genomic characterization of felis catus papillomavirus-3: a novel papillomavirus detected in a...
TRANSCRIPT

GeA
in
Joha Colb Bio
1. I
stra(Beinflimphigtha
Veterinary Microbiology 165 (2013) 319–325
A R
Artic
Rece
Rece
Acce
Keyw
Cat
Papi
Bow
Tau
Squ
Skin
*
Palm
fax:
037
http
nomic characterization of Felis catus papillomavirus-3:novel papillomavirus detected in a feline Bowenoid
situ carcinoma
n S. Munday a,*, Magda Dunowska a, Simon F. Hills a, Rebecca E. Laurie b
lege of Sciences, Massey University, Palmerston North, New Zealand
chemistry Department, Otago University, 710 Cumberland Street, Dunedin, New Zealand
ntroduction
Papillomaviruses (PVs) are small circular double-nded DNA viruses that generally infect epitheliumrnard et al., 2010). Papillomaviruses have the ability touence cell growth and differentiation and some areortant causes of cancer. Papillomaviruses are typically
hly host specific and have been detected in most speciest have been extensively studied (Bernard et al., 2010;
Joh et al., 2011). Papillomaviruses are classified into generabased on the sequence of the highly conserved ORF L1
(Bernard et al., 2010).Prior to the present study, two PVs from domestic cats
had been fully sequenced and classified. These viruseswere designated as Felis domesticus PVs (FdPVs). Howeverthe correct taxonomic name for domestic cats is Felis
catus suggesting the most correct taxonomic names forPVs that infect the domestic cat is FcaPV. In the presentreport, PV sequences that have been previously describedare referred to as FdPV while the newly reported virus isdesignated FcaPV. FdPV-1 is a lambda-PV that is closelyrelated to canine PV (CPV)-1 (Tachezy et al., 2002). ThisPV was detected in a cutaneous viral plaque; however, ithas rarely been detected subsequently and appears to be
T I C L E I N F O
le history:
ived 18 October 2012
ived in revised form 27 March 2013
pted 7 April 2013
ords:
llomavirus
enoid in situ carcinoma
papillomavirus
amous cell carcinoma
A B S T R A C T
There is increasing evidence that papillomaviruses (PVs) may cause skin cancer in cats.
Neoplasms most frequently contain Felis domesticus PV type 2 (FdPV-2) DNA, but other PV
DNA sequences have also been detected suggesting multiple PVs could cause disease. One
of these sequences, FdPV-MY2, was previously detected in 5 of a series of 70 feline skin
cancers. The aim was to determine the genome sequence of this PV. Using the circular
nature of PV DNA, ‘outward facing’ primers specific for FdPV-MY2 were designed and
amplified a 7300 bp length of DNA from a feline Bowenoid in situ carcinoma (BISC) that
showed microscopic evidence of a viral etiology and tested positive for FdPV-MY2 DNA.
The PCR product was sequenced using next generation sequencing technology. The full
genomic sequence of the virus, comprising 7583 bp, was assembled and analyzed. As this
is the third PV from a domestic cat, the virus was designated Felis catus PV type 3 (FcaPV-3).
Consistent with other PVs, the putative coding regions of FcaPV-3 were predicted to
produce 6 early proteins and 2 late ones. Classification was difficult as the virus contained
over 60% nucleotide similarity within the ORF L1 with PVs from 3 different genera.
However, based on phylogenetic analysis of ORF L1, FcaPV-3 was most closely related to
the tau-PVs CPV-2 and CPV-7. As FcaPV-3 has over 60% nucleotide similarity with the ORF
L1 of both tau-PVs, it is proposed that FcaPV-3 is classified in the genus Taupapillomavirus
and is the first non-canine PV in this genus.
� 2013 Elsevier B.V. All rights reserved.
Corresponding author at: Pathobiology, IVABS, Massey University,
erston North, New Zealand. Tel.: +64 6 356 9099;
+64 6 350 5714.
E-mail address: [email protected] (J.S. Munday).
Contents lists available at SciVerse ScienceDirect
Veterinary Microbiology
jo u rn al ho m epag e: ww w.els evier .c o m/lo cat e/vetmic
8-1135/$ – see front matter � 2013 Elsevier B.V. All rights reserved.
://dx.doi.org/10.1016/j.vetmic.2013.04.006

J.S. Munday et al. / Veterinary Microbiology 165 (2013) 319–325320
an uncommon cause of skin disease in cats. The fullsequence of FdPV-2 was published in 2009 and this virushas been classified in the genus Dyothetapapillomavirus
(Bernard et al., 2010; Lange et al., 2009b). This PV can bedetected in most feline cutaneous viral plaques andBowenoid in situ carcinomas (BISCs) and is thought to bethe cause of these pre-neoplastic lesions (Lange et al.,2009b; Munday et al., 2007; Munday and Peters-Kennedy, 2010). Additionally, FdPV-2 DNA was detectedmore frequently in feline SCCs than in non-neoplasticsamples of skin, suggesting a possible role in thedevelopment of these common skin cancers (Mundayet al., 2011; Munday et al., 2008). While FdPV-2 is the PVthat has been detected most frequently in feline BISCsand SCCs, short sequences of DNA from other PVs havealso been amplified from these lesions (Munday et al.,2011; O’Neill et al., 2011), including a novel PV sequencethat was designated FdPV-MY2. This PV sequence wasdetected in 5 of 70 feline cutaneous SCCs, suggesting apossible role of FdPV-MY2 in the development of cancer(Munday et al., 2011). In the present report we describethe full genomic sequence of the PV that contains theFdPV-MY2 sequence. This PV was amplified from a catwith multiple BISCs. As this is the third fully sequencedpapillomavirus that has been amplified from a domesticcat, it was designated FcaPV-3.
