gp -c r -...
TRANSCRIPT

14 Jan 2003 15:1 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC10.1146/annurev.physiol.65.092101.142611
Annu. Rev. Physiol. 2003. 65:851–79doi: 10.1146/annurev.physiol.65.092101.142611
Copyright c© 2003 by Annual Reviews. All rights reservedFirst published online as a Review in Advance on November 20, 2002
G PROTEIN-COUPLED RECEPTOR
RHODOPSIN: A Prospectus
SÃlawomir Filipek1, Ronald E. Stenkamp2,7,David C. Teller3,7, and Krzysztof Palczewski4,5,61Department of Chemistry, University of Warsaw, 1 Pasteur St, PL-02093 Warsaw, Poland;2Departments of Biological Structure,3Biochemistry,4Ophthalmology,5Chemistry,6Pharmacology, and7Biomolecular Structure Center, University of Washington, Seattle,Washington 98195; e-mail: [email protected]
Key Words crystal structure, phototransduction, vitamin A, transmembraneprotein, vision, signal transduction
■ Abstract Rhodopsin is a retinal photoreceptor protein of bipartite structure con-sisting of the transmembrane protein opsin and a light-sensitive chromophore 11-cis-retinal, linked to opsin via a protonated Schiff base. Studies on rhodopsin have unveiledmany structural and functional features that are common to a large and pharmaco-logically important group of proteins from the G protein-coupled receptor (GPCR)superfamily, of which rhodopsin is the best-studied member. In this work, we focuson structural features of rhodopsin as revealed by many biochemical and structuralinvestigations. In particular, the high-resolution structure of bovine rhodopsin pro-vides a template for understanding how GPCRs work. We describe the sensitivity andcomplexity of rhodopsin that lead to its important role in vision.
INTRODUCTION: HISTORICAL PERSPECTIVE
The visual pigment rhodopsin was originally extracted from bovine retina usingbile salts by Kuhne (1). He found that the color of rhodopsin resulted from a chro-mophore different from xanthophylls (egg yolk) orβ-carotene (corpus luteum).It faded upon exposure to light, heating, or organic solvents and could be saltedout from the extract by ammonium sulfate. Regeneration of bleached rhodopsinrequired interaction of the retina with adjacent retinal pigment epithelium cells(RPE). The need for vitamin A in vision triggered an interest in retinoids as apotential source of the chromophore. Wald provided evidence that visual pigmentis composed of a protein conjugated with an unidentified retinoid (2). Vitamin A(all-trans-retinol) is a precursor of this retinoid and a product of visual pigmentphotobleaching. Successive work demonstrated that bleaching and regeneration ofthe visual pigment involves a cycle of stereoisomerization of all-trans-retinal/all-trans-retinol. The reddish color of rhodopsin results from the protonated Schiff
0066-4278/03/0315-0851$14.00 851

14 Jan 2003 15:1 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
852 FILIPEK ET AL.
base linkage of 11-cis-retinal to the membrane protein opsin (3, 4). Therefore, ametabolic cycle of production and recycling of the photoisomerized visual chro-mophore is essential for our vision. Bleaching of rhodopsin introduces a succes-sion of quasi-stable intermediates that within milliseconds reach an equilibriumof metarhodopsin I (Meta I) and metarhodopsin II (Meta II) (5). Meta II is fa-vorable in higher temperatures and pH, salt, and glycerol, and its formation isaccompanied by a large increase in entropy. Subsequent work focused on bovinerhodopsin, where the naturally high abundance of this interesting protein was uti-lized in a number of biochemical and biophysical approaches [early work reviewedin (6)].
The first protein amino acid sequence of the G protein-coupled receptor (GPCR),opsin, was determined in the laboratories of Hargrave et al. and Ovchinnikov (7, 8),and the first models of the seven-transmembrane helix topology, based partly onproteolysis, antibody-recognition, and chemical and posttranslational (glycosy-lation) modifications, were introduced (9). The overall topology of rhodopsin issimilar to another retinylidene-binding protein, bacteriorhodopsin (8). In parallelwith the improved understanding of the properties of rhodopsin, functional studiesprovided a solid basis for the way the light signal is translated into biochemi-cal reactions, a process termed phototransduction. Fung & Stryer demonstratedthat light-activated rhodopsin catalyzes the nucleotide exchange of GDP to GTPin an interacting protein called transducin, a G protein, which in turn activatesa photoreceptor cGMP-specific phosphodiesterase (10). Hundreds of transducinmolecules are activated per photoactivated rhodopsin in this first amplification stepduring phototransduction (11). Deprotonation of Meta II’s Schiff base between thechromophore and opsin appeared to be critical for the activating property of light-exposed rhodopsin (12). A fortuitous discovery, made in the early 1970s, thatrhodopsin is phosphorylated (13–15), has been connected with observations thatATP mediates quenching of phosphodiesterase activity (16, 17). The responsiblerhodopsin kinase has been purified (18) and cloned (19). For complete quenching ofthe light-activated phosphorylated rhodopsin activity, specific binding of a 48-kDaprotein is critical (20). Arrestin-like molecules play a key role in the signaling andcellular processes of a large number of agonist-stimulated GPCRs (21).
Nathans & Hogness were the first to clone opsin from the bovine cDNA library(22). Subsequently, the generated bovine DNA sequence was essential in the de-termination of the bovine opsin genomic structure (22); the determination of thecDNA and genomic structures of human opsin and blue, green, and red visualpigments (23, 24); the cloning of the first invertebrate rhodopsin and its gene fromDrosophila(25); and the total chemical synthesis of cDNA for bovine rhodopsin(26).
The crystal structure of rhodopsin isolated from bovine retinas has been solved(27) and refined (28), providing significant insight into the function of this im-portant molecule. In this review, we describe the major structural properties ofrhodopsin. As with any rich subject, not all of the nuances of rhodopsin structureand function can be covered here. Therefore, the authors encourage readers to

14 Jan 2003 15:1 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
G PROTEIN-COUPLED RECEPTOR RHODOPSIN 853
peruse other reviews dedicated to rhodopsin and different aspects of phototrans-duction (6, 21, 29–36).
RHODOPSIN
Expression
Retinal rod cells are specialized neurons that function in capturing photons andcommunicating with secondary neurons about the presence of light. These eventsare initiated by conformational changes in the light-sensitive pigment, rhodopsin,followed by a biochemical cascade of reactions, termed phototransduction (30, 31).In mammalian retina there are∼108 photoreceptor cells. Rods are highly differ-entiated cells with outer segments (ROS) containing all components necessaryfor phototransduction. ROS are composed of stacks of 1000–2000 independentdisk membranes surrounded by plasma membrane (Figure 1, see SupplementalMaterials: Follow the Supplemental Material link on the Annual Reviews home-page at http://annualreviews.org/). The main component of the disk membranes isrhodopsin (>90% of the membrane’s proteins). Opsin, the protein component ofrhodopsin, is specifically expressed in retinal rod photoreceptors and in some cellsof the pineal gland (37). In adult mice, 0.06% of the total retinal RNA encodesfor opsin (photoreceptors constitute∼80–90% of all cells in the retina) (38). Thistranslates into the highest expression of any GPCR. About 0.5–1 mg of rhodopsincan be isolated from one bovine retina.
Purification
The absorption spectrum of rhodopsin in detergent solutions displays two maximain the UV-Vis region: the protein band around 280 nm and the chromophore-relatedpeak at 498 nm. The ratio of the absorption A280 nm/A498 nm for pure rhodopsindevoid of opsin is∼1.6, with an extinction coefficient of 42,000 cm−1 M−1 at498 nm. Rhodopsin is stable at room temperature for days when extracted intomany detergents, including Chaps, dodecyl-β-maltoside, or Tween 80. The mostfrequent source of native rhodopsin is bovine eyes. Native rhodopsin is initiallypre-purified by isolation of ROS from the retina using a sucrose gradient method(50–70% rhodopsin content in ROS) [reviewed in (6)]. The purification methodsrequire separation of rhodopsin from contaminating proteins and from bleachedopsin. All methods require detergent for rhodopsin solubilization. The first methodtakes advantage of rhodopsin glycosylation and employs concanavalin A affinitychromatography (39). A large percent of opsin binds irreversibly to the resin.Concanavalin A chromatography is scalable, and large amounts of material can beprepared, although the purified rhodopsin is contaminated with other glycoproteinsand with concanavalin A, which leaches from the resin. The second method useshydroxyapatite chromatography and yields only partially purified protein (40).Rhodopsin is also efficiently purified by immunoaffinity chromatography using

14 Jan 2003 15:1 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
854 FILIPEK ET AL.
Molday’s 1D4 monoclonal antibody, which recognizes seven to nine C-terminalamino acids of bovine rhodopsin. This method is particularly useful for heterolo-gously expressed rhodopsin and other visual pigments (26, 41). The fourth methodtakes advantage of highly enriched rhodopsin in ROS and the instability of opsinand contaminating protein during prolonged incubation in mild detergent in thepresence of divalent metal ions (43). It is an effective and scalable method thatdoes not use any chromatographic columns. This method also enriches rhodopsinpreparations with native ROS lipids and produces highly concentrated rhodopsinwith controllable concentrations of detergent suitable for crystallization studies.This method is not applicable to heterologous systems due to the low expressionlevels of rhodopsin.
Composition
Rhodopsin is composed of a transmembrane apoprotein, opsin, and 11-cis-retinalbound to the protein through a Schiff base linkage to a lysine side-chain. Bovineopsin (Swiss-Prot: P02699) is composed of 348 amino acids with a molecular massof 39,007. The total molecular mass increases to 42,002 when posttranslationalmodifications are included (palmitoylation, acetylation of N terminus, and glyco-sylation). The most prevalent amino acids are Phe (8.9%), Val (8.9), Ala (8.3), andLeu (8.0), suggesting a major hydrophobic character for this protein.
Rhodopsin is extensively modified by posttranslational modifications. The chro-mophore is attached through a protonated Schiff base to Lys296(44). The N-terminalMet is acetylated (45), as is found frequently for other eukaryotic proteins at theirinitiation Met residue. Using ex vivo [3H]-labeling, it was shown that opsin ispalmitoylated (46), a modification that is frequently observed among GPCRs.Two Cys residues, Cys322 and Cys323 located at the C terminus, are palmitoylated(47). Two Cys residues from helix III and E III (the loop connecting helices IVand V) are cross-linked by a disulfide bond (48). Two (Man)3(GlcNAc)3 groupsmodify opsin through an asparagine-linkage (Figure 2, see Supplemental Materi-als: Follow the Supplemental Material link on the Annual Reviews homepage athttp://annualreviews.org/) at the N terminus (Asn2 and Asn15) (49). Glycosyla-tion is not homogenous and other, but minor, compositions of the carbohydratemoieties were identified (50). Rhodopsin undergoes a light-dependent phospho-rylation on six to seven Ser/Thr residues at the C-terminal end (51). In vivo, thereare three sites, Ser336, Ser338, and Ser343, that are phosphorylated by direct andquantitative methods after 20–40% bleaching of the protein (52, 53). The phos-phorylation processes, at lower bleaches such as 100–10,000 photons/s per rod,are not yet accessible to currently available technologies. At these bleach levels,our rods are functional before they saturate at more intense light. Heterogeneityand multiple rhodopsin phosphorylation have been observed in vitro [reviewed in(54)].
Rhodopsin, when isolated from ROS, contains variable amounts of tightlybound phospholipids. These lipids possibly stabilize rhodopsin and coat the

14 Jan 2003 15:1 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
G PROTEIN-COUPLED RECEPTOR RHODOPSIN 855
hydrophobic transmembrane regions of the protein. We found, using radioactivephospholipids, that they can be removed completely only under very harsh con-ditions, such as 1% SDS in formic acid and trifluoroethanol (1:1) (X. Zhou &K. Palczewski, unpublished data). It is reasonable to speculate that the integrityof rhodopsin and other membrane proteins may be lost by removal of all tightlyassociated lipids.
Regeneration and Photobleaching Pathway
The key reaction of visual excitation is the ultra-fast (femtoseconds) photochem-ical reaction of rhodopsin’s chromophore, 11-cis-retinal forming all-trans-retinal(55). From a theoretical organic chemistry point of view, this isomerization mostlikely involves the “Hula-Twist” mechanism that preserves, at first approximation,the positions of theβ-ionone ring and the Schiff base (56). A conventional “one-bond-flip” mechanism would predict a large rotation of theβ-ionone ring withinthe rhodopsin molecules (Figure 3, see Supplemental Materials: Follow the Sup-plemental Material link on the Annual Reviews homepage at http://annualreviews.org/). In milliseconds, the signaling Meta II is established and this catalyt-ically active form of rhodopsin binds and activates transducin (T, Gt).
The photoisomerization of 11-cis-retinylidene to all-trans-retinylidene triggersconformational changes of opsin through multiple intermediates, such as photo-rhodopsin, bathorhodopsin, lumirhodopsin, Meta I, Meta II, and Meta III, beforethe chromophore hydrolyzes and leaves the binding pocket (34). These interme-diates are short-lived, but can be trapped by low temperature and distinguishedby specific absorption maxima in the range of visible light (57). The final steps,namely hydrolysis of all-trans-retinylidene and its relation to Meta III, are poorlyunderstood at the mechanistic level. The hydrolysis of the Schiff base appears tobe the rate-limiting step in the release of the chromophore (58). In addition, twoforms of Meta II were identified, Meta IIa and Meta IIb, that differ in the protona-tion state at the cytoplasmic surface, whereas the Schiff base is deprotonated witha characteristicλmax at 380 nm (59). The transducin-activating form is Meta IIb(34).
Free all-trans-retinal also binds to opsin and forms partially active receptorstoward activation of transducin [reviewed in (60)]. This activity is enhanced com-pared with the activity of free opsin. From the crystallographic model, a poten-tial binding site for the retinal has been identified in the hydrophobic domain ofrhodopsin (D.C. Teller, unpublished data). For 11-cis-retinal, binding to this sitecould be the first step in the regeneration of rhodopsin, before the chromophorebinds non-covalently to the retinylidene cavity and prior to stable formation ofthe Schiff base (61). In the Meta II-transducin complex, all-trans-retinylidene canbe photoisomerized back to 11-cis-retinylidene without transducin dissociation,suggesting that the G protein imposes conformational changes on the rhodopsinsurface that cannot be reversed even upon photoisomerization (62). However,when Meta II is photolyzed by blue light, a product is formed that has absorption

