glycoconjugates glycobiology
TRANSCRIPT

Glycoconjugates
Structural diversity
Diversity of functions
• Structural / modulation
• Recognition
• Glycoproteins
• Glycolipids
• Proteoglycans
Altered glycosylation and glycosylation
defects are observed in many pathological
situations (inflammation, cancer, congenital
disorders of glycosylation, �)
GLYCOBIOLOGYStudy of structure, synthesis and biology of
sugars, particularly the sugar chains that are
attached to proteins and lipids
1

:
Although proteins comprise the largest fraction of a cell's dry mass, it is
estimated that more than half are modified with glycans, lipids or other
metabolites.
Prescher and Bertozzi, 2005, Nat. Chem. Biol. 1, 13-21
Post-translational modifications of proteins are important regulators
of protein function
2

ER
Golgi
M.E. Taylor & K. Drickamer (Introduction to Glycobiology -2003- Oxford University Press)
Location of glycoconjugates in multicellular organisms
3

.
BIOSYNTHESIS OF GLYCANS : FOCUS ON THE
LARGE GLYCOSYLTRANSFERASE FAMILY
Structural and Functional aspects
Prof. Christelle BRETON
4

Photo credit : Barbieri, Chenu, Francillon, Vinçon, D.R. AEPI
This slide shoud always be used with the AEPI logo. Any modification of the slide has to be expressly notified to the AEPI
Grenoble
5

Centre de Recherches sur les
Macromolécules Végétales
Glycosciences
6

Structure-Function
Studies of Lectins and
Glycosyltransferases
Molecular Glycobiology at CERMAV
7

.
1- General features
• General reaction
• Cellular localization & topology
• Glycosylation events in the Golgi
• Take home message (1)
2- Classification
• Processivity / stereochemistry
• Classification
• GT repertoire
• Take home message (2)
3- Structure
• Structural information (current picture)
• GT-A fold / DxD motif
• GT-B fold
• Other folds
• Take home message (3)
8
GLYCOSYLTRANSFERASE course : Contents
4- Reaction mechanisms
• Inverting/retaining reaction mechanisms
• Ordered binding in GT-A enzymes
• additional difficulties
• Take home message (4)
5- Future directions

