handbook of enology (the microbiology of wine and vinifications) || biochemistry of alcoholic...
TRANSCRIPT

2
Biochemistry of Alcoholic Fermentationand Metabolic Pathways of Wine Yeasts
2.1 Introduction 532.2 Sugar degradation pathways 542.3 Regulation of sugar-utilizing metabolic pathways 612.4 Metabolism of nitrogen compounds 69
2.1 INTRODUCTION
The synthesis of living material is endergonic,requiring the consumption of energy. Chloro-phyllous plants, called phototrophs, collect solarenergy. Some bacteria obtain energy from the oxi-dation of minerals: they are chemolithotrophs. Likemost animals and bacteria, fungi, including yeast,are chemoorganotrophs: they draw their necessaryenergy from the degradation of organic nutrients.
In a growing organism, energy produced bydegradation reactions (catabolism) is transferred tothe chain of synthesis reactions (anabolism). Con-forming to the laws of thermodynamics, energy
furnished by the degradation of a substrate is onlypartially converted into work; this is called freeenergy (the rest is dissipated in the form of heat).Part of this free energy can be used for transport,movement, or synthesis. In most cases, the freeenergy transporter particular to biological systemsis adenosine triphosphate (ATP). This molecule isrich in energy because its triphosphate unit con-tains two phosphoanhydride bonds (Figure 2.1).The hydrolysis of ATP into adenosine diphosphate(ADP) results in the liberation of a large quan-tity of free energy (7.3 kcal/mol). Biosynthesis andthe active transport of metabolites make use offree energy.
Handbook of Enology Volume 1 The Microbiology of Wine and Vinifications 2nd Edition P. Ribereau-Gayon, D. Dubourdieu, B. Doneche andA. Lonvaud 2006 John Wiley & Sons, Ltd ISBN: 0-470-01034-7

54 Handbook of Enology: The Microbiology of Wine and Vinifications
−O P O
−O
O
P
O
−O
O P
O
−O
OCH2O
HO OH
H HH
N
CHN
C
CC
N
CH
N
NH2
H
Fig. 2.1. Structure of adenosine triphosphate (ATP)
ATP + H2O −−−→←−−− ADP + Pi + H+�G◦′
= −7.3 kcal/mol (2.1)
In this reaction, �G◦′ is the change in free energy.ATP is considered to be ‘the universal money offree energy in biological systems’ (Stryer, 1992).In reality, microorganism growth or, in this case,yeast growth is directly related to the quantity ofATP furnished by metabolic pathways used fordegrading a substrate. It is indirectly related to thequantity of substrate degraded.
In the living cell, there are two processes whichproduce ATP: substrate-level phosphorylation andoxidative phosphorylation. Both of these pathwaysexist in wine yeasts.
Substrate-level phosphorylation can be eitheraerobic or anaerobic. During oxidation by electronloss, an ester–phosphoric bond is formed. It isan energy-rich bond between the oxidized carbonof the substrate and a molecule of inorganicphosphate. This bond is then transferred to theADP by transphosphorylation, thus forming ATP.This process takes place during glycolysis.
Oxidative phosphorylation is an aerobic pro-cess. The production of ATP is linked to thetransport of electrons to an oxygen molecule bythe cytochromic respiratory chain. This oxygenmolecule is the final acceptor of the electrons.These reactions occur in the mitochondria.
This chapter describes the principal biochemicalreactions occurring during grape must fermenta-tion by wine yeasts. It covers sugar metabolisms,i.e. the biochemistry of alcoholic fermentation, and
nitrogen metabolisms. Volatile sulfur-containingcompounds and volatile phenol formation mecha-nisms will be discussed in Volume 2, Chapter 8 inthe section concerning olfactory flaws. The influ-ence of yeasts on varietal wine aromas will becovered in Volume 2, Chapter 7.
2.2 SUGAR DEGRADATIONPATHWAYS
Depending on aerobic conditions, yeast can de-grade sugars using two metabolic pathways: alco-holic fermentation and respiration. These twoprocesses begin in the same way, sharing the com-mon trunk of glycolysis.
2.2.1 Glycolysis
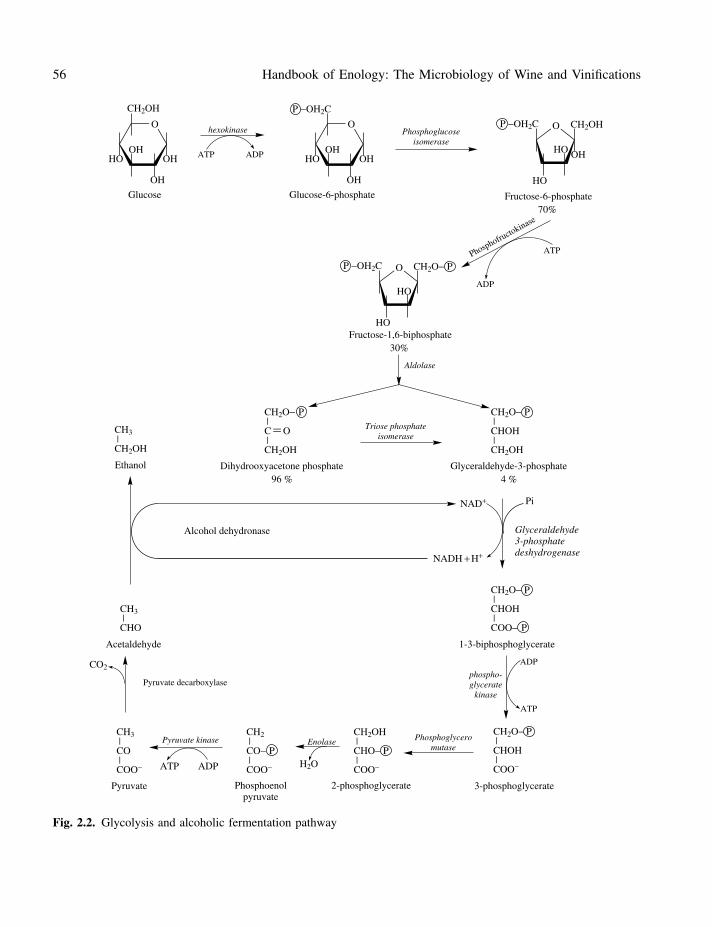
This series of reactions, transforming glucose intopyruvate with the formation of ATP, constitutesa quasi-universal pathway in biological systems.The elucidation of the different steps of glycolysisis intimately associated with the birth of modernbiochemistry. The starting point was the fortu-itous discovery by Hans and Eduard Buchner, in1897, of the fermentation of saccharose by an acel-lular yeast extract. Studying possible therapeuticapplications for their yeast extracts, the Buchnersdiscovered that the sugar used to preserve theiryeast extract was rapidly fermented into alcohol.Several years later, Harden and Young demon-strated that inorganic phosphate must be added toacellular yeast extract to assure a constant glu-cose fermentation speed. The depletion of inor-ganic phosphate during in vitro fermentation ledthem to believe that it was incorporated into asugar phosphate. They also observed that the yeastextract activity was due to a non-dialyzable com-ponent, denaturable by heat, and a thermostabledialyzable component. They named these two com-ponents ‘zymase’ and ‘cozymase’. Today, zymaseis known to be a series of enzymes and cozymaseis composed of their cofactors as well as metal ionsand ATP. The complete description of glycolysisdates back to the 1940s, due in particular to thecontributions of Embden, Meyerhoff and Neuberg.

