growth and nitrogen relations in reciprocal grafts of wild-type and
TRANSCRIPT

Journal of Experimental Botany, Vol. 48, No. 311, pp. 1241-1250, June 1997Journal ofExperimentalBotany
Growth and nitrogen relations in reciprocal grafts ofwild-type and nitrate reductase-deficient mutants of pea(Pisum sativum L. var. Juneau)
Matej Lexa1 and John M. Cheeseman
Department of Plant Biology, University of Illinois, 505 S. Goodwin Ave, Urbana, IL 61801, USA
Received 11 November 1996; Accepted 20 February 1997
Abstract
Plants growing on NO3 differ in the proportion ofNO3~ reduced in their roots and leaves. Using a versionof a two-compartment, two-resource plant growthsimulation model, the possible effects of changing theN assimilation site on plant growth were explored.Subsequently, using an experimental system designedto vary the site of NO3" reduction; chimeras (reciprocalgrafts) of Pisum sativum var. Juneau and its mutant,A317, deficient in nitrate reductase. Plant growth andN metabolism at varying NO3 levels were examined ina hydroponic system. In what is thought to be the firstdirect plant manipulation of this kind, total dry weight,root:shoot ratio, total N and NO3 concentrations, invitro nitrate reductase activity, NO3 in xylem exudates,and leaf chlorophyll content were compared. It wasfound that changing the location of nitrate reductiondid not change growth significantly and caused onlysubtle changes in N concentration. These indiffer-ences occurred in spite of marked changes in growthassociated with NO3 availability and useability. Toachieve this kind of plasticity the plants must havedramatically altered the flow and storage of NO3 andreduced N. The results are compared with existingviews of biomass partitioning and regulation of Nassimilation.
Key words: Nitrogen assimilation site, root:shoot ratio,grafting, mathematical model, biomass partitioning.
Introduction
The allocation of biomass to roots and shoots has longbeen considered a process of physiological and ecological
importance, at least partly for its association with nitrogenmetabolism. Plants have evolved with varying character-istic root-to-shoot ratios (RSR) and physiologicalresponses of nitrogen acquisition and allocation to theenvironment (Smith and Rice, 1983; Stewart et al, 1993;Andrews et al, 1992, 1984). Chapin (1980) summarizedthe response of crop plants and other rapidly growingruderal species to changes in N availability in a blockmodel which linked N uptake and C allocation to rootgrowth. Under N-limited conditions, the increased reten-tion of N in, and allocation of C to roots was recognizedas associated with increased root growth and N intercep-tion, partially compensating for the nutrient stress. Inother species, especially more slowly growing tolerantones, the situation is altered by greater constitutive alloca-tion of biomass to roots and variations in the form of N(ammonium or nitrate) available to the plants. In suchcases, more of the accumulated N, regardless of its form,tends to be incorporated into amino acids in the roots.Some authors have emphasized this difference, categor-izing plants according to their N assimilation strategies(Andrews et al, 1992, 1986). Others have noted thatfollowing forest clearing, pioneer plants tend to behighly responsive to NO3~ availability and to assimilateN, mostly as NO3~", in their leaves; members of climaxcommunities are less responsive to NO3" and are morelikely to be 'root assimilators' (Stewart et al, 1988, 1992).
In general, these categories are not rigid. The strategyemphasized in any individual depends, in most cases, onother environmental factors as well as evolutionarilydetermined physiological propensities. For example, thesite of assimilation depends on the form of N in the rootzone. Plants may use either NH^ or NO3" as their mainsource of N. With NH^", the N must be incorporatedinto amino acids before it is transported to the shoot;
1 To whom correspondence should be addressed: Fax: +1 217 244 7246. E-mail: m.lexa©uiuc.edu
6 Oxford University Press 1997
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/48/6/1241/654720 by guest on 04 April 2019

1242 Lexa and Cheeseman
assimilation occurs only in the root. On the other hand,NO^ may be transported in the xylem and reduced inthe shoot (Runge, 1983; Botella et al, 1993; Peuke et al,1996). The level of NO ~ in the root medium is anotherimportant factor. Oscarson and Larsson (1986), forexample, reported that in high NO3~ conditions whenuptake rates were high, pea plants could not reduce andassimilate all the N in their roots and more was trans-ported to the shoot for reduction.
Environmental modification of NR activity distributionhas also been demonstrated in studies such as that ofDeane-Drummond et al. (1980). In that case, when barleyroot zone temperatures were controlled to be lower thanair temperature (considered more realistic than the uni-form temperature in controlled environments) root NRactivity and the proportion of total NO3~" reduced in theroots increased. In similar conditions, Clarkson et al.(1986) reported adjustments in both relative and absoluterates of uptake of NO ~ and NH^. Using salinity as theenvironmental variable, Botella et al. (1993) and Peukeet al. (1996) reported that relatively more NO ~ wasreduced in Ricinus communis roots at high salt than undernon-saline conditions. This was attributed directly to thedecreased NR activity in the shoot.
Despite understanding several partial responses, thequestion of why some plant species are root reducerswhile others are shoot reducers remains open. One fre-quently suggested advantage of reducing nitrate in theshoot is its lower energy cost than root reduction whenthere is enough light available (Stewart et al, 1988).There are also advantages to reducing nitrate in the roots.Root reducers are less likely to export excess NO ~ to theshoot, thus avoiding potentially harmful effects of excessaccumulation in the leaves (Andrews, 1986a). It also hasbeen postulated that plants that rely on ammonium Nsources most of their lives may be better equipped toassimilate reduced N in the roots, since NH^ is nottransported to the shoot in large quantities. Such plantsmay preferably perform NOJ reduction in the same plantpart (Stewart et al, 1988). Whether and how theseadvantages and disadvantages translate into the presenceof certain species in certain ecosystems or habitats remainsto be seen. While Andrews (19866) recognized generaldifferences, mainly between tropical and temperatespecies, Stewart et al. (1992) concluded that no preciseboundary can be drawn in terms of the preferred habitatof root and leaf reducers. They mentioned, however, thatleaf reducers tend to be herbaceous early colonizers andforest gap species.