.UDP-sugars
Glucose
Galactose
N-acetylglucosamine
N-acetylgalactosamine
Glucuronic acid
Xylose
...
CMP-sugars
Sialic acids
GDP-sugars
Fucose
Mannose
XDP-sugar + OH-Acceptor sugar-O-Acceptor + XDP
GT
GLYCOSYLTRANSFERASES (Leloir enzymes*)
specificity for both the donor sugar and acceptor substrates
* In honor of Luis L. Leloir who discovered the first nucleotide sugar (Nobel Prize in Chemistry in 1970)9
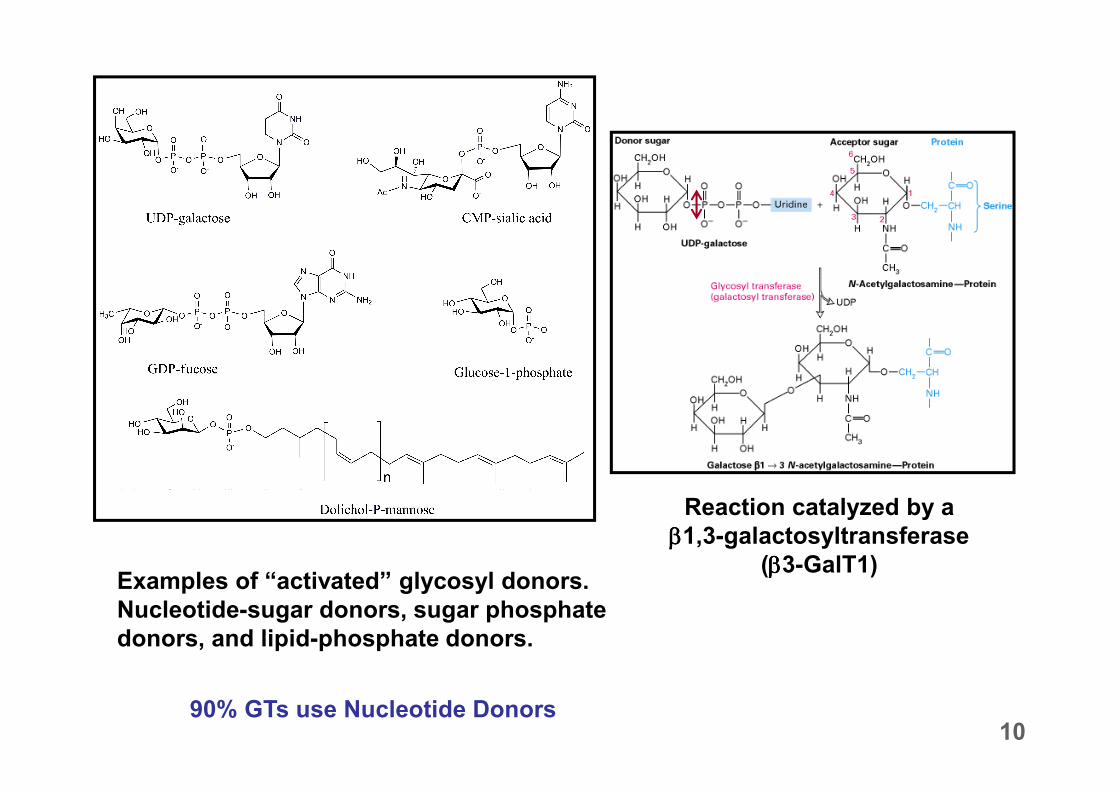
Examples of “activated” glycosyl donors.
Nucleotide-sugar donors, sugar phosphate
donors, and lipid-phosphate donors.
90% GTs use Nucleotide Donors
Reaction catalyzed by a
ββββ1,3-galactosyltransferase
(ββββ3-GalT1)
10

.
� ER, Golgi (membrane proteins )
� cytosol and nucleus (soluble forms)
� biological fluids (soluble forms) ??
� plasma membrane (membrane proteins)
Eukaryotes
Cellular localization and topology
GLYCOSYLTRANSFERASES
Secretory pathway
Most of the complex glycans are
built into the ER and GOLGI
11

.
Topologies of glycosyltransferases in the ER and Golgi
RE lumen
Cytoplasm
GT using a Dolichol-P-sugar
integral membrane protein
TMD
NH2
Cytoplasm
Golgi lumenstem
catalytic domain
Typical Golgi GT using a nucleotide-sugar
Type II membrane protein
12

.
TMD stem
Add-on domain
(i.e. Lectin domain)
lumen
N-
N-
N-
Catalytic domain
Topology of Golgi glycosyltransferases
Stem variable length
may modulate in vivo GT activity
« cleavable »
Catalytic domain globular, minimal active structure,
typically ~300-350 aa
Breton et al. (2000) Biochimie
����
13

퀀
Glycosylation events in Golgi
Nucleotide-sugars are
synthesized in the cytosol
and then transported into
the Golgi by specific
transporters
(CMP-NeuAc synthesized in
the nucleus)
GT
Cytoplasme
14

.
Glycosylation events in Golgi
GT localization ?
Supramolecular organization ?
De Graffenreid & Bertozzi, 2004Brockhausen, 2006
O-glycans N-glycans
15

.
Although the mechanisms of Golgi enzyme
localization are still under debate, both the
distribution of these enzymes among the Golgi
cisternae and specific association among
glycosylation enzymes can dictate the overall glycan
structures (glycome) produced by a cell.
TAKE HOME MESSAGE (1)
16

