h/d exchange mass spectrometry: applications in antibody
TRANSCRIPT

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 1
H/D Exchange Mass Spectrometry: Applications in Antibody Aggregation and Disordered Proteins
David Weis
Department of Chemistry
Instrumentation
Bio-pharmaceuticsCrowding
McGuffee, & Elcock, PLoSComput Biol 2010, 6, e1000694.
Disordered proteins
Ligand discovery
David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 2
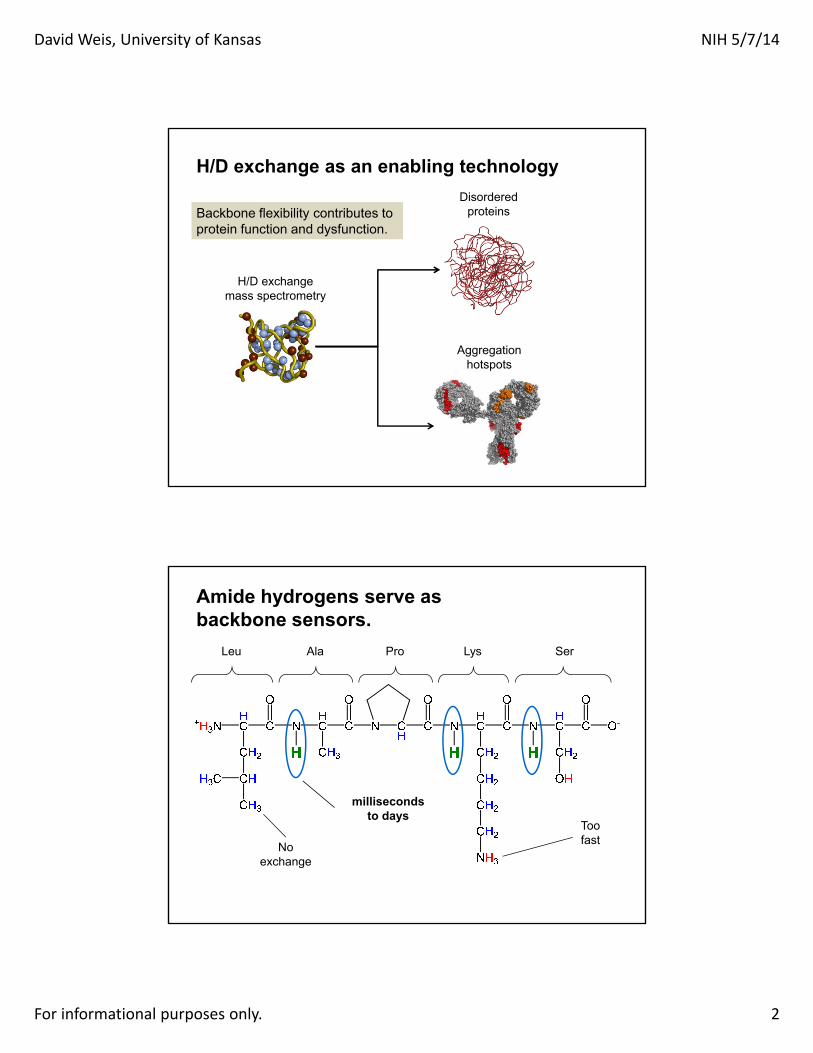
H/D exchange as an enabling technology
Backbone flexibility contributes toprotein function and dysfunction.
Disordered proteins
Aggregation hotspots
H/D exchangemass spectrometry
Amide hydrogens serve as backbone sensors.
Leu Ala Pro Lys Ser
Noexchange
Toofast
millisecondsto days

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 3
Major Histocompatibilty Complex (MHC) proteinhttp://www.youtube.com/watch?v=Y79Xl0LfYI4
H/D exchange reports on backbone conformation and dynamics.
O NH O O
kop
kcl
OD−
OH−
kint
opobs int
cl
kk k
k
Kaj Linderstrøm-LangCarlsberg Laboratory
Conformationand dynamics
Intrinsic

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 4
H/D exchange kinetics probes backbone dynamics.
D2O D2O
Flexible regions exchange rapidly
Rigid regions exchange slowly
MS approach uses quench and proteolysis.
Undeuterated
Quench
0 °CpH 2.5
Labeling
D2O
Deuterated
ProteolysisPepsin
Mass Analysis

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 5
Pepsin digestion gives localized coverage.
Average length: 14 residuesTotal coverage: 85%
676 680 684
m/z
Undeuterated
5 sec
4 hr
Peptides progressively gain mass.
Deuterium uptake curve
MassIncrease
(Da)
D2O Exposure (s)
100 101 102 103 104
0
3
6
9
Bound
Free

