ghost shrimp and blue mud shrimp - usgs ... shrimp and blue mud shrimp coastal ecology group fish...
TRANSCRIPT

Biological Report 82(11.93) January 1989
Species Profiles: Life Histories and Environmental Requirements of Coastal Fishes and Invertebrates (Pacific Northwest)
GHOST SHRIMP AND BLUE MUD SHRIMP
Coastal Ecology Group Fish and Wildlife Service Waterways ~xperiment Station U.S. Department of the Interior U.S. Army Corps of Engineers

B i o l o g i c a l Report 82(11.93) TR EL-82-4 January 1989
Species P ro f i l es : L i f e H i s t o r i e s and Environmental Requirements o f Coastal Fishes and Inver tebra tes ( P a c i f i c Northwest)
GHOST SHRIMP AND BLUE MUD SHRIMP
Susanna Horni g An i ta S t e r l i ng
Stanley D. Smith U.S. F i sh and W i l d l i f e Serv ice
Nat ional F ishery Research Center B u i l d i n g 204, Naval S t a t i o n
Sea t t l e , WA 98115
P r o j e c t O f f i c e r David Moran
U.S. F i sh and W i l d l i f e Serv ice Nat ional Wetlands Research Center
1010 Gause Boulevard S l i d e l l , LA 70458
Performed f o r U.S. Army Corps o f Engineers Waterways Experiment S t a t i o n
Coastal Ecology Group Vicksburg, MS 39180
and
U.S. Department o f t he I n t e r i o r F i sh and W i l d l i f e Serv ice Research and Development
Nat iona l Wetlands Research Center Washington, DC 20240

DISCLAIMER
The mention o f t r a d e names i n t h i s r e p o r t does n o t c o n s t i t u t e endorsement o r recommendation f o r use by t h e Federal Government.
T h i s s e r i e s may be re fe renced as f o l l ows :
U.S. F i s h and W i l d l i f e Service. 1983-19-. Species p r o f i l e s : l i f e h i s t o r i e s and environmental requirements of coas ta l f i s h e s and i nve r teb ra tes . U.S. F i s h W i l d l . Serv. B i o l . Rep. 82(11). U. S. Army Corps o f Engineers, TR EL-82-4.
Th i s p r o f i l e may be c i t e d as f o l l ows :
Hornig, S., A. S t e r l i n g , and S.D. Smith. 1989. Species p r o f i l e s : l i f e h i s t o r i e s and environmental requirements o f coas ta l f i s h e s and i n v e r t e b r a t e s ( P a c i f i c Northwest)--ghost shrimp and b l u e mud shrimp. U. S. F i s h W i l d l . Serv. B i o l . Rep. 82(11.93). U. S. Army Corps o f Engineers, TR EL-82-4. 14 pp.

PREFACE
Th is species p r o f i l e i s one o f a se r i es on coasta l aquat ic organisms, p r i n c i p a l l y f i s h , o f spo r t , commercial, o r eco log ica l importance. The p r o f i l e s are designed t o prov ide coasta l managers, engineers, and b i o l o g i s t s w i t h a b r i e f comprehensive sketch o f t he b i o l o g i c a l c h a r a c t e r i s t i c s and environmental requirements o f t h e species and t o descr ibe how popu la t ions o f t h e species may be expected t o r e a c t t o environmental changes caused by coasta l development. Each p r o f i l e has sec t ions on taxonomy, 1 i f e h i s t o r y , eco log i ca l r o l e , environmental requirements, and economic importance, i f app l icab le . A th ree - r i ng b inder i s used f o r t h i s se r i es so t h a t new p r o f i l e s can be added as they are prepared. Th is p r o j e c t i s j o i n t l y planned and f inanced by the U. S. Army Corps o f Engineers and t h e U.S. F i s h and W i l d l i f e Service.
Suggestions o r questions regard ing t h i s r e p o r t should be d i r e c t e d t o one of t he f o l l owing addresses.
I n fo rma t ion Trans fer S p e c i a l i s t Nat ional ' Wetlands Research Center U.S. F i s h and W i l d l i f e Serv ice NASA-Sl i d e l l Computer Complex 1010 Gause Boulevard S l i d e l l , LA 70458
U. S. Army Engineer Waterways Experiment S t a t i o n A t t e n t i on: WESER-C Post O f f i c e Box 631 Vicksburg, MS 39180

CONVERSION TABLE
M e t r i c t o U.S. Customary
M u l t i p l y
m i l l i m e t e r s (mm) cent imeters (cm) meters (m) meters (m) k i lometers (km) k i lometers (km)
square meters (m2) square k i lometers (km2) hectares (ha)
l i t e r s (1) cub ic meters (m3) cub ic meters (m3)
m i l l i g r ams (mg) grams (g) ki lograms (kg) me t r i c tons (t) me t r i c tons (t)
k i l o c a l o r i e s ( kca l ) Ce ls ius degrees (OC)
U.S. Customary t o M e t r i c
inches 25.40 inches 2.54 f e e t ( f t ) 0.3048 fathoms 1.829 s t a t u t e m i l es (mi) 1.609 n a u t i c a l m i les (nmi) 1.852
square f e e t (ft2) square m i l es (mi2) acres
ga l l ons ( ga l ) cub ic f e e t (ft3) ac re - f ee t
ounces (oz) ounces (oz) pounds ( l b ) pounds ( l b ) sho r t tons ( ton)
B r i t i s h thermal u n i t s (Btu) Fahrenhei t degrees (OF)
To Obta in
inches inches f e e t fathoms s t a t u t e m i l es n a u t i c a l m i les
square f e e t square m i l es acres
ga l 1 ons cub ic f e e t ac re - f ee t
ounces ounces pounds pounds sho r t tons
B r i t i s h thermal u n i t s Fahrenhei t degrees
m i 11 imeters cent imeters meters meters k i lometers k i lometers
square meters square k i lometers hectares
l i t e r s cubic meters cub ic meters
m i l l i g r ams grams k i lograms me t r i c tons me t r i c tons
k i l o c a l o r i e s Ce ls ius degrees

CONTENTS
Page
PREFACE ................................................................. iii CONVERSION TABLE ........................................................ i v ACKNOWLEDGMENTS ......................................................... v i
NOMENCLATURE/TAXONOMY/RANGE ............................................. 1 MORPHOLOGY AND IDENTIFICATION AIDS ...................................... 1 REASONS FOR INCLUSION I N SERIES ......................................... 5 L IFE HISTORY ............................................................ 5
Development o f Eggs and Larvae ........................................ 6 P o s t l a r v a l Development ................................................ 6 H a b i t a t ............................................................... 6
GROWTH CHARACTERISTICS .................................................. 6 THE FISHERY ............................................................. 7
......................................................... ECOLOGICAL ROLE 8 Food and Feeding H a b i t s ............................................... 8 Cooperat ion. Compet i t ion . and P r e d a t i o n ............................... 8
ENVIRONMENTAL REQUIREMENTS .............................................. 10
LITERATURE CITED ........................................................ 13

ACKNOWLEDGMENTS
We are g ra te fu l f o r the reviews and suggestions o f David Buchanan o f the Oregon Department o f Fish and W i l d l i f e and Jefferson Gonor o f Oregon State Univers i ty , and f o r the use o f the i l l u s t r a t i o n s by Paul and Lynn Rudy.