Biochemistry of Alcoholic Fermentation and Metabolic Pathways of Wine Yeasts 55
For that reason, glycolysis is often called the Emb-den–Meyerhoff pathway.
The transport of must hexose (glucose and fruc-tose) across the plasmic membrane activates acomplex system of proteinic transporters not fullyexplained (Section 1.3.2). This mechanism facili-tates the diffusion of must hexoses in the cyto-plasm, where they are rapidly metabolized. Sincesolute moves in the direction of the concentrationgradient, from the concentrated outer medium tothe diluted inner medium, it is not an active trans-port system requiring energy.
Next, glycolysis (Figure 2.2) is carried outentirely in the cytosol of the cell. It includes afirst stage which converts glucose into fructose1,6-biphosphate, requiring two ATP molecules.This transformation itself comprises three steps:an initial phosphorylation of glucose into glu-cose 6-phosphate, the isomerization of the latterinto fructose 6-phosphate and a second phospho-rylation forming fructose 1,6-biphosphate. Thesethree reactions are catalyzed by hexokinase, phos-phoglucose isomerase and phosphoglucokinase,respectively.
In fact, Saccharomyces cerevisiae has two hex-okinases (PI and PII) capable of phosphorylatingglucose and fructose. Hexokinase PII is essen-tial and is active predominantly during the yeastlog phase in a medium with a high sugar con-centration. Hexokinase PI, partially repressed byglucose, is not active until the stationary phase(Bisson, 1991).
Mutant strains devoid of phosphoglucoiso-merase have been isolated. Their inability todevelop on glucose suggests that glycolysis isthe only catabolic pathway of glucose in Sac-charomyces cerevisiae (Caubet et al., 1988). Theoxidative pentose phosphate pathway, by whichsome organisms utilize sugars, serves only as ameans of synthesizing ribose 5-phosphate, incorpo-rated in nucleic acids and in reduced nicotinamide-adenine dinucleotide phosphate (NADPH) inSaccharomyces.
The second stage of glycolysis forms glycer-aldehyde 3-phosphate. Under the catalytic actionof aldolase, fructose 1,6-biphosphate is cleavedthus forming two triose phosphate isomers:
dihydroxyacetone phosphate and glyceraldehyde3-phosphate. The triose phosphate isomerase cat-alyzes the isomerization of these two compounds.Although at equilibrium the ketonic form is moreabundant, the transformation of dihydroxyace-tone phosphate into glyceraldehyde 3-phosphate israpid, since this compound is continually elimi-nated by the ensuing glycolysis reactions. In otherwords, a molecule of glucose leads to the formationof two molecules of glyceraldehyde 3-phosphate.
The third phase of glycolysis comprises twosteps which recover part of the energy from glyc-eraldehyde 3-phosphate (G3P). Initially, GA3Pis converted into 1,3-biphosphoglycerate (1,3-BPG). This reaction is catalyzed by glyceralde-hyde 3-phosphate dehydrogenase. It is an oxidationcoupled with a substrate-level phosphorylation.Nicotinamide–adenine dinucleotide (NAD+) is thecofactor of the dehydrogenation. At this stage, it isin its oxidized form; nicotinamide is the reactivepart of the molecule (Figure 2.3). Simultaneously,an energy-rich bond is established between the oxi-dized carbon of the substrate and the inorganicphosphate. The NAD+ accepts two electrons anda hydrogen atom lost by the oxidized substrate.Next, phosphoglyceratekinase catalyzes the trans-fer of the phosphoryl group of the acylphosphatefrom 1,3-BPG to ADP; and 3-phosphoglycerateand ATP are formed.
The last phase of glycolysis transforms 3-phosphoglycerate into pyruvate. Phosphoglycero-mutase catalyzes the conversion of 3-phospho-glycerate into 2-phosphoglycerate. Enolasecatalyzes the dehydration of the latter, formingphosphoenolpyruvate. This compound has a highphosphoryl group transfer potential. By phospho-rylation of ADP, pyruvic acid and ATP are formed;the pyruvate kinase catalyzes this reaction. In thismanner, glycolysis creates four ATP molecules.Two are immediately used to activate a new hexosemolecule, and the net gain of glycolysis is there-fore two ATP molecules per molecule of hexosemetabolized. This stage marks the end of the com-mon trunk of glycolysis; alcoholic fermentation,glyceropyruvic fermentation or respiration follow,depending on various conditions.
56 Handbook of Enology: The Microbiology of Wine and Vinifications
O
P −OH2C
ATP ADP
hexokinase Phosphoglucoseisomerase
OP −OH2C
CH2O− P
Glucose Glucose-6-phosphate Fructose-6-phosphate70%
ADP
ATP
Fructose-1,6-biphosphate30%
Triose phosphateisomerase
CH2O− P
C O
CH2OH
CH2O− P
CHOH
CH2OH
Dihydrooxyacetone phosphate96 %
Glyceraldehyde-3-phosphate4 %
CH2O− P
CHOH
COO− P
CH3
CHO
CH3
CO
COO−
CH2
CO∼ P
COO−
CH2OH
CHO− P
COO−
CH2O− P
CHOH
COO−
Acetaldehyde 1-3-biphosphoglycerate
ATP
ADP
phospho-glycerate
kinase
NAD+
NADH + H+
Pyruvate decarboxylase
Phosphoglyceromutase
EnolasePyruvate kinase
ATP ADP
CH3
CH2OH
Ethanol
Alcohol dehydronase
O
OP −OH2CPhosphofru
ctokinase
Glyceraldehyde3-phosphatedeshydrogenase
Pyruvate Phosphoenolpyruvate
2-phosphoglycerate 3-phosphoglycerate
Aldolase
Pi
CH2OH
HOOH
OH
OH OH
CH2OH
HO
HO
CO2
H2O
HOOH
OH
OH
HO
HO
Fig. 2.2. Glycolysis and alcoholic fermentation pathway
Biochemistry of Alcoholic Fermentation and Metabolic Pathways of Wine Yeasts 57
N
C NH2
O
N
N
HCN
C
CC
CH
N
NH2
N
H
R
CONH2
N
HH
R
CONH2+ 2H+ + 2e−
• •
+ H+
NADHNAD+
(a)
(b)
Reactive site
+
+
O
HH
HO OHH
H
O
P O−O
O
CH2
P O−O
O
CH2 O
HH
HO OH
H
H
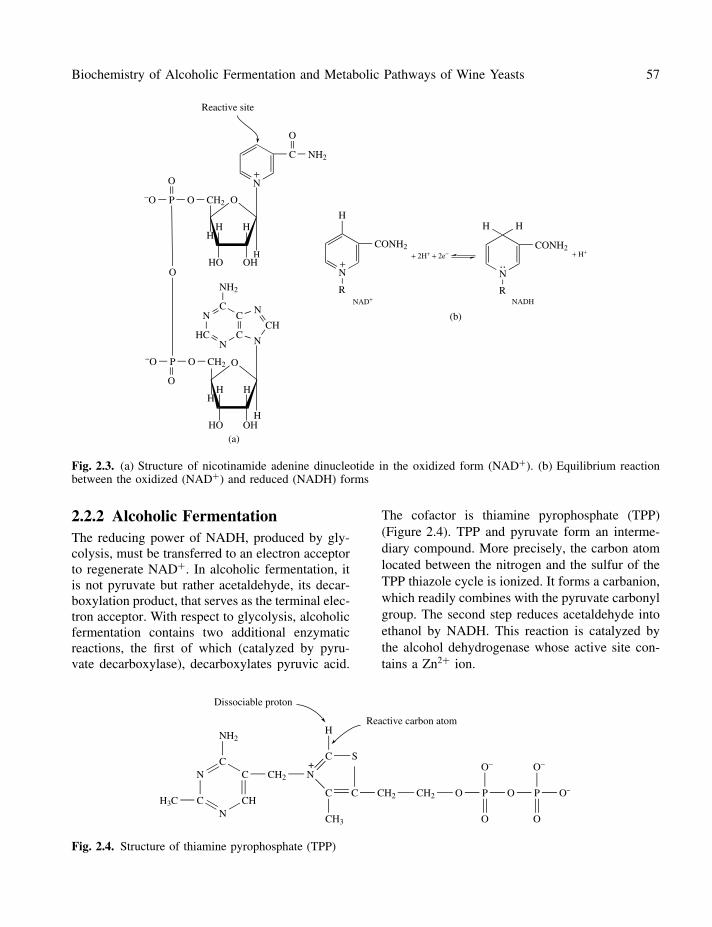
Fig. 2.3. (a) Structure of nicotinamide adenine dinucleotide in the oxidized form (NAD+). (b) Equilibrium reactionbetween the oxidized (NAD+) and reduced (NADH) forms
2.2.2 Alcoholic FermentationThe reducing power of NADH, produced by gly-colysis, must be transferred to an electron acceptorto regenerate NAD+. In alcoholic fermentation, itis not pyruvate but rather acetaldehyde, its decar-boxylation product, that serves as the terminal elec-tron acceptor. With respect to glycolysis, alcoholicfermentation contains two additional enzymaticreactions, the first of which (catalyzed by pyru-vate decarboxylase), decarboxylates pyruvic acid.
The cofactor is thiamine pyrophosphate (TPP)(Figure 2.4). TPP and pyruvate form an interme-diary compound. More precisely, the carbon atomlocated between the nitrogen and the sulfur of theTPP thiazole cycle is ionized. It forms a carbanion,which readily combines with the pyruvate carbonylgroup. The second step reduces acetaldehyde intoethanol by NADH. This reaction is catalyzed bythe alcohol dehydrogenase whose active site con-tains a Zn2+ ion.
N
CN
CH
CC
NH2
H3C
CH2 N
C
CH3
C
C S
H
CH2 CH2 O P
O
O−
O P
O
O−
O−
+
Dissociable proton
Reactive carbon atom
Fig. 2.4. Structure of thiamine pyrophosphate (TPP)

58 Handbook of Enology: The Microbiology of Wine and Vinifications
S. cerevisiae pyruvate decarboxylase (PDC)comprises two isoenzymes: a major form, PDC1,representing 80% of the decarboxylase activity,and a minor form, PDC5, whose function remainsuncertain.
From an energy viewpoint, glycolysis followedby alcoholic fermentation supplies the yeast withtwo molecules of ATP per molecule of glucosedegraded, or 14.6 biologically usable kcal/molof glucose fermented. From a thermodynamicviewpoint, the change in free energy during thedegradation of a mole of glucose into ethanol andCO2 is −40 kcal. The difference (25.4 kcal) isdissipated in the form of heat.
2.2.3 Glyceropyruvic Fermentation
In the presence of sulfite (Neuberg, 1946), thefermentation of glucose by yeasts produces equiv-alent quantities of glycerol, carbon dioxide, andacetaldehyde in its bisulfitic form. This glyceropy-ruvic fermentation takes place in the followingmanner. Since the acetaldehyde combined withsulfite cannot be reduced into ethanol, dihydroxy-acetone-1-phosphate becomes the terminal electronacceptor. It is derived from the oxidation of glyc-eraldehyde 3-phosphate and reduced to glycerol3-phosphate, which is itself dephosphorylated intoglycerol. This mechanism was used for the indus-trial production of glycerol. In this fermentation,only two molecules of ATP are produced for everymolecule of hexose degraded. ATP is required toactivate the glucose in the first step of glycolysis(Figure 2.5). Glyceropyruvic fermentation, whosenet gain in ATP is nil, does not furnish biologicallyassimilable energy for yeasts.
Glyceropyruvic fermentation does not occuruniquely in a highly sulfitic environment. In thebeginning of the alcoholic fermentation of grapemust, the inoculum consists of yeasts initiallygrown in the presence of oxygen. Their pyru-vate decarboxylase and alcohol dehydrogenaseare weakly expressed. As a result, ethanal accu-mulation is limited. The reoxidation of NADHdoes not involve ethanal, but rather dihydroxy-acetone. Glycerol, pyruvate and some secondaryfermentation products are formed. These secondary
Fructose 1,6-biphosphate
Dihydroxyacetone phosphate
Glycerol-3-phosphate
Glycerol Pyruvate
1,3-Biphosphoglycerate
2ADP
2ATP
NAD+
NADH, H+
H2O
H3PO4
Pl
Glyceraldehyde-3-phosphate
Secondary products(a-ketoglutaric acid, succinic acid,
butanediol, diacetyl, etc.)
Fig. 2.5. Glyceropyruvic fermentation pathway
products are pyruvate derivatives—including, butnot limited to, succinate and diacetyl.
2.2.4 RespirationWhen sugar is used by the respiratory pathway,pyruvic acid (originating in glycolysis) undergoesan oxidative decarboxylation in the presence ofcoenzyme A (CoA) (Figure 2.6) and NAD+. Thisprocess generates carbon dioxide, NADH andacetyl-CoA:
pyruvate + CoA + NAD+ −−−→acetyl CoA + CO2 + NADH + H+ (2.2)
The enzymatic system of the pyruvate dehydroge-nase catalyzes this reaction. It takes place in theinterior of the mitochondria. Thiamine pyrophos-phate (TPP), lipoamide and flavin–adenine din-ucleotide (FAD) participate in this reaction andserve as catalytic cofactors.
The acetyl unit issued from pyruvate is activatedin the form of acetyl CoA. The reactions of thecitric acid cycle, also called the tricarboxylic acidscycle and Krebs cycle (Figure 2.7), completelyoxidize the acetyl CoA into CO2. These reactionsalso occur in the mitochondria.
This cycle begins with the condensation ofa 2-carbon acetyl unit with a 4-carbon com-pound, oxaloacetate, to produce a tricarboxylic

