gluconeogenesis & control of the blood glucose
TRANSCRIPT

Gluconeogenesis & Control of the Blood Glucose
Al-Sham Private UniversityFaculty Of Pharmacy
Lecturer Prof. Abboud Al-Saleh

BIOMEDICAL IMPORTANCE• Gluconeogenesis is the term used to include all pathways
responsible for converting noncarbohydrate precursors to glucose or glycogen.
• The major substrates are the glucogenic amino acids , lactate, and glycerol.
• Liver and kidney are the major gluconeogenic tissues.
• Gluconeogenesis meets the needs of the body for glucose when carbohydrate is not available in sufficient amounts from the diet or from glycogen reserves.
• A supply of glucose is necessary especially for the nervous system and erythrocytes.
• Failure of gluconeogenesis is usually fatal.

• Hypoglycemia causes brain dysfunction, which can lead
to coma and death.
• Glucose is also important in maintaining the level of
intermediates of the citric acid cycle even when fatty
acids are the main source of acetyl-CoA in the tissues.
• gluconeogenesis clears lactate produced by muscle and
erythrocytes and glycerol produced by adipose tissue.

Thermodynamic Barriers Preventing a Simple Reversal of Glycolysis
• Three nonequilibrium reactions catalyzed by
• hexokinase,
• phosphofructokinase,
• pyruvate kinase
• prevent simple reversal of glycolysis for glucose synthesis

1- PYRUVATE & PHOSPHOENOLPYRUVATE
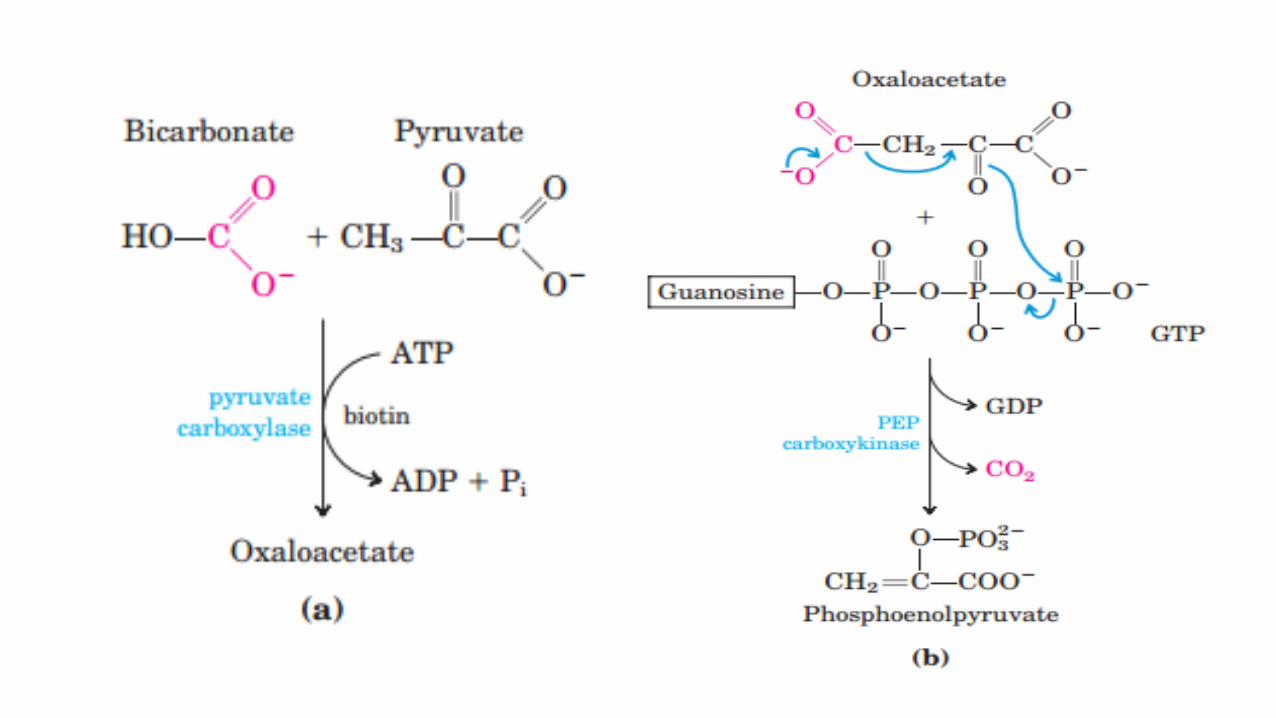
• Mitochondrial pyruvate carboxylase catalyzes the carboxylation of pyruvate to oxaloacetate, an ATP-requiring reaction in which the vitamin biotin is the coenzyme.
• Biotin binds CO2 from bicarbonate as carboxybiotin prior to the addition of the CO2 to pyruvate.
• A second enzyme, phosphoenolpyruvate carboxykinase, catalyzes the decarboxylation and phosphorylation of oxaloacetate to phosphoenolpyruvate using GTP as the phosphate donor.
• Thus, reversal of the reaction catalyzed by pyruvate kinase in glycolysis involves two endergonic reactions.