GHOST SHRIMP AND BLUE MUD SHRIMP
S c i e n t i f i c name .......... Cal l ianassa c a l i f o r n i e n s i s Dana
Pre fer red common name ... Ghost shrimp (F igure 1)
S c i e n t i f i c name ............. Upogebia puget tens is (Dana)
Pre fer red common name ....... Blue mud shrimp (Figure 2)
Other common names ..... Crawfish, mud prawns, ghost shrimp ( c o l l e c t i v e l y ) , burrowing shrimp (Stevens 1928); r e d ghost shrimp (C. c a l i f o r n i e n s i s ; P h i l l i p s 1984); orange mud shrimp (C. c a l i f o r n i e n s i s ; MacGini t i e 1935); mud shrimp (U. - puget tens is ; Hedgpeth 1970).
Class ...................... Crustacea Order ....................... Decapoda Family ................ Cal l ianass idae
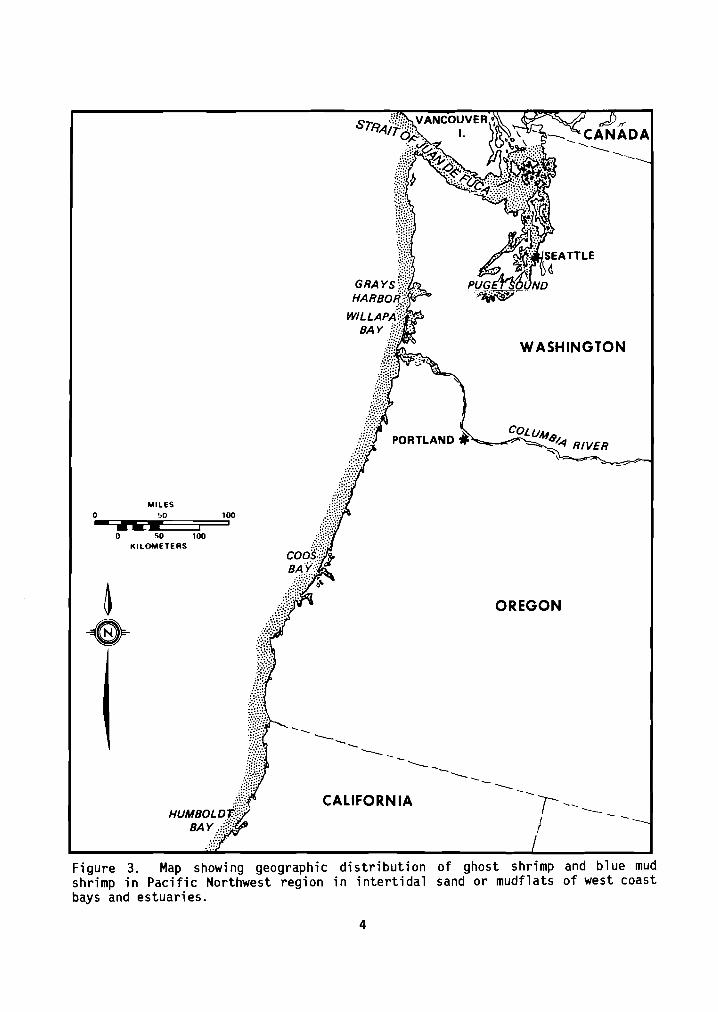
Geographic range: The ghost shrimp i s found i n i n t e r t i d a l areas a long the west coast o f North America from Mut iny Bay, Alaska, t o t h e mouth o f t he T i juana River , San Diego County, C a l i f o r n i a ; MacGin i t ie (1934) and R icke t t s and Ca lv in (1968) repor ted f i n d i n g specimens as f a r south as E l Es tua r io de Punto Banda, Baja C a l i - f o r n i a Norte, Mexico. The b lue mud shrimp i s found from southeastern Alaska t o San Quent in Bay (Bahia de San Quent in) i n Baja C a l i f o r n i a Norte. The general d i s t r i b u t i o n o f t h e two species i n t h e P a c i f i c Northwest i s i d e n t i c a l (F igure 3).
MORPHOLOGY AND IDENTIFICATION AIDS
The head and thorax o f t he ghost and b lue mud shrimps are u n i t e d i n t o a
cephalothorax. L i ke t h a t o f o the r arthropods, t h i s cephal othorax i s covered by a carapace o r exoskeleton o f hard, c h i t i n o u s ma te r i a l t h a t i s shed (molted) p e r i o d i c a l l y t o a1 low f o r growth. The g i 11s are l oca ted i n spec ia l chambers a t t h e s ides o f t he thorax under t he carapace. The b lue mud shrimp has a l a r g e rostrum ( f o r - ward extension o f t he carapace) and c y l i n d r i c a l eye s t a l k s ; t h e ghost shrimp has no rostrum o r a small one and f l a t t e n e d eye s ta l ks . Both have ex terna l mouthparts (maxi 11 ipeds) and antennae. Ha i r -1 i ke s t ruc tu res cover much o f t h e shrimps' bodies and serve such funct ions as r e c e i v i n g sensory s t i m u l i , ob ta in ing food, c l ean ing s e l f , c r e a t i n g water cur ren ts , and c lean ing and c a r r y i n g eggs (MacGini t i e 1934).
Both shrimps have f i v e p a i r s o f tho- r a c i c legs (periopoda). The f i r s t p a i r o f legs may be s l i g h t l y unequal and on l y somewhat p incer1 i ke (sub- che la te) , and the r e s t , simple as i n t h e b lue mud shrimp; o r , t he f i r s t p a i r may be very unequal and very p i nce r l i ke (chelate) , t h e second p a i r a l so p i n c e r l i k e , and the f i f t h p a i r somewhat p i n c e r l i k e as i n t he ghost shrimp (Schmi tt 1921). The asymmetry o f the f i r s t p a i r o f legs character- i s t i c o f t h e ghost shrimp i s more pronounced i n males, and the l a r g e r che l iped (p ince r leg) may be on e i t h e r t h e l e f t o r t he r i g h t s i de (MacGini t ie 1934).
Both shrimps have f i v e p a i r s of 1 ea f 1 i ke abdominal appendages ( p l eo- pods) o r swimmerets. They a l so have f 1 at tened t a i 1 appendages (uropods) adapted f o r swimming. 'The b lue mud shrimp has a shor t , square t e l s o n

