grassland bird response to …digital.library.okstate.edu/etd/umi-okstate-1715.pdflogistic...
TRANSCRIPT

GRASSLAND BIRD RESPONSE TO
DISKING/INTERSEEDING OF LEGUMES IN
CONSERVATION RESERVE PROGRAM LANDS IN
NORTHEAST NEBRASKA
By
LUCAS PAUL NEGUS
Bachelor of Science
University of Nebraska at Kearney
Kearney, Nebraska
2002
Submitted to the Faculty of the Graduate College of the
Oklahoma State University in partial fulfillment of
the requirements for the Degree of
MASTER OF SCIENCE May, 2006

ii
GRASSLAND BIRD RESPONSE TO
DISKING/INTERSEEDING OF LEGUMES IN
CONSERVATION RESERVE PROGRAM LANDS IN
NORTHEAST NEBRASKA
Thesis Approved:
Craig Davis
Timothy O’Connell
Tim McCoy
A. Gordon Emslie Dean of the Graduate College

iii
ACKNOWLEDGEMENTS
I would like to thank the Nebraska Game and Parks Commission for funding this
project through a State Wildlife Grant. I would specifically like to thank Scott Wessel for
preparing the grant, as well as for providing guidance, advice, and all levels of support
for the project. I also thank Pheasants Forever for providing field equipment and funding
for the project. The Lower Elkhorn Natural Resources District also provided funding for
field technicians, for which I am very thankful. I am also thankful to the Oklahoma State
University Zoology Department for administering the funds for the project.
This project could not have been completed without the assistance and guidance
of many individuals. I sincerely thank my major advisor, Dr. Craig A. Davis, for
guidance, advice, and support through all aspects of this project. I truly appreciate the
assistance and advice my graduate committee members, Dr. Timothy O’Connell and Dr.
Tim McCoy, provided during the study. This project could not have been completed in
entirety without the help of my field technicians, Jamie Bachmann, Cassidy Goc, Jordan
Johnson, and Adam Schole. I am very thankful for their enthusiasm and tireless work
ethic, enduring early, wet mornings and hot afternoons with no complaints. I would also
like to thank the graduate students of the Zoology department for providing help and
support when needed, and especially for the friendship and many good memories. I
would particularly like to thank Sabrina Rust, who lovingly stuck by my side and
supported me in everything I did the last several years. I would like to thank my family
for all the support and visits, as well as my dog, Tyson, for putting up with city life for

iv
the last couple of years. Finally, I would like to thank my parents for not only letting me
stay at home once again during the summers, but also for instilling the love of the
outdoors in me at an early age and supporting me through everything I have done.

v
TABLE OF CONTENTS
Chapter Page ACKNOWLEDGEMENTS ...................................................................................... iii LIST OF TABLES .................................................................................................... vii LIST OF FIGURES ................................................................................................... ix
I. GRASSLAND BIRD RESPONSE TO DISKING/INTERSEEDING LEGUMES IN CONSERVATION RESERVE PROGRAM LANDS IN NORTHEAST NEBRASKA ...........................................................................................................1
Introduction..............................................................................................................1
Grassland Avifauna...............................................................................................1 Habitat Loss and Degredation..........................................................................1 Habitat Fragmentation .....................................................................................3 Conservation Reserve Program.............................................................................5 Justification ..............................................................................................................8 Objectives ..............................................................................................................12 Methods..................................................................................................................12 Study Area ..........................................................................................................12 Disking and Interseeding ....................................................................................13 Bird Abundance Surveys ....................................................................................15 Nest Searches and Monitoring ............................................................................15 Vegetation Sampling...........................................................................................16 Statistical Analysis..............................................................................................17 Results....................................................................................................................21 Grassland Bird Community ................................................................................21 Nesting Success ..................................................................................................23 Vegetation Characteristics ..................................................................................26 Vegetation Influences .........................................................................................27 Discussion ..............................................................................................................28 Conservation Implications and Management.........................................................41 Literature Cited ......................................................................................................46 APPENDICES ............................................................................................................85

vi
Appendix A............................................................................................................85 Appendix B ............................................................................................................87 Appendix C ............................................................................................................89

vii
LIST OF TABLES Table Page 1.1. Overall relative abundance, species richness, and species diversity of breeding
grassland birds in treatment and reference fields in Stanton County, Nebraska, 2004-2005. ....................................................................................................................61
1.2. Relative abundance (birds/transect) of breeding grassland birds that were observed
in >1% of surveys in treatment and reference fields in Stanton County, Nebraska, 2004-2005. ....................................................................................................................62
1.3. Relative abundance (birds/transect) of breeding grassland birds that were observed
in >1% of surveys in 3 portions of Conservation Reserve Program fields managed by disking/interseeding in Stanton County, Nebraska, 2004-2005...............................64
1.4. Number of nesting species, number of nests, and nest density (nests/ha) of grassland
birds in treatment and reference fields in Stanton County, Nebraska, 2004-2005 .......66 1.5. Nest success probabilities for incubation, nestling, and overall nesting period for all
bird species in treatment and reference fields in Stanton County, Nebraska, 2004-2005...............................................................................................................................67
1.6. Nest success probabilities for all bird species, dickcissels, and red-winged
blackbirds in Conservation Reserve Program fields in Stanton County, Nebraska, 2004 and 2005...............................................................................................................68
1.7. Vegetation characteristics of successful and unsuccessful nests of all grassland bird
species in Conservation Reserve Program fields in Stanton County, Nebraska, 2004-2005. ....................................................................................................................69
1.8. Vegetation characteristics of successful and unsuccessful dickcissel nests in
Conservation Reserve Program fields in Stanton County, Nebraska, 2004-2005 ........70 1.9. Vegetation characteristics of successful and unsuccessful red-winged blackbird
nests in Conservation Reserve Program fields in Stanton County, Nebraska, 2004-2005. .............................................................................................................................71
1.10. Vegetation characteristics of successful and unsuccessful grasshopper sparrow nests
in Conservation Reserve Program fields in Stanton County, Nebraska, 2004-2005....72

viii
1.11. Vegetation characteristics of successful and unsuccessful bobolink nests in
Conservation Reserve Program fields in Stanton County, Nebraska, 2004-2005 ........73 1.12. Vegetation characteristics of treatment and reference fields in Conservation Reserve
Program fields in Stanton County, Nebraska, 2004-2005 ............................................74 1.13. Vegetation characteristics of 3 portions of Conservation Reserve Program fields
managed by disking/interseeding in Stanton County, Nebraska, 2004-2005 ...............75 1.14. Logistic regression models for vegetation variables that best predicted grassland
bird presence in treatment sites in Stanton County, Nebraska, 2004-2005 ..................77 1.15. Logistic regression models for vegetation variables that best predicted grassland
bird presence in reference sites in Stanton County, Nebraska, 2004-2005 ..................78

ix
LIST OF FIGURES
Figure Page 1.1. Location of study area (represented by black box) in Stanton County, Nebraska........79 1.2. Distance frequency histograms for all species, dickcissels, grasshopper sparrows,
and bobolinks in treatment and reference fields in Stanton County, Nebraska, 2004 and 2005........................................................................................................................81
1.3. Mean (+ S.E.) nest density (nests/ha) for all species combined for 0- (n = 2), 1- (n =
4), and 2-year (n = 2) post treatment and no treatment (n = 11) nest search plots in Conservation Reserve Program fields in Stanton County, Nebraska, 2004 and 2005. Means with different letters are significantly different (P < 0.05) ...............................83

1
CHAPTER I: GRASSLAND BIRD RESPONSE TO DISKING/INTERSEEDING OF LEGUMES IN CONSERVATION RESERVE PROGRAM LANDS IN
NORTHEAST NEBRASKA
INTRODUCTION
Grassland Avifauna
Throughout the Midwest, grassland bird populations are declining faster than any
other group of birds (Samson and Knopf 1994). From 1969 to 1991, grassland bird
populations in Illinois, Minnesota, Wyoming, Nebraska and Missouri declined from 24 to
91% (Samson and Knopf 1994). These declines have been attributed mainly to the loss
of prairie habitat. Samson and Knopf (1994) estimated that as little as 1% of native
prairie habitat remains in the Midwest. Unfortunately, the prairie that remains often
exists in small fragments and receives insufficient management. Additionally, modern
agricultural practices that favor reduced crop diversity and increased field sizes may
contribute to declines in grassland bird numbers (Best et al. 1998).
Habitat loss and degradation—Habitat loss is the primary cause for declines in
grassland bird numbers. There were approximately 162 million ha of prairie in the Great
Plains prior to European settlement, with the Tallgrass Prairie Ecoregion constituting
approximately 60 million of these hectares (Samson and Knopf 1994, Steinauer and
Collins 1996). Today, most of the tallgrass prairie has been plowed and converted to
agricultural lands, with losses as high as 99.9% in some states (Steinauer and Collins
1996). Habitat loss is not the only factor contributing to the decline in grassland birds.

2
Improper or inadequate management of remaining grasslands may also play a role in
grassland bird declines. Fire historically played a major role in the maintenance of
prairies. Periodic fires (every 2-5 years) set by lightning or Native Americans restricted
woody vegetation from encroaching into the prairies and increased plant species diversity
(Steinauer and Collins 1996). Since European settlement, fires have been suppressed
allowing woody vegetation to encroach on and often dominate prairies. The importance
of fire is evident by the response of grassland birds to this disturbance factor. Dechant et
al. (2003) recommended burning every 2-4 years to improve habitat for grasshopper
sparrows (Ammodramus savannarum). Bobolinks (Dolichonyx oryzivorus) also respond
positively to properly timed burns (2-4 yrs) (Herkert 1994). Johnson and Temple (1990)
found lower rates of nest depredation on grasshopper sparrow nests in recently burned
areas in Minnesota.
Many grasslands are included in farming and ranching operations, often to the
detriment of the ecological attributes of those grasslands. Specifically, many agricultural
grasslands and forage crops are mowed or hayed annually, with entire fields being cut in
mid-summer during the peak of nesting season for many grassland birds. Annual
mowing during the breeding season results in high rates of nest failure for dickcissels
(Spiza americana; Frawley and Best 1991). In a study in New York, mowing during the
breeding season accounted for 51% of nest failures for bobolinks (Bollinger et al. 1990).
Annual mowing of entire fields provides habitat for only a select few grassland bird
species, limits changes in vegetation structure, and promotes grass succession (Horn and
Koford 2000, McCoy et al. 2001a). Improper grazing strategies also adversely affect
grassland birds (Zimmerman 1997). The primary impact of grazing is reduction or

3
elimination of above-ground vegetation. Season-long grazing or intensive grazing during
the nesting period reduces vegetation height and density at a critical period, resulting in
reduced avian abundance and productivity (Zimmerman 1997). For example, Swanson
(2003) reported that nesting success of savannah sparrows (Passerculus sandwichensis)
was significantly higher on ungrazed grasslands than on grasslands grazed continuously.
Temple et al. (1999) reported dickcissels were more abundant in ungrazed grasslands
than in continuously or rotationally grazed pastures in southwestern Wisconsin.
Habitat fragmentation—Habitat fragmentation may also be a contributing factor
to declining grassland bird numbers (Herkert 1994, Winter and Faaborg 1999). Johnson
and Igl (2001) defined habitat fragmentation in prairies as the division of large,
contiguous areas of prairie habitat into smaller patches isolated from one another. Three
effects of fragmentation are reduced patch-size, increased edge, and increased isolation
(Johnson and Igl 2001). Several studies have found decreasing patch size to have
negative effects on the presence and nest success of grassland birds (Herkert 1994, Helzer
and Jelinski 1999, Winter and Faaborg 1999, Johnson and Igl 2001). Johnson and Igl
(2001) found 6 of 15 grassland bird species consistently favored larger patches of habitat
over smaller pathces. Herkert et al. (2003) studied nest predation in relation to patch size
for 4 grassland bird species (dickcissel, grasshopper sparrow, Henslow’s sparrow
[Ammodramus henslowii], and eastern meadowlark [Sturnella magna]) and reported
predation rates of these species consistently declined with increasing patch size.
Moreover, predation rates were consistently lowest in prairies larger than 1,000 ha.
Increased edge caused by habitat fragmentation also increases brood parasitism by
brown-headed cowbirds (Molothrus ater). Brown-headed cowbirds, one of the most

4
common edge species in the Great Plains, seem to favor small grassland patches (Johnson
and Igl 2001). Nest productivity is reduced by cowbird parasitism because fewer host
young fledge from parasitized nests than non-parasitized nests (Johnson and Temple
1986). Johnson and Temple (1990) found brood parasitism increased with decreasing
distance to edge. It is speculated that brown-headed cowbirds are more effective near
edges because they use high perches in trees to locate the host’s nests (Johnson and
Temple 1990). Davis and Sealy (2000) observed that cowbird parasitism was highest in
small, irregularly shaped plots with high amounts of edge. Two proposed management
strategies that could reduce cowbird parasitism in grasslands are providing and creating
large tracts of grassland habitat and removing woody edges and woody vegetation from
grasslands (Johnson and Temple 1990, Davis and Sealy 2000, Koford et al. 2000).
The effects of habitat isolation are difficult to quantify. In the field of landscape
ecology, biologists attempt to label isolated habitats as source or sink habitats for animal
populations. A source population is a population in which fecundity is greater than that
required to maintain a stable population. Conversely, a sink population occurs if
fecundity is below the level that is required to maintain a stable population (Pulliam
1988). Although some grasslands have been identified as source or sink habitats for
grassland birds, critical size or specific characteristics of the grassland habitat that
influence source/sink status are not easily identified. Management recommendations
generally encourage large, diverse grasslands to benefit grassland birds, but exact sizes
and habitat conditions required by many species still remain unclear. Johnson and Igl
(2001) suggest that creating grassland habitat near existing grasslands, or establishing 1
large field rather than several small ones would benefit more grassland bird species than

5
creating small, isolated fields. Johnson and Igl (2001) also noted regional differences
may play a role in the development of management strategies (i.e., habitat requirements
in 1 region may not be applicable in another region). Identifying the regional differences
in habitat requirements and developing management strategies is an important component
in the conservation of grassland birds.
Conservation Reserve Program
With the tremendous losses of native prairie throughout the Midwest, surrogate
grasslands such as Conservation Reserve Program (CRP) fields have become increasingly
important to grassland wildlife. The CRP was established as a provision of the Food
Security Act of 1985 (1985 Farm Bill) and has been retained in both the 1996 and 2002
Farm Bills (U.S. Department of Agriculture 2003a). The original goals of CRP were to
reduce erosion and improve water quality of highly erodible cropland, with a co-equal
objective of creating and enhancing wildlife and fish habitat added in 1996 (Johnson and
Schwartz 1993a). CRP pays farmers annual rental payments to retire highly erodible
cropland from production and plant it to grasses, trees, or other perennial cover for 10-15
years (U.S. Department of Agriculture 2003a). CRP differs from previous farm set-aside
programs (e.g., the Payment in Kind Program) because it is a long-term program that
retires cropland for greater than 10 years by planting the cropland to permanent
vegetation providing substantial wildlife benefits (Hays et al. 1989).
In the Midwest, CRP lands have been found to be important to a variety of
grassland wildlife species. Game species such as ring-necked pheasants (Phasianus
colchicus; King and Savidge 1995), northern bobwhite (Colinus virginianus; King and
Savidge 1995), white-tailed deer (Odocoileus virginianus; Luttschwager and Higgins

6
1992) and ducks (Reynolds et al. 1994) all use CRP fields. A study by King and Savidge
(1995) revealed pheasant abundance was higher in areas with a high percentage of CRP
than areas with a low percentage of CRP, probably due to the increased nesting and
brood-rearing habitat. Reynolds et al. (1994) found duck nest success in CRP fields to be
as high or higher than that of cover planted specifically for duck nesting. Nest success
for ducks in CRP fields was 2-9% greater than nest success rates needed to maintain
stable populations.
Non-game grassland songbirds also benefit from CRP (Johnson and Schwartz
1993b, Patterson and Best 1996). Several studies have attributed increases or at least
stable trends in specific grassland bird species to CRP (Igl and Johnson 1995, Herkert
1998, Ryan et al. 1998). Herkert (1998) reported grasshopper sparrow population trends
that were negative in 13 mid-continental states prior to CRP were positive following CRP
enrollment in those states. Igl and Johnson (1995) attributed a rapid increase in Le
Conte’s sparrow (Ammodramus leconteii) in North Dakota to the increased nesting
habitat offered by CRP when favorable wet weather conditions occurred. In an analysis
of North American Breeding Bird Survey results before and after CRP, Reynolds et al.
(1994) concluded that 4 of 8 declining grassland bird species showed positive population
trends, with only 1 species continuing to decline. Not surprisingly, Ryan et al. (1998)
found relative abundance of birds in CRP fields was as much as 10 times higher than in
crop fields. In a similar study, Best et al. (1997) also found the abundance of birds to be
as much as 10.5 times higher in CRP than crop fields. Moreover, CRP fields supported 3
times more nesting species and 13.5 times the total number of nests than row crop fields.