.
1- General features
• General reaction
• Cellular localization & topology
• Glycosylation events in the Golgi
• Take home message (1)
2- Classification
• Processivity / stereochemistry
• Classification
• GT repertoire
• Take home message (2)
3- Structure
• Structural information (current picture)
• GT-A fold / DxD motif
• GT-B fold
• Other folds
• Take home message (3)
17
GLYCOSYLTRANSFERASE course : Contents
4- Reaction mechanisms
• Inverting/retaining reaction mechanisms
• Ordered binding in GT-A enzymes
• additional difficulties
• Take home message (4)
5- Future directions

.
Processive enzymes : transfer multiple sugar residues to the acceptor
(polysaccharide synthases,.....)
Non-processive enzymes : transfer a single sugar residue to the
acceptor
PROCESSIVITY
GLYCOSYLTRANSFERASES N
18

. O
OOHHO
HO
HOP
OP
O
O O
O O
ON
NH
HO OH
O
O
OOHHO
HO
HO
RRRReeeettttaaaaiiiinnnniiiinnnngggg GGGGllllyyyyccccoooossssyyyyllllttttrrrraaaannnnssssffffeeeerrrraaaasssseeee
IIIInnnnvvvveeeerrrrttttiiiinnnnggggGGGGllllyyyyccccoooossssyyyyllllttttrrrraaaannnnssssffffeeeerrrraaaasssseeee
OOHHO
HOOH
O
O
HO
OH
OH
OH
HO
O
HO
OH
OH
OH
HO
O
HO
OH
OH
OH
O
O
HO
OH
OH
OH
Transfer of a monosaccharide
from a donor to an acceptor:
Reaction can occur with retention
or inversion of configuration of
the transferred sugar
STEREOCHEMISTRY OF THE
GLYCOSIDIC LINKAGE
19

.
Galactosyltransferases (αααα3-GalTs, ββββ4-GalTs, ...)
Fucosyltransferases (αααα2-FucTs, αααα3-FucTs, αααα6-FucTs, ...)
Sialyltransferases (ST6Gal, ST3Gal, ST8Sia, ....)
.........
There is no or limited sequence homology among the different GT families
or even, within one family, among members catalyzing different reactions
They are primarily classified according to the type of sugar that they transfer
GLYCOSYLTRANSFERASES N
20

.
Classification based on amino acid sequence similarity, correlates with
(i) protein folding
(ii) enzyme mechanism
Glycosyltransferases .... CAZy classification
▪ Two very large families (~half of total number of GT entries)
inverting GT2 and retaining GT4 – considered as the ancestral
families
▪ Many families are polyspecific
~ 100,000 known and
putative GT sequences
distributed over 90
families
http://www.cazy.org/
21
October 2012

훀
� New GT families are exclusively created based on the
availability of at least one biochemically-characterized protein
member
Discovery of new GT families
� Experimental investigation (i.e. Rumi GT90)
� Bioinformatics and biochemical characterization
(i.e. plant XylTs, GT77)
� The number of GT families in CAZy is continually increasing
with the discovery of new GT genes (27 families in 1997, more
than 90 in 2012)
22

훀
235 GT genes
43 families
~ 80 % annotated genes
The most populated families
GT1 : UGTs
GT7 : ββββ4-GTs
GT27 : ppGalNAcTs
GT29 : SiaTs
GT31 : ββββ3-GTs
A large repertoire of genes is required for glycan assembly and function in
multicellular organism : 1-2% of the genes of an organism code for GTs
GT repertoire ....
23

.
▪ Only 1 plant specific GT family (GT37)
~ 460 GT genes
42 families
20 % are annotated
▪ Several highly populated GT families
GT1 : UGT-family (glycosylation of secondary metabolites)
GT2, 8, 31, 47 : biosynthesis of cell wall PS
Plants tend to have far more GT genes than
any other organism sequenced to date
24