Paralleling these experimental and ecological studiesover the last 25 years has been the modelling of root:shootpartitioning and N and C allocation (Thornley 1972,1976; Hirose, 1986; Keulen and Wolf, 1986; Cheeseman,1993; Cheeseman et al, 1996; Agren and Ingestad, 1987).Regardless of the details or strategies of the models and
despite their success at describing realistic biomass parti-tioning, they differ from plants themselves by the condi-tion that all N be incorporated in the roots. None of theexisting models allows either for NO^ transport to shootsor its reduction there. One good reason for these limita-tions is uncertainty concerning actual or potential mech-anisms for the long-distance control of root activities byshoot N status (Cheeseman et al, 1996).
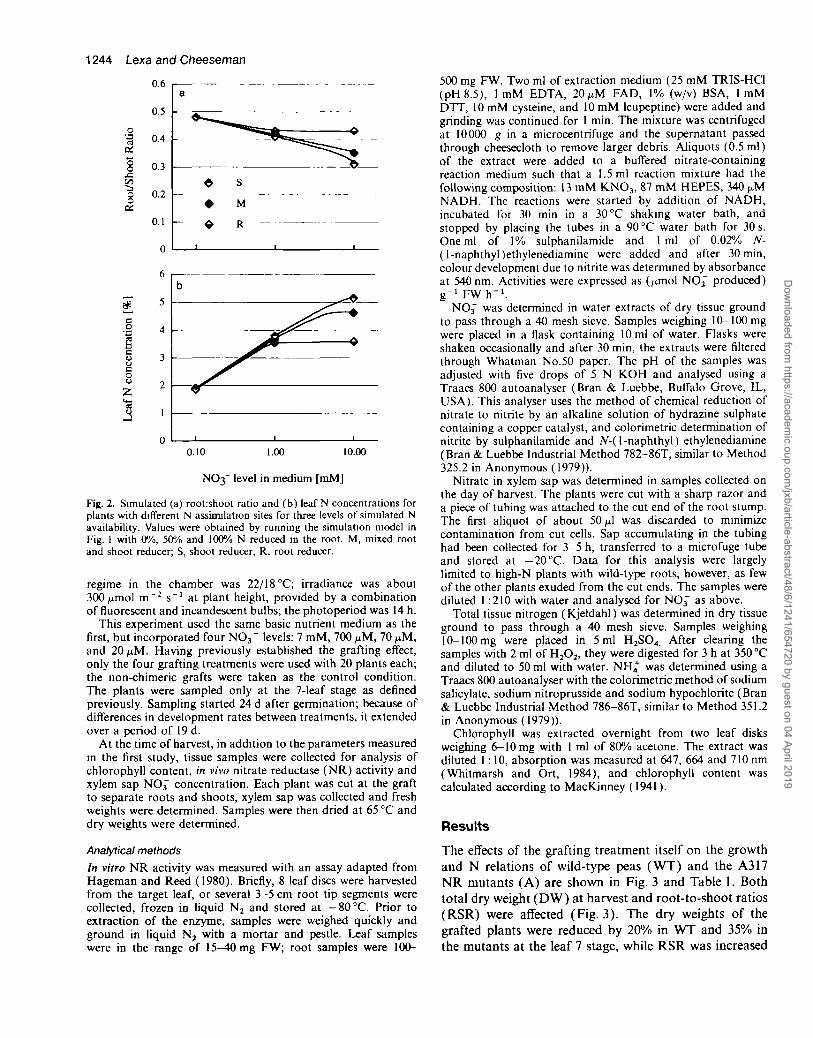
To address these deficiencies, a programme combininggrowth modelling with what is believed to be the firstdirect manipulation of the distribution of NR activitybetween roots and shoots was undertaken. For the simula-tions, the two-compartment, two-resource construct wasmodified from previous studies (Cheeseman, 1993)allowing manipulation of the site of N assimilation. Themodel is shown schematically in Fig. 1. By manipulatingNR activity coefficients, this model allows partitioningof NO3~ reduction in any proportions between roots andshoots. The results of the two extreme cases, full reductionin either the roots or shoots, and an intermediate one areshown in Fig. 2. Overall, the three simulated plants hadsimilar growth rates, but there were two responses whichcould be meaningful with respect to the importance of Nreduction site. With higher N input, the root reduceraccumulated relatively more biomass in the roots (Fig. 2a)and had a lower leaf N concentration than the shootreducer (Fig. 2b). A more detailed description and work-ing versions of the model (downloadable Stella formatand an HTML interface to Java code) are available electron-ically at http://www. life. uiuc. edu/cheeseman/NRsite
In this paper results of experiments to manipulate thesite of NO3~ reduction are reported and compared to themodelling results. For the manipulations, wild type andNR-deficient mutants of peas (Pisum sativum L. var.Juneau) were grafted to produce chimeras with differentpatterns and capacities for nitrogen assimilation. Thissystem was selected because of the ease with which peascan be grafted (Went, 1938, 1943) and the availabilityof the appropriate NR mutants (Warner et al, 1982;Kleinhofse/a/., 1978).
Materials and methods
Plant material
Wild-type and A317 mutant pea seeds (Pisum sativum L. var.Juneau) (Warner et al, 1982; KJeinhofs et al, 1978) wereobtained from Drs Barry Shelp and Brent Kaiser (Departmentof Horticultural Science, University of Guelph, Canada),propagated in our facilities and stored at 4 °C until needed. Forgermination, the seeds were soaked in concentrated sulphuricacid for 6 min, washed in aerated water for 24 h and planted inwet vermiculite in a dark growth chamber (22-18 °C). Inpreparation for grafting, the plants were kept in the dark aftergermination until their epicotyls were 2-4 cm long.