7
Although CRP fields provide habitat for grassland birds, the size and connectivity
of fields influence their suitability for grassland birds. McCoy et al. (1999) determined
whether CRP lands in Missouri were source or sink habitats for grassland bird species.
They found that source-sink status differed among grassland bird species using CRP
fields. Among their results, they found that CRP fields were source habitats for
grasshopper sparrows, field sparrows (Spizella pusilla), eastern meadowlarks, and
American goldfinches (Carduelis tristis) and sink habitats for dickcissels and red-winged
blackbirds (Agelaius phoeniceus). McCoy et al. (1999) concluded that CRP created
“source” habitats for many species, especially where alternative-breeding habitat was in
poor condition or absent from the area. Other studies, however, have documented
declines in grassland bird populations associated with CRP, indicating these lands may
act as a sink habitat (Reynolds et al. 1994). Source/sink dynamics of CRP may be
important to the conservation of grassland birds and need to be investigated more
thoroughly.
Although CRP has benefited numerous wildlife species, the types of plantings
used in CRP fields can have a major effect on grassland bird species. Conservation
Reserve Program fields are planted to a variety of cover types, referred to as conservation
practices (CPs). Two of the most commonly implemented CPs are CP1, a cool-season
grass mix, and CP2, a warm-season grass mix. CP1’s are frequently dominated by
smooth brome (Bromus inermis), orchard grass (Dactylis glomerata), or timothy (Phleum
pratense), while CP2s are frequently dominated by switchgrass (Panicum virgatum). The
vegetation in CP1s is typically short and dense compared to the tall CP2 vegetation.
Recently, natural resource agencies have promoted planting CP2s instead of CP1s to

8
benefit wildlife. However, several studies in the Midwest have found no difference in
total abundance or species richness of grassland birds between CP1 and CP2 plantings
(Johnson and Schwartz 1993b, King and Savidge 1995, Delisle and Savidge 1997,
McCoy et al. 2001b). Although avian richness and abundance may not differ between
CP1s and CP2s, some grassland bird species prefer certain habitat types that may not be
provided separately by each CP. For example, dickcissels prefer CRP fields with tall
grasses and high forb content (Patterson and Best 1996, Hughes et al. 1999), while
grasshopper sparrows prefer habitats of moderate height and density (McCoy et al.
2001a). Ideally, CRP fields should have separate portions planted to CP1 and CP2 to
benefit the full array of grassland bird species.
JUSTIFICATION
Similar to other states, CRP in Nebraska has become increasingly important to
grassland wildlife. The 472,000 ha of CRP in Nebraska is nearly four-times the 123,000
ha of remaining native tallgrass prairie in the state (Steinauer and Collins 1996, U.S.
Department of Agriculture 2003b). This CRP acreage provides additional grassland
habitat and complements the remaining native grasslands for grassland-dependent
species. However, nearly 90% of this CRP acreage in Nebraska is >5 years old (Scott
Wessel, Nebraska Game and Parks Commission, personal communication). As a result,
these older CRP fields have become senescent, providing limited habitat to wildlife.
In eastern Nebraska, typical CP1 plantings in the initial enrollment consisted of a
single cool season grass species, smooth brome, combined with several legume species
(Trifolium spp.) (Clayton Stalling, Nebraska Game and Parks Commission, personal
communication). In the early years of the program, these plantings provided a diverse

9
habitat, consisting of a mixture of grasses, and various forbs and legumes that provided
excellent habitat for many wildlife species. These diverse and heterogeneous grasslands
are desired because they provide multiple niches for a myriad of grassland species.
However, due to succession and the inhibiting nature of smooth brome, these once
productive grassland fields have become a monoculture of smooth brome that has a
limited value to wildlife (Millenbah et al. 1996, McCoy et al. 2001b). Specifically, these
monocultures of smooth brome are characterized by short and dense vegetation with high
amounts of litter accumulation that provide habitat for a limited number of grassland
species.
There have been many studies focused on the effects of mowing, grazing, fire, or
a combination of these management practices on grassland birds in CRP fields
(Zimmerman 1992, Knopf and Samson 1997, Swanson et al. 1999, Horn and Koford
2000, Madden et al. 2000, McCoy et al. 2001a). These management practices can be
effective in enhancing CRP, but are frequently ineffective due to logistical and/or social
constraints. Mowing and haying are often avoided because of the steep terrain and
presence of pocket gophers (Geomys bursarius) in the CRP fields. Pocket gophers make
large mounds of soil, which often damage haying equipment to a point that it may be too
costly to hay or mow (Dale Clark, CRP landowner, personal communication). Grazing is
often avoided because fences must be constructed, water sources provided, and cattle
must be monitored daily. Prescribed burning is rarely used for several reasons. First,
most landowners are not properly trained to conduct prescribed burns, and local fire
marshals are skeptical about providing burn permits to unqualified people. There are also
few burn crews that can provide professional burning services, and those that do are

10
expensive to hire. Finally, fire is still viewed by many landowners as being destructive
and dangerous, even on grasslands.
Because these management practices are rarely used, alternative management
practices have been explored. Plant diversity has been shown to increase through
mechanical means such as disking and interseeding legumes. Shallow disking can be an
effective tool for enhancing CRP fields, acting to set back succession in the fields.
Disking and interseeding is favored by wildlife managers because of the diverse habitat
created and is favored by landowners/farmers because of the availability of equipment
and convenience of the practice. However, disking and interseeding is an ephemeral
practice and provides diverse habitat for only 4-5 years before grass becomes the
dominant vegetation again (Scott Wessel, Nebraska Game and Parks Commission,
personal communication).
In May 2002, the Nebraska Game and Parks Commission initiated a program to
curb declining ring-necked pheasant populations in the state. The program, entitled
“Focus on Pheasants,” placed an emphasis on creating nesting and brood-rearing habitat
in aging CRP fields (Taylor 2002). The best habitats for nesting and brood rearing
include a weedy, diverse grassland. This can most easily be accomplished in CRP fields
by lightly disking the soil to break up the existing mature grass stand which promotes
growth of annual broad-leafed forbs (weeds), increases bare ground, and sets back
succession (Manley et al. 1994). As an added benefit, alfalfa and sweetclover (legumes)
are also interseeded after disking to increase forb abundance and encourage an influx of
insects (Whitmore et al. 1986). Manley et al. (1994) found arthropod biomass to be
significantly higher in disked portions of a field compared to undisked portions. A study

11
conducted by Leathers (2003) evaluated invertebrate abundance on CRP fields in eastern
Nebraska that had been disked and interseeded. Although his results were highly
variable, Leathers (2003) found invertebrate and forb abundance were often higher in
disked and interseeded portions of the fields than untreated CRP fields. The influx in
insects is important because they provide critical food resources for both game bird
chicks and grassland birds (Whitmore et al. 1986, Jackson et al. 1987, Kobal et al. 1998,
McIntyre and Thompson 2003).
Mid-term management, a new rule for CRP sign-up 26 and subsequent sign-ups,
requires management on CRP fields to enhance habitat diversity (U. S. Department of
Agriculture 2003a). Mowing, grazing, burning, and disking/interseeding are all approved
practices for this rule. According to the rule, management must be conducted at least
once on the entire CRP field during the life of the contract, and management can only be
conducted on a maximum of one-third of the field per year. A management regime that
includes disking one-third of a field in years 4, 5, and 6 of a contract could create
desirable field conditions for wildlife by having part of the field in different successional
stages. Millenbah (1993) found avian diversity and density was highest in 1-3 year old
CRP fields in Michigan, which was attributed to the diverse vegetation in the younger
fields.
Mid-term management is also cost-shared under U. S. Department of Agriculture
(USDA) rules. Up to 50% of the incurred cost for the performed management practice
will be reimbursed to the landowner. Cost-share payments combined with the
convenience of disking makes this management attractive to farmers. Resource
professionals in the area expect that mandated management on CRP along with the

12
efforts of conservation agencies and organizations will significantly increase the practice
of disking and interseeding not only in Nebraska, but also throughout the Midwest.
Currently, little knowledge exists on the effects of disking and interseeding on
grassland bird abundance and nest productivity. With CRP fields increasingly providing
key grassland habitat for wildlife throughout the Midwest, disking and interseeding has
the potential to improve the quality of existing CRP fields. While I studied the effects of
disking/interseeding legumes on a fine geographic scale, the response of the grassland
bird community may be extrapolated to a much broader scale to predict the effects of this
management practice throughout the Midwest. This study presents an opportunity to
evaluate grassland bird population response to different levels of succession in grassland
habitats, as well as evaluate the effects of a new management practice on grassland birds.
Moreover, information from this study will be important in guiding future decisions
concerning management of CRP fields and ultimately may influence future USDA
policies for the Conservation Reserve Program.
OBJECTIVES
1) Compare avian richness and abundance in disked/interseeded CRP fields to
unmanaged CRP fields.
2) Compare avian nest productivity in disked/interseeded CRP fields to unmanaged
CRP fields.
3) Evaluate differences in vegetation structure, composition, and cover between
disked/interseeded and unmanaged CRP fields.
METHODS
Study Area

13
This study was conducted in Stanton County, Nebraska during the summers of
2004 and 2005 (Figure 1). Stanton County lies in the Loess Uplands and Till Plains
Major Land Resource Area in northeast Nebraska. Precipitation in the area averages 50
to 65 cm per year, with most occurring during the growing season (Natural Resource
Conservation Service 2003). Soils in the area are of the Crofton-Nora and Nora-Crofton
complexes that include silty, loamy, and sandy textural classes (Hammond 1982).
Stanton County is mainly an agricultural county with corn and soybeans being the
predominant crops. Other common crops include oats and alfalfa (Hammond 1982).
Nearly 11,200 ha of CRP occur in Stanton County (U.S. Department of
Agriculture 2003b). This study was conducted on privately-owned CRP within a 51.5
km2 study area. The study area location was selected because of the large percentage of
older CRP fields in the area, the potential to improve habitat quality from management
practices, and landowner cooperation (Taylor 2002). Sixteen CRP fields ranging in size
from 16 to 64 ha were selected for the study. Eight of the fields were manipulated
(treatment fields), and 8 fields were unmanipulated and used as reference fields. A
portion (25-33%) of each manipulated field was disked in 2003 and 2004, resulting in 50-
66% of each field occurring in different successional stages. Conservation Reserve
Program fields in the study area were originally planted to smooth brome, alfalfa, and
yellow sweetclover, but are now dominated by smooth brome. No planned management
(disking, haying, or grazing) has occurred on the reference fields. Selection of study
fields was based on previous management history, field size, proximity to other fields,
and landowner cooperation.
Disking and Interseeding

14
Disking and interseeding of the treatment fields began in September 2002.
Disking was performed by a contractor using a tandem disc designed specifically for sod
breakup. Portions (up to one-third) of each field were selected and flagged prior to
disking. Selection of portions to be disked was based on topography and landowner
preference. Wherever possible, portions that had been hayed in August 2002 were
selected because the reduction of residual cover made disking more effective. Disking
was maintained at a depth of 7.6 to 10.2 cm. Additionally, each portion was disked twice
to effectively break-up the sod. After disking, a no-till drill was used to interseed
legumes. The legume mix, provided by Pheasants Forever, consisted of alfalfa, red
clover, and yellow sweetclover. The legumes were seeded at a rate of 6.75 kg/ha (1.69
kg of red clover, 1.69 kg of yellow sweetclover, and 3.38 kg of alfalfa). According to
USDA guidelines, interseeding was terminated on 15 September 2002 to avoid winter kill
of late germinating plants (Natural Resource Conservation Service 2002). Seeding was
continued after 1 November, when consistent freezes occurred at night. Consistent
freezes ensured a low soil temperature was maintained even during times with warm
daytime temperatures, preventing seed germination and winter-kill. Disking resumed in
April 2003 and continued until 1 May 2003. No disking or interseeding was conducted
between 1 May and 1 August 2003. Disking on another portion of each field resumed in
fall 2003 and was completed in the spring of 2004. All disking depths and seeding rates
were performed in accordance with USDA guidelines (Natural Resource Conservation
Service 2002).
Treatment study fields consisted of CRP fields that had portions disked and
interseeded over the span of 2 years. A typical treatment study field had 25-33% disked

15
in 2003 and 25-33% disked in 2004, with the remainder of the field undisked. The
reference study fields did not have any disking performed on any portion of the field.
Bird Abundance Surveys
I surveyed birds using the belt transect method similar to Best et al. (1997) and
McCoy et al. (2001b). Transects were 200-m long with a fixed 100-m width (Helzer and
Jelinski 1999). Transects were not located <50 m from any field edge or boundary
between disked portions of the field. By maintaining transects >50 m from the edge of
any field (including roads, tree lines, and agricultural fields), I minimized edge effects on
survey results (Helzer 1996). In the treatment fields, I established 3 transects with 1
transect located in each portion of the field (i.e., 2 transects in disked/interseeded portions
and 1 transect in the undisked portion). In the reference fields, I randomly established 2
transects in each field in 2004. In 2005, an additional transect was established in each
reference field to be more consistent with surveys in treatment fields.
I surveyed each transect for avian abundance once during 3 periods (25 May-14
June, 15 June-30 June, and 1 July-15 July) (Best et al. 1997, McCoy et al. 2001b). Avian
abundance was determined by counting birds seen and/or heard while walking along each
transect. All birds encountered within the belt transect were counted. Birds flying over
the transect were noted, but were not included in abundance data for the fields. In 2005, I
recorded the perpendicular distance from the transect to each bird to determine detection
probability for each species encountered. Surveys were conducted between sunrise and 3
hours after sunrise on days when the wind was <16 km/hr with no fog or rain.
Nest Searches and Monitoring

16
In 2 treatment fields, I established a 4-ha nest search plot centered around each
avian survey transect, resulting in 3 nest search plots per field. In 2004, I also established
2, 4-ha nest search plots in reference fields. In 2005, the number of search plots in
reference fields was increased to 6 to be more comparable to treatment fields. Nest
searches were conducted using 4 methods: 1) observing birds engaged in breeding
behavior indicating nest building, incubation, or feeding of nestlings (Martin and Geupel
1993, Giuliano and Daves 2002), 2) chain or rope dragging, 3) systematic walking with
or without a sweeping stick (Winter et al. 2003), and 4) random searches through field
plots (Giuliano and Daves 2002). Plots were searched periodically (>2 times per
sampling period) throughout the summer on a rotational basis to minimize disturbance.
Additional nests were located while conducting avian abundance surveys or sampling
vegetation.
Once a nest was located, it was marked with flagging 5 m to the north and 5 m to
the south of the nest (Giuliano and Daves 2002), and a description of the nest location
was recorded on a nest data sheet. Additionally, Universal Transverse Mercator (UTM)
coordinates for each nest were obtained using a Global Positioning System (GPS) unit.
Nests were checked every 2-5 days until a final outcome was determined (Martin and
Geupel 1993). Outcome was recorded as 1) successful (at least 1 young fledged), 2)
unsuccessful (no young fledged due to depredation, weather, nest parasitism, or unknown
causes), or 3) undetermined.
Vegetation Sampling
Vegetation sampling occurred along the avian abundance survey transects and at
each nest location. I located 4 sampling points (spaced evenly at 40 m, 80 m, 120 m, and

17
160 m) along each transect to record vegetation data. Vegetation data was collected
along survey transects immediately following avian abundance surveys during each
period and at nests within 10 days of nest completion.
At each sampling point on the transect and at each nest, I recorded the following
vegetative characteristics: percent cover, maximum vegetation height, litter depth, and
horizontal visual obstruction. Percent vegetation cover was estimated using a 20 x 50 cm
Daubenmire (1959) frame. Each frame was centered around each sampling point and
percent grass, forb, dead material, and bare ground were estimated (Hughes et al. 1999).
Planted legumes were included with all other forbs in the percent forb cover estimate.
Horizontal visual obstruction was measured using a Robel pole centered on each
sampling point and at each nest (Robel et al. 1970). Horizontal visual obstruction
readings were taken from a height of 1 m and a distance of 4 m from the 4 cardinal
directions. Litter depth and maximum vegetative height within the Daubenmire frame
were measured at each corner of the frame using a meter stick (Hughes et al. 1999).
Additionally, the plant species each nest occurred in and the height of each nest was
recorded.
Statistical Analysis
Prior to conducting statistical analysis, I determined total bird and individual
species relative abundances for treatment and reference fields by averaging count data
from each survey period for each year. Bobolink relative abundance was calculated from
the first and second survey periods because bobolinks were in large groups preparing for
migration during the third survey period. I only included bird species that occurred in
>1% of all surveys for the calculation of individual species abundances. Shannon-

18
Weiner species diversity index was used to calculate overall bird diversity for treatment
and reference fields for each year (Krebs 1999). To examine similarities in grassland
bird communities between treatment and reference fields and among portions of
treatment fields, I used Morisita’s index of similarity (Brower and Zar 1977): IM = 2Σxiyi
/ (λ1 + λ2) N1N2, where xi is the number of individuals in species i for community 1, yi is
the abundance of species i in community 2, λ1 is Simpson’s dominance index for
community 1, λ2 is Simpson’s dominance index for community 2, N1 is the total number
of individuals in community 1, and N2 is the total number of individuals in community 2.
The range of IM is from 0 (no similarity) to 1 (complete similarity). Avian species
richness was also calculated for each treatment and reference field for both years and
included all species observed at any time in study fields during each breeding season.
I used Levene’s test to test for homogeneity of variance for the avian abundance
variables (Zar 1999). Most data sets did not meet assumptions of homogeneity, so I used
a square-root transformation to correct for heteroscedacity (Zar 1999). Because only
total bird abundance met assumptions after transformation, I rank-transformed individual
species data sets that did not meet the assumptions of homogeneity (Conover and Iman
1981). I used a 2-way analysis of variance (ANOVA) to examine differences in total
bird abundance, individual species abundance, diversity, and richness between treatment
and reference fields and years (2004 and 2005) (SAS Institute 2003). To further evaluate
the effects of disking/interseeding on avian species, I also used a 2-way ANOVA to
examine differences in the total bird abundance and individual species abundance of 5
species (dickcissel, red-winged blackbird, bobolink, grasshopper sparrow, and common
yellowthroat) among treatments within each manipulated field (disked-2003, disked-

19
2004, and undisked) and year. I used Duncan’s Multiple Range Test to examine
differences between the 3 portions of treatment fields (SAS Institute 2003).
To assess differences in detectability of individual species between treatment and
reference fields, I constructed frequency histograms with 10-m increments using the
perpendicular distances for each bird observation that was recorded in 2005 (Rotella et al.
1999, McCoy et al. 2001b). Frequency histograms were constructed for all species
combined, as well as for dickcissels, bobolinks, and grasshopper sparrows individually
because they had >20 observations for both treatment and reference fields. Frequencies
were computed by calculating the proportion of observations in each distance increment
for treatment and reference fields. In general, comparisons of these histograms indicated
detectability for all species combined and individual species were similar between
treatment and reference fields (Figure 2).
Nearly all the vegetation data failed to meet assumptions of homogeneity.
Percent dead material was the only vegetation variable that was homogenous. All other
variables (maximum vegetation height; horizontal visual obstruction; litter depth; and
percent forb, grass, and bare ground cover) were square-root transformed (Zar 1999), but
still did not meet assumptions of homogeneity. Therefore, I rank-transformed those
variables (Conover and Iman 1981). I used a 2-way ANOVA to examine differences in
vegetation variables between treatment and reference fields and years. To further
evaluate effects of disking/interseeding on vegetation characteristics, I used a 2-way
ANOVA to examine differences in vegetation variables among different treatments
within each manipulated field and year. I used Duncan’s Multiple Range Test to examine
differences among the 3 portions of treatment fields (SAS Institute 2003).