�Ϟ
� A huge number of putative GT genes have been identified
through large genome sequencing projects (currently ~100,000)
� Large number of sequence-based GT families (> 90)
� The vast majority of these sequences (more than 90%) are
uncharacterized open-reading frames (particularly bacterial and
plant genomes)
� This is a challenging task for glycobiologists to assign
functions to the many uncharacterized GT sequences
TAKE HOME MESSAGE (2)
25

�Ϟ
1- General features
• General reaction
• Cellular localization & topology
• Glycosylation events in the Golgi
• Take home message (1)
2- Classification
• Processivity / stereochemistry
• Classification
• GT repertoire
• Take home message (2)
3- Structure
• Structural information (current picture)
• GT-A fold / DxD motif
• GT-B fold
• Other folds
• Take home message (3)
8
GLYCOSYLTRANSFERASE course : Contents
4- Reaction mechanisms
• Inverting/retaining reaction mechanisms
• Ordered binding in GT-A enzymes
• additional difficulties
• Take home message (4)
5- Future directions

.
���� Structural information for 39 CAZY families
���� Only two general folds, termed GT-A and GT-B (and variants), have
been observed for all structures of nucleotide-sugar-dependent GTs
solved to date
GT-A fold GT-B fold
Glycosyltransferases .... 3D structures
26

�Ϟ
GT-A and variants
2, 6, 7, 8, 13, 14, 15, 27,
29, 31, 42, 43, 44, 55,
64, 78, 81, 88
12, 16, 17, 21, 24, 25, 32,
34, 40, 45, 49, 54, 60, 62,
67, 69, 71, 73, 74, 75, 77,
82, 84, 92
GT-B and variants
1, 3, 4, 5, 9, 10, 20, 23
28, 30, 35, 41, 52, 63,
65, 68, 70, 72, 80
11, 18, 19, 33, 37, 38,
47, 56, 61, 90
~ 80 % of the current GT families are known, or predicted, to adopt a GT-A or GT-B fold (or variants)
3D
Predicted*
Breton et al. Biochem. Soc. Symp. (2002)
Breton et al. Glycobiology (2006)
Breton et al. COSB (2012)* Based on 3D-PSSM and PHYRE programs
(Bennett-Lovsey et al., 2008; Kelley et al., 2000)
(37)
(34)
27

͵
There is no correlation between the overall
fold of the GT and its mechanism of action
since both inverting and retaining
enzymes of both fold types (GT-A and GT-
B) are known.
28

�Ϟ
GT-A fold
a single domain
αααα/ββββ/αααα structure
mixed ββββ-sheet (3214657)
All of the strictly metal ion-dependent GTs for which structures have
been determined to date have been of this type and include a DxD
Metal Binding motif (Mg2+, Mn2+)* One exception (GT14)
29

眐Ϛ
Ubiquity and structural basis of the DxD motif in the GT-A fold
Variations in the DxD sequence : DVD, DID, DDD, EDD, EED, TDD, DxH, EPD, N.
cation-dependent GTs (Mn2+, Mg2+,…)
30

禠Ϝ
Cst II - sialyltransferase
Campylobacter jejuni
// ββββ-sheet (8712456)
no DxD motif
Canonical GT-A
mixed ββββ-sheet (3214657)
DxD motif
GT-A like
GT-A variant
GT42
New type of fold !31

禠Ϝ
GT-B fold
2 Rossmann-type
domain assembly
αααα/ββββ/αααα structure
// β-sheet (3214567)
The catalytic site is located in a cleft between the two domains
Metal ion-independent GTs
32

禠Ϝ
GT-B variant
Coiled coil region
(N-term)
SH3 domain
(Cterm)
Catalytic domain
Rossmann
domain in the C-
term of catalytic
domain
GT23
Human αααα6-Fucosyltransferase (FucT-VIII)
33

禠Ϝ
GT-B variant
Human O-ββββ-GlcNAcT (OGT)
Int-D
TPR
GT41• Unique and reversible
modification
• Found on a myriad of nuclear and
cytoplasmic proteins
• Extensive cross-talk with
phosphorylation
Hart et al., (2007) Nature, 446: 1017−1022
Lazarus et al., (2011) Nature, 469:564-56734