F i g u r e 1. Ghost shrimp male (a ) showing morphology o f l e g p a i r s ( a c t u a l t o t a l l e n g t h o f specimen from rostrum t o t e l s o n i s 5 cm (2 inches)) and (b) enlargement o f head a r e a (dorsal view). Reproduced w i t h permission from Rudy and Rudy 1983 (copyr ight Paul and Lynn Rudy).

rostrum
1
Figure 2. Ovigerous b lue mud shrimp female (a) ac tua l l eng th from rostrum t o t e l s o n (9 cm; 3.5 inches), (b) enlarged dorsal view o f head, and (c) f i r s t - s t a g e l a r v a l form (ac tua l t o t a l l eng th about 5 mm (0.2 inches)). Reproduced w i t h per- miss ion from Rudy and Rudy 1983 (copyr igh t Paul and Lynn Rudy).
( te rmina l segment); t he ghost shrimp has a longer, more po in ted one.
These two shrimps can be d i s t i n - guished from each o the r on t h e bas is o f t he d i f f e rences i n t he f i r s t p a i r o f legs and co lo r . The b lue mud shrimp i s u s u a l l y d i r t y blue-green and t h e ghost shrimp va r ies from wh i te
t o p ink , red, and orange. The carapace o f t h e ghost shrimp i s o f t e n t ransparent enough t o a l l ow observa- t i o n o f i t s i n t e r n a l organs (Johnson and Snook 1955), making i t an i n t e r e s t i n g study specimen. There are o the r Ca l l ianassa species besides t h e ghost shrimp on t h e west coast; how- ever, o n l y one, 5. gigas, i s s i m i l a r
WASHINGTON
KILOMETERS
CALIFORNIA
OREGON
Figure 3. Map showing geographic d i s t r i b u t i o n o f ghost shrimp and b lue mud shrimp i n P a c i f i c Northwest region i n i n t e r t i d a l sand o r mudflats o f west coast bays and estuaries.

i n d i s t r i b u t i o n . h a b i t a t , and hab i ts . Cal l ianassa gigas i s l a r g e r than the ahost s h r i m ~ (125-150 mm (5-6 inches) yong). ~ n d ' ai though the 'females and j uven i l es o f these two species are s i m i l a r i n appearance, t he l a r g e che l iped o f t h e C. g a s male i s longer and narrower than t h a t o f t h e male ghost shrimp (Morr is e t a l . 1980).
REASONS FOR INCLUSION I N THE SERIES
Although t h e ghost shrimp and b lue mud shrimp a re harvested as b a i t a long the e n t i r e west coast o f t he Un i ted States, they are considered by some t o be important pests o f commercial oyster-growi ng operat ions i n t h e P a c i f i c Northwest (R i cke t t s and Ca lv in 1968; McCrow 1972; Buchanan e t a l . 1985). They are be l ieved t o destabi - 1 i z e the subst ra te , smother oys ters w i t h debr is , and d r a i n o f f water (through t h e i r burrows) from d iked oys te r beds.
E i t h e r species can a l t e r t h e phys ica l c h a r a c t e r i s t i c s o f t h e h a b i t a t i t occupies and a f f e c t t h e composit ion o f t h e i n t e r t i d a l in fauna l community (Brenchley 1981; Posey 1986a). The ghost shrimp (Figure 1) i s t he more a c t i v e burrower o f t h e two and more severely a f f e c t s subst ra te consistency ( B i r d 1982). Both the b lue mud shrimp (Figure 2) and the ghost shrimp are associated w i t h a v a r i e t y o f commensal and p a r a s i t i c species (MacGi n i t i e 1930, 1934, 1935; MacGini ti e and MacGi n i ti e 1968; Rickets and Ca lv in 1968; K o z l o f f 1973). The ghost shrimp i s one o f t h e most abundant res idents o f marine sloughs o r bay mudf lats on t h e west coast o f Nor th America (MacGini t i e 1934).
LIFE HISTORY
Both the ghost shrimp and the b lue mud shrimp l i v e i n burrows i n the
i n t e r t i d a l sand o r mudf lats o f west coast bays and es tuar ies . Entrances t o ghost shrimp burrows may be observed i n t he center o f small con ica l h i l l s o f sand and small pebbles; those o f t he b lue mud shrimp are l ess conspicuous, w i t h much small e r , o r absent, surrounding h i 11s o f sand (Koz lo f f 1973).
Members o f t he b lue mud shrimp species nea r l y always l i v e i n male- and-female p a i r s ; each p a i r i n h a b i t s a smooth-wal 1 ed , permanent, branchi r(g burrow extending about 45 cm (18 inches) below the surface. The burrow general l y has several entrances, each about 1 cm (0.4 inches) i n diameter. The diameter o f the tunnel beyond the opening i s t oo narrow t o a1 low the shrimp t o t u r n around; consequently, spec ia l l y en1 arged chambers are re- qui red. The smooth w a l l s appear t o be cemented w i t h a sec re t i on produced by the shrimp. The b lue mud shrimp forms a "mud basket," w i t h i t s f i r s t two p a i r s o f legs, which i t uses as a scoop t o t ranspo r t mud and b u i l d i t s burrow, and as a s t r a i n e r t o c o l l e c t food. The t h i r d and f i f t h p a i r s o f legs are used f o r walking, and t h e f o u r t h p a i r i s braced aga ins t t he bur- row wa l ls . The t a i l - f a n can be used t o b lock the burrow tunnel so e f fec - t i v e l y t h a t t he f low o f water i s stopped; t h i s may poss ib l y be a pro- t e c t i v e maneuver t o ward o f f a t t acks from the r e a r (MacGini t ie 1934). The species feeds on d e t r i t u s and p lank ton s t r a i n e d from seawater, which i t forces through t h e burrow by us ing i t s f ou r pa i r s o f swimmerets ( p l eopods) (MacGi n i t i e 1930).
The ghost shrimp a l so i n h a b i t s burrows, b u t o f a l ess permanent char- a c t e r s ince t h i s species s i f t s most o f i t s food d i r e c t l y from t h e subst ra te and tunnels almost cons tant ly , rework- i n g the sediment t o a depth o f about 75 cm (30 inches) i n search o f food. Burrow s t ruc tu res o f ghost shrimp a re l ess cons i s ten t i n p a t t e r n than those o f b lue mud shrimp; t he ghost