14 Jan 2003 15:1 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
856 FILIPEK ET AL.
properties similar to that of rhodopsin withλmax at 500 nm, but with all-trans-retinal bound (63). These data indicate that all-trans-retinylidene can be bound inthe transmembrane segment of rhodopsin in two distinct conformations: one thatactivates the opsin moiety and another that inactivates it.
Mutagenesis
Availability of rhodopsin sequences and development of molecular biology tech-niques have had tremendous impacts in probing rhodopsin structure using muta-genesis and biochemical approaches. Findings from this research are discussed inthe subsequent paragraphs, where they strengthen observations and interpretationof rhodopsin’s structure and function.
A general method for rhodopsin expression and purification from monkey (orhuman) kidney cells (64), insect cells (65), and different yeast strains (66, 67)was introduced. In particular, we benefited from these studies in understandingthe spectral tuning of rhodopsin. Retinals in their Schiff base forms absorb in thenear ultraviolet range (360–380 nm, compared with∼320 nm for retinols). Thepredicted counterion for the protonated Schiff base has been identified as Glu113,which is highly conserved among all known vertebrate visual pigments (68–70).The addition of a proton to the Schiff base, together with critical placement ofintramolecular negative charges, results in a bathochromic shift in the absorptionspectrum to that characteristic of native rhodopsin (500 nm). The counterion inthe hydrophobic environment causes a∼sevenfold increase in the pKa of theprotonated Schiff base. Therefore, rhodopsin in its native environment will haveits chromophore in the protonated Schiff base linkage (29).
Glycosylation was shown to be dispensable for the proper folding and func-tion of rhodopsin. Mutant rhodopsins lacking Asn15-glycosylation exhibited poorfolding and were defective in transport to the cell surface. They were also poortransducin activators, perhaps owing to their intrinsic instability (71). These studieshave not yet been extended to mouse animal models.
Ridge et al. pioneered studies of rhodopsin using expressed polypeptide frag-ments in COS cells. Splitting rhodopsin in the second and third cytoplasmic loopsled to production of two-fragment rhodopsins that show properties similar to thewild-type (72, 73). These studies, and earlier limited proteolysis studies [reviewedin (6)], revealed several important aspects of rhodopsin function. For example,connecting subsequent loops II and III have only a small stabilizing effect onrhodopsin, whereas the chromophore is critical for the stabilization of the tertiarystructure. It appears that helices I–III and V–VII, with helix IV connected to eitherof these two fragments, form active rhodopsin, suggesting the existence of twoindependent intradomain interactions. These data are in agreement with anotherset of experiments. Rhodopsin retains its spectrum after extensive proteolysis ofits exposed loops by pronase in native membranes, or in detergent solutions, albeit,digested rhodopsin is more temperature-sensitive to denaturation (K. Palczewski,unpublished data).

21 Jan 2003 12:46 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
G PROTEIN-COUPLED RECEPTOR RHODOPSIN 857
CRYSTAL STRUCTURE OF RHODOPSIN
Crystallization and Data Analysis
Rhodopsin was selectively extracted from highly purified bovine ROS using a milddetergent (such as nonyl-β-glucoside) and high concentrations of divalent metalions (for example, 80 mM Zn2+) in the absence of Cl− ions. This method, devel-oped by a postdoctoral fellow, T. Okada, therefore does not involve any columnchromatography and maintains high concentrations of endogenous lipids in theextract (43). Importantly, pre-incubation of ROS at room temperature in detergentcauses opsin and other membranous proteins to precipitate and be efficiently sep-arated from rhodopsin. Opsin is present up to 15% of total opsin and rhodopsinin cow eyes obtained from a local slaughterhouse. The purified rhodopsin had anA280 nm/A500 nm ratio of 1.6, close to the theoretical minimum of 1.5, suggestinghigh purity of the protein and a lack of bleached rhodopsin, opsin (43, 74). Va-por diffusion crystallization techniques were used to crystallize rhodopsin, usingammonium sulfate as the precipitant and heptane-1,2,3-triol as an additive in nonyl-β-glucoside detergent solutions. Diffraction quality crystals were obtained and allwere merohedrally twinned to different degrees (43). The crystals were also sensi-tive to visible light (75). To avoid bleaching, all procedures were performed underdim red-light illumination. Initial diffraction data for the native protein were col-lected at Stanford Synchrotron Radiation Laboratory and Advanced Photon Sourcesynchrotrons; however, attempts to solve the structure using bacteriorhodopsin as amolecular replacement model failed. Hg-derivatives of rhodopsin were generatedfor multiple isomorphous replacement (MIR) and multiwave length anomolousdispersion (MAD) phasing experiments. The structure was solved using a six-wavelength mercury MAD data set obtained at SPring8 on a crystal with a 10%twinning ratio by Miyano and colleagues (27). Crystallographic refinement of thetwo molecules in the asymmetric unit generated the current model of rhodopsin at2.8A resolution (28).
Dimers and Crystallographic Lattice
Rhodopsin molecules, crystallized in space group P41, are packed in the crys-tal lattice to form an array of tube-like structures (Figure 1A). Eight rhodopsinmolecules fill the unit cell, with∼70% of the space occupied by solvent. Thedimensions of the unit cell are: a= b = 97.3 A, c = 149.5A. Two copiesof rhodopsin (denoted molecules A and B) make up the asymmetric unit and arerelated by a non-crystallographic twofold axis between helices IA and IB. The crys-tallographic lattice is formed by remarkably few contacts between the moleculeswhen compared with an unrelated, but also seven-transmembrane helical pro-tein, sensory rhodopsin, crystallized by a lipid cubic phase method (Figure 1B)(76).
The dimer of rhodopsin, as seen in the crystal structure, cannot be physiologi-cally relevant, as one molecule is rotated by∼160◦ in relation to the second. The

21 Jan 2003 12:46 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
858 FILIPEK ET AL.
two cytoplasmic surfaces (Figure 4, see Supplemental Materials: Follow Supple-mental Materials link on Annual Reviews homepage at http://annualreviews.org/)would project toward opposite sides of the membrane if this arrangement werefound in the bilayer. One of these two rhodopsins makes much fewer contacts thanthe other in the crystal. It appears that a strong hydrophobic interaction betweenrhodopsin molecules allows elongation of the dimers in the crystal, with only onemolecule promoting the tube-like structure while the other piggybacks in the dimer.The two sets of interacting residues found for molecules A and B are not identicalbecause of asymmetry in the dimer and slightly different protein conformationsfor the two crystallographically independent molecules (Figure 4A, Suppl. Figure).These interactions may possibly deform the structure of rhodopsin to some degree;however, preservation of rhodopsin’s spectral properties suggests that any disorderwould be localized and not affect the binding site of the chromophore. The contactsbetween dimers also occur with different residues in molecules A and B (Figure4A, Suppl. Figure). In addition, these contacts involve the oligosaccharides boundto the protein (Figure 4A–C, Suppl. Figure).
Three-Dimensional Model
The current model of bovine rhodopsin (Figure 2) includes>95% of its amino acidresidues. Missing residues in the structure are Gln236GlnGlnGluSer240
and Asp331GluAla333 in molecule A; and Lys141ProMetSerAsnPheArgPhe148, Val227PheThrValLysGluAlaAlaAlaGlnGlnGlnGluSerAlaThrThr243, andPro327LeuGlyAspAspGluAlaSerThrThrValSerLysThrGluThrSerGlnValAlaPro-Ala348in molecule B. The shape of the transmembrane domain is that of an ellipticcylinder of width∼41A, height∼75A. The length and width of the elliptic foot-print on the plane at the middle of the membrane is roughly 45 by 37A. Rhodopsin,when projected on the membrane, occupies a surface close to 1500A2 (28). Twopalmitoyl chains have been identified and two oligosaccharide units have beenmodeled. Rhodopsin folds into seven transmembrane helices, I–VII, varying inlength from 22 to 34 residues, and one cytoplasmic helix (Figure 2; helix VIII inred running parallel to imaginary membranes). These helices also differ on thelevel of irregularities and are tilted at various angles with respect to the expectedmembrane surface (28). The chromophore occupies a cavity approximately twothirds from the top or cytoplasmic side of the transmembrane segment, a locationthat had been previously predicted from biophysical studies (77).
The extracellular, doubly glycosylated domain is formed by 33 N-terminalamino acids and by interhelical loops between helices II and III (residues 101–105),IV and V (residues 173–198), and VI and VII (277–285). The region displays anoticeably compact structure. Four distorted shortβ-strands form a plug that limitsaccessibility to the chromophore-binding site from this extracellular side. The Gly3
to Pro12 region forms the firstβ-hairpin that runs parallel to the expected planeof the membrane. The third and fourthβ-strands, Arg177 to Asp190, form a secondtwistedβ-hairpin. This region contains the Cys187portion of the disulfide bond. Thefourthβ-strand is below the 11-cis-retinal and is a part of the chromophore-binding

21 Jan 2003 12:46 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
G PROTEIN-COUPLED RECEPTOR RHODOPSIN 859
pocket with the carbonyl oxygen atom of Cys187being only 3.0A from C12 of theretinal. An additional consequence of the plug is that Glu181, highly conservedin rhodopsins and in short- to mid-wavelength visual pigments (29), is in closeproximity (4.4 A) to retinal at the C12 position. Glu181 modulates the stabilityof Meta II and its accessibility to hydroxylamine (78), suggesting that the plugregion prevents rapid dissociation to the intradiscal space. In red visual pigments,a His residue occupies this position, forming a part of the binding site for a Cl−
ion (79). A Cl− ion modulates theλmax of the absorption spectrum, causing abathochromic shift by∼40 nm for these cone visual pigments, compared withpigments in mutants lacking this binding site (80).
The C-terminal structure of rhodopsin does not have any secondary structuralelements. From Gly324to Asp330, the polypeptide folds back over cytoplasmic helixVIII. Residues 331–333 have no discernable electron density; therefore, the con-formation of this region is uncertain. Functionally, rhodopsin’s C terminus playstwo roles. It is necessary for the vectorial transport of rhodopsin from the site ofsynthesis to the rod outer segment (81–83) and for desensitization of photoactivatedrhodopsin (35, 84).
The cytoplasmic surface contains two short loops connecting helices I and II(residues 65–70), and helices III and IV (residues 141–149). A third cytoplasmicloop is between helices V and VI (residues 226–243). In the current model thisloop is incomplete, lacking electron density for residues 236–240. The C terminusis unstructured in solution (85) but likely folds over a loop between helices Iand II (86). At the exit of transmembrane helix VII is helix VIII, which is fullycytoplasmic, formed by residues 311–323, and 16A in length.
Two-Dimensional Model
A two-dimensional model of rhodopsin (Figure 3) is based on a similar earliermodel (9). Fourteen positively charged residues are present on the cytoplasmicsurface of rhodopsin, whereas only three are on the extracellular face, in agree-ment with the positive inside rule for multi-spanning eukaryotic membrane pro-teins (87). In particular, three positive charges in the first cytoplasmic loop ofrhodopsin may be critical for proper insertion into the membranes (88). Thenegatively charged residues are similar on both sides of the membrane. Nomi-nally, rhodopsin has four charged residues buried in the transmembrane region ofrhodopsin: Asp83 in helix II, Glu113and Glu122 in helix III, and Lys296 in helix VII.Two of these residues are engaged in chromophore-binding, one by donating anamino group (Lys296) to form a protonated Schiff base and the other, Glu113, byacting as a counterion for this linkage. Two other charged residues, along withpositive and negative residues at the ends of the transmembrane helices, couldbe involved in the formation of water channels and water polarization within thehydrophobic interior of the molecule. This inference has yet to be substantiatedby strong experimental proof.
A set of residues facing the outside of the molecule are all highly hydrophobic, asexpected, either lacking side-chains (Gly), or having a small side chain such as Ala.