禠Ϝ
New folds are observed for GTs using lipid-phosphate sugar donors
No Rossmann fold !!
Peptidoglycan glycosyltransferase(Lovering et al., 2007, Science)
GT51
GT domain: bacteriophage λλλλ-lysozyme-like fold !!
Staphylococcus aureus
35

㐐
The process of N-linked protein glycosylation
Schwarz and Aebi (2011) COSB, 21:576-582 36

㐐
| N AT U R E | V O L 4 7 4 | 1 6 J U N E 2 0 1 1
GT66
The first structure of a membrane-embedded GT !
37

㐐
3D structure of PglB (C. jejuni) GT66
Lizak et al., (2011) Nature, 474, 350-355.38

㐐
Virtually nothing is known about the structures and mechanisms of
these large membrane-embedded GTs. The successful crystallization of
PglB now opens new avenues to fill the gap.
� The large GT family is characterized by a conserved 3D architecture
(GT-A, GT-B folds and variants)
The small variety of folds observed for GTs (constraint in nucleotide-sugar
binding ?) is compensated by a large structural variability in the acceptor
binding domain: «Functional plasticity» which allows fine-tuning with
respect to the acceptor.
� Novel folds are expected for GTs that use a lipid-phosphate donor
TAKE HOME MESSAGE (3)
39

㐐
1- General features
• General reaction
• Cellular localization & topology
• Glycosylation events in the Golgi
• Take home message (1)
2- Classification
• Processivity / stereochemistry
• Classification
• GT repertoire
• Take home message (2)
3- Structure
• Structural information (current picture)
• GT-A fold / DxD motif
• GT-B fold
• Other folds
• Take home message (3)
8
GLYCOSYLTRANSFERASE course : Contents
4- Reaction mechanisms
• Inverting/retaining reaction mechanisms
• Ordered binding in GT-A enzymes
• additional difficulties
• Take home message (4)
5- Future directions

㐐
Reaction mechanisms of GTs
In contrast to the well-characterized
catalytic mechanisms used for glycosidic
bond hydrolysis (glycosidases), the
mechanisms for glycoside bond formation
remain less clear.
For a review, see Lairson et al., (2008) Ann. Rev. Biochem., 77:521-555
Breton et al., (2012) COSB40

㐐
Inverting Glycosyltransferase Mechanism
A direct-displacement SN2-like reaction via a single oxycarbenium ion
Catalytic base (i.e. Asp, Glu, His) deprotonates acceptor OH
From Lairson et al. (2008) Ann Rev Biochem41

負Ϝ
Retaining Glycosyltransferase Mechanism: currently two hypotheses
SN2-like double displacement
mechanism with formation of a
covalently bound glycosyl-enzyme
intermediate (by analogy to
glycosylhydrolases)
Soya et al., (2011) Glycobiology, 21 (5)
From Lairson et al. (2008) Ann Rev Biochem
SNi-like mechanism
Front-face nucleophilic attack
involving hydrogen bonding between
leaving group and the acceptor
nucleophile
Lee et al., (2011) Nature Chem. Biol., 7
42

負Ϝ
The « closed » active conformation
of glycosyltransferases
LgtC
ααααGalT
Persson et al, Nature Struct. Biol., 8 (2001) 166
Base of UDP-Gal
Acceptordisaccharide
Reaction mechanism for GTs
of the GT-A family
Ordered binding of donor and
acceptor substrates, linked to
a donor substrate-induced
conformational change
Binding of a distorted conformation
of nucleotide sugar may be important
for catalysis
Ret Inv Inv
43