Biochemistry of Alcoholic Fermentation and Metabolic Pathways of Wine Yeasts 59
HS CH2 CH2 N
H
C
O
CH2 CH2 N
H
C
O
C
H
OH
C
CH2
CH3
O P
O−
O
O P
O−
O
O
HH
O OH
H H
O
PO O−
O−
N
HCN
C
C
N
CH
N
NH2
CH2
b -mercapto-ethylamine Pantothenate
Fig. 2.6. Structure of coenzyme A. The reactive site is the terminal thiol group
acid with 6 carbon atoms: citric acid. Four oxi-dation–reduction reactions regenerate the oxaloac-etate. The oxidative pathway decarboxylatesisocitrate, an isomer of citrate, into α-ketoglutarate.The isocitrate dehydrogenase catalyzes this reac-tion. The α-ketoglutarate, a 5-carbon atom com-pound, undergoes an oxidative decarboxylation tobecome succinate by the α-ketoglutarate dehy-drogenase. In these two reactions, NAD+ isthe hydrogen acceptor. The fumarate dehydroge-nase is responsible for the reduction of succinateinto fumarate; FAD is the hydrogen acceptor(Figure 2.8). Finally, fumarate is hydrated intoL-malate. The latter is reduced into oxaloacetate bythe malate dehydrogenase. In this case, the NAD+is an electron acceptor once again.
From acetate, each complete cycle produces twoCO2 molecules, three hydrogen ions transferred tothree NAD+ molecules (six electrons) and a pair of
hydrogen atoms (two electrons) transferred to anFAD molecule. The cytochrome chain transportsthese electrons towards the oxygen. ATP is formedduring this process. This oxidative phosphoryla-tion (Figure 2.9) takes place in the mitochondria.This process makes use of three enzymatic systems(the NADH-Q reductase, the cytochrome reduc-tase and the cytochrome oxidase). Two electrontransport systems (ubiquinone, or coenzyme Q, andcytochrome c) link these enzymatic systems.
Oxidative phosphorylation yields three ATPmolecules per pair of electrons transported betweenthe NADH and the oxygen—two ATP moleculeswith FADH2. In the Krebs cycle, substrate-levelphosphorylation forms an ATP molecule during thetransformation of succinyl CoA into succinate.
The respiration of a glucose molecule (Table2.1) produces 36 to 38 ATP molecules. Twooriginate from glycolysis, 28 from the oxidative
Table 2.1. Energy balance of oxidation of glucose in respiration
Stage Reduction coenzyme Number of moleculesof ATP formed
Glycolysis 2NADH 4 or 6Net gain in ATP from glycolysis 2Pyruvate −−−→ acetyl CoA NADH 6Isocitrate −−−→ α-Ketoglutarate NADH 6α-Ketoglutarate −−−→ succinyl CoA NADH 6Succinyl CoA −−−→ succinate 2Succinate −−−→ fumarate FADH2 4Malate −−−→ oxaloacetate NADH 6Net yield from glucose 36–38

60 Handbook of Enology: The Microbiology of Wine and Vinifications
COO−
Pyruvate
CH3
C
S
O
O
CoAAcetyl CoA
CO2
HSCoA
NADH, H+NAD+
COO−
COO−
Citrate
CH2
CHO COO−
CH2
COO−
COO−
Cis-aconitate
CH2
C COO−
CH
COO−
COO−
Isocitrate
CH2
CH
H OH
COO−
C
COO−
COO−
a-Ketogluturate
CH2
CH2
C O
COO−
OSuccinyl-CoA
CH2
CH2
C SCoA
COO−
COO−
Succinate
CH2
CH2
COO−
COO−
Fumarate
CH
HC
COO−
COO−
Malate
C H
CH2
HO
COO−
COO−
Oxaloacetate
C
CH2
O
H2O
H2O
1 2
3
4
5
6
7
8
9
NAD+
CO2
NADH, H+
HSCoANAD+
NADH, H+CO2
GDP
ADP
HSCoA
GTP
ATP
FAD
FADH2
H2O
NAD+
NADH, H+
HSCoA
C
CH3
Fig. 2.7. Tricarboxylic acid or Krebs cycle. 1 = citrate synthase; 2–3 = aconitase; 4 = isocitrate dehydroge-nase; 5 = complex α-ketoglutarate dehydrogenase; 6 = succinyl-CoA synthetase, 7 = succinate dehydrogenase;8 = fumarase; 9 = malate dehydrogenase; GTP = guanosine triphosphate; GDP = guanosine diphosphate

Biochemistry of Alcoholic Fermentation and Metabolic Pathways of Wine Yeasts 61
C
CC
C
CC
NC
CN
NC
NC
H3C
H3C
H
H O
CH2
O
H
CHOH
CHOH
CHOH
CH2
O
P
O−
O O P
O−
O
O CH2O
H
H
HO
H
OH
H
C
CC
C
CC
NC
CN
NC
NC
H3C
H3C
H O
O
H
H
H
R H(b)
(a)
Adenine
Fig. 2.8. Structure of flavin adenine dinucleotide (FAD): (a) oxidized form (FAD); (b) reduced form (FADH2)
ADP + Pi
ATP
ADP + Pi
ATP
NADH, H+
ADP + Pi
ATP
NADH - Qreductase
FADH2Coenzyme Q
Cytochromereductase
Cytochrome c
Cytochromeoxidase
½2 H2O
Fig. 2.9. Oxidative phosphorylation during electrontransport in the respiratory chain
phosphorylation of NADH and FADH2 generatedby the Krebs cycle, and two from substrate-level phosphorylation during the formation ofsuccinate. Four to six ATP molecules result from
the oxidative phosphorylation of two NADHmolecules produced in glycolysis. The precisenumber depends on the transport system used tomove the electrons of the cytosolic NADH tothe respiratory chain in the mitochondria. Therespiration of the same amount of sugar produces18 to 19 times more biologically usable energyavailable to yeasts than fermentation. Respirationis used for industrial yeast production.
2.3 REGULATION OFSUGAR-UTILIZINGMETABOLIC PATHWAYS
2.3.1 Regulation BetweenFermentation and Respiration:Pasteur Effect and CrabtreeEffect
Pasteur was the first to compare yeast growth inaerobiosis and anaerobiosis. For low concentrationsof glucose on culture media, yeasts utilize sugarsthrough either respiration or fermentation. Aera-tion induces an increase in biomass formed (totaland per unit of sugar degraded) and a decrease in

62 Handbook of Enology: The Microbiology of Wine and Vinifications
alcohol production and sugar consumption. Pas-teur therefore deduced that respiration inhibitsfermentation.
The ‘Pasteur effect’ has been interpreted inseveral ways. Two enzymes compete to catalyzeeither the respiration or fermentation of pyruvate.This competition explains the respiratory inhibitionof fermentation. The pyruvate decarboxylase isinvolved in the fermentative pathway. It has alower affinity towards pyruvate than pyruvatedehydrogenase. Furthermore, oxidative phosphory-lation consumes a lot of ADP and inorganic phos-phate, which migrate to the mitochondria. A lackof ADP and inorganic phosphate in the cytoplasmensues. This deficit can limit the phosphorylationand thus slow the glycolitic flux. The inhibitionof glycolysis enzymes by ATP explains the Pas-teur effect for the most part. The ATP issuedfrom oxidative phosphorylation inhibits phospho-fructokinase in particular. Phosphorylated hexosesaccumulate as a result. The transmembrane trans-port of sugars and thus glycolysis is slowed.
For high glucose concentrations—for example,in grape must—S. cerevisiae only metabolizessugars by the fermentative pathway. Even in thepresence of oxygen, respiration is impossible. Dis-covered by Crabtree (1929) on tumoral cells, thisphenomenon is known by several names: catabolicrepression by glucose, the Pasteur contrary effectand the Crabtree effect. Yeasts manifest the fol-lowing signs during this effect: a degeneration ofthe mitochondria, a decrease in the proportion ofcellular sterols and fatty acids, and a repressionof both the synthesis of Krebs cycle mitochondrialenzymes and constituents of the respiratory chain.With S. cerevisiae, there must be at least 9 g ofsugar per liter for the Crabtree effect to occur. Thecatabolic repression exerted by glucose on wineyeasts is very strong. In grape must, at any levelof aeration, yeasts are only capable of fermentingbecause of the high glucose and fructose concen-trations. From a technological viewpoint, yeastsconsume sugars by the respiratory pathway for theindustrial production of dry yeast, but not in wine-making. If must aeration helps the alcoholic fer-mentation process (Section 3.7.2), the fatty acidsand sterols synthesized by yeasts, proliferating in
the presence of oxygen, are responsible but notrespiration.
S. cerevisiae can metabolize ethanol by therespiratory pathway in the presence of smallquantities of glucose. After alcoholic fermentation,oxidative yeasts develop in a similar manner onthe surface of wine as part of the process ofmaking certain specialty wines (Sherry, YellowWine of Jura).
2.3.2 Regulation Between AlcoholicFermentation andGlyceropyruvic Fermentation;Glycerol Accumulation
Wines contain about 8 g of glycerol per 100 gof ethanol. During grape must fermentation, about8% of the sugar molecules undergo glyceropyru-vic fermentation and 92% undergo alcoholic fer-mentation. The fermentation of the first 100 g ofsugar forms the majority of glycerol, after whichglycerol production slows but is never nil. Glyc-eropyruvic fermentation is therefore more than aninductive fermentation which regenerates NAD+when acetaldehyde, normally reduced into ethanol,is not yet present. Alcoholic fermentation and glyc-eropyruvic fermentation overlap slightly through-out fermentation.
Pyruvic acid is derived from glycolysis. Whenthis molecule is not used by alcoholic fermentation,it participates in the formation of secondaryproducts. In this case, a molecule of glycerol isformed by the reduction of dihydroxyacetone.
Glycerol production therefore equilibrates theyeast endocellular oxidation–reduction potential,or NAD+/NADH balance. This ‘relief valve’eliminates surplus NADH which appears at the endof amino acid and protein synthesis.
Some winemakers place too much importanceon the organoleptical role of glycerol. This com-pound has a sugary flavor similar to glucose. In thepresence of other constituents of wine, however,the sweetness of glycerol is practically impercepti-ble. For the majority of tasters, even well trained,the addition of 3–6 g of glycerol per liter to ared wine is not discernible and so the pursuit ofwinemaking conditions that are more conducive