2. Materials and methods
2.1. Initial case summary and sample collection
In March 2010 a sphinx cat was observed to havemultiple slowly-enlarging hairless cutaneous plaques.Histology of one plaque revealed dysplasia that involvedthe full thickness of the epidermis that extended into hairfollicles and included groups of cells containing loss ofnuclear polarity (wind-blown appearance). There was nopenetration of the basement membrane by the dysplasticcells. Numerous enlarged keratinocytes with gray-bluecytoplasm and keratinocytes with small shrunken nucleithat were surrounded by a clear halo (koilocytes) werevisible within the lesion. These cytopathic changes areconsistently observed in cutaneous viral plaques in catsand are thought to be caused by PV infection (Mundayand Peters-Kennedy, 2010). Immunohistochemistryusing antibodies against p16CDKN2A protein (p16)revealed marked intranuclear and intracytoplasmicimmunostaining. Treatment using imiquimod cream(Aldara 5%, Meda AB, Solna, Sweeden) and a shampoocontaining 2% chlorhexidine and 2% miconazole nitrate(Dermcare Malaseb Shampoo, Pfizer Animal Health,Auckland, New Zealand) appeared to stabilize the lesionsand no further progression was observed. Six monthsafter the initial diagnosis of BISC, skin swabs were takenfor further evaluation of the PVs present. The swabs werecollected by rubbing a saline-soaked cotton bud onlesions on the right thorax, right neck, right flank,periocular region, chin, left eye, left shoulder, and dorsalneck. The swabs were stored in saline and the DNA wasextracted the day after the swabs were taken. At the timeof writing, the skin lesions have remained static over the
last 2 years. Interestingly, the cat shared a householdwith another sphinx cat that was euthanatized afterdeveloping multiple cutaneous SCCs. Neither cat wasallowed outdoors.
2.2. PCR and DNA sequencing
DNA was extracted from the formalin-fixed tissuesamples of the BISC and the skin swabs using a DNeasytissue kit (Qiagen GmbH, Hilden, Germany) according tomanufacturer’s instructions. Identification of PV sequencesin DNA extracted from the BISC was performed using theFAP59/64 and MY09/11 consensus primers and the primersets JMPF/R and JMY2F/R that were specifically designed toamplify sequences of FdPV-2 and FdPV-MY2, respectively.Only the JMPF/R and FdPV-MY2 specific primers were usedto amplify DNA from the swabs. Reaction conditions forthese primers sets have been previously reported (Mundayet al., 2011; Munday et al., 2008). The identity of the PCRproducts was confirmed by sequencing.
DNA extracted from a swab of a cutaneous lesion onthe right side of the neck was used to sequence FcaPV-3. Inorder to amplify the full genomic sequence of the novelvirus, ‘outward facing’ primers (JMY2InvF: 50-GAG GTTTCT TGC ATT TTG CAG C-30 and JMY2InvR: 50-GCT TGGTGT TTT GAG GAT CAA AGG-30) were designed based onthe previously determined FdPV-MY2 sequence. Theinverted primers were used to amplify the circular PVgenome using the Expand Long Template PCR System(Roche Diagnostics, Mannheim, Germany), according tothe manufacturer’s instructions. The resulting PCRproduct comprising approximately 7.3 kbp of the viralgenome was submitted to New Zealand Genomics Ltd. fornext generation sequencing (NGS). Briefly, an Illuminasequencing library was prepared by shearing all sub-mitted DNA into 300–400 bp fragments. Illumina adap-ters containing sequencing primer recognition sites werethen annealed to each DNA fragment using the IlluminaTruSeq DNA sample preparation v2 protocol (IlluminaInc., San Diego, CA). Paired-end 2 � 100 bp sequencing ofthe DNA library was then performed on an IlluminaHiSeq2000 sequencer. To assemble the viral genome, asubset of around 20,000 reads was assembled into a singlecontiguous sequence, which was further assembled withthe short PCR product amplified with the JMY2F/R primerset. All assemblies were performed using Geneious Prosoftware (Drummond et al., 2010). The NGS assembly wasconfirmed by amplification and end-sequencing of anoverlapping set of PCR products using DNA extracted fromthe same swab as a template (primer sequences availableon request).
2.3. DNA and protein sequence analysis
The putative coding regions in the PV sequence werepredicted using FGENESV0 (http://linux1.softberry.com).The characteristics of the putative viral proteins, thepresence of conserved protein domains and motifs, as wellas the presence of regulatory sequences, were predictedusing Geneious software including the third party plugins(InterProScan).

2.4.
eacwegenalig(httweme(Caamwaaligto cusinoutScieme
2.5.
und
3. R
3.1.
confixeseqdepdesPapsamdeswaDNFdPrigh
3.2.
conORF
Tab
Pred
poin
OR
E1
E2
E4
E5
E6
E7
L1
L2
J.S. Munday et al. / Veterinary Microbiology 165 (2013) 319–325 321
Phylogenetic analysis
Complete genomes of 63 PV representative species fromh of the currently recognized genera (Bernard et al., 2010)re obtained from Genbank. Nucleotide sequences for thees E1, E2, L1, and L2 were extracted and individuallyned using MAFFT (Katoh et al., 2002) in Geneoius v6.05p://www.geneious.com). Pairwise sequence similarities
re calculated from the complete L1 alignment. Align-nts of all 4 genes were further processed using GBlocksstresana, 2000) with default parameters to removebiguous sites. A concatenated alignment of 3013 bps constructed from the four cleaned gene alignments. Allnments were analyzed using jModeltest (Posada, 2008)alculate the best fitting nucleotide substitution modelg AIC. Maximum likelihood reconstruction was carried
using RaxML v7.2.8 (Stamatakis, 2006) on the CIPRESnce Gateway (Miller et al., 2010) using default para-
ters and 1000 bootstrap replicates.