The grafting protocol was modified from Derma (1962).Seedlings were cut with a sharp razor blade in the middle of
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/48/6/1241/654720 by guest on 04 April 2019

nitrate N
NRreduced N
nitrate N
NR
U
Nitrate reduction in reciprocal grafts 1243
SHOOT
reduced N
ROOT
Fig. 1. Simplified scheme of the C + N model, modified from Cheeseman (1993) to allow manipulation of the site of N reduction. Rectanglesrepresent state variables (NO^, C, N, protein and carbohydrate content). Valves represent points of control (U, nitrate uptake; NR, reduction; P,photosynthesis). Arrows represent flows of C and N as translocation, reduction or incorporation into macromolecules. The scheme presented herewas transformed into differential equations that were solved numerically for individual simulations using Stella modelling software (HighPerformance Systems, USA, 1990). Details and working versions of this model are available on the Internet at hitp://www.life.uiuc.edu/cheeseman/NRsite/
the elongated epicotyl at an angle of 20-30° to the axis of theplant creating a stock (a root with cotyledons attached) and ascion (consisting of the rest of the epicotyl and the hook).Segments of polyethylene tubing about a centimetre long withdiameter slightly smaller than that of the epicotyl were used tojoin the stocks and scions. The tubing was slit along its axis toallow for expansion as the plants grew. Following grafting, theseedlings were replanted in vermiculite, covered with perforated
transferred to each container at the beginning of the experiment.Solution flowed through the system at approximately 301min"1; this flow was evenly distributed to the plant containersusing a system of flow balancing valves. Parallel systems wereused for solutions at two NO^" levels, 70 and 700 fiM; thesolutions were changed twice a week. The day/night temper-atures in the greenhouse were set to 22/18 °C; daylight wassupplemented by metal halide lamps that were switched on
plastic film and placed under low light (~100 junol m 2 s ' ) . whenever light intensity fell below 1000/*mol m 2 s \ main-Thereafter, the plants were checked daily and lateral budsgrowing from the axils of the cotyledons were excised to preventtheir interference with the later studies. After 5 d, the cotyledonswere removed and the grafted seedlings were transferred tosolution culture.
The basic nutrient solution was 330 ^M NaCl, 140/xMKNO3, 280 ,xM Ca(NO3)2 110MMMgSO4 190 MM KH2PO4)
18/xM FeNa-EDTA, 3.2 MM H3BO3, 1.1 ^M MnSO4, 0.06 ^MZnSO4, 0.07 ^M CuSO4> and 0.04 ^M MoO3. The pH of thesolution was uncontrolled and varied between 6.5 and 8.5. Allnitrogen was in the form of NO^, initially at a concentrationof 700 ^M. To obtain lower levels of NO3"~, nitrate salts werereplaced with chloride salts of the same molarity. For growthat NOf above 700 /iM, the solution was supplementedwith KNO3.
Experimental protocols
Results will be presented for two separate experiments. Thefirst experiment was performed in a greenhouse using twoflowing solution hydroponic systems (Topa and Cheeseman,1992). Each system consisted of a solution tank (2001) and aset of 30 plant containers (51 each). Three plants were
taining a minimum irradiance at plant height of 900 ^mol m"s"1 and a 14 h photoperiod. The plants were placed in thegreenhouse on 10 April 1995.
The effects of grafting were assessed using six grafting andcontrol treatments designated as follows: W, ungrafted wild-type control plants; A, ungrafted A317 mutant controls; WW,wild-type roots and shoots grafted to each other; AA, mutantroots and shoots grafted to each other; AW, mutant shootgrafted to the wild-type root; and WA, wild-type shoot graftedto the mutant root. There were 30 plants in each treatment.The plants were harvested at two developmental stages, definedby the ages at which 7 or 12 leaves on the main stem were fullyopen and the leaflets of the following leaf were still folded.Sampling for leaf 7 started on 5 May and ended 10 May.Sampling for leaf 12 started 15 May and ended 21 May.
The second experiment started 20 September 1995, and wasdesigned to examine the effects of varying N availability over awider range. Plants were grown in a growth chamber (Conviron,Model El5, Pembina, ND) in a simple, frequent-solution-change hydroponic system. This consisted of individual 21compartments, each supporting five plants during the experi-ment. Solutions were changed daily. The day/night temperature
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/48/6/1241/654720 by guest on 04 April 2019
1244 Lexa and Cheeseman
0.6
0.5 —
o'ii °-4c6
8 0.3
I 0.206
0.1
0
a
-
—
*
1
s
M
R
• i
1Ioo
0.10 1.00 10.00
NO3" level in medium [mM]
Fig. 2. Simulated (a) root:shoot ratio and (b) leaf N concentrations forplants with different N assimilation sites for three levels of simulated Navailability. Values were obtained by running the simulation model inFig. 1 with 0%, 50% and 100% N reduced in the root. M, mixed rootand shoot reducer; S, shoot reducer, R, root reducer.
regime in the chamber was 22/18 °C; irradiance was about300/xmol m" 2 s"1 at plant height, provided by a combinationof fluorescent and incandescent bulbs; the photoperiod was 14 h.
This experiment used the same basic nutrient medium as thefirst, but incorporated four NO3~ levels: 7 mM, 700 M, 70 ^M,and 20 fjM. Having previously established the grafting effect,only the four grafting treatments were used with 20 plants each;the non-chimeric grafts were taken as the control condition.The plants were sampled only at the 7-leaf stage as definedpreviously. Sampling started 24 d after germination; because ofdifferences in development rates between treatments, it extendedover a period of 19 d.
At the time of harvest, in addition to the parameters measuredin the first study, tissue samples were collected for analysis ofchlorophyll content, in vivo nitrate reductase (NR) activity andxylem sap NO^ concentration. Each plant was cut at the graftto separate roots and shoots, xylem sap was collected and freshweights were determined. Samples were then dried at 65 °C anddry weights were determined.