21 Jan 2003 12:46 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
860 FILIPEK ET AL.
Surprisingly, five imino-acid residues, Pro, are also facing the lipids (Figure 3;the three-dimensional model is shown in Figure 5, see Supplemental Materials:Follow the Supplemental Material link on the Annual Reviews homepage athttp://physiol.annualreviews.org/). The composition of the lipid-facing residuescould be important for the formation of an organized oligomeric structure of thistransmembrane protein in the lipid bilayer (89).
Helices of Rhodopsin and Interhelical Interactions
Helix I, (encompassing residues 34 through 64 and having a length of 45A, istilted∼25◦ from the vector perpendicular to the plane of membranes and is bentby 12◦ around Pro53. The C-terminal tail of rhodopsin occupies the space overpart of the helical bundle, helix I and helix VII. Helix I forms multiple hydrogenbond interactions with helices II, VII, and VIII (Figure 4). One of the residuesthat exhibits the highest conservation among related proteins is Asn55 in helixI. Its side chain is responsible for two interhelical hydrogen bonds to Asp83 inhelix II and to the peptide carbonyl of Ala299. Additional interactions involveTyr43-Phe293 and Asn55-Ala299 with helix VII and Gln64-Thr320 with helix VIII(Figure 4). These and other hydrophobic interactions are depicted in Figure 6A(see Supplemental Materials: Follow the Supplemental Material link on the AnnualReviews homepage at http://physiol.annualreviews.org/).
Helix II, formed by residues 71 through 100, has a length of 40A, is tilted∼25◦
from the vector perpendicular to the plane of membranes, and is bent by 30◦ aroundtandem Gly residues, Gly89 and Gly90. As a consequence of this bending, Gly90
is in the vicinity of Glu113, the counterion for the protonated Schiff base of thechromophore. This helix forms multiple hydrogen bond interactions with helix I(see above), helix III (Asn78–Ser127), and helix IV (Tyr74-Glu150, Asn78-Thr160,Asn78-Trp161) (Figure 4). It appears that it forms a tight pair with helix I owingto extensive hydrophobic interactions, in addition to the ionic interaction betweenAsn55 and Asp83 (Figure 4). The cytoplasmic ends of helices II and IV are closeto each other.
Helix III, extraordinarily important for rhodopsin, encompasses residues 106 to139. It is the longest helix at 48A in length, tilted∼33◦ from the vector perpendicu-lar to the plane of membranes, and is bent twice, once by 12◦ around Gly120-Gly121
and the second by 11◦ at Ser127. The large tilt of this helix results in the helix runningacross the helical bundle and making contacts with five other helices (II, IV, V, VI,and VII) (Figure 4; Figure 6A, see Suppl. Figure). The ionic/hydrogen bond inter-actions include Asn78-Ser127, Glu122-Trp126-His211, Cys140-Thr229, Arg135-Glu247,and Glu113-Lys296(retinal). At the extracellular end of helix III, Cys110is engaged ina disulfide bond with Cys187of theβ-sheet. This disulfide bond is highly conservedamong GPCRs. In the middle of this helix, Glu113, Gly114, Ala117, Thr118, Gly120,Gly121, and Gly122 run below theβ-ionone ring to C15 of the chromophore. Glu113
is the counterion for the protonated Schiff base formed between retinal and Lys296.The C-terminal end of helix III at the cytoplasmic surface contains the E(D)RY

14 Jan 2003 15:1 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
G PROTEIN-COUPLED RECEPTOR RHODOPSIN 861
motif located in an extremely hydrophobic environment formed among residuesfrom helices II, IV, V and VI. The E(D)RY residues (although for rhodopsin thismotif is ERY, in a majority of GPCRs it is DRY and therefore referred to as theDRY motif ) have been implicated in the regulation of the receptor’s interactionwith its G protein. In this motif, there are salt bridges between Glu134, Glu247, andArg135. This organization of ionic interactions within the hydrophobic pocket couldbe one of the critical constraints in keeping rhodopsin in the inactive conforma-tion. The crystal structure in this region has high atomic displacement parameters(B-values), however, and the side chains may assume different orientations. TheDRY motif is highly conserved among GPCRs.
Helix IV, the shortest transmembrane helix, encompasses residues 150 through172 and is 33A in length. It runs almost perpendicular to the plane of the membraneand is bent by 30◦ around two neighboring proline residues, Pro170 and Pro171,near the C-terminal end of the helix. Furthermore, additional residues near thePro residues are involved in accommodating the perturbation of the helix by theseimino acids. The short nature of this helix results from the presence of the plug,which edges up close to the chromophore. The plug also includes a disulfide bondformed by Cys110 and Cys187. From helix IV, Cys167 participates in the bindingpocket of the chromophore. This helix is tightly coupled through ionic/hydrogenbond interactions with helix II (Tyr74-Glu150, Asn78-Thr160, Asn78-Trp161), interactsthrough hydrophobic interactions with helix III, and has an additional contact sitewith helix V (Ala166-Tyr206) (Figure 4; Figure 6A, see Suppl. Figure). Interestingly,the disulfide bond formed between Cys110 and Cys185 cannot substitute for thecorrect disulfide Cys110-Cys187 (90, 91). It is important to keep in mind that theCys110-Cys187 bridge is not absolutely essential for retinal-binding because Alamutants form visual pigments, although with modified bleaching properties (92).
Helix V, formed by residues 200–225 and having a length of 36A, is highly tiltedby∼26◦ from the vector perpendicular to the plane of the membrane, and is bentby 25◦ and 15◦ at Phe203and His211, respectively, surrounded by aromatic residuesat the bends. The bend at His211is caused by Pro215. Helix V forms ionic/hydrogenbond interactions with helix III (Cys140-Thr229, Glu122-Trp126-His211-Tyr206 net-work clustered about a cation or water molecule), helix IV (Ala166-Tyr206), andthrough hydrophobic interaction with helix VI (Figure 4; Figure 6A, see Suppl.Figure). At the cytoplasmic end, helix V breaks around Leu226, followed by apolypeptide chain connecting helix V and VI. This loop is critical in the bind-ing of transducin, arrestin and rhodopsin kinase. Furthermore, this helix providesresidues for the binding site for theβ-ionone ring of the chromophore (Met207,His211, and Phe212).
Helix VI, the second longest helix in rhodopsin, is composed of residues 244to 276, 47A in length, and only slightly tilted by∼5◦ from the vector perpen-dicular to the plane of membranes. It is strongly bent by 30◦ by Pro267. Helix VIappears to be most loosely associated with other helices in rhodopsin, a feature thatcould be very important during the activation process. Helix VI interacts throughionic/hydrogen bond interactions with helix III (Arg135-Glu247, important DRY

14 Jan 2003 15:1 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
862 FILIPEK ET AL.
region) and helix VII (Cys264-Thr297) (Figure 4; Figure 6A, see Suppl. Figure).Arg135 forms a salt bridge with the previous residue, Glu134, but is also connectedto Glu247and Thr251 in helix VI. These interactions could be critical constraints inkeeping rhodopsin in its inactive conformation. Interaction of helix VI with otherhelices is discussed further below. Hydrophobic residues from this helix (Val250,Met253) form a hydrophobic pocket for the ionic interaction within the DRY region.At the extracellular end of helix VI, Lys245, Lys248, and Arg252 form a positivelycharged cluster, whereas Tyr268, Phe261, Trp265, and Ala269 surround theβ-iononering. Lys245and Lys248residues are 4 and 6A, respectively, from the inner limit ofthe transmembrane domain. The indole ring of Trp265is very close to theβ-iononering and the C13-methyl group of the chromophore.
Helix VII is critical for rhodopsin function because it contains Lys296, whichforms the retinylidene linkage with the chromophore within the transmembrane re-gion. Helix VII is comprised of residues 286–309 and forms a 37A-long helix thatis minimally tilted with respect to the plane of the membrane (9◦), and is signifi-cantly bent by 24◦ and 21◦ to accommodate linkage with the chromophore aroundPro291and Pro303. It interacts with helix I (Tyr43-Phe293, Asn55-Ala299), helix III[Glu113-Lys296 (retinal)], and helix VI (Cys264-Thr297) through ionic and hydro-gen bonds, as well as hydrophobic interactions, and with helix II via hydrophobicinteractions (Figure 4; Figure 6A, see Suppl. Figure). Helix VII also stabilizescytoplasmic helix VIII (Ile307-Arg314) and provides residues that surround the pro-tonated Schiff base. The C-terminal end of this helix contains a highly conservedNPXXY motif in GPCRs. The side chain of a polar residue in the NPXXY region,Asn302, projects toward Asp83. The interaction between these residues may involvewater molecules (93). Tyr306 also projects inside the rhodopsin molecule and in-teracts with the conserved Asn73 of helix II. In addition, there is a hydrophobicinteraction between helices VII and I through Phe293 (helix VII) and Leu40 (helixI). Residues Lys296-Ala299 form a helix in the 310 conformation (three residues perhelical turn, and ten atoms in the ring closed by the hydrogen bond). All otherhelices in rhodopsin are regularα-helices. An additional kink in helix VII is atVal300. This distinct part of the helix (one helical turn) gives more freedom toLys296 because this residue has to accommodate big changes in retinal’s structureduring the first stages of isomerization. This accommodation greatly decreases theenergy barrier of isomerization.
The crystal structure of rhodopsin also revealed one cytoplasmicα-helix.Helix VIII, residues from 311 to 323, spans 16A, and forms a straight, short am-phiphilic helix connected through ionic/hydrogen bond interactions with helix I(Gln64-Thr320) and helix VII (Ile307-Arg314) (Figure 4; Figure 6A, see Suppl. Fig-ure). Elongation of the cytoplasmic surface by helix VIII may allow docking of thetransducin Gt trimer (α-, β-, γ -subunits) on the surface of a single photoactivatedrhodopsin (94). The C-terminal end of this protein contains Cys322and Cys323withthe attached palmitoyl moieties (Figure 3). The presence of these palmitoyl groups,attached enzymatically or spontaneously (46, 95), and the loop between helix VIIand VIII may affect rhodopsin regeneration (72, 96). In COS cells, Cys322 andCys323 mutants showed properties similar to those of wild-type rhodopsin (97). It

14 Jan 2003 15:1 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
G PROTEIN-COUPLED RECEPTOR RHODOPSIN 863
has been proposed that this region of helix VIII and palmitoylation is the site ofnon-covalent binding of all-trans-retinal to opsin (98), in agreement with rhodopsinmodeling (D.C. Teller, unpublished data).
Human Diseases Associated with Mutationin the Rhodopsin Gene
Mutations in the rhodopsin gene, mostly missense mutations and some frame-shiftmutations, correlate with the autosomal-dominant form of the blinding diseasetermed retinitis pigmentosa (RP) and account for∼30–40% of all cases of thisdisease (99, 100). The most relevant mutation is Pro23His, which accounts forabout 10% of autosomal-dominant RP, but∼100 other mutations were identifiedthroughout rhodopsin. The sites of mutation were previously presented in a two-dimensional model of rhodopsin (34). A null mutation in the rhodopsin geneproduces the recessive phenotype, but even heterozygotes have impaired rod vision(101, 102), suggesting the importance of a proper dosage of rhodopsin expressionfor healthy rod cells. These results are in an agreement with data obtained fromrhodopsin knockout mice (103–105). Mice lacking rhodopsin expression do notdevelop ROS, suggesting that it is also a structural protein necessary for the properformation of ROS in addition to being a receptor. Many mouse models generatedin recent years allow an understanding of how mutations in rhodopsin cause thedefect in rod cell biology.
Using the heterologous expression system, these mutants can be classifiedinto three classes: Class I is defective in photoactivation rearrangements, class IIremains in the endoplasmic reticulum and does not bind the chromophore, andclass III is expressed at low levels and poorly binds chromophore, suggesting aproblem with folding and stability (82, 106, 107). Here, we analyze some of thesemutants considering the crystal structure of rhodopsin. Mutations in rhodopsin’sextracelullar domain interfere with formation of the plug, destabilizing properfolding of rhodopsin and binding of the chromophore. The C-terminal mutationsare most likely associated with the breakdown of vectorial transport to rod outersegments (82). Owing to massive amounts of produced protein, inhibition of thisprocess would lead to blocking the inner segment as a consequence of inefficientdisposal of mutated opsin. Mutations in the transmembrane region of rhodopsinmay also lead to constitutive activity (108), metabolically overwhelming the rodphotoreceptor cells. Most of these mutations are centered on the chromophore-binding site and intense intramolecular stabilizing interactions are involved(Figure 6B, see Suppl. Figure). However, there are also mutations at the lipid-facing residues that most likely prevent proper packing of rhodopsin in ROS disksand/or formation of the organized structure of this protein within the disk.
Chromophore-Binding Site
The retinylidene-binding site is largely hydrophobic, but several charged residuesare also present around the chromophore and are necessary for the proper

