe widespread occurrence of algal endosymbiosis among
422-12 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
these protozoa in oceanic pelagic environments implies thatstrong selective forces appear to give rise to these associations(97 98)
Flagellated forms of protozoa exist across many protistanlineages Eukaryotic flagella come in various sizes numbersand forms (eg with or without tiny hairs) that are character-istic of the different lineagesMany flagellate species are appa-rently purely phototrophic or heterotrophic (phagotrophic)but a significant (still poorly known) fraction of flagellates aremixotrophic combining both phototrophy and phagotrophy(70 88) Flagella in phagotrophic flagellates are employed formotility and prey capture Most free-living flagellates possessone to four flagella (typically one or two) that can be manytimes the length of the cell itself Although there is tremen-dous species diversity among flagellated protozoa many ofthese species have broadly overlapping ecologies As a groupflagellates are the most numerically abundant protozoa ofboth benthic and pelagic ecosystems and they are fundamen-tally important as consumers of bacteria cyanobacteriaand other eukaryotes Flagellated protozoa within the nano-plankton size class (2ndash20 microm) are often counted as a singleassemblage in plankton studies because morphological details
apparent using light microscope are insufficient to distinguishamong the many species This assemblage has been variouslyreferred to by a variety of names and acronyms including het-erotrophic nanoplankton (HNAN HN) heterotrophic flag-ellates (Hflags) heterotrophic nanoflagellates (HNF) andmicroflagellates Heterotrophic nanoplankton is the mostaccurate term for most methodologies employed to countthese cells because it does not require visualization of flagella(which are often lost from these small cells during preparationfor microscopy) The confusion over terminology is partly aconsequence of the different methodologies that have beenused to count these species and partly a consequence of thefact that these species were first studied prior to the wideacceptance of the size convention of Sieburth et al (84)
Heterotrophic flagellates in the microplanktonic size class(20ndash200 microm mostly dinoflagellates) are important consum-ers of phytoplankton in pelagic ecosystems Many of thesespecies are capable of the production of large pseudopodialnets the pallium that can engulf prey (particularly diatoms)significantly larger than the diameter of the dinoflagellatetheca (176) This behavior and its ecological significance forenergy flow in plankton communities have been recognized
FIGURE 5 Body plans and size ranges of protozoa These micrographs depict amoeboid (andashg) flagellated (hndashj) and ciliated (kndashm) forms ofprotozoa From Caron et al (173) Markers bars are 5 (i) 10 (h) 20 (c j l) 30 (a) 50 (k m) 100 (b f) 500 (d) and 1000 (e g) micromdoi1011289781555818821ch422f5
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-13
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
only within the past few decades (177) These species areoften abundant in waters where diatoms dominate (178179) In such situations heterotrophic dinoflagellate biomasscan be similar to that of ciliated protozoa (180)
Ciliates are generally the most recognizable form of pro-tozoa to nonspecialists Nonetheless ciliates are a diversegroup morphologically as well as ecologically (181 182)The degree of ciliature on these species can range from uni-formly ciliated to totally devoid of cilia during most of theirlife cycle to ciliature restricted to specific regions of thecell In addition cilia can fuse to form various complexstructures (eg ciliary membranes or veils cirri) that assistin locomotion food capture or attachmentCiliates are abun-dant and ecologically important species in both benthic (8)and pelagic (183) ecosystems and can consume a variety ofprokaryotic and eukaryotic prey Together with the heterotro-phic dinoflagellates these species are the dominant consum-ers of phytoplankton in many pelagic ecosystems (184) andas a consequence they form an important trophic link to met-azoan zooplankton (185 186)
Diversity and Biogeography of Protists FromMorphology to DNA SequencesUntil relatively recently the biodiversity of marine protistanassemblages was not generally considered a controversialtopic It was generally accepted that while all species of pro-tists certainly have not been identified representatives ofmost types of algae and protozoa had been observed anddescribed if not actually brought into culture in the labora-tory Direct sequencing of 18S rRNA genes from environ-mental samples (as described in ldquoMolecular Phylogeny andMetagenomics Field Applicationsrdquo) has changed that viewInitial forays into environmental DNA indicated a muchgreater diversity of protists than previously documented usingclassical approaches of culture and microscopy (81 187ndash191) Numerous publications over the past decades haveexpanded these findings (see [192] for a recent global analy-sis) which are highly analogous to discoveries in marine pro-karyote research as described already
Previously uncharacterized protistan diversity has nowbeen documented at virtually every level of eukaryoticorganismal classification Some of these findings couldhave been expected but some have been very unexpectedFor example a much greater diversity than noted previouslyhas been observed among small (lt10 microm) protists in plank-tonic ecosystems These species generally possess few distinc-tive morphological features and one could expect that manycryptic species might be present among these small morpho-types The molecular ldquodiscoveryrdquo of this eukaryotic diversityhas stimulated progress on the isolation and descriptionof new species and genera of minute algae and protozoa(193ndash197)
Analyses of environmental samples have also indicatedthe unanticipated existence of novel 18S rRNA gene sequen-ces that imply the presence of novel lineages of eukaryotes innatural protistan communities (198ndash200) These are sequen-ces for which there are apparently no known described orcultured species The degree to which these sequences dif-fer from sequences of known sequenced eukaryotes impliesthat some of these lineages may be distinct at the level ofphylum (201)
These findings have raised basic questions and somedebate concerning the true diversity and biogeography ofprotistan assemblages in natural ecosystems (202ndash204) thevalidity of the many novel phylotypes or cryptic species that
are being documented (205) the potential importance andsignificance of the many rare taxa that characterize thesecommunities (206) and the implications of these findingsfor the ecologicalbiogeochemical roles that protists playin aquatic ecosystems For example it was postulated andsubsequently confirmed that some novel alveolate lineagesreported from marine ecosystems make up a suite of parasiticprotozoa whose ecological importance may have been signifi-cantly underestimated in the past (207) Deciphering theidentity and significance of these many unknown phylotypeswill constitute a significant effort for protistologists in thefuture (80 85)
Life Histories and Ecological Strategies of ProtozoaAbundance prey type and life histories all vary tremendouslyamongmarine protozoa The smallest species (eg many flag-ellates) tend to be the most abundant and widely distributedin the world ocean Indeed many of these species may beglobally distributed (208) Most nanoplanktonic flagellateshave potentially rapid rates of grazing and growth Underoptimal conditions these species can divide by binary fissionseveral times a day and thus dramatic increases in their pop-ulations can take place in response to favorable conditionswithin a few days (209) Many of these species are capableof surviving for limited periods of time without food andhave developed a variety of physiological or life cycle strat-egies to cope with these events (209) However in contrastto some bacteria that may remain viable through long periodsof starvation (210) protozoa will expire or encyst in responseto low food abundance and thus ldquoboom-and-bustrdquo popula-tion changes are characteristic of small flagellates
On the other end of the size spectrum from the ratherubiquitous nanoflagellate species many of the largest proto-zoan species (polycystine and phaeodarian radiolaria plank-tonic foraminifera) are exclusively oceanic (ie do notsurvive in most coastal environments) andor have specificlatitudinal and depth distributions Thus the abundancesof these latter species may range from undetectable to max-imal abundances of gt105 individualsm3 Dramatic changesin the abundance of these protozoan taxa also can be relatedto changes in prey abundance physicalbehavioral aggrega-tion or to periodicity of life cycle events For example theplanktonic foraminifer Hastigerina pelagica reproduces on alunar cycle and thus abundances (and life stages) of this spe-cies in oceanic waters can vary considerably over the courseof a month (211) In general life cycles for the large amoe-boid protozoa are lengthy and complex (for single-celledorganisms) with life spans unknown for many species(attempts to culture them in the lab have so far been unsuc-cessful) but estimated to be on the order of months to per-haps years (175)
The abundance and activities of microplanktonic hetero-trophic protists (mostly ciliates and heterotrophic dinoflagel-lates) tend to be somewhat intermediate to those ofnanoplanktonic flagellates and the larger amoeboid formsThese species are present in the majority of marine ecosys-tems and collectively play an important role in the controlof phytoplankton biomass (and probably the abundance ofnanoplanktonic protozoa although there is little informa-tion on this topic) in waters throughout the world oceanLike small flagellates ciliates and dinoflagellates reproduceprimarily by binary fission but their maximal growth ratesare typically slower (one division a day is typical) and theirlife cycles often include sexual phases that allow for geneticrecombination
422-14 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
VIRUSESViral Abundance and General PropertiesViruses are simple biological agents typically 20ndash200 nm indiameter composed of a nucleic acid genome in a proteincoat that infect cells and ldquocommandeerrdquo the cellrsquos machineryto make more viruses which are released into the environ-ment when the host cell lyses or bursts A particular virus isthought to be capable of infecting only a narrow range of hosts(usually one species sometimes a genus rarely broader)Although some early studies had isolated viruses from thesea therewere no data prior to the 1980s showing such viruseswere very abundant and more significantly no evidence thatinfection was occurring in any important part of the planktoncommunity It wasnrsquot until the late 1980s that electronmicro-scopy with suitable concentration methods showed thatviruses are extremely abundant similar to or even higherthan bacterial abundance (212 213) Interestingly themost suitable transmission electron microscopy (TEM)approach used by Bergh et al (described in detail by [214])was actually similar to a direct centrifugation method devel-oped in 1949 (215)
TEM studies of viruses permit high-resolution images andobservation of viral morphology showing features such ashead diameter tails sheaths and tail fibers (Fig 6 collage
in upper left) Each type of virus has a fixed morphology(unlike bacteria that are potentially more plastic in theirappearance) and hence a coarse measure of viral diversityhas been possible by cataloging morphologies Studies thathave done so have found dozens or more different morpholo-gies of marine viruses in a given sample (217ndash219) A largeproportion resemble bacteriophages which are viruses thatinfect bacteria
The availability of brightly fluorescent nucleic acid stainsand high-porosity fine pore size (002 μm) filters made fromAl2O3 have made it easy to accurately count viruses by epi-fluorescence microscopy (38 220 221) Epifluorescence per-mits abundance estimates but does not allow observation ofviral morphology as viruses are below the resolution limitof light microscopy viruses are visible only as sources of light(like stars in the night sky see Fig 6 lower micrograph) Anextension of manual epifluorescence counts is the use of flowcytometry to count viruses now commonly employed in somelabs (222)
Virus abundance has been found to be closely related tobacterial abundance with a virusbacteria ratio typically10ndash301 A typical oceanic profile of bacterial and viral abun-dance is shown in (Fig 6 panel on right) This tight ratioand the strong correlation to bacterial abundance in relationto weaker correlations to chlorophyll have been cited as
FIGURE 6 Marine viruses (upper left collage) viewed by transmission electron microscopy These are cyanophages in the groups myoviridae(right and bottom) siphoviridae (left) and podoviridae (top) Scale bars represent 100 nm From Sullivan (216) Epifluorescence micrograph(lower left) of SYBR green I stained viruses (small fluorescing objects) and bacteria + archaea (large fluorescing objects) Depth distribution ofvirus and bacteria + archaea abundances obtained by epifluorescence microscopy of SYBRGreen stained cells in the central San Pedro BasinCalifornia (11 August 2000) doi1011289781555818821ch422f6
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-15
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
evidence that most marine viruses infect bacteria rather thaneukaryotic phytoplankton (217 218 223 224) though thereare also many important viruses of phytoplankton and otherorganisms with significant impacts on primary productivityblooms and ecosystem function (225ndash227)
The first demonstration of the activity of marine viruseswas presented by Proctor and Fuhrman (228) who showedby electron microscopic examination that viruses wereactually infecting marine bacteria and cyanobacteria at ameasurable rate Because only the final portion of the viruslife cycle is visible by electron microscopy (when the virusesare assembled and ready to lyse the host cell) only a very smallfraction of the infected cells can be counted as infected at anygiven time Proctor and Fuhrman (228) reported that only afew percent of the total bacteria from coastal waters and theSargasso Sea were visibly infected but they interpreted thedata with a model that implied the actual fraction of the totalcommunity that was infected was much higher They con-cluded that the total fraction of bacterial mortality attribut-able to viruses is roughly 10ndash40 Subsequent studies haveused refined versions of that same general approach andmodel (229 230) and numerous studies have used a varietyof alternative approaches to estimate viral activitymdashall yield-ing basically the same general conclusion (although themodel parameters need to be adjusted for cyanobacteriawhich on average seem to be infected less than bacteria)These approaches include (a) calculation of virus turnoverfrom decay estimates (231) (b) measurement of viral DNAsynthesis by incorporation of tritiated thymidine (232) (c)observation of effects of added viruses (233 234) (d) estima-tion of bacterial mortality in the absence of protists (e) use offluorescent viruses to estimate production by an approachanalogous to isotope dilution (38) and (f) dilution of virusesin filtered samples to estimate viral production (235) Theoverall consensus of these studies is that the initial estimateswere basically correct that is that viruses are responsiblefor about 10ndash40 of the bacterial mortality in most marinesystems (reviewed by [218 223 226 236 237]) Howeverthere is still some disagreement about the higher estimatesand it is likely that 40 mortality from viruses alone is nottypical for most marine systems
Most of the foregoing work has focused on the viral infec-tion of bacteria primarily thought to be heterotrophicalthough the original report by Proctor and Fuhrman (228)also noted the occurrence of cyanobacterial infection Sev-eral subsequent studies focused on phytoplankton includingthe potential effect of viruses on the termination of phyto-plankton blooms of Emiliania and Phaeocystis and the likelyimpact this might have on release of climate-active gasessuch as dimethyl sulfide (225 227 238ndash247) Althoughdetails are beyond the scope of this chapter viruses arethought to infect virtually all marine organisms with poten-tially significant impacts from zooplankton to whales (226)Detailed studies of viruses infecting cyanobacteria such asSynechococcus have shown differences in viral host specific-ity particularly toward coastal and oceanic host strainsand some occasional high virus abundances (to 105ml) asmeasured by most probable number (MPN) cultivation tech-niques in the Gulf of Mexico near Texas (248 249) Othervirus cultivation studies with Prochlorococcus and Synecho-coccus in oligotrophic waters of the Sargasso Sea showedan interesting pattern of cross-infection between these gen-era by some virus types (suggesting gene flow among theseorganisms) but generally low MPN estimates of abundanceto 103ml even when cyanobacterial abundance was near105ml (216 250)
Overall the consensus emerging from direct comparisonsof viral-mediated mortality and grazer-mediated (ie micro-zooplankton) mortality have indicated that viral lysis of bac-teria constitutes a significant fraction of total mortality ofthis assemblage while the mortality of phytoplanktonappears to be dominated bymicrozooplankton inmost instan-ces (251 252)
As mentioned viruses have significant morphologicaldiversity as observed in TEM studies It is also possible toinvestigate aspects of their genetic diversity In early workon this topic Wommack et al (253) and Steward et al(254) observed the diversity of viral genome lengths in a fieldsample by pulsed field gel electrophoresis with viral genomesranging 25 kb to gt300 kb in length Field results show thatthe viral community composition is dynamic in space andtime with clear changes in the banding patterns over seasonsand locations in Chesapeake Bay (253) between oceanbasins and subsequent to dinoflagellate blooms (254) andwith depths to 500 m at one location (255)
Although viruses do not all share a set of core genesthat allows a universal viral phylogeny (comparable to SSUrRNA gene in cellular organisms) genetic diversity amonga single group of viruses can be examined by sequence analysisof shared genes within the group An example is the g20 geneinT4-like cyanophages that has shown extremely high diver-sity even among very closely related viruses and has demon-strated geographic and seasonal variation (256) A secondexample is g23 which is found broadly in diverse and wide-spread T4-like phages (257) and can have seasonally repeat-ing patterns (258) as well as short-term rapid dynamics thatcorrelate to those in bacteria (259)
Viral MetagenomicsAs with cellular organisms metagenomics can be used toexamine viral diversity and genetics without themany restric-tions of cultivation Viruses can be collected by selective fil-tration and concentrated by tangential flow filtration orflocculated with iron chloride (260 261) then their collec-tive metagenome can be extracted linker amplified and ana-lyzed by sequencing (262 263) Viral metagenomic studiesare particularly challenging because the large majority ofsequences have no annotated matches in any databases butso far results from marine samples around the world haveshown extremely high diversity and variations with depthlocation and time presumably with highly dispersed typesselected by local conditions (262ndash266) Because metage-nomes are best interpreted when there are representative cul-tures available the best matches of viral metagenomesoriginally tended to be to the few viral isolates infecting trulycommonmarine bacteria like cyanophage infecting Synecho-coccus and Prochlorococcus (267) However the developmentof dilution-to-extinction cultures representing common het-erotrophic marine taxa like Peligibacter (a member of theSAR11 clade) and SAR116 has allowed isolation from sea-water of viruses infecting these common organisms and theseisolates have indeed been found to be highly abundant inmarine viral metagenomes (268 269) New approaches tointerpret the results include clustering the proteins independ-ent of known proteins to compare samples to each other andtry to find environmental factors driving viral communitychanges (270) Such analysis of a large global data set (TaraOceans expedition) has shown that extensive sampling hascome close to reaching the total diversity in tropical and tem-perate waters of such viral protein clusters (which essentiallyrepresent various viral protein motifs not all viral proteintypes) and that viruses appear to be directionally dispersed
422-16 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquodownstreamrdquo in major ocean currents as one may expectfrom first principles but also supporting the ldquoseed bankrdquohypothesis of viral biogeography (271)
Viruses and Host DiversityViruses are themselves thought to be instrumental in drivingincreased diversity of their microbial hosts via a hypothesizedprocess often called ldquokill the winnerrdquo This is because viralinfection is host-specific and density-dependent the latterbecause viruses diffuse from host to host so an abundanthost is more likely to pass on infection than a rare oneThis means that if an organism becomes abundant andblooms winning the competition for resources it becomesmore susceptible to a viral epidemic This would benefit therarer organisms and thus help foster diversity (223 272273) However bloom scenarios as described above involvesystems far from steady state and the formal theory of Thing-stad and Lignell (274) has interesting steady-state solutionswhere several viruses infect several hosts stably over time(via trade-offs between growth rates and viral susceptibility)which may occur at the strain or species level (274) There issome experimental evidence that viruses have effects on nat-ural marine microbial community composition weaker thanthe kill the winner (bloom version) hypothesis would sug-gest (275 276) so something resembling the steady-statecoexistence described in the model may in fact be commonThere also appear to be processes that foster coexistencebetween viruses and hosts but the mechanisms are largelyspeculative (223 277 278)
Viruses may also be directly involved in host genetic diver-sity because they can be the agents of genetic exchangebetween microorganisms (223 273) This often involvesthe viral lifestyle known as lysogeny whereby viruses survivewithin host cells as DNA only integrated into the host chro-mosome and being reproduced each time the host divides Ahost harboring such a genome is called a lysogen becauseunder conditions of stress to the host cell a genetic switchmay cause the viral genome to initiate the lytic process pro-ducing many progeny viruses and bursting from the hostLysogeny is a very common property occurring in a signifi-cant part of the bacterial community (279 280) althoughthe incidence of induction of the lytic phase in nature is appa-rently low (281) Overall lysogeny is poorly understood butthought to have both positive and negative impacts on themicrobial community (282) Recent results suggest that ben-efits of being lysogenic in highly seasonal polar seas leads tofundamental difference between polar and other marine viralcommunities (283)
Viruses and the Microbial LoopAs part of the food web viruses occupy a unique positionThey infect host cells that are mostly thought to be heterotro-phic bacteria and by doing so they typically burst the hosts torelease progeny viruses and cellular debris But what is the fateof this material Viruses themselves do not last indefinitelyand a simple steady-state assumption implies that from eachburst of viruses (typically 20ndash100 per lytic event) only onesuccessfully infects another cell The rest are inactivatedand broken down by sunlight (UV and visible exposure)and enzymatic attack (284) or consumed byminute phagotro-phic protists (285) thus reentering the food web as substratefor bacteria or food for protistan consumers Experiments incontrolled laboratory systems and field studies with radioac-tively labeled viral lysis products have supported the conclu-sion that most of the organic matter released by the viralinfection is either taken up by bacteria or respired (286 287)
Modeling this process as part of the microbial loop showsthat viral lysis represent a sort of side loop that has the neteffect of remineralizing a significant amount of the carbonand nutrients that enter the bacteria-protist part of themicro-bial loop (Fig 7) A theoretical numerical steady-state modelcomparing a systemwith no viral activity to onewhere virusesare responsible for 50 of bacterial mortality showed that thesystem with viruses had 33 more bacterial production andrespiration than the virus-free system implying that theviruses had the effect of permitting the bacteria to processmore of the primary production than they would otherwise(223) Although 50 is a high number unlikely to be com-mon in the sea this model nevertheless illustrates that virusescan reduce the amount of energy reaching higher trophic lev-els The implication is that viruses lead to increased bacterialactivity at the expense of the larger organisms
MAJOR ENVIRONMENTAL CONTROLSLight Temperature and PressureTemperature has an important potential influence on bio-chemical reactions and therefore on biological processesin general Most ocean waters fall in the range of ndash2degC to30degC with obvious exceptions in hydrothermally heatedareas Temperature has long been known to be a regulatingfactor for the growth of heterotrophic microbes
In temperate waters it has been established that microbialactivity is generally much higher in warm summer waters thanin winter (55) The relationship is not simple howeverbecause multiple factors act at the same time Some contro-versy still exists regarding the highest and lowest extremesfor marine bacterial growth although there is broad agree-ment that bacteria grow gt100degC at hydrothermal vents andltminus5degC in sea ice brines Pomeroy et al (57) noted the inter-esting observation that bacteria seem particularly inhibitednear the freezing point of seawater (ca ndash22degC) comparedto eukaryotic phytoplankton This effect results in polarspring phytoplankton blooms that accumulate organic car-bon in advance of the response of the bacterial communityand development of the microbial loop and perhaps lead toenhanced benthic-pelagic coupling (288)
The relationship between temperature and the growth rateof marine phytoplankton was described broadly in a now
FIGURE 7 Modification of the microbial loop concept that incor-porates the functional role of viruses Export can be via predation orsinking From Fuhrman (223)doi1011289781555818821ch422f7
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-17
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
classic paper by Eppley (58) and later Goldman and Carpen-ter (289) Temperature was shown to exert a strong and directeffect on themaximal growth rates of these species withmax-imal intrinsic growth rates at 0degC generally lt1 divisiondaywhile growth rates at 30degC may be gt4day These relation-ships indicate the maximal rate that might be attained byphytoplankton at these temperatures but they do not takeother factors into account (nutrients light) While warmtemperature ostensibly allows more rapid growth it createshydrographic conditions that typically give rise to nutrientlimitation of algal growth Thus some of the coldest watersin nature witness some of the most massive phytoplanktonblooms (290) while warm oceanic gyres represent some ofthe most oligotrophic areas of the ocean The multiple andoften conflicting effects of temperature on the growth of nat-ural phytoplankton assemblages limit the accuracy of presentpredictions regarding how primary producers will respond toclimate change (291)
An analysis of the effect of temperature on the growth ofheterotrophic protists has indicated that the response is qual-itatively similar to that of phytoplankton but with a twistThe growth rates of at least some protozoa can exceed thoseof phytoplankton at warmer environmental temperaturesbut the opposite effect is apparent at very low environmentaltemperature This differential effect of temperature on thegrowth of phototrophic and heterotrophic protists was basedon a large meta-analysis of published protistan growth (59)That analysis demonstrated that the maximal growth ratesattained by phototrophic protists could exceed the maximalgrowth rates attained by heterotrophic protists (all otherpotential growth-limiting factors not considered) Thereforeprotozoan growth rates may be constrained to a greater degreeat low environmental temperature than rates for phytoplank-ton If so then phytoplankton bloomsmay get a head start ongrazers during spring in polar ecosystems This scenario is con-sistent with information on seasonal biomass changes andmicrozooplankton herbivory in the Ross Sea Antarctica(292 293) but there are still too few data to fully vet thishypothesis
The importance of high pressure on bacterial growthgained considerable attention in the late 1960s when thedeep sea submersible Alvin was accidentally lost overboardwith its hatch openWhile no lives were lost in this accidentsome workmenrsquos lunches sank to the bottom (sim1500 m)inside the submersible Alvin was recovered after 10 monthsand interestingly there was a waterlogged lunch containingapples bologna sandwiches and broken vacuum bottleswith broth that all appeared hardly degraded and tasted palat-able Yet when placed in a refrigerator on the ship these itemsdegraded relatively quickly Initially pressure was thought tobe the preserving factor as the sea floor temperature was sim-ilar to the refrigerator temperature and the only major differ-ence would be pressure (294) Following this observationa series of experiments to measure degradation of variousorganic materials left in the deep sea for extended periodsindicated that degradationwas typically significantly reducedimplying that pressure reduces the degradation rates (295)Nonetheless changes in the protozoan community of naturaldetrital material sinking to the deep ocean floor indicate thatthe microbial community can respond relatively quickly insome situations (296) Deep sea microbiology has advancedconsiderably yet it is still difficult to interpret results withrespect to actual in situ rates of naturally occurring organicmatter Deep sea bacteria adapted to high pressures havebeen isolated that are barophilic (also called piezophilic)meaning that they prefer high pressures and have reduced
activity at lower pressures whereas others are barotolerant(piezotolerant) tolerating but not preferring high pressuresfor example see (297 298)
Few data are available on barotolerantbarophilic marineprotozoa Protozoa certainly exist and grow at great oceanicdepths but measurements of in situ growth rates for these spe-cies do not yet exist Measurable protozoan numbers havebeen documented in the deep-sea sediments for more than30 years (299 300) and viable protozoa have occasionallybeen cultured from these environments (301ndash305) butvery few direct measurements of the activities of these speciesin situ have been reported (306) A few protozoa have beenisolated that will grow at high pressure (302 303 307) andprotists that appear to be unique to the deep ocean havebeen observed either directly or through the analysis ofDNA sequences (308ndash311) but possibly the best direct evi-dence that protozoan activity takes place at the high pressurescharacteristics of the deep sea are experimental and observa-tional work noting the stimulatory effect that detrital deposi-tion has on some components of the protozoan community(296 312) These observations indicate a diverse and activeprotozoan fauna of the deep ocean although their biogeo-chemical significance is largely uncharacterized at this time
Dissolved and Particulate Organic MatterBacteria and archaea are thought to be by far the most impor-tant organisms with respect to the processing of dissolvedorganic matter (DOM) and nonliving particulate organicmatter (POM also called detritus) in the ocean While theremay be some uptake of DOM by protists particularly forgrowth factors needed in trace amounts such as vitamins(313) the bulk of this material is probably utilized by bacteriaand archaea (314) including the smallest cyanobacteriumProchlorococcus (315 316) which thus may be considered amixotroph Due to their small size bacteria have extremelyhigh surfacevolume ratios and combined with their over-whelmingly high abundance an extremely high integratedsurface area Protozoa tend to obtain the organic materialsthat they require for growth from their prey rather thanthrough the uptake of DOM Overall protozoa tend to besources of dissolved and detrital organic substances throughthe excretion of unassimilated prey biomass in expelledfood vacuoles
Particulate organic matter is not directly available as sub-strate to bacteria These substances must first be reduced tosmall molecules that can be transported into the cell Thisis accomplished by the production of extracellular enzymes(note that few if any large polymers are directly taken up bybacteria with the possible exception of DNA) Hydrolyticenzymes produced by bacteria (and almost certainly archaea)break down polymers like proteins polysaccharides andnucleic acids As with DOM POM is composed of a complexmixture of compounds that vary in their susceptibility to bac-terial degradation and utilization
Particulate material in the water column serves not only asbacterial substrate but also as substratum POM occurs in thewater column across a huge size spectrum from micrometersup to some detrital aggregates more than 1 m in diameter(317) Much of this particulate material is in a constant stateof flux with colloidal material constantly coalescing andaggregating to form new or larger particles (318) as microbialdegradation acts simultaneously to remineralize this materialDetrital particles that attain macroscopic size either by directformation (317 319 320) or via accretion and aggregation(321) and are often called marine snow or macroaggregatesMarine snow particles are readily colonized by bacteria and
422-18 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
many other microorganisms and their abundances on theseaggregates can be orders of magnitude greater than abundan-ces in an equivalent volume of the water surrounding theaggregates (320 322ndash324) and their diversity and highlyconcentrated abundances on detrital aggregates implies thatthere may be stronger trophic coupling and efficient energyelemental cycling relative to these processes among free-living microorganisms in the surrounding waters This specu-lation has resulted in a number of formalizations of the poten-tial impact of aggregate microbiology on biogeochemicalprocesses in the ocean (325 326) Most definitions of bio-films include aggregated particulate and these particles areoften studied within the biofilm conceptual model (see chap-ter by Lawrence et al in this section)
Inorganic Nutrients
Macronutrients (N P)Virtually all heterotrophic organisms in the ocean contributeto the pool of available macronutrients via the excretion ofmetabolic wastes and protozoa contribute significantly tothis process Macronutrients are often in excess within proto-zoan prey relative to the consumersrsquo needs because much ofthe prey carbon is respired to produce energy for metabolismand growth ldquoExcessrdquo nutrients are eliminated as eitherorganic compounds or more commonly as ammonium (forN) and phosphate (for P) The importance of the match(or mismatch) between predator and prey stoichiometry hasbeen demonstrated experimentally and repeatedly in marineand freshwater ecosystems and has been elevated to a funda-mental ecological principle governing elemental and energyflow in aquatic food webs (327) Based on this reasoningCaron et al (328) concluded that bacterivorous protozoaplay an important role in nutrient remineralization andrelease in the ocean Bacterial biomass is typically rich in Nand P relative to other organisms so bacterial predators expe-rience large excesses of these elements relative to their growthneedsMoreover bacterivorous protozoa should be dispropor-tionately important in nutrient remineralization amongsingle-celled eukaryotes because as the smallest protozoa inaquatic ecosystems they have high weight-specific metabolicrates (329) Thus both stoichiometric and allometric rela-tionships implicate small bacterivorous protozoa as impor-tant sources of remineralized nutrients in the plankton
The reasoning also holds true for bacterial utilization oforganic compounds and it has led to some interestingfindings and conclusions regarding the role of bacteria innutrient cycling in the ocean Bacteria are important sourcesremineralized nutrients when they are consuming N- orP-rich substrate Contrary to this traditional view howeverheterotrophic bacteria are often strong competitors for inor-ganic nutrients under many situations largely because bacte-rial biomass is richer in N and P than much of the organicmatter they consume in nature so they require additional Nand P to produce their biomass (summarized by [330]) Exper-imental studies have demonstrated the uptake of ammoniumby bacteria other than cyanobacteria using stable isotope 15N(331) and also the short-lived radioisotope 13N (332) oftenfinding that bacteria may be responsible for one third of the Nor P uptake Additionally there is evidence that bacteria insome oceans like the Sargasso Sea are limited by phosphorus(333 334) Growth limitation of bacteria by phosphorusappears to be a common phenomenon of freshwater ecosys-tems (335 336)
N2 fixation is restricted to prokaryotes and in the oceanwater column it was thought for many years to be done
primarily byTrichodesmium a warm-water colonial cyanobac-terium that blooms sporadically andRichelia a cyanobacterialsymbiont that lives within certain diatoms (337) Some uni-cellular cyanobacteria also contribute significantly to globalN2 fixation (338) and molecular biological data suggestthat a variety of other bacteriamdashnot phytoplanktonmdashmayalso be fixing nitrogen in seawater (339) An exciting recentdiscovery in this field is that a bacterial nitrogen fixer origi-nally known only from nifH sequences UCYN-A has hadits genome sequenced by parallel pyrosequencing and wasfound be a cyanobacterium (Candidatus Atelocyanobacteriumthalassa) lacking the apparatus to generate oxygen as well asother pathways (340) Further study showed it to be symbioticwith a marine alga (341)
Micronutrients (Trace Metals Growth Factors)Bacterial growth may often be limited by macronutrients asdescribed and sometimes by trace nutrients particularlyiron that occur at very low concentrations in seawater(342 343) The importance of Fe as an element limiting pri-mary production in some oceanic regions has come underclose scrutiny and extensive experimental investigation inrecent years Many bacteria have special uptake mechanismsto utilize extremely low concentrations of iron includingsiderophores (released compounds that bind iron and arethen specifically taken up) Even if bacteria are capable ofgrowth at low Fe concentrations growth may be less efficientKirchman et al (344) found that Fe limitation stronglyreduced bacterial growth efficiency in a cultured marine bac-terium (ca 50 efficiency Fe replete but lt10 when Fedeplete) Thus the effects may be complex Marine N2 fixa-tion also may be limited by the availability of Fe becausenitrogen fixers have high Fe requirements
Protozoa tend to be a source rather than a sink for Fe in theocean analogous to their role in macronutrient remineraliza-tion Protozoan grazing activity has been shown to releaseiron from bacterial biomass thereby relieving Fe limitationin co-occurring phytoplankton (345) a mechanistic demon-stration of the process using protozoa growing on Fe-repletebacteria However it has also been reported that Fe-limitedbacteria may contain insufficient Fe for the bacterivorous pro-tozoa that consume them thus leading to Fe limitation in theprotozoan as well (346)
OxygenHypoxic and anoxic regions of the ocean are expected to sig-nificantly expand in the coming decades as a consequence ofchanges in ocean stratification resulting from global climatechange (347) As oxygen becomes depleted bacteria havethe capability to utilize a series of alternate electron acceptorsCompositional changes in the bacterial assemblage accom-pany these changes in metabolic activity but are beyondthe scope of this chapter Many of the studies on benthicmicrobiology have focused on redox reactions that occurthere Oxygen is usually consumed by sediment microbialactivity much faster than the rate at which it can be replacedby diffusion so most sediments especially organically richones are anaerobic below a few mm (fine-grained muds) orcm (coarser sediments) from the sedimentwater interfaceexcept where animals ventilate the benthos through tubesWhen oxygen is absent bacteria and archaea use alternateelectron acceptors such as nitrate nitrite oxidized Fe orMn organic matter or sulfate The use of nitrate or nitriteas an electron acceptor with concomitant production of N2
gas (denitrification) results in net loss of biologically availablenitrogen Bacteria that perform this reaction are typically
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-19
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquofacultativerdquo organisms that can switch from oxygen to nitrateor nitrite Organic matter may be reduced by fermentationreactions in which part of an organic substrate is oxidizedwhile another is reduced fermenter organisms are also typi-cally facultative Sulfate is a particularly important alternateelectron acceptor in marine systems because it occurs as thesecond most abundant anion in seawater (behind chloride)typically 28 mM Its use as an electron acceptor leads to theproduction of elemental sulfur or sulfide The use of sulfateas an electron acceptor yields much less energy than oxygenor nitrate and sulfate-reducing bacteria are strict anaerobesunable to use oxygen (often strongly inhibited by oxygen)Methane can be produced by methanogenic archaea whichare all strict anaerobes using CO2 as an electron acceptor(see Findlay and Battin this section for further discussion)
Reduced compounds such as sulfide are produced by theuse of these alternate electron acceptors and they diffuse awayfrom their source When they diffuse to environments with astrong oxidant like oxygen they may be oxidized by chemo-trophic bacteria in energy-yielding reactions Similarlyammonium can be oxidized to nitrite and then nitrate by che-motrophic nitrifying bacteria and the nitrate can diffuse intoanaerobic layers and become denitrified to N2 Such coupledoxidation-reduction reactions can lead to accelerated biogeo-chemical cycling particularly in sediments
It was originally believed that relatively few free-livingspecies of protozoa inhabit anaerobic and hypoxic marineenvironments This view has changed based on (largely)molecular analyses of some of these environments (308348ndash351) A significant number of previously undescribedeukaryotic phylotypes have been observed in anoxic environ-ments High protozoan biodiversity in the absence of molec-ular oxygen is not completely unexpected because otheranaerobic environments such as some gut environments(eg cockroach guts and cattle rumen) have notoriouslyabundant and diverse protozoan fauna but marine anaerobicenvironments have been difficult to access and sample prop-erly Many anaerobic protozoa exist together with symbioticbacteria (352 353) or possess unique organelles such as thehydrogenosome (354)
MAJOR BIOTIC CONTROLSThe Trophic Activities of Protozoa Predation andTop-Down ControlsPredator-prey relationships are fundamental interactionswithin biological communities linking production of bio-mass on the one hand with removal on the other hand Estab-lishing the trophic relationships among microorganisms andquantifying the rates at which they take place constitutemajor endeavors within marine microbial ecology As notedpreviously protozoa consume a wide variety of prokaryoticand protistan prey Numerous laboratory and field measure-ments have been made throughout the past quarter centuryto attempt to quantify the rates of bacterial and phytoplank-ton consumption The methodology for accomplishing thesestudies has varied and a number of approaches are presentlyemployed
Methodologies for estimating microbial grazing are diversebut generally fall into one of two experimental approachesperturbation experiments or tracer experiments Perturbationexperiments (size fractionation metabolic inhibitors anddilution technique) rely on manipulation of the consumeror prey assemblages in somemanner such that the effect is evi-dent from changes in the abundance of the prey population
during incubations relative to control treatments that receiveno manipulation Tracer experiments (radioactively labeledprey fluorescently labeled prey stable isotopendashlabeled pray)attempt to use labeled prey or a prey proxy to quantitativelyfollow the movement of prey biomass into consumers Forsummaries of these methods see (74 355)
Perturbation experiments have entailed a variety of mech-anisms to ldquodecouplerdquo predators from prey in natural microbialcommunities For example size fractionation uses various sizefilters to remove an entire size class of grazers from awater sam-ple leaving the prey to grow in the relative absence of thosepredators during subsequent incubations Unfiltered samples(controls) indicate the growth of prey in the presence of pre-dation Predation impact is obtained by the difference ingrowth of the prey between these treatments This methodhas been used to examine predation on cyanobacteria heter-otrophic bacteria and minute phototrophic eukaryotesAlternatively metabolic inhibitors have been employed tohalt predation (or in some cases prokaryote growth) insteadof physical removal by filtration
The dilution technique (356) has become a commonmethod for examining grazing mortality of phytoplanktonby microzooplankton (which is comprised predominantly ofprotozoa) This method relies on the dilution of herbivorouszooplankton in a series of subsamples (and concomitantmeasurements of phytoplankton growth in each subsample)to estimate the grazing mortality of the herbivores
Tracer experiments have used both radioactively labeledprey and fluorescently labeled prey to examine predation onbacteria and phytoplankton Radioactive labeling has beenaccomplished using 14C- or 3H-labeled organic compounds(for bacteria) or 14C-bicarbonate (for phytoplankton) Theuse of radioisotope approaches has diminished with improve-ments in the number and types of fluorescent compoundsavailable for this work Fluorescently labeled prey havebeen labeled with a variety of compounds that allow theprey to be observed readily (even inside predators) by epi-fluorescencemicroscopy (357 358) Alternatively a decreasein the total number of fluorescent prey in a sample indicatesthe rate of removal of the prey assemblage
Both perturbation experiments and tracer experimentsdescribed have inherent advantages and disadvantages Forexample perturbation experiments are fairly straightforwardto conduct but they may introduce artifact by removingpotentially important sources of nutrients or organic com-pounds released by predators (359) On the other hand thedilution technique is very labor-intensive but one advantageof this method is that it provides the opportunity to determineboth phytoplankton growth and grazing mortality in a singleprocedure Most tracer experiments are fairly easy to conductand generally do not perturb the food web being studied butfeeding selectivity for or against the tracer (relative to naturalprey) may produce spurious results In addition some of thesemethods provide a measurement of the activity of the entiregrazer assemblage and others are applicable for examiningthe activity of specific taxa For example size fractionationapproaches examine the effect of the removal of an entiresize class of organisms (a community-levelrdquo measurement)whereas fluorescently labeled prey can be used to examinethe ingestion rate of a particular species by observing theuptake of labeled prey into that species (a species-levelmeasurement)
Predation rates of numerous marine protozoa have beenexamined in the laboratory From these experiments a gen-eral knowledge of the rates of consumption by these specieshas been accumulated Feeding in most protozoa shows a
422-20 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
functional response to the availability of prey that is similar tothe response of many organisms (although there are a greatdeal of species-specific differences) That is ingestion rateincreases rapidly as prey abundance increases until a maximalingestion rate is attained (360)Manyof these curves have theform of a Type II functional response curve (361)
Based on the features of these curves it is clear that proto-zoa have the ability to ingest substantial numbers of prokar-yote and eukaryote prey in natural ecosystems Somebacterivorous flagellates can consume bacteria (and cyano-bacteria) at rates up to 100 s per individual per hour whereassome bacterivorous ciliates can consume thousands of bacte-ria per individual per hour Rates of consumption of eukary-otic phytoplankton by herbivorous ciliates and largeflagellates are considerably more variable and highly depend-ent of the feeding mechanism (eg filtration in ciliates versuspseudopodial net in dinoflagellates) and the size of the prey
These lab studies demonstrate that rarely are prey suffi-ciently abundant in nature to support the maximal ingestionrates of which many protozoa are capable Nevertheless pre-dation rates of protozoa in nature are sufficient to constitute amajor factor controlling the standing stocks of bacteria andphytoplankton (63 180 184 355) For example these sum-maries indicate that on average protozoa consume 20ndash80ofthe standing stock of phytoplankton (measured as chloro-phyll) in the world ocean and 60ndash75 of daily primary pro-duction a departure from the classical view of copepods as theprimary consumers of phytoplankton This large role of her-bivorous protozoa infers a large role for copepods as consumersof planktonic protozoa (185) Similarly the role of protozoaas major consumers of bacterial biomass is indicated by therelative balance between bacterial production on the onehand and bacterivory on the other (Fig 8) The approximateone-to-one balance between these measurements over a largenumber of studies implies that protozoan bacterivory must bea major factor determining the fate of bacteria in marine (andfreshwater) plankton ecosystems
It is interesting to note that some types of bacteria includ-ing marine Bdellovibrio are known to prey on other marine
bacteria (362) At the time of this writing the importanceof bacterial predation by other bacteria within the contextof oceanographic processes is not known and presumed tobe small While Bdellovibrio may be cultivated from seawaterit can also be found among sequences from cold sediments(363) and the water column Bdellovibrio happens to be thebest studied predatory bacterium but there are no doubtothers that we simply have not yet identified (364)
Species-Specific Interactions (MutualismParasitism Commensalisms)Interactions between marine microorganisms such as compe-tition allelopathy and symbiosis are fundamental aspectscontrolling community composition and activity but atpresent most are poorly characterized in natural marine sys-tems Bacteria must compete for dissolved and particulatecompounds with other bacteria andwith protistan taxa whilephagotrophic protists must compete with each other andmetazoa for available prey These interactions manifest them-selves in many forms including true competition for availableresources resource partitioning such that some taxa specializeon the utilization of certain substrates or prey while othersspecialize on different ones (reducing competition) or eventhe cooperative use of compounds (eg one bacteriumrsquoswaste products being utilized as substrate by another bacte-rium) These metabolic consortia are probably common inthe ocean but can be difficult to identify and characterizeConsortia of bacteria working together to perform complexreactions especially in anaerobic systems are well known inmicrobiology
Consortia in which there is a very high degree of integra-tion between species (ie symbiosis) are very common in theocean These associations may be beneficial to both partners(mutualism) beneficial to one part and inconsequential forthe other (commensalism) or beneficial to one part and det-rimental to the other (parasitism) Many highly visible andwell-characterized marine mutualisms exist between chemo-synthetic bacteria and animals such as hydrothermal ventor methane seep tubeworms clams mussels or other inverte-brates (365) and also between luminescent bacteria and fishor squids (366) Similarly many protozoa form mutualisticassociations with photosynthetic protists (367) The mostconspicuous of these associations involved large amoeboidprotozoa of oceanic ecosystems (planktonic foraminiferanspolycystine radiolaria and acantharia) which contain largenumbers of intracellular algae (74) but associations betweensome heterotrophic dinoflagellates and cyanobacteria are alsocommon (368) Studies have indicated that efficient nutrientcycling in these protozoan-algal associations confers an eco-logical advantage for both partners in surfacewaters of the oli-gotrophic ocean
Parasitism is also a common symbiotic relationship amongmarine protozoa Protozoan parasites have been reported for adiverse array of protists and animals in the ocean Heterotro-phic dinoflagellates and presently uncultured alveolate cladesrelated to alveolates appear to be particularly successful atevolving thismode of existence (369ndash371) In a common par-asitic group Amoebophrya spp the parasite invades photo-synthetic dinoflagellates where it undergoes reproductioneventually lysing the host and releasing dozens of infectiveswarmers The parasitic protist Pirsonia spp is a parasite ofdiatoms (372) The overall importance of these parasitismsin the population dynamics of their hosts is not clearly knownbut may constitute a significant source of mortality for somespecies (373 374) The frequency and diversity of ldquonewrdquo
FIGURE 8 Meta-analysis of experimental studies that have com-pared rates of bacterial productivity (converted to population growthrate units of dminus1) with rates of bacterivory by phagotrophic protists(expressed as equivalent units of bacterial growth rate dminus1) Thesolid line indicates where measurements of bacterial removal by graz-ers were equal tomeasurements of bacterial production Data redrawnfrom Sanders et al (63) doi1011289781555818821ch422f8
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-21
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-
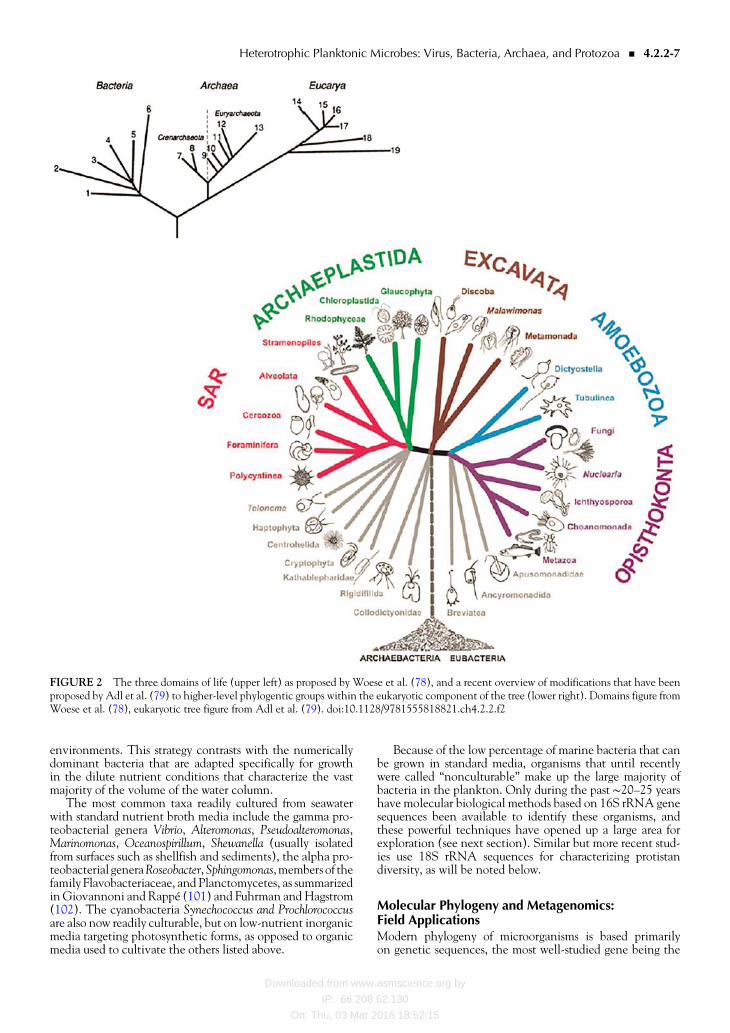
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
environments This strategy contrasts with the numericallydominant bacteria that are adapted specifically for growthin the dilute nutrient conditions that characterize the vastmajority of the volume of the water column
The most common taxa readily cultured from seawaterwith standard nutrient broth media include the gamma pro-teobacterial genera Vibrio Alteromonas PseudoalteromonasMarinomonas Oceanospirillum Shewanella (usually isolatedfrom surfaces such as shellfish and sediments) the alpha pro-teobacterial generaRoseobacterSphingomonasmembers of thefamily Flavobacteriaceae and Planctomycetes as summarizedin Giovannoni and Rappeacute (101) and Fuhrman and Hagstrom(102) The cyanobacteria Synechococcus and Prochlorococcusare also now readily culturable but on low-nutrient inorganicmedia targeting photosynthetic forms as opposed to organicmedia used to cultivate the others listed above
Because of the low percentage of marine bacteria that canbe grown in standard media organisms that until recentlywere called ldquononculturablerdquo make up the large majority ofbacteria in the plankton Only during the pastsim20ndash25 yearshave molecular biological methods based on 16S rRNA genesequences been available to identify these organisms andthese powerful techniques have opened up a large area forexploration (see next section) Similar but more recent stud-ies use 18S rRNA sequences for characterizing protistandiversity as will be noted below
Molecular Phylogeny and MetagenomicsField ApplicationsModern phylogeny of microorganisms is based primarilyon genetic sequences the most well-studied gene being the
FIGURE 2 The three domains of life (upper left) as proposed by Woese et al (78) and a recent overview of modifications that have beenproposed by Adl et al (79) to higher-level phylogentic groups within the eukaryotic component of the tree (lower right) Domains figure fromWoese et al (78) eukaryotic tree figure from Adl et al (79) doi1011289781555818821ch422f2
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-7
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
small subunit ribosomal RNA gene (16S rRNA in Bacteriaand Archaea and its larger homolog 18S rRNA in eukar-yotes) This molecule is strongly conserved over evolutionarytime so this single molecule has been used for constructingphylogenetic trees of all living organisms (httptolweborgtreephylogenyhtml) Analysis of 16S18S rRNA genesequences has been used to evaluate deep evolutionary rela-tionships among organisms and was instrumental in point-ing out that Archaea Bacteria and Eukarya should beconsidered different Domains of equivalent phylogeneticrank above kingdoms (78) However there are sufficientdifferences in 16S18S rRNA gene sequences to demonstrate
relationships (ormake distinctions) at the genus or sometimesspecies level
The first phylogenetic studies based on 16S18S rRNAgenes used sequences derived from cultures However onedoes not need cultures to obtain rRNA gene sequences (orany other sequences for that matter) An idea developed inthe lab of Norman Pace in themid-1980s involved extractionof DNA directly from natural samples and then cloning andsequencing of the DNA as a means of assaying the microbespresent in the samples (103 104) The original protocolscalled for cloning by creating what are called ldquophage librariesrdquofrom the natural DNA but since 1986 PCR has been applied
FIGURE 3 (a b) Plankton biomass in the Arabian Sea during the 1995 southwest monsoon (a) and intermonsoon period (b) Areas of theboxes indicate the relative magnitudes of the biomass in each category Categories within the dashed boxes in a b c are composed of protozoaArrows indicate the direction of energymaterial flow in the food web thicker arrows depicting greater flow Redrawn from (86)(c) Depth-integrated biomass (mgm2) in the upper 100 m of the Sargasso Sea near Bermuda and in the upper 200 m of the equatorial Pacificat 175degE Thewidth of the bars indicates the biomass in each size category Heterotrophs have been separated by size class while phytoplanktonhave not Size classes delineated by the dotted box are comprised of protozoa Redrawn from (65) doi1011289781555818821ch422f3
422-8 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
extensively for cloning and related studies The targetsequence can be almost instantly ldquoidentifiedrdquo to its closestphylogenetic neighbor by what have come to be standardonline sequence comparisons
Beyond the study of targeted genes (like the 16S rRNAgene) shotgun metagenomic studies have examined theentire genetic repertoire of the microbes in a given sampleThe metagenome is the collective genome of all organismsin the sample Initially these studies extracted DNA fromall organisms in a sample (usually prefiltered through a filterapproximately 1 microm to remove most organisms larger thanbacteria) sheared it to produce fragments and clonedthem into standard vectors either as small (thousands ofbases) or large (to hundreds of thousands of bases) insertsthat is fragments of DNA from the environment nowcloned into the vectors in a form suitable for sequencingThe best known early marine study of the former type isthe Global Ocean Survey with initial results published byVenter et al (105) which generated more than a billionbases of DNA sequence and reported 12 million previouslyunsequenced genes estimated to come from at least 1800different genomic species cumulatively in the many samplesthey analyzed
With the advent of next-generation sequencing that gen-erates millions ormore sequences in a run (known by a varietyof acronyms including 454 Illumina SOLiD etc) cloningof genes has largely been replaced by clone-free sequencingThe extent and power of such sequencing has recentlybeen demonstrated by the ability to construct essentiallythe entire genome of an uncultivated marine Group II Eur-yarchaeon that constituted only approximately 2 of amicrobial community sample using SOLiD sequencingmade possible by high coverage and the use of mate pairsequencing ofsim3000 bp fragments (106) (this length isneeded to span repeats and highly conserved genes that oth-erwise make genome construction difficult)
For 16S rRNA gene studies ldquotag sequencingrdquo pioneeredby Sogin et al (107) consists of amplifying a suitable sizedpart of the gene with broadly conserved primers (choice isimportantmdashfew are truly universal for the groups intended)often ldquobarcodedrdquo to allow multiple samples to be combinedinto a single run The amplified products are then sequencedand sequences processed en masse Many thousands of partialSSU rRNA gene sequences per sample are economically ana-lyzed this way though they are usually short (currently a fewhundred bases each depending on the sequencing platform)This way of analyzing the composition of microbial commun-ities is now standard These approaches provide so muchinformation even about very rare sequences that the resultshave led to the important concept of the ldquorare biosphererdquoorganisms that may be active or dormant and constituting avery small proportion of the community (eg often muchless than 01) but are potentially important for dispersionadaptation to changing conditions or even critical activitieslike nitrogen fixation or vitamin production (108) Howeverdue to the potential for artifacts like error sequences thisapproach requires rigorous application of quality filteringand clustering algorithms to avoid erroneous taxa and overes-timation of species richness (109 110)
Next-generation sequencing approaches have also allowedwork to begin metagenomics studies of microbial eukaryotes(111) These studies are still constrained by the much largergenomes of eukaryotes and therefore the difficulties ofobtaining sufficient sequences to reconstruct the genome ofa particular species Along with advances in sequencing abil-ity studies of eukaryotic metagenomics have been facilitated
by advances in single-cell isolation which when coupledwith high-throughput sequencing approaches reduce the tre-mendous complexity present in natural complex eukaryoticcommunities to a manageable task (112) Such single-celltechniques are also quite valuable in studies of bacteria andarchaea though the amplification technique tends to bevery uneven and typically generates less than half the genomeof each isolated cell regardless of domain (113 114)
Additionally similar to DNA mRNA is amenable toextraction and sequencing although greater care must betaken during extraction and purification as RNA shows agreater susceptibility to degradation during processing Copy-ing of mRNA by reverse transcription of RNA into cDNAfollowed by DNA sequencing has allowed insights into themetatranscriptomes of environmental samples Metatran-scriptomic studies provide information on gene expressionin an ecosystem and thereby indicates ldquoactivityrdquo of themicro-bial community rather than simply ldquopotentialrdquo represented bythe genomic DNA present in the sample with many applica-tions from showing which processes are being carried outby which organisms to fine-scaled diel studies (115ndash118)Nevertheless given variations in the lifetimes of differenttranscripts and protein molecules the transcriptome maynot be fully representative of the current activity of anorganism
Limitations of these analyses include sequencing errorsPCR mismatches or biases clustering and bioinformaticschallenges and chimeras generated during PCR Also thephylogenetic resolution of short sequences is limited giventhe high conservation of rRNA sequences Even with clone-free shotgun metagenomics there may be biases such as non-random losses of DNA during extraction and preparationor biases (eg from G +C content or secondary structure)in the sequencing procedures Determination of speciesdiversity by these approaches provides an example of theselimitations Shakya et al (119) working with synthetic com-munities (purified genomicDNA from16Archaea represent-ing 3 phyla and 48 Bacteria representing 16 phyla remixed tosimulate an environmental DNA extract) applied both meta-genomic analysis (454 and Illumina platforms) and PCRamplification followed by 454 sequencing of 16S rRNA genesto determine both species richness and relative abundanceThey found that PCR amplification454 sequencing of 16SrRNA genes yielded an accurate measure of species richness(providing that appropriate data processing was applied)but that the relative abundance of up to 94 of the species(depending on domain and variable region amplified) wasover- or underestimated by at least 15-fold (values rangedfrom not detected to 103-fold overestimation) In contrastboth metagenomic approaches yielded relative abundancesthat were within the authorsrsquo 15-fold accuracy cutoff forsim50 of the species However they concluded that addressingrichness overestimation in metagenomic analyses that isdistinguishing rare but real OTUs from experimental andcomputational artifacts awaits further computational andclassification improvements More recently Parada et al(120) used mock communities composed of 16S rRNAclones from 27 common marine taxa (from nine Bacterialand two Archaeal phyla) to show that small differences inPCR primers (and different clustering methods) can yieldlarge differences in apparent relative abundances of reportedtaxa However one primer pair and informatics pipelinethey tested using a particular version of 515F-926R (V4ndashV5) provided accurate estimates of relative clone abundance(r2 = 095) when comparing observed versus expected cloneabundance
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-9
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
Culture-Independent Diversity Studies
The first groups to be identified using cultivation-independ-ent molecular techniques from the marine plankton (121)were the bacterial SAR11 cluster (a group of closely relatedgene sequences or phylotypes) and marine picoplanktonicunicellular cyanobacteria Synechococcus and Prochlorococcus)Of these the SAR11 cluster was completely unknown butthe cyanobacteria had previously been recognized by theirunique pigment fluorescence Waterbury et al (122) andJohnson et al (123) used epifluorescence to observe Synecho-coccus and Chisholm et al (124) discovered Prochlorococcususing flow cytometry These cyanobacteria were later isolatedand grown in phytoplankton culturemedia These two groupsare generally common in the euphotic zone with SAR11 typ-ically comprising one third of the planktonic bacteria (125)and the cyanobacteria common everywhere but polar waters
Probably the biggest surprise to come from the applicationof molecular tools was the discovery by Fuhrman et al (126)of abundant archaea in the deep sea The archaea were foundto be in a unique phylogenetic cluster that was only distantlyrelated to any previously known archaea but the ldquoclosest rel-ativesrdquo (not really close at all) were extreme thermophiles Asubsequent study also found archaea to be present in near-surface coastal waters albeit relatively rare (lt2 relativeabundance) This study used a PCR technique specificallytargeted archaea and found ldquomarine Crenarchaeardquo plus a sec-ond group belonging to the phylum Euryarchaea (127)
Up until this time all known Archaea were thought to beldquoextremophilesrdquomdashadapted for either very high temperatures(thermophiles) extremely salty conditions (halophiles) orstrictly anaerobic environments (methanogens) Yet theseorganisms were present in cold or cool water at ordinary salin-ity and high oxygen concentrations Fluorescent in situhybridization (FISH) measurements from deep-sea sampleshave since showed that the archaea may make up appro-ximately 40 of the total countable prokaryotes with thepercentage reaching to 60 at 200 m depth in theMediterra-nean (35 36) An extensive time series of FISH measure-ments near Hawaii confirmed that the archaea are indeedvery abundant throughout the year from below the photiczone to at least 4000 m and typically constituting 30ndash40of the total prokaryotes present in waters deeper than a fewhundred meters (128) with similar results found elsewhere(129 130) (Fig 4) They have been reported from many pla-ces including the Atlantic Pacific and Southern Oceansand are dynamic components of the plankton the most com-mon type by far being the ldquomarine Crenarcheaeardquo (128 129131) While the marine archaea have been reported to bedominated by a few major ldquophylotypesrdquo (132) they alsohave been shown to have a great deal of microdiversity withinthese phylotypes suggesting there are many kinds of close rel-atives coexisting (133) Interestingly this group of archaeamay likely be the most abundant kind of organism on Earthgiven the huge volume of the deep sea and their high abun-dance there (134) It has recently been proposed that ldquomarineCrenarchaeardquo be elevated to their own major phylum outsidethe Crenarchaeota called the Thaumarchaeota on the basisof deep phylogenetic branching and fundamental differencesbetween them and the Crenarchaeota (135) The Thau-marchaeota possess the uniquely archaeal membrane lipidcrenarchaeol and are nowalso known to be abundant in soils
The physiology of the Thaumarchaeota has been anintriguing area of study Initially an autoradiography-FISHcombination approach demonstrated that they take upamino acids (130 136) But subsequent data showed that
crenarchaeol was derived largely from CO2 suggesting auto-trophic metabolism (137 138) An experiment showinguptake of 13C-labeled bicarbonate into these archaeal lipids(139) directly pointed to autotrophy in this group Chemoli-thoautotrophywas first hinted at byVenter et al (105) whosemetagenomic analysis showed an apparent archaeal scaffoldthat contained genes suggestive of ammonia oxidation Fur-ther evidence came from Schleper et al (140) who detectedseveral ammonia oxidation genes in order directly adjacent toa Thaumarchaeota 16S rRNA gene in a soil-derived metage-nomic clone The issue was directly resolved when Konnekeet al (141) isolated a related marine archaeon from sedimentof a marine aquarium and this organism Candidatus Nitroso-pumilus maritimus was found to have a chemoautotrophicmetabolism quantitatively oxidize ammonia to nitrite andcontain an archaeal ammonia monooxygenase gene (amoA)and interestingly did not grow heterotrophically Cultivationexperiments showed this organism has a high affinity forammonium allowing it to outcompete bacterial nitrifiers atlow (submicromolar) concentrations as are typical in thesea (142) The complete genome of the organism has shownnovel adaptations for nitrification and autotrophy (143)
Even if the Thaumarchaea are primarily chemolitho-autotrophs field data suggest a level of mixotrophy in thatsome organic substrates are being incorporated into biomassA stable isotope study using cells collected at 670 m depthoff Hawaii estimated that about 80 of the carbon incor-porated into archaea-specific lipids came from inorganic sour-ces and about 20 from organic compounds (144 145)Interestingly it has also been claimed that the genomesof deep-sea (gt2000 m depth) members of the Thaumarch-aea as well as those living in equatorial waters rarely containthe amoA gene and thus may be primarily chemoorgano-trophs (48)
Major bacterial groups that have been documentedfrom seawater using 16S rRNA characterization includesome that are also known from culture (eg AlteromonasRoseobacter) and several that are phylogenetically distantfrom standard cultures The most common groups are inrough order of their relative abundance in clone librariesfrom most to least abundant SAR11 (relatives of Pelagibac-ter ubique) Roseobacter SAR86 cyanobacteria SAR116SAR202 SAR234 andMarine Group A The SAR designa-tion followed by a number is an arbitrary sequential cloneidentifier from Sargasso Sea cloning studies done by the Gio-vannoni lab that did most of the early systematic cataloging ofclones Summaries of the data and phylogenetic relationshipsof these groups can be found in Fuhrman andHagstrom (102)and Giovannoni et al (101)
Molecular Genetic Discoveries in Bacterial andArchaeal Marine BiologyAs described earlier metagenomics is an extension of theideas used in the 16S rRNA cloning studies in that all genesfrom the native microorganisms are separated and clonedwithout having cultivated the organisms These methodsearly on started to find unexpected and very interestingresults The best examples involve unexpected marine photo-trophy the first of which is discovery of a nonchlorophyllphotosynthetic bacterial pigment called proteorhodopsin(146) The gene was found on a large environmentallyderived fragment of DNA that also had a gene coding for16S rRNA from the so-called SAR86 group (one ofthe groups common in seawater) This pigment can act as alight-driven proton pump thought to permit cells to generate
422-10 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
FIGURE 4 Distribution of Bacteria Euryarchaea andmarine Crenarchaea (recently renamed Thaumarchaea) along a transect in theNorthAtlantic Ocean as measured by CARD FISH and expressed as of total bacteria + archaea counts via DAPI stained epifluorescence fromTeira et al (130) Top panel shows station locations that are shown on the top of the lower three panels with the distance in km reported alongthe transect from lower to higher station numbers doi1011289781555818821ch422f4
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-11
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ATP from sunlight Interestingly different versions of this pig-ment are found at different depths apparently ldquotuningrdquo theabsorption to match the ambient wavelengths of light asthey change with depth (147) The proteorhodopsin genesare very widespread and diverse perhaps in half or more ofall marine bacteria including SAR11 and Euryarchaeota(105 148ndash150) However almost all of several culturedorganisms with proteorhodopsin studied to date do not showa growth benefit from light (150) with the exception of amember of the Flavobacteria Dokdonia sp strain MED134that grows faster in the light only under moderately lownutrient conditions (151) and a marine Vibrio has beenshown to survive starvation longer in the light than in dark-ness (152) These observation may explain the wide phyloge-netic distribution and high abundance of this gene in thatproteorhodopsin may often assist long-term survival of bacte-ria under extreme energy-limiting conditions yet not havemucheffect on growthduringmore energy-replete conditionsInterestingly a recent report suggests the growth benefit fromlight in Dokdonia MED134 is from enhanced uptake of itsrequired growth factor vitamin B1 which is transported by atonB-dependent transporter powered by a proton gradient(153) This pigment might also have sensory or other rolesnot yet well examined (150)
The second kind of phototrophy found by moleculargenetic and fluorescence techniques to be unexpectedlyimportant in marine plankton is anoxygenic aerobic bacterialphototrophy based on the pigment bacteriochlorophyll aCulturable aerobic anoxygenic photosynthetic (AAP) bacte-ria have been known from seawater for several years (154) andare widespread (155) Initial recent reports of direct countsclaimed they represent about 11 of the total bacterial com-munity in the euphotic zone (156) but those early counts didnot correct for the presence of other bacteria and such correc-tionoften yields estimates averaging closer to 2(157)Directmeasurements show the bacteriochlorophyll pigment is rela-tively rare (158) However studies in various ocean locationsincluding ocean gyres report that these AAP bacteriacan indeed on occasionmake up a quarter of the total prokar-yotes (159 160) A recent energetic modeling of AAP andproteorhodopsin-containing bacteria (161) has suggestedthat proteorhodopsin-containingones cangain approximately02asmuchenergy from sunlight asSynechococcus (commonmarine cyanobacterium) and AAP bacteria can gain approxi-mately 13 as much energy from sunlight as SynechococcusThey concluded the AAP bacteria may gain energy sufficientto meet maintenance costs but proteorhodopsin-containingbacteria were not expected to do so except at high light inten-sities andwith largenumbersof proteorhodopsinmoleculespercell They concluded the ease and low cost of maintainingPR-based phototrophy (a few genes required) may explainthe high incidence of proteorhodopsin genes
Other metagenomic studies have yielded interestinginsights about organisms and processes that would otherwisebe difficult or impossible to establish For example thesequence of an archaeal gene fragment isolated from seawaterrevealed extensive evidence of genetic exchange with othertypes of archaea and even bacteria (162) Such geneticexchange was also quite evident from the complete genesequences of different strains of Prochlorococcus and Synecho-coccus which was attributed in part to virus-mediated geneflow (163ndash166)
Dilution Cultures of Bacteria and ArchaeaSome recent cultivation techniques that permit growth ofldquotypicalrdquo oligotrophic marine bacteria in pure culture are
based on the simple concept that bacteria living in seawateron dissolved organic matter might best be grown in the labo-ratory in ordinary filtered seawater This conclusion followsfrom similar techniques to grow mixed ldquoseawater culturesrdquoof marine bacteria (167) but the seawater inoculum in thiscase is diluted so that only one or a few bacteria are addedto the initial culture vessel (168) A rapid throughput versionof this method has been used to cultivate members of bacte-rial groups thought to be conventionally uncultivable such asthe SAR11 clade (169 170) albeit often at very low den-sities Such cultures permit focused studies of the propertiesof those organisms including genomic analysis (169 170)This valuable work provides insight into the roles and activ-ities of some of the most common bacterial phylotypesobserved in the ocean (171) showing for example that culti-vated SAR11 requires a source of reduced sulfur as well asproviding templates aiding the interpretation of metage-nomic analysis (172)
PROTOZOADiversity and DistributionHeterotrophic protists have been identified historically fromtheir morphological features that are apparent at the levelof the light or electronmicroscope Features of cell size shapetype and pattern of flagellationciliation skeletal structurescharacteristics of the nucleus and other cellular structuresand organelles have been used to differentiate themany thou-sands of described species A description of the extent of thisdiversity of form and function is well beyond the scope of thisbook (see [79] for more information) However much of thisdiversity can be reduced to three basic body plans that dictatethe broadest ecological roles of these cells amoeboid flagel-lated and ciliated forms (Fig 5) Among the lineages possess-ing one of these three forms only the last group the ciliatedprotists constitute a monophyletic group within the domainEukarya
One of the simplest body plans for protozoa is the amoe-boid cell exemplified by the gymnamoebae or ldquonakedrdquoamoebae Motile nonphotosynthetic cells lacking flagellaor cilia occur as life stages in a number of phylogeneticallydiverse taxa but for many free-living species of protozoathis form constitutes the only life stage Motility is largelyconfined to movement along surfaces by means of pseudopo-dia that can take on a variety of (species-specific) shapes Sig-nificant abundances of amoebae are largely confined tobenthic and epibiotic environments (174) and to suspendedparticulate material where they can occasionally be highlyenriched (27) Most amoebae consume bacteria and otherminute prokaryotes and eukaryotes
Several heterotrophic protistan groups possess complexamoeboid body plans most notably the foraminifera polycys-tine and phaeodarian radiolaria and the acantharia Plank-tonic forms are predominantly oceanic in their distributions(although a large number of benthic species of foraminiferaexist) These species are heterotrophic and many are visibleto the naked eye (individual cells can bege1 cm gelatinouscolonies can form ribbon-like structuresge1 m in length)They feed on a wide variety of bacterial protistan and meta-zoan prey using pseudopodial networks have rather longcomplex life cycles for individual cells (weeks to months)and are extensively used in studies of paleoclimatologicalreconstruction (82 175) In addition many of these speciespossess intracellular symbiotic usually eukaryotic algaeThe widespread occurrence of algal endosymbiosis among
422-12 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
these protozoa in oceanic pelagic environments implies thatstrong selective forces appear to give rise to these associations(97 98)
Flagellated forms of protozoa exist across many protistanlineages Eukaryotic flagella come in various sizes numbersand forms (eg with or without tiny hairs) that are character-istic of the different lineagesMany flagellate species are appa-rently purely phototrophic or heterotrophic (phagotrophic)but a significant (still poorly known) fraction of flagellates aremixotrophic combining both phototrophy and phagotrophy(70 88) Flagella in phagotrophic flagellates are employed formotility and prey capture Most free-living flagellates possessone to four flagella (typically one or two) that can be manytimes the length of the cell itself Although there is tremen-dous species diversity among flagellated protozoa many ofthese species have broadly overlapping ecologies As a groupflagellates are the most numerically abundant protozoa ofboth benthic and pelagic ecosystems and they are fundamen-tally important as consumers of bacteria cyanobacteriaand other eukaryotes Flagellated protozoa within the nano-plankton size class (2ndash20 microm) are often counted as a singleassemblage in plankton studies because morphological details
apparent using light microscope are insufficient to distinguishamong the many species This assemblage has been variouslyreferred to by a variety of names and acronyms including het-erotrophic nanoplankton (HNAN HN) heterotrophic flag-ellates (Hflags) heterotrophic nanoflagellates (HNF) andmicroflagellates Heterotrophic nanoplankton is the mostaccurate term for most methodologies employed to countthese cells because it does not require visualization of flagella(which are often lost from these small cells during preparationfor microscopy) The confusion over terminology is partly aconsequence of the different methodologies that have beenused to count these species and partly a consequence of thefact that these species were first studied prior to the wideacceptance of the size convention of Sieburth et al (84)
Heterotrophic flagellates in the microplanktonic size class(20ndash200 microm mostly dinoflagellates) are important consum-ers of phytoplankton in pelagic ecosystems Many of thesespecies are capable of the production of large pseudopodialnets the pallium that can engulf prey (particularly diatoms)significantly larger than the diameter of the dinoflagellatetheca (176) This behavior and its ecological significance forenergy flow in plankton communities have been recognized
FIGURE 5 Body plans and size ranges of protozoa These micrographs depict amoeboid (andashg) flagellated (hndashj) and ciliated (kndashm) forms ofprotozoa From Caron et al (173) Markers bars are 5 (i) 10 (h) 20 (c j l) 30 (a) 50 (k m) 100 (b f) 500 (d) and 1000 (e g) micromdoi1011289781555818821ch422f5
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-13
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
only within the past few decades (177) These species areoften abundant in waters where diatoms dominate (178179) In such situations heterotrophic dinoflagellate biomasscan be similar to that of ciliated protozoa (180)
Ciliates are generally the most recognizable form of pro-tozoa to nonspecialists Nonetheless ciliates are a diversegroup morphologically as well as ecologically (181 182)The degree of ciliature on these species can range from uni-formly ciliated to totally devoid of cilia during most of theirlife cycle to ciliature restricted to specific regions of thecell In addition cilia can fuse to form various complexstructures (eg ciliary membranes or veils cirri) that assistin locomotion food capture or attachmentCiliates are abun-dant and ecologically important species in both benthic (8)and pelagic (183) ecosystems and can consume a variety ofprokaryotic and eukaryotic prey Together with the heterotro-phic dinoflagellates these species are the dominant consum-ers of phytoplankton in many pelagic ecosystems (184) andas a consequence they form an important trophic link to met-azoan zooplankton (185 186)
Diversity and Biogeography of Protists FromMorphology to DNA SequencesUntil relatively recently the biodiversity of marine protistanassemblages was not generally considered a controversialtopic It was generally accepted that while all species of pro-tists certainly have not been identified representatives ofmost types of algae and protozoa had been observed anddescribed if not actually brought into culture in the labora-tory Direct sequencing of 18S rRNA genes from environ-mental samples (as described in ldquoMolecular Phylogeny andMetagenomics Field Applicationsrdquo) has changed that viewInitial forays into environmental DNA indicated a muchgreater diversity of protists than previously documented usingclassical approaches of culture and microscopy (81 187ndash191) Numerous publications over the past decades haveexpanded these findings (see [192] for a recent global analy-sis) which are highly analogous to discoveries in marine pro-karyote research as described already
Previously uncharacterized protistan diversity has nowbeen documented at virtually every level of eukaryoticorganismal classification Some of these findings couldhave been expected but some have been very unexpectedFor example a much greater diversity than noted previouslyhas been observed among small (lt10 microm) protists in plank-tonic ecosystems These species generally possess few distinc-tive morphological features and one could expect that manycryptic species might be present among these small morpho-types The molecular ldquodiscoveryrdquo of this eukaryotic diversityhas stimulated progress on the isolation and descriptionof new species and genera of minute algae and protozoa(193ndash197)
Analyses of environmental samples have also indicatedthe unanticipated existence of novel 18S rRNA gene sequen-ces that imply the presence of novel lineages of eukaryotes innatural protistan communities (198ndash200) These are sequen-ces for which there are apparently no known described orcultured species The degree to which these sequences dif-fer from sequences of known sequenced eukaryotes impliesthat some of these lineages may be distinct at the level ofphylum (201)
These findings have raised basic questions and somedebate concerning the true diversity and biogeography ofprotistan assemblages in natural ecosystems (202ndash204) thevalidity of the many novel phylotypes or cryptic species that
are being documented (205) the potential importance andsignificance of the many rare taxa that characterize thesecommunities (206) and the implications of these findingsfor the ecologicalbiogeochemical roles that protists playin aquatic ecosystems For example it was postulated andsubsequently confirmed that some novel alveolate lineagesreported from marine ecosystems make up a suite of parasiticprotozoa whose ecological importance may have been signifi-cantly underestimated in the past (207) Deciphering theidentity and significance of these many unknown phylotypeswill constitute a significant effort for protistologists in thefuture (80 85)
Life Histories and Ecological Strategies of ProtozoaAbundance prey type and life histories all vary tremendouslyamongmarine protozoa The smallest species (eg many flag-ellates) tend to be the most abundant and widely distributedin the world ocean Indeed many of these species may beglobally distributed (208) Most nanoplanktonic flagellateshave potentially rapid rates of grazing and growth Underoptimal conditions these species can divide by binary fissionseveral times a day and thus dramatic increases in their pop-ulations can take place in response to favorable conditionswithin a few days (209) Many of these species are capableof surviving for limited periods of time without food andhave developed a variety of physiological or life cycle strat-egies to cope with these events (209) However in contrastto some bacteria that may remain viable through long periodsof starvation (210) protozoa will expire or encyst in responseto low food abundance and thus ldquoboom-and-bustrdquo popula-tion changes are characteristic of small flagellates
On the other end of the size spectrum from the ratherubiquitous nanoflagellate species many of the largest proto-zoan species (polycystine and phaeodarian radiolaria plank-tonic foraminifera) are exclusively oceanic (ie do notsurvive in most coastal environments) andor have specificlatitudinal and depth distributions Thus the abundancesof these latter species may range from undetectable to max-imal abundances of gt105 individualsm3 Dramatic changesin the abundance of these protozoan taxa also can be relatedto changes in prey abundance physicalbehavioral aggrega-tion or to periodicity of life cycle events For example theplanktonic foraminifer Hastigerina pelagica reproduces on alunar cycle and thus abundances (and life stages) of this spe-cies in oceanic waters can vary considerably over the courseof a month (211) In general life cycles for the large amoe-boid protozoa are lengthy and complex (for single-celledorganisms) with life spans unknown for many species(attempts to culture them in the lab have so far been unsuc-cessful) but estimated to be on the order of months to per-haps years (175)
The abundance and activities of microplanktonic hetero-trophic protists (mostly ciliates and heterotrophic dinoflagel-lates) tend to be somewhat intermediate to those ofnanoplanktonic flagellates and the larger amoeboid formsThese species are present in the majority of marine ecosys-tems and collectively play an important role in the controlof phytoplankton biomass (and probably the abundance ofnanoplanktonic protozoa although there is little informa-tion on this topic) in waters throughout the world oceanLike small flagellates ciliates and dinoflagellates reproduceprimarily by binary fission but their maximal growth ratesare typically slower (one division a day is typical) and theirlife cycles often include sexual phases that allow for geneticrecombination
422-14 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
VIRUSESViral Abundance and General PropertiesViruses are simple biological agents typically 20ndash200 nm indiameter composed of a nucleic acid genome in a proteincoat that infect cells and ldquocommandeerrdquo the cellrsquos machineryto make more viruses which are released into the environ-ment when the host cell lyses or bursts A particular virus isthought to be capable of infecting only a narrow range of hosts(usually one species sometimes a genus rarely broader)Although some early studies had isolated viruses from thesea therewere no data prior to the 1980s showing such viruseswere very abundant and more significantly no evidence thatinfection was occurring in any important part of the planktoncommunity It wasnrsquot until the late 1980s that electronmicro-scopy with suitable concentration methods showed thatviruses are extremely abundant similar to or even higherthan bacterial abundance (212 213) Interestingly themost suitable transmission electron microscopy (TEM)approach used by Bergh et al (described in detail by [214])was actually similar to a direct centrifugation method devel-oped in 1949 (215)
TEM studies of viruses permit high-resolution images andobservation of viral morphology showing features such ashead diameter tails sheaths and tail fibers (Fig 6 collage
in upper left) Each type of virus has a fixed morphology(unlike bacteria that are potentially more plastic in theirappearance) and hence a coarse measure of viral diversityhas been possible by cataloging morphologies Studies thathave done so have found dozens or more different morpholo-gies of marine viruses in a given sample (217ndash219) A largeproportion resemble bacteriophages which are viruses thatinfect bacteria
The availability of brightly fluorescent nucleic acid stainsand high-porosity fine pore size (002 μm) filters made fromAl2O3 have made it easy to accurately count viruses by epi-fluorescence microscopy (38 220 221) Epifluorescence per-mits abundance estimates but does not allow observation ofviral morphology as viruses are below the resolution limitof light microscopy viruses are visible only as sources of light(like stars in the night sky see Fig 6 lower micrograph) Anextension of manual epifluorescence counts is the use of flowcytometry to count viruses now commonly employed in somelabs (222)
Virus abundance has been found to be closely related tobacterial abundance with a virusbacteria ratio typically10ndash301 A typical oceanic profile of bacterial and viral abun-dance is shown in (Fig 6 panel on right) This tight ratioand the strong correlation to bacterial abundance in relationto weaker correlations to chlorophyll have been cited as
FIGURE 6 Marine viruses (upper left collage) viewed by transmission electron microscopy These are cyanophages in the groups myoviridae(right and bottom) siphoviridae (left) and podoviridae (top) Scale bars represent 100 nm From Sullivan (216) Epifluorescence micrograph(lower left) of SYBR green I stained viruses (small fluorescing objects) and bacteria + archaea (large fluorescing objects) Depth distribution ofvirus and bacteria + archaea abundances obtained by epifluorescence microscopy of SYBRGreen stained cells in the central San Pedro BasinCalifornia (11 August 2000) doi1011289781555818821ch422f6
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-15
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
evidence that most marine viruses infect bacteria rather thaneukaryotic phytoplankton (217 218 223 224) though thereare also many important viruses of phytoplankton and otherorganisms with significant impacts on primary productivityblooms and ecosystem function (225ndash227)
The first demonstration of the activity of marine viruseswas presented by Proctor and Fuhrman (228) who showedby electron microscopic examination that viruses wereactually infecting marine bacteria and cyanobacteria at ameasurable rate Because only the final portion of the viruslife cycle is visible by electron microscopy (when the virusesare assembled and ready to lyse the host cell) only a very smallfraction of the infected cells can be counted as infected at anygiven time Proctor and Fuhrman (228) reported that only afew percent of the total bacteria from coastal waters and theSargasso Sea were visibly infected but they interpreted thedata with a model that implied the actual fraction of the totalcommunity that was infected was much higher They con-cluded that the total fraction of bacterial mortality attribut-able to viruses is roughly 10ndash40 Subsequent studies haveused refined versions of that same general approach andmodel (229 230) and numerous studies have used a varietyof alternative approaches to estimate viral activitymdashall yield-ing basically the same general conclusion (although themodel parameters need to be adjusted for cyanobacteriawhich on average seem to be infected less than bacteria)These approaches include (a) calculation of virus turnoverfrom decay estimates (231) (b) measurement of viral DNAsynthesis by incorporation of tritiated thymidine (232) (c)observation of effects of added viruses (233 234) (d) estima-tion of bacterial mortality in the absence of protists (e) use offluorescent viruses to estimate production by an approachanalogous to isotope dilution (38) and (f) dilution of virusesin filtered samples to estimate viral production (235) Theoverall consensus of these studies is that the initial estimateswere basically correct that is that viruses are responsiblefor about 10ndash40 of the bacterial mortality in most marinesystems (reviewed by [218 223 226 236 237]) Howeverthere is still some disagreement about the higher estimatesand it is likely that 40 mortality from viruses alone is nottypical for most marine systems
Most of the foregoing work has focused on the viral infec-tion of bacteria primarily thought to be heterotrophicalthough the original report by Proctor and Fuhrman (228)also noted the occurrence of cyanobacterial infection Sev-eral subsequent studies focused on phytoplankton includingthe potential effect of viruses on the termination of phyto-plankton blooms of Emiliania and Phaeocystis and the likelyimpact this might have on release of climate-active gasessuch as dimethyl sulfide (225 227 238ndash247) Althoughdetails are beyond the scope of this chapter viruses arethought to infect virtually all marine organisms with poten-tially significant impacts from zooplankton to whales (226)Detailed studies of viruses infecting cyanobacteria such asSynechococcus have shown differences in viral host specific-ity particularly toward coastal and oceanic host strainsand some occasional high virus abundances (to 105ml) asmeasured by most probable number (MPN) cultivation tech-niques in the Gulf of Mexico near Texas (248 249) Othervirus cultivation studies with Prochlorococcus and Synecho-coccus in oligotrophic waters of the Sargasso Sea showedan interesting pattern of cross-infection between these gen-era by some virus types (suggesting gene flow among theseorganisms) but generally low MPN estimates of abundanceto 103ml even when cyanobacterial abundance was near105ml (216 250)
Overall the consensus emerging from direct comparisonsof viral-mediated mortality and grazer-mediated (ie micro-zooplankton) mortality have indicated that viral lysis of bac-teria constitutes a significant fraction of total mortality ofthis assemblage while the mortality of phytoplanktonappears to be dominated bymicrozooplankton inmost instan-ces (251 252)
As mentioned viruses have significant morphologicaldiversity as observed in TEM studies It is also possible toinvestigate aspects of their genetic diversity In early workon this topic Wommack et al (253) and Steward et al(254) observed the diversity of viral genome lengths in a fieldsample by pulsed field gel electrophoresis with viral genomesranging 25 kb to gt300 kb in length Field results show thatthe viral community composition is dynamic in space andtime with clear changes in the banding patterns over seasonsand locations in Chesapeake Bay (253) between oceanbasins and subsequent to dinoflagellate blooms (254) andwith depths to 500 m at one location (255)
Although viruses do not all share a set of core genesthat allows a universal viral phylogeny (comparable to SSUrRNA gene in cellular organisms) genetic diversity amonga single group of viruses can be examined by sequence analysisof shared genes within the group An example is the g20 geneinT4-like cyanophages that has shown extremely high diver-sity even among very closely related viruses and has demon-strated geographic and seasonal variation (256) A secondexample is g23 which is found broadly in diverse and wide-spread T4-like phages (257) and can have seasonally repeat-ing patterns (258) as well as short-term rapid dynamics thatcorrelate to those in bacteria (259)
Viral MetagenomicsAs with cellular organisms metagenomics can be used toexamine viral diversity and genetics without themany restric-tions of cultivation Viruses can be collected by selective fil-tration and concentrated by tangential flow filtration orflocculated with iron chloride (260 261) then their collec-tive metagenome can be extracted linker amplified and ana-lyzed by sequencing (262 263) Viral metagenomic studiesare particularly challenging because the large majority ofsequences have no annotated matches in any databases butso far results from marine samples around the world haveshown extremely high diversity and variations with depthlocation and time presumably with highly dispersed typesselected by local conditions (262ndash266) Because metage-nomes are best interpreted when there are representative cul-tures available the best matches of viral metagenomesoriginally tended to be to the few viral isolates infecting trulycommonmarine bacteria like cyanophage infecting Synecho-coccus and Prochlorococcus (267) However the developmentof dilution-to-extinction cultures representing common het-erotrophic marine taxa like Peligibacter (a member of theSAR11 clade) and SAR116 has allowed isolation from sea-water of viruses infecting these common organisms and theseisolates have indeed been found to be highly abundant inmarine viral metagenomes (268 269) New approaches tointerpret the results include clustering the proteins independ-ent of known proteins to compare samples to each other andtry to find environmental factors driving viral communitychanges (270) Such analysis of a large global data set (TaraOceans expedition) has shown that extensive sampling hascome close to reaching the total diversity in tropical and tem-perate waters of such viral protein clusters (which essentiallyrepresent various viral protein motifs not all viral proteintypes) and that viruses appear to be directionally dispersed
422-16 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquodownstreamrdquo in major ocean currents as one may expectfrom first principles but also supporting the ldquoseed bankrdquohypothesis of viral biogeography (271)
Viruses and Host DiversityViruses are themselves thought to be instrumental in drivingincreased diversity of their microbial hosts via a hypothesizedprocess often called ldquokill the winnerrdquo This is because viralinfection is host-specific and density-dependent the latterbecause viruses diffuse from host to host so an abundanthost is more likely to pass on infection than a rare oneThis means that if an organism becomes abundant andblooms winning the competition for resources it becomesmore susceptible to a viral epidemic This would benefit therarer organisms and thus help foster diversity (223 272273) However bloom scenarios as described above involvesystems far from steady state and the formal theory of Thing-stad and Lignell (274) has interesting steady-state solutionswhere several viruses infect several hosts stably over time(via trade-offs between growth rates and viral susceptibility)which may occur at the strain or species level (274) There issome experimental evidence that viruses have effects on nat-ural marine microbial community composition weaker thanthe kill the winner (bloom version) hypothesis would sug-gest (275 276) so something resembling the steady-statecoexistence described in the model may in fact be commonThere also appear to be processes that foster coexistencebetween viruses and hosts but the mechanisms are largelyspeculative (223 277 278)
Viruses may also be directly involved in host genetic diver-sity because they can be the agents of genetic exchangebetween microorganisms (223 273) This often involvesthe viral lifestyle known as lysogeny whereby viruses survivewithin host cells as DNA only integrated into the host chro-mosome and being reproduced each time the host divides Ahost harboring such a genome is called a lysogen becauseunder conditions of stress to the host cell a genetic switchmay cause the viral genome to initiate the lytic process pro-ducing many progeny viruses and bursting from the hostLysogeny is a very common property occurring in a signifi-cant part of the bacterial community (279 280) althoughthe incidence of induction of the lytic phase in nature is appa-rently low (281) Overall lysogeny is poorly understood butthought to have both positive and negative impacts on themicrobial community (282) Recent results suggest that ben-efits of being lysogenic in highly seasonal polar seas leads tofundamental difference between polar and other marine viralcommunities (283)
Viruses and the Microbial LoopAs part of the food web viruses occupy a unique positionThey infect host cells that are mostly thought to be heterotro-phic bacteria and by doing so they typically burst the hosts torelease progeny viruses and cellular debris But what is the fateof this material Viruses themselves do not last indefinitelyand a simple steady-state assumption implies that from eachburst of viruses (typically 20ndash100 per lytic event) only onesuccessfully infects another cell The rest are inactivatedand broken down by sunlight (UV and visible exposure)and enzymatic attack (284) or consumed byminute phagotro-phic protists (285) thus reentering the food web as substratefor bacteria or food for protistan consumers Experiments incontrolled laboratory systems and field studies with radioac-tively labeled viral lysis products have supported the conclu-sion that most of the organic matter released by the viralinfection is either taken up by bacteria or respired (286 287)
Modeling this process as part of the microbial loop showsthat viral lysis represent a sort of side loop that has the neteffect of remineralizing a significant amount of the carbonand nutrients that enter the bacteria-protist part of themicro-bial loop (Fig 7) A theoretical numerical steady-state modelcomparing a systemwith no viral activity to onewhere virusesare responsible for 50 of bacterial mortality showed that thesystem with viruses had 33 more bacterial production andrespiration than the virus-free system implying that theviruses had the effect of permitting the bacteria to processmore of the primary production than they would otherwise(223) Although 50 is a high number unlikely to be com-mon in the sea this model nevertheless illustrates that virusescan reduce the amount of energy reaching higher trophic lev-els The implication is that viruses lead to increased bacterialactivity at the expense of the larger organisms
MAJOR ENVIRONMENTAL CONTROLSLight Temperature and PressureTemperature has an important potential influence on bio-chemical reactions and therefore on biological processesin general Most ocean waters fall in the range of ndash2degC to30degC with obvious exceptions in hydrothermally heatedareas Temperature has long been known to be a regulatingfactor for the growth of heterotrophic microbes
In temperate waters it has been established that microbialactivity is generally much higher in warm summer waters thanin winter (55) The relationship is not simple howeverbecause multiple factors act at the same time Some contro-versy still exists regarding the highest and lowest extremesfor marine bacterial growth although there is broad agree-ment that bacteria grow gt100degC at hydrothermal vents andltminus5degC in sea ice brines Pomeroy et al (57) noted the inter-esting observation that bacteria seem particularly inhibitednear the freezing point of seawater (ca ndash22degC) comparedto eukaryotic phytoplankton This effect results in polarspring phytoplankton blooms that accumulate organic car-bon in advance of the response of the bacterial communityand development of the microbial loop and perhaps lead toenhanced benthic-pelagic coupling (288)