Nucleotide sequence accession number
The sequence of the novel PV was deposited in GenBanker accession number JX972168.
esults
Initial PV DNA amplification
Papillomaviral DNA was amplified using the MY09/11sensus primers from DNA extracted from the formalin-d sample of BISC. Sequencing revealed that this DNAuence was identical to one that had been previouslyosited in GenBank (accession number HM130736) andignated FdPV-MY2 (Munday and Peters-Kennedy, 2010).illomavirus DNA was also amplified from the sameple using the JMY2F/R primers that were specifically
igned to amplify the FdPV-MY2 sequence. No PV DNAs amplified by the FAP59/64 and JMPF/R primers fromA extracted from the BISC. However, both FdPV-2 andV-MY2 sequences were amplified from swabs of thet neck, left shoulder, around the mouth, and right flank.
FcaPV-3 complete gene sequence
The complete genome counts 7583 bp with a GCtent of 46.7%. The ‘‘A’’ in the first ATG codon of the
E6 was assigned position 1 in the sequence.
3.3. Open reading frame organization of FcaPV-3 genes
As shown in Fig. 1, the FcaPV-3 genome was predictedto contain 8 ORFs coding for six early (E) proteins (E1, E2,E4, E5, E6, and E7) and two late (L) proteins (L1 and L2). Thepredicted ORFs and characteristics of their putative proteinproducts are shown in Table 1.
Papillomaviral E6 and E7 proteins bind to cellularproteins and are important in promoting host cell growthand division (Stenlund, 2007). The E6 protein of FcaPV-3contains two conserved zinc-binding domains (CXXC-X29-CXXC, aa 25–62 and 98–135), which are separated by 36amino acids. A single zinc-binding domain is also presentat aa 50–86 within the E7 protein. However, the E7 proteinlacks the retinoblastoma (Rb) protein-binding site(LXCXE), that is present in high-risk oncogenic PVs (Dysonet al., 1989).
The E1 and E2 genes are mainly involved in viralreplication and transcription. The predicted E1 protein ofFcaPV-3 contains two helicase domains (aa1-129 andaa171-597). The C-terminal helicase domain contains theconserved ATP-binding site of an ATP-dependent helicaseGPPDTGKS (aa 439–444) and the RBD-like origin ofreplication binding domain (aa160-307). The C-terminalhelicase domain contains the conserved ATP-binding site
Fig. 1. Schematic genomic organization of Felis catus papillomavirus
type 3.
le 1
icted open reading frames (ORFs) in the genome of Felis catus papillomavirus type 3. nt is nucleotides, aa is amino acids, and pI indicates the isoelectric
t.
F ORF location Length (nt) Length (aa) Molecular mass (kDa) pI
688–2484 1797 599 68.170 5.34
2459–3784 1326 422 49.117 9.64
3045–3539 495 165 17.898 4.47
3845–4498 654 218 24.488 7.86
1–414 414 138 14.853 8.09
411–704 294 98 10.758 4.36
5670–7175 1506 502 56.780 7.87
4111–5652 1542 514 54.571 5.05

J.S. Munday et al. / Veterinary Microbiology 165 (2013) 319–325322
of an ATP-dependent helicase GPPDTGKS (aa 439–444) andthe RBD-like origin of replication binding domain (aa160-307). The FcaPV-3 E1 also contains cyclin A interactionmotifs (RXL) at positions 116, 273, 404, 537, and 596. Thebinding of cyclin/cyclin-dependent kinase complexes to E1has been shown to be important for initiation ofpapillomavirus replication (Ma et al., 1999). Conservedprotein domains identified within the E2 gene include aviral DNA binding domain (aa359-439) and the N-terminaltransactivation domain (aa 1–203). The leucine-zipperdomain present in E2 of some other PVs (Van Doorslaeret al., 2007) was not identified in FcaPV-3.
The FcaPV-3 genome was predicted to encode a 218 aaprotein downstream of E2 that partially overlaps L2. Basedon the position of this predicted ORF within the E2-L2
region, it was annotated as ORF E5 (Bravo and Alonso,2004). The putative ORF E5 of FcaPV-3 shared 51.3% and49.2% identity at the nucleotide level with the E5 of rabbitoral papillomavirus (NC_002232) and CPV-2 (NC_006564)over 228 and 128 bp fragments respectively.
The E4 ORF was identified within the ORF E2 region, butin a different translation frame. It has high proline content(15.2%) as reported for other PVs (Van Doorslaer et al.,2007).
The late region encodes two viral capsid proteins L1 andL2. Both proteins contain high proportion of positivelycharged residues (K and R) in their C-terminal end, whichlikely play a role in the nuclear translocation of L1 and L2during the viral life cycle.