Biochemistry of Alcoholic Fermentation and Metabolic Pathways of Wine Yeasts 63
to glyceropyruvic fermentation has no enologicalinterest. On the contrary, the winemaker shouldfavor a pure alcoholic fermentation and shouldlimit glyceropyruvic fermentation. The produc-tion of glycerol is accompanied by the formationof other secondary products, derived from pyru-vic acid, the increased presence of which (suchas carbonyl function compounds and acetic acid)decreases wine quality.
2.3.3 Secondary Products Formedfrom Pyruvate by GlyceropyruvicFermentation
When a molecule of glycerol is formed, a moleculeof pyruvate cannot be transformed into ethanol fol-lowing its decarboxylation into ethanal. In anaero-bic conditions, oxaloacetate is the means of entryof pyruvate into the cytosolic citric acid cycle.Although the mitochondria are no longer func-tional, the enzymes of the tricarboxylic acids cycleare present in the cytoplasm. Pyruvate carboxylase(PC) catalyzes the carboxylation of pyruvate intooxaloacetate. The prosthetic group of this enzymeis biotin; it serves as a CO2 transporter. The reactionmakes use of an ATP molecule:
biotin–PC + ATP + CO2 −−−→CO2 –biotin–PC + ADP + [iP] (2.3)
CO2 –biotin–PC + pyruvate −−−→biotin–PC + oxaloacetate (2.4)
In these anaerobic conditions, the citric acid cyclecannot be completed since the succinodehydro-genase activity requires the presence of FAD, astrictly respiratory coenzyme. The chain of reac-tions is therefore interrupted at succinate, whichaccumulates (0.5–1.5 g/l). The NADH generatedby this portion of the Krebs cycle (from oxaloac-etate to succinate) is reoxidized by the formationof glycerol from dihydroxyacetone.
The α-ketoglutarate dehydrogenase has a verylow activity in anaerobiosis; some authors there-fore believe that the oxidative reactions of theKrebs cycle are interrupted at α-ketoglutarate.In their opinion, a reductive pathway of the
citric acid cycle forms succinic acid in anaer-obiosis: oxaloacetate → malate → fumarate →succinate. Bacteria have a similar mechanism. Inyeast, this is probably a minor pathway since onlythe oxidative pathway of the Krebs cycle can main-tain the NAD+/NADH redox balance during fer-mentation (Oura, 1977). Furthermore, additionalsuccinate is formed during alcoholic fermentationon a glutamate-enriched medium. The glutamateis deaminated to form α-ketoglutarate, which isoxidized into succinate.
Among secondary products, ketonic functioncompounds (pyruvic acid, α-ketoglutaric acid) andacetaldehyde predominantly combine with sulfurdioxide in wines made from healthy grapes. Theirexcretion is significant during the yeast prolifera-tion phase and decreases towards the end of fer-mentation. Additional acetaldehyde is liberated inthe presence of excessive quantities of sulfur diox-ide in must. An elevated pH and fermentation tem-perature, anaerobic conditions, and a deficiency inthiamine and pantothenic acid increase productionof ketonic acids. Thiamine supplementing of mustlimits the accumulation of ketonic compounds inwine (Figure 2.10).
0
50
100
150
200
250
0 50 100 150 200
I
II
Fermented sugar (g/l)
Pyru
vic
acid
(m
g/l)
Fig. 2.10. Effect of a thiamine addition on pyru-vic acid production during alcoholic fermentation(Lafon-Lafourcade, 1983). I = control must; II = thia-mine supplemented must

64 Handbook of Enology: The Microbiology of Wine and Vinifications
Other secondary products of fermentation arealso derived from pyruvic acid: acetic acid,lactic acid, butanediol, diacetyl and acetoin. Theirformation processes are described in the followingparagraphs.
2.3.4 Formation and Accumulationof Acetic Acid by Yeasts
Acetic acid is the principal volatile acid of wine. Itis produced in particular during bacterial spoilage(acetic spoilage and lactic disease) but is alwaysformed by yeasts during fermentation. Beyonda certain limit, which varies depending on thewine, acetic acid has a detrimental organolepticaleffect on wine quality. In healthy grape mustwith a moderate sugar concentration (less than220 g/l), S. cerevisiae produces relatively smallquantities (100–300 mg/l), varying according tothe strain. In certain winemaking conditions, evenwithout bacterial contamination, yeast acetic acidproduction can be abnormally high and become aproblem for the winemaker.
The biochemical pathway for the formationof acetic acid in wine yeasts has not yet beenclearly identified. The hydrolysis of acetyl CoA canproduce acetic acid. The pyruvate dehydrogenaseproduces acetyl CoA beforehand by the oxidativedecarboxylation of pyruvic acid. This reaction takesplace in the matrices of the mitochondria but islimited in anaerobiosis. Aldehyde dehydrogenasecan also form acetic acid by the oxidation ofacetaldehyde (Figure 2.11). This enzyme, whosecofactor is NADP+, is active during alcoholicfermentation. The NADPH thus formed can be usedto synthesize lipids. When pyruvate dehydrogenaseis repressed, this pathway forms acetyl CoA throughthe use of acetyl CoA synthetase. In anaerobiosis ona model medium, yeast strains producing the leastamount of acetic acid have the highest acetyl CoAsynthetase activity (Verdhuyn et al., 1990).
The acetaldehyde dehydrogenase in S. cerevisiaehas five isoforms, three located in the cytosol(Section 1.4.1) (Ald6p, Ald2p, and Ald3p) and theremaining two (Ald4p and Ald5p) in the mito-chondria (Section 1.4.3). These enzymes differ bytheir specific use of the NAD+ or NADP+ cofactor(Table 2.2).
EthanolNAD+
NAD+
NADP+ NADPH, H+NADH, H+
NADH, H+
2
1
3
4Acetaldehyde Acetate
Pyruvate
CO2
5
HS-CoA
HS-CoAHS-CoA
6
H2O
Acetyl-CoA
Lipid synthesis
CO2
Fig. 2.11. Acetic acid formation pathways in yeasts.1 = pyruvate decarboxylase; 2 = alcohol dehydroge-nase; 3 = pyruvate dehydrogenase; 4 = aldehyde dehy-drogenase; 5 = acetyl-CoA hydrolase; 6 = acetyl-CoAsynthetase
Table 2.2. Isoforms of acetaldehyde dehydrogenase inS. cerevisiae (Navarro-Avino et al., 1999)
Chromosome Gene Location Cofactor
XIII ALD2 Cytosol NAD+XIII ALD3 Cytosol NAD+XV ALD4 Mitochondria NAD+ and
NADP+V ALD5 Mitochondria NADP+XVI ALD6 Cytosol NADP+
Remize et al. (2000) and Blondin et al. (2002)studied the impact of the deletion of each geneand demonstrated that the NADP-dependent cyto-plasmic isoform ALD6 played a major role in theformation of acetic acid during the fermentationof dry wines, while the ALD5 isoform was alsoinvolved, but to a lesser extent (Figure 2.12).
Practical winemaking conditions likely to leadto abnormally high acetic acid production byS. cerevisiae are well known. As is the casewith glycerol formation, acetic acid productionis closely dependent on the initial sugar levelof the must, independent of the quantity of sug-ars fermented (Table 2.3). The higher the sugarcontent of the must, the more acetic acid (andglycerol) the yeast produces during fermentation.This is due to the yeast’s mechanism for adapt-ing to a medium with a high sugar concentration:

Biochemistry of Alcoholic Fermentation and Metabolic Pathways of Wine Yeasts 65
0.8
0.7
0.6
0.5
0.4
0.3
0.2
0.1
0.0
Témoin ALD6 ALD3 ALD2 ALD4 ALD5
Ace
tate
(g/
l)
Fig. 2.12. Acetate production by strains of S. cerevisiae(V5) following deletion of different genes coding forisoforms of acetaldehyde dehydrogenase (Blondin et al.,2002)
Table 2.3. Effect of initial sugar concentration of themust on the formation of secondary products of thefermentation (Lafon-Lafourcade, 1983)
Initial Fermen- Secondary productssugar ted sugar
Acetic acid Glycerol Succinic acid(g/l) (g/l)(g/l) (g/l) (g/l)
224 211 0.26 4.77 0.26268 226 0.45 5.33 0.25318 211 0.62 5.70 0.26324 179 0.84 5.95 0.26348 152 1.12 7.09 0.28
S. cerevisiae increases its intracellular accumula-tion of glycerol to counterbalance the osmotic pres-sure of the medium (Blomberg and Alder, 1992).This regulation mechanism is controlled by a cas-cade of signal transmissions leading to an increasein the transcription level of genes involved in theproduction of glycerol (GPD1 ), but also of acetate(ALD2 and ALD3 ) (Attfield et al., 2000). Acetateformation plays an important physiological rolein the intracellular redox balance by regeneratingreduced equivalents of NADH. Thus, it is clearthat an increase in acetate production is inherentto the fermentation of high-sugar musts. However,Bely et al. (2003) demonstrated that it was possi-ble to reduce acetate production by supplying moreNADH to the redox balance process. This may be
Max. population (A610 max.)
1.4
1.2
1.0
Vol
atile
aci
dity
(g
/l ac
etic
aci
d)
0.8
0.6
0.4
0.2
5 10 15 20
Fig. 2.13. Correlation between volatile acidity produc-tion and the maximum cell population in high-sugar,botrytized musts
1.50
1.25
1.00
0.75
0.50
0.250 100 200
Available nitrogen (mg/l)
300 400
Vol
atile
aci
dity
(g
/l ac
etic
aci
d)
Fig. 2.14. Effect of the available nitrogen content inmust (with or without ammonium supplements) on theproduction of volatile acidity (initial sugar content:350 g/l)
done indirectly by stimulating biomass formation,which generates an excess of NADH during aminoacid synthesis. Available nitrogen in the must playsa key role in this process. Thus, in high-sugarmusts, acetate production is inversely correlatedwith the maximum cell population (Figure 2.13),which is, in turn, related to the available nitrogencontent of the must. It is strongly recommendedto monitor the available nitrogen content of botry-tized musts and supplement them with ammoniumsulfate, if necessary. The optimum available nitro-gen concentration in this type of must to minimizeacetic acid production is approximately 190 mg/l(Figure 2.14). The best time for adding nitrogensupplements is at the very beginning of fermenta-tion, as later additions are less effective and may