The relationship between temperature and the growth rateof marine phytoplankton was described broadly in a now
FIGURE 7 Modification of the microbial loop concept that incor-porates the functional role of viruses Export can be via predation orsinking From Fuhrman (223)doi1011289781555818821ch422f7
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-17
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
classic paper by Eppley (58) and later Goldman and Carpen-ter (289) Temperature was shown to exert a strong and directeffect on themaximal growth rates of these species withmax-imal intrinsic growth rates at 0degC generally lt1 divisiondaywhile growth rates at 30degC may be gt4day These relation-ships indicate the maximal rate that might be attained byphytoplankton at these temperatures but they do not takeother factors into account (nutrients light) While warmtemperature ostensibly allows more rapid growth it createshydrographic conditions that typically give rise to nutrientlimitation of algal growth Thus some of the coldest watersin nature witness some of the most massive phytoplanktonblooms (290) while warm oceanic gyres represent some ofthe most oligotrophic areas of the ocean The multiple andoften conflicting effects of temperature on the growth of nat-ural phytoplankton assemblages limit the accuracy of presentpredictions regarding how primary producers will respond toclimate change (291)
An analysis of the effect of temperature on the growth ofheterotrophic protists has indicated that the response is qual-itatively similar to that of phytoplankton but with a twistThe growth rates of at least some protozoa can exceed thoseof phytoplankton at warmer environmental temperaturesbut the opposite effect is apparent at very low environmentaltemperature This differential effect of temperature on thegrowth of phototrophic and heterotrophic protists was basedon a large meta-analysis of published protistan growth (59)That analysis demonstrated that the maximal growth ratesattained by phototrophic protists could exceed the maximalgrowth rates attained by heterotrophic protists (all otherpotential growth-limiting factors not considered) Thereforeprotozoan growth rates may be constrained to a greater degreeat low environmental temperature than rates for phytoplank-ton If so then phytoplankton bloomsmay get a head start ongrazers during spring in polar ecosystems This scenario is con-sistent with information on seasonal biomass changes andmicrozooplankton herbivory in the Ross Sea Antarctica(292 293) but there are still too few data to fully vet thishypothesis
The importance of high pressure on bacterial growthgained considerable attention in the late 1960s when thedeep sea submersible Alvin was accidentally lost overboardwith its hatch openWhile no lives were lost in this accidentsome workmenrsquos lunches sank to the bottom (sim1500 m)inside the submersible Alvin was recovered after 10 monthsand interestingly there was a waterlogged lunch containingapples bologna sandwiches and broken vacuum bottleswith broth that all appeared hardly degraded and tasted palat-able Yet when placed in a refrigerator on the ship these itemsdegraded relatively quickly Initially pressure was thought tobe the preserving factor as the sea floor temperature was sim-ilar to the refrigerator temperature and the only major differ-ence would be pressure (294) Following this observationa series of experiments to measure degradation of variousorganic materials left in the deep sea for extended periodsindicated that degradationwas typically significantly reducedimplying that pressure reduces the degradation rates (295)Nonetheless changes in the protozoan community of naturaldetrital material sinking to the deep ocean floor indicate thatthe microbial community can respond relatively quickly insome situations (296) Deep sea microbiology has advancedconsiderably yet it is still difficult to interpret results withrespect to actual in situ rates of naturally occurring organicmatter Deep sea bacteria adapted to high pressures havebeen isolated that are barophilic (also called piezophilic)meaning that they prefer high pressures and have reduced
activity at lower pressures whereas others are barotolerant(piezotolerant) tolerating but not preferring high pressuresfor example see (297 298)
Few data are available on barotolerantbarophilic marineprotozoa Protozoa certainly exist and grow at great oceanicdepths but measurements of in situ growth rates for these spe-cies do not yet exist Measurable protozoan numbers havebeen documented in the deep-sea sediments for more than30 years (299 300) and viable protozoa have occasionallybeen cultured from these environments (301ndash305) butvery few direct measurements of the activities of these speciesin situ have been reported (306) A few protozoa have beenisolated that will grow at high pressure (302 303 307) andprotists that appear to be unique to the deep ocean havebeen observed either directly or through the analysis ofDNA sequences (308ndash311) but possibly the best direct evi-dence that protozoan activity takes place at the high pressurescharacteristics of the deep sea are experimental and observa-tional work noting the stimulatory effect that detrital deposi-tion has on some components of the protozoan community(296 312) These observations indicate a diverse and activeprotozoan fauna of the deep ocean although their biogeo-chemical significance is largely uncharacterized at this time
Dissolved and Particulate Organic MatterBacteria and archaea are thought to be by far the most impor-tant organisms with respect to the processing of dissolvedorganic matter (DOM) and nonliving particulate organicmatter (POM also called detritus) in the ocean While theremay be some uptake of DOM by protists particularly forgrowth factors needed in trace amounts such as vitamins(313) the bulk of this material is probably utilized by bacteriaand archaea (314) including the smallest cyanobacteriumProchlorococcus (315 316) which thus may be considered amixotroph Due to their small size bacteria have extremelyhigh surfacevolume ratios and combined with their over-whelmingly high abundance an extremely high integratedsurface area Protozoa tend to obtain the organic materialsthat they require for growth from their prey rather thanthrough the uptake of DOM Overall protozoa tend to besources of dissolved and detrital organic substances throughthe excretion of unassimilated prey biomass in expelledfood vacuoles
Particulate organic matter is not directly available as sub-strate to bacteria These substances must first be reduced tosmall molecules that can be transported into the cell Thisis accomplished by the production of extracellular enzymes(note that few if any large polymers are directly taken up bybacteria with the possible exception of DNA) Hydrolyticenzymes produced by bacteria (and almost certainly archaea)break down polymers like proteins polysaccharides andnucleic acids As with DOM POM is composed of a complexmixture of compounds that vary in their susceptibility to bac-terial degradation and utilization
Particulate material in the water column serves not only asbacterial substrate but also as substratum POM occurs in thewater column across a huge size spectrum from micrometersup to some detrital aggregates more than 1 m in diameter(317) Much of this particulate material is in a constant stateof flux with colloidal material constantly coalescing andaggregating to form new or larger particles (318) as microbialdegradation acts simultaneously to remineralize this materialDetrital particles that attain macroscopic size either by directformation (317 319 320) or via accretion and aggregation(321) and are often called marine snow or macroaggregatesMarine snow particles are readily colonized by bacteria and
422-18 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
many other microorganisms and their abundances on theseaggregates can be orders of magnitude greater than abundan-ces in an equivalent volume of the water surrounding theaggregates (320 322ndash324) and their diversity and highlyconcentrated abundances on detrital aggregates implies thatthere may be stronger trophic coupling and efficient energyelemental cycling relative to these processes among free-living microorganisms in the surrounding waters This specu-lation has resulted in a number of formalizations of the poten-tial impact of aggregate microbiology on biogeochemicalprocesses in the ocean (325 326) Most definitions of bio-films include aggregated particulate and these particles areoften studied within the biofilm conceptual model (see chap-ter by Lawrence et al in this section)
Inorganic Nutrients
Macronutrients (N P)Virtually all heterotrophic organisms in the ocean contributeto the pool of available macronutrients via the excretion ofmetabolic wastes and protozoa contribute significantly tothis process Macronutrients are often in excess within proto-zoan prey relative to the consumersrsquo needs because much ofthe prey carbon is respired to produce energy for metabolismand growth ldquoExcessrdquo nutrients are eliminated as eitherorganic compounds or more commonly as ammonium (forN) and phosphate (for P) The importance of the match(or mismatch) between predator and prey stoichiometry hasbeen demonstrated experimentally and repeatedly in marineand freshwater ecosystems and has been elevated to a funda-mental ecological principle governing elemental and energyflow in aquatic food webs (327) Based on this reasoningCaron et al (328) concluded that bacterivorous protozoaplay an important role in nutrient remineralization andrelease in the ocean Bacterial biomass is typically rich in Nand P relative to other organisms so bacterial predators expe-rience large excesses of these elements relative to their growthneedsMoreover bacterivorous protozoa should be dispropor-tionately important in nutrient remineralization amongsingle-celled eukaryotes because as the smallest protozoa inaquatic ecosystems they have high weight-specific metabolicrates (329) Thus both stoichiometric and allometric rela-tionships implicate small bacterivorous protozoa as impor-tant sources of remineralized nutrients in the plankton
The reasoning also holds true for bacterial utilization oforganic compounds and it has led to some interestingfindings and conclusions regarding the role of bacteria innutrient cycling in the ocean Bacteria are important sourcesremineralized nutrients when they are consuming N- orP-rich substrate Contrary to this traditional view howeverheterotrophic bacteria are often strong competitors for inor-ganic nutrients under many situations largely because bacte-rial biomass is richer in N and P than much of the organicmatter they consume in nature so they require additional Nand P to produce their biomass (summarized by [330]) Exper-imental studies have demonstrated the uptake of ammoniumby bacteria other than cyanobacteria using stable isotope 15N(331) and also the short-lived radioisotope 13N (332) oftenfinding that bacteria may be responsible for one third of the Nor P uptake Additionally there is evidence that bacteria insome oceans like the Sargasso Sea are limited by phosphorus(333 334) Growth limitation of bacteria by phosphorusappears to be a common phenomenon of freshwater ecosys-tems (335 336)
N2 fixation is restricted to prokaryotes and in the oceanwater column it was thought for many years to be done
primarily byTrichodesmium a warm-water colonial cyanobac-terium that blooms sporadically andRichelia a cyanobacterialsymbiont that lives within certain diatoms (337) Some uni-cellular cyanobacteria also contribute significantly to globalN2 fixation (338) and molecular biological data suggestthat a variety of other bacteriamdashnot phytoplanktonmdashmayalso be fixing nitrogen in seawater (339) An exciting recentdiscovery in this field is that a bacterial nitrogen fixer origi-nally known only from nifH sequences UCYN-A has hadits genome sequenced by parallel pyrosequencing and wasfound be a cyanobacterium (Candidatus Atelocyanobacteriumthalassa) lacking the apparatus to generate oxygen as well asother pathways (340) Further study showed it to be symbioticwith a marine alga (341)
Micronutrients (Trace Metals Growth Factors)Bacterial growth may often be limited by macronutrients asdescribed and sometimes by trace nutrients particularlyiron that occur at very low concentrations in seawater(342 343) The importance of Fe as an element limiting pri-mary production in some oceanic regions has come underclose scrutiny and extensive experimental investigation inrecent years Many bacteria have special uptake mechanismsto utilize extremely low concentrations of iron includingsiderophores (released compounds that bind iron and arethen specifically taken up) Even if bacteria are capable ofgrowth at low Fe concentrations growth may be less efficientKirchman et al (344) found that Fe limitation stronglyreduced bacterial growth efficiency in a cultured marine bac-terium (ca 50 efficiency Fe replete but lt10 when Fedeplete) Thus the effects may be complex Marine N2 fixa-tion also may be limited by the availability of Fe becausenitrogen fixers have high Fe requirements
Protozoa tend to be a source rather than a sink for Fe in theocean analogous to their role in macronutrient remineraliza-tion Protozoan grazing activity has been shown to releaseiron from bacterial biomass thereby relieving Fe limitationin co-occurring phytoplankton (345) a mechanistic demon-stration of the process using protozoa growing on Fe-repletebacteria However it has also been reported that Fe-limitedbacteria may contain insufficient Fe for the bacterivorous pro-tozoa that consume them thus leading to Fe limitation in theprotozoan as well (346)
OxygenHypoxic and anoxic regions of the ocean are expected to sig-nificantly expand in the coming decades as a consequence ofchanges in ocean stratification resulting from global climatechange (347) As oxygen becomes depleted bacteria havethe capability to utilize a series of alternate electron acceptorsCompositional changes in the bacterial assemblage accom-pany these changes in metabolic activity but are beyondthe scope of this chapter Many of the studies on benthicmicrobiology have focused on redox reactions that occurthere Oxygen is usually consumed by sediment microbialactivity much faster than the rate at which it can be replacedby diffusion so most sediments especially organically richones are anaerobic below a few mm (fine-grained muds) orcm (coarser sediments) from the sedimentwater interfaceexcept where animals ventilate the benthos through tubesWhen oxygen is absent bacteria and archaea use alternateelectron acceptors such as nitrate nitrite oxidized Fe orMn organic matter or sulfate The use of nitrate or nitriteas an electron acceptor with concomitant production of N2
gas (denitrification) results in net loss of biologically availablenitrogen Bacteria that perform this reaction are typically
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-19
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquofacultativerdquo organisms that can switch from oxygen to nitrateor nitrite Organic matter may be reduced by fermentationreactions in which part of an organic substrate is oxidizedwhile another is reduced fermenter organisms are also typi-cally facultative Sulfate is a particularly important alternateelectron acceptor in marine systems because it occurs as thesecond most abundant anion in seawater (behind chloride)typically 28 mM Its use as an electron acceptor leads to theproduction of elemental sulfur or sulfide The use of sulfateas an electron acceptor yields much less energy than oxygenor nitrate and sulfate-reducing bacteria are strict anaerobesunable to use oxygen (often strongly inhibited by oxygen)Methane can be produced by methanogenic archaea whichare all strict anaerobes using CO2 as an electron acceptor(see Findlay and Battin this section for further discussion)
Reduced compounds such as sulfide are produced by theuse of these alternate electron acceptors and they diffuse awayfrom their source When they diffuse to environments with astrong oxidant like oxygen they may be oxidized by chemo-trophic bacteria in energy-yielding reactions Similarlyammonium can be oxidized to nitrite and then nitrate by che-motrophic nitrifying bacteria and the nitrate can diffuse intoanaerobic layers and become denitrified to N2 Such coupledoxidation-reduction reactions can lead to accelerated biogeo-chemical cycling particularly in sediments
It was originally believed that relatively few free-livingspecies of protozoa inhabit anaerobic and hypoxic marineenvironments This view has changed based on (largely)molecular analyses of some of these environments (308348ndash351) A significant number of previously undescribedeukaryotic phylotypes have been observed in anoxic environ-ments High protozoan biodiversity in the absence of molec-ular oxygen is not completely unexpected because otheranaerobic environments such as some gut environments(eg cockroach guts and cattle rumen) have notoriouslyabundant and diverse protozoan fauna but marine anaerobicenvironments have been difficult to access and sample prop-erly Many anaerobic protozoa exist together with symbioticbacteria (352 353) or possess unique organelles such as thehydrogenosome (354)
MAJOR BIOTIC CONTROLSThe Trophic Activities of Protozoa Predation andTop-Down ControlsPredator-prey relationships are fundamental interactionswithin biological communities linking production of bio-mass on the one hand with removal on the other hand Estab-lishing the trophic relationships among microorganisms andquantifying the rates at which they take place constitutemajor endeavors within marine microbial ecology As notedpreviously protozoa consume a wide variety of prokaryoticand protistan prey Numerous laboratory and field measure-ments have been made throughout the past quarter centuryto attempt to quantify the rates of bacterial and phytoplank-ton consumption The methodology for accomplishing thesestudies has varied and a number of approaches are presentlyemployed
Methodologies for estimating microbial grazing are diversebut generally fall into one of two experimental approachesperturbation experiments or tracer experiments Perturbationexperiments (size fractionation metabolic inhibitors anddilution technique) rely on manipulation of the consumeror prey assemblages in somemanner such that the effect is evi-dent from changes in the abundance of the prey population
during incubations relative to control treatments that receiveno manipulation Tracer experiments (radioactively labeledprey fluorescently labeled prey stable isotopendashlabeled pray)attempt to use labeled prey or a prey proxy to quantitativelyfollow the movement of prey biomass into consumers Forsummaries of these methods see (74 355)
Perturbation experiments have entailed a variety of mech-anisms to ldquodecouplerdquo predators from prey in natural microbialcommunities For example size fractionation uses various sizefilters to remove an entire size class of grazers from awater sam-ple leaving the prey to grow in the relative absence of thosepredators during subsequent incubations Unfiltered samples(controls) indicate the growth of prey in the presence of pre-dation Predation impact is obtained by the difference ingrowth of the prey between these treatments This methodhas been used to examine predation on cyanobacteria heter-otrophic bacteria and minute phototrophic eukaryotesAlternatively metabolic inhibitors have been employed tohalt predation (or in some cases prokaryote growth) insteadof physical removal by filtration
The dilution technique (356) has become a commonmethod for examining grazing mortality of phytoplanktonby microzooplankton (which is comprised predominantly ofprotozoa) This method relies on the dilution of herbivorouszooplankton in a series of subsamples (and concomitantmeasurements of phytoplankton growth in each subsample)to estimate the grazing mortality of the herbivores
Tracer experiments have used both radioactively labeledprey and fluorescently labeled prey to examine predation onbacteria and phytoplankton Radioactive labeling has beenaccomplished using 14C- or 3H-labeled organic compounds(for bacteria) or 14C-bicarbonate (for phytoplankton) Theuse of radioisotope approaches has diminished with improve-ments in the number and types of fluorescent compoundsavailable for this work Fluorescently labeled prey havebeen labeled with a variety of compounds that allow theprey to be observed readily (even inside predators) by epi-fluorescencemicroscopy (357 358) Alternatively a decreasein the total number of fluorescent prey in a sample indicatesthe rate of removal of the prey assemblage
Both perturbation experiments and tracer experimentsdescribed have inherent advantages and disadvantages Forexample perturbation experiments are fairly straightforwardto conduct but they may introduce artifact by removingpotentially important sources of nutrients or organic com-pounds released by predators (359) On the other hand thedilution technique is very labor-intensive but one advantageof this method is that it provides the opportunity to determineboth phytoplankton growth and grazing mortality in a singleprocedure Most tracer experiments are fairly easy to conductand generally do not perturb the food web being studied butfeeding selectivity for or against the tracer (relative to naturalprey) may produce spurious results In addition some of thesemethods provide a measurement of the activity of the entiregrazer assemblage and others are applicable for examiningthe activity of specific taxa For example size fractionationapproaches examine the effect of the removal of an entiresize class of organisms (a community-levelrdquo measurement)whereas fluorescently labeled prey can be used to examinethe ingestion rate of a particular species by observing theuptake of labeled prey into that species (a species-levelmeasurement)
Predation rates of numerous marine protozoa have beenexamined in the laboratory From these experiments a gen-eral knowledge of the rates of consumption by these specieshas been accumulated Feeding in most protozoa shows a
422-20 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
functional response to the availability of prey that is similar tothe response of many organisms (although there are a greatdeal of species-specific differences) That is ingestion rateincreases rapidly as prey abundance increases until a maximalingestion rate is attained (360)Manyof these curves have theform of a Type II functional response curve (361)
Based on the features of these curves it is clear that proto-zoa have the ability to ingest substantial numbers of prokar-yote and eukaryote prey in natural ecosystems Somebacterivorous flagellates can consume bacteria (and cyano-bacteria) at rates up to 100 s per individual per hour whereassome bacterivorous ciliates can consume thousands of bacte-ria per individual per hour Rates of consumption of eukary-otic phytoplankton by herbivorous ciliates and largeflagellates are considerably more variable and highly depend-ent of the feeding mechanism (eg filtration in ciliates versuspseudopodial net in dinoflagellates) and the size of the prey
These lab studies demonstrate that rarely are prey suffi-ciently abundant in nature to support the maximal ingestionrates of which many protozoa are capable Nevertheless pre-dation rates of protozoa in nature are sufficient to constitute amajor factor controlling the standing stocks of bacteria andphytoplankton (63 180 184 355) For example these sum-maries indicate that on average protozoa consume 20ndash80ofthe standing stock of phytoplankton (measured as chloro-phyll) in the world ocean and 60ndash75 of daily primary pro-duction a departure from the classical view of copepods as theprimary consumers of phytoplankton This large role of her-bivorous protozoa infers a large role for copepods as consumersof planktonic protozoa (185) Similarly the role of protozoaas major consumers of bacterial biomass is indicated by therelative balance between bacterial production on the onehand and bacterivory on the other (Fig 8) The approximateone-to-one balance between these measurements over a largenumber of studies implies that protozoan bacterivory must bea major factor determining the fate of bacteria in marine (andfreshwater) plankton ecosystems
It is interesting to note that some types of bacteria includ-ing marine Bdellovibrio are known to prey on other marine
bacteria (362) At the time of this writing the importanceof bacterial predation by other bacteria within the contextof oceanographic processes is not known and presumed tobe small While Bdellovibrio may be cultivated from seawaterit can also be found among sequences from cold sediments(363) and the water column Bdellovibrio happens to be thebest studied predatory bacterium but there are no doubtothers that we simply have not yet identified (364)
Species-Specific Interactions (MutualismParasitism Commensalisms)Interactions between marine microorganisms such as compe-tition allelopathy and symbiosis are fundamental aspectscontrolling community composition and activity but atpresent most are poorly characterized in natural marine sys-tems Bacteria must compete for dissolved and particulatecompounds with other bacteria andwith protistan taxa whilephagotrophic protists must compete with each other andmetazoa for available prey These interactions manifest them-selves in many forms including true competition for availableresources resource partitioning such that some taxa specializeon the utilization of certain substrates or prey while othersspecialize on different ones (reducing competition) or eventhe cooperative use of compounds (eg one bacteriumrsquoswaste products being utilized as substrate by another bacte-rium) These metabolic consortia are probably common inthe ocean but can be difficult to identify and characterizeConsortia of bacteria working together to perform complexreactions especially in anaerobic systems are well known inmicrobiology
Consortia in which there is a very high degree of integra-tion between species (ie symbiosis) are very common in theocean These associations may be beneficial to both partners(mutualism) beneficial to one part and inconsequential forthe other (commensalism) or beneficial to one part and det-rimental to the other (parasitism) Many highly visible andwell-characterized marine mutualisms exist between chemo-synthetic bacteria and animals such as hydrothermal ventor methane seep tubeworms clams mussels or other inverte-brates (365) and also between luminescent bacteria and fishor squids (366) Similarly many protozoa form mutualisticassociations with photosynthetic protists (367) The mostconspicuous of these associations involved large amoeboidprotozoa of oceanic ecosystems (planktonic foraminiferanspolycystine radiolaria and acantharia) which contain largenumbers of intracellular algae (74) but associations betweensome heterotrophic dinoflagellates and cyanobacteria are alsocommon (368) Studies have indicated that efficient nutrientcycling in these protozoan-algal associations confers an eco-logical advantage for both partners in surfacewaters of the oli-gotrophic ocean
Parasitism is also a common symbiotic relationship amongmarine protozoa Protozoan parasites have been reported for adiverse array of protists and animals in the ocean Heterotro-phic dinoflagellates and presently uncultured alveolate cladesrelated to alveolates appear to be particularly successful atevolving thismode of existence (369ndash371) In a common par-asitic group Amoebophrya spp the parasite invades photo-synthetic dinoflagellates where it undergoes reproductioneventually lysing the host and releasing dozens of infectiveswarmers The parasitic protist Pirsonia spp is a parasite ofdiatoms (372) The overall importance of these parasitismsin the population dynamics of their hosts is not clearly knownbut may constitute a significant source of mortality for somespecies (373 374) The frequency and diversity of ldquonewrdquo
FIGURE 8 Meta-analysis of experimental studies that have com-pared rates of bacterial productivity (converted to population growthrate units of dminus1) with rates of bacterivory by phagotrophic protists(expressed as equivalent units of bacterial growth rate dminus1) Thesolid line indicates where measurements of bacterial removal by graz-ers were equal tomeasurements of bacterial production Data redrawnfrom Sanders et al (63) doi1011289781555818821ch422f8
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-21
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
small subunit ribosomal RNA gene (16S rRNA in Bacteriaand Archaea and its larger homolog 18S rRNA in eukar-yotes) This molecule is strongly conserved over evolutionarytime so this single molecule has been used for constructingphylogenetic trees of all living organisms (httptolweborgtreephylogenyhtml) Analysis of 16S18S rRNA genesequences has been used to evaluate deep evolutionary rela-tionships among organisms and was instrumental in point-ing out that Archaea Bacteria and Eukarya should beconsidered different Domains of equivalent phylogeneticrank above kingdoms (78) However there are sufficientdifferences in 16S18S rRNA gene sequences to demonstrate
relationships (ormake distinctions) at the genus or sometimesspecies level
The first phylogenetic studies based on 16S18S rRNAgenes used sequences derived from cultures However onedoes not need cultures to obtain rRNA gene sequences (orany other sequences for that matter) An idea developed inthe lab of Norman Pace in themid-1980s involved extractionof DNA directly from natural samples and then cloning andsequencing of the DNA as a means of assaying the microbespresent in the samples (103 104) The original protocolscalled for cloning by creating what are called ldquophage librariesrdquofrom the natural DNA but since 1986 PCR has been applied
FIGURE 3 (a b) Plankton biomass in the Arabian Sea during the 1995 southwest monsoon (a) and intermonsoon period (b) Areas of theboxes indicate the relative magnitudes of the biomass in each category Categories within the dashed boxes in a b c are composed of protozoaArrows indicate the direction of energymaterial flow in the food web thicker arrows depicting greater flow Redrawn from (86)(c) Depth-integrated biomass (mgm2) in the upper 100 m of the Sargasso Sea near Bermuda and in the upper 200 m of the equatorial Pacificat 175degE Thewidth of the bars indicates the biomass in each size category Heterotrophs have been separated by size class while phytoplanktonhave not Size classes delineated by the dotted box are comprised of protozoa Redrawn from (65) doi1011289781555818821ch422f3
422-8 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
extensively for cloning and related studies The targetsequence can be almost instantly ldquoidentifiedrdquo to its closestphylogenetic neighbor by what have come to be standardonline sequence comparisons
Beyond the study of targeted genes (like the 16S rRNAgene) shotgun metagenomic studies have examined theentire genetic repertoire of the microbes in a given sampleThe metagenome is the collective genome of all organismsin the sample Initially these studies extracted DNA fromall organisms in a sample (usually prefiltered through a filterapproximately 1 microm to remove most organisms larger thanbacteria) sheared it to produce fragments and clonedthem into standard vectors either as small (thousands ofbases) or large (to hundreds of thousands of bases) insertsthat is fragments of DNA from the environment nowcloned into the vectors in a form suitable for sequencingThe best known early marine study of the former type isthe Global Ocean Survey with initial results published byVenter et al (105) which generated more than a billionbases of DNA sequence and reported 12 million previouslyunsequenced genes estimated to come from at least 1800different genomic species cumulatively in the many samplesthey analyzed
With the advent of next-generation sequencing that gen-erates millions ormore sequences in a run (known by a varietyof acronyms including 454 Illumina SOLiD etc) cloningof genes has largely been replaced by clone-free sequencingThe extent and power of such sequencing has recentlybeen demonstrated by the ability to construct essentiallythe entire genome of an uncultivated marine Group II Eur-yarchaeon that constituted only approximately 2 of amicrobial community sample using SOLiD sequencingmade possible by high coverage and the use of mate pairsequencing ofsim3000 bp fragments (106) (this length isneeded to span repeats and highly conserved genes that oth-erwise make genome construction difficult)
For 16S rRNA gene studies ldquotag sequencingrdquo pioneeredby Sogin et al (107) consists of amplifying a suitable sizedpart of the gene with broadly conserved primers (choice isimportantmdashfew are truly universal for the groups intended)often ldquobarcodedrdquo to allow multiple samples to be combinedinto a single run The amplified products are then sequencedand sequences processed en masse Many thousands of partialSSU rRNA gene sequences per sample are economically ana-lyzed this way though they are usually short (currently a fewhundred bases each depending on the sequencing platform)This way of analyzing the composition of microbial commun-ities is now standard These approaches provide so muchinformation even about very rare sequences that the resultshave led to the important concept of the ldquorare biosphererdquoorganisms that may be active or dormant and constituting avery small proportion of the community (eg often muchless than 01) but are potentially important for dispersionadaptation to changing conditions or even critical activitieslike nitrogen fixation or vitamin production (108) Howeverdue to the potential for artifacts like error sequences thisapproach requires rigorous application of quality filteringand clustering algorithms to avoid erroneous taxa and overes-timation of species richness (109 110)
Next-generation sequencing approaches have also allowedwork to begin metagenomics studies of microbial eukaryotes(111) These studies are still constrained by the much largergenomes of eukaryotes and therefore the difficulties ofobtaining sufficient sequences to reconstruct the genome ofa particular species Along with advances in sequencing abil-ity studies of eukaryotic metagenomics have been facilitated
by advances in single-cell isolation which when coupledwith high-throughput sequencing approaches reduce the tre-mendous complexity present in natural complex eukaryoticcommunities to a manageable task (112) Such single-celltechniques are also quite valuable in studies of bacteria andarchaea though the amplification technique tends to bevery uneven and typically generates less than half the genomeof each isolated cell regardless of domain (113 114)
Additionally similar to DNA mRNA is amenable toextraction and sequencing although greater care must betaken during extraction and purification as RNA shows agreater susceptibility to degradation during processing Copy-ing of mRNA by reverse transcription of RNA into cDNAfollowed by DNA sequencing has allowed insights into themetatranscriptomes of environmental samples Metatran-scriptomic studies provide information on gene expressionin an ecosystem and thereby indicates ldquoactivityrdquo of themicro-bial community rather than simply ldquopotentialrdquo represented bythe genomic DNA present in the sample with many applica-tions from showing which processes are being carried outby which organisms to fine-scaled diel studies (115ndash118)Nevertheless given variations in the lifetimes of differenttranscripts and protein molecules the transcriptome maynot be fully representative of the current activity of anorganism
Limitations of these analyses include sequencing errorsPCR mismatches or biases clustering and bioinformaticschallenges and chimeras generated during PCR Also thephylogenetic resolution of short sequences is limited giventhe high conservation of rRNA sequences Even with clone-free shotgun metagenomics there may be biases such as non-random losses of DNA during extraction and preparationor biases (eg from G +C content or secondary structure)in the sequencing procedures Determination of speciesdiversity by these approaches provides an example of theselimitations Shakya et al (119) working with synthetic com-munities (purified genomicDNA from16Archaea represent-ing 3 phyla and 48 Bacteria representing 16 phyla remixed tosimulate an environmental DNA extract) applied both meta-genomic analysis (454 and Illumina platforms) and PCRamplification followed by 454 sequencing of 16S rRNA genesto determine both species richness and relative abundanceThey found that PCR amplification454 sequencing of 16SrRNA genes yielded an accurate measure of species richness(providing that appropriate data processing was applied)but that the relative abundance of up to 94 of the species(depending on domain and variable region amplified) wasover- or underestimated by at least 15-fold (values rangedfrom not detected to 103-fold overestimation) In contrastboth metagenomic approaches yielded relative abundancesthat were within the authorsrsquo 15-fold accuracy cutoff forsim50 of the species However they concluded that addressingrichness overestimation in metagenomic analyses that isdistinguishing rare but real OTUs from experimental andcomputational artifacts awaits further computational andclassification improvements More recently Parada et al(120) used mock communities composed of 16S rRNAclones from 27 common marine taxa (from nine Bacterialand two Archaeal phyla) to show that small differences inPCR primers (and different clustering methods) can yieldlarge differences in apparent relative abundances of reportedtaxa However one primer pair and informatics pipelinethey tested using a particular version of 515F-926R (V4ndashV5) provided accurate estimates of relative clone abundance(r2 = 095) when comparing observed versus expected cloneabundance
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-9
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
Culture-Independent Diversity Studies
The first groups to be identified using cultivation-independ-ent molecular techniques from the marine plankton (121)were the bacterial SAR11 cluster (a group of closely relatedgene sequences or phylotypes) and marine picoplanktonicunicellular cyanobacteria Synechococcus and Prochlorococcus)Of these the SAR11 cluster was completely unknown butthe cyanobacteria had previously been recognized by theirunique pigment fluorescence Waterbury et al (122) andJohnson et al (123) used epifluorescence to observe Synecho-coccus and Chisholm et al (124) discovered Prochlorococcususing flow cytometry These cyanobacteria were later isolatedand grown in phytoplankton culturemedia These two groupsare generally common in the euphotic zone with SAR11 typ-ically comprising one third of the planktonic bacteria (125)and the cyanobacteria common everywhere but polar waters
Probably the biggest surprise to come from the applicationof molecular tools was the discovery by Fuhrman et al (126)of abundant archaea in the deep sea The archaea were foundto be in a unique phylogenetic cluster that was only distantlyrelated to any previously known archaea but the ldquoclosest rel-ativesrdquo (not really close at all) were extreme thermophiles Asubsequent study also found archaea to be present in near-surface coastal waters albeit relatively rare (lt2 relativeabundance) This study used a PCR technique specificallytargeted archaea and found ldquomarine Crenarchaeardquo plus a sec-ond group belonging to the phylum Euryarchaea (127)
Up until this time all known Archaea were thought to beldquoextremophilesrdquomdashadapted for either very high temperatures(thermophiles) extremely salty conditions (halophiles) orstrictly anaerobic environments (methanogens) Yet theseorganisms were present in cold or cool water at ordinary salin-ity and high oxygen concentrations Fluorescent in situhybridization (FISH) measurements from deep-sea sampleshave since showed that the archaea may make up appro-ximately 40 of the total countable prokaryotes with thepercentage reaching to 60 at 200 m depth in theMediterra-nean (35 36) An extensive time series of FISH measure-ments near Hawaii confirmed that the archaea are indeedvery abundant throughout the year from below the photiczone to at least 4000 m and typically constituting 30ndash40of the total prokaryotes present in waters deeper than a fewhundred meters (128) with similar results found elsewhere(129 130) (Fig 4) They have been reported from many pla-ces including the Atlantic Pacific and Southern Oceansand are dynamic components of the plankton the most com-mon type by far being the ldquomarine Crenarcheaeardquo (128 129131) While the marine archaea have been reported to bedominated by a few major ldquophylotypesrdquo (132) they alsohave been shown to have a great deal of microdiversity withinthese phylotypes suggesting there are many kinds of close rel-atives coexisting (133) Interestingly this group of archaeamay likely be the most abundant kind of organism on Earthgiven the huge volume of the deep sea and their high abun-dance there (134) It has recently been proposed that ldquomarineCrenarchaeardquo be elevated to their own major phylum outsidethe Crenarchaeota called the Thaumarchaeota on the basisof deep phylogenetic branching and fundamental differencesbetween them and the Crenarchaeota (135) The Thau-marchaeota possess the uniquely archaeal membrane lipidcrenarchaeol and are nowalso known to be abundant in soils
The physiology of the Thaumarchaeota has been anintriguing area of study Initially an autoradiography-FISHcombination approach demonstrated that they take upamino acids (130 136) But subsequent data showed that
crenarchaeol was derived largely from CO2 suggesting auto-trophic metabolism (137 138) An experiment showinguptake of 13C-labeled bicarbonate into these archaeal lipids(139) directly pointed to autotrophy in this group Chemoli-thoautotrophywas first hinted at byVenter et al (105) whosemetagenomic analysis showed an apparent archaeal scaffoldthat contained genes suggestive of ammonia oxidation Fur-ther evidence came from Schleper et al (140) who detectedseveral ammonia oxidation genes in order directly adjacent toa Thaumarchaeota 16S rRNA gene in a soil-derived metage-nomic clone The issue was directly resolved when Konnekeet al (141) isolated a related marine archaeon from sedimentof a marine aquarium and this organism Candidatus Nitroso-pumilus maritimus was found to have a chemoautotrophicmetabolism quantitatively oxidize ammonia to nitrite andcontain an archaeal ammonia monooxygenase gene (amoA)and interestingly did not grow heterotrophically Cultivationexperiments showed this organism has a high affinity forammonium allowing it to outcompete bacterial nitrifiers atlow (submicromolar) concentrations as are typical in thesea (142) The complete genome of the organism has shownnovel adaptations for nitrification and autotrophy (143)
Even if the Thaumarchaea are primarily chemolitho-autotrophs field data suggest a level of mixotrophy in thatsome organic substrates are being incorporated into biomassA stable isotope study using cells collected at 670 m depthoff Hawaii estimated that about 80 of the carbon incor-porated into archaea-specific lipids came from inorganic sour-ces and about 20 from organic compounds (144 145)Interestingly it has also been claimed that the genomesof deep-sea (gt2000 m depth) members of the Thaumarch-aea as well as those living in equatorial waters rarely containthe amoA gene and thus may be primarily chemoorgano-trophs (48)
Major bacterial groups that have been documentedfrom seawater using 16S rRNA characterization includesome that are also known from culture (eg AlteromonasRoseobacter) and several that are phylogenetically distantfrom standard cultures The most common groups are inrough order of their relative abundance in clone librariesfrom most to least abundant SAR11 (relatives of Pelagibac-ter ubique) Roseobacter SAR86 cyanobacteria SAR116SAR202 SAR234 andMarine Group A The SAR designa-tion followed by a number is an arbitrary sequential cloneidentifier from Sargasso Sea cloning studies done by the Gio-vannoni lab that did most of the early systematic cataloging ofclones Summaries of the data and phylogenetic relationshipsof these groups can be found in Fuhrman andHagstrom (102)and Giovannoni et al (101)
Molecular Genetic Discoveries in Bacterial andArchaeal Marine BiologyAs described earlier metagenomics is an extension of theideas used in the 16S rRNA cloning studies in that all genesfrom the native microorganisms are separated and clonedwithout having cultivated the organisms These methodsearly on started to find unexpected and very interestingresults The best examples involve unexpected marine photo-trophy the first of which is discovery of a nonchlorophyllphotosynthetic bacterial pigment called proteorhodopsin(146) The gene was found on a large environmentallyderived fragment of DNA that also had a gene coding for16S rRNA from the so-called SAR86 group (one ofthe groups common in seawater) This pigment can act as alight-driven proton pump thought to permit cells to generate
422-10 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
FIGURE 4 Distribution of Bacteria Euryarchaea andmarine Crenarchaea (recently renamed Thaumarchaea) along a transect in theNorthAtlantic Ocean as measured by CARD FISH and expressed as of total bacteria + archaea counts via DAPI stained epifluorescence fromTeira et al (130) Top panel shows station locations that are shown on the top of the lower three panels with the distance in km reported alongthe transect from lower to higher station numbers doi1011289781555818821ch422f4
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-11
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ATP from sunlight Interestingly different versions of this pig-ment are found at different depths apparently ldquotuningrdquo theabsorption to match the ambient wavelengths of light asthey change with depth (147) The proteorhodopsin genesare very widespread and diverse perhaps in half or more ofall marine bacteria including SAR11 and Euryarchaeota(105 148ndash150) However almost all of several culturedorganisms with proteorhodopsin studied to date do not showa growth benefit from light (150) with the exception of amember of the Flavobacteria Dokdonia sp strain MED134that grows faster in the light only under moderately lownutrient conditions (151) and a marine Vibrio has beenshown to survive starvation longer in the light than in dark-ness (152) These observation may explain the wide phyloge-netic distribution and high abundance of this gene in thatproteorhodopsin may often assist long-term survival of bacte-ria under extreme energy-limiting conditions yet not havemucheffect on growthduringmore energy-replete conditionsInterestingly a recent report suggests the growth benefit fromlight in Dokdonia MED134 is from enhanced uptake of itsrequired growth factor vitamin B1 which is transported by atonB-dependent transporter powered by a proton gradient(153) This pigment might also have sensory or other rolesnot yet well examined (150)
The second kind of phototrophy found by moleculargenetic and fluorescence techniques to be unexpectedlyimportant in marine plankton is anoxygenic aerobic bacterialphototrophy based on the pigment bacteriochlorophyll aCulturable aerobic anoxygenic photosynthetic (AAP) bacte-ria have been known from seawater for several years (154) andare widespread (155) Initial recent reports of direct countsclaimed they represent about 11 of the total bacterial com-munity in the euphotic zone (156) but those early counts didnot correct for the presence of other bacteria and such correc-tionoften yields estimates averaging closer to 2(157)Directmeasurements show the bacteriochlorophyll pigment is rela-tively rare (158) However studies in various ocean locationsincluding ocean gyres report that these AAP bacteriacan indeed on occasionmake up a quarter of the total prokar-yotes (159 160) A recent energetic modeling of AAP andproteorhodopsin-containing bacteria (161) has suggestedthat proteorhodopsin-containingones cangain approximately02asmuchenergy from sunlight asSynechococcus (commonmarine cyanobacterium) and AAP bacteria can gain approxi-mately 13 as much energy from sunlight as SynechococcusThey concluded the AAP bacteria may gain energy sufficientto meet maintenance costs but proteorhodopsin-containingbacteria were not expected to do so except at high light inten-sities andwith largenumbersof proteorhodopsinmoleculespercell They concluded the ease and low cost of maintainingPR-based phototrophy (a few genes required) may explainthe high incidence of proteorhodopsin genes
Other metagenomic studies have yielded interestinginsights about organisms and processes that would otherwisebe difficult or impossible to establish For example thesequence of an archaeal gene fragment isolated from seawaterrevealed extensive evidence of genetic exchange with othertypes of archaea and even bacteria (162) Such geneticexchange was also quite evident from the complete genesequences of different strains of Prochlorococcus and Synecho-coccus which was attributed in part to virus-mediated geneflow (163ndash166)
Dilution Cultures of Bacteria and ArchaeaSome recent cultivation techniques that permit growth ofldquotypicalrdquo oligotrophic marine bacteria in pure culture are
based on the simple concept that bacteria living in seawateron dissolved organic matter might best be grown in the labo-ratory in ordinary filtered seawater This conclusion followsfrom similar techniques to grow mixed ldquoseawater culturesrdquoof marine bacteria (167) but the seawater inoculum in thiscase is diluted so that only one or a few bacteria are addedto the initial culture vessel (168) A rapid throughput versionof this method has been used to cultivate members of bacte-rial groups thought to be conventionally uncultivable such asthe SAR11 clade (169 170) albeit often at very low den-sities Such cultures permit focused studies of the propertiesof those organisms including genomic analysis (169 170)This valuable work provides insight into the roles and activ-ities of some of the most common bacterial phylotypesobserved in the ocean (171) showing for example that culti-vated SAR11 requires a source of reduced sulfur as well asproviding templates aiding the interpretation of metage-nomic analysis (172)
PROTOZOADiversity and DistributionHeterotrophic protists have been identified historically fromtheir morphological features that are apparent at the levelof the light or electronmicroscope Features of cell size shapetype and pattern of flagellationciliation skeletal structurescharacteristics of the nucleus and other cellular structuresand organelles have been used to differentiate themany thou-sands of described species A description of the extent of thisdiversity of form and function is well beyond the scope of thisbook (see [79] for more information) However much of thisdiversity can be reduced to three basic body plans that dictatethe broadest ecological roles of these cells amoeboid flagel-lated and ciliated forms (Fig 5) Among the lineages possess-ing one of these three forms only the last group the ciliatedprotists constitute a monophyletic group within the domainEukarya
One of the simplest body plans for protozoa is the amoe-boid cell exemplified by the gymnamoebae or ldquonakedrdquoamoebae Motile nonphotosynthetic cells lacking flagellaor cilia occur as life stages in a number of phylogeneticallydiverse taxa but for many free-living species of protozoathis form constitutes the only life stage Motility is largelyconfined to movement along surfaces by means of pseudopo-dia that can take on a variety of (species-specific) shapes Sig-nificant abundances of amoebae are largely confined tobenthic and epibiotic environments (174) and to suspendedparticulate material where they can occasionally be highlyenriched (27) Most amoebae consume bacteria and otherminute prokaryotes and eukaryotes
Several heterotrophic protistan groups possess complexamoeboid body plans most notably the foraminifera polycys-tine and phaeodarian radiolaria and the acantharia Plank-tonic forms are predominantly oceanic in their distributions(although a large number of benthic species of foraminiferaexist) These species are heterotrophic and many are visibleto the naked eye (individual cells can bege1 cm gelatinouscolonies can form ribbon-like structuresge1 m in length)They feed on a wide variety of bacterial protistan and meta-zoan prey using pseudopodial networks have rather longcomplex life cycles for individual cells (weeks to months)and are extensively used in studies of paleoclimatologicalreconstruction (82 175) In addition many of these speciespossess intracellular symbiotic usually eukaryotic algaeThe widespread occurrence of algal endosymbiosis among
422-12 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
these protozoa in oceanic pelagic environments implies thatstrong selective forces appear to give rise to these associations(97 98)
Flagellated forms of protozoa exist across many protistanlineages Eukaryotic flagella come in various sizes numbersand forms (eg with or without tiny hairs) that are character-istic of the different lineagesMany flagellate species are appa-rently purely phototrophic or heterotrophic (phagotrophic)but a significant (still poorly known) fraction of flagellates aremixotrophic combining both phototrophy and phagotrophy(70 88) Flagella in phagotrophic flagellates are employed formotility and prey capture Most free-living flagellates possessone to four flagella (typically one or two) that can be manytimes the length of the cell itself Although there is tremen-dous species diversity among flagellated protozoa many ofthese species have broadly overlapping ecologies As a groupflagellates are the most numerically abundant protozoa ofboth benthic and pelagic ecosystems and they are fundamen-tally important as consumers of bacteria cyanobacteriaand other eukaryotes Flagellated protozoa within the nano-plankton size class (2ndash20 microm) are often counted as a singleassemblage in plankton studies because morphological details
apparent using light microscope are insufficient to distinguishamong the many species This assemblage has been variouslyreferred to by a variety of names and acronyms including het-erotrophic nanoplankton (HNAN HN) heterotrophic flag-ellates (Hflags) heterotrophic nanoflagellates (HNF) andmicroflagellates Heterotrophic nanoplankton is the mostaccurate term for most methodologies employed to countthese cells because it does not require visualization of flagella(which are often lost from these small cells during preparationfor microscopy) The confusion over terminology is partly aconsequence of the different methodologies that have beenused to count these species and partly a consequence of thefact that these species were first studied prior to the wideacceptance of the size convention of Sieburth et al (84)
Heterotrophic flagellates in the microplanktonic size class(20ndash200 microm mostly dinoflagellates) are important consum-ers of phytoplankton in pelagic ecosystems Many of thesespecies are capable of the production of large pseudopodialnets the pallium that can engulf prey (particularly diatoms)significantly larger than the diameter of the dinoflagellatetheca (176) This behavior and its ecological significance forenergy flow in plankton communities have been recognized
FIGURE 5 Body plans and size ranges of protozoa These micrographs depict amoeboid (andashg) flagellated (hndashj) and ciliated (kndashm) forms ofprotozoa From Caron et al (173) Markers bars are 5 (i) 10 (h) 20 (c j l) 30 (a) 50 (k m) 100 (b f) 500 (d) and 1000 (e g) micromdoi1011289781555818821ch422f5
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-13
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
only within the past few decades (177) These species areoften abundant in waters where diatoms dominate (178179) In such situations heterotrophic dinoflagellate biomasscan be similar to that of ciliated protozoa (180)
Ciliates are generally the most recognizable form of pro-tozoa to nonspecialists Nonetheless ciliates are a diversegroup morphologically as well as ecologically (181 182)The degree of ciliature on these species can range from uni-formly ciliated to totally devoid of cilia during most of theirlife cycle to ciliature restricted to specific regions of thecell In addition cilia can fuse to form various complexstructures (eg ciliary membranes or veils cirri) that assistin locomotion food capture or attachmentCiliates are abun-dant and ecologically important species in both benthic (8)and pelagic (183) ecosystems and can consume a variety ofprokaryotic and eukaryotic prey Together with the heterotro-phic dinoflagellates these species are the dominant consum-ers of phytoplankton in many pelagic ecosystems (184) andas a consequence they form an important trophic link to met-azoan zooplankton (185 186)
Diversity and Biogeography of Protists FromMorphology to DNA SequencesUntil relatively recently the biodiversity of marine protistanassemblages was not generally considered a controversialtopic It was generally accepted that while all species of pro-tists certainly have not been identified representatives ofmost types of algae and protozoa had been observed anddescribed if not actually brought into culture in the labora-tory Direct sequencing of 18S rRNA genes from environ-mental samples (as described in ldquoMolecular Phylogeny andMetagenomics Field Applicationsrdquo) has changed that viewInitial forays into environmental DNA indicated a muchgreater diversity of protists than previously documented usingclassical approaches of culture and microscopy (81 187ndash191) Numerous publications over the past decades haveexpanded these findings (see [192] for a recent global analy-sis) which are highly analogous to discoveries in marine pro-karyote research as described already
Previously uncharacterized protistan diversity has nowbeen documented at virtually every level of eukaryoticorganismal classification Some of these findings couldhave been expected but some have been very unexpectedFor example a much greater diversity than noted previouslyhas been observed among small (lt10 microm) protists in plank-tonic ecosystems These species generally possess few distinc-tive morphological features and one could expect that manycryptic species might be present among these small morpho-types The molecular ldquodiscoveryrdquo of this eukaryotic diversityhas stimulated progress on the isolation and descriptionof new species and genera of minute algae and protozoa(193ndash197)
Analyses of environmental samples have also indicatedthe unanticipated existence of novel 18S rRNA gene sequen-ces that imply the presence of novel lineages of eukaryotes innatural protistan communities (198ndash200) These are sequen-ces for which there are apparently no known described orcultured species The degree to which these sequences dif-fer from sequences of known sequenced eukaryotes impliesthat some of these lineages may be distinct at the level ofphylum (201)
These findings have raised basic questions and somedebate concerning the true diversity and biogeography ofprotistan assemblages in natural ecosystems (202ndash204) thevalidity of the many novel phylotypes or cryptic species that
are being documented (205) the potential importance andsignificance of the many rare taxa that characterize thesecommunities (206) and the implications of these findingsfor the ecologicalbiogeochemical roles that protists playin aquatic ecosystems For example it was postulated andsubsequently confirmed that some novel alveolate lineagesreported from marine ecosystems make up a suite of parasiticprotozoa whose ecological importance may have been signifi-cantly underestimated in the past (207) Deciphering theidentity and significance of these many unknown phylotypeswill constitute a significant effort for protistologists in thefuture (80 85)
Life Histories and Ecological Strategies of ProtozoaAbundance prey type and life histories all vary tremendouslyamongmarine protozoa The smallest species (eg many flag-ellates) tend to be the most abundant and widely distributedin the world ocean Indeed many of these species may beglobally distributed (208) Most nanoplanktonic flagellateshave potentially rapid rates of grazing and growth Underoptimal conditions these species can divide by binary fissionseveral times a day and thus dramatic increases in their pop-ulations can take place in response to favorable conditionswithin a few days (209) Many of these species are capableof surviving for limited periods of time without food andhave developed a variety of physiological or life cycle strat-egies to cope with these events (209) However in contrastto some bacteria that may remain viable through long periodsof starvation (210) protozoa will expire or encyst in responseto low food abundance and thus ldquoboom-and-bustrdquo popula-tion changes are characteristic of small flagellates
On the other end of the size spectrum from the ratherubiquitous nanoflagellate species many of the largest proto-zoan species (polycystine and phaeodarian radiolaria plank-tonic foraminifera) are exclusively oceanic (ie do notsurvive in most coastal environments) andor have specificlatitudinal and depth distributions Thus the abundancesof these latter species may range from undetectable to max-imal abundances of gt105 individualsm3 Dramatic changesin the abundance of these protozoan taxa also can be relatedto changes in prey abundance physicalbehavioral aggrega-tion or to periodicity of life cycle events For example theplanktonic foraminifer Hastigerina pelagica reproduces on alunar cycle and thus abundances (and life stages) of this spe-cies in oceanic waters can vary considerably over the courseof a month (211) In general life cycles for the large amoe-boid protozoa are lengthy and complex (for single-celledorganisms) with life spans unknown for many species(attempts to culture them in the lab have so far been unsuc-cessful) but estimated to be on the order of months to per-haps years (175)
The abundance and activities of microplanktonic hetero-trophic protists (mostly ciliates and heterotrophic dinoflagel-lates) tend to be somewhat intermediate to those ofnanoplanktonic flagellates and the larger amoeboid formsThese species are present in the majority of marine ecosys-tems and collectively play an important role in the controlof phytoplankton biomass (and probably the abundance ofnanoplanktonic protozoa although there is little informa-tion on this topic) in waters throughout the world oceanLike small flagellates ciliates and dinoflagellates reproduceprimarily by binary fission but their maximal growth ratesare typically slower (one division a day is typical) and theirlife cycles often include sexual phases that allow for geneticrecombination
422-14 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
VIRUSESViral Abundance and General PropertiesViruses are simple biological agents typically 20ndash200 nm indiameter composed of a nucleic acid genome in a proteincoat that infect cells and ldquocommandeerrdquo the cellrsquos machineryto make more viruses which are released into the environ-ment when the host cell lyses or bursts A particular virus isthought to be capable of infecting only a narrow range of hosts(usually one species sometimes a genus rarely broader)Although some early studies had isolated viruses from thesea therewere no data prior to the 1980s showing such viruseswere very abundant and more significantly no evidence thatinfection was occurring in any important part of the planktoncommunity It wasnrsquot until the late 1980s that electronmicro-scopy with suitable concentration methods showed thatviruses are extremely abundant similar to or even higherthan bacterial abundance (212 213) Interestingly themost suitable transmission electron microscopy (TEM)approach used by Bergh et al (described in detail by [214])was actually similar to a direct centrifugation method devel-oped in 1949 (215)
TEM studies of viruses permit high-resolution images andobservation of viral morphology showing features such ashead diameter tails sheaths and tail fibers (Fig 6 collage
in upper left) Each type of virus has a fixed morphology(unlike bacteria that are potentially more plastic in theirappearance) and hence a coarse measure of viral diversityhas been possible by cataloging morphologies Studies thathave done so have found dozens or more different morpholo-gies of marine viruses in a given sample (217ndash219) A largeproportion resemble bacteriophages which are viruses thatinfect bacteria
The availability of brightly fluorescent nucleic acid stainsand high-porosity fine pore size (002 μm) filters made fromAl2O3 have made it easy to accurately count viruses by epi-fluorescence microscopy (38 220 221) Epifluorescence per-mits abundance estimates but does not allow observation ofviral morphology as viruses are below the resolution limitof light microscopy viruses are visible only as sources of light(like stars in the night sky see Fig 6 lower micrograph) Anextension of manual epifluorescence counts is the use of flowcytometry to count viruses now commonly employed in somelabs (222)
Virus abundance has been found to be closely related tobacterial abundance with a virusbacteria ratio typically10ndash301 A typical oceanic profile of bacterial and viral abun-dance is shown in (Fig 6 panel on right) This tight ratioand the strong correlation to bacterial abundance in relationto weaker correlations to chlorophyll have been cited as
FIGURE 6 Marine viruses (upper left collage) viewed by transmission electron microscopy These are cyanophages in the groups myoviridae(right and bottom) siphoviridae (left) and podoviridae (top) Scale bars represent 100 nm From Sullivan (216) Epifluorescence micrograph(lower left) of SYBR green I stained viruses (small fluorescing objects) and bacteria + archaea (large fluorescing objects) Depth distribution ofvirus and bacteria + archaea abundances obtained by epifluorescence microscopy of SYBRGreen stained cells in the central San Pedro BasinCalifornia (11 August 2000) doi1011289781555818821ch422f6
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-15
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
evidence that most marine viruses infect bacteria rather thaneukaryotic phytoplankton (217 218 223 224) though thereare also many important viruses of phytoplankton and otherorganisms with significant impacts on primary productivityblooms and ecosystem function (225ndash227)
The first demonstration of the activity of marine viruseswas presented by Proctor and Fuhrman (228) who showedby electron microscopic examination that viruses wereactually infecting marine bacteria and cyanobacteria at ameasurable rate Because only the final portion of the viruslife cycle is visible by electron microscopy (when the virusesare assembled and ready to lyse the host cell) only a very smallfraction of the infected cells can be counted as infected at anygiven time Proctor and Fuhrman (228) reported that only afew percent of the total bacteria from coastal waters and theSargasso Sea were visibly infected but they interpreted thedata with a model that implied the actual fraction of the totalcommunity that was infected was much higher They con-cluded that the total fraction of bacterial mortality attribut-able to viruses is roughly 10ndash40 Subsequent studies haveused refined versions of that same general approach andmodel (229 230) and numerous studies have used a varietyof alternative approaches to estimate viral activitymdashall yield-ing basically the same general conclusion (although themodel parameters need to be adjusted for cyanobacteriawhich on average seem to be infected less than bacteria)These approaches include (a) calculation of virus turnoverfrom decay estimates (231) (b) measurement of viral DNAsynthesis by incorporation of tritiated thymidine (232) (c)observation of effects of added viruses (233 234) (d) estima-tion of bacterial mortality in the absence of protists (e) use offluorescent viruses to estimate production by an approachanalogous to isotope dilution (38) and (f) dilution of virusesin filtered samples to estimate viral production (235) Theoverall consensus of these studies is that the initial estimateswere basically correct that is that viruses are responsiblefor about 10ndash40 of the bacterial mortality in most marinesystems (reviewed by [218 223 226 236 237]) Howeverthere is still some disagreement about the higher estimatesand it is likely that 40 mortality from viruses alone is nottypical for most marine systems
Most of the foregoing work has focused on the viral infec-tion of bacteria primarily thought to be heterotrophicalthough the original report by Proctor and Fuhrman (228)also noted the occurrence of cyanobacterial infection Sev-eral subsequent studies focused on phytoplankton includingthe potential effect of viruses on the termination of phyto-plankton blooms of Emiliania and Phaeocystis and the likelyimpact this might have on release of climate-active gasessuch as dimethyl sulfide (225 227 238ndash247) Althoughdetails are beyond the scope of this chapter viruses arethought to infect virtually all marine organisms with poten-tially significant impacts from zooplankton to whales (226)Detailed studies of viruses infecting cyanobacteria such asSynechococcus have shown differences in viral host specific-ity particularly toward coastal and oceanic host strainsand some occasional high virus abundances (to 105ml) asmeasured by most probable number (MPN) cultivation tech-niques in the Gulf of Mexico near Texas (248 249) Othervirus cultivation studies with Prochlorococcus and Synecho-coccus in oligotrophic waters of the Sargasso Sea showedan interesting pattern of cross-infection between these gen-era by some virus types (suggesting gene flow among theseorganisms) but generally low MPN estimates of abundanceto 103ml even when cyanobacterial abundance was near105ml (216 250)
Overall the consensus emerging from direct comparisonsof viral-mediated mortality and grazer-mediated (ie micro-zooplankton) mortality have indicated that viral lysis of bac-teria constitutes a significant fraction of total mortality ofthis assemblage while the mortality of phytoplanktonappears to be dominated bymicrozooplankton inmost instan-ces (251 252)
As mentioned viruses have significant morphologicaldiversity as observed in TEM studies It is also possible toinvestigate aspects of their genetic diversity In early workon this topic Wommack et al (253) and Steward et al(254) observed the diversity of viral genome lengths in a fieldsample by pulsed field gel electrophoresis with viral genomesranging 25 kb to gt300 kb in length Field results show thatthe viral community composition is dynamic in space andtime with clear changes in the banding patterns over seasonsand locations in Chesapeake Bay (253) between oceanbasins and subsequent to dinoflagellate blooms (254) andwith depths to 500 m at one location (255)
Although viruses do not all share a set of core genesthat allows a universal viral phylogeny (comparable to SSUrRNA gene in cellular organisms) genetic diversity amonga single group of viruses can be examined by sequence analysisof shared genes within the group An example is the g20 geneinT4-like cyanophages that has shown extremely high diver-sity even among very closely related viruses and has demon-strated geographic and seasonal variation (256) A secondexample is g23 which is found broadly in diverse and wide-spread T4-like phages (257) and can have seasonally repeat-ing patterns (258) as well as short-term rapid dynamics thatcorrelate to those in bacteria (259)
Viral MetagenomicsAs with cellular organisms metagenomics can be used toexamine viral diversity and genetics without themany restric-tions of cultivation Viruses can be collected by selective fil-tration and concentrated by tangential flow filtration orflocculated with iron chloride (260 261) then their collec-tive metagenome can be extracted linker amplified and ana-lyzed by sequencing (262 263) Viral metagenomic studiesare particularly challenging because the large majority ofsequences have no annotated matches in any databases butso far results from marine samples around the world haveshown extremely high diversity and variations with depthlocation and time presumably with highly dispersed typesselected by local conditions (262ndash266) Because metage-nomes are best interpreted when there are representative cul-tures available the best matches of viral metagenomesoriginally tended to be to the few viral isolates infecting trulycommonmarine bacteria like cyanophage infecting Synecho-coccus and Prochlorococcus (267) However the developmentof dilution-to-extinction cultures representing common het-erotrophic marine taxa like Peligibacter (a member of theSAR11 clade) and SAR116 has allowed isolation from sea-water of viruses infecting these common organisms and theseisolates have indeed been found to be highly abundant inmarine viral metagenomes (268 269) New approaches tointerpret the results include clustering the proteins independ-ent of known proteins to compare samples to each other andtry to find environmental factors driving viral communitychanges (270) Such analysis of a large global data set (TaraOceans expedition) has shown that extensive sampling hascome close to reaching the total diversity in tropical and tem-perate waters of such viral protein clusters (which essentiallyrepresent various viral protein motifs not all viral proteintypes) and that viruses appear to be directionally dispersed
422-16 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquodownstreamrdquo in major ocean currents as one may expectfrom first principles but also supporting the ldquoseed bankrdquohypothesis of viral biogeography (271)
Viruses and Host DiversityViruses are themselves thought to be instrumental in drivingincreased diversity of their microbial hosts via a hypothesizedprocess often called ldquokill the winnerrdquo This is because viralinfection is host-specific and density-dependent the latterbecause viruses diffuse from host to host so an abundanthost is more likely to pass on infection than a rare oneThis means that if an organism becomes abundant andblooms winning the competition for resources it becomesmore susceptible to a viral epidemic This would benefit therarer organisms and thus help foster diversity (223 272273) However bloom scenarios as described above involvesystems far from steady state and the formal theory of Thing-stad and Lignell (274) has interesting steady-state solutionswhere several viruses infect several hosts stably over time(via trade-offs between growth rates and viral susceptibility)which may occur at the strain or species level (274) There issome experimental evidence that viruses have effects on nat-ural marine microbial community composition weaker thanthe kill the winner (bloom version) hypothesis would sug-gest (275 276) so something resembling the steady-statecoexistence described in the model may in fact be commonThere also appear to be processes that foster coexistencebetween viruses and hosts but the mechanisms are largelyspeculative (223 277 278)
Viruses may also be directly involved in host genetic diver-sity because they can be the agents of genetic exchangebetween microorganisms (223 273) This often involvesthe viral lifestyle known as lysogeny whereby viruses survivewithin host cells as DNA only integrated into the host chro-mosome and being reproduced each time the host divides Ahost harboring such a genome is called a lysogen becauseunder conditions of stress to the host cell a genetic switchmay cause the viral genome to initiate the lytic process pro-ducing many progeny viruses and bursting from the hostLysogeny is a very common property occurring in a signifi-cant part of the bacterial community (279 280) althoughthe incidence of induction of the lytic phase in nature is appa-rently low (281) Overall lysogeny is poorly understood butthought to have both positive and negative impacts on themicrobial community (282) Recent results suggest that ben-efits of being lysogenic in highly seasonal polar seas leads tofundamental difference between polar and other marine viralcommunities (283)
Viruses and the Microbial LoopAs part of the food web viruses occupy a unique positionThey infect host cells that are mostly thought to be heterotro-phic bacteria and by doing so they typically burst the hosts torelease progeny viruses and cellular debris But what is the fateof this material Viruses themselves do not last indefinitelyand a simple steady-state assumption implies that from eachburst of viruses (typically 20ndash100 per lytic event) only onesuccessfully infects another cell The rest are inactivatedand broken down by sunlight (UV and visible exposure)and enzymatic attack (284) or consumed byminute phagotro-phic protists (285) thus reentering the food web as substratefor bacteria or food for protistan consumers Experiments incontrolled laboratory systems and field studies with radioac-tively labeled viral lysis products have supported the conclu-sion that most of the organic matter released by the viralinfection is either taken up by bacteria or respired (286 287)
Modeling this process as part of the microbial loop showsthat viral lysis represent a sort of side loop that has the neteffect of remineralizing a significant amount of the carbonand nutrients that enter the bacteria-protist part of themicro-bial loop (Fig 7) A theoretical numerical steady-state modelcomparing a systemwith no viral activity to onewhere virusesare responsible for 50 of bacterial mortality showed that thesystem with viruses had 33 more bacterial production andrespiration than the virus-free system implying that theviruses had the effect of permitting the bacteria to processmore of the primary production than they would otherwise(223) Although 50 is a high number unlikely to be com-mon in the sea this model nevertheless illustrates that virusescan reduce the amount of energy reaching higher trophic lev-els The implication is that viruses lead to increased bacterialactivity at the expense of the larger organisms
MAJOR ENVIRONMENTAL CONTROLSLight Temperature and PressureTemperature has an important potential influence on bio-chemical reactions and therefore on biological processesin general Most ocean waters fall in the range of ndash2degC to30degC with obvious exceptions in hydrothermally heatedareas Temperature has long been known to be a regulatingfactor for the growth of heterotrophic microbes
In temperate waters it has been established that microbialactivity is generally much higher in warm summer waters thanin winter (55) The relationship is not simple howeverbecause multiple factors act at the same time Some contro-versy still exists regarding the highest and lowest extremesfor marine bacterial growth although there is broad agree-ment that bacteria grow gt100degC at hydrothermal vents andltminus5degC in sea ice brines Pomeroy et al (57) noted the inter-esting observation that bacteria seem particularly inhibitednear the freezing point of seawater (ca ndash22degC) comparedto eukaryotic phytoplankton This effect results in polarspring phytoplankton blooms that accumulate organic car-bon in advance of the response of the bacterial communityand development of the microbial loop and perhaps lead toenhanced benthic-pelagic coupling (288)
The relationship between temperature and the growth rateof marine phytoplankton was described broadly in a now
FIGURE 7 Modification of the microbial loop concept that incor-porates the functional role of viruses Export can be via predation orsinking From Fuhrman (223)doi1011289781555818821ch422f7
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-17
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
classic paper by Eppley (58) and later Goldman and Carpen-ter (289) Temperature was shown to exert a strong and directeffect on themaximal growth rates of these species withmax-imal intrinsic growth rates at 0degC generally lt1 divisiondaywhile growth rates at 30degC may be gt4day These relation-ships indicate the maximal rate that might be attained byphytoplankton at these temperatures but they do not takeother factors into account (nutrients light) While warmtemperature ostensibly allows more rapid growth it createshydrographic conditions that typically give rise to nutrientlimitation of algal growth Thus some of the coldest watersin nature witness some of the most massive phytoplanktonblooms (290) while warm oceanic gyres represent some ofthe most oligotrophic areas of the ocean The multiple andoften conflicting effects of temperature on the growth of nat-ural phytoplankton assemblages limit the accuracy of presentpredictions regarding how primary producers will respond toclimate change (291)
An analysis of the effect of temperature on the growth ofheterotrophic protists has indicated that the response is qual-itatively similar to that of phytoplankton but with a twistThe growth rates of at least some protozoa can exceed thoseof phytoplankton at warmer environmental temperaturesbut the opposite effect is apparent at very low environmentaltemperature This differential effect of temperature on thegrowth of phototrophic and heterotrophic protists was basedon a large meta-analysis of published protistan growth (59)That analysis demonstrated that the maximal growth ratesattained by phototrophic protists could exceed the maximalgrowth rates attained by heterotrophic protists (all otherpotential growth-limiting factors not considered) Thereforeprotozoan growth rates may be constrained to a greater degreeat low environmental temperature than rates for phytoplank-ton If so then phytoplankton bloomsmay get a head start ongrazers during spring in polar ecosystems This scenario is con-sistent with information on seasonal biomass changes andmicrozooplankton herbivory in the Ross Sea Antarctica(292 293) but there are still too few data to fully vet thishypothesis
The importance of high pressure on bacterial growthgained considerable attention in the late 1960s when thedeep sea submersible Alvin was accidentally lost overboardwith its hatch openWhile no lives were lost in this accidentsome workmenrsquos lunches sank to the bottom (sim1500 m)inside the submersible Alvin was recovered after 10 monthsand interestingly there was a waterlogged lunch containingapples bologna sandwiches and broken vacuum bottleswith broth that all appeared hardly degraded and tasted palat-able Yet when placed in a refrigerator on the ship these itemsdegraded relatively quickly Initially pressure was thought tobe the preserving factor as the sea floor temperature was sim-ilar to the refrigerator temperature and the only major differ-ence would be pressure (294) Following this observationa series of experiments to measure degradation of variousorganic materials left in the deep sea for extended periodsindicated that degradationwas typically significantly reducedimplying that pressure reduces the degradation rates (295)Nonetheless changes in the protozoan community of naturaldetrital material sinking to the deep ocean floor indicate thatthe microbial community can respond relatively quickly insome situations (296) Deep sea microbiology has advancedconsiderably yet it is still difficult to interpret results withrespect to actual in situ rates of naturally occurring organicmatter Deep sea bacteria adapted to high pressures havebeen isolated that are barophilic (also called piezophilic)meaning that they prefer high pressures and have reduced
activity at lower pressures whereas others are barotolerant(piezotolerant) tolerating but not preferring high pressuresfor example see (297 298)
Few data are available on barotolerantbarophilic marineprotozoa Protozoa certainly exist and grow at great oceanicdepths but measurements of in situ growth rates for these spe-cies do not yet exist Measurable protozoan numbers havebeen documented in the deep-sea sediments for more than30 years (299 300) and viable protozoa have occasionallybeen cultured from these environments (301ndash305) butvery few direct measurements of the activities of these speciesin situ have been reported (306) A few protozoa have beenisolated that will grow at high pressure (302 303 307) andprotists that appear to be unique to the deep ocean havebeen observed either directly or through the analysis ofDNA sequences (308ndash311) but possibly the best direct evi-dence that protozoan activity takes place at the high pressurescharacteristics of the deep sea are experimental and observa-tional work noting the stimulatory effect that detrital deposi-tion has on some components of the protozoan community(296 312) These observations indicate a diverse and activeprotozoan fauna of the deep ocean although their biogeo-chemical significance is largely uncharacterized at this time
Dissolved and Particulate Organic MatterBacteria and archaea are thought to be by far the most impor-tant organisms with respect to the processing of dissolvedorganic matter (DOM) and nonliving particulate organicmatter (POM also called detritus) in the ocean While theremay be some uptake of DOM by protists particularly forgrowth factors needed in trace amounts such as vitamins(313) the bulk of this material is probably utilized by bacteriaand archaea (314) including the smallest cyanobacteriumProchlorococcus (315 316) which thus may be considered amixotroph Due to their small size bacteria have extremelyhigh surfacevolume ratios and combined with their over-whelmingly high abundance an extremely high integratedsurface area Protozoa tend to obtain the organic materialsthat they require for growth from their prey rather thanthrough the uptake of DOM Overall protozoa tend to besources of dissolved and detrital organic substances throughthe excretion of unassimilated prey biomass in expelledfood vacuoles
Particulate organic matter is not directly available as sub-strate to bacteria These substances must first be reduced tosmall molecules that can be transported into the cell Thisis accomplished by the production of extracellular enzymes(note that few if any large polymers are directly taken up bybacteria with the possible exception of DNA) Hydrolyticenzymes produced by bacteria (and almost certainly archaea)break down polymers like proteins polysaccharides andnucleic acids As with DOM POM is composed of a complexmixture of compounds that vary in their susceptibility to bac-terial degradation and utilization
Particulate material in the water column serves not only asbacterial substrate but also as substratum POM occurs in thewater column across a huge size spectrum from micrometersup to some detrital aggregates more than 1 m in diameter(317) Much of this particulate material is in a constant stateof flux with colloidal material constantly coalescing andaggregating to form new or larger particles (318) as microbialdegradation acts simultaneously to remineralize this materialDetrital particles that attain macroscopic size either by directformation (317 319 320) or via accretion and aggregation(321) and are often called marine snow or macroaggregatesMarine snow particles are readily colonized by bacteria and
422-18 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
many other microorganisms and their abundances on theseaggregates can be orders of magnitude greater than abundan-ces in an equivalent volume of the water surrounding theaggregates (320 322ndash324) and their diversity and highlyconcentrated abundances on detrital aggregates implies thatthere may be stronger trophic coupling and efficient energyelemental cycling relative to these processes among free-living microorganisms in the surrounding waters This specu-lation has resulted in a number of formalizations of the poten-tial impact of aggregate microbiology on biogeochemicalprocesses in the ocean (325 326) Most definitions of bio-films include aggregated particulate and these particles areoften studied within the biofilm conceptual model (see chap-ter by Lawrence et al in this section)
Inorganic Nutrients
Macronutrients (N P)Virtually all heterotrophic organisms in the ocean contributeto the pool of available macronutrients via the excretion ofmetabolic wastes and protozoa contribute significantly tothis process Macronutrients are often in excess within proto-zoan prey relative to the consumersrsquo needs because much ofthe prey carbon is respired to produce energy for metabolismand growth ldquoExcessrdquo nutrients are eliminated as eitherorganic compounds or more commonly as ammonium (forN) and phosphate (for P) The importance of the match(or mismatch) between predator and prey stoichiometry hasbeen demonstrated experimentally and repeatedly in marineand freshwater ecosystems and has been elevated to a funda-mental ecological principle governing elemental and energyflow in aquatic food webs (327) Based on this reasoningCaron et al (328) concluded that bacterivorous protozoaplay an important role in nutrient remineralization andrelease in the ocean Bacterial biomass is typically rich in Nand P relative to other organisms so bacterial predators expe-rience large excesses of these elements relative to their growthneedsMoreover bacterivorous protozoa should be dispropor-tionately important in nutrient remineralization amongsingle-celled eukaryotes because as the smallest protozoa inaquatic ecosystems they have high weight-specific metabolicrates (329) Thus both stoichiometric and allometric rela-tionships implicate small bacterivorous protozoa as impor-tant sources of remineralized nutrients in the plankton
The reasoning also holds true for bacterial utilization oforganic compounds and it has led to some interestingfindings and conclusions regarding the role of bacteria innutrient cycling in the ocean Bacteria are important sourcesremineralized nutrients when they are consuming N- orP-rich substrate Contrary to this traditional view howeverheterotrophic bacteria are often strong competitors for inor-ganic nutrients under many situations largely because bacte-rial biomass is richer in N and P than much of the organicmatter they consume in nature so they require additional Nand P to produce their biomass (summarized by [330]) Exper-imental studies have demonstrated the uptake of ammoniumby bacteria other than cyanobacteria using stable isotope 15N(331) and also the short-lived radioisotope 13N (332) oftenfinding that bacteria may be responsible for one third of the Nor P uptake Additionally there is evidence that bacteria insome oceans like the Sargasso Sea are limited by phosphorus(333 334) Growth limitation of bacteria by phosphorusappears to be a common phenomenon of freshwater ecosys-tems (335 336)
N2 fixation is restricted to prokaryotes and in the oceanwater column it was thought for many years to be done
primarily byTrichodesmium a warm-water colonial cyanobac-terium that blooms sporadically andRichelia a cyanobacterialsymbiont that lives within certain diatoms (337) Some uni-cellular cyanobacteria also contribute significantly to globalN2 fixation (338) and molecular biological data suggestthat a variety of other bacteriamdashnot phytoplanktonmdashmayalso be fixing nitrogen in seawater (339) An exciting recentdiscovery in this field is that a bacterial nitrogen fixer origi-nally known only from nifH sequences UCYN-A has hadits genome sequenced by parallel pyrosequencing and wasfound be a cyanobacterium (Candidatus Atelocyanobacteriumthalassa) lacking the apparatus to generate oxygen as well asother pathways (340) Further study showed it to be symbioticwith a marine alga (341)
Micronutrients (Trace Metals Growth Factors)Bacterial growth may often be limited by macronutrients asdescribed and sometimes by trace nutrients particularlyiron that occur at very low concentrations in seawater(342 343) The importance of Fe as an element limiting pri-mary production in some oceanic regions has come underclose scrutiny and extensive experimental investigation inrecent years Many bacteria have special uptake mechanismsto utilize extremely low concentrations of iron includingsiderophores (released compounds that bind iron and arethen specifically taken up) Even if bacteria are capable ofgrowth at low Fe concentrations growth may be less efficientKirchman et al (344) found that Fe limitation stronglyreduced bacterial growth efficiency in a cultured marine bac-terium (ca 50 efficiency Fe replete but lt10 when Fedeplete) Thus the effects may be complex Marine N2 fixa-tion also may be limited by the availability of Fe becausenitrogen fixers have high Fe requirements
Protozoa tend to be a source rather than a sink for Fe in theocean analogous to their role in macronutrient remineraliza-tion Protozoan grazing activity has been shown to releaseiron from bacterial biomass thereby relieving Fe limitationin co-occurring phytoplankton (345) a mechanistic demon-stration of the process using protozoa growing on Fe-repletebacteria However it has also been reported that Fe-limitedbacteria may contain insufficient Fe for the bacterivorous pro-tozoa that consume them thus leading to Fe limitation in theprotozoan as well (346)
OxygenHypoxic and anoxic regions of the ocean are expected to sig-nificantly expand in the coming decades as a consequence ofchanges in ocean stratification resulting from global climatechange (347) As oxygen becomes depleted bacteria havethe capability to utilize a series of alternate electron acceptorsCompositional changes in the bacterial assemblage accom-pany these changes in metabolic activity but are beyondthe scope of this chapter Many of the studies on benthicmicrobiology have focused on redox reactions that occurthere Oxygen is usually consumed by sediment microbialactivity much faster than the rate at which it can be replacedby diffusion so most sediments especially organically richones are anaerobic below a few mm (fine-grained muds) orcm (coarser sediments) from the sedimentwater interfaceexcept where animals ventilate the benthos through tubesWhen oxygen is absent bacteria and archaea use alternateelectron acceptors such as nitrate nitrite oxidized Fe orMn organic matter or sulfate The use of nitrate or nitriteas an electron acceptor with concomitant production of N2
gas (denitrification) results in net loss of biologically availablenitrogen Bacteria that perform this reaction are typically
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-19
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquofacultativerdquo organisms that can switch from oxygen to nitrateor nitrite Organic matter may be reduced by fermentationreactions in which part of an organic substrate is oxidizedwhile another is reduced fermenter organisms are also typi-cally facultative Sulfate is a particularly important alternateelectron acceptor in marine systems because it occurs as thesecond most abundant anion in seawater (behind chloride)typically 28 mM Its use as an electron acceptor leads to theproduction of elemental sulfur or sulfide The use of sulfateas an electron acceptor yields much less energy than oxygenor nitrate and sulfate-reducing bacteria are strict anaerobesunable to use oxygen (often strongly inhibited by oxygen)Methane can be produced by methanogenic archaea whichare all strict anaerobes using CO2 as an electron acceptor(see Findlay and Battin this section for further discussion)
Reduced compounds such as sulfide are produced by theuse of these alternate electron acceptors and they diffuse awayfrom their source When they diffuse to environments with astrong oxidant like oxygen they may be oxidized by chemo-trophic bacteria in energy-yielding reactions Similarlyammonium can be oxidized to nitrite and then nitrate by che-motrophic nitrifying bacteria and the nitrate can diffuse intoanaerobic layers and become denitrified to N2 Such coupledoxidation-reduction reactions can lead to accelerated biogeo-chemical cycling particularly in sediments
It was originally believed that relatively few free-livingspecies of protozoa inhabit anaerobic and hypoxic marineenvironments This view has changed based on (largely)molecular analyses of some of these environments (308348ndash351) A significant number of previously undescribedeukaryotic phylotypes have been observed in anoxic environ-ments High protozoan biodiversity in the absence of molec-ular oxygen is not completely unexpected because otheranaerobic environments such as some gut environments(eg cockroach guts and cattle rumen) have notoriouslyabundant and diverse protozoan fauna but marine anaerobicenvironments have been difficult to access and sample prop-erly Many anaerobic protozoa exist together with symbioticbacteria (352 353) or possess unique organelles such as thehydrogenosome (354)
MAJOR BIOTIC CONTROLSThe Trophic Activities of Protozoa Predation andTop-Down ControlsPredator-prey relationships are fundamental interactionswithin biological communities linking production of bio-mass on the one hand with removal on the other hand Estab-lishing the trophic relationships among microorganisms andquantifying the rates at which they take place constitutemajor endeavors within marine microbial ecology As notedpreviously protozoa consume a wide variety of prokaryoticand protistan prey Numerous laboratory and field measure-ments have been made throughout the past quarter centuryto attempt to quantify the rates of bacterial and phytoplank-ton consumption The methodology for accomplishing thesestudies has varied and a number of approaches are presentlyemployed
Methodologies for estimating microbial grazing are diversebut generally fall into one of two experimental approachesperturbation experiments or tracer experiments Perturbationexperiments (size fractionation metabolic inhibitors anddilution technique) rely on manipulation of the consumeror prey assemblages in somemanner such that the effect is evi-dent from changes in the abundance of the prey population
during incubations relative to control treatments that receiveno manipulation Tracer experiments (radioactively labeledprey fluorescently labeled prey stable isotopendashlabeled pray)attempt to use labeled prey or a prey proxy to quantitativelyfollow the movement of prey biomass into consumers Forsummaries of these methods see (74 355)
Perturbation experiments have entailed a variety of mech-anisms to ldquodecouplerdquo predators from prey in natural microbialcommunities For example size fractionation uses various sizefilters to remove an entire size class of grazers from awater sam-ple leaving the prey to grow in the relative absence of thosepredators during subsequent incubations Unfiltered samples(controls) indicate the growth of prey in the presence of pre-dation Predation impact is obtained by the difference ingrowth of the prey between these treatments This methodhas been used to examine predation on cyanobacteria heter-otrophic bacteria and minute phototrophic eukaryotesAlternatively metabolic inhibitors have been employed tohalt predation (or in some cases prokaryote growth) insteadof physical removal by filtration
The dilution technique (356) has become a commonmethod for examining grazing mortality of phytoplanktonby microzooplankton (which is comprised predominantly ofprotozoa) This method relies on the dilution of herbivorouszooplankton in a series of subsamples (and concomitantmeasurements of phytoplankton growth in each subsample)to estimate the grazing mortality of the herbivores
Tracer experiments have used both radioactively labeledprey and fluorescently labeled prey to examine predation onbacteria and phytoplankton Radioactive labeling has beenaccomplished using 14C- or 3H-labeled organic compounds(for bacteria) or 14C-bicarbonate (for phytoplankton) Theuse of radioisotope approaches has diminished with improve-ments in the number and types of fluorescent compoundsavailable for this work Fluorescently labeled prey havebeen labeled with a variety of compounds that allow theprey to be observed readily (even inside predators) by epi-fluorescencemicroscopy (357 358) Alternatively a decreasein the total number of fluorescent prey in a sample indicatesthe rate of removal of the prey assemblage
Both perturbation experiments and tracer experimentsdescribed have inherent advantages and disadvantages Forexample perturbation experiments are fairly straightforwardto conduct but they may introduce artifact by removingpotentially important sources of nutrients or organic com-pounds released by predators (359) On the other hand thedilution technique is very labor-intensive but one advantageof this method is that it provides the opportunity to determineboth phytoplankton growth and grazing mortality in a singleprocedure Most tracer experiments are fairly easy to conductand generally do not perturb the food web being studied butfeeding selectivity for or against the tracer (relative to naturalprey) may produce spurious results In addition some of thesemethods provide a measurement of the activity of the entiregrazer assemblage and others are applicable for examiningthe activity of specific taxa For example size fractionationapproaches examine the effect of the removal of an entiresize class of organisms (a community-levelrdquo measurement)whereas fluorescently labeled prey can be used to examinethe ingestion rate of a particular species by observing theuptake of labeled prey into that species (a species-levelmeasurement)
Predation rates of numerous marine protozoa have beenexamined in the laboratory From these experiments a gen-eral knowledge of the rates of consumption by these specieshas been accumulated Feeding in most protozoa shows a
422-20 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
functional response to the availability of prey that is similar tothe response of many organisms (although there are a greatdeal of species-specific differences) That is ingestion rateincreases rapidly as prey abundance increases until a maximalingestion rate is attained (360)Manyof these curves have theform of a Type II functional response curve (361)
Based on the features of these curves it is clear that proto-zoa have the ability to ingest substantial numbers of prokar-yote and eukaryote prey in natural ecosystems Somebacterivorous flagellates can consume bacteria (and cyano-bacteria) at rates up to 100 s per individual per hour whereassome bacterivorous ciliates can consume thousands of bacte-ria per individual per hour Rates of consumption of eukary-otic phytoplankton by herbivorous ciliates and largeflagellates are considerably more variable and highly depend-ent of the feeding mechanism (eg filtration in ciliates versuspseudopodial net in dinoflagellates) and the size of the prey
These lab studies demonstrate that rarely are prey suffi-ciently abundant in nature to support the maximal ingestionrates of which many protozoa are capable Nevertheless pre-dation rates of protozoa in nature are sufficient to constitute amajor factor controlling the standing stocks of bacteria andphytoplankton (63 180 184 355) For example these sum-maries indicate that on average protozoa consume 20ndash80ofthe standing stock of phytoplankton (measured as chloro-phyll) in the world ocean and 60ndash75 of daily primary pro-duction a departure from the classical view of copepods as theprimary consumers of phytoplankton This large role of her-bivorous protozoa infers a large role for copepods as consumersof planktonic protozoa (185) Similarly the role of protozoaas major consumers of bacterial biomass is indicated by therelative balance between bacterial production on the onehand and bacterivory on the other (Fig 8) The approximateone-to-one balance between these measurements over a largenumber of studies implies that protozoan bacterivory must bea major factor determining the fate of bacteria in marine (andfreshwater) plankton ecosystems
It is interesting to note that some types of bacteria includ-ing marine Bdellovibrio are known to prey on other marine
bacteria (362) At the time of this writing the importanceof bacterial predation by other bacteria within the contextof oceanographic processes is not known and presumed tobe small While Bdellovibrio may be cultivated from seawaterit can also be found among sequences from cold sediments(363) and the water column Bdellovibrio happens to be thebest studied predatory bacterium but there are no doubtothers that we simply have not yet identified (364)
Species-Specific Interactions (MutualismParasitism Commensalisms)Interactions between marine microorganisms such as compe-tition allelopathy and symbiosis are fundamental aspectscontrolling community composition and activity but atpresent most are poorly characterized in natural marine sys-tems Bacteria must compete for dissolved and particulatecompounds with other bacteria andwith protistan taxa whilephagotrophic protists must compete with each other andmetazoa for available prey These interactions manifest them-selves in many forms including true competition for availableresources resource partitioning such that some taxa specializeon the utilization of certain substrates or prey while othersspecialize on different ones (reducing competition) or eventhe cooperative use of compounds (eg one bacteriumrsquoswaste products being utilized as substrate by another bacte-rium) These metabolic consortia are probably common inthe ocean but can be difficult to identify and characterizeConsortia of bacteria working together to perform complexreactions especially in anaerobic systems are well known inmicrobiology
Consortia in which there is a very high degree of integra-tion between species (ie symbiosis) are very common in theocean These associations may be beneficial to both partners(mutualism) beneficial to one part and inconsequential forthe other (commensalism) or beneficial to one part and det-rimental to the other (parasitism) Many highly visible andwell-characterized marine mutualisms exist between chemo-synthetic bacteria and animals such as hydrothermal ventor methane seep tubeworms clams mussels or other inverte-brates (365) and also between luminescent bacteria and fishor squids (366) Similarly many protozoa form mutualisticassociations with photosynthetic protists (367) The mostconspicuous of these associations involved large amoeboidprotozoa of oceanic ecosystems (planktonic foraminiferanspolycystine radiolaria and acantharia) which contain largenumbers of intracellular algae (74) but associations betweensome heterotrophic dinoflagellates and cyanobacteria are alsocommon (368) Studies have indicated that efficient nutrientcycling in these protozoan-algal associations confers an eco-logical advantage for both partners in surfacewaters of the oli-gotrophic ocean
Parasitism is also a common symbiotic relationship amongmarine protozoa Protozoan parasites have been reported for adiverse array of protists and animals in the ocean Heterotro-phic dinoflagellates and presently uncultured alveolate cladesrelated to alveolates appear to be particularly successful atevolving thismode of existence (369ndash371) In a common par-asitic group Amoebophrya spp the parasite invades photo-synthetic dinoflagellates where it undergoes reproductioneventually lysing the host and releasing dozens of infectiveswarmers The parasitic protist Pirsonia spp is a parasite ofdiatoms (372) The overall importance of these parasitismsin the population dynamics of their hosts is not clearly knownbut may constitute a significant source of mortality for somespecies (373 374) The frequency and diversity of ldquonewrdquo