The long control region (LCR) comprises 408 bp (nt7176–7583) between L1 and E6. This region does notcode for any proteins, but regulates viral gene transcrip-tion. The FcaPV-3 LCR contains the expected regulatorysequences, including three polyadenylation sites (nt7176, 7216 and 7220) upstream of a CA dinucleotide (nt7262) and a GT cluster (nt 7245–7261), that have beenshown to be necessary for the processing of the L1 and L2
mRNA (Birnstiel et al., 1985). The TATA box (nt 7316–7321) for a putative promoter at position 7345 was alsoidentified. An E1 binding site (E1BS) was reported torequire the sequence motif ATNGTTNNNAACNAT. Therecognition sequence for E1 binding has been furtherdefined as hexanucleotides 50-ATTGTT -30, which areseparated by three nucleotides (Chen and Stenlund,1998). Some variations to this sequence, particularlytransitions, have only a small effect on E1 binding (Holtand Wilson, 1995). An E1BS was predicted in the FcaPV-3genome at nt positions 7492–7506. This putative E1BS isidentical to that previously described (Chen and Sten-lund, 1998), with the exception of C instead of T in theposition 7506. However, it does not form a palindrome,as described for other papillomaviruses. Three putativeE2BS with a consensus sequence ACCN6GGT (Androphyet al., 1987) were identified at nt positions 7440, 7456and 7533. The relative position of E1 and E2 bindingsites, with E1BS flanked on either side by the E2BS, istypical for the papillomavirus origin of replication(Stenlund, 2007). Unexpectedly, an additional ORF (nt7244–7492) was predicted within the LCR region ofFcaPV-3. No ORF has previously been described in thisposition of a PV genome and it is unknown whether or
not this ORF is expressed during the life cycle of FcaPV-3.As this ORF has not been described in other PVs, it wasnot annotated to the sequence of FcaPV-3 submitted toGenBank.
3.4. Phylogenetic analysis of FcaPV-3
All the alignments (E1, E2, L1, L2, and the concatenatedset) recovered GTR + I + G as the best fitting model ofnucleotide substitution. This result indicates that there areno significant differences in the patterns of DNA evolutionin each of the four genes. Therefore the concatenateddataset was treated as a single partition in subsequentphylogenetic reconstruction.
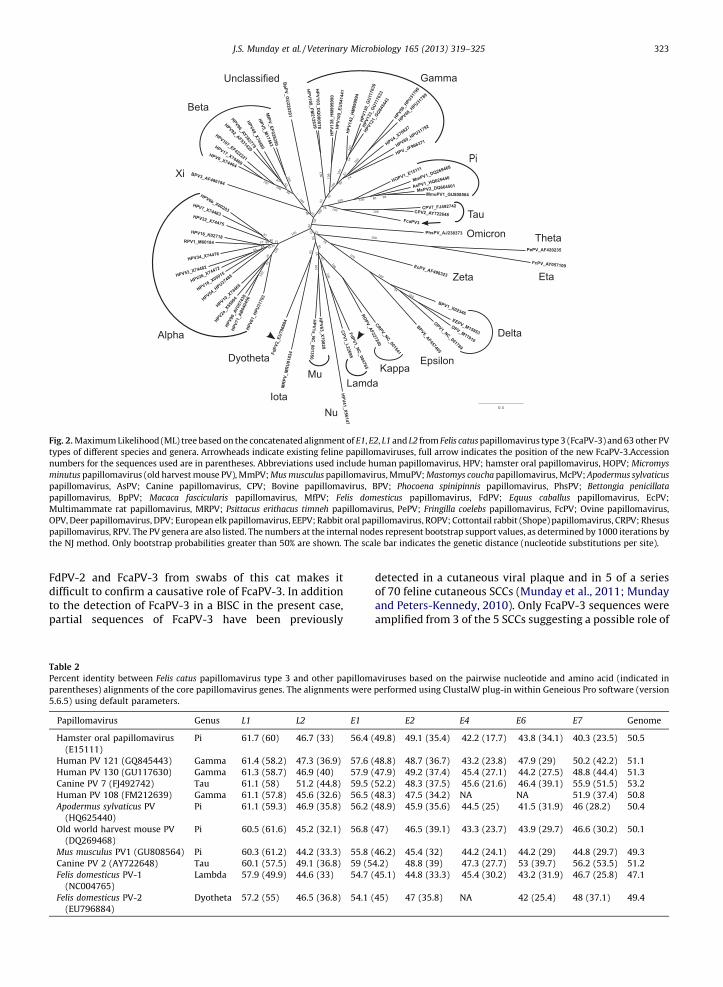
The Maximum Likelihood tree of the 64 papillomavirusspecies based on the concatenated dataset is shown inFig. 2. All of the major papillomavirus clades weresuccessfully recovered, most with high bootstrap support.Most of the low support branches were associated withdeep diverging lineages that are only represented in thetree by single species. These singletons are likely subject toartifacts such as long-branch attraction that leads totopological instability and low bootstrap support. FcaPV-3forms a well-supported clade with CPV-2 and CPV-7,although it represents an early divergence from the caninetaupapillomaviruses.
3.5. FcaPV-3 sequence similarity to other papillomaviruses
The pairwise nucleotide identity scores generated frommultiple alignments ranged from 45.7% for Fringilla coelebs
PV (eta-PV) and Psittacus erithacus timneh PVs (theta-PV) to61.1% for CPV-7 (tau-PV). The identity scores to CPV-2(another tau-PV) and Hamster Oral PV (pi-PV) were 59.5and 59.6% respectively. The % identity scores based onpairwise alignments of core PV ORFs from FcaPV-3 andselected other PVs are shown in Table 2. The ORF L1
nucleotide sequence of FcaPV-3 was more than 60%identical to sequences from viruses belonging to threedifferent genera (Gamma-, Pi- and Taupapillomaviruses).Percent identity between the ORF L1 nucleotide sequencesof FcaPV-3 and the two previously described FdPVs rangedbetween 57 and 58%.