負Ϝ
Superimposition of open (free
enzyme) and closed (upon UDP-Gal
binding) forms
openclose
Additional difficulties for glycosyltransferases
Loop movement domain movementGT-A GT-B
Superimposition of open state
(solid) and closed conformation
(transparent)
Buschiazzo et al. (2004)Gastinel et al. (2001); Boix et al., (2001)
αααα3-GalT Glycogen synthase GT5 GT6
44

負Ϝ
Crystallographic and modelling studies should
allow for a better understanding of molecular
bases responsible for substrate specificityN
The blood group synthases as a case of study !
GlycosyltransferasesN still puzzling
45

負Ϝ
OOHHO
OO
OR
OOHHO
HOOH
O
HOOH
OH
OOHHO
OO
OR
OOHHO
HONH
O
HOOH
OH
OOHHO
HOO
OR
O
HOOH
OH
OH3C
O
OOHHO
HONH
OH3C
P
O
OO
P
O
OO
ON
NH
O
O
OHHO
O
OOHHO
HOOH P
O
OO
P
O
OO
ON
NH
O
O
OHHO
UDP-GalNAc UDP-Gal
GTA GTB
O(H) Precursor
Blood Group A Blood Group B
Blood Group A & B Structures Made by Closely Related GTs
Yamamoto, Hakomori (Nature, 1990)Both enzymes adopt a GT-A fold
46

負Ϝ
– Highly homologous enzymes differing at only four critical amino acids
out of a total of 354 residues
– Alteration of these four crucial amino acid residues converts the
specificity from GTA to GTB
AAAA
BBBB
Blood group synthases GTA/GTB
Molecular basis for nucleotide sugar specificity ?
UDP-Gal vs UDP-GalNAc
47

負Ϝ
L266
E303
D302
G268
M266
E303
D302
A268
GTA GTB
S185 S185
αααα3-GalNAcT αααα3-GalT
Homology Modelling
UDP-Gal vs UDP-GalNAc
Only 2 of the 4 critical residues in binding site (L266M, G268A)
M266 + A268 excludes UDP-GalNAc (steric conflict)
G268 accomodation of N-acetamido group
GTA should utilize UDP-Gal ???
Heissigerova et al. Glycobiology, 13 (2002) 377
GT6 (GT-A fold)
48

負Ϝ
UDP-Gal vs UDP-GalNAc
Still unclear !!!
What can we learn about substrate binding and catalysis
of blood group GTs ?
Crystallographic studies
Modelling studies
Natural mutations in blood banks
Mutagenesis (chimeric enzymes)
Flexibles enzymes
Mutagenesis unpredictable
Much work to be done to understand catalysis
Monica Palcic and coll (in Alberta and Copenhagen)49

.A better understanding of catalytic mechanisms of GTs together with
much more structural information is needed, for the rational design of
specific inhibitors as well as for engineering purposes to expand the
biotechnological uses of GTs.
� Catalytic mechanisms are still poorly understood
• Movements of loops and/or domains involved in the binding
• Flexibility
TAKE HOME MESSAGE (4)
50

負Ϝ
Glycosyltransferases
“in vivo”
- Functional organization in Golgi
* supramolecular organisation
- Spatial and temporal expression of
GTs in physiological and pathological
situations (Glycomics)
“in vitro”
- “Structure/function” relationships
* new 3D structures
* mechanism of action, inhibitors
- “ORFeome” of GTs
* heterologous expresssion
* library of acceptors
Biotechnological applications
- Enzymatic synthesis of bioactive oligosaccharides
- Production of recombinant GP for therapeutic use
- GTs and medicine (diagnostic tools, new therapies )
- GT engineering (new catalysts with improved capabilities)
- Oligo/polysaccharide engineering (modification of functional
properties)
.....
Future directions N
51