Analytical methods
In vitro NR activity was measured with an assay adapted fromHageman and Reed (1980). Briefly, 8 leaf discs were harvestedfrom the target leaf, or several 3-5 cm root tip segments werecollected, frozen in liquid N2 and stored at - 8 0 °C. Prior toextraction of the enzyme, samples were weighed quickly andground in liquid N2 with a mortar and pestle. Leaf sampleswere in the range of 15^40 mg FW; root samples were 100-
500 mg FW. Two ml of extraction medium (25 mM TRIS-HC1(pH8.5), 1 mM EDTA, 20 MM FAD, 1% (w/v) BSA, 1 mMDTT, 10 mM cysteine, and 10 mM leupeptine) were added andgrinding was continued for 1 min. The mixture was centrifugedat 10000 g in a microcentrifuge and the supernatant passedthrough cheesecloth to remove larger debris. AJiquots (0.5 ml)of the extract were added to a buffered nitrate-containingreaction medium such that a 1.5 ml reaction mixture had thefollowing composition: 13 mM KN0 3 , 87 mM HEPES, 340 /xMNADH. The reactions were started by addition of NADH,incubated for 30 min in a 30 °C shaking water bath, andstopped by placing the tubes in a 90 °C water bath for 30 s.One ml of 1% sulphanilamide and 1 ml of 0.02% N-(l-naphthyl)ethylenediamine were added and after 30 min,colour development due to nitrite was determined by absorbanceat 540 nm. Activities were expressed as (^mol NO^~ produced)g"1 F W h " 1 .
NO3- was determined in water extracts of dry tissue groundto pass through a 40 mesh sieve. Samples weighing 10-100 mgwere placed in a flask containing 10 ml of water. Flasks wereshaken occasionally and after 30 min, the extracts were filteredthrough Whatman No. 50 paper. The pH of the samples wasadjusted with five drops of 5 N KOH and analysed using aTraacs 800 autoanalyser (Bran & Luebbe, Buffalo Grove, IL,USA). This analyser uses the method of chemical reduction ofnitrate to nitrite by an alkaline solution of hydrazine sulphatecontaining a copper catalyst, and colorimetric determination ofnitrite by sulphanilamide and TV-(l-naphthyl) ethylenediamine(Bran & Luebbe Industrial Method 782-86T, similar to Method325.2 in Anonymous (1979)).
Nitrate in xylem sap was determined in samples collected onthe day of harvest. The plants were cut with a sharp razor anda piece of tubing was attached to the cut end of the root stump.The first aliquot of about 50 /A was discarded to minimizecontamination from cut cells. Sap accumulating in the tubinghad been collected for 3-5 h, transferred to a microfuge tubeand stored at — 20 °C. Data for this analysis were largelylimited to high-N plants with wild-type roots, however, as fewof the other plants exuded from the cut ends. The samples werediluted 1:210 with water and analysed for NO3~ as above.
Total tissue nitrogen (Kjeldahl) was determined in dry tissueground to pass through a 40 mesh sieve. Samples weighing10-100 mg were placed in 5 ml H2SO4. After clearing thesamples with 2 ml of H2O2, they were digested for 3 h at 350 °Cand diluted to 50 ml with water. NH^ was determined using aTraacs 800 autoanalyser with the colorimetric method of sodiumsaJicylate, sodium nitroprusside and sodium hypochlorite (Bran& Luebbe Industrial Method 786-86T, similar to Method 351.2in Anonymous (1979)).
Chlorophyll was extracted overnight from two leaf disksweighing 6-10 mg with 1 ml of 80% acetone. The extract wasdiluted 1:10, absorption was measured at 647, 664 and 710 nm(Whitmarsh and Ort, 1984), and chlorophyll content wascalculated according to MacKinney (1941).
Results
The effects of the grafting treatment itself on the growthand N relations of wild-type peas (WT) and the A317NR mutants (A) are shown in Fig. 3 and Table 1. Bothtotal dry weight (DW) at harvest and root-to-shoot ratios(RSR) were affected (Fig. 3). The dry weights of thegrafted plants were reduced by 20% in WT and 35% inthe mutants at the leaf 7 stage, while RSR was increased
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/48/6/1241/654720 by guest on 04 April 2019

Nitrate reduction in reciprocal grafts 1245
1250
£/e
ight
dry
vta
l1000
750
500
250
0
• Ungrafted
D Grafted
WT
0.80
§ 040
Qi 0.20
0.00WT A
Grafting treatment
Fig. 3. The effect of grafting on (a) dry weight and (b) root.shoot ratioof wild type and A317 mutants. WT, wild type; A, A317 NR-deficientmutant of Piston sativum L. var. Juneau. Grafted plants were geneticallyuniform. Error bars indicate the standard error of the mean.
by 15% and 30% respectively. However, there were nografting effects on total N contents, unreduced NO3~concentrations or specific NR activities (Table 1). Theshoots of both grafted and ungrafted mutants containedan order of magnitude more unreduced NO^~ than thewild-type plants; root concentrations were even higherthough they were also highly variable. As expected, NRactivity in the mutants was very low and the WT activitieswere largely associated with the shoots. When total plantNR activities were compared, differences were more
4000 r
r r 3000
a.E
'5
2000
1000
ww WA
Grafting treatment
AW
Fig. 4. The effect of age and N assimilation site on dry weightaccumulation in grafted pea seedlings. Total dry weights were determinedat the leaf 7 and leaf 12 stages. WW, wild-type control; WA, shootreducer; AW, root reducer, AA, A317 mutant control. Error barsindicate the standard error of the mean.
apparent (Table 1), reflecting the size reductions associ-ated with grafting (Fig. 3).
The growth patterns of the grafted wild-type plants,the mutants and two chimeras at the leaf 7 and leaf 12stages are shown in Fig. 4 for the plants growing in700 /xM NOf medium. As expected, the mutants (AA)were the smallest, while the grafted wild-types (WW)were the largest. The chimeras had sizes more comparableto those of the wild-type (WW), with no significantdifferences at the leaf 7 stage. At the leaf 12 stage, bothchimeras (AW, WA) were smaller than the WW control.In the low N (70 ^M) medium, the chimeras were slightlybigger than WW control, though the differences were notstatistically significant.
The differences in growth rates between the chimeras(Fig. 4) were consistent with the major localization ofNR to the shoots in the wild-type peas (Table 1), andthe effect of NR localization was examined in more detail
Table 1. The effect of grafting and presence of nitrate reductase on nitrogen and nitrate contents and specific and total nitrate reductaseactivity (NRA) in leaves (L) and roots (R) of peas
Wild type = Pisum sativum L. var. Juneau; mutant = A317 mutant of var. Juneau. Total NRA was calculated as a product of the dry weight andspecific NRA of a given plant part. Data are means (SEM).