21 Jan 2003 12:48 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
864 FILIPEK ET AL.
functioning of rhodopsin. The chromophore of rhodopsin, 11-cis-retinal, is linkedto opsin through a Schiff base (Figure 5). There is clear electron density fromthe crystal structure analysis that corresponds to the Schiff base and theβ-iononeportion of the chromophore (27). The helical nature of the transmembrane seg-ment in the vicinity of Lys296 is greatly distorted by Pro291 and Pro303. Lys296
is located just below the NPXXY region, which forms the C-terminal region ofhelix VII. The chromophore is in a twisted 6-s-cis-conformation within the bind-ing pocket of rhodopsin (Figure 5). The torsion angles for the chromophore aregiven in a previous publication (28). The Schiff base linkage is protonated, and thecounterion on helix III, Glu113, is 3.26A away. The vicinity of the Schiff base isformed by residues from helix II (Phe91, Thr94), helix III (Glu113, Ala117), the loopconnecting helices IV and V that forms an antiparallelβ-sheet and runs underthe chromophore (Ser186, Cys187), and helix VII (Ala292, Phe293, Ala295). The kinkin the chromophore at C11 (the cis conformation) is close to Gly114, Ala117, andThr118of helix III, and to Cys187, and Gly188 in the loop connecting helices IV andV. Theβ-ionone is surrounded by residues from helix III (Thr118, Gly121, Glu122,Leu125), helix IV (Cys167), helix V (Met207, Phe208, His211, Phe212), and helix VI(Phe261, Trp265, Tyr268, Ala269). Thr118, Gly188, Ile189, Tyr191, Tyr268 are critical forthe proper conformation around the C9-methyl of the chromophore. Lack of the C9
methyl group impedes photoactivation (109). Furthermore, Glu122 has been pro-posed to determine the rate of Meta II decay (110) (Figure 5B). Water also playsan important part in the binding site, although its location and role are not fully de-fined based on the crystallographic studies. The only well-defined water moleculeis near the retinal chromophore near C13 and is hydrogen-bonded to Glu181 andSer186 (28). The localization of the chromophore is in reasonable agreement withthe cross-linking studies of retinal analogs and identification of Phe115, Ala117,Glu122, Trp126, Ser127, and Trp265as residues in the binding pocket of theβ-iononering (111, 112)1.
Over the years, retinoid analogs have provided useful information on variousphotochemical steps during photoactivation, mapping of the chromophore-bindingsite, and the conformation of the chromophore within rhodopsin [reviewed in(113)]. Opsin reacts within minutes with 11-cis-retinal to form rhodopsin. Simi-larly, 9-cis-retinal (also called isorhodopsin) and 7-cis-retinal form visual pigmentswithλmaxat 483 nm (114) and 450 nm (115), respectively. When illuminated, 7-cis-,9-cis- and 11-cis-rhodopsin converge to the same signaling Meta II intermediate(116, 117) with quantum efficiencies one seventh, one third, and equal to thatof native rhodopsin, respectively. Double and triplecis-retinals and a number ofanalogs are known to reconstitute with opsin. Conversely, all-trans-retinal and13-cis-retinal do not regenerate the visual pigment. For rhodopsin, it is generally
1Cα of Phe115 is 6.7A from C11 of retinal, Phe115 (ring center) is 10.0A from C11 retinal,Trp126 (closest atom from ring) is 8.8A from C3 of retinal, and Ser127 is 12.0A from C3 ofretinal, distances that appear too far for direct contact. It is possible that the analog usedbinds in an alternative conformation to rhodopsin, thus satisfying the geometry of identifiedresidues.

14 Jan 2003 15:1 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
G PROTEIN-COUPLED RECEPTOR RHODOPSIN 865
accepted that 11-cis-retinal keeps this receptor in an inactive conformation. Themechanism of these structural constraints imposed by 11-cis-retinal may involvemultiple hydrophobic and ionic interactions, as described above, that bring helicestogether, sequester Arg135from the DRY region in a hydrophobic environment, andtighten up the interaction between helices VII and VIII. Light induces thecis-transisomerization and rhodopsin activation. However, the active site contains sufficientplasticity to accommodate 7-cis- and 9-cis-retinal and multiple doublecis-isomers.This fact suggests either that each of these retinoids induces a different confor-mation of the protein or that the active site has sufficient fluidity to allow isomersto fit identical or similar binding sites. From the crystal structure of rhodopsin,it appears that the binding site has well-defined binding properties. One involvesthe fixed position of the Schiff base and the second, the binding ofβ-ionone inthe active site. These two constraints on the chromophore determine the lengthof the chromophore that will fit into the binding site; any shortening or extendingof the chromophore through addition or removal of carbon atoms is non-productiveand does not lead to pigment formation (118). Furthermore, rhodopsin regeneratedwith a retinal with the 11-cis-bond constrained by cyclohexanen (6-lock-retinal)or cycloheptanen rings undergoes photoisomerization around other double bonds;however, rhodopsin remains marginally active (119; K. Palczewski, unpublisheddata). We believe that the isomerization occurs without the chromophore-inducedconformational change of the opsin moiety because theβ-ionone ring rotatesbut is largely confined within the binding site of the natural 11-cis-retinal chro-mophore. Isomerization of 6-lock-rhodopsin opens up the chromophore-bindingsite to nucleophiles, leading to hydrolysis of the chromophore (120, 121). Muta-tion of Trp265 to a smaller Phe and regeneration with the six-membered ring of11-cis-retinal leads to formation of truly photostable rhodopsin (122). Whetherisomerization of the chromophore is still taking place when bound to the mutanthas not been explored. The covalent linkage of the chromophore to opsin is notnecessary for the characteristic visible spectrum of rhodopsin or for photoactiva-tion of rhodopsin, as has been elegantly shown using opsin mutants and diffusible11-cis-retinylidene alkylamine analogs (123).
Structural Constraints and Functional Regions
The protein structure must impose the stringent properties that are required forrhodopsin to be a single-photon detector. In a milieu of 108 rhodopsin molecules,detection of a single photon requires that noise from all other non-photoactivatedrhodopsin be exceedingly low. Therefore, the rhodopsin structure evolved to fulfillthis requirement with multiple structural constraints. (a) The extracellular plugis highly structured with multiple intradomain interactions fixed by seven trans-membrane helices that likely will not allow significant spontaneous conforma-tional changes within this region to take place in the dark or during activation.However, it was noted by others that Cys185 is labeled only upon illumination(91, 124), suggesting that some conformational changes are possible. This domainis further stabilized by a disulfide bond within this plug (48, 90, 92, 125). (b) The

21 Jan 2003 12:49 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
866 FILIPEK ET AL.
chromophore tightens up the structure through multiple hydrophobic interactions,as well as ionic interactions between the protonated Schiff base (helix VII) and itscounterion, Glu113 (helix III). Mutation of Lys296 or Glu113 results in opsins thatdisplay constitutive activity toward transducin in accordance with the idea thatopsin is constrained in an inactive conformation by a salt bridge between Lys296
and Glu113 (108). (c) Asp83, with other residues and in addition to hydrophobicinteractions that involve neighboring residues, forms a tight network of interheli-cal interactions with helix I (Asn55 in helix I) and helix VII in the NPXXY region(Asn302 via a putative water molecule). (d) Highly tilted and central to the bundleof helices, the long helix III interacts with most of the other helices (with the ex-ception of helix I, which is a tightly paired with helix II), especially with helix V(Trp126-His211, Cys140-Thr229, Glu122-Trp126-His211-Tyr206). (e) Sequestered Arg135
within the hydrophobic pocket is stabilized by ionic interaction with Glu134 (thesame helix III) and Glu247 (helix VI), and further changes the Glu225 and Tyr136
hydrogen bonding (Figure 6A). It appears that Arg135 is critical for the interactionof photoactivated rhodopsin with transducin (126, 127); therefore, sequesteringthese residues may eliminate one of the key interactions between these two pro-teins. Arg135 is highly conserved among GPCRs in the so-called DRY motif. (f )The NPXXY region interacts with the double-palmitoylated helix VIII and bringsit close to the rest of the protein. The movement of helix VIII is observed duringrhodopsin activation (128) (Figure 6B). Furthermore, an anti-Meta II antibody thatrecognizes residues 304–311 (a hinge between helix VII and helix VIII) does notreact with rhodopsin or opsin (129).
Previous Models: Prediction, Bacteriorhodopsinand Cryo-Electron Microscopy
For years, the rhodopsin structure has been compared with that of bacteriorhodop-sin, also a retinylidene seven-transmembrane protein. However, the arrangementsof the seven helices were found to be different on the basis of earlier low-resolutionstudies on rhodopsin using cryo-electron microscopy at 7.5A resolution in theplane of the membrane and at 16.5A resolution perpendicular to the membrane(130–132) [see structural comparison in (28)]. The projection map of inverte-brate rhodopsin is also similar to maps previously determined for bovine and frogrhodopsins (133). In rhodopsin, the helices are slightly longer than those of bacte-riorhodopsin and differently arranged. Helices IV and V of the two proteins do notsuperimpose (whereas helices I, II, and III are similar). The helices in rhodopsin aremore tilted and kinked, and its extramembrane regions are larger and more orga-nized. Rhodopsin also contains an extra helix (helix VIII). The retinal chromophoreis in the twisted 6-s-cisconformation in rhodopsin, whereas in bacteriorhodopsin itis 6-s-trans-conformation. In addition, the retinylidene groups in the two proteinsare in different positions relative to the membrane plane, although the ionone ringsare placed in similar positions. Because G protein receptors were not amenable forcrystallographic studies, three-dimensional models based on the modeling stud-ies were proposed. A particular model of rhodopsin was computed that closely

14 Jan 2003 15:1 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
G PROTEIN-COUPLED RECEPTOR RHODOPSIN 867
resembles the transmembrane region structure found from the actual crystal struc-ture (134). Similar models were also generated for other GPCRs (135).
A superimposition of the crystal structure of bovine rhodopsin and a model offrog rhodopsin from cryo-electron microscopy (gray balls), obtained under differ-ent resolutions and in different packing environments, are generally in agreement(Figure 7, see Supplemental Materials: Follow the Supplemental Material link onthe Annual Reviews homepage at http://annualreviews.org/). However, the largestdifferences are seen within helices I, III, and VII. Remarkably, several of thekinked helices are observed in both structures. In addition to these structural stud-ies, UV, FTIR, NMR, and EPR spectroscopic investigations of rhodopsin and itsmutants in inactive and photoactivated states provide additional validation of thecrystallographic model and further significant insights into rhodopsin structureand activation. Owing to space limitation, these spectroscopic studies cannot befully evaluated herein.
ACTIVATION MECHANISM OF RHODOPSIN
Unified Model
Although the activation of rhodopsin is still roughly sketched, the crystal structureof rhodopsin provides the opportunity to summarize our current understanding ofthis process. The present view is inevitably oversimplified because of a vast amountof information existing in the literature. Therefore, the objective is to present a sim-plified, but coherent, hypothesis that unites major findings on rhodopsin activation.The reader should also consult recent publications (34, 75, 136, 137). One shouldalso reflect on the complexity of the interactions with the transmembrane segmentof rhodopsin (Figure 6A, see Suppl. Figure). Our activation model discusses onlythe major determinants of the activation process.
A photon of visible light absorbed by rhodopsin provides sufficient energy forthe structural transformation from quiescence into a signaling protein. First, photo-isomerization causes transformation of 11-cis-retinylidene to all-trans-retinylidene.The quantum yield for this reaction is∼0.67 (138). The reaction is too fast forany significant conformational change in opsin (139) (Figure 8, see SupplementalMaterials: Follow the Supplemental Material link on the Annual Reviews home-page at http://annualreviews.org/). Twisting of all-trans-retinylidene within theoriginal 11-cis-retinylidene binding site of opsin is a way by which two thirds of the55 kcal/mol energy of the photon is stored in the all-trans-retinylidene-opsin com-plex (140). This energy is then released during relaxation of earlier intermediates tothe signaling stage of the receptor, Meta II. Formation of Meta II leads to relocationof theβ-ionone ring to a new position, as proposed by cross-linking studies (141)2
(Figure 8, see Suppl. Figure). Photoisomerization of the chromophore may initially
2Agreement with cross-linking studies with the crystal structure of rhodopsin requires thatduring the photoactivating process, helix IV undergoes a large rotation. Independent meth-ods are needed to verify the cross-link observations.