負ϜTHE END

負Ϝ
Database development at Cermav : Glyco3D

�ҕ
Collaborations
Eric Maréchal
(CEA-Grenoble)
Emmanuel Bettler
(IBCP Lyon)
Jan Mucha
(Bratislava)
Soren B. Engelsen
(Univ. Copenhagen)
Monica Palcic & Ole Hindsagaul
(CRL- Copenhagen)
CERMAV - GT team
Christelle Breton
Anne Imberty
Aline Thomas
Olivier Lerouxel
Valerie Chazalet
Post-doc and PhDs
Magali Audry
Gaelle Batot
Peter Both
Sara Fasmer Hansen
Charlotte Jeanneau
Joana Rocha
Funding
French ANR
CEE

�ҕ
Test Arabidopsis proteome
HMMSearch
(1D level)
PSI-BLAST
+
Structural overlap
(2D level)
ProFit
(3D level)
> 150 new candidate sequences
Hansen et al. (2009) J. Proteome Res.
Arabidopsis proteome
30690 sequences
1- Removal of large
sequences(>1000 aa)
29457 sequences
2- Removal of
sequences with a cTP or
mTP (TargetP)
21755 sequences
3- Removal of proteins
with no TMD (TMHMM)
5666 sequences
4- Removal of CAZy
accessions (460 seqs)
Test Arabidopsis proteome
(5315 protein sequences)
Bioinformatics strategy
Use of several remote homology detection methods to search the
« test Arabidopsis proteome » for new candidate GT genes
+ Chemometrics
Hansen et al., (2010) Mol. Biosystems

�ҕ
Evaluation of candidate GT sequences (~150)…
Close examination of each sequence
� Annotation in databases
� Blast and Psi-BLAST
� TMHMM
� Phyre (fold recognition)
� Hydrophobic Cluster Analysis method (HCA)
~ 20 strong candidate genes*
harbouring a clear GT signature
* Distantly related to GT4, GT14, GT23, GT65
In any case, the validation step requires the critical expertise of the biologist

犐ҕ
The populations of the various GT families show
wide variations, with two large families, the
inverting GT2 and retaining GT4, accounting for
about half of the total number of GT entries, and
which can be considered as the most ancestral
families for which enzymes of both
stereochemistries have evolved
(Martinez-Fleites et al., 2006).

犐ҕ
number of sequences
GT2 (> 13,000 seqs)
GT4 (> 10,000 seqs)
GT63 (1 seq)
origin and function
GT2 Cellulose synthase, chitin synthase, Dol-P-Man/Glc synthase
GlcTs, GalTs, GalfTs, GnTs, RhaT, ...
animal, plant, yeast, bacteria,Q
GT27 pp-αααα-GalNAcTs (mucin-type glycosylation)
animal
Glycosyltransferases …. CAZy
Large differences among GT families
8 families have > 1000 sequences
13 families have < 20 sequences
50% of total seqs

犐ҕ
GLYCOSYLTRANSFERASES and biosynthesis of glycans
- Structure-function studies and engineering of GTs
- Biogenesis of PCW polysaccharides
Specific recognition of carbohydrates by LECTINS
- Lectins and bacterial infections
- Development of anti-adhesive drugs
- Lectins and biodiversity
New tools in Glycosciences
- Functionalized surfaces, gold nanoparticles
- Glyco-bioinformatics
- Modelling sulfated polysaccharides
Group Leader
A. ImbertyMOLECULAR GLYCOBIOLOGY at CERMAV

�Ϣ
A better understanding of catalytic mechanisms of GTs together with much more
structural information is needed, however, for the rational design of specific inhibitors as
well as for engineering purposes to expand the biotechnological uses of GTs.
the limiting factor in developing carbohydrate-based compounds for clinical application
was the high cost and complexity of producing glycans in large quantities.
the use of metabolically engineered bacteria that overexpress heterologous GTs has
become a powerful method for the large-scale production of complex glycans at low
cost (Endo and Koizumi, 2000; Fierfort and Samain, 2008). Glycobiology research will
greatly benefit from these major advances in cost-effective technologies for
carbohydrate synthesis.
� Catalytic mechanisms are still poorly understood
Movements of loops and/or domains involved in the binding