Wild type Mutant
LRLRLRLR
Ungrafted
5.2 (0.2)3.8(0.1)
16(7)33(6)8.6 (0.7)1.0(0.2)
37.85.7
Grafted
4.8 (0.1)3.8(0.1)
26(7)43(14)6.3 (0.7)0.5 (0.1)
20.22.3
Ungrafted
4.0 (0.2)3.1 (0.1)
246 (87)512(215)
0.2(0.1)0.1 (0.1)0.40.3
Grafted
3.9 (0.1)3.0 (0.2)
270 (115)1209 (673)
0.2(0.1)0.0(0.1)0.30.0
Nitrogen[%DW]
Nitrate[ppm]
NRA
Total NRA
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/48/6/1241/654720 by guest on 04 April 2019
1246 Lexa and Cheeseman
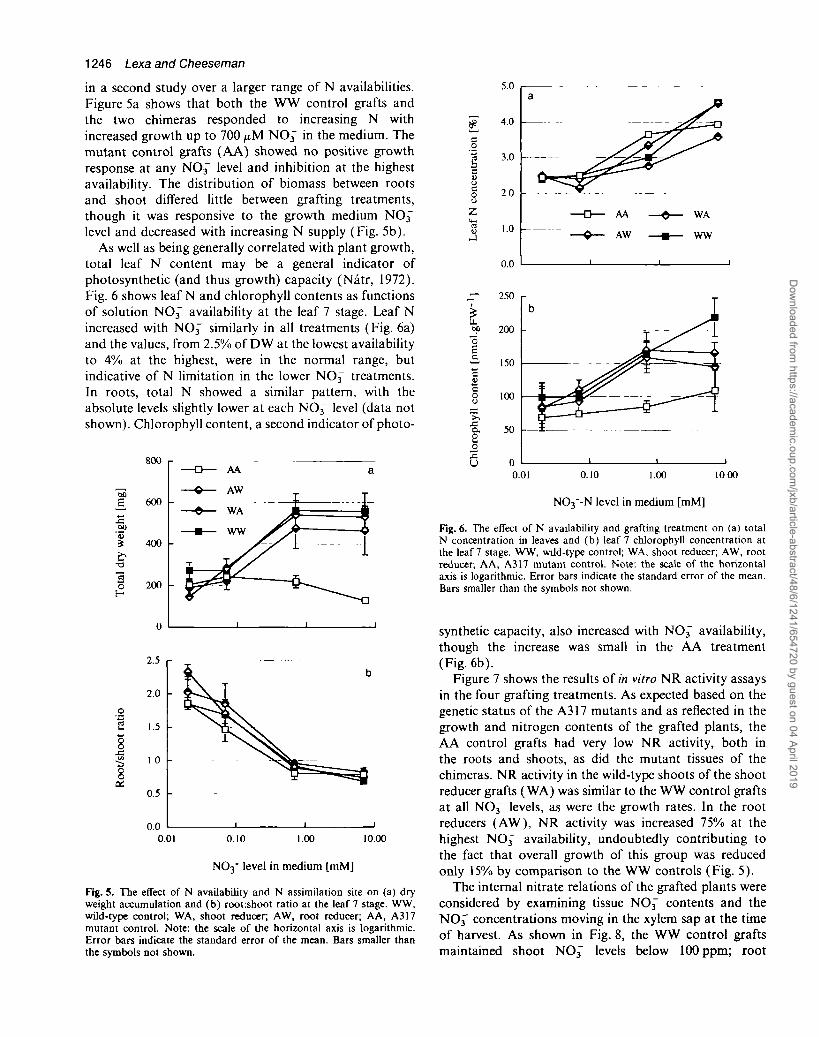
in a second study over a larger range of N availabilities.Figure 5a shows that both the WW control grafts andthe two chimeras responded to increasing N withincreased growth up to 700 M NO3~" in the medium. Themutant control grafts (AA) showed no positive growthresponse at any NO3~ level and inhibition at the highestavailability. The distribution of biomass between rootsand shoot differed little between grafting treatments,though it was responsive to the growth medium NO3~level and decreased with increasing N supply (Fig. 5b).
As well as being generally correlated with plant growth,total leaf N content may be a general indicator ofphotosynthetic (and thus growth) capacity (Natr, 1972).Fig. 6 shows leaf N and chlorophyll contents as functionsof solution NO3~ availability at the leaf 7 stage. Leaf Nincreased with NO3" similarly in all treatments (Fig. 6a)and the values, from 2.5% of DW at the lowest availabilityto 4% at the highest, were in the normal range, butindicative of N limitation in the lower NO3~ treatments.In roots, total N showed a similar pattern, with theabsolute levels slightly lower at each NO3~ level (data notshown). Chlorophyll content, a second indicator of photo-
00
'5
600
400
200
n
— D — AA
— « — AW
— $ — WA
_»_ ww^X
I
a
TA— 1
2.5
2.0
o
e 1.5
80.5
0.00.01 0.10 1.00 10.00
NO3" level in medium [mM]
Fig. 5. The effect of N availability and N assimilation site on (a) dryweight accumulation and (b) rootishoot ratio at the leaf 7 stage. WW,wild-type control; WA, shoot reducer; AW, root reducer; AA, A317mutant control. Note: the scale of the horizontal axis is logarithmic.Error bars indicate the standard error of the mean. Bars smaller thanthe symbols not shown.
5.0
4.0
3.0
20
1.0
0.0
JT 250 r-
o.
U0.01 0.10 1.00 1000
NO3~-N level in medium [mM]
Fig. 6. The effect of N availability and grafting treatment on (a) totalN concentration in leaves and (b) leaf 7 chlorophyll concentration atthe leaf 7 stage. WW, wild-type control; WA, shoot reducer; AW, rootreducer; AA, A317 mutant control. Note: the scale of the horizontalaxis is logarithmic. Error bars indicate the standard error of the mean.Bars smaller than the symbols not shown.
synthetic capacity, also increased with NO3 availability,though the increase was small in the AA treatment(Fig. 6b).