FIGURE 8 Meta-analysis of experimental studies that have com-pared rates of bacterial productivity (converted to population growthrate units of dminus1) with rates of bacterivory by phagotrophic protists(expressed as equivalent units of bacterial growth rate dminus1) Thesolid line indicates where measurements of bacterial removal by graz-ers were equal tomeasurements of bacterial production Data redrawnfrom Sanders et al (63) doi1011289781555818821ch422f8
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-21
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
extensively for cloning and related studies The targetsequence can be almost instantly ldquoidentifiedrdquo to its closestphylogenetic neighbor by what have come to be standardonline sequence comparisons
Beyond the study of targeted genes (like the 16S rRNAgene) shotgun metagenomic studies have examined theentire genetic repertoire of the microbes in a given sampleThe metagenome is the collective genome of all organismsin the sample Initially these studies extracted DNA fromall organisms in a sample (usually prefiltered through a filterapproximately 1 microm to remove most organisms larger thanbacteria) sheared it to produce fragments and clonedthem into standard vectors either as small (thousands ofbases) or large (to hundreds of thousands of bases) insertsthat is fragments of DNA from the environment nowcloned into the vectors in a form suitable for sequencingThe best known early marine study of the former type isthe Global Ocean Survey with initial results published byVenter et al (105) which generated more than a billionbases of DNA sequence and reported 12 million previouslyunsequenced genes estimated to come from at least 1800different genomic species cumulatively in the many samplesthey analyzed
With the advent of next-generation sequencing that gen-erates millions ormore sequences in a run (known by a varietyof acronyms including 454 Illumina SOLiD etc) cloningof genes has largely been replaced by clone-free sequencingThe extent and power of such sequencing has recentlybeen demonstrated by the ability to construct essentiallythe entire genome of an uncultivated marine Group II Eur-yarchaeon that constituted only approximately 2 of amicrobial community sample using SOLiD sequencingmade possible by high coverage and the use of mate pairsequencing ofsim3000 bp fragments (106) (this length isneeded to span repeats and highly conserved genes that oth-erwise make genome construction difficult)
For 16S rRNA gene studies ldquotag sequencingrdquo pioneeredby Sogin et al (107) consists of amplifying a suitable sizedpart of the gene with broadly conserved primers (choice isimportantmdashfew are truly universal for the groups intended)often ldquobarcodedrdquo to allow multiple samples to be combinedinto a single run The amplified products are then sequencedand sequences processed en masse Many thousands of partialSSU rRNA gene sequences per sample are economically ana-lyzed this way though they are usually short (currently a fewhundred bases each depending on the sequencing platform)This way of analyzing the composition of microbial commun-ities is now standard These approaches provide so muchinformation even about very rare sequences that the resultshave led to the important concept of the ldquorare biosphererdquoorganisms that may be active or dormant and constituting avery small proportion of the community (eg often muchless than 01) but are potentially important for dispersionadaptation to changing conditions or even critical activitieslike nitrogen fixation or vitamin production (108) Howeverdue to the potential for artifacts like error sequences thisapproach requires rigorous application of quality filteringand clustering algorithms to avoid erroneous taxa and overes-timation of species richness (109 110)
Next-generation sequencing approaches have also allowedwork to begin metagenomics studies of microbial eukaryotes(111) These studies are still constrained by the much largergenomes of eukaryotes and therefore the difficulties ofobtaining sufficient sequences to reconstruct the genome ofa particular species Along with advances in sequencing abil-ity studies of eukaryotic metagenomics have been facilitated
by advances in single-cell isolation which when coupledwith high-throughput sequencing approaches reduce the tre-mendous complexity present in natural complex eukaryoticcommunities to a manageable task (112) Such single-celltechniques are also quite valuable in studies of bacteria andarchaea though the amplification technique tends to bevery uneven and typically generates less than half the genomeof each isolated cell regardless of domain (113 114)
Additionally similar to DNA mRNA is amenable toextraction and sequencing although greater care must betaken during extraction and purification as RNA shows agreater susceptibility to degradation during processing Copy-ing of mRNA by reverse transcription of RNA into cDNAfollowed by DNA sequencing has allowed insights into themetatranscriptomes of environmental samples Metatran-scriptomic studies provide information on gene expressionin an ecosystem and thereby indicates ldquoactivityrdquo of themicro-bial community rather than simply ldquopotentialrdquo represented bythe genomic DNA present in the sample with many applica-tions from showing which processes are being carried outby which organisms to fine-scaled diel studies (115ndash118)Nevertheless given variations in the lifetimes of differenttranscripts and protein molecules the transcriptome maynot be fully representative of the current activity of anorganism
Limitations of these analyses include sequencing errorsPCR mismatches or biases clustering and bioinformaticschallenges and chimeras generated during PCR Also thephylogenetic resolution of short sequences is limited giventhe high conservation of rRNA sequences Even with clone-free shotgun metagenomics there may be biases such as non-random losses of DNA during extraction and preparationor biases (eg from G +C content or secondary structure)in the sequencing procedures Determination of speciesdiversity by these approaches provides an example of theselimitations Shakya et al (119) working with synthetic com-munities (purified genomicDNA from16Archaea represent-ing 3 phyla and 48 Bacteria representing 16 phyla remixed tosimulate an environmental DNA extract) applied both meta-genomic analysis (454 and Illumina platforms) and PCRamplification followed by 454 sequencing of 16S rRNA genesto determine both species richness and relative abundanceThey found that PCR amplification454 sequencing of 16SrRNA genes yielded an accurate measure of species richness(providing that appropriate data processing was applied)but that the relative abundance of up to 94 of the species(depending on domain and variable region amplified) wasover- or underestimated by at least 15-fold (values rangedfrom not detected to 103-fold overestimation) In contrastboth metagenomic approaches yielded relative abundancesthat were within the authorsrsquo 15-fold accuracy cutoff forsim50 of the species However they concluded that addressingrichness overestimation in metagenomic analyses that isdistinguishing rare but real OTUs from experimental andcomputational artifacts awaits further computational andclassification improvements More recently Parada et al(120) used mock communities composed of 16S rRNAclones from 27 common marine taxa (from nine Bacterialand two Archaeal phyla) to show that small differences inPCR primers (and different clustering methods) can yieldlarge differences in apparent relative abundances of reportedtaxa However one primer pair and informatics pipelinethey tested using a particular version of 515F-926R (V4ndashV5) provided accurate estimates of relative clone abundance(r2 = 095) when comparing observed versus expected cloneabundance
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-9
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
Culture-Independent Diversity Studies
The first groups to be identified using cultivation-independ-ent molecular techniques from the marine plankton (121)were the bacterial SAR11 cluster (a group of closely relatedgene sequences or phylotypes) and marine picoplanktonicunicellular cyanobacteria Synechococcus and Prochlorococcus)Of these the SAR11 cluster was completely unknown butthe cyanobacteria had previously been recognized by theirunique pigment fluorescence Waterbury et al (122) andJohnson et al (123) used epifluorescence to observe Synecho-coccus and Chisholm et al (124) discovered Prochlorococcususing flow cytometry These cyanobacteria were later isolatedand grown in phytoplankton culturemedia These two groupsare generally common in the euphotic zone with SAR11 typ-ically comprising one third of the planktonic bacteria (125)and the cyanobacteria common everywhere but polar waters
Probably the biggest surprise to come from the applicationof molecular tools was the discovery by Fuhrman et al (126)of abundant archaea in the deep sea The archaea were foundto be in a unique phylogenetic cluster that was only distantlyrelated to any previously known archaea but the ldquoclosest rel-ativesrdquo (not really close at all) were extreme thermophiles Asubsequent study also found archaea to be present in near-surface coastal waters albeit relatively rare (lt2 relativeabundance) This study used a PCR technique specificallytargeted archaea and found ldquomarine Crenarchaeardquo plus a sec-ond group belonging to the phylum Euryarchaea (127)
Up until this time all known Archaea were thought to beldquoextremophilesrdquomdashadapted for either very high temperatures(thermophiles) extremely salty conditions (halophiles) orstrictly anaerobic environments (methanogens) Yet theseorganisms were present in cold or cool water at ordinary salin-ity and high oxygen concentrations Fluorescent in situhybridization (FISH) measurements from deep-sea sampleshave since showed that the archaea may make up appro-ximately 40 of the total countable prokaryotes with thepercentage reaching to 60 at 200 m depth in theMediterra-nean (35 36) An extensive time series of FISH measure-ments near Hawaii confirmed that the archaea are indeedvery abundant throughout the year from below the photiczone to at least 4000 m and typically constituting 30ndash40of the total prokaryotes present in waters deeper than a fewhundred meters (128) with similar results found elsewhere(129 130) (Fig 4) They have been reported from many pla-ces including the Atlantic Pacific and Southern Oceansand are dynamic components of the plankton the most com-mon type by far being the ldquomarine Crenarcheaeardquo (128 129131) While the marine archaea have been reported to bedominated by a few major ldquophylotypesrdquo (132) they alsohave been shown to have a great deal of microdiversity withinthese phylotypes suggesting there are many kinds of close rel-atives coexisting (133) Interestingly this group of archaeamay likely be the most abundant kind of organism on Earthgiven the huge volume of the deep sea and their high abun-dance there (134) It has recently been proposed that ldquomarineCrenarchaeardquo be elevated to their own major phylum outsidethe Crenarchaeota called the Thaumarchaeota on the basisof deep phylogenetic branching and fundamental differencesbetween them and the Crenarchaeota (135) The Thau-marchaeota possess the uniquely archaeal membrane lipidcrenarchaeol and are nowalso known to be abundant in soils
The physiology of the Thaumarchaeota has been anintriguing area of study Initially an autoradiography-FISHcombination approach demonstrated that they take upamino acids (130 136) But subsequent data showed that
crenarchaeol was derived largely from CO2 suggesting auto-trophic metabolism (137 138) An experiment showinguptake of 13C-labeled bicarbonate into these archaeal lipids(139) directly pointed to autotrophy in this group Chemoli-thoautotrophywas first hinted at byVenter et al (105) whosemetagenomic analysis showed an apparent archaeal scaffoldthat contained genes suggestive of ammonia oxidation Fur-ther evidence came from Schleper et al (140) who detectedseveral ammonia oxidation genes in order directly adjacent toa Thaumarchaeota 16S rRNA gene in a soil-derived metage-nomic clone The issue was directly resolved when Konnekeet al (141) isolated a related marine archaeon from sedimentof a marine aquarium and this organism Candidatus Nitroso-pumilus maritimus was found to have a chemoautotrophicmetabolism quantitatively oxidize ammonia to nitrite andcontain an archaeal ammonia monooxygenase gene (amoA)and interestingly did not grow heterotrophically Cultivationexperiments showed this organism has a high affinity forammonium allowing it to outcompete bacterial nitrifiers atlow (submicromolar) concentrations as are typical in thesea (142) The complete genome of the organism has shownnovel adaptations for nitrification and autotrophy (143)
Even if the Thaumarchaea are primarily chemolitho-autotrophs field data suggest a level of mixotrophy in thatsome organic substrates are being incorporated into biomassA stable isotope study using cells collected at 670 m depthoff Hawaii estimated that about 80 of the carbon incor-porated into archaea-specific lipids came from inorganic sour-ces and about 20 from organic compounds (144 145)Interestingly it has also been claimed that the genomesof deep-sea (gt2000 m depth) members of the Thaumarch-aea as well as those living in equatorial waters rarely containthe amoA gene and thus may be primarily chemoorgano-trophs (48)
Major bacterial groups that have been documentedfrom seawater using 16S rRNA characterization includesome that are also known from culture (eg AlteromonasRoseobacter) and several that are phylogenetically distantfrom standard cultures The most common groups are inrough order of their relative abundance in clone librariesfrom most to least abundant SAR11 (relatives of Pelagibac-ter ubique) Roseobacter SAR86 cyanobacteria SAR116SAR202 SAR234 andMarine Group A The SAR designa-tion followed by a number is an arbitrary sequential cloneidentifier from Sargasso Sea cloning studies done by the Gio-vannoni lab that did most of the early systematic cataloging ofclones Summaries of the data and phylogenetic relationshipsof these groups can be found in Fuhrman andHagstrom (102)and Giovannoni et al (101)
Molecular Genetic Discoveries in Bacterial andArchaeal Marine BiologyAs described earlier metagenomics is an extension of theideas used in the 16S rRNA cloning studies in that all genesfrom the native microorganisms are separated and clonedwithout having cultivated the organisms These methodsearly on started to find unexpected and very interestingresults The best examples involve unexpected marine photo-trophy the first of which is discovery of a nonchlorophyllphotosynthetic bacterial pigment called proteorhodopsin(146) The gene was found on a large environmentallyderived fragment of DNA that also had a gene coding for16S rRNA from the so-called SAR86 group (one ofthe groups common in seawater) This pigment can act as alight-driven proton pump thought to permit cells to generate
422-10 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
FIGURE 4 Distribution of Bacteria Euryarchaea andmarine Crenarchaea (recently renamed Thaumarchaea) along a transect in theNorthAtlantic Ocean as measured by CARD FISH and expressed as of total bacteria + archaea counts via DAPI stained epifluorescence fromTeira et al (130) Top panel shows station locations that are shown on the top of the lower three panels with the distance in km reported alongthe transect from lower to higher station numbers doi1011289781555818821ch422f4
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-11
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ATP from sunlight Interestingly different versions of this pig-ment are found at different depths apparently ldquotuningrdquo theabsorption to match the ambient wavelengths of light asthey change with depth (147) The proteorhodopsin genesare very widespread and diverse perhaps in half or more ofall marine bacteria including SAR11 and Euryarchaeota(105 148ndash150) However almost all of several culturedorganisms with proteorhodopsin studied to date do not showa growth benefit from light (150) with the exception of amember of the Flavobacteria Dokdonia sp strain MED134that grows faster in the light only under moderately lownutrient conditions (151) and a marine Vibrio has beenshown to survive starvation longer in the light than in dark-ness (152) These observation may explain the wide phyloge-netic distribution and high abundance of this gene in thatproteorhodopsin may often assist long-term survival of bacte-ria under extreme energy-limiting conditions yet not havemucheffect on growthduringmore energy-replete conditionsInterestingly a recent report suggests the growth benefit fromlight in Dokdonia MED134 is from enhanced uptake of itsrequired growth factor vitamin B1 which is transported by atonB-dependent transporter powered by a proton gradient(153) This pigment might also have sensory or other rolesnot yet well examined (150)
The second kind of phototrophy found by moleculargenetic and fluorescence techniques to be unexpectedlyimportant in marine plankton is anoxygenic aerobic bacterialphototrophy based on the pigment bacteriochlorophyll aCulturable aerobic anoxygenic photosynthetic (AAP) bacte-ria have been known from seawater for several years (154) andare widespread (155) Initial recent reports of direct countsclaimed they represent about 11 of the total bacterial com-munity in the euphotic zone (156) but those early counts didnot correct for the presence of other bacteria and such correc-tionoften yields estimates averaging closer to 2(157)Directmeasurements show the bacteriochlorophyll pigment is rela-tively rare (158) However studies in various ocean locationsincluding ocean gyres report that these AAP bacteriacan indeed on occasionmake up a quarter of the total prokar-yotes (159 160) A recent energetic modeling of AAP andproteorhodopsin-containing bacteria (161) has suggestedthat proteorhodopsin-containingones cangain approximately02asmuchenergy from sunlight asSynechococcus (commonmarine cyanobacterium) and AAP bacteria can gain approxi-mately 13 as much energy from sunlight as SynechococcusThey concluded the AAP bacteria may gain energy sufficientto meet maintenance costs but proteorhodopsin-containingbacteria were not expected to do so except at high light inten-sities andwith largenumbersof proteorhodopsinmoleculespercell They concluded the ease and low cost of maintainingPR-based phototrophy (a few genes required) may explainthe high incidence of proteorhodopsin genes
Other metagenomic studies have yielded interestinginsights about organisms and processes that would otherwisebe difficult or impossible to establish For example thesequence of an archaeal gene fragment isolated from seawaterrevealed extensive evidence of genetic exchange with othertypes of archaea and even bacteria (162) Such geneticexchange was also quite evident from the complete genesequences of different strains of Prochlorococcus and Synecho-coccus which was attributed in part to virus-mediated geneflow (163ndash166)
Dilution Cultures of Bacteria and ArchaeaSome recent cultivation techniques that permit growth ofldquotypicalrdquo oligotrophic marine bacteria in pure culture are
based on the simple concept that bacteria living in seawateron dissolved organic matter might best be grown in the labo-ratory in ordinary filtered seawater This conclusion followsfrom similar techniques to grow mixed ldquoseawater culturesrdquoof marine bacteria (167) but the seawater inoculum in thiscase is diluted so that only one or a few bacteria are addedto the initial culture vessel (168) A rapid throughput versionof this method has been used to cultivate members of bacte-rial groups thought to be conventionally uncultivable such asthe SAR11 clade (169 170) albeit often at very low den-sities Such cultures permit focused studies of the propertiesof those organisms including genomic analysis (169 170)This valuable work provides insight into the roles and activ-ities of some of the most common bacterial phylotypesobserved in the ocean (171) showing for example that culti-vated SAR11 requires a source of reduced sulfur as well asproviding templates aiding the interpretation of metage-nomic analysis (172)
PROTOZOADiversity and DistributionHeterotrophic protists have been identified historically fromtheir morphological features that are apparent at the levelof the light or electronmicroscope Features of cell size shapetype and pattern of flagellationciliation skeletal structurescharacteristics of the nucleus and other cellular structuresand organelles have been used to differentiate themany thou-sands of described species A description of the extent of thisdiversity of form and function is well beyond the scope of thisbook (see [79] for more information) However much of thisdiversity can be reduced to three basic body plans that dictatethe broadest ecological roles of these cells amoeboid flagel-lated and ciliated forms (Fig 5) Among the lineages possess-ing one of these three forms only the last group the ciliatedprotists constitute a monophyletic group within the domainEukarya
One of the simplest body plans for protozoa is the amoe-boid cell exemplified by the gymnamoebae or ldquonakedrdquoamoebae Motile nonphotosynthetic cells lacking flagellaor cilia occur as life stages in a number of phylogeneticallydiverse taxa but for many free-living species of protozoathis form constitutes the only life stage Motility is largelyconfined to movement along surfaces by means of pseudopo-dia that can take on a variety of (species-specific) shapes Sig-nificant abundances of amoebae are largely confined tobenthic and epibiotic environments (174) and to suspendedparticulate material where they can occasionally be highlyenriched (27) Most amoebae consume bacteria and otherminute prokaryotes and eukaryotes
Several heterotrophic protistan groups possess complexamoeboid body plans most notably the foraminifera polycys-tine and phaeodarian radiolaria and the acantharia Plank-tonic forms are predominantly oceanic in their distributions(although a large number of benthic species of foraminiferaexist) These species are heterotrophic and many are visibleto the naked eye (individual cells can bege1 cm gelatinouscolonies can form ribbon-like structuresge1 m in length)They feed on a wide variety of bacterial protistan and meta-zoan prey using pseudopodial networks have rather longcomplex life cycles for individual cells (weeks to months)and are extensively used in studies of paleoclimatologicalreconstruction (82 175) In addition many of these speciespossess intracellular symbiotic usually eukaryotic algaeThe widespread occurrence of algal endosymbiosis among
422-12 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
these protozoa in oceanic pelagic environments implies thatstrong selective forces appear to give rise to these associations(97 98)
Flagellated forms of protozoa exist across many protistanlineages Eukaryotic flagella come in various sizes numbersand forms (eg with or without tiny hairs) that are character-istic of the different lineagesMany flagellate species are appa-rently purely phototrophic or heterotrophic (phagotrophic)but a significant (still poorly known) fraction of flagellates aremixotrophic combining both phototrophy and phagotrophy(70 88) Flagella in phagotrophic flagellates are employed formotility and prey capture Most free-living flagellates possessone to four flagella (typically one or two) that can be manytimes the length of the cell itself Although there is tremen-dous species diversity among flagellated protozoa many ofthese species have broadly overlapping ecologies As a groupflagellates are the most numerically abundant protozoa ofboth benthic and pelagic ecosystems and they are fundamen-tally important as consumers of bacteria cyanobacteriaand other eukaryotes Flagellated protozoa within the nano-plankton size class (2ndash20 microm) are often counted as a singleassemblage in plankton studies because morphological details
apparent using light microscope are insufficient to distinguishamong the many species This assemblage has been variouslyreferred to by a variety of names and acronyms including het-erotrophic nanoplankton (HNAN HN) heterotrophic flag-ellates (Hflags) heterotrophic nanoflagellates (HNF) andmicroflagellates Heterotrophic nanoplankton is the mostaccurate term for most methodologies employed to countthese cells because it does not require visualization of flagella(which are often lost from these small cells during preparationfor microscopy) The confusion over terminology is partly aconsequence of the different methodologies that have beenused to count these species and partly a consequence of thefact that these species were first studied prior to the wideacceptance of the size convention of Sieburth et al (84)
Heterotrophic flagellates in the microplanktonic size class(20ndash200 microm mostly dinoflagellates) are important consum-ers of phytoplankton in pelagic ecosystems Many of thesespecies are capable of the production of large pseudopodialnets the pallium that can engulf prey (particularly diatoms)significantly larger than the diameter of the dinoflagellatetheca (176) This behavior and its ecological significance forenergy flow in plankton communities have been recognized
FIGURE 5 Body plans and size ranges of protozoa These micrographs depict amoeboid (andashg) flagellated (hndashj) and ciliated (kndashm) forms ofprotozoa From Caron et al (173) Markers bars are 5 (i) 10 (h) 20 (c j l) 30 (a) 50 (k m) 100 (b f) 500 (d) and 1000 (e g) micromdoi1011289781555818821ch422f5
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-13
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
only within the past few decades (177) These species areoften abundant in waters where diatoms dominate (178179) In such situations heterotrophic dinoflagellate biomasscan be similar to that of ciliated protozoa (180)
Ciliates are generally the most recognizable form of pro-tozoa to nonspecialists Nonetheless ciliates are a diversegroup morphologically as well as ecologically (181 182)The degree of ciliature on these species can range from uni-formly ciliated to totally devoid of cilia during most of theirlife cycle to ciliature restricted to specific regions of thecell In addition cilia can fuse to form various complexstructures (eg ciliary membranes or veils cirri) that assistin locomotion food capture or attachmentCiliates are abun-dant and ecologically important species in both benthic (8)and pelagic (183) ecosystems and can consume a variety ofprokaryotic and eukaryotic prey Together with the heterotro-phic dinoflagellates these species are the dominant consum-ers of phytoplankton in many pelagic ecosystems (184) andas a consequence they form an important trophic link to met-azoan zooplankton (185 186)
Diversity and Biogeography of Protists FromMorphology to DNA SequencesUntil relatively recently the biodiversity of marine protistanassemblages was not generally considered a controversialtopic It was generally accepted that while all species of pro-tists certainly have not been identified representatives ofmost types of algae and protozoa had been observed anddescribed if not actually brought into culture in the labora-tory Direct sequencing of 18S rRNA genes from environ-mental samples (as described in ldquoMolecular Phylogeny andMetagenomics Field Applicationsrdquo) has changed that viewInitial forays into environmental DNA indicated a muchgreater diversity of protists than previously documented usingclassical approaches of culture and microscopy (81 187ndash191) Numerous publications over the past decades haveexpanded these findings (see [192] for a recent global analy-sis) which are highly analogous to discoveries in marine pro-karyote research as described already
Previously uncharacterized protistan diversity has nowbeen documented at virtually every level of eukaryoticorganismal classification Some of these findings couldhave been expected but some have been very unexpectedFor example a much greater diversity than noted previouslyhas been observed among small (lt10 microm) protists in plank-tonic ecosystems These species generally possess few distinc-tive morphological features and one could expect that manycryptic species might be present among these small morpho-types The molecular ldquodiscoveryrdquo of this eukaryotic diversityhas stimulated progress on the isolation and descriptionof new species and genera of minute algae and protozoa(193ndash197)
Analyses of environmental samples have also indicatedthe unanticipated existence of novel 18S rRNA gene sequen-ces that imply the presence of novel lineages of eukaryotes innatural protistan communities (198ndash200) These are sequen-ces for which there are apparently no known described orcultured species The degree to which these sequences dif-fer from sequences of known sequenced eukaryotes impliesthat some of these lineages may be distinct at the level ofphylum (201)
These findings have raised basic questions and somedebate concerning the true diversity and biogeography ofprotistan assemblages in natural ecosystems (202ndash204) thevalidity of the many novel phylotypes or cryptic species that
are being documented (205) the potential importance andsignificance of the many rare taxa that characterize thesecommunities (206) and the implications of these findingsfor the ecologicalbiogeochemical roles that protists playin aquatic ecosystems For example it was postulated andsubsequently confirmed that some novel alveolate lineagesreported from marine ecosystems make up a suite of parasiticprotozoa whose ecological importance may have been signifi-cantly underestimated in the past (207) Deciphering theidentity and significance of these many unknown phylotypeswill constitute a significant effort for protistologists in thefuture (80 85)
Life Histories and Ecological Strategies of ProtozoaAbundance prey type and life histories all vary tremendouslyamongmarine protozoa The smallest species (eg many flag-ellates) tend to be the most abundant and widely distributedin the world ocean Indeed many of these species may beglobally distributed (208) Most nanoplanktonic flagellateshave potentially rapid rates of grazing and growth Underoptimal conditions these species can divide by binary fissionseveral times a day and thus dramatic increases in their pop-ulations can take place in response to favorable conditionswithin a few days (209) Many of these species are capableof surviving for limited periods of time without food andhave developed a variety of physiological or life cycle strat-egies to cope with these events (209) However in contrastto some bacteria that may remain viable through long periodsof starvation (210) protozoa will expire or encyst in responseto low food abundance and thus ldquoboom-and-bustrdquo popula-tion changes are characteristic of small flagellates
On the other end of the size spectrum from the ratherubiquitous nanoflagellate species many of the largest proto-zoan species (polycystine and phaeodarian radiolaria plank-tonic foraminifera) are exclusively oceanic (ie do notsurvive in most coastal environments) andor have specificlatitudinal and depth distributions Thus the abundancesof these latter species may range from undetectable to max-imal abundances of gt105 individualsm3 Dramatic changesin the abundance of these protozoan taxa also can be relatedto changes in prey abundance physicalbehavioral aggrega-tion or to periodicity of life cycle events For example theplanktonic foraminifer Hastigerina pelagica reproduces on alunar cycle and thus abundances (and life stages) of this spe-cies in oceanic waters can vary considerably over the courseof a month (211) In general life cycles for the large amoe-boid protozoa are lengthy and complex (for single-celledorganisms) with life spans unknown for many species(attempts to culture them in the lab have so far been unsuc-cessful) but estimated to be on the order of months to per-haps years (175)
The abundance and activities of microplanktonic hetero-trophic protists (mostly ciliates and heterotrophic dinoflagel-lates) tend to be somewhat intermediate to those ofnanoplanktonic flagellates and the larger amoeboid formsThese species are present in the majority of marine ecosys-tems and collectively play an important role in the controlof phytoplankton biomass (and probably the abundance ofnanoplanktonic protozoa although there is little informa-tion on this topic) in waters throughout the world oceanLike small flagellates ciliates and dinoflagellates reproduceprimarily by binary fission but their maximal growth ratesare typically slower (one division a day is typical) and theirlife cycles often include sexual phases that allow for geneticrecombination
422-14 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
VIRUSESViral Abundance and General PropertiesViruses are simple biological agents typically 20ndash200 nm indiameter composed of a nucleic acid genome in a proteincoat that infect cells and ldquocommandeerrdquo the cellrsquos machineryto make more viruses which are released into the environ-ment when the host cell lyses or bursts A particular virus isthought to be capable of infecting only a narrow range of hosts(usually one species sometimes a genus rarely broader)Although some early studies had isolated viruses from thesea therewere no data prior to the 1980s showing such viruseswere very abundant and more significantly no evidence thatinfection was occurring in any important part of the planktoncommunity It wasnrsquot until the late 1980s that electronmicro-scopy with suitable concentration methods showed thatviruses are extremely abundant similar to or even higherthan bacterial abundance (212 213) Interestingly themost suitable transmission electron microscopy (TEM)approach used by Bergh et al (described in detail by [214])was actually similar to a direct centrifugation method devel-oped in 1949 (215)
TEM studies of viruses permit high-resolution images andobservation of viral morphology showing features such ashead diameter tails sheaths and tail fibers (Fig 6 collage
in upper left) Each type of virus has a fixed morphology(unlike bacteria that are potentially more plastic in theirappearance) and hence a coarse measure of viral diversityhas been possible by cataloging morphologies Studies thathave done so have found dozens or more different morpholo-gies of marine viruses in a given sample (217ndash219) A largeproportion resemble bacteriophages which are viruses thatinfect bacteria
The availability of brightly fluorescent nucleic acid stainsand high-porosity fine pore size (002 μm) filters made fromAl2O3 have made it easy to accurately count viruses by epi-fluorescence microscopy (38 220 221) Epifluorescence per-mits abundance estimates but does not allow observation ofviral morphology as viruses are below the resolution limitof light microscopy viruses are visible only as sources of light(like stars in the night sky see Fig 6 lower micrograph) Anextension of manual epifluorescence counts is the use of flowcytometry to count viruses now commonly employed in somelabs (222)
Virus abundance has been found to be closely related tobacterial abundance with a virusbacteria ratio typically10ndash301 A typical oceanic profile of bacterial and viral abun-dance is shown in (Fig 6 panel on right) This tight ratioand the strong correlation to bacterial abundance in relationto weaker correlations to chlorophyll have been cited as
FIGURE 6 Marine viruses (upper left collage) viewed by transmission electron microscopy These are cyanophages in the groups myoviridae(right and bottom) siphoviridae (left) and podoviridae (top) Scale bars represent 100 nm From Sullivan (216) Epifluorescence micrograph(lower left) of SYBR green I stained viruses (small fluorescing objects) and bacteria + archaea (large fluorescing objects) Depth distribution ofvirus and bacteria + archaea abundances obtained by epifluorescence microscopy of SYBRGreen stained cells in the central San Pedro BasinCalifornia (11 August 2000) doi1011289781555818821ch422f6
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-15
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
evidence that most marine viruses infect bacteria rather thaneukaryotic phytoplankton (217 218 223 224) though thereare also many important viruses of phytoplankton and otherorganisms with significant impacts on primary productivityblooms and ecosystem function (225ndash227)
The first demonstration of the activity of marine viruseswas presented by Proctor and Fuhrman (228) who showedby electron microscopic examination that viruses wereactually infecting marine bacteria and cyanobacteria at ameasurable rate Because only the final portion of the viruslife cycle is visible by electron microscopy (when the virusesare assembled and ready to lyse the host cell) only a very smallfraction of the infected cells can be counted as infected at anygiven time Proctor and Fuhrman (228) reported that only afew percent of the total bacteria from coastal waters and theSargasso Sea were visibly infected but they interpreted thedata with a model that implied the actual fraction of the totalcommunity that was infected was much higher They con-cluded that the total fraction of bacterial mortality attribut-able to viruses is roughly 10ndash40 Subsequent studies haveused refined versions of that same general approach andmodel (229 230) and numerous studies have used a varietyof alternative approaches to estimate viral activitymdashall yield-ing basically the same general conclusion (although themodel parameters need to be adjusted for cyanobacteriawhich on average seem to be infected less than bacteria)These approaches include (a) calculation of virus turnoverfrom decay estimates (231) (b) measurement of viral DNAsynthesis by incorporation of tritiated thymidine (232) (c)observation of effects of added viruses (233 234) (d) estima-tion of bacterial mortality in the absence of protists (e) use offluorescent viruses to estimate production by an approachanalogous to isotope dilution (38) and (f) dilution of virusesin filtered samples to estimate viral production (235) Theoverall consensus of these studies is that the initial estimateswere basically correct that is that viruses are responsiblefor about 10ndash40 of the bacterial mortality in most marinesystems (reviewed by [218 223 226 236 237]) Howeverthere is still some disagreement about the higher estimatesand it is likely that 40 mortality from viruses alone is nottypical for most marine systems
Most of the foregoing work has focused on the viral infec-tion of bacteria primarily thought to be heterotrophicalthough the original report by Proctor and Fuhrman (228)also noted the occurrence of cyanobacterial infection Sev-eral subsequent studies focused on phytoplankton includingthe potential effect of viruses on the termination of phyto-plankton blooms of Emiliania and Phaeocystis and the likelyimpact this might have on release of climate-active gasessuch as dimethyl sulfide (225 227 238ndash247) Althoughdetails are beyond the scope of this chapter viruses arethought to infect virtually all marine organisms with poten-tially significant impacts from zooplankton to whales (226)Detailed studies of viruses infecting cyanobacteria such asSynechococcus have shown differences in viral host specific-ity particularly toward coastal and oceanic host strainsand some occasional high virus abundances (to 105ml) asmeasured by most probable number (MPN) cultivation tech-niques in the Gulf of Mexico near Texas (248 249) Othervirus cultivation studies with Prochlorococcus and Synecho-coccus in oligotrophic waters of the Sargasso Sea showedan interesting pattern of cross-infection between these gen-era by some virus types (suggesting gene flow among theseorganisms) but generally low MPN estimates of abundanceto 103ml even when cyanobacterial abundance was near105ml (216 250)
Overall the consensus emerging from direct comparisonsof viral-mediated mortality and grazer-mediated (ie micro-zooplankton) mortality have indicated that viral lysis of bac-teria constitutes a significant fraction of total mortality ofthis assemblage while the mortality of phytoplanktonappears to be dominated bymicrozooplankton inmost instan-ces (251 252)
As mentioned viruses have significant morphologicaldiversity as observed in TEM studies It is also possible toinvestigate aspects of their genetic diversity In early workon this topic Wommack et al (253) and Steward et al(254) observed the diversity of viral genome lengths in a fieldsample by pulsed field gel electrophoresis with viral genomesranging 25 kb to gt300 kb in length Field results show thatthe viral community composition is dynamic in space andtime with clear changes in the banding patterns over seasonsand locations in Chesapeake Bay (253) between oceanbasins and subsequent to dinoflagellate blooms (254) andwith depths to 500 m at one location (255)
Although viruses do not all share a set of core genesthat allows a universal viral phylogeny (comparable to SSUrRNA gene in cellular organisms) genetic diversity amonga single group of viruses can be examined by sequence analysisof shared genes within the group An example is the g20 geneinT4-like cyanophages that has shown extremely high diver-sity even among very closely related viruses and has demon-strated geographic and seasonal variation (256) A secondexample is g23 which is found broadly in diverse and wide-spread T4-like phages (257) and can have seasonally repeat-ing patterns (258) as well as short-term rapid dynamics thatcorrelate to those in bacteria (259)
Viral MetagenomicsAs with cellular organisms metagenomics can be used toexamine viral diversity and genetics without themany restric-tions of cultivation Viruses can be collected by selective fil-tration and concentrated by tangential flow filtration orflocculated with iron chloride (260 261) then their collec-tive metagenome can be extracted linker amplified and ana-lyzed by sequencing (262 263) Viral metagenomic studiesare particularly challenging because the large majority ofsequences have no annotated matches in any databases butso far results from marine samples around the world haveshown extremely high diversity and variations with depthlocation and time presumably with highly dispersed typesselected by local conditions (262ndash266) Because metage-nomes are best interpreted when there are representative cul-tures available the best matches of viral metagenomesoriginally tended to be to the few viral isolates infecting trulycommonmarine bacteria like cyanophage infecting Synecho-coccus and Prochlorococcus (267) However the developmentof dilution-to-extinction cultures representing common het-erotrophic marine taxa like Peligibacter (a member of theSAR11 clade) and SAR116 has allowed isolation from sea-water of viruses infecting these common organisms and theseisolates have indeed been found to be highly abundant inmarine viral metagenomes (268 269) New approaches tointerpret the results include clustering the proteins independ-ent of known proteins to compare samples to each other andtry to find environmental factors driving viral communitychanges (270) Such analysis of a large global data set (TaraOceans expedition) has shown that extensive sampling hascome close to reaching the total diversity in tropical and tem-perate waters of such viral protein clusters (which essentiallyrepresent various viral protein motifs not all viral proteintypes) and that viruses appear to be directionally dispersed
422-16 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquodownstreamrdquo in major ocean currents as one may expectfrom first principles but also supporting the ldquoseed bankrdquohypothesis of viral biogeography (271)
Viruses and Host DiversityViruses are themselves thought to be instrumental in drivingincreased diversity of their microbial hosts via a hypothesizedprocess often called ldquokill the winnerrdquo This is because viralinfection is host-specific and density-dependent the latterbecause viruses diffuse from host to host so an abundanthost is more likely to pass on infection than a rare oneThis means that if an organism becomes abundant andblooms winning the competition for resources it becomesmore susceptible to a viral epidemic This would benefit therarer organisms and thus help foster diversity (223 272273) However bloom scenarios as described above involvesystems far from steady state and the formal theory of Thing-stad and Lignell (274) has interesting steady-state solutionswhere several viruses infect several hosts stably over time(via trade-offs between growth rates and viral susceptibility)which may occur at the strain or species level (274) There issome experimental evidence that viruses have effects on nat-ural marine microbial community composition weaker thanthe kill the winner (bloom version) hypothesis would sug-gest (275 276) so something resembling the steady-statecoexistence described in the model may in fact be commonThere also appear to be processes that foster coexistencebetween viruses and hosts but the mechanisms are largelyspeculative (223 277 278)
Viruses may also be directly involved in host genetic diver-sity because they can be the agents of genetic exchangebetween microorganisms (223 273) This often involvesthe viral lifestyle known as lysogeny whereby viruses survivewithin host cells as DNA only integrated into the host chro-mosome and being reproduced each time the host divides Ahost harboring such a genome is called a lysogen becauseunder conditions of stress to the host cell a genetic switchmay cause the viral genome to initiate the lytic process pro-ducing many progeny viruses and bursting from the hostLysogeny is a very common property occurring in a signifi-cant part of the bacterial community (279 280) althoughthe incidence of induction of the lytic phase in nature is appa-rently low (281) Overall lysogeny is poorly understood butthought to have both positive and negative impacts on themicrobial community (282) Recent results suggest that ben-efits of being lysogenic in highly seasonal polar seas leads tofundamental difference between polar and other marine viralcommunities (283)
Viruses and the Microbial LoopAs part of the food web viruses occupy a unique positionThey infect host cells that are mostly thought to be heterotro-phic bacteria and by doing so they typically burst the hosts torelease progeny viruses and cellular debris But what is the fateof this material Viruses themselves do not last indefinitelyand a simple steady-state assumption implies that from eachburst of viruses (typically 20ndash100 per lytic event) only onesuccessfully infects another cell The rest are inactivatedand broken down by sunlight (UV and visible exposure)and enzymatic attack (284) or consumed byminute phagotro-phic protists (285) thus reentering the food web as substratefor bacteria or food for protistan consumers Experiments incontrolled laboratory systems and field studies with radioac-tively labeled viral lysis products have supported the conclu-sion that most of the organic matter released by the viralinfection is either taken up by bacteria or respired (286 287)
Modeling this process as part of the microbial loop showsthat viral lysis represent a sort of side loop that has the neteffect of remineralizing a significant amount of the carbonand nutrients that enter the bacteria-protist part of themicro-bial loop (Fig 7) A theoretical numerical steady-state modelcomparing a systemwith no viral activity to onewhere virusesare responsible for 50 of bacterial mortality showed that thesystem with viruses had 33 more bacterial production andrespiration than the virus-free system implying that theviruses had the effect of permitting the bacteria to processmore of the primary production than they would otherwise(223) Although 50 is a high number unlikely to be com-mon in the sea this model nevertheless illustrates that virusescan reduce the amount of energy reaching higher trophic lev-els The implication is that viruses lead to increased bacterialactivity at the expense of the larger organisms
MAJOR ENVIRONMENTAL CONTROLSLight Temperature and PressureTemperature has an important potential influence on bio-chemical reactions and therefore on biological processesin general Most ocean waters fall in the range of ndash2degC to30degC with obvious exceptions in hydrothermally heatedareas Temperature has long been known to be a regulatingfactor for the growth of heterotrophic microbes
In temperate waters it has been established that microbialactivity is generally much higher in warm summer waters thanin winter (55) The relationship is not simple howeverbecause multiple factors act at the same time Some contro-versy still exists regarding the highest and lowest extremesfor marine bacterial growth although there is broad agree-ment that bacteria grow gt100degC at hydrothermal vents andltminus5degC in sea ice brines Pomeroy et al (57) noted the inter-esting observation that bacteria seem particularly inhibitednear the freezing point of seawater (ca ndash22degC) comparedto eukaryotic phytoplankton This effect results in polarspring phytoplankton blooms that accumulate organic car-bon in advance of the response of the bacterial communityand development of the microbial loop and perhaps lead toenhanced benthic-pelagic coupling (288)
The relationship between temperature and the growth rateof marine phytoplankton was described broadly in a now
FIGURE 7 Modification of the microbial loop concept that incor-porates the functional role of viruses Export can be via predation orsinking From Fuhrman (223)doi1011289781555818821ch422f7
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-17
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
classic paper by Eppley (58) and later Goldman and Carpen-ter (289) Temperature was shown to exert a strong and directeffect on themaximal growth rates of these species withmax-imal intrinsic growth rates at 0degC generally lt1 divisiondaywhile growth rates at 30degC may be gt4day These relation-ships indicate the maximal rate that might be attained byphytoplankton at these temperatures but they do not takeother factors into account (nutrients light) While warmtemperature ostensibly allows more rapid growth it createshydrographic conditions that typically give rise to nutrientlimitation of algal growth Thus some of the coldest watersin nature witness some of the most massive phytoplanktonblooms (290) while warm oceanic gyres represent some ofthe most oligotrophic areas of the ocean The multiple andoften conflicting effects of temperature on the growth of nat-ural phytoplankton assemblages limit the accuracy of presentpredictions regarding how primary producers will respond toclimate change (291)
An analysis of the effect of temperature on the growth ofheterotrophic protists has indicated that the response is qual-itatively similar to that of phytoplankton but with a twistThe growth rates of at least some protozoa can exceed thoseof phytoplankton at warmer environmental temperaturesbut the opposite effect is apparent at very low environmentaltemperature This differential effect of temperature on thegrowth of phototrophic and heterotrophic protists was basedon a large meta-analysis of published protistan growth (59)That analysis demonstrated that the maximal growth ratesattained by phototrophic protists could exceed the maximalgrowth rates attained by heterotrophic protists (all otherpotential growth-limiting factors not considered) Thereforeprotozoan growth rates may be constrained to a greater degreeat low environmental temperature than rates for phytoplank-ton If so then phytoplankton bloomsmay get a head start ongrazers during spring in polar ecosystems This scenario is con-sistent with information on seasonal biomass changes andmicrozooplankton herbivory in the Ross Sea Antarctica(292 293) but there are still too few data to fully vet thishypothesis
The importance of high pressure on bacterial growthgained considerable attention in the late 1960s when thedeep sea submersible Alvin was accidentally lost overboardwith its hatch openWhile no lives were lost in this accidentsome workmenrsquos lunches sank to the bottom (sim1500 m)inside the submersible Alvin was recovered after 10 monthsand interestingly there was a waterlogged lunch containingapples bologna sandwiches and broken vacuum bottleswith broth that all appeared hardly degraded and tasted palat-able Yet when placed in a refrigerator on the ship these itemsdegraded relatively quickly Initially pressure was thought tobe the preserving factor as the sea floor temperature was sim-ilar to the refrigerator temperature and the only major differ-ence would be pressure (294) Following this observationa series of experiments to measure degradation of variousorganic materials left in the deep sea for extended periodsindicated that degradationwas typically significantly reducedimplying that pressure reduces the degradation rates (295)Nonetheless changes in the protozoan community of naturaldetrital material sinking to the deep ocean floor indicate thatthe microbial community can respond relatively quickly insome situations (296) Deep sea microbiology has advancedconsiderably yet it is still difficult to interpret results withrespect to actual in situ rates of naturally occurring organicmatter Deep sea bacteria adapted to high pressures havebeen isolated that are barophilic (also called piezophilic)meaning that they prefer high pressures and have reduced
activity at lower pressures whereas others are barotolerant(piezotolerant) tolerating but not preferring high pressuresfor example see (297 298)
Few data are available on barotolerantbarophilic marineprotozoa Protozoa certainly exist and grow at great oceanicdepths but measurements of in situ growth rates for these spe-cies do not yet exist Measurable protozoan numbers havebeen documented in the deep-sea sediments for more than30 years (299 300) and viable protozoa have occasionallybeen cultured from these environments (301ndash305) butvery few direct measurements of the activities of these speciesin situ have been reported (306) A few protozoa have beenisolated that will grow at high pressure (302 303 307) andprotists that appear to be unique to the deep ocean havebeen observed either directly or through the analysis ofDNA sequences (308ndash311) but possibly the best direct evi-dence that protozoan activity takes place at the high pressurescharacteristics of the deep sea are experimental and observa-tional work noting the stimulatory effect that detrital deposi-tion has on some components of the protozoan community(296 312) These observations indicate a diverse and activeprotozoan fauna of the deep ocean although their biogeo-chemical significance is largely uncharacterized at this time
Dissolved and Particulate Organic MatterBacteria and archaea are thought to be by far the most impor-tant organisms with respect to the processing of dissolvedorganic matter (DOM) and nonliving particulate organicmatter (POM also called detritus) in the ocean While theremay be some uptake of DOM by protists particularly forgrowth factors needed in trace amounts such as vitamins(313) the bulk of this material is probably utilized by bacteriaand archaea (314) including the smallest cyanobacteriumProchlorococcus (315 316) which thus may be considered amixotroph Due to their small size bacteria have extremelyhigh surfacevolume ratios and combined with their over-whelmingly high abundance an extremely high integratedsurface area Protozoa tend to obtain the organic materialsthat they require for growth from their prey rather thanthrough the uptake of DOM Overall protozoa tend to besources of dissolved and detrital organic substances throughthe excretion of unassimilated prey biomass in expelledfood vacuoles
Particulate organic matter is not directly available as sub-strate to bacteria These substances must first be reduced tosmall molecules that can be transported into the cell Thisis accomplished by the production of extracellular enzymes(note that few if any large polymers are directly taken up bybacteria with the possible exception of DNA) Hydrolyticenzymes produced by bacteria (and almost certainly archaea)break down polymers like proteins polysaccharides andnucleic acids As with DOM POM is composed of a complexmixture of compounds that vary in their susceptibility to bac-terial degradation and utilization
Particulate material in the water column serves not only asbacterial substrate but also as substratum POM occurs in thewater column across a huge size spectrum from micrometersup to some detrital aggregates more than 1 m in diameter(317) Much of this particulate material is in a constant stateof flux with colloidal material constantly coalescing andaggregating to form new or larger particles (318) as microbialdegradation acts simultaneously to remineralize this materialDetrital particles that attain macroscopic size either by directformation (317 319 320) or via accretion and aggregation(321) and are often called marine snow or macroaggregatesMarine snow particles are readily colonized by bacteria and
422-18 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
many other microorganisms and their abundances on theseaggregates can be orders of magnitude greater than abundan-ces in an equivalent volume of the water surrounding theaggregates (320 322ndash324) and their diversity and highlyconcentrated abundances on detrital aggregates implies thatthere may be stronger trophic coupling and efficient energyelemental cycling relative to these processes among free-living microorganisms in the surrounding waters This specu-lation has resulted in a number of formalizations of the poten-tial impact of aggregate microbiology on biogeochemicalprocesses in the ocean (325 326) Most definitions of bio-films include aggregated particulate and these particles areoften studied within the biofilm conceptual model (see chap-ter by Lawrence et al in this section)
Inorganic Nutrients
Macronutrients (N P)Virtually all heterotrophic organisms in the ocean contributeto the pool of available macronutrients via the excretion ofmetabolic wastes and protozoa contribute significantly tothis process Macronutrients are often in excess within proto-zoan prey relative to the consumersrsquo needs because much ofthe prey carbon is respired to produce energy for metabolismand growth ldquoExcessrdquo nutrients are eliminated as eitherorganic compounds or more commonly as ammonium (forN) and phosphate (for P) The importance of the match(or mismatch) between predator and prey stoichiometry hasbeen demonstrated experimentally and repeatedly in marineand freshwater ecosystems and has been elevated to a funda-mental ecological principle governing elemental and energyflow in aquatic food webs (327) Based on this reasoningCaron et al (328) concluded that bacterivorous protozoaplay an important role in nutrient remineralization andrelease in the ocean Bacterial biomass is typically rich in Nand P relative to other organisms so bacterial predators expe-rience large excesses of these elements relative to their growthneedsMoreover bacterivorous protozoa should be dispropor-tionately important in nutrient remineralization amongsingle-celled eukaryotes because as the smallest protozoa inaquatic ecosystems they have high weight-specific metabolicrates (329) Thus both stoichiometric and allometric rela-tionships implicate small bacterivorous protozoa as impor-tant sources of remineralized nutrients in the plankton
The reasoning also holds true for bacterial utilization oforganic compounds and it has led to some interestingfindings and conclusions regarding the role of bacteria innutrient cycling in the ocean Bacteria are important sourcesremineralized nutrients when they are consuming N- orP-rich substrate Contrary to this traditional view howeverheterotrophic bacteria are often strong competitors for inor-ganic nutrients under many situations largely because bacte-rial biomass is richer in N and P than much of the organicmatter they consume in nature so they require additional Nand P to produce their biomass (summarized by [330]) Exper-imental studies have demonstrated the uptake of ammoniumby bacteria other than cyanobacteria using stable isotope 15N(331) and also the short-lived radioisotope 13N (332) oftenfinding that bacteria may be responsible for one third of the Nor P uptake Additionally there is evidence that bacteria insome oceans like the Sargasso Sea are limited by phosphorus(333 334) Growth limitation of bacteria by phosphorusappears to be a common phenomenon of freshwater ecosys-tems (335 336)
N2 fixation is restricted to prokaryotes and in the oceanwater column it was thought for many years to be done
primarily byTrichodesmium a warm-water colonial cyanobac-terium that blooms sporadically andRichelia a cyanobacterialsymbiont that lives within certain diatoms (337) Some uni-cellular cyanobacteria also contribute significantly to globalN2 fixation (338) and molecular biological data suggestthat a variety of other bacteriamdashnot phytoplanktonmdashmayalso be fixing nitrogen in seawater (339) An exciting recentdiscovery in this field is that a bacterial nitrogen fixer origi-nally known only from nifH sequences UCYN-A has hadits genome sequenced by parallel pyrosequencing and wasfound be a cyanobacterium (Candidatus Atelocyanobacteriumthalassa) lacking the apparatus to generate oxygen as well asother pathways (340) Further study showed it to be symbioticwith a marine alga (341)
Micronutrients (Trace Metals Growth Factors)Bacterial growth may often be limited by macronutrients asdescribed and sometimes by trace nutrients particularlyiron that occur at very low concentrations in seawater(342 343) The importance of Fe as an element limiting pri-mary production in some oceanic regions has come underclose scrutiny and extensive experimental investigation inrecent years Many bacteria have special uptake mechanismsto utilize extremely low concentrations of iron includingsiderophores (released compounds that bind iron and arethen specifically taken up) Even if bacteria are capable ofgrowth at low Fe concentrations growth may be less efficientKirchman et al (344) found that Fe limitation stronglyreduced bacterial growth efficiency in a cultured marine bac-terium (ca 50 efficiency Fe replete but lt10 when Fedeplete) Thus the effects may be complex Marine N2 fixa-tion also may be limited by the availability of Fe becausenitrogen fixers have high Fe requirements
Protozoa tend to be a source rather than a sink for Fe in theocean analogous to their role in macronutrient remineraliza-tion Protozoan grazing activity has been shown to releaseiron from bacterial biomass thereby relieving Fe limitationin co-occurring phytoplankton (345) a mechanistic demon-stration of the process using protozoa growing on Fe-repletebacteria However it has also been reported that Fe-limitedbacteria may contain insufficient Fe for the bacterivorous pro-tozoa that consume them thus leading to Fe limitation in theprotozoan as well (346)
OxygenHypoxic and anoxic regions of the ocean are expected to sig-nificantly expand in the coming decades as a consequence ofchanges in ocean stratification resulting from global climatechange (347) As oxygen becomes depleted bacteria havethe capability to utilize a series of alternate electron acceptorsCompositional changes in the bacterial assemblage accom-pany these changes in metabolic activity but are beyondthe scope of this chapter Many of the studies on benthicmicrobiology have focused on redox reactions that occurthere Oxygen is usually consumed by sediment microbialactivity much faster than the rate at which it can be replacedby diffusion so most sediments especially organically richones are anaerobic below a few mm (fine-grained muds) orcm (coarser sediments) from the sedimentwater interfaceexcept where animals ventilate the benthos through tubesWhen oxygen is absent bacteria and archaea use alternateelectron acceptors such as nitrate nitrite oxidized Fe orMn organic matter or sulfate The use of nitrate or nitriteas an electron acceptor with concomitant production of N2
gas (denitrification) results in net loss of biologically availablenitrogen Bacteria that perform this reaction are typically
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-19
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquofacultativerdquo organisms that can switch from oxygen to nitrateor nitrite Organic matter may be reduced by fermentationreactions in which part of an organic substrate is oxidizedwhile another is reduced fermenter organisms are also typi-cally facultative Sulfate is a particularly important alternateelectron acceptor in marine systems because it occurs as thesecond most abundant anion in seawater (behind chloride)typically 28 mM Its use as an electron acceptor leads to theproduction of elemental sulfur or sulfide The use of sulfateas an electron acceptor yields much less energy than oxygenor nitrate and sulfate-reducing bacteria are strict anaerobesunable to use oxygen (often strongly inhibited by oxygen)Methane can be produced by methanogenic archaea whichare all strict anaerobes using CO2 as an electron acceptor(see Findlay and Battin this section for further discussion)
Reduced compounds such as sulfide are produced by theuse of these alternate electron acceptors and they diffuse awayfrom their source When they diffuse to environments with astrong oxidant like oxygen they may be oxidized by chemo-trophic bacteria in energy-yielding reactions Similarlyammonium can be oxidized to nitrite and then nitrate by che-motrophic nitrifying bacteria and the nitrate can diffuse intoanaerobic layers and become denitrified to N2 Such coupledoxidation-reduction reactions can lead to accelerated biogeo-chemical cycling particularly in sediments
It was originally believed that relatively few free-livingspecies of protozoa inhabit anaerobic and hypoxic marineenvironments This view has changed based on (largely)molecular analyses of some of these environments (308348ndash351) A significant number of previously undescribedeukaryotic phylotypes have been observed in anoxic environ-ments High protozoan biodiversity in the absence of molec-ular oxygen is not completely unexpected because otheranaerobic environments such as some gut environments(eg cockroach guts and cattle rumen) have notoriouslyabundant and diverse protozoan fauna but marine anaerobicenvironments have been difficult to access and sample prop-erly Many anaerobic protozoa exist together with symbioticbacteria (352 353) or possess unique organelles such as thehydrogenosome (354)
MAJOR BIOTIC CONTROLSThe Trophic Activities of Protozoa Predation andTop-Down ControlsPredator-prey relationships are fundamental interactionswithin biological communities linking production of bio-mass on the one hand with removal on the other hand Estab-lishing the trophic relationships among microorganisms andquantifying the rates at which they take place constitutemajor endeavors within marine microbial ecology As notedpreviously protozoa consume a wide variety of prokaryoticand protistan prey Numerous laboratory and field measure-ments have been made throughout the past quarter centuryto attempt to quantify the rates of bacterial and phytoplank-ton consumption The methodology for accomplishing thesestudies has varied and a number of approaches are presentlyemployed
Methodologies for estimating microbial grazing are diversebut generally fall into one of two experimental approachesperturbation experiments or tracer experiments Perturbationexperiments (size fractionation metabolic inhibitors anddilution technique) rely on manipulation of the consumeror prey assemblages in somemanner such that the effect is evi-dent from changes in the abundance of the prey population
during incubations relative to control treatments that receiveno manipulation Tracer experiments (radioactively labeledprey fluorescently labeled prey stable isotopendashlabeled pray)attempt to use labeled prey or a prey proxy to quantitativelyfollow the movement of prey biomass into consumers Forsummaries of these methods see (74 355)
Perturbation experiments have entailed a variety of mech-anisms to ldquodecouplerdquo predators from prey in natural microbialcommunities For example size fractionation uses various sizefilters to remove an entire size class of grazers from awater sam-ple leaving the prey to grow in the relative absence of thosepredators during subsequent incubations Unfiltered samples(controls) indicate the growth of prey in the presence of pre-dation Predation impact is obtained by the difference ingrowth of the prey between these treatments This methodhas been used to examine predation on cyanobacteria heter-otrophic bacteria and minute phototrophic eukaryotesAlternatively metabolic inhibitors have been employed tohalt predation (or in some cases prokaryote growth) insteadof physical removal by filtration
The dilution technique (356) has become a commonmethod for examining grazing mortality of phytoplanktonby microzooplankton (which is comprised predominantly ofprotozoa) This method relies on the dilution of herbivorouszooplankton in a series of subsamples (and concomitantmeasurements of phytoplankton growth in each subsample)to estimate the grazing mortality of the herbivores
Tracer experiments have used both radioactively labeledprey and fluorescently labeled prey to examine predation onbacteria and phytoplankton Radioactive labeling has beenaccomplished using 14C- or 3H-labeled organic compounds(for bacteria) or 14C-bicarbonate (for phytoplankton) Theuse of radioisotope approaches has diminished with improve-ments in the number and types of fluorescent compoundsavailable for this work Fluorescently labeled prey havebeen labeled with a variety of compounds that allow theprey to be observed readily (even inside predators) by epi-fluorescencemicroscopy (357 358) Alternatively a decreasein the total number of fluorescent prey in a sample indicatesthe rate of removal of the prey assemblage
Both perturbation experiments and tracer experimentsdescribed have inherent advantages and disadvantages Forexample perturbation experiments are fairly straightforwardto conduct but they may introduce artifact by removingpotentially important sources of nutrients or organic com-pounds released by predators (359) On the other hand thedilution technique is very labor-intensive but one advantageof this method is that it provides the opportunity to determineboth phytoplankton growth and grazing mortality in a singleprocedure Most tracer experiments are fairly easy to conductand generally do not perturb the food web being studied butfeeding selectivity for or against the tracer (relative to naturalprey) may produce spurious results In addition some of thesemethods provide a measurement of the activity of the entiregrazer assemblage and others are applicable for examiningthe activity of specific taxa For example size fractionationapproaches examine the effect of the removal of an entiresize class of organisms (a community-levelrdquo measurement)whereas fluorescently labeled prey can be used to examinethe ingestion rate of a particular species by observing theuptake of labeled prey into that species (a species-levelmeasurement)
Predation rates of numerous marine protozoa have beenexamined in the laboratory From these experiments a gen-eral knowledge of the rates of consumption by these specieshas been accumulated Feeding in most protozoa shows a
422-20 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
functional response to the availability of prey that is similar tothe response of many organisms (although there are a greatdeal of species-specific differences) That is ingestion rateincreases rapidly as prey abundance increases until a maximalingestion rate is attained (360)Manyof these curves have theform of a Type II functional response curve (361)
Based on the features of these curves it is clear that proto-zoa have the ability to ingest substantial numbers of prokar-yote and eukaryote prey in natural ecosystems Somebacterivorous flagellates can consume bacteria (and cyano-bacteria) at rates up to 100 s per individual per hour whereassome bacterivorous ciliates can consume thousands of bacte-ria per individual per hour Rates of consumption of eukary-otic phytoplankton by herbivorous ciliates and largeflagellates are considerably more variable and highly depend-ent of the feeding mechanism (eg filtration in ciliates versuspseudopodial net in dinoflagellates) and the size of the prey
These lab studies demonstrate that rarely are prey suffi-ciently abundant in nature to support the maximal ingestionrates of which many protozoa are capable Nevertheless pre-dation rates of protozoa in nature are sufficient to constitute amajor factor controlling the standing stocks of bacteria andphytoplankton (63 180 184 355) For example these sum-maries indicate that on average protozoa consume 20ndash80ofthe standing stock of phytoplankton (measured as chloro-phyll) in the world ocean and 60ndash75 of daily primary pro-duction a departure from the classical view of copepods as theprimary consumers of phytoplankton This large role of her-bivorous protozoa infers a large role for copepods as consumersof planktonic protozoa (185) Similarly the role of protozoaas major consumers of bacterial biomass is indicated by therelative balance between bacterial production on the onehand and bacterivory on the other (Fig 8) The approximateone-to-one balance between these measurements over a largenumber of studies implies that protozoan bacterivory must bea major factor determining the fate of bacteria in marine (andfreshwater) plankton ecosystems
It is interesting to note that some types of bacteria includ-ing marine Bdellovibrio are known to prey on other marine
bacteria (362) At the time of this writing the importanceof bacterial predation by other bacteria within the contextof oceanographic processes is not known and presumed tobe small While Bdellovibrio may be cultivated from seawaterit can also be found among sequences from cold sediments(363) and the water column Bdellovibrio happens to be thebest studied predatory bacterium but there are no doubtothers that we simply have not yet identified (364)
Species-Specific Interactions (MutualismParasitism Commensalisms)Interactions between marine microorganisms such as compe-tition allelopathy and symbiosis are fundamental aspectscontrolling community composition and activity but atpresent most are poorly characterized in natural marine sys-tems Bacteria must compete for dissolved and particulatecompounds with other bacteria andwith protistan taxa whilephagotrophic protists must compete with each other andmetazoa for available prey These interactions manifest them-selves in many forms including true competition for availableresources resource partitioning such that some taxa specializeon the utilization of certain substrates or prey while othersspecialize on different ones (reducing competition) or eventhe cooperative use of compounds (eg one bacteriumrsquoswaste products being utilized as substrate by another bacte-rium) These metabolic consortia are probably common inthe ocean but can be difficult to identify and characterizeConsortia of bacteria working together to perform complexreactions especially in anaerobic systems are well known inmicrobiology
Consortia in which there is a very high degree of integra-tion between species (ie symbiosis) are very common in theocean These associations may be beneficial to both partners(mutualism) beneficial to one part and inconsequential forthe other (commensalism) or beneficial to one part and det-rimental to the other (parasitism) Many highly visible andwell-characterized marine mutualisms exist between chemo-synthetic bacteria and animals such as hydrothermal ventor methane seep tubeworms clams mussels or other inverte-brates (365) and also between luminescent bacteria and fishor squids (366) Similarly many protozoa form mutualisticassociations with photosynthetic protists (367) The mostconspicuous of these associations involved large amoeboidprotozoa of oceanic ecosystems (planktonic foraminiferanspolycystine radiolaria and acantharia) which contain largenumbers of intracellular algae (74) but associations betweensome heterotrophic dinoflagellates and cyanobacteria are alsocommon (368) Studies have indicated that efficient nutrientcycling in these protozoan-algal associations confers an eco-logical advantage for both partners in surfacewaters of the oli-gotrophic ocean
Parasitism is also a common symbiotic relationship amongmarine protozoa Protozoan parasites have been reported for adiverse array of protists and animals in the ocean Heterotro-phic dinoflagellates and presently uncultured alveolate cladesrelated to alveolates appear to be particularly successful atevolving thismode of existence (369ndash371) In a common par-asitic group Amoebophrya spp the parasite invades photo-synthetic dinoflagellates where it undergoes reproductioneventually lysing the host and releasing dozens of infectiveswarmers The parasitic protist Pirsonia spp is a parasite ofdiatoms (372) The overall importance of these parasitismsin the population dynamics of their hosts is not clearly knownbut may constitute a significant source of mortality for somespecies (373 374) The frequency and diversity of ldquonewrdquo
FIGURE 8 Meta-analysis of experimental studies that have com-pared rates of bacterial productivity (converted to population growthrate units of dminus1) with rates of bacterivory by phagotrophic protists(expressed as equivalent units of bacterial growth rate dminus1) Thesolid line indicates where measurements of bacterial removal by graz-ers were equal tomeasurements of bacterial production Data redrawnfrom Sanders et al (63) doi1011289781555818821ch422f8
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-21
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
Culture-Independent Diversity Studies
The first groups to be identified using cultivation-independ-ent molecular techniques from the marine plankton (121)were the bacterial SAR11 cluster (a group of closely relatedgene sequences or phylotypes) and marine picoplanktonicunicellular cyanobacteria Synechococcus and Prochlorococcus)Of these the SAR11 cluster was completely unknown butthe cyanobacteria had previously been recognized by theirunique pigment fluorescence Waterbury et al (122) andJohnson et al (123) used epifluorescence to observe Synecho-coccus and Chisholm et al (124) discovered Prochlorococcususing flow cytometry These cyanobacteria were later isolatedand grown in phytoplankton culturemedia These two groupsare generally common in the euphotic zone with SAR11 typ-ically comprising one third of the planktonic bacteria (125)and the cyanobacteria common everywhere but polar waters
Probably the biggest surprise to come from the applicationof molecular tools was the discovery by Fuhrman et al (126)of abundant archaea in the deep sea The archaea were foundto be in a unique phylogenetic cluster that was only distantlyrelated to any previously known archaea but the ldquoclosest rel-ativesrdquo (not really close at all) were extreme thermophiles Asubsequent study also found archaea to be present in near-surface coastal waters albeit relatively rare (lt2 relativeabundance) This study used a PCR technique specificallytargeted archaea and found ldquomarine Crenarchaeardquo plus a sec-ond group belonging to the phylum Euryarchaea (127)
Up until this time all known Archaea were thought to beldquoextremophilesrdquomdashadapted for either very high temperatures(thermophiles) extremely salty conditions (halophiles) orstrictly anaerobic environments (methanogens) Yet theseorganisms were present in cold or cool water at ordinary salin-ity and high oxygen concentrations Fluorescent in situhybridization (FISH) measurements from deep-sea sampleshave since showed that the archaea may make up appro-ximately 40 of the total countable prokaryotes with thepercentage reaching to 60 at 200 m depth in theMediterra-nean (35 36) An extensive time series of FISH measure-ments near Hawaii confirmed that the archaea are indeedvery abundant throughout the year from below the photiczone to at least 4000 m and typically constituting 30ndash40of the total prokaryotes present in waters deeper than a fewhundred meters (128) with similar results found elsewhere(129 130) (Fig 4) They have been reported from many pla-ces including the Atlantic Pacific and Southern Oceansand are dynamic components of the plankton the most com-mon type by far being the ldquomarine Crenarcheaeardquo (128 129131) While the marine archaea have been reported to bedominated by a few major ldquophylotypesrdquo (132) they alsohave been shown to have a great deal of microdiversity withinthese phylotypes suggesting there are many kinds of close rel-atives coexisting (133) Interestingly this group of archaeamay likely be the most abundant kind of organism on Earthgiven the huge volume of the deep sea and their high abun-dance there (134) It has recently been proposed that ldquomarineCrenarchaeardquo be elevated to their own major phylum outsidethe Crenarchaeota called the Thaumarchaeota on the basisof deep phylogenetic branching and fundamental differencesbetween them and the Crenarchaeota (135) The Thau-marchaeota possess the uniquely archaeal membrane lipidcrenarchaeol and are nowalso known to be abundant in soils
The physiology of the Thaumarchaeota has been anintriguing area of study Initially an autoradiography-FISHcombination approach demonstrated that they take upamino acids (130 136) But subsequent data showed that
crenarchaeol was derived largely from CO2 suggesting auto-trophic metabolism (137 138) An experiment showinguptake of 13C-labeled bicarbonate into these archaeal lipids(139) directly pointed to autotrophy in this group Chemoli-thoautotrophywas first hinted at byVenter et al (105) whosemetagenomic analysis showed an apparent archaeal scaffoldthat contained genes suggestive of ammonia oxidation Fur-ther evidence came from Schleper et al (140) who detectedseveral ammonia oxidation genes in order directly adjacent toa Thaumarchaeota 16S rRNA gene in a soil-derived metage-nomic clone The issue was directly resolved when Konnekeet al (141) isolated a related marine archaeon from sedimentof a marine aquarium and this organism Candidatus Nitroso-pumilus maritimus was found to have a chemoautotrophicmetabolism quantitatively oxidize ammonia to nitrite andcontain an archaeal ammonia monooxygenase gene (amoA)and interestingly did not grow heterotrophically Cultivationexperiments showed this organism has a high affinity forammonium allowing it to outcompete bacterial nitrifiers atlow (submicromolar) concentrations as are typical in thesea (142) The complete genome of the organism has shownnovel adaptations for nitrification and autotrophy (143)
Even if the Thaumarchaea are primarily chemolitho-autotrophs field data suggest a level of mixotrophy in thatsome organic substrates are being incorporated into biomassA stable isotope study using cells collected at 670 m depthoff Hawaii estimated that about 80 of the carbon incor-porated into archaea-specific lipids came from inorganic sour-ces and about 20 from organic compounds (144 145)Interestingly it has also been claimed that the genomesof deep-sea (gt2000 m depth) members of the Thaumarch-aea as well as those living in equatorial waters rarely containthe amoA gene and thus may be primarily chemoorgano-trophs (48)
Major bacterial groups that have been documentedfrom seawater using 16S rRNA characterization includesome that are also known from culture (eg AlteromonasRoseobacter) and several that are phylogenetically distantfrom standard cultures The most common groups are inrough order of their relative abundance in clone librariesfrom most to least abundant SAR11 (relatives of Pelagibac-ter ubique) Roseobacter SAR86 cyanobacteria SAR116SAR202 SAR234 andMarine Group A The SAR designa-tion followed by a number is an arbitrary sequential cloneidentifier from Sargasso Sea cloning studies done by the Gio-vannoni lab that did most of the early systematic cataloging ofclones Summaries of the data and phylogenetic relationshipsof these groups can be found in Fuhrman andHagstrom (102)and Giovannoni et al (101)
Molecular Genetic Discoveries in Bacterial andArchaeal Marine BiologyAs described earlier metagenomics is an extension of theideas used in the 16S rRNA cloning studies in that all genesfrom the native microorganisms are separated and clonedwithout having cultivated the organisms These methodsearly on started to find unexpected and very interestingresults The best examples involve unexpected marine photo-trophy the first of which is discovery of a nonchlorophyllphotosynthetic bacterial pigment called proteorhodopsin(146) The gene was found on a large environmentallyderived fragment of DNA that also had a gene coding for16S rRNA from the so-called SAR86 group (one ofthe groups common in seawater) This pigment can act as alight-driven proton pump thought to permit cells to generate
422-10 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
FIGURE 4 Distribution of Bacteria Euryarchaea andmarine Crenarchaea (recently renamed Thaumarchaea) along a transect in theNorthAtlantic Ocean as measured by CARD FISH and expressed as of total bacteria + archaea counts via DAPI stained epifluorescence fromTeira et al (130) Top panel shows station locations that are shown on the top of the lower three panels with the distance in km reported alongthe transect from lower to higher station numbers doi1011289781555818821ch422f4
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-11
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ATP from sunlight Interestingly different versions of this pig-ment are found at different depths apparently ldquotuningrdquo theabsorption to match the ambient wavelengths of light asthey change with depth (147) The proteorhodopsin genesare very widespread and diverse perhaps in half or more ofall marine bacteria including SAR11 and Euryarchaeota(105 148ndash150) However almost all of several culturedorganisms with proteorhodopsin studied to date do not showa growth benefit from light (150) with the exception of amember of the Flavobacteria Dokdonia sp strain MED134that grows faster in the light only under moderately lownutrient conditions (151) and a marine Vibrio has beenshown to survive starvation longer in the light than in dark-ness (152) These observation may explain the wide phyloge-netic distribution and high abundance of this gene in thatproteorhodopsin may often assist long-term survival of bacte-ria under extreme energy-limiting conditions yet not havemucheffect on growthduringmore energy-replete conditionsInterestingly a recent report suggests the growth benefit fromlight in Dokdonia MED134 is from enhanced uptake of itsrequired growth factor vitamin B1 which is transported by atonB-dependent transporter powered by a proton gradient(153) This pigment might also have sensory or other rolesnot yet well examined (150)
The second kind of phototrophy found by moleculargenetic and fluorescence techniques to be unexpectedlyimportant in marine plankton is anoxygenic aerobic bacterialphototrophy based on the pigment bacteriochlorophyll aCulturable aerobic anoxygenic photosynthetic (AAP) bacte-ria have been known from seawater for several years (154) andare widespread (155) Initial recent reports of direct countsclaimed they represent about 11 of the total bacterial com-munity in the euphotic zone (156) but those early counts didnot correct for the presence of other bacteria and such correc-tionoften yields estimates averaging closer to 2(157)Directmeasurements show the bacteriochlorophyll pigment is rela-tively rare (158) However studies in various ocean locationsincluding ocean gyres report that these AAP bacteriacan indeed on occasionmake up a quarter of the total prokar-yotes (159 160) A recent energetic modeling of AAP andproteorhodopsin-containing bacteria (161) has suggestedthat proteorhodopsin-containingones cangain approximately02asmuchenergy from sunlight asSynechococcus (commonmarine cyanobacterium) and AAP bacteria can gain approxi-mately 13 as much energy from sunlight as SynechococcusThey concluded the AAP bacteria may gain energy sufficientto meet maintenance costs but proteorhodopsin-containingbacteria were not expected to do so except at high light inten-sities andwith largenumbersof proteorhodopsinmoleculespercell They concluded the ease and low cost of maintainingPR-based phototrophy (a few genes required) may explainthe high incidence of proteorhodopsin genes
Other metagenomic studies have yielded interestinginsights about organisms and processes that would otherwisebe difficult or impossible to establish For example thesequence of an archaeal gene fragment isolated from seawaterrevealed extensive evidence of genetic exchange with othertypes of archaea and even bacteria (162) Such geneticexchange was also quite evident from the complete genesequences of different strains of Prochlorococcus and Synecho-coccus which was attributed in part to virus-mediated geneflow (163ndash166)
Dilution Cultures of Bacteria and ArchaeaSome recent cultivation techniques that permit growth ofldquotypicalrdquo oligotrophic marine bacteria in pure culture are
based on the simple concept that bacteria living in seawateron dissolved organic matter might best be grown in the labo-ratory in ordinary filtered seawater This conclusion followsfrom similar techniques to grow mixed ldquoseawater culturesrdquoof marine bacteria (167) but the seawater inoculum in thiscase is diluted so that only one or a few bacteria are addedto the initial culture vessel (168) A rapid throughput versionof this method has been used to cultivate members of bacte-rial groups thought to be conventionally uncultivable such asthe SAR11 clade (169 170) albeit often at very low den-sities Such cultures permit focused studies of the propertiesof those organisms including genomic analysis (169 170)This valuable work provides insight into the roles and activ-ities of some of the most common bacterial phylotypesobserved in the ocean (171) showing for example that culti-vated SAR11 requires a source of reduced sulfur as well asproviding templates aiding the interpretation of metage-nomic analysis (172)
PROTOZOADiversity and DistributionHeterotrophic protists have been identified historically fromtheir morphological features that are apparent at the levelof the light or electronmicroscope Features of cell size shapetype and pattern of flagellationciliation skeletal structurescharacteristics of the nucleus and other cellular structuresand organelles have been used to differentiate themany thou-sands of described species A description of the extent of thisdiversity of form and function is well beyond the scope of thisbook (see [79] for more information) However much of thisdiversity can be reduced to three basic body plans that dictatethe broadest ecological roles of these cells amoeboid flagel-lated and ciliated forms (Fig 5) Among the lineages possess-ing one of these three forms only the last group the ciliatedprotists constitute a monophyletic group within the domainEukarya
One of the simplest body plans for protozoa is the amoe-boid cell exemplified by the gymnamoebae or ldquonakedrdquoamoebae Motile nonphotosynthetic cells lacking flagellaor cilia occur as life stages in a number of phylogeneticallydiverse taxa but for many free-living species of protozoathis form constitutes the only life stage Motility is largelyconfined to movement along surfaces by means of pseudopo-dia that can take on a variety of (species-specific) shapes Sig-nificant abundances of amoebae are largely confined tobenthic and epibiotic environments (174) and to suspendedparticulate material where they can occasionally be highlyenriched (27) Most amoebae consume bacteria and otherminute prokaryotes and eukaryotes
Several heterotrophic protistan groups possess complexamoeboid body plans most notably the foraminifera polycys-tine and phaeodarian radiolaria and the acantharia Plank-tonic forms are predominantly oceanic in their distributions(although a large number of benthic species of foraminiferaexist) These species are heterotrophic and many are visibleto the naked eye (individual cells can bege1 cm gelatinouscolonies can form ribbon-like structuresge1 m in length)They feed on a wide variety of bacterial protistan and meta-zoan prey using pseudopodial networks have rather longcomplex life cycles for individual cells (weeks to months)and are extensively used in studies of paleoclimatologicalreconstruction (82 175) In addition many of these speciespossess intracellular symbiotic usually eukaryotic algaeThe widespread occurrence of algal endosymbiosis among
422-12 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
these protozoa in oceanic pelagic environments implies thatstrong selective forces appear to give rise to these associations(97 98)
Flagellated forms of protozoa exist across many protistanlineages Eukaryotic flagella come in various sizes numbersand forms (eg with or without tiny hairs) that are character-istic of the different lineagesMany flagellate species are appa-rently purely phototrophic or heterotrophic (phagotrophic)but a significant (still poorly known) fraction of flagellates aremixotrophic combining both phototrophy and phagotrophy(70 88) Flagella in phagotrophic flagellates are employed formotility and prey capture Most free-living flagellates possessone to four flagella (typically one or two) that can be manytimes the length of the cell itself Although there is tremen-dous species diversity among flagellated protozoa many ofthese species have broadly overlapping ecologies As a groupflagellates are the most numerically abundant protozoa ofboth benthic and pelagic ecosystems and they are fundamen-tally important as consumers of bacteria cyanobacteriaand other eukaryotes Flagellated protozoa within the nano-plankton size class (2ndash20 microm) are often counted as a singleassemblage in plankton studies because morphological details
apparent using light microscope are insufficient to distinguishamong the many species This assemblage has been variouslyreferred to by a variety of names and acronyms including het-erotrophic nanoplankton (HNAN HN) heterotrophic flag-ellates (Hflags) heterotrophic nanoflagellates (HNF) andmicroflagellates Heterotrophic nanoplankton is the mostaccurate term for most methodologies employed to countthese cells because it does not require visualization of flagella(which are often lost from these small cells during preparationfor microscopy) The confusion over terminology is partly aconsequence of the different methodologies that have beenused to count these species and partly a consequence of thefact that these species were first studied prior to the wideacceptance of the size convention of Sieburth et al (84)
Heterotrophic flagellates in the microplanktonic size class(20ndash200 microm mostly dinoflagellates) are important consum-ers of phytoplankton in pelagic ecosystems Many of thesespecies are capable of the production of large pseudopodialnets the pallium that can engulf prey (particularly diatoms)significantly larger than the diameter of the dinoflagellatetheca (176) This behavior and its ecological significance forenergy flow in plankton communities have been recognized
FIGURE 5 Body plans and size ranges of protozoa These micrographs depict amoeboid (andashg) flagellated (hndashj) and ciliated (kndashm) forms ofprotozoa From Caron et al (173) Markers bars are 5 (i) 10 (h) 20 (c j l) 30 (a) 50 (k m) 100 (b f) 500 (d) and 1000 (e g) micromdoi1011289781555818821ch422f5
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-13
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
only within the past few decades (177) These species areoften abundant in waters where diatoms dominate (178179) In such situations heterotrophic dinoflagellate biomasscan be similar to that of ciliated protozoa (180)
Ciliates are generally the most recognizable form of pro-tozoa to nonspecialists Nonetheless ciliates are a diversegroup morphologically as well as ecologically (181 182)The degree of ciliature on these species can range from uni-formly ciliated to totally devoid of cilia during most of theirlife cycle to ciliature restricted to specific regions of thecell In addition cilia can fuse to form various complexstructures (eg ciliary membranes or veils cirri) that assistin locomotion food capture or attachmentCiliates are abun-dant and ecologically important species in both benthic (8)and pelagic (183) ecosystems and can consume a variety ofprokaryotic and eukaryotic prey Together with the heterotro-phic dinoflagellates these species are the dominant consum-ers of phytoplankton in many pelagic ecosystems (184) andas a consequence they form an important trophic link to met-azoan zooplankton (185 186)
Diversity and Biogeography of Protists FromMorphology to DNA SequencesUntil relatively recently the biodiversity of marine protistanassemblages was not generally considered a controversialtopic It was generally accepted that while all species of pro-tists certainly have not been identified representatives ofmost types of algae and protozoa had been observed anddescribed if not actually brought into culture in the labora-tory Direct sequencing of 18S rRNA genes from environ-mental samples (as described in ldquoMolecular Phylogeny andMetagenomics Field Applicationsrdquo) has changed that viewInitial forays into environmental DNA indicated a muchgreater diversity of protists than previously documented usingclassical approaches of culture and microscopy (81 187ndash191) Numerous publications over the past decades haveexpanded these findings (see [192] for a recent global analy-sis) which are highly analogous to discoveries in marine pro-karyote research as described already
Previously uncharacterized protistan diversity has nowbeen documented at virtually every level of eukaryoticorganismal classification Some of these findings couldhave been expected but some have been very unexpectedFor example a much greater diversity than noted previouslyhas been observed among small (lt10 microm) protists in plank-tonic ecosystems These species generally possess few distinc-tive morphological features and one could expect that manycryptic species might be present among these small morpho-types The molecular ldquodiscoveryrdquo of this eukaryotic diversityhas stimulated progress on the isolation and descriptionof new species and genera of minute algae and protozoa(193ndash197)
Analyses of environmental samples have also indicatedthe unanticipated existence of novel 18S rRNA gene sequen-ces that imply the presence of novel lineages of eukaryotes innatural protistan communities (198ndash200) These are sequen-ces for which there are apparently no known described orcultured species The degree to which these sequences dif-fer from sequences of known sequenced eukaryotes impliesthat some of these lineages may be distinct at the level ofphylum (201)
These findings have raised basic questions and somedebate concerning the true diversity and biogeography ofprotistan assemblages in natural ecosystems (202ndash204) thevalidity of the many novel phylotypes or cryptic species that
are being documented (205) the potential importance andsignificance of the many rare taxa that characterize thesecommunities (206) and the implications of these findingsfor the ecologicalbiogeochemical roles that protists playin aquatic ecosystems For example it was postulated andsubsequently confirmed that some novel alveolate lineagesreported from marine ecosystems make up a suite of parasiticprotozoa whose ecological importance may have been signifi-cantly underestimated in the past (207) Deciphering theidentity and significance of these many unknown phylotypeswill constitute a significant effort for protistologists in thefuture (80 85)
Life Histories and Ecological Strategies of ProtozoaAbundance prey type and life histories all vary tremendouslyamongmarine protozoa The smallest species (eg many flag-ellates) tend to be the most abundant and widely distributedin the world ocean Indeed many of these species may beglobally distributed (208) Most nanoplanktonic flagellateshave potentially rapid rates of grazing and growth Underoptimal conditions these species can divide by binary fissionseveral times a day and thus dramatic increases in their pop-ulations can take place in response to favorable conditionswithin a few days (209) Many of these species are capableof surviving for limited periods of time without food andhave developed a variety of physiological or life cycle strat-egies to cope with these events (209) However in contrastto some bacteria that may remain viable through long periodsof starvation (210) protozoa will expire or encyst in responseto low food abundance and thus ldquoboom-and-bustrdquo popula-tion changes are characteristic of small flagellates
On the other end of the size spectrum from the ratherubiquitous nanoflagellate species many of the largest proto-zoan species (polycystine and phaeodarian radiolaria plank-tonic foraminifera) are exclusively oceanic (ie do notsurvive in most coastal environments) andor have specificlatitudinal and depth distributions Thus the abundancesof these latter species may range from undetectable to max-imal abundances of gt105 individualsm3 Dramatic changesin the abundance of these protozoan taxa also can be relatedto changes in prey abundance physicalbehavioral aggrega-tion or to periodicity of life cycle events For example theplanktonic foraminifer Hastigerina pelagica reproduces on alunar cycle and thus abundances (and life stages) of this spe-cies in oceanic waters can vary considerably over the courseof a month (211) In general life cycles for the large amoe-boid protozoa are lengthy and complex (for single-celledorganisms) with life spans unknown for many species(attempts to culture them in the lab have so far been unsuc-cessful) but estimated to be on the order of months to per-haps years (175)
The abundance and activities of microplanktonic hetero-trophic protists (mostly ciliates and heterotrophic dinoflagel-lates) tend to be somewhat intermediate to those ofnanoplanktonic flagellates and the larger amoeboid formsThese species are present in the majority of marine ecosys-tems and collectively play an important role in the controlof phytoplankton biomass (and probably the abundance ofnanoplanktonic protozoa although there is little informa-tion on this topic) in waters throughout the world oceanLike small flagellates ciliates and dinoflagellates reproduceprimarily by binary fission but their maximal growth ratesare typically slower (one division a day is typical) and theirlife cycles often include sexual phases that allow for geneticrecombination
422-14 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
VIRUSESViral Abundance and General PropertiesViruses are simple biological agents typically 20ndash200 nm indiameter composed of a nucleic acid genome in a proteincoat that infect cells and ldquocommandeerrdquo the cellrsquos machineryto make more viruses which are released into the environ-ment when the host cell lyses or bursts A particular virus isthought to be capable of infecting only a narrow range of hosts(usually one species sometimes a genus rarely broader)Although some early studies had isolated viruses from thesea therewere no data prior to the 1980s showing such viruseswere very abundant and more significantly no evidence thatinfection was occurring in any important part of the planktoncommunity It wasnrsquot until the late 1980s that electronmicro-scopy with suitable concentration methods showed thatviruses are extremely abundant similar to or even higherthan bacterial abundance (212 213) Interestingly themost suitable transmission electron microscopy (TEM)approach used by Bergh et al (described in detail by [214])was actually similar to a direct centrifugation method devel-oped in 1949 (215)
TEM studies of viruses permit high-resolution images andobservation of viral morphology showing features such ashead diameter tails sheaths and tail fibers (Fig 6 collage
in upper left) Each type of virus has a fixed morphology(unlike bacteria that are potentially more plastic in theirappearance) and hence a coarse measure of viral diversityhas been possible by cataloging morphologies Studies thathave done so have found dozens or more different morpholo-gies of marine viruses in a given sample (217ndash219) A largeproportion resemble bacteriophages which are viruses thatinfect bacteria
The availability of brightly fluorescent nucleic acid stainsand high-porosity fine pore size (002 μm) filters made fromAl2O3 have made it easy to accurately count viruses by epi-fluorescence microscopy (38 220 221) Epifluorescence per-mits abundance estimates but does not allow observation ofviral morphology as viruses are below the resolution limitof light microscopy viruses are visible only as sources of light(like stars in the night sky see Fig 6 lower micrograph) Anextension of manual epifluorescence counts is the use of flowcytometry to count viruses now commonly employed in somelabs (222)
Virus abundance has been found to be closely related tobacterial abundance with a virusbacteria ratio typically10ndash301 A typical oceanic profile of bacterial and viral abun-dance is shown in (Fig 6 panel on right) This tight ratioand the strong correlation to bacterial abundance in relationto weaker correlations to chlorophyll have been cited as
FIGURE 6 Marine viruses (upper left collage) viewed by transmission electron microscopy These are cyanophages in the groups myoviridae(right and bottom) siphoviridae (left) and podoviridae (top) Scale bars represent 100 nm From Sullivan (216) Epifluorescence micrograph(lower left) of SYBR green I stained viruses (small fluorescing objects) and bacteria + archaea (large fluorescing objects) Depth distribution ofvirus and bacteria + archaea abundances obtained by epifluorescence microscopy of SYBRGreen stained cells in the central San Pedro BasinCalifornia (11 August 2000) doi1011289781555818821ch422f6
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-15
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
evidence that most marine viruses infect bacteria rather thaneukaryotic phytoplankton (217 218 223 224) though thereare also many important viruses of phytoplankton and otherorganisms with significant impacts on primary productivityblooms and ecosystem function (225ndash227)
The first demonstration of the activity of marine viruseswas presented by Proctor and Fuhrman (228) who showedby electron microscopic examination that viruses wereactually infecting marine bacteria and cyanobacteria at ameasurable rate Because only the final portion of the viruslife cycle is visible by electron microscopy (when the virusesare assembled and ready to lyse the host cell) only a very smallfraction of the infected cells can be counted as infected at anygiven time Proctor and Fuhrman (228) reported that only afew percent of the total bacteria from coastal waters and theSargasso Sea were visibly infected but they interpreted thedata with a model that implied the actual fraction of the totalcommunity that was infected was much higher They con-cluded that the total fraction of bacterial mortality attribut-able to viruses is roughly 10ndash40 Subsequent studies haveused refined versions of that same general approach andmodel (229 230) and numerous studies have used a varietyof alternative approaches to estimate viral activitymdashall yield-ing basically the same general conclusion (although themodel parameters need to be adjusted for cyanobacteriawhich on average seem to be infected less than bacteria)These approaches include (a) calculation of virus turnoverfrom decay estimates (231) (b) measurement of viral DNAsynthesis by incorporation of tritiated thymidine (232) (c)observation of effects of added viruses (233 234) (d) estima-tion of bacterial mortality in the absence of protists (e) use offluorescent viruses to estimate production by an approachanalogous to isotope dilution (38) and (f) dilution of virusesin filtered samples to estimate viral production (235) Theoverall consensus of these studies is that the initial estimateswere basically correct that is that viruses are responsiblefor about 10ndash40 of the bacterial mortality in most marinesystems (reviewed by [218 223 226 236 237]) Howeverthere is still some disagreement about the higher estimatesand it is likely that 40 mortality from viruses alone is nottypical for most marine systems