21 Jan 2003 12:52 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
868 FILIPEK ET AL.
involve reorganization of the 310 helical region of helix VII. The consequence ofthis movement is the breakage of a salt bridge between Glu113 and the protonatedSchiff base between Lys296 and the retinal (108). This involves movement of aproton from the donor to the acceptor (142) in conjunction with the motion of thetransmembrane helices (136, 143). The movement of helix VII away from helix Iby 2–4A and the pushing of helix VI (64)∼3 A by the photolyzed chromophorefrom the first cytoplasmic loop lead to removal of Glu247from the ionic interactionin the DRY region (Figure 6) [summarized in (136)]. Subsequent movement ofArg135 to the cytosol allows photoactivated rhodopsin to attract and interact withtransducin. Protonation of Glu134, which is left in a very hydrophobic milieu, com-pletes this transformation (34). This would reflect changes attributed to the centralhelix III (144). It is also important to note that helix VI is only loosely associatedwith the remaining bundle of helices (28) (Figures 4 and 7). The residues in he-lix VI, particularly Pro267, affect the assembly of rhodopsin from two fragmentsand the conformation of the cytoplasmic loop III (72). For theα1B-adrenergic re-ceptor, this region has been identified as key to the coupling process with its Gprotein (145). Therefore, the charge-neutralizing mutation Glu134 to Gln134 pro-duces hyperactivity in the activated state and produces constitutive activity in opsin(144). The consequences of photoactivation are also transmitted toward a secondregion, the NPXXY cluster in helix VII. Structural changes connected with ac-tivation have been detected using spin labels at residues 306, 313, and 316. Thechanges at side-chain 313 can be accounted for by movements in the adjacent helixVII (146). Movement of this helix is critical for efficient coupling with transducin(147, 148). Methyl groups, in particular the C9-methyl group on retinal, are criticalfor efficient movement of this helix (O.P. Ernst, K.P. Hofmann & K. Palczewski,unpublished data). The separation of helices on the cytoplasmic side of Meta IIopens up a cavity into which transducin can dock (149). The formation of Meta IIis transiently stabilized by rearrangement of the Glu122-Trp126-His211 triad(helices III–V) (150) before Meta II decays to opsin. Therefore, the activationprocess could be considered a removal of structural constraints in rhodopsin, andit is not surprising that a soluble cytoplasmic domain grafted into soluble proteinsis very effective in the stimulation of transducin (151). It is important to note thatcertain phospholipids may impede these conformational changes (152).
Cavities and the Second Chromophore-Binding Site
The resolution of the rhodopsin structure is 2.8A and does not allow precise lo-calization of all water molecules associated with the receptor. We have identifiedseveral regions around the chromophore and intracellular face that could be oc-cupied by water molecules in the dark state, as well as large cavities suitablefor protein-protein interactions (Figure 9, see Supplemental Materials: Followthe Supplemental Material link on the Annual Reviews homepage at http://annualreviews. org/). Water molecules could be involved in the activation processsimilar to the role of water in the bacteriorhodopsin photocycle. The reorganization

21 Jan 2003 12:52 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
G PROTEIN-COUPLED RECEPTOR RHODOPSIN 869
of water molecules during activation has been detected by FTIR spectroscopy(153). Furthermore, one of these cavities could be involved in the non-covalentbinding of all-trans-retinal, which activates opsin in transducin, arrestin, andrhodopsin kinase assays [reviewed in (60)]. One such binding site may be presentaround helix VIII and the palmitoylation sites (D.C. Teller, unpublished data). Arecent refinement of rhodopsin at 2.6A resolution has provided an updated viewof the contributions of water molecules to the activity of the protein (93).
CONE VISUAL PIGMENTS
Although a full analysis of spectral tuning and the properties of the cone pig-ments is outside the scope of this review, a few points are worth mentioning.Rhodopsin appears to be an ideal structural template for cone pigments. We car-ried out homology modeling for cone pigments that revealed several interestingfeatures related to the DRY region (Figure 8A) and the chromophore-bindingsites (Figure 8B) (154). In contrast, the NPXXY region is highly homologousamong cone pigments and rhodopsin (data not shown). The only difference in thechromophore-binding site between green and red pigments is in the immediatevicinity of retinal, where Phe261 is replaced by Tyr261 (red), as predicted fromstudies on the spectral tuning of primates aimed at understanding the molecularproperties underlying red-green color vision (155, 156). These studies were ex-tended to identify residues responsible in in vitro experiments [157, 158; reviewedin (159)]. Furthermore, it has been convincingly documented that these in vitroanalyses correlate with the in vivo data based on psychophysical measurements(160). The difference inλmax between the green and red pigments has been pro-posed to result from other hydroxyl-bearing amino acids (Ser164and Thr269) that arefarther away from the chromophore [(161); reviewed in (159)]. These earlierfindings were confirmed by more recent studies (162). The difference betweenrhodopsin and green pigments (30 nm blue shift) could be accounted for by the in-creased hydrogen bonding of the protonated Schiff base group in rhodopsin (163).In the chromophore-binding site, Ser202 is shown to hydrogen bond with Glu113 inthe blue pigment, but points toward a Cl− ion in the red/green pigment. Throughmutagenesis studies, two residues, His197 and Lys200, were identified in the Cl−-binding site of red and green visual pigments (79). In our model, Lys200 does notparticipate in the binding of this ion.
OTHER G PROTEIN-COUPLED RECEPTORS
An enormous set of information must flow across the cell surface to the cell toensure survival, and transmembrane proteins play a key role in this process. Mem-brane proteins, among them rhodopsin-like proteins, GPCRs, represent a large andversatile group of protein sensors involved in nearly all physiological processes invertebrates. In all organisms, from the most primitive to mammals, but perhaps not

21 Jan 2003 12:52 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
870 FILIPEK ET AL.
in plants, GPCRs are present in multiple molecular forms. For example, 5–10%of theHomo sapiensgenome encodes GPCRs, which translates into>600 non-sensory and 1000s of sensory GPCRs. These molecules are involved in decodinginformation through the olfaction, vision, and taste systems, and comprise oneof the largest families of proteins encoded by our genome. Understanding howrhodopsin, the best-studied member of this family, works reaches far beyond itskey role in vision. Parallel information that can be learned from phototransduc-tion is directly implemented into a better picture of other signal transduction viaGPCRs, one of the most important topics in current neurobiology/pharmacology.
GPCRs transduce an extracellular signal by coupling with G proteins on the cy-toplasmic side of the cell; the binding site for the activating ligand is either withinthe hydrophobic bundle of helices or at an extracellular domain of these receptors.There is a growing body of evidence that these receptors may also modulate the in-tracellular processes independently of G proteins (164). On the basis of the conser-vation pattern of the primary amino acid sequence, three major families of GPCRshave been described (165). Family 1 (or A), the largest of the three, is defined byreceptors whose sequences are homologous to rhodopsin, including opiate, adren-ergic, cannabinoid, and muscarinic receptors. Family 2 (or B) consists of GPCRshomologous in sequence to the secretin receptor, and Family 3 (or C) includesGPCRs homologous in sequence to the GABA receptor. The overall homology be-tween receptors even within the same family is low (frequently<35%). However,some regions are highly conserved, suggesting that their function is either struc-tural or related to the molecular mechanism of receptor activation. Some of theseconserved residues, clustered in space, define conserved three-dimensional motifsreferred to as functional microdomains (167, 169). The functional microdomainsinclude the DRY region (Figure 9B, in rhodopsin ERY), helix VIII and palmi-toyl moieties, the NPXXY region, a ligand-binding site, a Pro kink in helix VI, adisulfide bond, and oligosaccharide moieties. Therefore, rhodopsin is a very use-ful template for all GPCRs, at least until more high-resolution structures becomeavailable, especially for Family 1 (163).
The structure and organization of rhodopsin and other GPCRs in native mem-branes is not known. For many GPCRs, including adenosine, adrenergic, dopamine,opioid, and other receptors, dimeric structures have been more or less stringentlydocumented (166).
FINAL REMARKS
In coming years, further advances will be made by understanding all of the proteinsand complexes in different signaling states on a structural level and by givingreliable and logical chemical/physical models of visual transduction at an atomicresolution that will be coherent and consistent with thermodynamic laws. Thestructures of G proteins, arrestins, and now rhodopsin are the first steps towardthese goals. Similar to research on the metabolic pathways in the past 10–20 years,signal transduction by GPCRs has become a very mature field. However, many

14 Jan 2003 15:1 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
G PROTEIN-COUPLED RECEPTOR RHODOPSIN 871
fundamental questions remain in relation to the activation mechanism of GPCRs,the specificity and recognition of interacting proteins and their mechanisms ofactivation by GPCRs, and the structure of receptors and interacting proteins inthe native setting of phospholipids in in vivo conditions. The great interest inthese topics from academic and industrial laboratories makes it certain that furtherprogress will be made.
ACKNOWLEDGMENTS
We thank Yunie Kim for her outstanding help during manuscript preparation. Thisresearch was supported in part by U.S. Public Health Service grants EY01730,EY08061 and EY0861; from the National Eye Institute, National Institutes ofHealth, grant GM63020; an unrestricted grant from Research to Prevent Blindness,Inc. (RPB) to the Department of Ophthalmology at the University of Washington;and a grant from the E.K. Bishop Foundation. Calculations and visualizations werepartly done in the ICM Computer Center, University of Warsaw, Poland.
The Annual Review of Physiologyis online at http://physiol.annualreviews.org
LITERATURE CITED
1. Kuhne W. 1878.Ueber den Sehpurpur.Heidelberg: Unters. Physiol. Inst. Univ.Heidelberg. 113 pp.
2. Wald G. 1935–1936. Carotenoids and thevisual cycle.J. Gen. Physiol.19:351–71
3. Hubbard R, Kropf A. 1967. Molecular iso-mers in vision.Sci. Am.216:64–70
4. Wald G. 1968. The molecular basis of vi-sual excitation.Nature219:800–7
5. Matthews RG, Hubbard R, Brown PK,Wald G. 1963. Tautomeric forms of meta-rhodopsin.J. Gen. Physiol.47:215–40
6. Hargrave PA. 1982. Rhodopsin chemistry,structure and topography.Prog. RetinalRes.1:1–51
7. Hargrave PA, McDowell JH, Curtis DR,Wang JK, Juszczak E, et al. 1983. Thestructure of bovine rhodopsin.Biophys.Struct. Mech.9:235–44
8. Ovchinnikov YA. 1982. Rhodopsin andbacteriorhodopsin: structure-function re-lationships.FEBS Lett.148:179–91
9. Argos P, Rao JK, Hargrave PA. 1982.Structural prediction of membrane-boundproteins.Eur. J. Biochem.128:565–75
10. Fung BK-K, Stryer L. 1980. Photolyzedrhodopsin catalyzes the exchange of GTPfor bound GDP in retinal.Proc. Natl.Acad. Sci. USA77:2500–4
11. Kuhn H, Bennett N, Michel Villaz M,Chabre M. 1981. Interactions betweenphotoexcited rhodopsin and GTP-bindingprotein. Proc. Natl. Acad. Sci. USA78:6873–77
12. Longstaff C, Calhoon RD, Rando RR.1986. Deprotonation of the Schiff base ofrhodopsin is obligate in the activation ofthe G protein.Proc. Natl. Acad. Sci. USA83:4209–13
13. Bownds D, Dawes J, Miller J, StahlmanM. 1972. Phosphorylation of frog photo-receptor membranes induced by light.Nat. New Biol.237:125–27
14. Frank RN, Cavanagh HD, Kenyon KR.1973. Light-stimulated phosphorylationof bovine visual pigments by adenosinetriphosphate.J. Biol. Chem.248:596–609
15. Kuhn H, Dreyer WJ. 1972. Light de-pendent phosphorylation of rhodopsin byATP. FEBS Lett.20:1–6