Figure 7 shows the results of in vitro NR activity assaysin the four grafting treatments. As expected based on thegenetic status of the A317 mutants and as reflected in thegrowth and nitrogen contents of the grafted plants, theAA control grafts had very low NR activity, both inthe roots and shoots, as did the mutant tissues of thechimeras. NR activity in the wild-type shoots of the shootreducer grafts (WA) was similar to the WW control graftsat all NO3~ levels, as were the growth rates. In the rootreducers (AW), NR. activity was increased 75% at thehighest NO3" availability, undoubtedly contributing tothe fact that overall growth of this group was reducedonly 15% by comparison to the WW controls (Fig. 5).
The internal nitrate relations of the grafted plants wereconsidered by examining tissue NO3" contents and theNO3~ concentrations moving in the xylem sap at the timeof harvest. As shown in Fig. 8, the WW control graftsmaintained shoot NO3" levels below 100 ppm; root
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/48/6/1241/654720 by guest on 04 April 2019

—f.
Oil
imol
a£Z
Leaf
in
SCI
E
tro
NR
A
r*
Roo
t ii
8.0
7.0
6.0
5.0
4.0
3.0
2.0
1.0
0.0
1.8
1.6
1.4
1.2
1.0
0.8
0.6
0.4
0.2
0.0
WW AW WA AA
• 7.0 mM NO3"
• 0.7mMNO3-
• 0.07 mM NO3-
• 0.02 mM NO3-
WW AW WA
Grafting treatment
Fig. 7. In vitro nitrate reductase activity in (a) leaves and (b) roots ofgrafted pea seedlings at the leaf 7 stage. Plants were grown at fourdifferent N availabilities. WW, wild-type control; WA, shoot reducer;AW, root reducer; AA, A317 mutant control. Note: the vertical scalesof the two graphs are not identical. Error bars indicate the standarderror of the mean.
NO3 levels were linearly related to the levels of NO3 inthe medium. The AA control grafts accumulated muchmore NO^, the difference being more apparent in theshoots. In chimeric grafts, the wild-type part of the plantmimicked the response of the WW control. The mutantpart, however, had an intermediate response. At low N,the concentration of NO^ in the tissue was similar to theWW controls, but at high N it was similar to the AAgrafts. Plant parts that accumulated large amounts ofNO3~ occasionally displayed necrosis; most of these wereAA plants (data not shown).
Because spontaneous exudation of xylem sap fromdetopped root systems occurred primarily from the WWcontrols and AW chimeras, rather than from all treat-ments (i.e. mostly wild-type roots) and mostly at high N,analysis of xylem sap provided only limited, though,useful information. In the highest NO3" treatment, thelevels in both WW and AW sap ranged from 10 to15 mM NO^. They were slighly lower (6 to 12 mM) in
O.a.
O
100000
10000
1000
100
10
1
Nitrate reduction in reciprocal grafts 1247
8
a.a.
i
oz
100000 r-
10000 -
1000
0.10 1.00 10.00
NO3" level in medium [mM]
Fig. 8. The effect of N availability and grafting treatment on tissuenitrate concentration of (a) leaves and (b) roots at the leaf 7 stage.WW, wild-type control; WA, shoot reducer; AW, root reducer; AA,A317 mutant control. Note: the scale of the horizontal axis islogarithmic. Error bars indicate the standard error of the mean. Barssmaller than the symbols not shown.
the 700 fiM treatment. The shoot reducer (WA) in 7 mMNO^, had xylem NO^ more than twice as high (34 mM).These data made it possible to estimate, at least crudely,the proportion of NO^ reduced in the roots. Using theexperimental data on DW and N concentration at harvestaveraged over the entire study period, we estimated thegrowth rate and N uptake, and from that calculated theaverage daily N flow to the shoot. Assuming a transpir-ation coefficient of 100 I/kg, the average NO3" concentra-tions in the transpiration stream are summarized inTable 2.
Discussion
In this study, the objective was to explore the relationshipbetween allocation of biomass to roots and shoots andthe site of primary nitrate reduction by directly manipulat-ing the latter. In what is believed to be the first study ofthis sort, using wild-type and NR mutant pea, growthand N use by plants that were either root- or shoot-reducers as a result of grafting were compared.
The grafting procedure itself changed growth (Fig. 3).
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/48/6/1241/654720 by guest on 04 April 2019

1248 Lexa and Cheeseman
Table 2. Estimated proportion of nitrate uptake reduced in theroots of grafted pea plants
The amount of N needed for the observed growth was calculated fromDW and tissue N contents at the time of harvest. The respectiveconcentrations of N (in ppm) in the transpiration stream needed tosupply that N were calculated to a first approximation based on anestimated transpiration coefficient of 1001 kg"1. The transpirationcoefficient is a measure of the water transpired for a unit increase indry mass. This value is reasonable, though low for C3 plants in general,given the high humidity (80-90%) and the absence of wind in thegrowth chamber. The estimated values were compared with themeasured xylem sap nitrate to yield an estimate of the proportion of Nreduced in the root. Data shown only for those treatments from whichxylem exudate could be collected.
NO3"
7mM
700 MM
70 MM
Treatment
WWAWWAWWAAWW
Sap nitrate(calculated)
500490415354354275
Sap nitrate(measured)
160196469
9213822
Reduced in theroot [%]
6860
<0746192
Comparison of grafted and ungrafted plants showed thatgrafting increased the RSR of the experimental plantsand slightly decreased their DW when compared toungrafted plants. Based on observations of the younggrafts, this was a direct result of the interruption of shoot(scion) growth for several days immediately after grafting.The roots (stock), being attached to the seed and havingresources available, kept growing, whereas the shootresumed full growth only after establishing a graft connec-tion. On the other hand, by the time that the plants wereharvested, there were no grafting-related differencesbetween N content, unreduced NO ~ concentration orNR activity. It was therefore concluded that grafting is areasonable approach to the question of N assimilation inroots and shoots, provided non-chimeric (same genotype)grafts are used as control.