Most of the foregoing work has focused on the viral infec-tion of bacteria primarily thought to be heterotrophicalthough the original report by Proctor and Fuhrman (228)also noted the occurrence of cyanobacterial infection Sev-eral subsequent studies focused on phytoplankton includingthe potential effect of viruses on the termination of phyto-plankton blooms of Emiliania and Phaeocystis and the likelyimpact this might have on release of climate-active gasessuch as dimethyl sulfide (225 227 238ndash247) Althoughdetails are beyond the scope of this chapter viruses arethought to infect virtually all marine organisms with poten-tially significant impacts from zooplankton to whales (226)Detailed studies of viruses infecting cyanobacteria such asSynechococcus have shown differences in viral host specific-ity particularly toward coastal and oceanic host strainsand some occasional high virus abundances (to 105ml) asmeasured by most probable number (MPN) cultivation tech-niques in the Gulf of Mexico near Texas (248 249) Othervirus cultivation studies with Prochlorococcus and Synecho-coccus in oligotrophic waters of the Sargasso Sea showedan interesting pattern of cross-infection between these gen-era by some virus types (suggesting gene flow among theseorganisms) but generally low MPN estimates of abundanceto 103ml even when cyanobacterial abundance was near105ml (216 250)
Overall the consensus emerging from direct comparisonsof viral-mediated mortality and grazer-mediated (ie micro-zooplankton) mortality have indicated that viral lysis of bac-teria constitutes a significant fraction of total mortality ofthis assemblage while the mortality of phytoplanktonappears to be dominated bymicrozooplankton inmost instan-ces (251 252)
As mentioned viruses have significant morphologicaldiversity as observed in TEM studies It is also possible toinvestigate aspects of their genetic diversity In early workon this topic Wommack et al (253) and Steward et al(254) observed the diversity of viral genome lengths in a fieldsample by pulsed field gel electrophoresis with viral genomesranging 25 kb to gt300 kb in length Field results show thatthe viral community composition is dynamic in space andtime with clear changes in the banding patterns over seasonsand locations in Chesapeake Bay (253) between oceanbasins and subsequent to dinoflagellate blooms (254) andwith depths to 500 m at one location (255)
Although viruses do not all share a set of core genesthat allows a universal viral phylogeny (comparable to SSUrRNA gene in cellular organisms) genetic diversity amonga single group of viruses can be examined by sequence analysisof shared genes within the group An example is the g20 geneinT4-like cyanophages that has shown extremely high diver-sity even among very closely related viruses and has demon-strated geographic and seasonal variation (256) A secondexample is g23 which is found broadly in diverse and wide-spread T4-like phages (257) and can have seasonally repeat-ing patterns (258) as well as short-term rapid dynamics thatcorrelate to those in bacteria (259)
Viral MetagenomicsAs with cellular organisms metagenomics can be used toexamine viral diversity and genetics without themany restric-tions of cultivation Viruses can be collected by selective fil-tration and concentrated by tangential flow filtration orflocculated with iron chloride (260 261) then their collec-tive metagenome can be extracted linker amplified and ana-lyzed by sequencing (262 263) Viral metagenomic studiesare particularly challenging because the large majority ofsequences have no annotated matches in any databases butso far results from marine samples around the world haveshown extremely high diversity and variations with depthlocation and time presumably with highly dispersed typesselected by local conditions (262ndash266) Because metage-nomes are best interpreted when there are representative cul-tures available the best matches of viral metagenomesoriginally tended to be to the few viral isolates infecting trulycommonmarine bacteria like cyanophage infecting Synecho-coccus and Prochlorococcus (267) However the developmentof dilution-to-extinction cultures representing common het-erotrophic marine taxa like Peligibacter (a member of theSAR11 clade) and SAR116 has allowed isolation from sea-water of viruses infecting these common organisms and theseisolates have indeed been found to be highly abundant inmarine viral metagenomes (268 269) New approaches tointerpret the results include clustering the proteins independ-ent of known proteins to compare samples to each other andtry to find environmental factors driving viral communitychanges (270) Such analysis of a large global data set (TaraOceans expedition) has shown that extensive sampling hascome close to reaching the total diversity in tropical and tem-perate waters of such viral protein clusters (which essentiallyrepresent various viral protein motifs not all viral proteintypes) and that viruses appear to be directionally dispersed
422-16 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquodownstreamrdquo in major ocean currents as one may expectfrom first principles but also supporting the ldquoseed bankrdquohypothesis of viral biogeography (271)
Viruses and Host DiversityViruses are themselves thought to be instrumental in drivingincreased diversity of their microbial hosts via a hypothesizedprocess often called ldquokill the winnerrdquo This is because viralinfection is host-specific and density-dependent the latterbecause viruses diffuse from host to host so an abundanthost is more likely to pass on infection than a rare oneThis means that if an organism becomes abundant andblooms winning the competition for resources it becomesmore susceptible to a viral epidemic This would benefit therarer organisms and thus help foster diversity (223 272273) However bloom scenarios as described above involvesystems far from steady state and the formal theory of Thing-stad and Lignell (274) has interesting steady-state solutionswhere several viruses infect several hosts stably over time(via trade-offs between growth rates and viral susceptibility)which may occur at the strain or species level (274) There issome experimental evidence that viruses have effects on nat-ural marine microbial community composition weaker thanthe kill the winner (bloom version) hypothesis would sug-gest (275 276) so something resembling the steady-statecoexistence described in the model may in fact be commonThere also appear to be processes that foster coexistencebetween viruses and hosts but the mechanisms are largelyspeculative (223 277 278)
Viruses may also be directly involved in host genetic diver-sity because they can be the agents of genetic exchangebetween microorganisms (223 273) This often involvesthe viral lifestyle known as lysogeny whereby viruses survivewithin host cells as DNA only integrated into the host chro-mosome and being reproduced each time the host divides Ahost harboring such a genome is called a lysogen becauseunder conditions of stress to the host cell a genetic switchmay cause the viral genome to initiate the lytic process pro-ducing many progeny viruses and bursting from the hostLysogeny is a very common property occurring in a signifi-cant part of the bacterial community (279 280) althoughthe incidence of induction of the lytic phase in nature is appa-rently low (281) Overall lysogeny is poorly understood butthought to have both positive and negative impacts on themicrobial community (282) Recent results suggest that ben-efits of being lysogenic in highly seasonal polar seas leads tofundamental difference between polar and other marine viralcommunities (283)
Viruses and the Microbial LoopAs part of the food web viruses occupy a unique positionThey infect host cells that are mostly thought to be heterotro-phic bacteria and by doing so they typically burst the hosts torelease progeny viruses and cellular debris But what is the fateof this material Viruses themselves do not last indefinitelyand a simple steady-state assumption implies that from eachburst of viruses (typically 20ndash100 per lytic event) only onesuccessfully infects another cell The rest are inactivatedand broken down by sunlight (UV and visible exposure)and enzymatic attack (284) or consumed byminute phagotro-phic protists (285) thus reentering the food web as substratefor bacteria or food for protistan consumers Experiments incontrolled laboratory systems and field studies with radioac-tively labeled viral lysis products have supported the conclu-sion that most of the organic matter released by the viralinfection is either taken up by bacteria or respired (286 287)
Modeling this process as part of the microbial loop showsthat viral lysis represent a sort of side loop that has the neteffect of remineralizing a significant amount of the carbonand nutrients that enter the bacteria-protist part of themicro-bial loop (Fig 7) A theoretical numerical steady-state modelcomparing a systemwith no viral activity to onewhere virusesare responsible for 50 of bacterial mortality showed that thesystem with viruses had 33 more bacterial production andrespiration than the virus-free system implying that theviruses had the effect of permitting the bacteria to processmore of the primary production than they would otherwise(223) Although 50 is a high number unlikely to be com-mon in the sea this model nevertheless illustrates that virusescan reduce the amount of energy reaching higher trophic lev-els The implication is that viruses lead to increased bacterialactivity at the expense of the larger organisms
MAJOR ENVIRONMENTAL CONTROLSLight Temperature and PressureTemperature has an important potential influence on bio-chemical reactions and therefore on biological processesin general Most ocean waters fall in the range of ndash2degC to30degC with obvious exceptions in hydrothermally heatedareas Temperature has long been known to be a regulatingfactor for the growth of heterotrophic microbes
In temperate waters it has been established that microbialactivity is generally much higher in warm summer waters thanin winter (55) The relationship is not simple howeverbecause multiple factors act at the same time Some contro-versy still exists regarding the highest and lowest extremesfor marine bacterial growth although there is broad agree-ment that bacteria grow gt100degC at hydrothermal vents andltminus5degC in sea ice brines Pomeroy et al (57) noted the inter-esting observation that bacteria seem particularly inhibitednear the freezing point of seawater (ca ndash22degC) comparedto eukaryotic phytoplankton This effect results in polarspring phytoplankton blooms that accumulate organic car-bon in advance of the response of the bacterial communityand development of the microbial loop and perhaps lead toenhanced benthic-pelagic coupling (288)
The relationship between temperature and the growth rateof marine phytoplankton was described broadly in a now
FIGURE 7 Modification of the microbial loop concept that incor-porates the functional role of viruses Export can be via predation orsinking From Fuhrman (223)doi1011289781555818821ch422f7
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-17
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
classic paper by Eppley (58) and later Goldman and Carpen-ter (289) Temperature was shown to exert a strong and directeffect on themaximal growth rates of these species withmax-imal intrinsic growth rates at 0degC generally lt1 divisiondaywhile growth rates at 30degC may be gt4day These relation-ships indicate the maximal rate that might be attained byphytoplankton at these temperatures but they do not takeother factors into account (nutrients light) While warmtemperature ostensibly allows more rapid growth it createshydrographic conditions that typically give rise to nutrientlimitation of algal growth Thus some of the coldest watersin nature witness some of the most massive phytoplanktonblooms (290) while warm oceanic gyres represent some ofthe most oligotrophic areas of the ocean The multiple andoften conflicting effects of temperature on the growth of nat-ural phytoplankton assemblages limit the accuracy of presentpredictions regarding how primary producers will respond toclimate change (291)
An analysis of the effect of temperature on the growth ofheterotrophic protists has indicated that the response is qual-itatively similar to that of phytoplankton but with a twistThe growth rates of at least some protozoa can exceed thoseof phytoplankton at warmer environmental temperaturesbut the opposite effect is apparent at very low environmentaltemperature This differential effect of temperature on thegrowth of phototrophic and heterotrophic protists was basedon a large meta-analysis of published protistan growth (59)That analysis demonstrated that the maximal growth ratesattained by phototrophic protists could exceed the maximalgrowth rates attained by heterotrophic protists (all otherpotential growth-limiting factors not considered) Thereforeprotozoan growth rates may be constrained to a greater degreeat low environmental temperature than rates for phytoplank-ton If so then phytoplankton bloomsmay get a head start ongrazers during spring in polar ecosystems This scenario is con-sistent with information on seasonal biomass changes andmicrozooplankton herbivory in the Ross Sea Antarctica(292 293) but there are still too few data to fully vet thishypothesis
The importance of high pressure on bacterial growthgained considerable attention in the late 1960s when thedeep sea submersible Alvin was accidentally lost overboardwith its hatch openWhile no lives were lost in this accidentsome workmenrsquos lunches sank to the bottom (sim1500 m)inside the submersible Alvin was recovered after 10 monthsand interestingly there was a waterlogged lunch containingapples bologna sandwiches and broken vacuum bottleswith broth that all appeared hardly degraded and tasted palat-able Yet when placed in a refrigerator on the ship these itemsdegraded relatively quickly Initially pressure was thought tobe the preserving factor as the sea floor temperature was sim-ilar to the refrigerator temperature and the only major differ-ence would be pressure (294) Following this observationa series of experiments to measure degradation of variousorganic materials left in the deep sea for extended periodsindicated that degradationwas typically significantly reducedimplying that pressure reduces the degradation rates (295)Nonetheless changes in the protozoan community of naturaldetrital material sinking to the deep ocean floor indicate thatthe microbial community can respond relatively quickly insome situations (296) Deep sea microbiology has advancedconsiderably yet it is still difficult to interpret results withrespect to actual in situ rates of naturally occurring organicmatter Deep sea bacteria adapted to high pressures havebeen isolated that are barophilic (also called piezophilic)meaning that they prefer high pressures and have reduced
activity at lower pressures whereas others are barotolerant(piezotolerant) tolerating but not preferring high pressuresfor example see (297 298)
Few data are available on barotolerantbarophilic marineprotozoa Protozoa certainly exist and grow at great oceanicdepths but measurements of in situ growth rates for these spe-cies do not yet exist Measurable protozoan numbers havebeen documented in the deep-sea sediments for more than30 years (299 300) and viable protozoa have occasionallybeen cultured from these environments (301ndash305) butvery few direct measurements of the activities of these speciesin situ have been reported (306) A few protozoa have beenisolated that will grow at high pressure (302 303 307) andprotists that appear to be unique to the deep ocean havebeen observed either directly or through the analysis ofDNA sequences (308ndash311) but possibly the best direct evi-dence that protozoan activity takes place at the high pressurescharacteristics of the deep sea are experimental and observa-tional work noting the stimulatory effect that detrital deposi-tion has on some components of the protozoan community(296 312) These observations indicate a diverse and activeprotozoan fauna of the deep ocean although their biogeo-chemical significance is largely uncharacterized at this time
Dissolved and Particulate Organic MatterBacteria and archaea are thought to be by far the most impor-tant organisms with respect to the processing of dissolvedorganic matter (DOM) and nonliving particulate organicmatter (POM also called detritus) in the ocean While theremay be some uptake of DOM by protists particularly forgrowth factors needed in trace amounts such as vitamins(313) the bulk of this material is probably utilized by bacteriaand archaea (314) including the smallest cyanobacteriumProchlorococcus (315 316) which thus may be considered amixotroph Due to their small size bacteria have extremelyhigh surfacevolume ratios and combined with their over-whelmingly high abundance an extremely high integratedsurface area Protozoa tend to obtain the organic materialsthat they require for growth from their prey rather thanthrough the uptake of DOM Overall protozoa tend to besources of dissolved and detrital organic substances throughthe excretion of unassimilated prey biomass in expelledfood vacuoles
Particulate organic matter is not directly available as sub-strate to bacteria These substances must first be reduced tosmall molecules that can be transported into the cell Thisis accomplished by the production of extracellular enzymes(note that few if any large polymers are directly taken up bybacteria with the possible exception of DNA) Hydrolyticenzymes produced by bacteria (and almost certainly archaea)break down polymers like proteins polysaccharides andnucleic acids As with DOM POM is composed of a complexmixture of compounds that vary in their susceptibility to bac-terial degradation and utilization
Particulate material in the water column serves not only asbacterial substrate but also as substratum POM occurs in thewater column across a huge size spectrum from micrometersup to some detrital aggregates more than 1 m in diameter(317) Much of this particulate material is in a constant stateof flux with colloidal material constantly coalescing andaggregating to form new or larger particles (318) as microbialdegradation acts simultaneously to remineralize this materialDetrital particles that attain macroscopic size either by directformation (317 319 320) or via accretion and aggregation(321) and are often called marine snow or macroaggregatesMarine snow particles are readily colonized by bacteria and
422-18 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
many other microorganisms and their abundances on theseaggregates can be orders of magnitude greater than abundan-ces in an equivalent volume of the water surrounding theaggregates (320 322ndash324) and their diversity and highlyconcentrated abundances on detrital aggregates implies thatthere may be stronger trophic coupling and efficient energyelemental cycling relative to these processes among free-living microorganisms in the surrounding waters This specu-lation has resulted in a number of formalizations of the poten-tial impact of aggregate microbiology on biogeochemicalprocesses in the ocean (325 326) Most definitions of bio-films include aggregated particulate and these particles areoften studied within the biofilm conceptual model (see chap-ter by Lawrence et al in this section)
Inorganic Nutrients
Macronutrients (N P)Virtually all heterotrophic organisms in the ocean contributeto the pool of available macronutrients via the excretion ofmetabolic wastes and protozoa contribute significantly tothis process Macronutrients are often in excess within proto-zoan prey relative to the consumersrsquo needs because much ofthe prey carbon is respired to produce energy for metabolismand growth ldquoExcessrdquo nutrients are eliminated as eitherorganic compounds or more commonly as ammonium (forN) and phosphate (for P) The importance of the match(or mismatch) between predator and prey stoichiometry hasbeen demonstrated experimentally and repeatedly in marineand freshwater ecosystems and has been elevated to a funda-mental ecological principle governing elemental and energyflow in aquatic food webs (327) Based on this reasoningCaron et al (328) concluded that bacterivorous protozoaplay an important role in nutrient remineralization andrelease in the ocean Bacterial biomass is typically rich in Nand P relative to other organisms so bacterial predators expe-rience large excesses of these elements relative to their growthneedsMoreover bacterivorous protozoa should be dispropor-tionately important in nutrient remineralization amongsingle-celled eukaryotes because as the smallest protozoa inaquatic ecosystems they have high weight-specific metabolicrates (329) Thus both stoichiometric and allometric rela-tionships implicate small bacterivorous protozoa as impor-tant sources of remineralized nutrients in the plankton
The reasoning also holds true for bacterial utilization oforganic compounds and it has led to some interestingfindings and conclusions regarding the role of bacteria innutrient cycling in the ocean Bacteria are important sourcesremineralized nutrients when they are consuming N- orP-rich substrate Contrary to this traditional view howeverheterotrophic bacteria are often strong competitors for inor-ganic nutrients under many situations largely because bacte-rial biomass is richer in N and P than much of the organicmatter they consume in nature so they require additional Nand P to produce their biomass (summarized by [330]) Exper-imental studies have demonstrated the uptake of ammoniumby bacteria other than cyanobacteria using stable isotope 15N(331) and also the short-lived radioisotope 13N (332) oftenfinding that bacteria may be responsible for one third of the Nor P uptake Additionally there is evidence that bacteria insome oceans like the Sargasso Sea are limited by phosphorus(333 334) Growth limitation of bacteria by phosphorusappears to be a common phenomenon of freshwater ecosys-tems (335 336)
N2 fixation is restricted to prokaryotes and in the oceanwater column it was thought for many years to be done
primarily byTrichodesmium a warm-water colonial cyanobac-terium that blooms sporadically andRichelia a cyanobacterialsymbiont that lives within certain diatoms (337) Some uni-cellular cyanobacteria also contribute significantly to globalN2 fixation (338) and molecular biological data suggestthat a variety of other bacteriamdashnot phytoplanktonmdashmayalso be fixing nitrogen in seawater (339) An exciting recentdiscovery in this field is that a bacterial nitrogen fixer origi-nally known only from nifH sequences UCYN-A has hadits genome sequenced by parallel pyrosequencing and wasfound be a cyanobacterium (Candidatus Atelocyanobacteriumthalassa) lacking the apparatus to generate oxygen as well asother pathways (340) Further study showed it to be symbioticwith a marine alga (341)
Micronutrients (Trace Metals Growth Factors)Bacterial growth may often be limited by macronutrients asdescribed and sometimes by trace nutrients particularlyiron that occur at very low concentrations in seawater(342 343) The importance of Fe as an element limiting pri-mary production in some oceanic regions has come underclose scrutiny and extensive experimental investigation inrecent years Many bacteria have special uptake mechanismsto utilize extremely low concentrations of iron includingsiderophores (released compounds that bind iron and arethen specifically taken up) Even if bacteria are capable ofgrowth at low Fe concentrations growth may be less efficientKirchman et al (344) found that Fe limitation stronglyreduced bacterial growth efficiency in a cultured marine bac-terium (ca 50 efficiency Fe replete but lt10 when Fedeplete) Thus the effects may be complex Marine N2 fixa-tion also may be limited by the availability of Fe becausenitrogen fixers have high Fe requirements
Protozoa tend to be a source rather than a sink for Fe in theocean analogous to their role in macronutrient remineraliza-tion Protozoan grazing activity has been shown to releaseiron from bacterial biomass thereby relieving Fe limitationin co-occurring phytoplankton (345) a mechanistic demon-stration of the process using protozoa growing on Fe-repletebacteria However it has also been reported that Fe-limitedbacteria may contain insufficient Fe for the bacterivorous pro-tozoa that consume them thus leading to Fe limitation in theprotozoan as well (346)
OxygenHypoxic and anoxic regions of the ocean are expected to sig-nificantly expand in the coming decades as a consequence ofchanges in ocean stratification resulting from global climatechange (347) As oxygen becomes depleted bacteria havethe capability to utilize a series of alternate electron acceptorsCompositional changes in the bacterial assemblage accom-pany these changes in metabolic activity but are beyondthe scope of this chapter Many of the studies on benthicmicrobiology have focused on redox reactions that occurthere Oxygen is usually consumed by sediment microbialactivity much faster than the rate at which it can be replacedby diffusion so most sediments especially organically richones are anaerobic below a few mm (fine-grained muds) orcm (coarser sediments) from the sedimentwater interfaceexcept where animals ventilate the benthos through tubesWhen oxygen is absent bacteria and archaea use alternateelectron acceptors such as nitrate nitrite oxidized Fe orMn organic matter or sulfate The use of nitrate or nitriteas an electron acceptor with concomitant production of N2
gas (denitrification) results in net loss of biologically availablenitrogen Bacteria that perform this reaction are typically
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-19
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquofacultativerdquo organisms that can switch from oxygen to nitrateor nitrite Organic matter may be reduced by fermentationreactions in which part of an organic substrate is oxidizedwhile another is reduced fermenter organisms are also typi-cally facultative Sulfate is a particularly important alternateelectron acceptor in marine systems because it occurs as thesecond most abundant anion in seawater (behind chloride)typically 28 mM Its use as an electron acceptor leads to theproduction of elemental sulfur or sulfide The use of sulfateas an electron acceptor yields much less energy than oxygenor nitrate and sulfate-reducing bacteria are strict anaerobesunable to use oxygen (often strongly inhibited by oxygen)Methane can be produced by methanogenic archaea whichare all strict anaerobes using CO2 as an electron acceptor(see Findlay and Battin this section for further discussion)
Reduced compounds such as sulfide are produced by theuse of these alternate electron acceptors and they diffuse awayfrom their source When they diffuse to environments with astrong oxidant like oxygen they may be oxidized by chemo-trophic bacteria in energy-yielding reactions Similarlyammonium can be oxidized to nitrite and then nitrate by che-motrophic nitrifying bacteria and the nitrate can diffuse intoanaerobic layers and become denitrified to N2 Such coupledoxidation-reduction reactions can lead to accelerated biogeo-chemical cycling particularly in sediments
It was originally believed that relatively few free-livingspecies of protozoa inhabit anaerobic and hypoxic marineenvironments This view has changed based on (largely)molecular analyses of some of these environments (308348ndash351) A significant number of previously undescribedeukaryotic phylotypes have been observed in anoxic environ-ments High protozoan biodiversity in the absence of molec-ular oxygen is not completely unexpected because otheranaerobic environments such as some gut environments(eg cockroach guts and cattle rumen) have notoriouslyabundant and diverse protozoan fauna but marine anaerobicenvironments have been difficult to access and sample prop-erly Many anaerobic protozoa exist together with symbioticbacteria (352 353) or possess unique organelles such as thehydrogenosome (354)
MAJOR BIOTIC CONTROLSThe Trophic Activities of Protozoa Predation andTop-Down ControlsPredator-prey relationships are fundamental interactionswithin biological communities linking production of bio-mass on the one hand with removal on the other hand Estab-lishing the trophic relationships among microorganisms andquantifying the rates at which they take place constitutemajor endeavors within marine microbial ecology As notedpreviously protozoa consume a wide variety of prokaryoticand protistan prey Numerous laboratory and field measure-ments have been made throughout the past quarter centuryto attempt to quantify the rates of bacterial and phytoplank-ton consumption The methodology for accomplishing thesestudies has varied and a number of approaches are presentlyemployed
Methodologies for estimating microbial grazing are diversebut generally fall into one of two experimental approachesperturbation experiments or tracer experiments Perturbationexperiments (size fractionation metabolic inhibitors anddilution technique) rely on manipulation of the consumeror prey assemblages in somemanner such that the effect is evi-dent from changes in the abundance of the prey population
during incubations relative to control treatments that receiveno manipulation Tracer experiments (radioactively labeledprey fluorescently labeled prey stable isotopendashlabeled pray)attempt to use labeled prey or a prey proxy to quantitativelyfollow the movement of prey biomass into consumers Forsummaries of these methods see (74 355)
Perturbation experiments have entailed a variety of mech-anisms to ldquodecouplerdquo predators from prey in natural microbialcommunities For example size fractionation uses various sizefilters to remove an entire size class of grazers from awater sam-ple leaving the prey to grow in the relative absence of thosepredators during subsequent incubations Unfiltered samples(controls) indicate the growth of prey in the presence of pre-dation Predation impact is obtained by the difference ingrowth of the prey between these treatments This methodhas been used to examine predation on cyanobacteria heter-otrophic bacteria and minute phototrophic eukaryotesAlternatively metabolic inhibitors have been employed tohalt predation (or in some cases prokaryote growth) insteadof physical removal by filtration
The dilution technique (356) has become a commonmethod for examining grazing mortality of phytoplanktonby microzooplankton (which is comprised predominantly ofprotozoa) This method relies on the dilution of herbivorouszooplankton in a series of subsamples (and concomitantmeasurements of phytoplankton growth in each subsample)to estimate the grazing mortality of the herbivores
Tracer experiments have used both radioactively labeledprey and fluorescently labeled prey to examine predation onbacteria and phytoplankton Radioactive labeling has beenaccomplished using 14C- or 3H-labeled organic compounds(for bacteria) or 14C-bicarbonate (for phytoplankton) Theuse of radioisotope approaches has diminished with improve-ments in the number and types of fluorescent compoundsavailable for this work Fluorescently labeled prey havebeen labeled with a variety of compounds that allow theprey to be observed readily (even inside predators) by epi-fluorescencemicroscopy (357 358) Alternatively a decreasein the total number of fluorescent prey in a sample indicatesthe rate of removal of the prey assemblage
Both perturbation experiments and tracer experimentsdescribed have inherent advantages and disadvantages Forexample perturbation experiments are fairly straightforwardto conduct but they may introduce artifact by removingpotentially important sources of nutrients or organic com-pounds released by predators (359) On the other hand thedilution technique is very labor-intensive but one advantageof this method is that it provides the opportunity to determineboth phytoplankton growth and grazing mortality in a singleprocedure Most tracer experiments are fairly easy to conductand generally do not perturb the food web being studied butfeeding selectivity for or against the tracer (relative to naturalprey) may produce spurious results In addition some of thesemethods provide a measurement of the activity of the entiregrazer assemblage and others are applicable for examiningthe activity of specific taxa For example size fractionationapproaches examine the effect of the removal of an entiresize class of organisms (a community-levelrdquo measurement)whereas fluorescently labeled prey can be used to examinethe ingestion rate of a particular species by observing theuptake of labeled prey into that species (a species-levelmeasurement)
Predation rates of numerous marine protozoa have beenexamined in the laboratory From these experiments a gen-eral knowledge of the rates of consumption by these specieshas been accumulated Feeding in most protozoa shows a
422-20 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
functional response to the availability of prey that is similar tothe response of many organisms (although there are a greatdeal of species-specific differences) That is ingestion rateincreases rapidly as prey abundance increases until a maximalingestion rate is attained (360)Manyof these curves have theform of a Type II functional response curve (361)
Based on the features of these curves it is clear that proto-zoa have the ability to ingest substantial numbers of prokar-yote and eukaryote prey in natural ecosystems Somebacterivorous flagellates can consume bacteria (and cyano-bacteria) at rates up to 100 s per individual per hour whereassome bacterivorous ciliates can consume thousands of bacte-ria per individual per hour Rates of consumption of eukary-otic phytoplankton by herbivorous ciliates and largeflagellates are considerably more variable and highly depend-ent of the feeding mechanism (eg filtration in ciliates versuspseudopodial net in dinoflagellates) and the size of the prey
These lab studies demonstrate that rarely are prey suffi-ciently abundant in nature to support the maximal ingestionrates of which many protozoa are capable Nevertheless pre-dation rates of protozoa in nature are sufficient to constitute amajor factor controlling the standing stocks of bacteria andphytoplankton (63 180 184 355) For example these sum-maries indicate that on average protozoa consume 20ndash80ofthe standing stock of phytoplankton (measured as chloro-phyll) in the world ocean and 60ndash75 of daily primary pro-duction a departure from the classical view of copepods as theprimary consumers of phytoplankton This large role of her-bivorous protozoa infers a large role for copepods as consumersof planktonic protozoa (185) Similarly the role of protozoaas major consumers of bacterial biomass is indicated by therelative balance between bacterial production on the onehand and bacterivory on the other (Fig 8) The approximateone-to-one balance between these measurements over a largenumber of studies implies that protozoan bacterivory must bea major factor determining the fate of bacteria in marine (andfreshwater) plankton ecosystems
It is interesting to note that some types of bacteria includ-ing marine Bdellovibrio are known to prey on other marine
bacteria (362) At the time of this writing the importanceof bacterial predation by other bacteria within the contextof oceanographic processes is not known and presumed tobe small While Bdellovibrio may be cultivated from seawaterit can also be found among sequences from cold sediments(363) and the water column Bdellovibrio happens to be thebest studied predatory bacterium but there are no doubtothers that we simply have not yet identified (364)
Species-Specific Interactions (MutualismParasitism Commensalisms)Interactions between marine microorganisms such as compe-tition allelopathy and symbiosis are fundamental aspectscontrolling community composition and activity but atpresent most are poorly characterized in natural marine sys-tems Bacteria must compete for dissolved and particulatecompounds with other bacteria andwith protistan taxa whilephagotrophic protists must compete with each other andmetazoa for available prey These interactions manifest them-selves in many forms including true competition for availableresources resource partitioning such that some taxa specializeon the utilization of certain substrates or prey while othersspecialize on different ones (reducing competition) or eventhe cooperative use of compounds (eg one bacteriumrsquoswaste products being utilized as substrate by another bacte-rium) These metabolic consortia are probably common inthe ocean but can be difficult to identify and characterizeConsortia of bacteria working together to perform complexreactions especially in anaerobic systems are well known inmicrobiology
Consortia in which there is a very high degree of integra-tion between species (ie symbiosis) are very common in theocean These associations may be beneficial to both partners(mutualism) beneficial to one part and inconsequential forthe other (commensalism) or beneficial to one part and det-rimental to the other (parasitism) Many highly visible andwell-characterized marine mutualisms exist between chemo-synthetic bacteria and animals such as hydrothermal ventor methane seep tubeworms clams mussels or other inverte-brates (365) and also between luminescent bacteria and fishor squids (366) Similarly many protozoa form mutualisticassociations with photosynthetic protists (367) The mostconspicuous of these associations involved large amoeboidprotozoa of oceanic ecosystems (planktonic foraminiferanspolycystine radiolaria and acantharia) which contain largenumbers of intracellular algae (74) but associations betweensome heterotrophic dinoflagellates and cyanobacteria are alsocommon (368) Studies have indicated that efficient nutrientcycling in these protozoan-algal associations confers an eco-logical advantage for both partners in surfacewaters of the oli-gotrophic ocean
Parasitism is also a common symbiotic relationship amongmarine protozoa Protozoan parasites have been reported for adiverse array of protists and animals in the ocean Heterotro-phic dinoflagellates and presently uncultured alveolate cladesrelated to alveolates appear to be particularly successful atevolving thismode of existence (369ndash371) In a common par-asitic group Amoebophrya spp the parasite invades photo-synthetic dinoflagellates where it undergoes reproductioneventually lysing the host and releasing dozens of infectiveswarmers The parasitic protist Pirsonia spp is a parasite ofdiatoms (372) The overall importance of these parasitismsin the population dynamics of their hosts is not clearly knownbut may constitute a significant source of mortality for somespecies (373 374) The frequency and diversity of ldquonewrdquo
FIGURE 8 Meta-analysis of experimental studies that have com-pared rates of bacterial productivity (converted to population growthrate units of dminus1) with rates of bacterivory by phagotrophic protists(expressed as equivalent units of bacterial growth rate dminus1) Thesolid line indicates where measurements of bacterial removal by graz-ers were equal tomeasurements of bacterial production Data redrawnfrom Sanders et al (63) doi1011289781555818821ch422f8
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-21
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
FIGURE 4 Distribution of Bacteria Euryarchaea andmarine Crenarchaea (recently renamed Thaumarchaea) along a transect in theNorthAtlantic Ocean as measured by CARD FISH and expressed as of total bacteria + archaea counts via DAPI stained epifluorescence fromTeira et al (130) Top panel shows station locations that are shown on the top of the lower three panels with the distance in km reported alongthe transect from lower to higher station numbers doi1011289781555818821ch422f4
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-11
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ATP from sunlight Interestingly different versions of this pig-ment are found at different depths apparently ldquotuningrdquo theabsorption to match the ambient wavelengths of light asthey change with depth (147) The proteorhodopsin genesare very widespread and diverse perhaps in half or more ofall marine bacteria including SAR11 and Euryarchaeota(105 148ndash150) However almost all of several culturedorganisms with proteorhodopsin studied to date do not showa growth benefit from light (150) with the exception of amember of the Flavobacteria Dokdonia sp strain MED134that grows faster in the light only under moderately lownutrient conditions (151) and a marine Vibrio has beenshown to survive starvation longer in the light than in dark-ness (152) These observation may explain the wide phyloge-netic distribution and high abundance of this gene in thatproteorhodopsin may often assist long-term survival of bacte-ria under extreme energy-limiting conditions yet not havemucheffect on growthduringmore energy-replete conditionsInterestingly a recent report suggests the growth benefit fromlight in Dokdonia MED134 is from enhanced uptake of itsrequired growth factor vitamin B1 which is transported by atonB-dependent transporter powered by a proton gradient(153) This pigment might also have sensory or other rolesnot yet well examined (150)
The second kind of phototrophy found by moleculargenetic and fluorescence techniques to be unexpectedlyimportant in marine plankton is anoxygenic aerobic bacterialphototrophy based on the pigment bacteriochlorophyll aCulturable aerobic anoxygenic photosynthetic (AAP) bacte-ria have been known from seawater for several years (154) andare widespread (155) Initial recent reports of direct countsclaimed they represent about 11 of the total bacterial com-munity in the euphotic zone (156) but those early counts didnot correct for the presence of other bacteria and such correc-tionoften yields estimates averaging closer to 2(157)Directmeasurements show the bacteriochlorophyll pigment is rela-tively rare (158) However studies in various ocean locationsincluding ocean gyres report that these AAP bacteriacan indeed on occasionmake up a quarter of the total prokar-yotes (159 160) A recent energetic modeling of AAP andproteorhodopsin-containing bacteria (161) has suggestedthat proteorhodopsin-containingones cangain approximately02asmuchenergy from sunlight asSynechococcus (commonmarine cyanobacterium) and AAP bacteria can gain approxi-mately 13 as much energy from sunlight as SynechococcusThey concluded the AAP bacteria may gain energy sufficientto meet maintenance costs but proteorhodopsin-containingbacteria were not expected to do so except at high light inten-sities andwith largenumbersof proteorhodopsinmoleculespercell They concluded the ease and low cost of maintainingPR-based phototrophy (a few genes required) may explainthe high incidence of proteorhodopsin genes
Other metagenomic studies have yielded interestinginsights about organisms and processes that would otherwisebe difficult or impossible to establish For example thesequence of an archaeal gene fragment isolated from seawaterrevealed extensive evidence of genetic exchange with othertypes of archaea and even bacteria (162) Such geneticexchange was also quite evident from the complete genesequences of different strains of Prochlorococcus and Synecho-coccus which was attributed in part to virus-mediated geneflow (163ndash166)
Dilution Cultures of Bacteria and ArchaeaSome recent cultivation techniques that permit growth ofldquotypicalrdquo oligotrophic marine bacteria in pure culture are
based on the simple concept that bacteria living in seawateron dissolved organic matter might best be grown in the labo-ratory in ordinary filtered seawater This conclusion followsfrom similar techniques to grow mixed ldquoseawater culturesrdquoof marine bacteria (167) but the seawater inoculum in thiscase is diluted so that only one or a few bacteria are addedto the initial culture vessel (168) A rapid throughput versionof this method has been used to cultivate members of bacte-rial groups thought to be conventionally uncultivable such asthe SAR11 clade (169 170) albeit often at very low den-sities Such cultures permit focused studies of the propertiesof those organisms including genomic analysis (169 170)This valuable work provides insight into the roles and activ-ities of some of the most common bacterial phylotypesobserved in the ocean (171) showing for example that culti-vated SAR11 requires a source of reduced sulfur as well asproviding templates aiding the interpretation of metage-nomic analysis (172)
PROTOZOADiversity and DistributionHeterotrophic protists have been identified historically fromtheir morphological features that are apparent at the levelof the light or electronmicroscope Features of cell size shapetype and pattern of flagellationciliation skeletal structurescharacteristics of the nucleus and other cellular structuresand organelles have been used to differentiate themany thou-sands of described species A description of the extent of thisdiversity of form and function is well beyond the scope of thisbook (see [79] for more information) However much of thisdiversity can be reduced to three basic body plans that dictatethe broadest ecological roles of these cells amoeboid flagel-lated and ciliated forms (Fig 5) Among the lineages possess-ing one of these three forms only the last group the ciliatedprotists constitute a monophyletic group within the domainEukarya
One of the simplest body plans for protozoa is the amoe-boid cell exemplified by the gymnamoebae or ldquonakedrdquoamoebae Motile nonphotosynthetic cells lacking flagellaor cilia occur as life stages in a number of phylogeneticallydiverse taxa but for many free-living species of protozoathis form constitutes the only life stage Motility is largelyconfined to movement along surfaces by means of pseudopo-dia that can take on a variety of (species-specific) shapes Sig-nificant abundances of amoebae are largely confined tobenthic and epibiotic environments (174) and to suspendedparticulate material where they can occasionally be highlyenriched (27) Most amoebae consume bacteria and otherminute prokaryotes and eukaryotes
Several heterotrophic protistan groups possess complexamoeboid body plans most notably the foraminifera polycys-tine and phaeodarian radiolaria and the acantharia Plank-tonic forms are predominantly oceanic in their distributions(although a large number of benthic species of foraminiferaexist) These species are heterotrophic and many are visibleto the naked eye (individual cells can bege1 cm gelatinouscolonies can form ribbon-like structuresge1 m in length)They feed on a wide variety of bacterial protistan and meta-zoan prey using pseudopodial networks have rather longcomplex life cycles for individual cells (weeks to months)and are extensively used in studies of paleoclimatologicalreconstruction (82 175) In addition many of these speciespossess intracellular symbiotic usually eukaryotic algaeThe widespread occurrence of algal endosymbiosis among
422-12 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
these protozoa in oceanic pelagic environments implies thatstrong selective forces appear to give rise to these associations(97 98)
Flagellated forms of protozoa exist across many protistanlineages Eukaryotic flagella come in various sizes numbersand forms (eg with or without tiny hairs) that are character-istic of the different lineagesMany flagellate species are appa-rently purely phototrophic or heterotrophic (phagotrophic)but a significant (still poorly known) fraction of flagellates aremixotrophic combining both phototrophy and phagotrophy(70 88) Flagella in phagotrophic flagellates are employed formotility and prey capture Most free-living flagellates possessone to four flagella (typically one or two) that can be manytimes the length of the cell itself Although there is tremen-dous species diversity among flagellated protozoa many ofthese species have broadly overlapping ecologies As a groupflagellates are the most numerically abundant protozoa ofboth benthic and pelagic ecosystems and they are fundamen-tally important as consumers of bacteria cyanobacteriaand other eukaryotes Flagellated protozoa within the nano-plankton size class (2ndash20 microm) are often counted as a singleassemblage in plankton studies because morphological details
apparent using light microscope are insufficient to distinguishamong the many species This assemblage has been variouslyreferred to by a variety of names and acronyms including het-erotrophic nanoplankton (HNAN HN) heterotrophic flag-ellates (Hflags) heterotrophic nanoflagellates (HNF) andmicroflagellates Heterotrophic nanoplankton is the mostaccurate term for most methodologies employed to countthese cells because it does not require visualization of flagella(which are often lost from these small cells during preparationfor microscopy) The confusion over terminology is partly aconsequence of the different methodologies that have beenused to count these species and partly a consequence of thefact that these species were first studied prior to the wideacceptance of the size convention of Sieburth et al (84)
Heterotrophic flagellates in the microplanktonic size class(20ndash200 microm mostly dinoflagellates) are important consum-ers of phytoplankton in pelagic ecosystems Many of thesespecies are capable of the production of large pseudopodialnets the pallium that can engulf prey (particularly diatoms)significantly larger than the diameter of the dinoflagellatetheca (176) This behavior and its ecological significance forenergy flow in plankton communities have been recognized
FIGURE 5 Body plans and size ranges of protozoa These micrographs depict amoeboid (andashg) flagellated (hndashj) and ciliated (kndashm) forms ofprotozoa From Caron et al (173) Markers bars are 5 (i) 10 (h) 20 (c j l) 30 (a) 50 (k m) 100 (b f) 500 (d) and 1000 (e g) micromdoi1011289781555818821ch422f5
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-13
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
only within the past few decades (177) These species areoften abundant in waters where diatoms dominate (178179) In such situations heterotrophic dinoflagellate biomasscan be similar to that of ciliated protozoa (180)
Ciliates are generally the most recognizable form of pro-tozoa to nonspecialists Nonetheless ciliates are a diversegroup morphologically as well as ecologically (181 182)The degree of ciliature on these species can range from uni-formly ciliated to totally devoid of cilia during most of theirlife cycle to ciliature restricted to specific regions of thecell In addition cilia can fuse to form various complexstructures (eg ciliary membranes or veils cirri) that assistin locomotion food capture or attachmentCiliates are abun-dant and ecologically important species in both benthic (8)and pelagic (183) ecosystems and can consume a variety ofprokaryotic and eukaryotic prey Together with the heterotro-phic dinoflagellates these species are the dominant consum-ers of phytoplankton in many pelagic ecosystems (184) andas a consequence they form an important trophic link to met-azoan zooplankton (185 186)
Diversity and Biogeography of Protists FromMorphology to DNA SequencesUntil relatively recently the biodiversity of marine protistanassemblages was not generally considered a controversialtopic It was generally accepted that while all species of pro-tists certainly have not been identified representatives ofmost types of algae and protozoa had been observed anddescribed if not actually brought into culture in the labora-tory Direct sequencing of 18S rRNA genes from environ-mental samples (as described in ldquoMolecular Phylogeny andMetagenomics Field Applicationsrdquo) has changed that viewInitial forays into environmental DNA indicated a muchgreater diversity of protists than previously documented usingclassical approaches of culture and microscopy (81 187ndash191) Numerous publications over the past decades haveexpanded these findings (see [192] for a recent global analy-sis) which are highly analogous to discoveries in marine pro-karyote research as described already
Previously uncharacterized protistan diversity has nowbeen documented at virtually every level of eukaryoticorganismal classification Some of these findings couldhave been expected but some have been very unexpectedFor example a much greater diversity than noted previouslyhas been observed among small (lt10 microm) protists in plank-tonic ecosystems These species generally possess few distinc-tive morphological features and one could expect that manycryptic species might be present among these small morpho-types The molecular ldquodiscoveryrdquo of this eukaryotic diversityhas stimulated progress on the isolation and descriptionof new species and genera of minute algae and protozoa(193ndash197)
Analyses of environmental samples have also indicatedthe unanticipated existence of novel 18S rRNA gene sequen-ces that imply the presence of novel lineages of eukaryotes innatural protistan communities (198ndash200) These are sequen-ces for which there are apparently no known described orcultured species The degree to which these sequences dif-fer from sequences of known sequenced eukaryotes impliesthat some of these lineages may be distinct at the level ofphylum (201)
These findings have raised basic questions and somedebate concerning the true diversity and biogeography ofprotistan assemblages in natural ecosystems (202ndash204) thevalidity of the many novel phylotypes or cryptic species that
are being documented (205) the potential importance andsignificance of the many rare taxa that characterize thesecommunities (206) and the implications of these findingsfor the ecologicalbiogeochemical roles that protists playin aquatic ecosystems For example it was postulated andsubsequently confirmed that some novel alveolate lineagesreported from marine ecosystems make up a suite of parasiticprotozoa whose ecological importance may have been signifi-cantly underestimated in the past (207) Deciphering theidentity and significance of these many unknown phylotypeswill constitute a significant effort for protistologists in thefuture (80 85)
Life Histories and Ecological Strategies of ProtozoaAbundance prey type and life histories all vary tremendouslyamongmarine protozoa The smallest species (eg many flag-ellates) tend to be the most abundant and widely distributedin the world ocean Indeed many of these species may beglobally distributed (208) Most nanoplanktonic flagellateshave potentially rapid rates of grazing and growth Underoptimal conditions these species can divide by binary fissionseveral times a day and thus dramatic increases in their pop-ulations can take place in response to favorable conditionswithin a few days (209) Many of these species are capableof surviving for limited periods of time without food andhave developed a variety of physiological or life cycle strat-egies to cope with these events (209) However in contrastto some bacteria that may remain viable through long periodsof starvation (210) protozoa will expire or encyst in responseto low food abundance and thus ldquoboom-and-bustrdquo popula-tion changes are characteristic of small flagellates
On the other end of the size spectrum from the ratherubiquitous nanoflagellate species many of the largest proto-zoan species (polycystine and phaeodarian radiolaria plank-tonic foraminifera) are exclusively oceanic (ie do notsurvive in most coastal environments) andor have specificlatitudinal and depth distributions Thus the abundancesof these latter species may range from undetectable to max-imal abundances of gt105 individualsm3 Dramatic changesin the abundance of these protozoan taxa also can be relatedto changes in prey abundance physicalbehavioral aggrega-tion or to periodicity of life cycle events For example theplanktonic foraminifer Hastigerina pelagica reproduces on alunar cycle and thus abundances (and life stages) of this spe-cies in oceanic waters can vary considerably over the courseof a month (211) In general life cycles for the large amoe-boid protozoa are lengthy and complex (for single-celledorganisms) with life spans unknown for many species(attempts to culture them in the lab have so far been unsuc-cessful) but estimated to be on the order of months to per-haps years (175)
The abundance and activities of microplanktonic hetero-trophic protists (mostly ciliates and heterotrophic dinoflagel-lates) tend to be somewhat intermediate to those ofnanoplanktonic flagellates and the larger amoeboid formsThese species are present in the majority of marine ecosys-tems and collectively play an important role in the controlof phytoplankton biomass (and probably the abundance ofnanoplanktonic protozoa although there is little informa-tion on this topic) in waters throughout the world oceanLike small flagellates ciliates and dinoflagellates reproduceprimarily by binary fission but their maximal growth ratesare typically slower (one division a day is typical) and theirlife cycles often include sexual phases that allow for geneticrecombination
422-14 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
VIRUSESViral Abundance and General PropertiesViruses are simple biological agents typically 20ndash200 nm indiameter composed of a nucleic acid genome in a proteincoat that infect cells and ldquocommandeerrdquo the cellrsquos machineryto make more viruses which are released into the environ-ment when the host cell lyses or bursts A particular virus isthought to be capable of infecting only a narrow range of hosts(usually one species sometimes a genus rarely broader)Although some early studies had isolated viruses from thesea therewere no data prior to the 1980s showing such viruseswere very abundant and more significantly no evidence thatinfection was occurring in any important part of the planktoncommunity It wasnrsquot until the late 1980s that electronmicro-scopy with suitable concentration methods showed thatviruses are extremely abundant similar to or even higherthan bacterial abundance (212 213) Interestingly themost suitable transmission electron microscopy (TEM)approach used by Bergh et al (described in detail by [214])was actually similar to a direct centrifugation method devel-oped in 1949 (215)
TEM studies of viruses permit high-resolution images andobservation of viral morphology showing features such ashead diameter tails sheaths and tail fibers (Fig 6 collage
in upper left) Each type of virus has a fixed morphology(unlike bacteria that are potentially more plastic in theirappearance) and hence a coarse measure of viral diversityhas been possible by cataloging morphologies Studies thathave done so have found dozens or more different morpholo-gies of marine viruses in a given sample (217ndash219) A largeproportion resemble bacteriophages which are viruses thatinfect bacteria
The availability of brightly fluorescent nucleic acid stainsand high-porosity fine pore size (002 μm) filters made fromAl2O3 have made it easy to accurately count viruses by epi-fluorescence microscopy (38 220 221) Epifluorescence per-mits abundance estimates but does not allow observation ofviral morphology as viruses are below the resolution limitof light microscopy viruses are visible only as sources of light(like stars in the night sky see Fig 6 lower micrograph) Anextension of manual epifluorescence counts is the use of flowcytometry to count viruses now commonly employed in somelabs (222)
Virus abundance has been found to be closely related tobacterial abundance with a virusbacteria ratio typically10ndash301 A typical oceanic profile of bacterial and viral abun-dance is shown in (Fig 6 panel on right) This tight ratioand the strong correlation to bacterial abundance in relationto weaker correlations to chlorophyll have been cited as
FIGURE 6 Marine viruses (upper left collage) viewed by transmission electron microscopy These are cyanophages in the groups myoviridae(right and bottom) siphoviridae (left) and podoviridae (top) Scale bars represent 100 nm From Sullivan (216) Epifluorescence micrograph(lower left) of SYBR green I stained viruses (small fluorescing objects) and bacteria + archaea (large fluorescing objects) Depth distribution ofvirus and bacteria + archaea abundances obtained by epifluorescence microscopy of SYBRGreen stained cells in the central San Pedro BasinCalifornia (11 August 2000) doi1011289781555818821ch422f6
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-15
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
evidence that most marine viruses infect bacteria rather thaneukaryotic phytoplankton (217 218 223 224) though thereare also many important viruses of phytoplankton and otherorganisms with significant impacts on primary productivityblooms and ecosystem function (225ndash227)
The first demonstration of the activity of marine viruseswas presented by Proctor and Fuhrman (228) who showedby electron microscopic examination that viruses wereactually infecting marine bacteria and cyanobacteria at ameasurable rate Because only the final portion of the viruslife cycle is visible by electron microscopy (when the virusesare assembled and ready to lyse the host cell) only a very smallfraction of the infected cells can be counted as infected at anygiven time Proctor and Fuhrman (228) reported that only afew percent of the total bacteria from coastal waters and theSargasso Sea were visibly infected but they interpreted thedata with a model that implied the actual fraction of the totalcommunity that was infected was much higher They con-cluded that the total fraction of bacterial mortality attribut-able to viruses is roughly 10ndash40 Subsequent studies haveused refined versions of that same general approach andmodel (229 230) and numerous studies have used a varietyof alternative approaches to estimate viral activitymdashall yield-ing basically the same general conclusion (although themodel parameters need to be adjusted for cyanobacteriawhich on average seem to be infected less than bacteria)These approaches include (a) calculation of virus turnoverfrom decay estimates (231) (b) measurement of viral DNAsynthesis by incorporation of tritiated thymidine (232) (c)observation of effects of added viruses (233 234) (d) estima-tion of bacterial mortality in the absence of protists (e) use offluorescent viruses to estimate production by an approachanalogous to isotope dilution (38) and (f) dilution of virusesin filtered samples to estimate viral production (235) Theoverall consensus of these studies is that the initial estimateswere basically correct that is that viruses are responsiblefor about 10ndash40 of the bacterial mortality in most marinesystems (reviewed by [218 223 226 236 237]) Howeverthere is still some disagreement about the higher estimatesand it is likely that 40 mortality from viruses alone is nottypical for most marine systems
Most of the foregoing work has focused on the viral infec-tion of bacteria primarily thought to be heterotrophicalthough the original report by Proctor and Fuhrman (228)also noted the occurrence of cyanobacterial infection Sev-eral subsequent studies focused on phytoplankton includingthe potential effect of viruses on the termination of phyto-plankton blooms of Emiliania and Phaeocystis and the likelyimpact this might have on release of climate-active gasessuch as dimethyl sulfide (225 227 238ndash247) Althoughdetails are beyond the scope of this chapter viruses arethought to infect virtually all marine organisms with poten-tially significant impacts from zooplankton to whales (226)Detailed studies of viruses infecting cyanobacteria such asSynechococcus have shown differences in viral host specific-ity particularly toward coastal and oceanic host strainsand some occasional high virus abundances (to 105ml) asmeasured by most probable number (MPN) cultivation tech-niques in the Gulf of Mexico near Texas (248 249) Othervirus cultivation studies with Prochlorococcus and Synecho-coccus in oligotrophic waters of the Sargasso Sea showedan interesting pattern of cross-infection between these gen-era by some virus types (suggesting gene flow among theseorganisms) but generally low MPN estimates of abundanceto 103ml even when cyanobacterial abundance was near105ml (216 250)
Overall the consensus emerging from direct comparisonsof viral-mediated mortality and grazer-mediated (ie micro-zooplankton) mortality have indicated that viral lysis of bac-teria constitutes a significant fraction of total mortality ofthis assemblage while the mortality of phytoplanktonappears to be dominated bymicrozooplankton inmost instan-ces (251 252)
As mentioned viruses have significant morphologicaldiversity as observed in TEM studies It is also possible toinvestigate aspects of their genetic diversity In early workon this topic Wommack et al (253) and Steward et al(254) observed the diversity of viral genome lengths in a fieldsample by pulsed field gel electrophoresis with viral genomesranging 25 kb to gt300 kb in length Field results show thatthe viral community composition is dynamic in space andtime with clear changes in the banding patterns over seasonsand locations in Chesapeake Bay (253) between oceanbasins and subsequent to dinoflagellate blooms (254) andwith depths to 500 m at one location (255)
Although viruses do not all share a set of core genesthat allows a universal viral phylogeny (comparable to SSUrRNA gene in cellular organisms) genetic diversity amonga single group of viruses can be examined by sequence analysisof shared genes within the group An example is the g20 geneinT4-like cyanophages that has shown extremely high diver-sity even among very closely related viruses and has demon-strated geographic and seasonal variation (256) A secondexample is g23 which is found broadly in diverse and wide-spread T4-like phages (257) and can have seasonally repeat-ing patterns (258) as well as short-term rapid dynamics thatcorrelate to those in bacteria (259)
Viral MetagenomicsAs with cellular organisms metagenomics can be used toexamine viral diversity and genetics without themany restric-tions of cultivation Viruses can be collected by selective fil-tration and concentrated by tangential flow filtration orflocculated with iron chloride (260 261) then their collec-tive metagenome can be extracted linker amplified and ana-lyzed by sequencing (262 263) Viral metagenomic studiesare particularly challenging because the large majority ofsequences have no annotated matches in any databases butso far results from marine samples around the world haveshown extremely high diversity and variations with depthlocation and time presumably with highly dispersed typesselected by local conditions (262ndash266) Because metage-nomes are best interpreted when there are representative cul-tures available the best matches of viral metagenomesoriginally tended to be to the few viral isolates infecting trulycommonmarine bacteria like cyanophage infecting Synecho-coccus and Prochlorococcus (267) However the developmentof dilution-to-extinction cultures representing common het-erotrophic marine taxa like Peligibacter (a member of theSAR11 clade) and SAR116 has allowed isolation from sea-water of viruses infecting these common organisms and theseisolates have indeed been found to be highly abundant inmarine viral metagenomes (268 269) New approaches tointerpret the results include clustering the proteins independ-ent of known proteins to compare samples to each other andtry to find environmental factors driving viral communitychanges (270) Such analysis of a large global data set (TaraOceans expedition) has shown that extensive sampling hascome close to reaching the total diversity in tropical and tem-perate waters of such viral protein clusters (which essentiallyrepresent various viral protein motifs not all viral proteintypes) and that viruses appear to be directionally dispersed
422-16 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquodownstreamrdquo in major ocean currents as one may expectfrom first principles but also supporting the ldquoseed bankrdquohypothesis of viral biogeography (271)
Viruses and Host DiversityViruses are themselves thought to be instrumental in drivingincreased diversity of their microbial hosts via a hypothesizedprocess often called ldquokill the winnerrdquo This is because viralinfection is host-specific and density-dependent the latterbecause viruses diffuse from host to host so an abundanthost is more likely to pass on infection than a rare oneThis means that if an organism becomes abundant andblooms winning the competition for resources it becomesmore susceptible to a viral epidemic This would benefit therarer organisms and thus help foster diversity (223 272273) However bloom scenarios as described above involvesystems far from steady state and the formal theory of Thing-stad and Lignell (274) has interesting steady-state solutionswhere several viruses infect several hosts stably over time(via trade-offs between growth rates and viral susceptibility)which may occur at the strain or species level (274) There issome experimental evidence that viruses have effects on nat-ural marine microbial community composition weaker thanthe kill the winner (bloom version) hypothesis would sug-gest (275 276) so something resembling the steady-statecoexistence described in the model may in fact be commonThere also appear to be processes that foster coexistencebetween viruses and hosts but the mechanisms are largelyspeculative (223 277 278)
Viruses may also be directly involved in host genetic diver-sity because they can be the agents of genetic exchangebetween microorganisms (223 273) This often involvesthe viral lifestyle known as lysogeny whereby viruses survivewithin host cells as DNA only integrated into the host chro-mosome and being reproduced each time the host divides Ahost harboring such a genome is called a lysogen becauseunder conditions of stress to the host cell a genetic switchmay cause the viral genome to initiate the lytic process pro-ducing many progeny viruses and bursting from the hostLysogeny is a very common property occurring in a signifi-cant part of the bacterial community (279 280) althoughthe incidence of induction of the lytic phase in nature is appa-rently low (281) Overall lysogeny is poorly understood butthought to have both positive and negative impacts on themicrobial community (282) Recent results suggest that ben-efits of being lysogenic in highly seasonal polar seas leads tofundamental difference between polar and other marine viralcommunities (283)
Viruses and the Microbial LoopAs part of the food web viruses occupy a unique positionThey infect host cells that are mostly thought to be heterotro-phic bacteria and by doing so they typically burst the hosts torelease progeny viruses and cellular debris But what is the fateof this material Viruses themselves do not last indefinitelyand a simple steady-state assumption implies that from eachburst of viruses (typically 20ndash100 per lytic event) only onesuccessfully infects another cell The rest are inactivatedand broken down by sunlight (UV and visible exposure)and enzymatic attack (284) or consumed byminute phagotro-phic protists (285) thus reentering the food web as substratefor bacteria or food for protistan consumers Experiments incontrolled laboratory systems and field studies with radioac-tively labeled viral lysis products have supported the conclu-sion that most of the organic matter released by the viralinfection is either taken up by bacteria or respired (286 287)
Modeling this process as part of the microbial loop showsthat viral lysis represent a sort of side loop that has the neteffect of remineralizing a significant amount of the carbonand nutrients that enter the bacteria-protist part of themicro-bial loop (Fig 7) A theoretical numerical steady-state modelcomparing a systemwith no viral activity to onewhere virusesare responsible for 50 of bacterial mortality showed that thesystem with viruses had 33 more bacterial production andrespiration than the virus-free system implying that theviruses had the effect of permitting the bacteria to processmore of the primary production than they would otherwise(223) Although 50 is a high number unlikely to be com-mon in the sea this model nevertheless illustrates that virusescan reduce the amount of energy reaching higher trophic lev-els The implication is that viruses lead to increased bacterialactivity at the expense of the larger organisms
MAJOR ENVIRONMENTAL CONTROLSLight Temperature and PressureTemperature has an important potential influence on bio-chemical reactions and therefore on biological processesin general Most ocean waters fall in the range of ndash2degC to30degC with obvious exceptions in hydrothermally heatedareas Temperature has long been known to be a regulatingfactor for the growth of heterotrophic microbes
In temperate waters it has been established that microbialactivity is generally much higher in warm summer waters thanin winter (55) The relationship is not simple howeverbecause multiple factors act at the same time Some contro-versy still exists regarding the highest and lowest extremesfor marine bacterial growth although there is broad agree-ment that bacteria grow gt100degC at hydrothermal vents andltminus5degC in sea ice brines Pomeroy et al (57) noted the inter-esting observation that bacteria seem particularly inhibitednear the freezing point of seawater (ca ndash22degC) comparedto eukaryotic phytoplankton This effect results in polarspring phytoplankton blooms that accumulate organic car-bon in advance of the response of the bacterial communityand development of the microbial loop and perhaps lead toenhanced benthic-pelagic coupling (288)
The relationship between temperature and the growth rateof marine phytoplankton was described broadly in a now
FIGURE 7 Modification of the microbial loop concept that incor-porates the functional role of viruses Export can be via predation orsinking From Fuhrman (223)doi1011289781555818821ch422f7
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-17
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
classic paper by Eppley (58) and later Goldman and Carpen-ter (289) Temperature was shown to exert a strong and directeffect on themaximal growth rates of these species withmax-imal intrinsic growth rates at 0degC generally lt1 divisiondaywhile growth rates at 30degC may be gt4day These relation-ships indicate the maximal rate that might be attained byphytoplankton at these temperatures but they do not takeother factors into account (nutrients light) While warmtemperature ostensibly allows more rapid growth it createshydrographic conditions that typically give rise to nutrientlimitation of algal growth Thus some of the coldest watersin nature witness some of the most massive phytoplanktonblooms (290) while warm oceanic gyres represent some ofthe most oligotrophic areas of the ocean The multiple andoften conflicting effects of temperature on the growth of nat-ural phytoplankton assemblages limit the accuracy of presentpredictions regarding how primary producers will respond toclimate change (291)
An analysis of the effect of temperature on the growth ofheterotrophic protists has indicated that the response is qual-itatively similar to that of phytoplankton but with a twistThe growth rates of at least some protozoa can exceed thoseof phytoplankton at warmer environmental temperaturesbut the opposite effect is apparent at very low environmentaltemperature This differential effect of temperature on thegrowth of phototrophic and heterotrophic protists was basedon a large meta-analysis of published protistan growth (59)That analysis demonstrated that the maximal growth ratesattained by phototrophic protists could exceed the maximalgrowth rates attained by heterotrophic protists (all otherpotential growth-limiting factors not considered) Thereforeprotozoan growth rates may be constrained to a greater degreeat low environmental temperature than rates for phytoplank-ton If so then phytoplankton bloomsmay get a head start ongrazers during spring in polar ecosystems This scenario is con-sistent with information on seasonal biomass changes andmicrozooplankton herbivory in the Ross Sea Antarctica(292 293) but there are still too few data to fully vet thishypothesis
The importance of high pressure on bacterial growthgained considerable attention in the late 1960s when thedeep sea submersible Alvin was accidentally lost overboardwith its hatch openWhile no lives were lost in this accidentsome workmenrsquos lunches sank to the bottom (sim1500 m)inside the submersible Alvin was recovered after 10 monthsand interestingly there was a waterlogged lunch containingapples bologna sandwiches and broken vacuum bottleswith broth that all appeared hardly degraded and tasted palat-able Yet when placed in a refrigerator on the ship these itemsdegraded relatively quickly Initially pressure was thought tobe the preserving factor as the sea floor temperature was sim-ilar to the refrigerator temperature and the only major differ-ence would be pressure (294) Following this observationa series of experiments to measure degradation of variousorganic materials left in the deep sea for extended periodsindicated that degradationwas typically significantly reducedimplying that pressure reduces the degradation rates (295)Nonetheless changes in the protozoan community of naturaldetrital material sinking to the deep ocean floor indicate thatthe microbial community can respond relatively quickly insome situations (296) Deep sea microbiology has advancedconsiderably yet it is still difficult to interpret results withrespect to actual in situ rates of naturally occurring organicmatter Deep sea bacteria adapted to high pressures havebeen isolated that are barophilic (also called piezophilic)meaning that they prefer high pressures and have reduced
activity at lower pressures whereas others are barotolerant(piezotolerant) tolerating but not preferring high pressuresfor example see (297 298)
Few data are available on barotolerantbarophilic marineprotozoa Protozoa certainly exist and grow at great oceanicdepths but measurements of in situ growth rates for these spe-cies do not yet exist Measurable protozoan numbers havebeen documented in the deep-sea sediments for more than30 years (299 300) and viable protozoa have occasionallybeen cultured from these environments (301ndash305) butvery few direct measurements of the activities of these speciesin situ have been reported (306) A few protozoa have beenisolated that will grow at high pressure (302 303 307) andprotists that appear to be unique to the deep ocean havebeen observed either directly or through the analysis ofDNA sequences (308ndash311) but possibly the best direct evi-dence that protozoan activity takes place at the high pressurescharacteristics of the deep sea are experimental and observa-tional work noting the stimulatory effect that detrital deposi-tion has on some components of the protozoan community(296 312) These observations indicate a diverse and activeprotozoan fauna of the deep ocean although their biogeo-chemical significance is largely uncharacterized at this time
Dissolved and Particulate Organic MatterBacteria and archaea are thought to be by far the most impor-tant organisms with respect to the processing of dissolvedorganic matter (DOM) and nonliving particulate organicmatter (POM also called detritus) in the ocean While theremay be some uptake of DOM by protists particularly forgrowth factors needed in trace amounts such as vitamins(313) the bulk of this material is probably utilized by bacteriaand archaea (314) including the smallest cyanobacteriumProchlorococcus (315 316) which thus may be considered amixotroph Due to their small size bacteria have extremelyhigh surfacevolume ratios and combined with their over-whelmingly high abundance an extremely high integratedsurface area Protozoa tend to obtain the organic materialsthat they require for growth from their prey rather thanthrough the uptake of DOM Overall protozoa tend to besources of dissolved and detrital organic substances throughthe excretion of unassimilated prey biomass in expelledfood vacuoles
Particulate organic matter is not directly available as sub-strate to bacteria These substances must first be reduced tosmall molecules that can be transported into the cell Thisis accomplished by the production of extracellular enzymes(note that few if any large polymers are directly taken up bybacteria with the possible exception of DNA) Hydrolyticenzymes produced by bacteria (and almost certainly archaea)break down polymers like proteins polysaccharides andnucleic acids As with DOM POM is composed of a complexmixture of compounds that vary in their susceptibility to bac-terial degradation and utilization
Particulate material in the water column serves not only asbacterial substrate but also as substratum POM occurs in thewater column across a huge size spectrum from micrometersup to some detrital aggregates more than 1 m in diameter(317) Much of this particulate material is in a constant stateof flux with colloidal material constantly coalescing andaggregating to form new or larger particles (318) as microbialdegradation acts simultaneously to remineralize this materialDetrital particles that attain macroscopic size either by directformation (317 319 320) or via accretion and aggregation(321) and are often called marine snow or macroaggregatesMarine snow particles are readily colonized by bacteria and
422-18 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
many other microorganisms and their abundances on theseaggregates can be orders of magnitude greater than abundan-ces in an equivalent volume of the water surrounding theaggregates (320 322ndash324) and their diversity and highlyconcentrated abundances on detrital aggregates implies thatthere may be stronger trophic coupling and efficient energyelemental cycling relative to these processes among free-living microorganisms in the surrounding waters This specu-lation has resulted in a number of formalizations of the poten-tial impact of aggregate microbiology on biogeochemicalprocesses in the ocean (325 326) Most definitions of bio-films include aggregated particulate and these particles areoften studied within the biofilm conceptual model (see chap-ter by Lawrence et al in this section)
Inorganic Nutrients
Macronutrients (N P)Virtually all heterotrophic organisms in the ocean contributeto the pool of available macronutrients via the excretion ofmetabolic wastes and protozoa contribute significantly tothis process Macronutrients are often in excess within proto-zoan prey relative to the consumersrsquo needs because much ofthe prey carbon is respired to produce energy for metabolismand growth ldquoExcessrdquo nutrients are eliminated as eitherorganic compounds or more commonly as ammonium (forN) and phosphate (for P) The importance of the match(or mismatch) between predator and prey stoichiometry hasbeen demonstrated experimentally and repeatedly in marineand freshwater ecosystems and has been elevated to a funda-mental ecological principle governing elemental and energyflow in aquatic food webs (327) Based on this reasoningCaron et al (328) concluded that bacterivorous protozoaplay an important role in nutrient remineralization andrelease in the ocean Bacterial biomass is typically rich in Nand P relative to other organisms so bacterial predators expe-rience large excesses of these elements relative to their growthneedsMoreover bacterivorous protozoa should be dispropor-tionately important in nutrient remineralization amongsingle-celled eukaryotes because as the smallest protozoa inaquatic ecosystems they have high weight-specific metabolicrates (329) Thus both stoichiometric and allometric rela-tionships implicate small bacterivorous protozoa as impor-tant sources of remineralized nutrients in the plankton
The reasoning also holds true for bacterial utilization oforganic compounds and it has led to some interestingfindings and conclusions regarding the role of bacteria innutrient cycling in the ocean Bacteria are important sourcesremineralized nutrients when they are consuming N- orP-rich substrate Contrary to this traditional view howeverheterotrophic bacteria are often strong competitors for inor-ganic nutrients under many situations largely because bacte-rial biomass is richer in N and P than much of the organicmatter they consume in nature so they require additional Nand P to produce their biomass (summarized by [330]) Exper-imental studies have demonstrated the uptake of ammoniumby bacteria other than cyanobacteria using stable isotope 15N(331) and also the short-lived radioisotope 13N (332) oftenfinding that bacteria may be responsible for one third of the Nor P uptake Additionally there is evidence that bacteria insome oceans like the Sargasso Sea are limited by phosphorus(333 334) Growth limitation of bacteria by phosphorusappears to be a common phenomenon of freshwater ecosys-tems (335 336)
N2 fixation is restricted to prokaryotes and in the oceanwater column it was thought for many years to be done
primarily byTrichodesmium a warm-water colonial cyanobac-terium that blooms sporadically andRichelia a cyanobacterialsymbiont that lives within certain diatoms (337) Some uni-cellular cyanobacteria also contribute significantly to globalN2 fixation (338) and molecular biological data suggestthat a variety of other bacteriamdashnot phytoplanktonmdashmayalso be fixing nitrogen in seawater (339) An exciting recentdiscovery in this field is that a bacterial nitrogen fixer origi-nally known only from nifH sequences UCYN-A has hadits genome sequenced by parallel pyrosequencing and wasfound be a cyanobacterium (Candidatus Atelocyanobacteriumthalassa) lacking the apparatus to generate oxygen as well asother pathways (340) Further study showed it to be symbioticwith a marine alga (341)
Micronutrients (Trace Metals Growth Factors)Bacterial growth may often be limited by macronutrients asdescribed and sometimes by trace nutrients particularlyiron that occur at very low concentrations in seawater(342 343) The importance of Fe as an element limiting pri-mary production in some oceanic regions has come underclose scrutiny and extensive experimental investigation inrecent years Many bacteria have special uptake mechanismsto utilize extremely low concentrations of iron includingsiderophores (released compounds that bind iron and arethen specifically taken up) Even if bacteria are capable ofgrowth at low Fe concentrations growth may be less efficientKirchman et al (344) found that Fe limitation stronglyreduced bacterial growth efficiency in a cultured marine bac-terium (ca 50 efficiency Fe replete but lt10 when Fedeplete) Thus the effects may be complex Marine N2 fixa-tion also may be limited by the availability of Fe becausenitrogen fixers have high Fe requirements
Protozoa tend to be a source rather than a sink for Fe in theocean analogous to their role in macronutrient remineraliza-tion Protozoan grazing activity has been shown to releaseiron from bacterial biomass thereby relieving Fe limitationin co-occurring phytoplankton (345) a mechanistic demon-stration of the process using protozoa growing on Fe-repletebacteria However it has also been reported that Fe-limitedbacteria may contain insufficient Fe for the bacterivorous pro-tozoa that consume them thus leading to Fe limitation in theprotozoan as well (346)
OxygenHypoxic and anoxic regions of the ocean are expected to sig-nificantly expand in the coming decades as a consequence ofchanges in ocean stratification resulting from global climatechange (347) As oxygen becomes depleted bacteria havethe capability to utilize a series of alternate electron acceptorsCompositional changes in the bacterial assemblage accom-pany these changes in metabolic activity but are beyondthe scope of this chapter Many of the studies on benthicmicrobiology have focused on redox reactions that occurthere Oxygen is usually consumed by sediment microbialactivity much faster than the rate at which it can be replacedby diffusion so most sediments especially organically richones are anaerobic below a few mm (fine-grained muds) orcm (coarser sediments) from the sedimentwater interfaceexcept where animals ventilate the benthos through tubesWhen oxygen is absent bacteria and archaea use alternateelectron acceptors such as nitrate nitrite oxidized Fe orMn organic matter or sulfate The use of nitrate or nitriteas an electron acceptor with concomitant production of N2
gas (denitrification) results in net loss of biologically availablenitrogen Bacteria that perform this reaction are typically
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-19
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquofacultativerdquo organisms that can switch from oxygen to nitrateor nitrite Organic matter may be reduced by fermentationreactions in which part of an organic substrate is oxidizedwhile another is reduced fermenter organisms are also typi-cally facultative Sulfate is a particularly important alternateelectron acceptor in marine systems because it occurs as thesecond most abundant anion in seawater (behind chloride)typically 28 mM Its use as an electron acceptor leads to theproduction of elemental sulfur or sulfide The use of sulfateas an electron acceptor yields much less energy than oxygenor nitrate and sulfate-reducing bacteria are strict anaerobesunable to use oxygen (often strongly inhibited by oxygen)Methane can be produced by methanogenic archaea whichare all strict anaerobes using CO2 as an electron acceptor(see Findlay and Battin this section for further discussion)
Reduced compounds such as sulfide are produced by theuse of these alternate electron acceptors and they diffuse awayfrom their source When they diffuse to environments with astrong oxidant like oxygen they may be oxidized by chemo-trophic bacteria in energy-yielding reactions Similarlyammonium can be oxidized to nitrite and then nitrate by che-motrophic nitrifying bacteria and the nitrate can diffuse intoanaerobic layers and become denitrified to N2 Such coupledoxidation-reduction reactions can lead to accelerated biogeo-chemical cycling particularly in sediments
It was originally believed that relatively few free-livingspecies of protozoa inhabit anaerobic and hypoxic marineenvironments This view has changed based on (largely)molecular analyses of some of these environments (308348ndash351) A significant number of previously undescribedeukaryotic phylotypes have been observed in anoxic environ-ments High protozoan biodiversity in the absence of molec-ular oxygen is not completely unexpected because otheranaerobic environments such as some gut environments(eg cockroach guts and cattle rumen) have notoriouslyabundant and diverse protozoan fauna but marine anaerobicenvironments have been difficult to access and sample prop-erly Many anaerobic protozoa exist together with symbioticbacteria (352 353) or possess unique organelles such as thehydrogenosome (354)
MAJOR BIOTIC CONTROLSThe Trophic Activities of Protozoa Predation andTop-Down ControlsPredator-prey relationships are fundamental interactionswithin biological communities linking production of bio-mass on the one hand with removal on the other hand Estab-lishing the trophic relationships among microorganisms andquantifying the rates at which they take place constitutemajor endeavors within marine microbial ecology As notedpreviously protozoa consume a wide variety of prokaryoticand protistan prey Numerous laboratory and field measure-ments have been made throughout the past quarter centuryto attempt to quantify the rates of bacterial and phytoplank-ton consumption The methodology for accomplishing thesestudies has varied and a number of approaches are presentlyemployed
Methodologies for estimating microbial grazing are diversebut generally fall into one of two experimental approachesperturbation experiments or tracer experiments Perturbationexperiments (size fractionation metabolic inhibitors anddilution technique) rely on manipulation of the consumeror prey assemblages in somemanner such that the effect is evi-dent from changes in the abundance of the prey population
during incubations relative to control treatments that receiveno manipulation Tracer experiments (radioactively labeledprey fluorescently labeled prey stable isotopendashlabeled pray)attempt to use labeled prey or a prey proxy to quantitativelyfollow the movement of prey biomass into consumers Forsummaries of these methods see (74 355)
Perturbation experiments have entailed a variety of mech-anisms to ldquodecouplerdquo predators from prey in natural microbialcommunities For example size fractionation uses various sizefilters to remove an entire size class of grazers from awater sam-ple leaving the prey to grow in the relative absence of thosepredators during subsequent incubations Unfiltered samples(controls) indicate the growth of prey in the presence of pre-dation Predation impact is obtained by the difference ingrowth of the prey between these treatments This methodhas been used to examine predation on cyanobacteria heter-otrophic bacteria and minute phototrophic eukaryotesAlternatively metabolic inhibitors have been employed tohalt predation (or in some cases prokaryote growth) insteadof physical removal by filtration
The dilution technique (356) has become a commonmethod for examining grazing mortality of phytoplanktonby microzooplankton (which is comprised predominantly ofprotozoa) This method relies on the dilution of herbivorouszooplankton in a series of subsamples (and concomitantmeasurements of phytoplankton growth in each subsample)to estimate the grazing mortality of the herbivores
Tracer experiments have used both radioactively labeledprey and fluorescently labeled prey to examine predation onbacteria and phytoplankton Radioactive labeling has beenaccomplished using 14C- or 3H-labeled organic compounds(for bacteria) or 14C-bicarbonate (for phytoplankton) Theuse of radioisotope approaches has diminished with improve-ments in the number and types of fluorescent compoundsavailable for this work Fluorescently labeled prey havebeen labeled with a variety of compounds that allow theprey to be observed readily (even inside predators) by epi-fluorescencemicroscopy (357 358) Alternatively a decreasein the total number of fluorescent prey in a sample indicatesthe rate of removal of the prey assemblage
Both perturbation experiments and tracer experimentsdescribed have inherent advantages and disadvantages Forexample perturbation experiments are fairly straightforwardto conduct but they may introduce artifact by removingpotentially important sources of nutrients or organic com-pounds released by predators (359) On the other hand thedilution technique is very labor-intensive but one advantageof this method is that it provides the opportunity to determineboth phytoplankton growth and grazing mortality in a singleprocedure Most tracer experiments are fairly easy to conductand generally do not perturb the food web being studied butfeeding selectivity for or against the tracer (relative to naturalprey) may produce spurious results In addition some of thesemethods provide a measurement of the activity of the entiregrazer assemblage and others are applicable for examiningthe activity of specific taxa For example size fractionationapproaches examine the effect of the removal of an entiresize class of organisms (a community-levelrdquo measurement)whereas fluorescently labeled prey can be used to examinethe ingestion rate of a particular species by observing theuptake of labeled prey into that species (a species-levelmeasurement)
Predation rates of numerous marine protozoa have beenexamined in the laboratory From these experiments a gen-eral knowledge of the rates of consumption by these specieshas been accumulated Feeding in most protozoa shows a
422-20 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
functional response to the availability of prey that is similar tothe response of many organisms (although there are a greatdeal of species-specific differences) That is ingestion rateincreases rapidly as prey abundance increases until a maximalingestion rate is attained (360)Manyof these curves have theform of a Type II functional response curve (361)
Based on the features of these curves it is clear that proto-zoa have the ability to ingest substantial numbers of prokar-yote and eukaryote prey in natural ecosystems Somebacterivorous flagellates can consume bacteria (and cyano-bacteria) at rates up to 100 s per individual per hour whereassome bacterivorous ciliates can consume thousands of bacte-ria per individual per hour Rates of consumption of eukary-otic phytoplankton by herbivorous ciliates and largeflagellates are considerably more variable and highly depend-ent of the feeding mechanism (eg filtration in ciliates versuspseudopodial net in dinoflagellates) and the size of the prey
These lab studies demonstrate that rarely are prey suffi-ciently abundant in nature to support the maximal ingestionrates of which many protozoa are capable Nevertheless pre-dation rates of protozoa in nature are sufficient to constitute amajor factor controlling the standing stocks of bacteria andphytoplankton (63 180 184 355) For example these sum-maries indicate that on average protozoa consume 20ndash80ofthe standing stock of phytoplankton (measured as chloro-phyll) in the world ocean and 60ndash75 of daily primary pro-duction a departure from the classical view of copepods as theprimary consumers of phytoplankton This large role of her-bivorous protozoa infers a large role for copepods as consumersof planktonic protozoa (185) Similarly the role of protozoaas major consumers of bacterial biomass is indicated by therelative balance between bacterial production on the onehand and bacterivory on the other (Fig 8) The approximateone-to-one balance between these measurements over a largenumber of studies implies that protozoan bacterivory must bea major factor determining the fate of bacteria in marine (andfreshwater) plankton ecosystems
It is interesting to note that some types of bacteria includ-ing marine Bdellovibrio are known to prey on other marine
bacteria (362) At the time of this writing the importanceof bacterial predation by other bacteria within the contextof oceanographic processes is not known and presumed tobe small While Bdellovibrio may be cultivated from seawaterit can also be found among sequences from cold sediments(363) and the water column Bdellovibrio happens to be thebest studied predatory bacterium but there are no doubtothers that we simply have not yet identified (364)
Species-Specific Interactions (MutualismParasitism Commensalisms)Interactions between marine microorganisms such as compe-tition allelopathy and symbiosis are fundamental aspectscontrolling community composition and activity but atpresent most are poorly characterized in natural marine sys-tems Bacteria must compete for dissolved and particulatecompounds with other bacteria andwith protistan taxa whilephagotrophic protists must compete with each other andmetazoa for available prey These interactions manifest them-selves in many forms including true competition for availableresources resource partitioning such that some taxa specializeon the utilization of certain substrates or prey while othersspecialize on different ones (reducing competition) or eventhe cooperative use of compounds (eg one bacteriumrsquoswaste products being utilized as substrate by another bacte-rium) These metabolic consortia are probably common inthe ocean but can be difficult to identify and characterizeConsortia of bacteria working together to perform complexreactions especially in anaerobic systems are well known inmicrobiology
Consortia in which there is a very high degree of integra-tion between species (ie symbiosis) are very common in theocean These associations may be beneficial to both partners(mutualism) beneficial to one part and inconsequential forthe other (commensalism) or beneficial to one part and det-rimental to the other (parasitism) Many highly visible andwell-characterized marine mutualisms exist between chemo-synthetic bacteria and animals such as hydrothermal ventor methane seep tubeworms clams mussels or other inverte-brates (365) and also between luminescent bacteria and fishor squids (366) Similarly many protozoa form mutualisticassociations with photosynthetic protists (367) The mostconspicuous of these associations involved large amoeboidprotozoa of oceanic ecosystems (planktonic foraminiferanspolycystine radiolaria and acantharia) which contain largenumbers of intracellular algae (74) but associations betweensome heterotrophic dinoflagellates and cyanobacteria are alsocommon (368) Studies have indicated that efficient nutrientcycling in these protozoan-algal associations confers an eco-logical advantage for both partners in surfacewaters of the oli-gotrophic ocean
Parasitism is also a common symbiotic relationship amongmarine protozoa Protozoan parasites have been reported for adiverse array of protists and animals in the ocean Heterotro-phic dinoflagellates and presently uncultured alveolate cladesrelated to alveolates appear to be particularly successful atevolving thismode of existence (369ndash371) In a common par-asitic group Amoebophrya spp the parasite invades photo-synthetic dinoflagellates where it undergoes reproductioneventually lysing the host and releasing dozens of infectiveswarmers The parasitic protist Pirsonia spp is a parasite ofdiatoms (372) The overall importance of these parasitismsin the population dynamics of their hosts is not clearly knownbut may constitute a significant source of mortality for somespecies (373 374) The frequency and diversity of ldquonewrdquo
FIGURE 8 Meta-analysis of experimental studies that have com-pared rates of bacterial productivity (converted to population growthrate units of dminus1) with rates of bacterivory by phagotrophic protists(expressed as equivalent units of bacterial growth rate dminus1) Thesolid line indicates where measurements of bacterial removal by graz-ers were equal tomeasurements of bacterial production Data redrawnfrom Sanders et al (63) doi1011289781555818821ch422f8
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-21
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ATP from sunlight Interestingly different versions of this pig-ment are found at different depths apparently ldquotuningrdquo theabsorption to match the ambient wavelengths of light asthey change with depth (147) The proteorhodopsin genesare very widespread and diverse perhaps in half or more ofall marine bacteria including SAR11 and Euryarchaeota(105 148ndash150) However almost all of several culturedorganisms with proteorhodopsin studied to date do not showa growth benefit from light (150) with the exception of amember of the Flavobacteria Dokdonia sp strain MED134that grows faster in the light only under moderately lownutrient conditions (151) and a marine Vibrio has beenshown to survive starvation longer in the light than in dark-ness (152) These observation may explain the wide phyloge-netic distribution and high abundance of this gene in thatproteorhodopsin may often assist long-term survival of bacte-ria under extreme energy-limiting conditions yet not havemucheffect on growthduringmore energy-replete conditionsInterestingly a recent report suggests the growth benefit fromlight in Dokdonia MED134 is from enhanced uptake of itsrequired growth factor vitamin B1 which is transported by atonB-dependent transporter powered by a proton gradient(153) This pigment might also have sensory or other rolesnot yet well examined (150)
The second kind of phototrophy found by moleculargenetic and fluorescence techniques to be unexpectedlyimportant in marine plankton is anoxygenic aerobic bacterialphototrophy based on the pigment bacteriochlorophyll aCulturable aerobic anoxygenic photosynthetic (AAP) bacte-ria have been known from seawater for several years (154) andare widespread (155) Initial recent reports of direct countsclaimed they represent about 11 of the total bacterial com-munity in the euphotic zone (156) but those early counts didnot correct for the presence of other bacteria and such correc-tionoften yields estimates averaging closer to 2(157)Directmeasurements show the bacteriochlorophyll pigment is rela-tively rare (158) However studies in various ocean locationsincluding ocean gyres report that these AAP bacteriacan indeed on occasionmake up a quarter of the total prokar-yotes (159 160) A recent energetic modeling of AAP andproteorhodopsin-containing bacteria (161) has suggestedthat proteorhodopsin-containingones cangain approximately02asmuchenergy from sunlight asSynechococcus (commonmarine cyanobacterium) and AAP bacteria can gain approxi-mately 13 as much energy from sunlight as SynechococcusThey concluded the AAP bacteria may gain energy sufficientto meet maintenance costs but proteorhodopsin-containingbacteria were not expected to do so except at high light inten-sities andwith largenumbersof proteorhodopsinmoleculespercell They concluded the ease and low cost of maintainingPR-based phototrophy (a few genes required) may explainthe high incidence of proteorhodopsin genes
Other metagenomic studies have yielded interestinginsights about organisms and processes that would otherwisebe difficult or impossible to establish For example thesequence of an archaeal gene fragment isolated from seawaterrevealed extensive evidence of genetic exchange with othertypes of archaea and even bacteria (162) Such geneticexchange was also quite evident from the complete genesequences of different strains of Prochlorococcus and Synecho-coccus which was attributed in part to virus-mediated geneflow (163ndash166)
Dilution Cultures of Bacteria and ArchaeaSome recent cultivation techniques that permit growth ofldquotypicalrdquo oligotrophic marine bacteria in pure culture are
based on the simple concept that bacteria living in seawateron dissolved organic matter might best be grown in the labo-ratory in ordinary filtered seawater This conclusion followsfrom similar techniques to grow mixed ldquoseawater culturesrdquoof marine bacteria (167) but the seawater inoculum in thiscase is diluted so that only one or a few bacteria are addedto the initial culture vessel (168) A rapid throughput versionof this method has been used to cultivate members of bacte-rial groups thought to be conventionally uncultivable such asthe SAR11 clade (169 170) albeit often at very low den-sities Such cultures permit focused studies of the propertiesof those organisms including genomic analysis (169 170)This valuable work provides insight into the roles and activ-ities of some of the most common bacterial phylotypesobserved in the ocean (171) showing for example that culti-vated SAR11 requires a source of reduced sulfur as well asproviding templates aiding the interpretation of metage-nomic analysis (172)
PROTOZOADiversity and DistributionHeterotrophic protists have been identified historically fromtheir morphological features that are apparent at the levelof the light or electronmicroscope Features of cell size shapetype and pattern of flagellationciliation skeletal structurescharacteristics of the nucleus and other cellular structuresand organelles have been used to differentiate themany thou-sands of described species A description of the extent of thisdiversity of form and function is well beyond the scope of thisbook (see [79] for more information) However much of thisdiversity can be reduced to three basic body plans that dictatethe broadest ecological roles of these cells amoeboid flagel-lated and ciliated forms (Fig 5) Among the lineages possess-ing one of these three forms only the last group the ciliatedprotists constitute a monophyletic group within the domainEukarya