66 Handbook of Enology: The Microbiology of Wine and Vinifications
even increase acetate production. Indeed, in viewof the unpredictable increase in acetic acid produc-tion that sometimes occurred in botrytized mustssupplemented with ammonium sulfate, many enol-ogists had given up the practice entirely. It is nowknown that, provided the supplement is added atthe very beginning of fermentation, adjusting theavailable nitrogen content to the optimum level(190 mg/l) always minimizes acetic acid produc-tion in botrytized wines.
In wines made from noble rotted grapes, certainsubstances in the must inhibit yeast growth andincrease the production of acetic acid and glyc-erol during fermentation. Botrytis cinerea secretesthese ‘botryticine’ substances (Ribereau-Gayonet al., 1952, 1979). Fractional precipitation withethanol partially purifies these compounds frommust and culture media of Botrytis cinerea. Theseheat stable glycoproteins have molecular weightsbetween 10 and 50 000. They comprise a peptidic(10%) and glucidic part containing mostly man-nose and galactose and some rhamnose and glu-cose (Dubourdieu, 1982). When added to healthygrape must, these compounds provoke an increasein glyceropyruvic fermentation and a significantexcretion of acetic acid at the end of fermentation(Figure 2.15). The mode of action of these glyco-proteins on yeasts has not yet been identified. Thephysiological state of yeast populations at the timeof inoculation seems to play an important role inthe fermentative development of botrytized grapemust. Industrial dry yeast preparations are muchmore sensitive to alcoholic fermentation inhibitorsthan yeast starters obtained by preculture in healthygrape musts.
Other winemaking factors favor the productionof acetic acid by S. cerevisiae: anaerobiosis, verylow pH (<3.1) or very high pH (>4), certainamino acid or vitamin deficiencies in the must,and too high of a temperature (25–30◦C) dur-ing the yeast multiplication phase. In red wine-making, temperature is the most important factor,especially when the must has a high sugar concen-tration. In hot climates, the grapes should be cooledwhen filling the vats. The temperature should notexceed 20◦C at the beginning of fermentation. The
Ace
tic a
cid
(g/l)
Gly
cero
l (g/
l)
1
1
0 5 10 20
1
2′
2
1′
Days
Fig. 2.15. Effect of an alcohol-induced precipitate ofa botrytized grape must on glycerol and acetic acidformation during the alcoholic fermentation of healthygrape must (Dubourdieu, 1982). (1) Evolution of acidacetic concentration in the control must; (2) evolution ofacid acetic concentration in the must supplemented withthe freeze-dried precipitate; (1′) evolution of glycerolconcentration in the control must; (2′) evolution ofglycerol concentration in the must supplemented withthe freeze-dried precipitate
same procedure should be followed in thermovini-fication immediately following the heating of thegrapes.
In dry white and rose winemaking, excessivemust clarification can also lead to the exces-sive production of volatile acidity by yeast. Thisphenomenon can be particularly pronounced withcertain yeast strains. Therefore, must turbidityshould be adjusted to the lowest possible levelwhich permits a complete and rapid fermen-tation (Chapter 13). Solids sedimentation (mustlees) furnishes long-chain unsaturated fatty acids(C18:1, C18:2). Yeast lipidic alimentation greatly
Biochemistry of Alcoholic Fermentation and Metabolic Pathways of Wine Yeasts 67
0,4
0,3
0,2
0,1
Vol
atile
aci
dity
(g/
l H2S
O4)
Must andfresh lees
Must and lipidicextract from lees
Must and micro-granular cellulose
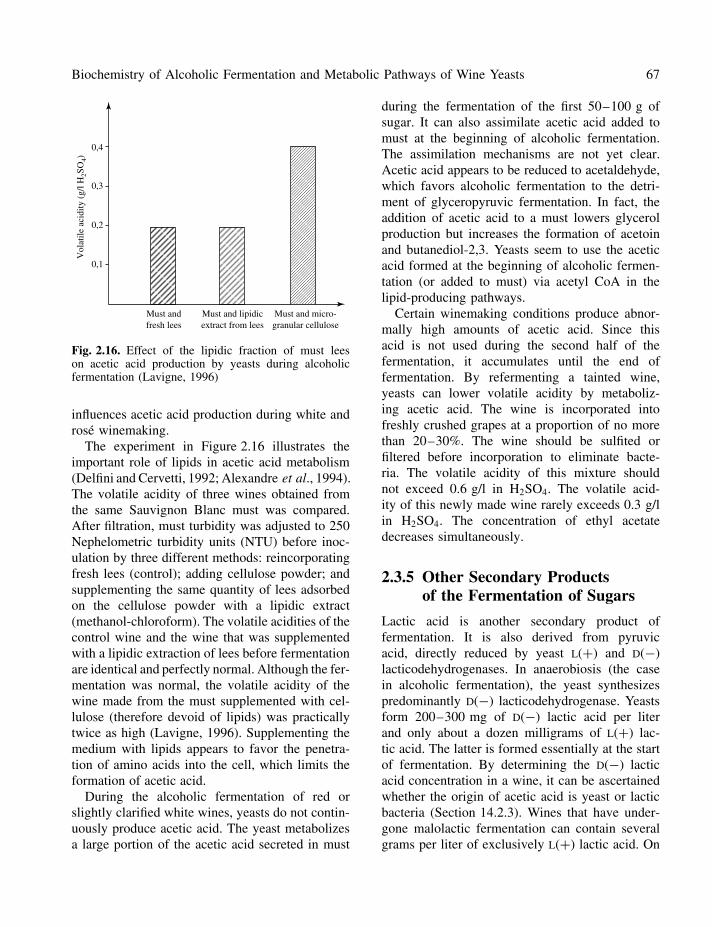
Fig. 2.16. Effect of the lipidic fraction of must leeson acetic acid production by yeasts during alcoholicfermentation (Lavigne, 1996)
influences acetic acid production during white androse winemaking.
The experiment in Figure 2.16 illustrates theimportant role of lipids in acetic acid metabolism(Delfini and Cervetti, 1992; Alexandre et al., 1994).The volatile acidity of three wines obtained fromthe same Sauvignon Blanc must was compared.After filtration, must turbidity was adjusted to 250Nephelometric turbidity units (NTU) before inoc-ulation by three different methods: reincorporatingfresh lees (control); adding cellulose powder; andsupplementing the same quantity of lees adsorbedon the cellulose powder with a lipidic extract(methanol-chloroform). The volatile acidities of thecontrol wine and the wine that was supplementedwith a lipidic extraction of lees before fermentationare identical and perfectly normal. Although the fer-mentation was normal, the volatile acidity of thewine made from the must supplemented with cel-lulose (therefore devoid of lipids) was practicallytwice as high (Lavigne, 1996). Supplementing themedium with lipids appears to favor the penetra-tion of amino acids into the cell, which limits theformation of acetic acid.
During the alcoholic fermentation of red orslightly clarified white wines, yeasts do not contin-uously produce acetic acid. The yeast metabolizesa large portion of the acetic acid secreted in must
during the fermentation of the first 50–100 g ofsugar. It can also assimilate acetic acid added tomust at the beginning of alcoholic fermentation.The assimilation mechanisms are not yet clear.Acetic acid appears to be reduced to acetaldehyde,which favors alcoholic fermentation to the detri-ment of glyceropyruvic fermentation. In fact, theaddition of acetic acid to a must lowers glycerolproduction but increases the formation of acetoinand butanediol-2,3. Yeasts seem to use the aceticacid formed at the beginning of alcoholic fermen-tation (or added to must) via acetyl CoA in thelipid-producing pathways.
Certain winemaking conditions produce abnor-mally high amounts of acetic acid. Since thisacid is not used during the second half of thefermentation, it accumulates until the end offermentation. By refermenting a tainted wine,yeasts can lower volatile acidity by metaboliz-ing acetic acid. The wine is incorporated intofreshly crushed grapes at a proportion of no morethan 20–30%. The wine should be sulfited orfiltered before incorporation to eliminate bacte-ria. The volatile acidity of this mixture shouldnot exceed 0.6 g/l in H2SO4. The volatile acid-ity of this newly made wine rarely exceeds 0.3 g/lin H2SO4. The concentration of ethyl acetatedecreases simultaneously.
2.3.5 Other Secondary Productsof the Fermentation of Sugars
Lactic acid is another secondary product offermentation. It is also derived from pyruvicacid, directly reduced by yeast L(+) and D(−)lacticodehydrogenases. In anaerobiosis (the casein alcoholic fermentation), the yeast synthesizespredominantly D(−) lacticodehydrogenase. Yeastsform 200–300 mg of D(−) lactic acid per literand only about a dozen milligrams of L(+) lac-tic acid. The latter is formed essentially at the startof fermentation. By determining the D(−) lacticacid concentration in a wine, it can be ascertainedwhether the origin of acetic acid is yeast or lacticbacteria (Section 14.2.3). Wines that have under-gone malolactic fermentation can contain severalgrams per liter of exclusively L(+) lactic acid. On