20
To determine the effect of vegetation characteristics on nest success, I used a 2-
way ANOVA to examine differences in vegetation variables for years and nest fate
(successful and unsuccessful) of all bird species combined, as well as individual
dickcissel, red-winged blackbird, bobolink, and grasshopper sparrow nests. Nest success
for all bird species combined, dickcissels, and red-winged blackbirds was determined
using the Mayfield method (Mayfield 1975, Johnson 1979). Nest success probability
between reference and treatment fields was examined using only all species combined for
2004 and 2005 because <20 nests were found in reference fields in 2004. Dickcissel and
red-winged blackbird nest success probability could not be compared between treatment
and reference fields because <20 nests for each species were found in reference fields
during both years. I used a Chi-square contingency table to examine differences in nest
success, number of nests, and number of species nesting between treatment and reference
fields (Dow 1978, SAS Institute 2003). I used a 2-way ANOVA to test for differences in
nest density between treatment and reference fields and years (SAS Institute 2003).
Additionally, I used a 1-way ANOVA to test for differences in nest density among plots
that were in different post-treatment stages (1- and 2- years post-treatment, current-year
treatment, and no treatment). For all analyses performed, I inferred a significance level
at P < 0.05.
I used logistic regression to develop explanatory models for the occurrence of
individual grassland bird species occurring in >1% of surveys in treatment and reference
fields based on vegetation characteristics (i.e., forb, grass, dead material, and bare ground
cover; horizontal visual obstruction; litter depth; and maximum vegetation height). I
selected logistic regression over linear regression because the individual species

21
abundance data were heavily weighted with zeros and violated assumptions of linear
regression (Zar 1999, Madden et al. 2000). I used the logistic model: P(presence) = 1/ (1
+ exp{ - [bo + b1(x)]}) where P(presence) was the probability that a bird species was
present, bo and b1 were intercept and slope coefficients, and x was the predictor variable
(vegetation variable). I used a backward-elimination routine to create the best model for
each bird species using all vegetation variables. A variable was eliminated from the
model if its observed significance level for the regression coefficient (based on Wald chi-
square) was P > 0.05. I used the Hosmer and Lemeshow (1989) test to assess the
goodness-of-fit of each model.
RESULTS
Grassland Bird Community
Over the 2 years of the study, I observed 28 bird species in treatment fields and 25
species in reference fields (Appendix A). There was high overlap in species assemblages
(IM = 0.72) between treatment and reference fields, yet several species were unique to
treatment and reference fields. Orchard orioles (Icterus spurious), American robins
(Turdus migratorius), cedar waxwings (Bombycilla cedrorum), house sparrows (Passer
domesticus), and killdeer (Charadrius vociferous) were only observed in treatment fields,
while eastern meadowlarks and Baltimore orioles (Icterus galbula) were only observed in
reference fields. Mean abundance of all grassland birds and species diversity were higher
in treatment than reference fields (Table 1). There was a treatment x year interaction for
species richness (F1,28 = 4.47, P = 0.044). In 2004, species richness was higher in
treatment fields, but in 2005, there was no significant difference for species richness
between treatment and reference fields.

22
During the study, 2 species (common yellowthroats [Geothlypis trichas] and red-
winged blackbirds) were significantly more abundant in treatment fields, 2 species
(bobolinks and Henslow’s sparrows) were significantly more abundant in reference
fields, and 1 species (sedge wren [Cistothorus platensis]) had similar relative abundances
in treatment and reference fields (Table 2). Three species (dickcissels [F1,260 = 5.13, P =
0.024], grasshopper sparrows [F1,260 = 8.55, P = 0.004], and western meadowlarks
[Sturnella neglecta] [F1, 260 = 5.29, P = 0.022]), showed significant treatment x year
interactions. Dickcissels were more abundant in treatment fields for each year.
Grasshopper sparrow abundance did not significantly differ between treatment and
reference fields in 2004, but they were more abundant in reference than treatment fields
in 2005. Western meadowlark abundance was significantly higher in treatment fields in
2004, but was not significantly different between treatment and reference fields in 2005.
Dickcissels, grasshopper sparrows, and bobolinks were the most abundant species for
both years in treatment and reference fields, accounting for 78% of the total bird
abundance.
High overlap occurred in grassland bird communities among portions of treatment
fields. Species assemblages between disked-2003 and disked-2004 portions (IM = 0.90)
and between disked-2004 and undisked portions (IM = 0.84) exhibited high overlap. The
least overlap occurred between disked-2003 and undisked portions (IM = 0.66) of
treatment fields. Though Morista’s index of similarity indicated grassland bird
communities in different portions of treatment fields were similar, there were several
differences in bird abundances within treatment fields (Table 3). Bobolinks were
significantly more abundant in the undisked portion than the disked-2003 and disked-

23
2004 portions (P < 0.001). Treatment x year interaction effects were observed for all
other species (all species combined [F2, 138 = 16.76, P < 0.001], dickcissel [F2,138 = 47.98,
P < 0.001], red-winged blackbird [F2,138 = 10.41, P < 0.001], grasshopper sparrow [F2,138
= 5.77, P = 0.004], and common yellowthroat [F2,138 = 10.12, P < 0.001]) (Table 3).
Therefore, subsequent analyses for those species were performed within years. In 2004,
all species combined, dickcissels, red-winged blackbirds, and common yellowthroats
were significantly more abundant in the disked-2003 portion (P < 0.001) than the disked-
2004 or undisked portions of the fields. Grasshopper sparrows were significantly more
abundant in the disked-2004 portion (P = 0.002) of the fields than the disked-2003 and
undisked portions of the fields in 2004. In 2005, all species combined, dickcissels, and
common yellowthroats were significantly more abundant in the disked-2004 portion (P <
0.001) than the disked-2003 or undisked portions. Red-winged blackbirds did not
significantly differ among treatment types (P = 0.522) in 2005. In 2005, grasshopper
sparrows were significantly more abundant in the undisked portion than the disked-2003
portion (P = 0.008), but abundance in the disked-2004 portion did not differ significantly
from the other 2 portions (Table 3). Dickcissels dominated the observations in the
disked-2003 and disked-2004 portions of treatment fields, accounting for 56% and 43%
of bird observations, respectively. In the undisked portions of treatment fields,
grasshopper sparrows were observed most often (34%), followed closely by dickcissels
(27%) and bobolinks (26%).
Nesting Success
I located a total of 247 nests (112 nests in 2004, 135 nests in 2005) during the 2
years of the study. Of those 247 nests, 206 nests of 8 species occurred in treatment fields,

24
and 41 nests of 10 species occurred in reference fields (Table 4). The majority of the
nests located were dickcissel (89) and red-winged blackbird (107) nests. Nest density did
not differ between treatment and reference fields (P = 0.3) or years (P = 0.850) (Table 4).
When further evaluated by years post-treatment instead of by treatment or reference field,
nest density was significantly higher in the 1-year post-treatment plots than the current-
year post-treatment and no treatment plots (P = 0.008). Nest density in the 2-year post-
treatment plots did not differ from any other plots (Figure 3).
Overall nest success probability for all species was similar between treatment and
reference fields (Table 5). Mayfield nest success probability for all bird species was
higher in 2004 than 2005 (Table 6). Dickcissels had nearly twice the nest success
probability in 2004 than 2005, and red-winged blackbirds were nearly 3 times more
successful in 2004 than 2005 (Table 6). Apparent nest success for all birds was 31% in
2004 and 19% in 2005. For dickcissels and red-winged blackbirds, apparent nest success
was 33% and 29% in 2004, and 16% and 9% in 2005, respectively. During both years,
nest failures for all birds were attributed to several causes: depredation (64% - 2004,
75% - 2005), abandonment (8% - 2004, 13% - 2005), weather (7% - 2004, 9% - 2005),
and unknown causes (21% - 2004, 3% - 2005). Dickcissel nest failures were attributed to
depredation (52% - 2004, 73% - 2005), abandonment (7% - 2004, 19% - 2005), weather
(15% - 2004, 3% - 2005), and unknown causes (26% - 2004, 5% - 2005). Red-winged
blackbird nest failures were attributed to depredation (71% - 2004, 75% - 2005),
abandonment (9% - 2004, 12% - 2005), weather (3% - 2004, 2% - 2005), and unknown
causes (17% - 2004, 12% - 2005).

25
I observed many differences in vegetation characteristics between successful and
unsuccessful nests for all species combined (Table 7). Litter depth and percent grass
cover were significantly higher at successful nests. Unsuccessful nests had a
significantly higher percentage of forb cover than successful nests. Litter depth, percent
grass cover, and percent dead material cover were significantly higher at all nests in 2005
than 2004. The percentage of forb cover and bare ground was higher at all nests in 2004
than 2005 (Table 7). A fate x year interaction was observed in nest height (F1,234 = 5.55,
P = 0.019), horizontal visual obstruction (F1,236 = 5.48, P = 0.020), and maximum
vegetation height (F1,236 = 8.19, P = 0.005). Subsequent analyses for these characteristics
were performed within years. In 2004, there was no difference in nest height, horizontal
visual obstruction, or maximum vegetation height between successful and unsuccessful
nests. In 2005, nest height, horizontal visual obstruction, and maximum vegetation
height were significantly higher at unsuccessful nests than successful nests.
There were few differences in vegetation characteristics between successful and
unsuccessful dickcissel nests (Table 8). Fate x year interactions were observed for
maximum vegetation height (F1,78 = 6.26, P = 0.014) and percent dead material (F1,78 =
4.09, P = 0.047). Further analyses were performed within years for these characteristics.
In 2004, neither maximum vegetation height nor percent dead material differed between
successful and unsuccessful dickcissel nests. In 2005, maximum vegetation height was
higher at unsuccessful than successful dickcissel nests. Nest height, horizontal visual
obstruction, percent forb cover, and percent bare ground were significantly higher at
dickcissel nests in 2004 than 2005. Litter depth and percent grass cover were
significantly higher at dickcissel nests in 2005 than 2004. Nest height did not differ

26
between successful and unsuccessful dickcissel nests, but dickcissel nests were higher in
2004 than 2005 (Table 8). Red-winged blackbird nests showed several differences
between successful and unsuccessful nests (Table 9). No fate x year interactions were
observed. Litter depth and percent grass cover were higher at successful than
unsuccessful red-winged blackbird nests (Table 9). Percent forb cover was higher at
unsuccessful than successful red-winged blackbird nests. Horizontal visual obstruction,
maximum vegetation height, percent forb cover, and percent bare ground were higher at
all nests in 2004 than 2005. Litter depth and percent grass cover were higher at red-
winged blackbird nests in 2005 than 2004. Grasshopper sparrow nests exhibited 1
difference between successful and unsuccessful nests; percent bare ground was higher at
successful than unsuccessful grasshopper sparrow nests (Table 10). There were no
significant differences between successful and unsuccessful bobolink nests (Table 11).
Vegetation Characteristics
Many vegetation characteristics differed between treatment and reference fields
(Table 12). Forb cover was significantly higher in treatment fields, while grass cover was
significantly higher in reference fields. Grass cover was higher in all fields (treatment
and reference) in 2005 than 2004. Treatment x year interactions occurred in horizontal
visual obstruction (F1,1040 = 8.65, P < 0.001), litter depth (F1,1040 = 30.17, P < 0.001),
maximum vegetation height (F1,1040 = 61.70, P < 0.001), percent dead material (F1,1040 =
25.20, P < 0.001), and percent bare ground (F1,1040 = 47.13, P < 0.001). Further analyses
of these vegetation characteristics were performed within years. Maximum vegetation
height, horizontal visual obstruction, and percent bare ground were all significantly
higher in treatment fields than reference fields in 2004 and 2005. Litter depth and

27
percent dead material were significantly higher in reference than treatment fields in 2004
and 2005 (Table 12).
There were also many differences in vegetation characteristics within treatment
fields (Table 13). Treatment x year interactions occurred in all analyses (horizontal
visual obstruction [F2,570 = 104.12, P < 0.001]; litter depth [F2,570 = 81.33, P < 0.001];
maximum vegetation height [F2,570 = 170.57, P < 0.001]; percent forb [F2,570 = 128.05, P
< 0.001], grass [F2,570 = 46.53, P < 0.001], dead material [F2,570 = 18.59, P < 0.001], and
bare ground [F2,570 = 56.38, P < 0.001] cover). Thus, further analyses were performed
within years. Horizontal visual obstruction was highest in the disked-2003 portion of the
treatment fields for both years (P < 0.001) (Table 13). Litter depth and percent dead
material were highest in the undisked portion of the treatment fields for 2004 and 2005 (P
< 0.001). In 2004, litter depth was similar between disked-2003 and disked-2004
portions, while in 2005 disked-2003 portions had more litter than disked-2004 portions.
In 2004, maximum vegetation height was highest in the disked-2003 portion, but in 2005,
maximum vegetation height was highest in disked-2003 and disked-2004 portions (P <
0.001). Forb cover was highest in the disked-2003 portion of treatment fields in 2004 (P
< 0.001), but shifted to being highest in the disked-2004 portion in 2005 (P < 0.001). In
2004, grass cover was higher in the undisked portion of the fields than the other 2
portions (P < 0.001). But, in 2005, grass cover was highest in undisked and disked-2003
portions (P < 0.001). Percent bare ground was highest in the disked-2004 portion of the
fields in 2004 and 2005 (P < 0.001) (Table 13).
Vegetation Influences

28
Vegetation had considerable influence on the presence of individual grassland
bird species in CRP fields. In treatment fields, 7 of the 8 most common grassland bird
species had significant explanatory models (Table 14). Presence of all the species, except
red-winged blackbirds, was best explained by more than 1 variable. Red-winged
blackbird occurrence was explained by increasing maximum vegetation height.
Dickcissel occurrence was best explained by 2 variables, increasing forb cover and visual
obstruction. Common yellowthroat occurrence was explained by increasing forb cover
and vegetation height, while sedge wrens were associated with increasing forb cover and
litter depth. Bobolinks were associated with increasing dead material and decreasing
bare ground. Western meadowlarks were associated with decreasing forb and grass
cover. Grasshopper sparrows were associated with increasing forb cover and litter depth,
and decreasing vegetation height (Table 14). Henslow’s sparrows did not have a
significant model in treatment fields.
In reference fields, 3 of the 8 most common grassland bird species had significant
explanatory models (Table 15). Common yellowthroats were associated with increasing
vegetation height, and red-winged blackbirds were associated with increasing litter depth.
Bobolink occurrence was explained by 2 variables, increasing grass and dead material
cover. Dickcissels, grasshopper sparrows, Henslow’s sparrows, sedge wrens, and
western meadowlarks did not have significant explanatory models in reference fields.
DISCUSSION
Overall, CRP grasslands in northeastern Nebraska provided habitat for a wide
variety of grassland birds. Of the 30 bird species observed in my study fields, 8 were
grassland bird species that commonly occurred in the fields and used the fields for

29
nesting, foraging, and raising young. In an analysis of North American Breeding Bird
Surveys (BBS) from 1966-1993, Herkert (1995) reported that over 75% of all grassland
bird species, including the 8 most commonly observed species in my study, have shown
significant declines throughout the Midwest. Recent BBS data (1966-2004) indicate that
grasshopper sparrows, western meadowlarks, dickcissels, and common yellowthroats
continue to decline (Sauer et al. 2005). However, these more recent data also show an
increasing trend for bobolinks and sedge wrens and a stable or a slightly decreasing trend
for red-winged blackbirds. Henslow’s sparrows, which were only observed in my
reference fields, may be one of the fastest declining grassland birds (Herkert 1997),
especially in the east and northeast portions of its range (Pruitt 1996). Recently,
however, local Henslow’s sparrow populations in the Midwest have increased (Sauer et
al. 2005), possibly due to habitat created by CRP (Herkert et al. 2002). These recent BBS
results, along with my findings, suggest that CRP grasslands will continue to be
important for these grassland bird species into the future and may be critical in
maintaining or increasing current populations.
Although CRP fields appear to provide critical habitat for grassland birds, the lack
of appropriate management often reduces their value to individual species. Disking and
interseeding creates a diverse mosaic of vegetation that positively benefits many
grassland bird species. Avian species diversity, richness (in 2004), and mean abundance
of all species combined were higher in the treatment than reference fields, indicating the
vegetation structure resulting from treatment of fields accommodated more species than
the reference fields. Millenbah et al. (1996) found similar results in newly planted CRP
fields, presumably because of the vegetation changes from the disturbance of planting