Surprisingly, the experiments showed that distributionof NR activity between roots and shoots had no effecton the resulting RSR of the manipulated plants. Theseresults are difficult to reconcile with existing models ofRSR control (Thornley, 1972; Cheeseman, 1993).Simulations with the model predicted differences betweenreciprocal grafts (Fig. 2) that did not materialize in theexperiments. This lack of match could be resolved, how-ever, if each of the reduced C and N pools were modelledas a single pool. Having just one pool with rapid mixingwould eliminate the significance of the N assimilationsite. A one compartment model would also reflect theexperimental results of Cooper and Clarkson (1989), whohave shown nutrient cycling between roots and shootsusing 15N. Similarly, Heuvelink (1995) has shown thatselective removal of fruits in tomato results in the samefruit size regardless of which ones are removed. He
interpreted this result as indicative of one commonassimilate pool.
Alternatively, the controlling mechanism for RSRcould bypass the internal N pool by a secondary signal,perhaps hormonal, such as cytokinin (Werf and Nagel,1996), if its production rate was dependent on externalrather than internal N. As noted by Marschner et al.(1996), in models of this category, conditions leading toincreases in N concentration in the plant should favourbiomass partitioning towards the shoot. The results haveshown that regardless of the location or presence of NR,all the plants in a given N treatment had the same RSR(Fig. 5b), but not DW (Fig. 5a). It appears, therefore,that while overall growth is a function of the internalavailability of N to the plant (which depends on externalavailability), RSR is not. This, of course, is not what onewould expect by applying a Thornley type model (includ-ing ours, Fig. 1), where RSR is determined internally byavailability of reduced N to biochemical processes.Interestingly, other authors have perhaps recognized thesedifficulties and circumvented them by using empiricalfunctions to describe partitioning (Hirose, 1986).
Higher values of root NR activity in root reducers(AW) than in wild-type controls (WW) show that theroots can also integrate their physiological response withthe shoot; a low shoot NR activity is compensated forpartly by an increase in root NR activity. Though in thisstudy NO3~ translocated to the shoot may be physiologic-ally lost for the root reducer, only in the very high Ntreatment did its accumulation in the leaves match thatof mutant plants (Fig. 8a). Instead, the plants apparentlyadapted by increasing root NR activity by 75% (Fig. 7b)compared to WW roots. If it is assumed, for now, thatthe amount of N reduced by a plant is directly propor-tional to its NR activity, then, without this up-regulationof NR, the plants would have reduced about 45% lesstotal N. This compensatory response must, presumably,be due to a signal from the shoot generated in responseto the overflowing NO^~. Although Ben-Zioni et al. (1971)proposed a model by which malate from the shoot couldbe involved in regulation of NO " uptake, and the workof Cooper and Clarkson (1989) provides for regulationof NR by circulating amino acids, the exact mechanismresponsible for the effect has yet to be determined.Functioning of a similar regulatory mechanism has beenrecently reported in respect to pea iron reductase. Grusakand Pezeshgi (1996) demonstrated that the dgl gene inpeas was responsible for the production of a transmissiblesignal involved in regulating root Fe(III) reductase.
While this study will make it possible to modify theway plant growth models are studied and the way theytreat partitioning, root:shoot communication and regula-tion, it did not provide an answer to the question posedin the Introduction of why some plants are root reducersand others are not. The initial simulation results suggested
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/48/6/1241/654720 by guest on 04 April 2019

that DW and RSR should be sensitive to the site of Nassimilation. However, as no significant differences werefound in either parameter between root and shootassimilators, it is difficult at this point to discuss furtherevolutionary advantages of either strategy. Potentialdifferences in the efficiency of the use of NH^ assimilationenzymes or advantages in relation to photosyntheticproduction of carbon skeletons are alternative hypothesesthat were beyond the scope of the present study, butwairant further investigation.
In conclusion, the results of this study indicate thatplants can regulate NR activity and flows of NO3~" andorganic N between roots and shoots in a way thatminimizes differences in growth based on the assimilationsite. The most important seems to be the ability toincrease assimilation in the roots when the shoot is gettingtoo much NO^~. This regulation is reflected in consider-able differences of NR activity and NO^~ concentrationin the xylem sap, leading to small changes in the overallcapacity to reduce N. A mechanism ensuring that bothorgans have equal access to resources (in this case N)regardless of the reduction site is apparently present inthe plant. As a result, very little difference in overallgrowth is seen. Originally, a question was posed aboutwhy some plants are shoot reducers while others are not.Because it was found that the assimilation site had littleeffect on growth and partitioning, these are unlikely tobe the factors responsible for evolution of root and shootreducers and this problem clearly requires further exam-ination. Finally, while most of the interest in recent yearshas been in signalling from the root to the shoot, theresults indicate the need to also address communicationin the opposite direction.
Acknowledgements
We thank Barry Shelp and Brent Kaiser from University ofGuelph, Ontario, Canada for kindly providing seeds of peasused in this work. We also thank Tom Jacobs and Don Ort foruseful comments in the preparatory stage of this work. Partialsupport for this project was provided by Sigma Xi Grant-in-Aid-of-Research
References
Agren GI, Ingest ad T. 1987. Root:shoot ratio as a balancebetween nitrogen productivity and photosynthesis. Plant, Celland Environment 10, 579-86.
Andrews M. 1986a. Nitrate and reduced-N concentrations inthe xylem sap of Stellaria media, Xanthium strumarium andsix legume species. Plant, Cell and Environment 9, 605-8.
Andrews M. 19866. The partitioning of nitrate assimilationbetween root and shoot of higher plants. Plant, Cell andEnvironment 9, 511-19.
Andrews M, Morton JD, Lieffering M, Bisset L. 1992. Thepartitioning of nitrate assimilation between root and shoot
Nitrate reduction in reciprocal grafts 1249
of a range of temperate cereals and pasture grasses. Annalsof Botany 70, 271-6.
Andrews M, Sutherland JM, Thomas RJ, Sprent JI. 1984.Distribution of nitrate reductase activity in six legumes: theimportance of the stem. New Phytologist 98, 301-10.
Anonymous. 1979. Methods for tlie chemical analysis of waterand wastes, 3rd edn. Washington DC: US EnvironmentalProtection Agency.