One of the simplest body plans for protozoa is the amoe-boid cell exemplified by the gymnamoebae or ldquonakedrdquoamoebae Motile nonphotosynthetic cells lacking flagellaor cilia occur as life stages in a number of phylogeneticallydiverse taxa but for many free-living species of protozoathis form constitutes the only life stage Motility is largelyconfined to movement along surfaces by means of pseudopo-dia that can take on a variety of (species-specific) shapes Sig-nificant abundances of amoebae are largely confined tobenthic and epibiotic environments (174) and to suspendedparticulate material where they can occasionally be highlyenriched (27) Most amoebae consume bacteria and otherminute prokaryotes and eukaryotes
Several heterotrophic protistan groups possess complexamoeboid body plans most notably the foraminifera polycys-tine and phaeodarian radiolaria and the acantharia Plank-tonic forms are predominantly oceanic in their distributions(although a large number of benthic species of foraminiferaexist) These species are heterotrophic and many are visibleto the naked eye (individual cells can bege1 cm gelatinouscolonies can form ribbon-like structuresge1 m in length)They feed on a wide variety of bacterial protistan and meta-zoan prey using pseudopodial networks have rather longcomplex life cycles for individual cells (weeks to months)and are extensively used in studies of paleoclimatologicalreconstruction (82 175) In addition many of these speciespossess intracellular symbiotic usually eukaryotic algaeThe widespread occurrence of algal endosymbiosis among
422-12 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
these protozoa in oceanic pelagic environments implies thatstrong selective forces appear to give rise to these associations(97 98)
Flagellated forms of protozoa exist across many protistanlineages Eukaryotic flagella come in various sizes numbersand forms (eg with or without tiny hairs) that are character-istic of the different lineagesMany flagellate species are appa-rently purely phototrophic or heterotrophic (phagotrophic)but a significant (still poorly known) fraction of flagellates aremixotrophic combining both phototrophy and phagotrophy(70 88) Flagella in phagotrophic flagellates are employed formotility and prey capture Most free-living flagellates possessone to four flagella (typically one or two) that can be manytimes the length of the cell itself Although there is tremen-dous species diversity among flagellated protozoa many ofthese species have broadly overlapping ecologies As a groupflagellates are the most numerically abundant protozoa ofboth benthic and pelagic ecosystems and they are fundamen-tally important as consumers of bacteria cyanobacteriaand other eukaryotes Flagellated protozoa within the nano-plankton size class (2ndash20 microm) are often counted as a singleassemblage in plankton studies because morphological details
apparent using light microscope are insufficient to distinguishamong the many species This assemblage has been variouslyreferred to by a variety of names and acronyms including het-erotrophic nanoplankton (HNAN HN) heterotrophic flag-ellates (Hflags) heterotrophic nanoflagellates (HNF) andmicroflagellates Heterotrophic nanoplankton is the mostaccurate term for most methodologies employed to countthese cells because it does not require visualization of flagella(which are often lost from these small cells during preparationfor microscopy) The confusion over terminology is partly aconsequence of the different methodologies that have beenused to count these species and partly a consequence of thefact that these species were first studied prior to the wideacceptance of the size convention of Sieburth et al (84)
Heterotrophic flagellates in the microplanktonic size class(20ndash200 microm mostly dinoflagellates) are important consum-ers of phytoplankton in pelagic ecosystems Many of thesespecies are capable of the production of large pseudopodialnets the pallium that can engulf prey (particularly diatoms)significantly larger than the diameter of the dinoflagellatetheca (176) This behavior and its ecological significance forenergy flow in plankton communities have been recognized
FIGURE 5 Body plans and size ranges of protozoa These micrographs depict amoeboid (andashg) flagellated (hndashj) and ciliated (kndashm) forms ofprotozoa From Caron et al (173) Markers bars are 5 (i) 10 (h) 20 (c j l) 30 (a) 50 (k m) 100 (b f) 500 (d) and 1000 (e g) micromdoi1011289781555818821ch422f5
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-13
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
only within the past few decades (177) These species areoften abundant in waters where diatoms dominate (178179) In such situations heterotrophic dinoflagellate biomasscan be similar to that of ciliated protozoa (180)
Ciliates are generally the most recognizable form of pro-tozoa to nonspecialists Nonetheless ciliates are a diversegroup morphologically as well as ecologically (181 182)The degree of ciliature on these species can range from uni-formly ciliated to totally devoid of cilia during most of theirlife cycle to ciliature restricted to specific regions of thecell In addition cilia can fuse to form various complexstructures (eg ciliary membranes or veils cirri) that assistin locomotion food capture or attachmentCiliates are abun-dant and ecologically important species in both benthic (8)and pelagic (183) ecosystems and can consume a variety ofprokaryotic and eukaryotic prey Together with the heterotro-phic dinoflagellates these species are the dominant consum-ers of phytoplankton in many pelagic ecosystems (184) andas a consequence they form an important trophic link to met-azoan zooplankton (185 186)
Diversity and Biogeography of Protists FromMorphology to DNA SequencesUntil relatively recently the biodiversity of marine protistanassemblages was not generally considered a controversialtopic It was generally accepted that while all species of pro-tists certainly have not been identified representatives ofmost types of algae and protozoa had been observed anddescribed if not actually brought into culture in the labora-tory Direct sequencing of 18S rRNA genes from environ-mental samples (as described in ldquoMolecular Phylogeny andMetagenomics Field Applicationsrdquo) has changed that viewInitial forays into environmental DNA indicated a muchgreater diversity of protists than previously documented usingclassical approaches of culture and microscopy (81 187ndash191) Numerous publications over the past decades haveexpanded these findings (see [192] for a recent global analy-sis) which are highly analogous to discoveries in marine pro-karyote research as described already
Previously uncharacterized protistan diversity has nowbeen documented at virtually every level of eukaryoticorganismal classification Some of these findings couldhave been expected but some have been very unexpectedFor example a much greater diversity than noted previouslyhas been observed among small (lt10 microm) protists in plank-tonic ecosystems These species generally possess few distinc-tive morphological features and one could expect that manycryptic species might be present among these small morpho-types The molecular ldquodiscoveryrdquo of this eukaryotic diversityhas stimulated progress on the isolation and descriptionof new species and genera of minute algae and protozoa(193ndash197)
Analyses of environmental samples have also indicatedthe unanticipated existence of novel 18S rRNA gene sequen-ces that imply the presence of novel lineages of eukaryotes innatural protistan communities (198ndash200) These are sequen-ces for which there are apparently no known described orcultured species The degree to which these sequences dif-fer from sequences of known sequenced eukaryotes impliesthat some of these lineages may be distinct at the level ofphylum (201)
These findings have raised basic questions and somedebate concerning the true diversity and biogeography ofprotistan assemblages in natural ecosystems (202ndash204) thevalidity of the many novel phylotypes or cryptic species that
are being documented (205) the potential importance andsignificance of the many rare taxa that characterize thesecommunities (206) and the implications of these findingsfor the ecologicalbiogeochemical roles that protists playin aquatic ecosystems For example it was postulated andsubsequently confirmed that some novel alveolate lineagesreported from marine ecosystems make up a suite of parasiticprotozoa whose ecological importance may have been signifi-cantly underestimated in the past (207) Deciphering theidentity and significance of these many unknown phylotypeswill constitute a significant effort for protistologists in thefuture (80 85)
Life Histories and Ecological Strategies of ProtozoaAbundance prey type and life histories all vary tremendouslyamongmarine protozoa The smallest species (eg many flag-ellates) tend to be the most abundant and widely distributedin the world ocean Indeed many of these species may beglobally distributed (208) Most nanoplanktonic flagellateshave potentially rapid rates of grazing and growth Underoptimal conditions these species can divide by binary fissionseveral times a day and thus dramatic increases in their pop-ulations can take place in response to favorable conditionswithin a few days (209) Many of these species are capableof surviving for limited periods of time without food andhave developed a variety of physiological or life cycle strat-egies to cope with these events (209) However in contrastto some bacteria that may remain viable through long periodsof starvation (210) protozoa will expire or encyst in responseto low food abundance and thus ldquoboom-and-bustrdquo popula-tion changes are characteristic of small flagellates
On the other end of the size spectrum from the ratherubiquitous nanoflagellate species many of the largest proto-zoan species (polycystine and phaeodarian radiolaria plank-tonic foraminifera) are exclusively oceanic (ie do notsurvive in most coastal environments) andor have specificlatitudinal and depth distributions Thus the abundancesof these latter species may range from undetectable to max-imal abundances of gt105 individualsm3 Dramatic changesin the abundance of these protozoan taxa also can be relatedto changes in prey abundance physicalbehavioral aggrega-tion or to periodicity of life cycle events For example theplanktonic foraminifer Hastigerina pelagica reproduces on alunar cycle and thus abundances (and life stages) of this spe-cies in oceanic waters can vary considerably over the courseof a month (211) In general life cycles for the large amoe-boid protozoa are lengthy and complex (for single-celledorganisms) with life spans unknown for many species(attempts to culture them in the lab have so far been unsuc-cessful) but estimated to be on the order of months to per-haps years (175)
The abundance and activities of microplanktonic hetero-trophic protists (mostly ciliates and heterotrophic dinoflagel-lates) tend to be somewhat intermediate to those ofnanoplanktonic flagellates and the larger amoeboid formsThese species are present in the majority of marine ecosys-tems and collectively play an important role in the controlof phytoplankton biomass (and probably the abundance ofnanoplanktonic protozoa although there is little informa-tion on this topic) in waters throughout the world oceanLike small flagellates ciliates and dinoflagellates reproduceprimarily by binary fission but their maximal growth ratesare typically slower (one division a day is typical) and theirlife cycles often include sexual phases that allow for geneticrecombination
422-14 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
VIRUSESViral Abundance and General PropertiesViruses are simple biological agents typically 20ndash200 nm indiameter composed of a nucleic acid genome in a proteincoat that infect cells and ldquocommandeerrdquo the cellrsquos machineryto make more viruses which are released into the environ-ment when the host cell lyses or bursts A particular virus isthought to be capable of infecting only a narrow range of hosts(usually one species sometimes a genus rarely broader)Although some early studies had isolated viruses from thesea therewere no data prior to the 1980s showing such viruseswere very abundant and more significantly no evidence thatinfection was occurring in any important part of the planktoncommunity It wasnrsquot until the late 1980s that electronmicro-scopy with suitable concentration methods showed thatviruses are extremely abundant similar to or even higherthan bacterial abundance (212 213) Interestingly themost suitable transmission electron microscopy (TEM)approach used by Bergh et al (described in detail by [214])was actually similar to a direct centrifugation method devel-oped in 1949 (215)
TEM studies of viruses permit high-resolution images andobservation of viral morphology showing features such ashead diameter tails sheaths and tail fibers (Fig 6 collage
in upper left) Each type of virus has a fixed morphology(unlike bacteria that are potentially more plastic in theirappearance) and hence a coarse measure of viral diversityhas been possible by cataloging morphologies Studies thathave done so have found dozens or more different morpholo-gies of marine viruses in a given sample (217ndash219) A largeproportion resemble bacteriophages which are viruses thatinfect bacteria
The availability of brightly fluorescent nucleic acid stainsand high-porosity fine pore size (002 μm) filters made fromAl2O3 have made it easy to accurately count viruses by epi-fluorescence microscopy (38 220 221) Epifluorescence per-mits abundance estimates but does not allow observation ofviral morphology as viruses are below the resolution limitof light microscopy viruses are visible only as sources of light(like stars in the night sky see Fig 6 lower micrograph) Anextension of manual epifluorescence counts is the use of flowcytometry to count viruses now commonly employed in somelabs (222)
Virus abundance has been found to be closely related tobacterial abundance with a virusbacteria ratio typically10ndash301 A typical oceanic profile of bacterial and viral abun-dance is shown in (Fig 6 panel on right) This tight ratioand the strong correlation to bacterial abundance in relationto weaker correlations to chlorophyll have been cited as
FIGURE 6 Marine viruses (upper left collage) viewed by transmission electron microscopy These are cyanophages in the groups myoviridae(right and bottom) siphoviridae (left) and podoviridae (top) Scale bars represent 100 nm From Sullivan (216) Epifluorescence micrograph(lower left) of SYBR green I stained viruses (small fluorescing objects) and bacteria + archaea (large fluorescing objects) Depth distribution ofvirus and bacteria + archaea abundances obtained by epifluorescence microscopy of SYBRGreen stained cells in the central San Pedro BasinCalifornia (11 August 2000) doi1011289781555818821ch422f6
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-15
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
evidence that most marine viruses infect bacteria rather thaneukaryotic phytoplankton (217 218 223 224) though thereare also many important viruses of phytoplankton and otherorganisms with significant impacts on primary productivityblooms and ecosystem function (225ndash227)
The first demonstration of the activity of marine viruseswas presented by Proctor and Fuhrman (228) who showedby electron microscopic examination that viruses wereactually infecting marine bacteria and cyanobacteria at ameasurable rate Because only the final portion of the viruslife cycle is visible by electron microscopy (when the virusesare assembled and ready to lyse the host cell) only a very smallfraction of the infected cells can be counted as infected at anygiven time Proctor and Fuhrman (228) reported that only afew percent of the total bacteria from coastal waters and theSargasso Sea were visibly infected but they interpreted thedata with a model that implied the actual fraction of the totalcommunity that was infected was much higher They con-cluded that the total fraction of bacterial mortality attribut-able to viruses is roughly 10ndash40 Subsequent studies haveused refined versions of that same general approach andmodel (229 230) and numerous studies have used a varietyof alternative approaches to estimate viral activitymdashall yield-ing basically the same general conclusion (although themodel parameters need to be adjusted for cyanobacteriawhich on average seem to be infected less than bacteria)These approaches include (a) calculation of virus turnoverfrom decay estimates (231) (b) measurement of viral DNAsynthesis by incorporation of tritiated thymidine (232) (c)observation of effects of added viruses (233 234) (d) estima-tion of bacterial mortality in the absence of protists (e) use offluorescent viruses to estimate production by an approachanalogous to isotope dilution (38) and (f) dilution of virusesin filtered samples to estimate viral production (235) Theoverall consensus of these studies is that the initial estimateswere basically correct that is that viruses are responsiblefor about 10ndash40 of the bacterial mortality in most marinesystems (reviewed by [218 223 226 236 237]) Howeverthere is still some disagreement about the higher estimatesand it is likely that 40 mortality from viruses alone is nottypical for most marine systems
Most of the foregoing work has focused on the viral infec-tion of bacteria primarily thought to be heterotrophicalthough the original report by Proctor and Fuhrman (228)also noted the occurrence of cyanobacterial infection Sev-eral subsequent studies focused on phytoplankton includingthe potential effect of viruses on the termination of phyto-plankton blooms of Emiliania and Phaeocystis and the likelyimpact this might have on release of climate-active gasessuch as dimethyl sulfide (225 227 238ndash247) Althoughdetails are beyond the scope of this chapter viruses arethought to infect virtually all marine organisms with poten-tially significant impacts from zooplankton to whales (226)Detailed studies of viruses infecting cyanobacteria such asSynechococcus have shown differences in viral host specific-ity particularly toward coastal and oceanic host strainsand some occasional high virus abundances (to 105ml) asmeasured by most probable number (MPN) cultivation tech-niques in the Gulf of Mexico near Texas (248 249) Othervirus cultivation studies with Prochlorococcus and Synecho-coccus in oligotrophic waters of the Sargasso Sea showedan interesting pattern of cross-infection between these gen-era by some virus types (suggesting gene flow among theseorganisms) but generally low MPN estimates of abundanceto 103ml even when cyanobacterial abundance was near105ml (216 250)
Overall the consensus emerging from direct comparisonsof viral-mediated mortality and grazer-mediated (ie micro-zooplankton) mortality have indicated that viral lysis of bac-teria constitutes a significant fraction of total mortality ofthis assemblage while the mortality of phytoplanktonappears to be dominated bymicrozooplankton inmost instan-ces (251 252)
As mentioned viruses have significant morphologicaldiversity as observed in TEM studies It is also possible toinvestigate aspects of their genetic diversity In early workon this topic Wommack et al (253) and Steward et al(254) observed the diversity of viral genome lengths in a fieldsample by pulsed field gel electrophoresis with viral genomesranging 25 kb to gt300 kb in length Field results show thatthe viral community composition is dynamic in space andtime with clear changes in the banding patterns over seasonsand locations in Chesapeake Bay (253) between oceanbasins and subsequent to dinoflagellate blooms (254) andwith depths to 500 m at one location (255)
Although viruses do not all share a set of core genesthat allows a universal viral phylogeny (comparable to SSUrRNA gene in cellular organisms) genetic diversity amonga single group of viruses can be examined by sequence analysisof shared genes within the group An example is the g20 geneinT4-like cyanophages that has shown extremely high diver-sity even among very closely related viruses and has demon-strated geographic and seasonal variation (256) A secondexample is g23 which is found broadly in diverse and wide-spread T4-like phages (257) and can have seasonally repeat-ing patterns (258) as well as short-term rapid dynamics thatcorrelate to those in bacteria (259)
Viral MetagenomicsAs with cellular organisms metagenomics can be used toexamine viral diversity and genetics without themany restric-tions of cultivation Viruses can be collected by selective fil-tration and concentrated by tangential flow filtration orflocculated with iron chloride (260 261) then their collec-tive metagenome can be extracted linker amplified and ana-lyzed by sequencing (262 263) Viral metagenomic studiesare particularly challenging because the large majority ofsequences have no annotated matches in any databases butso far results from marine samples around the world haveshown extremely high diversity and variations with depthlocation and time presumably with highly dispersed typesselected by local conditions (262ndash266) Because metage-nomes are best interpreted when there are representative cul-tures available the best matches of viral metagenomesoriginally tended to be to the few viral isolates infecting trulycommonmarine bacteria like cyanophage infecting Synecho-coccus and Prochlorococcus (267) However the developmentof dilution-to-extinction cultures representing common het-erotrophic marine taxa like Peligibacter (a member of theSAR11 clade) and SAR116 has allowed isolation from sea-water of viruses infecting these common organisms and theseisolates have indeed been found to be highly abundant inmarine viral metagenomes (268 269) New approaches tointerpret the results include clustering the proteins independ-ent of known proteins to compare samples to each other andtry to find environmental factors driving viral communitychanges (270) Such analysis of a large global data set (TaraOceans expedition) has shown that extensive sampling hascome close to reaching the total diversity in tropical and tem-perate waters of such viral protein clusters (which essentiallyrepresent various viral protein motifs not all viral proteintypes) and that viruses appear to be directionally dispersed
422-16 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquodownstreamrdquo in major ocean currents as one may expectfrom first principles but also supporting the ldquoseed bankrdquohypothesis of viral biogeography (271)
Viruses and Host DiversityViruses are themselves thought to be instrumental in drivingincreased diversity of their microbial hosts via a hypothesizedprocess often called ldquokill the winnerrdquo This is because viralinfection is host-specific and density-dependent the latterbecause viruses diffuse from host to host so an abundanthost is more likely to pass on infection than a rare oneThis means that if an organism becomes abundant andblooms winning the competition for resources it becomesmore susceptible to a viral epidemic This would benefit therarer organisms and thus help foster diversity (223 272273) However bloom scenarios as described above involvesystems far from steady state and the formal theory of Thing-stad and Lignell (274) has interesting steady-state solutionswhere several viruses infect several hosts stably over time(via trade-offs between growth rates and viral susceptibility)which may occur at the strain or species level (274) There issome experimental evidence that viruses have effects on nat-ural marine microbial community composition weaker thanthe kill the winner (bloom version) hypothesis would sug-gest (275 276) so something resembling the steady-statecoexistence described in the model may in fact be commonThere also appear to be processes that foster coexistencebetween viruses and hosts but the mechanisms are largelyspeculative (223 277 278)
Viruses may also be directly involved in host genetic diver-sity because they can be the agents of genetic exchangebetween microorganisms (223 273) This often involvesthe viral lifestyle known as lysogeny whereby viruses survivewithin host cells as DNA only integrated into the host chro-mosome and being reproduced each time the host divides Ahost harboring such a genome is called a lysogen becauseunder conditions of stress to the host cell a genetic switchmay cause the viral genome to initiate the lytic process pro-ducing many progeny viruses and bursting from the hostLysogeny is a very common property occurring in a signifi-cant part of the bacterial community (279 280) althoughthe incidence of induction of the lytic phase in nature is appa-rently low (281) Overall lysogeny is poorly understood butthought to have both positive and negative impacts on themicrobial community (282) Recent results suggest that ben-efits of being lysogenic in highly seasonal polar seas leads tofundamental difference between polar and other marine viralcommunities (283)
Viruses and the Microbial LoopAs part of the food web viruses occupy a unique positionThey infect host cells that are mostly thought to be heterotro-phic bacteria and by doing so they typically burst the hosts torelease progeny viruses and cellular debris But what is the fateof this material Viruses themselves do not last indefinitelyand a simple steady-state assumption implies that from eachburst of viruses (typically 20ndash100 per lytic event) only onesuccessfully infects another cell The rest are inactivatedand broken down by sunlight (UV and visible exposure)and enzymatic attack (284) or consumed byminute phagotro-phic protists (285) thus reentering the food web as substratefor bacteria or food for protistan consumers Experiments incontrolled laboratory systems and field studies with radioac-tively labeled viral lysis products have supported the conclu-sion that most of the organic matter released by the viralinfection is either taken up by bacteria or respired (286 287)
Modeling this process as part of the microbial loop showsthat viral lysis represent a sort of side loop that has the neteffect of remineralizing a significant amount of the carbonand nutrients that enter the bacteria-protist part of themicro-bial loop (Fig 7) A theoretical numerical steady-state modelcomparing a systemwith no viral activity to onewhere virusesare responsible for 50 of bacterial mortality showed that thesystem with viruses had 33 more bacterial production andrespiration than the virus-free system implying that theviruses had the effect of permitting the bacteria to processmore of the primary production than they would otherwise(223) Although 50 is a high number unlikely to be com-mon in the sea this model nevertheless illustrates that virusescan reduce the amount of energy reaching higher trophic lev-els The implication is that viruses lead to increased bacterialactivity at the expense of the larger organisms
MAJOR ENVIRONMENTAL CONTROLSLight Temperature and PressureTemperature has an important potential influence on bio-chemical reactions and therefore on biological processesin general Most ocean waters fall in the range of ndash2degC to30degC with obvious exceptions in hydrothermally heatedareas Temperature has long been known to be a regulatingfactor for the growth of heterotrophic microbes
In temperate waters it has been established that microbialactivity is generally much higher in warm summer waters thanin winter (55) The relationship is not simple howeverbecause multiple factors act at the same time Some contro-versy still exists regarding the highest and lowest extremesfor marine bacterial growth although there is broad agree-ment that bacteria grow gt100degC at hydrothermal vents andltminus5degC in sea ice brines Pomeroy et al (57) noted the inter-esting observation that bacteria seem particularly inhibitednear the freezing point of seawater (ca ndash22degC) comparedto eukaryotic phytoplankton This effect results in polarspring phytoplankton blooms that accumulate organic car-bon in advance of the response of the bacterial communityand development of the microbial loop and perhaps lead toenhanced benthic-pelagic coupling (288)
The relationship between temperature and the growth rateof marine phytoplankton was described broadly in a now
FIGURE 7 Modification of the microbial loop concept that incor-porates the functional role of viruses Export can be via predation orsinking From Fuhrman (223)doi1011289781555818821ch422f7
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-17
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
classic paper by Eppley (58) and later Goldman and Carpen-ter (289) Temperature was shown to exert a strong and directeffect on themaximal growth rates of these species withmax-imal intrinsic growth rates at 0degC generally lt1 divisiondaywhile growth rates at 30degC may be gt4day These relation-ships indicate the maximal rate that might be attained byphytoplankton at these temperatures but they do not takeother factors into account (nutrients light) While warmtemperature ostensibly allows more rapid growth it createshydrographic conditions that typically give rise to nutrientlimitation of algal growth Thus some of the coldest watersin nature witness some of the most massive phytoplanktonblooms (290) while warm oceanic gyres represent some ofthe most oligotrophic areas of the ocean The multiple andoften conflicting effects of temperature on the growth of nat-ural phytoplankton assemblages limit the accuracy of presentpredictions regarding how primary producers will respond toclimate change (291)
An analysis of the effect of temperature on the growth ofheterotrophic protists has indicated that the response is qual-itatively similar to that of phytoplankton but with a twistThe growth rates of at least some protozoa can exceed thoseof phytoplankton at warmer environmental temperaturesbut the opposite effect is apparent at very low environmentaltemperature This differential effect of temperature on thegrowth of phototrophic and heterotrophic protists was basedon a large meta-analysis of published protistan growth (59)That analysis demonstrated that the maximal growth ratesattained by phototrophic protists could exceed the maximalgrowth rates attained by heterotrophic protists (all otherpotential growth-limiting factors not considered) Thereforeprotozoan growth rates may be constrained to a greater degreeat low environmental temperature than rates for phytoplank-ton If so then phytoplankton bloomsmay get a head start ongrazers during spring in polar ecosystems This scenario is con-sistent with information on seasonal biomass changes andmicrozooplankton herbivory in the Ross Sea Antarctica(292 293) but there are still too few data to fully vet thishypothesis
The importance of high pressure on bacterial growthgained considerable attention in the late 1960s when thedeep sea submersible Alvin was accidentally lost overboardwith its hatch openWhile no lives were lost in this accidentsome workmenrsquos lunches sank to the bottom (sim1500 m)inside the submersible Alvin was recovered after 10 monthsand interestingly there was a waterlogged lunch containingapples bologna sandwiches and broken vacuum bottleswith broth that all appeared hardly degraded and tasted palat-able Yet when placed in a refrigerator on the ship these itemsdegraded relatively quickly Initially pressure was thought tobe the preserving factor as the sea floor temperature was sim-ilar to the refrigerator temperature and the only major differ-ence would be pressure (294) Following this observationa series of experiments to measure degradation of variousorganic materials left in the deep sea for extended periodsindicated that degradationwas typically significantly reducedimplying that pressure reduces the degradation rates (295)Nonetheless changes in the protozoan community of naturaldetrital material sinking to the deep ocean floor indicate thatthe microbial community can respond relatively quickly insome situations (296) Deep sea microbiology has advancedconsiderably yet it is still difficult to interpret results withrespect to actual in situ rates of naturally occurring organicmatter Deep sea bacteria adapted to high pressures havebeen isolated that are barophilic (also called piezophilic)meaning that they prefer high pressures and have reduced
activity at lower pressures whereas others are barotolerant(piezotolerant) tolerating but not preferring high pressuresfor example see (297 298)
Few data are available on barotolerantbarophilic marineprotozoa Protozoa certainly exist and grow at great oceanicdepths but measurements of in situ growth rates for these spe-cies do not yet exist Measurable protozoan numbers havebeen documented in the deep-sea sediments for more than30 years (299 300) and viable protozoa have occasionallybeen cultured from these environments (301ndash305) butvery few direct measurements of the activities of these speciesin situ have been reported (306) A few protozoa have beenisolated that will grow at high pressure (302 303 307) andprotists that appear to be unique to the deep ocean havebeen observed either directly or through the analysis ofDNA sequences (308ndash311) but possibly the best direct evi-dence that protozoan activity takes place at the high pressurescharacteristics of the deep sea are experimental and observa-tional work noting the stimulatory effect that detrital deposi-tion has on some components of the protozoan community(296 312) These observations indicate a diverse and activeprotozoan fauna of the deep ocean although their biogeo-chemical significance is largely uncharacterized at this time
Dissolved and Particulate Organic MatterBacteria and archaea are thought to be by far the most impor-tant organisms with respect to the processing of dissolvedorganic matter (DOM) and nonliving particulate organicmatter (POM also called detritus) in the ocean While theremay be some uptake of DOM by protists particularly forgrowth factors needed in trace amounts such as vitamins(313) the bulk of this material is probably utilized by bacteriaand archaea (314) including the smallest cyanobacteriumProchlorococcus (315 316) which thus may be considered amixotroph Due to their small size bacteria have extremelyhigh surfacevolume ratios and combined with their over-whelmingly high abundance an extremely high integratedsurface area Protozoa tend to obtain the organic materialsthat they require for growth from their prey rather thanthrough the uptake of DOM Overall protozoa tend to besources of dissolved and detrital organic substances throughthe excretion of unassimilated prey biomass in expelledfood vacuoles
Particulate organic matter is not directly available as sub-strate to bacteria These substances must first be reduced tosmall molecules that can be transported into the cell Thisis accomplished by the production of extracellular enzymes(note that few if any large polymers are directly taken up bybacteria with the possible exception of DNA) Hydrolyticenzymes produced by bacteria (and almost certainly archaea)break down polymers like proteins polysaccharides andnucleic acids As with DOM POM is composed of a complexmixture of compounds that vary in their susceptibility to bac-terial degradation and utilization
Particulate material in the water column serves not only asbacterial substrate but also as substratum POM occurs in thewater column across a huge size spectrum from micrometersup to some detrital aggregates more than 1 m in diameter(317) Much of this particulate material is in a constant stateof flux with colloidal material constantly coalescing andaggregating to form new or larger particles (318) as microbialdegradation acts simultaneously to remineralize this materialDetrital particles that attain macroscopic size either by directformation (317 319 320) or via accretion and aggregation(321) and are often called marine snow or macroaggregatesMarine snow particles are readily colonized by bacteria and
422-18 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
many other microorganisms and their abundances on theseaggregates can be orders of magnitude greater than abundan-ces in an equivalent volume of the water surrounding theaggregates (320 322ndash324) and their diversity and highlyconcentrated abundances on detrital aggregates implies thatthere may be stronger trophic coupling and efficient energyelemental cycling relative to these processes among free-living microorganisms in the surrounding waters This specu-lation has resulted in a number of formalizations of the poten-tial impact of aggregate microbiology on biogeochemicalprocesses in the ocean (325 326) Most definitions of bio-films include aggregated particulate and these particles areoften studied within the biofilm conceptual model (see chap-ter by Lawrence et al in this section)
Inorganic Nutrients
Macronutrients (N P)Virtually all heterotrophic organisms in the ocean contributeto the pool of available macronutrients via the excretion ofmetabolic wastes and protozoa contribute significantly tothis process Macronutrients are often in excess within proto-zoan prey relative to the consumersrsquo needs because much ofthe prey carbon is respired to produce energy for metabolismand growth ldquoExcessrdquo nutrients are eliminated as eitherorganic compounds or more commonly as ammonium (forN) and phosphate (for P) The importance of the match(or mismatch) between predator and prey stoichiometry hasbeen demonstrated experimentally and repeatedly in marineand freshwater ecosystems and has been elevated to a funda-mental ecological principle governing elemental and energyflow in aquatic food webs (327) Based on this reasoningCaron et al (328) concluded that bacterivorous protozoaplay an important role in nutrient remineralization andrelease in the ocean Bacterial biomass is typically rich in Nand P relative to other organisms so bacterial predators expe-rience large excesses of these elements relative to their growthneedsMoreover bacterivorous protozoa should be dispropor-tionately important in nutrient remineralization amongsingle-celled eukaryotes because as the smallest protozoa inaquatic ecosystems they have high weight-specific metabolicrates (329) Thus both stoichiometric and allometric rela-tionships implicate small bacterivorous protozoa as impor-tant sources of remineralized nutrients in the plankton
The reasoning also holds true for bacterial utilization oforganic compounds and it has led to some interestingfindings and conclusions regarding the role of bacteria innutrient cycling in the ocean Bacteria are important sourcesremineralized nutrients when they are consuming N- orP-rich substrate Contrary to this traditional view howeverheterotrophic bacteria are often strong competitors for inor-ganic nutrients under many situations largely because bacte-rial biomass is richer in N and P than much of the organicmatter they consume in nature so they require additional Nand P to produce their biomass (summarized by [330]) Exper-imental studies have demonstrated the uptake of ammoniumby bacteria other than cyanobacteria using stable isotope 15N(331) and also the short-lived radioisotope 13N (332) oftenfinding that bacteria may be responsible for one third of the Nor P uptake Additionally there is evidence that bacteria insome oceans like the Sargasso Sea are limited by phosphorus(333 334) Growth limitation of bacteria by phosphorusappears to be a common phenomenon of freshwater ecosys-tems (335 336)
N2 fixation is restricted to prokaryotes and in the oceanwater column it was thought for many years to be done
primarily byTrichodesmium a warm-water colonial cyanobac-terium that blooms sporadically andRichelia a cyanobacterialsymbiont that lives within certain diatoms (337) Some uni-cellular cyanobacteria also contribute significantly to globalN2 fixation (338) and molecular biological data suggestthat a variety of other bacteriamdashnot phytoplanktonmdashmayalso be fixing nitrogen in seawater (339) An exciting recentdiscovery in this field is that a bacterial nitrogen fixer origi-nally known only from nifH sequences UCYN-A has hadits genome sequenced by parallel pyrosequencing and wasfound be a cyanobacterium (Candidatus Atelocyanobacteriumthalassa) lacking the apparatus to generate oxygen as well asother pathways (340) Further study showed it to be symbioticwith a marine alga (341)
Micronutrients (Trace Metals Growth Factors)Bacterial growth may often be limited by macronutrients asdescribed and sometimes by trace nutrients particularlyiron that occur at very low concentrations in seawater(342 343) The importance of Fe as an element limiting pri-mary production in some oceanic regions has come underclose scrutiny and extensive experimental investigation inrecent years Many bacteria have special uptake mechanismsto utilize extremely low concentrations of iron includingsiderophores (released compounds that bind iron and arethen specifically taken up) Even if bacteria are capable ofgrowth at low Fe concentrations growth may be less efficientKirchman et al (344) found that Fe limitation stronglyreduced bacterial growth efficiency in a cultured marine bac-terium (ca 50 efficiency Fe replete but lt10 when Fedeplete) Thus the effects may be complex Marine N2 fixa-tion also may be limited by the availability of Fe becausenitrogen fixers have high Fe requirements
Protozoa tend to be a source rather than a sink for Fe in theocean analogous to their role in macronutrient remineraliza-tion Protozoan grazing activity has been shown to releaseiron from bacterial biomass thereby relieving Fe limitationin co-occurring phytoplankton (345) a mechanistic demon-stration of the process using protozoa growing on Fe-repletebacteria However it has also been reported that Fe-limitedbacteria may contain insufficient Fe for the bacterivorous pro-tozoa that consume them thus leading to Fe limitation in theprotozoan as well (346)
OxygenHypoxic and anoxic regions of the ocean are expected to sig-nificantly expand in the coming decades as a consequence ofchanges in ocean stratification resulting from global climatechange (347) As oxygen becomes depleted bacteria havethe capability to utilize a series of alternate electron acceptorsCompositional changes in the bacterial assemblage accom-pany these changes in metabolic activity but are beyondthe scope of this chapter Many of the studies on benthicmicrobiology have focused on redox reactions that occurthere Oxygen is usually consumed by sediment microbialactivity much faster than the rate at which it can be replacedby diffusion so most sediments especially organically richones are anaerobic below a few mm (fine-grained muds) orcm (coarser sediments) from the sedimentwater interfaceexcept where animals ventilate the benthos through tubesWhen oxygen is absent bacteria and archaea use alternateelectron acceptors such as nitrate nitrite oxidized Fe orMn organic matter or sulfate The use of nitrate or nitriteas an electron acceptor with concomitant production of N2
gas (denitrification) results in net loss of biologically availablenitrogen Bacteria that perform this reaction are typically
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-19
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquofacultativerdquo organisms that can switch from oxygen to nitrateor nitrite Organic matter may be reduced by fermentationreactions in which part of an organic substrate is oxidizedwhile another is reduced fermenter organisms are also typi-cally facultative Sulfate is a particularly important alternateelectron acceptor in marine systems because it occurs as thesecond most abundant anion in seawater (behind chloride)typically 28 mM Its use as an electron acceptor leads to theproduction of elemental sulfur or sulfide The use of sulfateas an electron acceptor yields much less energy than oxygenor nitrate and sulfate-reducing bacteria are strict anaerobesunable to use oxygen (often strongly inhibited by oxygen)Methane can be produced by methanogenic archaea whichare all strict anaerobes using CO2 as an electron acceptor(see Findlay and Battin this section for further discussion)
Reduced compounds such as sulfide are produced by theuse of these alternate electron acceptors and they diffuse awayfrom their source When they diffuse to environments with astrong oxidant like oxygen they may be oxidized by chemo-trophic bacteria in energy-yielding reactions Similarlyammonium can be oxidized to nitrite and then nitrate by che-motrophic nitrifying bacteria and the nitrate can diffuse intoanaerobic layers and become denitrified to N2 Such coupledoxidation-reduction reactions can lead to accelerated biogeo-chemical cycling particularly in sediments
It was originally believed that relatively few free-livingspecies of protozoa inhabit anaerobic and hypoxic marineenvironments This view has changed based on (largely)molecular analyses of some of these environments (308348ndash351) A significant number of previously undescribedeukaryotic phylotypes have been observed in anoxic environ-ments High protozoan biodiversity in the absence of molec-ular oxygen is not completely unexpected because otheranaerobic environments such as some gut environments(eg cockroach guts and cattle rumen) have notoriouslyabundant and diverse protozoan fauna but marine anaerobicenvironments have been difficult to access and sample prop-erly Many anaerobic protozoa exist together with symbioticbacteria (352 353) or possess unique organelles such as thehydrogenosome (354)
MAJOR BIOTIC CONTROLSThe Trophic Activities of Protozoa Predation andTop-Down ControlsPredator-prey relationships are fundamental interactionswithin biological communities linking production of bio-mass on the one hand with removal on the other hand Estab-lishing the trophic relationships among microorganisms andquantifying the rates at which they take place constitutemajor endeavors within marine microbial ecology As notedpreviously protozoa consume a wide variety of prokaryoticand protistan prey Numerous laboratory and field measure-ments have been made throughout the past quarter centuryto attempt to quantify the rates of bacterial and phytoplank-ton consumption The methodology for accomplishing thesestudies has varied and a number of approaches are presentlyemployed
Methodologies for estimating microbial grazing are diversebut generally fall into one of two experimental approachesperturbation experiments or tracer experiments Perturbationexperiments (size fractionation metabolic inhibitors anddilution technique) rely on manipulation of the consumeror prey assemblages in somemanner such that the effect is evi-dent from changes in the abundance of the prey population
during incubations relative to control treatments that receiveno manipulation Tracer experiments (radioactively labeledprey fluorescently labeled prey stable isotopendashlabeled pray)attempt to use labeled prey or a prey proxy to quantitativelyfollow the movement of prey biomass into consumers Forsummaries of these methods see (74 355)
Perturbation experiments have entailed a variety of mech-anisms to ldquodecouplerdquo predators from prey in natural microbialcommunities For example size fractionation uses various sizefilters to remove an entire size class of grazers from awater sam-ple leaving the prey to grow in the relative absence of thosepredators during subsequent incubations Unfiltered samples(controls) indicate the growth of prey in the presence of pre-dation Predation impact is obtained by the difference ingrowth of the prey between these treatments This methodhas been used to examine predation on cyanobacteria heter-otrophic bacteria and minute phototrophic eukaryotesAlternatively metabolic inhibitors have been employed tohalt predation (or in some cases prokaryote growth) insteadof physical removal by filtration
The dilution technique (356) has become a commonmethod for examining grazing mortality of phytoplanktonby microzooplankton (which is comprised predominantly ofprotozoa) This method relies on the dilution of herbivorouszooplankton in a series of subsamples (and concomitantmeasurements of phytoplankton growth in each subsample)to estimate the grazing mortality of the herbivores
Tracer experiments have used both radioactively labeledprey and fluorescently labeled prey to examine predation onbacteria and phytoplankton Radioactive labeling has beenaccomplished using 14C- or 3H-labeled organic compounds(for bacteria) or 14C-bicarbonate (for phytoplankton) Theuse of radioisotope approaches has diminished with improve-ments in the number and types of fluorescent compoundsavailable for this work Fluorescently labeled prey havebeen labeled with a variety of compounds that allow theprey to be observed readily (even inside predators) by epi-fluorescencemicroscopy (357 358) Alternatively a decreasein the total number of fluorescent prey in a sample indicatesthe rate of removal of the prey assemblage
Both perturbation experiments and tracer experimentsdescribed have inherent advantages and disadvantages Forexample perturbation experiments are fairly straightforwardto conduct but they may introduce artifact by removingpotentially important sources of nutrients or organic com-pounds released by predators (359) On the other hand thedilution technique is very labor-intensive but one advantageof this method is that it provides the opportunity to determineboth phytoplankton growth and grazing mortality in a singleprocedure Most tracer experiments are fairly easy to conductand generally do not perturb the food web being studied butfeeding selectivity for or against the tracer (relative to naturalprey) may produce spurious results In addition some of thesemethods provide a measurement of the activity of the entiregrazer assemblage and others are applicable for examiningthe activity of specific taxa For example size fractionationapproaches examine the effect of the removal of an entiresize class of organisms (a community-levelrdquo measurement)whereas fluorescently labeled prey can be used to examinethe ingestion rate of a particular species by observing theuptake of labeled prey into that species (a species-levelmeasurement)
Predation rates of numerous marine protozoa have beenexamined in the laboratory From these experiments a gen-eral knowledge of the rates of consumption by these specieshas been accumulated Feeding in most protozoa shows a
422-20 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
functional response to the availability of prey that is similar tothe response of many organisms (although there are a greatdeal of species-specific differences) That is ingestion rateincreases rapidly as prey abundance increases until a maximalingestion rate is attained (360)Manyof these curves have theform of a Type II functional response curve (361)
Based on the features of these curves it is clear that proto-zoa have the ability to ingest substantial numbers of prokar-yote and eukaryote prey in natural ecosystems Somebacterivorous flagellates can consume bacteria (and cyano-bacteria) at rates up to 100 s per individual per hour whereassome bacterivorous ciliates can consume thousands of bacte-ria per individual per hour Rates of consumption of eukary-otic phytoplankton by herbivorous ciliates and largeflagellates are considerably more variable and highly depend-ent of the feeding mechanism (eg filtration in ciliates versuspseudopodial net in dinoflagellates) and the size of the prey
These lab studies demonstrate that rarely are prey suffi-ciently abundant in nature to support the maximal ingestionrates of which many protozoa are capable Nevertheless pre-dation rates of protozoa in nature are sufficient to constitute amajor factor controlling the standing stocks of bacteria andphytoplankton (63 180 184 355) For example these sum-maries indicate that on average protozoa consume 20ndash80ofthe standing stock of phytoplankton (measured as chloro-phyll) in the world ocean and 60ndash75 of daily primary pro-duction a departure from the classical view of copepods as theprimary consumers of phytoplankton This large role of her-bivorous protozoa infers a large role for copepods as consumersof planktonic protozoa (185) Similarly the role of protozoaas major consumers of bacterial biomass is indicated by therelative balance between bacterial production on the onehand and bacterivory on the other (Fig 8) The approximateone-to-one balance between these measurements over a largenumber of studies implies that protozoan bacterivory must bea major factor determining the fate of bacteria in marine (andfreshwater) plankton ecosystems
It is interesting to note that some types of bacteria includ-ing marine Bdellovibrio are known to prey on other marine
bacteria (362) At the time of this writing the importanceof bacterial predation by other bacteria within the contextof oceanographic processes is not known and presumed tobe small While Bdellovibrio may be cultivated from seawaterit can also be found among sequences from cold sediments(363) and the water column Bdellovibrio happens to be thebest studied predatory bacterium but there are no doubtothers that we simply have not yet identified (364)
Species-Specific Interactions (MutualismParasitism Commensalisms)Interactions between marine microorganisms such as compe-tition allelopathy and symbiosis are fundamental aspectscontrolling community composition and activity but atpresent most are poorly characterized in natural marine sys-tems Bacteria must compete for dissolved and particulatecompounds with other bacteria andwith protistan taxa whilephagotrophic protists must compete with each other andmetazoa for available prey These interactions manifest them-selves in many forms including true competition for availableresources resource partitioning such that some taxa specializeon the utilization of certain substrates or prey while othersspecialize on different ones (reducing competition) or eventhe cooperative use of compounds (eg one bacteriumrsquoswaste products being utilized as substrate by another bacte-rium) These metabolic consortia are probably common inthe ocean but can be difficult to identify and characterizeConsortia of bacteria working together to perform complexreactions especially in anaerobic systems are well known inmicrobiology
Consortia in which there is a very high degree of integra-tion between species (ie symbiosis) are very common in theocean These associations may be beneficial to both partners(mutualism) beneficial to one part and inconsequential forthe other (commensalism) or beneficial to one part and det-rimental to the other (parasitism) Many highly visible andwell-characterized marine mutualisms exist between chemo-synthetic bacteria and animals such as hydrothermal ventor methane seep tubeworms clams mussels or other inverte-brates (365) and also between luminescent bacteria and fishor squids (366) Similarly many protozoa form mutualisticassociations with photosynthetic protists (367) The mostconspicuous of these associations involved large amoeboidprotozoa of oceanic ecosystems (planktonic foraminiferanspolycystine radiolaria and acantharia) which contain largenumbers of intracellular algae (74) but associations betweensome heterotrophic dinoflagellates and cyanobacteria are alsocommon (368) Studies have indicated that efficient nutrientcycling in these protozoan-algal associations confers an eco-logical advantage for both partners in surfacewaters of the oli-gotrophic ocean
Parasitism is also a common symbiotic relationship amongmarine protozoa Protozoan parasites have been reported for adiverse array of protists and animals in the ocean Heterotro-phic dinoflagellates and presently uncultured alveolate cladesrelated to alveolates appear to be particularly successful atevolving thismode of existence (369ndash371) In a common par-asitic group Amoebophrya spp the parasite invades photo-synthetic dinoflagellates where it undergoes reproductioneventually lysing the host and releasing dozens of infectiveswarmers The parasitic protist Pirsonia spp is a parasite ofdiatoms (372) The overall importance of these parasitismsin the population dynamics of their hosts is not clearly knownbut may constitute a significant source of mortality for somespecies (373 374) The frequency and diversity of ldquonewrdquo
FIGURE 8 Meta-analysis of experimental studies that have com-pared rates of bacterial productivity (converted to population growthrate units of dminus1) with rates of bacterivory by phagotrophic protists(expressed as equivalent units of bacterial growth rate dminus1) Thesolid line indicates where measurements of bacterial removal by graz-ers were equal tomeasurements of bacterial production Data redrawnfrom Sanders et al (63) doi1011289781555818821ch422f8
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-21
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-
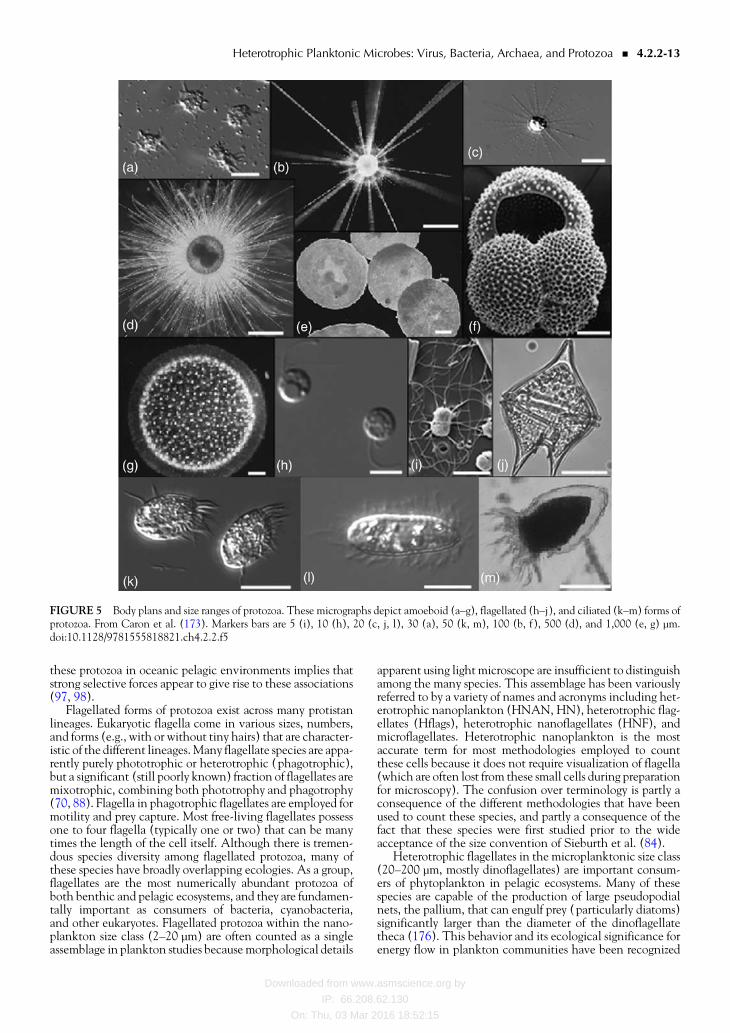
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
these protozoa in oceanic pelagic environments implies thatstrong selective forces appear to give rise to these associations(97 98)
Flagellated forms of protozoa exist across many protistanlineages Eukaryotic flagella come in various sizes numbersand forms (eg with or without tiny hairs) that are character-istic of the different lineagesMany flagellate species are appa-rently purely phototrophic or heterotrophic (phagotrophic)but a significant (still poorly known) fraction of flagellates aremixotrophic combining both phototrophy and phagotrophy(70 88) Flagella in phagotrophic flagellates are employed formotility and prey capture Most free-living flagellates possessone to four flagella (typically one or two) that can be manytimes the length of the cell itself Although there is tremen-dous species diversity among flagellated protozoa many ofthese species have broadly overlapping ecologies As a groupflagellates are the most numerically abundant protozoa ofboth benthic and pelagic ecosystems and they are fundamen-tally important as consumers of bacteria cyanobacteriaand other eukaryotes Flagellated protozoa within the nano-plankton size class (2ndash20 microm) are often counted as a singleassemblage in plankton studies because morphological details
apparent using light microscope are insufficient to distinguishamong the many species This assemblage has been variouslyreferred to by a variety of names and acronyms including het-erotrophic nanoplankton (HNAN HN) heterotrophic flag-ellates (Hflags) heterotrophic nanoflagellates (HNF) andmicroflagellates Heterotrophic nanoplankton is the mostaccurate term for most methodologies employed to countthese cells because it does not require visualization of flagella(which are often lost from these small cells during preparationfor microscopy) The confusion over terminology is partly aconsequence of the different methodologies that have beenused to count these species and partly a consequence of thefact that these species were first studied prior to the wideacceptance of the size convention of Sieburth et al (84)
Heterotrophic flagellates in the microplanktonic size class(20ndash200 microm mostly dinoflagellates) are important consum-ers of phytoplankton in pelagic ecosystems Many of thesespecies are capable of the production of large pseudopodialnets the pallium that can engulf prey (particularly diatoms)significantly larger than the diameter of the dinoflagellatetheca (176) This behavior and its ecological significance forenergy flow in plankton communities have been recognized
FIGURE 5 Body plans and size ranges of protozoa These micrographs depict amoeboid (andashg) flagellated (hndashj) and ciliated (kndashm) forms ofprotozoa From Caron et al (173) Markers bars are 5 (i) 10 (h) 20 (c j l) 30 (a) 50 (k m) 100 (b f) 500 (d) and 1000 (e g) micromdoi1011289781555818821ch422f5
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-13
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
only within the past few decades (177) These species areoften abundant in waters where diatoms dominate (178179) In such situations heterotrophic dinoflagellate biomasscan be similar to that of ciliated protozoa (180)
Ciliates are generally the most recognizable form of pro-tozoa to nonspecialists Nonetheless ciliates are a diversegroup morphologically as well as ecologically (181 182)The degree of ciliature on these species can range from uni-formly ciliated to totally devoid of cilia during most of theirlife cycle to ciliature restricted to specific regions of thecell In addition cilia can fuse to form various complexstructures (eg ciliary membranes or veils cirri) that assistin locomotion food capture or attachmentCiliates are abun-dant and ecologically important species in both benthic (8)and pelagic (183) ecosystems and can consume a variety ofprokaryotic and eukaryotic prey Together with the heterotro-phic dinoflagellates these species are the dominant consum-ers of phytoplankton in many pelagic ecosystems (184) andas a consequence they form an important trophic link to met-azoan zooplankton (185 186)
Diversity and Biogeography of Protists FromMorphology to DNA SequencesUntil relatively recently the biodiversity of marine protistanassemblages was not generally considered a controversialtopic It was generally accepted that while all species of pro-tists certainly have not been identified representatives ofmost types of algae and protozoa had been observed anddescribed if not actually brought into culture in the labora-tory Direct sequencing of 18S rRNA genes from environ-mental samples (as described in ldquoMolecular Phylogeny andMetagenomics Field Applicationsrdquo) has changed that viewInitial forays into environmental DNA indicated a muchgreater diversity of protists than previously documented usingclassical approaches of culture and microscopy (81 187ndash191) Numerous publications over the past decades haveexpanded these findings (see [192] for a recent global analy-sis) which are highly analogous to discoveries in marine pro-karyote research as described already
Previously uncharacterized protistan diversity has nowbeen documented at virtually every level of eukaryoticorganismal classification Some of these findings couldhave been expected but some have been very unexpectedFor example a much greater diversity than noted previouslyhas been observed among small (lt10 microm) protists in plank-tonic ecosystems These species generally possess few distinc-tive morphological features and one could expect that manycryptic species might be present among these small morpho-types The molecular ldquodiscoveryrdquo of this eukaryotic diversityhas stimulated progress on the isolation and descriptionof new species and genera of minute algae and protozoa(193ndash197)
Analyses of environmental samples have also indicatedthe unanticipated existence of novel 18S rRNA gene sequen-ces that imply the presence of novel lineages of eukaryotes innatural protistan communities (198ndash200) These are sequen-ces for which there are apparently no known described orcultured species The degree to which these sequences dif-fer from sequences of known sequenced eukaryotes impliesthat some of these lineages may be distinct at the level ofphylum (201)
These findings have raised basic questions and somedebate concerning the true diversity and biogeography ofprotistan assemblages in natural ecosystems (202ndash204) thevalidity of the many novel phylotypes or cryptic species that
are being documented (205) the potential importance andsignificance of the many rare taxa that characterize thesecommunities (206) and the implications of these findingsfor the ecologicalbiogeochemical roles that protists playin aquatic ecosystems For example it was postulated andsubsequently confirmed that some novel alveolate lineagesreported from marine ecosystems make up a suite of parasiticprotozoa whose ecological importance may have been signifi-cantly underestimated in the past (207) Deciphering theidentity and significance of these many unknown phylotypeswill constitute a significant effort for protistologists in thefuture (80 85)
Life Histories and Ecological Strategies of ProtozoaAbundance prey type and life histories all vary tremendouslyamongmarine protozoa The smallest species (eg many flag-ellates) tend to be the most abundant and widely distributedin the world ocean Indeed many of these species may beglobally distributed (208) Most nanoplanktonic flagellateshave potentially rapid rates of grazing and growth Underoptimal conditions these species can divide by binary fissionseveral times a day and thus dramatic increases in their pop-ulations can take place in response to favorable conditionswithin a few days (209) Many of these species are capableof surviving for limited periods of time without food andhave developed a variety of physiological or life cycle strat-egies to cope with these events (209) However in contrastto some bacteria that may remain viable through long periodsof starvation (210) protozoa will expire or encyst in responseto low food abundance and thus ldquoboom-and-bustrdquo popula-tion changes are characteristic of small flagellates
On the other end of the size spectrum from the ratherubiquitous nanoflagellate species many of the largest proto-zoan species (polycystine and phaeodarian radiolaria plank-tonic foraminifera) are exclusively oceanic (ie do notsurvive in most coastal environments) andor have specificlatitudinal and depth distributions Thus the abundancesof these latter species may range from undetectable to max-imal abundances of gt105 individualsm3 Dramatic changesin the abundance of these protozoan taxa also can be relatedto changes in prey abundance physicalbehavioral aggrega-tion or to periodicity of life cycle events For example theplanktonic foraminifer Hastigerina pelagica reproduces on alunar cycle and thus abundances (and life stages) of this spe-cies in oceanic waters can vary considerably over the courseof a month (211) In general life cycles for the large amoe-boid protozoa are lengthy and complex (for single-celledorganisms) with life spans unknown for many species(attempts to culture them in the lab have so far been unsuc-cessful) but estimated to be on the order of months to per-haps years (175)
The abundance and activities of microplanktonic hetero-trophic protists (mostly ciliates and heterotrophic dinoflagel-lates) tend to be somewhat intermediate to those ofnanoplanktonic flagellates and the larger amoeboid formsThese species are present in the majority of marine ecosys-tems and collectively play an important role in the controlof phytoplankton biomass (and probably the abundance ofnanoplanktonic protozoa although there is little informa-tion on this topic) in waters throughout the world oceanLike small flagellates ciliates and dinoflagellates reproduceprimarily by binary fission but their maximal growth ratesare typically slower (one division a day is typical) and theirlife cycles often include sexual phases that allow for geneticrecombination
422-14 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
VIRUSESViral Abundance and General PropertiesViruses are simple biological agents typically 20ndash200 nm indiameter composed of a nucleic acid genome in a proteincoat that infect cells and ldquocommandeerrdquo the cellrsquos machineryto make more viruses which are released into the environ-ment when the host cell lyses or bursts A particular virus isthought to be capable of infecting only a narrow range of hosts(usually one species sometimes a genus rarely broader)Although some early studies had isolated viruses from thesea therewere no data prior to the 1980s showing such viruseswere very abundant and more significantly no evidence thatinfection was occurring in any important part of the planktoncommunity It wasnrsquot until the late 1980s that electronmicro-scopy with suitable concentration methods showed thatviruses are extremely abundant similar to or even higherthan bacterial abundance (212 213) Interestingly themost suitable transmission electron microscopy (TEM)approach used by Bergh et al (described in detail by [214])was actually similar to a direct centrifugation method devel-oped in 1949 (215)
TEM studies of viruses permit high-resolution images andobservation of viral morphology showing features such ashead diameter tails sheaths and tail fibers (Fig 6 collage
in upper left) Each type of virus has a fixed morphology(unlike bacteria that are potentially more plastic in theirappearance) and hence a coarse measure of viral diversityhas been possible by cataloging morphologies Studies thathave done so have found dozens or more different morpholo-gies of marine viruses in a given sample (217ndash219) A largeproportion resemble bacteriophages which are viruses thatinfect bacteria
The availability of brightly fluorescent nucleic acid stainsand high-porosity fine pore size (002 μm) filters made fromAl2O3 have made it easy to accurately count viruses by epi-fluorescence microscopy (38 220 221) Epifluorescence per-mits abundance estimates but does not allow observation ofviral morphology as viruses are below the resolution limitof light microscopy viruses are visible only as sources of light(like stars in the night sky see Fig 6 lower micrograph) Anextension of manual epifluorescence counts is the use of flowcytometry to count viruses now commonly employed in somelabs (222)
Virus abundance has been found to be closely related tobacterial abundance with a virusbacteria ratio typically10ndash301 A typical oceanic profile of bacterial and viral abun-dance is shown in (Fig 6 panel on right) This tight ratioand the strong correlation to bacterial abundance in relationto weaker correlations to chlorophyll have been cited as
FIGURE 6 Marine viruses (upper left collage) viewed by transmission electron microscopy These are cyanophages in the groups myoviridae(right and bottom) siphoviridae (left) and podoviridae (top) Scale bars represent 100 nm From Sullivan (216) Epifluorescence micrograph(lower left) of SYBR green I stained viruses (small fluorescing objects) and bacteria + archaea (large fluorescing objects) Depth distribution ofvirus and bacteria + archaea abundances obtained by epifluorescence microscopy of SYBRGreen stained cells in the central San Pedro BasinCalifornia (11 August 2000) doi1011289781555818821ch422f6
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-15
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
evidence that most marine viruses infect bacteria rather thaneukaryotic phytoplankton (217 218 223 224) though thereare also many important viruses of phytoplankton and otherorganisms with significant impacts on primary productivityblooms and ecosystem function (225ndash227)
The first demonstration of the activity of marine viruseswas presented by Proctor and Fuhrman (228) who showedby electron microscopic examination that viruses wereactually infecting marine bacteria and cyanobacteria at ameasurable rate Because only the final portion of the viruslife cycle is visible by electron microscopy (when the virusesare assembled and ready to lyse the host cell) only a very smallfraction of the infected cells can be counted as infected at anygiven time Proctor and Fuhrman (228) reported that only afew percent of the total bacteria from coastal waters and theSargasso Sea were visibly infected but they interpreted thedata with a model that implied the actual fraction of the totalcommunity that was infected was much higher They con-cluded that the total fraction of bacterial mortality attribut-able to viruses is roughly 10ndash40 Subsequent studies haveused refined versions of that same general approach andmodel (229 230) and numerous studies have used a varietyof alternative approaches to estimate viral activitymdashall yield-ing basically the same general conclusion (although themodel parameters need to be adjusted for cyanobacteriawhich on average seem to be infected less than bacteria)These approaches include (a) calculation of virus turnoverfrom decay estimates (231) (b) measurement of viral DNAsynthesis by incorporation of tritiated thymidine (232) (c)observation of effects of added viruses (233 234) (d) estima-tion of bacterial mortality in the absence of protists (e) use offluorescent viruses to estimate production by an approachanalogous to isotope dilution (38) and (f) dilution of virusesin filtered samples to estimate viral production (235) Theoverall consensus of these studies is that the initial estimateswere basically correct that is that viruses are responsiblefor about 10ndash40 of the bacterial mortality in most marinesystems (reviewed by [218 223 226 236 237]) Howeverthere is still some disagreement about the higher estimatesand it is likely that 40 mortality from viruses alone is nottypical for most marine systems
Most of the foregoing work has focused on the viral infec-tion of bacteria primarily thought to be heterotrophicalthough the original report by Proctor and Fuhrman (228)also noted the occurrence of cyanobacterial infection Sev-eral subsequent studies focused on phytoplankton includingthe potential effect of viruses on the termination of phyto-plankton blooms of Emiliania and Phaeocystis and the likelyimpact this might have on release of climate-active gasessuch as dimethyl sulfide (225 227 238ndash247) Althoughdetails are beyond the scope of this chapter viruses arethought to infect virtually all marine organisms with poten-tially significant impacts from zooplankton to whales (226)Detailed studies of viruses infecting cyanobacteria such asSynechococcus have shown differences in viral host specific-ity particularly toward coastal and oceanic host strainsand some occasional high virus abundances (to 105ml) asmeasured by most probable number (MPN) cultivation tech-niques in the Gulf of Mexico near Texas (248 249) Othervirus cultivation studies with Prochlorococcus and Synecho-coccus in oligotrophic waters of the Sargasso Sea showedan interesting pattern of cross-infection between these gen-era by some virus types (suggesting gene flow among theseorganisms) but generally low MPN estimates of abundanceto 103ml even when cyanobacterial abundance was near105ml (216 250)
Overall the consensus emerging from direct comparisonsof viral-mediated mortality and grazer-mediated (ie micro-zooplankton) mortality have indicated that viral lysis of bac-teria constitutes a significant fraction of total mortality ofthis assemblage while the mortality of phytoplanktonappears to be dominated bymicrozooplankton inmost instan-ces (251 252)
As mentioned viruses have significant morphologicaldiversity as observed in TEM studies It is also possible toinvestigate aspects of their genetic diversity In early workon this topic Wommack et al (253) and Steward et al(254) observed the diversity of viral genome lengths in a fieldsample by pulsed field gel electrophoresis with viral genomesranging 25 kb to gt300 kb in length Field results show thatthe viral community composition is dynamic in space andtime with clear changes in the banding patterns over seasonsand locations in Chesapeake Bay (253) between oceanbasins and subsequent to dinoflagellate blooms (254) andwith depths to 500 m at one location (255)
Although viruses do not all share a set of core genesthat allows a universal viral phylogeny (comparable to SSUrRNA gene in cellular organisms) genetic diversity amonga single group of viruses can be examined by sequence analysisof shared genes within the group An example is the g20 geneinT4-like cyanophages that has shown extremely high diver-sity even among very closely related viruses and has demon-strated geographic and seasonal variation (256) A secondexample is g23 which is found broadly in diverse and wide-spread T4-like phages (257) and can have seasonally repeat-ing patterns (258) as well as short-term rapid dynamics thatcorrelate to those in bacteria (259)
Viral MetagenomicsAs with cellular organisms metagenomics can be used toexamine viral diversity and genetics without themany restric-tions of cultivation Viruses can be collected by selective fil-tration and concentrated by tangential flow filtration orflocculated with iron chloride (260 261) then their collec-tive metagenome can be extracted linker amplified and ana-lyzed by sequencing (262 263) Viral metagenomic studiesare particularly challenging because the large majority ofsequences have no annotated matches in any databases butso far results from marine samples around the world haveshown extremely high diversity and variations with depthlocation and time presumably with highly dispersed typesselected by local conditions (262ndash266) Because metage-nomes are best interpreted when there are representative cul-tures available the best matches of viral metagenomesoriginally tended to be to the few viral isolates infecting trulycommonmarine bacteria like cyanophage infecting Synecho-coccus and Prochlorococcus (267) However the developmentof dilution-to-extinction cultures representing common het-erotrophic marine taxa like Peligibacter (a member of theSAR11 clade) and SAR116 has allowed isolation from sea-water of viruses infecting these common organisms and theseisolates have indeed been found to be highly abundant inmarine viral metagenomes (268 269) New approaches tointerpret the results include clustering the proteins independ-ent of known proteins to compare samples to each other andtry to find environmental factors driving viral communitychanges (270) Such analysis of a large global data set (TaraOceans expedition) has shown that extensive sampling hascome close to reaching the total diversity in tropical and tem-perate waters of such viral protein clusters (which essentiallyrepresent various viral protein motifs not all viral proteintypes) and that viruses appear to be directionally dispersed
422-16 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquodownstreamrdquo in major ocean currents as one may expectfrom first principles but also supporting the ldquoseed bankrdquohypothesis of viral biogeography (271)
Viruses and Host DiversityViruses are themselves thought to be instrumental in drivingincreased diversity of their microbial hosts via a hypothesizedprocess often called ldquokill the winnerrdquo This is because viralinfection is host-specific and density-dependent the latterbecause viruses diffuse from host to host so an abundanthost is more likely to pass on infection than a rare oneThis means that if an organism becomes abundant andblooms winning the competition for resources it becomesmore susceptible to a viral epidemic This would benefit therarer organisms and thus help foster diversity (223 272273) However bloom scenarios as described above involvesystems far from steady state and the formal theory of Thing-stad and Lignell (274) has interesting steady-state solutionswhere several viruses infect several hosts stably over time(via trade-offs between growth rates and viral susceptibility)which may occur at the strain or species level (274) There issome experimental evidence that viruses have effects on nat-ural marine microbial community composition weaker thanthe kill the winner (bloom version) hypothesis would sug-gest (275 276) so something resembling the steady-statecoexistence described in the model may in fact be commonThere also appear to be processes that foster coexistencebetween viruses and hosts but the mechanisms are largelyspeculative (223 277 278)
Viruses may also be directly involved in host genetic diver-sity because they can be the agents of genetic exchangebetween microorganisms (223 273) This often involvesthe viral lifestyle known as lysogeny whereby viruses survivewithin host cells as DNA only integrated into the host chro-mosome and being reproduced each time the host divides Ahost harboring such a genome is called a lysogen becauseunder conditions of stress to the host cell a genetic switchmay cause the viral genome to initiate the lytic process pro-ducing many progeny viruses and bursting from the hostLysogeny is a very common property occurring in a signifi-cant part of the bacterial community (279 280) althoughthe incidence of induction of the lytic phase in nature is appa-rently low (281) Overall lysogeny is poorly understood butthought to have both positive and negative impacts on themicrobial community (282) Recent results suggest that ben-efits of being lysogenic in highly seasonal polar seas leads tofundamental difference between polar and other marine viralcommunities (283)
Viruses and the Microbial LoopAs part of the food web viruses occupy a unique positionThey infect host cells that are mostly thought to be heterotro-phic bacteria and by doing so they typically burst the hosts torelease progeny viruses and cellular debris But what is the fateof this material Viruses themselves do not last indefinitelyand a simple steady-state assumption implies that from eachburst of viruses (typically 20ndash100 per lytic event) only onesuccessfully infects another cell The rest are inactivatedand broken down by sunlight (UV and visible exposure)and enzymatic attack (284) or consumed byminute phagotro-phic protists (285) thus reentering the food web as substratefor bacteria or food for protistan consumers Experiments incontrolled laboratory systems and field studies with radioac-tively labeled viral lysis products have supported the conclu-sion that most of the organic matter released by the viralinfection is either taken up by bacteria or respired (286 287)
Modeling this process as part of the microbial loop showsthat viral lysis represent a sort of side loop that has the neteffect of remineralizing a significant amount of the carbonand nutrients that enter the bacteria-protist part of themicro-bial loop (Fig 7) A theoretical numerical steady-state modelcomparing a systemwith no viral activity to onewhere virusesare responsible for 50 of bacterial mortality showed that thesystem with viruses had 33 more bacterial production andrespiration than the virus-free system implying that theviruses had the effect of permitting the bacteria to processmore of the primary production than they would otherwise(223) Although 50 is a high number unlikely to be com-mon in the sea this model nevertheless illustrates that virusescan reduce the amount of energy reaching higher trophic lev-els The implication is that viruses lead to increased bacterialactivity at the expense of the larger organisms
MAJOR ENVIRONMENTAL CONTROLSLight Temperature and PressureTemperature has an important potential influence on bio-chemical reactions and therefore on biological processesin general Most ocean waters fall in the range of ndash2degC to30degC with obvious exceptions in hydrothermally heatedareas Temperature has long been known to be a regulatingfactor for the growth of heterotrophic microbes
In temperate waters it has been established that microbialactivity is generally much higher in warm summer waters thanin winter (55) The relationship is not simple howeverbecause multiple factors act at the same time Some contro-versy still exists regarding the highest and lowest extremesfor marine bacterial growth although there is broad agree-ment that bacteria grow gt100degC at hydrothermal vents andltminus5degC in sea ice brines Pomeroy et al (57) noted the inter-esting observation that bacteria seem particularly inhibitednear the freezing point of seawater (ca ndash22degC) comparedto eukaryotic phytoplankton This effect results in polarspring phytoplankton blooms that accumulate organic car-bon in advance of the response of the bacterial communityand development of the microbial loop and perhaps lead toenhanced benthic-pelagic coupling (288)
The relationship between temperature and the growth rateof marine phytoplankton was described broadly in a now
FIGURE 7 Modification of the microbial loop concept that incor-porates the functional role of viruses Export can be via predation orsinking From Fuhrman (223)doi1011289781555818821ch422f7
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-17
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
classic paper by Eppley (58) and later Goldman and Carpen-ter (289) Temperature was shown to exert a strong and directeffect on themaximal growth rates of these species withmax-imal intrinsic growth rates at 0degC generally lt1 divisiondaywhile growth rates at 30degC may be gt4day These relation-ships indicate the maximal rate that might be attained byphytoplankton at these temperatures but they do not takeother factors into account (nutrients light) While warmtemperature ostensibly allows more rapid growth it createshydrographic conditions that typically give rise to nutrientlimitation of algal growth Thus some of the coldest watersin nature witness some of the most massive phytoplanktonblooms (290) while warm oceanic gyres represent some ofthe most oligotrophic areas of the ocean The multiple andoften conflicting effects of temperature on the growth of nat-ural phytoplankton assemblages limit the accuracy of presentpredictions regarding how primary producers will respond toclimate change (291)
An analysis of the effect of temperature on the growth ofheterotrophic protists has indicated that the response is qual-itatively similar to that of phytoplankton but with a twistThe growth rates of at least some protozoa can exceed thoseof phytoplankton at warmer environmental temperaturesbut the opposite effect is apparent at very low environmentaltemperature This differential effect of temperature on thegrowth of phototrophic and heterotrophic protists was basedon a large meta-analysis of published protistan growth (59)That analysis demonstrated that the maximal growth ratesattained by phototrophic protists could exceed the maximalgrowth rates attained by heterotrophic protists (all otherpotential growth-limiting factors not considered) Thereforeprotozoan growth rates may be constrained to a greater degreeat low environmental temperature than rates for phytoplank-ton If so then phytoplankton bloomsmay get a head start ongrazers during spring in polar ecosystems This scenario is con-sistent with information on seasonal biomass changes andmicrozooplankton herbivory in the Ross Sea Antarctica(292 293) but there are still too few data to fully vet thishypothesis
The importance of high pressure on bacterial growthgained considerable attention in the late 1960s when thedeep sea submersible Alvin was accidentally lost overboardwith its hatch openWhile no lives were lost in this accidentsome workmenrsquos lunches sank to the bottom (sim1500 m)inside the submersible Alvin was recovered after 10 monthsand interestingly there was a waterlogged lunch containingapples bologna sandwiches and broken vacuum bottleswith broth that all appeared hardly degraded and tasted palat-able Yet when placed in a refrigerator on the ship these itemsdegraded relatively quickly Initially pressure was thought tobe the preserving factor as the sea floor temperature was sim-ilar to the refrigerator temperature and the only major differ-ence would be pressure (294) Following this observationa series of experiments to measure degradation of variousorganic materials left in the deep sea for extended periodsindicated that degradationwas typically significantly reducedimplying that pressure reduces the degradation rates (295)Nonetheless changes in the protozoan community of naturaldetrital material sinking to the deep ocean floor indicate thatthe microbial community can respond relatively quickly insome situations (296) Deep sea microbiology has advancedconsiderably yet it is still difficult to interpret results withrespect to actual in situ rates of naturally occurring organicmatter Deep sea bacteria adapted to high pressures havebeen isolated that are barophilic (also called piezophilic)meaning that they prefer high pressures and have reduced
activity at lower pressures whereas others are barotolerant(piezotolerant) tolerating but not preferring high pressuresfor example see (297 298)
Few data are available on barotolerantbarophilic marineprotozoa Protozoa certainly exist and grow at great oceanicdepths but measurements of in situ growth rates for these spe-cies do not yet exist Measurable protozoan numbers havebeen documented in the deep-sea sediments for more than30 years (299 300) and viable protozoa have occasionallybeen cultured from these environments (301ndash305) butvery few direct measurements of the activities of these speciesin situ have been reported (306) A few protozoa have beenisolated that will grow at high pressure (302 303 307) andprotists that appear to be unique to the deep ocean havebeen observed either directly or through the analysis ofDNA sequences (308ndash311) but possibly the best direct evi-dence that protozoan activity takes place at the high pressurescharacteristics of the deep sea are experimental and observa-tional work noting the stimulatory effect that detrital deposi-tion has on some components of the protozoan community(296 312) These observations indicate a diverse and activeprotozoan fauna of the deep ocean although their biogeo-chemical significance is largely uncharacterized at this time
Dissolved and Particulate Organic MatterBacteria and archaea are thought to be by far the most impor-tant organisms with respect to the processing of dissolvedorganic matter (DOM) and nonliving particulate organicmatter (POM also called detritus) in the ocean While theremay be some uptake of DOM by protists particularly forgrowth factors needed in trace amounts such as vitamins(313) the bulk of this material is probably utilized by bacteriaand archaea (314) including the smallest cyanobacteriumProchlorococcus (315 316) which thus may be considered amixotroph Due to their small size bacteria have extremelyhigh surfacevolume ratios and combined with their over-whelmingly high abundance an extremely high integratedsurface area Protozoa tend to obtain the organic materialsthat they require for growth from their prey rather thanthrough the uptake of DOM Overall protozoa tend to besources of dissolved and detrital organic substances throughthe excretion of unassimilated prey biomass in expelledfood vacuoles
Particulate organic matter is not directly available as sub-strate to bacteria These substances must first be reduced tosmall molecules that can be transported into the cell Thisis accomplished by the production of extracellular enzymes(note that few if any large polymers are directly taken up bybacteria with the possible exception of DNA) Hydrolyticenzymes produced by bacteria (and almost certainly archaea)break down polymers like proteins polysaccharides andnucleic acids As with DOM POM is composed of a complexmixture of compounds that vary in their susceptibility to bac-terial degradation and utilization
Particulate material in the water column serves not only asbacterial substrate but also as substratum POM occurs in thewater column across a huge size spectrum from micrometersup to some detrital aggregates more than 1 m in diameter(317) Much of this particulate material is in a constant stateof flux with colloidal material constantly coalescing andaggregating to form new or larger particles (318) as microbialdegradation acts simultaneously to remineralize this materialDetrital particles that attain macroscopic size either by directformation (317 319 320) or via accretion and aggregation(321) and are often called marine snow or macroaggregatesMarine snow particles are readily colonized by bacteria and
422-18 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
many other microorganisms and their abundances on theseaggregates can be orders of magnitude greater than abundan-ces in an equivalent volume of the water surrounding theaggregates (320 322ndash324) and their diversity and highlyconcentrated abundances on detrital aggregates implies thatthere may be stronger trophic coupling and efficient energyelemental cycling relative to these processes among free-living microorganisms in the surrounding waters This specu-lation has resulted in a number of formalizations of the poten-tial impact of aggregate microbiology on biogeochemicalprocesses in the ocean (325 326) Most definitions of bio-films include aggregated particulate and these particles areoften studied within the biofilm conceptual model (see chap-ter by Lawrence et al in this section)
Inorganic Nutrients
Macronutrients (N P)Virtually all heterotrophic organisms in the ocean contributeto the pool of available macronutrients via the excretion ofmetabolic wastes and protozoa contribute significantly tothis process Macronutrients are often in excess within proto-zoan prey relative to the consumersrsquo needs because much ofthe prey carbon is respired to produce energy for metabolismand growth ldquoExcessrdquo nutrients are eliminated as eitherorganic compounds or more commonly as ammonium (forN) and phosphate (for P) The importance of the match(or mismatch) between predator and prey stoichiometry hasbeen demonstrated experimentally and repeatedly in marineand freshwater ecosystems and has been elevated to a funda-mental ecological principle governing elemental and energyflow in aquatic food webs (327) Based on this reasoningCaron et al (328) concluded that bacterivorous protozoaplay an important role in nutrient remineralization andrelease in the ocean Bacterial biomass is typically rich in Nand P relative to other organisms so bacterial predators expe-rience large excesses of these elements relative to their growthneedsMoreover bacterivorous protozoa should be dispropor-tionately important in nutrient remineralization amongsingle-celled eukaryotes because as the smallest protozoa inaquatic ecosystems they have high weight-specific metabolicrates (329) Thus both stoichiometric and allometric rela-tionships implicate small bacterivorous protozoa as impor-tant sources of remineralized nutrients in the plankton
The reasoning also holds true for bacterial utilization oforganic compounds and it has led to some interestingfindings and conclusions regarding the role of bacteria innutrient cycling in the ocean Bacteria are important sourcesremineralized nutrients when they are consuming N- orP-rich substrate Contrary to this traditional view howeverheterotrophic bacteria are often strong competitors for inor-ganic nutrients under many situations largely because bacte-rial biomass is richer in N and P than much of the organicmatter they consume in nature so they require additional Nand P to produce their biomass (summarized by [330]) Exper-imental studies have demonstrated the uptake of ammoniumby bacteria other than cyanobacteria using stable isotope 15N(331) and also the short-lived radioisotope 13N (332) oftenfinding that bacteria may be responsible for one third of the Nor P uptake Additionally there is evidence that bacteria insome oceans like the Sargasso Sea are limited by phosphorus(333 334) Growth limitation of bacteria by phosphorusappears to be a common phenomenon of freshwater ecosys-tems (335 336)
N2 fixation is restricted to prokaryotes and in the oceanwater column it was thought for many years to be done
primarily byTrichodesmium a warm-water colonial cyanobac-terium that blooms sporadically andRichelia a cyanobacterialsymbiont that lives within certain diatoms (337) Some uni-cellular cyanobacteria also contribute significantly to globalN2 fixation (338) and molecular biological data suggestthat a variety of other bacteriamdashnot phytoplanktonmdashmayalso be fixing nitrogen in seawater (339) An exciting recentdiscovery in this field is that a bacterial nitrogen fixer origi-nally known only from nifH sequences UCYN-A has hadits genome sequenced by parallel pyrosequencing and wasfound be a cyanobacterium (Candidatus Atelocyanobacteriumthalassa) lacking the apparatus to generate oxygen as well asother pathways (340) Further study showed it to be symbioticwith a marine alga (341)
Micronutrients (Trace Metals Growth Factors)Bacterial growth may often be limited by macronutrients asdescribed and sometimes by trace nutrients particularlyiron that occur at very low concentrations in seawater(342 343) The importance of Fe as an element limiting pri-mary production in some oceanic regions has come underclose scrutiny and extensive experimental investigation inrecent years Many bacteria have special uptake mechanismsto utilize extremely low concentrations of iron includingsiderophores (released compounds that bind iron and arethen specifically taken up) Even if bacteria are capable ofgrowth at low Fe concentrations growth may be less efficientKirchman et al (344) found that Fe limitation stronglyreduced bacterial growth efficiency in a cultured marine bac-terium (ca 50 efficiency Fe replete but lt10 when Fedeplete) Thus the effects may be complex Marine N2 fixa-tion also may be limited by the availability of Fe becausenitrogen fixers have high Fe requirements
Protozoa tend to be a source rather than a sink for Fe in theocean analogous to their role in macronutrient remineraliza-tion Protozoan grazing activity has been shown to releaseiron from bacterial biomass thereby relieving Fe limitationin co-occurring phytoplankton (345) a mechanistic demon-stration of the process using protozoa growing on Fe-repletebacteria However it has also been reported that Fe-limitedbacteria may contain insufficient Fe for the bacterivorous pro-tozoa that consume them thus leading to Fe limitation in theprotozoan as well (346)
OxygenHypoxic and anoxic regions of the ocean are expected to sig-nificantly expand in the coming decades as a consequence ofchanges in ocean stratification resulting from global climatechange (347) As oxygen becomes depleted bacteria havethe capability to utilize a series of alternate electron acceptorsCompositional changes in the bacterial assemblage accom-pany these changes in metabolic activity but are beyondthe scope of this chapter Many of the studies on benthicmicrobiology have focused on redox reactions that occurthere Oxygen is usually consumed by sediment microbialactivity much faster than the rate at which it can be replacedby diffusion so most sediments especially organically richones are anaerobic below a few mm (fine-grained muds) orcm (coarser sediments) from the sedimentwater interfaceexcept where animals ventilate the benthos through tubesWhen oxygen is absent bacteria and archaea use alternateelectron acceptors such as nitrate nitrite oxidized Fe orMn organic matter or sulfate The use of nitrate or nitriteas an electron acceptor with concomitant production of N2
gas (denitrification) results in net loss of biologically availablenitrogen Bacteria that perform this reaction are typically
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-19
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquofacultativerdquo organisms that can switch from oxygen to nitrateor nitrite Organic matter may be reduced by fermentationreactions in which part of an organic substrate is oxidizedwhile another is reduced fermenter organisms are also typi-cally facultative Sulfate is a particularly important alternateelectron acceptor in marine systems because it occurs as thesecond most abundant anion in seawater (behind chloride)typically 28 mM Its use as an electron acceptor leads to theproduction of elemental sulfur or sulfide The use of sulfateas an electron acceptor yields much less energy than oxygenor nitrate and sulfate-reducing bacteria are strict anaerobesunable to use oxygen (often strongly inhibited by oxygen)Methane can be produced by methanogenic archaea whichare all strict anaerobes using CO2 as an electron acceptor(see Findlay and Battin this section for further discussion)
Reduced compounds such as sulfide are produced by theuse of these alternate electron acceptors and they diffuse awayfrom their source When they diffuse to environments with astrong oxidant like oxygen they may be oxidized by chemo-trophic bacteria in energy-yielding reactions Similarlyammonium can be oxidized to nitrite and then nitrate by che-motrophic nitrifying bacteria and the nitrate can diffuse intoanaerobic layers and become denitrified to N2 Such coupledoxidation-reduction reactions can lead to accelerated biogeo-chemical cycling particularly in sediments
It was originally believed that relatively few free-livingspecies of protozoa inhabit anaerobic and hypoxic marineenvironments This view has changed based on (largely)molecular analyses of some of these environments (308348ndash351) A significant number of previously undescribedeukaryotic phylotypes have been observed in anoxic environ-ments High protozoan biodiversity in the absence of molec-ular oxygen is not completely unexpected because otheranaerobic environments such as some gut environments(eg cockroach guts and cattle rumen) have notoriouslyabundant and diverse protozoan fauna but marine anaerobicenvironments have been difficult to access and sample prop-erly Many anaerobic protozoa exist together with symbioticbacteria (352 353) or possess unique organelles such as thehydrogenosome (354)
MAJOR BIOTIC CONTROLSThe Trophic Activities of Protozoa Predation andTop-Down ControlsPredator-prey relationships are fundamental interactionswithin biological communities linking production of bio-mass on the one hand with removal on the other hand Estab-lishing the trophic relationships among microorganisms andquantifying the rates at which they take place constitutemajor endeavors within marine microbial ecology As notedpreviously protozoa consume a wide variety of prokaryoticand protistan prey Numerous laboratory and field measure-ments have been made throughout the past quarter centuryto attempt to quantify the rates of bacterial and phytoplank-ton consumption The methodology for accomplishing thesestudies has varied and a number of approaches are presentlyemployed
Methodologies for estimating microbial grazing are diversebut generally fall into one of two experimental approachesperturbation experiments or tracer experiments Perturbationexperiments (size fractionation metabolic inhibitors anddilution technique) rely on manipulation of the consumeror prey assemblages in somemanner such that the effect is evi-dent from changes in the abundance of the prey population
during incubations relative to control treatments that receiveno manipulation Tracer experiments (radioactively labeledprey fluorescently labeled prey stable isotopendashlabeled pray)attempt to use labeled prey or a prey proxy to quantitativelyfollow the movement of prey biomass into consumers Forsummaries of these methods see (74 355)
Perturbation experiments have entailed a variety of mech-anisms to ldquodecouplerdquo predators from prey in natural microbialcommunities For example size fractionation uses various sizefilters to remove an entire size class of grazers from awater sam-ple leaving the prey to grow in the relative absence of thosepredators during subsequent incubations Unfiltered samples(controls) indicate the growth of prey in the presence of pre-dation Predation impact is obtained by the difference ingrowth of the prey between these treatments This methodhas been used to examine predation on cyanobacteria heter-otrophic bacteria and minute phototrophic eukaryotesAlternatively metabolic inhibitors have been employed tohalt predation (or in some cases prokaryote growth) insteadof physical removal by filtration
The dilution technique (356) has become a commonmethod for examining grazing mortality of phytoplanktonby microzooplankton (which is comprised predominantly ofprotozoa) This method relies on the dilution of herbivorouszooplankton in a series of subsamples (and concomitantmeasurements of phytoplankton growth in each subsample)to estimate the grazing mortality of the herbivores
Tracer experiments have used both radioactively labeledprey and fluorescently labeled prey to examine predation onbacteria and phytoplankton Radioactive labeling has beenaccomplished using 14C- or 3H-labeled organic compounds(for bacteria) or 14C-bicarbonate (for phytoplankton) Theuse of radioisotope approaches has diminished with improve-ments in the number and types of fluorescent compoundsavailable for this work Fluorescently labeled prey havebeen labeled with a variety of compounds that allow theprey to be observed readily (even inside predators) by epi-fluorescencemicroscopy (357 358) Alternatively a decreasein the total number of fluorescent prey in a sample indicatesthe rate of removal of the prey assemblage
Both perturbation experiments and tracer experimentsdescribed have inherent advantages and disadvantages Forexample perturbation experiments are fairly straightforwardto conduct but they may introduce artifact by removingpotentially important sources of nutrients or organic com-pounds released by predators (359) On the other hand thedilution technique is very labor-intensive but one advantageof this method is that it provides the opportunity to determineboth phytoplankton growth and grazing mortality in a singleprocedure Most tracer experiments are fairly easy to conductand generally do not perturb the food web being studied butfeeding selectivity for or against the tracer (relative to naturalprey) may produce spurious results In addition some of thesemethods provide a measurement of the activity of the entiregrazer assemblage and others are applicable for examiningthe activity of specific taxa For example size fractionationapproaches examine the effect of the removal of an entiresize class of organisms (a community-levelrdquo measurement)whereas fluorescently labeled prey can be used to examinethe ingestion rate of a particular species by observing theuptake of labeled prey into that species (a species-levelmeasurement)
Predation rates of numerous marine protozoa have beenexamined in the laboratory From these experiments a gen-eral knowledge of the rates of consumption by these specieshas been accumulated Feeding in most protozoa shows a
422-20 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
functional response to the availability of prey that is similar tothe response of many organisms (although there are a greatdeal of species-specific differences) That is ingestion rateincreases rapidly as prey abundance increases until a maximalingestion rate is attained (360)Manyof these curves have theform of a Type II functional response curve (361)
Based on the features of these curves it is clear that proto-zoa have the ability to ingest substantial numbers of prokar-yote and eukaryote prey in natural ecosystems Somebacterivorous flagellates can consume bacteria (and cyano-bacteria) at rates up to 100 s per individual per hour whereassome bacterivorous ciliates can consume thousands of bacte-ria per individual per hour Rates of consumption of eukary-otic phytoplankton by herbivorous ciliates and largeflagellates are considerably more variable and highly depend-ent of the feeding mechanism (eg filtration in ciliates versuspseudopodial net in dinoflagellates) and the size of the prey
These lab studies demonstrate that rarely are prey suffi-ciently abundant in nature to support the maximal ingestionrates of which many protozoa are capable Nevertheless pre-dation rates of protozoa in nature are sufficient to constitute amajor factor controlling the standing stocks of bacteria andphytoplankton (63 180 184 355) For example these sum-maries indicate that on average protozoa consume 20ndash80ofthe standing stock of phytoplankton (measured as chloro-phyll) in the world ocean and 60ndash75 of daily primary pro-duction a departure from the classical view of copepods as theprimary consumers of phytoplankton This large role of her-bivorous protozoa infers a large role for copepods as consumersof planktonic protozoa (185) Similarly the role of protozoaas major consumers of bacterial biomass is indicated by therelative balance between bacterial production on the onehand and bacterivory on the other (Fig 8) The approximateone-to-one balance between these measurements over a largenumber of studies implies that protozoan bacterivory must bea major factor determining the fate of bacteria in marine (andfreshwater) plankton ecosystems
It is interesting to note that some types of bacteria includ-ing marine Bdellovibrio are known to prey on other marine
bacteria (362) At the time of this writing the importanceof bacterial predation by other bacteria within the contextof oceanographic processes is not known and presumed tobe small While Bdellovibrio may be cultivated from seawaterit can also be found among sequences from cold sediments(363) and the water column Bdellovibrio happens to be thebest studied predatory bacterium but there are no doubtothers that we simply have not yet identified (364)