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 6
Efficient and robust platformsare now available
Informatics
TOF-MS
HPLC
Automation
Year
Weeks
Disordered proteins
Aggregation hotspots
H/D exchangemass spectrometry
H/D exchange as an enabling technology

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 7
Acid blobsNegative noodles
Intrinsically disorderedIntrinsically unstructured
Natively denaturedNatively unfolded
Mostly unstructuredFloppy
Dynamic
The disorder continuum
PONDR:Predictor Of Natively Disordered Regions
Romero, P.; Obradovic, Z.; Li, X.; Garner, E. C.; Brown, C. J.; Dunker, A. K. Proteins 2001, 42, 38-48. www.pondr.com
Spa32 (S. flexneri)
Residue Number
0 50 100 150 200 250 300
PO
ND
R s
core
0.0
0.2
0.4
0.6
0.8
1.0
Disordered
Ordered
David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 8
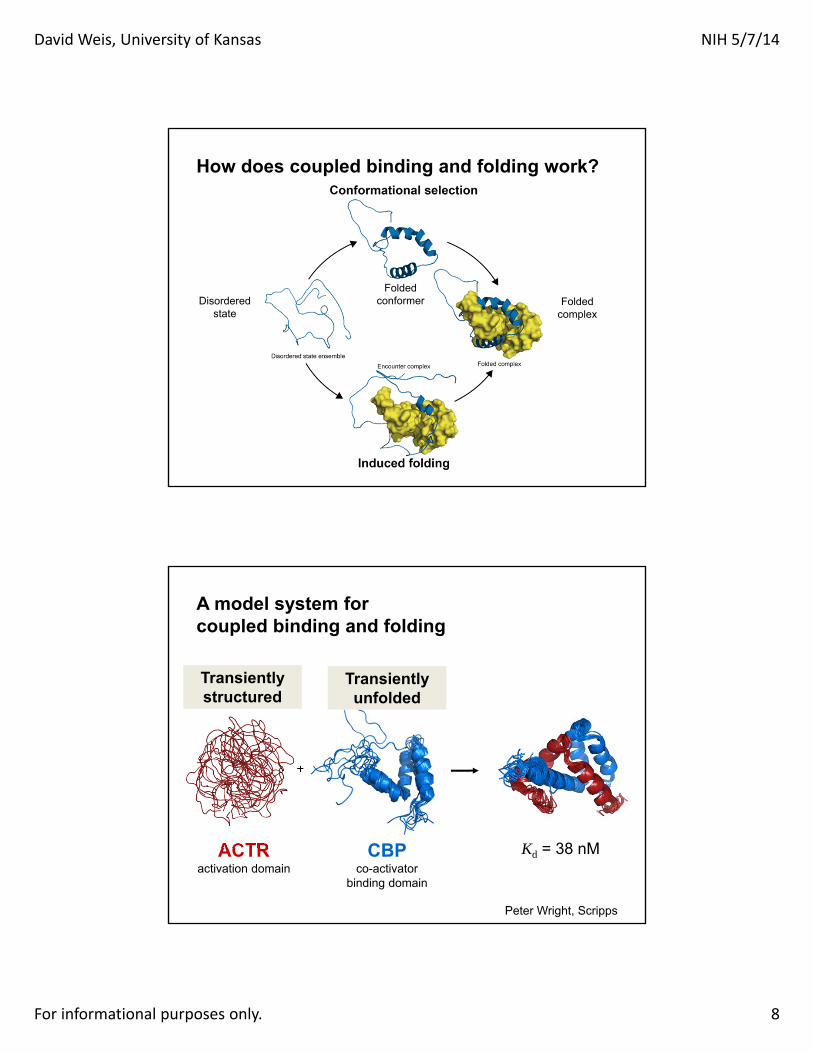
How does coupled binding and folding work?
Disorderedstate
Foldedconformer Folded
complex
CBPco-activator
binding domain
ACTRactivation domain
Kd = 38 nM
+
Unstructured Molten globule
A model system for coupled binding and folding
Transiently structured
Transiently unfolded
Peter Wright, Scripps

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 9
The speed limit of H/D exchange
R A L L D Q L
0
1
2
3
kint
(s−1)
int
ex
kPF
kProtection factor:
maximum rate
measured rate
D2O Exposure (s)
10-3 10-2 10-1 100 101 102 103 104
Deu
teri
um
Lev
el (
Da)
0
1
2
3
4
5
Kinetic analysis of H/D exchange
CBP ACTR
free
bound
1 ktD t N e