30
them. In my study, disked/interseeded portions of treatment fields had a diverse
vegetation composition and structure, consisting of various heights of forbs, weeds, and
grasses. Additionally, treatment fields consisted of 3 portions that were in different
vegetation successional stages. This diverse vegetation composition and structure within
disked/interseeded portions and variety of successional stages throughout treatment fields
is likely responsible for the high avian diversity, richness, and abundance.
Dickcissels were the most common grassland birds in my study area and were
most abundant in treatment fields. Dickcissels prefer grasslands that have dense, tall
cover that provide many elevated song perches (Hughes et al. 1999, Temple 2002,
Dechant et al. 2003). Additionally, areas with high proportions of forbs, especially
legumes, provide nesting cover, nest support (Appendix B), and an increased abundance
of invertebrate foods (Frawley and Best 1991, Patterson and Best 1996, Temple 2002).
Treatment fields provided the dense, tall legumes important to dickcissels for nesting and
foraging, along with live and dead forbs (sunflowers [Asteraceae], ragweed [Asteraceae],
hemp [Cannabaceae], and thistles [Asteraceae]) that were frequently used for perches
(Lucas Negus, personal observation). More specifically, the disked/interseeded portions
of the treatment fields, particularly the 1-year post-treatment portions, provided
vegetation structure that was nearly ideal for dickcissels. This preference for the
vegetation structure in the 1-year post-treatment portions within treatment fields is shown
by the consistent high abundance of dickcissels in the 1-year post-treatment portions of
treatment fields over the 2 years of the study. Dickcissels were significantly more
abundant in the 1-year post-treatment portion of treatment fields during both years of the
study. Red-winged blackbirds and common yellowthroats were also more abundant in

31
treatment fields. Like dickcissels, both species showed preferences for the 1-year post-
treatment portion of the treatment fields during both years of the study. Red-winged
blackbirds, which generally breed in a wide range of wetland and upland habitats
(Yasukawa and Searcy 1995), were observed almost exclusively in treatment fields.
They commonly placed their nests high (> 50 cm) in the legumes, and used live and dead
forbs for perching. Common yellowthroats also used the tall vegetation for perching, but
were more commonly observed hidden in the lower, dense legumes. This preference for
the low, dense vegetation by common yellowthroats is consistent with that reported by
Stewart (1953) for common yellowthroats in Michigan.
Grasshopper sparrows showed no preference between treatment and reference
fields in 2004, but preferred reference fields in 2005. Grasshopper sparrows generally
prefer grasslands of moderate height and density with patchy bare ground (Vickery 1996,
McCoy et al. 2001a). With the exception of bare ground, reference fields and undisked
portions of the treatment fields offered this suitable grassland habitat of moderate height
and density. However, within treatment fields in 2004, grasshopper sparrows displayed a
preference for areas with bare ground and were significantly more abundant in the
portions of treatment field that had been disked in the spring 2004 than any other field
portions. In the disked portions, grasshopper sparrows were frequently observed perched
on dirt clumps, where vegetation was sparse and short or intermediate in height
throughout the season and bare ground was abundant (Lucas Negus, personal
observation). In 2005, disked/interseeded portions of treatment fields had undergone at
least 1 full growing season, resulting in the majority (50 – 66%) of each field having tall,

32
dense vegetation that was less suitable for grasshopper sparrows compared to reference
fields.
Western meadowlarks were more abundant in treatment fields in 2004, but
showed no difference between treatment and reference fields in 2005. The preference of
treatment fields in 2004 was somewhat unexpected and contradictory to the findings of
Wiens and Rotenberry (1981), who reported western meadowlarks preferred areas with
high amounts of grass and litter cover. Reference fields were characterized more by high
amounts of grass and litter than treatment fields. Perhaps the treatment fields provided
adequate areas of grass and litter cover for nesting and were more attractive for foraging
than reference fields. In 2005, western meadowlarks showed no preference between
treatment and reference fields, possibly indicating that the vegetation changes in
treatment fields between 2004 and 2005 resulted in less desirable vegetation in 2005.
Bobolinks and Henslow’s sparrows were more abundant in reference fields than
treatment fields during both years of the study. Both species prefer habitats with high
percentages of grass and dead material, and high amounts of litter (Bollinger and Gavin
1992, Herkert and Glass 1999). This habitat was primarily found in reference fields and
the portion of treatment fields that was undisked. Henslow’s sparrows were most often
observed perched and nesting on hilltops where vegetation was not as high or dense as it
was in valleys. This is contrary to the findings of Zimmerman (1988) who found
Henslow’s sparrow males in Kansas set up territories in areas with taller vegetation than
surrounding areas that were unused. This may indicate Henslow’s sparrows use a wider
range of habitats than originally thought. Bollinger and Gavin (1992) found bobolinks in
New York preferred old hayfields (>8 years old), and fields >30 ha supported twice the

33
bobolink density of fields <10 ha. The reference fields in my study were all >10 ha and
had not been disturbed for >10 years; it is not surprising bobolink abundance was higher
in these fields than the treatment fields that had smaller undisked portions. Although
results from my study primarily support Bollinger and Gavins’ (1992) findings, bobolinks
were also observed nesting in small (<10 ha) undisked portions of treatment fields, as
well as disked/interseeded portions of the fields. This indicates that although treatment
fields were not used as much as reference fields were used by bobolinks, they still
provided adequate nesting habitat for a reduced number of bobolinks.
Not surprisingly, vegetation characteristics significantly affected presence of
many of the grassland bird species in CRP fields. Nearly every species (7 of 8 species)
had significant explanatory models in treatment fields, while only 3 species (red-winged
blackbirds, bobolinks, and common yellowthroats) had significant models in reference
fields. Reference fields likely had few explanatory models because vegetation was very
uniform throughout the entire fields, and species presence could not be attributed to
distinct vegetation characteristics. Dickcissels and common yellowthroats were
associated with similar vegetation variables in treatment fields. Dickcissels were
positively associated with vegetation height and forb cover, while common yellowthroats
were positively associated with visual obstruction and forb cover in treatment fields.
Both species were most commonly observed in disked/interseeded portions of the
treatment fields that had undergone at least 1 full growing season. Although these 2
species were associated with similar vegetation variables and were commonly observed
in the same portions of the treatment fields, they were likely able to coexist without
competition in these portions because the vertical vegetation structure present provided a

34
separate foraging space for each species. Cody (1968) concluded that vegetation height
was important because avian species foraging strategies are stratified by the vertical
vegetation structure. Dickcissels used the upper levels of the vertical structure in CRP
fields, while common yellowthroats used the lower levels. Maximum vegetation height
has been reported as being an important factor for dickcissels (Zimmerman 1966) and
was likely a key variable in my study because dickcissels were commonly observed
perching and nesting high in the vegetation. In addition to the height of vegetation, the
amount of forb cover present was also important for dickcissels. Delisle and Savidge
(1997) concluded dickcissel abundance was positively correlated to forb occurrence.
Forbs were used by dickcissels for both perching and nesting during my study. Visual
obstruction was likely a key variable for common yellowthroats because they prefer the
low, dense vegetation (Stewart 1953). A high visual obstruction measurement would
indicate the vegetation is both tall and dense, providing larger amounts of the preferred
dense vegetation. Common yellowthroats were most likely associated with increasing
forbs because they provided this tall, dense vegetation.
Red-winged blackbirds were positively associated with maximum vegetation
height in treatment fields and litter depth in reference fields. It was not unexpected that
red-winged blackbirds would be associated with increasing vegetation height in treatment
fields since they frequently selected the tallest vegetation for nest placement (Lucas
Negus, personal observation). The tall vegetation also provided prominent perch sites for
males to display and to alert others when potential predators are near (Yasukawa and
Searcy 1995). In reference fields, red-winged blackbirds were quite often observed in
transects parallel to or crossing valleys or water-ways in fields. These valleys and water-

35
ways generally consisted of dense, lush stands of grass compared to other areas in the
fields, presumably due to the higher amounts of moisture in these areas. Because of the
density of the vegetation in these areas, litter accumulation was considerably higher in
these areas than other areas of the field, explaining the positive association with litter
depth in reference fields.
Grasshopper sparrow presence in treatment fields was associated with increasing
forb cover and litter depth, and decreasing vegetation height. This model fits well with
observations reported by others. While habitat preferences of grasshopper sparrows vary
by region, they generally prefer somewhat disturbed areas with short or intermediate
vegetation height (Vickery 1996, Dechant et al. 2003). Whitmore (1981) noted
grasshopper sparrows preferred sparse, patchy habitat and nested in vegetation clumps
with high amounts of litter, and Schneider (1998) found increasing litter to be one of the
strongest predictors of grasshopper sparrow presence in grasslands.
Bobolinks were associated with increasing dead material cover and decreasing
bare ground in treatment fields and increasing grass and dead material cover in reference
fields. These predictive models seem to fit very well with the abundance surveys and
observations from this study, as well as other studies. In New York, bobolink density
was higher in areas with low legume cover and high amounts of litter and grass cover
(Bollinger and Gavin 1992). Similar to Henslow’s sparrows, bobolinks used undisked
CRP more often than disked/interseeded CRP. With no disturbance, the undisked CRP
had high amounts of litter, dead material, and grass cover.
Sedge wrens were associated with 2 vegetation variables in treatment fields,
increasing forb cover and litter depth. Sedge wrens prefer areas with tall, dense

36
vegetation (Johnson and Schwartz 1993b, Herkert et al. 2001). Delisle and Savidge
(1997) also found sedge wrens were positively associated with litter depth in Nebraska.
In treatment fields, the disked/interseeded portions had high amounts of forbs that likely
provided the tall, dense vegetation that was preferred by sedge wrens rather than the
undisked portions.
Western meadowlarks in treatment fields were negatively associated with forb
and grass cover, which is somewhat puzzling. Western meadowlarks occur in a diversity
of grassland habitats, but are generally thought to prefer areas of short, less dense grasses
(Lanyon 1994, McCoy 1996). One explanation may be that the forbs were too tall and
dense in disked portions of treatment fields. In a study on Montana grasslands, Dieni and
Jones (2003) reported that western meadowlarks avoided nesting in patches with high
percentages of forb cover. Another explanation may be that landscape characteristics of
treatment fields such as patch size, core area, and edge area, which were not included in
the model, were responsible for the presence of western meadowlarks rather than the
measured vegetation characteristics that were used to predict presence. Perhaps the best
explanation for the model comes from Wiens and Rotenberry (1981), who reported that
western meadowlarks did not correlate well with any habitat characteristics.
Vegetation characteristics also likely influenced nest densities in my study fields.
The nest density in treatment plots was nearly 3 times the nest density in reference plots.
Additionally, the nest plots in the undisked portion of the treatments fields had very low
densities (0.77 nests/ha), while the plots in disked/interseeded portions had moderate to
very high nest densities (3.00 – 6.25 nests/ha). A further evaluation of nest densities by
years post-treatment may better represent the differences in nest density. The density of

37
nests peaked in the 1-year post-treatment plots and decreased in all other stages of
treatment (no treatment, current-year, and 2-year post-treatment) (Figure 3). This trend is
due to the high number of red-winged blackbird and dickcissel nests that were observed
nearly exclusively in the disked/interseeded portions of treatment fields. As mentioned
above, the vegetation characteristics of disked/interseeded portions of treatment fields
were nearly ideal for dickcissels, and red-winged blackbirds were attracted to the same
characteristics. Dickcissels and red-winged blackbirds are both polygynous and as many
as 15 red-winged blackbird females have been reported nesting in 1 male territory,
resulting in high nest densities (Zimmerman 1966, Yasukawa and Searcy 1995).
Additionally, it appeared both species re-nested in the same area soon after a nest failure
rather than leaving the disked/interseeded portions of fields (Lucas Negus, personal
observation). This is somewhat contrary to that reported by Zimmerman (1982), who
reported only 17% of dickcissel females re-nested in the same territory they originally
nested in. This use of the same field portions for re-nesting may have also contributed to
the high nest density in the disked/interseeded portions of treatment fields.
Although differences in nest density between treatment and reference fields were
detected, there were no differences in nest success probability between treatment and
reference fields. The vegetation characteristics of treatment fields positively influenced
both abundance and nest density of many grassland bird species, but appear to not
enhance the nest success. The lack of difference in nest success suggests that vegetation
characteristics that were attractive for nesting did not offer more protection from
predators than reference field vegetation.

38
As mentioned, there was no difference in nest success probability between
treatment and reference fields, but nest success probability of all species combined and
red-winged blackbirds in 2005 was only half of that in 2004. Possible factors that may
have caused the decline in nest success include climate/weather differences, increased
search effort during the 2005 field season, increased nest densities, increased predator
numbers, and improved predator search images. Climate/weather differences were not
likely responsible; the weather conditions in 2004 and 2005 were similar and nest failure
due to weather was similar between the 2 years. In 2005, I hired 2 additional field
technicians that increased our nest searching efforts. Although we had more people in
the fields in 2005, the same methods for finding and checking nests were used as were
used the previous summer, so there should not have been an increase in disturbance
affecting nest success probability in 2005.
Although previous research is inconclusive, nest density may be correlated to
depredation rates. Robertson (1972) reported predation rates decreased with increasing
nest density, while Fretwell (1977) reported increased predation rates with increased
density. Zimmerman (1984) found no correlation between dickcissel nest density and
predation rates. Regardless, nest densities did not differ between 2004 and 2005, thus
nest densities likely had little influence on the nest success probability difference between
the 2 years.
Because the percentage of nests that were depredated increased from 64% in 2004
to 75% in 2005, it is possible that predators may have increased in abundance or
developed better search images in the 2005 field season. Although nest success
probability was similar between treatment and reference fields, more nests were

39
depredated in treatment fields simply because there were many more nests (primarily red-
winged blackbird and dickcissel nests) in these fields. Because treatment fields were first
disked/interseeded in 2004, it was the first time a large number of red-winged blackbirds
and dickcissels nested in these CRP fields. Therefore, it is possible that predator
populations may have been low or had not developed the search image to find these
particular nests. After the influx of nests in 2004, predator populations may have
increased or predators may have developed a search image for nests by the 2005 field
season.
Others have suggested yearly predation differences are due to fluctuations in
predator populations (Davis 2003). In a study on Saskatchewan grasslands, Davis (2003)
partially attributed abnormally low nest success in 1997 to an influx of prairie voles
(Microtus ochrogaster) during that year. While the prairie voles were not always the nest
predator, they likely also attracted other predators that foraged opportunistically on
grassland bird nests. Additionally, predators select prey from a large prey community
that includes grassland birds. Annual changes in grassland bird nest predation may be
reflective of changes in the overall prey community. Schmidt (1999) predicted that
increasing abundance of alternative foods (prey) would decrease grassland bird nest
depredation. A study on artificial nest success in CRP fields by Vander Lee et al. (1999)
supported this prediction, reporting increased nest success in plots in which supplemental
prey was supplied. While I did not evaluate changes in the prey community during this
study, it is likely that the prey community, like the bird and vegetation communities,
changed over the years of the study due to the disking/interseeding.

40
Overall, 71% of the nests were depredated over the 2 years of this study. This is
similar to findings of other studies in the Midwest (Patterson and Best 1996, Winter
1999). In my study, few of the nests were disturbed or destroyed, suggesting the
predators involved were most likely snakes or small mammals (Thompson et al. 1999,
Pietz and Granfors 2000). A depredated field sparrow nest was observed with 1 dead,
decapitated young, suggesting a mouse likely depredated the nest (Pietz and Granfors
2000). Thirteen-lined ground squirrels (Spermophilus tridecemlineatus) and Franklin’s
ground squirrels (Spermophilus franklinii), known nest predators, were also frequently
observed in the fields.
Other nests were unsuccessful due to abandonment, weather, or unknown causes.
Surprisingly, no nests were unsuccessful due to cowbird parasitism. Only a few nests
were parasitized over the 2 years of the study, but all of these nests were depredated
before the young fledged. This is surprising, however, because power lines, shelter belts,
and farm buildings, all reported as perch fields for cowbirds (Johnson and Temple 1990,
Schaffer et al. 2003), are abundant in the study area. Most likely, very few nests were
parasitized simply because there were very few cowbirds in the area. My survey results
indicated there was a very low abundance of cowbirds throughout the study area.
Vegetation characteristics also may have affected nest success of individual
species. For dickcissels, the only vegetation difference between successful and
unsuccessful nests occurred in 2005. Contrary to what was expected, maximum
vegetation height around unsuccessful nests was higher than that around successful nests.
It is expected that higher vegetation around nests would provide better nest concealment,
increasing the probability of nest success. However, this pattern may indicate predators

41
that use olfactory senses or audio cues rather than visual cues, such as snakes, were
responsible for depredation. Zimmerman (1984) concluded in his study on dickcissels in
Kansas that snakes were most likely a major predator on nests and nest density or
concealment of nests were not important factors influencing the rate of predation.
Instead, snake density was likely the most important factor influencing depredation. In
my study, most depredated nests were not disturbed or destroyed indicating snakes could
have been the primary nest predator (Thompson et al. 1999).
Nest success of the 2 most abundant ground-nesting species (grasshopper
sparrows and bobolinks) did not appear to be significantly affected by vegetation.
Successful grasshopper sparrow nests had more bare ground surrounding them than
unsuccessful nests. Whitmore (1981) reported grasshopper sparrow territories had higher
amounts of bare ground than areas not used for territories. Grasshopper sparrows
generally nested in clumps of vegetation, usually surrounded by bare ground. This also
indicates grasshopper sparrows may be more successful in areas that have a diversity of
grasses, forbs, and bare ground, rather than an evenly distributed monoculture of
vegetation. Bobolink nests exhibited no vegetation differences between successful and
unsuccessful nests. Bobolinks primarily nested in undisked CRP, where there were few
forbs and vegetation was uniform throughout the field. Contrary to other studies,
bobolinks in my study concealed their nests in litter surrounded by evenly-distributed
grasses and not in clumps of forbs or grasses (Martin and Gavin 1995). This is likely the
reason there were no vegetation differences between successful and unsuccessful
bobolink nests.
CONSERVATION IMPLICATIONS AND RECOMMENDATIONS