Ben-Zioni A, Vaadia Y, Lips SH. 1971. Nitrate uptake by rootsas regulated by nitrate reduction in the shoot. PhysiologiaPlantarum 24, 288-90.
Botella MA, Cruz C, Martins Loucao MA, Cerda A. 1993.Nitrate reductase activity in wheat seedlings as affected byNO^~/NH^ ratio and salinity. Journal of Plant Physiology142, 531-6.
Chapin HI SF. 1980. The mineral nutrition of wild plants.Annual Review of Ecology and Systematic^ 11, 233-60.
Cbeeseman JM. 1993. Plant growth modelling without integrat-ing mechanisms. Plant, Cell and Environment 16, 137—47.
Cheeseman JM, Barreiro R, Lexa M. 1996. Plant growthmodelling and the integration of shoot and root activitieswithout communicating messenger-opinion. Plant and Soil185, 51-64.
Clarkson DT, Hopper MJ, Jones LHP. 1986. The effect of roottemperature on the uptake of nitrogen and the relative sizeof the root system in Lolium perenne. I. Solutions containingboth NH4
+ and NO3". Plant, Cell and Environment 9, 535^15.Cooper HD, Clarkson DT. 1989. Cycling of amino-nitrogen and
other nutrients between shoots and roots in cereals: a possiblemechanism integrating shoot and root in the regulation ofnutrient uptake. Journal of Experimental Botany 40, 753-62.
Deane-Dnimmond CE, Clarkson DT, Johnson CB. 1980. Theeffect of differential root and shoot temperature on the nitratereductase activity, assayed in vivo and in vitro in roots ofHordeum vulgare (barley). Relationship with diurnal changesin endogenous malate and sugar. Planta 148, 455-61.
Denna DW. 1962. A simple grafting technique for cucurbits.Proceedings of the American Society of Horticultural Science81, 369-70.
Grusak MA, Pezeshgi S. 1996. Shoot-to-root signal transmissionregulates root Fe(III) reductase activity in the dgl mutant ofpea. Plant Physiology 110, 329-34.
Hageman RH, Reed AJ. 1980. Nitrate reductase from higherplants. Methods in Enzymology 69, 270—80.
Heuvelink E. 1995. Dry matter partitioning in a tomato plant:one common assimilate pool? Journal of Experimental Botany46, 1025-33.
Hirose T. 1986. Nitrogen uptake and plant growth. II. Anempirical model of vegetative growth and partitioning. Annalsof Botany 58, 487-96.
van Kculen H, Wolf J. 1986. Modelling of agriculturalproduction: weather, soils and crop. Wageningen: Pudoc.
Kleinhofs A, Warner RL, Muehlbauer FJ, Nilan RA. 1978.Induction and selection of specific gene mutations in Hordeumand Pisum. Mutation Research 51, 29-35.
MacKinney G. 1941. Absorption of light by chlorophyllsolutions. Journal of Biological Chemistry 140, 315-22.
Marschner H, Kirkby EA, Cakmak I. 1996. Effect of mineralnutritional status on shoot-root partitioning of photoassim-ilates and cycling of mineral nutrients. Journal of ExperimentalBotany 47, 1255-63.
Natr L. 1972. Influence of mineral nutrients on photosynthesisof higher plants. Photosynthetica 6, 80-99.
Oscarson P, Larsson C-M. 1986. Relations between uptake andutilization of NO^~ in Pisum growing exponentially undernitrogen limitation. Physiologia Plantarum 67, 109-17.
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/48/6/1241/654720 by guest on 04 April 2019

1250 Lexa and Cheeseman
Peuke AD, Glaab J, Kaiser WM, Jeschke WD. 1996. Theuptake and flow of C, N and ions between roots and shootsin Ricinus communis L. IV. Flow and metabolism of inorganicnitrogen and malate depending on nitrogen nutrition and salttreatment. Journal of Experimental Botany 47, 377-85.
Runge M. 1983. Physiology and ecology of nitrogen nutrition.In: Lange OL, ed. Encyclopedia of Plant Physiology. Berlin:Springer Verlag, 163-200.
Smith JL, Rice EL. 1983. Differences in nitrate reductaseactivity between species of different stages in old fieldsuccession. Oecologia 57, 43^4-.
Stewart GR, Hegarty EE, Specht RL. 1988. Inorganic nitrogenassimilation in plants of Australian rainforest communities.Physiologia Plantarum 74, 26-33.
Stewart GR, Joly CA, Smirnoff N. 1992. Partitioning ofinorganic nitrogen assimilation between the roots and shootsof cerrado and forest trees of contrasting plant communitiesof South East Brasil. Oecologia 91, 511-17.
Stewart GR, Pate JS, Unkovich M. 1993. Characteristics ofinorganic nitrogen assimilation of plants in fire-proneMediterranean-type vegetation. Plant, Cell and Environment16, 351-63.
Thoraley JHM. 1972. A balanced quantitative model for
root:shoot ratios in vegetative plants. Annals of Botany36, 431^11.
Thornley JHM. 1976. Mathematical models in plant physiology:a quantitative approach to problems in plant and cropphysiology. London: Academic Press.
Topa MA, Cheeseman JM. 1992. Effects of root hypoxia and alow P supply on relative growth, carbon dioxide exchangerates and carbon partitioning in Pinus serotina seedlings.Physiologia Plantarum 86, 136-44.
Warner RL, Kleinhofs A, Muehlbauer FJ. 1982. Characterizationof nitrate reductase-deficient mutants in pea. Crop Science22, 389-93.
Went FW. 1938. Transplantation experiments with peas.American Journal of Botany 25, 44-55.
Went FW. 1943. Transplantation experiments with peas. III.Botanical Gazette 104, 460-74.
van der Werf A, Nagel OW. 1996. Carbon allocation toshoots and roots in relation to nitrogen supply is medi-ated by cytokinins and sucrose: opinion. Plant and Soil (inpress).
Whltmarsh J, Ort D. 1984. Stoichiometries of electron transportcomplexes in spinach chloroplasts. Archives of Biochemistryand Biophysics 231, 378-89.
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/48/6/1241/654720 by guest on 04 April 2019