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 10
Mapping protection factors
G T Q N R P L L R N S L D D L V G P P S N L E G Q S D E R A L L D Q L H T L L S N T D A T G L E E I D R A L G I P E L V N Q G Q A L E P K Q D
1-12 36-47 60-7113-22 48-59
Aα1 Aα2 Aα3
33-42
29-3533-47
00 0
33-47
36-4729-35
33-42
48-5960-711-12
13-22
P N R S I S P S A L Q D L L R T L K S P S S P Q Q Q Q Q V L N I L K S N P Q L M A A F I K Q R T A K Y V A N Q P G M Q
Cα1
1-14 30-38 43-5931-40
431-10 17-30 40-
Cα2 Cα3000
1-141-10 17-30
30-38 43-5931-40
40- 43
<33-9
9-2020-50
>50
PF
Free
Complex
Complex
Free
CBP
ACTR
CBP is a molten globule
NMR secondary shifts
Ebert, Bae, Dyson, & Wright Biochemistry 2008, 47, 1299.
Free CBP
Kjaergaard, Teilum, & Poulsen, PNAS 2010, 107, 12535 (2KKJ)
Structure by NMR
∆Gu = 1.5 kcal mol−1
~8% Unstructured

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 11
CBP transiently populates an unfolded state.
Can H/D exchange reveal transiently foldedstates?

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 12
Exchange is too fast
10−3 10−2 10−1 100 101 102 103 104
Exchange time (sec)
Millisecond quench-flow H/D exchange
40-3500 msec

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 13
Kinetic analysis of exchangeby an unstructured protein
Exchange (s)
0.0 3.5
Deu
tera
tio
n (
%)
0
50
100
1017-10241029-10381039-1048 1045-10621059-10631059-10631064-10661072-1080 1081-1087
t50%
Peptide-level half-life reveals residual structure
α-helix α-helixabsentSecondary structure in complex

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 14
Localizing structure by residue-level averaging
it
2
2
j jn
ij
n
t Lt
L
weighted half-life
normalization factor
t1t2t3t4
Localized half-life correlates well withresidual structure.
Kjaergaard, 2010
α-helix α-helixabsent
H/D exchange
NMR
Helical propensity AGADIR

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 15
H/D exchange can capture both extremes of disorder
ACTR
CBP
Molten globule
Unstructured
Disordered proteins
Aggregation hotspots
H/D exchangemass spectrometry
H/D exchange as an enabling technology

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 16
49 biologics
There has been rapid growth in the useof therapeutic monoclonal antibodies.
0
31
1985 2011
$20 billionin sales
FDA approvedmAbs

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 17
mAbs are the Cadillacs of biotherapeutics.
• 150 kDa• IgG1• Glycosylated• 12 disulfide bonds• 50 mg/mL, pH 6
Antigenbinding
SS
SS
Antigenbinding
VH
VL
CL
CH1
CH2
CH3
VH
VL
CL
CH1
CH2
CH3
glycan glycan
Maintaining the physical stability ofprotein therapeutics is a critical problem.
Solution: Develop a stabilizing formulation.
Loss of efficacy
Immunogenicity
Conformationalinstability
Aggregation
David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 18
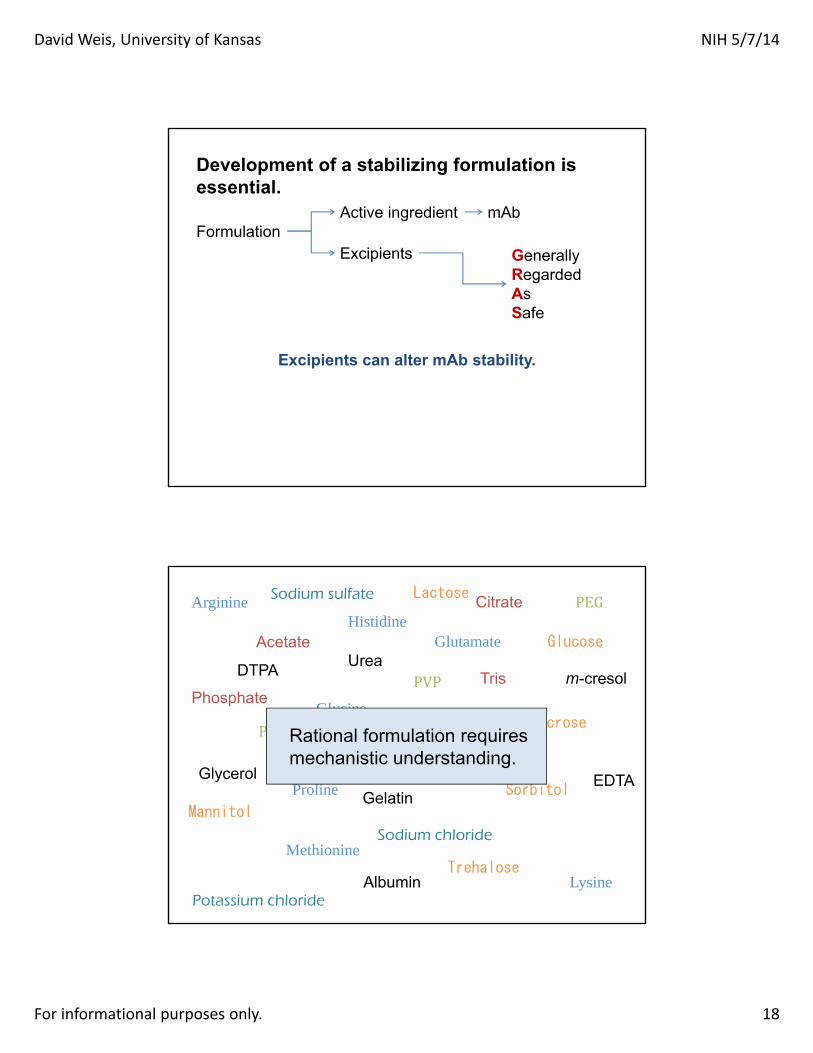
Development of a stabilizing formulation is essential.
GenerallyRegardedAsSafe
Formulation Active ingredient
Excipients
mAb
Excipients can alter mAb stability.
Citrate
Acetate Histidine
Phosphate
Arginine
Glycine
Proline
Lysine
Methionine
Sucrose
Trehalose
Sorbitol
Glutamate
Glycerol
Urea
Mannitol
Glucose
Lactose
Albumin
Gelatin
PVP
PLGA
PEG
Sodium chloride
Potassium chloride
Sodium sulfate
Polysorbate
EDTA
DTPA
Ethanol
m-cresol Tris
Benzylalcohol Rational formulation requires
mechanistic understanding.