68 Handbook of Enology: The Microbiology of Wine and Vinifications
TPP
Pyruvate
Pyruvate
TPP-C2α-Acetolactate
CO2
CO2
CH3
CH3
CO
CO
CH3
CH3
CHOH
CO
CH3
CH3
CHOH
CHOH
NADH, H+
NADH, H+
NADH, H+
NAD+
NAD+
NAD+
Diacetyl Acetoin 2,3-Butanediol
CO2
Fig. 2.17. Acetoin, diacetyl and 2,3-butanediol formation by yeasts in anaerobiosis. TPP = thiamine pyrophosphate;TPP-C2 = active acetaldehyde
the other hand, the lactic fermentation of sugars(lactic disease) forms D(−) lactic acid. Lactic bac-teria have transformed substrates other than malicacid, when D(−) lactic acid concentrations exceed200 to 300 mg/l.
Yeasts also make use of pyruvic acid to formacetoin, diacetyl and 2,3-butanediol (Figure 2.17).This process begins with the condensation of apyruvate molecule and active acetaldehyde boundto thiamine pyrophosphate, leading to the forma-tion of α-acetolactic acid. The oxidative decar-boxylation of α-acetolactic acid produces diacetyl.Acetoin is produced by either the non-oxidativedecarboxylation of α-acetolactic acid or the reduc-tion of diacetyl. The reduction of acetoin leads tothe formation of 2,3-butanediol; this last reactionis reversible.
From the start of alcoholic fermentation, yeastsproduce diacetyl, which is rapidly reduced toacetoin and 2,3-butanediol. This reduction takesplace in the days that follow the end of alcoholicfermentation, when wines are conserved on theyeast biomass (de Revel et al., 1996). Acetoin andespecially diacetyl are strong-smelling compoundswhich evoke a buttery aroma. Above a certain
concentration, they have a negative effect on winearoma. The concentration of wines that have notundergone malolactic fermentation is too low (afew milligrams per liter for diacetyl) to havean olfactive influence. On the other hand, lacticbacteria can degrade citric acid to produce muchhigher quantities of these carbonyl compoundsthan yeasts (Section 5.3.2).
Finally, yeasts condense acetic acid (in theform of acetyl CoA) and pyruvic acid to pro-duce citramalic acid (0–300 mg/l) and dimethyl-glyceric acid (0–600 mg/l) (Figure 2.18). Thesecompounds have little organoleptic incidence.
H3C C
COOH
CH2
OH
COOH
HO CH
C
CH3
COOHHO
CH3
(a)
(b)
Fig. 2.18. (a) Citramalic acid and (b) dimethylglycericacid

Biochemistry of Alcoholic Fermentation and Metabolic Pathways of Wine Yeasts 69
COOH
CH2
CHOH
COOH
CH3
C
COOH
O
CH3
CHO
CH3
CH2OH
Malate Pyruvate Acetaldehyde Ethanol
Malicenzyme
Pyruvatedecarboxylase
Alcoholdehydrogenase
NADH, H+
CO2 CO2NAD+
Fig. 2.19. Decomposition of malic acid by yeasts during alcoholic fermentation
2.3.6 Degradation of Malic Acidby Yeast
Saccharomyces cerevisiae partially degrades mustmalic acid (10–25%) during alcoholic fermenta-tion. Different strains degrade varying amountsof this acid, and degradation is more signifi-cant when the pH is low. Alcoholic fermentationis the principal pathway degrading malic acid.The pyruvic acid resulting from this transforma-tion is decarboxylated into ethanal, which is thenreduced to ethanol. The malic enzyme is respon-sible for the transformation of malic acid intopyruvic acid (Figure 2.19). This oxidative decar-boxylation requires NAD+ (Fuck and Radler,1972). This maloalcoholic fermentation lowerswine acidity significantly more than malolacticfermentation.
Schizosaccharomyces differs from wine yeasts.The alcoholic fermentation of malic acid is com-plete in yeasts of this genus, which possess anactive malate transport system. (In S. cerevisiae,malic acid penetrates the cell by simple diffu-sion.) Yet at present no attempts to use Schizosac-charomyces in winemaking have been successful(Peynaud et al., 1964; Carre et al., 1983). First ofall, the implantation of these yeasts in the pres-ence of S. cerevisiae is difficult in a non-sterilizedmust. Secondly, their optimum growth tempera-ture (30◦C) higher than for S. cerevisiae, imposeswarmer fermentation conditions. Sometimes, thehigher temperature adversely affects the organolep-tical quality of wine. Finally, some grape varietiesfermented by Schizosaccharomyces do not expresstheir varietal aromas. The acidic Gros Mansengvariety produces a very fruity wine when correctly
vinified with S. cerevisiae, but has no varietalaroma when fermented by Schizosaccharomyces.To resolve these problems, some researchers haveused non-proliferating populations of Schizosac-charomyces enclosed in alginate balls. These pop-ulations degrade the malic acid in wines hav-ing already completed their alcoholic fermentation(Magyar and Panyid, 1989; Taillandier and Streha-iano, 1990). Although no organoleptical defect isfound in these wines, the techniques have not yetbeen developed for practical use.
Today, molecular biology permits another strat-egy for making use of the ability of Schizosac-charomyces to ferment malic acid. It consists ofintegrating Schizosaccharomyces malate permeasegenes and the malic enzyme (mae 1 and mae 2)in the S. cerevisiae genome (Van Vuuren et al.,1996). The technological interest of a wine yeastgenetically modified in this manner is not yet clear,nor are the risks of its proliferation in wineries andnature.
2.4 METABOLISM OF NITROGENCOMPOUNDS
The nitrogen requirements of wine yeasts andthe nitrogen supply in grape musts are dis-cussed in Chapter 3 (Section 3.4.2). The fol-lowing section covers the general mechanismsof assimilation, biosynthesis, and degradation ofamino acids in yeasts. The consequences of thesemetabolisms, which occur during alcoholic fer-mentation and affect the production of higher alco-hols and their associated esters in wine, are alsodiscussed.

70 Handbook of Enology: The Microbiology of Wine and Vinifications
2.4.1 Amino Acid Synthesis Pathways
The ammonium ion and amino acids found ingrape must supply the yeast with nitrogen. Theyeast can also synthesize most of the aminoacids necessary for constructing its proteins. Itfixes an ammonium ion on a carbon skeletonderived from the metabolism of sugars. The yeastuses the same reactional pathways as all organ-isms. Glutamate and glutamine play an importantrole in this process (Cooper, 1982; Magasanik,1992).
The NADP+ glutamate dehydrogenase (NADP+-GDH), product of the GDH1 gene, produces gluta-mate (Figure 2.20) from an ammonium ion and anα-ketoglutarate molecule. The latter is an interme-diary product of the citric acid cycle. The yeastalso possesses an NAD+ glutamate dehydroge-nase (NAD+-GDH), product of the GDH2 gene.This dehydrogenase is involved in the oxidative
catabolism of glutamate. It produces the inversereaction of the precedent, liberating the ammoniumion used in the synthesis of glutamine. NADP+-GDH activity is at its maximum when the yeastis cultivated on a medium containing exclusivelyammonium as its source of nitrogen. The NAD+-GDH activity, however, is at its highest levelwhen the principal source of nitrogen is gluta-mate. Glutamine synthetase (GS) produces glu-tamine from glutamate and ammonium. This ami-nation requires the hydrolysis of an ATP molecule(Figure 2.21).
Through transamination reactions, glutamatethen serves as an amino group donor in the bio-synthesis of different amino acids. Pyridoxal phos-phate is the transaminase cofactor (Figure 2.22); itis derived from pyridoxine (vitamin B6).
The carbon skeleton of amino acids originatesfrom glycolysis intermediary products (pyruvate,3-phosphoglycerate, phosphoenolpyruvate), the
NH4+ + CH2
C
CH2
COO−
COO−
NADP GDH
NADPH + H+ NADP+
COO−
C
CH2
H+H3N
CH2
COO+
+ H2O
α -ketoglutarate Glutamate
O
Fig. 2.20. Incorporation of the ammonium ion in α-ketoglutarate catalyzed by NADP glutamate dehydrogenase(NADP-GDP)
+NH3 C H
COO−
CH2 + NH4+ + ATP
CH2
COO−
GS
COO−
C H+H3N
CH2
CH2
C O
NH2
+ ADP + Pi + H+
Glutamine
Glutamate
Fig. 2.21. Amidation of glutamate into glutamine by glutamine synthetase (GS)

Biochemistry of Alcoholic Fermentation and Metabolic Pathways of Wine Yeasts 71
−O P
O
O−
NH
OH2C
C
OH
CH3
H O
+NH
H2C
CH2
OH
CH3
NH3+
+
PLD PMP
−O P
O
O−
O
Fig. 2.22. Pyridoxal phosphate (PLP) and pyridoxamine phosphate (PMP)
citric acid cycle (α-ketoglutarate, oxaloacetate) orthe pentose phosphate cycle (ribose 5-phosphate,erythrose 4-phosphate). Some of these reactionsare very simple, such as the formation of aspartateor alanine by transamination of glutamate intooxaloacetate or pyruvate:
oxaloacetate + glutamate −−−→aspartate + α-ketoglutarate (2.5)
pyruvate + glutamate −−−→alanine + α-ketoglutarate (2.6)
Other biosynthetic pathways are more complex,but still occur in yeasts as in the rest of the livingworld. The amino acids can be classified into sixbiosynthetic families depending on their nature andtheir carbon precursor (Figure 2.23):
1. In addition to glutamate and glutamine, prolineand arginine are formed from α-ketoglutarate.
2. Asparagine, methionine, lysine, threonine andisoleucine are derived from aspartate, whichis issued from oxaloacetate. ATP can acti-vate methionine to form S-adenosylmethionine,which can be demethylated to form S-adenosylhomocysteine, the hydrolysis of whichliberates adenine to produce homocysteine.
3. Pyruvate is the starting point for the synthesisof alanine, valine and leucine.
4. 3-Phosphoglycerate leads to the formationof serine and glycine. The condensation ofhomocysteine and serine produces cystathion-ine, a precursor of cysteine.
Oxaloacetate
GlutamateGlutamine
AsparagineLysineThreonine Isoleucine
HomocysteineMethionine
ProlineArginine
3-phosphoglycerate
Ribose 5-phosphate
Pyruvate
PhosphoenolpyruvateErythrose 4-phosphate
α-ketoglutarate
Aspartate
Histidine
Cysteine
PhenylalanineTyrosineTryptophane
AlanineValineLeucine
CystathionineGlycine
Serine
Fig. 2.23. General biosynthesis pathways of aminoacids
5. The imidazole cycle of histidine is formed fromribose 5-phosphate and adenine of ATP.
6. The amino acids possessing an aromaticcycle (tyrosine, phenylalanine, trytophan) arederived from erythrose 4-phosphate and phos-phoenolpyruvate. These two compounds areintermediaries of the pentose cycle and gly-colysis, respectively. Their condensation formsshikimate. The condensation of this compoundwith another molecule of phosphoenolpyruvateproduces chorismate, a precursor of aromaticamino acids.