shrimp d igs tunne ls branching i n a l l d i r e c t i o n s , forming complex burrows w i t h var ious numbers o f openings (MacGini t ie 1934). The second and t h i r d p a i r s o f legs are used f o r d ig - g ing and the f o u r t h and f i f t h f o r c lean ing i t s appendages, g i 11 s, and back, and f o r c lean ing and manipulat- i n g i t s eggs (MacGini t ie 1934). The t h i r d , f ou r th , and f i f t h p a i r s o f legs are used i n walking; t h e f o u r t h p a i r i s extended outward aga ins t t he burrow w a l l f o r support. The swimmerets o f t he ghost shrimp cons tan t l y c i r c u l a t e water through t h e burrow, f a c i l i t a t i n g r e s p i r a t i o n . I t s t a i l - f a n , 1 i ke the b lue mud shrimp's, can be used (proba- b l y p r o t e c t i v e l y ) t o b lock the burrow. The l a rge che l iped o f t h e male i s a weapon used i n d isputes over t e r r i t o r y and dur ing the mating season (MacGini t ie 1934).
Development o f Eggs and Larvae
Female ghost shrimp are ovigerous (capable o f producing f e r t i 1 e eggs) throughout t h e year, b u t t he p r i n c i p a l spawning season i s i n l a t e June and e a r l y J u l y (MacGini t ie 1935). In ten- s i v e breeding probably begins i n spr ing, bu t ovigerous females may s t i l l be p l e n t i f u l as l a t e as August. Spr ing warming appears t o be t h e t r i g - ger f o r egg development. Three t o f o u r broods are produced a t about 6-week i n t e r v a l s . The 1 arvae develop as p lank ton i n coasta l waters through f i v e zoeal stages, which are d i s t i n - guishable from one another p r imar i l y on the bas is o f s i z e (McCrow 1972). A t o t a l o f 6-8 weeks i s spent as nearshore oceanic p lank ton (through the f i v e zoeal and one megalopal stage); zoeal l a r vae are usua l l y re - leased on ebb t i d e s i n June and Ju l y , and the f i r s t megalopae appear i n ear- l y August. Recruitment t o t he estuary i s probably f a c i l i t a t e d by f l o o d t i d e s occu r r i ng i n l a t e summer and fa1 1. Larval d r i f t i n g dur ing t h i s p lank ton i c phase very 1 i k e l y serves as a mecha- nism o f genet ic exchange among popula- t i o n s i n d i f f e r e n t es tuar ies (Johnson and Gonor 1982).
Blue mud shrimp females are known t o be ovigerous i n January, February, and p a r t o f March (MacGini t ie 1935), bu t l a r v a l development o f t h i s species has no t been s tud ied ex tens ive ly .
Pos t l a rva l Development
Juven i l e ghost shrimp are presumed t o metamorphose r a p i d l y t o a s t a t e adapted f o r l i f e on t h e bottom j u s t before recru i tment t o t h e estuary (Johnson and Gonor 1982). Mor ta l i t y due t o predat ion i s probably substan- t i a l du r i ng the sho r t p e r i o d (minutes t o hours) between the moment t he or - ganism drops t o t he subst ra te and i t s successful burrowing beneath the sur- face (MacGini t ie 1934). The l i f e s p a n o f t he ghost shrimp has been v a r i o u s l y est imated a t 3-5 years ( B i r d 1982), 10 years (MacGi n i ti e 1935), and 15-16 years (R icke t t s and Ca lv in 1968). The b lue mud shrimp i s a l so be l ieved t o be r e l a t i v e l y l ong - l i ved (MacGini t ie 1930).
Hab i ta t
Both o f these species are commonly found i n i n t e r t i d a l areas o f mixed sand and mud. The b lue mud shrimp l i v e s i n muddier areas than does the ghost shrimp; observat ions w i t h re - spect t o t i d a l he igh t preferences vary (Table 1). I n Oregon es tuar ies , ghost shrimp were c o n s i s t e n t l y found i n t i d e f l a t s c lose r t o t he ocean than were b lue mud shrimp ( B i r d 1982). Both species are common res iden ts o f eelgrass beds i n t h e P a c i f i c Northwest (Phi 11 i p s 1984).
GROWTH CHARACTERISTICS
Typ ica l l y , l eng th o f a d u l t s i s 5-10 cm (2-4 inches) i n t h e ghost shrimp and 7.5-10 cm (3-4 inches) i n t he b lue mud shrimp (MacGini t ie and MacGi n i t i e 1968). However, 1 ength may reach 11.5 cm (4.5 inches) i n t h e ghost shrimp and 15 cm (6 inches) i n t he b lue mud shrimp (Morr is e t a l . 1980). MacGin i t ie ( 1930, 1935 )

Table 1. Hab i ta t preferences o f ghost shrimp and b lue mud shrimp repor ted a t d i f f e r e n t 1 ocations.
Locat ion Ghost shrimp Blue mud shrimp Source
P a c i f i c NW Muddy sand Muddier sand
Yaquina Bay, OR 0 t o 1 f t 0 t o 1 f t
Oregon es tuar ies T i d e f l a t s c lose T ide f l a t s f u r t h e r t o ocean from ocean
N. C a l i f o r n i a Sandier m id - t i da l Lower, muddier areas f 1 a t s
Tomales Bay & E l khorn Slough, CA Muddy sand So f te r mud
Monterey Bay, CA General l y lower General l y h igher t i d a l areas; t i d a l areas; mud, mixed sand and sandy mud w i t h mud c l ay
Kozl o f f 1973
Thompson and P r i t cha rd 1969
B i r d 1982
Hedgpeth 1970
Smith and Car l t on 1975
repor ted f i n d i n g the l a r g e s t b lue mud shrimp i n t h e muddiest, l e a s t rocky areas.
Ghost shrimp mature a t 18-24 months and some reproduct ive females may be l ess than 3 cm (1.2 inches) long; b lue mud shrimp take 3 o r more years t o mature and reproduct ive females exceed 6 cm (2.4 inches) i n length. Est imated growth i n l eng th averages approximately 15-30 mm/yr (0.6-1.2 inches/yr) i n ghost shrimp and 18-26 mm/yr (0.7-1.0 inches/yr) i n b lue mud shrimp ( B i r d 1982). Densi ty w i t h i n a ghost shrimp colony and the co lony 's l o c a t i o n appear t o i n f l uence both growth and s i z e a t sexual ma tu r i t y ; ghost shrimp i n t h e less dense co lon ies c l o s e s t t o t he ocean grow f a s t e r , and the females become sexual- l y mature a t l a r g e r s izes and produce more and l a r g e r eggs ( B i r d 1982).
Dens i t i es o f ghost shrimp have been est imated a t 700-1 ,400/m2 (2.8-5.6 m i 1 1 ion/acre) i n Yaqui na Bay, Oregon
(McCrow 1972) ; 420-770/m2 (1.7-3.1 m i 11 ion/acre) i n high-densi t y areas o f Sand Lake Estuary, Oregon; and l e s s than 300/m2 (1.2 m i l l ion/acre) i n o ther areas on t h e Oregon coast ( B i r d 1982). Blue mud shrimp d e n s i t i e s i n Oregon es tuar ies range from 330 t o 660/m2 (1.3-2.7 m i 11 ion/acre) ( B i r d 1982). Biomass o f e i t h e r species sometimes exceeds 2.0 kg/m2 (18,000 lb /acre (wet weight)).
THE FISHERY
Ghost and b lue mud shrimp a re harvested by commercial b a i t f ishermen and rec rea t i ona l f ishermen i n Cal i f o r - n ia , Oregon, and Washington. Peterson (1977) descr ibed a method used i n southern C a l i f o r n i a i n which water i s pumped i n t o the subst ra te under pres- sure, f o r c i n g the animals ou t o f t h e i r burrows; i n t he area he studied, har- ves t no t i ceab ly reduced the ghost shrimp popu la t ion . I n t he P a c i f i c Northwest, attempts have been made t o