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 19
thiocyanatearginine
Thermal stability Aggregation Backbone dynamics
Do local dynamics correlate with stability?
chloride sucrosesulfate
mAb-BIgG1
Sucrose increased thermal stability.Arginine decreased thermal stability.
Temperature (°C)
Cp
(cal
mol
−1
K−
1)
sucrose+6 °C
Temperature (°C)
arginine−4 °C

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 20
mAb-B aggregated at 40 °Cunder accelerated storage conditions.
8.0 9.0 10.0 11.0 12.0 13.0 14.0 15.0 min
Ab
sorb
ance
(28
0 n
m)
Dimer
Monomer
FragmentsMultimer
0.5 M NaSCN40 °C
initial
2 months
Sulfate slowed aggregation.Thiocyanate accelerated aggregation.
0 2 4 6 8 10 12
30
40
50
60
70
80
90
100
Monomer(%)
4 C
25 C
Months
NaSCN
Na2SO4
Control
40 °C

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 21
Stabilizers and destabilizersacted as expected.
∆Tm1 Aggregation
thiocyanate −9.0 °C Faster (++)
arginine −1.9 °C Faster (+)
chloride +0.3 °C Negligible
sucrose +1.5 °C Slower (−)
sulfate +1.8 °C Slower (−)How do these excipients workat the molecular level?
The effects of excipients are not uniform.
∆m
CH1 CH2 CH2
CH2 VL VL
Arg, 0.5 M
NaCl, 0.1 M
sucrose, 0.5 M

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 22
The effects are excipient-dependent.
Peptide (N to C)
±0.59 Da (99% CI)120 s
103 s
104 s
105 s
Differentialdeuteriumuptake (Da)
arginine, 0.5 M sucrose, 0.5 M
Arg+SCN−
sucrose SO
faster
slowerno effect
no data
The correlation between stability andaltered H/D exchange is not obvious.
Homology model based on [Saphire, 2001] 1HZH

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 23
SCN− SO
Destabilizer Stabilizer
Extremes have nearly-identical effects.
Arg+SCN−
sucrose SO
faster
slowerno effect
no data
The correlation with altered H/D exchangeis not obvious.

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 24
Arg+SCN−
DestabilizerDestabilizer
Destabilizers have very different effects.
A hydrophobic segment of the CH2 domainmay mediate aggregation.
CH3 CH3
CH2 CH2
VFLFPPKPDTLMI
Destabilizers and oxidation increased backbone flexibility. [Houde, 2010]
Protein A binding inhibits aggregation. [Zhang, 2012]
Disulfide bond increased thermal stability. [Gong, 2009]

David Weis, University of Kansas NIH 5/7/14
For informational purposes only. 25
New insights into function and dysfunction
Increased flexibility accelerates aggregation
H/D exchange
Detecting transient states in disordered proteins
CAREERMCB-1149538
University of Kansas: David Volkin & Russ Middaugh
MedImmune: Hardeep Samra, Hasige Sathish, Steven Bishop, Prakash Manikwar