72 Handbook of Enology: The Microbiology of Wine and Vinifications
2.4.2 Assimilation Mechanisms ofAmmonium and Amino Acids
The penetration of ammonium and amino acidsinto the yeast cell activates numerous membraneproteinic transporters or permeases (Section 1.3.2).S. cerevisiae has at least two specific ammoniumion transporters (Dubois and Grenson, 1979). Theiractivity is inhibited by several amino acids, in anon-competitive manner.
Two distinct categories of transporters ensureamino acid transport:
1. A general amino acid permease (GAP) trans-ports all of the amino acids. The ammonium ioninhibits and represses the GAP. The GAP there-fore only appears to be active during the secondhalf of fermentation, when the must no longercontains ammonium. It acts as a ‘nitrogenscavenger’ towards amino acids (Cartwrightet al., 1989).
2. S. cerevisiae also has many specific aminoacid permeases (at least 11). Each one ensuresthe transport of one or more amino acids.In Contrast to GAP, the ammonium ion doesnot limit their activity. From the beginning ofthe yeast log phase during the first stages offermentation, these transporters ensure the rapidassimilation of must amino acids.
Glutamate and glutamine, crossroads of amino acidsynthesis, are not the only amino acids rapidlyassimilated. Most of the amino acids are practicallydepleted from the must by the time the first 30 g ofsugar have been fermented. Alanine and arginineare the principal amino acids found in must. Yeastsmake use of these two compounds and ammoniumslightly after the depletion of other amino acids.Furthermore, yeasts massively assimilate arginineonly after the disappearance of ammonium fromthe medium. Sometimes, yeasts do not completelyconsume γ -aminobutyric acid. Yeasts do not utilizeproline during fermentation, although it is one ofthe principal amino acids found in must.
During fermentation, yeasts assimilate between1 and 2 g/l of amino acids. Towards the end offermentation, yeasts excrete significant but variable
amounts of different amino acids. Finally, at theend of alcoholic fermentation, a few hundredmilligrams of amino acids per liter remain; prolinegenerally represents half.
Contrary to must hexoses that penetrate thecell by facilitated diffusion, ammonium and aminoacids require active transport. Their concentrationin the cell is generally higher than in the externalmedium. The permease involved couples the trans-port of an amino acid molecule (or ammonium ion)with the transport of a hydrogen ion. The hydro-gen ion moves in the direction of the concentrationgradient: the concentration of protons in the mustis higher than in the cytoplasm. The amino acidand the proton are linked to the same transportprotein and penetrate the cell simultaneously. Thisconcerted transport of two substances in the samedirection is called symport (Figure 2.24). Obvi-ously, the proton that penetrates the cell must thenbe exported to avoid acidification of the cytoplasm.This movement is made against the concentra-tion gradient and requires energy. The membraneATPase ensures the excretion of the hydrogen ionacross the plasmic membrane, acting as a pro-ton pump.
P
ATPase
Plasmic membrane
ADP + Pi
ATP
H+ H+
H+ H+
Amino acidAmino acid
cytosol External medium
Fig. 2.24. Active amino acid transfer mechanisms in theyeast plasma membrane. P = protein playing the role ofan amino acid ‘symporter’

Biochemistry of Alcoholic Fermentation and Metabolic Pathways of Wine Yeasts 73
Ethanol strongly limits amino acid transport. Itmodifies the composition and the properties ofthe phospholipids of the plasmic membrane. Themembrane becomes more permeable. The H+ ionsof the medium massively penetrate the interiorof the cell by simple diffusion. The membraneATPase must increase its operation to control theintracellular pH. As soon as this task monopolizesthe ATPase, the symport of the amino acids nolonger functions. In other words, at the beginningof fermentation, and for as long as the ethanolconcentration in the must is low, yeasts can rapidlyassimilate amino acids and concentrate them inthe vacuoles for later use, according to theirbiosynthesis needs.
2.4.3 Catabolism of Amino AcidsThe ammonium ion is essential for the synthesisof amino acids necessary for building proteins,but yeasts cannot always find sufficient quantitiesin their environment. Fortunately, they can obtainammonium from available amino acids throughvarious reactions.
The most common pathway is the transfer ofan α-amino group, originating from one of many
different amino acids, onto α-ketoglutaric acidto form glutamate. Aminotransferases or transam-inases catalyze this reaction, whose prostheticgroup is pyridoxal phosphate (PLP). Glutamate isthen deaminated by oxidative pathways to formNH4
+ (Figure 2.25). These two reactions can besummarized as follows:
α-amino acid + NAD+ + H2O −−−→α-ketonic acid + NH4
+ + NADH + H+ (2.7)
During transamination, pyridoxal phosphate istemporarily transformed into pyridoxamine phos-phate (PMP). The PLP aldehydic group is linked toa lysine residue ε-amino group on the active site ofthe aminotransferase to form an intermediary prod-uct (E-PLP) (Figure 2.26). The α-amino group of
NADH + NH4+
NAD+ + H2O
Glutamatedehydrogenase
transaminase
α-ketonic acid
α-ketoglutarate
Glutamate
α-amino acid
Fig. 2.25. Oxidative deamination of an amino acid, cat-alyzed by a transaminase and glutamate dehydrogenase
NH
C
OHOH2C
CH3
N
H
(CH2)4
+
P−O
O
O−
Aminotransferase
PLP−aminotransferase
intermediary substance
NH
OH2C
C
N
C
H
COO−R
OH
CH3
P
O
O−
−O
H
+(CH2)4
Aminotransferase
NH3+
NH
CH2
OH
CH
OH2C
NH3+
+
P
O
−O
−O
R CO COO−
PMPKetonic acid
PLP−amino acid intermediary
H2O
Amino acid
+
(E-PLP)
Fig. 2.26. Mode of action of pyridoxal phosphate (PLP) in transamination reactions. Formation of intermediaryproducts between PLP and aminotransferase or the amino acid substrate

74 Handbook of Enology: The Microbiology of Wine and Vinifications
+H3N C
COO−
CH2OH
HH2O
+H3N C
COO−
CH2
CO
COO−
CH3
+ NH4+
H2O
Serine Aminoacylate Pyruvate
Fig. 2.27. Deamination of serine by a dehydratase
HC NH2
COOH
R
C O
COOH
R
C OH
R
C
H
OHHNADH + H+ NAD+
NH3 CO2
Deamination
α-ketonic acid
Reduction
Alcohol
DecarboxylationR
Amino acid Aldehyde
Fig. 2.28. Formation of higher alcohols from amino acids (Ehrlich reactions)
the amino acid substrate of the transamination dis-places the lysine residue ε-amino group linked toPLP. The cleavage of this intermediary product lib-erates PMP and ketonic acid, corresponding to theamino acid substrate. PMP can in turn react withanother ketonic acid to furnish a second amino acidand regenerate pyridoxal phosphate. The partialreactions can be written in the following manner:
amino acid 1 + E-PLP −−−→ketonic acid 1 + E-PMP (2.8)
ketonic acid 2 + E-PMP −−−→amino acid 2 + E-PLP (2.9)
the balance sheet for which is:
amino acid 1 + ketonic acid 2 −−−→ketonic acid 1 + amino acid 2 (2.10)
Some amino acids, such as serine and threonine,possess a hydroxyl group on their β carbon.They can be directly deaminated by dehydration.A dehydratase catalyses this reaction, producingthe corresponding ketonic acid and ammonium(Figure 2.27).
2.4.4 Formation of Higher Alcoholsand Esters
Yeasts can excrete ketonic acids originating fromthe deamination of amino acids only after their
decarboxylation into aldehyde and reduction intoalcohol (Figure 2.28). This mechanism, known asthe Ehrlich reaction, explains in part the formationof higher alcohols in wine. Table 2.4 lists theprincipal higher alcohols and their correspondingamino acids, possible precursors of these alcohols.
Several experiments clearly indicate, however,that the degradation of amino acids is not the onlypathway for forming higher alcohols in wine. Infact, certain ones, such as propan-1-ol and butan-1-ol, do not have amino acid precursors. Moreover,certain mutants deficient in the synthesis of specificamino acids do not produce the correspondinghigher alcohol, even if the amino acid is presentin the culture medium. There is no relationshipbetween the amount of amino acids in must and theamount of corresponding higher alcohols in wine.
Higher alcohol production by yeasts appears tobe linked not only to the catabolism of amino acidsbut also to their synthesis via the correspondingketonic acids. These acids are derived from themetabolism of sugars. For example, propan-1-olhas no corresponding amino acid. It is derived fromα-ketobutyrate which can be formed from pyruvateand acetyl coenzyme A. α-Ketoisocaproate is aprecursor of isoamylic alcohol and an intermediaryproduct in the synthesis of leucine. It too can beproduced from α-acetolactate, which is derivedfrom pyruvate. Most higher alcohols in wine canalso be formed by the metabolism of glucosewithout the involvement of amino acids.