4. Discussion
Papillomaviruses are an established cause of humancancer and there is accumulating evidence that they mayalso influence the development of cancer in domesticanimals (Borzacchiello and Roperto, 2008; Munday andKiupel, 2010). In cats, PVs are thought to cause cutaneousviral plaques and PVs are strongly associated with thedevelopment of BISCs (Lange et al., 2009b; Munday et al.,2008; Munday and Peters-Kennedy, 2010). Multiplestudies by independent researchers have revealed FdPV-2 to be the most commonly detected PV in these pre-neoplastic feline skin lesions (Lange et al., 2009b; Mundayet al., 2007; O’Neill et al., 2011). In the present case, onlyFcaPV-3 DNA sequences were detected in the initial BISCsample. While this could be evidence that FcaPV-3 was thecause of the BISC, the subsequent amplification of both
FdPdiffito tpar
Fig.
type
num
min
pap
pap
Mul
OPV
pap
the
Tab
Perc
pare
5.6.5
Pa
Ha
Hu
Hu
Ca
Hu
Ap
Ol
M
Ca
Fe
Fe
J.S. Munday et al. / Veterinary Microbiology 165 (2013) 319–325 323
V-2 and FcaPV-3 from swabs of this cat makes itcult to confirm a causative role of FcaPV-3. In additionhe detection of FcaPV-3 in a BISC in the present case,tial sequences of FcaPV-3 have been previously
detected in a cutaneous viral plaque and in 5 of a seriesof 70 feline cutaneous SCCs (Munday et al., 2011; Mundayand Peters-Kennedy, 2010). Only FcaPV-3 sequences wereamplified from 3 of the 5 SCCs suggesting a possible role of
2. Maximum Likelihood (ML) tree based on the concatenated alignment of E1, E2, L1 and L2 from Felis catus papillomavirus type 3 (FcaPV-3) and 63 other PV
s of different species and genera. Arrowheads indicate existing feline papillomaviruses, full arrow indicates the position of the new FcaPV-3.Accession
bers for the sequences used are in parentheses. Abbreviations used include human papillomavirus, HPV; hamster oral papillomavirus, HOPV; Micromys
utus papillomavirus (old harvest mouse PV), MmPV; Mus musculus papillomavirus, MmuPV; Mastomys coucha papillomavirus, McPV; Apodermus sylvaticus
illomavirus, AsPV; Canine papillomavirus, CPV; Bovine papillomavirus, BPV; Phocoena spinipinnis papillomavirus, PhsPV; Bettongia penicillata
illomavirus, BpPV; Macaca fascicularis papillomavirus, MfPV; Felis domesticus papillomavirus, FdPV; Equus caballus papillomavirus, EcPV;
timammate rat papillomavirus, MRPV; Psittacus erithacus timneh papillomavirus, PePV; Fringilla coelebs papillomavirus, FcPV; Ovine papillomavirus,
, Deer papillomavirus, DPV; European elk papillomavirus, EEPV; Rabbit oral papillomavirus, ROPV; Cottontail rabbit (Shope) papillomavirus, CRPV; Rhesus
illomavirus, RPV. The PV genera are also listed. The numbers at the internal nodes represent bootstrap support values, as determined by 1000 iterations by
NJ method. Only bootstrap probabilities greater than 50% are shown. The scale bar indicates the genetic distance (nucleotide substitutions per site).
le 2
ent identity between Felis catus papillomavirus type 3 and other papillomaviruses based on the pairwise nucleotide and amino acid (indicated in
ntheses) alignments of the core papillomavirus genes. The alignments were performed using ClustalW plug-in within Geneious Pro software (version
) using default parameters.
pillomavirus Genus L1 L2 E1 E2 E4 E6 E7 Genome
mster oral papillomavirus
(E15111)
Pi 61.7 (60) 46.7 (33) 56.4 (49.8) 49.1 (35.4) 42.2 (17.7) 43.8 (34.1) 40.3 (23.5) 50.5
man PV 121 (GQ845443) Gamma 61.4 (58.2) 47.3 (36.9) 57.6 (48.8) 48.7 (36.7) 43.2 (23.8) 47.9 (29) 50.2 (42.2) 51.1
man PV 130 (GU117630) Gamma 61.3 (58.7) 46.9 (40) 57.9 (47.9) 49.2 (37.4) 45.4 (27.1) 44.2 (27.5) 48.8 (44.4) 51.3
nine PV 7 (FJ492742) Tau 61.1 (58) 51.2 (44.8) 59.5 (52.2) 48.3 (37.5) 45.6 (21.6) 46.4 (39.1) 55.9 (51.5) 53.2
man PV 108 (FM212639) Gamma 61.1 (57.8) 45.6 (32.6) 56.5 (48.3) 47.5 (34.2) NA NA 51.9 (37.4) 50.8
odermus sylvaticus PV
(HQ625440)
Pi 61.1 (59.3) 46.9 (35.8) 56.2 (48.9) 45.9 (35.6) 44.5 (25) 41.5 (31.9) 46 (28.2) 50.4
d world harvest mouse PV
(DQ269468)
Pi 60.5 (61.6) 45.2 (32.1) 56.8 (47) 46.5 (39.1) 43.3 (23.7) 43.9 (29.7) 46.6 (30.2) 50.1
us musculus PV1 (GU808564) Pi 60.3 (61.2) 44.2 (33.3) 55.8 (46.2) 45.4 (32) 44.2 (24.1) 44.2 (29) 44.8 (29.7) 49.3
nine PV 2 (AY722648) Tau 60.1 (57.5) 49.1 (36.8) 59 (54.2) 48.8 (39) 47.3 (27.7) 53 (39.7) 56.2 (53.5) 51.2
lis domesticus PV-1
(NC004765)
Lambda 57.9 (49.9) 44.6 (33) 54.7 (45.1) 44.8 (33.3) 45.4 (30.2) 43.2 (31.9) 46.7 (25.8) 47.1
lis domesticus PV-2 Dyotheta 57.2 (55) 46.5 (36.8) 54.1 (45) 47 (35.8) NA 42 (25.4) 48 (37.1) 49.4
(EU796884)

J.S. Munday et al. / Veterinary Microbiology 165 (2013) 319–325324
this virus in the development of feline skin cancer(Munday et al., 2011).