2- FRUCTOSE 1,6-BISPHOSPHATE & FRUCTOSE 6-PHOSPHATE
conversion of fructose 1,6-bisphosphate to fructose 6-phosphate
fructose-1,6-bisphosphatase is an allosteric enzyme, inhibited by AMP andactivated by ATP.
It is present in liver, kidney, and skeletal muscle but is probably absent fromheart and smooth muscle.
Fructose-6-phosphate
C
CH2 OP O32 -
O
HHO
OHH
OHH
CH2 OH
Fructose-1,6-bisphosphate
C
CH2 OP O32 -
O
HHO
OHH
OHH
CH2 OP O32 -
fructose-1,6-bis-
phosphatase
H2 O P i

3- GLUCOSE 6-PHOSPHATE & GLUCOSE
• It is catalyzed by glucose-6-phosphatase.
• It is present in liver and kidney but absent from muscle and adipose tissue, which, therefore, cannot export glucose into the
bloodstream.

GLUCOSE1-PHOSPHATE & GLYCOGEN
• The breakdown of glycogen to glucose 1-phosphate is catalyzed by phosphorylase.
• Glycogen synthesis involves a different pathway via uridine diphosphate glucose and glycogen synthase(figure).


Propionate metabolism
• Propionate is esterified with CoA, then Propinoyl-CoA, is carboxylated to D-Methylmalonyl-CoA, catalyzed by Propinoyl-CoA carboxylase, a biotin-dependent enzyme .
• Methylmalonyl-CoA racemase catalyzes the conversion of D-Methylmalonyl-CoA to L-Methylmalonyl CoA, which then undergoes isomerization to Succinyl CoA catalyzed by Methylmalonyl-CoA isomerase.
• This enzyme requires vitamin B12 as a coenzyme, and deficiency of this vitamin results in the excretion of methylmalonate (methylmalonic aciduria).
• Dietary odd-carbon fatty acids upon oxidation yield propionate, which is a substrate for gluconeogenesis in human liver.


Glycerol catabolism
•Glycerol is released from adipose tissue as a result of lipolysis.
•Only tissues such as liver and kidney that possess glycerol kinase ,which catalyzes the conversion of glycerol to glycerol 3-phosphate, can utilize it.
•Glycerol 3-phosphate may be oxidized to dihydroxyacetone phosphate by NAD+ catalyzed by glycerol-3-phosphate dehydrogenase.

Cori cycle
• lactate, formed by glycolysis in skeletal muscle and erythrocytes, is transported to the liver and kidney where it re-forms glucose.
• Glucose again becomes available via the circulation for oxidation in the tissues.
• This process is known as the Cori cycle, or lactic acid cycle (Figure).

Glucose-alanine cycle
• The glucose-alanine cycle (Figure) transports glucose from liver to muscle with formation of pyruvate, followed by transamination to alanine, then transports alanine to the liver, followed by gluconeogenesis back to glucose.
• A net transfer of amino nitrogen from muscle to liver and of free energy from liver to muscle is effected.


Regulation of Gluconeogenesis
Control of glycolysis and gluconeogenesis is reciprocal
Gluconeogenesis and Glycolysis are regulated by similar effector molecules but in the opposite direction
When one pathway is activated , the other is inhibited
Gluconeogenesis is subject to both:
- Hormonal control by Glucagon, Cortisol, Adrenaline and Insulin
- Allosteric regulation of gluconeogenic enzymes

Hormonal Regulation
Glucagon, Cortisol, Adrenaline
Are secreted during fasting, stress and sever muscular exercise
Induce gluconeogenic enzymes
Repress glycolytic enzymes
Insulin
Secreted after CHO meal
Induce glycolytic enzymes
Repress gluconeogenic enzymes

Allosteric regulation

Metabolic & Hormonal Mechanisms Regulate the Concentration of the Blood Glucose
• The maintenance of stable levels of glucose in the blood is one of the most finely regulated of all homeostatic mechanisms, involving the liver, extrahepatic tissues, and several hormones.
• Liver cells are freely permeable to glucose (via the GLUT 2 transporter), whereas cells of extrahepatic tissues (apart from pancreatic B islets) are relatively impermeable, and their glucose transporters are regulated by insulin.
• As a result, uptake from the bloodstream is the rate-limiting step in the utilization of glucose in extrahepatic tissues.
• The role of various glucose transporter proteins found in cell membranes is shown in the table.