42
As indicated by my results and those of many others, CRP fields provide critical
habitat for grassland breeding birds (Johnson and Schwartz 1993b, Igl and Johnson 1995,
Best et al. 1997, Delisle and Savidge 1997). However, as CRP fields age without
management, their benefits to grassland birds may decrease (Millenbah et al. 1996).
Aging CRP fields can be enhanced using several management methods including haying,
burning, grazing, and disking/interseeding legumes (McCoy et al. 2001b). In my study,
disking/interseeding legumes was promoted primarily because of the diverse vegetation
created in CRP fields dominated by smooth brome compared to other practices.
Although the costs of management vary by region and practice, disking/interseeding is
generally more expensive than the other practices, with fuel costs contributing greatly.
However, if entire CRP fields are hayed or grazed, a 25% reduction in CRP payments is
imposed (U. S. Department of Agriculture 2003a). Depending on land rental rates, this
reduction in CRP payments may make haying or grazing more costly than
disking/interseeding. Burning is the most cost effective of the practices, but is rarely
used in the region because of lack of burn crews, landowner experience, and social
factors (fear of burning). Additionally, burning smooth brome does not create the
vegetation diversity that can be created using disking/interseeding (Scott Wessel,
Nebraska Game and Parks Commission, personal communication).
According to my results, disking/interseeding legumes benefited many grassland
bird species in several ways. Disked/interseeded fields accommodated higher
abundances, more species, and higher diversities of grassland birds than the reference
fields. The manner in which the fields were disked/interseeded seemed to create a range
of vegetation characteristics, or successional stages, that were desirable to a wide array of

43
grassland bird species. Disking/interseeding produced the weedy, sparse vegetation that
is similar to that of newly seeded CRP (Millenbah et al. 1996). In the first growing
season following disking, typical vegetation characteristics included little to no litter,
high amounts of bare ground, sparse and short stature legumes (alfalfa and red clover)
early in the season, moderately dense and medium height planted legumes later in the
season, with patches of unplanted forbs (sunflowers, ragweed, hemp, and thistles)
occurring throughout the field (Lucas Negus, personal observation). Grasses present in
the first growing season were typically annuals such as foxtails (Setaria spp.).
Vegetation in the 1-year post-treatment portions was predominantly tall, dense planted
legumes. Yellow sweetclover provided tall vegetation stature, whereas the red clover and
alfalfa filled in the lower levels of the vertical structure. There were few other forbs and
grasses present after 1 full growing season. However, many unplanted forb stalks
remained from the previous season and were frequently used for perching. In the 2-years
post treatment portions, the vegetation consisted mainly of tall smooth brome, with sparse
alfalfa and red clover interspersed in the lower levels of the vegetation. There was little
bare ground in the third year after disking. Reference fields were typically dominated by
short to medium height smooth brome, few or no forbs, moderate amounts of standing
dead material, high amounts of litter, and no bare ground. Vegetation characteristics in
reference fields did not change during the 2 years of the study.
It is likely factors other than vegetation, such as patch characteristics and
invertebrate communities, had impacts on the grassland bird community in the CRP
fields. Core area, field or patch size, edge area, and distance to edge have been reported
to affect grassland bird communities throughout the Midwest (Herkert 1994, Helzer and

44
Jelinski 1999, Winter and Faaborg 1999, Sporrong 2004). While I did not examine patch
characteristics such as field or patch size and edge area, it is possible that these factors
influenced the grassland bird communities in my study. Moreover, it is also possible that
disked/interseeded portions were too small to accommodate some area sensitive species
such as grasshopper sparrows, bobolinks, and western meadowlarks (Helzer and Jelinski
1999, Winter and Faaborg 1999). Additionally, the increased amount of edge created in
treatment fields by disking/interseeding portions of the fields may have reduced nest
success due to increased predation. Winter et al. (2000) suggested that dickcissels and
other habitat generalists are affected by decreased nest success near edges due to
increased depredation (primarily by mesocarnivores) near these edges. However, nearly
all “edge” studies focused on “hard” edges such as tree rows or roads, whereas the edges
in my project were “soft” edges, representing changes in herbaceous vegetation structure.
Further research on the effects of “soft” edges on grassland birds is needed. I attempted
as best as I could to control for field size by selecting similar sized reference and
treatment fields, as well as selecting treatment fields in which portions were
disked/interseeded in similar sizes and shapes, but ultimately could not select fields that
were identical in size and shape. Although not likely, these small differences in size and
shape may have influenced bird communities in my treatment and reference fields.
Invertebrates are important food resources for breeding grassland birds, as well as
many other bird species (Hull et al. 1996). Several studies have reported that grassland
birds opportunistically forage on invertebrates from the orders Coleoptera, Lepidoptera,
Orthoptera, and Aranae (Kaspari and Joern 1993, Kobal et al. 1998, McIntyre and
Thompson 2003). An examination of the invertebrate community in my study fields in

45
2004 indicated treatment fields provided higher abundances and biomasses of these
orders than reference fields (Appendix C). Grassland birds are likely not only responding
to the vegetation differences in the treatment fields, but also to the invertebrate
community that is influenced by the vegetation changes.
To maximize vegetation and grassland bird diversity in CRP fields, I recommend
using disking/interseeding legumes as a management tool. Moreover, I recommend
establishing an annual rotation of this management practice to maximize benefits. An
annual rotation would create a mosaic of vegetation characteristics, which would provide
habitat for several species simultaneously (Herkert 1996, Madden et al. 2000, Winter et
al. 2005). Ideally, fields could be divided into fourths, with one-fourth of the field being
disked/interseeded annually. To accommodate the suite of grassland birds, I also
recommend maintaining one-fourth of the field out of the annual rotation to provide the
mature grassland habitat that is preferred by several species (e.g., Henslow’s sparrows
and bobolinks) (Herkert 1996). Additionally, fields that are difficult to disk/interseed
because of rough terrain, poor accessibility, or other reasons should not be
disked/interseeded, thus providing important habitat to those species that may be
displaced by disking/interseeding.
This study was relatively short in duration, and the benefits of
disking/interseeding beyond 2 years after the initial disking/interseeding are unknown.
Petersen and Best (1999) concluded that 2-3 years of post-disturbance data for
perturbation experiments may be insufficient for grassland birds. Vegetation
characteristics indicate that in the third year after disking/interseeding, vegetation
becomes more similar to that in reference fields. It can be predicted that the grassland

46
bird community will likely reflect these vegetation changes as the field matures.
However, exactly how long it takes for grassland bird communities in treatment fields to
become similar to those of reference fields is not known. Researching the
disked/interseeded fields for several years (> 5) following management would yield more
confident conclusions.
Future grassland bird studies in CRP should not only focus on vegetation
characteristics, but should also focus on the landscape characteristics and invertebrate
communities as mentioned previously. Additionally, studies should be long-term (> 3
years) (Petersen and Best 1999) and cover a wide geographic range (Winter et al. 2005).
Grassland habitats vary highly between years and regions, as do grassland bird
populations. Thus it is important that studies extend over wide geographic regions and
years to accurately measure grassland bird communities (Igl and Johnson 1999). These
types of studies are important to better understand grassland bird populations and trends
in the future, especially as required management of CRP becomes more common and
widespread. Future studies on management practices, specifically disking/interseeding,
should also examine source-sink population implications. Although disked/interseeded
portions had higher abundances and nest densities than undisked portions, they may be
acting as sink habitats rather than source habitats. Finally, future studies of CRP
vegetation management should focus on techniques that will increase the longevity of the
vegetation created by disking/interseeding. Disking/interseeding legumes may be more
effective when used in conjunction with other management techniques such as herbicide
application, burning, haying, or grazing.
LITERATURE CITED

47
Best, L. B., H. Campa, III, K. E. Kemp, R. J. Robel, M. R. Ryan, J. A. Savidge, H. P.
Weeks, Jr., and S. R. Winterstein. 1997. Bird abundance and nesting in CRP
fields and cropland in the Midwest: a regional approach. Wildife Society
Bulletin 25:864-877.
__________. 1998. Avian abundance in CRP and crop fields during winter in the
Midwest. American Midland Naturalist 139:311-324.
Bollinger, E. K., P. B. Bollinger, and T. A. Gavin. 1990. Effects of cropping on eastern
populations of the bobolink. Wildlife Society Bulletin 18:142-150.
__________, and T. A. Gavin. 1992. Eastern bobolink populations: ecology and
conservation in an agricultural landscape. Pages 497-506 in J. M. Hagan III and
D. W. Johnston, editors, Ecology and conservation of neotropical migrant
landbirds, Smithsonian Institute Press, Washington, D. C., USA.
Brower, J. E., and J. H. Zar. 1977. Field and laboratory methods for general ecology.
William C. Brown Publishers, Dubuque, Iowa, USA.
Cody, M. L. 1968. On the methods of resource division in grassland bird communities.
American Midland Naturalist 102:107-147.
Conover, W. J., and R. L. Iman. 1981. Rank transformations as a bridge between
parametric and nonparametric statistics. American Statistician 35:124-129.
Daubenmire, R. F. 1959. A canopy coverage method of vegetational analysis.
Northwest Science 33:43-64.
Davis, S. K., and S. G. Sealy. 2000. Cowbird parasitism and nest predation in
fragmented grasslands of southwestern Manitoba. Pages 220-228 in J. N. Smith,

48
T. L. Cook, S. I. Rothstein, S. K. Robinson, and S. G. Sealy, editors, Ecology and
management of cowbirds and their hosts. University of Texas Press, Austin, USA.
__________. 2003. Nesting ecology of mixed-grass prairie songbirds in southern
Saskatchewan. Wilson Bulletin 115:119-130.
Dechant, J. A., M. L. Sondreal, D. H. Johnson, L. D. Igl, C. M. Goldade, M. P.
Nenneman, and B. R. Euliss. 2003. Effects of management practices on
grassland birds: Grasshopper Sparrow. Northern Prairie Wildlife Research
Center, Jamestown, ND. Jamestown, ND: Northern Prairie Wildlife Research
Center Home Page.
http://www.npwrc.usgs.gov/resource/literatr/grasbird/grsp/grsp.htm
(Version 12AUG2004). Accessed 5 December, 2005.
Delisle, J. M., and J. A. Savidge. 1997. Avian use and vegetation characteristics of
Conservation Reserve Program fields. Journal of Wildlife Management 61:318-
325.
Dieni, J. S., and S.L. Jones. 2003. Grassland songbird nest site selection patterns in
northcentral Montana. Wilson Bulletin 115:388-396.
Dow, D. D. 1978. A test of significance for Mayfield’s method of calculating nest
success. Wilson Bulletin 90:291-295.
Frawley, B. J., and L. B. Best. 1991. Effects of mowing on breeding bird abundance and
species composition in alfalfa fields. Wildlife Society Bulletin 19:135-142.
Fretwell, S. D. 1977. Is the dickcissel a threatened species? American Birds 31:923-
932.

49
Guiliano, W. M., and S. E. Daves. 2002. Avian responses to warm-season grass use in
pasture and hayfield management. Biological Conservation 106:1-9.
Hammond, C. L. 1982. Soil Survey of Stanton County Nebraska. United States
Department of Agriculture- Soil Conservation Service in cooperation with
University of Nebraska, Conservation and Survey Division.
Hays, R. L., R. P. Webb, and A .H. Farmer. 1989. Effects of the Conservation Reserve
Program on wildlife habitat: results of 1988 monitoring. Transactions of the
North American Wildlife and Natural Resources Conference 54:365-376.
Helzer, C. J. 1996. The effects of wet meadow fragmentation on grassland birds.
Masters Thesis, University of Nebraska, Lincoln.
__________, and D. E. Jelinski. 1999. The relative importance of patch area and
perimeter-area ratio to grassland breeding birds. Ecological Applications 9:1448-
1458.
Herkert, J. R. 1994. The effects of habitat fragmentation on Midwestern grassland bird
communities. Ecological Applications 4:461-471.
__________. 1995. An analysis of Midwestern breeding bird population trends: 1966-
1993. American Midland Naturalist 134:41-50.
__________. 1997. Population trends of the Henslow’s sparrow in relation to the
Conservation Reserve Program in Illinois, 1975-1995. Journal of Field
Ornithology 68:235-244.
__________. 1998. The influence of the CRP on grasshopper sparrow population trends
in the mid-continental United States. Wildlife Society Bulletin 26:227-231.

50
__________, and W. D. Glass. 1999. Henslow’s sparrow response to prescribed fire in
an Illinois prairie remnant. Studies in Avian Biology 19:160-164.
__________, D. E. Kroodsma, and J. P. Gibbs. 2001. Sedge wren (Cistothorus
platensis). No. 582 in A. Poole and F. Gill, editors, The birds of North America,
The Birds of North America, Inc., Philadelphia, Pennsylvania, USA.
__________, P. D. Vickery, and D. E. Kroodsma. 2002. Henslow’s sparrow
(Ammomdramus henslowii). No. 672 in A. Poole and F. Gill, editors, The birds of
North America, The Birds of North America, Inc., Philadelphia, Pennsylvania,
USA.
__________, D. L. Reinking, D. A. Wiedenfeld, M. Winter, J. L. Zimmerman, W. E.
Jensen, E. J. Finck, R. R. Koford, D. H. Wolfe, S. K. Sherrod, M. A. Jenkins, J.
Faaborg, and S. K. Robison. 2003. Effects of prairie fragmentation on the nest
success of breeding birds in the Midcontinental United States. Conservation
Biology 17:587-594.
Horn, D. J., and R. R. Koford. 2000. Relation of grassland bird abundance to mowing of
Conservation Reserve Program fields in North Dakota. Wildife Society Bulletin
28:653-659.
Hosmer, D. W., Jr., and S. Lemeshow. 1989. Applied logistic regression. John Willey
and Sons, New York, New York, USA.
Hughes, J. P., R. J. Robel, K. E. Kemp, and J. L. Zimmerman. 1999. Effects of habitat
on dickcissel abundance and nest success in Conservation Reserve Program fields
in Kansas. Journal of Wildlife Management 63:523-529.

51
Hull, S. D., R. J. Robel, and K. E. Kemp. 1996. Summer avian abundance, invertebrate
biomass, and forbs in Kansas CRP. Prairie Naturalist 28:1-12.
Igl, L. D., and D. H. Johnson. 1995. Dramatic increase of Le Conte’s Sparrow in
Conservation Reserve Program fields in northern Great Plains. Prairie Naturalist
27:89-94.
_________, and _________. 1999. Le Conte’s sparrows breeding in Conservation
Reserve Program fields: precipitation and patterns of population change. Studies
in Avian Biology 19:178-186.
Jackson, J. R., G. A. Hurst, and E. A. Gluesing. 1987. Abundance and selection of
invertebrates by northern bobwhite chicks. Proceedings Annual Conference
Southeast Association Fish and Wildlife Agencies 41:303-310.
Johnson, D. H. 1979. Estimating nest success: the Mayfield method and an alternative.
Auk 96:651-661.
__________, and L. D. Igl. 2001. Area requirements of grassland birds: a regional
perspective. Auk 118:24-34.
__________, and M. D. Schwartz. 1993a. The Conservation Reserve Program and
grassland birds. Conservation Biology 7:934-937.
__________, and M. D. Schwartz. 1993b. The Conservation Reserve Program: habitat
for grassland birds. Great Plains Research 3:273-295.
Johnson, R. G., and S. A. Temple. 1986. Assessing habitat quality for birds nesting in
fragmented tallgrass prairies. Pages 245-249 in J. Verner, M.L. Morrison, and C.

52
J. Ralph, editors. Modeling habitat relationships of terrestrial vertebrates.
University of Wisconsin Press, Madison, USA.
__________, and S. A. Temple. 1990. Nest predation and brood parasitism of tallgrass
prairie birds. Journal of Wildlife Management 54:106-111.
Kaspari, M., and A. Joern. 1993. Prey choice by three insectivorous grassland birds:
reevaluating opportunism. Oikos 68:414-430.
King, J. W., and J. A. Savidge. 1995. Effects of the Conservation Reserve Program on
wildlife in southeast Nebraska. Wildlife Society Bulletin 23:377-385.
Knopf, F. L., and F. B. Samson. 1997. Conservation of grassland vertebrates. Pages
273-289 in F. L. Knopf and F. B. Samson, editors. Ecology and conservation of
Great Plains vertebrates. Springer-Verlag, New York, USA.
Kobal, S. N., N. F. Payne, and D. R. Ludwig. 1998. Nestling food habits of 7 grassland
bird species and insect abundance in grassland habitats in northern Illinois.
Transactions of the Illinois State Academy of Science 91:69-75.
Koford, R. R., B. S. Bowen, J. T. Lokemoen, and A. D. Kruse. 2000. Cowbird
parasitism in grassland and cropland in the northern Great Plains. Pages 229-235
in J. N. M. Smith, T. L. Cook, S. I. Rothstein, S. K. Robison, and S. G. Sealy,
editors. Ecology and management of cowbirds and their hosts. University of
Texas Press, Austin, USA.
Krebs, C. J. 1999. Ecological methodology, second edition. Addison-Welsey
Educational Publishers, Inc., Menlo Park, California, USA.