14 Jan 2003 15:1 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
872 FILIPEK ET AL.
16. Liebman PA, Pugh EN Jr. 1980. ATP me-diates rapid reversal of cyclic GMP phos-phodiesterase activation in visual receptormembranes.Nature287:734–36
17. Sitaramayya A, Liebman PA. 1983. Phos-phorylation of rhodopsin and quenchingof cyclic GMP phosphodiesterase activa-tion by ATP at weak bleaches.J. Biol.Chem.258:12106–9
18. Palczewski K, McDowell JH, HargravePA. 1988. Purification and characteriza-tion of rhodopsin kinase.J. Biol. Chem.263:14067–73
19. Lorenz W, Inglese J, Palczewski K, Ono-rato JJ, Caron MG, Lefkowitz RJ. 1991.The receptor kinase family: Primary struc-ture of rhodopsin kinase reveals simi-larities to the beta-adrenergic receptorkinase.Proc. Natl. Acad. Sci. USA88:8715–19
20. Wilden U, Hall SW, Kuhn H. 1986. Phos-phodiesterase activation by photoexcitedrhodopsin is quenched when rhodopsinis phosphorylated and binds the intrinsic48-kDa protein of rod outer segments.Proc. Natl. Acad. Sci. USA83:1174–78
21. Palczewski K. 1994. Structure and func-tions of arrestins.Protein Sci.3:1355–61
22. Nathans J, Hogness DS. 1983. Isolation,sequence analysis, and intron-exon ar-rangement of the gene encoding bovinerhodopsin.Cell 34:807–14
23. Nathans J, Hogness DS. 1984. Isola-tion and nucleotide sequence of the geneencoding human rhodopsin.Proc. Natl.Acad. Sci. USA81:4851–55
24. Nathans J, Thomas D, Hogness DS. 1986.Molecular genetics of human color vision:the genes encoding blue, green, and redpigments.Science232:193–202
25. Zuker CS, Cowman AF, Rubin GM. 1985.Isolation and structure of a rhodopsin genefrom D. melanogaster. Cell40:851–58
26. Ferretti L, Karnik SS, Khorana HG, Nas-sal M, Oprian DD. 1986. Total synthesisof a gene for bovine rhodopsin.Proc. Natl.Acad. Sci. USA83:599–603
27. Palczewski K, Kumasaka T, Hori T, Beh-nke CA, Motoshima H, et al. 2000. Crys-tal structure of rhodopsin: a G protein-coupled receptor.Science289:739–45
28. Teller DC, Okada T, Behnke CA, Palc-zewski K, Stenkamp RE. 2001. Advancesin determination of a high-resolutionthree-dimensional structure of rhodopsin,a model of G-protein-coupled receptors(GPCRs).Biochemistry40:7761–72
29. Ebrey T, Koutalos Y. 2001. Vertebratephotoreceptors.Prog. Retina Eye Res.20:49–94
30. Fain GL, Matthews HR, Cornwall MC,Koutalos Y. 2001. Adaptation in verte-brate photoreceptors.Physiol. Rev.81:117–51
31. Polans A, Baehr W, Palczewski K. 1996.Turned on by Ca2+! The physiology andpathology of Ca2+-binding proteins in theretina.Trends Neurosci.19:547–54
32. Khorana HG. 1999. Molecular biology oflight transduction by the mammalian pho-toreceptor, rhodopsin.J. Biomol. Struct.Des.1:1–16
33. Menon ST, Han M, Sakmar TP. 2001.Rhodopsin: structural basis of molecularphysiology.Physiol. Rev.81:1659–88
34. Okada T, Ernst OP, Palczewski K, Hof-mann KP. 2001. Activation of rhodopsin:new insights from structural and biochem-ical studies.Trends Biochem. Sci.26:318–24
35. Palczewski K, Benovic JL. 1991. G-pro-tein-coupled receptor kinases.TrendsBiochem. Sci.16:387–91
36. Palczewski K, Polans AS, Baehr W, AmesJB. 2000. Ca2+-binding proteins in theretina: structure, function, and the etiol-ogy of human visual diseases.BioEssays22:337–50
37. Zhao X, Yokoyama K, Whitten ME,Huang J, Gelb MH, Palczewski K. 1999.A novel form of rhodopsin kinase fromchicken retina and pineal gland.FEBSLett.454:115–21
38. Treisman JE, Morabito MA, BarnstableCJ. 1988. Opsin expression in the rat

14 Jan 2003 15:1 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
G PROTEIN-COUPLED RECEPTOR RHODOPSIN 873
retina is developmentally regulated bytranscriptional activation.Mol. Cell. Biol.8:1570–79
39. Litman BJ. 1982. Purification of rhod-opsin by concanavalin A affinity chro-matography.Methods Enzymol.81:150–53
40. Hong K, Knudsen PJ, Hubbell WL. 1982.Purification of rhodopsin on hydroxya-patite columns, detergent exchange, andrecombination with phospholipids.Meth-ods Enzymol.81:144–50
41. Oprian DD, Asenjo AB, Lee N, PelletierSL. 1991. Design, chemical synthesis,and expression of genes for the three hu-man color vision pigments.Biochemistry30:11367–72
42. Deleted in proof43. Okada T, Le Trong I, Fox BA, Behnke CA,
Stenkamp RE, Palczewski K. 2000. X-raydiffraction analysis of three-dimensionalcrystals of bovine rhodopsin obtainedfrom mixed micelles.J. Struct. Biol.130:73–80
44. Bownds D. 1967. Site of attachment ofretinal in rhodopsin.Nature216:1178–81
45. Hargrave PA, Fong SL. 1977. The amino-and carboxyl-terminal sequence of bovinerhodopsin.J. Supramol. Struct.6:559–70
46. O’Brien PJ, Zatz M. 1984. Acylation ofbovine rhodopsin by [3H]palmitic acid.J.Biol. Chem.259:5054–57
47. Ovchinnikov YA, Abdulaev NG, Bo-gachuk AS. 1988. Two adjacent cysteineresidues in the C-terminal cytoplasmicfragment of bovine rhodopsin are palmity-lated.FEBS Lett.230:1–5
48. Karnik SS, Sakmar TP, Chen HB, Kho-rana HG. 1988. Cysteine residues 110and 187 are essential for the formationof correct structure in bovine rhodopsin.Proc. Natl. Acad. Sci. USA85:8459–63
49. Fukuda MN, Papermaster DS, HargravePA. 1982. Structural analysis of carbohy-drate moiety of bovine rhodopsin.Meth-ods Enzymol.81:214–23
50. Schey KL, Papac DI, Knapp DR, CrouchRK. 1992. Matrix-assisted laser desorp-tion mass spectrometry of rhodopsin andbacteriorhodopsin.Biophys. J.63:1240–43
51. Palczewski K, Buczylko J, Kaplan MW,Polans AS, Crabb JW. 1991. Mechanismof rhodopsin kinase activation.J. Biol.Chem.266:12949–55
52. Ohguro H, Rudnicka-Nawrot M, Buc-zylko J, Zhao X, Taylor JA, et al. 1996.Structural and enzymatic aspects of rhod-opsin phosphorylation.J. Biol. Chem.271:5215–24
53. Ohguro H, Van Hooser JP, Milam AH,Palczewski K. 1995. Rhodopsin phospho-rylation and dephosphorylation in vivo.J.Biol. Chem.270:14259–62
54. Sokal I, Pulvermuller A, Buczylko J, Hof-mann KP, Palczewski K. 2002. Rhodopsinand its kinase.Methods Enzymol.343:578–600
55. Wang Q, Schoenlein RW, Peteanu LA,Mathies RA, Shank CV. 1994. Vibra-tionally coherent photochemistry in thefemtosecond primary event of vision.Sci-ence266:422–24
56. Liu RS. 2001. Photoisomerization byhula-twist: a fundamental supramolecularphotochemical reaction.Acc. Chem. Res.34:555–62
57. Shichida Y, Imai H. 1998. Visual pigment:G-protein-coupled receptor for light sig-nals.Cell. Mol. Life Sci.54:1299–315
58. Janz JM, Farrens DL. 2001. Engineeringa functional blue-wavelength-shifted rho-dopsin mutant.Biochemistry40:7219–27
59. Szundi I, Mah TL, Lewis JW, Jager S,Ernst OP, et al. 1998. Proton transfer reac-tions linked to rhodopsin activation.Bio-chemistry37:14237–44
60. McBee JK, Palczewski K, Baehr W, Pep-perberg DR. 2001. Confronting complex-ity: the interlink of phototransductionand retinoid metabolism in the verte-brate retina.Prog. Retin. Eye Res.20:469–529

14 Jan 2003 15:1 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
874 FILIPEK ET AL.
61. Kefalov VJ, Cornwall MC, Crouch RK.1999. Occupancy of the chromophorebinding site of opsin activates visual trans-duction in rod photoreceptors.J. Gen.Physiol.113:491–503
62. Arnis S, Hofmann KP. 1995. Photoregen-eration of bovine rhodopsin from its sig-naling state.Biochemistry34:9333–40
63. Bartl FJ, Ritter E, Hofmann KP. 2001. Sig-naling states of rhodopsin: absorption oflight in active metarhodopsin II generatesan all-trans-retinal bound inactive state.J.Biol. Chem.276:30161–66
64. Oprian DD, Molday RS, Kaufman RJ,Khorana HG. 1987. Expression of a syn-thetic bovine rhodopsin gene in monkeykidney cells.Proc. Natl. Acad. Sci. USA84:8874–78
65. Klaassen CH, DeGrip WJ. 2000. Bac-ulovirus expression system for expressionand characterization of functional recom-binant visual pigments.Methods Enzy-mol.315:12–29
66. Abdulaev NG, Ridge KD. 2000. Heterolo-gous expression of bovine opsin inPichiapastoris. Methods Enzymol.315:3–11
67. Mollaaghababa R, Davidson FF, KaiserC, Khorana HG. 1996. Structure andfunction in rhodopsin: expression offunctional mammalian opsin inSaccha-romyces cerevisiae. Proc. Natl. Acad. Sci.USA93:11482–86
68. Nathans J. 1990. Determinants of visualpigment absorbance: identification of theretinylidene Schiff’s base counterion inbovine rhodopsin.Biochemistry29:9746–52
69. Zhukovsky EA, Oprian DD. 1989. Ef-fect of carboxylic acid side chains on theabsorption maximum of visual pigments.Science246:928–30
70. Sakmar TP, Franke RR, Khorana HG.1989. Glutamic acid-113 serves as theretinylidene Schiff base counterion inbovine rhodopsin.Proc. Natl. Acad. Sci.USA86:8309–13
71. Kaushal S, Ridge KD, Khorana HG. 1994.Structure and function in rhodopsin: the
role of asparagine-linked glycosylation.Proc. Natl. Acad. Sci. USA91:4024–28
72. Ridge KD, Ngo T, Lee SS, Abdulaev NG.1999. Folding and assembly in rhodopsin.Effect of mutations in the sixth trans-membrane helix on the conformation ofthe third cytoplasmic loop.J. Biol. Chem.274:21437–42
73. Ridge KD, Lee SS, Yao LL. 1995. In vivoassembly of rhodopsin from expressedpolypeptide fragments.Proc. Natl. Acad.Sci. USA92:3204–8
74. Okada T, Takeda K, Kouyama T. 1998.Highly selective separation of rhodopsinfrom bovine rod outer segment mem-branes using combination of divalentcation and alkyl(thio)glucoside.Photo-chem. Photobiol.67:495–99
75. Okada T, Palczewski K. 2001. Crystalstructure of rhodopsin: implications forvision and beyond.Curr. Opin. Struct.Biol. 11:420–26
76. Royant A, Nollert P, Edman K, Neutze R,Landau EM, et al. 2001. X-ray structure ofsensory rhodopsin II at 2.1-A resolution.Proc. Natl. Acad. Sci. USA98:10131–36
77. Thomas DD, Stryer L. 1982. Transverselocation of the retinal chromophore ofrhodopsin in rod outer segment disc mem-branes.J. Mol. Biol.154:145–57
78. Yan EC, Kazmi MA, De S, Chang BS,Seibert C, et al. 2002. Function of ex-tracellular loop 2 in rhodopsin: glutamicacid 181 modulates stability and absorp-tion wavelength of metarhodopsin II.Bio-chemistry41:3620–27
79. Wang Z, Asenjo AB, Oprian DD. 1993.Identification of the Cl(−)-binding site inthe human red and green color vision pig-ments.Biochemistry32:2125–30
80. Crescitelli F. 1977. The visual pigmentsof geckos and other vertebrates: an es-say in comparative biology. InHandbookof Sensory Physiology, pp. 391–449. NewYork: Springer-Verlag
81. Moritz OL, Tam BM, Papermaster DS,Nakayama T. 2001. A functional rhod-opsin-green fluorescent protein fusion

14 Jan 2003 15:1 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
G PROTEIN-COUPLED RECEPTOR RHODOPSIN 875
protein localizes correctly in transgenicXenopus laevisretinal rods and is ex-pressed in a time-dependent pattern.J.Biol. Chem.276:28242–51
82. Sung CH, Makino C, Baylor D, Nathans J.1994. A rhodopsin gene mutation respon-sible for autosomal dominant retinitis.J.Neurosci.14:5818–33
83. Tam BM, Moritz OL, Hurd LB, Paper-master DS. 2000. Identification of an outersegment targeting signal in the COOHterminus of rhodopsin using transgenicXenopus laevis. J. Cell Biol.151:1369–80
84. Cideciyan AV, Zhao X, Nielsen L, KhaniSC, Jacobson SG, Palczewski K. 1998.Null mutation in the rhodopsin kinasegene slows recovery kinetics of rod andcone phototransduction in man.Proc.Natl. Acad. Sci. USA95:328–33
85. Langen R, Cai K, Altenbach C, KhoranaHG, Hubbell WL. 1999. Structural fea-tures of the C-terminal domain of bovinerhodopsin: a site-directed spin-labelingstudy.Biochemistry38:7918–24
86. Mielke T, Villa C, Edwards PC, SchertlerGF, Heyn MP. 2002. X-ray diffractionof heavy-atom labelled two-dimensionalcrystals of rhodopsin identifies the posi-tion of cysteine 140 in helix 3 and cysteine316 in helix 8.J. Mol. Biol.316:693–709
87. Sipos L, von Heijne G. 1993. Predictingthe topology of eukaryotic membrane pro-teins.Eur. J. Biochem.213:1333–40
88. Hartmann E, Rapoport TA, Lodish HF.1989. Predicting the orientation of eu-karyotic membrane-spanning proteins.Proc. Natl. Acad. Sci. USA86:5786–90
89. Popot JL, Engelman DM. 2000. Helicalmembrane protein folding, stability, andevolution.Annu. Rev. Biochem.69:881–922
90. Hwa J, Reeves PJ, Klein-Seetharaman J,Davidson F, Khorana HG. 1999. Structureand function in rhodopsin: further elucida-tion of the role of the intradiscal cysteines,Cys-110, -185, and -187, in rhodopsinfolding and function.Proc. Natl. Acad.Sci. USA96:1932–35
91. Ridge KD, Lu Z, Liu X, Khorana HG.1995. Structure and function in rhodopsin.Separation and characterization of the cor-rectly folded and misfolded opsins pro-duced on expression of an opsin mutantgene containing only the native intradiscalcysteine codons.Biochemistry34:3261–67
92. Davidson FF, Loewen PC, Khorana HG.1994. Structure and function in rhodopsin:replacement by alanine of cysteine re-sidues 110 and 187, components of a con-served disulfide bond in rhodopsin, affectsthe light-activated metarhodopsin II state.Proc. Natl. Acad. Sci. USA91:4029–33
93. Okada T, Fujiyoshi Y, Silow M, NavarroJ, Landau EM, Shichida Y. 2002. Func-tional role of internal water molecules inrhodopsin revealed by X-ray crystallogra-phy.Proc. Natl. Acad. Sci. USA99:5982–87
94. Kisselev OG, Meyer CK, Heck M, ErnstOP, Hofmann KP. 1999. Signal transferfrom rhodopsin to the G-protein: evidencefor a two-site sequential fit mechanism.Proc. Natl. Acad. Sci. USA96:4898–903
95. Veit M, Sachs K, Heckelmann M, Maret-zki D, Hofmann KP, Schmidt MF. 1998.Palmitoylation of rhodopsin with S-protein acyltransferase: enzyme catalyzedreaction versus autocatalytic acylation.Biochim. Biophys. Acta1394:90–98
96. Weiss ER, Osawa S, Shi W, DickersonCD. 1994. Effects of carboxyl-terminaltruncation on the stability and G protein-coupling activity of bovine rhodopsin.Biochemistry33:7587–93
97. Karnik SS, Ridge KD, Bhattacharya S,Khorana HG. 1993. Palmitoylation ofbovine opsin and its cysteine mutants inCOS cells.Proc. Natl. Acad. Sci. USA90:40–44
98. Sachs K, Maretzki D, Meyer CK, Hof-mann KP. 2000. Diffusible ligand all-trans-retinal activates opsin via a palmi-toylation-dependent mechanism.J. Biol.Chem.275:6189–94