c o n t r o l t he shrimp on commercial Japanese oys ter ( ~ r a s s o s t r e a g igas) grounds w i t h t h e i n s e c t i c i d e Sevin (carbaryl ). This p e s t i c i d e has been used t o c o n t r o l ghost and mud shrimp i n Washington s ince 1963 (Washington Department of F isher ies and Washington Department o f Ecology 1985), and a l - though i t has been used on oys te r grounds i n Oregon, such use i s cur- r e n t l y unlawful t he re (L. Fredd, Oregon Department o f F i sh and Wild- l i f e , Port land, OR; pers. comm.). Dur ing i t s use i n Oregon, b a i t f i s h e r - men noted ghost shrimp m o r t a l i t i e s i n untreated areas soon a f t e r nearby oys te r grounds were sprayed (Buchanan e t a1 . 1985).
Washington oys ter growers est imate t h a t oys ter p roduct ion would drop 70%- 80%, r e s u l t i n g i n a $5 m i l l i o n annual l oss i n P a c i f i c and Grays Harbor Counties, w i thou t ghost shrimp c o n t r o l (Washington Department o f F i she r ies and Washington Department o f Ecology 1985). However, questions have been ra i sed about t he e f f e c t s o f Sevin on o the r organisms, i n c l u d i n g the commercial l y important ~ u n ~ e n e s s crab (Cancer magister), and on the es tuar ine ecosystem as a whole (Lindsay 1961; Stewart e t a l . 1967; Buchanan e t a l . 1985). Although the b lue mud shrimp i s be l i eved t o d i s t u r b t h e sediment f a r l ess ex tens i ve l y than t h e ghost shrimp ( B i r d 1982), bo th have been t h e ob jec ts o f c o n t r o l programs.
ECOLOGICAL ROLE
Food and Feeding Habi ts
The ghost shrimp was once thought t o feed exc lus i ve l y by s i f t i n g organic d e t r i t u s from the f l o o r o f i t s burrow through t h e h a i r s on the second and t h i r d p a i r s of 1 egs , r e j e c t i n g coarse ma te r i a l , and then i n g e s t i n g the re- t a ined f i n e p a r t i c l e s by the use o f t he maxi 11 ipeds (MacGini t i e 1934). And al though i t i s s t i l l thought t o ob ta in most o f i t s food i n t h i s
manner, t he re i s evidence t h a t i t a l so f i l t e r s d e t r i t u s and p lank ton from the water moving through i t s burrow as does t h e b lue mud shrimp (Morr is e t a l . 1980). Rejected ma te r i a l i s deposi ted outs ide the burrow. Burrowing a c t i v i t y i s heaviest i n t h e upper 45-50 cm (18-20 inches), where the a v a i l a b i l i t y o f food i s g rea tes t (MacGi n i t i e 1934). The burrowing and feeding behavior o f t he ghost shrimp i s vigorous enough t o cause substan- t i a l a1 t e r a t i o n s i n sur face sediment c h a r a c t e r i s t i c s over t ime, decreasing organic content and s h i f t i n g the par- t i c l e s i z e d i s t r i b u t i o n upwards ( B i r d 1982). Sediment i n dense ghost shrimp beds o f t e n has a s o f t , quicksand qual- i t y (Posey 1985). The burrowing a c t i v i t y o f both the ghost and b lue mud shrimp aerates t h e subsurface s o i l (MacGi n i ti e 1930, 1934).
The b lue mud shrimp i s a suspension feeder, s t r a i n i n g d e t r i t a l p a r t i c l e s and p lank ton from seawater kept moving through i t s burrow by the a c t i o n o f i t s swimmerets. To feed, the animal p o s i t i o n s i t s e l f near a burrow entrance and increases the movement o f t h e swimmerets t o increase t h e c u r r e n t of seawater through the burrow. The t h i r d max i l l i peds are used t o p e r i o d i - c a l l y sweep t h e food p a r t i c l e s c o l - l e c t e d i n t o the an imal 's mouth. P a r t i c l e s t h a t are too b i g are e jec ted (MacGi n i t i e 1930).
Cooperation, Competi t ion, and Predat ion
By ae ra t i ng the subsurface sediment and d igg ing burrows p ro tec ted from most predators, ghost shrimp and b lue mud shrimp prov ide an environment a t t r a c t i v e t o commensal s. Commensal and p a r a s i t i c species associated w i t h these shrimp inc lude a b l i n d goby, t h ree species o f pea crabs, two species o f clams, a copepod, a shrimp, po lyno id worms, and isopods (see Tab1 e 2).
Species t h a t might compete w i t h these shrimp f o r e i t h e r food o r space