Species-Specific Interactions (MutualismParasitism Commensalisms)Interactions between marine microorganisms such as compe-tition allelopathy and symbiosis are fundamental aspectscontrolling community composition and activity but atpresent most are poorly characterized in natural marine sys-tems Bacteria must compete for dissolved and particulatecompounds with other bacteria andwith protistan taxa whilephagotrophic protists must compete with each other andmetazoa for available prey These interactions manifest them-selves in many forms including true competition for availableresources resource partitioning such that some taxa specializeon the utilization of certain substrates or prey while othersspecialize on different ones (reducing competition) or eventhe cooperative use of compounds (eg one bacteriumrsquoswaste products being utilized as substrate by another bacte-rium) These metabolic consortia are probably common inthe ocean but can be difficult to identify and characterizeConsortia of bacteria working together to perform complexreactions especially in anaerobic systems are well known inmicrobiology
Consortia in which there is a very high degree of integra-tion between species (ie symbiosis) are very common in theocean These associations may be beneficial to both partners(mutualism) beneficial to one part and inconsequential forthe other (commensalism) or beneficial to one part and det-rimental to the other (parasitism) Many highly visible andwell-characterized marine mutualisms exist between chemo-synthetic bacteria and animals such as hydrothermal ventor methane seep tubeworms clams mussels or other inverte-brates (365) and also between luminescent bacteria and fishor squids (366) Similarly many protozoa form mutualisticassociations with photosynthetic protists (367) The mostconspicuous of these associations involved large amoeboidprotozoa of oceanic ecosystems (planktonic foraminiferanspolycystine radiolaria and acantharia) which contain largenumbers of intracellular algae (74) but associations betweensome heterotrophic dinoflagellates and cyanobacteria are alsocommon (368) Studies have indicated that efficient nutrientcycling in these protozoan-algal associations confers an eco-logical advantage for both partners in surfacewaters of the oli-gotrophic ocean
Parasitism is also a common symbiotic relationship amongmarine protozoa Protozoan parasites have been reported for adiverse array of protists and animals in the ocean Heterotro-phic dinoflagellates and presently uncultured alveolate cladesrelated to alveolates appear to be particularly successful atevolving thismode of existence (369ndash371) In a common par-asitic group Amoebophrya spp the parasite invades photo-synthetic dinoflagellates where it undergoes reproductioneventually lysing the host and releasing dozens of infectiveswarmers The parasitic protist Pirsonia spp is a parasite ofdiatoms (372) The overall importance of these parasitismsin the population dynamics of their hosts is not clearly knownbut may constitute a significant source of mortality for somespecies (373 374) The frequency and diversity of ldquonewrdquo
FIGURE 8 Meta-analysis of experimental studies that have com-pared rates of bacterial productivity (converted to population growthrate units of dminus1) with rates of bacterivory by phagotrophic protists(expressed as equivalent units of bacterial growth rate dminus1) Thesolid line indicates where measurements of bacterial removal by graz-ers were equal tomeasurements of bacterial production Data redrawnfrom Sanders et al (63) doi1011289781555818821ch422f8
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-21
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
only within the past few decades (177) These species areoften abundant in waters where diatoms dominate (178179) In such situations heterotrophic dinoflagellate biomasscan be similar to that of ciliated protozoa (180)
Ciliates are generally the most recognizable form of pro-tozoa to nonspecialists Nonetheless ciliates are a diversegroup morphologically as well as ecologically (181 182)The degree of ciliature on these species can range from uni-formly ciliated to totally devoid of cilia during most of theirlife cycle to ciliature restricted to specific regions of thecell In addition cilia can fuse to form various complexstructures (eg ciliary membranes or veils cirri) that assistin locomotion food capture or attachmentCiliates are abun-dant and ecologically important species in both benthic (8)and pelagic (183) ecosystems and can consume a variety ofprokaryotic and eukaryotic prey Together with the heterotro-phic dinoflagellates these species are the dominant consum-ers of phytoplankton in many pelagic ecosystems (184) andas a consequence they form an important trophic link to met-azoan zooplankton (185 186)
Diversity and Biogeography of Protists FromMorphology to DNA SequencesUntil relatively recently the biodiversity of marine protistanassemblages was not generally considered a controversialtopic It was generally accepted that while all species of pro-tists certainly have not been identified representatives ofmost types of algae and protozoa had been observed anddescribed if not actually brought into culture in the labora-tory Direct sequencing of 18S rRNA genes from environ-mental samples (as described in ldquoMolecular Phylogeny andMetagenomics Field Applicationsrdquo) has changed that viewInitial forays into environmental DNA indicated a muchgreater diversity of protists than previously documented usingclassical approaches of culture and microscopy (81 187ndash191) Numerous publications over the past decades haveexpanded these findings (see [192] for a recent global analy-sis) which are highly analogous to discoveries in marine pro-karyote research as described already
Previously uncharacterized protistan diversity has nowbeen documented at virtually every level of eukaryoticorganismal classification Some of these findings couldhave been expected but some have been very unexpectedFor example a much greater diversity than noted previouslyhas been observed among small (lt10 microm) protists in plank-tonic ecosystems These species generally possess few distinc-tive morphological features and one could expect that manycryptic species might be present among these small morpho-types The molecular ldquodiscoveryrdquo of this eukaryotic diversityhas stimulated progress on the isolation and descriptionof new species and genera of minute algae and protozoa(193ndash197)
Analyses of environmental samples have also indicatedthe unanticipated existence of novel 18S rRNA gene sequen-ces that imply the presence of novel lineages of eukaryotes innatural protistan communities (198ndash200) These are sequen-ces for which there are apparently no known described orcultured species The degree to which these sequences dif-fer from sequences of known sequenced eukaryotes impliesthat some of these lineages may be distinct at the level ofphylum (201)
These findings have raised basic questions and somedebate concerning the true diversity and biogeography ofprotistan assemblages in natural ecosystems (202ndash204) thevalidity of the many novel phylotypes or cryptic species that
are being documented (205) the potential importance andsignificance of the many rare taxa that characterize thesecommunities (206) and the implications of these findingsfor the ecologicalbiogeochemical roles that protists playin aquatic ecosystems For example it was postulated andsubsequently confirmed that some novel alveolate lineagesreported from marine ecosystems make up a suite of parasiticprotozoa whose ecological importance may have been signifi-cantly underestimated in the past (207) Deciphering theidentity and significance of these many unknown phylotypeswill constitute a significant effort for protistologists in thefuture (80 85)
Life Histories and Ecological Strategies of ProtozoaAbundance prey type and life histories all vary tremendouslyamongmarine protozoa The smallest species (eg many flag-ellates) tend to be the most abundant and widely distributedin the world ocean Indeed many of these species may beglobally distributed (208) Most nanoplanktonic flagellateshave potentially rapid rates of grazing and growth Underoptimal conditions these species can divide by binary fissionseveral times a day and thus dramatic increases in their pop-ulations can take place in response to favorable conditionswithin a few days (209) Many of these species are capableof surviving for limited periods of time without food andhave developed a variety of physiological or life cycle strat-egies to cope with these events (209) However in contrastto some bacteria that may remain viable through long periodsof starvation (210) protozoa will expire or encyst in responseto low food abundance and thus ldquoboom-and-bustrdquo popula-tion changes are characteristic of small flagellates
On the other end of the size spectrum from the ratherubiquitous nanoflagellate species many of the largest proto-zoan species (polycystine and phaeodarian radiolaria plank-tonic foraminifera) are exclusively oceanic (ie do notsurvive in most coastal environments) andor have specificlatitudinal and depth distributions Thus the abundancesof these latter species may range from undetectable to max-imal abundances of gt105 individualsm3 Dramatic changesin the abundance of these protozoan taxa also can be relatedto changes in prey abundance physicalbehavioral aggrega-tion or to periodicity of life cycle events For example theplanktonic foraminifer Hastigerina pelagica reproduces on alunar cycle and thus abundances (and life stages) of this spe-cies in oceanic waters can vary considerably over the courseof a month (211) In general life cycles for the large amoe-boid protozoa are lengthy and complex (for single-celledorganisms) with life spans unknown for many species(attempts to culture them in the lab have so far been unsuc-cessful) but estimated to be on the order of months to per-haps years (175)
The abundance and activities of microplanktonic hetero-trophic protists (mostly ciliates and heterotrophic dinoflagel-lates) tend to be somewhat intermediate to those ofnanoplanktonic flagellates and the larger amoeboid formsThese species are present in the majority of marine ecosys-tems and collectively play an important role in the controlof phytoplankton biomass (and probably the abundance ofnanoplanktonic protozoa although there is little informa-tion on this topic) in waters throughout the world oceanLike small flagellates ciliates and dinoflagellates reproduceprimarily by binary fission but their maximal growth ratesare typically slower (one division a day is typical) and theirlife cycles often include sexual phases that allow for geneticrecombination
422-14 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
VIRUSESViral Abundance and General PropertiesViruses are simple biological agents typically 20ndash200 nm indiameter composed of a nucleic acid genome in a proteincoat that infect cells and ldquocommandeerrdquo the cellrsquos machineryto make more viruses which are released into the environ-ment when the host cell lyses or bursts A particular virus isthought to be capable of infecting only a narrow range of hosts(usually one species sometimes a genus rarely broader)Although some early studies had isolated viruses from thesea therewere no data prior to the 1980s showing such viruseswere very abundant and more significantly no evidence thatinfection was occurring in any important part of the planktoncommunity It wasnrsquot until the late 1980s that electronmicro-scopy with suitable concentration methods showed thatviruses are extremely abundant similar to or even higherthan bacterial abundance (212 213) Interestingly themost suitable transmission electron microscopy (TEM)approach used by Bergh et al (described in detail by [214])was actually similar to a direct centrifugation method devel-oped in 1949 (215)
TEM studies of viruses permit high-resolution images andobservation of viral morphology showing features such ashead diameter tails sheaths and tail fibers (Fig 6 collage
in upper left) Each type of virus has a fixed morphology(unlike bacteria that are potentially more plastic in theirappearance) and hence a coarse measure of viral diversityhas been possible by cataloging morphologies Studies thathave done so have found dozens or more different morpholo-gies of marine viruses in a given sample (217ndash219) A largeproportion resemble bacteriophages which are viruses thatinfect bacteria
The availability of brightly fluorescent nucleic acid stainsand high-porosity fine pore size (002 μm) filters made fromAl2O3 have made it easy to accurately count viruses by epi-fluorescence microscopy (38 220 221) Epifluorescence per-mits abundance estimates but does not allow observation ofviral morphology as viruses are below the resolution limitof light microscopy viruses are visible only as sources of light(like stars in the night sky see Fig 6 lower micrograph) Anextension of manual epifluorescence counts is the use of flowcytometry to count viruses now commonly employed in somelabs (222)
Virus abundance has been found to be closely related tobacterial abundance with a virusbacteria ratio typically10ndash301 A typical oceanic profile of bacterial and viral abun-dance is shown in (Fig 6 panel on right) This tight ratioand the strong correlation to bacterial abundance in relationto weaker correlations to chlorophyll have been cited as
FIGURE 6 Marine viruses (upper left collage) viewed by transmission electron microscopy These are cyanophages in the groups myoviridae(right and bottom) siphoviridae (left) and podoviridae (top) Scale bars represent 100 nm From Sullivan (216) Epifluorescence micrograph(lower left) of SYBR green I stained viruses (small fluorescing objects) and bacteria + archaea (large fluorescing objects) Depth distribution ofvirus and bacteria + archaea abundances obtained by epifluorescence microscopy of SYBRGreen stained cells in the central San Pedro BasinCalifornia (11 August 2000) doi1011289781555818821ch422f6
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-15
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
evidence that most marine viruses infect bacteria rather thaneukaryotic phytoplankton (217 218 223 224) though thereare also many important viruses of phytoplankton and otherorganisms with significant impacts on primary productivityblooms and ecosystem function (225ndash227)
The first demonstration of the activity of marine viruseswas presented by Proctor and Fuhrman (228) who showedby electron microscopic examination that viruses wereactually infecting marine bacteria and cyanobacteria at ameasurable rate Because only the final portion of the viruslife cycle is visible by electron microscopy (when the virusesare assembled and ready to lyse the host cell) only a very smallfraction of the infected cells can be counted as infected at anygiven time Proctor and Fuhrman (228) reported that only afew percent of the total bacteria from coastal waters and theSargasso Sea were visibly infected but they interpreted thedata with a model that implied the actual fraction of the totalcommunity that was infected was much higher They con-cluded that the total fraction of bacterial mortality attribut-able to viruses is roughly 10ndash40 Subsequent studies haveused refined versions of that same general approach andmodel (229 230) and numerous studies have used a varietyof alternative approaches to estimate viral activitymdashall yield-ing basically the same general conclusion (although themodel parameters need to be adjusted for cyanobacteriawhich on average seem to be infected less than bacteria)These approaches include (a) calculation of virus turnoverfrom decay estimates (231) (b) measurement of viral DNAsynthesis by incorporation of tritiated thymidine (232) (c)observation of effects of added viruses (233 234) (d) estima-tion of bacterial mortality in the absence of protists (e) use offluorescent viruses to estimate production by an approachanalogous to isotope dilution (38) and (f) dilution of virusesin filtered samples to estimate viral production (235) Theoverall consensus of these studies is that the initial estimateswere basically correct that is that viruses are responsiblefor about 10ndash40 of the bacterial mortality in most marinesystems (reviewed by [218 223 226 236 237]) Howeverthere is still some disagreement about the higher estimatesand it is likely that 40 mortality from viruses alone is nottypical for most marine systems
Most of the foregoing work has focused on the viral infec-tion of bacteria primarily thought to be heterotrophicalthough the original report by Proctor and Fuhrman (228)also noted the occurrence of cyanobacterial infection Sev-eral subsequent studies focused on phytoplankton includingthe potential effect of viruses on the termination of phyto-plankton blooms of Emiliania and Phaeocystis and the likelyimpact this might have on release of climate-active gasessuch as dimethyl sulfide (225 227 238ndash247) Althoughdetails are beyond the scope of this chapter viruses arethought to infect virtually all marine organisms with poten-tially significant impacts from zooplankton to whales (226)Detailed studies of viruses infecting cyanobacteria such asSynechococcus have shown differences in viral host specific-ity particularly toward coastal and oceanic host strainsand some occasional high virus abundances (to 105ml) asmeasured by most probable number (MPN) cultivation tech-niques in the Gulf of Mexico near Texas (248 249) Othervirus cultivation studies with Prochlorococcus and Synecho-coccus in oligotrophic waters of the Sargasso Sea showedan interesting pattern of cross-infection between these gen-era by some virus types (suggesting gene flow among theseorganisms) but generally low MPN estimates of abundanceto 103ml even when cyanobacterial abundance was near105ml (216 250)
Overall the consensus emerging from direct comparisonsof viral-mediated mortality and grazer-mediated (ie micro-zooplankton) mortality have indicated that viral lysis of bac-teria constitutes a significant fraction of total mortality ofthis assemblage while the mortality of phytoplanktonappears to be dominated bymicrozooplankton inmost instan-ces (251 252)
As mentioned viruses have significant morphologicaldiversity as observed in TEM studies It is also possible toinvestigate aspects of their genetic diversity In early workon this topic Wommack et al (253) and Steward et al(254) observed the diversity of viral genome lengths in a fieldsample by pulsed field gel electrophoresis with viral genomesranging 25 kb to gt300 kb in length Field results show thatthe viral community composition is dynamic in space andtime with clear changes in the banding patterns over seasonsand locations in Chesapeake Bay (253) between oceanbasins and subsequent to dinoflagellate blooms (254) andwith depths to 500 m at one location (255)
Although viruses do not all share a set of core genesthat allows a universal viral phylogeny (comparable to SSUrRNA gene in cellular organisms) genetic diversity amonga single group of viruses can be examined by sequence analysisof shared genes within the group An example is the g20 geneinT4-like cyanophages that has shown extremely high diver-sity even among very closely related viruses and has demon-strated geographic and seasonal variation (256) A secondexample is g23 which is found broadly in diverse and wide-spread T4-like phages (257) and can have seasonally repeat-ing patterns (258) as well as short-term rapid dynamics thatcorrelate to those in bacteria (259)
Viral MetagenomicsAs with cellular organisms metagenomics can be used toexamine viral diversity and genetics without themany restric-tions of cultivation Viruses can be collected by selective fil-tration and concentrated by tangential flow filtration orflocculated with iron chloride (260 261) then their collec-tive metagenome can be extracted linker amplified and ana-lyzed by sequencing (262 263) Viral metagenomic studiesare particularly challenging because the large majority ofsequences have no annotated matches in any databases butso far results from marine samples around the world haveshown extremely high diversity and variations with depthlocation and time presumably with highly dispersed typesselected by local conditions (262ndash266) Because metage-nomes are best interpreted when there are representative cul-tures available the best matches of viral metagenomesoriginally tended to be to the few viral isolates infecting trulycommonmarine bacteria like cyanophage infecting Synecho-coccus and Prochlorococcus (267) However the developmentof dilution-to-extinction cultures representing common het-erotrophic marine taxa like Peligibacter (a member of theSAR11 clade) and SAR116 has allowed isolation from sea-water of viruses infecting these common organisms and theseisolates have indeed been found to be highly abundant inmarine viral metagenomes (268 269) New approaches tointerpret the results include clustering the proteins independ-ent of known proteins to compare samples to each other andtry to find environmental factors driving viral communitychanges (270) Such analysis of a large global data set (TaraOceans expedition) has shown that extensive sampling hascome close to reaching the total diversity in tropical and tem-perate waters of such viral protein clusters (which essentiallyrepresent various viral protein motifs not all viral proteintypes) and that viruses appear to be directionally dispersed
422-16 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquodownstreamrdquo in major ocean currents as one may expectfrom first principles but also supporting the ldquoseed bankrdquohypothesis of viral biogeography (271)
Viruses and Host DiversityViruses are themselves thought to be instrumental in drivingincreased diversity of their microbial hosts via a hypothesizedprocess often called ldquokill the winnerrdquo This is because viralinfection is host-specific and density-dependent the latterbecause viruses diffuse from host to host so an abundanthost is more likely to pass on infection than a rare oneThis means that if an organism becomes abundant andblooms winning the competition for resources it becomesmore susceptible to a viral epidemic This would benefit therarer organisms and thus help foster diversity (223 272273) However bloom scenarios as described above involvesystems far from steady state and the formal theory of Thing-stad and Lignell (274) has interesting steady-state solutionswhere several viruses infect several hosts stably over time(via trade-offs between growth rates and viral susceptibility)which may occur at the strain or species level (274) There issome experimental evidence that viruses have effects on nat-ural marine microbial community composition weaker thanthe kill the winner (bloom version) hypothesis would sug-gest (275 276) so something resembling the steady-statecoexistence described in the model may in fact be commonThere also appear to be processes that foster coexistencebetween viruses and hosts but the mechanisms are largelyspeculative (223 277 278)
Viruses may also be directly involved in host genetic diver-sity because they can be the agents of genetic exchangebetween microorganisms (223 273) This often involvesthe viral lifestyle known as lysogeny whereby viruses survivewithin host cells as DNA only integrated into the host chro-mosome and being reproduced each time the host divides Ahost harboring such a genome is called a lysogen becauseunder conditions of stress to the host cell a genetic switchmay cause the viral genome to initiate the lytic process pro-ducing many progeny viruses and bursting from the hostLysogeny is a very common property occurring in a signifi-cant part of the bacterial community (279 280) althoughthe incidence of induction of the lytic phase in nature is appa-rently low (281) Overall lysogeny is poorly understood butthought to have both positive and negative impacts on themicrobial community (282) Recent results suggest that ben-efits of being lysogenic in highly seasonal polar seas leads tofundamental difference between polar and other marine viralcommunities (283)
Viruses and the Microbial LoopAs part of the food web viruses occupy a unique positionThey infect host cells that are mostly thought to be heterotro-phic bacteria and by doing so they typically burst the hosts torelease progeny viruses and cellular debris But what is the fateof this material Viruses themselves do not last indefinitelyand a simple steady-state assumption implies that from eachburst of viruses (typically 20ndash100 per lytic event) only onesuccessfully infects another cell The rest are inactivatedand broken down by sunlight (UV and visible exposure)and enzymatic attack (284) or consumed byminute phagotro-phic protists (285) thus reentering the food web as substratefor bacteria or food for protistan consumers Experiments incontrolled laboratory systems and field studies with radioac-tively labeled viral lysis products have supported the conclu-sion that most of the organic matter released by the viralinfection is either taken up by bacteria or respired (286 287)
Modeling this process as part of the microbial loop showsthat viral lysis represent a sort of side loop that has the neteffect of remineralizing a significant amount of the carbonand nutrients that enter the bacteria-protist part of themicro-bial loop (Fig 7) A theoretical numerical steady-state modelcomparing a systemwith no viral activity to onewhere virusesare responsible for 50 of bacterial mortality showed that thesystem with viruses had 33 more bacterial production andrespiration than the virus-free system implying that theviruses had the effect of permitting the bacteria to processmore of the primary production than they would otherwise(223) Although 50 is a high number unlikely to be com-mon in the sea this model nevertheless illustrates that virusescan reduce the amount of energy reaching higher trophic lev-els The implication is that viruses lead to increased bacterialactivity at the expense of the larger organisms
MAJOR ENVIRONMENTAL CONTROLSLight Temperature and PressureTemperature has an important potential influence on bio-chemical reactions and therefore on biological processesin general Most ocean waters fall in the range of ndash2degC to30degC with obvious exceptions in hydrothermally heatedareas Temperature has long been known to be a regulatingfactor for the growth of heterotrophic microbes
In temperate waters it has been established that microbialactivity is generally much higher in warm summer waters thanin winter (55) The relationship is not simple howeverbecause multiple factors act at the same time Some contro-versy still exists regarding the highest and lowest extremesfor marine bacterial growth although there is broad agree-ment that bacteria grow gt100degC at hydrothermal vents andltminus5degC in sea ice brines Pomeroy et al (57) noted the inter-esting observation that bacteria seem particularly inhibitednear the freezing point of seawater (ca ndash22degC) comparedto eukaryotic phytoplankton This effect results in polarspring phytoplankton blooms that accumulate organic car-bon in advance of the response of the bacterial communityand development of the microbial loop and perhaps lead toenhanced benthic-pelagic coupling (288)
The relationship between temperature and the growth rateof marine phytoplankton was described broadly in a now
FIGURE 7 Modification of the microbial loop concept that incor-porates the functional role of viruses Export can be via predation orsinking From Fuhrman (223)doi1011289781555818821ch422f7
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-17
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
classic paper by Eppley (58) and later Goldman and Carpen-ter (289) Temperature was shown to exert a strong and directeffect on themaximal growth rates of these species withmax-imal intrinsic growth rates at 0degC generally lt1 divisiondaywhile growth rates at 30degC may be gt4day These relation-ships indicate the maximal rate that might be attained byphytoplankton at these temperatures but they do not takeother factors into account (nutrients light) While warmtemperature ostensibly allows more rapid growth it createshydrographic conditions that typically give rise to nutrientlimitation of algal growth Thus some of the coldest watersin nature witness some of the most massive phytoplanktonblooms (290) while warm oceanic gyres represent some ofthe most oligotrophic areas of the ocean The multiple andoften conflicting effects of temperature on the growth of nat-ural phytoplankton assemblages limit the accuracy of presentpredictions regarding how primary producers will respond toclimate change (291)
An analysis of the effect of temperature on the growth ofheterotrophic protists has indicated that the response is qual-itatively similar to that of phytoplankton but with a twistThe growth rates of at least some protozoa can exceed thoseof phytoplankton at warmer environmental temperaturesbut the opposite effect is apparent at very low environmentaltemperature This differential effect of temperature on thegrowth of phototrophic and heterotrophic protists was basedon a large meta-analysis of published protistan growth (59)That analysis demonstrated that the maximal growth ratesattained by phototrophic protists could exceed the maximalgrowth rates attained by heterotrophic protists (all otherpotential growth-limiting factors not considered) Thereforeprotozoan growth rates may be constrained to a greater degreeat low environmental temperature than rates for phytoplank-ton If so then phytoplankton bloomsmay get a head start ongrazers during spring in polar ecosystems This scenario is con-sistent with information on seasonal biomass changes andmicrozooplankton herbivory in the Ross Sea Antarctica(292 293) but there are still too few data to fully vet thishypothesis
The importance of high pressure on bacterial growthgained considerable attention in the late 1960s when thedeep sea submersible Alvin was accidentally lost overboardwith its hatch openWhile no lives were lost in this accidentsome workmenrsquos lunches sank to the bottom (sim1500 m)inside the submersible Alvin was recovered after 10 monthsand interestingly there was a waterlogged lunch containingapples bologna sandwiches and broken vacuum bottleswith broth that all appeared hardly degraded and tasted palat-able Yet when placed in a refrigerator on the ship these itemsdegraded relatively quickly Initially pressure was thought tobe the preserving factor as the sea floor temperature was sim-ilar to the refrigerator temperature and the only major differ-ence would be pressure (294) Following this observationa series of experiments to measure degradation of variousorganic materials left in the deep sea for extended periodsindicated that degradationwas typically significantly reducedimplying that pressure reduces the degradation rates (295)Nonetheless changes in the protozoan community of naturaldetrital material sinking to the deep ocean floor indicate thatthe microbial community can respond relatively quickly insome situations (296) Deep sea microbiology has advancedconsiderably yet it is still difficult to interpret results withrespect to actual in situ rates of naturally occurring organicmatter Deep sea bacteria adapted to high pressures havebeen isolated that are barophilic (also called piezophilic)meaning that they prefer high pressures and have reduced
activity at lower pressures whereas others are barotolerant(piezotolerant) tolerating but not preferring high pressuresfor example see (297 298)
Few data are available on barotolerantbarophilic marineprotozoa Protozoa certainly exist and grow at great oceanicdepths but measurements of in situ growth rates for these spe-cies do not yet exist Measurable protozoan numbers havebeen documented in the deep-sea sediments for more than30 years (299 300) and viable protozoa have occasionallybeen cultured from these environments (301ndash305) butvery few direct measurements of the activities of these speciesin situ have been reported (306) A few protozoa have beenisolated that will grow at high pressure (302 303 307) andprotists that appear to be unique to the deep ocean havebeen observed either directly or through the analysis ofDNA sequences (308ndash311) but possibly the best direct evi-dence that protozoan activity takes place at the high pressurescharacteristics of the deep sea are experimental and observa-tional work noting the stimulatory effect that detrital deposi-tion has on some components of the protozoan community(296 312) These observations indicate a diverse and activeprotozoan fauna of the deep ocean although their biogeo-chemical significance is largely uncharacterized at this time
Dissolved and Particulate Organic MatterBacteria and archaea are thought to be by far the most impor-tant organisms with respect to the processing of dissolvedorganic matter (DOM) and nonliving particulate organicmatter (POM also called detritus) in the ocean While theremay be some uptake of DOM by protists particularly forgrowth factors needed in trace amounts such as vitamins(313) the bulk of this material is probably utilized by bacteriaand archaea (314) including the smallest cyanobacteriumProchlorococcus (315 316) which thus may be considered amixotroph Due to their small size bacteria have extremelyhigh surfacevolume ratios and combined with their over-whelmingly high abundance an extremely high integratedsurface area Protozoa tend to obtain the organic materialsthat they require for growth from their prey rather thanthrough the uptake of DOM Overall protozoa tend to besources of dissolved and detrital organic substances throughthe excretion of unassimilated prey biomass in expelledfood vacuoles
Particulate organic matter is not directly available as sub-strate to bacteria These substances must first be reduced tosmall molecules that can be transported into the cell Thisis accomplished by the production of extracellular enzymes(note that few if any large polymers are directly taken up bybacteria with the possible exception of DNA) Hydrolyticenzymes produced by bacteria (and almost certainly archaea)break down polymers like proteins polysaccharides andnucleic acids As with DOM POM is composed of a complexmixture of compounds that vary in their susceptibility to bac-terial degradation and utilization
Particulate material in the water column serves not only asbacterial substrate but also as substratum POM occurs in thewater column across a huge size spectrum from micrometersup to some detrital aggregates more than 1 m in diameter(317) Much of this particulate material is in a constant stateof flux with colloidal material constantly coalescing andaggregating to form new or larger particles (318) as microbialdegradation acts simultaneously to remineralize this materialDetrital particles that attain macroscopic size either by directformation (317 319 320) or via accretion and aggregation(321) and are often called marine snow or macroaggregatesMarine snow particles are readily colonized by bacteria and
422-18 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
many other microorganisms and their abundances on theseaggregates can be orders of magnitude greater than abundan-ces in an equivalent volume of the water surrounding theaggregates (320 322ndash324) and their diversity and highlyconcentrated abundances on detrital aggregates implies thatthere may be stronger trophic coupling and efficient energyelemental cycling relative to these processes among free-living microorganisms in the surrounding waters This specu-lation has resulted in a number of formalizations of the poten-tial impact of aggregate microbiology on biogeochemicalprocesses in the ocean (325 326) Most definitions of bio-films include aggregated particulate and these particles areoften studied within the biofilm conceptual model (see chap-ter by Lawrence et al in this section)
Inorganic Nutrients
Macronutrients (N P)Virtually all heterotrophic organisms in the ocean contributeto the pool of available macronutrients via the excretion ofmetabolic wastes and protozoa contribute significantly tothis process Macronutrients are often in excess within proto-zoan prey relative to the consumersrsquo needs because much ofthe prey carbon is respired to produce energy for metabolismand growth ldquoExcessrdquo nutrients are eliminated as eitherorganic compounds or more commonly as ammonium (forN) and phosphate (for P) The importance of the match(or mismatch) between predator and prey stoichiometry hasbeen demonstrated experimentally and repeatedly in marineand freshwater ecosystems and has been elevated to a funda-mental ecological principle governing elemental and energyflow in aquatic food webs (327) Based on this reasoningCaron et al (328) concluded that bacterivorous protozoaplay an important role in nutrient remineralization andrelease in the ocean Bacterial biomass is typically rich in Nand P relative to other organisms so bacterial predators expe-rience large excesses of these elements relative to their growthneedsMoreover bacterivorous protozoa should be dispropor-tionately important in nutrient remineralization amongsingle-celled eukaryotes because as the smallest protozoa inaquatic ecosystems they have high weight-specific metabolicrates (329) Thus both stoichiometric and allometric rela-tionships implicate small bacterivorous protozoa as impor-tant sources of remineralized nutrients in the plankton
The reasoning also holds true for bacterial utilization oforganic compounds and it has led to some interestingfindings and conclusions regarding the role of bacteria innutrient cycling in the ocean Bacteria are important sourcesremineralized nutrients when they are consuming N- orP-rich substrate Contrary to this traditional view howeverheterotrophic bacteria are often strong competitors for inor-ganic nutrients under many situations largely because bacte-rial biomass is richer in N and P than much of the organicmatter they consume in nature so they require additional Nand P to produce their biomass (summarized by [330]) Exper-imental studies have demonstrated the uptake of ammoniumby bacteria other than cyanobacteria using stable isotope 15N(331) and also the short-lived radioisotope 13N (332) oftenfinding that bacteria may be responsible for one third of the Nor P uptake Additionally there is evidence that bacteria insome oceans like the Sargasso Sea are limited by phosphorus(333 334) Growth limitation of bacteria by phosphorusappears to be a common phenomenon of freshwater ecosys-tems (335 336)
N2 fixation is restricted to prokaryotes and in the oceanwater column it was thought for many years to be done
primarily byTrichodesmium a warm-water colonial cyanobac-terium that blooms sporadically andRichelia a cyanobacterialsymbiont that lives within certain diatoms (337) Some uni-cellular cyanobacteria also contribute significantly to globalN2 fixation (338) and molecular biological data suggestthat a variety of other bacteriamdashnot phytoplanktonmdashmayalso be fixing nitrogen in seawater (339) An exciting recentdiscovery in this field is that a bacterial nitrogen fixer origi-nally known only from nifH sequences UCYN-A has hadits genome sequenced by parallel pyrosequencing and wasfound be a cyanobacterium (Candidatus Atelocyanobacteriumthalassa) lacking the apparatus to generate oxygen as well asother pathways (340) Further study showed it to be symbioticwith a marine alga (341)
Micronutrients (Trace Metals Growth Factors)Bacterial growth may often be limited by macronutrients asdescribed and sometimes by trace nutrients particularlyiron that occur at very low concentrations in seawater(342 343) The importance of Fe as an element limiting pri-mary production in some oceanic regions has come underclose scrutiny and extensive experimental investigation inrecent years Many bacteria have special uptake mechanismsto utilize extremely low concentrations of iron includingsiderophores (released compounds that bind iron and arethen specifically taken up) Even if bacteria are capable ofgrowth at low Fe concentrations growth may be less efficientKirchman et al (344) found that Fe limitation stronglyreduced bacterial growth efficiency in a cultured marine bac-terium (ca 50 efficiency Fe replete but lt10 when Fedeplete) Thus the effects may be complex Marine N2 fixa-tion also may be limited by the availability of Fe becausenitrogen fixers have high Fe requirements
Protozoa tend to be a source rather than a sink for Fe in theocean analogous to their role in macronutrient remineraliza-tion Protozoan grazing activity has been shown to releaseiron from bacterial biomass thereby relieving Fe limitationin co-occurring phytoplankton (345) a mechanistic demon-stration of the process using protozoa growing on Fe-repletebacteria However it has also been reported that Fe-limitedbacteria may contain insufficient Fe for the bacterivorous pro-tozoa that consume them thus leading to Fe limitation in theprotozoan as well (346)
OxygenHypoxic and anoxic regions of the ocean are expected to sig-nificantly expand in the coming decades as a consequence ofchanges in ocean stratification resulting from global climatechange (347) As oxygen becomes depleted bacteria havethe capability to utilize a series of alternate electron acceptorsCompositional changes in the bacterial assemblage accom-pany these changes in metabolic activity but are beyondthe scope of this chapter Many of the studies on benthicmicrobiology have focused on redox reactions that occurthere Oxygen is usually consumed by sediment microbialactivity much faster than the rate at which it can be replacedby diffusion so most sediments especially organically richones are anaerobic below a few mm (fine-grained muds) orcm (coarser sediments) from the sedimentwater interfaceexcept where animals ventilate the benthos through tubesWhen oxygen is absent bacteria and archaea use alternateelectron acceptors such as nitrate nitrite oxidized Fe orMn organic matter or sulfate The use of nitrate or nitriteas an electron acceptor with concomitant production of N2
gas (denitrification) results in net loss of biologically availablenitrogen Bacteria that perform this reaction are typically
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-19
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquofacultativerdquo organisms that can switch from oxygen to nitrateor nitrite Organic matter may be reduced by fermentationreactions in which part of an organic substrate is oxidizedwhile another is reduced fermenter organisms are also typi-cally facultative Sulfate is a particularly important alternateelectron acceptor in marine systems because it occurs as thesecond most abundant anion in seawater (behind chloride)typically 28 mM Its use as an electron acceptor leads to theproduction of elemental sulfur or sulfide The use of sulfateas an electron acceptor yields much less energy than oxygenor nitrate and sulfate-reducing bacteria are strict anaerobesunable to use oxygen (often strongly inhibited by oxygen)Methane can be produced by methanogenic archaea whichare all strict anaerobes using CO2 as an electron acceptor(see Findlay and Battin this section for further discussion)
Reduced compounds such as sulfide are produced by theuse of these alternate electron acceptors and they diffuse awayfrom their source When they diffuse to environments with astrong oxidant like oxygen they may be oxidized by chemo-trophic bacteria in energy-yielding reactions Similarlyammonium can be oxidized to nitrite and then nitrate by che-motrophic nitrifying bacteria and the nitrate can diffuse intoanaerobic layers and become denitrified to N2 Such coupledoxidation-reduction reactions can lead to accelerated biogeo-chemical cycling particularly in sediments
It was originally believed that relatively few free-livingspecies of protozoa inhabit anaerobic and hypoxic marineenvironments This view has changed based on (largely)molecular analyses of some of these environments (308348ndash351) A significant number of previously undescribedeukaryotic phylotypes have been observed in anoxic environ-ments High protozoan biodiversity in the absence of molec-ular oxygen is not completely unexpected because otheranaerobic environments such as some gut environments(eg cockroach guts and cattle rumen) have notoriouslyabundant and diverse protozoan fauna but marine anaerobicenvironments have been difficult to access and sample prop-erly Many anaerobic protozoa exist together with symbioticbacteria (352 353) or possess unique organelles such as thehydrogenosome (354)
MAJOR BIOTIC CONTROLSThe Trophic Activities of Protozoa Predation andTop-Down ControlsPredator-prey relationships are fundamental interactionswithin biological communities linking production of bio-mass on the one hand with removal on the other hand Estab-lishing the trophic relationships among microorganisms andquantifying the rates at which they take place constitutemajor endeavors within marine microbial ecology As notedpreviously protozoa consume a wide variety of prokaryoticand protistan prey Numerous laboratory and field measure-ments have been made throughout the past quarter centuryto attempt to quantify the rates of bacterial and phytoplank-ton consumption The methodology for accomplishing thesestudies has varied and a number of approaches are presentlyemployed
Methodologies for estimating microbial grazing are diversebut generally fall into one of two experimental approachesperturbation experiments or tracer experiments Perturbationexperiments (size fractionation metabolic inhibitors anddilution technique) rely on manipulation of the consumeror prey assemblages in somemanner such that the effect is evi-dent from changes in the abundance of the prey population
during incubations relative to control treatments that receiveno manipulation Tracer experiments (radioactively labeledprey fluorescently labeled prey stable isotopendashlabeled pray)attempt to use labeled prey or a prey proxy to quantitativelyfollow the movement of prey biomass into consumers Forsummaries of these methods see (74 355)
Perturbation experiments have entailed a variety of mech-anisms to ldquodecouplerdquo predators from prey in natural microbialcommunities For example size fractionation uses various sizefilters to remove an entire size class of grazers from awater sam-ple leaving the prey to grow in the relative absence of thosepredators during subsequent incubations Unfiltered samples(controls) indicate the growth of prey in the presence of pre-dation Predation impact is obtained by the difference ingrowth of the prey between these treatments This methodhas been used to examine predation on cyanobacteria heter-otrophic bacteria and minute phototrophic eukaryotesAlternatively metabolic inhibitors have been employed tohalt predation (or in some cases prokaryote growth) insteadof physical removal by filtration
The dilution technique (356) has become a commonmethod for examining grazing mortality of phytoplanktonby microzooplankton (which is comprised predominantly ofprotozoa) This method relies on the dilution of herbivorouszooplankton in a series of subsamples (and concomitantmeasurements of phytoplankton growth in each subsample)to estimate the grazing mortality of the herbivores
Tracer experiments have used both radioactively labeledprey and fluorescently labeled prey to examine predation onbacteria and phytoplankton Radioactive labeling has beenaccomplished using 14C- or 3H-labeled organic compounds(for bacteria) or 14C-bicarbonate (for phytoplankton) Theuse of radioisotope approaches has diminished with improve-ments in the number and types of fluorescent compoundsavailable for this work Fluorescently labeled prey havebeen labeled with a variety of compounds that allow theprey to be observed readily (even inside predators) by epi-fluorescencemicroscopy (357 358) Alternatively a decreasein the total number of fluorescent prey in a sample indicatesthe rate of removal of the prey assemblage
Both perturbation experiments and tracer experimentsdescribed have inherent advantages and disadvantages Forexample perturbation experiments are fairly straightforwardto conduct but they may introduce artifact by removingpotentially important sources of nutrients or organic com-pounds released by predators (359) On the other hand thedilution technique is very labor-intensive but one advantageof this method is that it provides the opportunity to determineboth phytoplankton growth and grazing mortality in a singleprocedure Most tracer experiments are fairly easy to conductand generally do not perturb the food web being studied butfeeding selectivity for or against the tracer (relative to naturalprey) may produce spurious results In addition some of thesemethods provide a measurement of the activity of the entiregrazer assemblage and others are applicable for examiningthe activity of specific taxa For example size fractionationapproaches examine the effect of the removal of an entiresize class of organisms (a community-levelrdquo measurement)whereas fluorescently labeled prey can be used to examinethe ingestion rate of a particular species by observing theuptake of labeled prey into that species (a species-levelmeasurement)
Predation rates of numerous marine protozoa have beenexamined in the laboratory From these experiments a gen-eral knowledge of the rates of consumption by these specieshas been accumulated Feeding in most protozoa shows a
422-20 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
functional response to the availability of prey that is similar tothe response of many organisms (although there are a greatdeal of species-specific differences) That is ingestion rateincreases rapidly as prey abundance increases until a maximalingestion rate is attained (360)Manyof these curves have theform of a Type II functional response curve (361)
Based on the features of these curves it is clear that proto-zoa have the ability to ingest substantial numbers of prokar-yote and eukaryote prey in natural ecosystems Somebacterivorous flagellates can consume bacteria (and cyano-bacteria) at rates up to 100 s per individual per hour whereassome bacterivorous ciliates can consume thousands of bacte-ria per individual per hour Rates of consumption of eukary-otic phytoplankton by herbivorous ciliates and largeflagellates are considerably more variable and highly depend-ent of the feeding mechanism (eg filtration in ciliates versuspseudopodial net in dinoflagellates) and the size of the prey
These lab studies demonstrate that rarely are prey suffi-ciently abundant in nature to support the maximal ingestionrates of which many protozoa are capable Nevertheless pre-dation rates of protozoa in nature are sufficient to constitute amajor factor controlling the standing stocks of bacteria andphytoplankton (63 180 184 355) For example these sum-maries indicate that on average protozoa consume 20ndash80ofthe standing stock of phytoplankton (measured as chloro-phyll) in the world ocean and 60ndash75 of daily primary pro-duction a departure from the classical view of copepods as theprimary consumers of phytoplankton This large role of her-bivorous protozoa infers a large role for copepods as consumersof planktonic protozoa (185) Similarly the role of protozoaas major consumers of bacterial biomass is indicated by therelative balance between bacterial production on the onehand and bacterivory on the other (Fig 8) The approximateone-to-one balance between these measurements over a largenumber of studies implies that protozoan bacterivory must bea major factor determining the fate of bacteria in marine (andfreshwater) plankton ecosystems
It is interesting to note that some types of bacteria includ-ing marine Bdellovibrio are known to prey on other marine
bacteria (362) At the time of this writing the importanceof bacterial predation by other bacteria within the contextof oceanographic processes is not known and presumed tobe small While Bdellovibrio may be cultivated from seawaterit can also be found among sequences from cold sediments(363) and the water column Bdellovibrio happens to be thebest studied predatory bacterium but there are no doubtothers that we simply have not yet identified (364)
Species-Specific Interactions (MutualismParasitism Commensalisms)Interactions between marine microorganisms such as compe-tition allelopathy and symbiosis are fundamental aspectscontrolling community composition and activity but atpresent most are poorly characterized in natural marine sys-tems Bacteria must compete for dissolved and particulatecompounds with other bacteria andwith protistan taxa whilephagotrophic protists must compete with each other andmetazoa for available prey These interactions manifest them-selves in many forms including true competition for availableresources resource partitioning such that some taxa specializeon the utilization of certain substrates or prey while othersspecialize on different ones (reducing competition) or eventhe cooperative use of compounds (eg one bacteriumrsquoswaste products being utilized as substrate by another bacte-rium) These metabolic consortia are probably common inthe ocean but can be difficult to identify and characterizeConsortia of bacteria working together to perform complexreactions especially in anaerobic systems are well known inmicrobiology
Consortia in which there is a very high degree of integra-tion between species (ie symbiosis) are very common in theocean These associations may be beneficial to both partners(mutualism) beneficial to one part and inconsequential forthe other (commensalism) or beneficial to one part and det-rimental to the other (parasitism) Many highly visible andwell-characterized marine mutualisms exist between chemo-synthetic bacteria and animals such as hydrothermal ventor methane seep tubeworms clams mussels or other inverte-brates (365) and also between luminescent bacteria and fishor squids (366) Similarly many protozoa form mutualisticassociations with photosynthetic protists (367) The mostconspicuous of these associations involved large amoeboidprotozoa of oceanic ecosystems (planktonic foraminiferanspolycystine radiolaria and acantharia) which contain largenumbers of intracellular algae (74) but associations betweensome heterotrophic dinoflagellates and cyanobacteria are alsocommon (368) Studies have indicated that efficient nutrientcycling in these protozoan-algal associations confers an eco-logical advantage for both partners in surfacewaters of the oli-gotrophic ocean
Parasitism is also a common symbiotic relationship amongmarine protozoa Protozoan parasites have been reported for adiverse array of protists and animals in the ocean Heterotro-phic dinoflagellates and presently uncultured alveolate cladesrelated to alveolates appear to be particularly successful atevolving thismode of existence (369ndash371) In a common par-asitic group Amoebophrya spp the parasite invades photo-synthetic dinoflagellates where it undergoes reproductioneventually lysing the host and releasing dozens of infectiveswarmers The parasitic protist Pirsonia spp is a parasite ofdiatoms (372) The overall importance of these parasitismsin the population dynamics of their hosts is not clearly knownbut may constitute a significant source of mortality for somespecies (373 374) The frequency and diversity of ldquonewrdquo
FIGURE 8 Meta-analysis of experimental studies that have com-pared rates of bacterial productivity (converted to population growthrate units of dminus1) with rates of bacterivory by phagotrophic protists(expressed as equivalent units of bacterial growth rate dminus1) Thesolid line indicates where measurements of bacterial removal by graz-ers were equal tomeasurements of bacterial production Data redrawnfrom Sanders et al (63) doi1011289781555818821ch422f8
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-21
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
VIRUSESViral Abundance and General PropertiesViruses are simple biological agents typically 20ndash200 nm indiameter composed of a nucleic acid genome in a proteincoat that infect cells and ldquocommandeerrdquo the cellrsquos machineryto make more viruses which are released into the environ-ment when the host cell lyses or bursts A particular virus isthought to be capable of infecting only a narrow range of hosts(usually one species sometimes a genus rarely broader)Although some early studies had isolated viruses from thesea therewere no data prior to the 1980s showing such viruseswere very abundant and more significantly no evidence thatinfection was occurring in any important part of the planktoncommunity It wasnrsquot until the late 1980s that electronmicro-scopy with suitable concentration methods showed thatviruses are extremely abundant similar to or even higherthan bacterial abundance (212 213) Interestingly themost suitable transmission electron microscopy (TEM)approach used by Bergh et al (described in detail by [214])was actually similar to a direct centrifugation method devel-oped in 1949 (215)
TEM studies of viruses permit high-resolution images andobservation of viral morphology showing features such ashead diameter tails sheaths and tail fibers (Fig 6 collage
in upper left) Each type of virus has a fixed morphology(unlike bacteria that are potentially more plastic in theirappearance) and hence a coarse measure of viral diversityhas been possible by cataloging morphologies Studies thathave done so have found dozens or more different morpholo-gies of marine viruses in a given sample (217ndash219) A largeproportion resemble bacteriophages which are viruses thatinfect bacteria
The availability of brightly fluorescent nucleic acid stainsand high-porosity fine pore size (002 μm) filters made fromAl2O3 have made it easy to accurately count viruses by epi-fluorescence microscopy (38 220 221) Epifluorescence per-mits abundance estimates but does not allow observation ofviral morphology as viruses are below the resolution limitof light microscopy viruses are visible only as sources of light(like stars in the night sky see Fig 6 lower micrograph) Anextension of manual epifluorescence counts is the use of flowcytometry to count viruses now commonly employed in somelabs (222)
Virus abundance has been found to be closely related tobacterial abundance with a virusbacteria ratio typically10ndash301 A typical oceanic profile of bacterial and viral abun-dance is shown in (Fig 6 panel on right) This tight ratioand the strong correlation to bacterial abundance in relationto weaker correlations to chlorophyll have been cited as
FIGURE 6 Marine viruses (upper left collage) viewed by transmission electron microscopy These are cyanophages in the groups myoviridae(right and bottom) siphoviridae (left) and podoviridae (top) Scale bars represent 100 nm From Sullivan (216) Epifluorescence micrograph(lower left) of SYBR green I stained viruses (small fluorescing objects) and bacteria + archaea (large fluorescing objects) Depth distribution ofvirus and bacteria + archaea abundances obtained by epifluorescence microscopy of SYBRGreen stained cells in the central San Pedro BasinCalifornia (11 August 2000) doi1011289781555818821ch422f6
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-15
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
evidence that most marine viruses infect bacteria rather thaneukaryotic phytoplankton (217 218 223 224) though thereare also many important viruses of phytoplankton and otherorganisms with significant impacts on primary productivityblooms and ecosystem function (225ndash227)
The first demonstration of the activity of marine viruseswas presented by Proctor and Fuhrman (228) who showedby electron microscopic examination that viruses wereactually infecting marine bacteria and cyanobacteria at ameasurable rate Because only the final portion of the viruslife cycle is visible by electron microscopy (when the virusesare assembled and ready to lyse the host cell) only a very smallfraction of the infected cells can be counted as infected at anygiven time Proctor and Fuhrman (228) reported that only afew percent of the total bacteria from coastal waters and theSargasso Sea were visibly infected but they interpreted thedata with a model that implied the actual fraction of the totalcommunity that was infected was much higher They con-cluded that the total fraction of bacterial mortality attribut-able to viruses is roughly 10ndash40 Subsequent studies haveused refined versions of that same general approach andmodel (229 230) and numerous studies have used a varietyof alternative approaches to estimate viral activitymdashall yield-ing basically the same general conclusion (although themodel parameters need to be adjusted for cyanobacteriawhich on average seem to be infected less than bacteria)These approaches include (a) calculation of virus turnoverfrom decay estimates (231) (b) measurement of viral DNAsynthesis by incorporation of tritiated thymidine (232) (c)observation of effects of added viruses (233 234) (d) estima-tion of bacterial mortality in the absence of protists (e) use offluorescent viruses to estimate production by an approachanalogous to isotope dilution (38) and (f) dilution of virusesin filtered samples to estimate viral production (235) Theoverall consensus of these studies is that the initial estimateswere basically correct that is that viruses are responsiblefor about 10ndash40 of the bacterial mortality in most marinesystems (reviewed by [218 223 226 236 237]) Howeverthere is still some disagreement about the higher estimatesand it is likely that 40 mortality from viruses alone is nottypical for most marine systems
Most of the foregoing work has focused on the viral infec-tion of bacteria primarily thought to be heterotrophicalthough the original report by Proctor and Fuhrman (228)also noted the occurrence of cyanobacterial infection Sev-eral subsequent studies focused on phytoplankton includingthe potential effect of viruses on the termination of phyto-plankton blooms of Emiliania and Phaeocystis and the likelyimpact this might have on release of climate-active gasessuch as dimethyl sulfide (225 227 238ndash247) Althoughdetails are beyond the scope of this chapter viruses arethought to infect virtually all marine organisms with poten-tially significant impacts from zooplankton to whales (226)Detailed studies of viruses infecting cyanobacteria such asSynechococcus have shown differences in viral host specific-ity particularly toward coastal and oceanic host strainsand some occasional high virus abundances (to 105ml) asmeasured by most probable number (MPN) cultivation tech-niques in the Gulf of Mexico near Texas (248 249) Othervirus cultivation studies with Prochlorococcus and Synecho-coccus in oligotrophic waters of the Sargasso Sea showedan interesting pattern of cross-infection between these gen-era by some virus types (suggesting gene flow among theseorganisms) but generally low MPN estimates of abundanceto 103ml even when cyanobacterial abundance was near105ml (216 250)
Overall the consensus emerging from direct comparisonsof viral-mediated mortality and grazer-mediated (ie micro-zooplankton) mortality have indicated that viral lysis of bac-teria constitutes a significant fraction of total mortality ofthis assemblage while the mortality of phytoplanktonappears to be dominated bymicrozooplankton inmost instan-ces (251 252)
As mentioned viruses have significant morphologicaldiversity as observed in TEM studies It is also possible toinvestigate aspects of their genetic diversity In early workon this topic Wommack et al (253) and Steward et al(254) observed the diversity of viral genome lengths in a fieldsample by pulsed field gel electrophoresis with viral genomesranging 25 kb to gt300 kb in length Field results show thatthe viral community composition is dynamic in space andtime with clear changes in the banding patterns over seasonsand locations in Chesapeake Bay (253) between oceanbasins and subsequent to dinoflagellate blooms (254) andwith depths to 500 m at one location (255)
Although viruses do not all share a set of core genesthat allows a universal viral phylogeny (comparable to SSUrRNA gene in cellular organisms) genetic diversity amonga single group of viruses can be examined by sequence analysisof shared genes within the group An example is the g20 geneinT4-like cyanophages that has shown extremely high diver-sity even among very closely related viruses and has demon-strated geographic and seasonal variation (256) A secondexample is g23 which is found broadly in diverse and wide-spread T4-like phages (257) and can have seasonally repeat-ing patterns (258) as well as short-term rapid dynamics thatcorrelate to those in bacteria (259)
Viral MetagenomicsAs with cellular organisms metagenomics can be used toexamine viral diversity and genetics without themany restric-tions of cultivation Viruses can be collected by selective fil-tration and concentrated by tangential flow filtration orflocculated with iron chloride (260 261) then their collec-tive metagenome can be extracted linker amplified and ana-lyzed by sequencing (262 263) Viral metagenomic studiesare particularly challenging because the large majority ofsequences have no annotated matches in any databases butso far results from marine samples around the world haveshown extremely high diversity and variations with depthlocation and time presumably with highly dispersed typesselected by local conditions (262ndash266) Because metage-nomes are best interpreted when there are representative cul-tures available the best matches of viral metagenomesoriginally tended to be to the few viral isolates infecting trulycommonmarine bacteria like cyanophage infecting Synecho-coccus and Prochlorococcus (267) However the developmentof dilution-to-extinction cultures representing common het-erotrophic marine taxa like Peligibacter (a member of theSAR11 clade) and SAR116 has allowed isolation from sea-water of viruses infecting these common organisms and theseisolates have indeed been found to be highly abundant inmarine viral metagenomes (268 269) New approaches tointerpret the results include clustering the proteins independ-ent of known proteins to compare samples to each other andtry to find environmental factors driving viral communitychanges (270) Such analysis of a large global data set (TaraOceans expedition) has shown that extensive sampling hascome close to reaching the total diversity in tropical and tem-perate waters of such viral protein clusters (which essentiallyrepresent various viral protein motifs not all viral proteintypes) and that viruses appear to be directionally dispersed
422-16 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquodownstreamrdquo in major ocean currents as one may expectfrom first principles but also supporting the ldquoseed bankrdquohypothesis of viral biogeography (271)
Viruses and Host DiversityViruses are themselves thought to be instrumental in drivingincreased diversity of their microbial hosts via a hypothesizedprocess often called ldquokill the winnerrdquo This is because viralinfection is host-specific and density-dependent the latterbecause viruses diffuse from host to host so an abundanthost is more likely to pass on infection than a rare oneThis means that if an organism becomes abundant andblooms winning the competition for resources it becomesmore susceptible to a viral epidemic This would benefit therarer organisms and thus help foster diversity (223 272273) However bloom scenarios as described above involvesystems far from steady state and the formal theory of Thing-stad and Lignell (274) has interesting steady-state solutionswhere several viruses infect several hosts stably over time(via trade-offs between growth rates and viral susceptibility)which may occur at the strain or species level (274) There issome experimental evidence that viruses have effects on nat-ural marine microbial community composition weaker thanthe kill the winner (bloom version) hypothesis would sug-gest (275 276) so something resembling the steady-statecoexistence described in the model may in fact be commonThere also appear to be processes that foster coexistencebetween viruses and hosts but the mechanisms are largelyspeculative (223 277 278)
Viruses may also be directly involved in host genetic diver-sity because they can be the agents of genetic exchangebetween microorganisms (223 273) This often involvesthe viral lifestyle known as lysogeny whereby viruses survivewithin host cells as DNA only integrated into the host chro-mosome and being reproduced each time the host divides Ahost harboring such a genome is called a lysogen becauseunder conditions of stress to the host cell a genetic switchmay cause the viral genome to initiate the lytic process pro-ducing many progeny viruses and bursting from the hostLysogeny is a very common property occurring in a signifi-cant part of the bacterial community (279 280) althoughthe incidence of induction of the lytic phase in nature is appa-rently low (281) Overall lysogeny is poorly understood butthought to have both positive and negative impacts on themicrobial community (282) Recent results suggest that ben-efits of being lysogenic in highly seasonal polar seas leads tofundamental difference between polar and other marine viralcommunities (283)
Viruses and the Microbial LoopAs part of the food web viruses occupy a unique positionThey infect host cells that are mostly thought to be heterotro-phic bacteria and by doing so they typically burst the hosts torelease progeny viruses and cellular debris But what is the fateof this material Viruses themselves do not last indefinitelyand a simple steady-state assumption implies that from eachburst of viruses (typically 20ndash100 per lytic event) only onesuccessfully infects another cell The rest are inactivatedand broken down by sunlight (UV and visible exposure)and enzymatic attack (284) or consumed byminute phagotro-phic protists (285) thus reentering the food web as substratefor bacteria or food for protistan consumers Experiments incontrolled laboratory systems and field studies with radioac-tively labeled viral lysis products have supported the conclu-sion that most of the organic matter released by the viralinfection is either taken up by bacteria or respired (286 287)
Modeling this process as part of the microbial loop showsthat viral lysis represent a sort of side loop that has the neteffect of remineralizing a significant amount of the carbonand nutrients that enter the bacteria-protist part of themicro-bial loop (Fig 7) A theoretical numerical steady-state modelcomparing a systemwith no viral activity to onewhere virusesare responsible for 50 of bacterial mortality showed that thesystem with viruses had 33 more bacterial production andrespiration than the virus-free system implying that theviruses had the effect of permitting the bacteria to processmore of the primary production than they would otherwise(223) Although 50 is a high number unlikely to be com-mon in the sea this model nevertheless illustrates that virusescan reduce the amount of energy reaching higher trophic lev-els The implication is that viruses lead to increased bacterialactivity at the expense of the larger organisms
MAJOR ENVIRONMENTAL CONTROLSLight Temperature and PressureTemperature has an important potential influence on bio-chemical reactions and therefore on biological processesin general Most ocean waters fall in the range of ndash2degC to30degC with obvious exceptions in hydrothermally heatedareas Temperature has long been known to be a regulatingfactor for the growth of heterotrophic microbes
In temperate waters it has been established that microbialactivity is generally much higher in warm summer waters thanin winter (55) The relationship is not simple howeverbecause multiple factors act at the same time Some contro-versy still exists regarding the highest and lowest extremesfor marine bacterial growth although there is broad agree-ment that bacteria grow gt100degC at hydrothermal vents andltminus5degC in sea ice brines Pomeroy et al (57) noted the inter-esting observation that bacteria seem particularly inhibitednear the freezing point of seawater (ca ndash22degC) comparedto eukaryotic phytoplankton This effect results in polarspring phytoplankton blooms that accumulate organic car-bon in advance of the response of the bacterial communityand development of the microbial loop and perhaps lead toenhanced benthic-pelagic coupling (288)
The relationship between temperature and the growth rateof marine phytoplankton was described broadly in a now
FIGURE 7 Modification of the microbial loop concept that incor-porates the functional role of viruses Export can be via predation orsinking From Fuhrman (223)doi1011289781555818821ch422f7
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-17
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
classic paper by Eppley (58) and later Goldman and Carpen-ter (289) Temperature was shown to exert a strong and directeffect on themaximal growth rates of these species withmax-imal intrinsic growth rates at 0degC generally lt1 divisiondaywhile growth rates at 30degC may be gt4day These relation-ships indicate the maximal rate that might be attained byphytoplankton at these temperatures but they do not takeother factors into account (nutrients light) While warmtemperature ostensibly allows more rapid growth it createshydrographic conditions that typically give rise to nutrientlimitation of algal growth Thus some of the coldest watersin nature witness some of the most massive phytoplanktonblooms (290) while warm oceanic gyres represent some ofthe most oligotrophic areas of the ocean The multiple andoften conflicting effects of temperature on the growth of nat-ural phytoplankton assemblages limit the accuracy of presentpredictions regarding how primary producers will respond toclimate change (291)
An analysis of the effect of temperature on the growth ofheterotrophic protists has indicated that the response is qual-itatively similar to that of phytoplankton but with a twistThe growth rates of at least some protozoa can exceed thoseof phytoplankton at warmer environmental temperaturesbut the opposite effect is apparent at very low environmentaltemperature This differential effect of temperature on thegrowth of phototrophic and heterotrophic protists was basedon a large meta-analysis of published protistan growth (59)That analysis demonstrated that the maximal growth ratesattained by phototrophic protists could exceed the maximalgrowth rates attained by heterotrophic protists (all otherpotential growth-limiting factors not considered) Thereforeprotozoan growth rates may be constrained to a greater degreeat low environmental temperature than rates for phytoplank-ton If so then phytoplankton bloomsmay get a head start ongrazers during spring in polar ecosystems This scenario is con-sistent with information on seasonal biomass changes andmicrozooplankton herbivory in the Ross Sea Antarctica(292 293) but there are still too few data to fully vet thishypothesis
The importance of high pressure on bacterial growthgained considerable attention in the late 1960s when thedeep sea submersible Alvin was accidentally lost overboardwith its hatch openWhile no lives were lost in this accidentsome workmenrsquos lunches sank to the bottom (sim1500 m)inside the submersible Alvin was recovered after 10 monthsand interestingly there was a waterlogged lunch containingapples bologna sandwiches and broken vacuum bottleswith broth that all appeared hardly degraded and tasted palat-able Yet when placed in a refrigerator on the ship these itemsdegraded relatively quickly Initially pressure was thought tobe the preserving factor as the sea floor temperature was sim-ilar to the refrigerator temperature and the only major differ-ence would be pressure (294) Following this observationa series of experiments to measure degradation of variousorganic materials left in the deep sea for extended periodsindicated that degradationwas typically significantly reducedimplying that pressure reduces the degradation rates (295)Nonetheless changes in the protozoan community of naturaldetrital material sinking to the deep ocean floor indicate thatthe microbial community can respond relatively quickly insome situations (296) Deep sea microbiology has advancedconsiderably yet it is still difficult to interpret results withrespect to actual in situ rates of naturally occurring organicmatter Deep sea bacteria adapted to high pressures havebeen isolated that are barophilic (also called piezophilic)meaning that they prefer high pressures and have reduced
activity at lower pressures whereas others are barotolerant(piezotolerant) tolerating but not preferring high pressuresfor example see (297 298)
Few data are available on barotolerantbarophilic marineprotozoa Protozoa certainly exist and grow at great oceanicdepths but measurements of in situ growth rates for these spe-cies do not yet exist Measurable protozoan numbers havebeen documented in the deep-sea sediments for more than30 years (299 300) and viable protozoa have occasionallybeen cultured from these environments (301ndash305) butvery few direct measurements of the activities of these speciesin situ have been reported (306) A few protozoa have beenisolated that will grow at high pressure (302 303 307) andprotists that appear to be unique to the deep ocean havebeen observed either directly or through the analysis ofDNA sequences (308ndash311) but possibly the best direct evi-dence that protozoan activity takes place at the high pressurescharacteristics of the deep sea are experimental and observa-tional work noting the stimulatory effect that detrital deposi-tion has on some components of the protozoan community(296 312) These observations indicate a diverse and activeprotozoan fauna of the deep ocean although their biogeo-chemical significance is largely uncharacterized at this time
Dissolved and Particulate Organic MatterBacteria and archaea are thought to be by far the most impor-tant organisms with respect to the processing of dissolvedorganic matter (DOM) and nonliving particulate organicmatter (POM also called detritus) in the ocean While theremay be some uptake of DOM by protists particularly forgrowth factors needed in trace amounts such as vitamins(313) the bulk of this material is probably utilized by bacteriaand archaea (314) including the smallest cyanobacteriumProchlorococcus (315 316) which thus may be considered amixotroph Due to their small size bacteria have extremelyhigh surfacevolume ratios and combined with their over-whelmingly high abundance an extremely high integratedsurface area Protozoa tend to obtain the organic materialsthat they require for growth from their prey rather thanthrough the uptake of DOM Overall protozoa tend to besources of dissolved and detrital organic substances throughthe excretion of unassimilated prey biomass in expelledfood vacuoles
Particulate organic matter is not directly available as sub-strate to bacteria These substances must first be reduced tosmall molecules that can be transported into the cell Thisis accomplished by the production of extracellular enzymes(note that few if any large polymers are directly taken up bybacteria with the possible exception of DNA) Hydrolyticenzymes produced by bacteria (and almost certainly archaea)break down polymers like proteins polysaccharides andnucleic acids As with DOM POM is composed of a complexmixture of compounds that vary in their susceptibility to bac-terial degradation and utilization
Particulate material in the water column serves not only asbacterial substrate but also as substratum POM occurs in thewater column across a huge size spectrum from micrometersup to some detrital aggregates more than 1 m in diameter(317) Much of this particulate material is in a constant stateof flux with colloidal material constantly coalescing andaggregating to form new or larger particles (318) as microbialdegradation acts simultaneously to remineralize this materialDetrital particles that attain macroscopic size either by directformation (317 319 320) or via accretion and aggregation(321) and are often called marine snow or macroaggregatesMarine snow particles are readily colonized by bacteria and
422-18 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
many other microorganisms and their abundances on theseaggregates can be orders of magnitude greater than abundan-ces in an equivalent volume of the water surrounding theaggregates (320 322ndash324) and their diversity and highlyconcentrated abundances on detrital aggregates implies thatthere may be stronger trophic coupling and efficient energyelemental cycling relative to these processes among free-living microorganisms in the surrounding waters This specu-lation has resulted in a number of formalizations of the poten-tial impact of aggregate microbiology on biogeochemicalprocesses in the ocean (325 326) Most definitions of bio-films include aggregated particulate and these particles areoften studied within the biofilm conceptual model (see chap-ter by Lawrence et al in this section)
Inorganic Nutrients
Macronutrients (N P)Virtually all heterotrophic organisms in the ocean contributeto the pool of available macronutrients via the excretion ofmetabolic wastes and protozoa contribute significantly tothis process Macronutrients are often in excess within proto-zoan prey relative to the consumersrsquo needs because much ofthe prey carbon is respired to produce energy for metabolismand growth ldquoExcessrdquo nutrients are eliminated as eitherorganic compounds or more commonly as ammonium (forN) and phosphate (for P) The importance of the match(or mismatch) between predator and prey stoichiometry hasbeen demonstrated experimentally and repeatedly in marineand freshwater ecosystems and has been elevated to a funda-mental ecological principle governing elemental and energyflow in aquatic food webs (327) Based on this reasoningCaron et al (328) concluded that bacterivorous protozoaplay an important role in nutrient remineralization andrelease in the ocean Bacterial biomass is typically rich in Nand P relative to other organisms so bacterial predators expe-rience large excesses of these elements relative to their growthneedsMoreover bacterivorous protozoa should be dispropor-tionately important in nutrient remineralization amongsingle-celled eukaryotes because as the smallest protozoa inaquatic ecosystems they have high weight-specific metabolicrates (329) Thus both stoichiometric and allometric rela-tionships implicate small bacterivorous protozoa as impor-tant sources of remineralized nutrients in the plankton
The reasoning also holds true for bacterial utilization oforganic compounds and it has led to some interestingfindings and conclusions regarding the role of bacteria innutrient cycling in the ocean Bacteria are important sourcesremineralized nutrients when they are consuming N- orP-rich substrate Contrary to this traditional view howeverheterotrophic bacteria are often strong competitors for inor-ganic nutrients under many situations largely because bacte-rial biomass is richer in N and P than much of the organicmatter they consume in nature so they require additional Nand P to produce their biomass (summarized by [330]) Exper-imental studies have demonstrated the uptake of ammoniumby bacteria other than cyanobacteria using stable isotope 15N(331) and also the short-lived radioisotope 13N (332) oftenfinding that bacteria may be responsible for one third of the Nor P uptake Additionally there is evidence that bacteria insome oceans like the Sargasso Sea are limited by phosphorus(333 334) Growth limitation of bacteria by phosphorusappears to be a common phenomenon of freshwater ecosys-tems (335 336)
N2 fixation is restricted to prokaryotes and in the oceanwater column it was thought for many years to be done
primarily byTrichodesmium a warm-water colonial cyanobac-terium that blooms sporadically andRichelia a cyanobacterialsymbiont that lives within certain diatoms (337) Some uni-cellular cyanobacteria also contribute significantly to globalN2 fixation (338) and molecular biological data suggestthat a variety of other bacteriamdashnot phytoplanktonmdashmayalso be fixing nitrogen in seawater (339) An exciting recentdiscovery in this field is that a bacterial nitrogen fixer origi-nally known only from nifH sequences UCYN-A has hadits genome sequenced by parallel pyrosequencing and wasfound be a cyanobacterium (Candidatus Atelocyanobacteriumthalassa) lacking the apparatus to generate oxygen as well asother pathways (340) Further study showed it to be symbioticwith a marine alga (341)
Micronutrients (Trace Metals Growth Factors)Bacterial growth may often be limited by macronutrients asdescribed and sometimes by trace nutrients particularlyiron that occur at very low concentrations in seawater(342 343) The importance of Fe as an element limiting pri-mary production in some oceanic regions has come underclose scrutiny and extensive experimental investigation inrecent years Many bacteria have special uptake mechanismsto utilize extremely low concentrations of iron includingsiderophores (released compounds that bind iron and arethen specifically taken up) Even if bacteria are capable ofgrowth at low Fe concentrations growth may be less efficientKirchman et al (344) found that Fe limitation stronglyreduced bacterial growth efficiency in a cultured marine bac-terium (ca 50 efficiency Fe replete but lt10 when Fedeplete) Thus the effects may be complex Marine N2 fixa-tion also may be limited by the availability of Fe becausenitrogen fixers have high Fe requirements
Protozoa tend to be a source rather than a sink for Fe in theocean analogous to their role in macronutrient remineraliza-tion Protozoan grazing activity has been shown to releaseiron from bacterial biomass thereby relieving Fe limitationin co-occurring phytoplankton (345) a mechanistic demon-stration of the process using protozoa growing on Fe-repletebacteria However it has also been reported that Fe-limitedbacteria may contain insufficient Fe for the bacterivorous pro-tozoa that consume them thus leading to Fe limitation in theprotozoan as well (346)
OxygenHypoxic and anoxic regions of the ocean are expected to sig-nificantly expand in the coming decades as a consequence ofchanges in ocean stratification resulting from global climatechange (347) As oxygen becomes depleted bacteria havethe capability to utilize a series of alternate electron acceptorsCompositional changes in the bacterial assemblage accom-pany these changes in metabolic activity but are beyondthe scope of this chapter Many of the studies on benthicmicrobiology have focused on redox reactions that occurthere Oxygen is usually consumed by sediment microbialactivity much faster than the rate at which it can be replacedby diffusion so most sediments especially organically richones are anaerobic below a few mm (fine-grained muds) orcm (coarser sediments) from the sedimentwater interfaceexcept where animals ventilate the benthos through tubesWhen oxygen is absent bacteria and archaea use alternateelectron acceptors such as nitrate nitrite oxidized Fe orMn organic matter or sulfate The use of nitrate or nitriteas an electron acceptor with concomitant production of N2
gas (denitrification) results in net loss of biologically availablenitrogen Bacteria that perform this reaction are typically
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-19
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquofacultativerdquo organisms that can switch from oxygen to nitrateor nitrite Organic matter may be reduced by fermentationreactions in which part of an organic substrate is oxidizedwhile another is reduced fermenter organisms are also typi-cally facultative Sulfate is a particularly important alternateelectron acceptor in marine systems because it occurs as thesecond most abundant anion in seawater (behind chloride)typically 28 mM Its use as an electron acceptor leads to theproduction of elemental sulfur or sulfide The use of sulfateas an electron acceptor yields much less energy than oxygenor nitrate and sulfate-reducing bacteria are strict anaerobesunable to use oxygen (often strongly inhibited by oxygen)Methane can be produced by methanogenic archaea whichare all strict anaerobes using CO2 as an electron acceptor(see Findlay and Battin this section for further discussion)
Reduced compounds such as sulfide are produced by theuse of these alternate electron acceptors and they diffuse awayfrom their source When they diffuse to environments with astrong oxidant like oxygen they may be oxidized by chemo-trophic bacteria in energy-yielding reactions Similarlyammonium can be oxidized to nitrite and then nitrate by che-motrophic nitrifying bacteria and the nitrate can diffuse intoanaerobic layers and become denitrified to N2 Such coupledoxidation-reduction reactions can lead to accelerated biogeo-chemical cycling particularly in sediments
It was originally believed that relatively few free-livingspecies of protozoa inhabit anaerobic and hypoxic marineenvironments This view has changed based on (largely)molecular analyses of some of these environments (308348ndash351) A significant number of previously undescribedeukaryotic phylotypes have been observed in anoxic environ-ments High protozoan biodiversity in the absence of molec-ular oxygen is not completely unexpected because otheranaerobic environments such as some gut environments(eg cockroach guts and cattle rumen) have notoriouslyabundant and diverse protozoan fauna but marine anaerobicenvironments have been difficult to access and sample prop-erly Many anaerobic protozoa exist together with symbioticbacteria (352 353) or possess unique organelles such as thehydrogenosome (354)
MAJOR BIOTIC CONTROLSThe Trophic Activities of Protozoa Predation andTop-Down ControlsPredator-prey relationships are fundamental interactionswithin biological communities linking production of bio-mass on the one hand with removal on the other hand Estab-lishing the trophic relationships among microorganisms andquantifying the rates at which they take place constitutemajor endeavors within marine microbial ecology As notedpreviously protozoa consume a wide variety of prokaryoticand protistan prey Numerous laboratory and field measure-ments have been made throughout the past quarter centuryto attempt to quantify the rates of bacterial and phytoplank-ton consumption The methodology for accomplishing thesestudies has varied and a number of approaches are presentlyemployed
Methodologies for estimating microbial grazing are diversebut generally fall into one of two experimental approachesperturbation experiments or tracer experiments Perturbationexperiments (size fractionation metabolic inhibitors anddilution technique) rely on manipulation of the consumeror prey assemblages in somemanner such that the effect is evi-dent from changes in the abundance of the prey population
during incubations relative to control treatments that receiveno manipulation Tracer experiments (radioactively labeledprey fluorescently labeled prey stable isotopendashlabeled pray)attempt to use labeled prey or a prey proxy to quantitativelyfollow the movement of prey biomass into consumers Forsummaries of these methods see (74 355)
Perturbation experiments have entailed a variety of mech-anisms to ldquodecouplerdquo predators from prey in natural microbialcommunities For example size fractionation uses various sizefilters to remove an entire size class of grazers from awater sam-ple leaving the prey to grow in the relative absence of thosepredators during subsequent incubations Unfiltered samples(controls) indicate the growth of prey in the presence of pre-dation Predation impact is obtained by the difference ingrowth of the prey between these treatments This methodhas been used to examine predation on cyanobacteria heter-otrophic bacteria and minute phototrophic eukaryotesAlternatively metabolic inhibitors have been employed tohalt predation (or in some cases prokaryote growth) insteadof physical removal by filtration
The dilution technique (356) has become a commonmethod for examining grazing mortality of phytoplanktonby microzooplankton (which is comprised predominantly ofprotozoa) This method relies on the dilution of herbivorouszooplankton in a series of subsamples (and concomitantmeasurements of phytoplankton growth in each subsample)to estimate the grazing mortality of the herbivores
Tracer experiments have used both radioactively labeledprey and fluorescently labeled prey to examine predation onbacteria and phytoplankton Radioactive labeling has beenaccomplished using 14C- or 3H-labeled organic compounds(for bacteria) or 14C-bicarbonate (for phytoplankton) Theuse of radioisotope approaches has diminished with improve-ments in the number and types of fluorescent compoundsavailable for this work Fluorescently labeled prey havebeen labeled with a variety of compounds that allow theprey to be observed readily (even inside predators) by epi-fluorescencemicroscopy (357 358) Alternatively a decreasein the total number of fluorescent prey in a sample indicatesthe rate of removal of the prey assemblage
Both perturbation experiments and tracer experimentsdescribed have inherent advantages and disadvantages Forexample perturbation experiments are fairly straightforwardto conduct but they may introduce artifact by removingpotentially important sources of nutrients or organic com-pounds released by predators (359) On the other hand thedilution technique is very labor-intensive but one advantageof this method is that it provides the opportunity to determineboth phytoplankton growth and grazing mortality in a singleprocedure Most tracer experiments are fairly easy to conductand generally do not perturb the food web being studied butfeeding selectivity for or against the tracer (relative to naturalprey) may produce spurious results In addition some of thesemethods provide a measurement of the activity of the entiregrazer assemblage and others are applicable for examiningthe activity of specific taxa For example size fractionationapproaches examine the effect of the removal of an entiresize class of organisms (a community-levelrdquo measurement)whereas fluorescently labeled prey can be used to examinethe ingestion rate of a particular species by observing theuptake of labeled prey into that species (a species-levelmeasurement)
Predation rates of numerous marine protozoa have beenexamined in the laboratory From these experiments a gen-eral knowledge of the rates of consumption by these specieshas been accumulated Feeding in most protozoa shows a
422-20 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
functional response to the availability of prey that is similar tothe response of many organisms (although there are a greatdeal of species-specific differences) That is ingestion rateincreases rapidly as prey abundance increases until a maximalingestion rate is attained (360)Manyof these curves have theform of a Type II functional response curve (361)
Based on the features of these curves it is clear that proto-zoa have the ability to ingest substantial numbers of prokar-yote and eukaryote prey in natural ecosystems Somebacterivorous flagellates can consume bacteria (and cyano-bacteria) at rates up to 100 s per individual per hour whereassome bacterivorous ciliates can consume thousands of bacte-ria per individual per hour Rates of consumption of eukary-otic phytoplankton by herbivorous ciliates and largeflagellates are considerably more variable and highly depend-ent of the feeding mechanism (eg filtration in ciliates versuspseudopodial net in dinoflagellates) and the size of the prey
These lab studies demonstrate that rarely are prey suffi-ciently abundant in nature to support the maximal ingestionrates of which many protozoa are capable Nevertheless pre-dation rates of protozoa in nature are sufficient to constitute amajor factor controlling the standing stocks of bacteria andphytoplankton (63 180 184 355) For example these sum-maries indicate that on average protozoa consume 20ndash80ofthe standing stock of phytoplankton (measured as chloro-phyll) in the world ocean and 60ndash75 of daily primary pro-duction a departure from the classical view of copepods as theprimary consumers of phytoplankton This large role of her-bivorous protozoa infers a large role for copepods as consumersof planktonic protozoa (185) Similarly the role of protozoaas major consumers of bacterial biomass is indicated by therelative balance between bacterial production on the onehand and bacterivory on the other (Fig 8) The approximateone-to-one balance between these measurements over a largenumber of studies implies that protozoan bacterivory must bea major factor determining the fate of bacteria in marine (andfreshwater) plankton ecosystems
It is interesting to note that some types of bacteria includ-ing marine Bdellovibrio are known to prey on other marine
bacteria (362) At the time of this writing the importanceof bacterial predation by other bacteria within the contextof oceanographic processes is not known and presumed tobe small While Bdellovibrio may be cultivated from seawaterit can also be found among sequences from cold sediments(363) and the water column Bdellovibrio happens to be thebest studied predatory bacterium but there are no doubtothers that we simply have not yet identified (364)
Species-Specific Interactions (MutualismParasitism Commensalisms)Interactions between marine microorganisms such as compe-tition allelopathy and symbiosis are fundamental aspectscontrolling community composition and activity but atpresent most are poorly characterized in natural marine sys-tems Bacteria must compete for dissolved and particulatecompounds with other bacteria andwith protistan taxa whilephagotrophic protists must compete with each other andmetazoa for available prey These interactions manifest them-selves in many forms including true competition for availableresources resource partitioning such that some taxa specializeon the utilization of certain substrates or prey while othersspecialize on different ones (reducing competition) or eventhe cooperative use of compounds (eg one bacteriumrsquoswaste products being utilized as substrate by another bacte-rium) These metabolic consortia are probably common inthe ocean but can be difficult to identify and characterizeConsortia of bacteria working together to perform complexreactions especially in anaerobic systems are well known inmicrobiology
Consortia in which there is a very high degree of integra-tion between species (ie symbiosis) are very common in theocean These associations may be beneficial to both partners(mutualism) beneficial to one part and inconsequential forthe other (commensalism) or beneficial to one part and det-rimental to the other (parasitism) Many highly visible andwell-characterized marine mutualisms exist between chemo-synthetic bacteria and animals such as hydrothermal ventor methane seep tubeworms clams mussels or other inverte-brates (365) and also between luminescent bacteria and fishor squids (366) Similarly many protozoa form mutualisticassociations with photosynthetic protists (367) The mostconspicuous of these associations involved large amoeboidprotozoa of oceanic ecosystems (planktonic foraminiferanspolycystine radiolaria and acantharia) which contain largenumbers of intracellular algae (74) but associations betweensome heterotrophic dinoflagellates and cyanobacteria are alsocommon (368) Studies have indicated that efficient nutrientcycling in these protozoan-algal associations confers an eco-logical advantage for both partners in surfacewaters of the oli-gotrophic ocean
Parasitism is also a common symbiotic relationship amongmarine protozoa Protozoan parasites have been reported for adiverse array of protists and animals in the ocean Heterotro-phic dinoflagellates and presently uncultured alveolate cladesrelated to alveolates appear to be particularly successful atevolving thismode of existence (369ndash371) In a common par-asitic group Amoebophrya spp the parasite invades photo-synthetic dinoflagellates where it undergoes reproductioneventually lysing the host and releasing dozens of infectiveswarmers The parasitic protist Pirsonia spp is a parasite ofdiatoms (372) The overall importance of these parasitismsin the population dynamics of their hosts is not clearly knownbut may constitute a significant source of mortality for somespecies (373 374) The frequency and diversity of ldquonewrdquo
FIGURE 8 Meta-analysis of experimental studies that have com-pared rates of bacterial productivity (converted to population growthrate units of dminus1) with rates of bacterivory by phagotrophic protists(expressed as equivalent units of bacterial growth rate dminus1) Thesolid line indicates where measurements of bacterial removal by graz-ers were equal tomeasurements of bacterial production Data redrawnfrom Sanders et al (63) doi1011289781555818821ch422f8
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-21
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
evidence that most marine viruses infect bacteria rather thaneukaryotic phytoplankton (217 218 223 224) though thereare also many important viruses of phytoplankton and otherorganisms with significant impacts on primary productivityblooms and ecosystem function (225ndash227)
The first demonstration of the activity of marine viruseswas presented by Proctor and Fuhrman (228) who showedby electron microscopic examination that viruses wereactually infecting marine bacteria and cyanobacteria at ameasurable rate Because only the final portion of the viruslife cycle is visible by electron microscopy (when the virusesare assembled and ready to lyse the host cell) only a very smallfraction of the infected cells can be counted as infected at anygiven time Proctor and Fuhrman (228) reported that only afew percent of the total bacteria from coastal waters and theSargasso Sea were visibly infected but they interpreted thedata with a model that implied the actual fraction of the totalcommunity that was infected was much higher They con-cluded that the total fraction of bacterial mortality attribut-able to viruses is roughly 10ndash40 Subsequent studies haveused refined versions of that same general approach andmodel (229 230) and numerous studies have used a varietyof alternative approaches to estimate viral activitymdashall yield-ing basically the same general conclusion (although themodel parameters need to be adjusted for cyanobacteriawhich on average seem to be infected less than bacteria)These approaches include (a) calculation of virus turnoverfrom decay estimates (231) (b) measurement of viral DNAsynthesis by incorporation of tritiated thymidine (232) (c)observation of effects of added viruses (233 234) (d) estima-tion of bacterial mortality in the absence of protists (e) use offluorescent viruses to estimate production by an approachanalogous to isotope dilution (38) and (f) dilution of virusesin filtered samples to estimate viral production (235) Theoverall consensus of these studies is that the initial estimateswere basically correct that is that viruses are responsiblefor about 10ndash40 of the bacterial mortality in most marinesystems (reviewed by [218 223 226 236 237]) Howeverthere is still some disagreement about the higher estimatesand it is likely that 40 mortality from viruses alone is nottypical for most marine systems
Most of the foregoing work has focused on the viral infec-tion of bacteria primarily thought to be heterotrophicalthough the original report by Proctor and Fuhrman (228)also noted the occurrence of cyanobacterial infection Sev-eral subsequent studies focused on phytoplankton includingthe potential effect of viruses on the termination of phyto-plankton blooms of Emiliania and Phaeocystis and the likelyimpact this might have on release of climate-active gasessuch as dimethyl sulfide (225 227 238ndash247) Althoughdetails are beyond the scope of this chapter viruses arethought to infect virtually all marine organisms with poten-tially significant impacts from zooplankton to whales (226)Detailed studies of viruses infecting cyanobacteria such asSynechococcus have shown differences in viral host specific-ity particularly toward coastal and oceanic host strainsand some occasional high virus abundances (to 105ml) asmeasured by most probable number (MPN) cultivation tech-niques in the Gulf of Mexico near Texas (248 249) Othervirus cultivation studies with Prochlorococcus and Synecho-coccus in oligotrophic waters of the Sargasso Sea showedan interesting pattern of cross-infection between these gen-era by some virus types (suggesting gene flow among theseorganisms) but generally low MPN estimates of abundanceto 103ml even when cyanobacterial abundance was near105ml (216 250)
Overall the consensus emerging from direct comparisonsof viral-mediated mortality and grazer-mediated (ie micro-zooplankton) mortality have indicated that viral lysis of bac-teria constitutes a significant fraction of total mortality ofthis assemblage while the mortality of phytoplanktonappears to be dominated bymicrozooplankton inmost instan-ces (251 252)
As mentioned viruses have significant morphologicaldiversity as observed in TEM studies It is also possible toinvestigate aspects of their genetic diversity In early workon this topic Wommack et al (253) and Steward et al(254) observed the diversity of viral genome lengths in a fieldsample by pulsed field gel electrophoresis with viral genomesranging 25 kb to gt300 kb in length Field results show thatthe viral community composition is dynamic in space andtime with clear changes in the banding patterns over seasonsand locations in Chesapeake Bay (253) between oceanbasins and subsequent to dinoflagellate blooms (254) andwith depths to 500 m at one location (255)
Although viruses do not all share a set of core genesthat allows a universal viral phylogeny (comparable to SSUrRNA gene in cellular organisms) genetic diversity amonga single group of viruses can be examined by sequence analysisof shared genes within the group An example is the g20 geneinT4-like cyanophages that has shown extremely high diver-sity even among very closely related viruses and has demon-strated geographic and seasonal variation (256) A secondexample is g23 which is found broadly in diverse and wide-spread T4-like phages (257) and can have seasonally repeat-ing patterns (258) as well as short-term rapid dynamics thatcorrelate to those in bacteria (259)
Viral MetagenomicsAs with cellular organisms metagenomics can be used toexamine viral diversity and genetics without themany restric-tions of cultivation Viruses can be collected by selective fil-tration and concentrated by tangential flow filtration orflocculated with iron chloride (260 261) then their collec-tive metagenome can be extracted linker amplified and ana-lyzed by sequencing (262 263) Viral metagenomic studiesare particularly challenging because the large majority ofsequences have no annotated matches in any databases butso far results from marine samples around the world haveshown extremely high diversity and variations with depthlocation and time presumably with highly dispersed typesselected by local conditions (262ndash266) Because metage-nomes are best interpreted when there are representative cul-tures available the best matches of viral metagenomesoriginally tended to be to the few viral isolates infecting trulycommonmarine bacteria like cyanophage infecting Synecho-coccus and Prochlorococcus (267) However the developmentof dilution-to-extinction cultures representing common het-erotrophic marine taxa like Peligibacter (a member of theSAR11 clade) and SAR116 has allowed isolation from sea-water of viruses infecting these common organisms and theseisolates have indeed been found to be highly abundant inmarine viral metagenomes (268 269) New approaches tointerpret the results include clustering the proteins independ-ent of known proteins to compare samples to each other andtry to find environmental factors driving viral communitychanges (270) Such analysis of a large global data set (TaraOceans expedition) has shown that extensive sampling hascome close to reaching the total diversity in tropical and tem-perate waters of such viral protein clusters (which essentiallyrepresent various viral protein motifs not all viral proteintypes) and that viruses appear to be directionally dispersed