53
Lanyon, W. E. 1994. Western Meadowlark (Sturnella neglecta). No. 104 in A. Poole
and F. Gill, editors, The birds of North America, The Academy of Natural
Sciences, Philadelphia, Pennsylvania, and The American Ornithologists’ Union,
Washington, D.C., USA.
Leathers, R. J. 2003. Relative invertebrate availability in Nebraska’s Conservation
Reserve Program Management Access Program. Master’s Thesis. South Dakota
State University, Brookings.
Luttschwager, K. A., and K. F. Higgins. 1992. Nongame bird, game bird, and deer use
of Conservation Reserve Program fields in eastern South Dakota. Proceedings of
South Dakota Academy of Science 71:31-36.
Madden, E. M., R. K. Murphy, A. J. Hansen, and L. Murray. 2000. Models for guiding
management of prairie bird habitat in northwestern North Dakota. American
Midland Naturalist 144:377-392.
Manley, S. W., R. S. Fuller, J. M. Lee, and L. E. Brennan. 1994. Arthropod response to
strip disking in old fields managed for bobwhites. Proceedings Annual
Conference Southeast Association Fish and Wildlife Agencies 48:227-235.
Martin, S. G., and T. A. Gavin. 1995. Bobolink (Dolichonyx oryzivorus). No. 176 in A.
Poole and F. Gill, editors, The birds of North America, The Academy of Natural
Sciences, Philadelphia, Pennsylvania, and The American Ornithologists’ Union,
Washington, D.C., USA.
Martin, T. E., and G. R. Geupel. 1993. Nest-monitoring plots: methods for locating
nests and monitoring success. Journal of Field Ornithology 64:507-519.

54
Mayfield, H. F. 1975. Suggestions for calculating nest success. Wilson Bulletin 87:456-
466.
McCoy, T. D. 1996. Avian abundance, composition, and reproductive success on
Conservation Reserve Program fields in northern Missouri. Master’s Thesis.
University of Missouri, Columbia.
__________, M. R. Ryan, E. W. Kurzejeski, and L. W. Burger, Jr. 1999. Conservation
Reserve Program: Source or sink habitat for grassland birds in Missouri? Journal
of Wildlife Management 63:530-538.
__________, M. R. Ryan, L. W. Burger, Jr., and E. W. Kurzejeski. 2001a. Effects of
conservation practice, mowing, and temporal changes on vegetation structure on
CRP fields in northern Missouri. Wildlife Society Bulletin 29:979-987.
__________, E. W. Kurzejeski, L. W. Burger, Jr., and M. R. Ryan. 2001b. Grassland
bird conservation: CP1 vs. CP2 plantings in Conservation Reserve Program fields
in Missouri. American Midland Naturalist 145:1-17.
McIntyre, N. E., and T. R. Thompson. 2003. A comparison of Conservation Reserve
Program habitat plantings with respect to arthropod prey for grassland birds.
American Midland Naturalist 150:291-301.
Millenbah, K. F. 1993. The effects of different age classes of fields enrolled in the
Conservation Reserve Program in Michigan on avian diversity, density, and
productivity. Master’s Thesis, Michigan State University, East Lansing.

55
__________, S. R. Winterstein, H. Campa III, L. T. Furrow, and R. B. Minnis. 1996.
Effects of Conservation Reserve Program field age on avian relative abundance,
diversity, and productivity. Wilson Bulletin 108:760-770.
Natural Resource Conservation Service. 2002. Practice specification-early successional
habitat development/management-disking. Nebraska Field Office Technical
Guide 525. S-647a.
__________. 2003. Nebraska’s major land resource areas.
http://www.ne.nrcs.usda.gov/technical/MLRA_MAP.html. Accessed 29 April
2004.
Patterson, M. P., and L. B. Best. 1996. Bird abundance and nesting success in Iowa CRP
fields: the importance of vegetation structure and composition. American
Midland Naturalist 135:153-167.
Petersen, K. L., and L. B. Best. 1999. Design and duration of perturbation experiments:
implications for data interpretation. Studies in Avian Biology 19:230-236.
Pietz, P. J., and D. A. Granfors. 2000. Identifying predators and fates of grassland
passerine nests using miniature video cameras. Journal of Wildlife Management
64:71-87.
Pruitt, L. 1996. Henslow’s sparrow status assessment. U. S. Fish and Wildlife Service,
Bloomington Field Office, Region 3, Bloomington, Indiana, USA.
Pulliam, H. R. 1988. Sources, sinks, and population regulation. American Midland
Naturalist 132:652-661.

56
Reynolds, R. E., T. L. Shaffer, J. R. Sauer, and B. G. Peterjohn. 1994. Conservation
Reserve Program: benefit for grassland birds in the northern plains. Transactions
of the North American Wildlife and Natural Resources Conference 59:328-336.
Robel, R. J., J. N. Briggs, A. D. Dayton, and L. C. Hulbert. 1970. Relationships between
visual obstruction measurements and weight of grassland vegetation. Journal of
Range Management 23:295-297.
Robertson, R. J. 1972. Optimal niche space of the redwinged blackbird (Agelaius
phoeniceus). I. Nesting success in marsh and upland habitat. Canadian Journal of
Zoology 50:247-263.
Rotella, J. J., E. M. Madden, and A. J. Hansen. 1999. Sampling considerations for
estimating density of passerines in grasslands. Studies in Avian Biology 19:237-
243.
Ryan, M. R., L. W. Burger, and E. W. Kurzejeski. 1998. The impact of CRP on avian
wildlife: a review. Journal of Production Agriculture 11:61-66.
Samson, F. B., and F. L. Knopf. 1994. Prairie conservation in North America.
BioScience 44:418-421.
SAS Institute. 2003. SAS 9.1 for windows. SAS Institute Inc., Cary, North Carolina,
USA.
Sauer, J. R., J. E. Hines, and J. Fallon. 2005. The North American breeding bird survey,
results and analysis 1966 - 2004. Version 2005.2. USGS Patuxent Wildlife
Research Center, Laurel, Maryland, USA.

57
Schmidt, K. A. 1999. Foraging theory as a conceptual framework for studying nest
predation. Oikos 85:151-160.
Schneider, N. A. 1998. Passerine use of grasslands managed with two grazing regimes on
the Missouri Coteau in North Dakota. Masters’ Thesis, South Dakota State
University, Brookings, South Dakota.
Shaffer, J. A., C. M. Goldade, M. F. Dinkins, D. H. Johnson, L. D. Igl, and B. R. Euliss.
2003. Brown-headed cowbirds in grasslands: their habitats, hosts, and response to
management. Prairie Naturalist 35:145-186.
Sporrong, J. M. 2004. Response of nongame birds and terrestrial invertebrates to
restoration of upland grasslands in the Rainwater Basin Region, Nebraska.
Masters’ Thesis, Oklahoma State University, Stillwater.
Stewart, R. E. 1953. A life history study of the yellow-throat. Wilson Bulletin 65:99-
115.
Steinauer, E. M., and S. L. Collins. 1996. Prairie ecology-the Tallgrass Prairie. Pages
39-52 in F. B. Samson and F. L. Knopf, editors. Prairie conservation-preserving
North America’s most endangered ecosystem. Island Press, Washington DC.,
USA.
Swanson, D. A., D. P. Scott, and D. L. Risley. 1999. Wildlife benefits of the
Conservation Reserve Program in Ohio. Journal of Soil and Water Conservation
54:390-394.
__________. 2003. Effects of management practices on grassland birds: savannah
sparrow. Northern Prairie Wildlife Research Center, Jamestown, ND.

58
Jamestown, ND: Northern Prairie Wildlife Research Center Home Page.
http://www.npwrc.usgs.gov/resource/literatr/grasbird/savs/savs.htm
(Version 12DEC2003). Accessed 29 April 2004.
Taylor, S. 2002. Focus on pheasants- proposed activities. Nebraska Game and Parks
Commission. Unpublished report. Lincoln, Nebraska, USA.
Temple, S. A., B. M. Fevold, L. K. Paine, D. J. Undersander, and D. W. Sample.
1999. Nesting birds and grazing cattle: accommodating both on midwestern
pastures. Studies in Avian Biology 19:196-202.
__________. 2002. Dickcissel (Spiza americana). No. 703 in A. Poole and F. Gill,
editors, The birds of North America, The Birds of North America, Inc.,
Philadelphia, Pennsylvania, USA.
Thompson, F. R., III, W. Dijak, and D. E. Burhans. 1999. Video identification of
predators at songbird nests in old fields. Auk 116:259-264.
U.S. Department of Agriculture. 2003a. 2002 Farm Bill—Conservation Reserve
Program—long-term policy; interim rule. Federal Register, 7 CFR Part 1410.
_________. 2003b. The Conservation Reserve Program statistics.
http://www.fsa.usda.gov/dafp/cepd/crp.htm. Accessed 29 April 2004.
Vander Lee, B. A., R. S. Lutz, L. A. Hansen, and N. E. Mathews. 1999. Effects of
supplemental prey, vegetation, and time on success of artificial nests. Journal of
Wildlife Management 63:1299-1305.

59
Vickery, P. D. 1996. Grasshopper sparrow (Ammodramus savannarum). No. 239 in A.
Poole and F. Gill, editors, The birds of North America, The Academy of Natural
Sciences, Philadelphia, Pennsylvania, and The American Ornithologists’ Union,
Washington, D.C., USA.
Whitmore, R. C. 1981. Structural characteristics of grasshopper sparrow habitat.
Journal of Wildlife Management 45:811-814.
Whitmore, R. W., K. P. Pruess, and R. E. Gold. 1986. Insect food selection by 2-week-
old ring-necked pheasant chicks. Journal of Wildlife Management 50:223-228.
Wiens, J. A., and J. T. Rotenberry. 1981. Habitat associations and community structure
of birds in shrub steppe environments. Ecological Monographs 51:21-41.
__________. 1999. Nesting biology of dickcissels and Henslow’s sparrows in
southwestern Missouri prairie fragments. Wilson Bulletin 111:515-527.
__________, and J. Faaborg. 1999. Patterns of area sensitivity in grassland-nesting
birds. Conservation Biology 13:1424-1436.
__________, D. H. Johnson, and J. Faaborg. 2000. Evidence for edge effects on
multiple levels in tallgrass prairie. Condor 102:256-266.
__________, S. E. Hawks, J. A. Shaffer, and D. H. Johnson. 2003. Guidelines for
finding nests of passerine birds in tallgrass prairie. Prairie Naturalist 35:197-211.
__________, D. H. Johnson, and J. A. Shaffer. 2005. Variability in vegetation effects on
density and nesting success of grassland birds. Journal of Wildlife Management
69:185-198.

60
Yasukawa, K., and W. A. Searcy. 1995. Red-winged blackbird (Agelaius phoeniceus).
No. 184 in A. Poole and F. Gill, editors, The birds of North America, The
Academy of Natural Sciences, Philadelphia, Pennsylvania, and The American
Ornithologists’ Union, Washington, D.C., USA.
Zar, J. H. 1999. Biostatistical analysis, fourth edition. Prentice-Hall, Upper Saddle
River, New Jersey, USA.
Zimmerman, J. L. 1966. Polygyny in the dickcissel. Auk 83:534-546.
Zimmerman, J. L. 1982. Nesting success of dickcissels (Spiza americana) in preferred
and less preferred habitats. Auk 99:292-298.
__________. 1984. Nest predation and its relationship to habitat and nest density in
dickcissels. Condor 86:68-72.
__________. 1988. Breeding season habitat selection by the Henslow’s sparrow
(Ammodramus henslowii) in Kansas. Wilson Bulletin 100:17-24.
__________. 1992. Density-independent factors affecting the avian diversity of the
tallgrass prairie community. Wilson Bulletin 104:85-94.
__________. 1997. Avian community responses to fire, grazing, and drought in the
tallgrass prairie. Pages 167-180 in F. L. Knopf and F. B. Samson, editors.
Ecology and conservation of Great Plains vertebrates. Springer-Verlag, New
York, USA.

61
Table 1. Overall relative abundancea, species richnessb, and species diversityc of
breeding grassland birds in treatment and reference fields in Stanton County, Nebraska,
2004-2005.
a Mean abundance of all bird species from 3 sampling periods during each year. b Number of avian species observed in each field. c Shannon-Weiner species diversity index (Krebs 1999). d P = P-value for treatment (treatment vs. reference) and year (2004 vs. 2005) effects
from a 2-way analysis of variance. Interaction effects (treatment x year) were observed
for species richness (F1,28 = 4.47, P = 0.044). Means for species richness are reported
separately for each year.
Treatment (n = 8)
Reference (n = 8)
Pd
Year Mean S.E. Mean S.E. Treatment Year Overall abundance (no./transect)
2004-2005
4.49 0.25 2.93 0.21 <0.001 0.150
2004 10.13 0.48 6.00 0.53 <0.001 Species richness
2005 10.38 0.63 8.75 0.70 0.106 Species diversity
2004-2005
1.41 0.06 1.23 0.05 0.034 0.229
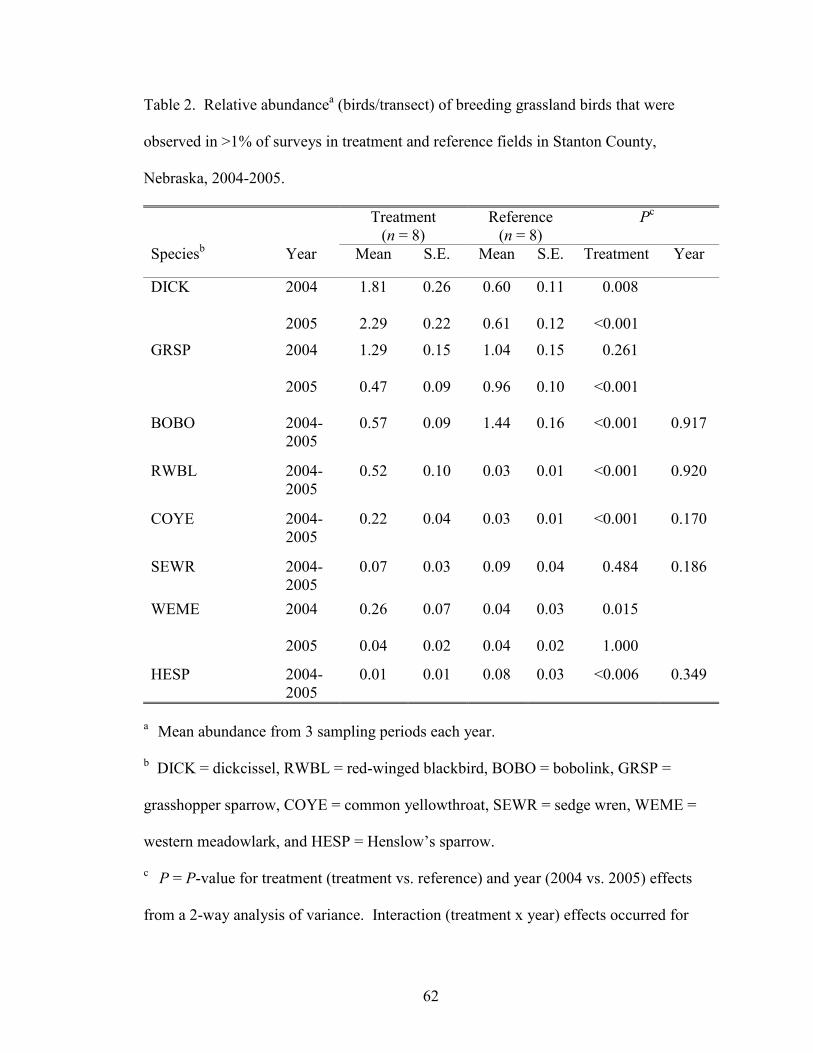
62
Table 2. Relative abundancea (birds/transect) of breeding grassland birds that were
observed in >1% of surveys in treatment and reference fields in Stanton County,
Nebraska, 2004-2005.
Treatment (n = 8)
Reference (n = 8)
Pc
Speciesb Year Mean S.E. Mean S.E. Treatment Year DICK 2004
2005
1.81
2.29
0.26
0.22
0.60
0.61
0.11
0.12
0.008
<0.001
GRSP 2004 2005
1.29
0.47
0.15
0.09
1.04
0.96
0.15
0.10
0.261
<0.001
BOBO 2004- 2005
0.57
0.09 1.44 0.16 <0.001 0.917
RWBL 2004-2005
0.52 0.10 0.03 0.01 <0.001 0.920
COYE 2004-2005
0.22 0.04 0.03 0.01 <0.001 0.170
SEWR 2004-2005
0.07 0.03 0.09 0.04 0.484 0.186
WEME 2004 2005
0.26
0.04
0.07
0.02
0.04
0.04
0.03
0.02
0.015
1.000
HESP 2004-2005
0.01 0.01 0.08 0.03 <0.006 0.349
a Mean abundance from 3 sampling periods each year. b DICK = dickcissel, RWBL = red-winged blackbird, BOBO = bobolink, GRSP =
grasshopper sparrow, COYE = common yellowthroat, SEWR = sedge wren, WEME =
western meadowlark, and HESP = Henslow’s sparrow. c P = P-value for treatment (treatment vs. reference) and year (2004 vs. 2005) effects
from a 2-way analysis of variance. Interaction (treatment x year) effects occurred for

63
dickcissels (F1,260 = 5.13, P = 0.024), grasshopper sparrows (F1,260 = 8.55, P = 0.004), and
western meadowlarks (F1,260 = 5.29, P = 0.022).