Table 2. Commensal (c) and p a r a s i t i c (p) species repor ted i n burrows o f ghost shrimp and b lue mud shrimp (compiled from MacGini t ie and MacGin i t ie 1968; R icke t t s and Ca lv in 1968; Koz lo f f 1973).
Species
- -
Found w i t h ghost shrimp
-
Found w i t h b lue mud shrimp
Goby Clevelandia ios (c) I n burrow
Pea crabs Abundant i n burrow Abundant i n burrow I n burrow ( ra re )
Clams
I n burrow
Abundant i n burrow --- - - -
Pseudopythina r u g i f e r a (c) Cryptomya ca l i f o r n i c a (c)
Copepods Hemicyclops ca l l ianassae (c) Clausidium vancouverense (c)
Shrimp Betaeus ense:adensi s (c)
Polynoid worms Hesperonoe spp. (c )
Isopods ( u n i d e n t i f ied--p) Phyl lodurus abdominal is (p)
--- Extends s y p h o n ~ i n t o burrow
On g i l l s Underside carapace (common)
On g i l l s
I n burrow
Under carapace -- -
Underside o f abdomen Extends syphons i n t o burrow
On g i l l s Under carapace
I n burrow
--- Underside o f abdomen
are r a r e i n ghost shrimp co lon ies because o f the con t i nua l rework ing o f t h e sediment by t h i s species. Infauna are both more v a r i e d and more abundant i n b lue mud shrimp co lon ies because t h i s species less severely a f f e c t s t h e sediment s t r u c t u r e ( B i r d 1982).
A1 though ghost shrimp t y p i c a l l y i n h a b i t deep burrows, they are suscep- t i b l e t o predat ion by o ther animals because they sometimes venture outs ide t h e i r burrow entrances. Under t e s t cond i t ions , ghost shrimp spent over 25% o f t h e t ime w i t h i n 2 cm o f t he burrow entrance; t h e shrimp were a l so
observed t o move from one burrow t o another and were o f t e n found w i t h p a r t o f an appendage exposed above the sur face (Posey 1985).
The seaward boundary o f dense shrimp beds coinc ided w i t h a f o u r f o l d seaward increase i n t he dens i t y o f t he major predator , t he P a c i f i c staghorn scu lp in (Leptocot tus armatus) i n Coos Bay, Oregon (Posey Caging experiments i n Coos Bay i n d i c a t e d t h a t p redat ion by t h i s f i s h , which was most in tense i n summer, probably r e s t r i c t s t he seaward d i s t r i b u t i o n o f ghost shrimp (Posey 1986b).

Mud and ghost shrimps are sometimes k i l l e d by t h e leopard shark, T r i a k i s semifasciata, and by the brown smooth- hound shark, Mustelus hen le i . The leopard shark. whose ranae extends n o r t h t o Oregon, apparentlyWcan shovel o r burrow i n t o the subst ra te t o prey on benth ic species (Russo 1975). Dungeness crabs are known t o e a t ghost shrimp, b u t t he shrimp does no t appear t o be a major component o f t h e c rab ' s d i e t (Stevens e t a l . 1982; Posey 1985). Sea-run c u t t h r o a t t r o u t (Sal mo c l a r k i c l a r k i ) a l so commonly e a t ghost -- shrimp, b u t a re no t considered a major p redator (Posey 1985). Posey a l so suggests t h a t i n t e r t i d a l l y f o rag ing b i r d s may occas iona l ly e a t ghost shrimp.
ENVIRONMENTAL REQUIREMENTS
The opt imal temperature range f o r ghost shrimp appears t o be 10 t o 13 OC, depending on depth below t h e surface. Egg-beari ng females seem t o p r e f e r t he coo le r water a t t he g rea te r depths; immature specimens are found h igher up i n t he burrow. Water tem- pera ture i n ghost shrimp h a b i t a t i n Yaquina Bay, Oregon, va r i es season- a l l y from 9 t o 15 O C (McCrow 1972). A c t i v i t y of ghost shrimp decreased s l i g h t l y w i t h increas ing maximum d a i l y a i r temperature i n an outdoor aquarium ( Posey 1987).
Ghost shrimp tend t o be most abundant a t t h e seaward end o f bays w i t h subs tan t i a l f reshwater i n f l o w (McCrow 1972) and to1 e ra te sa l i n i t i e s from about 25% t o 125% the s a l i n i t y o f normal seawater (33 ppt) . Blood s a l i n i t y changes a long w i t h water s a l i n i t y . I n a l abo ra to ry t e s t , s a l i n i t i e s o f 8-9 p p t were l e t h a l t o 75%-100% o f ghost shrimp (Posey 1987). A c t i v i t y o f ghost shrimp decreased w i t h decreasing s a l i n i t y between 33 and 10 p p t (Posey 1987). The b lue mud shrimp t o l e r a t e s s a l i n i t i e s as low as 10% t h a t o f seawater and regu- l a t e s osmot i ca l l y when sa l i n i t y fa1 1 s
below 75% t h a t o f seawater (Morr is e t a1 . 1980).
Oxygen a v a i l a b i l i t y i s no doubt a 1 i m i t i n g f a c t o r f o r a1 1 i n t e r t i d a l species, i n c l u d i n g the ghost and b lue mud shrimps (MacGi n i t i e 1935). Thomp- son and P r i t c h a r d (1969) measured oxy- gen l e v e l s i n burrows i n Yaquina Bay, Oregon, and found t h a t du r i ng ebb t i d e , oxygen l e v e l s were occas iona l ly zero. They a l so found t h a t under lab- o ra to ry cond i t ions , t he ghost shrimp cou ld su rv i ve anoxia ( l a c k o f oxygen) f o r 5.7 days and the b lue mud shrimp could surv ive f o r 3.3 days, f a r longer than they would normal ly be subjected t o anoxia i n t he environment.
Although the ghost shrimp has a lower normal metabol ic r a t e and sur- v ives anoxia and hypoxi a (1 ow oxygen) b e t t e r than does t h e b lue mud shrimp, bo th appear t o have resp i ra to ry adaptat ions t h a t a l l o w them t o t o l e r a t e the low oxygen cond i t ions under which they l i v e . Laboratory experiments have shown t h a t both species are ab le t o lower t h e i r metabol ic r a t e s once oxygen l e v e l s become c r i t i c a l l y low. A d d i t i o n a l l y , s tud ies o f t h e ghost shrimp have demonstrated the f o l l owing adaptat ions t o hypoxia/anoxia: when oxygen 1 eve1 s become low, hea r t r a t e i s lowered (Thompson and P r i t c h a r d 1969); a r e s p i r a t o r y pigment, hemocyanin, l i b e r a t e s more bound oxygen t o t he t i ssues (Morr is e t a l . 1980); and t h e shrimp i s ab le t o swi tch t o an a1 te rnate , anaerobic metabol ism (P r i t cha rd and Eddy 1979; Mor r i s e t a l . 1980).
C l i f t o n e t a l . (1984) s tud ied t h e e f f e c t o f s p i l l e d o i l on ghost shrimp co lon ies i n W i l lapa Bay, Washington. They concluded t h a t small amounts of o i l c a r r i e d i n on the t i d e s and tempo- r a r i l y stranded i n i n t e r t i d a l areas are u n l i k e l y t o have a ser ious long- term impact. However, stranded o i 1 t h a t i s bu r i ed by a subsequent depos i t ion o f o i 1 - f ree sediment cre- a tes a b a r r i e r t o burrowing a c t i v i t y

t h a t can be expected t o p e r s i s t f o r nature, t h e ghost and b l u e mud shrimp years. They a l so concluded t h a t t he appear t o be an i n t e g r a l p a r t of t h e burrowing a c t i v i t y o f t he shrimp con- nearshore environments. And f o r t u - t r i b u t e s t o t he i n t r o d u c t i o n o f o i l na te l y f o r t h e shrimp, t h e i r wide- i n t o t h e sub-surface. spread d i s t r i b u t i o n should a1 low them
t o sus ta in t h e i r popu la t ions desp i te Although t h e i r e f f e c t s on the the c u r r e n t attempts t o e l im ina te them
environment a re con t rove rs ia l i n 1 ocal l y .