422-16 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquodownstreamrdquo in major ocean currents as one may expectfrom first principles but also supporting the ldquoseed bankrdquohypothesis of viral biogeography (271)
Viruses and Host DiversityViruses are themselves thought to be instrumental in drivingincreased diversity of their microbial hosts via a hypothesizedprocess often called ldquokill the winnerrdquo This is because viralinfection is host-specific and density-dependent the latterbecause viruses diffuse from host to host so an abundanthost is more likely to pass on infection than a rare oneThis means that if an organism becomes abundant andblooms winning the competition for resources it becomesmore susceptible to a viral epidemic This would benefit therarer organisms and thus help foster diversity (223 272273) However bloom scenarios as described above involvesystems far from steady state and the formal theory of Thing-stad and Lignell (274) has interesting steady-state solutionswhere several viruses infect several hosts stably over time(via trade-offs between growth rates and viral susceptibility)which may occur at the strain or species level (274) There issome experimental evidence that viruses have effects on nat-ural marine microbial community composition weaker thanthe kill the winner (bloom version) hypothesis would sug-gest (275 276) so something resembling the steady-statecoexistence described in the model may in fact be commonThere also appear to be processes that foster coexistencebetween viruses and hosts but the mechanisms are largelyspeculative (223 277 278)
Viruses may also be directly involved in host genetic diver-sity because they can be the agents of genetic exchangebetween microorganisms (223 273) This often involvesthe viral lifestyle known as lysogeny whereby viruses survivewithin host cells as DNA only integrated into the host chro-mosome and being reproduced each time the host divides Ahost harboring such a genome is called a lysogen becauseunder conditions of stress to the host cell a genetic switchmay cause the viral genome to initiate the lytic process pro-ducing many progeny viruses and bursting from the hostLysogeny is a very common property occurring in a signifi-cant part of the bacterial community (279 280) althoughthe incidence of induction of the lytic phase in nature is appa-rently low (281) Overall lysogeny is poorly understood butthought to have both positive and negative impacts on themicrobial community (282) Recent results suggest that ben-efits of being lysogenic in highly seasonal polar seas leads tofundamental difference between polar and other marine viralcommunities (283)
Viruses and the Microbial LoopAs part of the food web viruses occupy a unique positionThey infect host cells that are mostly thought to be heterotro-phic bacteria and by doing so they typically burst the hosts torelease progeny viruses and cellular debris But what is the fateof this material Viruses themselves do not last indefinitelyand a simple steady-state assumption implies that from eachburst of viruses (typically 20ndash100 per lytic event) only onesuccessfully infects another cell The rest are inactivatedand broken down by sunlight (UV and visible exposure)and enzymatic attack (284) or consumed byminute phagotro-phic protists (285) thus reentering the food web as substratefor bacteria or food for protistan consumers Experiments incontrolled laboratory systems and field studies with radioac-tively labeled viral lysis products have supported the conclu-sion that most of the organic matter released by the viralinfection is either taken up by bacteria or respired (286 287)
Modeling this process as part of the microbial loop showsthat viral lysis represent a sort of side loop that has the neteffect of remineralizing a significant amount of the carbonand nutrients that enter the bacteria-protist part of themicro-bial loop (Fig 7) A theoretical numerical steady-state modelcomparing a systemwith no viral activity to onewhere virusesare responsible for 50 of bacterial mortality showed that thesystem with viruses had 33 more bacterial production andrespiration than the virus-free system implying that theviruses had the effect of permitting the bacteria to processmore of the primary production than they would otherwise(223) Although 50 is a high number unlikely to be com-mon in the sea this model nevertheless illustrates that virusescan reduce the amount of energy reaching higher trophic lev-els The implication is that viruses lead to increased bacterialactivity at the expense of the larger organisms
MAJOR ENVIRONMENTAL CONTROLSLight Temperature and PressureTemperature has an important potential influence on bio-chemical reactions and therefore on biological processesin general Most ocean waters fall in the range of ndash2degC to30degC with obvious exceptions in hydrothermally heatedareas Temperature has long been known to be a regulatingfactor for the growth of heterotrophic microbes
In temperate waters it has been established that microbialactivity is generally much higher in warm summer waters thanin winter (55) The relationship is not simple howeverbecause multiple factors act at the same time Some contro-versy still exists regarding the highest and lowest extremesfor marine bacterial growth although there is broad agree-ment that bacteria grow gt100degC at hydrothermal vents andltminus5degC in sea ice brines Pomeroy et al (57) noted the inter-esting observation that bacteria seem particularly inhibitednear the freezing point of seawater (ca ndash22degC) comparedto eukaryotic phytoplankton This effect results in polarspring phytoplankton blooms that accumulate organic car-bon in advance of the response of the bacterial communityand development of the microbial loop and perhaps lead toenhanced benthic-pelagic coupling (288)
The relationship between temperature and the growth rateof marine phytoplankton was described broadly in a now
FIGURE 7 Modification of the microbial loop concept that incor-porates the functional role of viruses Export can be via predation orsinking From Fuhrman (223)doi1011289781555818821ch422f7
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-17
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
classic paper by Eppley (58) and later Goldman and Carpen-ter (289) Temperature was shown to exert a strong and directeffect on themaximal growth rates of these species withmax-imal intrinsic growth rates at 0degC generally lt1 divisiondaywhile growth rates at 30degC may be gt4day These relation-ships indicate the maximal rate that might be attained byphytoplankton at these temperatures but they do not takeother factors into account (nutrients light) While warmtemperature ostensibly allows more rapid growth it createshydrographic conditions that typically give rise to nutrientlimitation of algal growth Thus some of the coldest watersin nature witness some of the most massive phytoplanktonblooms (290) while warm oceanic gyres represent some ofthe most oligotrophic areas of the ocean The multiple andoften conflicting effects of temperature on the growth of nat-ural phytoplankton assemblages limit the accuracy of presentpredictions regarding how primary producers will respond toclimate change (291)
An analysis of the effect of temperature on the growth ofheterotrophic protists has indicated that the response is qual-itatively similar to that of phytoplankton but with a twistThe growth rates of at least some protozoa can exceed thoseof phytoplankton at warmer environmental temperaturesbut the opposite effect is apparent at very low environmentaltemperature This differential effect of temperature on thegrowth of phototrophic and heterotrophic protists was basedon a large meta-analysis of published protistan growth (59)That analysis demonstrated that the maximal growth ratesattained by phototrophic protists could exceed the maximalgrowth rates attained by heterotrophic protists (all otherpotential growth-limiting factors not considered) Thereforeprotozoan growth rates may be constrained to a greater degreeat low environmental temperature than rates for phytoplank-ton If so then phytoplankton bloomsmay get a head start ongrazers during spring in polar ecosystems This scenario is con-sistent with information on seasonal biomass changes andmicrozooplankton herbivory in the Ross Sea Antarctica(292 293) but there are still too few data to fully vet thishypothesis
The importance of high pressure on bacterial growthgained considerable attention in the late 1960s when thedeep sea submersible Alvin was accidentally lost overboardwith its hatch openWhile no lives were lost in this accidentsome workmenrsquos lunches sank to the bottom (sim1500 m)inside the submersible Alvin was recovered after 10 monthsand interestingly there was a waterlogged lunch containingapples bologna sandwiches and broken vacuum bottleswith broth that all appeared hardly degraded and tasted palat-able Yet when placed in a refrigerator on the ship these itemsdegraded relatively quickly Initially pressure was thought tobe the preserving factor as the sea floor temperature was sim-ilar to the refrigerator temperature and the only major differ-ence would be pressure (294) Following this observationa series of experiments to measure degradation of variousorganic materials left in the deep sea for extended periodsindicated that degradationwas typically significantly reducedimplying that pressure reduces the degradation rates (295)Nonetheless changes in the protozoan community of naturaldetrital material sinking to the deep ocean floor indicate thatthe microbial community can respond relatively quickly insome situations (296) Deep sea microbiology has advancedconsiderably yet it is still difficult to interpret results withrespect to actual in situ rates of naturally occurring organicmatter Deep sea bacteria adapted to high pressures havebeen isolated that are barophilic (also called piezophilic)meaning that they prefer high pressures and have reduced
activity at lower pressures whereas others are barotolerant(piezotolerant) tolerating but not preferring high pressuresfor example see (297 298)
Few data are available on barotolerantbarophilic marineprotozoa Protozoa certainly exist and grow at great oceanicdepths but measurements of in situ growth rates for these spe-cies do not yet exist Measurable protozoan numbers havebeen documented in the deep-sea sediments for more than30 years (299 300) and viable protozoa have occasionallybeen cultured from these environments (301ndash305) butvery few direct measurements of the activities of these speciesin situ have been reported (306) A few protozoa have beenisolated that will grow at high pressure (302 303 307) andprotists that appear to be unique to the deep ocean havebeen observed either directly or through the analysis ofDNA sequences (308ndash311) but possibly the best direct evi-dence that protozoan activity takes place at the high pressurescharacteristics of the deep sea are experimental and observa-tional work noting the stimulatory effect that detrital deposi-tion has on some components of the protozoan community(296 312) These observations indicate a diverse and activeprotozoan fauna of the deep ocean although their biogeo-chemical significance is largely uncharacterized at this time
Dissolved and Particulate Organic MatterBacteria and archaea are thought to be by far the most impor-tant organisms with respect to the processing of dissolvedorganic matter (DOM) and nonliving particulate organicmatter (POM also called detritus) in the ocean While theremay be some uptake of DOM by protists particularly forgrowth factors needed in trace amounts such as vitamins(313) the bulk of this material is probably utilized by bacteriaand archaea (314) including the smallest cyanobacteriumProchlorococcus (315 316) which thus may be considered amixotroph Due to their small size bacteria have extremelyhigh surfacevolume ratios and combined with their over-whelmingly high abundance an extremely high integratedsurface area Protozoa tend to obtain the organic materialsthat they require for growth from their prey rather thanthrough the uptake of DOM Overall protozoa tend to besources of dissolved and detrital organic substances throughthe excretion of unassimilated prey biomass in expelledfood vacuoles
Particulate organic matter is not directly available as sub-strate to bacteria These substances must first be reduced tosmall molecules that can be transported into the cell Thisis accomplished by the production of extracellular enzymes(note that few if any large polymers are directly taken up bybacteria with the possible exception of DNA) Hydrolyticenzymes produced by bacteria (and almost certainly archaea)break down polymers like proteins polysaccharides andnucleic acids As with DOM POM is composed of a complexmixture of compounds that vary in their susceptibility to bac-terial degradation and utilization
Particulate material in the water column serves not only asbacterial substrate but also as substratum POM occurs in thewater column across a huge size spectrum from micrometersup to some detrital aggregates more than 1 m in diameter(317) Much of this particulate material is in a constant stateof flux with colloidal material constantly coalescing andaggregating to form new or larger particles (318) as microbialdegradation acts simultaneously to remineralize this materialDetrital particles that attain macroscopic size either by directformation (317 319 320) or via accretion and aggregation(321) and are often called marine snow or macroaggregatesMarine snow particles are readily colonized by bacteria and
422-18 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
many other microorganisms and their abundances on theseaggregates can be orders of magnitude greater than abundan-ces in an equivalent volume of the water surrounding theaggregates (320 322ndash324) and their diversity and highlyconcentrated abundances on detrital aggregates implies thatthere may be stronger trophic coupling and efficient energyelemental cycling relative to these processes among free-living microorganisms in the surrounding waters This specu-lation has resulted in a number of formalizations of the poten-tial impact of aggregate microbiology on biogeochemicalprocesses in the ocean (325 326) Most definitions of bio-films include aggregated particulate and these particles areoften studied within the biofilm conceptual model (see chap-ter by Lawrence et al in this section)
Inorganic Nutrients
Macronutrients (N P)Virtually all heterotrophic organisms in the ocean contributeto the pool of available macronutrients via the excretion ofmetabolic wastes and protozoa contribute significantly tothis process Macronutrients are often in excess within proto-zoan prey relative to the consumersrsquo needs because much ofthe prey carbon is respired to produce energy for metabolismand growth ldquoExcessrdquo nutrients are eliminated as eitherorganic compounds or more commonly as ammonium (forN) and phosphate (for P) The importance of the match(or mismatch) between predator and prey stoichiometry hasbeen demonstrated experimentally and repeatedly in marineand freshwater ecosystems and has been elevated to a funda-mental ecological principle governing elemental and energyflow in aquatic food webs (327) Based on this reasoningCaron et al (328) concluded that bacterivorous protozoaplay an important role in nutrient remineralization andrelease in the ocean Bacterial biomass is typically rich in Nand P relative to other organisms so bacterial predators expe-rience large excesses of these elements relative to their growthneedsMoreover bacterivorous protozoa should be dispropor-tionately important in nutrient remineralization amongsingle-celled eukaryotes because as the smallest protozoa inaquatic ecosystems they have high weight-specific metabolicrates (329) Thus both stoichiometric and allometric rela-tionships implicate small bacterivorous protozoa as impor-tant sources of remineralized nutrients in the plankton
The reasoning also holds true for bacterial utilization oforganic compounds and it has led to some interestingfindings and conclusions regarding the role of bacteria innutrient cycling in the ocean Bacteria are important sourcesremineralized nutrients when they are consuming N- orP-rich substrate Contrary to this traditional view howeverheterotrophic bacteria are often strong competitors for inor-ganic nutrients under many situations largely because bacte-rial biomass is richer in N and P than much of the organicmatter they consume in nature so they require additional Nand P to produce their biomass (summarized by [330]) Exper-imental studies have demonstrated the uptake of ammoniumby bacteria other than cyanobacteria using stable isotope 15N(331) and also the short-lived radioisotope 13N (332) oftenfinding that bacteria may be responsible for one third of the Nor P uptake Additionally there is evidence that bacteria insome oceans like the Sargasso Sea are limited by phosphorus(333 334) Growth limitation of bacteria by phosphorusappears to be a common phenomenon of freshwater ecosys-tems (335 336)
N2 fixation is restricted to prokaryotes and in the oceanwater column it was thought for many years to be done
primarily byTrichodesmium a warm-water colonial cyanobac-terium that blooms sporadically andRichelia a cyanobacterialsymbiont that lives within certain diatoms (337) Some uni-cellular cyanobacteria also contribute significantly to globalN2 fixation (338) and molecular biological data suggestthat a variety of other bacteriamdashnot phytoplanktonmdashmayalso be fixing nitrogen in seawater (339) An exciting recentdiscovery in this field is that a bacterial nitrogen fixer origi-nally known only from nifH sequences UCYN-A has hadits genome sequenced by parallel pyrosequencing and wasfound be a cyanobacterium (Candidatus Atelocyanobacteriumthalassa) lacking the apparatus to generate oxygen as well asother pathways (340) Further study showed it to be symbioticwith a marine alga (341)
Micronutrients (Trace Metals Growth Factors)Bacterial growth may often be limited by macronutrients asdescribed and sometimes by trace nutrients particularlyiron that occur at very low concentrations in seawater(342 343) The importance of Fe as an element limiting pri-mary production in some oceanic regions has come underclose scrutiny and extensive experimental investigation inrecent years Many bacteria have special uptake mechanismsto utilize extremely low concentrations of iron includingsiderophores (released compounds that bind iron and arethen specifically taken up) Even if bacteria are capable ofgrowth at low Fe concentrations growth may be less efficientKirchman et al (344) found that Fe limitation stronglyreduced bacterial growth efficiency in a cultured marine bac-terium (ca 50 efficiency Fe replete but lt10 when Fedeplete) Thus the effects may be complex Marine N2 fixa-tion also may be limited by the availability of Fe becausenitrogen fixers have high Fe requirements
Protozoa tend to be a source rather than a sink for Fe in theocean analogous to their role in macronutrient remineraliza-tion Protozoan grazing activity has been shown to releaseiron from bacterial biomass thereby relieving Fe limitationin co-occurring phytoplankton (345) a mechanistic demon-stration of the process using protozoa growing on Fe-repletebacteria However it has also been reported that Fe-limitedbacteria may contain insufficient Fe for the bacterivorous pro-tozoa that consume them thus leading to Fe limitation in theprotozoan as well (346)
OxygenHypoxic and anoxic regions of the ocean are expected to sig-nificantly expand in the coming decades as a consequence ofchanges in ocean stratification resulting from global climatechange (347) As oxygen becomes depleted bacteria havethe capability to utilize a series of alternate electron acceptorsCompositional changes in the bacterial assemblage accom-pany these changes in metabolic activity but are beyondthe scope of this chapter Many of the studies on benthicmicrobiology have focused on redox reactions that occurthere Oxygen is usually consumed by sediment microbialactivity much faster than the rate at which it can be replacedby diffusion so most sediments especially organically richones are anaerobic below a few mm (fine-grained muds) orcm (coarser sediments) from the sedimentwater interfaceexcept where animals ventilate the benthos through tubesWhen oxygen is absent bacteria and archaea use alternateelectron acceptors such as nitrate nitrite oxidized Fe orMn organic matter or sulfate The use of nitrate or nitriteas an electron acceptor with concomitant production of N2
gas (denitrification) results in net loss of biologically availablenitrogen Bacteria that perform this reaction are typically
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-19
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquofacultativerdquo organisms that can switch from oxygen to nitrateor nitrite Organic matter may be reduced by fermentationreactions in which part of an organic substrate is oxidizedwhile another is reduced fermenter organisms are also typi-cally facultative Sulfate is a particularly important alternateelectron acceptor in marine systems because it occurs as thesecond most abundant anion in seawater (behind chloride)typically 28 mM Its use as an electron acceptor leads to theproduction of elemental sulfur or sulfide The use of sulfateas an electron acceptor yields much less energy than oxygenor nitrate and sulfate-reducing bacteria are strict anaerobesunable to use oxygen (often strongly inhibited by oxygen)Methane can be produced by methanogenic archaea whichare all strict anaerobes using CO2 as an electron acceptor(see Findlay and Battin this section for further discussion)
Reduced compounds such as sulfide are produced by theuse of these alternate electron acceptors and they diffuse awayfrom their source When they diffuse to environments with astrong oxidant like oxygen they may be oxidized by chemo-trophic bacteria in energy-yielding reactions Similarlyammonium can be oxidized to nitrite and then nitrate by che-motrophic nitrifying bacteria and the nitrate can diffuse intoanaerobic layers and become denitrified to N2 Such coupledoxidation-reduction reactions can lead to accelerated biogeo-chemical cycling particularly in sediments
It was originally believed that relatively few free-livingspecies of protozoa inhabit anaerobic and hypoxic marineenvironments This view has changed based on (largely)molecular analyses of some of these environments (308348ndash351) A significant number of previously undescribedeukaryotic phylotypes have been observed in anoxic environ-ments High protozoan biodiversity in the absence of molec-ular oxygen is not completely unexpected because otheranaerobic environments such as some gut environments(eg cockroach guts and cattle rumen) have notoriouslyabundant and diverse protozoan fauna but marine anaerobicenvironments have been difficult to access and sample prop-erly Many anaerobic protozoa exist together with symbioticbacteria (352 353) or possess unique organelles such as thehydrogenosome (354)
MAJOR BIOTIC CONTROLSThe Trophic Activities of Protozoa Predation andTop-Down ControlsPredator-prey relationships are fundamental interactionswithin biological communities linking production of bio-mass on the one hand with removal on the other hand Estab-lishing the trophic relationships among microorganisms andquantifying the rates at which they take place constitutemajor endeavors within marine microbial ecology As notedpreviously protozoa consume a wide variety of prokaryoticand protistan prey Numerous laboratory and field measure-ments have been made throughout the past quarter centuryto attempt to quantify the rates of bacterial and phytoplank-ton consumption The methodology for accomplishing thesestudies has varied and a number of approaches are presentlyemployed
Methodologies for estimating microbial grazing are diversebut generally fall into one of two experimental approachesperturbation experiments or tracer experiments Perturbationexperiments (size fractionation metabolic inhibitors anddilution technique) rely on manipulation of the consumeror prey assemblages in somemanner such that the effect is evi-dent from changes in the abundance of the prey population
during incubations relative to control treatments that receiveno manipulation Tracer experiments (radioactively labeledprey fluorescently labeled prey stable isotopendashlabeled pray)attempt to use labeled prey or a prey proxy to quantitativelyfollow the movement of prey biomass into consumers Forsummaries of these methods see (74 355)
Perturbation experiments have entailed a variety of mech-anisms to ldquodecouplerdquo predators from prey in natural microbialcommunities For example size fractionation uses various sizefilters to remove an entire size class of grazers from awater sam-ple leaving the prey to grow in the relative absence of thosepredators during subsequent incubations Unfiltered samples(controls) indicate the growth of prey in the presence of pre-dation Predation impact is obtained by the difference ingrowth of the prey between these treatments This methodhas been used to examine predation on cyanobacteria heter-otrophic bacteria and minute phototrophic eukaryotesAlternatively metabolic inhibitors have been employed tohalt predation (or in some cases prokaryote growth) insteadof physical removal by filtration
The dilution technique (356) has become a commonmethod for examining grazing mortality of phytoplanktonby microzooplankton (which is comprised predominantly ofprotozoa) This method relies on the dilution of herbivorouszooplankton in a series of subsamples (and concomitantmeasurements of phytoplankton growth in each subsample)to estimate the grazing mortality of the herbivores
Tracer experiments have used both radioactively labeledprey and fluorescently labeled prey to examine predation onbacteria and phytoplankton Radioactive labeling has beenaccomplished using 14C- or 3H-labeled organic compounds(for bacteria) or 14C-bicarbonate (for phytoplankton) Theuse of radioisotope approaches has diminished with improve-ments in the number and types of fluorescent compoundsavailable for this work Fluorescently labeled prey havebeen labeled with a variety of compounds that allow theprey to be observed readily (even inside predators) by epi-fluorescencemicroscopy (357 358) Alternatively a decreasein the total number of fluorescent prey in a sample indicatesthe rate of removal of the prey assemblage
Both perturbation experiments and tracer experimentsdescribed have inherent advantages and disadvantages Forexample perturbation experiments are fairly straightforwardto conduct but they may introduce artifact by removingpotentially important sources of nutrients or organic com-pounds released by predators (359) On the other hand thedilution technique is very labor-intensive but one advantageof this method is that it provides the opportunity to determineboth phytoplankton growth and grazing mortality in a singleprocedure Most tracer experiments are fairly easy to conductand generally do not perturb the food web being studied butfeeding selectivity for or against the tracer (relative to naturalprey) may produce spurious results In addition some of thesemethods provide a measurement of the activity of the entiregrazer assemblage and others are applicable for examiningthe activity of specific taxa For example size fractionationapproaches examine the effect of the removal of an entiresize class of organisms (a community-levelrdquo measurement)whereas fluorescently labeled prey can be used to examinethe ingestion rate of a particular species by observing theuptake of labeled prey into that species (a species-levelmeasurement)
Predation rates of numerous marine protozoa have beenexamined in the laboratory From these experiments a gen-eral knowledge of the rates of consumption by these specieshas been accumulated Feeding in most protozoa shows a
422-20 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
functional response to the availability of prey that is similar tothe response of many organisms (although there are a greatdeal of species-specific differences) That is ingestion rateincreases rapidly as prey abundance increases until a maximalingestion rate is attained (360)Manyof these curves have theform of a Type II functional response curve (361)
Based on the features of these curves it is clear that proto-zoa have the ability to ingest substantial numbers of prokar-yote and eukaryote prey in natural ecosystems Somebacterivorous flagellates can consume bacteria (and cyano-bacteria) at rates up to 100 s per individual per hour whereassome bacterivorous ciliates can consume thousands of bacte-ria per individual per hour Rates of consumption of eukary-otic phytoplankton by herbivorous ciliates and largeflagellates are considerably more variable and highly depend-ent of the feeding mechanism (eg filtration in ciliates versuspseudopodial net in dinoflagellates) and the size of the prey
These lab studies demonstrate that rarely are prey suffi-ciently abundant in nature to support the maximal ingestionrates of which many protozoa are capable Nevertheless pre-dation rates of protozoa in nature are sufficient to constitute amajor factor controlling the standing stocks of bacteria andphytoplankton (63 180 184 355) For example these sum-maries indicate that on average protozoa consume 20ndash80ofthe standing stock of phytoplankton (measured as chloro-phyll) in the world ocean and 60ndash75 of daily primary pro-duction a departure from the classical view of copepods as theprimary consumers of phytoplankton This large role of her-bivorous protozoa infers a large role for copepods as consumersof planktonic protozoa (185) Similarly the role of protozoaas major consumers of bacterial biomass is indicated by therelative balance between bacterial production on the onehand and bacterivory on the other (Fig 8) The approximateone-to-one balance between these measurements over a largenumber of studies implies that protozoan bacterivory must bea major factor determining the fate of bacteria in marine (andfreshwater) plankton ecosystems
It is interesting to note that some types of bacteria includ-ing marine Bdellovibrio are known to prey on other marine
bacteria (362) At the time of this writing the importanceof bacterial predation by other bacteria within the contextof oceanographic processes is not known and presumed tobe small While Bdellovibrio may be cultivated from seawaterit can also be found among sequences from cold sediments(363) and the water column Bdellovibrio happens to be thebest studied predatory bacterium but there are no doubtothers that we simply have not yet identified (364)
Species-Specific Interactions (MutualismParasitism Commensalisms)Interactions between marine microorganisms such as compe-tition allelopathy and symbiosis are fundamental aspectscontrolling community composition and activity but atpresent most are poorly characterized in natural marine sys-tems Bacteria must compete for dissolved and particulatecompounds with other bacteria andwith protistan taxa whilephagotrophic protists must compete with each other andmetazoa for available prey These interactions manifest them-selves in many forms including true competition for availableresources resource partitioning such that some taxa specializeon the utilization of certain substrates or prey while othersspecialize on different ones (reducing competition) or eventhe cooperative use of compounds (eg one bacteriumrsquoswaste products being utilized as substrate by another bacte-rium) These metabolic consortia are probably common inthe ocean but can be difficult to identify and characterizeConsortia of bacteria working together to perform complexreactions especially in anaerobic systems are well known inmicrobiology
Consortia in which there is a very high degree of integra-tion between species (ie symbiosis) are very common in theocean These associations may be beneficial to both partners(mutualism) beneficial to one part and inconsequential forthe other (commensalism) or beneficial to one part and det-rimental to the other (parasitism) Many highly visible andwell-characterized marine mutualisms exist between chemo-synthetic bacteria and animals such as hydrothermal ventor methane seep tubeworms clams mussels or other inverte-brates (365) and also between luminescent bacteria and fishor squids (366) Similarly many protozoa form mutualisticassociations with photosynthetic protists (367) The mostconspicuous of these associations involved large amoeboidprotozoa of oceanic ecosystems (planktonic foraminiferanspolycystine radiolaria and acantharia) which contain largenumbers of intracellular algae (74) but associations betweensome heterotrophic dinoflagellates and cyanobacteria are alsocommon (368) Studies have indicated that efficient nutrientcycling in these protozoan-algal associations confers an eco-logical advantage for both partners in surfacewaters of the oli-gotrophic ocean
Parasitism is also a common symbiotic relationship amongmarine protozoa Protozoan parasites have been reported for adiverse array of protists and animals in the ocean Heterotro-phic dinoflagellates and presently uncultured alveolate cladesrelated to alveolates appear to be particularly successful atevolving thismode of existence (369ndash371) In a common par-asitic group Amoebophrya spp the parasite invades photo-synthetic dinoflagellates where it undergoes reproductioneventually lysing the host and releasing dozens of infectiveswarmers The parasitic protist Pirsonia spp is a parasite ofdiatoms (372) The overall importance of these parasitismsin the population dynamics of their hosts is not clearly knownbut may constitute a significant source of mortality for somespecies (373 374) The frequency and diversity of ldquonewrdquo
FIGURE 8 Meta-analysis of experimental studies that have com-pared rates of bacterial productivity (converted to population growthrate units of dminus1) with rates of bacterivory by phagotrophic protists(expressed as equivalent units of bacterial growth rate dminus1) Thesolid line indicates where measurements of bacterial removal by graz-ers were equal tomeasurements of bacterial production Data redrawnfrom Sanders et al (63) doi1011289781555818821ch422f8
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-21
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquodownstreamrdquo in major ocean currents as one may expectfrom first principles but also supporting the ldquoseed bankrdquohypothesis of viral biogeography (271)
Viruses and Host DiversityViruses are themselves thought to be instrumental in drivingincreased diversity of their microbial hosts via a hypothesizedprocess often called ldquokill the winnerrdquo This is because viralinfection is host-specific and density-dependent the latterbecause viruses diffuse from host to host so an abundanthost is more likely to pass on infection than a rare oneThis means that if an organism becomes abundant andblooms winning the competition for resources it becomesmore susceptible to a viral epidemic This would benefit therarer organisms and thus help foster diversity (223 272273) However bloom scenarios as described above involvesystems far from steady state and the formal theory of Thing-stad and Lignell (274) has interesting steady-state solutionswhere several viruses infect several hosts stably over time(via trade-offs between growth rates and viral susceptibility)which may occur at the strain or species level (274) There issome experimental evidence that viruses have effects on nat-ural marine microbial community composition weaker thanthe kill the winner (bloom version) hypothesis would sug-gest (275 276) so something resembling the steady-statecoexistence described in the model may in fact be commonThere also appear to be processes that foster coexistencebetween viruses and hosts but the mechanisms are largelyspeculative (223 277 278)
Viruses may also be directly involved in host genetic diver-sity because they can be the agents of genetic exchangebetween microorganisms (223 273) This often involvesthe viral lifestyle known as lysogeny whereby viruses survivewithin host cells as DNA only integrated into the host chro-mosome and being reproduced each time the host divides Ahost harboring such a genome is called a lysogen becauseunder conditions of stress to the host cell a genetic switchmay cause the viral genome to initiate the lytic process pro-ducing many progeny viruses and bursting from the hostLysogeny is a very common property occurring in a signifi-cant part of the bacterial community (279 280) althoughthe incidence of induction of the lytic phase in nature is appa-rently low (281) Overall lysogeny is poorly understood butthought to have both positive and negative impacts on themicrobial community (282) Recent results suggest that ben-efits of being lysogenic in highly seasonal polar seas leads tofundamental difference between polar and other marine viralcommunities (283)
Viruses and the Microbial LoopAs part of the food web viruses occupy a unique positionThey infect host cells that are mostly thought to be heterotro-phic bacteria and by doing so they typically burst the hosts torelease progeny viruses and cellular debris But what is the fateof this material Viruses themselves do not last indefinitelyand a simple steady-state assumption implies that from eachburst of viruses (typically 20ndash100 per lytic event) only onesuccessfully infects another cell The rest are inactivatedand broken down by sunlight (UV and visible exposure)and enzymatic attack (284) or consumed byminute phagotro-phic protists (285) thus reentering the food web as substratefor bacteria or food for protistan consumers Experiments incontrolled laboratory systems and field studies with radioac-tively labeled viral lysis products have supported the conclu-sion that most of the organic matter released by the viralinfection is either taken up by bacteria or respired (286 287)
Modeling this process as part of the microbial loop showsthat viral lysis represent a sort of side loop that has the neteffect of remineralizing a significant amount of the carbonand nutrients that enter the bacteria-protist part of themicro-bial loop (Fig 7) A theoretical numerical steady-state modelcomparing a systemwith no viral activity to onewhere virusesare responsible for 50 of bacterial mortality showed that thesystem with viruses had 33 more bacterial production andrespiration than the virus-free system implying that theviruses had the effect of permitting the bacteria to processmore of the primary production than they would otherwise(223) Although 50 is a high number unlikely to be com-mon in the sea this model nevertheless illustrates that virusescan reduce the amount of energy reaching higher trophic lev-els The implication is that viruses lead to increased bacterialactivity at the expense of the larger organisms
MAJOR ENVIRONMENTAL CONTROLSLight Temperature and PressureTemperature has an important potential influence on bio-chemical reactions and therefore on biological processesin general Most ocean waters fall in the range of ndash2degC to30degC with obvious exceptions in hydrothermally heatedareas Temperature has long been known to be a regulatingfactor for the growth of heterotrophic microbes
In temperate waters it has been established that microbialactivity is generally much higher in warm summer waters thanin winter (55) The relationship is not simple howeverbecause multiple factors act at the same time Some contro-versy still exists regarding the highest and lowest extremesfor marine bacterial growth although there is broad agree-ment that bacteria grow gt100degC at hydrothermal vents andltminus5degC in sea ice brines Pomeroy et al (57) noted the inter-esting observation that bacteria seem particularly inhibitednear the freezing point of seawater (ca ndash22degC) comparedto eukaryotic phytoplankton This effect results in polarspring phytoplankton blooms that accumulate organic car-bon in advance of the response of the bacterial communityand development of the microbial loop and perhaps lead toenhanced benthic-pelagic coupling (288)
The relationship between temperature and the growth rateof marine phytoplankton was described broadly in a now
FIGURE 7 Modification of the microbial loop concept that incor-porates the functional role of viruses Export can be via predation orsinking From Fuhrman (223)doi1011289781555818821ch422f7
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-17
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
classic paper by Eppley (58) and later Goldman and Carpen-ter (289) Temperature was shown to exert a strong and directeffect on themaximal growth rates of these species withmax-imal intrinsic growth rates at 0degC generally lt1 divisiondaywhile growth rates at 30degC may be gt4day These relation-ships indicate the maximal rate that might be attained byphytoplankton at these temperatures but they do not takeother factors into account (nutrients light) While warmtemperature ostensibly allows more rapid growth it createshydrographic conditions that typically give rise to nutrientlimitation of algal growth Thus some of the coldest watersin nature witness some of the most massive phytoplanktonblooms (290) while warm oceanic gyres represent some ofthe most oligotrophic areas of the ocean The multiple andoften conflicting effects of temperature on the growth of nat-ural phytoplankton assemblages limit the accuracy of presentpredictions regarding how primary producers will respond toclimate change (291)
An analysis of the effect of temperature on the growth ofheterotrophic protists has indicated that the response is qual-itatively similar to that of phytoplankton but with a twistThe growth rates of at least some protozoa can exceed thoseof phytoplankton at warmer environmental temperaturesbut the opposite effect is apparent at very low environmentaltemperature This differential effect of temperature on thegrowth of phototrophic and heterotrophic protists was basedon a large meta-analysis of published protistan growth (59)That analysis demonstrated that the maximal growth ratesattained by phototrophic protists could exceed the maximalgrowth rates attained by heterotrophic protists (all otherpotential growth-limiting factors not considered) Thereforeprotozoan growth rates may be constrained to a greater degreeat low environmental temperature than rates for phytoplank-ton If so then phytoplankton bloomsmay get a head start ongrazers during spring in polar ecosystems This scenario is con-sistent with information on seasonal biomass changes andmicrozooplankton herbivory in the Ross Sea Antarctica(292 293) but there are still too few data to fully vet thishypothesis
The importance of high pressure on bacterial growthgained considerable attention in the late 1960s when thedeep sea submersible Alvin was accidentally lost overboardwith its hatch openWhile no lives were lost in this accidentsome workmenrsquos lunches sank to the bottom (sim1500 m)inside the submersible Alvin was recovered after 10 monthsand interestingly there was a waterlogged lunch containingapples bologna sandwiches and broken vacuum bottleswith broth that all appeared hardly degraded and tasted palat-able Yet when placed in a refrigerator on the ship these itemsdegraded relatively quickly Initially pressure was thought tobe the preserving factor as the sea floor temperature was sim-ilar to the refrigerator temperature and the only major differ-ence would be pressure (294) Following this observationa series of experiments to measure degradation of variousorganic materials left in the deep sea for extended periodsindicated that degradationwas typically significantly reducedimplying that pressure reduces the degradation rates (295)Nonetheless changes in the protozoan community of naturaldetrital material sinking to the deep ocean floor indicate thatthe microbial community can respond relatively quickly insome situations (296) Deep sea microbiology has advancedconsiderably yet it is still difficult to interpret results withrespect to actual in situ rates of naturally occurring organicmatter Deep sea bacteria adapted to high pressures havebeen isolated that are barophilic (also called piezophilic)meaning that they prefer high pressures and have reduced
activity at lower pressures whereas others are barotolerant(piezotolerant) tolerating but not preferring high pressuresfor example see (297 298)
Few data are available on barotolerantbarophilic marineprotozoa Protozoa certainly exist and grow at great oceanicdepths but measurements of in situ growth rates for these spe-cies do not yet exist Measurable protozoan numbers havebeen documented in the deep-sea sediments for more than30 years (299 300) and viable protozoa have occasionallybeen cultured from these environments (301ndash305) butvery few direct measurements of the activities of these speciesin situ have been reported (306) A few protozoa have beenisolated that will grow at high pressure (302 303 307) andprotists that appear to be unique to the deep ocean havebeen observed either directly or through the analysis ofDNA sequences (308ndash311) but possibly the best direct evi-dence that protozoan activity takes place at the high pressurescharacteristics of the deep sea are experimental and observa-tional work noting the stimulatory effect that detrital deposi-tion has on some components of the protozoan community(296 312) These observations indicate a diverse and activeprotozoan fauna of the deep ocean although their biogeo-chemical significance is largely uncharacterized at this time
Dissolved and Particulate Organic MatterBacteria and archaea are thought to be by far the most impor-tant organisms with respect to the processing of dissolvedorganic matter (DOM) and nonliving particulate organicmatter (POM also called detritus) in the ocean While theremay be some uptake of DOM by protists particularly forgrowth factors needed in trace amounts such as vitamins(313) the bulk of this material is probably utilized by bacteriaand archaea (314) including the smallest cyanobacteriumProchlorococcus (315 316) which thus may be considered amixotroph Due to their small size bacteria have extremelyhigh surfacevolume ratios and combined with their over-whelmingly high abundance an extremely high integratedsurface area Protozoa tend to obtain the organic materialsthat they require for growth from their prey rather thanthrough the uptake of DOM Overall protozoa tend to besources of dissolved and detrital organic substances throughthe excretion of unassimilated prey biomass in expelledfood vacuoles
Particulate organic matter is not directly available as sub-strate to bacteria These substances must first be reduced tosmall molecules that can be transported into the cell Thisis accomplished by the production of extracellular enzymes(note that few if any large polymers are directly taken up bybacteria with the possible exception of DNA) Hydrolyticenzymes produced by bacteria (and almost certainly archaea)break down polymers like proteins polysaccharides andnucleic acids As with DOM POM is composed of a complexmixture of compounds that vary in their susceptibility to bac-terial degradation and utilization
Particulate material in the water column serves not only asbacterial substrate but also as substratum POM occurs in thewater column across a huge size spectrum from micrometersup to some detrital aggregates more than 1 m in diameter(317) Much of this particulate material is in a constant stateof flux with colloidal material constantly coalescing andaggregating to form new or larger particles (318) as microbialdegradation acts simultaneously to remineralize this materialDetrital particles that attain macroscopic size either by directformation (317 319 320) or via accretion and aggregation(321) and are often called marine snow or macroaggregatesMarine snow particles are readily colonized by bacteria and
422-18 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
many other microorganisms and their abundances on theseaggregates can be orders of magnitude greater than abundan-ces in an equivalent volume of the water surrounding theaggregates (320 322ndash324) and their diversity and highlyconcentrated abundances on detrital aggregates implies thatthere may be stronger trophic coupling and efficient energyelemental cycling relative to these processes among free-living microorganisms in the surrounding waters This specu-lation has resulted in a number of formalizations of the poten-tial impact of aggregate microbiology on biogeochemicalprocesses in the ocean (325 326) Most definitions of bio-films include aggregated particulate and these particles areoften studied within the biofilm conceptual model (see chap-ter by Lawrence et al in this section)
Inorganic Nutrients
Macronutrients (N P)Virtually all heterotrophic organisms in the ocean contributeto the pool of available macronutrients via the excretion ofmetabolic wastes and protozoa contribute significantly tothis process Macronutrients are often in excess within proto-zoan prey relative to the consumersrsquo needs because much ofthe prey carbon is respired to produce energy for metabolismand growth ldquoExcessrdquo nutrients are eliminated as eitherorganic compounds or more commonly as ammonium (forN) and phosphate (for P) The importance of the match(or mismatch) between predator and prey stoichiometry hasbeen demonstrated experimentally and repeatedly in marineand freshwater ecosystems and has been elevated to a funda-mental ecological principle governing elemental and energyflow in aquatic food webs (327) Based on this reasoningCaron et al (328) concluded that bacterivorous protozoaplay an important role in nutrient remineralization andrelease in the ocean Bacterial biomass is typically rich in Nand P relative to other organisms so bacterial predators expe-rience large excesses of these elements relative to their growthneedsMoreover bacterivorous protozoa should be dispropor-tionately important in nutrient remineralization amongsingle-celled eukaryotes because as the smallest protozoa inaquatic ecosystems they have high weight-specific metabolicrates (329) Thus both stoichiometric and allometric rela-tionships implicate small bacterivorous protozoa as impor-tant sources of remineralized nutrients in the plankton
The reasoning also holds true for bacterial utilization oforganic compounds and it has led to some interestingfindings and conclusions regarding the role of bacteria innutrient cycling in the ocean Bacteria are important sourcesremineralized nutrients when they are consuming N- orP-rich substrate Contrary to this traditional view howeverheterotrophic bacteria are often strong competitors for inor-ganic nutrients under many situations largely because bacte-rial biomass is richer in N and P than much of the organicmatter they consume in nature so they require additional Nand P to produce their biomass (summarized by [330]) Exper-imental studies have demonstrated the uptake of ammoniumby bacteria other than cyanobacteria using stable isotope 15N(331) and also the short-lived radioisotope 13N (332) oftenfinding that bacteria may be responsible for one third of the Nor P uptake Additionally there is evidence that bacteria insome oceans like the Sargasso Sea are limited by phosphorus(333 334) Growth limitation of bacteria by phosphorusappears to be a common phenomenon of freshwater ecosys-tems (335 336)
N2 fixation is restricted to prokaryotes and in the oceanwater column it was thought for many years to be done
primarily byTrichodesmium a warm-water colonial cyanobac-terium that blooms sporadically andRichelia a cyanobacterialsymbiont that lives within certain diatoms (337) Some uni-cellular cyanobacteria also contribute significantly to globalN2 fixation (338) and molecular biological data suggestthat a variety of other bacteriamdashnot phytoplanktonmdashmayalso be fixing nitrogen in seawater (339) An exciting recentdiscovery in this field is that a bacterial nitrogen fixer origi-nally known only from nifH sequences UCYN-A has hadits genome sequenced by parallel pyrosequencing and wasfound be a cyanobacterium (Candidatus Atelocyanobacteriumthalassa) lacking the apparatus to generate oxygen as well asother pathways (340) Further study showed it to be symbioticwith a marine alga (341)
Micronutrients (Trace Metals Growth Factors)Bacterial growth may often be limited by macronutrients asdescribed and sometimes by trace nutrients particularlyiron that occur at very low concentrations in seawater(342 343) The importance of Fe as an element limiting pri-mary production in some oceanic regions has come underclose scrutiny and extensive experimental investigation inrecent years Many bacteria have special uptake mechanismsto utilize extremely low concentrations of iron includingsiderophores (released compounds that bind iron and arethen specifically taken up) Even if bacteria are capable ofgrowth at low Fe concentrations growth may be less efficientKirchman et al (344) found that Fe limitation stronglyreduced bacterial growth efficiency in a cultured marine bac-terium (ca 50 efficiency Fe replete but lt10 when Fedeplete) Thus the effects may be complex Marine N2 fixa-tion also may be limited by the availability of Fe becausenitrogen fixers have high Fe requirements
Protozoa tend to be a source rather than a sink for Fe in theocean analogous to their role in macronutrient remineraliza-tion Protozoan grazing activity has been shown to releaseiron from bacterial biomass thereby relieving Fe limitationin co-occurring phytoplankton (345) a mechanistic demon-stration of the process using protozoa growing on Fe-repletebacteria However it has also been reported that Fe-limitedbacteria may contain insufficient Fe for the bacterivorous pro-tozoa that consume them thus leading to Fe limitation in theprotozoan as well (346)
OxygenHypoxic and anoxic regions of the ocean are expected to sig-nificantly expand in the coming decades as a consequence ofchanges in ocean stratification resulting from global climatechange (347) As oxygen becomes depleted bacteria havethe capability to utilize a series of alternate electron acceptorsCompositional changes in the bacterial assemblage accom-pany these changes in metabolic activity but are beyondthe scope of this chapter Many of the studies on benthicmicrobiology have focused on redox reactions that occurthere Oxygen is usually consumed by sediment microbialactivity much faster than the rate at which it can be replacedby diffusion so most sediments especially organically richones are anaerobic below a few mm (fine-grained muds) orcm (coarser sediments) from the sedimentwater interfaceexcept where animals ventilate the benthos through tubesWhen oxygen is absent bacteria and archaea use alternateelectron acceptors such as nitrate nitrite oxidized Fe orMn organic matter or sulfate The use of nitrate or nitriteas an electron acceptor with concomitant production of N2
gas (denitrification) results in net loss of biologically availablenitrogen Bacteria that perform this reaction are typically
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-19
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquofacultativerdquo organisms that can switch from oxygen to nitrateor nitrite Organic matter may be reduced by fermentationreactions in which part of an organic substrate is oxidizedwhile another is reduced fermenter organisms are also typi-cally facultative Sulfate is a particularly important alternateelectron acceptor in marine systems because it occurs as thesecond most abundant anion in seawater (behind chloride)typically 28 mM Its use as an electron acceptor leads to theproduction of elemental sulfur or sulfide The use of sulfateas an electron acceptor yields much less energy than oxygenor nitrate and sulfate-reducing bacteria are strict anaerobesunable to use oxygen (often strongly inhibited by oxygen)Methane can be produced by methanogenic archaea whichare all strict anaerobes using CO2 as an electron acceptor(see Findlay and Battin this section for further discussion)
Reduced compounds such as sulfide are produced by theuse of these alternate electron acceptors and they diffuse awayfrom their source When they diffuse to environments with astrong oxidant like oxygen they may be oxidized by chemo-trophic bacteria in energy-yielding reactions Similarlyammonium can be oxidized to nitrite and then nitrate by che-motrophic nitrifying bacteria and the nitrate can diffuse intoanaerobic layers and become denitrified to N2 Such coupledoxidation-reduction reactions can lead to accelerated biogeo-chemical cycling particularly in sediments
It was originally believed that relatively few free-livingspecies of protozoa inhabit anaerobic and hypoxic marineenvironments This view has changed based on (largely)molecular analyses of some of these environments (308348ndash351) A significant number of previously undescribedeukaryotic phylotypes have been observed in anoxic environ-ments High protozoan biodiversity in the absence of molec-ular oxygen is not completely unexpected because otheranaerobic environments such as some gut environments(eg cockroach guts and cattle rumen) have notoriouslyabundant and diverse protozoan fauna but marine anaerobicenvironments have been difficult to access and sample prop-erly Many anaerobic protozoa exist together with symbioticbacteria (352 353) or possess unique organelles such as thehydrogenosome (354)
MAJOR BIOTIC CONTROLSThe Trophic Activities of Protozoa Predation andTop-Down ControlsPredator-prey relationships are fundamental interactionswithin biological communities linking production of bio-mass on the one hand with removal on the other hand Estab-lishing the trophic relationships among microorganisms andquantifying the rates at which they take place constitutemajor endeavors within marine microbial ecology As notedpreviously protozoa consume a wide variety of prokaryoticand protistan prey Numerous laboratory and field measure-ments have been made throughout the past quarter centuryto attempt to quantify the rates of bacterial and phytoplank-ton consumption The methodology for accomplishing thesestudies has varied and a number of approaches are presentlyemployed
Methodologies for estimating microbial grazing are diversebut generally fall into one of two experimental approachesperturbation experiments or tracer experiments Perturbationexperiments (size fractionation metabolic inhibitors anddilution technique) rely on manipulation of the consumeror prey assemblages in somemanner such that the effect is evi-dent from changes in the abundance of the prey population
during incubations relative to control treatments that receiveno manipulation Tracer experiments (radioactively labeledprey fluorescently labeled prey stable isotopendashlabeled pray)attempt to use labeled prey or a prey proxy to quantitativelyfollow the movement of prey biomass into consumers Forsummaries of these methods see (74 355)
Perturbation experiments have entailed a variety of mech-anisms to ldquodecouplerdquo predators from prey in natural microbialcommunities For example size fractionation uses various sizefilters to remove an entire size class of grazers from awater sam-ple leaving the prey to grow in the relative absence of thosepredators during subsequent incubations Unfiltered samples(controls) indicate the growth of prey in the presence of pre-dation Predation impact is obtained by the difference ingrowth of the prey between these treatments This methodhas been used to examine predation on cyanobacteria heter-otrophic bacteria and minute phototrophic eukaryotesAlternatively metabolic inhibitors have been employed tohalt predation (or in some cases prokaryote growth) insteadof physical removal by filtration
The dilution technique (356) has become a commonmethod for examining grazing mortality of phytoplanktonby microzooplankton (which is comprised predominantly ofprotozoa) This method relies on the dilution of herbivorouszooplankton in a series of subsamples (and concomitantmeasurements of phytoplankton growth in each subsample)to estimate the grazing mortality of the herbivores
Tracer experiments have used both radioactively labeledprey and fluorescently labeled prey to examine predation onbacteria and phytoplankton Radioactive labeling has beenaccomplished using 14C- or 3H-labeled organic compounds(for bacteria) or 14C-bicarbonate (for phytoplankton) Theuse of radioisotope approaches has diminished with improve-ments in the number and types of fluorescent compoundsavailable for this work Fluorescently labeled prey havebeen labeled with a variety of compounds that allow theprey to be observed readily (even inside predators) by epi-fluorescencemicroscopy (357 358) Alternatively a decreasein the total number of fluorescent prey in a sample indicatesthe rate of removal of the prey assemblage
Both perturbation experiments and tracer experimentsdescribed have inherent advantages and disadvantages Forexample perturbation experiments are fairly straightforwardto conduct but they may introduce artifact by removingpotentially important sources of nutrients or organic com-pounds released by predators (359) On the other hand thedilution technique is very labor-intensive but one advantageof this method is that it provides the opportunity to determineboth phytoplankton growth and grazing mortality in a singleprocedure Most tracer experiments are fairly easy to conductand generally do not perturb the food web being studied butfeeding selectivity for or against the tracer (relative to naturalprey) may produce spurious results In addition some of thesemethods provide a measurement of the activity of the entiregrazer assemblage and others are applicable for examiningthe activity of specific taxa For example size fractionationapproaches examine the effect of the removal of an entiresize class of organisms (a community-levelrdquo measurement)whereas fluorescently labeled prey can be used to examinethe ingestion rate of a particular species by observing theuptake of labeled prey into that species (a species-levelmeasurement)
Predation rates of numerous marine protozoa have beenexamined in the laboratory From these experiments a gen-eral knowledge of the rates of consumption by these specieshas been accumulated Feeding in most protozoa shows a
422-20 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
functional response to the availability of prey that is similar tothe response of many organisms (although there are a greatdeal of species-specific differences) That is ingestion rateincreases rapidly as prey abundance increases until a maximalingestion rate is attained (360)Manyof these curves have theform of a Type II functional response curve (361)
Based on the features of these curves it is clear that proto-zoa have the ability to ingest substantial numbers of prokar-yote and eukaryote prey in natural ecosystems Somebacterivorous flagellates can consume bacteria (and cyano-bacteria) at rates up to 100 s per individual per hour whereassome bacterivorous ciliates can consume thousands of bacte-ria per individual per hour Rates of consumption of eukary-otic phytoplankton by herbivorous ciliates and largeflagellates are considerably more variable and highly depend-ent of the feeding mechanism (eg filtration in ciliates versuspseudopodial net in dinoflagellates) and the size of the prey
These lab studies demonstrate that rarely are prey suffi-ciently abundant in nature to support the maximal ingestionrates of which many protozoa are capable Nevertheless pre-dation rates of protozoa in nature are sufficient to constitute amajor factor controlling the standing stocks of bacteria andphytoplankton (63 180 184 355) For example these sum-maries indicate that on average protozoa consume 20ndash80ofthe standing stock of phytoplankton (measured as chloro-phyll) in the world ocean and 60ndash75 of daily primary pro-duction a departure from the classical view of copepods as theprimary consumers of phytoplankton This large role of her-bivorous protozoa infers a large role for copepods as consumersof planktonic protozoa (185) Similarly the role of protozoaas major consumers of bacterial biomass is indicated by therelative balance between bacterial production on the onehand and bacterivory on the other (Fig 8) The approximateone-to-one balance between these measurements over a largenumber of studies implies that protozoan bacterivory must bea major factor determining the fate of bacteria in marine (andfreshwater) plankton ecosystems
It is interesting to note that some types of bacteria includ-ing marine Bdellovibrio are known to prey on other marine
bacteria (362) At the time of this writing the importanceof bacterial predation by other bacteria within the contextof oceanographic processes is not known and presumed tobe small While Bdellovibrio may be cultivated from seawaterit can also be found among sequences from cold sediments(363) and the water column Bdellovibrio happens to be thebest studied predatory bacterium but there are no doubtothers that we simply have not yet identified (364)
Species-Specific Interactions (MutualismParasitism Commensalisms)Interactions between marine microorganisms such as compe-tition allelopathy and symbiosis are fundamental aspectscontrolling community composition and activity but atpresent most are poorly characterized in natural marine sys-tems Bacteria must compete for dissolved and particulatecompounds with other bacteria andwith protistan taxa whilephagotrophic protists must compete with each other andmetazoa for available prey These interactions manifest them-selves in many forms including true competition for availableresources resource partitioning such that some taxa specializeon the utilization of certain substrates or prey while othersspecialize on different ones (reducing competition) or eventhe cooperative use of compounds (eg one bacteriumrsquoswaste products being utilized as substrate by another bacte-rium) These metabolic consortia are probably common inthe ocean but can be difficult to identify and characterizeConsortia of bacteria working together to perform complexreactions especially in anaerobic systems are well known inmicrobiology
Consortia in which there is a very high degree of integra-tion between species (ie symbiosis) are very common in theocean These associations may be beneficial to both partners(mutualism) beneficial to one part and inconsequential forthe other (commensalism) or beneficial to one part and det-rimental to the other (parasitism) Many highly visible andwell-characterized marine mutualisms exist between chemo-synthetic bacteria and animals such as hydrothermal ventor methane seep tubeworms clams mussels or other inverte-brates (365) and also between luminescent bacteria and fishor squids (366) Similarly many protozoa form mutualisticassociations with photosynthetic protists (367) The mostconspicuous of these associations involved large amoeboidprotozoa of oceanic ecosystems (planktonic foraminiferanspolycystine radiolaria and acantharia) which contain largenumbers of intracellular algae (74) but associations betweensome heterotrophic dinoflagellates and cyanobacteria are alsocommon (368) Studies have indicated that efficient nutrientcycling in these protozoan-algal associations confers an eco-logical advantage for both partners in surfacewaters of the oli-gotrophic ocean
Parasitism is also a common symbiotic relationship amongmarine protozoa Protozoan parasites have been reported for adiverse array of protists and animals in the ocean Heterotro-phic dinoflagellates and presently uncultured alveolate cladesrelated to alveolates appear to be particularly successful atevolving thismode of existence (369ndash371) In a common par-asitic group Amoebophrya spp the parasite invades photo-synthetic dinoflagellates where it undergoes reproductioneventually lysing the host and releasing dozens of infectiveswarmers The parasitic protist Pirsonia spp is a parasite ofdiatoms (372) The overall importance of these parasitismsin the population dynamics of their hosts is not clearly knownbut may constitute a significant source of mortality for somespecies (373 374) The frequency and diversity of ldquonewrdquo
FIGURE 8 Meta-analysis of experimental studies that have com-pared rates of bacterial productivity (converted to population growthrate units of dminus1) with rates of bacterivory by phagotrophic protists(expressed as equivalent units of bacterial growth rate dminus1) Thesolid line indicates where measurements of bacterial removal by graz-ers were equal tomeasurements of bacterial production Data redrawnfrom Sanders et al (63) doi1011289781555818821ch422f8
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-21
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
classic paper by Eppley (58) and later Goldman and Carpen-ter (289) Temperature was shown to exert a strong and directeffect on themaximal growth rates of these species withmax-imal intrinsic growth rates at 0degC generally lt1 divisiondaywhile growth rates at 30degC may be gt4day These relation-ships indicate the maximal rate that might be attained byphytoplankton at these temperatures but they do not takeother factors into account (nutrients light) While warmtemperature ostensibly allows more rapid growth it createshydrographic conditions that typically give rise to nutrientlimitation of algal growth Thus some of the coldest watersin nature witness some of the most massive phytoplanktonblooms (290) while warm oceanic gyres represent some ofthe most oligotrophic areas of the ocean The multiple andoften conflicting effects of temperature on the growth of nat-ural phytoplankton assemblages limit the accuracy of presentpredictions regarding how primary producers will respond toclimate change (291)
An analysis of the effect of temperature on the growth ofheterotrophic protists has indicated that the response is qual-itatively similar to that of phytoplankton but with a twistThe growth rates of at least some protozoa can exceed thoseof phytoplankton at warmer environmental temperaturesbut the opposite effect is apparent at very low environmentaltemperature This differential effect of temperature on thegrowth of phototrophic and heterotrophic protists was basedon a large meta-analysis of published protistan growth (59)That analysis demonstrated that the maximal growth ratesattained by phototrophic protists could exceed the maximalgrowth rates attained by heterotrophic protists (all otherpotential growth-limiting factors not considered) Thereforeprotozoan growth rates may be constrained to a greater degreeat low environmental temperature than rates for phytoplank-ton If so then phytoplankton bloomsmay get a head start ongrazers during spring in polar ecosystems This scenario is con-sistent with information on seasonal biomass changes andmicrozooplankton herbivory in the Ross Sea Antarctica(292 293) but there are still too few data to fully vet thishypothesis
The importance of high pressure on bacterial growthgained considerable attention in the late 1960s when thedeep sea submersible Alvin was accidentally lost overboardwith its hatch openWhile no lives were lost in this accidentsome workmenrsquos lunches sank to the bottom (sim1500 m)inside the submersible Alvin was recovered after 10 monthsand interestingly there was a waterlogged lunch containingapples bologna sandwiches and broken vacuum bottleswith broth that all appeared hardly degraded and tasted palat-able Yet when placed in a refrigerator on the ship these itemsdegraded relatively quickly Initially pressure was thought tobe the preserving factor as the sea floor temperature was sim-ilar to the refrigerator temperature and the only major differ-ence would be pressure (294) Following this observationa series of experiments to measure degradation of variousorganic materials left in the deep sea for extended periodsindicated that degradationwas typically significantly reducedimplying that pressure reduces the degradation rates (295)Nonetheless changes in the protozoan community of naturaldetrital material sinking to the deep ocean floor indicate thatthe microbial community can respond relatively quickly insome situations (296) Deep sea microbiology has advancedconsiderably yet it is still difficult to interpret results withrespect to actual in situ rates of naturally occurring organicmatter Deep sea bacteria adapted to high pressures havebeen isolated that are barophilic (also called piezophilic)meaning that they prefer high pressures and have reduced
activity at lower pressures whereas others are barotolerant(piezotolerant) tolerating but not preferring high pressuresfor example see (297 298)
Few data are available on barotolerantbarophilic marineprotozoa Protozoa certainly exist and grow at great oceanicdepths but measurements of in situ growth rates for these spe-cies do not yet exist Measurable protozoan numbers havebeen documented in the deep-sea sediments for more than30 years (299 300) and viable protozoa have occasionallybeen cultured from these environments (301ndash305) butvery few direct measurements of the activities of these speciesin situ have been reported (306) A few protozoa have beenisolated that will grow at high pressure (302 303 307) andprotists that appear to be unique to the deep ocean havebeen observed either directly or through the analysis ofDNA sequences (308ndash311) but possibly the best direct evi-dence that protozoan activity takes place at the high pressurescharacteristics of the deep sea are experimental and observa-tional work noting the stimulatory effect that detrital deposi-tion has on some components of the protozoan community(296 312) These observations indicate a diverse and activeprotozoan fauna of the deep ocean although their biogeo-chemical significance is largely uncharacterized at this time
Dissolved and Particulate Organic MatterBacteria and archaea are thought to be by far the most impor-tant organisms with respect to the processing of dissolvedorganic matter (DOM) and nonliving particulate organicmatter (POM also called detritus) in the ocean While theremay be some uptake of DOM by protists particularly forgrowth factors needed in trace amounts such as vitamins(313) the bulk of this material is probably utilized by bacteriaand archaea (314) including the smallest cyanobacteriumProchlorococcus (315 316) which thus may be considered amixotroph Due to their small size bacteria have extremelyhigh surfacevolume ratios and combined with their over-whelmingly high abundance an extremely high integratedsurface area Protozoa tend to obtain the organic materialsthat they require for growth from their prey rather thanthrough the uptake of DOM Overall protozoa tend to besources of dissolved and detrital organic substances throughthe excretion of unassimilated prey biomass in expelledfood vacuoles
Particulate organic matter is not directly available as sub-strate to bacteria These substances must first be reduced tosmall molecules that can be transported into the cell Thisis accomplished by the production of extracellular enzymes(note that few if any large polymers are directly taken up bybacteria with the possible exception of DNA) Hydrolyticenzymes produced by bacteria (and almost certainly archaea)break down polymers like proteins polysaccharides andnucleic acids As with DOM POM is composed of a complexmixture of compounds that vary in their susceptibility to bac-terial degradation and utilization
Particulate material in the water column serves not only asbacterial substrate but also as substratum POM occurs in thewater column across a huge size spectrum from micrometersup to some detrital aggregates more than 1 m in diameter(317) Much of this particulate material is in a constant stateof flux with colloidal material constantly coalescing andaggregating to form new or larger particles (318) as microbialdegradation acts simultaneously to remineralize this materialDetrital particles that attain macroscopic size either by directformation (317 319 320) or via accretion and aggregation(321) and are often called marine snow or macroaggregatesMarine snow particles are readily colonized by bacteria and
422-18 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
many other microorganisms and their abundances on theseaggregates can be orders of magnitude greater than abundan-ces in an equivalent volume of the water surrounding theaggregates (320 322ndash324) and their diversity and highlyconcentrated abundances on detrital aggregates implies thatthere may be stronger trophic coupling and efficient energyelemental cycling relative to these processes among free-living microorganisms in the surrounding waters This specu-lation has resulted in a number of formalizations of the poten-tial impact of aggregate microbiology on biogeochemicalprocesses in the ocean (325 326) Most definitions of bio-films include aggregated particulate and these particles areoften studied within the biofilm conceptual model (see chap-ter by Lawrence et al in this section)
Inorganic Nutrients
Macronutrients (N P)Virtually all heterotrophic organisms in the ocean contributeto the pool of available macronutrients via the excretion ofmetabolic wastes and protozoa contribute significantly tothis process Macronutrients are often in excess within proto-zoan prey relative to the consumersrsquo needs because much ofthe prey carbon is respired to produce energy for metabolismand growth ldquoExcessrdquo nutrients are eliminated as eitherorganic compounds or more commonly as ammonium (forN) and phosphate (for P) The importance of the match(or mismatch) between predator and prey stoichiometry hasbeen demonstrated experimentally and repeatedly in marineand freshwater ecosystems and has been elevated to a funda-mental ecological principle governing elemental and energyflow in aquatic food webs (327) Based on this reasoningCaron et al (328) concluded that bacterivorous protozoaplay an important role in nutrient remineralization andrelease in the ocean Bacterial biomass is typically rich in Nand P relative to other organisms so bacterial predators expe-rience large excesses of these elements relative to their growthneedsMoreover bacterivorous protozoa should be dispropor-tionately important in nutrient remineralization amongsingle-celled eukaryotes because as the smallest protozoa inaquatic ecosystems they have high weight-specific metabolicrates (329) Thus both stoichiometric and allometric rela-tionships implicate small bacterivorous protozoa as impor-tant sources of remineralized nutrients in the plankton
The reasoning also holds true for bacterial utilization oforganic compounds and it has led to some interestingfindings and conclusions regarding the role of bacteria innutrient cycling in the ocean Bacteria are important sourcesremineralized nutrients when they are consuming N- orP-rich substrate Contrary to this traditional view howeverheterotrophic bacteria are often strong competitors for inor-ganic nutrients under many situations largely because bacte-rial biomass is richer in N and P than much of the organicmatter they consume in nature so they require additional Nand P to produce their biomass (summarized by [330]) Exper-imental studies have demonstrated the uptake of ammoniumby bacteria other than cyanobacteria using stable isotope 15N(331) and also the short-lived radioisotope 13N (332) oftenfinding that bacteria may be responsible for one third of the Nor P uptake Additionally there is evidence that bacteria insome oceans like the Sargasso Sea are limited by phosphorus(333 334) Growth limitation of bacteria by phosphorusappears to be a common phenomenon of freshwater ecosys-tems (335 336)
N2 fixation is restricted to prokaryotes and in the oceanwater column it was thought for many years to be done
primarily byTrichodesmium a warm-water colonial cyanobac-terium that blooms sporadically andRichelia a cyanobacterialsymbiont that lives within certain diatoms (337) Some uni-cellular cyanobacteria also contribute significantly to globalN2 fixation (338) and molecular biological data suggestthat a variety of other bacteriamdashnot phytoplanktonmdashmayalso be fixing nitrogen in seawater (339) An exciting recentdiscovery in this field is that a bacterial nitrogen fixer origi-nally known only from nifH sequences UCYN-A has hadits genome sequenced by parallel pyrosequencing and wasfound be a cyanobacterium (Candidatus Atelocyanobacteriumthalassa) lacking the apparatus to generate oxygen as well asother pathways (340) Further study showed it to be symbioticwith a marine alga (341)
Micronutrients (Trace Metals Growth Factors)Bacterial growth may often be limited by macronutrients asdescribed and sometimes by trace nutrients particularlyiron that occur at very low concentrations in seawater(342 343) The importance of Fe as an element limiting pri-mary production in some oceanic regions has come underclose scrutiny and extensive experimental investigation inrecent years Many bacteria have special uptake mechanismsto utilize extremely low concentrations of iron includingsiderophores (released compounds that bind iron and arethen specifically taken up) Even if bacteria are capable ofgrowth at low Fe concentrations growth may be less efficientKirchman et al (344) found that Fe limitation stronglyreduced bacterial growth efficiency in a cultured marine bac-terium (ca 50 efficiency Fe replete but lt10 when Fedeplete) Thus the effects may be complex Marine N2 fixa-tion also may be limited by the availability of Fe becausenitrogen fixers have high Fe requirements
Protozoa tend to be a source rather than a sink for Fe in theocean analogous to their role in macronutrient remineraliza-tion Protozoan grazing activity has been shown to releaseiron from bacterial biomass thereby relieving Fe limitationin co-occurring phytoplankton (345) a mechanistic demon-stration of the process using protozoa growing on Fe-repletebacteria However it has also been reported that Fe-limitedbacteria may contain insufficient Fe for the bacterivorous pro-tozoa that consume them thus leading to Fe limitation in theprotozoan as well (346)
OxygenHypoxic and anoxic regions of the ocean are expected to sig-nificantly expand in the coming decades as a consequence ofchanges in ocean stratification resulting from global climatechange (347) As oxygen becomes depleted bacteria havethe capability to utilize a series of alternate electron acceptorsCompositional changes in the bacterial assemblage accom-pany these changes in metabolic activity but are beyondthe scope of this chapter Many of the studies on benthicmicrobiology have focused on redox reactions that occurthere Oxygen is usually consumed by sediment microbialactivity much faster than the rate at which it can be replacedby diffusion so most sediments especially organically richones are anaerobic below a few mm (fine-grained muds) orcm (coarser sediments) from the sedimentwater interfaceexcept where animals ventilate the benthos through tubesWhen oxygen is absent bacteria and archaea use alternateelectron acceptors such as nitrate nitrite oxidized Fe orMn organic matter or sulfate The use of nitrate or nitriteas an electron acceptor with concomitant production of N2
gas (denitrification) results in net loss of biologically availablenitrogen Bacteria that perform this reaction are typically
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-19
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquofacultativerdquo organisms that can switch from oxygen to nitrateor nitrite Organic matter may be reduced by fermentationreactions in which part of an organic substrate is oxidizedwhile another is reduced fermenter organisms are also typi-cally facultative Sulfate is a particularly important alternateelectron acceptor in marine systems because it occurs as thesecond most abundant anion in seawater (behind chloride)typically 28 mM Its use as an electron acceptor leads to theproduction of elemental sulfur or sulfide The use of sulfateas an electron acceptor yields much less energy than oxygenor nitrate and sulfate-reducing bacteria are strict anaerobesunable to use oxygen (often strongly inhibited by oxygen)Methane can be produced by methanogenic archaea whichare all strict anaerobes using CO2 as an electron acceptor(see Findlay and Battin this section for further discussion)
Reduced compounds such as sulfide are produced by theuse of these alternate electron acceptors and they diffuse awayfrom their source When they diffuse to environments with astrong oxidant like oxygen they may be oxidized by chemo-trophic bacteria in energy-yielding reactions Similarlyammonium can be oxidized to nitrite and then nitrate by che-motrophic nitrifying bacteria and the nitrate can diffuse intoanaerobic layers and become denitrified to N2 Such coupledoxidation-reduction reactions can lead to accelerated biogeo-chemical cycling particularly in sediments
It was originally believed that relatively few free-livingspecies of protozoa inhabit anaerobic and hypoxic marineenvironments This view has changed based on (largely)molecular analyses of some of these environments (308348ndash351) A significant number of previously undescribedeukaryotic phylotypes have been observed in anoxic environ-ments High protozoan biodiversity in the absence of molec-ular oxygen is not completely unexpected because otheranaerobic environments such as some gut environments(eg cockroach guts and cattle rumen) have notoriouslyabundant and diverse protozoan fauna but marine anaerobicenvironments have been difficult to access and sample prop-erly Many anaerobic protozoa exist together with symbioticbacteria (352 353) or possess unique organelles such as thehydrogenosome (354)
MAJOR BIOTIC CONTROLSThe Trophic Activities of Protozoa Predation andTop-Down ControlsPredator-prey relationships are fundamental interactionswithin biological communities linking production of bio-mass on the one hand with removal on the other hand Estab-lishing the trophic relationships among microorganisms andquantifying the rates at which they take place constitutemajor endeavors within marine microbial ecology As notedpreviously protozoa consume a wide variety of prokaryoticand protistan prey Numerous laboratory and field measure-ments have been made throughout the past quarter centuryto attempt to quantify the rates of bacterial and phytoplank-ton consumption The methodology for accomplishing thesestudies has varied and a number of approaches are presentlyemployed
Methodologies for estimating microbial grazing are diversebut generally fall into one of two experimental approachesperturbation experiments or tracer experiments Perturbationexperiments (size fractionation metabolic inhibitors anddilution technique) rely on manipulation of the consumeror prey assemblages in somemanner such that the effect is evi-dent from changes in the abundance of the prey population
during incubations relative to control treatments that receiveno manipulation Tracer experiments (radioactively labeledprey fluorescently labeled prey stable isotopendashlabeled pray)attempt to use labeled prey or a prey proxy to quantitativelyfollow the movement of prey biomass into consumers Forsummaries of these methods see (74 355)
Perturbation experiments have entailed a variety of mech-anisms to ldquodecouplerdquo predators from prey in natural microbialcommunities For example size fractionation uses various sizefilters to remove an entire size class of grazers from awater sam-ple leaving the prey to grow in the relative absence of thosepredators during subsequent incubations Unfiltered samples(controls) indicate the growth of prey in the presence of pre-dation Predation impact is obtained by the difference ingrowth of the prey between these treatments This methodhas been used to examine predation on cyanobacteria heter-otrophic bacteria and minute phototrophic eukaryotesAlternatively metabolic inhibitors have been employed tohalt predation (or in some cases prokaryote growth) insteadof physical removal by filtration
The dilution technique (356) has become a commonmethod for examining grazing mortality of phytoplanktonby microzooplankton (which is comprised predominantly ofprotozoa) This method relies on the dilution of herbivorouszooplankton in a series of subsamples (and concomitantmeasurements of phytoplankton growth in each subsample)to estimate the grazing mortality of the herbivores
Tracer experiments have used both radioactively labeledprey and fluorescently labeled prey to examine predation onbacteria and phytoplankton Radioactive labeling has beenaccomplished using 14C- or 3H-labeled organic compounds(for bacteria) or 14C-bicarbonate (for phytoplankton) Theuse of radioisotope approaches has diminished with improve-ments in the number and types of fluorescent compoundsavailable for this work Fluorescently labeled prey havebeen labeled with a variety of compounds that allow theprey to be observed readily (even inside predators) by epi-fluorescencemicroscopy (357 358) Alternatively a decreasein the total number of fluorescent prey in a sample indicatesthe rate of removal of the prey assemblage
Both perturbation experiments and tracer experimentsdescribed have inherent advantages and disadvantages Forexample perturbation experiments are fairly straightforwardto conduct but they may introduce artifact by removingpotentially important sources of nutrients or organic com-pounds released by predators (359) On the other hand thedilution technique is very labor-intensive but one advantageof this method is that it provides the opportunity to determineboth phytoplankton growth and grazing mortality in a singleprocedure Most tracer experiments are fairly easy to conductand generally do not perturb the food web being studied butfeeding selectivity for or against the tracer (relative to naturalprey) may produce spurious results In addition some of thesemethods provide a measurement of the activity of the entiregrazer assemblage and others are applicable for examiningthe activity of specific taxa For example size fractionationapproaches examine the effect of the removal of an entiresize class of organisms (a community-levelrdquo measurement)whereas fluorescently labeled prey can be used to examinethe ingestion rate of a particular species by observing theuptake of labeled prey into that species (a species-levelmeasurement)
Predation rates of numerous marine protozoa have beenexamined in the laboratory From these experiments a gen-eral knowledge of the rates of consumption by these specieshas been accumulated Feeding in most protozoa shows a
422-20 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
functional response to the availability of prey that is similar tothe response of many organisms (although there are a greatdeal of species-specific differences) That is ingestion rateincreases rapidly as prey abundance increases until a maximalingestion rate is attained (360)Manyof these curves have theform of a Type II functional response curve (361)
Based on the features of these curves it is clear that proto-zoa have the ability to ingest substantial numbers of prokar-yote and eukaryote prey in natural ecosystems Somebacterivorous flagellates can consume bacteria (and cyano-bacteria) at rates up to 100 s per individual per hour whereassome bacterivorous ciliates can consume thousands of bacte-ria per individual per hour Rates of consumption of eukary-otic phytoplankton by herbivorous ciliates and largeflagellates are considerably more variable and highly depend-ent of the feeding mechanism (eg filtration in ciliates versuspseudopodial net in dinoflagellates) and the size of the prey
These lab studies demonstrate that rarely are prey suffi-ciently abundant in nature to support the maximal ingestionrates of which many protozoa are capable Nevertheless pre-dation rates of protozoa in nature are sufficient to constitute amajor factor controlling the standing stocks of bacteria andphytoplankton (63 180 184 355) For example these sum-maries indicate that on average protozoa consume 20ndash80ofthe standing stock of phytoplankton (measured as chloro-phyll) in the world ocean and 60ndash75 of daily primary pro-duction a departure from the classical view of copepods as theprimary consumers of phytoplankton This large role of her-bivorous protozoa infers a large role for copepods as consumersof planktonic protozoa (185) Similarly the role of protozoaas major consumers of bacterial biomass is indicated by therelative balance between bacterial production on the onehand and bacterivory on the other (Fig 8) The approximateone-to-one balance between these measurements over a largenumber of studies implies that protozoan bacterivory must bea major factor determining the fate of bacteria in marine (andfreshwater) plankton ecosystems
It is interesting to note that some types of bacteria includ-ing marine Bdellovibrio are known to prey on other marine
bacteria (362) At the time of this writing the importanceof bacterial predation by other bacteria within the contextof oceanographic processes is not known and presumed tobe small While Bdellovibrio may be cultivated from seawaterit can also be found among sequences from cold sediments(363) and the water column Bdellovibrio happens to be thebest studied predatory bacterium but there are no doubtothers that we simply have not yet identified (364)
Species-Specific Interactions (MutualismParasitism Commensalisms)Interactions between marine microorganisms such as compe-tition allelopathy and symbiosis are fundamental aspectscontrolling community composition and activity but atpresent most are poorly characterized in natural marine sys-tems Bacteria must compete for dissolved and particulatecompounds with other bacteria andwith protistan taxa whilephagotrophic protists must compete with each other andmetazoa for available prey These interactions manifest them-selves in many forms including true competition for availableresources resource partitioning such that some taxa specializeon the utilization of certain substrates or prey while othersspecialize on different ones (reducing competition) or eventhe cooperative use of compounds (eg one bacteriumrsquoswaste products being utilized as substrate by another bacte-rium) These metabolic consortia are probably common inthe ocean but can be difficult to identify and characterizeConsortia of bacteria working together to perform complexreactions especially in anaerobic systems are well known inmicrobiology
Consortia in which there is a very high degree of integra-tion between species (ie symbiosis) are very common in theocean These associations may be beneficial to both partners(mutualism) beneficial to one part and inconsequential forthe other (commensalism) or beneficial to one part and det-rimental to the other (parasitism) Many highly visible andwell-characterized marine mutualisms exist between chemo-synthetic bacteria and animals such as hydrothermal ventor methane seep tubeworms clams mussels or other inverte-brates (365) and also between luminescent bacteria and fishor squids (366) Similarly many protozoa form mutualisticassociations with photosynthetic protists (367) The mostconspicuous of these associations involved large amoeboidprotozoa of oceanic ecosystems (planktonic foraminiferanspolycystine radiolaria and acantharia) which contain largenumbers of intracellular algae (74) but associations betweensome heterotrophic dinoflagellates and cyanobacteria are alsocommon (368) Studies have indicated that efficient nutrientcycling in these protozoan-algal associations confers an eco-logical advantage for both partners in surfacewaters of the oli-gotrophic ocean
Parasitism is also a common symbiotic relationship amongmarine protozoa Protozoan parasites have been reported for adiverse array of protists and animals in the ocean Heterotro-phic dinoflagellates and presently uncultured alveolate cladesrelated to alveolates appear to be particularly successful atevolving thismode of existence (369ndash371) In a common par-asitic group Amoebophrya spp the parasite invades photo-synthetic dinoflagellates where it undergoes reproductioneventually lysing the host and releasing dozens of infectiveswarmers The parasitic protist Pirsonia spp is a parasite ofdiatoms (372) The overall importance of these parasitismsin the population dynamics of their hosts is not clearly knownbut may constitute a significant source of mortality for somespecies (373 374) The frequency and diversity of ldquonewrdquo
FIGURE 8 Meta-analysis of experimental studies that have com-pared rates of bacterial productivity (converted to population growthrate units of dminus1) with rates of bacterivory by phagotrophic protists(expressed as equivalent units of bacterial growth rate dminus1) Thesolid line indicates where measurements of bacterial removal by graz-ers were equal tomeasurements of bacterial production Data redrawnfrom Sanders et al (63) doi1011289781555818821ch422f8
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-21
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
many other microorganisms and their abundances on theseaggregates can be orders of magnitude greater than abundan-ces in an equivalent volume of the water surrounding theaggregates (320 322ndash324) and their diversity and highlyconcentrated abundances on detrital aggregates implies thatthere may be stronger trophic coupling and efficient energyelemental cycling relative to these processes among free-living microorganisms in the surrounding waters This specu-lation has resulted in a number of formalizations of the poten-tial impact of aggregate microbiology on biogeochemicalprocesses in the ocean (325 326) Most definitions of bio-films include aggregated particulate and these particles areoften studied within the biofilm conceptual model (see chap-ter by Lawrence et al in this section)
Inorganic Nutrients
Macronutrients (N P)Virtually all heterotrophic organisms in the ocean contributeto the pool of available macronutrients via the excretion ofmetabolic wastes and protozoa contribute significantly tothis process Macronutrients are often in excess within proto-zoan prey relative to the consumersrsquo needs because much ofthe prey carbon is respired to produce energy for metabolismand growth ldquoExcessrdquo nutrients are eliminated as eitherorganic compounds or more commonly as ammonium (forN) and phosphate (for P) The importance of the match(or mismatch) between predator and prey stoichiometry hasbeen demonstrated experimentally and repeatedly in marineand freshwater ecosystems and has been elevated to a funda-mental ecological principle governing elemental and energyflow in aquatic food webs (327) Based on this reasoningCaron et al (328) concluded that bacterivorous protozoaplay an important role in nutrient remineralization andrelease in the ocean Bacterial biomass is typically rich in Nand P relative to other organisms so bacterial predators expe-rience large excesses of these elements relative to their growthneedsMoreover bacterivorous protozoa should be dispropor-tionately important in nutrient remineralization amongsingle-celled eukaryotes because as the smallest protozoa inaquatic ecosystems they have high weight-specific metabolicrates (329) Thus both stoichiometric and allometric rela-tionships implicate small bacterivorous protozoa as impor-tant sources of remineralized nutrients in the plankton
The reasoning also holds true for bacterial utilization oforganic compounds and it has led to some interestingfindings and conclusions regarding the role of bacteria innutrient cycling in the ocean Bacteria are important sourcesremineralized nutrients when they are consuming N- orP-rich substrate Contrary to this traditional view howeverheterotrophic bacteria are often strong competitors for inor-ganic nutrients under many situations largely because bacte-rial biomass is richer in N and P than much of the organicmatter they consume in nature so they require additional Nand P to produce their biomass (summarized by [330]) Exper-imental studies have demonstrated the uptake of ammoniumby bacteria other than cyanobacteria using stable isotope 15N(331) and also the short-lived radioisotope 13N (332) oftenfinding that bacteria may be responsible for one third of the Nor P uptake Additionally there is evidence that bacteria insome oceans like the Sargasso Sea are limited by phosphorus(333 334) Growth limitation of bacteria by phosphorusappears to be a common phenomenon of freshwater ecosys-tems (335 336)
N2 fixation is restricted to prokaryotes and in the oceanwater column it was thought for many years to be done
primarily byTrichodesmium a warm-water colonial cyanobac-terium that blooms sporadically andRichelia a cyanobacterialsymbiont that lives within certain diatoms (337) Some uni-cellular cyanobacteria also contribute significantly to globalN2 fixation (338) and molecular biological data suggestthat a variety of other bacteriamdashnot phytoplanktonmdashmayalso be fixing nitrogen in seawater (339) An exciting recentdiscovery in this field is that a bacterial nitrogen fixer origi-nally known only from nifH sequences UCYN-A has hadits genome sequenced by parallel pyrosequencing and wasfound be a cyanobacterium (Candidatus Atelocyanobacteriumthalassa) lacking the apparatus to generate oxygen as well asother pathways (340) Further study showed it to be symbioticwith a marine alga (341)
Micronutrients (Trace Metals Growth Factors)Bacterial growth may often be limited by macronutrients asdescribed and sometimes by trace nutrients particularlyiron that occur at very low concentrations in seawater(342 343) The importance of Fe as an element limiting pri-mary production in some oceanic regions has come underclose scrutiny and extensive experimental investigation inrecent years Many bacteria have special uptake mechanismsto utilize extremely low concentrations of iron includingsiderophores (released compounds that bind iron and arethen specifically taken up) Even if bacteria are capable ofgrowth at low Fe concentrations growth may be less efficientKirchman et al (344) found that Fe limitation stronglyreduced bacterial growth efficiency in a cultured marine bac-terium (ca 50 efficiency Fe replete but lt10 when Fedeplete) Thus the effects may be complex Marine N2 fixa-tion also may be limited by the availability of Fe becausenitrogen fixers have high Fe requirements
Protozoa tend to be a source rather than a sink for Fe in theocean analogous to their role in macronutrient remineraliza-tion Protozoan grazing activity has been shown to releaseiron from bacterial biomass thereby relieving Fe limitationin co-occurring phytoplankton (345) a mechanistic demon-stration of the process using protozoa growing on Fe-repletebacteria However it has also been reported that Fe-limitedbacteria may contain insufficient Fe for the bacterivorous pro-tozoa that consume them thus leading to Fe limitation in theprotozoan as well (346)
OxygenHypoxic and anoxic regions of the ocean are expected to sig-nificantly expand in the coming decades as a consequence ofchanges in ocean stratification resulting from global climatechange (347) As oxygen becomes depleted bacteria havethe capability to utilize a series of alternate electron acceptorsCompositional changes in the bacterial assemblage accom-pany these changes in metabolic activity but are beyondthe scope of this chapter Many of the studies on benthicmicrobiology have focused on redox reactions that occurthere Oxygen is usually consumed by sediment microbialactivity much faster than the rate at which it can be replacedby diffusion so most sediments especially organically richones are anaerobic below a few mm (fine-grained muds) orcm (coarser sediments) from the sedimentwater interfaceexcept where animals ventilate the benthos through tubesWhen oxygen is absent bacteria and archaea use alternateelectron acceptors such as nitrate nitrite oxidized Fe orMn organic matter or sulfate The use of nitrate or nitriteas an electron acceptor with concomitant production of N2
gas (denitrification) results in net loss of biologically availablenitrogen Bacteria that perform this reaction are typically
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-19
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquofacultativerdquo organisms that can switch from oxygen to nitrateor nitrite Organic matter may be reduced by fermentationreactions in which part of an organic substrate is oxidizedwhile another is reduced fermenter organisms are also typi-cally facultative Sulfate is a particularly important alternateelectron acceptor in marine systems because it occurs as thesecond most abundant anion in seawater (behind chloride)typically 28 mM Its use as an electron acceptor leads to theproduction of elemental sulfur or sulfide The use of sulfateas an electron acceptor yields much less energy than oxygenor nitrate and sulfate-reducing bacteria are strict anaerobesunable to use oxygen (often strongly inhibited by oxygen)Methane can be produced by methanogenic archaea whichare all strict anaerobes using CO2 as an electron acceptor(see Findlay and Battin this section for further discussion)
Reduced compounds such as sulfide are produced by theuse of these alternate electron acceptors and they diffuse awayfrom their source When they diffuse to environments with astrong oxidant like oxygen they may be oxidized by chemo-trophic bacteria in energy-yielding reactions Similarlyammonium can be oxidized to nitrite and then nitrate by che-motrophic nitrifying bacteria and the nitrate can diffuse intoanaerobic layers and become denitrified to N2 Such coupledoxidation-reduction reactions can lead to accelerated biogeo-chemical cycling particularly in sediments
It was originally believed that relatively few free-livingspecies of protozoa inhabit anaerobic and hypoxic marineenvironments This view has changed based on (largely)molecular analyses of some of these environments (308348ndash351) A significant number of previously undescribedeukaryotic phylotypes have been observed in anoxic environ-ments High protozoan biodiversity in the absence of molec-ular oxygen is not completely unexpected because otheranaerobic environments such as some gut environments(eg cockroach guts and cattle rumen) have notoriouslyabundant and diverse protozoan fauna but marine anaerobicenvironments have been difficult to access and sample prop-erly Many anaerobic protozoa exist together with symbioticbacteria (352 353) or possess unique organelles such as thehydrogenosome (354)
MAJOR BIOTIC CONTROLSThe Trophic Activities of Protozoa Predation andTop-Down ControlsPredator-prey relationships are fundamental interactionswithin biological communities linking production of bio-mass on the one hand with removal on the other hand Estab-lishing the trophic relationships among microorganisms andquantifying the rates at which they take place constitutemajor endeavors within marine microbial ecology As notedpreviously protozoa consume a wide variety of prokaryoticand protistan prey Numerous laboratory and field measure-ments have been made throughout the past quarter centuryto attempt to quantify the rates of bacterial and phytoplank-ton consumption The methodology for accomplishing thesestudies has varied and a number of approaches are presentlyemployed
Methodologies for estimating microbial grazing are diversebut generally fall into one of two experimental approachesperturbation experiments or tracer experiments Perturbationexperiments (size fractionation metabolic inhibitors anddilution technique) rely on manipulation of the consumeror prey assemblages in somemanner such that the effect is evi-dent from changes in the abundance of the prey population
during incubations relative to control treatments that receiveno manipulation Tracer experiments (radioactively labeledprey fluorescently labeled prey stable isotopendashlabeled pray)attempt to use labeled prey or a prey proxy to quantitativelyfollow the movement of prey biomass into consumers Forsummaries of these methods see (74 355)