Biochemistry of Alcoholic Fermentation and Metabolic Pathways of Wine Yeasts 75
Table 2.4. The principal alcohols found in wine and their amino acid precursors
Higher alcohol Amino acid precursor
CH3 CH CH2 CH2OH
CH3
CH
CH3
CH3 CH CH2OH
CH3
CH2CH3 CH2OH
CH2 CH
CH2 CH2OHHO
CH3 CH2 CH2OH
CH3 CH2 CH2 CH2OH
NH
CH2 CH2OH
CO CH2 CH2 CH2
O
CH3 S CH2 CH2 CH2OH
3-methyl-butan-1-olor isoamyl alcohol
2-methyl-butan-2-olor active amyl alcohol
2-methyl-propan-1-olor isobutyl alcohol
Phenylethanol
Tyrosol
Propan-1-ol
Butan-1-ol
Thyptophol
γ-Butyrolatone
Methionol
CH3 CH CH2 CH
CH3
COOH
NH2
CH3 CH CH
CH3
CH3 CH2 CH CH COOH
NH2CH3
COOH
NH2
NH2
COOH
CH2 CH
NH2
COOHHO
NH
CH2 CH
NH2
COOH
COOH CH2 CH2 CH COOH
NH2
CH3 S CH2 CH2 CH COOH
NH2
Leucine
Isoleucine
Valine
Phenylalanine
Tyrosine
?
?
Tryptophane
Glutamic acid
Methionine
CH2 CH2OH
80−300
30−100
50−150
10−100
20−50
10−50
1−10
0−1
0−5
0−5
Concentration in wine(mg/l)

76 Handbook of Enology: The Microbiology of Wine and Vinifications
The physiological function of higher alcoholproduction by yeasts is not clear. It may be asimple waste of sugars, a detoxification process ofthe intracellular medium, or a means of regulatingthe metabolism of amino acids.
With the exception of phenylethanol, whichhas a rose-like fragrance, higher alcohols smellbad. Most, such as isoamylic alcohol, have heavysolvent-like odors. Methanol is a peculiar alcoholbecause it contains a sulfur atom. Its cooked-cabbage odor has the lowest perception threshold(1.2 mg/l). It can be responsible for the mostpersistent and disagreeable olfactory flaws ofreduction, especially in white wines. In general, thewinemaker should avoid excessive higher alcoholodors. Fortunately, their organoleptical impact islimited at their usual concentrations in wine, butit depends on the overall aromatic intensity ofthe wine. Excessive yields and rain at the end ofmaturation can dilute the must, in which case thewine will have a low aromatic intensity and theheavy, common character of higher alcohols canbe pronounced.
The winemaking parameters that increase higheralcohol production by yeasts are well known:high pH, elevated fermentation temperature, andaeration. In red winemaking, the extraction ofpomace constituents and the concern for rapidand complete fermentations impose aeration andelevated temperatures, and in this case higheralcohol production by yeast cannot be limited.In white winemaking, a fermentation temperaturebetween 20 and 22◦C limits the formation of higheralcohols.
Ammonium and amino acid deficiencies inmust lead to an increased formation of higheralcohols. In these conditions, the yeast appears torecuperate all of the animated nitrogen availableby transamination. It abandons the unused carbonskeleton in the form of higher alcohols. Rackingwhite must also limits the production of higheralcohols (Chapter 13).
The nature of the yeast (species, strain)responsible for fermentation also affects theproduction of higher alcohols. Certain species,such as Hansenula anomala, have long beenknown to produce a lot, especially in aerobiosis
(Guymon et al., 1961). Yet production bywine yeasts is limited, even in spontaneousfermentation. More recently, various researchershave shown that most S. bayanus (ex uvarum)produce considerably more phenylethanol thanS. cerevisiae. Finally, higher alcohol productionin S. cerevisiae depends on the strain. A limitedhigher alcohol production (with the exception ofphenylethanol) should be among selection criteriafor wine yeasts.
Due to their esterase activities, yeasts form var-ious esters (a few milligrams per liter). The mostimportant acetates of higher alcohols are isoamylacetate (banana aroma) and phenylethyl acetate(rose aroma). Although they are not linked to nitro-gen metabolism, ethyl esters of medium-chain fattyacids are also involved. They are formed by thecondensation of acetyl coenzyme A. These estershave more interesting aromas than the others. Hex-anoate has a flowery and fruity aroma reminiscentof green apples. Ethyl decanoate has a soap-likeodor. In white winemaking, the production of theseesters can be increased by lowering the fermenta-tion temperature and increasing must clarification.Certain yeast strains (71B) produce large quantitiesof these compounds, which contribute to the fer-mentation aroma of young wines. They are rapidlyhydrolyzed during their first year in bottle and haveno long-term influence on the aromatic characterof white wines.
REFERENCES
Alexandre H., Nguyen Van Long T., Feuillat M. andCharpentier C., 1994, Rev. Fr. Œnol., cahiers scien-tifiques, 146, 11–20.
Bely M., Rinaldi A. and Dubourdieu D., 2003, J. Biosci.Bioeng. 96 (6), 507.
Bisson L.F., 1993, Yeasts-Metabolism of sugars, inWine microbiology and biotechnology, p. 55–75,G.H. Fleet., Harwood Academic Publishers, Chur,Switzerland.
Blomberg A. and Alder L., 1992, Adv. Microbiol. Phys-iol., 33, 145–212.
Blondin B., Dequin S., Saint Prix F. and SablayrollesJ.M., 2002, La formation d’acides volatils par les lev-ures, dans 13ieme Symposium international d’œnologie09–10 Juin 2002, INSA, INRA, Montpellier, France.
Carre E., Lafon-Lafourcade S. and Bertrand A., 1983,Conn. Vigne Vin, 17, 43–53.

Biochemistry of Alcoholic Fermentation and Metabolic Pathways of Wine Yeasts 77
Cartwright C.P., Rose A.H., Calderbank J. and KeenanM.H.J., 1989, Solute transport, in The Yeasts, vol. 3,p. 5–55. A.H. Rose, J.S. Harrison, Academic Press,London.
Caubet R., Guerin B. and Guerin M., 1988, Arch.Microbiol., 149, 324–329.
Cooper T.G., 1982, Nitrogen metabolism in Saccha-romyces cerevisiae, in The molecular biology ofthe yeast Saccharomyces: metabolism and geneexpression p. 39–99, J.N. Strathern, E.W. Jones, J.R.Broach., Cold Spring Harbor Laboratory, New York.
Crabtree H.G., 1929, Biochem. J., 23, 536–545.De Revel G., Lonvaud-Funel A. and Bertrand A., 1996,
Etude des composes dicarbonyles au cours des fer-mentations alcoolique et malolactique, in Œnolo-gie 95, 5 e Symposium international d’œnologie,p. 321–325, A. Lonvaud, Tec et Doc, Pairs.
Delfini C. and Cervetti F., 1992, Vitic. Enol. Sci., 46,142–150.
Dubois E. and Grenson M., 1979, Molecular and gen-eral genetics, 175, 67–76.
Dubourdieu D., 1982, Recherches sur les polysaccha-rides secretes par Botrytis cinerea dans la baie deraisin, These Doctorat es Sciences, Universite deBordeaux II.
Fuck E. and Radler F., 1972, Archiv, fur Mikrobiologie,87, 149–164.
Guymon J.F., Ingraham J.L. and Crowell E.A., 1961,Arch. Biochem. Biophys., 95, 163–168.
Kishimoto M., 1994, J. of Fermentation and Bio. engi-neering, 77, 4, 432–435.
Lafon-Lafourcade S., 1983, Wine and Brandy, in Bio-technology, 83–163 H.J. Rehm et G. Reed, VerlagChemie, Weinheim.
Lavigne V., 1996, Recherche sur les composes volatilsformes par la levure au cours de la vinification etde l’elevage des vins blancs secs, These de Doctorat,Universite de Bordeaux II.
Magasanik B., 1992, Regulation of nitrogen utilization,in The molecular biology of the yeast Saccharomyces.
Gene expression, p. 283–317, E.W. Jones, J.R.Pringle, J.R. Broach., Cold Spring Harbor Laboratory,New York.
Magyar I. and Panyik I., 1989, Am. J. Oenol. Vitic. 40,233–240.
Masneuf I., 1996, Recherches sur I’identification geneti-que des levures de vinification. Applications œnologi-ques, These de Doctorat de I’Universite de Bor-deaux II.
Navarro-Avino J.P., Prasad R., Miralles V.J., BenitoR.M. and Serrano R., 1999, Yeast, 15, 929–842.
Neuberg C., 1946, Ann. Rev. Biochem., 15, 435.Oura E., 1977, Process Biochem., 12, 19–21.Peynaud E., Domercq S. and Boidron A., Lafon-
Lafourcade S., Guimberteau G., 1964, Archiv. furMikrobiologie, 48, 150–165.
Remize F., Andrieu E. and Dequin S., 2000, Appl.Environ. Microbiol., 66, 3151–3159.
Ribereau-Gayon J., Peynaud E. and Lafourcade S., 1952,C. R. Acad. Sc. 234, 478.
Ribereau-Gayon P., Lafon-Lafourcade S., Dubour-dieu D., Lucmaret V. and Larue F., 1979, C. R. Acad.Sc., 289 D, 441.
Stryer L., 1992, Metabolisme: concepts et vue d’en-semble, in La biochimie de Lubert Stryer, p. 315–328,Flammarion, Paris.
Taillandier P., Strehaiano P., 1990, Degradation del’acide malique par Schizosaccharomyces, in Actu-alites œnologiques 89, C. R. du 4e Symposium inter-national d’œnologie, Bordeaux 1989, P. Ribereau-Gayon, A. Lonvaud, Dunod-Bordas, Paris.
Van Vuuren H.J.J., Viljoen M., Grobler J., Volschenk H.,Bauer F. and Subden R.E., 1996, Genetic analysisof the Schizosaccharomyces pombe malate perme-ases, Mael and malic enzyme, Maell, genes and theirexpression in Saccharomyces cerevisiae, in Œnolo-gie 95, 5 e Symposium international d’œnologie,p. 195–197, A. Lonvaud, Tech et Doc, Paris.
Verdhuyn C., Postma E., Scheffers W.A. and Van DijkenJ.P., 1990, J. Gen. Micro. 136, 359–403.