LITERATURE CITED
B i r d , E. M. 1982. Popu la t ion dynamics o f t h a l ass in idean shrimps and community e f f e c t s through sediment m o d i f i c a t i o n . Ph. 0. D i s s e r t a t i o n . U n i v e r s i t y o f Maryland, C o l l ege Park. 151 pp.
Brenchley, G. A. 1981. Dis turbance and c m u n i t y s t r u c t u r e : an exper imenta l s tudy o f b i o t u r b a t i o n i n marine so f t -bo t tom environments. J . Mar. Res. 39: 767-790.
Buchanan, D.V., D. L. Bottom, and D.A. Armstrong. 1985. The c o n t r o v e r s i a l use o f t h e i n s e c t i c i d e Sevin i n P a c i f i c Northwest es tua r i es : i t s e f f e c t s on Dungeness crab, P a c i f i c oys te r , and o the r species. Pages 401-407 i n Proceedings o f t h e symposium o n Dungeness c rab b i o l o g y and management, Oct. 9-11, 1984, Anchorage, AK. Alaska Sea Grant Rep. No. 85-3, U n i v e r s i t y o f Alaska, A1 aska Sea Grant Col 1 ege Program, Fairbanks.
C l i f t o n . H. E. . K.A. Kvenvolden. and
Es tuar ine Coastal Shel f Sci . 14: 501- 516.
Johnson, M.E. , and H.J. Snook. 1955. Seashore animals o f t h e P a c i f i c coast. Dover Pub l i ca t i ons I n c . , New York. 659 pp.
K o z l o f f , E.N. 1973. Seashore l i f e o f Puget Sound, t h e S t r a i t o f Georgia, and San Juan Archipelago. U n i v e r s i t y o f Washington Press, Sea t t l e . 282 pp. and 28 p l a t e s .
Lindsay, C. E. 1961. P e s t i c i d e t e s t s i n t h e marine environment i n t he s t a t e o f Washington. Proc. N a t l . Shel l f i s h . Assoc. 52:87-97.
MacGin i t ie , G. E. 1930. The n a t u r a l h i s t o r y o f t h e mud shrimp Upogebia
uge t t ens i s (Dana). Ann. Mag. Nat. { i s t . , Ser. 10, 6:36-44.
MacGini t i e , G. E. 1934. The n a t u r a l h i s t o r v o f C a l l ianassa c a l i f o r - niensi; Dana. Am. M id l . Nat. 15:166- 177.
J.B' R ~ P P . '1984' oii and M a c G i n i t i e , G.E. 1935. E c o l o g i c a ] i n f auna l a c t i v i ty--modi f i c a t i o n o f burrowing behavior and r e d i s t r i - aspects o f a C a l i f o r n i a marine
b u t i o n o f o i l . Mar. Environ. Res. es tuary . Am. Mid1 . Nat. 16: 629-765.
Hedgpeth, J . W. 1970. I n t r o d u c t i o n t o seashore l i f e o f t he San Francisco Bay reg ion and t h e coas t o f no r the rn C a l i f o r n i a . U n i v e r s i t y o f C a l i - f o r n i a Press, Berkeley. 136 pp.
Johnson, G. E. . and J.J. Gonor. 1982. Th6 t i d a l exchange o f Ca l l ianassa c a l i f o r n i e n s i s (Crustacea, Decapoda) l a r v a e between t h e ocean and t h e Salmon R iver Estuary, Oregon.
MacGin i t ie , G. E . , and N. MacGin i t ie . 1968. Pages 284-293 i n Natura l h i s t o r y o f marine animalF, 2nd ed. McGraw-Hi 11 , New York.
McCrow. L.T. 1972. The ahost s h r i m ~ . . . c a l l ianassa c a l i f o r n i e n s i s Dana, 1854, i n Yaquina Bay, Oregon. M.S. Thesis. 0regon S ta te u n i v e r s i t y , Corval 1 i s . 56 pp.
Mor r i s , R.H., D.P. Abbott , and E. C. Haderl i e . 1980. I n t e r t i d a l

i nver tebra tes o f C a l i f o r n i a . Stan- important i nve r teb ra te animals. f o r d U n i v e r s i t y Press, Stanford, U.S. F i sh Wi ld l . Serv. B i o l . Serv. C a l i f o r n i a . 690 pp. Program FWS/OBS-83/16. 225 pp.
Peterson, C. H. 1977. Competi t ive Russo, R.A. 1975. Observations on organ iza t ion o f t h e sof t -bot tom the food hab i t s o f leopard sharks macrobenthic communities o f southern ( T r i a k i s semi f asc ia ta ) and brown C a l i f o r n i a lagoons. Mar. B i o l . smooth-hounds (Mustel us hen le i ). (Ber l . ) 43: 343-359. C a l i f . F i sh Game 61: 95-103.
P h i l l i p s , R.C. 1984. The ecology o f eelgrass meadows i n t he P a c i f i c Northwest: a community p r o f i l e . U.S. F i sh Wi ld l . Serv. FWS/OBS- 84/24. 85 pp.
Posey, M.H. 1985. The e f f e c t s upon the macrofaunal community o f a dominant burrowing depos i t feeder, Ca l l ianassa c a l i f o r n i e n s i s , and the r o l e o f p redat ion i n determining i t s i n t e r t i d a l d i s t r i b u t i o n . Ph. D. D i sse r ta t i on . U n i v e r s i t y o f Oregon, Eugene. 119 pp.
Posey, M.H. 1986a. Changes i n a benth ic community associated w i t h dense beds o f a burrowing depos i t feeder, Ca l l ianassa c a l i f o r n i e n s i s . Mar. Ecol. Prog. Ser. 31:15-22.
Posey, M.H. 1986b. Predat ion on a burrowing shrimp: d i s t r i b u t i o n and community consequences. J. Exp. Mar. B i o l . Ecol. 103:143-161.
Posey, M.H. 1987. E f f e c t s o f lowered s a l i n i t y on a c t i v i t y o f t he ahost shrimp - Ca l l ianassa - c a l i fo rn iens i s . Northwest Sci. ( i n press).
Pr i tchard , A.W. , and S. Eddy. 1979. Lactate format ion i n c a l l ianassa c a l i f o r n i e n s i s and Upogebia puget- t e n s i s (Crustacea: Thal ass i nidea). Mar. B i o l . (Ber l . ) 50: 249-253.
R icke t t s , E.F., and J. Calv in. 1968. Between P a c i f i c t i des . Revised by J. W. Hedgpeth, S tanford U n i v e r s i t y Press, Stanford, CA. 614 pp.
Rudy, P. , J r . , and L.H. Rudy. 1983. Oregon es tuar ine inver tebra tes : an i l l u s t r a t e d guide t o t he common and
Schmitt , W.L. 1921. The marine decapod crustacea o f Cal i f o r n i a. Univ. Cal i f . Publ. Zoo1 . 23: 1-470.
Smith, R. , and J.T. Car l ton. 1975. L i g h t ' s manual : i n t e r t i d a l i nve r te - b ra tes o f t he c e n t r a l C a l i f o r n i a coast. U n i v e r s i t y o f C a l i f o r n i a Press, Berkeley. 716 pp.
Stevens, B. A. 1928. Ca l l ianassidae from the west coast o f North America. Pub'l. Puget Sound B i o l . (Ber l . ) Stn. 6: 315-369.
Stevens, B.G. , D.A. Armstrong, and R. Cusimano. 1982. Feeding hab i t s o f the Dungeness crab Cancer magister as determined by the Index o f Re1 a t i v e Importance. Mar. B i o l . 72: 132-145.
Stewart, N. E. , R. E. Millemann, and W. P. Breese. 1967. Acute t o x i c i t y o f t he i n s e c t i c i d e Sevin and i t s h y d r o l y t i c product 1-naphthol t o some marine organisms. Trans. Am. F i sh Soc. 96:25-30.
Thompson, R. K. , and A. W. P r i tchard. 1969. Respiratory adaptat ions o f two burrowing crustaceans, Ca l l ianassa c a l i f o r n i e n s i s and Upogebia puget tens i s (Decapoda, Thal ass i n i dea). B i o l . B u l l . (Woods Hole) 136: 274-287.
Washington Department o f F isher ies and Washington Department o f Ecology. 1985. F ina l environmental impact statement: use o f t he i n s e c t i c i d e Sevin t o c o n t r o l ghost and mud shrimp on oys ter beds i n W i l l apa Bay and Grays Harbor. 82 pp. p l u s appendices.