Perturbation experiments have entailed a variety of mech-anisms to ldquodecouplerdquo predators from prey in natural microbialcommunities For example size fractionation uses various sizefilters to remove an entire size class of grazers from awater sam-ple leaving the prey to grow in the relative absence of thosepredators during subsequent incubations Unfiltered samples(controls) indicate the growth of prey in the presence of pre-dation Predation impact is obtained by the difference ingrowth of the prey between these treatments This methodhas been used to examine predation on cyanobacteria heter-otrophic bacteria and minute phototrophic eukaryotesAlternatively metabolic inhibitors have been employed tohalt predation (or in some cases prokaryote growth) insteadof physical removal by filtration
The dilution technique (356) has become a commonmethod for examining grazing mortality of phytoplanktonby microzooplankton (which is comprised predominantly ofprotozoa) This method relies on the dilution of herbivorouszooplankton in a series of subsamples (and concomitantmeasurements of phytoplankton growth in each subsample)to estimate the grazing mortality of the herbivores
Tracer experiments have used both radioactively labeledprey and fluorescently labeled prey to examine predation onbacteria and phytoplankton Radioactive labeling has beenaccomplished using 14C- or 3H-labeled organic compounds(for bacteria) or 14C-bicarbonate (for phytoplankton) Theuse of radioisotope approaches has diminished with improve-ments in the number and types of fluorescent compoundsavailable for this work Fluorescently labeled prey havebeen labeled with a variety of compounds that allow theprey to be observed readily (even inside predators) by epi-fluorescencemicroscopy (357 358) Alternatively a decreasein the total number of fluorescent prey in a sample indicatesthe rate of removal of the prey assemblage
Both perturbation experiments and tracer experimentsdescribed have inherent advantages and disadvantages Forexample perturbation experiments are fairly straightforwardto conduct but they may introduce artifact by removingpotentially important sources of nutrients or organic com-pounds released by predators (359) On the other hand thedilution technique is very labor-intensive but one advantageof this method is that it provides the opportunity to determineboth phytoplankton growth and grazing mortality in a singleprocedure Most tracer experiments are fairly easy to conductand generally do not perturb the food web being studied butfeeding selectivity for or against the tracer (relative to naturalprey) may produce spurious results In addition some of thesemethods provide a measurement of the activity of the entiregrazer assemblage and others are applicable for examiningthe activity of specific taxa For example size fractionationapproaches examine the effect of the removal of an entiresize class of organisms (a community-levelrdquo measurement)whereas fluorescently labeled prey can be used to examinethe ingestion rate of a particular species by observing theuptake of labeled prey into that species (a species-levelmeasurement)
Predation rates of numerous marine protozoa have beenexamined in the laboratory From these experiments a gen-eral knowledge of the rates of consumption by these specieshas been accumulated Feeding in most protozoa shows a
422-20 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
functional response to the availability of prey that is similar tothe response of many organisms (although there are a greatdeal of species-specific differences) That is ingestion rateincreases rapidly as prey abundance increases until a maximalingestion rate is attained (360)Manyof these curves have theform of a Type II functional response curve (361)
Based on the features of these curves it is clear that proto-zoa have the ability to ingest substantial numbers of prokar-yote and eukaryote prey in natural ecosystems Somebacterivorous flagellates can consume bacteria (and cyano-bacteria) at rates up to 100 s per individual per hour whereassome bacterivorous ciliates can consume thousands of bacte-ria per individual per hour Rates of consumption of eukary-otic phytoplankton by herbivorous ciliates and largeflagellates are considerably more variable and highly depend-ent of the feeding mechanism (eg filtration in ciliates versuspseudopodial net in dinoflagellates) and the size of the prey
These lab studies demonstrate that rarely are prey suffi-ciently abundant in nature to support the maximal ingestionrates of which many protozoa are capable Nevertheless pre-dation rates of protozoa in nature are sufficient to constitute amajor factor controlling the standing stocks of bacteria andphytoplankton (63 180 184 355) For example these sum-maries indicate that on average protozoa consume 20ndash80ofthe standing stock of phytoplankton (measured as chloro-phyll) in the world ocean and 60ndash75 of daily primary pro-duction a departure from the classical view of copepods as theprimary consumers of phytoplankton This large role of her-bivorous protozoa infers a large role for copepods as consumersof planktonic protozoa (185) Similarly the role of protozoaas major consumers of bacterial biomass is indicated by therelative balance between bacterial production on the onehand and bacterivory on the other (Fig 8) The approximateone-to-one balance between these measurements over a largenumber of studies implies that protozoan bacterivory must bea major factor determining the fate of bacteria in marine (andfreshwater) plankton ecosystems
It is interesting to note that some types of bacteria includ-ing marine Bdellovibrio are known to prey on other marine
bacteria (362) At the time of this writing the importanceof bacterial predation by other bacteria within the contextof oceanographic processes is not known and presumed tobe small While Bdellovibrio may be cultivated from seawaterit can also be found among sequences from cold sediments(363) and the water column Bdellovibrio happens to be thebest studied predatory bacterium but there are no doubtothers that we simply have not yet identified (364)
Species-Specific Interactions (MutualismParasitism Commensalisms)Interactions between marine microorganisms such as compe-tition allelopathy and symbiosis are fundamental aspectscontrolling community composition and activity but atpresent most are poorly characterized in natural marine sys-tems Bacteria must compete for dissolved and particulatecompounds with other bacteria andwith protistan taxa whilephagotrophic protists must compete with each other andmetazoa for available prey These interactions manifest them-selves in many forms including true competition for availableresources resource partitioning such that some taxa specializeon the utilization of certain substrates or prey while othersspecialize on different ones (reducing competition) or eventhe cooperative use of compounds (eg one bacteriumrsquoswaste products being utilized as substrate by another bacte-rium) These metabolic consortia are probably common inthe ocean but can be difficult to identify and characterizeConsortia of bacteria working together to perform complexreactions especially in anaerobic systems are well known inmicrobiology
Consortia in which there is a very high degree of integra-tion between species (ie symbiosis) are very common in theocean These associations may be beneficial to both partners(mutualism) beneficial to one part and inconsequential forthe other (commensalism) or beneficial to one part and det-rimental to the other (parasitism) Many highly visible andwell-characterized marine mutualisms exist between chemo-synthetic bacteria and animals such as hydrothermal ventor methane seep tubeworms clams mussels or other inverte-brates (365) and also between luminescent bacteria and fishor squids (366) Similarly many protozoa form mutualisticassociations with photosynthetic protists (367) The mostconspicuous of these associations involved large amoeboidprotozoa of oceanic ecosystems (planktonic foraminiferanspolycystine radiolaria and acantharia) which contain largenumbers of intracellular algae (74) but associations betweensome heterotrophic dinoflagellates and cyanobacteria are alsocommon (368) Studies have indicated that efficient nutrientcycling in these protozoan-algal associations confers an eco-logical advantage for both partners in surfacewaters of the oli-gotrophic ocean
Parasitism is also a common symbiotic relationship amongmarine protozoa Protozoan parasites have been reported for adiverse array of protists and animals in the ocean Heterotro-phic dinoflagellates and presently uncultured alveolate cladesrelated to alveolates appear to be particularly successful atevolving thismode of existence (369ndash371) In a common par-asitic group Amoebophrya spp the parasite invades photo-synthetic dinoflagellates where it undergoes reproductioneventually lysing the host and releasing dozens of infectiveswarmers The parasitic protist Pirsonia spp is a parasite ofdiatoms (372) The overall importance of these parasitismsin the population dynamics of their hosts is not clearly knownbut may constitute a significant source of mortality for somespecies (373 374) The frequency and diversity of ldquonewrdquo
FIGURE 8 Meta-analysis of experimental studies that have com-pared rates of bacterial productivity (converted to population growthrate units of dminus1) with rates of bacterivory by phagotrophic protists(expressed as equivalent units of bacterial growth rate dminus1) Thesolid line indicates where measurements of bacterial removal by graz-ers were equal tomeasurements of bacterial production Data redrawnfrom Sanders et al (63) doi1011289781555818821ch422f8
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-21
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
ldquofacultativerdquo organisms that can switch from oxygen to nitrateor nitrite Organic matter may be reduced by fermentationreactions in which part of an organic substrate is oxidizedwhile another is reduced fermenter organisms are also typi-cally facultative Sulfate is a particularly important alternateelectron acceptor in marine systems because it occurs as thesecond most abundant anion in seawater (behind chloride)typically 28 mM Its use as an electron acceptor leads to theproduction of elemental sulfur or sulfide The use of sulfateas an electron acceptor yields much less energy than oxygenor nitrate and sulfate-reducing bacteria are strict anaerobesunable to use oxygen (often strongly inhibited by oxygen)Methane can be produced by methanogenic archaea whichare all strict anaerobes using CO2 as an electron acceptor(see Findlay and Battin this section for further discussion)
Reduced compounds such as sulfide are produced by theuse of these alternate electron acceptors and they diffuse awayfrom their source When they diffuse to environments with astrong oxidant like oxygen they may be oxidized by chemo-trophic bacteria in energy-yielding reactions Similarlyammonium can be oxidized to nitrite and then nitrate by che-motrophic nitrifying bacteria and the nitrate can diffuse intoanaerobic layers and become denitrified to N2 Such coupledoxidation-reduction reactions can lead to accelerated biogeo-chemical cycling particularly in sediments
It was originally believed that relatively few free-livingspecies of protozoa inhabit anaerobic and hypoxic marineenvironments This view has changed based on (largely)molecular analyses of some of these environments (308348ndash351) A significant number of previously undescribedeukaryotic phylotypes have been observed in anoxic environ-ments High protozoan biodiversity in the absence of molec-ular oxygen is not completely unexpected because otheranaerobic environments such as some gut environments(eg cockroach guts and cattle rumen) have notoriouslyabundant and diverse protozoan fauna but marine anaerobicenvironments have been difficult to access and sample prop-erly Many anaerobic protozoa exist together with symbioticbacteria (352 353) or possess unique organelles such as thehydrogenosome (354)
MAJOR BIOTIC CONTROLSThe Trophic Activities of Protozoa Predation andTop-Down ControlsPredator-prey relationships are fundamental interactionswithin biological communities linking production of bio-mass on the one hand with removal on the other hand Estab-lishing the trophic relationships among microorganisms andquantifying the rates at which they take place constitutemajor endeavors within marine microbial ecology As notedpreviously protozoa consume a wide variety of prokaryoticand protistan prey Numerous laboratory and field measure-ments have been made throughout the past quarter centuryto attempt to quantify the rates of bacterial and phytoplank-ton consumption The methodology for accomplishing thesestudies has varied and a number of approaches are presentlyemployed
Methodologies for estimating microbial grazing are diversebut generally fall into one of two experimental approachesperturbation experiments or tracer experiments Perturbationexperiments (size fractionation metabolic inhibitors anddilution technique) rely on manipulation of the consumeror prey assemblages in somemanner such that the effect is evi-dent from changes in the abundance of the prey population
during incubations relative to control treatments that receiveno manipulation Tracer experiments (radioactively labeledprey fluorescently labeled prey stable isotopendashlabeled pray)attempt to use labeled prey or a prey proxy to quantitativelyfollow the movement of prey biomass into consumers Forsummaries of these methods see (74 355)
Perturbation experiments have entailed a variety of mech-anisms to ldquodecouplerdquo predators from prey in natural microbialcommunities For example size fractionation uses various sizefilters to remove an entire size class of grazers from awater sam-ple leaving the prey to grow in the relative absence of thosepredators during subsequent incubations Unfiltered samples(controls) indicate the growth of prey in the presence of pre-dation Predation impact is obtained by the difference ingrowth of the prey between these treatments This methodhas been used to examine predation on cyanobacteria heter-otrophic bacteria and minute phototrophic eukaryotesAlternatively metabolic inhibitors have been employed tohalt predation (or in some cases prokaryote growth) insteadof physical removal by filtration
The dilution technique (356) has become a commonmethod for examining grazing mortality of phytoplanktonby microzooplankton (which is comprised predominantly ofprotozoa) This method relies on the dilution of herbivorouszooplankton in a series of subsamples (and concomitantmeasurements of phytoplankton growth in each subsample)to estimate the grazing mortality of the herbivores
Tracer experiments have used both radioactively labeledprey and fluorescently labeled prey to examine predation onbacteria and phytoplankton Radioactive labeling has beenaccomplished using 14C- or 3H-labeled organic compounds(for bacteria) or 14C-bicarbonate (for phytoplankton) Theuse of radioisotope approaches has diminished with improve-ments in the number and types of fluorescent compoundsavailable for this work Fluorescently labeled prey havebeen labeled with a variety of compounds that allow theprey to be observed readily (even inside predators) by epi-fluorescencemicroscopy (357 358) Alternatively a decreasein the total number of fluorescent prey in a sample indicatesthe rate of removal of the prey assemblage
Both perturbation experiments and tracer experimentsdescribed have inherent advantages and disadvantages Forexample perturbation experiments are fairly straightforwardto conduct but they may introduce artifact by removingpotentially important sources of nutrients or organic com-pounds released by predators (359) On the other hand thedilution technique is very labor-intensive but one advantageof this method is that it provides the opportunity to determineboth phytoplankton growth and grazing mortality in a singleprocedure Most tracer experiments are fairly easy to conductand generally do not perturb the food web being studied butfeeding selectivity for or against the tracer (relative to naturalprey) may produce spurious results In addition some of thesemethods provide a measurement of the activity of the entiregrazer assemblage and others are applicable for examiningthe activity of specific taxa For example size fractionationapproaches examine the effect of the removal of an entiresize class of organisms (a community-levelrdquo measurement)whereas fluorescently labeled prey can be used to examinethe ingestion rate of a particular species by observing theuptake of labeled prey into that species (a species-levelmeasurement)
Predation rates of numerous marine protozoa have beenexamined in the laboratory From these experiments a gen-eral knowledge of the rates of consumption by these specieshas been accumulated Feeding in most protozoa shows a
422-20 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
functional response to the availability of prey that is similar tothe response of many organisms (although there are a greatdeal of species-specific differences) That is ingestion rateincreases rapidly as prey abundance increases until a maximalingestion rate is attained (360)Manyof these curves have theform of a Type II functional response curve (361)
Based on the features of these curves it is clear that proto-zoa have the ability to ingest substantial numbers of prokar-yote and eukaryote prey in natural ecosystems Somebacterivorous flagellates can consume bacteria (and cyano-bacteria) at rates up to 100 s per individual per hour whereassome bacterivorous ciliates can consume thousands of bacte-ria per individual per hour Rates of consumption of eukary-otic phytoplankton by herbivorous ciliates and largeflagellates are considerably more variable and highly depend-ent of the feeding mechanism (eg filtration in ciliates versuspseudopodial net in dinoflagellates) and the size of the prey
These lab studies demonstrate that rarely are prey suffi-ciently abundant in nature to support the maximal ingestionrates of which many protozoa are capable Nevertheless pre-dation rates of protozoa in nature are sufficient to constitute amajor factor controlling the standing stocks of bacteria andphytoplankton (63 180 184 355) For example these sum-maries indicate that on average protozoa consume 20ndash80ofthe standing stock of phytoplankton (measured as chloro-phyll) in the world ocean and 60ndash75 of daily primary pro-duction a departure from the classical view of copepods as theprimary consumers of phytoplankton This large role of her-bivorous protozoa infers a large role for copepods as consumersof planktonic protozoa (185) Similarly the role of protozoaas major consumers of bacterial biomass is indicated by therelative balance between bacterial production on the onehand and bacterivory on the other (Fig 8) The approximateone-to-one balance between these measurements over a largenumber of studies implies that protozoan bacterivory must bea major factor determining the fate of bacteria in marine (andfreshwater) plankton ecosystems
It is interesting to note that some types of bacteria includ-ing marine Bdellovibrio are known to prey on other marine
bacteria (362) At the time of this writing the importanceof bacterial predation by other bacteria within the contextof oceanographic processes is not known and presumed tobe small While Bdellovibrio may be cultivated from seawaterit can also be found among sequences from cold sediments(363) and the water column Bdellovibrio happens to be thebest studied predatory bacterium but there are no doubtothers that we simply have not yet identified (364)
Species-Specific Interactions (MutualismParasitism Commensalisms)Interactions between marine microorganisms such as compe-tition allelopathy and symbiosis are fundamental aspectscontrolling community composition and activity but atpresent most are poorly characterized in natural marine sys-tems Bacteria must compete for dissolved and particulatecompounds with other bacteria andwith protistan taxa whilephagotrophic protists must compete with each other andmetazoa for available prey These interactions manifest them-selves in many forms including true competition for availableresources resource partitioning such that some taxa specializeon the utilization of certain substrates or prey while othersspecialize on different ones (reducing competition) or eventhe cooperative use of compounds (eg one bacteriumrsquoswaste products being utilized as substrate by another bacte-rium) These metabolic consortia are probably common inthe ocean but can be difficult to identify and characterizeConsortia of bacteria working together to perform complexreactions especially in anaerobic systems are well known inmicrobiology
Consortia in which there is a very high degree of integra-tion between species (ie symbiosis) are very common in theocean These associations may be beneficial to both partners(mutualism) beneficial to one part and inconsequential forthe other (commensalism) or beneficial to one part and det-rimental to the other (parasitism) Many highly visible andwell-characterized marine mutualisms exist between chemo-synthetic bacteria and animals such as hydrothermal ventor methane seep tubeworms clams mussels or other inverte-brates (365) and also between luminescent bacteria and fishor squids (366) Similarly many protozoa form mutualisticassociations with photosynthetic protists (367) The mostconspicuous of these associations involved large amoeboidprotozoa of oceanic ecosystems (planktonic foraminiferanspolycystine radiolaria and acantharia) which contain largenumbers of intracellular algae (74) but associations betweensome heterotrophic dinoflagellates and cyanobacteria are alsocommon (368) Studies have indicated that efficient nutrientcycling in these protozoan-algal associations confers an eco-logical advantage for both partners in surfacewaters of the oli-gotrophic ocean
Parasitism is also a common symbiotic relationship amongmarine protozoa Protozoan parasites have been reported for adiverse array of protists and animals in the ocean Heterotro-phic dinoflagellates and presently uncultured alveolate cladesrelated to alveolates appear to be particularly successful atevolving thismode of existence (369ndash371) In a common par-asitic group Amoebophrya spp the parasite invades photo-synthetic dinoflagellates where it undergoes reproductioneventually lysing the host and releasing dozens of infectiveswarmers The parasitic protist Pirsonia spp is a parasite ofdiatoms (372) The overall importance of these parasitismsin the population dynamics of their hosts is not clearly knownbut may constitute a significant source of mortality for somespecies (373 374) The frequency and diversity of ldquonewrdquo
FIGURE 8 Meta-analysis of experimental studies that have com-pared rates of bacterial productivity (converted to population growthrate units of dminus1) with rates of bacterivory by phagotrophic protists(expressed as equivalent units of bacterial growth rate dminus1) Thesolid line indicates where measurements of bacterial removal by graz-ers were equal tomeasurements of bacterial production Data redrawnfrom Sanders et al (63) doi1011289781555818821ch422f8
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-21
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
functional response to the availability of prey that is similar tothe response of many organisms (although there are a greatdeal of species-specific differences) That is ingestion rateincreases rapidly as prey abundance increases until a maximalingestion rate is attained (360)Manyof these curves have theform of a Type II functional response curve (361)
Based on the features of these curves it is clear that proto-zoa have the ability to ingest substantial numbers of prokar-yote and eukaryote prey in natural ecosystems Somebacterivorous flagellates can consume bacteria (and cyano-bacteria) at rates up to 100 s per individual per hour whereassome bacterivorous ciliates can consume thousands of bacte-ria per individual per hour Rates of consumption of eukary-otic phytoplankton by herbivorous ciliates and largeflagellates are considerably more variable and highly depend-ent of the feeding mechanism (eg filtration in ciliates versuspseudopodial net in dinoflagellates) and the size of the prey
These lab studies demonstrate that rarely are prey suffi-ciently abundant in nature to support the maximal ingestionrates of which many protozoa are capable Nevertheless pre-dation rates of protozoa in nature are sufficient to constitute amajor factor controlling the standing stocks of bacteria andphytoplankton (63 180 184 355) For example these sum-maries indicate that on average protozoa consume 20ndash80ofthe standing stock of phytoplankton (measured as chloro-phyll) in the world ocean and 60ndash75 of daily primary pro-duction a departure from the classical view of copepods as theprimary consumers of phytoplankton This large role of her-bivorous protozoa infers a large role for copepods as consumersof planktonic protozoa (185) Similarly the role of protozoaas major consumers of bacterial biomass is indicated by therelative balance between bacterial production on the onehand and bacterivory on the other (Fig 8) The approximateone-to-one balance between these measurements over a largenumber of studies implies that protozoan bacterivory must bea major factor determining the fate of bacteria in marine (andfreshwater) plankton ecosystems
It is interesting to note that some types of bacteria includ-ing marine Bdellovibrio are known to prey on other marine
bacteria (362) At the time of this writing the importanceof bacterial predation by other bacteria within the contextof oceanographic processes is not known and presumed tobe small While Bdellovibrio may be cultivated from seawaterit can also be found among sequences from cold sediments(363) and the water column Bdellovibrio happens to be thebest studied predatory bacterium but there are no doubtothers that we simply have not yet identified (364)
Species-Specific Interactions (MutualismParasitism Commensalisms)Interactions between marine microorganisms such as compe-tition allelopathy and symbiosis are fundamental aspectscontrolling community composition and activity but atpresent most are poorly characterized in natural marine sys-tems Bacteria must compete for dissolved and particulatecompounds with other bacteria andwith protistan taxa whilephagotrophic protists must compete with each other andmetazoa for available prey These interactions manifest them-selves in many forms including true competition for availableresources resource partitioning such that some taxa specializeon the utilization of certain substrates or prey while othersspecialize on different ones (reducing competition) or eventhe cooperative use of compounds (eg one bacteriumrsquoswaste products being utilized as substrate by another bacte-rium) These metabolic consortia are probably common inthe ocean but can be difficult to identify and characterizeConsortia of bacteria working together to perform complexreactions especially in anaerobic systems are well known inmicrobiology
Consortia in which there is a very high degree of integra-tion between species (ie symbiosis) are very common in theocean These associations may be beneficial to both partners(mutualism) beneficial to one part and inconsequential forthe other (commensalism) or beneficial to one part and det-rimental to the other (parasitism) Many highly visible andwell-characterized marine mutualisms exist between chemo-synthetic bacteria and animals such as hydrothermal ventor methane seep tubeworms clams mussels or other inverte-brates (365) and also between luminescent bacteria and fishor squids (366) Similarly many protozoa form mutualisticassociations with photosynthetic protists (367) The mostconspicuous of these associations involved large amoeboidprotozoa of oceanic ecosystems (planktonic foraminiferanspolycystine radiolaria and acantharia) which contain largenumbers of intracellular algae (74) but associations betweensome heterotrophic dinoflagellates and cyanobacteria are alsocommon (368) Studies have indicated that efficient nutrientcycling in these protozoan-algal associations confers an eco-logical advantage for both partners in surfacewaters of the oli-gotrophic ocean
Parasitism is also a common symbiotic relationship amongmarine protozoa Protozoan parasites have been reported for adiverse array of protists and animals in the ocean Heterotro-phic dinoflagellates and presently uncultured alveolate cladesrelated to alveolates appear to be particularly successful atevolving thismode of existence (369ndash371) In a common par-asitic group Amoebophrya spp the parasite invades photo-synthetic dinoflagellates where it undergoes reproductioneventually lysing the host and releasing dozens of infectiveswarmers The parasitic protist Pirsonia spp is a parasite ofdiatoms (372) The overall importance of these parasitismsin the population dynamics of their hosts is not clearly knownbut may constitute a significant source of mortality for somespecies (373 374) The frequency and diversity of ldquonewrdquo
FIGURE 8 Meta-analysis of experimental studies that have com-pared rates of bacterial productivity (converted to population growthrate units of dminus1) with rates of bacterivory by phagotrophic protists(expressed as equivalent units of bacterial growth rate dminus1) Thesolid line indicates where measurements of bacterial removal by graz-ers were equal tomeasurements of bacterial production Data redrawnfrom Sanders et al (63) doi1011289781555818821ch422f8
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-21
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
parasitic phylotypes from environmental gene surveys impliesthat there are probably many more parasitic relationships yetto be described in the ocean (371)
In addition to these organism-organism associations thereare numerous examples of ldquochemical warfarerdquo among marinemicroorganisms Allelopathy is defined as the production ofsubstances that adversely affect competing species althoughsubstances that are produced to deter grazers is also sometimesincluded under this rubricManymarinemicroorganisms pro-duce substances that adversely affect growth or survival ofother species The most obvious example is the existence ofnumerous classes of antibiotics produced by marine microor-ganisms One study showed that out of 86 marine microbialisolates tested more than half produced substances thatinhibited the growth of other isolates (375) Moreover itwas found that bacteria isolated from marine aggregateswere more likely to produce these substances then those iso-lated from free-living plankton Thus species living in closephysical proximity to one another may be more likely to pro-duce these substances Disruption of cell-cell communica-tion via production or degradation of chemical signals iscurrently an area of intensive research within marine ecologySee Roy et al (376) for a recent review of the chemical ecol-ogy of the marine plankton
Bacteria and phytoplankton in the planktonmay competeactively for various macro- and micronutrients and thesecompetitive interactions appear to be an important reasonfor the production of many allelopathic compounds (377)In addition a wide spectrum of protistan species produce sec-ondary metabolites that are believed to play a role in theinhibition of competing species (378 379) andor in reducingpredation by protozoa andor metazoa (380ndash382)
Balancing Microbial Growth and RemovalMicrobial communities are highly dynamic They can exhibitrapid shifts in community composition and significantincreases or decreases in biomass of species or whole assemb-lages Nevertheless over reasonably large scales of time andspace these communities are remarkably constant Thisobservation implies that processes of production and removalmust be in overall balance within the ocean
For heterotrophic bacteria production is dependent onavailable substrate for growth and the efficiency of conversioninto bacterial biomass Numerous measurements and esti-mates have been made of bacterial growth efficiency sincethe 1980s These estimates range broadly from approximately1 to more than 60 but typically average in the 20ndash30range (reviewed by del Giorgio and Cole [383]) indicatingthat the production of bacterial biomass is a relatively effi-cient process Nevertheless as noted already abundances ofbacteria in the ocean remain remarkably constant thereforeproduction must be closely coupled to losses from predation(protozoa viruses) because these cells are sufficiently smallthat sinking losses must be minimal The explanation forthis tight coupling between bacterial production and loss ratesis not completely clear but may relate to the fact that bacte-rial predators are capable of rapid increases in abundanceThis conclusion is also supported by summaries of large datasets that indicate an overall balance between bacteria produc-tion and bacterial mortality across a wide spectrum of envi-ronments (Fig 8)
Protozoan populations exhibit short-term (weeks) toseasonal variances in abundances that exceed the variancestypically observed for bacteria but massive ldquobloomsrdquo ofnano- or microzooplankton are not common This is notthe case for phytoplankton assemblages which exhibit
considerably more temporal and spatial variability than bac-teria or protozoa For example chlorophyll may vary two tothree orders of magnitude between rich coastal and oligotro-phic open ocean environments or seasonally in some coastalecosystems The less dramatic changes in the standing stock ofprotozoa cannot be attributed to low growth inefficiencyTypical gross growth efficiencies for these species are quitehigh ranging from 30 to 40 (329 384) It is probablythat many metazoan zooplankton prey heavily on protozoathereby exerting strong top-down control on these assemb-lages (and perhaps relieving phytoplankton from grazing pres-sure exerted by the microzooplankton)
THE MICROBIAL LOOP REVISITEDThe original concept of the microbial loop (12 34) depicteda large fraction of pelagic carbon flux passing from DOMPOM to bacteria and subsequently bacterial biomass beinggrazed by protists that enter the classic food web That depic-tion of microbial assemblages and energy flow has becomemore complicated in recent years with the inclusion ofviruses mixotrophs (both prokaryotic and eukaryotic) arch-aea bacterial and archaeal autotrophs phototrophs like cya-nobacteria and proteorhodopsin-based phototrophy (150)Our views regarding the small-scale distributions of microbesand their substrates have also changed For example Azamet al (325 385) have pictured the marine bacteria as beingembedded in a very loose gel-like matrix composed of thesematerials with various local ldquohot spotsrdquo of microbial activitywhere there may be a particularly rich source of nutrients(leaky organism degrading particle etc) These small-scalegradients and interactions are undoubtedly important forunderstanding how these systems function (386)
Despite these new wrinkles in our understanding or per-haps because of them it is now widely recognized that heter-otrophic microbes (viruses archaea bacteria protozoa) playpivotal roles in the ocean as agents of elemental transforma-tion and energy flow (85 387) Their activities may compli-cate how we model energy production and utilization andnutrient cycling but these populations are nonetheless recog-nized as integral components of global biogeochemical cyclesIn the 1980s a debate began about whether the microbialloop is primarily a link in marine food webs passing salvagedDOM and POM back up to larger organisms or a sink respir-ing almost all the carbon via microbial trophic interactions Itis clear now that the microbial loop (broadly defined) playsfundamental roles in both trophic transfer of energy and car-bonnutrient remineralization Which of these roles domi-nates is highly dependent on environmental conditions andcommunity composition For example the remineralizationof organic matter by heterotrophic bacteria constitutes a sub-stantial (albeit unavoidable) loss of primary production tohigher tropic levels At the same time photosynthetic cyano-bacteria lt2 microm in size make up the largest portion of total pri-mary production in many open ocean ecosystems The vastmajority of this biomassmust enter the foodweb via their con-sumption by minute protozoa Thus the only link for this bio-mass is via the microbial loop
RESEARCH TRENDS INCLUDING MICROBIALASSOCIATION NETWORKSMolecular approaches are going beyond just fueling aremarkable ldquoage of microbial discoveryrdquo that in many wayshas paralleled the level of discovery experienced by biologistsstudying macroorganisms during the late 1800s and early
422-22 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
1900s Molecular techniques are being employed to docu-ment the presence and abundance of ecologically importantor ldquoenvironmentally relevantrdquo species of microorganisms innatural aquatic communities to characterize the diversityand community structure of microbial communities and tobegin to understand how microbial diversity extrapolates tothe emergent properties of communities (eg primary pro-duction respiration food web structure) The developmentand application of these tools is enabling extensive anddetailed observations and experimental studies of microbesand their interactions A very recent example is the use ofmicrobial association networks to mathematically examinethe co-occurrence of multiple microbial types and environ-mental parameters to examine possible interactions typicallydone in time series studies and potentially including bacteria
archaea protists zooplankton and viruses (Fig 9) (388ndash390) These approaches are uncovering previously unrecog-nized correlations between taxa (positive negative simulta-neously or time-lagged) that hint at functional relationshipsbetween them for example predator-prey symbiotic or otherrelationships Such correlations are fodder for hypothesesregarding the nature of these relationships and experimentalstudies to test them For example association network analy-sis from the Tara Oceans expedition pointed to a specificmicrobial-animal photosymbiotic interaction between a flat-worm and a green microalga which was then verified bydirect observation (391)
As illustrated throughout this manual molecular methodsare used increasingly to investigate important biogeoche-mical processes such as the cycling reactions of N fixation
FIGURE9 Microbial association network showing co-occurrence patterns of near-surface planktonicmicrobes at the SanPedroOceanTimeSeries USCMicrobial Observatory sampled monthly for 3 years This network shows only organisms and parameters directly correlated to cya-nobacteria (green Prochlorococcus pink Synechococcus) Circles bacteria V-shape T4-like myoviruses blue diamonds protists hexagons andsquares environmental parameters and processes (nutrients salinity chlorophyll a [Chl_a] primary productivity [Prim_Prod]) solid lines pos-itive correlation asmeasured by local similarity analysis dashed line negative correlation arrows point to correlations lagged by 1month data inChow et al 2013 (388) Symbols sized to reflect relative abundances within each group doi1011289781555818821ch422f9
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-23
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
nitrification and denitrification (392 393) Given thepresent realization that microbial processes dominate biolog-ical productivity energy utilization and nutrient cycling inthe ocean these studies will provide fundamental knowledgeregarding how biological communities within the ocean arestructured and how they function
We thank our many students postdocs and colleagues for all their con-tributions over the years and support from the National Science Founda-tion including grants OCE 1031743 OCE1136818 OCE0962309MCB0703159 the National Oceanic and Atmospheric Administration(NA11NOS4780052) and the Gordon and Betty Moore Foundation
REFERENCES
1 Calkins GN 1901 Marine protozoa from Woods HoleBull Bur Fish 21413ndash468
2 Fischer B 1883 Bacteriologische Untersuchungen aufeiner Reise nach Westindien Zeitschr Hyg 1421ndash464
3 Haeckel E 1887 Report on radiolaria collected by HMS Challenger during the 1873ndash1876 pp 1ndash1760 InThompson CW Murray J (eds) The Voyage of the HMSChallenger vol 18 Her Majestyrsquos Stationary OfficeLondon
4 Jannasch HW Jones GE 1959 Bacterial populations insea water as determined by different methods of enumera-tion Limnol Oceanogr 4128ndash139
5 Parsons TR Strickland JDH 1961 On the production ofparticulate organic carbon by heterotrophic processes insea-water Deep Sea Res 8211ndash222
6 Wright RT Hobbie JE 1966 Use of glucose and acetateby bacteria and algae in aquatic ecosystems Ecology 47447ndash453
7 Beers JR Stewart GL 1969 Micro-zooplankton and itsabundance relative to the larger zooplankton and other ses-ton components Mar Biol 4182ndash189
8 Fenchel T 1967 The ecology of marine microbenthosI The quantitative importance of ciliates as comparedwith metazoans in various types of sediments Ophelia 4121ndash137
9 Lighthart B 1969 Planktonic and benthic bacteriovorousprotozoa at eleven stations in Puget Sound and adjacentPacific Ocean Can J Fish Aquat Sci 26299ndash304
10 Utermoumlhl H 1958 Zur Vervollkommung der quantita-tiven phytoplankton-methodik Mitt Int Ver Limnol 938
11 Steele JH 1974 The Structure of Marine Ecosystems Har-vard University Press Cambridge MA
12 Pomeroy LR 1974 The oceanrsquos food web a changingparadigm Bioscience 24499ndash504
13 Andrews PWilliams PJL 1971 Heterotrophic utilizationof dissolved compounds in the sea IIIMeasurements of theoxidation rates and concentrations of glucose and aminoacids in sea water J Mar Biol Assoc UK 51111ndash125
14 Williams PJL 1981 Microbial contribution to overallmarine plankton metabolismmdashdirect measurements of res-piration Oceanolog Acta 4359ndash364
15 Fenchel TM Jorgensen BB 1977 Detritus food chains ofaquatic ecosystems the role of bacteria pp 1ndash58 InAlexander M (ed) Adv microb ecol vol 1 Plenum PressNew York NY
16 Francisco DE Mah RA Rabin AC 1973 Acridineorange epifluorescence technique for counting bacteria innatural waters Trans Am Microsc Soc 92416ndash421
17 Hobbie JE Daley RJ Jasper S 1977 Use of Nuclepore fil-ters for counting bacteria by fluorescence microscopy ApplEnviron Microbiol 331225ndash1228
18 Beers JR Stewart GL 1971 Micro-zooplankters in theplankton communities of the upper waters of the easterntropical Pacific Deep Sea Res 18861ndash883
19 CaronDA 1983 Technique for enumeration of heterotro-phic and phototrophic nanoplankton using epifluores-cence microscopy and comparison with otherprocedures Appl Environ Microbiol 46491ndash498
20 Haas LW 1982 Improved epifluorescence microscopy forobserving planktonicmicro-organismsAnnls Inst Oceanogr58261ndash266
21 Sherr EB Caron DA Sherr BF 1993 Staining of hetero-trophic protists for visualization via epifluorescence micro-scopy pp 213ndash227 In Kemp P Sherr B Sherr E Cole J(eds) Handbook of Methods in Aquatic Microbial EcologyLewis Publishers Boca Raton FL
22 Montagnes DJS Lynn DH 1993 A quantitative protar-gol stain (QPS) for ciliates and other protists pp229ndash240 In Kemp PF Sherr BF Sherr EB Cole JJ (eds)Handbook of Methods in Aquatic Microbial Ecology LewisPublishers Boca Raton FL
23 Hagstroumlm A Larsson U Horstedt P Normark S 1979Frequency of dividing cells a new approach to the determi-nation of bacterial growth rates in aquatic environmentsAppl Environ Microbiol 37805ndash812
24 Fuhrman JA Azam F 1980 Bacterioplankton secondaryproduction estimates for coastal waters of British ColumbiaAntarctica and California Appl Environ Microbiol 391085ndash1095
25 Fuhrman JAAzam F 1982 Thymidine incorporation as ameasure of heterotrophic bacterioplankton production inmarine surface waters evaluation and field results MarBiol 66109ndash120
26 Kirchman Dl KrsquoNees E Hodson RE 1985 Leucineincorporation and its potential as a measure of protein syn-thesis by bacteria in natural aquatic systems Appl EnvironMicrobiol 49599ndash607
27 CaronDADavis PGMadinLP Sieburth JM 1982Het-erotrophic bacteria and bacterivorous protozoa in oceanicmacroaggregates Science 218795ndash797
28 FenchelT 1982 Ecology of heterotrophicmicroflagellatesI Some important forms and their functional morphologyMar Ecol Prog Ser 8211ndash223
29 Sherr BF Sherr EB Berman T 1982 Decomposition oforganic detritus a selective role for microflagellate proto-zoa Limnol Oceanogr 27765ndash769
30 Capriulo GM Carpenter EJ 1980 Grazing by 35 to 202micrommicro-zooplankton in Long Island SoundMar Biol 56319ndash326
31 Heinbokel JF Beers JR 1979 Studies on the functionalrole of tintinnids in the Southern California Bight IIIGrazing impact of natural assemblagesMar Biol 5223ndash32
32 Sherr BF Sherr EB Andrew TL Fallon RD Newell SY1986 Trophic interactions between heterotrophic Proto-zoa and bacterioplankton in estuarine water analyzedwith selective metabolic inhibitors Mar Ecol Prog Ser 32169ndash179
33 Stoecker DK Capuzzo JM 1990 Predation on protozoaits importance to zooplankton J Plankton Res 12891ndash908
34 Azam F Fenchel T Gray JG Meyer-Reil LA ThingstadT 1983 The ecological role of water-column microbes inthe sea Mar Ecol Prog Ser 10257ndash263
35 DeLong EF Taylor LT Marsh TL Preston CM 1999Visualization and enumeration of marine planktonic arch-aea and bacteria by using polyribonucleotide probes andfluorescent in situ hybridization Appl Environ Microbiol655554ndash5563
36 Fuhrman JA Ouverney CC 1998 Marine microbialdiversity studied via 16S rRNA sequences cloning resultsfrom coastal waters and counting of native archaeawith flu-orescent single cell probes Aq Ecol 323ndash15
37 Teira E Reinthaler T PernthalerA Pernthaler JHerndlGJ 2004 Combining catalyzed reporter deposition-
422-24 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
fluorescence in situ hybridization and microautoradio-graphy to detect substrate utilization by bacteria and arch-aea in the deep ocean Appl Environ Microbiol 704411ndash4414
38 Noble RT Fuhrman JA 1998 Use of SYBR Green I forrapid epifluorescence counts of marine viruses and bacteriaAq Microb Ecol 14113ndash118
39 Porter KG Feig YS 1980 The use of DAPI for identifyingand counting aquatic microflora Limnol Oceanogr 25943ndash948
40 Velji MI Albright LJ 1986 Microscopic enumeration ofattached marine bacteria of seawater marine sedimentfecal matter and kelp blade samples following pyrophos-phate and ultrasound treatments Can J Microbiol 32121ndash126
41 delGiorgio PA Gasol JM Vaque D Mura P Agusti SDuarte CM 1996 Bacterioplankton community structureprotists control net production and the proportion of activebacteria in a coastal marine community Limnol Oceanogr411169ndash1179
42 Monger BC Landry MR 1992 Size-selective grazing byheterotrophic nanoflagellates an analysis using live-stained bacteria and dual-beam flow cytometry ArchHydrobiol Beih 37173ndash185
43 Campbell L Nolla HA Vaulot D 1994 The importanceof prochlorococcus to community structure in the centralnorth Pacific-Ocean Limnol Oceanogr 39954ndash961
44 Ducklow HW 2000 Bacterial production and biomass inthe oceans pp 85ndash120 In Kirchman DL (ed) MicrobialEcology of the Oceans Wiley-Liss New York NY
45 Fuhrman JA Ammerman JW Azam F 1980 Bacterio-plankton in the coastal euphotic zone distribution activ-ity and possible relationships with phytoplankton MarBiol 60201ndash207
46 SimonM Azam F 1989 Protein content and protein syn-thesis rates of planktonicmarine bacteriaMar Ecol Prog Ser51201ndash213
47 Caron DA Lim EL Kunze H Cosper EM AndersonDM 1989 Trophic interactions between nano- andmicro-zooplankton and the ldquobrown tiderdquo pp 265ndash294 In CosperEM Bricelj VM Carpenter EJ (eds) Novel PhytoplanktonBlooms Causes and Impacts of Recurrent Brown Tides andOther Unusual Blooms vol 35 Springer Berlin
48 Agogue H Brink M Dinasquet J Herndl GJ2008 Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic Nature456788ndash791
49 Schmidt JL Deming JW Jumars PA Keil RG 1998Constancy of bacterial abundance in surficial marine sedi-ments Limnol Oceanogr 43976ndash982
50 Fuhrman JA Eppley RW Hagstrom A Azam F 1985Diel variation in bacterioplankton and related parametersin the Southern California Bight Mar Ecol Prog Ser 279ndash20
51 Wikner J Hagstrom A 1991 Annual study of bacterio-plankton community dynamics Limnol Oceanogr 361313ndash1324
52 CarlsonCADucklowHWSleeter TD 1996 Stocks anddynamics of bacterioplankton in the northwestern SargassoSea Deep Sea Res 43491ndash515
53 Burney CMDavis PG JohnsonKM Sieburth JM 1982Diel relationships of microbial trophic groups and in situdissolved carbohydrate dynamics in the Caribbean SeaMar Biol 67311ndash322
54 Gasol JMDovalMDPinhassi J Calderon-Paz JIGuixa-Boixareu N Vaque D Pedros-Alio C 1998 Diel varia-tions in bacterial heterotrophic activity and growth in thenorthwestern Mediterranean Sea Mar Ecol Prog Ser 164107ndash124
55 Hagstroumlm A Pinhassi J Zweifel UL 2001 Marine bac-terioplankton show bursts of rapid growth induced by sub-strate shifts Aq Microb Ecol 24109ndash115
56 Fuhrman JA 1987 Close coupling between release anduptake of dissolved free amino acids in seawater studied byan isotope dilution approachMar Ecol Prog Ser 3745ndash52
57 Pomeroy LRWiebeWJ Deibel D Thompson RJ RoweGT Pakulski JD 1991 Bacterial responses to temperatureand substrate concentration during the Newfoundlandspring bloom Mar Ecol Prog Ser 75143ndash159
58 Eppley RW 1972 Temperature and phytoplanktongrowth in the sea Fish Bull 701063ndash1085
59 Rose JM Caron DA 2007 Does low temperature con-strain the growth rates of heterotrophic protists Evidenceand implications for algal blooms in coldwater LimnolOce-anogr 52886ndash895
60 Jansson J Prosser J 1997 Quantification of the presenceand activity of specific microorganisms in natureMol Bio-technol 7103ndash120
61 Bird DF Kalff J 1984 Empirical relationships betweenbacterial abundance and chlorophyll concentration infresh and marine waters Can J Fish Aquat Sci 411015ndash1023
62 Cole JJ Findlay S PaceML 1988 Bacterial production infresh and saltwater ecosystems a cross-system overviewMarEcol Prog Ser 431ndash10
63 Sanders RWCaronDA BerningerU-G 1992 Relation-ships between bacteria and heterotrophic nanoplankton inmarine and fresh water an inter-ecosystem comparisonMar Ecol Prog Ser 861ndash14
64 Fuhrman JA Sleeter TD Carlson CA Proctor LM1989 Dominance of bacterial biomass in the SargassoSea and its ecological implications Mar Ecol Prog Ser 57207ndash217
65 RomanMRCaronDAKremer P Lessard EJMadin LPMalone TC Napp JM Peele ER Youngbluth MJ 1995Spatial and temporal changes in the partitioning of organiccarbon in the plankton community of the Sargasso Sea offBermuda Deep Sea Res 42973ndash992
66 Cho B Azam F 1990 Biogeochemical significance of bac-terial biomass in the oceanrsquos euphotic zone Mar Ecol ProgSer 63253ndash259
67 Sieracki ME Haugen EM Cucci TL 1995 Overestima-tion of heterotrophic bacteria in the Sargasso Sea directevidence by flow and imaging cytometry Deep Sea Res421399ndash1409
68 Whittaker RH 1969 New concepts of kingdoms of organ-isms Science 163150ndash160
69 Keeling PJ 2013 The number speed and impact of plastidendosymbioses on eukaryotic evolution Annu Rev PlantBiol 64583ndash607
70 Burkholder JM Glibert PM Skelton HM 2008 Mixo-trophy a major mode of nutrition for harmful algal speciesin eutrophic waters Harmful Algae 877ndash93
71 Hansen PJ 2011 The role of photosynthesis and fooduptake for the growth of marine mixotrophic dinoflagel-lates J Euk Microbiol 58203ndash214
72 Stoecker DK 1999 Mixotrophy among dinoflagellates JEuk Microbiol 46397ndash401
73 Skovgaard A 1998 Role of chloroplast retention in amarine dinoflagellate Aq Microb Ecol 15293ndash301
74 Caron DA 2000 Symbiosis and mixotrophy among pela-gic microorganisms pp 495ndash523 In Kirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
75 Mitra A Flynn KJ 2010 Modelling mixotrophy in harm-ful algal blooms more or less the sumof the parts JMar Sys-tems 83158ndash169
76 Sanders RW 1991 Mixotrophic protists in marine andfreshwater ecosystems J Protozool 3876ndash81
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-25
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
77 Stoecker DK 1998 Conceptual models of mixotrophy inplanktonic protists and some ecological and evolutionaryimplications Eur J Protistol 34281ndash290
78 Woese CR Kandler O Wheelis ML 1990 Towards anatural system of organisms proposal for the domainsArchaea Bacteria and Eukarya Proc Natl Acad Sci 874576ndash4579
79 Adl SM Simpson AGB Lane CE Lukeš J Bass DBowser SS Brown MW Burki F Dunthorn M HamplV Heiss A Hoppenrath M Lara E le Gall L LynnDH McManus H Mitchell EAD Mozley-StanridgeSE Parfrey LW Pawlowski J Rueckert S Shadwick LSchoch CL Smirnov A Spiegel FW 2012 The revisedclassification of eukaryotes J Euk Microbiol 59429ndash514
80 CaronDACountway PD JonesAC KimDY SchnetzerA 2012 Marine protistan diversity Ann Rev Mar Sci 4467ndash493
81 Guillou L Moon-van der Staay S-Y Claustre H Parten-sky F Vaulot D 1999 Diversity and abundance of Bolido-phyceae (Heterokonta) in two oceanic regions ApplEnviron Microbiol 654528ndash4536
82 Anderson OR 1983 Radiolaria Springer New York NY83 Swanberg NR Harbison GR 1980 The ecology ofCollo-
zoum longiforme sp nov a new colonial radiolarian fromthe equatorial Atlantic Ocean Deep Sea Res 27A715ndash732
84 Sieburth JM SmetacekV Lenz J 1978 Pelagic ecosystemstructure heterotrophic compartments of the plankton andtheir relationship to plankton size fractions Limnol Ocean-ogr 231256ndash1263
85 WordenAZ FollowsMJ Giovannoni SJWilken S Zim-merman AE Keeling PJ 2015 Environmental scienceRethinking themarine carbon cycle factoring in themulti-farious lifestyles of microbes Science 3471257594
86 Garrison DL Gowing MM Hughes MP Campbell LCaron DA Dennett MR Shalapyonok A Olson RJLandry MR Brown SL Liu HB Azam F Steward GFDucklowHW SmithDC 2000Microbial foodweb struc-ture in the Arabian Sea a US JGOFS studyDeep Sea Res II471387ndash1422
87 Arenovski AL Lim EL Caron DA 1995 Mixotrophicnanoplankton in oligotrophic surfacewaters of the SargassoSea may employ phagotrophy to obtain major nutrients JPlankton Res 17801ndash820
88 Sanders RW Gast RJ 2011 Bacterivory by phototrophicpicoplankton and nanoplankton in Arctic waters FEMSMicrobiol Ecol 82242ndash253 doi 101111j1574-6941201101253x242-252
89 Smalley GW Coats DW Adam EJ 1999 A new methodusing fluorescent microspheres to determine grazing on cil-iates by the mixotrophic dinoflagellate Ceratium furca AqMicrob Ecol 17167ndash179
90 Unrein F Gasol JM Not F Forn I Massana R 2014Mixotrophic haptophytes are key bacterial grazers in oligo-trophic coastal waters ISME J 8164ndash176
91 Bockstahler KR Coats DW 1993 Grazing of the mixo-trophic dinoflagellate Gymnodinium sangiuneum on ciliatepopulations of Chesapeake Bay Mar Biol 116477ndash487
92 Mitra A Flynn KJ Burkholder JM Berge T Calbet ARaven JA Graneacuteli E Glibert PM Hansen PJ StoeckerDKThingstad F TillmannUVaringge SWilken S ZubkovMV 2014 The role of mixotrophic protists in the biolog-ical carbon pump Biogeosciences 11995ndash1005
93 Stoecker D Taniguchi A Michaels AE 1989 Abun-dance of autotrophic mixotrophic and heterotrophicplanktonic ciliates in shelf and slope waters Mar EcolProg Ser 50241ndash254
94 Crawford DW 1989 Mesodinium rubrum the phyto-plankter that wasnrsquot Mar Ecol Prog Ser 58161ndash174
95 EstebanG Fenchel T Finlay BJ 2010Mixotrophy in cil-iates Protist 161621ndash641
96 Johnson MD 2011 Acquired phototrophy in ciliates areview of cellular interactions and structural adaptationsJ Euk Microbiol 58185ndash195
97 Caron DA Swanberg NR 1990 The ecology of plank-tonic sarcodines Rev Aq Sci 3147ndash180
98 Decelle J Probert I Bittner L Desdevises Y Colin S deVargas C Galiacute M SimoacuteRNot F 2012 An original modeof symbiosis in open ocean plankton Proc Nat Acad Sci10918000ndash18005
99 Caron DA Michaels AF Swanberg NR Howse FA1995 Primary productivity by symbiont-bearing plank-tonic sarcodines (acantharia radiolaria foraminifera) insurface waters near Bermuda J Plankton Res 17103ndash129
100 Evans GT Taylor FJR 1980 Phytoplankton accumula-tion in Langmuir cells Limnol Oceanogr 25840ndash845
101 Giovannoni SJ Rappe M 2000 Evolution diversity andmolecular ecology of marine prokaryotes pp 47ndash84 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
102 Fuhrman J Hagstroumlm Aring 2008 Bacterial and archaealcommunity structure and its patterns pp 45ndash90 In Kirch-manD (ed)Microbial Ecology of the Oceans 2nd edWileyHoboken New Jersey
103 Olsen GJ Lane DL Giovannoni SJ Pace NR 1986Microbial ecology and evolution a ribosomal RNAapproach Annu Rev Microbiol 40337ndash365
104 Pace NR Stahl DA Lane DL Olsen GJ 1986 The anal-ysis of natural microbial populations by rRNA sequencesAdv Microb Ecol 91ndash55
105 Venter JC Remington K Heidelberg JF Halpern ALRusch D Eisen JA Wu DY Paulsen I Nelson KE Nel-son W Fouts DE Levy S Knap AH Lomas MW Neal-son K White O Peterson J Hoffman J Parsons RBaden-Tillson H Pfannkoch C Rogers YH SmithHO 2004 Environmental genome shotgun sequencingof the Sargasso Sea Science 30466ndash74
106 Iverson V Morris RM Frazar CD Berthiaume CTMorales RL Armbrust EV 2012 Untangling genomesfrommetagenomes revealing an uncultured class of marineeuryarchaeota Science 335587ndash590
107 Sogin ML Morrison HG Huber JA Welch DM HuseSM Neal PR Arrieta JM Herndl GJ 2006 Microbialdiversity in the deep sea and the underexplored ldquorare bio-sphererdquo Proc Natl Acad Sci USA 10312115ndash12120
108 Lynch MDJ Neufeld JD 2015 Ecology and explorationof the rare biosphere Nat Rev Microbiol 13217ndash229
109 Huse S Welch D Morrison H Sogin M 2010 Ironingout the wrinkles in the rare biosphere through improvedOTU clustering Environ Microbiol 121889ndash1898
110 Kunin V Engelbrektson A Ochman H Hugenholtz P2010 Wrinkles in the rare biosphere pyrosequencingerrors can lead to artificial inflation of diversity estimatesEnviron Microbiol 12118ndash123
111 Cuvelier ML Allen AE Monier A McCrow JP MessieacuteM Tringe SG Woyke T Welsh RM Ishoey T LeeJ-T Binder BJ DuPont CL Latasa M Guigand CBuck KR Hilton J Thiagarajan M Caler E Read BLasken RS Chavez FP Worden AZ 2010 Targetedmetagenomics and ecology of globally important uncul-tured eukaryotic phytoplankton Proc Natl Acad Sci USA10714679ndash14684
112 YoonHS PriceDC StepanauskasRRajahVD SierackiME Wilson WH Yang EC Duffy S Bhattacharya D2011 Single-cell genomics reveals organismal interactionsin uncultivated marine protists Science 332714ndash717
113 Rinke C Schwientek P Sczyrba A IvanovaNNAnder-son IJ Cheng JF Darling A Malfatti S Swan BK Gies
422-26 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
EADodsworth JAHedlundBPTsiamis G Sievert SMLiu WT Eisen JA Hallam SJ Kyrpides NC Stepanaus-kasR Rubin EMHugenholtz PWoykeT 2013 Insightsinto the phylogeny and coding potential of microbial darkmatter Nature 499431ndash437
114 Stepanauskas R 2012 Single cell genomics an individuallook at microbes Curr Opin Microbiol 15613ndash620
115 Gifford SM Sharma S Booth M Moran MA 2013Expression patterns reveal niche diversification in a marinemicrobial assemblage ISME J 7281ndash298
116 Gilbert JA Field D Huang Y Edwards R Li W Gilna PJoint I 2008 Detection of large numbers of novel sequen-ces in themetatranscriptomes of complex marine microbialcommunities PLoS One 3e3042
117 Hewson I Poretsky RS Dyhrman ST Zielinski BWhite AE Tripp HJ Montoya JP Zehr JP 2009 Micro-bial community gene expression within colonies of thediazotroph Trichodesmium from the Southwest PacificOcean ISME J 31286ndash1300
118 Ottesen EA Young CR Gifford SM Eppley JMMarin R Schuster SC Scholin CA DeLong EF2014 Multispecies diel transcriptional oscillations inopen ocean heterotrophic bacterial assemblages Science345207ndash212
119 Shakya M Quince C Campbell JHYZK Schadt CWPodar M 2013 Comparative metagenomic and rRNAmicrobial diversity characterization using archaeal and bac-terial synthetic communities Environ Microbiol 151882ndash1899
120 Parada A Needham DM Fuhrman JA 2015 Every basematters assessing small subunit rRNA primers for marinemicrobiomes with mock communities time-series andglobal field samples Environ Microbiol doi 1011111462-292013023
121 Giovannoni SJ Britschgi TBMoyerCL FieldKG 1990Genetic diversity in Sargasso Sea bacterioplanktonNature34560ndash63
122 Waterbury JB Watson SW Guillard RLL Brand LE1979 Widespread occurrence of a unicellular marineplanktonic cyanobacterium Nature 227293ndash294
123 Johnson PW Sieburth JM 1979 Chroococcoid cyano-bacteria in the seamdashubiquitous and diverse phototropicbiomass Limnol Oceanogr 24928ndash935
124 Chisholm SW Olson RJ Zettler ERWaterbury J Goer-icke R Welschmeyer N 1988 A novel free-living pro-chlorophyte abundant in the oceanic euphotic zoneNature 334340ndash343
125 Morris RM Rappe MS Connon SA Vergin KL SieboldWA Carlson CA Giovannoni SJ 2002 SAR11 cladedominates ocean surface bacterioplankton communitiesNature 420806ndash810
126 Fuhrman JAMcCallumKDavis AA 1992 Novel majorarchaebacterial group from marine plankton Nature 356148ndash149
127 Delong EF 1992 Archaea in coastal marine environ-ments Proc Natl Acad Sci USA 895685ndash5689
128 Karner MB DeLong EF Karl DM 2001 Archaeal dom-inance in the mesopelagic zone of the Pacific OceanNature 409507ndash510
129 Kirchman DL Elifantz H Dittel AI Malmstrom RRCottrell MT 2007 Standing stocks and activity of archaeaand bacteria in the western Arctic Ocean Limnol Oceanogr52495ndash507
130 Teira E vanAkenH Veth CHerndl GJ 2006 Archaealuptake of enantiomeric amino acids in the meso- and bath-ypelagic waters of the North Atlantic Limnol Oceanogr 5160ndash69
131 Church MJ DeLong EF Ducklow HW Karner MBPreston CM Karl DM 2003 Abundance and distribution
of planktonic archaea and bacteria in the waters west of theAntarctic Peninsula Limnol Oceanogr 481893ndash1902
132 Massana R DeLong EF Pedros-Alio C 2000 A few cos-mopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces ApplEnviron Microbiol 661777ndash1787
133 Garcia-Martinez J Rodriguez-Valera F 2000Microdiver-sity of uncultured marine prokaryotes the SAR11 clusterand the marine Archaea of Group I Mol Ecol 9935ndash948
134 Fuhrman J 2011Oceans of Crenarchaeota a personal his-tory describing this paradigm shift Microbe 6531ndash537
135 Brochier-Armanet C Boussau B Gribaldo S Forterre P2008 Mesophilic crenarchaeota proposal for a third arch-aeal phylum the ThaumarchaeotaNature Rev Microbiol 6245ndash252
136 Ouverney CC Fuhrman JA 2000 Marine planktonicarchaea take up amino acids Appl Environ Microbiol 664829ndash4833
137 Kuypers MMM Blokker P Erbacher J Kinkel H Pan-cost RD Schouten S Damste JSS 2001 Massive expan-sion of marine archaea during a mid-Cretaceous oceanicanoxic event Science 29392ndash94
138 Pearson A McNichol AP Benitez-Nelson BC HayesJM Eglinton TI 2001 Origins of lipid biomarkers inSanta Monica Basin surface sediment a case study usingcompound-specific delta C-14 analysis Geochim Cosmo-chim Acta 653123ndash3137
139 Wuchter C Schouten S Boschker HTS Damste JSS2003 Bicarbonate uptake by marine CrenarchaeotaFEMS Microbiol Lett 219203ndash207
140 Schleper C Jurgens G Jonuscheit M 2005 Genomicstudies of uncultivated archaea Nature Rev Microbiol 3479ndash488
141 Konneke M Bernhard AE de la Torre JR Walker CBWaterbury JB Stahl DA 2005 Isolation of an autotro-phic ammonia-oxidizing marine archaeon Nature 437543ndash546
142 Martens-Habbena W Berube PM Urakawa H de laTorre JR Stahl DA 2009 Ammonia oxidation kineticsdetermine niche separation of nitrifying archaea and bacte-ria Nature 461976ndashU234
143 Walker CB de la Torre JR Klotz MGUrakawaH PinelN Arp DJ Brochier-Armanet C Chain PSG Chan PPGollabgir A Hemp J Hugler M Karr EA Konneke MShin M Lawton TJ Lowe T Martens-Habbena WSayavedra-Soto LA Lang D Sievert SM RosenzweigAC Manning G Stahl DA 2010 Nitrosopumilus mariti-mus genome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine crenarchaeaProc Natl Acad Sci USA 1078818ndash8823
144 IngallsAEShahSRHansmanRLAluwihareLI SantosGMDruffel ERMPearsonA 2006Quantifying archaealcommunity autotrophy in themesopelagic ocean using nat-ural radiocarbon Proc Natl Acad Sci USA 1036442ndash6447
145 Hansman RL Griffin S Watson JT Druffel ERMIngallsAE PearsonAAluwihare LI 2009 The radiocar-bon signature of microorganisms in the mesopelagic oceanProc Natl Acad Sci USA 1066513ndash6518
146 Beja O Aravind L Koonin EV Suzuki MT Hadd ANguyen LP Jovanovich SB Gates CM Feldman RASpudich JL Spudich ENDeLongEF 2000 Bacterial rho-dopsin evidence for a new type of phototrophy in the sea[see comments] Science 2891902ndash1906
147 Beja O Spudich EN Spudich JL LeclercM DeLong EF2001 Proteorhodopsin phototrophy in the ocean Nature411786ndash789
148 Campbell B Waidner L Cottrell M Kirchman D 2007Abundant proteorhodopsin genes in the North AtlanticOcean Environ Microbiol 1099ndash109
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-27
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
149 Frigaard NU Martinez A Mincer TJ DeLong EF 2006Proteorhodopsin lateral gene transfer between marineplanktonic bacteria and archaea Nature 439847ndash850
150 Fuhrman JA SchwalbachMS Stingl U 2008 Proteorho-dopsins an arrayof physiological rolesNature RevMicrobiol6488ndash494
151 Gomez-Consarnau L Gonzalez JM Coll-Llado MGourdon P Pascher T Neutze R Pedros-Alio C Pin-hassi J 2007 Light stimulates growth of proteorhodopsin-containing marine Flavobacteria Nature 445210ndash213
152 Gomez-Consarnau L Akram N Lindell K Pedersen ANeutze R Milton DL Gonzalez JM Pinhassi J 2010Proteorhodopsin phototrophy promotes survival of marinebacteria during starvation PLoS Biol 8e1000358
153 Goacutemez-Consarnau L Gonzaacutelez JM Riedel T Jaenicke SSantildeudo-Wilhelmy SA Wagner-Doumlbler I Fuhrman JA2015 Proteorhodopsin light-enhanced growth linked tovitamin-B1 acquisition in marine Flavobacteria ISME Jdoi 101038ismej2015196
154 Shiba T 1989 Taxonomy and ecology of marine bacteriapp 9ndash24 InHarashima K Shiba T Murata N (eds) Aero-bic Photosynthetic Bacteria Japan Scientific Societies PressTokyo
155 Yurkov VV Beatty JT 1998 Aerobic anoxygenic photo-trophic bacteria Microbiol Mol Biol Rev 62695ndash724
156 Kolber ZS Plumley FG Lang AS Beatty JT Blanken-ship RE VanDover CL Vetriani C Koblizek M Rath-geber C Falkowski PG 2001 Contribution of aerobicphotoheterotrophic bacteria to the carbon cycle in theocean Science 2922492ndash2495
157 Schwalbach MS Fuhrman JA 2005 Wide-ranging abun-dances of aerobic anoxygenic phototrophic bacteria in theworld ocean revealed by epifluorescence microscopy andquantitative PCR Limnol Oceanogr 50620ndash628
158 Goericke R 2002 Bacteriochlorophyll a in the ocean isanoxygenic bacterial photosynthesis important LimnolOceanogr 47290ndash295
159 Cottrell MT Mannino A Kirchman DL 2006 Aerobicanoxygenic phototrophic bacteria in the Mid-AtlanticBight and the North Pacific Gyre Appl Environ Microbiol72557ndash564
160 Lami R Cottrell MT Ras J Ulloa O Obernosterer IClaustre H Kirchman DL Lebaron P 2007 High abun-dances of aerobic anoxygenic photosynthetic bacteria inthe South Pacific Ocean Appl Environ Microbiol 734198ndash4205
161 Kirchman DL Hanson TE 2013 Bioenergetics of photo-heterotrophic bacteria in the oceans Environ Microbiol 5188ndash199
162 Lopez-Garcia P Brochier C Moreira D Rodriguez-Valera F 2004 Comparative analysis of a genome fragmentof an uncultivated mesopelagic crenarchaeote reveals mul-tiple horizontal gene transfers Environ Microbiol 619ndash34
163 Fuhrman J 2003 Genome sequences from the seaNature4241001ndash1002
164 Palenik B Brahamsha B Larimer FW Land M HauserL Chain P Lamerdin J Regala W Allen EE McCarrenJ Paulsen I Dufresne A Partensky FWebb EAWater-bury J 2003 The genome of a motile marine Synechococ-cus Nature 4241037ndash1042
165 Rocap G Larimer FW Lamerdin J Malfatti S Chain PAhlgren NA Arellano A Coleman M Hauser L HessWR Johnson ZI Land M Lindell D Post AF Regala WShah M Shaw SL Steglich C Sullivan MB Ting CSTolonen A Webb EA Zinser ER Chisholm SW 2003Genome divergence in two Prochlorococcus ecotypes reflectsoceanic niche differentiationNature 4241042ndash1047
166 Kashtan N Roggensack SE Rodrigue S Thompson JWBiller SJ Coe A Ding H Marttinen P Malmstrom RR
Stocker R Follows MJ Stepanauskas R Chisholm SW2014 Single-cell genomics reveals hundreds of coexistingsubpopulations in wild Prochlorococcus Science 344416ndash420
167 Ammerman JW Fuhrman JA Hagstroumlm Aring Azam F1984 Bacterioplankton growth in seawater I Growthkinetics and cellular characteristics in seawater culturesMar Ecol Prog Ser 1831ndash39
168 ButtonDK Schuts FQuang PMartinRRobertsonBR1993 Viability and isolation of marine bacteria by dilutionculture theory procedures and initial resultsAppl EnvironMicrobiol 59881ndash891
169 Rappeacute MS Connon SA Vergin KL Giovannoni SJ2002 Cultivation of the ubiquitous SAR11 marine bacter-ioplankton clade Nature 418630ndash633
170 Simu K Hagstroumlm A 2004 Oligotrophic bacterioplank-ton with a novel single-cell life strategy Appl EnvironMicrobiol 702445ndash2451
171 Giovannoni S Stingl U 2007 The importance of cultur-ing bacterioplankton in the ldquoomicsrdquo ageNature Rev Micro-biol 5820ndash826
172 Rappe MS 2013 Stabilizing the foundation of the housethat ldquoomicsrdquo builds the evolving value of cultured iso-lates to marine microbiology Curr Opin Microbiol 16618ndash624
173 Caron DA Countway PD Lambert S Rose J SchaffnerR 2002 Protozoa in marineestuarine waters pp2613ndash2626 In Bitton G (ed) Encyclopedia of Environmen-tal Microbiology John Wiley amp Sons New York NY
174 Butler H Rogerson A 1995 Temporal and spatial abun-dance of naked amoebae (Gymnamoebae) in marinebenthic sediments J Euk Microbiol 42724ndash730
175 Hemleben C Spindler M Anderson OR 1988 Modernplanktonic foraminifera Springer New York NY
176 Jacobson DM Anderson DM 1986 Thecate heterotro-phic dinoflagellates feeding behavior and mechanisms JPhycol 22249ndash258
177 Sherr EB Sherr BF 2009 Capacity of herbivorous protiststo control initiation and development of mass phytoplank-ton blooms Aq Microb Ecol 57253ndash262
178 Archer SD Leakey RJG Burkill PH Sleigh MA 1996Microbial dynamics in coastal waters of east Antarcticaherbivory by heterotrophic dinoflagellates Mar Ecol ProgSer 139239ndash255
179 Jeong HJ 1999 The ecological roles of heterotrophicdinoflagellates in marine planktonic community J EukMicrobiol 46390ndash396
180 Sherr E Sherr B 2002 Significance of predation by pro-tists in aquatic microbial food webs Antonie van Leeuwen-hoek 81293ndash308
181 Lynn DH 2008 The Ciliated Protozoa CharacterizationClassification and Guide to the Literature SpringerNew York NY
182 Dolan JR Pierce RW 2012 Diversity and distributions oftintinnids pp 214ndash243 InDolan JRMontagnes DJSAga-tha S Coats DW Stoecker DK (eds)The Biology and Ecol-ogy of Tintinnid Ciliates Models for Marine Plankton JohnWiley amp Sons New York NY
183 Pierce RW Turner JT 1992 Ecology of planktonic cili-ates in marine food webs Rev Aq Sci 6139ndash181
184 Calbet A Landry MR 2004 Phytoplankton growthmicrozooplankton grazing and carbon cycling in marinesystems Limnol Oceanogr 4951ndash57
185 Calbet A Saiz E 2005 The ciliate-copepod link in marinefood ecosystems Aq Microb Ecol 38157ndash167
186 Schmoker C Hernaacutendez-Leoacuten S Calbet A 2013 Micro-zooplankton grazing in the oceans impacts data variabil-ity knowledge gaps and future directions J Plankton Res35691ndash706
422-28 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
187 Caron DA Countway PD Brown MV 2004 The grow-ing contributions of molecular biology and immunologyto protistan ecology molecular signatures as ecologicaltools J Euk Microbiol 5138ndash48
188 Countway PD Gast RJ Savai P Caron DA 2005 Proti-stan diversity estimates based on 18S rDNA from seawaterincubations in the western North Atlantic J Euk Microbiol5295ndash106
189 Diez B Pedros-Alio CMassana R 2001 Study of geneticdiversity of eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning and sequenc-ing Appl Environ Microbiol 672932ndash2941
190 Guillou L Eikrem W Chretiennot-Dinet MJ Le Gall FMassana R Romari K Pedros-Alio C Vaulot D 2004Diversity of picoplanktonic prasinophytes assessed bydirect nuclear SSU rDNA sequencing of environmentalsamples and novel isolates retrieved from oceanic andcoastal marine ecosystems Protist 155193ndash214
191 Stoeck T hayward B Taylor GT Varela R Epstein SS2006 A multiple PCR-primer approach to access themicroeukaryotic diversity in environmental samples Protist15731ndash43
192 de Vargas C Audic S Henry N Decelle J Maheacute F Log-ares R Lara E Berney C Le Bescot N Probert I Carmi-chael M Poulain J Romac S Colin S Aury J-M BittnerL Chaffron S Dunthorn M Engelen S Flegontova OGuidi L Horaacutek A Jaillon O Lima-Mendez G Lukeš JMalviya S Morard R Mulot M Scalco E Siano R Vin-cent F Zingone A Dimier C Picheral M Searson SKandels-Lewis S Coordinators TO Acinas SG BorkP Bowler C Gorsky G Grimsley N Hingamp P Iudi-cone D Not F Ogata H Pesant S Raes J Sieracki MESpeich S Stemmann L Sunagawa S Weissenbach JWincker P Karsenti E 2015 Eukaryotic plankton diver-sity in the sunlit ocean Science 3481261605
193 Andersen RA Saunders GW Paskind MP Sexton JP1993 Ultrastructure and 18S rRNA gene sequence forPelagomonas calceolata gen et sp nov and the descriptionof a new algal class the Pelagophyceae classis nov J Phycol29701ndash715
194 del Campo J Massana R 2011 Emerging diversity withinchrysophytes choanoflagellates and bicosoecids based onmolecular surveys Protist 162435ndash448
195 Guillou L Chreacutetiennot-Dinet M-J Medlin LK ClaustreH Loiseaux-de Goer S Vaulot D 1999 Bolidomonas anew genus with two species belonging to a new algal classthe Bolidophyceae (Heterokonta) J Phycol 35368ndash381
196 Guillou LR Chreacutetiennot-Dinet M-J Boulben S Moon-van der Staay SY Vaulot D 1999 Symbiomonas scintillansgen et sp nov and Picophagus flagellatus gen et sp nov(Heterokonta) two new heterotrophic flagellates of pico-planktonic size Protist 150383ndash398
197 Massana R Pernice M Bunge JA Campo Jd 2011Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean ISME J 5184ndash195
198 CaronDA Gast RJ 2008 The diversity of free-living pro-tists seen and unseen cultured and uncultured pp 67ndash93In Zengler K (ed) Accessing uncultivated microorganismsfrom the environment to organisms and genomes and backASM Press Washington DC
199 Massana R Guillou L Diez B Pedros-Alio C 2002Unveiling the organisms behind novel eukaryotic riboso-mal DNA sequences from the oceanAppl EnvironMicrobiol684554ndash4558
200 Stoeck T Epstein S 2003 Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments Appl Environ Microbiol692657ndash2663
201 Shalchian-Tabrizi K EikremW Klaveness D Vaulot DMingeMA LaGall F Romari K Throndsen J BotnenAMassana R Thomsen HA Jakobsen KS 2006 Telone-mia a new protist phylymwith affinity to chromist lineagesProc R Soc Londonr B 2731833ndash1842
202 Caron DA 2009 Protistan biogeography why all the fussJ Euk Microbiol 56105ndash112
203 Finlay BJ Esteban G Fenchel T 2004 Protist diversity isdifferent Protist 15515ndash22
204 Foissner W 2006 Biogeography and dispersal of micro-organisms a review emphasizing protists Acta Protozool45111ndash136
205 Stoeck T Jost S Boenigk J 2008 Multigene phylogeniesof clonal Spumella-like strains a cryptic heterotrophicnanoflagellate isolated from different geographical regionsInt J Syst Evol Microbiol 58716ndash724
206 Caron DA Countway PD 2009 Hypotheses on the roleof the protistan rare biosphere in a changing world AqMicrob Ecol 57227ndash238
207 Dolven JK Lindqvist C Albert VA Bjoslashrklund KRYuasa T Takahashi OMayama S 2007Molecular diver-sity of alveolates associated with neritic North Atlanticradiolarians Protist 15865ndash76
208 Fenchel T Finlay BJ 2004 The ubiquity of small speciespatterns of local and global diversity Bioscience 54777ndash784
209 FenchelT 1982 Ecology of heterotrophicmicroflagellatesIII Adaptations to heterogeneous environmentsMar EcolProg Ser 925ndash33
210 MatinAAuger EA Blum PH Schultz JE 1989 Geneticbasis of starvation survival in nondifferentiating bacteriaAnn Rev Microbiol 43292ndash316
211 Spindler M Bayer U Hemleben C Beacute AWH AndersonOR 1979 Lunar periodicity in the planktonic foraminiferHastigerina pelagica Mar Ecol Prog Ser 161ndash64
212 Proctor LM Fuhrman JA Ledbetter MC 1988 Marinebacteriophages and bacterial mortality EOS Trans AmGeophys Union 691111ndash1112
213 BerghO BorsheimKY BratbakGHeldalM 1989Highabundance of viruses found in aquatic environmentsNature 340467ndash468
214 Borsheim KY Bratbak G Heldal M 1990 Enumerationand biomass estimation of planktonic bacteria and virusesby transmission electron microscopy Appl Environ Micro-biol 56352ndash356
215 Sharp GD 1949 Enumeration of virus particles by elec-tron micrography Proc Soc Exp Biol Med 7054ndash59
216 SullivanMBWaterbury JBChisholmSW 2003Cyano-phages infecting the oceanic cyanobacterium Prochloro-coccus Nature 4241047ndash1051
217 BorsheimKY 1993 Native marine bacteriophages FEMSMicrobiol Ecol 102141ndash159
218 Wommack KE Colwell RR 2000 Virioplanktonviruses in aquatic ecosystems Microbiolo Mol Biol Rev 6469ndash114
219 Brum J SchenckR SullivanM 2013Globalmorpholog-ical analysis of marine viruses shows minimal regional var-iation and dominance of non-tailed viruses ISME J 71738ndash1751
220 Chen F Lu JR Binder BJ Liu YC Hodson RE 2001Application of digital image analysis and flow cytometryto enumerate marine viruses stained with SYBR goldAppl Environ Microbiol 67539ndash545
221 Hennes KP Suttle CA 1995 Direct counts of viruses innatural waters and laboratory cultures by epifluorescencemicroscopy Limnol Oceanogr 401050ndash1055
222 Brussaard CPD 2004 Optimization of procedures forcounting viruses by flow cytometry Appl Environ Microbiol701506ndash1513
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-29
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
223 Fuhrman JA 1999 Marine viruses biogeochemical andecological effects Nature 399541ndash548
224 Fuhrman JA Suttle CA 1993 Viruses in marine plank-tonic systems Oceanography 651ndash63
225 Suttle CA Chan AM Cottrell MT 1990 Infection ofphytoplankton by viruses and reduction of primary produc-tivity Nature 387467ndash469
226 Suttle CA 2007 Marine virusesmdashmajor players in theglobal ecosystem Nature Rev Microbiol 5801ndash812
227 Bidle KD Vardi A 2011 A chemical arms race at seamediates algal host-virus interactions Curr Opin Microbiol14449ndash457
228 Proctor LM Fuhrman JA 1990 Viral mortality of marinebacteria and cyanobacteria Nature 34360ndash62
229 Proctor LM Okubo A Fuhrman JA 1993 Calibratingestimates of phage induced mortality in marine bacteriaultrastructural studies of marine bacteriophage develop-ment from one-step growth experiments Microb Ecol 25161ndash182
230 Weinbauer MG Peduzzi P 1995 Significance of virusesversus heterotrophic nanoflagellates for controlling bacte-rial abundance in the northern Adriatic Sea J PlanktonRes 171851ndash1856
231 Bratbak G Heldal M Thingstad TF Riemann B Has-lund OH 1992 Incorporation of viruses into the budgetof microbial C-transfer A first approach Mar Ecol ProgSer 83273ndash280
232 Steward GFWikner J Smith DC CochlanWP Azam F1992 Estimation of virus production in the sea I Methoddevelopment Mar Microb Food Webs 6(2)57ndash78
233 Proctor LM Fuhrman JA 1992 Mortality of marine bac-teria in response to enrichments of the virus size fractionfrom seawater Mar Ecol Prog Ser 87283ndash293
234 Suttle CA 1994 The significance of viruses to morta-lity in aquatic microbial communities Microb Ecol 28237ndash243
235 Wilhelm SW Brigden SM Suttle CA 2002 A dilutiontechnique for the direct measurement of viral productiona comparison in stratified and tidally mixed coastal watersMicrob Ecol 43168ndash173
236 Suttle CA 2005 Viruses in the seaNature 437356ndash361237 Weinbauer MG 2004 Ecology of prokaryotic viruses
FEMS Microbiol Rev 28127ndash181238 Baudoux AC Veldhuis MJW Noordeloos AAM van
Noort G Brussaard CPD 2008 Estimates of virus- vsgrazing-induced mortality of picophytoplankton in theNorth Sea during summer Aq Microb Ecol 5269ndash82
239 Bratbak G Egge JK Heldal M 1993 Viral mortalityof the marine alga Emiliana huxleyi (Haptophyceae)and termination of algal blooms Mar Ecol Prog Ser 9339ndash48
240 Bratbak G Jacobsen A Heldal M 1998 Viral lysis ofPhaeocystis pouchetti and bacterial secondary productionAq Microb Ecol 1611ndash16
241 Bratbak G Levasseur M Michaud S Cantin G Fernan-dez E Heimdal BR Heldal M 1995 Viral activity in rela-tion to Emiliania huxleyi bloomsmdasha mechanism of DMSPrelease Mar Ecol Prog Ser 128133ndash142
242 Bratbak G Wilson W Heldal M 1996 Viral control ofEmiliania huxleyi blooms J Mar Systems 975ndash81
243 Brussaard CPD Bratbak G Baudoux AC Ruardij P2007 Phaeocystis and its interaction with viruses Biogeo-chemistry 83201ndash215
244 Evans C Brussaard CPD 2012 Regional variation inlytic and lysogenic viral Infection in the Southern Oceanandits contribution to biogeochemical cycling Appl Envi-ron Microbiol 786741ndash6748
245 Gastrich MD Anderson OR Benmayor SS Cosper EM1998 Ultrastructural analysis of viral infection in the
brown-tide alga Aureococcus anophagefferens Phycologia37300ndash306
246 Suttle CA 1992 Inhibition of photosynthesis in phyto-plankton by the submicron size fraction concentratedfrom seawater Mar Ecol Prog Ser 87105ndash112
247 Suttle CA Chan AM 1995 Viruses infecting the marineprymnesiophyte Chrysochromulina spp isolation prelimi-nary characterization and natural abundance Mar EcolProg Ser 118275ndash282
248 Suttle CA Chan AM 1993 Marine cyanophages infect-ing oceanic and coastal strains of Synechococcus abun-dance morphology cross-infectivity and growthcharacteristics Mar Ecol Prog Ser 9299ndash109
249 Suttle CA Chan AM 1994 Dynamics and distribution ofcyanophages and their effect on marine Synechococcus sppAppl Environ Microbiol 603167ndash3174
250 Mann NH 2003 Phages of the marine cyanobacterialpicophytoplankton FEMS Microbiol Rev 2717ndash34
251 Ortmann AC Metzger RC Liefer JD Novoveska L2012 Grazing and viral lysis vary for different componentsof the microbial community across an estuarine gradientAq Microb Ecol 65143ndash157
252 Ory P HartmannHJ Jude F Dupuy C Del Amo Y Cat-ala P Mornet F Huet V Jan B Vincent D Sautour BMontanieacute H 2010 Pelagic food web patterns do theymodulate virus and nanoflagellate effects on picoplanktonduring the phytoplankton spring bloom Environ Microbiol122755ndash2772
253 Wommack KE Ravel J Hill RT Chun J Colwell RR1999 Population dynamics of Chesapeake Bay virioplank-ton total-community analysis by pulsed-field gel electro-phoresis Appl Environ Microbiol 65231ndash240
254 Steward GF Montiel JL Azam F 2000 Genome size dis-tributions indicate variability and similarities amongmarine viral assemblages from diverse environments Lim-nol Oceanogr 451697ndash1706
255 Fuhrman JA Griffith JF SchwalbachMS 2002 Prokary-otic and viral diversity patterns in marine plankton EcolRes 17183ndash194
256 Fuller NJ Wilson WH Joint IR Mann NH 1998Occurrence of a sequence in marine cyanophages similarto that of T4 g20 and its application to PCR-based detec-tion and quantification techniques Appl Environ Microbiol642051ndash2060
257 Comeau AM Krisch HM 2008 The capsid of the T4phage superfamily the evolution diversity and structureof some of the most prevalent proteins in the biosphereMol Biol Evol 251321ndash1332
258 Chow CE Fuhrman JA 2012 Seasonality and monthlydynamics ofmarinemyovirus communitiesEnvironMicrobiol142171ndash2183 doi 101111j1462-2920201202744x
259 Needham DM Chow CE Cram JA Sachdeva R ParadaA Fuhrman JA 2013 Short-term observations of marinebacterial and viral communities patterns connections andresilience ISME J 71274ndash1285
260 Hurwitz BL Deng L Poulos BT Sullivan MB2012 Evaluation of methods to concentrate and purifyocean virus communities through comparative replicatedmetagenomics Environ Microbiol 151428ndash1440
261 John SG Mendez CB Deng L Poulos B KauffmanAKM Kern S Brum J Polz MF Boyle EA SullivanMB 2011 A simple and efficient method for concentra-tion of ocean viruses by chemical flocculation EnvironMicrobiol Rep 3195ndash202
262 Duhaime MB Deng L Poulos BT Sullivan MB 2012Towards quantitative metagenomics of wild viruses andother ultra-low concentration DNA samples a rigorousassessment and optimization of the linker amplifica-tion method Environ Microbiol 142526ndash2537
422-30 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
263 Thurber RV HaynesM Breitbart MWegley L RohwerF 2009 Laboratory procedures to generate viral metage-nomes Nat Protoc 4470ndash483
264 Angly FE Felts B Breitbart M Salamon P Edwards RACarlson C Chan AM Haynes M Kelley S Liu HMahaffy JM Mueller JE Nulton J Olson R ParsonsR Rayhawk S Suttle CA Rohwer F 2006 The marineviromes of four oceanic regions PLoS Biol 42121ndash2131
265 Bench SRHansonTEWilliamson KE GhoshD Rado-sovich M Wang K Wommack KE 2007 Metagenomiccharacterization of Chesapeake Bay virioplankton ApplEnviron Microbiol 737629ndash7641
266 Rodriguez-Brito B Li LL Wegley L Furlan M Angly FBreitbart M Buchanan J Desnues C Dinsdale EEdwardsR Felts BHaynesM LiuH LipsonDMahaffyJ Martin-Cuadrado AB Mira A Nulton J Pasic L Ray-hawk S Rodriguez-Mueller J Rodriguez-Valera F Sala-mon P Srinagesh S Thingstad TF Tran T ThurberRV Willner D Youle M Rohwer F 2010 Viral andmicrobial community dynamics in four aquatic environ-ments ISME J 4739ndash751
267 Sullivan MB Coleman ML Weigele P Rohwer F Chis-holm SW 2005 Three Prochlorococcus cyanophagegenomes signature features and ecological interpretationsPLoS Biol 3790ndash806
268 Kang I Oh HM Kang D Cho JC 2013 Genome of aSAR116 bacteriophage shows the prevalence of thisphage type in the oceans Proc Natl Acad Sci USA 11012343ndash12348
269 Zhao YL Temperton B Thrash JC Schwalbach MSVergin KL Landry ZC Ellisman M Deerinck T Sulli-van MB Giovannoni SJ 2013 Abundant SAR11 virusesin the ocean Nature 494357ndash360
270 Hurwitz BL Westveld AH Brum JR Sullivan MB2014 Modeling ecological drivers in marine viral com-munities using comparative metagenomics and networkanalyses Proc Nat Acad Sci 11110714ndash10719
271 Brum JR Ignacio-Espinoza JC Roux S Doulcier G Aci-nas SG Alberti A Chaffron S Cruaud C de Vargas CGasol JM Gorsky G Gregory AC Guidi L HingampP Iudicone D Not F Ogata H Pesant S Poulos BTSchwenck SM Speich S Dimier C Kandels-Lewis SPicheral M Searson S Coordinators TO Bork P BowlerC Sunagawa S Wincker P Karsenti E Sullivan MB2015 Patterns and ecological drivers of ocean viral com-munities Science 348
272 Thingstad TF Lignell R 1997 Theoretical models for thecontrol of bacterial growth rate abundance diversity andcarbon demand Aq Microb Ecol 1319ndash27
273 Weinbauer MG Rassoulzadegan F 2004 Are virusesdriving microbial diversification and diversity EnvironMicrobiol 61ndash11
274 Thingstad TF Vage S Storesund JE Sandaa RA GiskeJ 2014 A theoretical analysis of how strain-specific virusescan control microbial species diversity Proc Natl Acad SciUSA 1117813ndash7818
275 Middelboe M HagstromA Blackburn N Sinn B FischerUBorchNHPinhassi JSimuKLorenzMG2001Effectsof bacteriophages on the population dynamics of four strainsof pelagic marine bacteriaMicrob Ecol 42395ndash406
276 Schwalbach MS Hewson I Fuhrman JA 2004 Viraleffects on bacterial community composition in marineplankton microcosms Aq Microb Ecol 34117ndash127
277 Mojica KD Brussaard CP 2014 Factors affecting virusdynamics and microbial host-virus interactions in marineenvironments FEMS Microbiol Ecol 89495ndash515
278 Forterre P Prangishvili D 2013 The major role of virusesin cellular evolution facts and hypothesesCurr Opin Virol3558ndash565
279 Jiang SC Paul JH 1994 Seasonal and diel abundanceof viruses and occurrence of lysogenybacteriocinogenyin the marine environment Mar Ecol Prog Ser 104163ndash172
280 Jiang SC Paul JH 1996 Occurrence of lysogenic bacteriain marine microbial communities as determined by pro-phage induction Mar Ecol Prog Ser 14227ndash38
281 Wilcox RM Fuhrman JA 1994 Bacterial viruses incoastal seawater lytic rather than lysogenic productionMar Ecol Prog Ser 11435ndash45
282 Paul JH 2008 Prophages in marine bacteria dangerousmolecular time bombs or the key to survival in the seasISME J 2579ndash589
283 Brum JR Hurwitz BL Schofield O Ducklow HW Sul-livan MB 2015 Seasonal time bombs dominant temper-ate viruses affect Southern Ocean microbial dynamicsISME J doi 101038ismej2015125
284 Noble RT Fuhrman JA 1997 Virus decay and its causesin coastal waters Appl Environ Microbiol 6377ndash83
285 Gonzaacutelez JM Suttle CA 1994 Grazing by marine nano-fiagellates on viruses and virus-sized particles ingestionand digestion Mar Ecol Prog Ser 941ndash10
286 Middelboe M Jorgensen NOG Kroer N 1996 Effects ofviruses on nutrient turnover and growth efficiency of non-infected marine bacterioplankton Appl Environ Microbiol621991ndash1997
287 Noble RT Fuhrman JA 1999 Breakdown and microbialuptake ofmarine viruses and other lysis productsAqMicrobEcol 201ndash11
288 Townsend DW Cammen LM 1988 Potential impor-tance of the timing of spring plankton blooms to benthic-pelagic coupling and recruitment of juvenile demersalfishes Biol Oceanog 5215ndash228
289 Goldman JC Carpenter EJ 1974 A kinetic approach tothe effect of temperature on algal growth Limnol Oceanogr19756ndash766
290 SmithWO Jr Gordon LI 1997 Hyperproductivity of theRoss Sea (Antarctica) polynya during austral spring Geo-phys Res Lett 24233ndash236
291 Caron DA Hutchins DA 2013 The effects of changingclimate on microzooplankton grazing and communitystructure drivers predictions and knowledge gaps J Plank-ton Res 35235ndash252
292 Caron DA Dennett MR Lonsdale DJ Moran DM Sha-lapyonok L 2000 Microzooplankton herbivory in theRoss Sea Antarctica Deep Sea Res 4715ndash16
293 DennettMRMathot S CaronDA SmithWO LonsdaleDJ 2001 Abundance and distribution of phototrophicand heterotrophic nano- and microplankton in the south-ern Ross Sea Deep Sea Res 484019ndash4037
294 Jannasch HW Eimhjellen K Wirsen CO Farmanfar-maian A 1971 Microbial degradation of organic matterin the deep sea Science 171672ndash675
295 Jannasch HW Wirsen CO 1973 Deep-sea microorgan-isms in situ response to nutrient enrichment Science180641ndash643
296 Gooday AJ Lambshead PJD 1989 Influence of season-ally deposited phytodetritus on benthic foraminiferal pop-ulations in the bathyal Northeast Atlantic the speciesresponse Mar Ecol Prog Ser 5853ndash67
297 Eloe EA Malfatti F Gutierrez J Hardy K Schmidt WEPogliano K Pogliano J Azam F Bartlett DH 2011Isolation and characterization of a psychropiezophilicalphaproteobacterium Appl Environ Microbiol 778145ndash8153
298 Kato C Bartlett DH 1997 The molecular biology of bar-ophilic bacteria Extremophiles 1111ndash116
299 Burnett BR 1977 Quantitative sampling of microbiota ofthe deep-sea benthosmdashI Sampling techniques and some
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-31
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
data from the abyssal central North Pacific Deep Sea Res24781ndash789
300 Burnett BR 1981 Quantitative sampling of microbiota ofthe deep-sea benthosmdashIII The bathyal San Diego TroughDeep Sea Res 28A649ndash663
301 Atkins MS Teske AP Anderson OR 2000 A survey offlagellate diversity at four deep-sea hydrothermal vents inthe eastern Pacific Ocean using structural and molecularapproaches J Euk Microbiol 47400ndash411
302 Turley CM GoodayAJ Green JC 1993 Maintenance ofabyssal benthic foraminifera under high pressure and lowtemperature some preliminary results Deep Sea Res 40643ndash652
303 Turley CM Lochte K Patterson DJ 1988 A barophilicflagellate isolated from 4500 m in the mid-North AtlanticDeep Sea Res 351079ndash1092
304 Weinberg JR 1990 High rates of long-term survival ofdeep-sea infauna in the laboratory Deep Sea Res 371375ndash1379
305 Buck KR Barry JP Simpson AGB 2000 Monterey Baycold seep biota euglenozoa with chemoautotrophic bacte-rial epibionts Europ J Protistol 36117ndash126
306 Cho BC Na SC Choi DH 2000 Active ingestion of flu-orescently labeled bacteria by mesopelagic heterotrophicnanoflagellates in the East Sea Korea Mar Ecol Prog Ser20623ndash32
307 Turley CM Carstens M 1991 Pressure tolerance of oce-anic flagellates implications for remineralization of organicmatter Deep Sea Res 38403ndash413
308 Edgcomb V Orsi W Taylor GT Vdacny P Taylor CSuarez P Epstein S 2011 Accessing marine protistsfrom the anoxic Cariaco Basin ISME J 51237ndash1241
309 Edgcomb VP Kysela DT Teske A Gomez AD SoginML 2002 Benthic eukaryotic diversity in the GuaymasBasin hydrothermal vent environment Proc Natl Acad SciUSA 997658ndash7662
310 Moreira D Lopez-Garcia P 2003 Are hydrothermal ventsoases for parasitic protistsTrends inParasitology19556ndash558
311 Orsi W Edgcomb V Faria J Foissner W Fowle WHHohmann T Suarez P Taylor C Taylor GT VdrsquoacnyacuteP Epstein SS 2012 Class Cariacotrichea a novel ciliatetaxon from the anoxic Cariaco Basin Venezuela Int JSyst Evol Microbiol 621425ndash1433
312 Turley CM Lochte K 1990 Microbial response to theinput of fresh detritus to the deep-sea bed Palaeogeog Palae-oclimatol Palaeoecol 893ndash23
313 CroftMTWarrenMJ Smith AG 2006 Algae need theirvitamins Eukar Cell 51175ndash1183
314 Williams PJL 2000 Heterotrophic bacteria and thedynamics of dissolved organic matter pp 153ndash200 InKirchman DL (ed)Microbial Ecology of the Oceans Wiley-Liss New York NY
315 ZubkovMV FuchsBMTarranGA Burkill PHAmannR 2003 High rate of uptake of organic nitrogen com-pounds by Prochlorococcus cyanobacteria as a key to theirdominance in oligotrophic oceanic waters Appl EnvironMicrobiol 691299ndash1304
316 ZubkovMV Tarran GA Fuchs BM 2004 Depth relatedamino acid uptake by Prochlorococcus cyanobacteria in theSouthern Atlantic tropical gyre FEMS Microbiol Ecol 50153ndash161
317 Silver MW Alldredge AL 1981 Bathypelagic marinesnow deep-sea algal and detrital community J Mar Res39501ndash530
318 RiebesellU 1992 The formation of largemarine snowandits sustained residence in surface waters Limnol Oceanogr3763ndash76
319 Alldredge AL 1976 Discarded appendicularian houses assources of food surface habitats and particulate organic
matter in planktonic environments Limnol Oceanogr 2114ndash23
320 Herndl GJ Peduzzi P 1988 The ecology of amorphousaggregations (marine snow) in the Northern AdriaticSea PSZNI Mar Ecol 979ndash90
321 Kioslashrboe T Andersen KP Dam HG 1990 Coagulationefficiency and aggregate formation in marine phytoplank-ton Mar Biol 107235ndash245
322 Caron DA Madin LP Davis PG Sieburth JM 1982Marine snow as a micro-environment for protozoan growthin oceanic plankton communities J Protozool 29484ndash485
323 Caron DA Davis PG Madin LP Sieburth JM 1986Enrichment of microbial populations in macroaggregates(marine snow) from the surface waters of the North Atlan-tic J Mar Res 44543ndash565
324 Davoll PJ Silver MW 1986 Marine snow aggregates lifehistory sequence and microbial community of abandonedlarvacean houses from Monterey Bay California MarEcol Prog Ser 33111ndash120
325 Azam F Worden AZ 2004 Oceanography microbesmolecules and marine ecosystems Science 3031622ndash1624
326 Goldman JC 1984 Conceptual role for microaggregates inpelagic waters Bull Mar Sci 35462ndash476
327 Elser JJ Loladze I Peace AL Kuang Y 2012 Lotkare-loaded modeling trophic interactions under stoichio-metric constraints Ecol Model 2453ndash11
328 Caron DA 1994 Inorganic nutrients bacteria and themicrobial loop Microb Ecol 28295ndash298
329 Caron DA Goldman JC 1990 Protozoan nutrient regen-eration pp 283ndash306 In Capriulo GM (ed) Ecology ofMarine Protozoa Oxford University Press New York NY
330 KirchmanDL 2000Uptake and regeneration of inorganicnutrients by marine heterotrophic bacteria pp 261ndash288 InKirchman DL (ed)Microbial Ecology of the Oceans WileyNew York NY
331 Wheeler PAKirchmanDL 1986Utilization of inorganicand organic nitrogen by bacteria in marine systems LimnolOceanogr 31998ndash1099
332 Fuhrman JAHorrigan SG CaponeDG 1988 The use of13N as tracer for bacterial and algal uptake of ammoniumfrom seawater Mar Ecol Prog Ser 45271ndash278
333 Caron DA Lim EL Sanders RW Dennett MR Ber-ninger UG 2000 Responses of bacterioplankton and phy-toplankton to organic carbon and inorganic nutrientadditions in contrasting oceanic ecosystems Aq MicrobEcol 22175ndash184
334 Cotner JB Ammerman JW Peele ER Bentzen E 1997Phosphorus-limited bacterioplankton growth in the Sar-gasso Sea Aq Microb Ecol 13141ndash149
335 Carlsson P Caron DA 2001 Seasonal variation of phos-phorus limitation of bacterial growth in a small lake LimnolOceanogr 46108ndash120
336 Currie DJ Kalff J 1984 The relative importance of bac-terioplankton and phytoplankton in phosphorus uptakein freshwater Limnol Oceanogr 29311ndash321
337 Capone DG Zehr JP Paerl HW Bergman B CarpenterEJ 1997 Trichodesmium a globally significant marine cya-nobacterium Science 2761221ndash1229
338 Zehr JP Waterbury JB Turner PJ Montoya JP Omore-gie E StewardGFHansenAKarlDM 2001Unicellularcyanobacteria fix N-2 in the subtropical North PacificOcean Nature 412635ndash638
339 Jenkins BD Steward GF Short SM Ward BB Zehr JP2004 Fingerprinting diazotroph communities in the Ches-apeake Bay by using a DNA macroarray Appl EnvironMicrobiol 701767ndash1776
340 Tripp HJ Bench SR Turk KA Foster RA Desany BANiazi F Affourtit JP Zehr JP 2010 Metabolic
422-32 AQUATIC ENVIRONMENTS
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
streamlining in an open-ocean nitrogen-fixing cyanobacte-rium Nature 46490ndash94
341 Thompson AW Foster RA Krupke A Carter BJ MusatN Vaulot D Kuypers MMM Zehr JP 2012 Unicellularcyanobacterium symbiotic with a single-celled eukaryoticalga Science 3371546ndash1550
342 Kirchman DL Meon B Cottrell MT Hutchins DAWeeks D Bruland KW 2000 Carbon versus iron limita-tion of bacterial growth in the California upwelling regimeLimnol Oceanogr 451681ndash1688
343 Martin JH Fitzwater SE 1988 Iron deficiency limits phy-toplankton growth in the north-east Pacific subarcticNature 331341ndash344
344 Kirchman DL Hoffman KA Weaver R Hutchins DA2003 Regulation of growth and energetics of a marine bac-terium by nitrogen source and iron availability Mar EcolProg Ser 250291ndash296
345 Barbeau K Moffett JW Caron DA Croot PL ErdnerDL 1996 Role of protozoan grazing in relieving iron lim-itation of phytoplankton Nature 38061ndash64
346 Chase Z PriceNM 1997Metabolic consequences of irondeficiency in heterotrophic marine protozoa Limnol Oce-anogr 421673ndash1684
347 Gruber N 2011 Warming up turning sour losing breathocean biogeochemistry under global change Phil TransRoyal Soc A Math Phys Eng Sci 3691980ndash1996
348 Edgcomb V Orsi W Bunge J Jeon S Christen R LeslinC Holder M Taylor GT Suarez P Varela R Epstein S2011 Protistanmicrobial observatory in the Cariaco BasinCaribbean I Pyrosequencing vs Sanger insights into spe-cies richness ISME J 51344ndash1356
349 EdgcombVOrsiW Leslin C Epstein S Bunge J Jeon SYakimov M Behnke A Stoeck T 2009 Protistan com-munity patterns within the brine and halocline of deephypersaline anoxic basins in the eastern MediterraneanSea Extremophiles 13151ndash167
350 Schnetzer A Moorthi SD Countway PD Gast RJ GilgIC Caron DA 2011 Depth matters microbial eukaryotediversity and community structure in the eastern NorthPacific revealed through environmental gene librariesDeep Sea Res 5816ndash26
351 Stoeck T Behnke A Christen R Amaral-Zettler LARodriguez-Mora MJ Chistoserdov A Orsi W EdgcombVP 2009 Massively parallel tag sequencing reveals thecomplexity of anaerobic marine protistan communitiesBMC Biol 772
352 Buck KR Bernhard JM 2001 Protistan-prokaryotic sym-bioses in deep-sea sulfidic sediments pp 507ndash517 In Seck-bach J (ed) Symbiosis Kluwer Academic Dordrecht TheNetherlands
353 Fenchel T Finlay BJ 1994 The evolution of life withoutoxygen Am Sci 8222ndash29
354 Muumlller M 1980 The hydrogenosome pp 127ndash142 InGooday GW LLoyd D Trinci APJ (eds) The EukaryoticMicrobialCellCambridgeUniversityPressCambridgeUK
355 Strom SL 2000 Bacterivory interactions betweenbacteria and their grazers pp 351ndash386 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
356 Landry MR Kirshtein J Constantinou J 1995 A refineddilution technique for measuring the community grazingimpact of microzooplankton with experimental tests inthe central equatorial PacificMarEcol Prog Ser12053ndash63
357 Rublee PA Gallegos CL 1989 Use of fluorescentlylabelled algae (FLA) to estimatemicrozooplankton grazingMar Ecol Prog Ser 51221ndash227
358 Sherr BF Sherr EB Fallon RD 1987 Use of mono-dispersed fluorescently labeled bacteria to estimate in situprotozoan bacterivoryAppl EnvironMicrobiol 53958ndash965
359 Sherr BF Sherr EB Hopkinson CS 1988 Trophic inter-actions within pelagic microbial communities indicationsof feedback regulation of carbon flow Hydrobiologia 15919ndash26
360 FenchelT 1982 Ecology of heterotrophicmicroflagellatesII Bioenergetics and growthMar Ecol Prog Ser 8225ndash231
361 Holling CS 1959 Some characteristics of simple types ofpredation and parasitism Can Entomol 91385ndash398
362 Rice TDWilliamsHNTurng BF 1998 Susceptibility ofbacteria in estuarine environments to autochthonous bdel-lovibrios Microb Ecol 35256ndash264
363 Ravenschlag K Sahm K Pernthaler J Amann R 1999High bacterial diversity in permanently cold marine sedi-ments Appl Environ Microbiol 653982ndash3989
364 Martin MO 2002 Predatory prokaryotes an emerg-ing research opportunity J Mol Microbiol Biotechnol 4467ndash477
365 Cavanaugh CM 1994 Microbial symbiosismdashpatterns ofdiversity in the marine environment Am Zool 3479ndash89
366 McFall-Ngai MJ 2000 Negotiations between animalsand bacteria the ldquodiplomacyrdquo of the squid-vibrio symbiosisComp Biochem Physiol A Mol Integr Physiol 126471ndash480
367 Gast RJ Sanders RW Caron DA 2009 Ecological strat-egies of protists and their symbiotic relationships with pro-karyotic microbes Trends Microbiol 17563ndash569
368 GordonN Angel DL Neori A Kress N Kimor B 1994Heterotrophic dinoflagellates with symbiotic cyanobacte-ria and nitrogen limitation in the Gulf of Aqaba MarEcol Prog Ser 10783ndash88
369 Coats DW 1999 Parasitic life styles of marine dinoflagel-lates J Euk Microbiol 46402ndash409
370 Coats DW Park MG 2002 Parasitism of photosyntheticdinoflagellates by three strains of Amoebophrya (Dino-phyta) parasite survival infectivity generation time andhost specificity J Phycol 38520ndash528
371 Guillou L Viprey M Chambouvet A Welsh RM Kirk-ham AR Massana R Scanlan DJ Worden AZ 2008Widespread occurrence and genetic diversity ofmarine par-asitoids belonging to Syndiniales (Alveolata) EnvironMicrobiol 103349ndash3365
372 Kuumlhn SF 1998 Infection ofCoscinodiscus spp by the para-sitoid nanoflagellate Pirsonia diadema II Selective infec-tion behaviour for host species and individual host cells JPlankton Res 20443ndash454
373 ParkMGCooney SKKim JSCoatsDW 2002 Effects ofparasitism on diel vertical migration phototaxisgeotaxisand swimming speed of the bloom-forming dinoflagellateAkashiwo sanguinea Aq Microb Ecol 2911ndash18
374 ParkMG Cooney SK YihW Coats DW 2002 Effects oftwo strains of the parastic dinoflagellate Amoebophrys ongrowth photosynthesis light absorption and quantumyield of bloom-forming dinoflagellates Mar Ecol Prog Ser227281ndash292
375 LongRA Azam F 2001 Antagonistic interactions amongmarine pelagic bacteria Appl Environ Microbiol 674975ndash4983
376 Roy JS Poulson-Ellestad KL Drew Sieg R Poulin RXKubanek J 2013 Chemical ecology of the marine plank-ton Nat Prod Rep 301364ndash1379
377 Imai I Ishida Y Sakaguchi K Hata Y 1995 Algicidalmarine bacteria isolated from northern Hiroshima BayJapan Fish Sci 61628ndash636
378 Fistarol GO Legrand C Graneli E 2003 Allelopathiceffect of Prymnesium parvum on a natural plankton com-munity Mar Ecol Prog Ser 255115ndash125
379 Hulot FD Huisman J 2004 Allelopathic interactionsbetween phytoplankton species the roles of heterotrophicbacteria and mixing intensity Limnol Oceanogr 491424ndash1434
Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa 422-33
Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-

Downloaded from wwwasmscienceorg by
IP 6620862130
On Thu 03 Mar 2016 185215
380 Strom SWolfe GV Slajer A Lambert S Clough J 2003Chemical defenses in the microplankton II inhibition ofprotist feeding by B-dimethylsulfoniopropionate (DMSP)Limnol Oceanogr 48230ndash237
381 Turner JT Tester PA 1997 Toxic marine phytoplank-ton zooplankton grazers and pelagic food webs LimnolOceanogr 421203ndash1214
382 Wolfe GV Steinke M Kirst GO 1997 Grazing-activatedchemical defense in a unicellular marine algaNature 387894ndash897
383 del Giorgio PA Cole JJ 2000 Bacterial energeticsand growth efficiency pp 289ndash325 In Kirchman DL(ed) Microbial Ecology of the Oceans Wiley-LissNew York NY
384 Hansen PJ Bjoslashrnsen PK Hansen BW 1997 Zooplank-ton grazing and growth scaling within the 2ndash2000-micrombody size range Limnol Oceanogr 42687ndash704
385 Azam F 1998 Microbial control of oceanic carbon fluxthe plot thickens Science 280694ndash696
386 Stocker R 2012 Marine microbes see a sea of gradientsScience 338628ndash633
387 Karl DM 2007 Microbial oceanography paradigms proc-esses and promise Nat Rev Microbiol 5759ndash769
388 Chow C-ET Kim DY Sachdeva R Caron DA FuhrmanJA 2014 Top-down controls on bacterial communitystructure microbial network analysis of bacteria T4-likeviruses and protists ISME J 8816ndash829
389 Fuhrman JA Steele JA 2008 Community structure ofmarine bacterioplankton patterns networks and relation-ships to function Aq Microb Ecol 5369ndash81
390 Gilbert JA Steele J Caporaso JG Steinbruumlck L ReederJ TempertonBHuse S Joint IMcHardyACKnightRSomerfield P Fuhrman JA Field D 2011 Defining sea-sonal marine microbial community dynamics ISME J 6298ndash308
391 Lima-Mendez G Faust K Henry N Decelle J Colin SCarcillo F Chaffron S Ignacio-Espinosa JC Roux SVincent F Bittner L Darzi Y Wang J Audic S BerlineL Bontempi G Cabello AM Coppola L Cornejo-Castillo FM drsquoOvidio F De Meester L Ferrera I Garet-Delmas M-J Guidi L Lara E Pesant S Royo-Llonch MSalazar G Saacutenchez P Sebastian M Souffreau C DimierC Picheral M Searson S Kandels-Lewis S coordinatorsTO Gorsky G Not F Ogata H Speich S Stemmann LWeissenbach J Wincker P Acinas SG Sunagawa SBork P Sullivan MB Karsenti E Bowler C de VargasC Raes J 2015 Determinants of community structure inthe global plankton interactome Science 348126073
392 Zehr JP Kudela RM 2011 Nitrogen cycle of the openocean from genes to ecosystems Annu Rev Mar Sci 3197ndash225
393 Zehr JP Ward BB 2002 Nitrogen cycling in the oceannew perspectives on processes and paradigms Appl EnvironMicrobiol 681015ndash1024
422-34 AQUATIC ENVIRONMENTS
- Heterotrophic Planktonic Microbes Virus Bacteria Archaea and Protozoa
-
- Background and Historical Development
-
- Marine Microbial Ecology into the 1970s
- The Microbial Loop Revolution
- Definitions and Concepts
- Estimating ``Bacterial Biomass and ``Bacterial Production Definitions and Methods
- Geographic and Temporal Distributions of Microbial Biomass and Activity
- The Changing and Complex World of Eukaryote Phylogeny
-
- Bacteria and Archaea
-
- ``Culturable versus ``Nonculturable Cells
- Molecular Phylogeny and Metagenomics Field Applications
- Culture-Independent Diversity Studies
- Molecular Genetic Discoveries in Bacterial and Archaeal Marine Biology
- Dilution Cultures of Bacteria and Archaea
-
- Protozoa
-
- Diversity and Distribution
- Diversity and Biogeography of Protists From Morphology to DNA Sequences
- Life Histories and Ecological Strategies of Protozoa
-
- Viruses
-
- Viral Abundance and General Properties
- Viral Metagenomics
- Viruses and Host Diversity
- Viruses and the Microbial Loop
-
- Major Environmental Controls
-
- Light Temperature and Pressure
- Dissolved and Particulate Organic Matter
- Inorganic Nutrients
-
- Macronutrients (N P)
- Micronutrients (Trace Metals Growth Factors)
- Oxygen
-
- Major Biotic Controls
-
- The Trophic Activities of Protozoa Predation and Top-Down Controls
- Species-Specific Interactions (Mutualism Parasitism Commensalisms)
- Balancing Microbial Growth and Removal
-
- The Microbial Loop Revisited
- Research Trends Including Microbial Association Networks
- References
-