Human alpha-PVs cause cancer, in part, by PV E7-protein-mediated degradation of retinoblastoma protein(pRb). This protein controls entry of the cell into the S-phaseof replication and degeneration of cellular pRb abolishes animportant cell cycle control mechanism (McLaughlin-Drubin et al., 2012). The normal function of pRb is regulatedby mediators such as p16. When pRb is lost, the cell tries toprevent cell cycling by increasing cellular p16. While theincreased p16 cannot influence cell cycling in the absence ofpRb, the increased p16 can be detected immunohisto-chemically. The loss of pRb and subsequent increase in p16is so consistent in PV-infected cells that p16 immunostain-ing is used as a biomarker for PV-induced cancer in humans(Smeets et al., 2007). In cats, loss of pRb and increased p16 isvisible within all viral plaques and BISCs and in 84% of SCCsthat contain PV DNA (Munday and Aberdein, 2012; Mundayet al., 2011). Increased p16 was visible in the BISC thatcontained FcaPV-3 in the present study and in all 3 SCCs thathad been previously found to contain only FcaPV-3 DNAsequences (Munday et al., 2011). Unlike FdPV-2, FcaPV-3does not have an identifiable pRb binding motif within theE7 gene. While this could suggest that FcaPV-3 does notcause the p16-positive neoplasms, PVs without a pRb-binding motif have been reported to cause cancer inhumans, horses, cattle, and dogs possibly as these PVs havepoorly-defined alternative pRb binding sites within the E7carboxyl-terminal domain (Borzacchiello and Roperto,2008; Wang et al., 2010).
Classification of FcaPV-3 is complicated by someoverlap in the criteria used to classify PVs. The mostrecently proposed classification system states that PVsfrom different genera share less than 60% nucleotidesequence identity within the ORF L1. However, there isflexibility within this classification and some PVs areclassified within the same genus with just 56% identitywhile others with 62% identity are classified in separategenera (Bernard et al., 2010). Based on the pairwisealignments of ORF L1 sequences, FcaPV-3 shared more than60% identity with PVs from 3 different genera (Tau-, Pi-,and Gammapapillomavirus) making it difficult to propose ataxonomic classification based on this feature alone. Thepairwise nucleotide alignment scores generated from themultiple alignments of ORF L1 sequences from 64 PVs alsoproduced ambivalent results. Percent identity values ofFcaPV-3 ORF L1 to selected pi-, gamma-, and tau-PV rangedfrom 57.9 to 61.1%. Because it can be difficult to classifynovel PVs based solely on the ORF L1 sequence, it iscurrently recommended that other factors, such asphylogeny and biological behavior are also taken intoaccount (Bernard et al., 2010). Based on phylogeneticanalysis, FcaPV-3 was most closely related to the tau-PVsCPV-2 and CPV-7, although a branch length separatingFcaPV-3 from CPV-2 and CPV-7 was longer than the branchseparating members of two separate genera Epsilonpapil-
lomavirus and Deltapapillomavirus. This highlights thedifficulties in groupings of the genera that currently havefew representatives.
Both CPV-2 and CPV-7 have been associated with caninepapillomas, and less commonly, cutaneous neoplasia
(Goldschmidt et al., 2006; Lange et al., 2009a). Cutaneouspapillomas are extremely uncommon in cats and there is noevidence of FcaPV-3 involvement in the small number ofpapillomas reported. However, similarly to both CPV-2 andCPV-7, the results of the present study suggest that FcaPV-3could be a rare cause of cutaneous neoplasia. As FcaPV-3 hasover 60% nucleotide sequence identity with the ORF L1 ofboth tau-PVs, clusters with both viruses on the phylogenetictree, and may share some similar behavioral characteristics,it is proposed that FcaPV-3 is classified within theTaupapillomavirus genus. It is noted that, similarly toFcaPV-3, there is evidence that both CPV-2 and CPV-7 areassociated with neoplasia without either containing anyidentifiable pRb binding domain.
An E5 protein was predicted to be encoded by theFcaPV-3 genome However, in contrast to the ORF E5 ofother PVs, the FcaPV-3 E5 ORF overlapped with ORF L2.Additionally, the predicted E5 product was less hydro-phobic (GAVY hydropathicity index of 0.46) than the E5proteins of CPV-2, bovine PV-1 and human PV-16 (GAVYhydropathicity index of 1.98, 1.63 and 1.79, respectively),although its GAVY scores were still higher than thoseexpected of soluble proteins (-0.4) (Kyte and Doolittle,1982). This, together with the presence of a predictedsignal peptide (aa1-23) and trans-membrane domains(aa10-30 and 67–87) at the N-terminal, suggests that theputative E5 product is a membrane bound protein.Whether or not the putative E5 product is expressedduring the viral life cycle and its role remains to beestablished.