64
Table 3. Relative abundancea (birds/transect) of breeding grassland birds occurring in
>1% of surveys in 3 portions of Conservation Reserve Program fields managed by
disking/interseeding in Stanton County, Nebraska, 2004-2005.
Disked-2003 (n = 8)
Disked-2004 (n = 8)
Undisked (n = 8)
Speciesc Year Mean S.E. Mean S.E. Mean S.E. All Species
2004 2005
7.96ab
3.88b
0.69
0.45
3.33b
6.08a
0.21
0.52
3.00b
2.71b
0.43
0.43 DICK
2004 2005
4.25a
2.42b
0.40
0.31
0.33b
3.75a
0.16
0.31
0.83b
0.71c
0.24
0.19 RWBL 2004
2005
1.75a
0.38a
0.41
0.15
0.04b
0.59a
0.04
0.20
0.13b
0.25a
0.09
0.11 BOBO 2004-
2005 0.41b 0.14 0.22b 0.14 1.09a 0.16
GRSP 2004 2005
0.75b
0.17b
0.22
0.08
2.00a
0.42ab
0.23
0.15
1.13b
0.83a
0.26
0.20 COYE 2004
2005
0.42a
0.25b
0.12
0.09
0.00b
0.58a
0.00
0.15
0.04b
0.00c
0.04
0.00 a Mean abundance from 3 sampling periods each year. b Means in rows with different letters were different (P < 0.05). c DICK = dickcissel, RWBL = red-winged blackbird, BOBO = bobolink, GRSP =
grasshopper sparrow, and COYE = common yellowthroat. d P = P-value for treatment (disked 2003 vs. disked 2004 vs. undisked) effects from a 2-
way analysis of variance. Treatment x year interaction effects occurred in all species
combined (F2, 138 = 16.76, P < 0.001), dickcissels (F2,138 = 47.98, P < 0.001), red-winged
blackbirds (F2,138 = 10.41, P < 0.001), grasshopper sparrows (F2,138 = 5.77, P = 0.004),

65
and common yellowthroats (F2,138 = 10.12, P < 0.001). Means are reported separately for
each year.

66
Table 4. Number of nesting species, number of nests, and nest density (nests/ha) of
grassland birds in treatment and reference fields in Stanton County, Nebraska, 2004-
2005.
Treatment (n = 6)
Reference (n = 6)
Year Mean S.E. Mean S.E. Number of species
2004 2005
5
8
--
--
4 9
--
-- Number of nests
2004 2005
100
106
--
--
12
29
--
-- Nest density (nests/ha)
2004- 2005
2.79 1.10 0.91 0.28

67
Table 5. Nest success probabilitiesa for incubation, nestling, and overall nesting period
for all bird species in treatment and reference fields in Stanton County, Nebraska, 2004-
2005.
a Determined using Mayfield (1975) method. b P = P-value for chi-square analysis of overall nest success probability (Dow 1978).
Treatment (n = 159)
Reference (n = 30)
Pb
Incubation(%)
Nestling(%)
Overall(%)
Incubation(%)
Nestling(%)
Overall(%)
All bird species
47.64 43.29 20.62 52.55 38.57 20.27
0.971

68
Table 6. Nest success probabilitiesa for all bird species, dickcissels, and red-winged
blackbirds in Conservation Reserve Program fields in Stanton County, Nebraska, 2004-
2005.
a Determined using Mayfield (1975) method. b DICK = dickcissel and RWBL = red-winged blackbird. c P = P-value for chi-square analysis of overall nest success probability (Dow 1978).
2004 (n = 93)
2005 (n = 96)
Pc
Speciesb Incubation(%)
Nestling(%)
Overall(%)
Incubation(%)
Nestling(%)
Overall(%)
All bird species
57.77 46.43 26.82 41.32 37.98 15.28 0.053
DICK 64.19 40.36 25.91 31.77 44.64 14.18 0.232
RWBL 52.81 51.12 27.00 41.94 23.52 9.87 0.045

69
Table 7. Vegetation characteristics of successful and unsuccessful nests of all grassland
bird species in Conservation Reserve Program fields in Stanton County, Nebraska, 2004-
2005.
Successful (n = 60)
Unsuccessful (n = 180)
Pa
Vegetation characteristic
Year Mean S.E. Mean S.E. Fate Year
Nest height (cm) 2004 2005
36.06
16.00
4.68
3.96
37.71
34.22
2.76
2.14
0.749
<0.001
Visual obstruction (dm)
2004 2005
6.55
3.84
0.47
0.34
6.79
5.69
0.29
0.20
0.649
<0.001
Litter depth (cm) 2004-2005
1.95 0.24 1.64 0.13 0.036 <0.001
Maximum vegetation height (cm)
2004 2005
100.93
67.12
5.46
3.99
100.44
87.79
2.79
2.20
0.690
<0.001
Forb (%) 2004-2005
34.00 4.47 47.71 2.71 0.001 <0.001
Grass (%) 2004-2005
49.50 3.66 41.96 2.23 0.015 <0.001
Dead material (%) 2004-2005
12.83 2.52 9.21 1.15 0.057 0.010
Bare ground (%) 2004-2005
4.17 1.14 1.57 0.39 0.602 <0.001
a P = P-value for fate (successful vs. unsuccessful) and year (2004 vs. 2005) effects from
2-way analysis of variance. Interaction (fate x year) effects occurred in maximum
vegetation height (F1,236 = 8.19, P = 0.005), nest height (F1,234 = 5.55, P = 0.019), and
visual obstruction (F1,236 = 5.48, P = 0.020). Means are reported separately for each year
for these variables.

70
Table 8. Vegetation characteristics of successful and unsuccessful dickcissel nests in
Conservation Reserve Program fields in Stanton County, Nebraska, 2004-2005.
Successful (n = 20)
Unsuccessful (n = 62)
Pa
Vegetation characteristic
Year Mean S.E. Mean S.E. Fate Year
Nest height (cm) 2004-2005
31.25 3.64 33.31 1.73 0.171 0.002
Visual obstruction (dm)
2004-2005
6.80 0.50 6.46 0.24 0.560 <0.001
Litter depth (cm) 2004-2005
1.31 0.37 1.40 0.24 0.438 <0.001
Maximum vegetation height (cm)
2004 2005
112.62
72.86
6.27
7.23
102.27
89.80
4.12
3.03
0.148
0.043
Forb (%) 2004-2005
60.50 7.27 56.77 4.29 0.751 0.005
Grass (%) 2004-2005
31.00 5.75 37.10 3.77 0.928 0.006
Dead material (%) 2004 2005
0.00
12.86
0.00
7.14
2.31
6.11
1.28
1.34
0.213
0.123
Bare ground (%) 2004-2005
3.00 2.52 1.45 0.64 0.755 0.032
a P = P-value for fate (successful vs. unsuccessful) and year (2004 vs. 2005) effects from
2-way analysis of variance. Interaction (fate x year) effects occurred in maximum
vegetation height (F1,78 = 6.26, P = 0.014) and dead material cover (F1,78 = 4.09, P =
0.047). Means are reported separately for each year for these variables.

71
Table 9. Vegetation characteristics of successful and unsuccessful red-winged blackbird
nests in Conservation Reserve Program fields in Stanton County, Nebraska, 2004-2005.
Successful (n = 18)
Unsuccessful (n = 86)
Pa
Vegetation characteristic
Year Mean S.E. Mean S.E. Fate Year
Nest height (cm) 2004-2005
55.17 4.14 49.37 1.95 0.483 0.257
Visual obstruction (dm)
2004-2005
6.94 0.40 7.04 0.20 0.226 0.003
Litter depth (cm) 2004-2005
1.39 0.40 1.19 0.15 0.031 <0.001
Maximum vegetation height (cm)
2004-2005
111.61 4.21 103.88 1.70 0.903 <0.001
Forb (%) 2004-2005
41.11 6.36 56.98 3.39 0.013 0.041
Grass (%) 2004-2005
53.89 6.67 37.67 3.12 0.007 0.030
Dead material (%) 2004-2005
1.11 0.76 4.42 0.80 0.281 0.078
Bare ground (%) 2004-2005
4.44 2.46 1.41 0.44 0.912 0.005
a P= P-value for fate (successful vs. unsuccessful) and year (2004 vs. 2005) effects from
2-way analysis of variance. No interaction (fate x year) effects occurred.
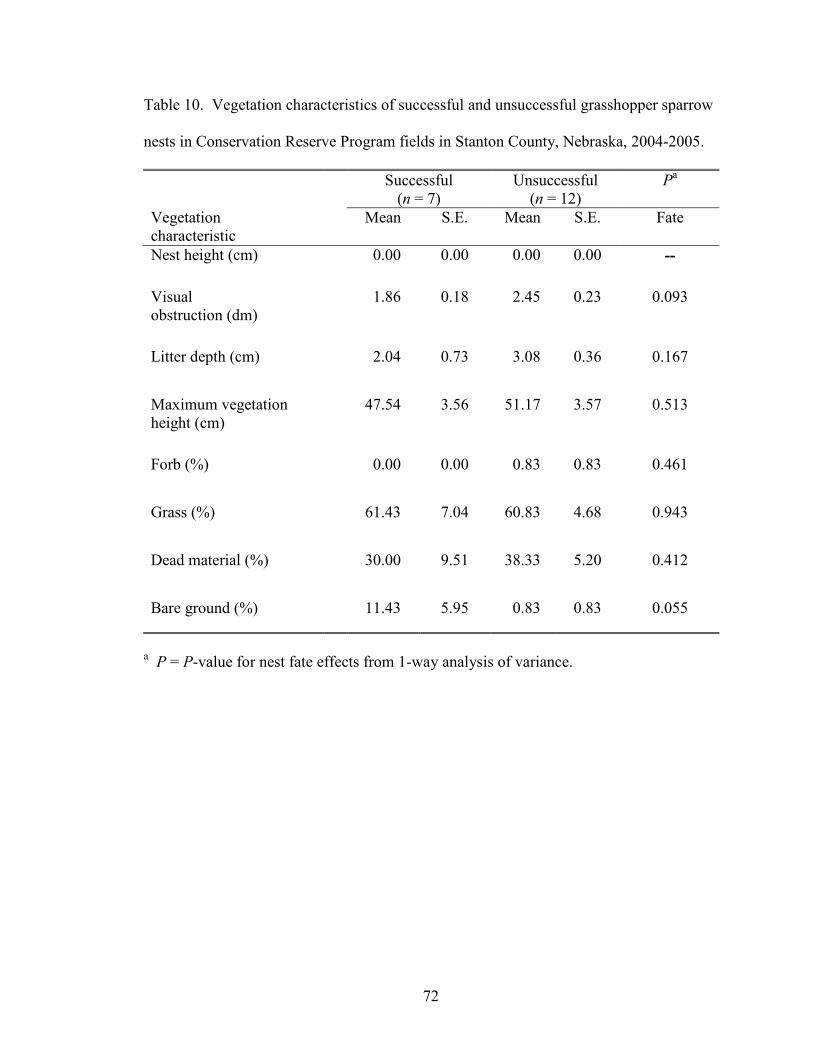
72
Table 10. Vegetation characteristics of successful and unsuccessful grasshopper sparrow
nests in Conservation Reserve Program fields in Stanton County, Nebraska, 2004-2005.
Successful (n = 7)
Unsuccessful (n = 12)
Pa
Vegetation characteristic
Mean S.E. Mean S.E. Fate
Nest height (cm) 0.00 0.00 0.00 0.00 --
Visual obstruction (dm)
1.86 0.18 2.45 0.23 0.093
Litter depth (cm) 2.04 0.73 3.08 0.36 0.167
Maximum vegetation height (cm)
47.54 3.56 51.17 3.57 0.513
Forb (%) 0.00 0.00 0.83 0.83 0.461
Grass (%) 61.43 7.04 60.83 4.68 0.943
Dead material (%) 30.00 9.51 38.33 5.20 0.412
Bare ground (%) 11.43 5.95 0.83 0.83 0.055
a P = P-value for nest fate effects from 1-way analysis of variance.

73
Table 11. Vegetation characteristics of successful and unsuccessful bobolink nests in
Conservation Reserve Program fields in Stanton County, Nebraska, 2004-2005.
Successful (n = 7)
Unsuccessful (n = 8)
Pa
Vegetation characteristic
Mean S.E. Mean S.E. Fate
Nest height (cm) 0.00 0.00 0.00 0.00 -- Visual obstruction (dm)
3.50 0.22 3.16 0.13 0.190
Litter depth (cm) 3.57 0.30 3.94 0.33 0.430
Maximum vegetation height (cm)
56.29 2.99 61.41 3.56 0.298
Forb (%) 0.00 0.00 0.00 0.00 --
Grass (%) 70.00 5.77 65.00 5.67 0.549 Dead material (%) 32.86 7.14 36.25 7.30 0.747
Bare ground (%) 0.00 0.00 2.50 1.64 0.179
a P = P-value for nest fate effects from 1-way analysis of variance.
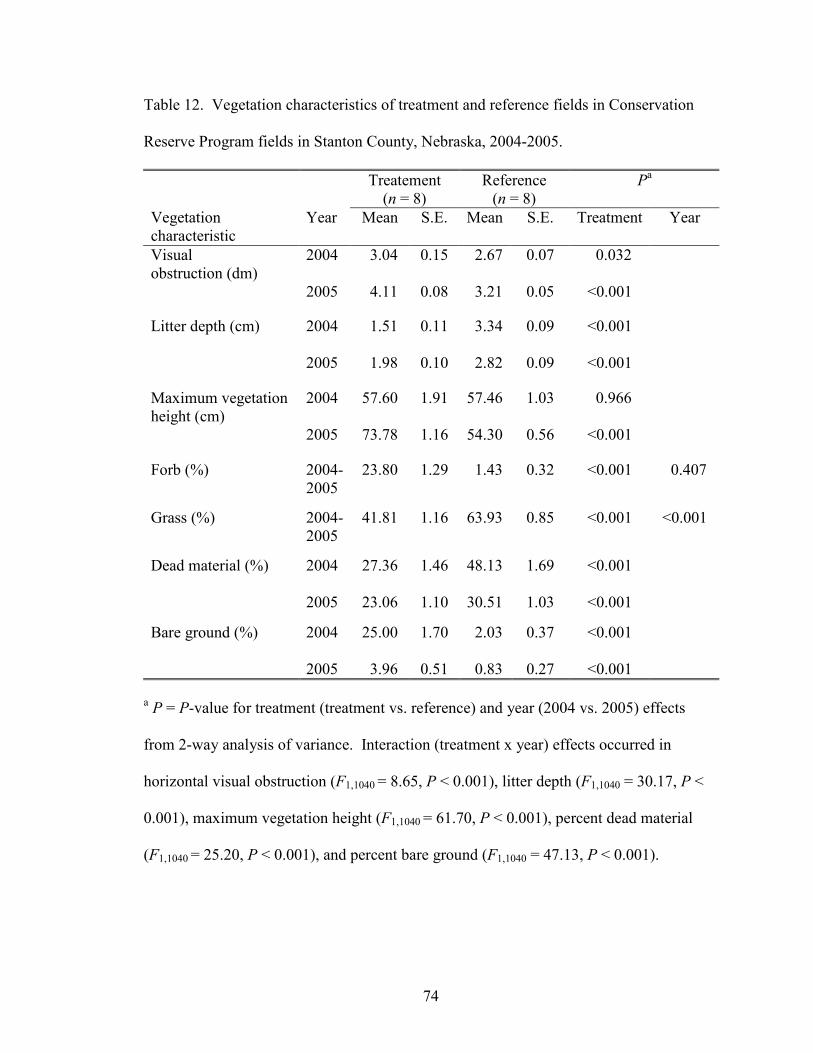
74
Table 12. Vegetation characteristics of treatment and reference fields in Conservation
Reserve Program fields in Stanton County, Nebraska, 2004-2005.
Treatement (n = 8)
Reference (n = 8)
Pa
Vegetation characteristic
Year Mean S.E. Mean S.E. Treatment Year
Visual obstruction (dm)
2004 2005
3.04
4.11
0.15
0.08
2.67
3.21
0.07
0.05
0.032
<0.001
Litter depth (cm) 2004 2005
1.51
1.98
0.11
0.10
3.34
2.82
0.09
0.09
<0.001
<0.001
Maximum vegetation height (cm)
2004 2005
57.60
73.78
1.91
1.16
57.46
54.30
1.03
0.56
0.966
<0.001
Forb (%) 2004-2005
23.80 1.29 1.43 0.32 <0.001 0.407
Grass (%) 2004-2005
41.81 1.16 63.93 0.85 <0.001 <0.001
Dead material (%) 2004 2005
27.36
23.06
1.46
1.10
48.13
30.51
1.69
1.03
<0.001
<0.001
Bare ground (%) 2004 2005
25.00
3.96
1.70
0.51
2.03
0.83
0.37
0.27
<0.001
<0.001
a P = P-value for treatment (treatment vs. reference) and year (2004 vs. 2005) effects
from 2-way analysis of variance. Interaction (treatment x year) effects occurred in
horizontal visual obstruction (F1,1040 = 8.65, P < 0.001), litter depth (F1,1040 = 30.17, P <
0.001), maximum vegetation height (F1,1040 = 61.70, P < 0.001), percent dead material
(F1,1040 = 25.20, P < 0.001), and percent bare ground (F1,1040 = 47.13, P < 0.001).

75
Table 13. Vegetation characteristics of 3 portions of Conservation Reserve Program
fields managed by disking/interseeding in Stanton County, Nebraska, 2004-2005.
Disked-2003 (n = 8)
Disked-2004 (n = 8)
Undisked (n = 8)
Vegetation characteristics
Year Mean S.E. Mean S.E. Mean S.E.
Visual obstruction (dm)
2004 2005
5.44aa
4.70a
0.26
0.13
0.50c
4.31b
0.08
0.17
3.17b
3.32c
0.10
0.09
Litter depth (cm)
2004 2005
0.34b
2.33b
0.05
0.19
0.39b
0.71c
0.06
0.10
3.80a
2.89a
0.12
0.15
Maximum vegetation height (cm)
2004 2005
90.31a
81.79a
2.38
1.98
23.96c
80.72a
1.49
1.85
58.54b
58.83b
1.42
1.17
Forb (%) 2004 2005
58.44a
17.29b
2.96
1.98
16.15b
49.27a
2.36
3.20
0.42c
1.25c
0.25
0.60 Grass (%) 2004
2005
21.88b
59.90a
2.11
2.39
14.90c
29.27b
2.01
2.09
62.50a
62.40a
1.40
1.64 Dead material (%)
2004 2005
11.04c
19.48b
1.47
1.61
22.92b
14.48c
2.24
1.77
48.13a
35.21a
2.17
1.68 Bare ground (%)
2004 2005
19.48b
3.33b
1.70
0.78
53.54a
7.40a
2.88
1.16
1.98c
1.15c
0.72
0.44 a Means in rows with different letters were different (P < 0.05). b P = P-value for treatment (disked 2003 vs. disked 2004 vs. undisked) effects from a 2-
way analysis of variance. Treatment x year interaction effects occurred in all analyses
(horizontal visual obstruction [F2,570 = 104.12, P < 0.001]; litter depth [F2,570 = 81.33, P <
0.001]; maximum vegetation height [F2,570 = 170.57, P < 0.001]; percent forb [F2,570 =

76
128.05, P < 0.001], grass [F2,570 = 46.53, P < 0.001], dead material [F2,570 = 18.59, P <
0.001], and bare ground [F2,570 = 56.38, P < 0.001] cover).