THE CONCENTRATION OF BLOOD GLUCOSE IS REGULATED WITHIN NARROW LIMITS
• In the post absorptive state, the concentration of bloodglucose in most mammals is maintained between 4.5 and 5.5mmole/L. After the ingestion of a carbohydrate meal, it mayrise to 6.5–7.2 mmole/L, and in starvation, it may fall to 3.3–3.9 mmole/L.
• A sudden decrease in blood glucose will cause convulsions,as in insulin overdose


Regulation of Blood Glucose levels
• Glucose is the major source of energy for cells
• Blood Glucose (BG) regulated by Insulin & Glucagon

Glucosuria Occurs When the Renal Threshold for Glucose Is Exceeded• When the blood glucose rises to relatively high levels, the kidney exerts
a regulatory effect.
• Glucose is continuously filtered by the glomeruli but is normally completely reabsorbed in the renal tubules by active transport.
• The capacity of the tubular system to reabsorb glucose is limited to a rate of about 350 mg/min, and in hyperglycemia (as occurs in poorly controlled diabetes mellitus) the glomerular filtrate may contain more glucose than can be reabsorbed, resulting in Glucosuria.
• Glucosuria occurs when the venous blood glucose concentration exceeds 9.5–10.0 mmol/L; this is termed the renal threshold for glucose.

Glucose Tolerance
• Glucose tolerance is the ability to regulate the blood glucoseconcentration after the administration of a test dose of glucose(normally 1 g/kg body weight).
• Diabetes mellitus (type 1, or insulin-dependent diabetesmellitus; IDDM) is characterized by decreased glucose tolerancedue to decreased secretion of insulin in response to the glucosechallenge.
• Glucose tolerance is also impaired in type 2 diabetes mellitus(NIDDM),which is often associated with obesity and raised levelsof plasma free fatty acids and in conditions where the liver isdamaged; in some infections; and in response to some drugs.

• Poor glucose tolerance can also be expected due to hyperactivity of the pituitary or adrenal cortex because of the antagonism of the hormones secreted by these glands to the action of insulin.
• Administration of insulin lowers the blood glucose and increases its utilization and storage in the liver and muscle as glycogen.
• An excess of insulin may cause hypoglycemia, resulting in convulsions and even in death unless glucose is administered promptly.
• Increased tolerance to glucose is observed in pituitary oradrenocortical insufficiency—attributable to a decrease in the antagonism to insulin by the hormones normally secreted by these glands.


SYNTHESIS OF LACTOSE• In the synthesis of lactose in the mammary gland, UDPGal condenses with
glucose to yield lactose, catalyzed by lactose synthase(figure).

Uronic Acid Pathway

Overview
It is an alternative oxidative pathway for glucose
that doesn’t lead to ATP generation.
It includes oxidation of glucose to
1. Glucuronic acid
2. Ascorbic acid
It occurs mainly in the liver cytoplasm.

Metabolic reactions
1.Glucose 6-phosphate is isomerized to glucose 1-phosphate
2.Glucose 1-phosphate reacts with uridine triphosphate(UTP) to form uridine diphosphate glucose (UDPGlu) in areaction catalyzed by UDPGlu Pyrophosphorylase
3.UDPGlu is oxidized at carbon 6 by NAD-dependent UDPGludehydrogenase in a two-step reaction to yield UDP-glucuronate


UDP Glucuronic acid (active form) is needed in:
Conjugation to less polar compounds as bilirubin, steroids andsome drugs making them more water soluble (detoxicated) .
Synthesis of Glycosaminoglycans (Mucopolysaccharides) asheparin, hyaluronic acid.
In plants and some animals (not Human) glucuronic acidserves as a precursor of L-ascorbic acid.
The Uronic acid pathway also provides a mechanism by whichdietary D-Xylulose enter the central pathway.

Formula to calculate mmol/l from mg/dl: mmol/l = mg/dl / 18 Formula to calculate mg/dl from mmol/l: mg/dl = 18 × mmol/lmmol/L (UK standard) to mg/dL (US standard)