4. Title and Subtitle S. Rapon oat. Species P r o f i l e s : L i f e H i s t o r i es and Environmental Requirements of Coastal Fishes and Inver tebrates (Pac i f i c Northwest)--Ghost Shrimp
30272 -101 3. Ruiplant's Accasslon NO.
and Blue Mud Shrimp 7. Author(s)
Susanna Hornig. An i ta S te r l i ng , and Stanley 0. Smith 9. Parforming O~n lza t ion Name and Address
U.S. F ish and W i l d l i f e Service
- -
U.S. Department o f the I n t e r i o r U.S. Army Corps o f Engineers 1% T Y ~ . Of RapoR 4 h r i o d Cowrod
F ish and W i l d l i f e Service Waterways Experiment S ta t i on Nat ional Wetlands Research Center P.O. Box 631 Washington, DC 20240 Vicksburg, MS 39180
2. REPORT DOCUMENTATION PAGE
8. Parforming O~anlzation Rapt. No.
10. Pmi.ct/Task/Worh Unlt No.
I National F ishery Research Center Bu i l d i ng 204, Naval S ta t i on Puget Sound Seat t le , WA 98115-5007 --
12. Smnsorins Onanlzatlon Nama and Address
I IS. Supplamentar)' Notes
*U.S. Army Corps o f Engineers Report No. TR EL-82-4
16. -tract (Limi(: 200 Mrh)
1. REPOW
B io l og i ca l Report 82(11.93)*
11. bntnctfc) or Gnnt(G) NO.
(c)
(0)
Species p r o f i l e s are l i t e r a t u r e summaries o f the taxonomy, morphology, range, l i f e h i s t o r y , and environmental requirements o f coastal aquat ic species. The p r o f i l e s are prepared t o a s s i s t i n environmental impact assessments. The ghost shrimp 1 . . (Ca l l ianassa c a l i f o r n i e n s i s ) and b lue mud shrimp (Upogebia pugettensis) are common I res iden ts o f i n t e r t i d a l mudf lats o f the P a c i f i c Northwest. as we l l as o f the e n t i r e I west coast o f t he contiguous Uni ted States. These species are decapod crustaceans, bu t no t t r u e shrimp. They are harvested as b a i t by rec rea t iona l and commercial oyster-growing operat ions. Ghost shrimp la rvae develop i n summer i n nearshore coasta l waters and s e t t l e t o the subst ra te surface, where they r a p i d l y metamorphose; t he l i f e cyc le o f t he b lue mud shrimp i s presumed t o be s im i l a r . Both species spend t h e i r l i v e s i n burrows i n t he mudf lat , where the ghost shrimp i s p r i m a r i l y a deposi t feeder and t he b lue mud shrimp i s a suspension feeder.
17. Dosumant Analyais a. Daurlpton
Fishery Sal i n i t y Sediments Predators Growth Temperature L i f e Cycles Competitors Estuar ies Oxygen Shrimps Commensals Feeding hab i t s Depth O i l s p i l l s
b. ldentlfian/Own-End& Terms
Ghost Shrimp Cal l ianassa c a l i f o r n i e n s i s Dana
Blue Mud Shrimp Upogebia pugettensis (Dana)
1 L Arallablllty Statamant 1 19. Sacunty Class (This Ramrl)
Unc lass i f i ed Un l im i ted release 2a Sacuritv Class (This Pama)
Unc lass i f i ed
21. No. of Pages
14 22. Prlca
(s- ms1-u9. in r-
OPTIONAL CORY 272 (4-72) (Forrmrly NTtS-35) Oapaamant of Commma

As the Nation's principal conservation agency, the Department of the Interior has responsibility for most of our nationally owned public lands and natural resources. This includes fostering the wisest use of our land and water resources, protecting our fish and wildlife, preserving the environmental and cultural values of our national parks and historical places, and providing for the enjoy- ment of life through outdoor recreation. The Department assesses our energy and mineral resources and works to assure that their development is in the best interests of all our people. The Depart- ment also has a major responsibility for American Indian reservation communities and for people who live in island territories under U.S. administration.
U.S. DEPARTMENT OF THE INTERIOR FlSH AND WILDLIFE SERVICE
TAKE PRIDE in America
UNITED STATES DEPARTMENT OF THE INTERIOR
FlSH AND WILDLIFE SERVICE National Wetlands Research Center NASA-Slidell Computer Complex
101 0 Gause Boulevard Slidell, LA 70458
M A Q E AND RE8 PAID ua DEPARTMENT OF THE INTERIOR
INT-421
OFFICIAL BUSINESS PENALTY FOR PRIVATE USE, $300