77
Table 14. Logistic regression models for vegetation variables that best predicted
grassland bird presence in treatment fields in Stanton County, Nebraska, 2004-2005.
Variables were selected from a set of vegetation variables using a backward-elimination
routine.
Speciesa Fitted logistic modelbc Pd
DICK -3.59 + 0.07 (forb) + 1.13 (vo)
0.313
RWBL -3.94 + 0.04 (max ht)
0.837
GRSP 2.78 + 0.04 (forb) + 0.41 (litter) – 0.07 (max ht)
0.409
BOBO -1.22 + 0.04 (dead) - 0.08 (bare)
0.816
COYE -6.13 + 0.02 (forb) + 0.05 (max ht)
0.621
WEME 0.98 – 0.03 (forb) – 0.07 (grass)
0.587
SEWR -6.65 + 0.05 (forb) + 1.00 (litter)
0.944
HESP No Significant Model
--
a DICK = dickcissel, RWBL = red-winged blackbird, BOBO = bobolink, GRSP =
grasshopper sparrow, COYE = common yellowthroat, SEWR = sedge wren, WEME =
western meadowlark, HESP = Henslow’s sparrow. b P(presence) = 1/ (1 + exp{ - [bo + b1(x)]}) where P(presence) was the probability that a
bird species was present, bo and b1 were intercept and slope coefficients, and x was the
predictor variable (vegetation variable), and Absence(x) = 1 – Presence(x). c forb = forb cover, grass = grass cover, dead = dead material cover, bare = bare ground,
vo = horizontal visual obstruction, max ht = maximum vegetation height, and litter =
litter depth. d P = P-value for Hosmer and Lemeshow goodness-of-fit test of overall model.
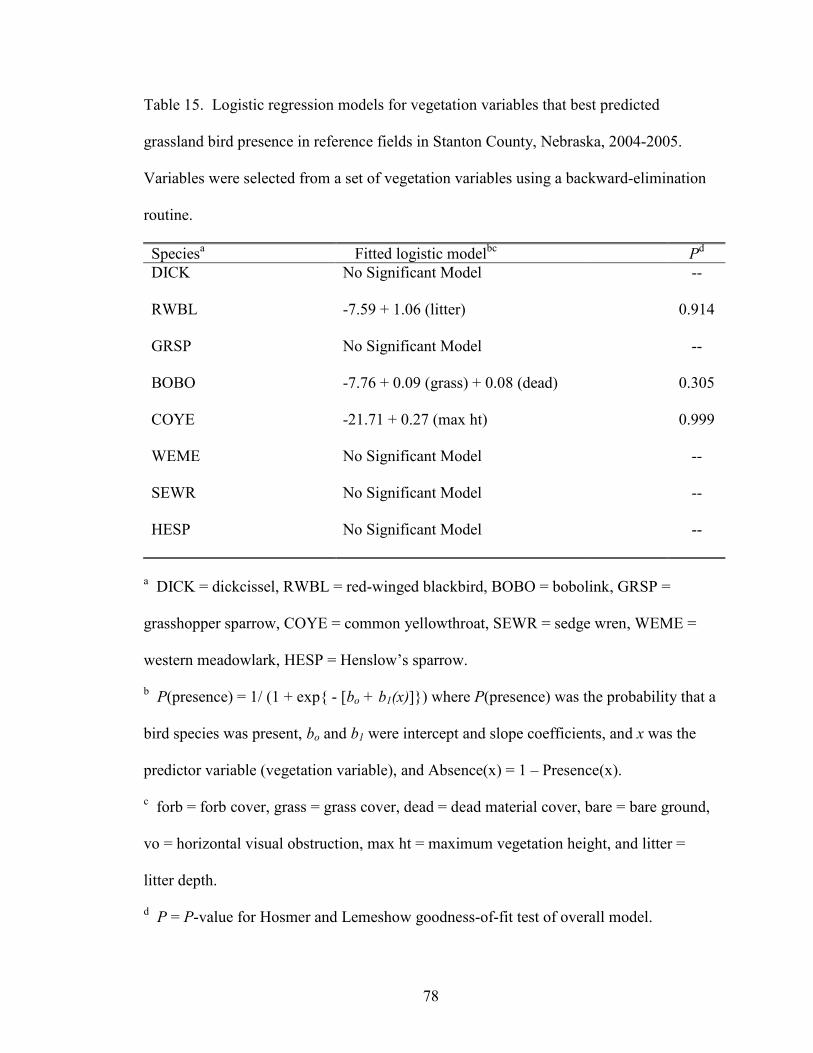
78
Table 15. Logistic regression models for vegetation variables that best predicted
grassland bird presence in reference fields in Stanton County, Nebraska, 2004-2005.
Variables were selected from a set of vegetation variables using a backward-elimination
routine.
Speciesa Fitted logistic modelbc Pd
DICK No Significant Model
--
RWBL -7.59 + 1.06 (litter)
0.914
GRSP No Significant Model
--
BOBO -7.76 + 0.09 (grass) + 0.08 (dead)
0.305
COYE -21.71 + 0.27 (max ht)
0.999
WEME No Significant Model
--
SEWR No Significant Model
--
HESP No Significant Model
--
a DICK = dickcissel, RWBL = red-winged blackbird, BOBO = bobolink, GRSP =
grasshopper sparrow, COYE = common yellowthroat, SEWR = sedge wren, WEME =
western meadowlark, HESP = Henslow’s sparrow. b P(presence) = 1/ (1 + exp{ - [bo + b1(x)]}) where P(presence) was the probability that a
bird species was present, bo and b1 were intercept and slope coefficients, and x was the
predictor variable (vegetation variable), and Absence(x) = 1 – Presence(x). c forb = forb cover, grass = grass cover, dead = dead material cover, bare = bare ground,
vo = horizontal visual obstruction, max ht = maximum vegetation height, and litter =
litter depth. d P = P-value for Hosmer and Lemeshow goodness-of-fit test of overall model.

79
Figure 1. Location of study area (represented by the black box) in Stanton County,
Nebraska.

80
NEBRASKA

81
Figure 2. Distance frequency histograms for all species combined, dickcissels,
grasshopper sparrows, and bobolinks in treatment and reference fields in Stanton County,
Nebraska, 2005.
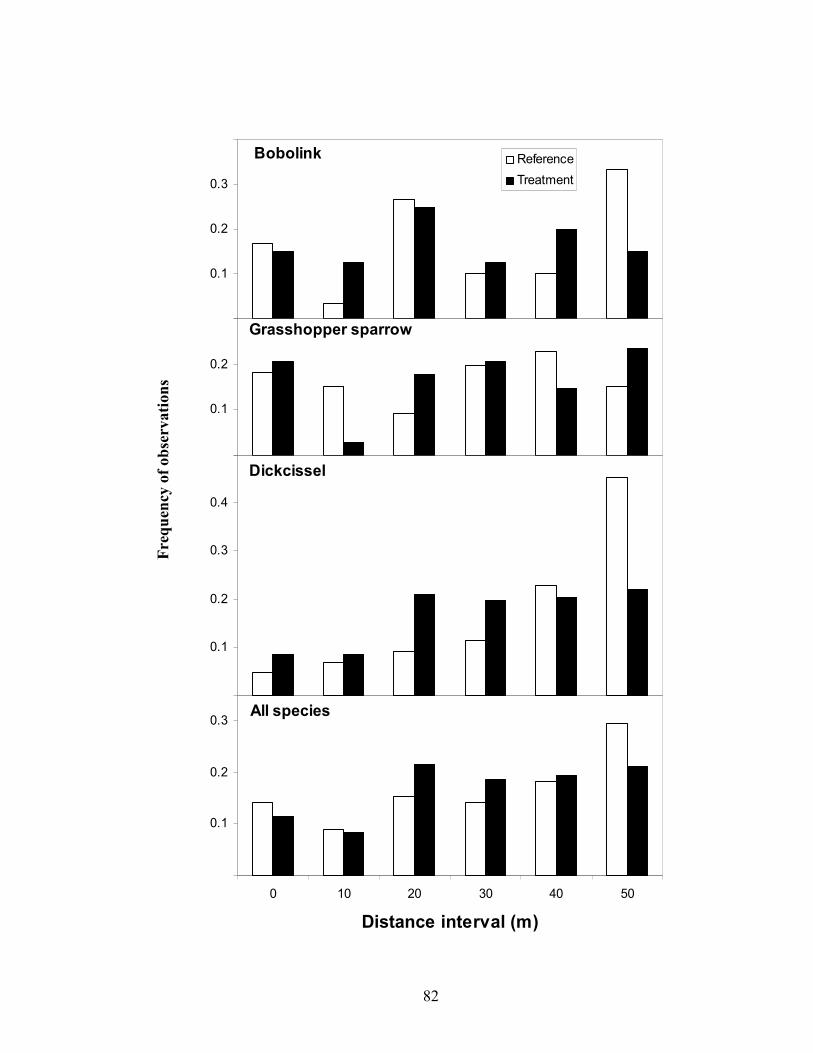
82
Freq
uenc
yofo
bserv
ation
s
All species
0
0.1
0.2
0.3
0 10 20 30 40 50
Distance interval (m)
Bobolink
0
0.1
0.2
0.3
0.4ReferenceTreatment
Grasshopper sparrow
0
0.1
0.2
0.3
Dickcissel
0
0.1
0.2
0.3
0.4
0.5

83
Figure 3. Mean (+ S.E.) nest density (nests/ha) for all species combined for 0- (n = 2), 1-
(n = 4), and 2-year (n = 2) post treatment and no treatment (n = 11) nest search plots in
Conservation Reserve Program fields in Stanton County, Nebraska, 2004 and 2005.
Means with different letters are significantly different (P < 0.05).

84
0123456789
10
0 1 2 No treatmentYears post treatment
Nest
s/ha
A
AB
BB

85
APPENDIX A
Bird species observed in Conservation Reserve Program fields in Stanton County,
Nebraska, 2004-2005.
Species
Treatment Reference
Grasshopper sparrow (Ammodramus savannarum)
X X
Dickcissel (Spiza Americana) X X
Bobolink (Dolichonyx oryzivorus) X X
Common yellowthroat (Geothlypis trichas) X X
Red-winged blackbird (Agelaius phoeniceus) X X
Western meadowlark (Sturnella neglecta) X X
Eastern kingbird (Tyrannus tyrannus) X X
Sedge wren (Cistothorus platensis) X X
Henslow’s sparrow (Ammodramus henslowii) X X
Barn swallow (Hirundo rustica) X X
Tree swallow (Tachycineta bicolor) X X
Northern rough-winged swallow (Stelgidopteryx serripennis)
X X
Mourning dove (Zenaida macroura) X X
Common grackle (Quiscalus quiscula) X X
Mallard (Anas platyrhychos) X XAmerican goldfinch (Carduelis tristis) X X
Field sparrow (Spizella pusilla) X X
Killdeer (Charadrius vociferous) X
House sparrow (Passer domesticus) X

86
Species
Treatment Reference
Blue-winged teal (Anus discors) X X
Brown-headed cowbird (Molothrus ater) X X
Ring-necked pheasant (Phasianus colchicus) X X
European starling (Sturnus vulgaris) X X
Northern flicker (Colaptes auratus) X X
Northern harrier (Circus cyaneus) X X
Orchard oriole (Icterus spurious) X
American robin (Turdus migratorius) X
Cedar waxwing (Bombycilla cedrorum) X
Eastern meadowlark (Sturnella magna) X
Baltimore oriole (Icterus galbula) X

87
APPENDIX B
Percentage of each plant species used as a primary vegetationa support for nest placement
by the 4 most common nesting grassland bird species in Conservation Reserve Program
fields in Stanton County, Nebraska, 2004-2005.
Plant Species Dickcissel (n = 89)
Red-winged blackbird (n = 106)
Grasshopper sparrow (n = 19)
Bobolink (n = 15)
Yellow sweetclover (Melilotus officinalis)
36 52
Alfalfa (Medicago sativa)
17 9
Red clover (Trifolium pratense)
11 4
Mustard spp. (Brassica spp.)
2 9
Smooth brome (Bromus inermis)
21 14 84 100
Thistle spp. (Carduus spp.) or (Cirsium spp.)
9 4
Curly doc (Rumex crispus)
5
Kentucky bluegrass (Poa pratensis)
5
Orchard grass (Dactylis glomerata)
1 11
Nettle spp. (Urtica spp.)
1
Sunflower spp. (Helianthus spp.)
1
Milkweed spp. (Asclepias spp.)
1
Wild plum (Prunus americana)
1

88
Plant Species Dickcissel
Red-winged blackbird
Grasshopper sparrow
Bobolink
Chinese elm (Ulmus parvifolia)
1
a In addition to the primary plant species used for nest placement, 36% of red-winged
blackbirds and 39% of dickcissels used a secondary plant species to help support the nest.
Secondary plant species were most commonly alfalfa (25%), smooth brome (23%),
yellow sweetclover (22%), and red clover (18%).

89
APPENDIX C
Relative abundance and biomass of invertebrates in disked/interseeded and undisked
portions of Conservation Reserve Program fields in Stanton County, Nebraska, 2004.
Disked-2003 (n = 8)
Disked-2004 (n = 8)
Undisked (n = 24)
Pa
Mean S.E. Mean S.E. Mean S.E. Period TreatmentAbundance 176.00ab 63.38 192.88a 50.94 79.58b 21.07 0.11 0.05 Biomass 1.29ab 0.38 1.53a 0.59 0.64b 0.09 0.02 0.03
a P = P-value for period (1 vs. 2) and treatment (disked-2003 vs. disked-2004 vs.
undisked) effects from 2-way analysis of variance. No interaction (period x treatment)
effects occurred. b Means in rows with different letters were different (P < 0.05).

VITA
Lucas Paul Negus
Candidate for the Degree of
Master of Science
Thesis: GRASSLAND BIRD RESPONSE TO DISKING/INTERSEEDING OF LEGUMES IN CONSERVATION RESERVE PROGRAM LANDS IN NORTHEAST NEBRASKA
Major Field: Wildlife and Fisheries Ecology Biographical:
Personal Data: Born in Tilden, Nebraska, 1980 Education: Elkhorn Valley High School, 1998; Bachelors of Science from
University of Nebraska-Kearney in May, 2002; Masters of Science candidate at Oklahoma State University in May, 2006.
Experience: Fish and Wildlife Biologist, Nebraska Game and Parks
Commission, 2002-2003; Conservation Technician, Nebraska Game and Parks Commission, 1999-2002; Work Study, Nebraska Game and Parks Commission, 1998-2002; Family farm employee.
Professional Memberships: Member of National chapter, Nebraska, and
Oklahoma state chapters of The Wildlife Society, Pheasants Forever, Oklahoma State University Zoology Graduate Student Society, Oklahoma State University student chapters of The Wildlife Society and American Fisheries Society.

Name: Lucas Paul Negus Date of Degree: May, 2006 Institution: Oklahoma State University Location: Stillwater, Oklahoma Title of Study: GRASSLAND BIRD RESPONSE TO DISKING/INTERSEEDING OF
LEGUMES IN CONSERVATION RESERVE PROGRAM LANDS IN NORTHEAST NEBRASKA
Pages in Study: 89 Candidate for the Degree of Master of Science
Major Field: Wildlife and Fisheries Ecology Scope and Method of Study: Throughout the Midwest, grassland birds have been
declining faster than any other group of birds, with the main cause for these declines being the extensive loss of native prairies. During the last 25 years, surrogate grasslands, such as Conservation Reserve Program (CRP) lands, have become increasingly important as an alternative habitat for grassland birds. However, many CRP grasslands that once provided excellent habitat are now dominated by monoculture stands of grass that have reduced wildlife benefits. In summer 2000, the Nebraska Game and Parks Commission and Pheasants Forever partnered with the United States Department of Agriculture’s Farm Service Agency and Natural Resource Conservation Service to initiate a program that promotes disking and interseeding legumes in CRP to improve nesting and brood-rearing habitat for ring-necked pheasants (Phasianus colchicus). Disking and interseeding will also likely affect grassland birds, yet there has been no research on their response to this practice. The objectives of this study were to determine grassland bird abundance and nest-productivity in disked and interseeded CRP, evaluate vegetative responses to disking and interseeding, and provide management recommendations that may influence future farm bill policy regarding CRP grasslands.
Findings and Conclusions: Findings indicate experimental fields had higher overall avian
species richness, abundance, and diversity than reference fields. Dickcissels, red-winged blackbirds, and common yellowthroats were more abundant in treatment fields while bobolinks and Henslow’s sparrows were more abundant in reference fields. Nest densities in experimental fields were nearly 3 times greater than nest densities in reference fields. To increase grassland bird diversity and abundance, I recommend establishing an annual rotation of disking/interseeding to create a mosaic of vegetation types, while leaving portions of CRP fields undisked to accommodate species that don’t respond positively to management.
ADVISER’S APPROVAL: Craig A. Davis