Of the three classified feline PVs, two are most closelyrelated to canine PVs with FdPV-1 and FcaPV-3 most closelyrelated to CPV-1 and CPV-7 respectively. The phylogenetictree suggests that FcaPV-3, CPV-2, and CPV-7 may haveshared a common ancestor. The split between FcaPV-3 andthe canine taupapillomaviruses might have happened whenthe Caniformia split from the Feliformia, which would be inagreement with the view that PVs co-evolved with theirhosts (Gottschling et al., 2011; Rector et al., 2007). Thedivergence of the members of Canidae and Felidae wasinitially estimated to be between 38 and 50 million yearsago. Using this time, Tachezy et al. (Tachezy et al., 2002)suggested a mutation rate in the papillomaviruses of 0.73–0.96 � 10�8 substitutions per site per year based on thesequence divergence of FdPV-1 and CPV-1. Evaluation of thedivergence of FcaPV-3 and CPV-7 reveals a similar rate of0.74–0.97 � 10�8 substitutions per site per year.
While this data suggests that the divergence of FdPV-1and CPV-1 and the divergence of FcaPV-3 and CPV-7occurred at a similar time, exactly when this was is difficultto determine. A more recent analysis if the divergencetimes within the order Carnivora suggested that the split ofthe Caniformia from the Feliformia occurred between 62.3and 67.5 million years ago (Nyakatura and Bininda-Emonds, 2012). Recalculating the nucleotide substitutionrate of taupapillomaviruses results in a mutation rate of0.55–0.59 � 10�8 substitutions per site per year. Withoutknowing for certain either the time that the Caniformiasplit from the Feliformia or the true mutation rate of thepapillomaviruses, it is not possible to definitively deter-mine the time of divergence of FcaPV-3 from the canine

tauaccma
5. C
domexpthealthdiseclaswobe
Ack
(Smfor
Ref
And
Bern
Birn
Borz
Brav
Cast
Che
Dru
Dys
Gold
Gott
Holt
Joh,
Kato
J.S. Munday et al. / Veterinary Microbiology 165 (2013) 319–325 325
papillomaviruses. Clearly there is a need to moreurately characterize the rate of evolution in Papillo-viruses, but that is beyond the scope of this study
onclusion
This is the third PV to be fully sequenced from aestic cat. The results of the present, and previous
eriments, suggest that FcaPV-3 may be associated with development of in situ and invasive skin carcinomas,ough the exact role of this PV in development ofase remains uncertain. It is proposed that FcaPV-3 issified within the Taupapillomavirus genus and FcaPV-3
uld represent the third PV in this genus and the first toidentified in a non-canine species.
nowledgements
The authors thank Dave Baumberg BVSc, MANZCVScall animal medicine), Pet Doctors, Dinsdale, Hamiltonassistance with this case.
erences
rophy, E.J., Lowy, D.R., Schiller, J.T., 1987. Bovine papillomavirus E2trans-activating gene product binds to specific sites in papillomavirusDNA. Nature 325, 70–73.ard, H.U., Burk, R.D., Chen, Z., van Doorslaer, K., Hausen, H., de Villiers,
E.M., 2010. Classification of papillomaviruses (PVs) based on 189 PVtypes and proposal of taxonomic amendments. Virology 401, 70–79.stiel, M.L., Busslinger, M., Strub, K., 1985. Transcription terminationand 30 processing: the end is in site! Cell 41, 349–359.acchiello, G., Roperto, F., 2008. Bovine papillomaviruses, papillomas
and cancer in cattle. Vet. Res. 39, 45.o, I.G., Alonso, A., 2004. Mucosal human papillomaviruses encode
four different E5 proteins whose chemistry and phylogeny correlatewith malignant or benign growth. J. Virol. 78, 13613–13626.resana, J., 2000. Selection of conserved blocks from multiple align-ments for their use in phylogenetic analysis. Mol. Biol. Evol. 17, 540–552.n, G., Stenlund, A., 1998. Characterization of the DNA-binding domainof the bovine papillomavirus replication initiator E1. J. Virol. 72,2567–2576.mmond, A.J., Ashton, B., Buxton, S., Cheung, M., Cooper, A., Duran, C.,Field, M., Heled, J., Kearse, M., Markowitz, S., Moir, R., Stones-Havas, S.,Sturrock, S., Thierer, T., Wilson, A., 2010. Geneious v5.3.on, N., Howley, P.M., Munger, K., Harlow, E., 1989. The human papil-loma virus-16 E7 oncoprotein is able to bind to the retinoblastomagene product. Science 243, 934–937.schmidt, M.H., Kennedy, J.S., Kennedy, D.R., Yuan, H., Holt, D.E., Casal,
M.L., Traas, A.M., Mauldin, E.A., Moore, P.F., Henthorn, P.S., Hartnett,B.J., Weinberg, K.I., Schlegel, R., Felsburg, P.J., 2006. Severe papilloma-virus infection progressing to metastatic squamous cell carcinoma inbone marrow-transplanted X-linked SCID dogs. J. Virol. 80, 6621–6628.schling, M., Goker, M., Stamatakis, A., Bininda-Emonds, O.R., Nindl, I.,Bravo, I.G., 2011. Quantifying the phylodynamic forces driving papil-lomavirus evolution. Mol. Biol. Evol. 28, 2101–2113., S.E., Wilson, V.G., 1995. Mutational analysis of the 18-base-pairinverted repeat element at the bovine papillomavirus origin of repli-cation – identification of critical sequences for E1 binding and in-vivoreplication. J. Virol. 69, 6525–6532.
J., Jenson, A.B., King, W., Proctor, M., Ingle, A., Sundberg, J.P., Ghim, S.J.,2011. Genomic analysis of the first laboratory-mouse papillomavirus.J. Gen. Virol. 92, 692–698.h, K., Misawa, K., Kuma K.-i. Miyata, T., 2002. MAFFT: a novel method
for rapid multiple sequence alignment based on fast Fourier trans-form. Nucleic Acids Res. 30, 3059–3066.