14 Jan 2003 15:1 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
876 FILIPEK ET AL.
99. Rattner A, Sun H, Nathans J. 1999. Molec-ular genetics of human retinal disease.Annu. Rev. Genet.33:89–131
100. Dryja TP, Li T. 1995. Molecular geneticsof retinitis pigmentosa.Hum. Mol. Genet.4:1739–43
101. Rosenfeld PJ, Cowley GS, McGee TL,Sandberg MA, Berson EL, Dryja TP.1992. A null mutation in the rhodopsingene causes rod photoreceptor dysfunc-tion and autosomal recessive retinitis pig-mentosa.Nat. Genet.1:209–13
102. Kumaramanickavel G, Maw M, DentonMJ, John S, Srikumari CR, et al. 1994.Missense rhodopsin mutation in a familywith recessive RP.Nat. Genet.8:10–11
103. Toda K, Bush RA, Humphries P, SievingPA. 1999. The electroretinogram of therhodopsin knockout mouse.Vis. Neurosci.16:391–98
104. Humphries MM, Rancourt D, Farrar GJ,Kenna P, Hazel M, et al. 1997. Retinopa-thy induced in mice by targeted disrup-tion of the rhodopsin gene.Nat. Genet.15:216–19
105. Lem J, Krasnoperova NV, Calvert PD,Kosaras B, Cameron DA, et al. 1999. Mor-phological, physiological, and biochemi-cal changes in rhodopsin knockout mice.Proc. Natl. Acad. Sci. USA96:736–41
106. Jacobson SG, Kemp CM, Sung CH,Nathans J. 1991. Retinal function andrhodopsin levels in autosomal dominantretinitis.Am. J. Ophthalmol.112:256–71
107. Kaushal S, Khorana HG. 1994. Structureand function in rhodopsin. 7. Point mu-tations associated with autosomal dom-inant retinitis pigmentosa.Biochemistry33:6121–28
108. Rao VR, Oprian DD. 1996. Activat-ing mutations of rhodopsin and otherG protein-coupled receptors.Annu. Rev.Biophys. Biomol. Struct.25:287–314
109. Vogel R, Fan GB, Sheves M, Siebert F.2000. The molecular origin of the inhibi-tion of transducin activation in rhodopsinlacking the 9-methyl group of the retinalchromophore: a UV-Vis and FTIR spec-
troscopic study.Biochemistry39:8895–908
110. Imai H, Kojima D, Oura T, TachibanakiS, Terakita A, Shichida Y. 1997. Singleamino acid residue as a functional deter-minant of rod and cone visual pigments.Proc. Natl. Acad. Sci. USA94:2322–26
111. Nakayama TA, Khorana HG. 1991. Map-ping of the amino acids in membrane-embedded helices that interact with theretinal chromophore in bovine rhodopsin.J. Biol. Chem.266:4269–75
112. Nakayama TA, Khorana HG. 1990. Orien-tation of retinal in bovine rhodopsin deter-mined by cross-linking using a photoac-tivatable analog of 11-cis-retinal.J. Biol.Chem.265:15762–69
113. Lou J, Tan Q, Karnaukhova E, Berova N,Nakanishi K, Crouch RK. 2000. Syntheticretinals: convenient probes of rhodopsinand visual transduction process.MethodsEnzymol.315:219–37
114. Okada T, Kandori H, Shichida Y, Yoshi-zawa T, Denny M, et al. 1991. Spectro-scopic study of the batho-to-lumi tran-sition during the photobleaching ofrhodopsin using ring-modified retinalanalogues.Biochemistry30:4796–802
115. DeGrip WJ, Liu RS, Ramamurthy V,Asato A. 1976. Rhodopsin analoguesfrom highly hindered 7-cisisomers of reti-nal.Nature262:416–18
116. Kawamura S, Miyatani S, MatsumotoH, Yoshizawa T, Liu RS. 1980. Photo-chemical studies of 7-cis-rhodopsin atlow temperatures. Nature and propertiesof the bathointermediate.Biochemistry19:1549–53
117. Shichida Y, Kandori H, Okada T,Yoshizawa T, Nakashima N, YoshiharaK. 1991. Differences in the photobleach-ing process between 7-cis- and 11-cis-rhodopsins: a unique interaction changebetween the chromophore and the pro-tein during the lumi-meta I transition.Bio-chemistry30:5918–26
118. Buczylko J, Saari JC, Crouch RK, Pal-czewski K. 1996. Mechanisms of opsin

14 Jan 2003 15:1 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
G PROTEIN-COUPLED RECEPTOR RHODOPSIN 877
activation. J. Biol. Chem.271:20621–30
119. Jang GF, Kuksa V, Filipek S, Bartl F,Ritter E, et al. 2001. Mechanism ofrhodopsin activation as examined withring-constrained retinal analogs and thecrystal structure of the ground state pro-tein.J. Biol. Chem.276:26148–53
120. Bhattacharya S, Ridge KD, Knox BE,Khorana HG. 1992. Light-stable rhodo-psin. I. A rhodopsin analog reconsti-tuted with a nonisomerizable 11-cisretinal derivative.J. Biol. Chem.267:6763–69
121. Jang GF, McBee JK, Alekseev AM, Hae-seleer F, Palczewski K. 2000. Stereoiso-meric specificity of the retinoid cyclein the vertebrate retina.J. Biol. Chem.275:28128–38
122. Ridge KD, Bhattacharya S, NakayamaTA, Khorana HG. 1992. Light-stablerhodopsin. II. An opsin mutant (TRP-265-Phe) and a retinal analog with a non-isomerizable 11-cis configuration form aphotostable chromophore.J. Biol. Chem.267:6770–75
123. Zhukovsky EA, Robinson PR, OprianDD. 1991. Transducin activation by rho-dopsin without a covalent bond to the11-cis-retinal chromophore.Science251:558–60
124. McDowell JH, Williams TP. 1976. Oxida-tion states of four sulfurs of rhodopsin be-fore and after bleaching.Vis. Res.16:643–46
125. Karnik SS, Khorana HG. 1990. Assemblyof functional rhodopsin requires a disul-fide bond between cysteine residues 110and 187.J. Biol. Chem.265:17520–24
126. Ernst OP, Hofmann KP, Sakmar TP. 1995.Characterization of rhodopsin mutantsthat bind transducin but fail to induce GTPnucleotide uptake. Classification of mu-tant pigments by fluorescence, nucleotiderelease, and flash-induced light-scatteringassays.J. Biol. Chem.270:10580–86
127. Franke RR, Konig B, Sakmar TP, Kho-rana HG, Hofmann KP. 1990. Rhodopsin
mutants that bind but fail to activate trans-ducin.Science250:123–25
128. Imamoto Y, Kataoka M, Tokunaga F, Pal-czewski K. 2000. Light-induced confor-mational changes of rhodopsin probedby fluorescent Alexa594 immobilized onthe cytoplasmic surface.Biochemistry39:15225–33
129. Abdulaev NG, Ridge KD. 1998. Light-induced exposure of the cytoplasmic endof transmembrane helix seven.Proc. Natl.Acad. Sci. USA95:12854–59
130. Schertler GF, Hargrave PA. 1995. Projec-tion structure of frog rhodopsin in twocrystal forms.Proc. Natl. Acad. Sci. USA92:11578–82
131. Schertler GF, Hargrave PA. 2000. Prepa-ration and analysis of two-dimensionalcrystals of rhodopsin.Methods Enzymol.315:91–107
132. Schertler GF, Villa C, Henderson R. 1993.Projection structure of rhodopsin.Nature362:770–72
133. Davies A, Gowen BE, Krebs AM,Schertler GF, Saibil HR. 2001. Three-dimensional structure of an invertebraterhodopsin and basis for ordered alignmentin the photoreceptor membrane.J. Mol.Biol. 314:455–63
134. Pogozheva ID, Lomize AL, Mosberg HI.1997. The transmembrane 7-alpha-bundleof rhodopsin: distance geometry calcula-tions with hydrogen bonding constraints.Biophys. J.72:1963–85
135. Baldwin JM, Schertler GF, Unger VM.1997. An alpha-carbon template for thetransmembrane helices in the rhodopsinfamily of G-protein-coupled receptors.J.Mol. Biol. 272:144–64
136. Altenbach C, Klein-Seetharaman J, Cai K,Khorana HG, Hubbell WL. 2001. Struc-ture and function in rhodopsin: mappinglight-dependent changes in distance be-tween residue 316 in helix 8 and residuesin the sequence 60–75, covering the cyto-plasmic end of helices TM1 and TM2 andtheir connection loop CL1.Biochemistry40:15493–500

14 Jan 2003 15:1 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
878 FILIPEK ET AL.
137. Meng EC, Bourne HR. 2001. Recep-tor activation: What does the rhodopsinstructure tell us?Trends Pharmacol. Sci.22:587–93
138. Dartnall HJA. 1968. The photosensitivi-ties of visual pigments in the presence ofhydroxylamine.Vis. Res.8:339–58
139. Honig B, Ebrey T, Callender RH, DinurU, Ottolenghi M. 1979. Photoisomeriza-tion, energy storage, and charge separa-tion: a model for light energy transductionin visual pigments and bacteriorhodopsin.Proc. Natl. Acad. Sci. USA76:2503–7
140. Cooper A. 1979. Energy uptake in the firststep of visual excitation.Nature282:531–33
141. Borhan B, Souto ML, Imai H, ShichidaY, Nakanishi K. 2000. Movement of reti-nal along the visual transduction path.Sci-ence288:2209–12
142. Jager F, Fahmy K, Sakmar TP, SiebertF. 1994. Identification of glutamic acid113 as the Schiff base proton acceptorin the metarhodopsin II photointermediateof rhodopsin.Biochemistry33:10878–82
143. Farrens DL, Altenbach C, Yang K,Hubbell WL, Khorana HG. 1996. Re-quirement of rigid-body motion of trans-membrane helices for light activation ofrhodopsin.Science274:768–70
144. Kim JM, Altenbach C, Thurmond RL,Khorana HG, Hubbell WL. 1997. Struc-ture and function in rhodopsin: rhodopsinmutants with a neutral amino acid at E134have a partially activated conformation inthe dark state.Proc. Natl. Acad. Sci. USA94:14273–78
145. Chen S, Lin F, Xu M, Hwa J, GrahamRM. 2000. Dominant-negative activity ofan alpha(1B)-adrenergic receptor signal-inactivating point mutation.EMBO J.19:4265–71
146. Altenbach C, Cai K, Klein-Seetharaman J,Khorana HG, Hubbell WL. 2001. Struc-ture and function in rhodopsin: map-ping light-dependent changes in distancebetween residue 65 in helix TM1 andresidues in the sequence 306–319 at the
cytoplasmic end of helix TM7 and in he-lix H8. Biochemistry40:15483–92
147. Ernst OP, Meyer CK, Marin EP, HenkleinP, Fu WY, et al. 2000. Mutation of thefourth cytoplasmic loop of rhodopsin af-fects binding of transducin and peptidesderived from the carboxyl-terminal se-quences of transducin alpha and gammasubunits.J. Biol. Chem.275:1937–43
148. Marin EP, Krishna AG, Zvyaga TA,Isele J, Siebert F, Sakmar TP. 2000.The amino terminus of the fourth cyto-plasmic loop of rhodopsin modulatesrhodopsin-transducin interaction.J. Biol.Chem.275:1930–36
149. Hamm HE. 2001. How activated receptorscouple to G proteins.Proc. Natl. Acad.Sci. USA98:4819–21
150. Weitz CJ, Nathans J. 1992. Histidineresidues regulate the transition of pho-toexcited rhodopsin to its active confor-mation, metarhodopsin II.Neuron8:465–72
151. Abdulaev NG, Ngo T, Chen R, Lu Z,Ridge KD. 2000. Functionally discretemimics of light-activated rhodopsin iden-tified through expression of soluble cy-toplasmic domains.J. Biol. Chem.275:39354–63
152. Baldwin PA, Hubbell WL. 1985. Effectsof lipid environment on the light-inducedconformational changes of rhodopsin. 2.Roles of lipid chain length, unsaturation,and phase state.Biochemistry24:2633–39
153. Rath P, DeGrip WJ, Rothschild KJ. 1998.Photoactivation of rhodopsin causes anincreased hydrogen-deuterium exchangeof buried peptide groups.Biophys. J.74:192–98
154. Stenkamp RE, Filipek S, Driessen CAGG,Teller DC, Palczewski K. 2002. Crys-tal structure of rhodopsin: a templatefor cone visual pigments and other Gprotein-coupled receptors.Biochim. Bio-phys. Acta1565:168–82
155. Neitz M, Neitz J, Jacobs GH. 1991.Spectral tuning of pigments underlying