Kyte, J., Doolittle, R.F., 1982. A simple method for displaying the hydro-pathic character of a protein. J. Mol. Biol. 157, 105–132.
Lange, C.E., Tobler, K., Ackermann, M., Panakova, L., Thoday, K.L., Favrot, C.,2009a. Three novel canine papillomaviruses support taxonomic cladeformation. J. Gen. Virol. 90, 2615–2621.
Lange, C.E., Tobler, K., Markau, T., Alhaidari, Z., Bornand, V., Stockli, R.,Trussel, M., Ackermann, M., Favrot, C., 2009b. Sequence and classi-fication of FdPV2, a papillomavirus isolated from feline Bowenoid insitu carcinomas. Vet. Microbiol. 137, 60–65.
Ma, T.L., Zou, N.X., Lin, B.Y., Chow, L.T., Harper, J.W., 1999. Interactionbetween cyclin-dependent kinases and human papillomavirus repli-cation-initiation protein E1 is required for efficient viral replication.Proc. Natl. Acad. Sci. U.S.A. 96, 382–387.
McLaughlin-Drubin, M.E., Meyers, J., Munger, K., 2012. Cancer associatedhuman papillomaviruses. Curr. Opin. Virol. 2, 459–466.
Miller, M.A., Pfeiffer, W., Schwartz, T., 2010. Creating the CIPRES ScienceGateway for inference of large phylogenetic trees. In: Gateway Com-puting Environments Workshop (GCE). pp. 1–8.
Munday, J.S., Aberdein, D., 2012. Loss of retinoblastoma protein, but notp53, is associated with the presence of papillomaviral DNA in felineviral plaques, Bowenoid in situ carcinomas, and squamous cell carci-nomas. Vet. Pathol. 49, 538–545.
Munday, J.S., Gibson, I., French, A.F., 2011. Papillomaviral DNA andincreased p16(CDKN2A) protein are frequently present within felinecutaneous squamous cell carcinomas in ultraviolet-protected skin.Vet. Dermatol. 22, 360–366.
Munday, J.S., Kiupel, M., 2010. Papillomavirus-associated cutaneous neo-plasia in mammals. Vet. Pathol. 47, 254–264.
Munday, J.S., Kiupel, M., French, A.F., Howe, L., 2008. Amplification ofpapillomaviral DNA sequences from a high proportion of felinecutaneous in situ and invasive squamous cell carcinomas using anested polymerase chain reaction. Vet. Dermatol. 19, 259–263.
Munday, J.S., Kiupel, M., French, A.F., Howe, L., Squires, R.A., 2007. Detec-tion of papillomaviral sequences in feline Bowenoid in situ carcinomausing consensus primers. Vet. Dermatol. 18, 241–245.
Munday, J.S., Peters-Kennedy, J., 2010. Consistent detection of Felis domes-ticus papillomavirus 2 DNA sequences within feline viral plaques. J.Vet. Diagn. Invest. 22, 946–949.
Nyakatura, K., Bininda-Emonds, O.R., 2012. Updating the evolutionaryhistory of Carnivora (Mammalia): a new species-level supertreecomplete with divergence time estimates. BMC Biol. 10, 12.
O’Neill, S.H., Newkirk, K.M., Anis, E.A., Brahmbhatt, R., Frank, L.A., Kania,S.A., 2011. Detection of human papillomavirus DNA in feline pre-malignant and invasive squamous cell carcinoma. Vet. Dermatol. 22,68–74.
Posada, D., 2008. jModelTest: phylogenetic model averaging. Mol. Biol.Evol. 25, 1253–1256.
Rector, A., Lemey, P., Tachezy, R., Mostmans, S., Ghim, S.J., Van Doorslaer,K., Roelke, M., Bush, M., Montali, R.J., Joslin, J., Burk, R.D., Jenson, A.B.,Sundberg, J.P., Shapiro, B., Van Ranst, M., 2007. Ancient papilloma-virus-host co-speciation in Felidae. Genome Biol. 8, R57.
Smeets, S.J., Hesselink, A.T., Speel, E.J., Haesevoets, A., Snijders, P.J.,Pawlita, M., Meijer, C.J., Braakhuis, B.J., Leemans, C.R., Brakenhoff,R.H., 2007. A novel algorithm for reliable detection of human papil-lomavirus in paraffin embedded head and neck cancer specimen. Int.J. Cancer 121, 2465–2472.
Stamatakis, A., 2006. RAxML-VI-HPC: maximum likelihood-based phylo-genetic analyses with thousands of taxa and mixed models. Bioinfor-matics 22, 2688–2690.
Stenlund, A., 2007. DNA replication of papillomaviruses. In: Garcea, R.L.,DiMaio, D. (Eds.), The Papillomaviruses. Springer, New York, pp. 145–175.
Tachezy, R., Duson, G., Rector, A., Jenson, A.B., Sundberg, J.P., Van Ranst, M.,2002. Cloning and genomic characterization of Felis domesticus papil-lomavirus type 1. Virology 301, 313–321.
Van Doorslaer, K., Rector, A., Jenson, A.B., Sundberg, J.P., Van Ranst, M.,Ghim, S.J., 2007. Complete genomic characterization of a murinepapillomavirus isolated from papillomatous lesions of a Europeanharvest mouse (Micromys minutus). J. Gen. Virol. 88, 1484–1488.
Wang, J., Zhou, D., Prabhu, A., Schlegel, R., Yuan, H., 2010. The caninepapillomavirus and gamma HPV E7 proteins use an alternativedomain to bind and destabilize the retinoblastoma protein. PLoSPathog. 6, e1001089.