14 Jan 2003 15:1 AR AR177-PH65-35.tex AR177-PH65-35.sgm LaTeX2e(2002/01/18)P1: IBC
G PROTEIN-COUPLED RECEPTOR RHODOPSIN 879
red-green color vision.Science252:971–74
156. Stockman A, Sharpe LT, Merbs S,Nathans J. 2000. Spectral sensitivities ofhuman cone visual pigments determinedin vivo and in vitro. Methods Enzymol.316:626–50
157. Merbs SL, Nathans J. 1992. Absorptionspectra of human cone pigments.Nature356:433–35
158. Merbs SL, Nathans J. 1992. Absorptionspectra of the hybrid pigments responsi-ble for anomalous color vision.Science258:464–66
159. Nathans J. 1994. In the eye of the be-holder: visual pigments and inherited vari-ation in human vision.Cell 78:357–60
160. Sharpe LT, Stockman A, Jagle H, KnauH, Klausen G, et al. 1998. Red, green,and red-green hybrid pigments in the hu-man retina: correlations between deducedprotein sequences and psychophysicallymeasured spectral sensitivities.J. Neu-rosci.18:10053–69
161. Merbs SL, Nathans J. 1993. Role ofhydroxyl-bearing amino acids in differ-entially tuning the absorption spectra ofthe human red and green cone pigments.Photochem. Photobiol.58:706–10
162. Asenjo AB, Rim J, Oprian DD. 1994.Molecular determinants of human red/green color discrimination.Neuron 12:1131–38
163. Kochendoerfer GG, Kaminaka S, MathiesRA. 1997. Ultraviolet resonance Ramanexamination of the light-induced protein.Biochemistry36:13153–59
164. Marinissen MJ, Gutkind JS. 2001. G–protein-coupled receptors and signalingnetworks: emerging paradigms.TrendsPharmacol. Sci.22:368–76
165. Gether U. 2000. Uncovering molecularmechanisms involved in activation of Gprotein-coupled receptors.Endocr. Rev.21:90–113
166. Angers S, Salahpour A, Bouvier M. 2002.Dimerization: an emerging concept forG protein-coupled receptor ontogeny andfunction.Annu. Rev. Pharmacol. Toxicol.42:409–35
167. Ballesteros JA, Shi L, Javitch JA. 2001.Structural mimicry in G protein-coupledreceptors: implications of the high-re-solution structure of rhodopsin forstructure-function analysis of rhodopsin-like receptors.Mol. Pharmacol. 60:1–19
168. Brady JGP, Stouten PFW. 2000. Fast pre-diction and visualization of protein bind-ing pockets with PASS.J. Comp.-AidedMol. Des.14:383–401
169. Ballesteros J, Palczewski K. 2001. Gprotein-coupled receptor drug discovery:implication from the crystal structure ofrhodopsin.Curr. Opin. Drug Disc. Dev.4:561–74

22 Jan 2003 21:38 AR AR177-35-COLOR.tex AR177-35-COLOR.SGM LaTeX2e(2002/01/18)P1: GDL
Fig
ure
1M
olec
ular
pack
ing
ofrh
odop
sin
(A
)an
dse
nsor
yrh
odop
sin
(B
)in
thei
rcr
ysta
ls.
One
asym
met
ricun
itco
ntai
nstw
orh
odop
sin
mol
ecul
es.T
hece
ntra
ldim
eris
show
nas
aco
ntin
uous
surf
ace,
whe
reas
the
rest
are
show
nas
ribbo
ns.V
iew
sar
efr
omdi
ffere
ntcr
ysta
llogr
aphi
cX
-,Y-
,Z-a
xes
for
rhod
opsi
n(2
7),a
ndX
-,an
dZ
-for
sens
ory
rhod
opsi
n.T
heda
tafo
rsen
sory
rhod
opsi
nw
ere
take
nfr
omP
rote
inD
ata
Bas
e(1
H68
).C
oord
inat
es:M
odel
sof
rhod
opsi
nan
dth
eho
mol
ogy
mod
els
ofco
nepi
gmen
tsha
vebe
ende
posi
ted
inth
eP
rote
inD
ata
Ban
k(P
DB
)(id
entifi
ers
1F88
,1H
ZX
,1K
PN
,1K
PW
,1K
PX
).

22 Jan 2003 21:38 AR AR177-35-COLOR.tex AR177-35-COLOR.SGM LaTeX2e(2002/01/18)P1: GDL
Fig
ure
2R
ibbo
ndr
awin
gsof
rhod
opsi
n.H
elic
esI–
VIII
are
colo
red
asa
spec
trum
ofvi
sibl
elig
htfr
ombl
ue(h
elix
I)to
red
(hel
ixV
III),
and
two
orie
ntat
ions
are
show
n.P
alm
itoyl
chai
nsan
dol
igos
acch
arid
egr
oups
show
nus
ing
ball-
and-
stic
km
odel
s.

22 Jan 2003 21:38 AR AR177-35-COLOR.tex AR177-35-COLOR.SGM LaTeX2e(2002/01/18)P1: GDL
Fig
ure
3Tw
o-di
men
sion
alm
odel
ofbo
vine
rhod
opsi
n.B
urie
dre
sidu
esar
esh
own
ingr
ay.O
rang
e,re
d,an
dvi
olet
deno
tesu
rfac
ere
sidu
es.
Res
idue
sin
cont
actw
ithpo
lar
head
grou
psof
the
mem
bran
eon
the
cyto
plas
mic
side
are
oran
ge.S
imila
rre
sidu
eson
the
extr
acel
lula
rsi
dear
ere
d.A
llot
her
surf
ace
resi
dues
are
viol
et.

22 Jan 2003 21:38 AR AR177-35-COLOR.tex AR177-35-COLOR.SGM LaTeX2e(2002/01/18)P1: GDL
Figure 4 Hydrophobic and hydrophilic interactions between rhodopsin helices (loopsnot included) viewed from the intracellular side. Hydrophobic interactions are shownas orange fuzzy bars together with numbers denoting their strength. Hydrophilic inter-actions are shown as red lines. List of hydrophilic interactions between helices includehelices I–II, Asn55-Asp83; helices I–VII, Tyr43-Phe293, Asn55-Ala299; helices I–VIII,Gln64-Thr320; helices II–III, Asn78-Ser127; helices II–IV: Tyr74-Glu150, Asn78-Thr160,Asn78-Trp161; helices III–V, Cys140-Thr229, Glu122-Trp126-His211-Tyr206 network; he-lices III–VI, Arg135-Glu247; helices III–VII, Glu113-Lys296 (retinal); helices IV–V,Ala166-Tyr206; helices VI–VII, Cys264-Thr297; and helices VII–VIII, Ile307-Arg314.
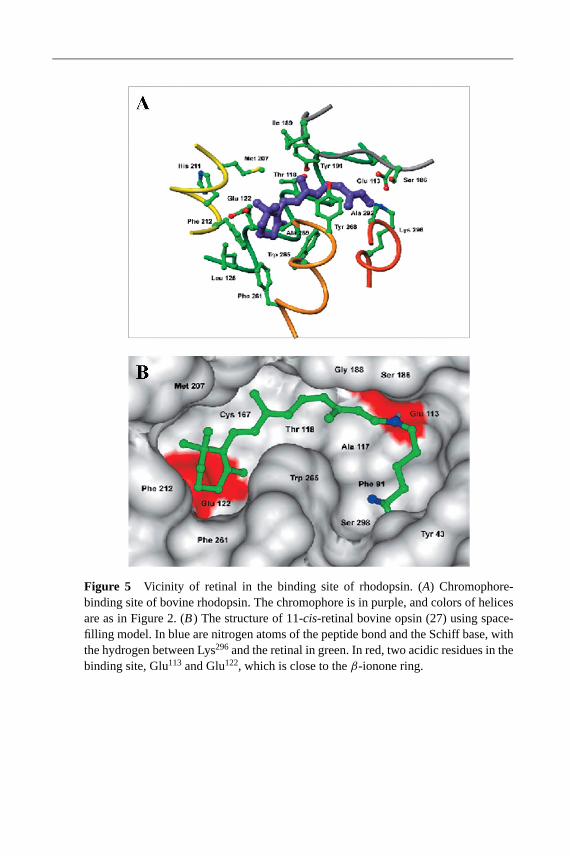
22 Jan 2003 21:38 AR AR177-35-COLOR.tex AR177-35-COLOR.SGM LaTeX2e(2002/01/18)P1: GDL
Figure 5 Vicinity of retinal in the binding site of rhodopsin. (A) Chromophore-binding site of bovine rhodopsin. The chromophore is in purple, and colors of helicesare as in Figure 2. (B) The structure of 11-cis-retinal bovine opsin (27) using space-filling model. In blue are nitrogen atoms of the peptide bond and the Schiff base, withthe hydrogen between Lys296and the retinal in green. In red, two acidic residues in thebinding site, Glu113 and Glu122, which is close to theβ-ionone ring.

22 Jan 2003 21:38 AR AR177-35-COLOR.tex AR177-35-COLOR.SGM LaTeX2e(2002/01/18)P1: GDL
Figure 6 The DRY region (A) and NPXXY region (B) are believed to be involved inthe activation and transformation of photoactivated rhodopsin. Note that in rhodopsin, thesequence of the DRY region is ERY.
Figure 7 Interactions of helix VI. Hydrophobic contacts of helix VI shown as whiteresidues and hydrophilic interactions represented by lines. (A) Interaction of helix VIwith helices II and III. (B) Interaction of helix VI with helices V and VII.

22 Jan 2003 21:38 AR AR177-35-COLOR.tex AR177-35-COLOR.SGM LaTeX2e(2002/01/18)P1: GDL
Fig
ure
8D
iffer
entr
egio
nsof
cone
pigm
ents
.(A)
DR
Yre
gion
and
the
chro
mop
hore
-bin
ding
site
sof
blue
and
red/
gree
nco
neop
sins
.(B
)The
vici
nity
ofth
ech
rom
opho
refr
omle
ftto
right
:blu
e,gr
een,
and
red
pigm
ents
.The
num
berin
gof
corr
espo
ndin
gre
gion
sis
base
don
the
bovi
nerh
odop
sin
prim
ary
sequ
ence
.

22 Jan 2003 21:38 AR AR177-35-COLOR.tex AR177-35-COLOR.SGM LaTeX2e(2002/01/18)P1: GDL
Fig
ure
9F
unct
iona
ldom
ains
ofrh
odop
sin
that
are
high
lyco
nser
ved
amon
gm
embe
rsof
the
GP
CR
supe
rfam
ily.
(A
)Lo
catio
nof
thes
edo
mai
nsin
the
thre
e-di
men
sion
alst
ruct
ure
ofrh
odop
sin.
(B
)C
lose
-up
ofth
ecr
itica
lreg
ions
:DR
Yre
gion
(a,
pa
ne
lA,i
nrh
odop
sin
ER
Y,bl
ue);
palm
itoyl
grou
ps(b,
pa
ne
lA,
red)
;N
PX
XY
regi
on(c,
pa
ne
lA,
light
blue
);ch
rom
opho
re;
Pro
kink
inhe
lixV
I,Ly
s29
6(d
,pa
ne
lA,
viol
et);
disu
lfide
brid
ge(e,
pa
ne
lA,y
ello
w);
and
olig
osac
char
ide
moi
etie
s(
f,p
an
elA
,bro
wn)
.